细胞周期的调控 南通大学基础医学院 陈 莉
DESCRIPTION
细胞周期的调控 南通大学基础医学院 陈 莉. 细胞周期的准确调控对生物的生存、繁殖、发育和遗传均是十分重要的。 简单生物调控细胞周期主要是为了适应自然环境,以便根据环境状况调节繁殖速度,以保证物种的繁衍。 复杂生物的细胞则需面对来自自然环境和其他细胞、组织的信号,作出正确的应答,以保证组织、器官和个体的形成、生长以及创伤愈合等过程能正常进行,需要更为精细的细胞周期调控机制。. 第一节 细胞周期的基本概念 - PowerPoint PPT PresentationTRANSCRIPT

细胞周期的调控
南通大学基础医学院
陈 莉

细胞周期的准确调控对生物的生存、繁殖、发育和遗传均是十分重要的。
简单生物调控细胞周期主要是为了适应自然环境,以便根据环境状况调节繁殖速度,以保证物种的繁衍。
复杂生物的细胞则需面对来自自然环境和其他细胞、组织的信号,作出正确的应答,以保证组织、器官和个体的形成、生长以及创伤愈合等过程能正常进行,需要更为精细的细胞周期调控机制。

第一节 细胞周期的基本概念
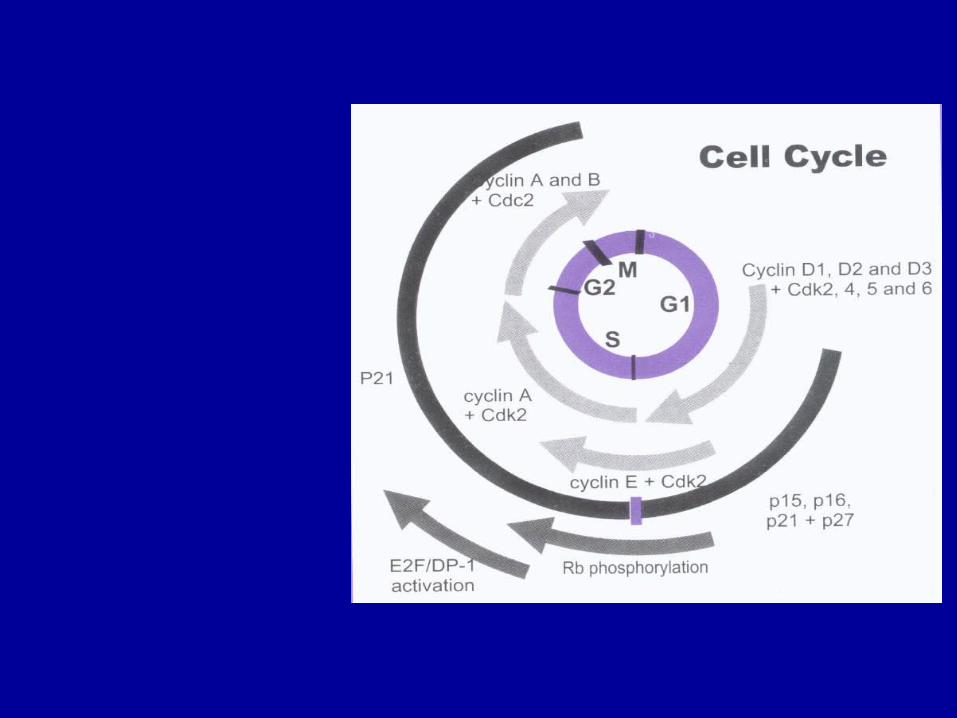
细胞周期的基本任务是保证 S 期的 DNA 复制和 M 期有同等的染色体分布到两个子细胞中去。在 DNA 合成期 (S 期 ) 和有丝分裂期 (M 期 ) 之间,M 期和下一个 S 期之间,分别存在着两个间期 (Gap , G) , S 期之前是第一间期 (G1 期 ) , S 期和M 期之间为第二间期 (G2 期 ) 。细胞生长、分裂时,依次经过 G1 、 S 、 G2 、 M 期而一分为二,周而复始,故称为细胞分裂周期 (cell division cycle) 。

当细胞中 DNA 损伤时,激活凋亡基因,使进入增殖周期的细胞停留在 G1→S 期,容许细胞修复 DNA 避免突变。如果损伤严重时,细胞则走向凋亡。

细胞周期素依赖激酶
(cyclin-dependent kinase , CDK)
细胞周期素 (cyclin)
细胞周期蛋白依赖性激酶抑制剂
( CDK inhibitory protein CKIS )
第二节 细胞周期调控中的重要元素
一、细胞周期蛋白( cyclin )
1953 年霍华德等人首先提出细胞分化是通过细胞周期完成的理论;
1983 年, Evans 等首次在海洋无脊椎动物中发现一组蛋白质呈周期性出现,并调节细胞的生长,其被确定为细胞周期蛋白。 1988 年科学家们发现细胞周期调节蛋白能与细胞分化周期编码蛋白结合并激活相应的蛋白激酶,从而促进细胞分裂。

至少发现有 11 种不同的 cyclin ,分别为 A 、 B1 、 B2 、 C 、 D1 、 D2 、 D3 、E 、 F 、 G 和 H 。其中 8 种主要的 cyclin己被分离。根据 cyclin 调控细胞周期时相的不同,可分为 G1 期和 M 期两大类。

(一) G1 期胞周期蛋白( G1—cyclin )
作用在 G1 期或 G1/S 交界期,启动细胞周期和促进 DNA 合成的 cyclin , G1 期是增殖细胞唯一能接受从外界传入的增殖或抑制增殖信号的时期。
1 cyclin D :
cyclin D1 , cyclinD2 , cyclin D3
2 cyclin C
3 cyclin E : cyclin E 1 , cyclin E 2

1 、 cyclin D
cyclin D 首先在酵母菌中被发现,它能激活 CDK6,驱动细胞通过 START。它有 3 个亚型,包括 D1 、 D2 、 D3 ,具组织特异性。cyclin D1 与 cyclin D2功能相似,都在酵母子细胞中起作用, cyclin D3 在酵母母细胞中起作用。

在细胞周期的调节中 cyclinDl 是一个比其它 cyclins 更加敏感的指标。
cyclin D1 的编码基因位于 11q13上,全长约 15kb ,与其他周期素相比最小,主要是因为其 N末端缺少一个“降解盒”片段 ,该蛋白半衰期很短,不足 25min 。
在有生长因子的情况下, cyclin D1 在细胞周期中首先被合成,并于 G1 中期合成达到高峰, cyclln D1 的功能主要是促进细胞增殖,是 G1 期细胞增殖信号的关键蛋白质,被视为癌基因,其过度表达可致细胞增殖失控而恶性化。
cyclinD2 的编码基因位于 12p13 ,称为 CCND2 ,在正常的二倍体细胞及 Rb阳性肿瘤细胞中 cyclin D2 的表达呈波动状态,其峰值在 G1晚期。 给 G1 期细胞微量注射 cyclinD2抗体,可使表达 cyclin D2 的淋巴细胞停滞在 G1期,说明 cyclin D2 是细胞从 G1 期向 S 期转移所必须的。

cyclin D3 的编码基因位于染色体 6p21 ,称为 CCND3 。
正常和恶性组织中未见 cyclinD3 基因异常及其蛋白的过度表达。 目前认为 cyclinD3似乎不直接反映恶性度,而是肿瘤发展到晚期的结果。

2 、 cyclin C
cyclin C 与所有 cyclin 的同源性最低,主要在果蝇及人类细胞中发现,它与其他 G1-cyclin 不同的是其mRNA 和蛋白质水平在 G1早期达最高,可能在 G1早期发挥作用。

3 、 cyclin E
cyclin E 在 cyclin D 之后出现,于 G1/ S转化过程中表达,人类 cyclin E 基因定位于染色体 19q12-q13 。cyclin E 中 1/ 3段为高度保守区,称为周期蛋白盒,此为 CDKs 结合所必须。在 G1晚期发挥正调控细胞周期的作用。 cyclin E 蛋白的 C端存在 PEST 序列 ( 一个富含脯氨酸( P)、谷氨酸( E )、天冬氨酸( S )、丝氨酸和苏氨酸( T)残基的 PEST 序列,在蛋白质转化和降解中起作用 ) 。
cyclin E 基因及其产物的表达在细胞周期的 G1 中期上升,至 G1晚期或 S早期达高峰,然后经与“PEST”序列有关的蛋白水解或与 S 期激酶相关蛋白 -2 ( S-phase kinase-associated protein SKP2 )泛素路径降解而迅速下降。 缺乏 SKP2 的细胞表现 cyclin E 蛋白降解不足并不断积累。

( 二 ) M 期 细 胞 周 期 蛋 白 ( M-cyclin ) 在 G2/M 交界期诱导细胞分裂的 cyclin 。
1 、 cyclin A
cyclin A 在 cyclin E 之后很快表达。 cyclin A 是 G1期向 S 期转移的限速因素,也可促进细胞从 G2 期向 M期的转化。它由 CCNA 基因编码。
2 、 cyclin B
cyclin B 是有丝分裂蛋白激酶的一个亚单位,能促进G2 期向 M 期的过渡。哺乳动物 cyclin B 在 S晚期合成。
cyclin A 、 cyclin B 在 M 期通过泛素途径降解,这是细胞脱离有丝分裂所必须。

cyclin A 与 cyclin B 之间存在多种差异( 1 )周期积累方式不同, cyclin A含量在 S 期及G2 期初最高, cyclin B 在 G2 期末含量最高;
( 2 )结合的催化亚基不同, cyclin A 与 p33cdc2结合, cyclin B 与 p34cdc2 结合;
( 3 )功能不同, cyclin A 在 S 期发挥作用,与 DNA 的复制完成有关, cyclin B 在 G2/ M 交界期发挥作用,诱发细胞分裂;
( 4)对细胞分裂的影响不同, cyclin B持续升高可使细胞停滞于分裂期,而 cyclin A 的持续升高并不影响细胞分裂的完成。
二、细胞周期素依赖激酶 (cyclin-dependent kinase , CDK)
CDK 是一类重要的丝氨酸 /苏氨酸蛋白激酶,包括 CDK1—7种。 CDK 的主要生物学作用是启动 DNA 的复制和诱发细胞的有丝分裂,以复合物形式出现。
催化亚基 CDK
复合物
调节亚基 cyclin 。 ATP的结合部位
该酶的活性部位
CDK 有三个重要的功能区 调节亚基的结合部位
P13sucl 的结合部位 ( P13sucl 能抑制激酶的活性,阻止细胞进入或退出 M 期)

三、细胞周期蛋白依赖性激酶抑制剂( CDK inhibitory protein CKIS )
CKIS 是 CDK 抑制蛋白,通过竞争性地抑制 cyclin 或 cyclin—CDK 复合物,导致 cyclin 生物学功能丧失;对细胞生长起负调控作用。
G1 期 CKIS 有 2 个蛋白家族:
(一) INKs : p15、 p16、 p18 、 p19
是 cDK4和 cDK6的特异性抑制物。
(二) CIP/ KIPs : p21 、 P27、 p57等,
抑制各种 cyclin-CDK 复合物,阻止 CDK激
酶的激活,或阻止活化的 CDK 激酶活性。

1 、 P16
p16INK4 位于染色体 9p21 ,又称多肿瘤抑制基因(multiple tumor suppressor MTSI ) ,是 CDK4的特异性抑制物,可与 cyclin D竞争与 CDK4或 CDK6的结合,抑制 CDK4对细胞生长分裂的正向作用,参与抑制细胞周期 G1/ S 的转化。 p16在缺乏功能性 Rb的细胞中水平上升,提示 Rb 可能抑制 p16的表达,同时 Rb刺激 cyclin D 的表达。
2 、 p15INKB
p15INKB位于 9号染色体紧邻 p16的区域,它与 p16一样属抑癌基因。

P16
CDK4 cyclinD1
一
P cdk 介导的磷酸化
Rb Rb P
一
G1 S
DNA转录

3 、 P27
P27可能是最直接地影响 G1/S 期限制位点的调控。广泛抑制 cyclin—CDK 复合物。
正常情况下 P27在 G0 / G1 时表达增高,进入 S期后表达下降。其基因定位于染色体 12p13.1 及 12p13.2处 ,人的 p27cDNA全长 594bp ,编码 198个氨基酸,是高度保守的蛋白分子,在人、鼠、貂中 p27的氨基酸主序列有 90%同源性,其 C末端均含有一个双枝核定位信号。其 N末端介导抑制 CDK ,约 12-87 氨基酸主序列与 p21 同源。 P27与p21 在 N端序列上有 42%相同,但是 P27 介导抑制 CDK 的区域与 p21 不尽相同。

p27 还参与对细胞分化的调控:
同 p21 一样它可诱导未成熟细胞进行分化;
p27 也可诱导肿瘤细胞分化(如外源性 p27可诱导原巨核细胞白血病细胞分化);
p27不能诱导成熟正常细胞的衰老。
p27 表达水平受多种因素调控,如有丝分裂原、抗增殖信号因子、细胞因子、癌基因子及接触抑制等。
TGF-β和接触抑制能共同调控转录 p27和 p15,其负调节信息的共同通路是抑制 CDK 和 G1—cyclin功能,发挥 CKI 抑制作用。
TGF-β对 p27 表达的影响是双相的,在大多数细胞中, TGF-β可诱导 p27的表达,但是在正常垂体前部和垂体瘤细胞中, TGF-β可下调 p27mRNA 及蛋白的表达。
PDGF 、 EGF 等也可下调 p27的表达。
p27对细胞周期的调控主要依赖于其蛋白表达水平,而非基因突变。 p27的表达下降或缺失会引起基因组不稳定,甚至导致肿瘤发生。

4 、 p21
p21 基因位于染色体 6p21. 2 ,第 17-71氨基酸含有 cyclin 结合抑制区。 p21 可能阻碍细胞进入 S 期;能抑制 SAPK(stress-activated protein kinase) ,参与细胞应激状态时的信号转导级联系统的调节。


第三节 细胞周期调控中各元素间的相互作用
细胞周期的调控可分为外源和内源性调控 ,
外源性调控主要是细胞因子以及其它外界刺
激引起;
内源性调控主要是通过 Cyclin—CDK—CDI的网络调控来实现。

各种细胞周期蛋白随特定细胞时相而出现。
G1早期, cyclinD表达并与 CDK2 或 CDK4结合,成为 始动细胞周期的启动子;
G1晚期、进入 S早期后 cyclinE表达,并与 CDK2 结合,推动细胞进入 S 期;
进入 S 期后, cyclin A表达, cyclinD 、 cyclin E降解;
S晚期、 G2早期, cyclIin A 、 cyclin B表达,并与 cdc2 结合,促进细胞进入 M 期。

cyclin A 和 CDK2 相结合可以调节 S 期进入 G2期;
cyclin Bl—2 可与 CDKl 结合并在 G2/ M转化期间活性达到最高峰;
与 cyclin C匹配的 CDK 及其酶解底物尚不清楚;
cyclinH 与 cyclinC 有较高的同源序列,可以和 CDK7装配成全酶对细胞周期各阶段行使调节作用。

细胞转录因子( E2F )
在许多 DNA 合成基因和细胞生长调控基因的启动子中均含有 E2F 的位点, E2F 可以直接活化这些基因,启动 DNA 合成使细胞进入 S 期。
在 E2F 基因活化转录功能区内有一段 18 个氨基酸的序列可与 Rb 结合, Rb 通过与 E2F功能区的结合遮盖其功能区,抑制其活性转录功能,抑制 DNA 合成。
E1A 的介入,释放出更多的游离 E2F ,影响着一些与转化表型有关的特定基因表达。

Rb 基因 Rb 基因位于人类染色体 13q14,其转录产物 Rb蛋白是主要的转录信号连接物,在细胞周期中起制动器功能。
它能与转录因子 E2F 结合并阻止相应基因转录表达,从而抑制细胞生长。
cyclin D 是 Rb 调节细胞周期的基础。 cyclin D1-CDK4复合物可看做 G1 期 Rb 蛋白激酶,它能结合 Rb 的N末端,磷酸化 Rb 蛋白,使转录因子释放,导致 G1/ S转化。
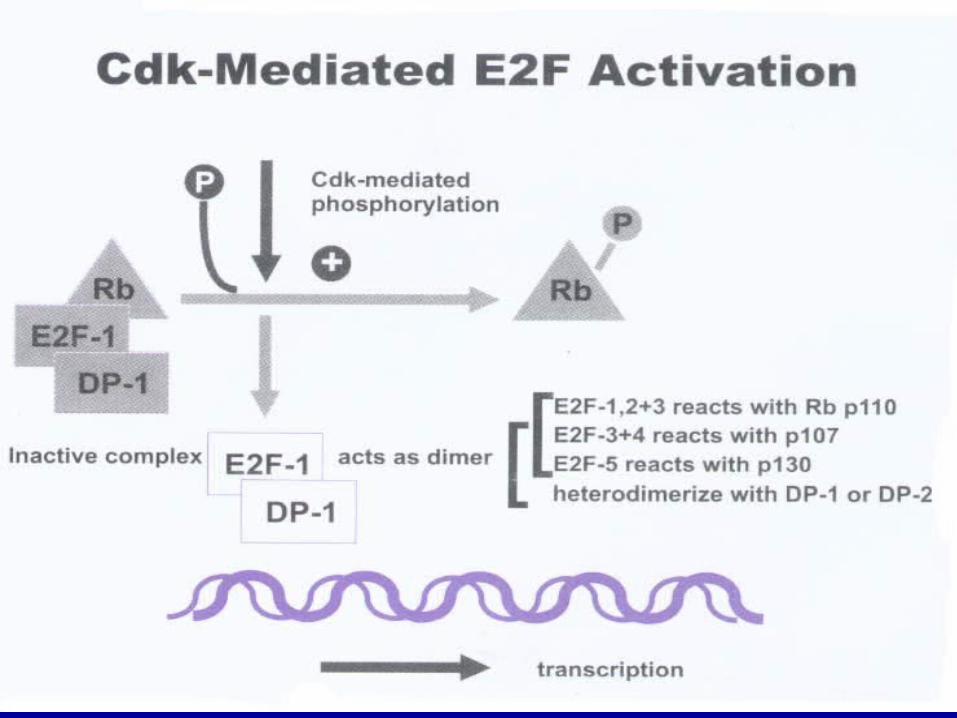
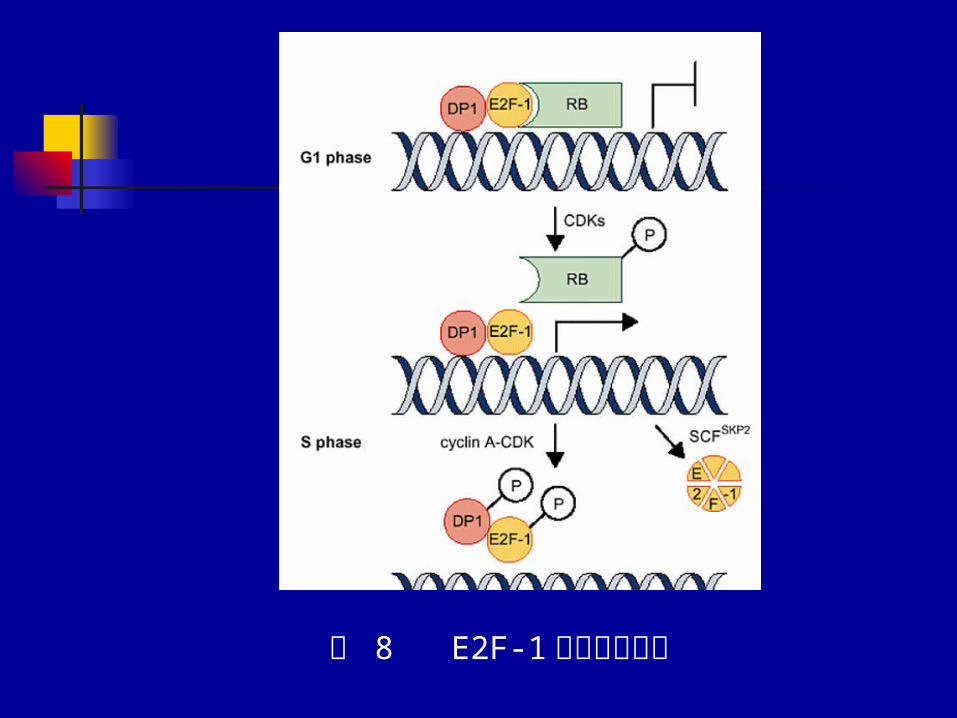
图 8 E2F-1功能调节机制

cyclin D1 与 Rb 的功能是相互依赖的
低磷酸化的 Rb还可刺激 cyclin D1 的转录,使其合成增加,并活化再导致Rb磷酸化,这样形成负反馈环以调节 cyclin D1 的表达。 CyclinE-CDK2 的作用是通过正反馈以促进Rb磷酸化和 E2F 的释放。

P21 和 p27 抑制 cyclin-CDK 复合物的负性调节作用
P21 结合并抑制多种 cyclin-CDK 复合物,负性调节 CDK功能,实验证明,正常细胞多数 cyclin-CDK复合物都与 p21 结合,而多数转化细胞中则不结合。
P21 是 P53 作用的靶点, p21 启动子含有 P53 结合位点。 G1 期 DNA 损伤可激活 p53 ,诱导P21转录,导致 cyclin D-CDK4和 cyclin E-CDK2 抑制,从而阻止细胞进入 S 期,使损伤 DNA得到修复。 p21在 p53介导的 DNA 损伤所致的 G1 期停滞中起重要作用。

P53 P16
介导 抑制 CDK4
p21 CDK4 cyclinD1
抑制 CDK4 活化 CDK4
P cdk 介导的磷酸化
Rb Rb P
E2F
无活性复合物
E2F 启动
DNA转录
G1 S
DNA转录

CDK 活性状态是以磷酸化形式存在的。
CAK ( CDK 激活酶)可诱导 CDK磷酸化,而 p27 通过与 CDK 亚单位的结合,使 CAK 不能与 CDK直接发生作用。非活化的 CDK 不能使 Rb 蛋白磷酸化,使细胞停留在 G1 期,对细胞周期进行负调控。
P27 还可阻止 Rb 蛋白磷酸化,其过度表达能抑制细胞进入 G1 期。
p27在调节细胞进入和退出 M 期中起重要作用,抗丝裂原环境中细胞生长停滞与 p27—CDK2 复合物的量相关。


细胞周期阶段 cyclin-CDK 复合物 抑制因子
P16 P21 P27
G1 cyclinD1-CDK4 - - -
G2/S cyclinE-CDK2 - + -
S cyclinA-CDK2 - + -
G2/M cyclin-CDK2 - + -

第四节 细胞周期检测点的调控 细胞在长期的进化过程中发展出了一套保证细胞周期中 DNA 复制和染色体分配质量的检查机制 , 通常被称为细胞周期检测点 (check point) 又称为限制点 (restriction point) 。这是一类负反馈调节机制。当细胞周期进程中出现异常事件 , 如 DNA 损伤或 DNA 复制受阻 ,这类调节机制就被激活 , 及时地中断细胞周期的运行 , 待细胞修复或排除了故障后 ,细胞周期才能恢复运转。保证了在细胞周期中上一期事件完成以后才开始下一期的事件。

细胞周期检测点: G1-S 期检测点、 S 期检测点、
G2 期检测点、 M 期检测点。
检测点的组成:发现或传感 (detect 或 sensor) 、
制动或扣留 (stop 或 arrest) 、
修复 (repair) 、
继续分裂或死亡。
检测点功能缺陷会导致基因突变和染色体结构异常细胞增殖 , 导致肿瘤发生 。

检测点的任何一部分出了问题,如发现不了 DNA损伤 ( 如 ATM 突变 ) 、不能使细胞周期停下来 ( 如 p53突变 ) 、 DNA 修复错误 ( 如 MLHl/ PSM 突变 ) 、决定错误 ( 如 BCL—2 突变 ) 等都会导致遗传的不稳定性、( 基因 ) 受损细胞的存活和复制或细胞遗传物质的改编(adaptation) 。

细胞周期检测点的作用细胞周期检测点 (checkpoints)构成了 DNA 修复的完整元件。
检测点通过延缓细胞周期的进展,为 DNA 复制前的修复、基因组的复制、有丝分裂及基因组的分离提供更多的时间。
检测点功能的丢失或减弱可能通过降低 DNA 复制效率来增加和诱导基因突变和染色体畸变。在某些遗传性癌症和细胞转化早期中,已经观察到检测点调控的缺失,后者可能导致遗传失稳态,促使向新生物转化。


一、 G1-S 期检测点 G1-S 期检测点是最重要的检测点。细胞在该检测点对各类生长因子、分裂原以及 DNA 损伤等复杂的细胞内外信号进行整合和传递 ,决定细胞是否进行分裂、发生凋亡或是进入 G0 期。
细胞顺利完成其周期需经过若干检测点,其中最重要的控制点是 G1晚期的 START, START调节失灵,将导致细胞越过正常的程序限制进入 s 期,并允许细胞复制未修复的突变 DNA ,从而积累形成肿瘤表型的基因改变。
G1-S 期检测点缺陷导致肿瘤的原因主要是 p53 缺失和 cyclinD1上调。
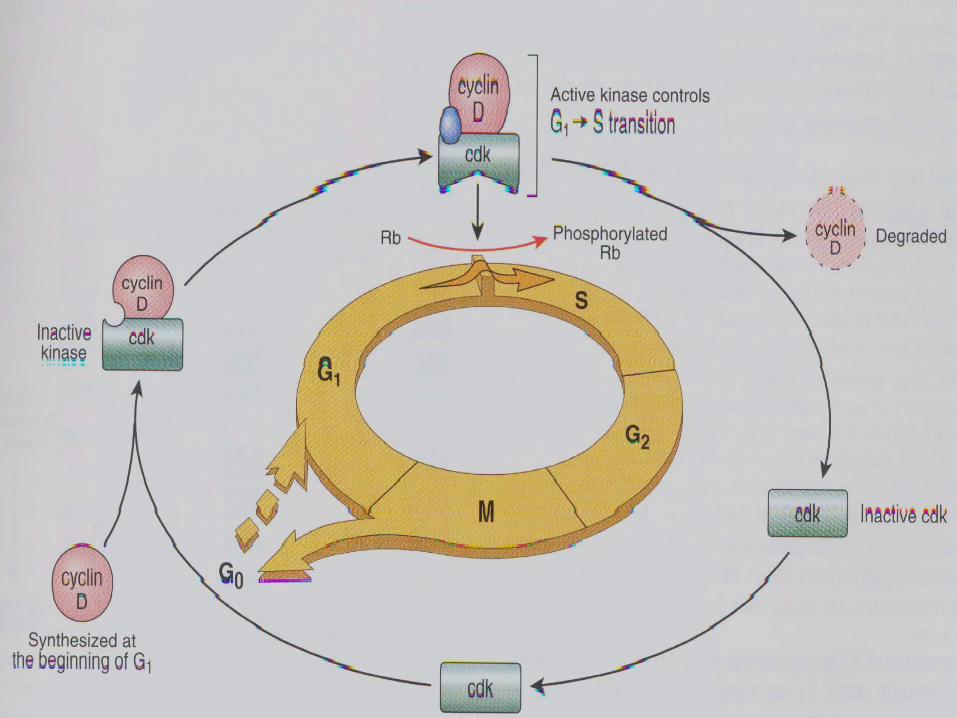

失去调控的癌细胞倾向于保留在细胞周期中持续循环;一旦细胞通过 G1晚期限制点,将对胞外生长调控信号产生不应期而代之以自律性程序,并带着这些信息进入有丝分裂。对限制点调控的研究是搞清癌细胞是怎样和为何持续进入循环的关键所在。

二、 S 期检测点 控制进入 S 期的检测点 (Gl 期检测点 ) 可防止 DNA 受损的细胞进入 S 期。
三、 G2 期检测点 控制进入 M 期的检测点 (G2 期检测点 ) 可防止受损的 DNA 和未完成复制的 DNA 进入有丝分裂。 p53 是 DNA 损伤诱导 G2 期阻滞的关键机制 , 因此 p53 缺失导致的 G2 期检测点缺陷与许多肿瘤的发生有关。


四、 M 期检测点 M 期检测点又叫纺锤体组装检测点。主要是阻止细胞分裂、阻止细胞两极形成纺锤体、阻止染色体附着到纺锤体上。

细胞周期检测点功能的减弱,会导致突变基因的累积和遗传的不稳定性,但只有当累积的突变基因破坏了细胞周期驱动机制时,细胞才能进入失控性生长。

第五节 细胞周期的调控与肿瘤一、细胞周期素与原癌基因与 G0 / G1 期转化有关的原癌基因有myc 和 ras ,它们在转化细胞中与 cyclin D共同作用。
E2F刺激 c-myc 和 cyclin A 的表达。 E2F释放可以进一步增加 c-myc 的表达,诱导 cyclin E 和 cyclin A 。
cyclin E—CDK 的激活则依赖于 ras 和myc 的共同作用。
c-myc表达增加可防止 cyclinE-CDK2灭活,允许细胞在 p27存在的情况下继续增殖。 C-myc 在此过程中诱导 p27以一种不能与 cyclinE-CDK2 结合的形式存在。
c-fos高表达可下调 p27 水平而使 cylcinE-CDK2 复合物活性增强,促使细胞增殖。
C-myc 与 ras 在细胞内共表达时可使细胞内 p27消失。

SKP2(S 期激酶相关蛋白 -2) 是新的肿瘤标记物,在正常组织中只表达于扁桃体和胎盘组织中,但在肿瘤组织中 SKP2表达广泛,包括部分结肠癌、前列腺癌、胰腺癌和皮肤癌,尤其在肺癌、乳腺癌、卵巢癌和子宫内膜癌高表达,并在淋巴瘤和乳腺癌肿瘤发生中起重要作用。在结肠癌和口腔上皮癌中SKP2 的表达与 p27 表达水平下降有一定的相关性。
尽管 cycIin D1 与肿瘤的关系最为密切。但单独的 cyclin D1 过度表达不足以造成细胞的恶性转化,还必须与其他能使细胞退出 G0期的原癌基因共同作用,如与 ras 、myc共同作用。

二、 cyclinDl 在人类恶性肿瘤中研究的意义 在正常组织中除了扁桃体(检测 Cyclin D1最理想的组织)、血管内皮细胞、一些组织细胞和鳞状上皮的基底细胞表达 cyclin D1 外其他组织均不表达 cyclin D1 。
cyclinDl由于基因的扩增或重排,在多种肿瘤中有过度表达,但在有些肿瘤中,如乳腺癌、肝癌、食管癌及胰腺癌中, cyclinDl RNA 和/或蛋白出现了过度表达,但相应的基因却没有出现扩增,这种扩增与表达的不一致的现象。在肿瘤中 cyclin D1 基因扩增约11%— 20 %, cyclin D1 蛋白过度表达率约为 50 %或更高,这可能是基因启动子区域的改变或调节转录因子如 c-jun 的异常表达所致,或是由于 cyclinDl mRNA3’非翻译区的截短导致其稳定性增加引起,或mRNA半衰期延长也可能导致这种结果。

陈莉的研究结果显示鼻咽癌中有 Cyclin D1 过表达( 56% ),在巢性膨胀性生长中 Cyclin D1 阳性率显著高于其它生长类型。在鼻咽癌中 Cyclin D1 、 P16 、RB 三者异常可同时存在。 CyclinD1 、 P16 、 RB 表达异常是鼻咽癌中常见的分子异常,并与癌生长、分化、转移有关; LMP1感染与 CyclinD1 过表达呈正相关,而与 P16 、 RB 表达呈负相关。
陈莉,宗永生《》癌症 1996年

p16缺失与 cyclin D1 过度表达联合作用可使肿瘤细胞获得更大的生长趋势。有报道认为缺乏 Rb功能的细胞对 p16敏感性下降。某些存在 p16 的细胞株伴有功能性 Rb 的缺乏;相反,缺乏 p16 的细胞株则常伴有功能性 Rb 的存在。
陈莉等在鼻咽癌的研究中指出:在鼻咽癌中 P16 表达减少,特别是鼻咽癌转移时,说明 P16改变在鼻咽癌发生中是晚期事件。 Rb 表达在非角化性鼻咽癌(WHOⅡ型)中显著高于未分化(WHO Ⅲ型), RB 阳性时 P16 表达减少,两者呈显著负相关。
p16 在很多肿瘤中都有重排、缺失或突变,提示 p16灭活与许多肿瘤的发生有关。 p16甲基化可能是家族性黑色素瘤的遗传基础。

在肿瘤中 cyclinD1 过表达的意义有:
cyclinD1 过表达与某些肿瘤的组织类型相关,在不同肿瘤中阳性检出率不同;
与组织分化程度相关,在分化差的肿瘤中表达增强;
与预后相关,在多种肿瘤中是独立的预后指标

套细胞淋巴瘤:
套细胞淋巴瘤除了有特征性的 t(11 , 14)(q13 ,q32) 外,还发现有 cyclin D1 基因 3’端的重排。对大多数套细胞淋巴瘤的研究表明均有 cyclinDl 蛋白的过表达。因此,国际淋巴瘤协会认为套细胞淋巴瘤染色体 11q13 的易位和 cyclinD1 的过表达都是很重要的特征。在套细胞淋巴瘤中,由于 cyclinD1具有高度的敏感性和相对的特异性,可能成为区别其他淋巴瘤的特异性标记。

套细胞淋巴瘤 CD5+ 、 CD45+
Cyclin D+

三、 cyclin A 在人类恶性肿瘤中研究的意义
最早使人们将肿瘤与细胞周期机制联系在一起的是 Cyclin A ,当时有两条线索提示 CyclinA 的改变可能与细胞转化 (癌变 ) 有关。如人们发现,在肝细胞癌的细胞中,乙肝病毒的 DNA片段整合到 cyclin A 的基因中;这是克隆肝细胞中 HBV的唯一插入点。病毒插入后整合人宿主 DNA 形成嵌合体, cyclin A 的 N末端直至 cyclin盒 (cyclin box) 的区域被病毒的基因序列取代,所形成的蛋白质不能在正常的有丝分裂期被降解。同样从腺病毒转化细胞中,看到 Cyclin A 与腺病毒转化蛋白 ElA 结合在一起。

四、 P27 在人类恶性肿瘤中研究的意义:
陈莉等研究 100 例胃癌组织中 p27Kip1 的表达与胃癌浸润、转移和预后的关系中指出,胃癌组织中 p27Kip1 蛋白阳性表达率为 44% 。在胃癌深部浸润组、淋巴结转移组和 5年内死亡组中 p27Kip1呈显著低表达( P<0.05 );经单变量分析结果显示 p27Kip1 高表达组 5年生存率为 70.59%, 显著高于低表达组 54.55% 和阴性组26%;多变量 Cox 回归模型分析显示 p27Kip1 是独立的预后指标。
( Li Chen De-chuen Li,Hong-jie Son The Relationship between the Expression of p27kip1,p53 and the Infiltration,Metastasis and Prognosis in Gastric Carcinoma, the Chinese-German Journal of Clinical Oncology 2005; 4(3) : 155-160)



问题与展望
细胞周期通常受到信号传导途径和反馈环路的精确调控 ,其正常与否和细胞以及个体的生长、分化、衰老和癌变密切相关。经过一个完整的细胞周期后,有的细胞继续增殖 ,有的细胞可能暂不增殖或永不增殖。决定细胞周期后各类细胞命运的关键则是细胞周期的调控。目前对细胞周期的调控研究已作为生命科学研究的热点,随着研究的深入将进一步揭示细胞周期调控的分子机制、细胞信号传导途径、恶性肿瘤癌变机制,将对认识细胞生长的动力学、有目的的调控细胞周期、对肿瘤的防治干预具有重要的指导意义。
细胞周期的调控与肿瘤
细胞周期检测点的调控
细胞周期调控中各元素间的相互作用
细胞周期调控中的重要元素
细胞周期的基本概念
