-galactosylceramide promotes killing of listeria ...iai.asm.org/content/78/6/2667.full.pdfpopulation...
TRANSCRIPT

INFECTION AND IMMUNITY, June 2010, p. 2667–2676 Vol. 78, No. 60019-9567/10/$12.00 doi:10.1128/IAI.01441-09Copyright © 2010, American Society for Microbiology. All Rights Reserved.
�-Galactosylceramide Promotes Killing of Listeria monocytogeneswithin the Macrophage Phagosome through Invariant
NKT-Cell Activation�
Masashi Emoto,1* Tomomi Yoshida,2 Toshio Fukuda,2 Ikuo Kawamura,3 Masao Mitsuyama,3Eiji Kita,4 Robert Hurwitz,5 Stefan H. E. Kaufmann,6 and Yoshiko Emoto1
Laboratory of Immunology, Department of Laboratory Sciences, Gunma University School of Health Sciences, Gunma 371-8511, Japan1;Laboratory of Pathology and Diagnostic Cytology, Department of Laboratory Sciences, Gunma University School of
Health Sciences, Gunma 371-8511, Japan2; Department of Microbiology, Kyoto University Graduate School ofMedicine, Kyoto 606-8501, Japan3; Department of Bacteriology, Nara Medical University, Nara 634-8521,
Japan4; Core Facilities Biochemistry, Max Planck Institute for Infection Biology, Berlin 10117,Germany5; and Department of Immunology, Max Planck Institute for
Infection Biology, Berlin 10117, Germany6
Received 23 December 2009/Returned for modification 25 January 2010/Accepted 17 March 2010
�-Galactosylceramide (�-GalCer) has been exploited for the treatment of microbial infections. Althoughamelioration of infection by �-GalCer involves invariant natural killer T (iNKT)-cell activation, it remains tobe determined whether macrophages (M�) participate in the control of microbial pathogens. In the presentstudy, we examined the participation of M� in immune intervention in infection by �-GalCer using a murinemodel of listeriosis. Phagocytic and bactericidal activities of peritoneal M� from C57BL/6 mice, but not iNKTcell-deficient mice, were enhanced after intraperitoneal injection of �-GalCer despite the absence of iNKT cellsin the peritoneal cavity. High levels of gamma interferon (IFN-�) and nitric oxide (NO) were detected in theperitoneal cavities of mice treated with �-GalCer and in culture supernatants of peritoneal M� from micetreated with �-GalCer, respectively. Although enhanced bactericidal activity of peritoneal M� by �-GalCer wasabrogated by endogenous IFN-� neutralization, this was only marginally affected by NO inhibition. Similarresults were obtained by using a listeriolysin O-deficient strain of Listeria monocytogenes. Moreover, respiratoryburst in M� was increased after �-GalCer treatment. Our results suggest that amelioration of listeriosis by�-GalCer is, in part, caused by enhanced killing of L. monocytogenes within phagosomes of M� activated byIFN-� from iNKT cells residing in an organ(s) other than the peritoneal cavity.
Listeria monocytogenes, a Gram-positive facultative intracel-lular bacterium, is the causative agent of listeriosis, with anoverall mortality rate of 30% (76). A major virulence factor ofL. monocytogenes is listeriolysin O (LLO), a 58-kDa proteinencoded by the hly gene (26, 42, 65). LLO promotes intracel-lular survival of L. monocytogenes in professional phagocytessuch as macrophages (M�) by promoting listerial escape fromthe phagosome into the cytosol (10, 22, 26, 42, 62, 65). Cells ofthe innate immune system play a pivotal role as a first line ofdefense against L. monocytogenes infection and among these,mononuclear phagocytes are critical (56, 61). Activation of M�by gamma interferon (IFN-�) is mandatory for elimination ofL. monocytogenes (31, 35). Nitric oxide (NO) synthesized byinducible NO synthase, which is localized in the cytosol ofprofessional phagocytes, participates in killing of L. monocy-togenes (48, 52, 69, 71). Similarly, reactive oxygen intermedi-ates (ROI) play a role in killing of L. monocytogenes within thephagosome (52, 53, 59).
Natural killer T (NKT) cells represent a unique T-lymphocytepopulation expressing NKR-P1B/C (NK1.1; CD161), which is a
type 2 membrane glycoprotein of the C-type lectin superfamily(6). In the mouse, the majority of NKT cells express aninvariant T-cell receptor (TCR) � chain encoded by V�14/J�18 gene segments and a TCRV� highly biased towardV�8.2, V�7, and V�2 (invariant NKT [iNKT] cells) (6). Incontrast to conventional T cells, which recognize antigenic pep-tides presented by polymorphic major histocompatibility complexclass I or class II molecules, iNKT cells recognize glycolipid an-tigens, including �-galactosylceramide (�-GalCer), a syntheticglycolipid originally isolated from a marine sponge, presented bythe nonpolymorphic antigen presentation molecule CD1d (6, 40).iNKT cells are highly versatile and promptly produce both type1 and type 2 cytokines, such as IFN-� and interleukin-4 (IL-4),respectively, upon activation through their TCRs (1, 15–17,79). IL-15 is an essential growth factor of both iNKT cells andNK cells and, hence, both cell populations are absent in IL-15-deficient (IL-15�/�) mice (58). The numbers of iNKT cellsare also markedly reduced in SJL mice because of a largedeletion in their TCRV� genetic region (5, 78).
In vivo administration of �-GalCer causes prompt release ofvarious cytokines by iNKT cells, which are involved in the controlof various diseases, e.g., tumor rejection and prevention of auto-immune diseases (33, 41, 67, 70). Although �-GalCer has beenreported to enhance host resistance to some microbial patho-gens (27–29, 37, 39, 44, 55, 64), its potential role in protectionagainst intracellular bacterial infections remains enigmatic.
* Corresponding author. Mailing address: Laboratory of Immu-nology, Department of Laboratory Sciences, Gunma UniversitySchool of Health Sciences, 3-39-22 Showa-machi, Maebashi,Gunma 371-8511, Japan. Phone and fax: 81-27-220-8935. E-mail:[email protected].
� Published ahead of print on 29 March 2010.
2667
on May 14, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

We have recently described that �-GalCer ameliorates mu-rine listeriosis, which is, in part, caused by accelerated infiltra-tion of inflammatory cells into the liver (18), although iNKTcells themselves exacerbate disease (19). Because M� play acentral role in the elimination of L. monocytogenes, we consid-ered the possibility that M� participate in enhanced resistanceto L. monocytogenes infection caused by �-GalCer treatment.In the present study, we examined the influence of �-GalCeron listericidal activities of M� using a virulent and an avirulentstrain of L. monocytogenes.
MATERIALS AND METHODS
Mice. C57BL/6, IL-15�/�, SJL, and recombination-activating gene 1�/�
(RAG-1�/�) mice were purchased from Japan SLC (Hamamatsu, Japan), theJackson Laboratory (Bar Harbor, ME), Charles River Laboratories Japan(Yokohama, Japan), and the Jackson Laboratory, respectively, and maintainedunder specific-pathogen-free conditions at our animal facilities. Weight-matched female mice were used at 8 to 12 weeks of age in accordance withinstitutional guidelines of Gunma University, Max Planck Institute for Infec-tion Biology and Nara Medical University. Unless otherwise stated, C57BL/6mice were used.
Bacteria and infection. The parental wild-type strain of L. monocytogenes usedin the present study was EGD. The �hly mutant strain was constructed from thewild-type strain of L. monocytogenes by homologous recombination as describedpreviously (30). L. monocytogenes EGD and �hly strains recovered from infectedliver were grown in tryptic soy broth (Difco Laboratories, Detroit, MI) at 37°Cfor 18 h, and aliquots were frozen at –80°C for later use. The final concentrationof viable bacteria was enumerated by plate counts on tryptic soy agar (DifcoLaboratories). Mice were infected intraperitoneally (i.p.) with 2 � 104
L. monocytogenes bacteria. Unless otherwise stated, strain EGD was used.Antibodies. Monoclonal antibodies (MAbs) against Fc� receptor (Fc�R) and
IFN-� (2.4G2 and XMG1.2, respectively) were purified from hybridoma culturesupernatants by ammonium sulfate precipitation and affinity chromatography onprotein A- or G-Sepharose (Amersham Biosciences, Freiburg, Germany). Flu-orescein isothiocyanate (FITC)-conjugated anti-CD3ε MAb (145-2C11) was pur-chased from BD PharMingen (Hamburg, Germany). Alexa 594-conjugated anti-rabbit IgG antibody (Ab) and rabbit anti-Listeria sp. Ab were obtained fromViroStat (Portland, ME) and Molecular Probes (Eugene, OR), respectively.
�-GalCer-loaded CD1d tetramer. �-GalCer-loaded CD1d (�-GalCer/CD1d)tetramers were prepared by using the baculovirus expression system, using aCD1d construct with a BirA biotinylation site, followed by a His6 tag, as de-scribed previously (19, 50).
�-GalCer treatment. �-GalCer was kindly provided by Kirin Pharma Co. Ltd.(Tokyo, Japan). Polysorbate 20 was purchased from Wako Pure Chemical In-dustries (Osaka, Japan). Mice were treated i.p. with �-GalCer dissolved in 1%polysorbate 20 (1 �g/200 �l) or with 1% polysorbate 20 (200 �l) as a vehicle. Inanother experiment, cells were incubated in the presence of �-GalCer dissolvedin 1% polysorbate 20 (100 ng/ml) or polysorbate 20.
Preparation of cells and peritoneal fluids. Mice were anesthetized with ether,and peritoneal cells (PC) were harvested from the peritoneal cavity of mice afteri.p. injection with RPMI 1640 (Nissui Pharmaceutical Co., Ltd., Tokyo, Japan)containing 10% fetal calf serum (FCS; Bio West, France) (5 ml). After a washingstep, cells were incubated in six-well tissue culture plates (Corning, Inc., Canton,NY) at 37°C for 2 h, and nonadherent and adherent cells (regarded as M�) werecollected separately. To obtain thioglycolate (TG)-induced M�, mice were in-jected i.p. with 3 ml of TG (Eiken Chemical Co., Ltd., Tokyo, Japan) 4 daysbefore cell preparation. To obtain peritoneal washout, mice were injected i.p.with 3 ml of RPMI 1640 containing 10% FCS, peritoneal fluids containing PCwere harvested from the peritoneal cavity, and supernatants were collected aftercentrifugation. Unless otherwise stated, TG-uninduced M� were used. Hepaticleukocytes (HL) were prepared as described previously (15).
Determination of bactericidal activity of M�. M� were incubated in 24-welltissue culture plates (Corning) at 106/well for 2 h and infected with L. monocy-togenes at 107 bacteria/well (multiplicity of infection [MOI] 10). After 60 minof incubation, the cells were washed three times with prewarmed RPMI 1640containing 10% FCS and gentamicin (Sigma-Aldrich, Tokyo, Japan) (5 �g/ml) toremove noningested bacteria. A portion of cells was treated with 0.5% saponin(Sigma-Aldrich) and plated on tryptic soy agar plates after sonication, and CFUwere determined after 48 h of culture at 37°C (P60). For determination ofbactericidal activity, cells were further incubated in RPMI 1640 containing 10%
FCS and gentamicin (5 �g/ml) for 30 min, treated with 0.5% saponin (Sigma-Aldrich), and plated on tryptic soy agar plates after sonication. CFU counts weredetermined after 48 h of culture at 37°C (B30). Bactericidal activity was calcu-lated as follows: 100 – [(B30/P60) � 100] (%). The data are expressed as a killingindex calculated as follows: experimental group {100 – [(B30/P60) � 100]}/control group {100 – [(B30/P60) � 100]}.
Flow cytometry. Cells were incubated with anti-Fc�R MAb and then stainedwith conjugated MAb at 4°C for 15 min. Stained cells were washed with phos-phate-buffered saline containing 0.1% bovine serum albumin (Thermo, Hamil-ton, New Zealand) and 0.1% sodium azide (Wako Pure Chemical Industries),fixed with 1% paraformaldehyde (Merck, Darmstadt, Germany), acquired byFACSCalibur (BD Biosciences, Mountain View, CA), and analyzed withCellQuest software (BD Biosciences). Cells were stained with phycoerythrin(PE)-labeled �-GalCer/CD1d tetramer for 15 min at room temperature afterblocking.
Enzyme-linked immunosorbent assay. IFN-� levels were determined by Quan-tikine (R&D Systems, Inc., Minneapolis, MN) according to the manufacturer’sinstructions. The detection limit was 2 pg/ml.
NO measurement. NO was measured using the Griess reagent system(Promega, Madison, WI) according to the manufacturer’s instructions. Thedetection limit was 2.5 �M.
Immunohistochemistry. M� (105/well) incubated in a Lab-Tek Chamber Slide(Nalge Nunc International, Naperville, IL) were infected with L. monocytogenes(106 bacteria/well; MOI 10) for 1, 2, or 4 h. After a wash with prewarmedRPMI 1640 containing 10% FCS and gentamicin (5 �g/ml), the cells werestained with Alexa 488-conjugated phalloidin (Molecular Probes) and/or rabbitanti-Listeria Ab at room temperature for 2 h, followed by Alexa 594-conjugatedanti-rabbit IgG Ab at room temperature for a further 2 h. Specimens wereobserved under fluorescence microscope and photographed.
Inhibition experiment. M� were incubated in 24-well tissue culture plates(Corning) at 106/well in the presence or absence of NG-monomethyl-L-arginine(500 �M; Calbiochem, Darmstadt, Germany) or anti-IFN-� neutralizing MAb(50 �g/ml) for 24 h.
Reverse transcription-PCR. TCRV�14 mRNA expression was analyzed asdescribed previously (20), with slight modifications. Briefly, total RNA extractedwith an RNAgents total RNA isolation system (Promega) was primed for reversetranscription with random hexamers. To normalize cDNA content for furthercomparative analysis, �-actin-specific PCR was performed with serial dilutions ofcDNA. The �-actin-specific primers were 5-TGGAATCCTGTGGCATCCATGAAC-3 (forward) and 5-TAAAACGCAGCTCAGTAACAGTCCG-3 (re-verse).
After a denaturation step at 94°C for 3 min, PCR cycles were run at 94°C for30 s, 58°C for 30 s, and 72°C for 1 min for 30 cycles, followed by an extension stepat 72°C for 7 min. �-Actin PCR products were analyzed by 1.2% agarose gelelectrophoresis and ethidium bromide staining. For analysis of TCRV�14mRNA expression, a normalized amount of cDNA yielding equivalent amountsof �-actin-specific PCR products was applied. The cDNA was denatured at 94°Cfor 4 min, and PCR cycles were run at 94°C for 30 s, 55°C for 30 s, and 72°C for40 s for 30 cycles. The PCR products were subjected to electrophoresis on a 1.8%agarose gel. The 3 and 5 primers for the TCRV�14 PCR were (3) C� (5-GAAGCTTGTCTGGTTGCTCCAG-3) and (5) V�14 (5-CTAAGCACAGCACGCTGCACA-3). All PCRs were performed in a PTC-150 MiniCycler (Bio-Rad, Tokyo, Japan).
Luminol-dependent chemiluminescence. The activity of respiratory burst inperitoneal M� was evaluated by using luminol-dependent chemiluminescence(CL) as described previously (23, 46). In brief, a peritoneal M� suspension(1.2 � 106 to 1.5 � 106 cells in 400 �l of Hanks balanced salt solution [HBSS]containing 1% gelatin [GHBSS]) was mixed with 30 �l of luminol solution (10�6
M; Laboscience Co., Tokyo, Japan) in a counting tube, which was kept at 37°Cfor 5 min with gentle agitation. L. monocytogenes grown in tryptic soy broth at37°C for 18 h was sedimented (1,200 � g) for 20 min and washed twice in HBSSbefore being adjusted to a concentration of 6 � 108 CFU/ml. The bacteria wereopsonized for 60 min at 37°C with 10% normal pooled mouse serum in GHBSSand washed twice with GHBSS before use for the CL assay. To generate CL,opsonized L. monocytogenes (108 CFU in 150 �l of GHBSS) was added to thecell mixture. Light release was measured by using a Lumiphotometer TD-4000(Laboscience) over a 15-min period, and the peak response (in relative light units[RLU]) was noted.
Statistical analysis. The statistical significance was determined by using aMann-Whitney U test, and a P value of �0.05 was regarded as significant.
2668 EMOTO ET AL. INFECT. IMMUN.
on May 14, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

RESULTS
�-GalCer treatment ameliorates murine listeriosis. C57BL/6mice were left untreated or were treated with �-GalCer orvehicle and then infected with L. monocytogenes. CFU in peri-toneal cavities, livers, and spleens were determined at differenttime points thereafter. Considerable numbers of viable bacte-ria were detected in peritoneal cavities of vehicle-treated anduntreated mice 1 h postinfection (p.i.), and these were reducedby 1 order of magnitude after �-GalCer treatment (Fig. 1).Viable bacteria in peritoneal cavities became undetectable (�50CFU) at 4 h p.i. regardless of �-GalCer treatment (data notshown). High numbers of viable bacteria were detected in thelivers and spleens of vehicle-treated and untreated mice on day4 p.i., and these were reduced up to 100-fold by �-GalCer treat-ment (Fig. 1). Thus, �-GalCer treatment ameliorated listeriosis.
In vivo administration of �-GalCer enhances bactericidaland phagocytic activities of M� in an iNKT-cell-dependentmanner. M� play a central role in elimination of L. monocy-togenes (56, 61). We therefore examined the influence of�-GalCer on the bactericidal activity of M�. Peritoneal M�from mice left untreated or treated with �-GalCer or vehiclewere infected with L. monocytogenes, and their bactericidalactivities were compared. Since it has been shown that genta-micin has the potential to kill L. monocytogenes in M� (13), wefirst examined whether L. monocytogenes in M� was killed bygentamicin. To address this issue, we compared numbers ofviable bacteria in M� cultured in the presence or absence ofgentamicin. In contrast to previous findings (13), the numbersof viable bacteria in M� were comparable between the twogroups (data not shown). Consistent with this, many actin tailswere found in TG-induced M� 4 h after in vitro culture in thepresence of gentamicin (see Fig. 3D). Thus, the viability of L.monocytogenes in M� was apparently not affected by gentami-cin in our experimental system. Because viable L. monocyto-genes bacteria in resident M� from C57BL/6 mice were max-imal at 60 min after phagocytosis and viable bacteria becamevirtually undetectable a further 60 min after incubation regard-less of mouse strains, we determined phagocytic and bacteri-cidal activities of M� at 60 and 30 min after infection, respec-tively. The data are expressed as a killing index and not asnumbers of killed bacteria to standardize bactericidal activity,because it is possible that numbers of killed bacteria are higherin M� that phagocytose many bacteria than in those thatphagocytose few bacteria. The bactericidal activity (for 30 min
after 60 min of phagocytosis) of M� from �-GalCer-treatedC57BL/6 mice was significantly higher than that from vehicle-treated and untreated C57BL/6 mice (Fig. 2A, upper panel).Accordingly, M� showed morphological changes after �-Gal-Cer treatment (see Fig. 3A). In contrast to C57BL/6 mice,bactericidal activities of M� from IL-15�/� and SJL mice werevirtually unchanged after �-GalCer treatment (Fig. 2A, upperpanel). Bactericidal activities of M� from untreated C57BL/6,IL-15�/�, and SJL mice were comparable, suggesting thatM� from these mouse strains have similar listericidal po-tentials (data not shown). Hence, �-GalCer enhanced bacte-ricidal activity of M� in an iNKT-cell-dependent fashion.
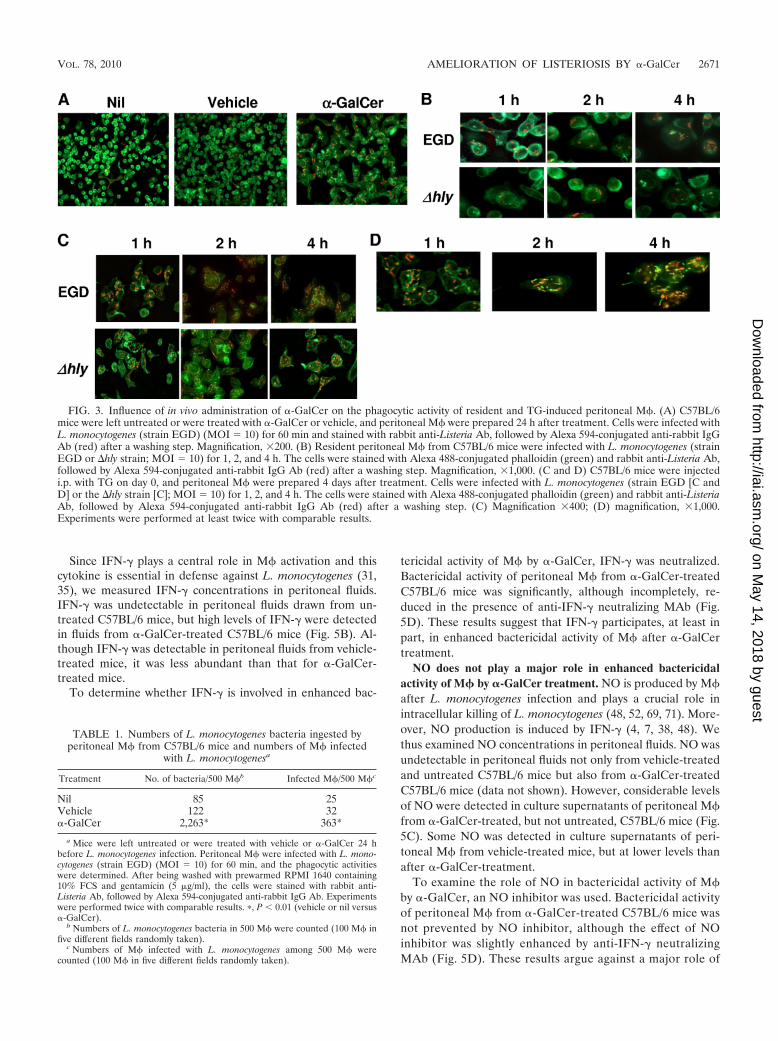
We next examined the influence of �-GalCer on phagocyticactivity of M� by counting viable bacteria (colony countmethod) after 60 min of phagocytosis. Considerable numbers,but low efficacy of phagocytosis (ca. 0.1%), of viable bacteriawere detected in M� from untreated and vehicle-treatedC57BL/6 mice, and these numbers were significantly lower inM� from �-GalCer-treated C57BL/6 mice (Fig. 2B). To verifywhether phagocytic activity of M� from C57BL/6 mice wasindeed reduced by �-GalCer treatment, peritoneal M� fromC57BL/6 mice that were left untreated or were treated with�-GalCer or vehicle were infected with L. monocytogenes, andthe numbers of L. monocytogenes bacteria phagocytosed byM� were determined immunohistochemically using anti-List-eria Ab. In contrast to the colony count method, the numbersof L. monocytogenes bacteria ingested by M� from �-GalCer-treated C57BL/6 mice were markedly higher than those fromvehicle-treated and untreated C57BL/6 mice (Fig. 3A and Ta-ble 1). No alterations were found in the numbers of L. mono-cytogenes bacteria ingested by M� from IL-15�/� and SJL miceafter �-GalCer treatment (Fig. 2B). Hence, �-GalCer en-hanced the phagocytic activity of M� in an iNKT-cell-depen-dent manner.
iNKT cells in organs other than peritoneal cavity partici-pate in enhanced bactericidal activity of peritoneal M� by�-GalCer treatment. We wondered whether enhancement ofbactericidal activity of peritoneal M� by �-GalCer involvediNKT cells present in the peritoneal cavity. Total PC fromC57BL/6 mice were stimulated in vitro with �-GalCer andinfected with L. monocytogenes, and bactericidal activity of M�was determined. In contrast to in vivo treatment, bactericidalactivity of M� was unchanged by in vitro stimulation with�-GalCer (data not shown). We therefore rule out the pos-
FIG. 1. Influence of in vivo administration of �-GalCer on listerial growth in the peritoneal cavities, livers, and spleens of C57BL/6 mice. Micewere left untreated or were treated with �-GalCer or vehicle on day �1 and infected with L. monocytogenes (strain EGD) on day 0. CFU weredetermined 1 h (peritoneal cavity) or 4 days (liver and spleen) p.i. Each symbol represents CFU in an individual animal, and horizontal linesindicate the mean CFU. The data from four mice per group are shown. Experiments were performed three times with comparable results. *,P � 0.01 (vehicle or nil versus �-GalCer).
VOL. 78, 2010 AMELIORATION OF LISTERIOSIS BY �-GalCer 2669
on May 14, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

sibility that enhancement of bactericidal activity of peritonealM� by �-GalCer involved iNKT cells in the peritoneal cavity.We wondered whether the lack of enhanced bactericidal activ-ity of peritoneal M� by �-GalCer was caused by the absence ofiNKT cells in the peritoneal cavity. We therefore attemptedto detect iNKT cells in PC using �-GalCer/CD1d tetramers.HL were used as a positive control, because iNKT cells aremost abundant in the liver compared to other organs (16).�-GalCer/CD1d tetramer-reactive T cells were undetectablein peritoneal cavities not only of IL-15�/� and SJL mice (datanot shown) but also of C57BL/6 mice regardless of �-GalCertreatment (Fig. 4A), although high numbers of �-GalCer/CD1d tetramer-reactive T cells were detected in the livers ofuntreated C57BL/6 mice, and these became undetectable after�-GalCer treatment (Fig. 4A) (18, 19). To verify whetheriNKT cells were indeed absent in the peritoneal cavity, V�14mRNA expression in PC was analyzed. As positive and nega-tive controls, V�14 mRNA expression of HL from C57BL/6mice and RAG-1�/� mice lacking T cells, including iNKT cells,was also analyzed. TCRV�14 mRNA was detected in HL from
C57BL/6 mice but was undetectable in PC from C57BL/6 miceregardless of �-GalCer treatment, as well as in HL from RAG-1�/� mice (Fig. 4B). These results indicate that iNKT cellsresponsible for enhanced bactericidal activity of M� after�-GalCer treatment reside in organs other than the peritonealcavity.
IFN-� from iNKT cells participates in enhanced bacteri-cidal activity of M� by �-GalCer treatment. We wonderedwhether cytokines secreted by iNKT cells participate in en-hanced bactericidal activity of M� by �-GalCer. Residentperitoneal M� from C57BL/6 mice were incubated in thepresence of peritoneal fluids from C57BL/6 mice left un-treated or treated with �-GalCer or vehicle, and the bacteri-cidal activities were determined. Bactericidal activities of M�were unchanged by peritoneal fluids from vehicle-treated oruntreated mice, whereas those of M� were significantly en-hanced by peritoneal fluids from �-GalCer-treated mice (Fig.5A). These results suggest that cytokines enhance bactericidalactivity of M� in the peritoneal cavities of �-GalCer-treatedmice.
FIG. 2. Influence of in vivo administration of �-GalCer on listericidal and phagocytic activities of peritoneal M�. C57BL/6, IL-15�/�, and SJLmice were left untreated or were treated with �-GalCer or vehicle, and peritoneal M� were prepared 24 h after treatment. Cells were infected withL. monocytogenes (strain EGD or �hly mutant; MOI 10), and the bactericidal and phagocytic activities were determined. (A) Data representthe killing index (nil 1) and are expressed as means � the standard deviations (SD) of three mice. (B) Data represent the phagocytic index (nil 1) and are expressed as means � the SD of three mice. Experiments were performed at least three times with comparable results. *, P � 0.05(vehicle or nil versus �-GalCer).
2670 EMOTO ET AL. INFECT. IMMUN.
on May 14, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
Since IFN-� plays a central role in M� activation and thiscytokine is essential in defense against L. monocytogenes (31,35), we measured IFN-� concentrations in peritoneal fluids.IFN-� was undetectable in peritoneal fluids drawn from un-treated C57BL/6 mice, but high levels of IFN-� were detectedin fluids from �-GalCer-treated C57BL/6 mice (Fig. 5B). Al-though IFN-� was detectable in peritoneal fluids from vehicle-treated mice, it was less abundant than that for �-GalCer-treated mice.
To determine whether IFN-� is involved in enhanced bac-
tericidal activity of M� by �-GalCer, IFN-� was neutralized.Bactericidal activity of peritoneal M� from �-GalCer-treatedC57BL/6 mice was significantly, although incompletely, re-duced in the presence of anti-IFN-� neutralizing MAb (Fig.5D). These results suggest that IFN-� participates, at least inpart, in enhanced bactericidal activity of M� after �-GalCertreatment.
NO does not play a major role in enhanced bactericidalactivity of M� by �-GalCer treatment. NO is produced by M�after L. monocytogenes infection and plays a crucial role inintracellular killing of L. monocytogenes (48, 52, 69, 71). More-over, NO production is induced by IFN-� (4, 7, 38, 48). Wethus examined NO concentrations in peritoneal fluids. NO wasundetectable in peritoneal fluids not only from vehicle-treatedand untreated C57BL/6 mice but also from �-GalCer-treatedC57BL/6 mice (data not shown). However, considerable levelsof NO were detected in culture supernatants of peritoneal M�from �-GalCer-treated, but not untreated, C57BL/6 mice (Fig.5C). Some NO was detected in culture supernatants of peri-toneal M� from vehicle-treated mice, but at lower levels thanafter �-GalCer-treatment.
To examine the role of NO in bactericidal activity of M�by �-GalCer, an NO inhibitor was used. Bactericidal activityof peritoneal M� from �-GalCer-treated C57BL/6 mice wasnot prevented by NO inhibitor, although the effect of NOinhibitor was slightly enhanced by anti-IFN-� neutralizingMAb (Fig. 5D). These results argue against a major role of
FIG. 3. Influence of in vivo administration of �-GalCer on the phagocytic activity of resident and TG-induced peritoneal M�. (A) C57BL/6mice were left untreated or were treated with �-GalCer or vehicle, and peritoneal M� were prepared 24 h after treatment. Cells were infected withL. monocytogenes (strain EGD) (MOI 10) for 60 min and stained with rabbit anti-Listeria Ab, followed by Alexa 594-conjugated anti-rabbit IgGAb (red) after a washing step. Magnification, �200. (B) Resident peritoneal M� from C57BL/6 mice were infected with L. monocytogenes (strainEGD or �hly strain; MOI 10) for 1, 2, and 4 h. The cells were stained with Alexa 488-conjugated phalloidin (green) and rabbit anti-Listeria Ab,followed by Alexa 594-conjugated anti-rabbit IgG Ab (red) after a washing step. Magnification, �1,000. (C and D) C57BL/6 mice were injectedi.p. with TG on day 0, and peritoneal M� were prepared 4 days after treatment. Cells were infected with L. monocytogenes (strain EGD [C andD] or the �hly strain [C]; MOI 10) for 1, 2, and 4 h. The cells were stained with Alexa 488-conjugated phalloidin (green) and rabbit anti-ListeriaAb, followed by Alexa 594-conjugated anti-rabbit IgG Ab (red) after a washing step. (C) Magnification �400; (D) magnification, �1,000.Experiments were performed at least twice with comparable results.
TABLE 1. Numbers of L. monocytogenes bacteria ingested byperitoneal M� from C57BL/6 mice and numbers of M� infected
with L. monocytogenesa
Treatment No. of bacteria/500 M�b Infected M�/500 M�c
Nil 85 25Vehicle 122 32�-GalCer 2,263* 363*
a Mice were left untreated or were treated with vehicle or �-GalCer 24 hbefore L. monocytogenes infection. Peritoneal M� were infected with L. mono-cytogenes (strain EGD) (MOI 10) for 60 min, and the phagocytic activitieswere determined. After being washed with prewarmed RPMI 1640 containing10% FCS and gentamicin (5 �g/ml), the cells were stained with rabbit anti-Listeria Ab, followed by Alexa 594-conjugated anti-rabbit IgG Ab. Experimentswere performed twice with comparable results. �, P � 0.01 (vehicle or nil versus�-GalCer).
b Numbers of L. monocytogenes bacteria in 500 M� were counted (100 M� infive different fields randomly taken).
c Numbers of M� infected with L. monocytogenes among 500 M� werecounted (100 M� in five different fields randomly taken).
VOL. 78, 2010 AMELIORATION OF LISTERIOSIS BY �-GalCer 2671
on May 14, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

NO in enhanced bactericidal activity of M� by �-GalCertreatment.
Respiratory burst within the phagosome is promoted by�-GalCer. ROI are produced by M� in response to L.monocytogenes infection, and they play a central role inlisterial killing within phagosomes (52, 53, 59). To deter-mine the role of ROI experimentally, the respiratory burstsin M� from �-GalCer- and vehicle-treated C57BL/6 micewere compared. The maximal CL responses were signifi-cantly higher in peritoneal M� from �-GalCer-treated micethan in those from vehicle-treated mice (Table 2). The CLemission during phagocytosis by peritoneal M� from �-Gal-Cer-treated mice reached a maximum within 160 s (seeTable 2) and then gradually decayed (data not shown). Incontrast, the emission by peritoneal M� from vehicle-treated mice was slower (see Table 2) and rapidly decayedafter reaching its maximum (data not shown). Thus, �-Gal-Cer promoted a profound respiratory burst in M� afteringestion of L. monocytogenes.
�-GalCer promotes killing of L. monocytogenes within thephagosomes of peritoneal M�. To evade phagosomal destruc-tion, L. monocytogenes escapes from the phagosome into thecytosol by means of LLO (10, 22, 26, 42, 62, 65). However, inthe cytosol, NO is capable of killing L. monocytogenes (48, 52,69, 71). We therefore determined whether �-GalCer promotesthe killing of L. monocytogenes after escape into the cytosol ofM�. Subsequent to listerial escape from the phagosome, actintails are formed in the cytosol of M� (12, 30, 32, 63, 74). Weused immunohistochemistry, specifically fluorescein-labeledphalloidin, for detection of actin tails in peritoneal M�. Incontrast to previous findings (12, 30, 32, 63, 74), the actin tailwas undetectable in M� even 4 h p.i. regardless of �-GalCertreatment (Fig. 3B, upper panel). This not only indicates thatL. monocytogenes does not escape from the phagosome into
the cytosol but also suggests that the killing of L. monocyto-genes occurs in the phagosome, prior to entry into the cytosol.To decide whether �-GalCer promotes killing of L. monocy-togenes within the phagosome, the �hly L. monocytogenesstrain, which has lost the capacity to escape from the phago-some into the cytosol due to the absence of LLO, was used.Bactericidal and phagocytic activities of M� from C57BL/6mice, but not from IL-15�/� and SJL mice, were enhanced by�-GalCer treatment (Fig. 2, lower panel, and Fig. 3B, lowerpanel). We wondered whether our failure to obtain evidencefor phagosomal escape of L. monocytogenes was related to thesource of M� used in our experiments. We thus used TG-induced peritoneal M�. In contrast to resident M�, the actintail was detected in TG-induced M� infected with strain EGDbut not with the �hly strain (Fig. 3C and D). The proportion ofM� with the actin tail was rare, although detectable, 1 h afterinfection. However, the numbers of M� with actin tails wereincreased thereafter, and actin tails became detectable in mostof infected M� 4 h after infection (see Fig. 3D). Our resultssuggest that �-GalCer promotes the killing of L. monocyto-genes within the phagosomes of M�.
DISCUSSION
We show here that M� activation by iNKT cells is in-volved in the amelioration of listeriosis after �-GalCer in-tervention. Listericidal activity of peritoneal M� was en-hanced by �-GalCer, at least in part, via IFN-� secreted byiNKT cells. We conclude that effects of �-GalCer in listeriosisare influenced by enhanced bactericidal M� activities and me-diated by IFN-� from iNKT cells.
In our experiments, IL-15�/� mice were used as iNKT cell-deficient mice. Since IL-15�/� mice are devoid not only ofiNKT cells but also of NK cells, we cannot formally exclude the
FIG. 4. Proportions of iNKT cells in the peritoneal cavity and liver, and TCRV�14 mRNA expression in nonadherent PC and HL before andafter in vivo administration of �-GalCer. Mice were left untreated or treated with �-GalCer. (A) Nonadherent PC and HL from C57BL/6 micewere stained with FITC-conjugated anti-CD3ε MAb and PE-labeled �-GalCer/CD1d tetramer. The data are expressed as dot plots after gatingon lymphoid cells defined by forward-scatter and side-scatter profiles. The numbers in dot plots represent the percentages of CD3ε �-GalCer/CD1d tetramer-reactive T cells. Representative data from three mice are shown. (B) TCRV�14 mRNA expression in HL from RAG-1�/� miceand in HL and PC from C57BL/6 mice was analyzed. Total RNA extracted from cells was reverse transcribed into cDNA and amplified by PCRusing primers for V�14 and C� segments. �-Actin-specific PCR was also performed. Experiments were performed twice with comparable results.
2672 EMOTO ET AL. INFECT. IMMUN.
on May 14, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

possible contribution of NK cells to amelioration of listeriosisand to enhanced bactericidal activity of M� by �-GalCer.However, amelioration of listeriosis and enhanced bactericidalactivity of M� by �-GalCer were not found in SJL mice, witha genetic background different from that of C57BL/6 mice, andamelioration of listeriosis by �-GalCer was not found in
J�18�/� mice, which harbor NK cells but not iNKT cells (19).Thus, we assume that iNKT cells are essential for ameliorationof listeriosis and M� activation by �-GalCer.
Numbers of viable L. monocytogenes bacteria in M� from�-GalCer-treated C57BL/6 mice were significantly lowerthan those from vehicle-treated and untreated mice 60 minafter infection (as determined by the colony count method).Nevertheless, by immunohistochemical analysis, consider-able numbers of L. monocytogenes bacteria were detected inM� from �-GalCer-treated C57BL/6 mice, and these weremarkedly higher than those from vehicle-treated and untreatedmice. The discrepancy is probably caused by the difference inkilling activity of each M�, i.e., L. monocytogenes organismsare killed by M� from �-GalCer-treated mice immediatelyafter infection. Indeed, the numbers of viable L. monocyto-genes bacteria in M� from �-GalCer-treated C57BL/6 micewere markedly higher than those from vehicle-treated anduntreated mice 20 min after infection (M. Emoto et al., un-published observations). These results indicate that �-GalCerhas a great potential to ingest and kill L. monocytogenes by M�through iNKT cell activation. However, because it is possible
FIG. 5. IFN-� levels in peritoneal fluids and NO concentrations in M� culture supernatants after �-GalCer stimulation, and the influence ofIFN-� neutralization or NO inhibition on bactericidal activity of M�. (A) C57BL/6 mice were left untreated or were treated with �-GalCer orvehicle, and the peritoneal fluids were collected 24 h after treatment. Resident peritoneal M� from C57BL/6 mice were incubated with theperitoneal fluids. Cells were then infected with L. monocytogenes (strain EGD; MOI 10), and the bactericidal activities were determined. Thedata represent the killing index (nil 1) and are expressed as means � the SD of three mice. Experiments were performed twice with comparableresults. *, P � 0.05 (vehicle or nil versus �-GalCer). (B) C57BL/6 mice were left untreated or were treated with �-GalCer or vehicle, and theperitoneal fluids were collected 24 h after treatment. The IFN-� levels in the peritoneal fluids were determined by enzyme-linked immunosorbentassay. The data are expressed as means � the SD of three mice. Experiments were performed twice with comparable results. *, P � 0.01 (vehicleor nil versus �-GalCer). (C) C57BL/6 mice were left untreated or were treated with �-GalCer or vehicle, and the peritoneal M� were prepared24 h after treatment. Cells were incubated for 2 h in a 96-well flat-bottom tissue culture plate, and the NO concentrations in the culturesupernatants were determined. The data are expressed as means � the SD of three mice. Experiments were performed twice with comparableresults. *, P � 0.01 (vehicle or nil versus �-GalCer). (D) Resident peritoneal M� from C57BL/6 mice were incubated with peritoneal fluids fromC57BL/6 mice treated with �-GalCer or vehicle in the presence or absence of anti-IFN-� MAb and/or NG-monomethyl-L-arginine for 24 h. Thecells were then infected with L. monocytogenes (strains EGD; MOI 10), and the bactericidal activities were determined. The data represent thekilling index (nil 1) and are expressed as means � the SD of three mice. Experiments were performed twice with comparable results. *, P �0.05 (�-GalCer versus �-GalCer anti-IFN-� MAb); **, P � 0.01 (�-GalCer versus �-GalCer anti-IFN-� MAb NO inhibitor).
TABLE 2. CL response to L. monocytogenes by peritoneal M�from C57BL/6 mice treated with �-GalCer or vehiclea
Treatment
Mean � SD
Intensity of CLb Peak(RLU, 10�3) Peak time (s)
�-GalCer 11.01 � 0.85* 1.80 � 0.33 163 � 12*Vehicle 4.57 � 1.32 1.71 � 0.17 375 � 27
a Resident peritoneal M� (1.2 � 106 to 1.5 � 106 cells) were mixed withluminol solution (10�6 M) and opsonized L. monocytogenes (108 CFU) at 37°C.Light emission was measured at 37°C for 15 min. Experiments were performedfour times with comparable results. In each assay, the CL measurement wasperformed in duplicate for the same sample (n 8). �, P � 0.05 (vehicle versus�-GalCer).
b Intensities are expressed as peak RLU values.
VOL. 78, 2010 AMELIORATION OF LISTERIOSIS BY �-GalCer 2673
on May 14, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

that pinocytosis is accelerated by �-GalCer, we cannot formallyexclude the possibility that this effect is influenced by genta-micin, although comparable numbers of viable L. monocyto-genes bacteria were detected in M� cultured in the presence orabsence of gentamicin.
Abundant numbers of iNKT cells have been detected in theperitoneal cavities of C57BL/6 mice by flow cytometry (36). Incontrast, we did not detect iNKT cells in the peritoneal cavityeven at mRNA levels. The discrepancy is probably caused bynonspecific staining in the study of Ito et al. (36), because cellsare diagonally stained and a distinct population of NKT cells isinvisible. Since (i) iNKT cells secrete IFN-� in response to�-GalCer (6, 40), (ii) high levels of IFN-� were detected inperitoneal fluids of �-GalCer-treated C57BL/6, but not IL-15�/� and SJL, mice, and (iii) iNKT cells were undetectable inthe peritoneal cavity even at mRNA levels, we assume thatIFN-� was derived from iNKT cells residing in an organ(s)other than the peritoneal cavity. Indeed, high numbers of IFN-�-secreting cells were detected among liver iNKT cells 1 h after�-GalCer treatment (18). Consistent with this notion, IFN-�was detected in the peripheral blood of C57BL/6, but notIL-15�/� and SJL, mice after in vivo administration of�-GalCer (21, 43, 54; Emoto et al., unpublished). It is pos-sible that iNKT cells infiltrate the peritoneal cavity after�-GalCer treatment. Because iNKT cells were undetectable inthe peritoneal cavity after �-GalCer treatment not only at thecellular level but also at the mRNA level (this study) andbecause most L. monocytogenes bacteria are trapped in theliver and spleen immediately after i.p. infection (see Fig. 1), weassume that IFN-� detected in the peritoneal cavity is derivedfrom other organs, such as the liver and spleen.
NO participates in the killing of L. monocytogenes in M�(48, 52, 69, 71). Consistent with this, considerable NO levelswere detected in culture supernatants of peritoneal M� from�-GalCer-treated mice. However, the enhanced listericidal ac-tivity of peritoneal M� by �-GalCer was only marginally af-fected by NO inhibitor. At first sight, these findings seem toargue against a major role of NO in killing of L. monocyto-genes. However, as shown here, the escape of L. monocytogenesfrom the phagosome into the cytosol was not detectable inperitoneal M�. Since (i) NO is produced in the cytosol (45, 60)and (ii) killing of L. monocytogenes in TG-induced peritonealM� that permit escape of L. monocytogenes from the phago-some into the cytosol was inhibited by NO inhibitor (Emoto etal., unpublished), it is possible that NO plays only a minor rolein prompt phagosomal killing as observed in our model.
�-GalCer promoted a profound respiratory burst in M�after ingestion of opsonized L. monocytogenes. ROI are pro-duced by M� in response to L. monocytogenes infection, andthey play a central role in listerial killing within phagosomes(52, 53, 59). We therefore assume that enhanced listerial kill-ing in M� by �-GalCer is similarly caused by rapid and abun-dant production of ROI.
Further, in our hands, actin tails were undetectable in resi-dent peritoneal M� even 4 h after L. monocytogenes infectionin vitro. This finding could be taken as evidence against listerialescape from the phagosome into the cytosol (10, 22, 26, 42, 62,65). However, actin tails were detected in TG-induced M�already 1 h p.i. in vitro. We thus consider the possibility that L.monocytogenes escapes from the phagosome in inflammatory,
but not resident, peritoneal M�. Resident and inflammatoryM� differ in maturation, differentiation, and activation status(14, 34, 57, 68, 72, 73, 75). Bone marrow-derived monocytesare immature compared to resident M� (8, 11, 47, 51, 77), andM� colony-stimulating factor provides an essential signal forM� maturation (8, 11, 47, 51, 77). After the i.p. administrationof TG or proteose peptone, monocytes from bone marrowinfiltrate the peritoneal cavity (2, 9). We assume that L. mono-cytogenes escapes from the phagosome into the cytosol infreshly immigrant monocytes, but not in resident peritonealM�. Consistent with this notion, escape of L. monocytogenesfrom the phagosome has been described to occur in bonemarrow-derived M� (32, 63). LLO is essential for the escape ofL. monocytogenes from the phagosome into the cytosol (10, 22,26, 42, 62, 65), and its activity is highly dependent on low pH(3, 24, 25, 66) and on the abundance of ROI (49, 59). Bacterialnumbers ingested by M� were comparable for the hly-express-ing strain L. monocytogenes EGD and for the L. monocytogenes�hly strain (see Fig. 3B and C), indicating that phagocytosisoccurred independently of LLO. The numbers of engulfed L.monocytogenes bacteria were higher in TG-induced M� than inresident peritoneal M� (see Fig. 3B and C). We assume thatinflammatory M� have a higher phagocytic potential than res-ident peritoneal M�. Since L. monocytogenes escaped from thephagosome into the cytosol in TG-induced M� but not resi-dent peritoneal M�, we assume that killing capacities are in-fluenced by the degree of escape from the phagosome into thecytosol. Because bacterial escape from the phagosome into thecytosol is determined by whether the actin tail can be detectedor not, it would be better to verify this with a confocal micro-scope to obtain direct evidence.
In conclusion, we describe here profound bactericidal activ-ities of M� after �-GalCer treatment. M� activation was me-diated by IFN-� from iNKT cells. Our findings therefore define�-GalCer as an anti-intracellular bacterial agent.
ACKNOWLEDGMENTS
This study was supported by a Grant-in-Aid for Scientific Research(no. 17590383) from the Japan Society for the Promotion of Science;The Waksman Foundation of Japan, Inc.; the Japan Research Foun-dation of Clinical Pharmacology; and the German Science Foundation(SFB421).
We are grateful to M. Kronenberg and Kirin Pharma Co., Ltd., forproviding mouse CD1d/�2-microglobulin-expressing baculovirus and�-GalCer, respectively. We thank Y. Shimozawa for technical assis-tance and M. Staber for MAb purification. We also thank M. L.Grossman for critical reading of the manuscript.
REFERENCES
1. Arase, H., N. Arase, K. Nakagawa, R. A. Good, and K. Onoe. 1993. NK1.1
CD4 CD8� thymocytes with specific lymphokine secretion. Eur. J. Immu-nol. 23:307–310.
2. Baker, L. A., and P. A. Campbell. 1980. Thioglycolate medium decreasesresistance to bacterial infection in mice. Infect. Immun. 27:455–460.
3. Beauregard, K. E., K.-D. Lee, R. J. Collier, and J. A. Swanson. 1997. pH-dependent perforation of macrophage phagosomes by listeriolysin O fromListeria monocytogenes. J. Exp. Med. 186:1159–1163.
4. Beckerman, K. P., H. W. Rogers, J. A. Corbett, R. D. Schreiber, M. L.McDaniel, and E. R. Unanue. 1993. Release of nitric oxide during the Tcell-independent pathway of macrophage activation. J. Immunol. 150:888–895.
5. Behlke, A. M., T. J. Henkel, S. J. Anderson, N. C. Lan, L. Hood, V. L.Braciale, T. J. Braciale, and D. Y. Loh. 1987. Expression of a murine poly-clonal T-cell receptor marker correlates with the use of specific members ofthe V�8 gene segment subfamily. J. Exp. Med. 165:257–262.
6. Bendelac, A., P. B. Savage, and L. Teyton. 2007. The biology of NKT cells.Annu. Rev. Immunol. 25:297–336.
2674 EMOTO ET AL. INFECT. IMMUN.
on May 14, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

7. Bogdan, C., Y. Vodovotz, J. Paik, Q. W. Xie, and C. Nathan. 1993. Traces ofbacterial lipopolysaccharide suppress IFN-�-induced nitric oxide synthasegene expression in primary mouse macrophages. J. Immunol. 151:301–309.
8. Cinatl, J., E. Paluska, V. Chudomel, V. Malaskova, and M. Elleder. 1982.Culture of macrophage cell lines from normal mouse bone marrow. Nature298:388–389.
9. Cohn, Z. A. 1978. Activation of mononuclear phagocytes: fact, fancy, andfuture. J. Immunol. 121:813–816.
10. Cossart, P., M. F. Vicente, J. Mengaud, F. Baquero, J. C. Perez-Diaz, and P.Berche. 1989. Listeriolysin O is essential for virulence of Listeria monocyto-genes: direct evidence obtained by gene complementation. Infect. Immun.57:3629–3636.
11. Dai, X.-M., G. R. Ryan, A. J. Hapel, M. G. Dominguez, R. G. Russell, V.Sylvestre, and E. R. Stanley. 2002. Targeted disruption of the mouse colony-stimulating factor 1 receptor gene results in osteopetrosis, mononuclearphagocyte deficiency, increased primitive progenitor cell frequencies, andreproductive defects. Blood 99:111–120.
12. Drevets, D. A., B. P. Canono, and P. A. Campbell. 1992. Listericidal andnonlistericidal mouse macrophages differ in complement receptor type 3-me-diated phagocytosis of L. monocytogenes and in preventing escape of thebacteria into the cytoplasm. J. Leukoc. Biol. 52:70–79.
13. Drevets, D. A., B. P. Canono, P. J. M. Leenen, and P. A. Campbell. 1994.Gentamicin kills intracellular Listeria monocytogenes. Infect. Immun. 62:2222–2228.
14. Edelson, P. J., and Z. A. Cohn. 1976. 5-Nucleotidase activity of mouseperitoneal macrophages. I. Synthesis and degradation in resident and inflam-matory populations. J. Exp. Med. 144:1581–1595.
15. Emoto, M., Y. Emoto, I. B. Buchwalow, and S. H. E. Kaufmann. 1999.Induction of IFN-�-producing CD4 natural killer T cells by Mycobacteriumbovis bacillus Calmette-Guerin. Eur. J. Immunol. 29:650–659.
16. Emoto, M., Y. Emoto, and S. H. E. Kaufmann. 1995. IL-4 producing CD4
TCR��int liver lymphocytes: influence of thymus, �2-microglobulin, andNK1.1 expression. Int. Immunol. 7:1729–1739.
17. Emoto, M., Y. Emoto, and S. H. E. Kaufmann. 1995. Interleukin-4-producingCD4 NK1.1 TCR�/�intermediate liver lymphocytes are downregulated byListeria monocytogenes. Eur. J. Immunol. 25:3321–3325.
18. Emoto, M., Y. Emoto, I. Yoshizawa, E. Kita, T. Shimizu, R. Hurwitz, V.Brinkmann, and S. H. E. Kaufmann. 2010. �-GalCer ameliorates listeriosisby accelerating infiltration of Gr-1 cells into the liver. Eur. J. Immunol.40:1–15.
19. Emoto, M., I. Yoshizawa, Y. Emoto, M. Miamoto, R. Hurwitz, and S. H. E.Kaufmann. 2006. Rapid development of a gamma interferon-secreting gly-colipid/CD1d-specific V�14 NK1.1� T-cell subset after bacterial infection.Infect. Immun. 74:5903–5913.
20. Emoto, M., J. Zerrahn, M. Miyamoto, B. Perarnau, and S. H. E. Kaufmann.2000. Phenotypic characterization of CD8 NKT cells. Eur. J. Immunol.30:2300–2311.
21. Fujii, S., K. Shimizu, M. Kronenberg, and R. M. Steinman. 2002. ProlongedIFN-�-producing NKT response induced with �-galactosylceramide-loadedDCs. Nat. Immunol. 3:867–874.
22. Gaillard, J. L., P. Berche, and P. Sansonetti. 1986. Transposon mutagenesisas a tool to study the role of hemolysin in the virulence of Listeria monocy-togenes. Infect. Immun. 52:50–55.
23. Gemmell, C. G., and C. W. Ford. 2002. Virulence factor expression byGram-positive cocci exposed to subinhibitory concentrations of linezolid. J.Antimicrob. Chemother. 50:665–672.
24. Geoffroy, C., J.-L. Gaillard, J. E. Alouf, and P. Berche. 1987. Purification,characterization, and toxicity of the sulfhydryl-activated hemolysin listerio-lysin O from Listeria monocytogenes. Infect. Immun. 55:1641–1646.
25. Glomski, I. J., M. M. Gedde, A. W. Tsang, J. A. Swanson, and D. A. Portnoy.2002. The Listeria monocytogenes hemolysin has an acidic pH optimum tocompartmentalize activity and prevent damage to infected host cells. J. CellBiol. 156:1029–1038.
26. Goldfine, H., and S. J. Wadsworth. 2002. Macrophage intracellular signalinginduced by Listeria monocytogenes. Microbes Infect. 4:1335–1343.
27. Gonzalez-Aseguinolaza, G., C. de Oliveira, M. Tomaska, S. Hong, O. Bruna-Romero, T. Nakayama, M. Taniguchi, A. Bendelac, L. van Kaer, Y. Koezuka,and M. Tsuji. 2000. �-Galactosylceramide-activated V�14 natural killer Tcells mediate protection against murine malaria. Proc. Natl. Acad. Sci.U. S. A. 97:8461–8466.
28. Gonzalez-Aseguinolaza, G., L. van Kaer, C. C. Bergmann, J. M. Wilson, J.Schmieg, M. Kronenberg, T. Nakayama, M. Taniguchi, Y. Koezuka, and M.Tsuji. 2002. Natural killer T-cell ligand �-galactosylceramide enhances pro-tective immunity induced by malaria vaccines. J. Exp. Med. 195:617–624.
29. Grubor-Bauk, B., A. Simmons, G. Mayrhofer, and P. G. Speck. 2003. Im-paired clearance of herpes simplex virus type 1 from mice lacking CD1d orNKT cells expressing the semivariant V�14-J.�281 TCR. J. Immunol. 170:1430–1434.
30. Hara, H., I. Kawamura, T. Nomura, T. Tominaga, K. Tsuchiya, and M.Mitsuyama. 2007. Cytolysin-dependent escape of the bacterium from thephagosome is required but not sufficient for induction of the Th1 immune
response against Listeria monocytogenes infection: distinct role of listeriolysinO determined by cytolysin gene replacement. Infect. Immun. 75:3791–3801.
31. Harty, J. T., and M. J. Bevan. 1995. Specific immunity to Listeria monocy-togenes in the absence of IFN-�. Immunity 3:109–117.
32. Higginbotham, J. N., and S. B. Pruett. 1994. Assessment of the correlationbetween nitrite concentration and listericidal activity in cultures of residentand elicited murine macrophages. Clin. Exp. Immunol. 97:100–106.
33. Hong, S., M. T. Wilson, I. Serizawa, L. Wu, N. Singh, O. V. Naidenko, T.Miura, T. Haba, D. C. Scherer, J. Wei, M. Kronenberg, Y. Koezuka, and L.van Kaer. 2001. The natural killer T-cell ligand �-galactosylceramide pre-vents autoimmune diabetes in non-obese diabetic mice. Nat. Med. 7:1052–1056.
34. Hoover, D. L., and C. A. Nacy. 1984. Macrophage activation to kill Leish-mania tropica: defective intracellular killing of amastigotes by macrophageselicited with sterile inflammatory agents. J. Immunol. 132:1487–1493.
35. Huang, S., W. Hendriks, A. Althage, S. Hemmi, H. Bluethmann, R. Kamijo,J. Vilcek, R. M. Zinkernagel, and M. Aguet. 1993. Immune response in micethat lack the interferon-� receptor. Science 259:1693–1694.
36. Ito, H., N. Koide, A. Morikawa, F. Hassan, S. Islam, G. Tumurkhuu, I. Mori,T. Yoshida, S. Kakumu, H. Moriwaki, and T. Yokochi. 2005. Augmentationof lipopolysaccharide-induced nitric oxide production by �-galactosylceram-ide in mouse peritoneal cells. J. Endotoxin Res. 11:213–219.
37. Kakimi, K., L. G. Guidotti, Y. Koezuka, and F. V. Chisari. 2000. Naturalkiller T-cell activation inhibits hepatitis B virus replication in vivo. J. Exp.Med. 192:921–930.
38. Karupiah, G., Q. W. Xie, R. M. Buller, C. Nathan, C. Duarte, and J. D.MacMicking. 1993. Inhibition of viral replication by interferon-�-inducednitric oxide synthase. Science 261:1445–1448.
39. Kawakami, K., Y. Kinjo, S. Yara, Y. Koguchi, K. Uezu, T. Nakayama, M.Taniguchi, and A. Saito. 2001. Activation of V�14 natural killer T cells by�-galactosylceramide results in development of Th1 response and local hostresistance in mice infected with Cryptococcus neoformans. Infect. Immun.69:213–220.
40. Kawano, T., J. Cui, Y. Koezuka, I. Toura, Y. Kaneko, K. Motoki, H. Ueno, R.Nakagawa, H. Sato, E. Kondo, H. Koseki, and M. Taniguchi. 1997. CD1d-restricted and TCR-mediated activation of V�14 NKT cells by glycosylcer-amides. Science 278:1626–1629.
41. Kawano, T., J. Cui, Y. Koezuka, I. Toura, Y. Kaneko, H. Sato, E. Kondo, M.Harada, H. Koseki, T. Nakayama, Y. Tanaka, and M. Taniguchi. 1998.Natural killer-like nonspecific tumor cell lysis mediated by specific ligand-activated V�14 NKT cells. Proc. Natl. Acad. Sci. U. S. A. 95:5690–5693.
42. Kayal, S., and A. Charbit. 2006. Listeriolysin O: a key protein of Listeriamonocytogenes with multiple functions. FEMS Microbiol. Rev. 30:514–529.
43. Kojo, S., C. Elly, Y. Harada, W. Y. Langdon, M. Kronenberg, and Y. Liu.2009. Mechanisms of NKT cell anergy induction involve Cbl-b-promotedmonoubiquitination of CARMA1. Proc. Natl. Acad. Sci. U. S. A. 106:17847–17851.
44. Kumar, H., A. Belperron, S. W. Barthold, and L. K. Bockenstedt. 2000.CD1d deficiency impairs murine host defense against the spirochete, Borreliaburgdorferi. J. Immunol. 165:4797–4801.
45. Lim, M. H., D. Xu, and S. J. Lippard. 2006. Visualization of nitric oxide inliving cells by a copper-based fluorescent probe. Nat. Chem. Biol. 2:375–380.
46. Liu, J., Z. Liu, S. Chuai, and X. Shen. 2003. Phospholipase C and phospha-tidylinositol 3-kinase signaling are involved in the exogenous arachidonicacid-stimulated respiratory burst in human neutrophils. J. Leukoc. Biol.74:428–437.
47. Loberas, J., C. Soler, and A. Celada. 1998. Dexamethasone enhances mac-rophage colony stimulating factor- and granulocyte macrophage colony stim-ulating factor-stimulated proliferation of bone marrow-derived macro-phages. Int. Immunol. 10:593–599.
48. MacMicking, D. J., C. Nathan, G. Hom, N. Chartrain, D. S. Fletcher, M.Trumbauer, K. Stevens, Q. Xie, K. Sokol, N. Hutchinson, H. Chen, and J. S.Mudgett. 1995. Altered responses to bacterial infection and endotoxic shockin mice lacking inducible nitric oxide synthase. Cell 81:641–650.
49. Makino, M., M. Kawai, I. Kawamura, M. Fujita, F. Gejo, and M. Mit-suyama. 2005. Involvement of reactive oxygen intermediate in the enhancedexpression of virulence-associated genes of Listeria monocytogenes insideactivated macrophages. Microbiol. Immunol. 49:805–811.
50. Matsuda, J. L., O. V. Naidenko, L. Gapin, T. Nakayama, M. Taniguchi, C.-R.Wang, Y. Koezuka, and M. Kronenberg. 2000. Tracking the response ofnatural killer T cells to glycolipid antigens using CD1d tetramer. J. Exp.Med. 192:741–753.
51. Melcher, M., B. Unger, U. Schmidt, I. A. Rajantie, K. Alitalo, and W.Ellmeier. 2008. Essential roles for the Tec family kinases Tec and Btk inM-CSF receptor signaling pathways that regulate macrophage survival.J. Immunol. 180:8048–8056.
52. Muller, M., R. Althaus, D. Frohlich, K. Frei, and H.-P. Eugster. 1999.Reduced antilisterial activity of TNF-deficient bone marrow-derived macro-phage is due to impaired superoxide production. Eur. J. Immunol. 29:3089–3097.
53. Myers, J. T., A. W. Tsang, and J. A. Swanson. 2003. Localized reactive
VOL. 78, 2010 AMELIORATION OF LISTERIOSIS BY �-GalCer 2675
on May 14, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

oxygen and nitrogen intermediates inhibit escape of Listeria monocytogenesfrom vacuoles in activated macrophages. J. Immunol. 171:5447–5453.
54. Nakaya, M., M. Hashimoto, R. Nakagawa, Y. Wakabayashi, T. Ishizaki, I.Takada, K. Komai, H. Yoshida, and A. Yoshimura. 2009. SOCS3 in T andNKT cells negatively regulates cytokine production and ameliorates ConA-induced hepatitis. 183:7047–7053.
55. Nieuwenhuis, E. E. S., T. Matsumoto, M. Exley, R. A. Schleipman, J. Glick-man, D. T. Bailey, N. Corazza, S. P. Colgan, A. B. Onderdonk, and R. S.Blumberg. 2002. CD1d-dependent macrophage-mediated clearance ofPseudomonas aeruginosa from lung. Nat. Med. 8:588–593.
56. North, R. J., and J. W. Conlan. 1998. Immunity to Listeria monocytogenes.Chem. Immunol. 70:1–20.
57. Oghiso, Y., Y. Yamada, and Y. Shibata. 1992. Exudation of proliferativemacrophages in local inflammation in the peritoneum. J. Leukoc. Biol.52:421–424.
58. Ohteki, T., S. Ho, H. Suzuki, T. W. Mak, and P. S. Ohashi. 1997. Role forIL-15/IL-15 receptor �-chain in natural killer 1.1 T-cell receptor-�� celldevelopment. J. Immunol. 159:5931–5935.
59. Ohya, S., Y. Tanabe, M. Makino, T. Nomura, H. Xiong, M. Arakawa, and M.Mitsuyama. 1998. The contributions of reactive oxygen intermediates andreactive nitrogen intermediates to listericidal mechanisms differ in macro-phages activated pre- and postinfection. Infect. Immun. 66:4043–4049.
60. Panda, K., M. Chawla-Sarkar, C. Santos, T. Koeck, S. C. Erzurum, J. F.Parkinson, and D. J. Stuehr. 2005. Visualizing inducible nitric-oxide syn-thase in living cells with a heme-binding fluorescent inhibitor. Proc. Natl.Acad. Sci. U. S. A. 102:10117–10122.
61. Pinto, A. J., D. Stewart, N. van Rooijen, and P. S. Morahan. 1991. Selectivedepletion of liver and splenic macrophages using liposomes encapsulatingthe drug dichloromethylene diphosphonate: effects on antimicrobial resis-tance. J. Leukoc. Biol. 49:579–586.
62. Portnoy, D. A., P. S. Jacks, and D. J. Hinrichs. 1988. Role of hemolysin forthe intracellular growth of Listeria monocytogenes. J. Exp. Med. 167:1459–1471.
63. Rouquette, C., C. de Chastellier, S. Nair, and P. Berche. 1998. The ClpCATPase of Listeria monocytogenes is a general stress protein required forvirulence and promoting early bacterial escape from the phagosome ofmacrophages. Mol. Microbiol. 27:1235–1245.
64. Schmieg, J., G. Yang, R. W. Franck, and M. Tsuji. 2003. Superior protectionagainst malaria and melanoma metastases by a C-glycoside analogue of thenatural killer T-cell ligand �-galactosylceramide. J. Exp. Med. 198:1631–1641.
65. Schnupf, P., and D. A. Portnoy. 2007. Listeriolysin O: a phagosome-specificlysin. Microb. Infect. 9:1176–1187.
66. Schuerch, D. W., E. M. Wilson-Kubalek, and R. K. Tweten. 2005. Molecularbasis of listeriolysin O pH dependence. Proc. Natl. Acad. Sci. U. S. A.102:12537–12542.
67. Sharif, S., G. A. Arreaza, P. Zucker, Q.-S. Mi, J. Sondhi, O. V. Naidenko, M.Kronenberg, Y. Koezuka, and T. L. Delovitch. 2001. Activation of naturalkiller T cells by �-galactosylceramide treatment prevents the onset andrecurrence of autoimmune type 1 diabetes. Nat. Med. 7:1057–1062.
68. Shaw, D. R., and F. M. Griffin, Jr. 1982. Thioglycolate-elicited mouse peri-toneal macrophages are less efficient than resident macrophages in antibody-dependent cell-mediated cytolysis. J. Immunol. 128:433–440.
69. Shiloh, M. U., J. D. MacMicking, S. Nicholson, J. E. Brause, S. Potter, M.Marino, F. Fang, M. Dinauer, and C. Nathan. 1999. Phenotype of mice andmacrophages deficient in both phagocyte oxidase and inducible nitric oxidesynthase. Immunity 10:29–38.
70. Singh, A. K., M. T. Wilson, S. Hong, D. Olivares-Villagomez, C. Du, A. K.Stanic, S. Joyce, S. Sriram, Y. Koezuka, and L. van Kaer. 2001. Natural killerT-cell activation protects mice against experimental autoimmune encepha-lomyelitis. J. Exp. Med. 194:1801–1811.
71. Stuehr, D. J., and C. F. Nathan. 1989. Nitric oxide: a macrophage productresponsible for cytostasis and respiratory inhibition in tumor target cells. J.Exp. Med. 169:1543–1555.
72. Takemura, R., and Z. Werb. 1984. Modulation of apoprotein E secretion inresponse to receptor-mediated endocytosis in resident and inflammatorymacrophages. J. Exp. Med. 159:167–178.
73. Takemura, R., and Z. Werb. 1984. Regulation of elastase and plasminogenactivator secretion in resident and inflammatory macrophages by receptorsfor the Fc domain of immunoglobulin G. J. Exp. Med. 159:152–166.
74. Tanabe, Y., H. Xiong, T. Nomura, M. Arakawa, and M. Mitsuyama. 1999.Induction of protective T cells against Listeria monocytogenes in mice byimmunization with a listeriolysin O-negative avirulent strain of bacteria andliposome-encapsulated listeriolysin O. Infect. Immun. 67:568–575.
75. Tannenbaum, S. C., L. Nurmi-McKernan, and M. T. Largen. 1987. Differ-ential protein synthesis by murine peritoneal macrophage elicited by variousstimuli. J. Leukoc. Biol. 41:527–538.
76. Vazquez-Boland, J. A., M. Kuhn, P. Berche, T. Chakraborty, G. Domínguez-Bernal, W. Goebel, B. Gonzalez-Zorn, J. Wehland, and J. Kreft. 2001. Lis-teria pathogenesis and molecular virulence determinants. Clin. Microbiol.Rev. 14:584–640.
77. Wiktor-Jedrzejczak, W., A. Bartocci, A. W. Ferrante, Jr., A. Ahmed-Ansari,K. W. Sell, J. W. Pollard, and E. R. Stanley. 1990. Total absence of colony-stimulating factor 1 in the macrophage-deficient osteopetrotic (op/op)mouse. Proc. Natl. Acad. Sci. U. S. A. 87:4828–4832.
78. Yoshimoto, T., A. Bendelac, J. Hu-Li, and W. E. Paul. 1995. Defective IgEproduction by SJL mice is linked to the absence of CD4 , NK1.1 T cellsthat promptly produce interleukin 4. Proc. Natl. Acad. Sci. U. S. A. 92:11931–11934.
79. Yoshimoto, T., and W. E. Paul. 1994. CD4pos, NK1.1pos T cells promptlyproduce interleukin 4 in response to in vivo challenge with anti-CD3. J. Exp.Med. 179:1285–1295.
Editor: J. B. Bliska
2676 EMOTO ET AL. INFECT. IMMUN.
on May 14, 2018 by guest
http://iai.asm.org/
Dow
nloaded from