caracterizaciÓn de la comunidad de aves de un … · para el muestreo de aves se utilizaron las...
TRANSCRIPT

i
UNIVERSIDAD NACIONAL
SISTEMA DE ESTUDIOS DE POSGRADO
PROGRAMA REGIONAL EN MANEJO DE VIDA SILVESTRE PARA
MESOAMÉRICA Y EL CARIBE
CARACTERIZACIÓN DE LA COMUNIDAD DE AVES DE UN BOSQUE
TROPICAL MUY HÚMEDO EN EL CARIBE NORTE DE COSTA RICA
María Alejandra Maglianesi
Heredia, Costa Rica
Noviembre del 2004
Tesis sometida a consideración del Tribunal Examinador de posgrado en
Conservación y Manejo de Vida Silvestre para optar al grado de Magister Scientiae

ii
CARACTERIZACIÓN DE LA COMUNIDAD DE AVES DE UN BOSQUE
TROPICAL MUY HÚMEDO EN EL CARIBE NORTE DE COSTA RICA
María Alejandra Maglianesi
Tesis en la modalidad de artículos científicos presentada para optar al grado de
Magister Scientiae en Conservación y Manejo de Vida Silvestre. La misma cumple
con los requisitos establecidos por el Sistema de Estudios de Posgrado de la
Universidad Nacional, Heredia, Costa Rica.

iii
Miembros del Tribunal Examinador
_____________________________________________
, Consejo Central de Posgrado
__________________________________ ______________________
M.Sc. Jorge Fallas, Director del PRMVS M.Sc. Michael McCoy, Tutor
__________________________ _______________________
Dr. Gilbert Barrantes, Asesor Dr. C. John Ralph, Asesor
_____________________________
Dr. Jaime Rau, Asesor
________________________________
María Alejandra Maglianesi, Sustentante

iv
RESUMEN
En la presente tesis se realizó una caracterización de la comunidad de aves
asociada a cinco sitios diferentes en los alrededores de Tortuguero, en el Caribe norte de
Costa Rica durante los meses de agosto hasta diciembre del 2002. Medidas de abundancia,
composición y riqueza de especies residentes y migratorias fueron tomadas y comparadas
entre sitios intervenidos y no intervenidos, con la finalidad de sugerir estrategias
apropiadas de manejo. Para el muestreo de aves se utilizaron las técnicas de captura con
redes de niebla y conteos de búsqueda de aves y se estimó el número de frutos presentes
para las aves en cuatro de los sitios. Se registraron un total de 128 especies de aves, de las
cuales el 28.1% fueron especies migratorias. Las especies residentes tienden a evitar más
los hábitats intervenidos que las especies migratorias. El mantenimiento de hábitats
primarios debería ser una prioridad debido al mayor tiempo que implica su formación y
porque muchas especies de aves, especialmente residentes son dependientes de este tipo
de hábitats. Sin embargo, los esfuerzos de conservación dirigidos a maximizar la riqueza
de especies de aves deberían incluir la preservación de los sitios intervenidos ya que
proveen hábitats importantes para las especies de aves, especialmente para migratorias.
Por otro lado se compararon los resultados obtenidos por las dos técnicas de muestreo
empleadas. Para las comparaciones entre métodos se consideraron 110 especies en los
cinco sitios, de las cuales 61 fueron comunes a ambos métodos. La tasa promedio de
detección de especies fue mayor con los conteos de búsqueda de aves que con capturas.
Para el total de especies registradas por ambos métodos, la abundancia relativa mostró una
correlación positiva significativa y para el 89% de las especies los estimados de
abundancia relativa fueron precisos para ambos métodos. Es necesario reconocer los
sesgos asociados a las técnicas de muestreo de aves e interpretar cuidadosamente los
resultados antes de tomar decisiones de manejo. A pesar de que el método de búsqueda de
aves resulta más eficiente en cuanto a tiempo y costos, debería ser utilizado en
combinación con otros métodos para obtener una representación más completa de la
comunidad de aves en los trópicos.

v
ABSTRACT
Bird community was examined in five sites, two disturbed and three undisturbed
wet lowland forest, since August until late December 2002 near Tortuguero, northern
Caribbean coast in Costa Rica. Relative abundance, composition and richness of resident
and migrant landbirds among different sites were measured and compared in order to
suggest appropriate management strategies. To sample birds I used mist-netting and area
search count techniques. Number of fruits was measured in four sites during the same
period that birds were netted and observed. I recorded a total of 128 species of birds by
both methods from the five sites. More resident species avoid disturbed habitats. In
contrast, migrant species more frequently occur at disturbed habitat. Maintenance of
undisturbed areas should be the highest priority, because the vegetation of mature forest
require relatively extensive periods of time to reach their former states after cutting, and
because many bird species are dependent of this type of habitat. However, conservation
efforts directed toward maximizing bird species richness should include disturbed areas
their protection from agricultural or urban areas, because these provide important habitat
to bird species. I compared the results of mist-net and area search bird surveys in order to
determine if differences in abundance estimated, composition and richness of landbirds
varied with the survey method. I considered 110 species across the five sites. There were
61 species detected by both methods. Mean species detection rate recorded with area
searches was greater than with mist nets. However, mist-netting detected a greater
percentage of migratory species than area searches. For the total species, relative
abundance estimates from mist-netting and area searches were significantly correlated.
Over 89% of the 61 species the two methods were consistent in measuring relative
abundance. Individuals planning bird surveys should recognize biases and to be careful in
interpret results before to take management decision involving bird populations. Despite
of area search alone would be the most time efficient and least costly technique, should be
used combining other methods for to have a more relatively complete description of the
bird community in the tropics.

vi
TABLA DE CONTENIDO
Página
RESUMEN .................................................................................................................. iv
ABSTRACT ................................................................................................................ v
LISTA DE CUADROS ............................................................................................... x
LISTA DE FIGURAS ................................................................................................ xii
PRESENTACIÓN ...................................................................................................... xv
PRESENTATION ...................................................................................................... xvi
INTRODUCCIÓN GENERAL ................................................................................... 1
LITERATURA CITADA ............................................................................................ 3
ARTÍCULO I: Influencia de las características de hábitat de
la comunidad de aves de un bosque tropical muy húmedo
en el Caribe norte de Costa Rica.
(Hoja titular, formato de tesis) ........................................................................... 7
(Hoja titular, formato de artículo científico) ...................................................... 8
RESUMEN .................................................................................................................. 9
ABSTRACT ............................................................................................................... 10
INTRODUCCIÓN ...................................................................................................... 11
MÉTODOS ................................................................................................................. 13
Área de estudio ....................................................................................................... 13
Sitios de estudio ................................................................................................. 13
Muestreo de aves .................................................................................................... 14
Captura con redes de niebla ................................................................................ 14
Conteos de búsqueda de aves ............................................................................. 15
Muestreo de frutos .................................................................................................. 15
Análisis estadísticos ............................................................................................... 16
Curvas de acumulación de especies ................................................................... 16
Riqueza de especies ............................................................................................ 16
Abundancia relativa ............................................................................................ 17

vii
TABLA DE CONTENIDO (continuación)
Categorías de abundancia ................................................................................... 17
Similitud entre comunidades .............................................................................. 17
Patrones de movimiento y recaptura .................................................................. 17
Abundancia de frutos y relación con la riqueza de especies .............................. 17
de aves
Uso de hábitat ..................................................................................................... 18
RESULTADOS .......................................................................................................... 18
Especies más comunes ....... .................................................................................. 18
Species-effort curves......................................................….……………............... 16
Riqueza de especies ............................................................................................... 20
Abundancia relativa............................................................................................... 20
Índices de similitud ............................................................................................... 21
Patrones de movimiento y recaptura ..................................................................... 21
Plantas y especies de aves frugívoras .................................................................... 22
Uso de hábitat ......................................................................................................... 22
DISCUSIÓN ............................................................................................................... 23
Curvas de acumulación de especies ...................................................................... 23
Patrones de distribución y abundancia .................................................................. 23
Influencia de la abundancia de frutos .................................................................... 24
Porcentajes de recaptura pueden indicar preferencia por hábitat .......................... 25
Especies migratorias más comunes en sitios intervenidos .................................... 26
Sobre las decisiones de manejo .............................................................................. 28
Conclusiones y recomendaciones ........................................................................... 29
AGRADECIMIENTOS ............................................................................................. 30
LITERATURA CITADA ........................................................................................... 31
CUADROS ................................................................................................................. 37
LISTA DE FIGURAS ................................................................................................ 45
APÉNDICE A ............................................................................................................ 63
ARTÍCULO II: Comparación de dos técnicas de muestreo:
Página

viii
TABLA DE CONTENIDO (continuación)
captura con redes de niebla y búsqueda de aves, en un bosque
tropical muy húmedo en el Caribe norte de Costa Rica.
(Hoja titular, formato de tesis) ........................................................................ 70
(Hoja titular, formato de artículo científico) .................................................. 71
ABSTRACT ............................................................................................................... 72
SINOPSIS ................................................................................................................... 73
INTRODUCCIÓN ..................................................................................................... 74
MÉTODOS ................................................................................................................. 76
Área de estudio ....................................................................................................... 76
Sitios de estudio ................................................................................................. 77
Muestreo de aves .................................................................................................... 78
Captura con redes de niebla ................................................................................ 78
Conteos de búsqueda de aves ............................................................................. 78
Análisis estadísticos ............................................................................................... 79
RESULTADOS .......................................................................................................... 81
Estimación de las especies por método .................................................................. 81
Curvas de acumulación de especies ....................................................................... 81
Tasa de detección de especies ................................................................................ 82
Porcentaje de especies migratorias ......................................................................... 82
Composición de especies por familia ..................................................................... 83
Consistencia entre métodos .................................................................................... 83
DISCUSIÓN ............................................................................................................... 84
Presencia vs. ausencia de especies .......................... .................................................. 84
Abundancia relativa .................................................................................................... 85
Conclusiones y recomendaciones ............................................................................... 86
AGRADECIMIENTOS ............................................................................................. 87
LITERATURA CITADA ........................................................................................... 88
CUADROS ................................................................................................................. 92
LISTA DE FIGURAS ................................................................................................ 97
Página

ix
TABLA DE CONTENIDO (continuación)
APÉNDICE A ........................................................................................................... 107
SÍNTESIS .................................................................................................................. 113
APÉNDICE A: Instrucciones para autores para la revista
Wilson Bulletin ....................................................................................................... 115
APÉNDICE B: Instrucciones para autores para la revista
Journal of Field Ornithology .........................................................................….…. 120
Página

x
LISTA DE CUADROS
ARTÍCULO I
Cuadro Página
1. Esfuerzo de muestreo, número de individuos y especies
capturadas en cinco sitios en el caribe norte de Costa Rica ........................... 37
2. Esfuerzo de muestreo y número medio de individuos y
especies observados durante conteos de búsqueda de aves
en cinco sitios en el Caribe norte de Costa Rica ............................................ 38
3. Estadísticos de regresión del modelo de Clench para cinco
sitios en Caribe norte de Costa Rica ............................................................... 39
4. Coeficientes de similitud de Jaccard en cinco sitios en el
Caribe norte de Costa Rica ............................................................................. 40
5. Número de recapturas en cinco sitios en el Caribe norte
de Costa Rica .................................................................................................. 41
6. Porcentaje de recapturas de especies de aves residentes y
migratorias en cinco sitios en el Caribe norte de Costa Rica ......................... 42
7. Recapturas de especies de aves residentes en sitios diferentes
al de la captura inicial ..................................................................................... 43
8. Especies de plantas registradas durante conteos de frutos
en cuatro sitios en el Caribe norte de Costa Rica ........................................... 44
ARTÍCULO II
Cuadro
1. Número de especies capturadas en redes de niebla y observadas
en conteos de búsqueda de aves en cinco sitios de estudio en
el Caribe norte de Costa Rica ......................................................................... 92
2. Especies de aves detectadas por los métodos de captura
con redes y conteos de búsqueda de aves en cinco sitios en el

xi
LISTA DE CUADROS (continuación)
Cuadro Página
Caribe norte de Costa Rica ............................................................................. 93
3. Parámetros de regresión del modelo de Clench para cinco
sitios de estudio en el Caribe norte de Costa Rica .......................................... 96

xii
LISTA DE FIGURAS
ARTÍCULO I
Figura Página
1. Área de estudio ................................................................................................... 46
2. Curvas de acumulación de especies para especies de aves
capturadas basada en el modelo de Clench ....................................................... 47
3. Curvas de acumulación de especies para especies de aves
observadas basada en el modelo de Clench ....................................................... 48
4. Número medio de especies de aves residentes capturadas en
cinco sitios (intervalos de confianza son + HSD intervalos
Tukey) .......................... .................................................................................... 49
5. Número medio de especies de aves migratorias en cinco sitios
(intervalos de confianza son + HSD intervalos Tukey) ................................... 50
6. Tasa media de captura de aves residentes en cinco sitios de
estudio (intervalos de confianza son + HSD intervalos
Tukey) ......... ..................................................................................................... 51
7. Tasa media de captura de aves migratorias en cinco sitios de
estudio (intervalos de confianza son + HSD intervalos
Tukey) ............................................................................................................... 52
8. Tasa media de conteo de aves residentes y migratorias en cinco
sitios de estudio (intervalos de confianza are + HSD intervalos
Tukey) ............................................................................................................... 53
9. Categorías de abundancia basadas en tasas de captura ...................................... 54
10. Categorías de abundancia basadas en tasas de conteo ...................................... 55
11. Dendrograma de la riqueza de especies en cinco sitios
basado en el vecino más cercano, distancia cuadrado
Eucladiana ......................................................................................................... 56
12. Porcentajes de especies de aves residentes y migratorias

xiii
LISTA DE FIGURAS (continuación)
Figura Página
recapturadas en cinco sitios ............................................................................... 57
13. Número medio de frutos maduros en 50 m2 en cuatro sitios
en el Caribe norte de Costa Rica ....................................................................... 58
14. Número medio de frutos maduros a lo largo de cuatro meses .......................... 59
15. Número medio de especies de aves frugívoras en cuatro
sitios en los alrededores de Tortuguero ............................................................ 60
16. Porcentaje de especies registradas en un único sitio en el
Caribe norte de Costa Rica ................................................................................ 61
17. Porcentaje de especies en hábitats intervenido y no
intervenido en el Caribe norte de Costa Rica ..................................................... 62
ARTÍCULO II
Figura
1. Área de estudio con la ubicación de los sitios muestreados ............................... 98
2. Curva de acumulación de especies para especies de aves
capturadas basada en el modelo de Clench ....................................................... 99
3. Curva de acumulación de especies para especies de aves
observadas basada en el modelo de Clench ...................................................... 100
4. Asociación de la tasa de detección de especies de aves
residentes entre métodos, en cinco sitios del
Caribe norte de Costa Rica .............................................................................. 101
5. Asociación de las tasas medias de detección de especies de
aves migratorias entre métodos, en cinco sitios del Caribe
norte de Costa Rica ........................................................................................... 102
6. Tasa media de detección de especies en cinco sitios por los
métodos de captura con redes y conteos de búsqueda de aves
(intervalos de confianza son + intervalos de Tukey) ....................................... 103
7. Porcentaje medio de especies migratorias detectadas por

xiv
LISTA DE FIGURAS (continuación)
Figura Página
medio de captura con redes y conteos de búsqueda de aves
en cinco sitios del Caribe norte de Costa Rica (intervalos
de confianza son + intervalos de Tukey) .......................................................... 104
8. Número total de especies por familia detectadas por los métodos
de captura con redes y conteos de búsqueda de aves ........................................ 105
9. Relación entre la proporción de días en que las especies
fueron detectadas (PDD) por los métodos de captura con redes
y búsqueda de aves. Los datos fueron estandarizados y luego
transformados logarítmicamente (base 10). Ver Tabla 2 para
los códigos de las especies .............................................................................. 106

xv
PRESENTACIÓN
Esta tesis titulada “Caracterización de la Comunidad de Aves de un Bosque Tropical
muy Húmedo en el Caribe Norte de Costa Rica” está compuesto de dos artículos
independientes basados en el reglamento del Sistema de Estudios de Posgrado de la
Universidad Nacional (SEPUNA) en Heredia, Costa Rica.
El primer artículo se titula “Influencias de las Características del Hábitat sobre la
Comunidad de Aves de un Bosque Tropical muy Húmedo en el Caribe Norte de Costa
Rica” y sigue el formato requerido para publicar en la revista Wilson Bulletin.
El segundo artículo se titula “Comparación de dos Técnicas de Muestreo de Aves en
un Bosque Tropical muy Húmedo en el Caribe Norte de Costa Rica” y sigue el formato
requerido para publicar en la revista Journal of Field Ornithology.

xvi
PRESENTATION
This thesis is named as “Population and Community Characteristics of Resident and
Neotropical Migrant Landbirds in a Tropical Rain Forest of the Northern Caribbean of
Costa Rica” contains two articles based on the rules of the Postgraduate Studies System
from Universidad Nacional (SEPUNA) in Heredia, Costa Rica. I used the required Journal
format of the Wilson Bulletin in the first article and Journal of Field Ornithology in the
second article.
The first article is titled “Implication of Habitat Characteristics on Bird Community
in a Tropical Rain Forest of the Northern Caribbean of Costa Rica”. The second article is
titled “Comparison of two bird survey techniques used in a wet lowland forest of the
northern Caribbean of Costa Rica”.

1
INTRODUCCIÓN GENERAL
Costa Rica se encuentra entre los países que poseen una gran riqueza de especies en
su avifauna (más de 850 especies identificadas), entre las cuales se incluyen alrededor de
600 especies de residentes permanentes y más de 200 especies migratorias que visitan el
país cada año (Stiles 1983, Stiles y Skutch 1989, Wunderle y Waide 1993).
El área de Tortuguero se encuentra ubicada en la costa norte del Caribe de Costa
Rica. El alto número de especies de aves (44% de la avifauna total del país) (Widdowson
y Widdowson 2000) y el gran volumen de aves migratorias que pasan por este sitio
reflejan la gran importancia que tiene para la avifauna.
La identificación de los factores que regulan el tamaño de las poblaciones resulta
clave para comprender la dinámica poblacional de cualquier organismo. No obstante, la
determinación de tales factores reguladores constituye una difícil tarea para muchos
organismos, especialmente para las aves migratorias, para las cuales los recursos de sus
hábitats desempeñan frecuentemente un rol central en la regulación de sus poblaciones
(Latta y Faaborg 2002).
Las altas tasas de deforestación en las áreas tropicales y subtropicales que
constituyen el hábitat para las especies migratorias, indican que los sucesos que ocurren en
el Neotrópico podrían llegar a tener un gran impacto sobre las aves migratorias en el
futuro (Rappole y Powell 1986). Sin embargo, los hábitats con vegetación alterada que
han ido reemplazando los bosques de tierras bajas en muchas partes del Neotrópico,
pueden proveer un importante hábitat para las especies de aves (Karr 1971, Blake y
Loiselle 1991, Jiménez 1999). El tiempo de permanencia en las áreas de invierno puede
constituir una evidencia del uso e importancia de tales áreas para las especies migratorias.
Las fluctuaciones en la abundancia poblacional de las especies de aves pueden
deberse a variaciones temporales y espaciales en la abundancia de alimento, la cual varía
entre diferentes sitios (Karr y Freemark 1983, Stiles 1980). La importancia del alimento
como una hipótesis para explicar la variación en la abundancia poblacional de las aves se
ve reflejada en un número creciente de estudios realizados en la última década (Loiselle y

2
Blake 1991, Levey 1988). Sin embargo, el rol que desempeña este factor ha resultado ser
controversial en muchos casos.
Otros estudios han comparado la avifauna entre hábitats en diferentes estados de
conservación (Jiménez 1999). En el área de Tortuguero se estudió la asociación del hábitat
con el número de capturas de dos especies de saltarines (Pipridae) y el comportamiento
reproductivo de estas especies (Herrera 1998). Desde 1994 las poblaciones de aves están
siendo monitoreadas en diferentes sitios de Tortuguero (Ralph et al. 2002). Sin embargo,
la influencia de los frutos sobre la distribución y abundancia de las poblaciones de aves no
ha sido investigada aún.
Por otro lado, la descripción tanto de la composición como de los patrones de
abundancia y riqueza de especies de aves en un área en particular no es una tarea simple y
depende en gran medida del método empleado (Terborgh et al. 1990, Robinson et al.
2000). Además, las comparaciones entre hábitats pueden ser especialmente influenciadas
por los métodos (Blake y Loiselle 2001).
La técnica de captura con redes de niebla se ha utilizado ampliamente para
determinar patrones de abundancia relativa, diversidad y distribución de especies de aves.
Un método que se ha empezado a utilizar es el conteo de búsqueda de aves (Ralph et al.
1993, Slater 1994). Tanto el método de captura con redes como los métodos de
observación poseen ventajas y desventajas (Karr 1981, Karr 1990, Remsen y Good 1996)
y cada uno puede proveer diferentes perspectivas sobre la estructura de una comunidad
(Whitman et al. 1997, Blake y Loiselle 2000). Aunque se ha sugerido que ambos métodos
son sesgados (Karr 1981, Dieni y Jones 2002), las comparaciones entre los mismos no se
encuentran bien documentadas.
Numerosos estudios han comparado resultados obtenidos por captura con redes de
niebla y puntos de conteo (Blake y Loiselle 1989, Nur y Geupel 1993, Ralph y Fancy
1995, Gram y Faaborg 1997, Whitman et al. 1997, Blake y Loiselle 2002, Rappole et al.
1998, Pagen et al. 2000, Pagen et al. 2002, Wang y Finch 2002). Otros estudios han
comparado resultados obtenidos a partir de captura con redes de niebla y conteos de
vocalizaciones (Rappole et al. 1993) y otros, de búsqueda de aves y mapeo de puntos
(Diene y Jones 2002), pero no se encuentran documentadas comparaciones entre los
métodos de captura con redes y conteos de búsqueda de aves.

3
En el primer artículo de este estudio se realiza una caracterización de la avifauna
asociada a cinco sitios diferentes en el área de Tortuguero, de los cuales dos han sido
intervenidos por el ser humano y tres se encuentran inalterados; y se analiza la influencia
de la abundancia de frutos sobre la riqueza de especies y abundancia de las poblaciones de
aves. En el segundo artículo se comparan los datos sobre composición, abundancia
poblacional y riqueza de especies obtenidos a partir de las dos técnicas de muestreo
empleadas.
LITERATURA CITADA
Blake, J. G., y B. A. Loiselle. 1989. Habitat use by neotropical migrants at La Selva
Biological Station y Braulio Carrillo National Park, Costa Rica. Ps. 257-272 in: J. M.
Hagan III y D. W. Johnston (eds.). Ecology y conservation of neotropical migratory
lybirds. Smithsonian Institute Press. Washington DC. USA.
Blake, J. G., y B. A. Loiselle. 1991. Variation in resource abundance affects capture rates
of birds in three lowly habitats in Costa Rica. Auk 108:114-130.
Blake, J. G., y B. A. Loiselle 2000. Diversity of birds along an elevational gradient in the
Cordillera Central, Costa Rica. Auk 117:663-686.
Blake, J. G., y B. A. Loiselle. 2001. Bird assemblages in second-growth y old-growth
forests, Costa Rica: erspectives from mist nets y point counts. Auk 118(2):304-326.
Blake, J. G., y B. A. Loiselle. 2002. Manakins (Pipridae) in second-growth y old-growth
forests: patterns of habitat use, movement, y survival. Auk 119(1):132-148.
Dieni, J. S. y S. L. Jones. 2002. A field test of the area search method for measuring
breeding bird populations. Journal of Field Ornithology 73(3):253-257.
Gram, W. K., y J. Faaborg.1997. The distribution of neotropical migrant birds wintering in
the Cielo Biosphere Reserve Tamaulipas, Mexico. Condor 99:658-670.
Herrera, H. 1998. Características del hábitat y de su uso por dos especies de saltarines
(Pipridae) en un humedal del bosque lluvioso neotropical. Una tesis presentada como
requisito parcial para optar al grado de Magister Scientiae en conservación y manejo
de vida silvestre. Programa Regional en Manejo de Vida Silvestre. Heredia, Costa
Rica.

4
Jiménez, B. 1999. Avifauna asociada a las áreas boscosas de la zona de amortiguamiento
del Refugio Nacional de Vida Silvestre Caño Negro, Los Chiles, Alajuela, Costa
Rica. Tesis presentada como requisito parcial par optar al grado de Magister Scientiae
en Conservación y Manejo de Vida Silvestre. Programa Regional en Manejo de Vida
Silvestre para Mesoamérica y el Caribe. Heredia, Costa Rica.
Karr, J. R. 1971. Wintering Kentucky Warblers (Oporornis formosus) y a warning to
byers. Bird Bying 42:299.
Karr, J. R. 1981. Surveying birds with mist nets. Studies in Avian Biology 6:62-67.
Karr, J. R. 1990. The avifauna of Barro Colorado Isly y the Pipeline Road, Panama. Ps.
183-198 in: A. H. Gentry (ed.). Four Neotropical rainforests. Yale University. Press,
New Haven, C.T.
Karr, J. R., y K. E. Freemark. 1983. Habitat selection y environmental gradients: dynamics
in the “stable” tropics. Ecology 64:1481-1494.
Latta, S. C., y J. Faaborg. 2002. Demographic y population responses of Cape May
Warblers wintering in multiple habitats. Ecology 83(9):2502-2515.
Levey, D. J. 1988. Spatial y temporal variation in Costa Rican fruit y fruit-eating bird
abundance. Ecological Monograph 58(4):251-269.
Loiselle, B. A., y J. G. Blake. 1991. Temporal variation in birds y fruits along an
elevational gradient in Costa Rica. Ecology 72(1):180-193.
Nur, N., y G. R, Geupel. 1993. Evaluating mist-netting, nest-searching, y other methods of
monitoring demographic processes in lybird populations. Ps. 237-244 in: D. M.
Finch. y P. W. Stangel (eds.). Status y management of Neotropical migratory birds.
Gen. Tech. Rep. RM-229. USDA Forest Service. USA.
Pagen, R. W., F. R. Thompson III, y D. E. Burhans. 2000. Breeding y post-breeding
habitat use by forest migrant songbirds in the Missouri ozarks. Condor 102:738-747.
Pagen, R. W., F. R. Thompson, III, y D. E. Burhans. 2002. A comparison of point-count y
mist-net detections of songbirds by habitat y time-of-season. Journal of Field
Ornithology 73(1): 53-59.
Ralph, C. J., G. R. Geupel, P. Pyle, T. Martin, D. F. DeSante y B. Milá. 1993. Hybook of
field methods for monitoring lybirds. Pacific Southwest Research Station. Albany,
California. 41 ps. USA.

5
Ralph, C. J., y S. G. Fancy. 1995. Demography y movements of Apapane y Iiwi in
Hawaii. Condor 97:729-742.
Ralph, C. J., M. Widdowson, B. Widdowson, B. O’Donnell, y R. I. Frey. 2002.
Tortuguero bird monitoring station protocol. The Tortuguero Integrated Bird
Monitoring Program. Redwood Sciences Laboratory. U.S. Forest Service. Arcata,
California. USA.
Rappole, J. H., y G. Powell. 1986. The hooded Warbler. Ps. 827-853. En: R. L. D.
Silvestro (ed.). Audubon Wildlife Report . National Audubon Society, New York.
USA.
Rappole, J. H, W. J. McShea, y J. Vega-Rivera. 1993. Evaluation of two survey methods
in uply avian breeding communities. Journal of Field Ornithology 64:55-70.
Rappole, J. H., K Winker, y G. V. N. Powell. 1998. Migratory bird habitat use in southern
Mexico: mist nets versus point counts. Journal of Field Ornithology 69:635-643.
Remsen, J. V., JR, y D. A. Good. 1996. Misuse of data from mist-net captures to assess
relative abundance in bird populations. The Auk 113(2):381-398.
Slater, P. J. 1994. Factors affecting the efficiency of the area search method of censusing
birds in open forests y woodlys. Emu 94:9-16.
Robinson, W. D., J. D. Brown, y S. K. Robinson. 2000. Forest bird community structure
in central Panama: influence of spatial scale y biogeography. Ecological
Monographs 70: 209-235.
Stiles, F. G. 1980. Ps. 421-436. in: A. Keast y E. S. Morton (eds.). Migrant birds in the
Neotropics: ecology, behavior, distribution y conservation. Smithsonian Inst. Press.
Washington, D. C. USA.
Stiles, F. G. 1983. Birds. Ps. 502-530. En: D. H. Janzen (ed.). Costa Rican Natural
History. San Jose, Costa Rica.
Stiles, F. G., y A. F. Skutch 1989. Guía de aves de Costa Rica. Instituto Nacional de
Biodiversidad. Heredia, Costa Rica.
Terborgh, J., S. K. Robinson, T. A. Parker III, C. A. Munn, y N. Pierpont. 1990. Structure
y organization of an Amazonian forest bird community. Ecological Monographs
60:213-238.
Whitman, A. A., J. M. Hagan III, y N. V. L. Brokaw. 1997. A comparison of two bird

6
survey techniques used in a subtropical forest. Condor 99:955-965.
Wunderle, J. M., Jr., y R. B. Waide 1993. Distribution of overwintering Nearctic migrants
in the Bahamas y Greater Antilles. Condor 95:904-933.

7
UNIVERSIDAD NACIONAL
SISTEMA DE ESTUDIOS DE POSGRADO
PROGRAMA REGIONAL EN MANEJO DE VIDA SILVESTRE PARA
MESOAMÉRICA Y EL CARIBE
INFLUENCIAS DE LAS CARACTERÍSTICAS DE HÁBITAT SOBRE LA
COMUNIDAD DE AVES DE UN BOSQUE TROPICAL MUY HÚMEDO EN EL
CARIBE NORTE DE COSTA RICA
María Alejandra Maglianesi
Programa Regional en Manejo de Vida Silvestre para Mesoamérica y el Caribe,
Universidad Nacional. Apartado 1350-3000, Heredia, Costa Rica.
Teléfono: 237-7039. Correo electrónico: [email protected]
Noviembre del 2004

8
INFLUENCIAS DE LAS CARACTERÍSTICAS DE HÁBITAT SOBRE LA
COMUNIDAD DE AVES DE UN BOSQUE TROPICAL MUY HÚMEDO EN EL
CARIBE NORTE DE COSTA RICA
MARÍA ALEJANDRA MAGLIANESI
Programa Regional en Manejo de Vida Silvestre para Mesoamérica y el Caribe,
Universidad Nacional. Apartado 1350-3000, Heredia, Costa Rica
María Alejandra Maglianesi
Programa Regional en Manejo de Vida Silvestre para Mesoamérica y el Caribe.
Universidad Nacional. Apartado 1350-3000, Heredia, Costa Rica. Teléfono: 237-7039.
Correo electrónico: [email protected]

9
RESUMEN
La fase de campo de este estudio fue realizado entre agosto y diciembre del 2002 en los
alrededores de Tortuguero, en la costa norte del Caribe de Costa Rica, provincia de Limón.
Se realizó una caracterización de la comunidad de aves en cinco sitios diferentes, de los
cuales dos corresponden a bosques intervenidos y tres a bosques primarios. Medidas de
abundancia, composición y riqueza de especies residentes y migratorias fueron tomadas y
comparadas entre sitios con la finalidad de sugerir estrategias apropiadas de manejo. En
este estudio se determinó cómo las aves están distribuidas entre los hábitats disponibles y
se analizaron patrones de recaptura de especies migratorias durante la estación de otoño
correspondiente al Hemisferio norte. La abundancia relativa y riqueza de especies fueron
determinadas a partir de datos obtenidos por medio de la técnica de captura con redes de
niebla y conteos de búsqueda de aves. Se estimó el número de frutos presentes para las
aves en cuatro de los sitios, en forma simultánea con las sesiones de captura y los conteos
de búsqueda de aves. Se registró un total de 128 especies de aves, de las cuales 93 especies
fueron capturadas y 100 especies observadas durante los conteos. La mitad de todas las
especies fueron detectadas tanto en hábitats intervenidos como en no intervenidos,
incluyendo un 43.4% de especies residentes y un 69.4% de especies migratorias. A su vez,
las especies migratorias fueron más comunes en los hábitats intervenidos (27.8%) que en
los no intervenidos (2.8%). En los sitios con mayor abundancia de frutos se registró un
mayor numero de especies de aves frugívoras. Las especies residentes tienden a evitar más
los hábitats intervenidos que las especies migratorias. El mantenimiento de hábitats
primarios debería ser una prioridad debido al mayor tiempo que implica su formación y
porque muchas especies de aves, especialmente residentes son dependientes de este tipo de
hábitats. Sin embargo, los esfuerzos de conservación dirigidos a maximizar la riqueza de
especies de aves deberían incluir la protección de las áreas intervenidas evitando que sean
convertidas en áreas agrícolas o urbanas. De esta forma se favorecería el mantenimiento de
un mosaico de hábitats que permite un ensamblaje diverso de especies residentes y
migratorias en Tortuguero.
PALABRAS CLAVES: aves residentes, aves migratorias, riqueza de especies, tasa de
captura, uso de hábitat, bosques alterados, bosques primarios, frutos

10
ABSTRACT
Field work was conducted since late July until late December 2002 near Tortuguero,
northern Caribbean coast in Costa Rica, Limon Province. Bird community was examined
in five sites, two disturbed and three undisturbed wet lowland forest. Relative abundance,
composition and richness of resident and migrant landbirds among different sites were
measured and compared in order to suggest appropriate management strategies. In this
study, was determined how resident and migrant birds are distributed among the available
habitats and it was analyzed recapture pattern of migrant species during the fall season
from temperate North America. Species richness and relative abundance were determined
by combining mist-netting and area search count data. Number of fruits available to birds
was measured in four sites during the same period that birds were netted and observed. I
recorded a total of 128 species of birds by both methods from the five sites, including 93
species captured and 100 species observed. In terms of total species richness (observed
and/or netted), migrants represented 28.1% of all species. Half of all species occurred in
both disturbed and undisturbed habitats including 43.4% of the resident species and 69.4%
of the migrant species. Migrant species were more common in disturbed habitats (27.8%)
than in undisturbed forest habitats (2.8%). Differences in capture rates among species and
sites reflected variation in patterns of habitat use. Results in this study suggest that more
resident species avoid disturbed habitats. In contrast, migrant species more frequently
occur at disturbed habitat. Maintenance of undisturbed areas should be the highest priority,
because the vegetation of mature forest require relatively extensive periods of time to
reach their former states after cutting, and because many migratory and specially, resident,
bird species are dependent of this type of habitat. However, conservation efforts directed
toward maximizing bird species richness should include disturbed areas their protection
from agricultural or urban areas. There should be a maintenance of mosaics of habitats that
support diverse assemblages of resident and migrant species in Tortuguero.
KEY WORDS: resident birds, neotropical migrants birds, bird richness, capture rate,
habitat use, disturbed forest, undisturbed forest, fruit availability

11
En el área de Tortuguero, localizada en la costa Caribe de Costa Rica, se han registrado
un total de 374 especies de aves, lo cual representa el 44% de las especies de aves del país
(Widdowson y Widdowson 2000). Gran parte de las especies de aves migratorias
neotropicales pasan por la costa del Caribe norte de Costa Rica durante la estación de
otoño del Hemisferio norte. Para estas especies de aves, los eventos que ocurren en sus
áreas de invierno podrían ser especialmente importantes porque muchas de ellas pasan más
de la mitad de su ciclo anual en dichas áreas (Keast 1980, Petit 2000). Las altas tasas de
deforestación en las áreas tropicales y subtropicales que constituyen el hábitat para las
especies migratorias, indican que los sucesos que ocurren en el Neotrópico podrían llegar a
tener un gran impacto sobre las aves migratorias en el futuro (Rappole y Powell 1986).
Sin embargo, los hábitats con vegetación alterada que han ido reemplazando los
bosques primarios de tierras bajas en muchas partes del Neotrópico, pueden proveer un
importante hábitat para las especies de aves (Karr 1971, Blake y Loiselle 1991, Jiménez
1999), aunque la disponibilidad de tales hábitats podría ser muy variable. Por otro lado, la
cuantificación de la riqueza de especies en una comunidad de aves ha ido ganando mayor
importancia en los estudios de impacto ambiental, planificaciones para la conservación e
investigaciones ecológicas (Herzog et al. 2002).
Las fluctuaciones en la abundancia poblacional de las especies de aves pueden ser el
resultado de una gran variedad de factores tales como el grado de depredación, la
competencia y variaciones temporales y espaciales en la abundancia de alimento (Karr y
Freemark 1983, Stiles 1980). Hutto (1980) sostiene que los hábitats alterados y de
crecimiento secundario poseen un mayor grado de depredación de nidos, por lo cual las
especies residentes tienden a ocupar más los hábitats primarios a diferencia de las especies
migratorias. Por otro lado, la importancia de la abundancia de alimento como una hipótesis
para explicar la variación en la abundancia poblacional de las aves se ve reflejada en un
número creciente de estudios realizados en la última década (Loiselle y Blake 1991, Levey
1988). El rol que desempeña el alimento para las aves ha llegado a quedar claro en algunos
casos, pero resulta controversial en otros casos.
Si las poblaciones de aves se ven afectadas por la abundancia de frutos, deberían
presentarse cambios concurrentes en los patrones de abundancia poblacional (Loiselle y
Blake 1991). Las evidencias que demuestren que las aves siguen el recurso frutos puede

12
llegar a indicar que el alimento podría actuar como un factor limitante en los bosques que
habitan (Wiens 1989).
Otros estudios han comparado la avifauna entre hábitats en diferentes estados de
conservación (Tramer 1974, Hutto 1989, Borges y Stouffer 1999, Jiménez 1999). Jiménez
(1999) evaluó el efecto de la degradación de bosques sobre la estructura de la comunidad
de aves comparando cuatro estados sucesionales en el norte de Costa Rica. Sin embargo, el
área considerada en este estudio se encuentra influenciada por dos zonas avifaunísticas:
Bosque Tropical Siempre Verde y Bosque Tropical Deciduo, las cuales muestran una alta
riqueza de especies de aves. Por lo tanto, la composición de la comunidad de aves es
diferente a la del Caribe norte y también los problemas que afectan la conservación de las
especies. Además, si estudios como el de Jiménez (1999) y otros similares pudieran ser
combinados con medidas simultáneas de factores ambientales, tales como abundancia de
frutos, podrían explicar algunas diferencias en la abundancia relativa de las especies de
aves.
En el área de Tortuguero, Herrera (1998) estudió la asociación del hábitat con el
número de capturas de dos especies de saltarines (Pipridae) y el comportamiento
reproductivo de estas especies. Desde 1994 las poblaciones de aves están siendo
monitoreadas en diferentes sitios de Tortuguero (Ralph et al. 2002). Sin embargo, la
influencia de los frutos sobre la distribución y abundancia de las poblaciones de aves no ha
sido investigada aún.
El presente estudio provee una caracterización de la avifauna asociada a diferentes
sitios en un Bosque Tropical muy Húmedo y analiza el alimento como una base potencial
para explicar las variaciones espaciales en las poblaciones de aves, con el propósito de
lograr una mejor comprensión de los patrones de distribución y abundancia de la avifauna
y sugerir recomendaciones para un manejo apropiado.
Los objetivos de este estudio fueron: 1) medir y comparar la abundancia relativa, la
composición y riqueza de especies de aves residentes y migratorias entre cinco sitios
diferentes (dos sitios alterados y tres sitios no alterados); 2) determinar el grado de
solapamiento en la composición de especies entre diferentes comunidades; 3) determinar
los patrones de movimiento y recaptura de especies residentes y migratorias; 4) analizar la

13
influencia de la abundancia de frutos sobre la distribución y riqueza de especies entre
cuatro sitios (dos sitios alterados y dos sitios no alterados).
MÉTODOS
ÁREA DE ESTUDIO. La fase de campo fue realizada entre agosto y diciembre del 2002
en la zona de Tortuguero, ubicada en la costa del Caribe norte en la Provincia de Limón,
Costa Rica (Fig. 1). La principal zona de vida en las tierras bajas del Caribe es el Bosque
Tropical muy Húmedo (Holdridge 1969), el cual se caracteriza por poseer una vegetación
siempreverde que refleja una breve estación seca (Stiles 1983). El Bosque Tropical muy
Húmedo presenta un dosel alto y una variedad de estratos de vegetación. Unas pocas
especies son caducifolias, pero esto no cambia el aspecto general siempreverde del bosque
(Janzen 1983). El área de estudio se ubica dentro del Área de Conservación Tortuguero
(ACTo), el cual incluye uno de los humedales más grandes del país (Oro 1996). La
temperatura media anual es de 29.9ºC y la precipitación media anual de 6,000 mm hace
esta área una de las más húmedas del país (Coen 1983, Oro 1996, Stiles y Skutch 1989).
La estación seca se extiende desde finales de diciembre hasta marzo. Durante los cinco
meses de este estudio se registró una precipitación de 2,138 mm y una temperatura
promedio de 26.9ºC, con una mínima de 24.5ºC y una máxima de 30.7ºC.
Tortuguero estaba ampliamente rodeado por bosque continuo hasta 1960, que
posteriormente fue transformado a un mosaico de diferentes estados sucesionales causado
por actividades antrópicas. La expansión de las pasturas para ganado y los cultivos de
banano provocó la pérdida de bosques en las tierras bajas del Atlántico (Boucher et al.
1983). En la actualidad la principal actividad económica en Tortuguero es el turismo,
donde las aves son uno de los grupos más importantes de vida silvestre que atraen a los
turistas, debido a su gran diversidad y a su relativamente fácil detección (Herrera 1998).
La infraestructura hotelera ha aumentado rápidamente en la zona de Tortuguero durante la
última década y ha contribuido a la fragmentación del bosque.
Sitios de estudio.—Los sitios estudiados se establecieron en dos tipos de bosques:
intervenido y no intervenido. Dos de los cinco sitios incluyen vegetación alterada por
influencia humana: Corporación para la Conservación del Caribe (CCC) y el Aeropuerto
Barra de Tortuguero (AERO). Los tres sitios restantes se encuentran en bosque primario:

14
Parque Nacional Tortuguero (PARQ), Tortuga Lodge (TORT) y la Estación Biológica
Caño Palma (CAÑO) (Fig. 1). En los sitios no intervenidos el dosel puede alcanzar 40-50
m de altura y muchos árboles pueden tener hasta 100-200 cm de diámetro a la altura del
pecho (DAP). Los árboles del subdosel miden alrededor de 25-35 m de altura y el
sotobosque mide entre 10 y 25 m de altura. La capa de arbustos en el sotobosque es de 1-
2.5 m de altura y allí se encuentran abundantes palmas. En los sitios intervenidos, donde
fueron talados selectivamente los árboles de mayor DAP hace aproximadamente 20 años,
los árboles del dosel son más bajos con una altura de 20-25 m aproximadamente y la capa
de arbustos del sotobosque es particularmente densa.
AERO se encuentra localizado adyacente al extremo norte de la pista de aterrizaje de
Tortuguero y TORT se encuentra en una propiedad privada del lado opuesto del río, ambos
sitios están a 10 min en lancha desde CCC. Mientras que CAÑO se encuentra localizado
adyacente a la estación canadiense de investigación, a unos 20-25 min en lancha desde
CCC, PARQ corresponde a la esquina noreste del Parque Nacional Tortuguero y CCC se
encuentra contiguo al pueblo de Tortuguero (Fig. 1).
MUESTREO DE AVES. Este estudio siguió el protocolo de campo usado por el Programa
Integrado de Monitoreo de Tortuguero (Ralph et al. 2002). Este programa se inició en
1994 y consiste en un esfuerzo cooperativo que involucra diferentes organizaciones,
colaboradores y voluntarios. Se incluyeron dos técnicas de muestreo de aves: captura con
redes de niebla y conteos de búsqueda de aves siguiendo la descripción hecha por Ralph et
al. (1993). Se tomaron registros de precipitación y temperatura en CCC. Las listas de aves
que se incluyen en este estudio siguen el orden establecido en la Unión Americana de
Ornitología (AOU 1998).
Captura con redes de niebla.—Las aves fueron capturadas con redes estándar (12 m x 3
m, 36 mm de malla), (Avinet, Box 1103, Dryden NY, 13053, USA. Telefax: 607-844-
3277. E-mail: [email protected]). Se colocaron 10-15 redes de niebla separadas por 40
m aproximadamente unas de otras, en el perímetro de un área circular, cubriendo un área
de 6 ha en cada unos de los cinco sitios. Las redes fueron operadas desde el nivel del suelo
1-3 veces por semana y revisadas a intervalos de 40 min como máximo (más
frecuentemente en condiciones de viento y calor extremos). Las redes no fueron operadas

15
en condiciones de lluvia, viento o calor extremos y si las redes ya estaban abiertas en tales
condiciones fueron inmediatamente cerradas. Las redes fueron abiertas desde las 5:30-5:40
(Central Standard Time) y cerradas a las 11:30, tratando en lo posible de mantenerlas
abiertas durante 6 hs por día. Las redes fueron cerradas en el mismo orden en que fueron
abiertas (Ralph et al. 1993).
Todas las aves capturadas, incluyendo aquellas recapturadas, fueron identificadas hasta
el nivel de especie a excepción de dos especies pertenecientes al género Empidonax. Los
individuos capturados fueron marcados con anillos de aluminio numerados (Bird Banding
Laboratory, Fish and Wildlife Service, Laurel, Maryland 20708, USA.). Luego de ser
procesadas, las aves fueron liberadas en el sitio de captura. Los individuos muy jóvenes
fueron liberados cerca de la misma red en que fueron capturados.
El número de aves capturadas en 100 hs red (1 red abierta durante 1 h = 1 h red) fue
usado como un índice de abundancia relativa para cada especie y se hará referencia a esta
variable como la “tasa de captura”. Los datos de captura y recaptura fueron utilizados para
analizar el uso de hábitat y los movimientos dentro y entre los diferentes sitios tanto para
aves residentes como para migratorias.
Conteos de búsqueda de aves.—Los conteos de búsqueda de aves consisten en
desplazarse dentro del área (6 ha) de capturas con redes de niebla registrando todas las
especies observadas y/o escuchadas durante un periodo de 20 min (Ralph et al. 1993). Se
realizaron dos conteos durante cada mañana en que las redes de niebla fueron operadas. El
primer conteo se llevó a cabo una vez finalizada la apertura de las redes (alrededor de las
6:00 hs) y el segundo conteo se realizó a las 9:30 hs aproximadamente. No se realizaron
conteos bajo condiciones climáticas desfavorables. Las especies de aves fueron
identificadas usando binoluares (10 x 50) y una guía local de aves (Stiles y Skutch 1989).
El número medio de individuos por especie observados por conteo fue utilizado como un
índice de abundancia relativa y definido como “tasa de conteo”.
MUESTREO DE FRUTOS. Se tomaron muestras de frutos en cuatro sitios (CCC, AERO,
PARQ y TORT) durante el mismo periodo de captura de aves, dentro de transectos de 2 m
de ancho x 12,5 m de largo, paralelos y a una distancia de 1 m de las redes de niebla
(Blake y Loiselle 1991). Cada 15 días se contaron todos los frutos maduros por especie de

16
planta por debajo de los 10 m de altura. Las especies de plantas fueron identificadas in situ
en la mayoría de los casos o se identificaron en el Hervario Juvenal Rodriguez de la
Universidad Nacional. Para el caso de CAÑO no fueron tomadas muestras de frutos
debido su difícil acceso. Se consideró como unidad de muestreo a cada par de trasectos,
considerando como variable el número total de frutos en 50 m2.
ANÁLISIS ESTADÍSTICO. El paquete estadístico STATGRAPHICS version 3.1
(Statistical Graphic Corp.) fue usado para los diferentes análisis que se presentan en este
estudio. Todas las variables fueron sometidas a pruebas de normalidad (Wilk-Shapiro test),
homogeneidad de varianza (Bartlett’s test) e independencia, antes de realizar las pruebas
estadísticas (Sokal y Rohlf 1979). Cuando las variables no cumplieron con los supuestos
de las pruebas paramétricas se realizaron transformaciones hasta alcanzar los supuestos.
Curvas de acumulación de especies.—Debido a que el esfuerzo de muestreo varió entre
sitios, curvas de acumulación de especies para las capturas y los conteos fueron
construidas usando el modelo de Clench derivado de la ecuación de Michaelis-Menten
(Soberón y Llorente 1993). Este modelo exponencial y asintótico, en el cual la
probabilidad de agregar nuevas especies a un inventario eventualmente se aproxima a cero,
fue usado para extrapolar un estimado de la riqueza de especies para la asíntota de la curva
y para determinar el esfuerzo de muestreo requerido para detectar una proporción del
número de especies que predice el modelo. El modelo de Clench utiliza la ecuación S (t) =
a* t / (1+ b* t) donde S (t) = número total de especies que predice el modelo, t = esfuerzo
de muestreo, a = tasa de incremento del número de especies al comienzo del muestreo y b
= tasa de incremento del número de especies. La asíntota está dada por el cociente a/b y el
tiempo tq requerido para registrar una proporción del total de especies q que predice el
modelo fue calculado a partir de q/(b*(1-q)). Las muestras fueron aleatorizadas 20 veces y
los datos fueron ajustados al modelo usando una regresión no lineal en el Programa
InfoStat Versión 1.1.
Riqueza de especies.—El número de especies de aves capturadas en cada sitio fue
definido como la “riqueza de especies”, la cual fue comparada entre sitios utilizando
Análisis de Varianza (ANDEVA) de 2 factores (Sokal y Rohlf 1979) (sitio y mes). Este
análisis se hizo independientemente para especies residentes y migratorias; y se utilizó

17
como unidad de muestreo las sesiones de captura (6 hs de esfuerzo-persona por cada
sesión).
Abundancia relativa.—Las tasas de captura fueron comparadas entre sitios usando
ANDEVA de dos factores (sitio y mes). Las recapturas no fueron incluidas en el cálculo de
la tasa de captura para alcanzar la independencia de los datos. Las tasas de conteo fueron
también comparadas usando ANDEVA de 2 factores (especie y sitio).
Categorías de abundancia.—Para analizar la rareza de especies en las comunidades de
aves, las especies fueron agrupadas en diferentes categorías de abundancia basadas en sus
tasas de captura y conteo. Estas categorías tuvieron un rango de 0.001 a 3.000 (valor
mínimo y máximo de las tasas), para los datos provenientes tanto de las capturas como de
los conteos.
Similitud entre comunidades de aves.—El grado de solapamiento en la composición de
especies entre pares de sitios es comúnmente medida mediante el uso de coeficientes de
similitud, y entre éstos el índice de Jaccard es ampliamente utilizado (Magurran 1988)..
Una matriz con coeficientes de Jaccard fue construida a partir de la fórmula: Sj = a / a + b
+ c, donde S = coeficiente de similitud de Jaccard, a = número de especies en el sitio A y
sitio B (especies comunes), b = número de especies presentes en el sitio B pero no en el
sitio A, y c = número de especies en presentes en el sitio A pero no en el sitio B (Krebs
1989). Un análisis de clúster fue realizado siguiendo el procedimiento del vecino más
cercano en el programa Statgraphics (Statistical Graphic Corp.) y representado
gráficamente a través de un dendrograma.
Patrones de movimiento y recaptura.—Los patrones de recaptura fueron analizados
usando porcentajes de recaptura por especie con respecto al total de individuos capturados,
con la finalidad de determinar el tiempo de permanencia de las especies migratorias en las
áreas de invierno. Los porcentajes de recaptura fueron comparados entre sitios a través de
ANDEVA de 1 factor.
Abundancia de frutos y relación con la riqueza de especies de aves.—Aunque los frutos
que se encuentran en el sotobosque varían en tamaño y contenido de nutrientes, se usó el
número total de frutos maduros en cada sitio como un estimado de la abundancia de frutos
para las aves. El número de frutos maduros fue comparado entre los 4 sitios usando
ANDEVA de dos factores (sitio y muestra). El análisis fue restringido a aquellas especies

18
de plantas que se conocen son visitadas por las aves capturadas (basado en observaciones
personales de comportamiento de alimentación de las aves y en reportes de la gente local).
Para relacionar la riqueza de especies de aves con la abundancia de frutos se consideraron
solamente a las especies de aves frugívoras, por lo cual se consideró la información sobre
dietas obtenida en investigaciones previas (Bierregaard 1990, Karr et al. 1990). El criterio
empleado para asignar a las especies dentro del gremio de las aves frugívoras fue que éstas
consuman únicamente frutos o la mayor parte de su dieta consista en frutos y también que
al menos ocasionalmente consuman frutos en sus dietas. El número de especies de aves
frugívoras fue comparado entre sitios y meses con un ANDEVA de dos factores.
Uso de hábitat.— Para analizar el uso de los sitios por las especies residentes y
migratorias, se calculó el porcentaje de especies presentes en sólo uno de los cinco sitios
con respecto al total de especies capturadas y/o obsevadas en conteos, y se realizaron
comparaciones utilizando ANDEVA. Además, los cinco sitios fueron agrupados dentro de
categorías de hábitats (intervenido y no intervenido) y se comparó el porcentaje de
especies presentes en cada categoría con ANDEVA. Las especies fueron agrupadas
dependiendo de su presencia sólo en hábitats intervenidos, sólo en hábitats no intervenidos
o en ambas categorías.
RESULTADOS
Un total de 128 especies de aves distribuidas en 32 familias fueron registradas por los
dos métodos combinados en los cinco sitios de muestreo. Fueron capturadas en redes de
niebla 93 especies distribuidas en 21 familias (1996 individuos capturados durante 7571
horas-red). Durante 186 conteos de búsqueda de aves (79.9 hs) se observó 100 especies
distribuidas en 31 familias (Apéndice A). El número total de especies detectadas en este
estudio representó el 34% de la avifauna registradas en la lista de aves para Tortuguero
(Widdowson y Widdowson 2000). Del total de especies registradas en este estudio el
28.1% fueron migratorias (Cuadro 1). Ara ambigua, una especie amenazada, fue
observada únicamente durante los conteos de búsqueda de aves y en los sitios no
intervenidos.
Especies más comunes.—La especie residente más comúnmente capturada fue Manacus
candei, la cual comprendió el 19% del total de individuos capturados. Tres de las cinco

19
especies más comunes en las redes de niebla fueron colibríes: Amazilia tzacatl, Glaucis
aenea, y Phaethornis superciliosus, Sporophila americana fue también muy común. Estas
cinco especies acumularon el 75% del total de individuos residentes capturados. Las cinco
especies migratorias más comúnmente capturadas fueron (en orden): Catharus ustulatus,
Empidonax sp (Empidonax alnorum y E. trailli ), Prothonotaria citrea, Catharus
fuscescens, y Catharus minimus. Estas especies comprendieron el 65% del total de
individuos migrantes capturados.
Las especies residentes con mayor tasa de conteo fueron (en orden): Pionus senilis,
Coragyps atratus, Psaracolius montezuma, Todirostrum cinereum, Myozetetes similis, y
Hylophilus decurtatus. Las especies migratorias con mayor tasa de conteo fueron (en
orden): Catharus ustulatus, Vireo olivaceus, Contopus virens, Dendroica petechia y
Dendroica pensylvanica.
Curvas de acumulación de especies.—Los parámetros del análisis de regresión del
modelo de Clench para los cinco sitios y para ambos métodos de muestreo se encuentran
en la Cuadro 3. Los parámetros calculados para especies capturadas (R2, a, b y la asíntota)
para CCC, PARQ y CAÑO fueron mayores que para AERO y TORT, mostrando que en
esos tres sitios hubo un incremento rápido en el porcentaje de especies capturadas en las
primeras muestras. Las curvas de acumulación de especies para cada uno de los sitios
indican que nuevas especies continuaron siendo registradas a lo largo del muestreo (Fig.
2).
La asíntota que predice el modelo de Clench fue mayor que los valores derivados de los
datos de captura para los cinco sitios. Tanto los parámetros como el esfuerzo de muestreo
se ajustaron únicamente para CAÑO, donde una proporción elevada del total de especies
de aves que predice el modelo fueron capturadas (77%) en comparación con los otros
sitios. En los cinco sitios se requiere un mayor esfuerzo de muestreo para alcanzar el 90%
de la asíntota, siendo CCC y TORT los sitios donde es necesario aumentar más el número
total de hs-red (Cuadro 3).
Durante los conteos de búsqueda de aves, PARQ aumentó rápidamente el porcentaje de
especies observadas en las primeras muestras y tuvo los mayores parámetros del modelo
de Clench. El porcentaje de especies representadas en los conteos con respecto al total de
especies que predice el modelo para CCC fue más bajo que para los datos provenientes de

20
capturas. Este porcentaje fue más alto en CCC para los conteos (67%), siendo en otros
sitios menor a 45% (Fig. 3). Las curvas obtenidas a partir de los conteos poseen mayores
pendientes con respecto a aquellas obtenidas a partir de capturas, lo cual indica que por
medio del método de conteo de aves se registran las especies más rápidamente con por
medio de capturas.
Riqueza de especies.—Se capturaron 1284 individuos pertenecientes a 62 especies
residentes (66.7%) y 712 individuos pertenecientes a 31 especies migratorias (33.3%). El
número de especies capturadas en diferentes sitios fue como mínimo 33 en TORT y como
máximo 62 en CCC (Cuadro 1). De las 100 especies observadas en conteos de búsqueda
de aves, 72 fueron residentes y 28 migratorias (28%). El número de especies observadas
fue como mínimo 41 en TORT y como máximo 65 en CCC (Cuadro 2).
La riqueza media de especies residentes capturadas varió entre sitios (F=2.68; gl=4,16;
P=0.037) siendo más alta en PARQ (x=1.80) durante la mayoría de los meses, seguido por
AERO (x=1.69). Sin embargo, la riqueza media mayor para especies residentes fue
durante agosto en CAÑO aunque las diferencias no fueron significativas entre meses (Fig.
4). La riqueza media de especies migratorias capturadas fue mayor en AERO (x=1.00) y
CCC (x=0.99) para la mayoría de los meses. Aunque en agosto y diciembre esta variable
fue mayor en PARQ (F=2.45; gl=4,16; P=0.055). Se alcanzó un pico en la riqueza media
de especies migratorias durante octubre para todos los sitios. (F=11.55; gl=4,16;
P<0.0001) (Fig. 5).
Abundancia relativa.—De las 93 especies capturadas, 33 (37%) estuvieron
representadas por sólo una o dos capturas y en conjunto reunieron el 2.5% del total de
individuos capturados. La tasa media de captura de residentes varió significativamente
entre sitios (F=7.12; gl=4,16; P=0.0001) siendo mayor para AERO (x=3.15) que para los
otros cuatro sitios durante todos los meses. Agosto fue el mes que tuvo las tasas de captura
de residentes más altas en la mayoría de los sitios (F=5.58; gl=4,16; P=0.0005) (Fig. 6)
(x=2.98).
La tasa media de captura de migratorias fue mayor en AERO (x=1.88) y CCC (x=1.77)
que en los otros sitios, aunque las diferencias no fueron significativas (F=2.0; gl=4,16;
P=0.106). La tasa media de captura de migratorias alcanzó un pico durante el mes de
octubre en todos los sitios (F=25.64; gl=4,16; P<0.001) (Fig. 7).

21
La tasa media de conteo fue mayor en CCC (x=0.50) y AERO (x=0.43) que en los otros
tres sitios, aunque las diferencias no fueron significativas (F=1.95; gl=4,137; P=0.10). Las
especies con mayor tasa media de conteo fueron Amazona autumnalis (x=2.44), Pionus
senilis (x=2.17), Hirudindo rustica (x=1.82) y Habia fuscicauda (x=1.32), (F=3.60;
gl=94,137; P<0.0001) (Fig.8).
Las categorías de abundancia son presentadas en la Fig. 9 para las especies capturadas y
en la Fig.10 para las especies observadas en conteos. Del total de especies capturadas hubo
57 especies residentes (92%) y 25 especies migratorias (81%) en la categoría de
abundancia más baja de abundancia relativa (tasas de captura < 0.500). Estas especies
representaron el 88% del total de especies capturadas. De las 100 especies observadas en
conteos, hubo 34 especies residentes (47%) y 16 especies migratorias (57%) en la
categoría mas baja de abundancia relativa, (<0.600) las cuales representaron la mitad de
todas las especies observadas.
Índices de similitud.—Los coeficientes de similitud entre sitios para las especies
capturadas son mostrados en la Cuadro 4. El dendrograma indica mayor similitud en la
composición de las comunidades de aves que se encuentran en los tres sitios no
intervenidos y entre las comunidades de aves de los dos sitios intervenidos (Fig. 11).
Patrones de movimiento y recaptura.—Hubo un total de 19 especies residentes y cuatro
especies migratorias con individuos que fueron recapturados. Del total de individuos
recapturados hubo 200 (15.6% del total de capturas) pertenecientes a especies residentes y
22 (3.2% del total de capturas) a especies migratorias en los cinco sitios estudiados
(Cuadro 5).
El porcentaje medio de individuos recapturados por especie varió entre sitios (F=3.23;
gl=4,41; P=0.02), siendo mayor en AERO (x=7.9) (Fig. 12). Aunque las diferencias entre
especies no resultaron significativas, Henicorhina leucosticta (50%, n=6 captures),
Cercomacra tyrannina (50%, n=2 captures), Thamnophilus atrinucha (40%, n=20
capturas), Manacus candei (29.1%, n=254 capturas) y Hylophilus decurtatus (28.6%, n=21
capturas) tuvieron mayores porcentajes de recaptura (Cuadro 6). Los individuos de esta
última especie fueron recapturados siempre dentro de CCC y los de M. candei fueron
recapturados dentro de CCC, AERO y TORT. Mientras que los individuos de T. atrinucha
fueron recapturados dentro de los tres sitios no intervenidos (PARQ, TORT y CAÑO).

22
La mayoría de los individuos residentes fueron recapturados dentro del mismo sitio
donde previamente fueron capturados inicialmente (84.7%). Del total de recapturas, hubo
7 especies (37%) y 31 individuos (15.5%) que fueron recapturados en diferentes sitios
(Cuadro 7). No hubo recapturas de especies migratorias en sitios diferentes al de la captura
inicial. Oporornis formosus tuvo un periodo máximo transcurrido entre recapturas en el
área de estudio de 101 días, seguido por Protonotaria citrea (94 días) y Seiurus
noveboracensis (39 días).
Plantas y especies de aves frugívoras.—Un total de 16 especies de plantas con frutos
maduros pertenecientes a 8 familias fueron registradas (Cuadro 8). La familia más común
fue Rubiaceae, la cual comprendió el 56% de todas las especies de plantas. En CCC se
registró el mayor número de especies de plantas con frutos. La abundancia de frutos varió
espacialmente entre sitios (F=2.54; gl=3,612; P=0.05) (Fig. 13) y temporalmente (F=7.44;
gl=6,612; P< 0.0001) (Fig. 14). El número medio de frutos/50 m2 fue mayor en los sitios
intervenidos (CCC y AERO) que en los sitios no intervenidos (TORT y CAÑO) y durante
las dos primeras muestras en el mes de setiembre.
El 53% de las especies de aves capturadas fueron incluidas en el gremio de los
frugívoros. El número medio de especies frugívoras varió entre sitios (F=6.88; gl=4,12;
P<0.0004) y a través del tiempo (F=13.27; gl=4,12; P<0.0001), siendo mayor en CCC en
la mayoría de los sitios. El mes con mayor número medio de especies de aves frugívoras
fue octubre en todos los sitios (Fig. 15).
Uso de hábitat.—De las 128 especies registradas por ambos métodos, un total de 44
especies (34.4%) estuvieron restringidas a sólo uno de los cinco sitios y la mayoría de
estas especies fueron residentes (88.6%). De las especies restringidas a un único sitio se
encuentran incluidas el 42% del total de especies residentes registradas en este estudio y el
14% del total de especies migratorias, y CCC tuvo el más alto porcentaje de especies
restringidas a un único sitio (48%) (Fig. 16).
La mitad de todas las especies capturadas y/o observadas a lo largo de este estudio se
encontraron en ambos tipos de hábitats: intervenidos y no intervenidos, incluyendo el
43.4% del total de especies residentes y el 69.4% del total de especies migratorias
(F=2.64; gl=2,2; P=0.27). Las especies migratorias fueron más comunes en los sitios

23
intervenidos (27.8% del total es especies migratorias) que en los sitios no intervenidos
(2.8% del total de especies migratorias) (Fig. 17).
DISCUSIÓN
Curvas de acumulación de especies.—El poder contar con una base teórica que permita
comprender la relación que existe entre el esfuerzo de muestreo y el número de especies
acumuladas sería de gran utilidad debido a que ello permitiría establecer comparaciones
cuantitativas más rigurosas entre deferentes comunidades de aves. Para elegir entre los
diferentes modelos disponibles se requiere de información sobre el tamaño de área a
muestrear y el grupo de fauna o flora.
Los modelos varían considerablemente en las extrapolaciones que se obtienen a partir
de un mismo grupo de datos (Soberón y Llorente 1993). En mi estudio utilicé el modelo de
Clench porque a pesar de que las aves son un taxón bien conocido, hay muchas especies
raras y el modelo de Clench como así también los modelos logarítmicos pueden ser más
adecuados para este tipo de situación (Soberón y Llorente 1993).
Sin embargo, los parámetros y el esfuerzo de muestreo que predicen el modelo de
Clench indicaron que se necesita un mayor esfuerzo de muestreo. Una posible explicación
para esto es que los datos fueron tomados solamente durante una estación determinada y la
extrapolación sería válida sólo para esta parte del año. Además, las redes de niebla
permiten obtener datos de las especies del sotobosque y de igual manera las
extrapolaciones serían válidas para esta porción de la comunidad de aves y no para su
totalidad.
Patrones de distribución y abundancia.—Las aves migratorias en el presente estudio
comprendieron un componente más importante de la avifauna de invierno que en otros
estudios similares en hábitats neotropicales (Lynch 1989). Sin embargo, las categorías de
abundancia en este estudio indican que la mayoría de las especies residentes y migratorias
tienen una abundancia relativa baja. Estos resultados son consistentes con reportes previos
sugiriendo que los bosques neotropicales poseen típicamente una alta riqueza de especies
pero la mayoría de estas son raras (Terborgh et al. 1990, Thiollay 1999).
En el presente estudio las diferencias en las tasas de captura entre especies y sitios
reflejan las preferencias de hábitat de las especies en el área de Tortuguero. Muchas

24
especies que se encuentran en un único sitio fueron capturadas en CCC y el sitio con
mayor tasas de captura tanto para especies residentes como migratorias fue AERO, en
ambos casos se trata de sitios intervenidos. Además, existe una tendencia a ser mayores las
tasas medias de conteo en CCC y AERO.
Los factores causantes de la distribución de las aves no están claros aún, pero se ha
sugerido ampliamente que entre estos se incluyen interacciones interespecíficas (Hutto
1980, Emlen 1980) y los recursos alimenticios (Hutto 1980). Existe actualmente poca o
ninguna evidencia de que las especies residentes excluyan a las migratorias neotropicales
de los bosques que habitan (Waide 1980). Por lo tanto, el alimento sería un factor más
probable afectando la distribución de las aves. Karr (1976) sugirió que las especies
migratorias están usualmente subordinadas a las especies residentes y que por lo general se
alimentan en áreas periféricas o subóptimas cuando las residentes se encuentran presentes
en un sitio.
La abundancia relativa de las poblaciones de aves en los sitios intervenidos puede haber
estado sesgado debido a la mayor probabilidad de detección de las aves, ya sea por
capturas o conteos, con respecto a los sitios no intervenidos (Stiles 1980). Sin embargo, es
probable que la mayor abundancia en los sitios intervenidos en este estudio no sea un
resultado de simplemente de un sesgo en los muestreos, sino que refleja una diferencia real
en la distribución de las aves.
Influencia de la abundancia de frutos.—Los sitios con mayores tasas de captura y
riqueza de especies de aves residentes y migratorias frugívoras capturadas, son los mismos
que aquellos con mayor abundancia de frutos: CCC y AERO. Esto sugiere una relación
entre la actividad de las aves y la abundancia de frutos, siendo consistente con Herrera
(1998), quien encontró una asociación entre las capturas y el alimento potencial para
saltarines (Pipridae) en Tortuguero. La identificación de los factores que promueven a las
fluctuaciones en los patrones de abundancia de las poblaciones de aves es fundamental
para comprender como estas comunidades están organizadas (Loiselle y Blake 1991).
Cabe considerar que entre un 63% y un 77% de los arbustos y árboles del sotobosque
producen frutos que son dispersados por aves en los bosques siempreverdes de Costa Rica
y las aves frugívoras frecuentemente constituyen una porción importante de la avifauna
tropical (Siltes 1985).

25
Por otro lado, el método por el cual la abundancia de frutos es estimada tiene gran
importancia para el análisis de los frutos como un recurso para las aves (Blake et al. 1990).
En este estudio se usó un método de muestreo basado en el área, el cual ha sido utilizado
para estimar la producción de frutos en una variedad de sitios tropicales de tierras bajas.
Mi estudio se enfocó a los arbustos, árboles, lianas y epífitas del sotobosque (<10 m del
suelo) y no se incluyeron estimados de la producción de frutos del dosel, debido a que el
análisis estuvo basado en las aves capturadas en redes de niebla y su actividad ocurre en
los estratos más bajos del bosque.
El presente estudio incluyó la influencia de la abundancia de alimento sobre un gremio
en particular de aves: los frugívoros. Sin embargo, debe quedar claro que los frugívoros no
reflejan las necesidades de otros gremios componentes de la avifauna tropical. Por lo tanto,
si bien los sitios intervenidos tuvieron más abundancia de frutos y mayor riqueza de
especies frugívoras, lo que ocurre con el resto de la comunidad de aves resta por conocer.
Porcentajes de recaptura pueden indicar preferencia por un hábitat.—Los altos
porcentajes de recaptura de algunas especies como Henicorhina leucosticta y Cercomacra
tyrannina pueden ser un reflejo del tamaño de muestra reducido. Sin embargo, para el caso
de otras especies como Thamnophilus atrinucha, Manacus candei y Hylophilus decurtatus
pueden reflejar una alta fidelidad al sitio. Los altos porcentajes de recaptura para
Thamnophilus atrinucha sólo dentro de PARQ, TORT y CAÑO indican una preferencia
por los bosques no intervenidos con respecto a aquellos intervenidos en Tortuguero.
Las especies migratorias en Costa Rica se encuentran principalmente de paso durante su
época no reproductiva, permaneciendo en una misma área por breves periodos. En
contraste, unas pocas especies tales como Oporornis formosus, Protonotaria citrea y
Seiurus noveboracensis aunque son consideradas como de paso en Costa Rica, fueron
recapturadas dentro del mismo sitio con un tiempo considerable entre recapturas. Esto
indica que los individuos de dichas especies hacen un uso más prolongado que lo esperado
de los hábitats en Tortuguero y pueden ser consideradas como especies residentes de
invierno en Costa Rica, coincidiendo con las observaciones hechas por G. Barrantes (com.
per. Escuela de Biología, Universidad de Costa Rica, San Pedro, Costa Rica).
Por otro lado, los patrones de recaptura en este estudio muestran que las especies
residentes se desplazan entre CCC y PARQ a través del pueblo de Tortuguero, lo cual

26
tiene implicancias desde el punto de vista del manejo y la conservación de las especies de
aves.
Especies migratorias más comunes en sitios intervenidos.— En el presente estudio, se
registraron más especies residentes en un único sitio con respecto a las especies
migratorias y estas últimas fueron más comunes en los sitios intervenidos. Estos resultados
coinciden con estudios previos que evidencian un mayor uso de los hábitats intervenidos
por parte de las especies migratorias en sus áreas de invierno en el Neotrópico (Karr 1976,
Hutto 1980, Martin 1985, Waide 1980, Willis 1980, Kricher y Davis 1989, Blake y
Loiselle 1992, Jiménez 1999).
El conocimiento sobre la importancia relativa o el uso que las especies de aves hacen de
los diferentes hábitats y cómo estos usos varían a través del tiempo y el espacio, tiene
implicaciones tanto ecológicas como para la conservación de dichas especies. Por ejemplo,
la pérdida de dispersores de semillas podría llegar a afectar la composición florística de los
bosques (Loiselle y Blake 2001). Mientras que la pérdida de hábitats específicos podría
tener consecuencias para la supervivencia de la aves, particularmente aquellas que
necesitan más de un hábitat durante su ciclo anual como es el caso de las especies de aves
migratorias (Stilesy Clark 1989).
Cabe considerar que muchas especies migratorias usan una variedad de hábitats en
Costa Rica (Stiles 1980). La deforestación reduce el número de especies de aves presente
en un área determinada (Terborgh 1980), pero a su vez algunos hábitats por los que son
reemplazados los bosques primarios pueden ofrecer refugio para ciertas especies de aves
(Mills y Rogers 1992).
Sin embargo, nuestro conocimiento sobre uso que las especies migratorias hacen de los
hábitat en el Neotrópico está lejos de ser completo y en diferentes estudios se han
encontrado resultados opuestos. Por ejemplo, se ha sugerido en numerosas ocasiones que
las especies de aves migratorias se encuentran principalmente en hábitats intervenidos y de
crecimiento secundario mientras se encuentran en sus áreas de invierno (Karr 1976,
Fitzpatrick 1980, Martin 1985, Hutto 1989, Jiménez 1999). En contraposición Lynch
(1989) y Tramer (1974) encontraron pocas diferencias en la abundancia de migratorias
entre una serie de diferentes estados sucesionales. Más aún, algunos autores han enfatizado
que muchas especies migratorias dependen de hábitats primarios y forman una parte

27
integral de los bosques tropicales primarios (Stiles 1980, Terborgh 1980, Rappole y
Morton 1985).
Los resultados obtenidos en el presente estudio sugieren que las especies residentes
evitan los sitios intervenidos, mientras que las especies migratorias fueron más comunes
en estos sitios. Cabe preguntar entonces, ¿por qué las especies migratorias deberían
encontrarse más probablemente restringidas a los hábitats intervenidos?.
Hutto (1980) argumentó que la tasa de depredación de nidos podría llegar a ser muy
elevada en los hábitats intervenidos como para tener una residencia permanente ventajosa.
Por lo tanto, la sub-utilización de dichos hábitats por parte de las especies residentes puede
ser explotada por las especies migratorias durante su época no reproductiva. En contraste,
debido a que los bosques no intervenidos son ventajosos para ser ocupados durante todo el
año por las especies residentes, las migratorias podrían ser competitivamente excluidas de
tales bosques (Slud 1960, Potti 1985).
Las aves migratorias deben balancear las influencias opuestas cuando seleccionan sus
áreas de invierno. Así por ejemplo, el mayor riesgo de depredación que existe en los sitios
intervenidos (Blake y Loiselle 1992) podría ser compensado por la gran abundancia de
alimento que hay en estos hábitats en comparación con los hábitats primarios, según los
resultados obtenidos en el presente estudio y por otros autores (Martin 1985, Levey 1988,
Loiselle y Blake 1990, Blake y Loiselle 1991).
El concepto del no equilibrio en la diversidad de los bosques tropicales (Connel 1978)
se encuentra probablemente reflejado por los patrones de riqueza de especies de aves
migratorias en el área de Tortuguero, apoyando la idea de que niveles moderados de
perturbación podrían beneficiar a un número considerable de especies migratorias
neotropicales que utilizan los bordes de los hábitats y la vegetación en diferentes estados
sucesionales. Esto puede ser explicado por la teoría de las perturbaciones intermedias, la
cual predice que los niveles mayores de diversidad son mantenidos a escalas intermedias
en intensidad y frecuencia de perturbaciones.
Algunos autores están de acuerdo en que ciertas especies migratorias en América
Central no se encuentran amenazadas debido a la destrucción de bosques primarios ya que
la mayoría utilizan los hábitats intervenidos. Esto puede ser cierto para la mayoría de las
especies, pero algunas tales como Oporornis formosus pueden ser seriamente afectadas por

28
la pérdida reciente y acelerada de bosques de tierras bajas, debido a que estas especies
necesitan grandes áreas de bosques para sobrevivir (Karr 1976, Lynch 1989). En mi
estudio los sitios no intervenidos fueron utilizados por Ara ambigua. Se ha reportado que
las poblaciones de esta especie ha declinado en otros sitios de Costa Rica en las últimas
décadas debido a la pérdida de hábitat principalmente, quedando actualmente pequeñas
subpoblaciones en el país (Blake et al. 1990).
Desde un punto de vista de la conservación, existen aspectos críticos que necesitan de
mayor investigación, incluyendo los factores que influyen la selección de hábitat (Blake y
Loiselle 1992). Por ejemplo, las especies actualmente “prefieren” los hábitats intervenidos
o solo los usan porque todos los sitios preferidos están ya ocupados (Rappole et al. 1989).
Sobre las decisiones de manejo.—Existe una amplia creencia, especialmente entre los
sectores gubernamentales, de que los bosques de crecimiento secundario y hábitats
intervenidos no tienen ninguna importancia para la conservación de las especies de aves.
Una evidencia de esto, es que durante el gobierno de Calderón en Costa Rica en el periodo
1990-94, una compañía trasnacional expandió las plantaciones de banano a través de las
tierras bajas en la zona de Río Frío (norte de Costa Rica). Esta situación provocó la
deforestación de una extensión de 400 has en la región Atlántica del país (Espinoza 1992).
El Ministro de Agricultura apoyó esta expansión porque para él los bosques jóvenes eran
unos “tacotales” con ninguna importancia ecológica desde un punto de vista de la
conservación de la vida silvestre.
Sin embargo, los resultados en este estudio y otros similares (Hutto 1989, Jiménez
1999) sugieren lo opuesto, debido a que los sitios intervenidos poseen una mayor
abundancia poblacional de especies residentes y migratorias de aves y una mayor riqueza
de especies de aves migratorias. Los esfuerzos de conservación dirigidos a maximizar la
riqueza de especies de aves deberían incluir la protección de las áreas intervenidas
evitando que sean convertidas en áreas agrícolas o urbanas. De esta forma se favorecería el
mantenimiento de un mosaico de hábitats que permite un ensamblaje diverso de especies
residentes y migratorias (Kricher y Davis 1989). Dicho mosaico debería tener un acceso
directo entre áreas intervenidas y bosques primarios, debido a que la mitad del total de
especies registradas en este estudio hacen uso de ambos tipos de hábitats. Podría ser

29
esencial para gran parte de la comunidad de aves de los sitios intervenidos, contar con
áreas adyacentes de parches de bosque maduro.
Conclusiones y recomendaciones.—Los hábitats en el área de Tortuguero apoyan un
ensamblaje diverso de especies residentes y migratorias, incluyendo una especie
amenazada. La prioridad más alta debería ser preservar los sitios primarios o áreas no
intervenidas, debido a que la vegetación de estos bosques requiere de considerables
periodos de tiempo para alcanzar la madurez, y porque ciertas especies de aves migratorias
y en particular las especies residentes dependen de este tipo de bosques.
Debido a que una aproximación basada en estudios autoecológicos podrían no ser
generalizables para la comunidad de aves, yo sugiero que se realicen estudios sobre uso de
hábitat de la comunidad entera o gremios de aves. Estos estudios debería realizarse en
combinación con medidas simultáneas de factores ambientales, tales como frutos,
artrópodos, estructura y composición de la vegetación, con la finalidad de comprender
mejor los factores que podrían influenciar la presencia o ausencia de las especies de aves
en una determinada comunidad local.
Asimismo, se requiere de más estudios sobre las fluctuaciones en las poblaciones de
aves y en la abundancia de alimento. Un análisis más directo de las respuestas que las
especies hacen a las variaciones de alimento es fundamental para resolver el rol
controversial de este recurso como un factor que influye en la organización de las
comunidades de aves (Wiens 1984, Martin 1986). Yo sugiero que se lleven a cabo estudios
fenológicos registrando la actividad de floración y fructificación de la vegetación a lo
largo de varios años, simultáneamente con el monitoreo de las poblaciones de aves, con el
objetivo de comprender mejor las relaciones entre el recurso alimenticio y las aves.
Las especies residentes y migratorias de aves se ven afectadas por los factores del
hábitat de diferentes maneras. Como conservacionistas, debemos apreciar el hecho de que
las especies son independientes unas de otras y únicas en sus necesidades (Hutto 1989).
Por razones prácticas, debemos enfocar los esfuerzos de monitoreo y conservación hacia
grupos o especies de aves “indicadoras”.
Para la conservación de la avifauna en el área de Tortuguero, los programas de
educación e incentivos deberían ser implementados para promover la protección por parte

30
del gobierno y propietarios de áreas de bosque primario como así también de áreas con
vegetación alterada o de crecimiento secundario.
AGRADECIMIENTOS
En primer lugar quisiera agradecer a los miembros de mi Comité de Tesis: Michael
McCoy, Gilbert Barrantes, C. John Ralph, y Jaime Rau, quienes me ofrecieron
constantemente sus comentarios y recomendaciones a lo largo de este estudio. Realmente
me siento afortunada de haber trabajado bajo la guía de estos renombrados especialistas.
Me encuentro muy agradecida con el Programa Regional en Manejo de Vida Silvestre para
Mesoamérica y el Caribe (PRMVS) por permitirme llevar a cabo un grado de maestría, a
partir del cual elaboré el presente artículo de tesis como un requisito para optar al título de
Master en Conservación y Manejo de Vida Silvestre. También agradezco al Laboratorio de
TeleSig del PRMVS por permitirme el uso de equipo y programas de cómputo
especializados, en particular le agradezco a Henry Chávez por su contribución en la
elaboración del mapa sobre el área de estudio.
Agradezco también a la Corporación para la Conservación del Caribe por permitirme el
acceso a sus instalaciones para llevar a cabo la fase de campo en su propiedad, a Pablo
Sánchez del Herbario Juvenal Valerio Rodríguez (Universidad Nacional) quien me brindó
su ayuda en la identificación de las especies de plantas y a la organización Idea Wild por
proveer equipo para este estudio.
Doy mi mas sincero agradecimiento a los voluntarios del Programa Integrado de
Monitoreo de Aves de Tortuguero, quienes contribuyeron en la toma de datos de campo:
Dai Shizuka, Devin Taylor, Gabriel David, Lucie Parker, Dan van den Broek, Ana Clarke
y Ernest Clarke. Ellos fueron excelentes compañeros y entusiastas asistentes de campo.
Doy un especial agradecimiento a Robert Frey, quien es un invaluable compañero en el
campo y me ofreció sus consejos, compartiendo su gran experiencia y pasión por las aves.
Este estudio fue posible gracias al Programa Integrado de Monitoreo de Aves de
Tortuguero (Redwood Sciences Laboratory, U.S. Forest Service), el cual facilitó el equipo
y los voluntarios para el trabajo de campo y a los observatorios de aves Reyes Point y
Klamath, quienes facilitaron los fondos para los aspectos logísticos de esta investigación.

31
LITERATURA CITADA
AOU. 1998. Check-list of North American Birds. 7th Edition. American Ornithologists’
Union. American Ornithologist Union (ed.), Washington, D.C.
Blake, J. G., B. A. Loiselle, T. C. Moermond, D. J. Levey y J. S. Denslow. 1990.
Quantifying abundance of fruits for birds in tropical habitats. Pp. 73-79 in Studies in
avian biology, vol. 13 (M. L. Morrison, C. J. Ralph, J. Verner y J. R. Jehl, Jr., Eds.).
Allen Press, Inc., Lawrence, Kansas.
Blake, J. G. y B. A. Loiselle. 1991. Variation in resource abundance affects capture rates
of birds in three lowland habitats in Costa Rica. Auk 108:114-130.
Blake, J. G. y B. A. Loiselle. 1992. Habitat use by Neotropical migrants at La Selva
Biological Station and Braulio Carrillo National Park, Costa Rica. Pp. 257-272 in
Ecology and conservation of neotropical migrant landbirds (J. M. Hagan III y D. W.
Johnston, Eds.). Smithsonian Inst. Press, Washington, D.C.
Bierregaard, Jr, R. O. 1990. Species composition and trophic organization of the
understory bird community in a Central Amazonian Terra Firme Forest. Pp. 217-236 in
Four neotropical rainforests (A. H. Gentry, Ed.). Yale University Press. New Haven.
Borges, S. H. y P. C. Stouffer. 1999. Bird communities in two types of anthropogenic
successional vegetation in Central Amazonia. Condor 101:529-536.
Boucher, D. H., M Hansen, S. Risch y J. H. Vandermeer. 1983. Pp. 66-117 in Costa Rican
natural history (D. H. Janzen, Ed.). The University of Chicago Press, Chicago and
London.
Coen, E. 1983. Climate. Pp. 35-46 in Costa Rican natural history (D. H. Janzen, Ed.). The
University of Chicago Press, Chicago and London.
Connel, J. H. 1978. Diversity in tropical rain forests and coral reefs. Science 199:1302-
1310.
Emlen, J. T. 1980. Interaction of migrant and resident land birds in Florida and Bahama
pinelands. Pp. 133-144 in Migrant birds in the neotropics: ecology, behavior,
distribution, and conservation (A. Keast y E. S. Morton, Eds.). Smithsonian Inst. Press.
Wasghington, D. C.
Espinoza, R. 1992. Calderón niega pugna entre ministros. La Nación. 15 de Julio. San
José, Costa Rica.

32
Fitzpatrick, J. W. 1980. Wintering of North American tyrant flycatcher in the Neotropics.
Pp. 67-78 in Migrant birds in the neotropics: ecology, behavior, distribution, and
conservation (A. Keast y E. S. Morton, Eds.). Smithsonian Inst. Press. Wasghington,
D. C.
Herrera, H. 1998. Características del hábitat y de su uso por dos especies de saltarines
(Pipridae) en un humedal del bosque lluvioso neotropical. Tesis de Maestría,
Universidad Nacional, Heredia, Costa Rica.
Herzog, S. K., M. Kessler y T. M. Cahill. 2002. Estimating species richness of tropical
bird communities from rapid assessment data. Auk 119:749-769.
Holdridge, L. R 1969. Life zone ecology. Rev. Ed. San José, Costa Rica. Tropical Science
Center. San José, Costa Rica.
Hutto, R. L. 1980. Winter habitat distribution of migratory land birds in western Mexico
with special reference to small, folige-gleaning insectivores. Pp. 181-204 in Migrant
birds in the neotropics: ecology, behavior, distribution, and conservation (A. Keast y E.
S. Morton, Eds.). Smithsonian Inst. Press. Wasghington, D. C.
Hutto, R. L. 1989. The effect of habitat alteration on migratory land birds in a west
Mexican tropical deciduous forest: a conservation perspective. Conserv. Biol. 3:138-
148.
Janzen, D. H. 1983. Costa Rican natural history. The University of Chicago Press, Chicago
and London.
Jiménez, B. 1999. Avifauna asociada a las áreas boscosas de la zona de amortiguamiento
del Refugio Nacional de Vida Silvestre Caño Negro, Los Chiles, Alajuela, Costa Rica.
Tesis de Maestría, Universidad Nacional, Heredia, Costa Rica.
Karr, J. R. 1971. Wintering Kentucky Warblers (Oporornis formosus) and a warning to
banders. Bird Banding 42:299.
Karr, J. R. 1976. On the relative abundance of migrants from the north temperate zone in
tropical habitats. Wilson Bull. 88:433-458.
Karr, J. R. y K. E. Freemark. 1983. Habitat selection and environmental gradients:
dynamics in the “stable” tropics. Ecology 64:1481-1494.

33
Karr, J. R., S. Robinson, J. G. Blake y R. O. Bierregaard, Jr. 1990. Birds of four
Neotropical forests. Pp. 237-272 in Four Neotropical rainforests (A. H. Gentry, Ed.).
Yale University. Press, New Haven, C.T.
Keast, A. 1980. Migratory parulidae: what can species co occurrence in the north reveal
about ecological plasticity and wintering patterns? Pp. 457-476 in Migrant birds in the
neotropics: ecology, behavior, distribution, and conservation (A. Keast y E. S. Morton,
Eds.). Smithsonian Inst. Press. Washington, D. C.
Krebs, C. J. 1989. Ecological Methodology. University of British Columbia. Harper
Collins Publishers.
Kricher, J. C y W. E. Davis, Jr. 1989. Patterns of avian species richness in disturbed and
undisturbed habitats in Belize. Pp. 240-246 in Ecology and conservation of
Neotropical migrant landbirds (J. M. Hagan, III y D. W. Johnston, Eds.). Smithsonian.
Inst. Press, Washington, D.C.
Levey, D. J. 1988. Spatial and temporal variation in Costa Rican fruit and fruit-eating bird
abundance. Ecological Monograph 58:251-269.
Loiselle, B. A. y J. G. Blake. 1990. Diets of understory fruit-eating birds in Costa Rica:
seasonality and resource abundance. Pp. 91-103 in Studies in avian biology, vol. 13
(M. L. Morrison, C. J. Ralph, J. Verner y J. R. Jehl, Jr., Eds.). Allen Press, Inc.,
Lawrence, Kansas.
Loiselle, B. A. y J. G. Blake. 1991. Temporal variation in birds and fruits along an
elevational gradient in Costa Rica. Ecology 72:180-193.
Loiselle, B. A. y J. G. Blake. 2001. Potential consequences of extinction of frugivorous
birds for shrubs of a tropical wet forest. Pp. 397-405 in Seed dispersal and frugivory:
ecology, evolution and conservation (D. J. Lovey, W. R. Silva y M. Galetti, Eds.).
CABI Publishing, Wallingford, Oxfordshire, UK.
Lynch, J. F. 1989. Distribution of overwintering nearctic migrants in the Yucatan
Peninsula. I: general patterns of occurrence. Condor 91:515-544.
Magurran, A. E. 1988. Ecological diversity and its measurement. Princeton University
Press. Princeton, New Jersey.
Martin, T. E. 1985. Selection of second-growth woodlands by frugivorous migrating birds
in Panama: an effect of fruit size and plant density? J. Trop. Ecol. 1:157-170.

34
Martin, T. E. 1986. Competition in breeding birds: on the importance of considering
processes at the level of the individuals. Curr. Ornithol. 4:181-210.
Mills, E. D. y D. T. Rogers, Jr. 1992. Ratios of neotropical migrant and neotropical
resident birds in winter in a citrus plantation in Central Belize. J. Field Ornithol.
63:109-116.
Oro, G. M. 1996. Ficha Técnica RAMSAR. Manuscrito. Proyecto de consolidación del
Área de conservación y desarrollo sostenible llanuras de Tortuguero (PACTO). San
José, Costa Rica.
Petit, D. R. 2000. Habitat use by landbirds along nearctic-neotropical migration routes:
implications for conservation of stopover habitats. Pp. 15-33 in Studies in avian
biology vol. 20
Potti, J. 1985. Sobre la distribución de los migrantes transaharianos en la Península
Ibérica. Ardeola 32:57-68.
Ralph, C. J., G. R. Geupel, P. Pyle, T. Martin, D. F. DeSante y B. Milá. 1993. Handbook
of field methods for monitoring landbirds. Pacific Southwest Research Station.
Albany, California. 41 p. USA.
Ralph, C. J., M. Widdowson, B. Widdowson, B. O’Donnell y R. I. Frey. 2002. Tortuguero
bird monitoring station protocol. The Tortuguero Integrated Bird Monitoring Program.
Redwood Sciences Laboratory. U.S. Forest Service. Arcata, California.
Rappole, J. H. y E. S. Morton 1985. Effects of habitat alteration on a tropical avian forest
community. Ornithological Monographs 36:1013-1021.
Rappole, J. H. y G. Powell. 1986. The hooded Warbler. Pp. 827-853 in Audubon Wildlife
Report (R. L. D. Silvestro, Ed.). National Audubon Society, New York.
Rappole, J. H., M. A. Ramos y K. Winker. 1989. Wintering Wood Thrush movements and
mortality in southern Veracruz. Auk 106:402-410.
Slud, P. 1960. The birds of finca “La Selva” Costa Rica: a tropical wet forest locality.
Bulletin of the American Museum of Natural History 121:49-148.
Soberón, J. M. y J. B. Llorente. 1993. The use of species accumulation functions for the
prediction of species richness. Conserv. Biol. 7:480-488.

35
Sokal, R. R. y F. J. Rohlf. 1979. Biometría. Primera edición en español. Principios y
métodos estadísticos en la investigación biológica. H. Blume Ediciones. Madrid,
España.
Stiles, F. G. 1980. Pp. 421-436 in Migrant birds in the neotropics: ecology, behavior,
distribution, and conservation (A. Keast y E. S. Morton, Eds.). Smithsonian Inst. Press.
Wasghington, D. C.
Stiles, F. G. 1983. Birds. Pp. 502-530 in Costa Rican natural history (D. H. Janzen, Ed.).
The University of Chicago Press, Chicago and London.
Stiles, F. G. 1985. Conservation of forest birds in Costa Rica: problems and perspectives.
Pp. 141-168 en Conservation of tropical forest birds (A. W. Diamond y T. E. Lovejoy,
Eds.). International Council Bird Preservation. Technical Publication Nº 4. Cambridge,
England.
Stiles, F. G. y A. F. Skutch 1989. Guía de aves de Costa Rica. Instituto Nacional de
Biodiversidad. Heredia, Costa Rica.
Stiles, F. G. y D. A. Clark 1989. Conservation of tropical rain forest birds: a case study
from Costa Rica. American Birds 43:420-428.
Terborgh, J. W. 1980. The conservation status of neotropical migrants: present and future.
Pp. 21-30 in Migrant birds in the neotropics: ecology, behavior, distribution, and
conservation (A. Keast y E. S. Morton, Eds.). Smithsonian Inst. Press. Washington,
D.C.
Terborgh, J. W., S. K. Robinson, T. A. Parker III, C. A. Munn y N. Pierpont. 1990.
Structure and organization of an Amazonian forest bird community. Ecological
Monographs 60:213-238.
Thiollay, J. M. 1999. Bird community structure of a primary rain forest in Guinea: changes
with scale and disturbance. Pp. 2580-2590 in Proceeding 22 International Congress (N.
J. Adams y R. H. Slowtow, Eds.). Birdlife. Durban, South Africa.
Tramer, E. 1974. Proportions of wintering North American birds in disturbed and
undisturbed dry tropical habitats. Condor 76:460-464.
Waide, R. B. 1980. Resource partitioning between migrant and resident birds: the use of
irregular resources. Pp. 337-352 in Migrant birds in the neotropics: ecology, behavior,
distribution, and conservation (A. Keast y E. S. Morton, Eds.). Smithsonian Inst. Press.

36
Washington, D.C.
Widdowson, W. P. y M. J. Widdowson. 2000. Checklist to the birds of Tortuguero. The
Tortuguero Integrated Bird Monitoring Program. Redwood Sciences Laboratory. U.S.
Forest Service. Arcata, California.
Wiens, J. 1984. Resource systems, populations and communities. Pp. 397-436 in A new
Ecology (P. W. Price, W. S. Gaud y C. N. Slobodchikoff, Eds.). New York: Wiley.
Wiens, J. 1989. The ecology of bird communities, vols. 1 y 2. Cambridge University Press,
Cambridge, England.
Willis, E. O. 1980. Ecological roles of migratory and resident birds on Barro Colorado
Island, Panama. Pp. 205-225 in Migrant birds in the neotropics: ecology, behavior,
distribution, and conservation (A. Keast y E. S. Morton, Eds.). Smithsonian Inst. Press.
Washington, D.C.

CUADRO 1. Esfuerzo de muestreo de captura, número de individuos y especies capturadas en cinco sitios en el Caribe norte de Costa Rica.
Número de Capturasa Número de Especies Capturadas
Sitiob
h-rc Residentes Migrantes Total % Migrantes Residentes Migrantes Total % Migrantes
CCC 3725.5 562 410 972 42.2 35 27 62 43.5
AERO 855.8 262 127 389 32.5 20 19 39 48.7
PARQ 904.0 152 45 197 22.8 24 14 38 36.8
TORT 1065.0 149 82 231 35.5 19 14 33 42.4
CAÑO 1021.0 159 48 207 23.2 28 12 41 29.3
Todos los Sitios 7571.0 1284 712 1996 35.7 62 31 93 33.3
a Capturas incluye todas las capturas de aves individuales, incluyendo recapturas. b CCC = Corporación para la Conservación del Caribe; PARQ = Parque Nacional Tortuguero; AERO: Aeropuerto Barra de Tortuguero; TORT = Tortuga Lodge; CAÑO = Estación Biológica Caño Palma.
c h-r = horas-red.
37
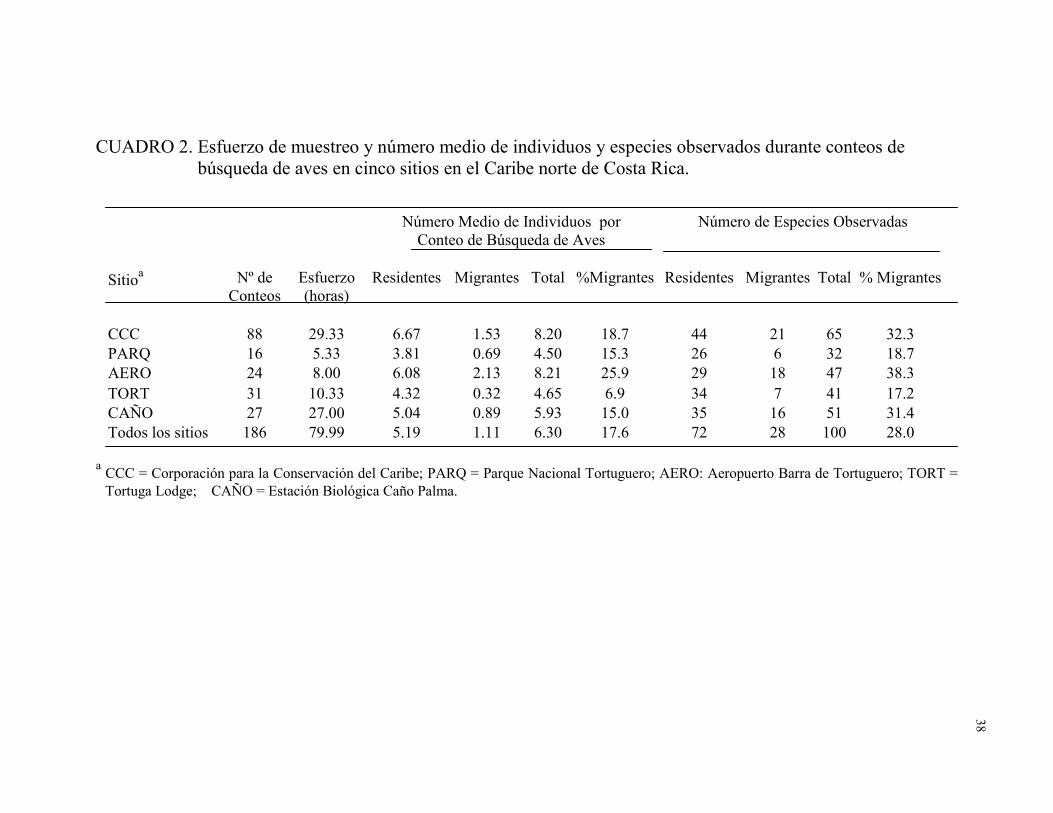
CUADRO 2. Esfuerzo de muestreo y número medio de individuos y especies observados durante conteos de búsqueda de aves en cinco sitios en el Caribe norte de Costa Rica.
Número Medio de Individuos por Conteo de Búsqueda de Aves
Número de Especies Observadas
Sitioa Nº de Conteos
Esfuerzo (horas)
Residentes Migrantes Total %Migrantes Residentes Migrantes Total % Migrantes
CCC 88 29.33 6.67 1.53 8.20 18.7 44 21 65 32.3 PARQ 16 5.33 3.81 0.69 4.50 15.3 26 6 32 18.7 AERO 24 8.00 6.08 2.13 8.21 25.9 29 18 47 38.3 TORT 31 10.33 4.32 0.32 4.65 6.9 34 7 41 17.2 CAÑO 27 27.00 5.04 0.89 5.93 15.0 35 16 51 31.4 Todos los sitios 186 79.99 5.19 1.11 6.30 17.6 72 28 100 28.0
a CCC = Corporación para la Conservación del Caribe; PARQ = Parque Nacional Tortuguero; AERO: Aeropuerto Barra de Tortuguero; TORT = Tortuga Lodge; CAÑO = Estación Biológica Caño Palma.
38

CUADRO 3. Estadísticos de regresión del modelo de Clench para cinco sitios en Caribe norte de Costa Rica. Captura con Redes de Niebla Conteos de Búsqueda de Áves
Sitioa
R2
a
b
Asíntota (a/b)
Porcentaje de Especies Capturadas
Esfuerzo para Capturar el 90% de las Especies (hr)b
R2
a
b
Asíntota (a/b)
Porcentaje de Especies Observadas
Esfuerzo para Contar el 90% de las Especies (Conteos)
CCC 0.96 0.04 3.1-4 129 48 29,032 0.96 1.94 0.02 97 67 450 PARQ 0.96 0.1 1.6-3 62 61 5,625 0.96 3.84 0.05 77 42 180 AERO 0.96 0.08 7.4-4 108 36 12,272 0.94 1.8 -1-2* 180 26 900 TORT 0.94 0.04 3.3-4 121 27 27,272 0.94 1.62 0.01 162 25 900 CAÑO 0.98 0.15 2.8-3 53 77 3,124 0.94 1.99 1.2-3* 1658 3 7,500 a CCC = Corporación para la Conservación del Caribe; PARQ = Parque Nacional Tortuguero; AERO: Aeropuerto Barra de Tortuguero; TORT = Tortuga Lodge; CAÑO = Estación Biológica Caño Palma.
b hr = horas-red. * Estadísticamente no significativo (P>0.05).
39
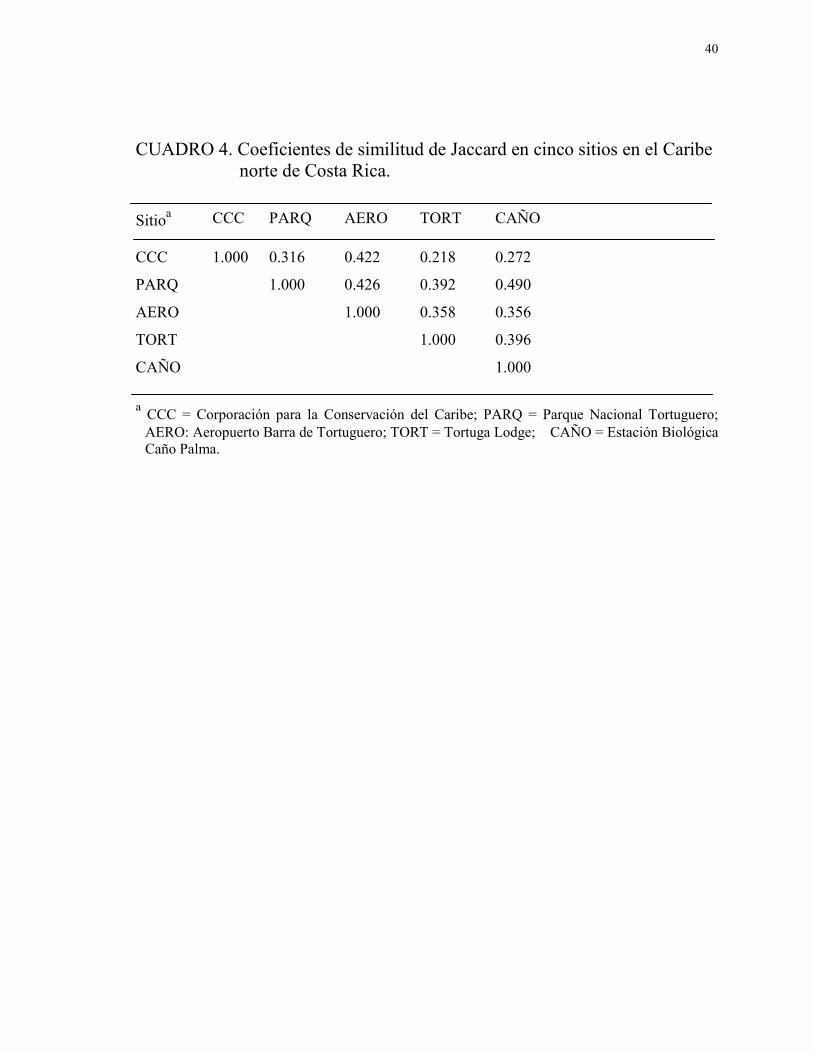
CUADRO 4. Coeficientes de similitud de Jaccard en cinco sitios en el Caribe
norte de Costa Rica.
Sitioa CCC PARQ AERO TORT CAÑO
CCC 1.000 0.316 0.422 0.218 0.272
PARQ 1.000 0.426 0.392 0.490
AERO 1.000 0.358 0.356
TORT 1.000 0.396
CAÑO 1.000
a CCC = Corporación para la Conservación del Caribe; PARQ = Parque Nacional Tortuguero; AERO: Aeropuerto Barra de Tortuguero; TORT = Tortuga Lodge; CAÑO = Estación Biológica Caño Palma.
40

CUADRO 5. Número y porcentajes de recapturas de aves en cinco sitios en el
Caribe norte de Costa Rica.
Residentes
Migrantes
Sitioa Captures Recapturas % Captures Recapturas %
CCC 562 91 16.19 410 10 2.44 PARQ 152 20 13.16 45 0 0 AERO 262 48 18.32 127 9 7.09 TORT 149 30 20.13 82 0 0 CAÑO 159 11 6.92 48 3 6.25
Total 1284 200 15.58 712 22 3.09 a CCC = Corporación para la Conservación del Caribe; PARQ = Parque Nacional Tortuguero; AERO: Aeropuerto Barra de Tortuguero; TORT = Tortuga Lodge; CAÑO = Estación Biológica Caño Palma.
41

CUADRO 6. Porcentaje de recapturas de especies de aves residentes y migratorias en cinco sitios en el Caribe norte de Costa Rica.
Especies Captures Porcentaje de Recaptura
Manacus candei 254 29.13 Amazilia tzcatl 230 10.43 Glaucis aenea 183 13.66 Sporophila americana 157 21.66 Phaethornis superciliosus 104 10.58 Protonotaria citrea 93 15.05 Seiurus noveboracensis 47 6.38 Contopus virens 31 3.23 Pipra mentalis 29 10.34 Anthracothorax prevostii 26 3.85 Hylophilus decurtatus 21 28.57 Turdus grayi 21 9.52 Thamnophilus atrinucha 20 40.00 Oporornis formosus 17 23.53 Chloroceryle aenea 15 13.33 Pitangus sulphuratus 12 25.00 Lepidocolaptes souleyetii 10 10.00 Habia fuscicauda 8 25.00 Henicorhina leucosticta 6 50.00 Dendrocolaptes certhia 6 16.67 Gymnopithys leucaspis 5 20.00 Trogon massena 4 25.00 Cercomacra tyrannina 2 50.00
42

CUADRO 7. Recapturas de especies de aves residentes en sitios diferentes al
de la captura inicial.
Especies Sitio de Captura Sitio de Recaptura
Manacus candei AERO CCC TORT CCC PARQ CCC
Sporophila americana CCC PARQ CCC AERO
Amazilia tzacatl CCC PARQ CCC AERO
Glaucis aenea CCC AERO PARQ CCC, AERO PARQ AERO
Lepidocolaptes souleyetii AERO CCC
Trogon massena TORT AERO
Dendrocolaptes certhia AERO PARQ
43

CUADRO 8. Especies de plantas registradas durante conteos de frutos en
cuatro sitios en el Caribe norte de Costa Rica. Taxon Sitioa
Costaceae Costus laevis
CCC, PARQ, AERO
Heliconidae Heliconia sp.
AERO, TORT
Melastomataceae Conostegia xalapensis
CCC, AERO
Myristicaceae Myristica fragans
TORT
Nyctaginaceae Neea amplifolia
PARQ, AERO
Polygonaceae Coccoloba uvifera
CCC
Rubiaceae Alibertia edulis
Alibertia garapatica
Chiococca alba
Palicourea guianensis
Posoqueria latifolia
Psycotria chagrensis
Psychotria glomerulata
Psycotria limonensis
Rondelethia sp.
TORT CCC CCC CCC, PARQ, AERO TORT PARQ CCC TORT PARQ
Solanaceae Witheringia sp.
TORT
a CCC = Corporación para la Conservación del Caribe; PARQ = Parque Nacional Tortuguero; AERO: Aeropuerto Barra de Tortuguero; TORT = Tortuga Lodge; CAÑO = Estación Biológica Caño Palma.
44


LISTA DE TÍTULOS DE FIGURAS
Figura
1. Área de estudio.
2. Curva de acumulación de especies para especies de aves capturadas basada en el
modelo de Clench.
3. Curva de acumulación de especies para especies de aves observadas basada en el
modelo de Clench.
4. Número medio de especies de aves residentes capturadas en cinco sitios (intervalos de
confianza son + HSD intervalos Tukey).
5. Número medio de especies de aves migratorias en cinco sitios (intervalos de confianza
son + HSD intervalos Tukey).
6. Tasa media de captura de aves residentes en cinco sitios de estudio (intervalos de
confianza son + HSD intervalos Tukey).
7. Tasa media de captura de aves migratorias en cinco sitios de estudio (intervalos de
confianza son + HSD intervalos Tukey).
8. Tasa media de conteo de aves residentes y migratorias en cinco sitios de estudio
(intervalos de confianza are + HSD intervalos Tukey).
9. Categorías de abundancia basadas en tasas de captura.
10. Categorías de abundancia basadas en tasas de conteo.
11. Dendrograma de la riqueza de especies en cinco sitios basado en el vecino más
cercano, distancia cuadrado Eucladiana.
12. Porcentajes de especies de aves residentes y migratorias recapturadas en cinco sitios.
13. Número medio de frutos maduros en 50 m2 en cuatro sitios en el Caribe norte de Costa
Rica.
14. Número medio de frutos maduros a lo largo de cuatro meses.
15. Número medio de especies de aves frugívoras en cuatro sitios en los alrededores de
Tortuguero.
16. Porcentaje de especies totales registradas en un único sitio en el Caribe norte de Costa
Rica.
17. Porcentaje de especies totales en hábitats intervenido y no intervenido en el Caribe
norte de Costa Rica.
45

Fig. 1; 46
Cerro Tortuguero Boca de las lagunas de Tortuguero
Tortuguero

Fig. 2; 47
0
10
20
30
40
50
60
70
1 5 9 13 17 21 25 29 33 37 41 45
Esfuerzo de Muestreo (Horas-Red)
Número Acumulado
de Especies Capturadas
CCC
PARQ
AERO
TORT
CAÑO
50 350 650 950 1250 1550 1850 2150 2450 2750 3050 3350

0
5
10
15
20
25
30
35
40
45
50
55
60
1 5 9 13 17 21 25 29 33 37 41 45 49 53 57 61 65 69 73 77 81 85
Esfuerzo de Muestreo (Número de Conteos)
Número Acumulado de
Especies Observadas
CCC
PARQ
AERO
TORT
CAÑO
Fig. 3; 48
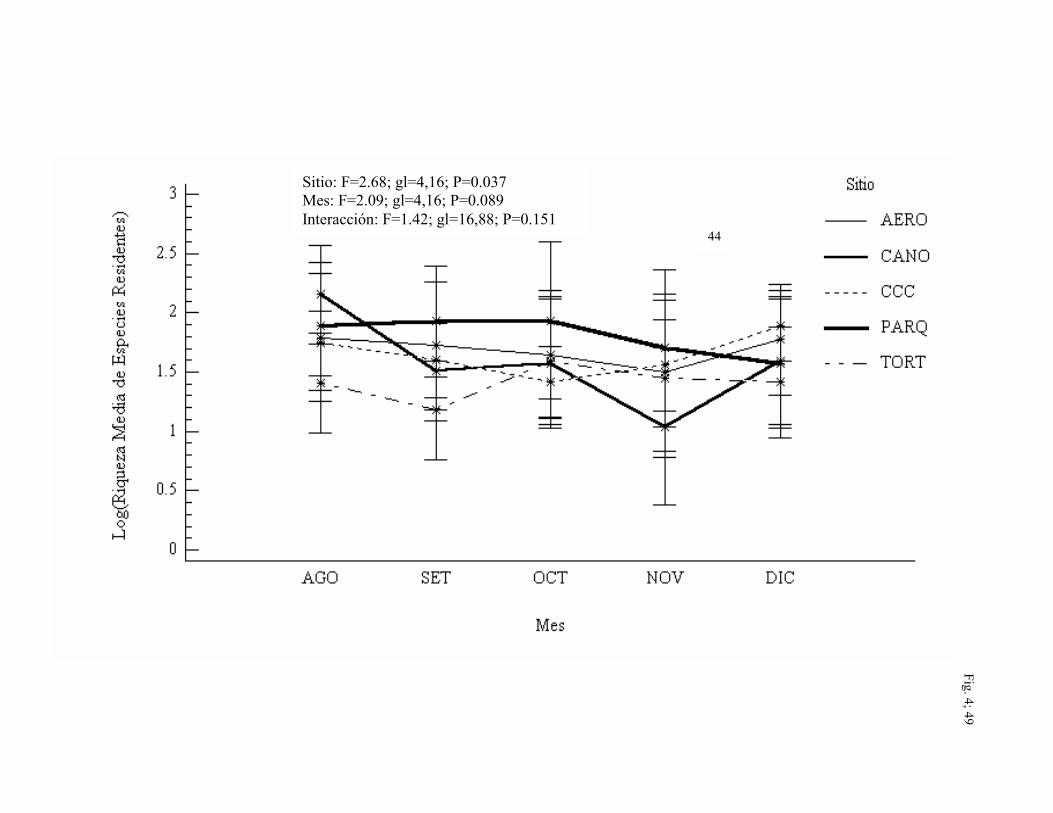
Fig. 4; 49
Sitio: F=2.68; gl=4,16; P=0.037 Mes: F=2.09; gl=4,16; P=0.089 Interacción: F=1.42; gl=16,88; P=0.151
44

Fig. 5; 50
Sitio: F=2.45; gl=4,16; P=0.055 Mes: F=11.55; gl=4,16; P<0.0001 Interacción: F=1.59; gl=16,88; P=0.096

Fig. 6; 51
Sitio: F=7.12; gl=4,16; P= 0.0001 Mes: F=5.58; gl=4,16; P=0.0005 Interacción: F=1.77; gl=16,63; P=0.047

Fig. 7; 52
Sitio: F=2.00; gl=4,16; P= 0.106 Mes: F=20.53; gl=4,16; P=0.0001 Interacción: F=2.45; gl=16,63; P=0.006
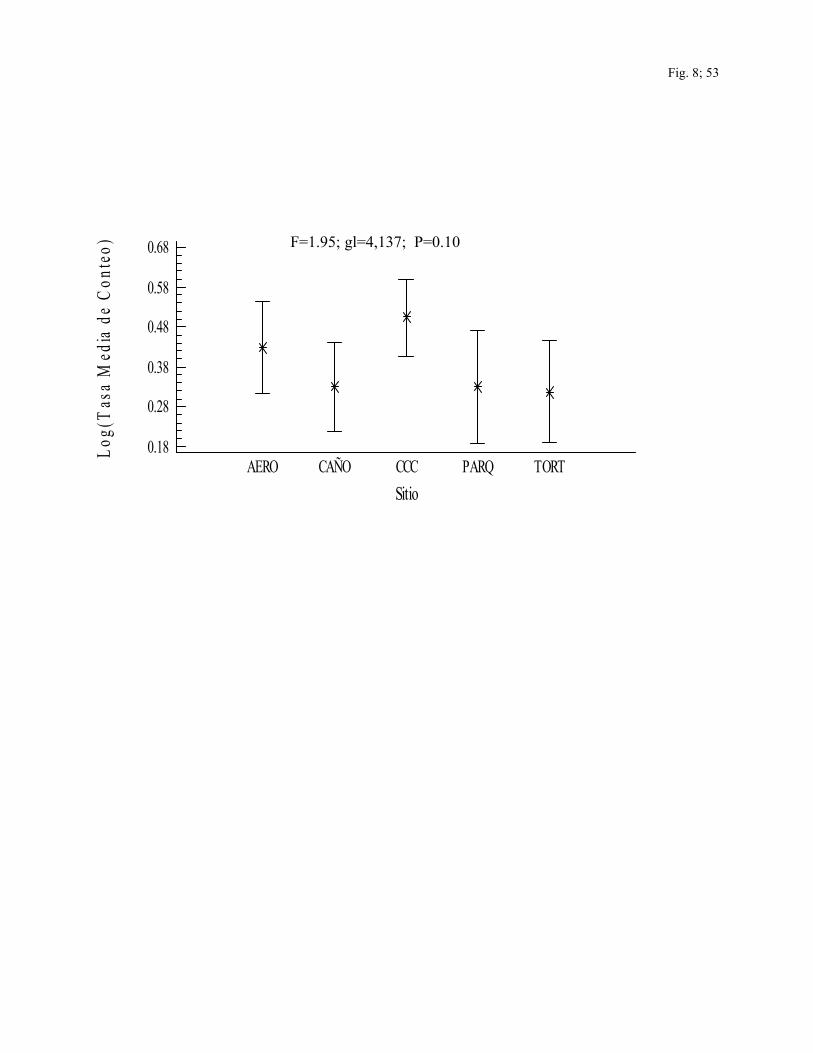
Sitio
Log(Tasa Media de Conteo)
AERO CAÑO CCC PARQ TORT0.18
0.28
0.38
0.48
0.58
0.68 F=1.95; gl=4,137; P=0.10
Fig. 8; 53

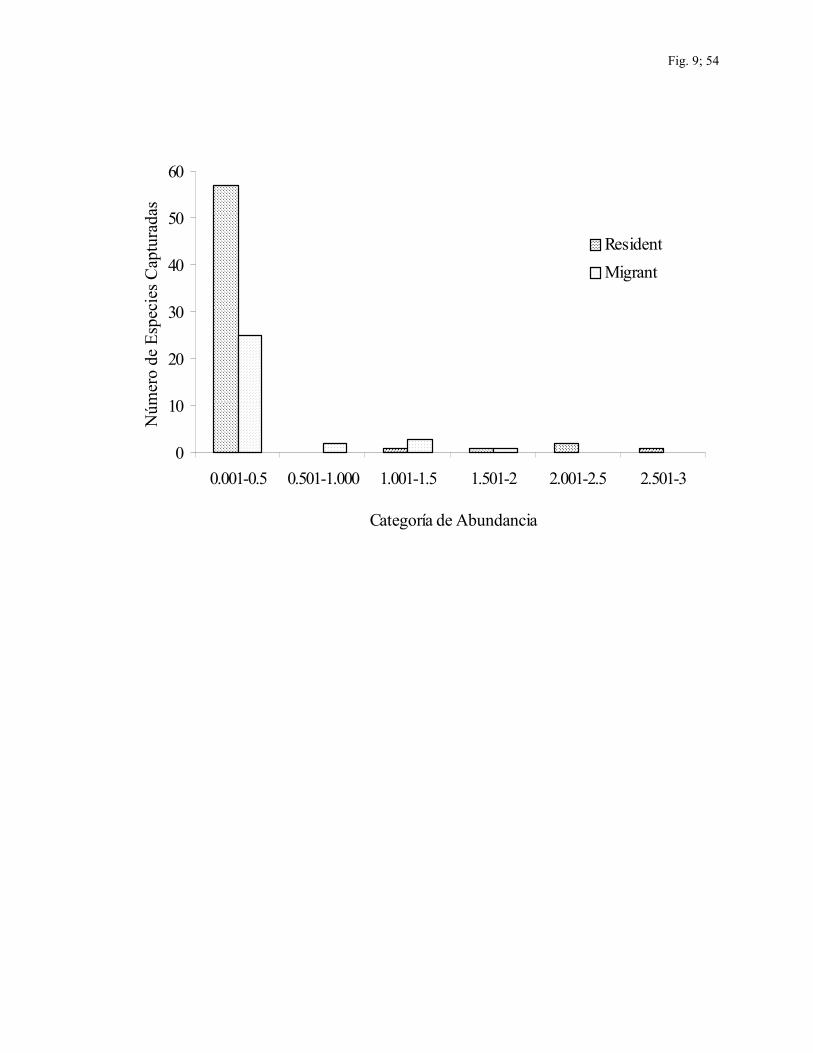
0
10
20
30
40
50
60
0.001-0.5 0.501-1.000 1.001-1.5 1.501-2 2.001-2.5 2.501-3
Categoría de Abundancia
Número de Especies Capturadas
Resident
Migrant
Fig. 9; 54
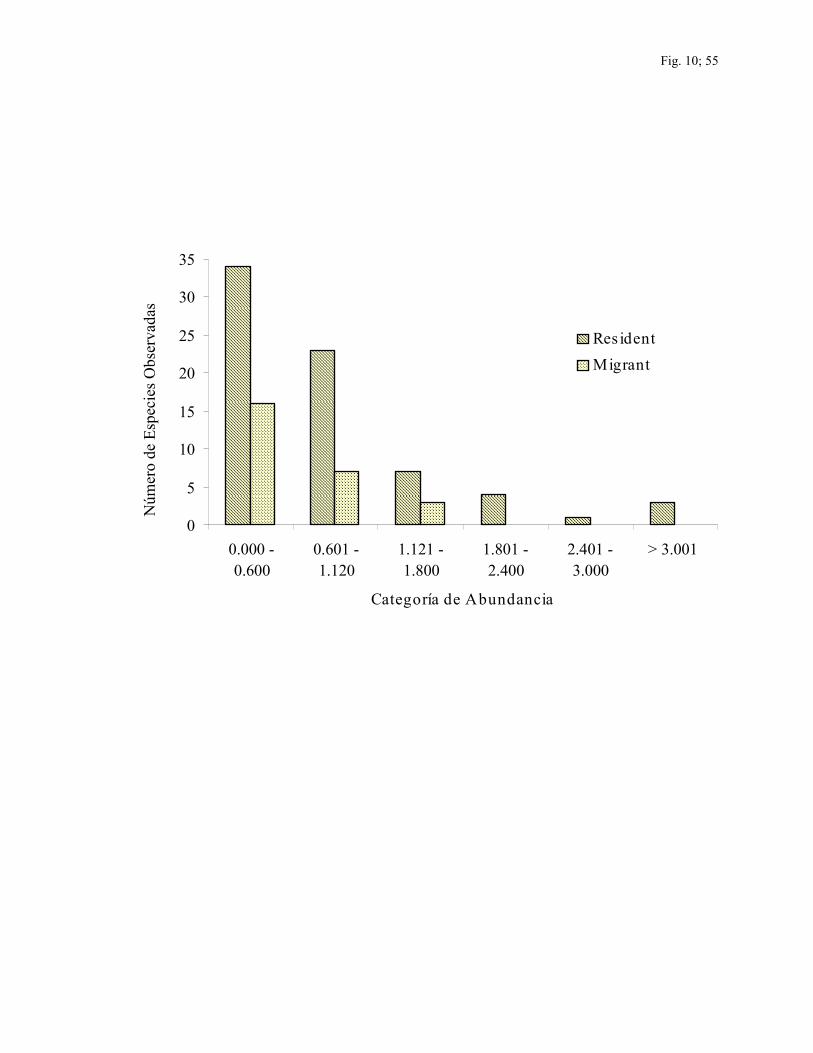
0
5
10
15
20
25
30
35
0.000 -0.600
0.601 -1.120
1.121 -1.800
1.801 -2.400
2.401 -3.000
> 3.001
Categoría de Abundancia
Número de Especies Observadas
Res ident
Migrant
Fig. 10; 55
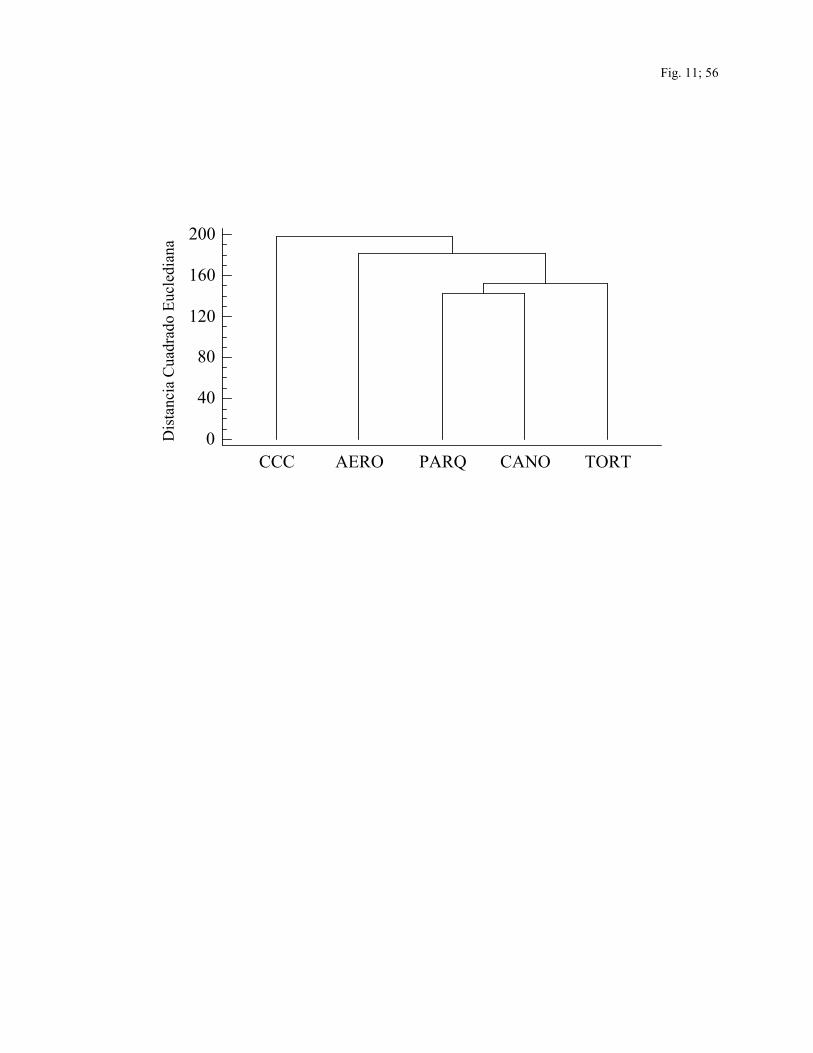
Distance
0
40
80
120
160
200
CCC AERO PARQ TORTCANO
Fig. 11; 56
Distancia Cuadrado Euclediana
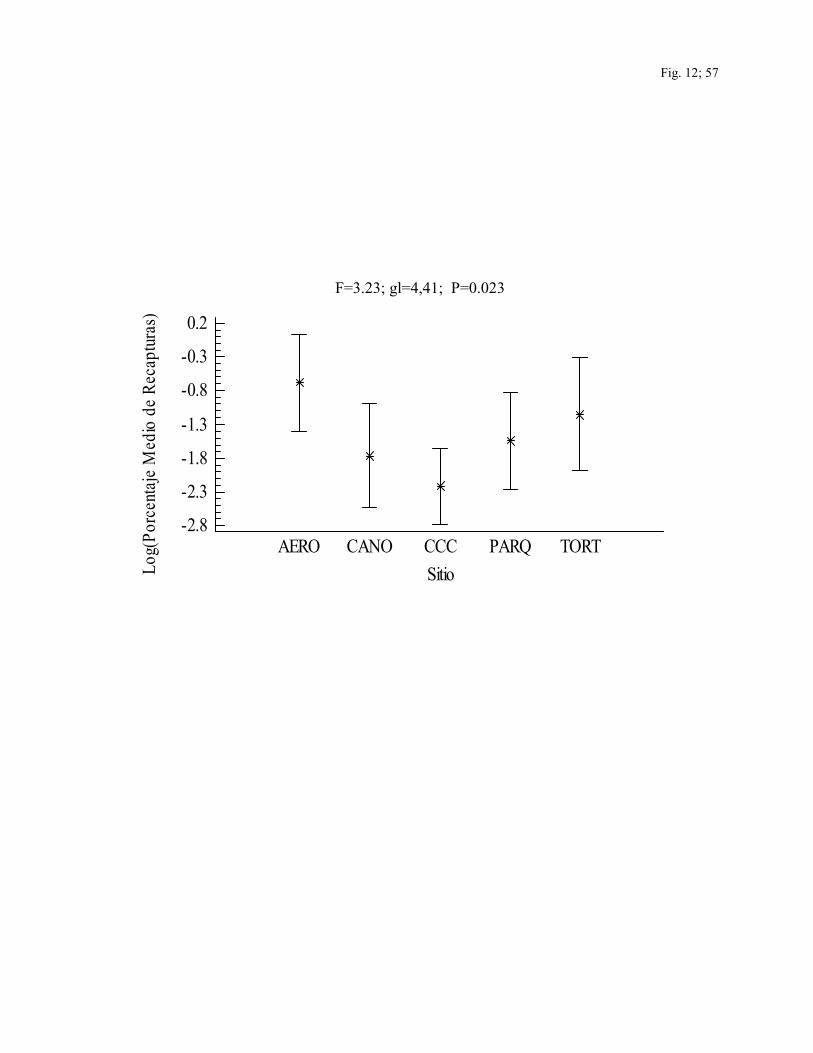
F=3.23; gl=4,41; P=0.023
SitioLog(Porcentaje Medio de Recapturas)
AERO CANO CCC PARQ TORT-2.8
-2.3
-1.8
-1.3
-0.8
-0.3
0.2
Fig. 12; 57
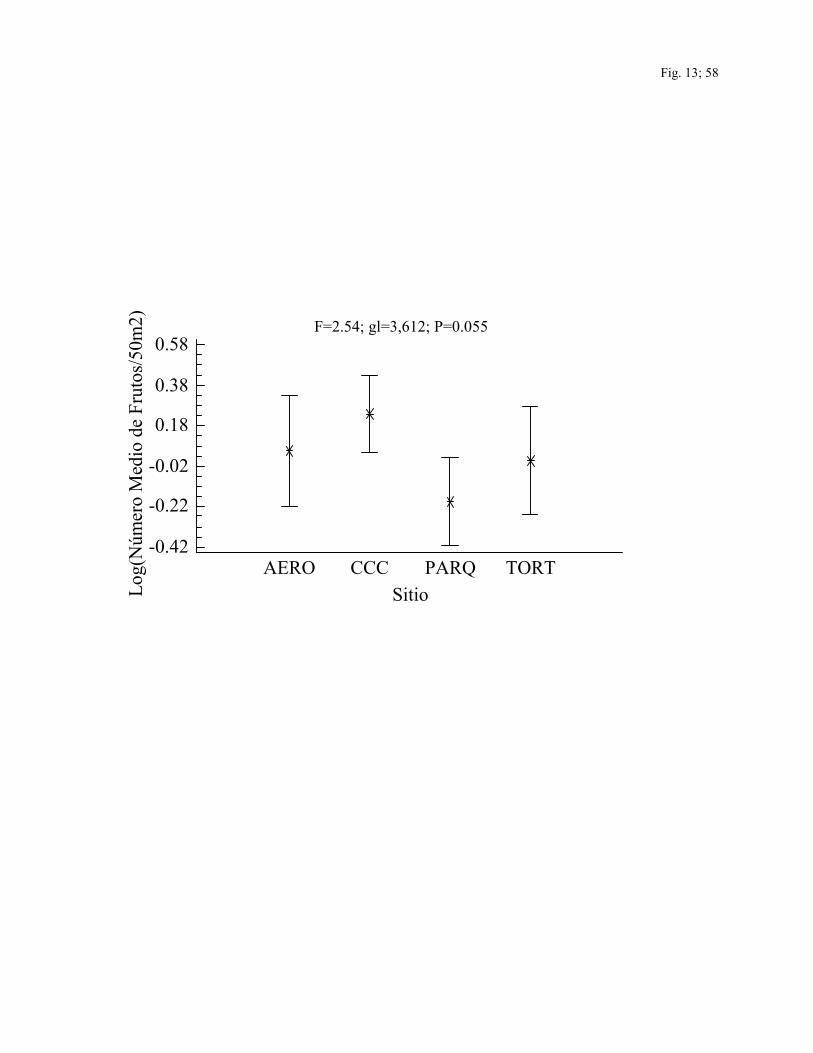
SitioLog(Número Medio de Frutos/50m2)
AERO CCC PARQ TORT-0.42
-0.22
-0.02
0.18
0.38
0.58F=2.54; gl=3,612; P=0.055
Fig. 13; 58

SitioLog(Número Medio de Frutos/50m2)
Set1 Set2 Oct1 Oct2 Nov1Nov2 Dic1-0.8
-0.4
0
0.4
0.8
1.2 F=7.44; gl=6,612; P<0.0001
Fig. 14; 59
Log(Número Medio de Frutos /50 m2

Sitio: F=6.88; gl=4,12; P=0.0004 Mes: F=13.27; gl=4,12; P<0.0001 Interacción: F= 0.86; gl=12,75; P=0.59
56 60
Fig. 15; 60
Número Medio de Especies de Aves Frugìvoras
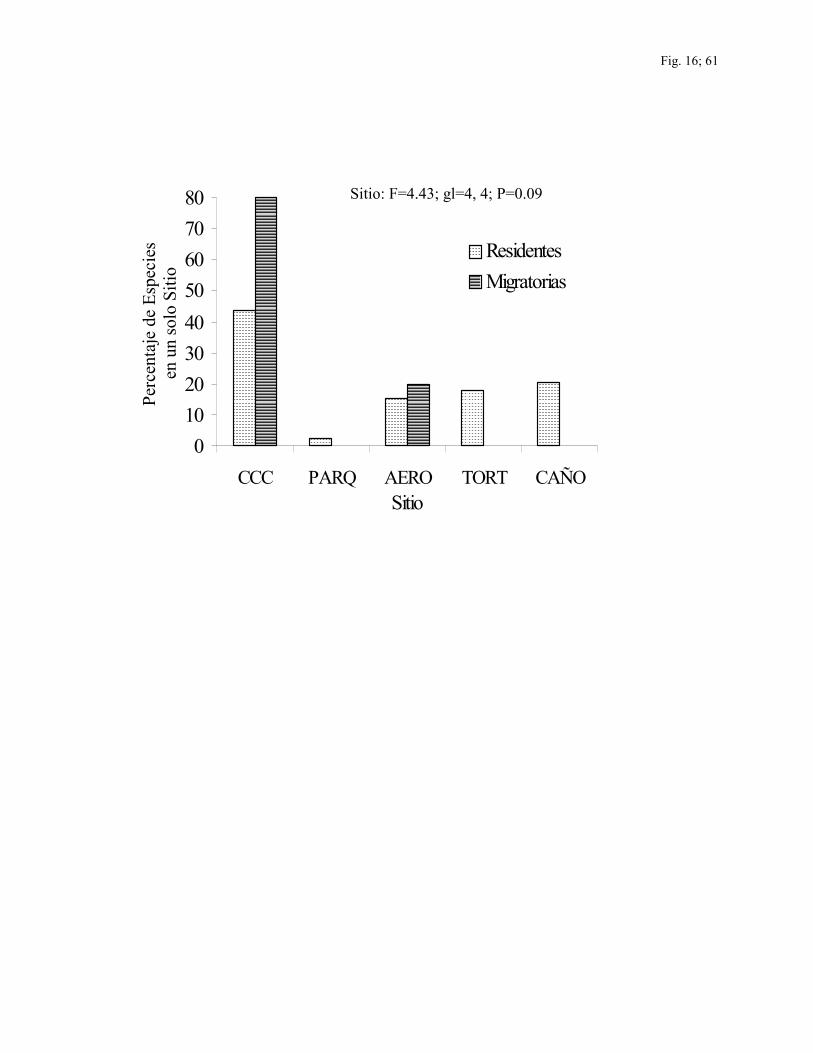
0
10
20
30
40
50
60
70
80
CCC PARQ AERO TORT CAÑOSitio
Percentaje de Especies
en un solo Sitio
Residentes
Migratorias
Sitio: F=4.43; gl=4, 4; P=0.09
Fig. 16; 61
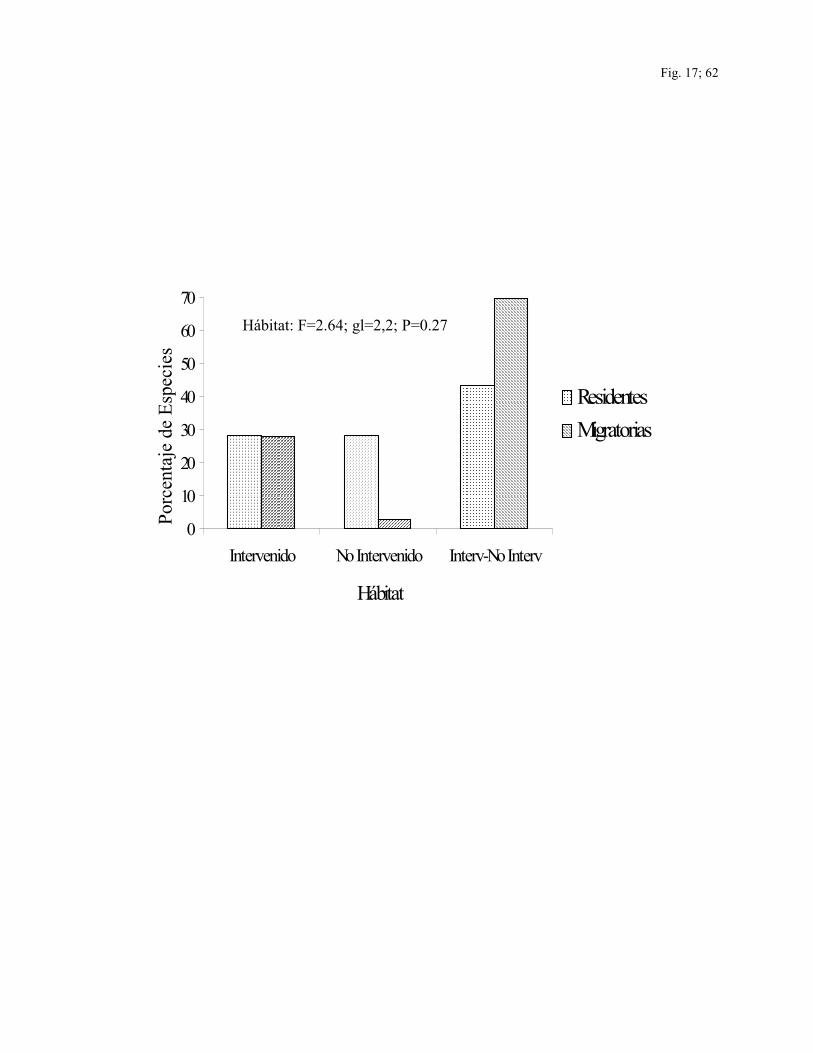
0
10
20
30
40
50
60
70
Intervenido No Intervenido Interv-No Interv
Hábitat
Porcentaje de Especies
Residentes
Migratorias
Hábitat: F=2.64; gl=2,2; P=0.27
Fig. 17; 62

APÉNDICE A. Especies, tipo de hábitats y sitios donde fueron detectadas las especies de aves durante captura con redes de niebla y conteos de búsqueda de aves en cinco sitios en el Caribe norte de Costa Rica.
Nº Taxon Nombre Común Estatusa Hábitatb Sitioc
1
PELECANIDAE Pelecanus occidentalis
Brown Pelican
R
I
CCC
2
FREGATIDAE Fregata magnificens
Magnificent Fregata
R
I, NI
CCC, CAÑO
3
ARDEIDAE Ardea alba
Great Egret
R
I
AERO
4 Egretta thula Snowy Egret R I CCC
5 Egretta caerulea Little-blue Heron R I, NI CCC, AERO, TORT
6 CATHARTIDAE Coragyps atratus
Black Vulture
R
I, NI
CCC, AERO, TORT, CAÑO
7 Cathartes aura Turkey Vulture R I, NI CCC, CAÑO
8
ACCIPITRIDAE Leucopternis semiplumbea
Semiplumbeous Hawk
R
NI
TORT
9 Buteogallus anthracinus Common Black Hawk R I CCC
10
FALCONIDAE Herpeotheres cachinnans
Laughing Falcon
R
I
CCC
11
SCOLOPACIDAE Numenius phaeopus
Whimbrel
M
I
AERO
12
COLUMBIDAE Columba flavirostris
Red-billed Pigeon
R
I
CCC
13 Columba cayennensis Pale-vented Pigeon R I CCC, AERO 14 Columba nigrirostris Short-billed Pigeon R NI CAÑO 15 Leptotila cassini Gray-chested Dove R I, NI PARQ, AERO, TORT, CAÑO 16 Geotrygon montana Ruddy Quail Dove R I CCC 63

APÉNDICE A. (Continuación)
Nº Taxon Nombre Común Estatus Hábitat Sitio
17
PSITTACIDAE Aratinga nana
Olive-throated Parrakeet
R
I
AERO
18 Ara ambigua Great Green Macaw R NI TORT 19 Pionus senilis White-crowned Parrot R I, NI CCC, AERO, TORT 20 Amazona autumnalis Red-lored Parrot R I AERO 21 Amazona farinosa Mearly Parrot R NI TORT
22
CUCULIDAE Piaya cayana
Squirrel Cuckoo
R
NI
CAÑO
23
TROCHILIDAE Glaucis aenea
Bronzy Hermit
R
I, NI
Todos los sitios
24 Threnetes ruckeri Band-tailed Barbthroat R NI CAÑO 25 Phaethornis superciliosus Long-tailed Hermit R I, NI Todos los sitios 26 Phaethornis longuemareus Little Hermit R I, NI CCC, PARQ, CAÑO 27 Anthracothorax prevostii Green-breasted Mango R I, NI CCC, PARQ, AERO 28 Thalurania colombica Violet-crowned Woodnymph R I, NI Todos los sitios 29 Hylocharis eliciae Blue-throated Goldentail R I, NI CCC, CAÑO 30 Amazilia amabilis Blue-chested Hummingbird R I, NI CCC, CAÑO 31 Amazilia rutila Cinnamon Hummingbird R I CCC 32 Amazilia tzacatl Rufous-tailed Hummingbird R I, NI Todos los sitios 33 Heliothryx barroti Purple-crowned Fair R NI CAÑO
34
TROGONIDAE Trogon rufus
Black-throated Trogon
R
NI
PARQ, TORT, CAÑO
35 Trogon massena Slaty-tailed Trogon R I, NI Todos los sitios
36
ALCEDINIDAE Ceryle torquata
Ringed Kingfisher
R
I
AERO
37 Chloroceryle americana Green Kingfisher R I AERO
64
APÉNDICE A. (Continuación)
Nº Taxon Nombre Común Estatus Hábitat Sitio
38 Chloroceryle aenea American Pygmy Kingfisher R NI TORT, CAÑO
39
BUCCONIDAE Monasa morphoeus
White-fronted Nunbird
R
NI
PARQ, TORT
40
RAMPHASTIDAE Pteroglossus torquatus
Collared Aracari
R
I, NI
PARQ, AERO, TORT, CAÑO
41 Ramphastos sulfuratus Keel-billed Toucan R I, NI PARQ, AERO, TORT, CAÑO 42 Ramphastos swainsonii Chestnut-mandible Toucan R NI TORT, CAÑO
43
PICIDAE Dryocopus lineatus
Lineated Woodpecker
R
I, NI
CCC, TORT, CAÑO
44 Campephilus guatemalensis Pale-billed Woodpecker R NI TORT
45
DENDROCOLAPTIDAE Dendrocolaptes certhia*
Barred Woodcreeper
R
I, NI
PARQ, AERO, TORT, CAÑO
46 Xiphorhynchus erythropygius Spotted Wodckreeper R I CCC 47 Lepidocolaptes souleyetii Streak-headed Woodcreeper R I CCC, AERO
48
THAMNOPHILIDAE Thamnophilus doliatus
Barred Ant-Shrike
R
I
CCC
49 Thamnophilus atrinucha Slaty-AntShrike R I, NI PARQ, AERO, TORT, CAÑO 50 Myrmotherula axillaris White-flanked Antwren R NI CAÑO 51 Cercomacra tyrannina Dusky Antbird R I CCC 52 Myrmeciza exsul Chestnut-backed Antbird R NI PARQ, TORT, CAÑO 53 Gymnopithys leucaspis Bicolored Antbird R NI PARQ, CAÑO 54
FORMICARIDAE Formicarius analis
Black-faced Antthrush
R
NI
TORT
55
TYRANNIDAE Elaenia flavogaster
Yellow-bellied Elaenia
R
I, NI
CCC, PARQ
65
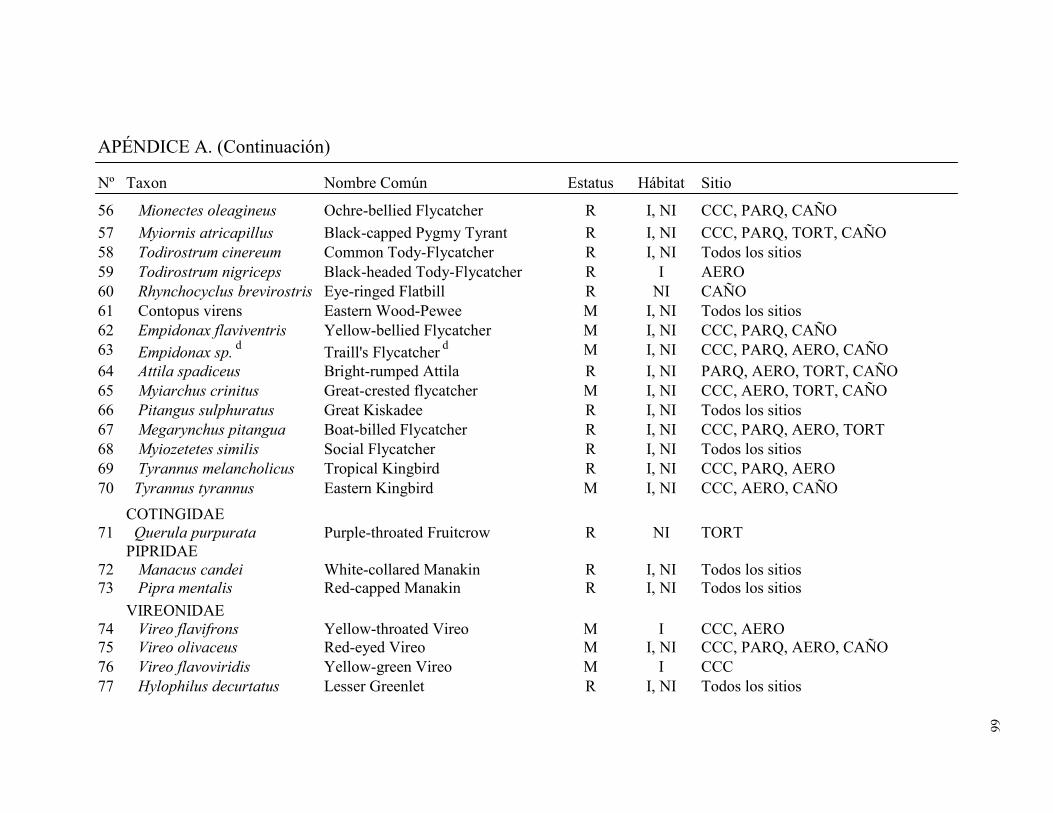
APÉNDICE A. (Continuación)
Nº Taxon Nombre Común Estatus Hábitat Sitio
56 Mionectes oleagineus Ochre-bellied Flycatcher R I, NI CCC, PARQ, CAÑO
57 Myiornis atricapillus Black-capped Pygmy Tyrant R I, NI CCC, PARQ, TORT, CAÑO 58 Todirostrum cinereum Common Tody-Flycatcher R I, NI Todos los sitios 59 Todirostrum nigriceps Black-headed Tody-Flycatcher R I AERO 60 Rhynchocyclus brevirostris Eye-ringed Flatbill R NI CAÑO 61 Contopus virens Eastern Wood-Pewee M I, NI Todos los sitios 62 Empidonax flaviventris Yellow-bellied Flycatcher M I, NI CCC, PARQ, CAÑO 63 Empidonax sp. d Traill's Flycatcher d M I, NI CCC, PARQ, AERO, CAÑO 64 Attila spadiceus Bright-rumped Attila R I, NI PARQ, AERO, TORT, CAÑO 65 Myiarchus crinitus Great-crested flycatcher M I, NI CCC, AERO, TORT, CAÑO 66 Pitangus sulphuratus Great Kiskadee R I, NI Todos los sitios 67 Megarynchus pitangua Boat-billed Flycatcher R I, NI CCC, PARQ, AERO, TORT 68 Myiozetetes similis Social Flycatcher R I, NI Todos los sitios 69 Tyrannus melancholicus Tropical Kingbird R I, NI CCC, PARQ, AERO 70 Tyrannus tyrannus Eastern Kingbird M I, NI CCC, AERO, CAÑO
71
COTINGIDAE Querula purpurata
Purple-throated Fruitcrow
R
NI
TORT
72
PIPRIDAE Manacus candei
White-collared Manakin
R
I, NI
Todos los sitios
73 Pipra mentalis Red-capped Manakin R I, NI Todos los sitios
74
VIREONIDAE Vireo flavifrons
Yellow-throated Vireo
M
I
CCC, AERO
75 Vireo olivaceus Red-eyed Vireo M I, NI CCC, PARQ, AERO, CAÑO 76 Vireo flavoviridis Yellow-green Vireo M I CCC 77 Hylophilus decurtatus Lesser Greenlet R I, NI Todos los sitios
66

APÉNDICE A. (Continuación)
Nº Taxon Nombre Común Estatus Hábitat Sitio
78
HIRUDINIDAE Hirudindo rustica
Barn Swallow
R
I
CCC
79
TROGLODYTIDAE Thryothorus nigricapillus
Bay Wren
R
I, NI
CCC, AERO, TORT, CAÑO
80 Thryothorus thoracicus Striped-breasted Wren R NI TORT 81 Troglodytes aedon House Wren R I, NI CCC, PARQ, AERO 82 Henicorhina leucosticta White-breasted Wood-Wren R NI TORT, CAÑO 83
SILVIIDAE Ramphocaenus melanurus
Long-billed Gnatwren
R
NI
CAÑO
84 Polioptila plumbea Tropical Gnatwren R I CCC 85
TURDIDAE Catharus fuscescens
Veery
M
I, NI
Todos los sitios
86 Catharus minimus Gray-cheeked Thrush M I, NI Todos los sitios 87 Catharus ustulatus Swainson's Thrush M I, NI Todos los sitios 88 Hylocichla mustelina Wood Thrush M NI PARQ, TORT, CAÑO 89 Turdus grayi Clay-colored Robin R I, NI CCC, PARQ, TORT, CAÑO 90
MIMIDAE Dumetella carolinensis
Gray Catbird
M
I, NI
CCC, AERO, TORT
91
PARULIDAE Vermivora peregrina
Tennessee Warbler
M
I
CCC, AERO
92 Dendroica petechia Yellow Warbler M I, NI Todos los sitios 93 Dendroica pensylvanica Chestnut-side Warbler M I, NI Todos los sitios 94 Dendroica magnolia Magnolia Warbler M I, NI CCC, PARQ, AERO, TORT 95 Dendroica fusca Blackburnian Warbler M I, NI AERO, CAÑO 96 Dendroica castanea Bay-breasted Warbler M I, NI Todos los sitios 97 Dendroica cerulea Cerulean Warbler M I CCC
67
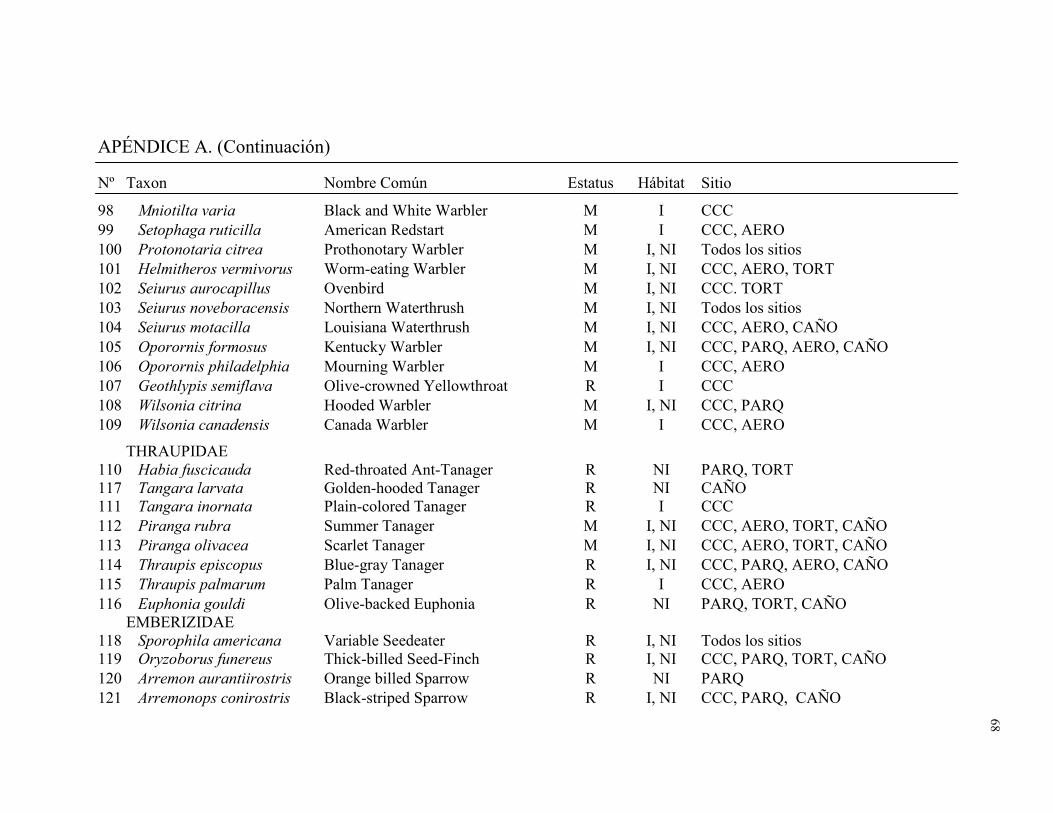
APÉNDICE A. (Continuación)
Nº Taxon Nombre Común Estatus Hábitat Sitio
98 Mniotilta varia Black and White Warbler M I CCC 99 Setophaga ruticilla American Redstart M I CCC, AERO 100 Protonotaria citrea Prothonotary Warbler M I, NI Todos los sitios 101 Helmitheros vermivorus Worm-eating Warbler M I, NI CCC, AERO, TORT 102 Seiurus aurocapillus Ovenbird M I, NI CCC. TORT 103 Seiurus noveboracensis Northern Waterthrush M I, NI Todos los sitios 104 Seiurus motacilla Louisiana Waterthrush M I, NI CCC, AERO, CAÑO 105 Oporornis formosus Kentucky Warbler M I, NI CCC, PARQ, AERO, CAÑO 106 Oporornis philadelphia Mourning Warbler M I CCC, AERO 107 Geothlypis semiflava Olive-crowned Yellowthroat R I CCC 108 Wilsonia citrina Hooded Warbler M I, NI CCC, PARQ 109 Wilsonia canadensis Canada Warbler M I CCC, AERO
110
THRAUPIDAE Habia fuscicauda
Red-throated Ant-Tanager
R
NI
PARQ, TORT
117 Tangara larvata Golden-hooded Tanager R NI CAÑO 111 Tangara inornata Plain-colored Tanager R I CCC 112 Piranga rubra Summer Tanager M I, NI CCC, AERO, TORT, CAÑO 113 Piranga olivacea Scarlet Tanager M I, NI CCC, AERO, TORT, CAÑO 114 Thraupis episcopus Blue-gray Tanager R I, NI CCC, PARQ, AERO, CAÑO 115 Thraupis palmarum Palm Tanager R I CCC, AERO 116 Euphonia gouldi Olive-backed Euphonia R NI PARQ, TORT, CAÑO 118
EMBERIZIDAE Sporophila americana
Variable Seedeater
R
I, NI
Todos los sitios
119 Oryzoborus funereus Thick-billed Seed-Finch R I, NI CCC, PARQ, TORT, CAÑO 120 Arremon aurantiirostris Orange billed Sparrow R NI PARQ 121 Arremonops conirostris Black-striped Sparrow R I, NI CCC, PARQ, CAÑO
68
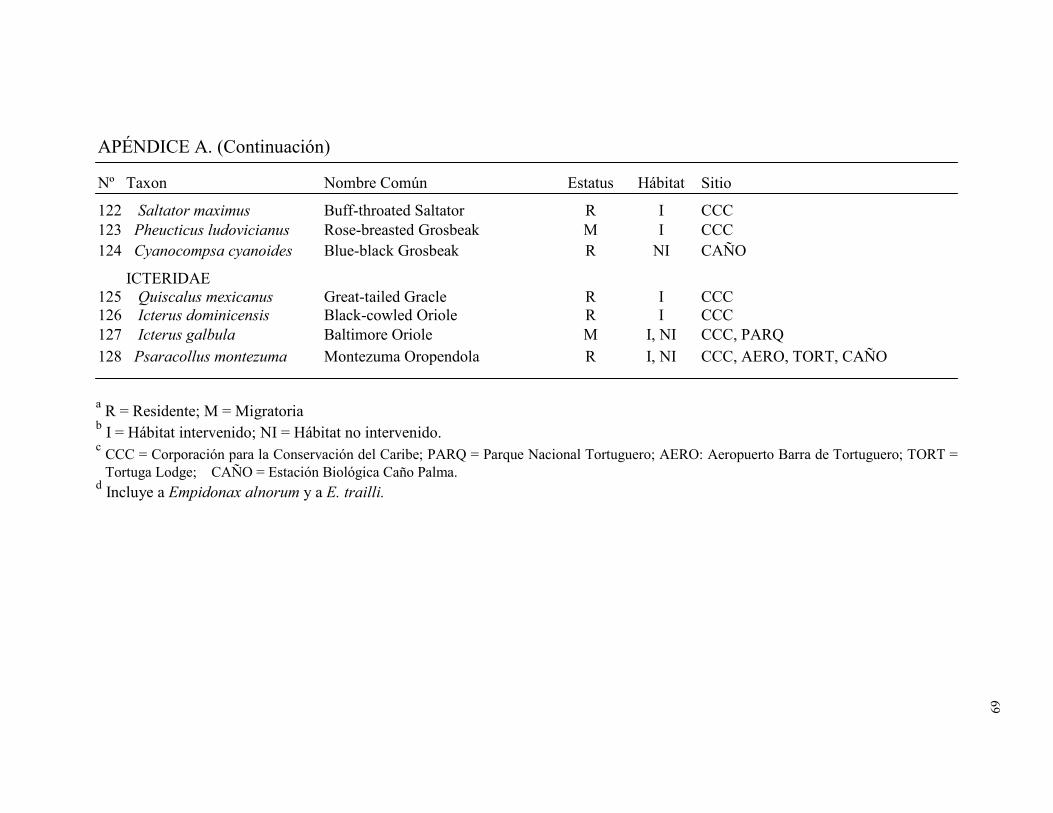
APÉNDICE A. (Continuación)
Nº Taxon Nombre Común Estatus Hábitat Sitio
122 Saltator maximus Buff-throated Saltator R I CCC 123 Pheucticus ludovicianus Rose-breasted Grosbeak M I CCC 124 Cyanocompsa cyanoides Blue-black Grosbeak R NI CAÑO
125
ICTERIDAE Quiscalus mexicanus
Great-tailed Gracle
R
I
CCC
126 Icterus dominicensis Black-cowled Oriole R I CCC 127 Icterus galbula Baltimore Oriole M I, NI CCC, PARQ
128 Psaracollus montezuma Montezuma Oropendola R I, NI CCC, AERO, TORT, CAÑO
a R = Residente; M = Migratoria b I = Hábitat intervenido; NI = Hábitat no intervenido. c CCC = Corporación para la Conservación del Caribe; PARQ = Parque Nacional Tortuguero; AERO: Aeropuerto Barra de Tortuguero; TORT = Tortuga Lodge; CAÑO = Estación Biológica Caño Palma.
d Incluye a Empidonax alnorum y a E. trailli.
69

Maglianesi
70
UNIVERSIDAD NACIONAL
SISTEMA DE ESTUDIOS DE POSGRADO
PROGRAMA REGIONAL EN MANEJO DE VIDA SILVESTRE PARA
MESOAMÉRICA Y EL CARIBE
COMPARACIÓN DE DOS TÉCNICAS DE MUESTREO DE AVES: CAPTURA CON
REDES DE NIEBLA Y BÚSQUEDA DE AVES, EN UN BOSQUE TROPICAL MUY
HÚMEDO EN EL CARIBE NORTE DE COSTA RICA
por
María Alejandra Maglianesi
Programa Regional en Manejo de Vida Silvestre para Mesoamérica y el Caribe,
Universidad Nacional. Apartado 1350-3000, Heredia, Costa Rica.
Teléfono: 237-7039. Correo electrónico: [email protected]
Noviembre del 2004

Maglianesi
71
M. A. Maglianesi María Alejandra Maglianesi. Programa Técnicas de muestreo de aves Regional en Manejo de Vida Silvestre.
Universidad Nacional. Apartado 1350-3000. Heredia – Costa Rica
Comparación de dos técnicas de muestreo de aves: captura con redes de niebla y búsqueda
de aves, en un Bosque Tropical muy Húmedo en el Caribe norte de Costa Rica
María Alejandra Maglianesi1
Programa Regional en Manejo de Vida Silvestre para Mesoamérica y el Caribe
Universidad Nacional. Apartado 1350 – 3000 Heredia, Costa Rica.
1Autor para correspondencia: E-mail: <[email protected]>

Maglianesi
72
ABSTRACT. I compared the results of mist-netting and area-search bird surveys
during the fall season over a 5-month period in five sites, two disturbed and three
undisturbed in wet lowland forest, near Tortuguero, northern Caribbean coast in Costa
Rica. I estimated differences in abundance, composition and richness of resident and
migrant landbirds with the two survey methods. I recorded 110 species across the five
sites, including 81 species (73.64%) captured in mist nets and 90 species (81.82%)
recorded during area searches. There were 61 species detected by both methods. Of the
total species detected, 29 species (26.4%) were not captured and 20 species (18.2%) were
not recorded during area searches. Number of resident species captured was slightly
correlated with number of resident species detected by area searches and the reverse was
true for migratory species with statistical significance. Mean species detection rate
recorded with area searches was greater than with mist nets and mean number of resident
species was greater with both methods. However, mist-netting detected a greater
percentage of migratory species than area searches. Species-accumulation curves
indicated that species increased more rapidly with area searching than mist-netting over
the first samples, but the two methods had a similar trend with more samples. For the total
species detected by both methods, relative abundance estimates from mist-netting and
area searches were significantly related. Over 89% of the 61 species the two methods
were consistent in measuring relative abundance. Only for three species area searches
tended to produce an extremely high relative abundance. Individuals planning bird
surveys should recognize biases and be careful in interpreting results before taking
management decisions involving bird populations. If they are going to survey the entire
bird community or guilds this may require using multiple survey methods. Despite of the
fact that area searches alone would be the most time efficient and least costly technique, it
should be used combining other methods to have a more relatively complete description
of the bird community in the tropics.
Key words: counts, mist-netting, area search, bird survey, bird methods, bird
communities

Maglianesi
73
SINOPSIS. En este estudio se compararon los resultados obtenidos a partir de captura
de aves con redes de niebla y búsqueda de aves durante cinco meses en cinco sitios
diferentes, de los cuales 2 corresponden a bosques alterados y 3 a bosques primarios, en
los alrededores de Tortuguero, Provincia de Limón, en la costa norte del Caribe de Costa
Rica. Se determinó si estimaciones de composición, riqueza y abundancia relativa
variaron con el método empleado. Se registraron 110 especies distribuidas en los cinco
sitios, incluyendo 81 especies (73.64%) capturadas y 90 especies (81.82%) detectadas en
búsquedas de aves. Se registraron 61 especies comunes a ambos métodos. Del total de
especies, 29 (26.4%) no estuvieron representadas en las capturas y 20 (18.2%) no
estuvieron representadas en las búsquedas de aves. El número de especies residentes
capturadas estuvo poco correlacionado con el número de especies residentes detectadas
durante búsqueda de aves, mientras que las especies migratorias tuvieron una correlación
significativa. La tasa promedio de detección de especies fue mayor con búsqueda de aves
que con capturas, sin embargo las capturas tuvieron un mayor porcentaje de especies
migratorias. La curva de acumulación de especies tuvo una mayor pendiente para el
método de búsqueda de aves, aunque en ambos casos alcanzaron a estabilizarse de
manera similar. Para el total de especies registradas por ambos métodos, la abundancia
relativa mostró una correlación positiva significativa para ambos métodos y para el 89%
de las especies los estimados de abundancia relativa fueron precisos para ambos métodos.
Solamente 3 especies tuvieron un estimado de abundancia relativa extremadamente alto a
partir del método de búsqueda de aves. Es necesario reconocer los sesgos asociados a las
técnicas de muestreo de aves e interpretar cuidadosamente los resultados antes de tomar
decisiones de manejo. A pesar de que el método de búsqueda de aves resulta más
eficiente en cuanto a tiempo y costos, debería ser utilizado en combinación con otros
métodos para obtener una representación más completa de la comunidad de aves en los
trópicos.
Palabras clave: captura de aves, búsqueda de aves, métodos, muestreo de aves,
comunidad, técnicas muestreo

Maglianesi
74
INTRODUCCIÓN
El muestreo de las comunidades de aves en los bosques tropicales ha llegado a ser
problemático debido a que estos bosques poseen una alta diversidad de especies y una
vegetación exuberante (Karr et al. 1990). La descripción de la composición de un
ensamblaje de aves en un área en particular no resulta fácil y depende en gran medida, de
los métodos empleados (Terborgh et al. 1990, Robinson et al. 2000) . Además, los
métodos pueden influenciar especialmente las comparaciones que se realizan entre
diferentes hábitats (Blake y Loiselle 2001).
El método de captura con redes de niebla ha sido ampliamente utilizado para
documentar la abundancia relativa, patrones de diversidad y distribución de las especies
de aves en muchos hábitats tropicales. Un método que se ha desarrollado más
recientemente en algunos países es el método de búsqueda de aves (Ralph et al. 1993),
una técnica que se caracteriza por poseer un tiempo y área restringidos (Slater 1994).
Posiblemente, el método de captura con redes de niebla ha tenido una historia más
larga y controversial en los estudios de las comunidades tropicales (Terborgh et al. 1990,
Remsen y Good 1996, Robinson et al. 2000), en comparación con otras técnicas de
muestreo de aves. Tanto el método de captura con redes como los métodos visuales-
auditivos poseen ventajas y desventajas (Karr 1981, Karr 1990, Remsen y Good 1996) y
cada uno puede proveer diferentes perspectivas sobre la estructura de la comunidad de
aves (Whitman et al. 1997, Blake y Loiselle 2000). Aunque se ha sugerido que ambas
técnicas son sesgadas (Karr 1981, Dieni y Jones 2002), las comparaciones entre estos
sesgos no se encuentran bien documentadas.
El método de captura con redes ofrece una posibilidad de acumular información sobre
la actividad de las especies de sotobosque en un periodo de tiempo relativamente corto.
Más aún, tanto el tamaño de muestra como el esfuerzo de muestreo pueden ser
estandarizados (Karr 1990). No obstante, debido a que el método de captura con redes
detecta a las aves mediante capturas, indudablemente posee la ventaja de detectar aquellas
especies poco conspicuas y quietas que pueden ser un componente importante de la
avifauna.
Sin embargo, los datos obtenidos a partir de capturas, sólo permiten monitorear
variaciones espaciales de especies de aves que usan un estrato específico dentro del

Maglianesi
75
bosque, pero por sí mismos, podrían no proveer información sobre otras especies de la
comunidad. A un nivel de población, existen diferentes factores que influyen sobre las
estimaciones de abundancia de las especies, como la distancia que se desplazan las aves
entre sitios de alimentación y los sistemas sociales que afectan fuertemente la
probabilidad de captura. Las especies que viajan largas distancias son sobreestimadas por
las capturas. Además, la eficiencia de captura de redes estándar depende del tamaño de
las especies. Así, las aves de mayor tamaño pueden escapar más fácilmente de las redes
en comparación con aquellas de pequeño a mediano tamaño (Jenni et al. 1996).
Los métodos visuales-auditivos registran una proporción considerablemente mayor de
la avifauna local que el de captura con redes de niebla, según algunos estudios (Whitman
et al. 1997), aunque estos métodos poseen sesgos asociados al observador, como la
familiaridad con las vocalizaciones y la habilidad para identificar especies que se mueven
muy rápidamente en el bosque por breves momentos (Karr 1990). Por lo tanto, las
técnicas de conteo requieren de ciertas habilidades del observador relacionadas con los
cantos y llamadas de, muchas veces, cientos de especies de aves (Beehler et al. 1995). El
conocimiento de las vocalizaciones tiene gran importancia en un boque muy húmedo
tropical en particular, donde la detección visual de las especies se vuelve difícil y aún así,
se podrían estar perdiendo aquellas especies quietas y no vocales (Karr 1981).
El método de búsqueda de aves, por su diseño, es menos rígido que otras técnicas
estandarizadas, debido a que el observador puede desplazarse libremente dentro de un
área determinada durante el transcurso del muestreo, en contraste con los métodos de
puntos de conteo y transectos, en donde el observador debe permanecer en un punto fijo o
sus movimientos están más restringidos (Dieni y Jones 2002). Una mayor movilidad del
observador permite a su vez, que muchas especies sean identificadas de forma más
precisa, a la vez que aquellas poco familiares e inconspicuas pueden ser detectadas
(Hewish y Loyn 1989, Ralph et al. 1993).
Existen muchos estudios que han comparado los resultados obtenidos por los métodos
de captura con redes y puntos de conteo (Blake y Loiselle 1989, Nur y Geupel 1993,
Ralph y Fancy 1995, Gram y Faaborg 1997, Whitman et al. 1997, Blake y Loiselle 2002,
Rappole et al. 1998, Pagen et al. 2000, Pagen et al. 2002, Wang y Finch 2002). Otro
estudio comparó el método de captura con el de conteos de cantos (Rappole et al. 1993) y

Maglianesi
76
otro comparó los métodos de búsqueda de aves con el de mapeo (Dieni y Jones 2002). Sin
embargo, comparaciones entre el método de captura con redes y el de búsqueda de aves,
no se encuentran documentados aún.
El presente estudio describe la composición, abundancia y riqueza de las comunidades
de aves residentes y migratorias en cinco sitios diferentes. Los resultados en este estudio
pueden ser de utilidad para aquellos investigadores que se encuentran en una fase de
selección de métodos de muestreo para el monitoreo de las poblaciones de aves.
El objetivo de este estudio fue responder a las siguientes preguntas:
(1) ¿Pueden los métodos de captura con redes o búsqueda de aves por sí solos producir
una estimación confiable de la presencia/ausencia de especies de aves? (2) ¿El número de
especies residentes y migratorias detectadas siguen un patrón similar entre ambos
métodos? (3) ¿El número de especies detectadas por familia depende del método
empleado? (4) ¿El número de especies y la proporción de especies migratorias por sitio
varían con el método utilizado? (5) ¿Los métodos difieren en la tasa de acumulación de
nuevas especies en cada sitio? y (6) ¿Existe una consistencia en la abundancia relativa
estimada por ambos métodos?.
MÉTODOS
Área de estudio. La fase de campo fue realizada entre agosto y diciembre del 2002
en la zona de Tortuguero, ubicada en la costa del Caribe norte en la Provincia de Limón,
Costa Rica (Fig. 1). La principal zona de vida en las tierras bajas del Caribe es el Bosque
Tropical muy Húmedo (Holdridge 1969), el cual se caracteriza por poseer una vegetación
siempreverde que refleja una breve estación seca (Stiles 1983). El Bosque Tropical muy
Húmedo presenta un dosel alto y una variedad de estratos de vegetación. Unas pocas
especies son caducifolias, pero esto no cambia el aspecto general siempreverde del
bosque (Janzen 1983). El área de estudio se ubica dentro del Área de Conservación
Tortuguero (ACTo), el cual incluye uno de los humedales más grandes del país (Oro
1996). La temperatura media anual es de 29.9ºC y la precipitación media anual de 6,000
mm hace esta área una de las más húmedas del país (Coen 1983, Oro 1996, Stiles y
Skutch 1989). La estación seca se extiende desde finales de diciembre hasta marzo.

Maglianesi
77
Durante los cinco meses de este estudio se registró una precipitación de 2,138 mm y una
temperatura promedio de 26.9ºC, con una mínima de 24.5ºC y una máxima de 30.7ºC.
Tortuguero estaba ampliamente rodeado por bosque continuo hasta 1960, que
posteriormente fue transformado a un mosaico de diferentes estados sucesionales causado
por actividades antrópicas. La expansión de las pasturas para ganado y los cultivos de
banano provocó la pérdida de bosques en las tierras bajas del Atlántico (Boucher et al.
1983). En la actualidad la principal actividad económica en Tortuguero es el turismo,
donde las aves son uno de los grupos más importantes de vida silvestre que atraen a los
turistas, debido a su gran diversidad y a su relativamente fácil detección (Herrera 1998).
La infraestructura hotelera ha aumentado rápidamente en la zona de Tortuguero durante la
última década y ha contribuido a la fragmentación del bosque.
Sitios de estudio. Los sitios estudiados se establecieron en dos tipos de bosques:
intervenido y no intervenido. Dos de los cinco sitios incluyen vegetación alterada por
influencia humana: Corporación para la Conservación del Caribe (CCC) y el Aeropuerto
Barra de Tortuguero (AERO). Los tres sitios restantes se encuentran en bosque primario:
Parque Nacional Tortuguero (PARQ), Tortuga Lodge (TORT) y la Estación Biológica
Caño Palma (CAÑO) (Fig. 1). En los sitios no intervenidos el dosel puede alcanzar 40-50
m de altura y muchos árboles pueden tener hasta 100-200 cm de diámetro a la altura del
pecho (DAP). Los árboles del subdosel miden alrededor de 25-35 m de altura y el
sotobosque mide entre 10 y 25 m de altura. La capa de arbustos en el sotobosque es de 1-
2.5 m de altura y allí se encuentran abundantes palmas. En los sitios intervenidos, donde
fueron talados selectivamente los árboles de mayor DAP hace aproximadamente 20 años,
los árboles del dosel son más bajos con una altura de 20-25 m aproximadamente y la capa
de arbustos del sotobosque es particularmente densa.
AERO se encuentra localizado adyacente al extremo norte de la pista de aterrizaje de
Tortuguero y TORT se encuentra en una propiedad privada del lado opuesto del río,
ambos sitios están a 10 min en lancha desde CCC. Mientras que CAÑO se encuentra
localizado adyacente a la estación canadiense de investigación, a unos 20-25 min en
lancha desde CCC, PARQ corresponde a la esquina noreste del Parque Nacional
Tortuguero y CCC se encuentra contiguo al pueblo de Tortuguero (Fig. 1).

Maglianesi
78
Muestreo de aves. Este estudio siguió el protocolo de campo usado por el Programa
Integrado de Monitoreo de Tortuguero (Ralph et al. 2002). Este programa se inició en
1994 y consiste en un esfuerzo cooperativo que involucra diferentes organizaciones,
colaboradores y voluntarios. Se incluyeron dos técnicas de muestreo de aves: captura con
redes de niebla y conteos de búsqueda de aves siguiendo la descripción hecha por Ralph
et al. (1993). Se tomaron registros de precipitación y temperatura en CCC. Las listas de
aves que se incluyen en este estudio siguen el orden establecido en la Unión Americana
de Ornitología (AOU 1998).
Captura con redes de niebla. Las aves fueron capturadas con redes estándar (12 m x
3 m, 36 mm de malla), (Avinet, Box 1103, Dryden NY, 13053, USA. Telefax: 607-844-
3277. E-mail: [email protected]). Se colocaron 10-15 redes de niebla separadas por
40 m aproximadamente unas de otras, en el perímetro de un área circular, cubriendo un
área de 6 ha en cada unos de los cinco sitios. Las redes fueron operadas desde el nivel del
suelo 1-3 veces por semana y revisadas a intervalos de 40 min como máximo (más
frecuentemente en condiciones de viento y calor extremos). Las redes no fueron operadas
en condiciones de lluvia, viento o calor extremos y si las redes ya estaban abiertas en tales
condiciones fueron inmediatamente cerradas. Las redes fueron abiertas desde las 5:30-
5:40 (Central Standard Time) y cerradas a las 11:30, tratando en lo posible de mantenerlas
abiertas durante 6 hs por día. Las redes fueron cerradas en el mismo orden en que fueron
abiertas (Ralph et al. 1993).
Todas las aves capturadas, incluyendo aquellas recapturadas, fueron identificadas hasta
el nivel de especie a excepción de dos especies pertenecientes al género Empidonax. Los
individuos capturados fueron marcados con anillos de aluminio numerados (Bird Banding
Laboratory, Fish and Wildlife Service, Laurel, Maryland 20708, USA.). Luego de ser
procesadas, las aves fueron liberadas en el sitio de captura. Los individuos muy jóvenes
fueron liberados cerca de la misma red en que fueron capturados.
Conteos de búsqueda de aves. Los conteos de búsqueda de aves consisten en
desplazarse dentro del área (6 ha) de capturas con redes de niebla registrando todas las
especies observadas y/o escuchadas durante un periodo de 20 min (Ralph et al. 1993). Se
realizaron dos conteos durante cada mañana en que las redes de niebla fueron operadas.
El primer conteo se llevó a cabo una vez finalizada la apertura de las redes (alrededor de

Maglianesi
79
las 6:00 hs) y el segundo conteo se realizó a las 9:30 hs aproximadamente. No se
realizaron conteos bajo condiciones climáticas desfavorables. Las especies de aves fueron
identificadas usando binoluares (10 x 50) y una guía local de aves (Stiles y Skutch 1989).
Análisis estadístico. Se compararon ambos métodos para estimar la similitud en
términos de a) número de especies, b) porcentaje de especies migratorias, C) composición
de la comunidad de aves (índice de Jaccard), d) eficiencia de detección de especies
(curvas de acumulación de especies) y e) abundancia relativa de las especies.
Debido a que el esfuerzo de muestreo varió entre métodos, los datos fueron
estandarizados. Para ello, se utilizó la variable número de especies detectadas (capturadas
y/o observadas) durante 40 min de persona-esfuerzo, la cual será mencionada en adelante
como “tasa de detección de especies”. Para comparar el patrón entre métodos, se realizó
un análisis de correlación de Pearson.
Para determinar el grado de solapamiento en la composición de especies entre ambos
métodos, se calculó el coeficiente de Jaccard basado en la presencia o ausencia de
especies a partir de la fórmula Sj = a / a + b + c, donde S = coeficiente de similitud de
Jaccard, a = número de especies detectado por los métodos A y B (especies comunes), b =
número de especies detectado por el método B pero no por el método A, y c = número de
especies detectado por el método A pero no por el método B (Krebs 1989).
Con la finalidad de analizar la proporción de especies migratorias detectadas entre
métodos, se utilizó el porcentaje de especies migratorias detectadas durante 40 min de
persona-esfuerzo. En todas las pruebas estadísticas se consideraron solamente las
muestras donde las aves fueron capturadas durante aproximadamente 6 hs y observadas
durante dos conteos de búsqueda de aves en la misma mañana. Debido a que se realizaron
comparaciones en el número de especies residentes y migratorias, se consideraron
solamente aquellas muestras posteriores a la fecha en que se detectó la primer especie
migratoria, ya sea por medio de captura u observación en cualquiera de los cinco sitios de
estudio.
Se utilizaron una serie de Análisis de Varianza (ANDEVAs) de 2 factores para evaluar
la variación en la tasa de detección de especies entre estatus (residente-migratoria) por
método y entre sitio por método. Se utilizó un ANDEVA de 2 factores para estimar la

Maglianesi
80
variación en el porcentaje medio de especies migratorias entre sitios y métodos. Se estimó
la dependencia entre el número de especies detectado por familia y el método de
detección con una tabla de contingencia (Sokal y Rohlf 1979).
Con la finalidad de estimar la eficiencia de detección de las especies por sitio, se
construyeron curvas de acumulación de especies a partir de la ecuación de Clench S(t) =
a* t / (1+ b* t). Donde S (t) = número total de especies que predice el modelo, t =
esfuerzo de muestreo, a = tasa de incremento del número de especies al comienzo del
muestreo, b = tasa de incremento del número de especies (Soberón y Llorente 1993).. La
asíntota está dada por el cociente a/b y el tiempo tq requerido para registrar una
proporción del total de especies q que predice el modelo fue calculado a partir de q/(b*(1-
q). Las muestras fueron aleatorizadas 20 veces y los datos fueron ajustados al modelo
usando una regresión no lineal en el Programa InfoStat Versión 1.1.
La variable utilizada fue la proporción del total de días en que cada una de las especies
fueron detectadas, como un indicador de la abundancia relativa y se hará referencia en
adelante como la “proporción de días en que las especies fueron detectadas” (PDD). Para
analizar la consistencia (precisión) en detectar la abundancia relativa por cada método, los
datos fueron estandarizados a través del cálculo de un valor estándar para cada una de las
especies y así ajustar las diferencias de escalas de ambos métodos, siguiendo el criterio
utilizado por Wang y Finch (2002).
Se realizó un análisis de regresión lineal con las variables PDD/40 minutos de conteo
como la variable dependiente y PDD/40 minutos de captura como la variable
independiente. Los valores residuales estandarizados que miden la desviación relativa de
los valores observados de los valores esperados del modelo de regresión, fueron usados
como una medida de consistencia entre ambos métodos (Wang y Finch 2002). Debido a
que arbitrariamente se asignó “PDD/40 minutos de conteo” como la variable dependiente,
un valor residual negativo por debajo de la línea de regresión indicaría que por medio del
método de búsqueda de aves se registró una abundancia relativa más baja que la esperada
para una determinada especie. Mientras que un valor residual positivo por encima de la
línea indicaría que por medio de dicho método se obtuvo un estimado de la abundancia
relativa mayor que la esperada.

Maglianesi
81
El paquete estadístico STATGRAPHICS version 3.1 (Statistical Graphic Corp.) fue
usado para los diferentes análisis realizados mencionados anteriormente. Todas las
variables fueron sometidas a pruebas de normalidad (Wilk-Shapiro test), homogeneidad
de varianza (Bartlett’s test) e independencia antes de realizar las pruebas estadísticas
(Sokal y Rohlf 1979). Cuando las variables no cumplieron con los supuestos de las
pruebas paramétricas se realizaron transformaciones hasta alcanzar los supuestos.
RESULTADOS
Estimación de las especies por método. Ni el método de captura con redes de
niebla, ni el de búsqueda de aves, ni ambos juntos detectaron el total de especies de aves
registradas en la lista de aves de Tortuguero (Widdowson y Widdowson 2000). El número
total de especies en este estudio representó solamente el 45.3% de las aves incluidas en
dicha lista (Tabla1). Se registraron un total de 110 especies de aves distribuidas en 26
familias en los cinco sitios de estudio, incluyendo 81 especies (73.64%) capturadas en
redes de niebla y 90 especies (81.82) observadas en conteos de búsqueda de aves. Hubo
una especie amenazada (Ara ambigua) observada durante los conteos y el 37% del total
de especies registradas por ambos métodos fueron especies migratorias (Cuadro 1,
Apéndice A).
Por medio de los conteos de búsqueda de aves se registraron nueve especies más
que por medio de capturas considerando todos los sitios en conjunto (Cuadro 1). Se
contaron entre 1 hasta 14 especies más en los cinco sitios que en capturas. Se registraron
61 especies en común (55.45%) por ambos métodos (Cuadro 2) y el coeficiente de
similitud de Jaccard entre métodos fue de 0.55. Del total de especies detectadas, 29
(26.4%) fueron no capturadas en redes y 20 (18.2%) fueron no observadas durante los
conteos.
Curvas de acumulación de especies. Los parámetros del modelo de Clench para los
cinco sitios estudiados y para ambos métodos son mostrados en la Tabla 3. Para el
método de captura con redes, los parámetros calculados (R2, a, b y la asíntota ) para
PARQ y CAÑO fueron más altos que aquellos obtenidos para los otros tres sitios,
mostrando un incremento rápido del porcentaje de especies capturadas en las primeras

Maglianesi
82
muestras. Los parámetros a y b para los diferentes sitios fueron estadísticamente
significativos excepto para AERO. Las curvas de acumulación de especies para cada uno
de los sitios indicaron que las especies capturadas continuaron aumentando en los cinco
sitios a lo largo de las muestras (Fig. 2). La asíntota que predijo el modelo de Clench fue
más alta que los valores derivados de los datos de captura para los cinco sitios. Los sitios
que tuvieron una mayor proporción del total de especies capturadas predichas por el
modelo fueron CAÑO (70%) y PARQ (60%). Para los cinco sitios estudiados se
requieren esfuerzos de muestreo mayores para alcanzar el 90% de la asíntota.
Para el método de búsqueda de aves, en PARQ aumentó rápidamente el porcentaje
de especies observadas en las primeras muestras y hubo los parámetros del modelo de
Clench más altos. En general, las proporciones de especies representadas en los conteos
con respecto al total de especies que predice el modelo fueron más bajas que aquellas
obtenidas para el método de captura. Esta proporción fue más alta para CCC (68%),
seguido por PARQ (55%), mientras que para los otros sitios resultó ser menor del 14%
(Fig. 3). Para los cinco sitios se requiere mayor cantidad de conteos para alcanzar el 90%
de la asíntota.
Tasa de detección de especies. Las tasas medias de detección de especies residentes
capturadas y observadas tuvieron un bajo coeficiente de correlación (r=0.133, N=71,
P=0.27), (Fig. 4), mientras que para las especies migratorias hubo una mayor asociación
(r=0.499, N=71, P=0.002), (Fig. 5). En los cinco sitios hubo una tasa media de detección
de especies mayor para los conteos que para las capturas (F1,132=870, P<0.0001) y hubo
consistencia en esta variable entre métodos para la mayoría de los sitios (F4,132=9.14,
P<0.0001) (Fig. 6).
Porcentaje de especies migratorias. Con el método de captura se obtuvo un mayor
porcentaje de especies migratorias que con los conteos en todos los sitios (F1,131=5.74,
P=0.019), pero no hubo significancia estadística entre los sitios individuales (Fig. 7). En
AERO (x=3.54) y CCC (x=3.52) hubo un mayor porcentaje de especies migratorias
capturadas, mientras que para los conteos esto ocurrió para AERO (x=3.34) y PARQ
(x=3.17).

Maglianesi
83
Composición de especies por familia. Con respecto a la composición de especies
por familia, las especies de la familia Mimidae fueron solamente capturadas en redes,
mientras que las especies de otras familias como Cathartidae, Accipitridae, Psittacidae,
Cuculidae, Bucconidae, Cotingidae, Hirudinidae y Sylviidae fueron solamente observadas
durante los conteos de búsqueda de aves. Las especies de las familias Trochilidae,
Thamnophilidae y Parulidae fueron más comunes en las redes que en los conteos,
mientras que lo contrario ocurrió con las especies de las familias Columbidae,
Ramphastidae y Thraupidae, aunque no hubo diferencias significativas (X225=18.82,
P=0.806) (Fig. 8).
Consistencia entre métodos. Para las 61 especies detectadas en común por ambos
métodos, las estimaciones de abundancia relativa (proporción de días en que las especies
son detectadas: PDD) obtenidas a partir de los método de captura y búsqueda de aves
estuvieron significativamente relacionados (R2 = 0.21; F1,59 = 16.17; P = 0.002) (Fig. 9).
Los nombres y los códigos de las 61 especies que se encuentran indicados en la Fig.9 se
muestran en el Cuadro 2. El 89% de las 61 especies tuvo un valor residual menor a 0.9
desviaciones estándar al modelo, indicando que en promedio los dos métodos fueron
consistentes en las estimaciones de abundancia relativa. Solamente tres especies
(Hylophilus decurtatus, Todirostrum cinereum y Pitangus sulphuratus) tuvieron valores
residuales altos (entre 2.5 y 3.9 desviaciones estándar), indicando que por medio de los
conteos de búsqueda de aves se obtuvo una abundancia relativa mucho mayor para estas
especies que por medio de capturas.
Algunas especies como Catharus fuscescens, Dendroica petechia, Myrmeciza exsul,
Turdus grayi y Phaethornis superciliosus tuvieron unos valores residuales muy cerca de la
línea de regresión, indicando una abundancia relativa similar obtenida por ambos
métodos. (Fig. 9). Mientras que otras especies como Glaucis aenea, Thalurania
colombica, Anthracothorax prevostii, Sporophila americana y Pipra mentalis tuvieron una
abundancia relativa estimada por medio de capturas mayor que la estimada por los
conteos. Esto ocurrió inversamente para Myozetetes similis, Megarynchus pitangua,
Thryothorus nigricapillus, Euphonia gouldi y Manacus candei.

Maglianesi
84
DISCUSIÓN
Existen varios factores que influyen la detección de las especies y su abundancia
relativa. Me referiré a aquellos que pueden explicar las diferencias en los resultados
obtenidos en este estudio. Estos factores son: tamaño, similitud morfológica entre
especies, movilidad, atributos del plumaje y uso del espacio vertical de las especies.
Presencia vs. ausencia de especies. Los resultados obtenidos a partir del método de
captura con redes o búsqueda de aves no describen el total de especies que componen la
comunidad de aves. Los patrones en composición y abundancia relativa variaron con el
método empleado, lo cual indica sesgos asociados a dichos métodos. El método de
captura falló en detectar más especies que el método de búsqueda de aves y este último
método resultó ser más eficiente en la detección de especies por poseer una tasa de
acumulación de nuevas especies más alta.
Por más de dos décadas, el análisis de los datos obtenidos de capturas de aves en redes
de niebla ha desempeñado un rol importante en estudios ecológicos (Remsen y Good
1996). La ventaja más evidente del método de captura para obtener información sobre la
composición de las comunidades de aves y la abundancia relativa de las especies es que
se evitan los sesgos propios de las técnicas de conteos relacionados con la habilidad
visual y auditiva del observador (Karr 1981, Karr 1990). Las especies de tamaño reducido
como miembros de la familia Trochilidae, Todirostrum nigriceps y Rynchocyclus
brevirostris, como así también aquellas especies con una morfología externa muy similar
como los miembros del género Catharus fueron detectadas solamente por medio de
capturas. Esto se debe probablemente a una mayor dificultad para la identificación
durante los conteos para algunos observadores.
Sin embargo, los resultados obtenidos a partir de las capturas no representan
cuantitativamente la composición de especies en una comunidad de aves, debido a que la
eficiencia en las capturas varían notablemente entre especies (Jenni et al. 1996). Los
resultados en este estudio coinciden con aquellos obtenidos por Lynch (1989), los cuales
evidencian que las redes no detectaron aquellas especies de gran tamaño ni aquellas que

Maglianesi
85
utilizan el dosel o subdosel del bosque. Por ejemplo, especies como Coragyps atratus,
Cathartes aura, Buteogallus anthracinus, Piaya cayana y Psaracolius montezuma
estuvieron representadas únicamente en los conteos, debido a que las redes de niebla
estándar no resultan eficientes para capturar estas especies de gran tamaño.
Especies como Hirudindo rustica, Ramphastos sp, Psittacidae, Polioptila plumbea, y
Tangara larvata también estuvieron representadas únicamente en los conteos debido a que
estas especies pasan mayor tiempo a ciertas alturas y por medio de las redes de niebla es
posible detectar la porción de la avifauna cuya actividad se desarrolla por debajo de los 2-
3 m de altura sobre el suelo. En contraste, Dumetella carolinensis, Oryzoborus funereus, y
Saltatos maximus, son especies que utilizan un estrato bajo del bosque, las cuales
detectadas únicamente por medio de capturas. Además, las redes fallan en detectar
aquellas especies como Monasa morphoeus sugiriendo que la probabilidad de captura
disminuye cuando las especies presentan menor movilidad.
Abundancia Relativa. El número de aves capturadas en redes durante muestreos
estandarizados es considerado frecuentemente como una medida superior de abundancia
relativa, particularmente debido a que permite comparaciones entre estudios que incluyen
diferentes investigadores (Karr et al. 1990). Más aún, la interpretación de los números
relativos de aves capturadas está dada como un reflejo de la abundancia relativa
solamente (Remsen y Parker, III 1983). Sin embargo, el número de aves capturadas en
redes de niebla durante un periodo de muestreo está determinado por otros aspectos,
además de la abundancia relativa (Remsen y Good 1996). El número de aves atrapadas
depende de los patrones espaciales y temporales específicos de cada especie (Jenni et al.
1996). Por lo tanto, las diferencias en movilidad y sistemas sociales de las especies que
afectan a los patrones de actividad pueden a su vez, influenciar fuertemente la abundancia
relativa de las especies.
Las especies que se mueven a través de largas distancias entre sitios de alimentación
en el sotobosque deberían estar representadas desproporcionadamente en las muestras de
capturas. Así, las especies que pertenecen a ciertos géneros como Pipra, Manacus,
Mionectes y Phaethornis están citadas generalmente como las especies más “abundantes”
en las muestras tomadas en comunidades tropicales. Aunque estas especies ciertamente se

Maglianesi
86
entran frecuentemente entre las más comunes en los bosques tropicales, algunos estudios
sugieren que los datos publicados sobre la abundancia relativa de estas especies están
drásticamente sobreestimados (Remsen y Parker, III 1983).
Los resultados en este estudio coinciden con los obtenidos por Remsen y Parker, III
(1983) pero sólo en parte. Mionectes oleagineus solamente estuvo representado en
muestras de capturas de acuerdo con otros estudios que comparan los métodos de captura
con puntos de conteo (Blake y Loiselle 2002) y Pipra mentalis tuvo una mayor
abundancia relativa con el método de captura. Sin embargo, a partir de los conteos de
búsqueda de aves se obtuvo una mayor abundancia relativa para Phaetornis y Manacus en
oposición con Remsen y Parker, III (1983).
En este estudio también se encontró que ciertas especies como Glaucis aenea y
Sporophila americana tuvieron una mayor abundancia relativa a partir de los datos de
capturas, probablemente porque estas especies utilizan un estrato bajo del bosque. En
contraste, Hylophilus decurtatus y Todirostrum cinereum tuvieron una abundancia
relativa desproporcionadamente mayor a partir de los conteos de búsqueda de aves, lo
cual puede deberse a que dichas especies se utilizan el dosel del bosque y son muy activas
en sus vocalizaciones. Los resultados en el presente estudio mostraron que muchas de las
especies bien detectadas por una técnica fueron pobremente detectadas por la otra,
conforme otros estudios (Whitman et al. 1997). Esto sugiere que algunos factores que
influyen en la determinación de la composición de la comunidad de aves y la abundancia
relativa de las especies por los métodos de captura y búsqueda de aves podrían eliminarse
unos con otros tendiendo a ser complementarios.
Por ejemplo, las especies silenciosas e inconspicuas con baja detectabilidad por el
método de conteos, pero con largas distancias de vuelo serían más capturadas. Mientras
que aquellas especies menos móviles o que se desplazan en un estrato alto del bosque y
que están poco representadas en las capturas, pero son vocales o de gran tamaño sería
bien detectadas durante los conteos, compensando en parte la influencia de diferencias en
la eficiencia relacionada con el tamaño y los patrones de actividad de las especies.
Conclusiones y recomendaciones. El monitoreo efectivo de las poblaciones de aves
es particularmente importante para la identificación de los problemas y necesidades de

Maglianesi
87
conservación de las especies (Baillie 1990). Un prerrequisito de los estudios que utilizan
el método de captura con redes de niebla para muestrear aves es estandarizar tanto como
sea posible la toma de datos a través del uso de un número constante de redes, del mismo
tipo y en las mismas ubicaciones por iguales periodos de tiempo, usando además un
sistema regular para revisar las redes y horas constantes para abrir y cerrar las redes
(Jenni et al. 1996).
Los estudios que utilizan principalmente datos obtenidos a partir de capturas para
analizar la estructura de la comunidad deberían tener bien presente los sesgos asociados a
los datos de capturas. En relación al uso directo de los datos de captura como una medida
de abundancia relativa, la interpretación de los resultados requiere de cuidado a la hora de
aplicar dichos resultados para la toma de decisiones de manejo.
Los investigadores que se encuentran planificando muestreos de aves debería
reconocer los sesgos y seleccionar el método que mejor muestree el segmento de la
comunidad que pretenden describir. Si se va a describir la comunidad entera o gremios de
aves, probablemente se requiera del uso de varios métodos de muestreo, como se hizo en
este estudio y en Pagen et al. (2000).
A pesar del hecho de que el método de búsqueda de aves por sí solo debería ser más
eficiente en cuanto al esfuerzo realizado y menos costoso, debería ser utilizado en
combinación con otros métodos para contar con una descripción más completa de la
comunidad de aves en las áreas tropicales. La composición exacta de la comunidad de
aves y la abundancia relativa puede ser solamente estimada de una manera cuasi-
cuantitativa a través de los métodos visuales-auditivos por observadores experimentados
en combinación con el método de captura con redes de niebla.
AGRADECIMIENTOS
En primer lugar quisiera agradecer a los miembros de mi Comité de Tesis: Michael
McCoy, Gilbert Barrantes, C. John Ralph, y Jaime Rau, quienes me ofrecieron
constantemente sus comentarios y recomendaciones a lo largo de este estudio. Realmente
me siento afortunada de haber trabajado bajo la guía de estos renombrados especialistas.
Me encuentro muy agradecida con el Programa Regional en Manejo de Vida Silvestre
para Mesoamérica y el Caribe (PRMVS) por permitirme llevar a cabo un grado de

Maglianesi
88
maestría a partir del cual surgió este artículo de tesis como un requisito para optar al título
de Master en Conservación y Manejo de Vida Silvestre. También agradezco al
Laboratorio de TeleSig del PRMVS por permitirme el uso de equipo y programas de
computo especializados, en particular le agradezco a Henry Chávez por su contribución
en la elaboración del mapa sobre el área de estudio.
Agradezco además a la Corporación para la Conservación del Caribe por permitirme el
acceso a sus instalaciones para llevar a cabo la fase de campo en su propiedad y a la
organización Idea Wild por proveer equipo para este estudio.
Doy mi mas sincero agradecimiento a los voluntarios del Programa Integrado de
Monitoreo de Aves de Tortuguero, quienes contribuyeron en la toma de datos de campo:
Dai Shizuka, Devin Taylor, Gabriel David, Lucie Parker, Dan van den Broek, Ana Clarke
y Ernest Clarke. Ellos fueron excelentes compañeros y entusiastas asistentes de campo.
Doy un especial agradecimiento a Robert Frey, quien es un invaluable compañero en el
campo y me ofreció sus consejos, compartiendo su gran experiencia y pasión por las aves.
Este estudio fue posible gracias al Programa Integrado de Monitoreo de Aves de
Tortuguero (Redwood Sciences Laboratory, U.S. Forest Service), el cual facilitó el equipo
y los voluntarios para el trabajo de campo y a los observatorios de aves Reyes Point y
Klamath, quien facilitó los fondos para los aspectos logísticos de esta investigación.
LITERATURA CITADA
AOU. American Ornithologist’ Union. 1998. The check-list of North American Birds.
Baillie, S. R. 1990. Integrated population monitoring of breeding birds in Britain and
Ireland. Ibis 132: 151-166.
Beehler, B. M., J. B. Senge, C. Filard y K. Merg. 1995. Documenting the lowland
rainforest avifauna in Papua New Guinea –effect of patchy distributions, survey effort,
and methodology. Emu 95: 149-161.
Blake, J. G. y B. A. Loiselle. 1989. Habitat use by neotropical migrants at La Selva
Biological Station and Braulio Carrillo National Park, Costa Rica. In: Ecology and
conservation of neotropical migratory landbirds (J. M. Hagan y D. W. Johnston, eds.),
pp. 257-272. Smithsonian Institute Press. Washington DC.
Blake, J. G. y B. A. Loiselle 2000. Diversity of birds along an elevational gradient in the

Maglianesi
89
Cordillera Central, Costa Rica. Auk 117: 663-686.
Blake, J. G. y B. A. Loiselle. 2001. Bird assemblages in second-growth and old-growth
forests, Costa Rica: perspectives from mist nets and point counts. Auk 118: 304-326.
Blake, J. G. y B. A. Loiselle. 2002. Manakins (Pipridae) in second-growth and old-growth
forests: patterns of habitat use, movement, and survival. Auk 119: 132-148.
Boucher, D. H., M Hansen, S. Risch y J. H. Vandermeer. 1983. In: Costa Rican Natural
History (D. H. Janzen, ed.), pp. 66-117. The University of Chicago Press Chicago and
London.
Coen, E. 1983. Climate. In: Costa Rican Natural History (D. H. Janzen, ed.), pp. 35-46.
The University of Chicago Press, Chicago and London.
Dieni, J. S. y S. L. Jones. 2002. A field test of the area search method for measuring
breeding bird populations. Journal of Field Ornithology 73: 253-257.
Gram, W. K. y J. Faaborg.1997. The distribution of neotropical migrant birds wintering in
the Cielo Biosphere Reserve Tamaulipas, Mexico. Condor 99: 658-670.
Herrera, H. 1998. Características del hábitat y de su uso por dos especies de saltarines
(Pipridae) en un humedal del bosque lluvioso neotropical. Tesis de Maestría.
Universidad Nacional. Heredia, Costa Rica.
Hewish, M. J. y R. H. Loyn. 1989. Popularity and effectiveness of four survey methods
for monitoring populations of Australian birds. R.A.O.U. Report Nº 55. Royal
Australasian Ornithologist Union, Moonee Ponds, Australia.
Holdridge, L. R 1969. Life zone ecology. Rev. Ed. San José, Costa Rica. Tropical Science
Center. San José, Costa Rica.
Janzen, D. H. 1983. Costa Rican Natural History. The University of Chicago Press,
Chicago and London.
Jenni, L., M. Leuenberger y F. Rampazzi. 1996. Capture efficiency of mist nets with
comments on their role in the assessment of passerine habitat use. Journal of Field
Ornithology: 67: 263-274.
Karr, J. R. 1981. Surveying birds with mist nets. In: Estimating numbers of terrestrial
birds (C. J. Ralph y J. M. Scott, eds.), pp. 62-67. Studies in Avian Biology 6. A
publication of the Cooper Ornithological Society.
Karr, J. R. 1990. The avifauna of Barro Colorado Island and the Pipeline Road, Panama.

Maglianesi
90
In: Four Neotropical rainforests (A. H. Gentry, ed.), pp. 183-198. Yale University.
Press, New Haven, C.T.
Karr, J. R., S. Robinson, J. G. Blake y R. O. Bierregaard, JR. 1990. Birds of four
Neotropical forests. In: Four Neotropical rainforests (A. H. Gentry, ed.), pp 237-272.
Yale University. Press, New Haven, C.T.
Krebs, C. J. 1989. Ecological Methodology. University of British Columbia. Harper
Collins Publishers.
Lynch, J. F. 1989. Distribution of overwintering nearctic migrants in the Yucatan
Peninsula. I: general patterns of occurrence. Condor 91: 515-544.
Nur, N. y G. R, Geupel. 1993. Evaluating mist-netting, nest-searching, and other methods
of monitoring demographic processes in landbird populations. In: Status and
management of Neotropical migratory birds (D. M. Finch y P. W. Stangel, eds.), pp.
237-244. Gen. Tech. Rep. RM-229. USDA Forest Service. USA.
Oro, G. M. 1996. Ficha Técnica RAMSAR. Manuscrito. Proyecto de consolidación del
Área de conservación y desarrollo sostenible llanuras de Tortuguero (PACTO).
Pagen, R. W., F. R. Thompson y D. E. Burhans. 2000. Breeding and post-breeding habitat
use by forest migrant songbirds in the Missouri ozarks. Condor 102: 738-747.
Pagen, R. W., F. R. Thompson y D. E. Burhans. 2002. A comparison of point-count and
mist-net detections of songbirds by habitat and time-of-season. Journal of Field
Ornithology 73: 53-59.
Ralph, C. J., G. R. Geupel, P. Pyle, T. Martin, D. F. DeSante y B. Milá. 1993. Handbook
of field methods for monitoring landbirds. Pacific Southwest Research Station.
Albany, California.
Ralph, C. J. y S. G. Fancy. 1995. Demography and movements of Apapane and Iiwi in
Hawaii. Condor 97: 729-742.
Ralph, C. J., M. Widdowson, B. Widdowson, B. O’Donnell y R. I. Frey. 2002.
Tortuguero bird monitoring station protocol. The Tortuguero Integrated Bird
Monitoring Program. Redwood Sciences Laboratory. U.S. Forest Service. Arcata,
California.
Rappole, J. H, W. J. McShea y J. Vega-Rivera. 1993. Evaluation of two survey methods
in upland avian breeding communities. Journal of Field Ornithology 64: 55-70.

Maglianesi
91
Rappole, J. H., K Winker y G. V. N. Powell. 1998. Migratory bird habitat use in southern
Mexico: mist nets versus point counts. Journal of Field Ornithology 69: 635-643.
Remsen, J. V. y T. A. Parker. 1983. Contribution of river-created habitats to bird species
richness in Amazonia. Biotropica 15: 223-231.
Remsen, J. V., Jr y D. A. Good. 1996. Misuse of data from mist-net captures to assess
relative abundance in bird populations. Auk 113: 381-398.
Robinson, W. D., J. D. Brown y S. K. Robinson. 2000. Forest bird community structure
in central Panama: influence of spatial scale and biogeography. Ecological
Monographs 70: 209-235.
Slater, P. J. 1994. Factors affecting the efficiency of the area search method of censusing
birds in open forests and woodlands. Emu 94: 9-16.
Soberón, J. M. and J. B. Llorente. 1993. The use of species accumulation functions for the
prediction of species richness. Conservation Biology 7:480-488.
Sokal, R. R. y F. J. Rohlf. 1979. Biometría. Principios y métodos estadísticos en la
investigación biológica. Primera edición española H. Blume Ediciones. Madrid,
España.
Stiles, F. G. 1983. Birds. In: Costa Rican Natural History (D. H. Janzen, ed.), pp. 502-
530. The University of Chicago Press, Chicago and London.
Stiles, F. G. y A. F. Skutch 1989. I guide to the birds of Costa Rica. Comstock Publishing
Associates. Cornell University Press. Ithaca, NY, USA.
Terborgh, J., S. K. Robinson, T. A. Parker III, C. A. Munn y N. Pierpont. 1990. Structure
and organization of an Amazonian forest bird community. Ecological Monographs 60:
213-238.
Wang, Y. y D. M. Finch. 2002. Consistency of mist netting and point counts in assessing
landbird species richness and relative abundance during migration. Condor 104: 59-72.
Whitman, A. A., J. M. Hagan III y N. V. L. Brokaw. 1997. A comparison of two bird
survey techniques used in a subtropical forest. Condor 99: 955-965.
Widdowson, W. P. y M. J. Widdowson. 2000. Checklist to the birds of Tortuguero. The
Tortuguero Integrated Bird Monitoring Program. Redwood Sciences Laboratory. U.S.
Forest Service. Arcata, California.

CUADRO 1. Número de especies capturadas en redes de niebla y observadas en conteos de búsqueda de aves en cinco sitios de estudio en el Caribe norte de Costa Rica.
Captura con Redes de Niebla Búsqueda de Aves
Sitioa
Días Muestreados
Esfuerzo (horas)
Residentes Migrantes Total % Migrantes
Esfuerzo (horas)
Residentes Migrantes Total % Migrantes
Todos los sitios 71 400.9 51 30 81 37.04 46.86 63 27 90 30.00
CCC 28 153.8 30 25 55 45.45 18.48 36 20 56 35.71
PARQ 7 38.8 18 12 30 40.00 4.62 26 5 31 16.13
AERO 10 58.2 19 17 36 47.22 6.6 25 17 42 40.48
TORT 14 79.6 18 11 29 37.93 9.24 31 5 36 13.89
CAÑO 12 70.5 22 11 33 33.33 7.92 34 13 47 27.66
a CCC = Corporación para la Conservación del Caribe; PARQ = Parque Nacional Tortuguero; AERO: Aeropuerto Barra de Tortuguero; TORT = Tortuga Lodge; CAÑO = Estación Biológica Caño Palma.
92

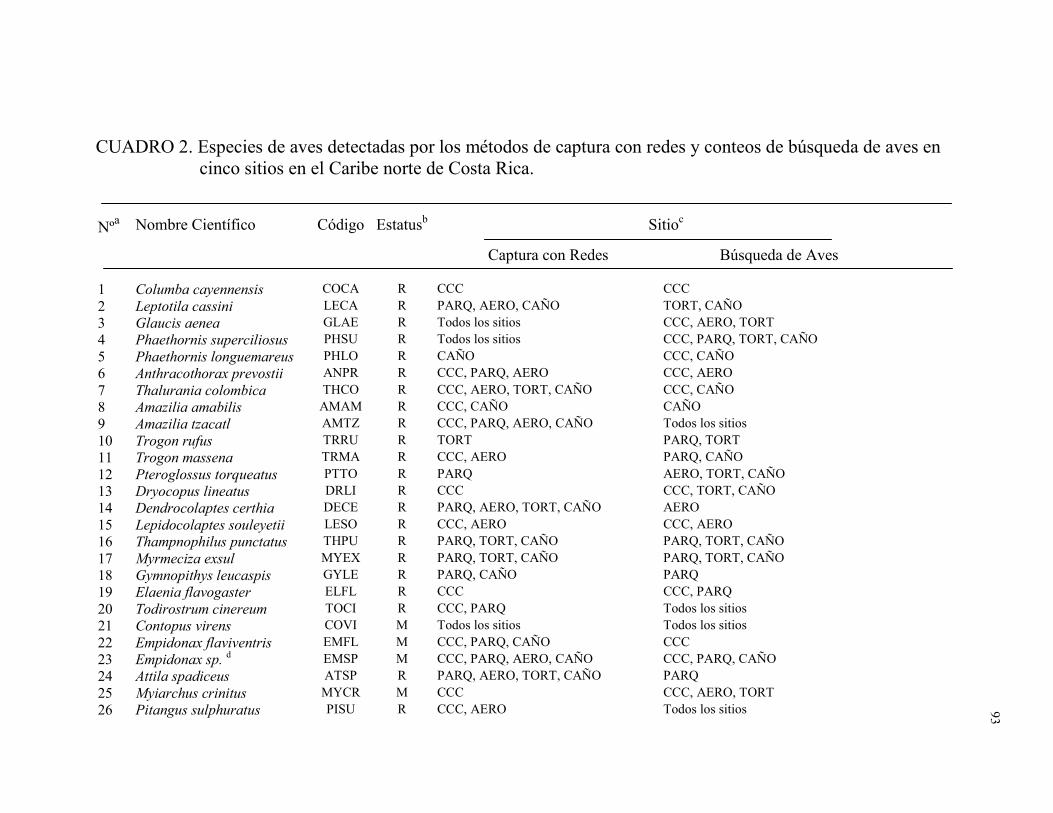
CUADRO 2. Especies de aves detectadas por los métodos de captura con redes y conteos de búsqueda de aves en cinco sitios en el Caribe norte de Costa Rica.
Nºa Nombre Científico Código Estatusb Sitioc
Captura con Redes Búsqueda de Aves
1 Columba cayennensis COCA R CCC CCC
2 Leptotila cassini LECA R PARQ, AERO, CAÑO TORT, CAÑO
3 Glaucis aenea GLAE R Todos los sitios CCC, AERO, TORT
4 Phaethornis superciliosus PHSU R Todos los sitios CCC, PARQ, TORT, CAÑO
5 Phaethornis longuemareus PHLO R CAÑO CCC, CAÑO
6 Anthracothorax prevostii ANPR R CCC, PARQ, AERO CCC, AERO
7 Thalurania colombica THCO R CCC, AERO, TORT, CAÑO CCC, CAÑO
8 Amazilia amabilis AMAM R CCC, CAÑO CAÑO
9 Amazilia tzacatl AMTZ R CCC, PARQ, AERO, CAÑO Todos los sitios
10 Trogon rufus TRRU R TORT PARQ, TORT
11 Trogon massena TRMA R CCC, AERO PARQ, CAÑO
12 Pteroglossus torqueatus PTTO R PARQ AERO, TORT, CAÑO
13 Dryocopus lineatus DRLI R CCC CCC, TORT, CAÑO
14 Dendrocolaptes certhia DECE R PARQ, AERO, TORT, CAÑO AERO
15 Lepidocolaptes souleyetii LESO R CCC, AERO CCC, AERO
16 Thampnophilus punctatus THPU R PARQ, TORT, CAÑO PARQ, TORT, CAÑO
17 Myrmeciza exsul MYEX R PARQ, TORT, CAÑO PARQ, TORT, CAÑO
18 Gymnopithys leucaspis GYLE R PARQ, CAÑO PARQ
19 Elaenia flavogaster ELFL R CCC CCC, PARQ
20 Todirostrum cinereum TOCI R CCC, PARQ Todos los sitios
21 Contopus virens COVI M Todos los sitios Todos los sitios
22 Empidonax flaviventris EMFL M CCC, PARQ, CAÑO CCC
23 Empidonax sp. d EMSP M CCC, PARQ, AERO, CAÑO CCC, PARQ, CAÑO
24 Attila spadiceus ATSP R PARQ, AERO, TORT, CAÑO PARQ
25 Myiarchus crinitus MYCR M CCC CCC, AERO, TORT
26 Pitangus sulphuratus PISU R CCC, AERO Todos los sitios
93

CUADRO 2. (Continuación)
Nºa Nombre Científico Código Estatusb Sitioc
Captura con Redes Búsqueda de Aves
27 Megarynchus pitangua MEPI R CCC CCC, PARQ, AERO, TORT
28 Myiozetetes similis MYSI R CCC Todos los sitios
29 Tyrannus melancholicus TYME R AERO CCC, PARQ, AERO
30 Manacus candei MACA R Todos los sitios Todos los sitios
31 Pipra mentalis PIME R PARQ, AERO, TORT, CAÑO CCC, PARQ, CAÑO
32 Vireo olivaceus VIOL M CCC, PARQ, AERO CCC, AERO, CAÑO
33 Vireo flavoviridis VIFL M CCC CCC
34 Hylophilus decurtatus HYDE R CCC Todos los sitios
35 Thryothorus nigricapillus THNI R AERO CCC, AERO, TORT, CAÑO
36 Thryothorus thoracicus THTH R TORT TORT
37 Troglodytes aedon TRAE R CCC, AERO CCC, PARQ
38 Henicorhina leucostica HELE R TORT, CAÑO TORT, CAÑO
39 Catharus fuscescens CAFU M CCC, PARQ, AERO, TORT CCC, TORT
40 Catharus ustulatus CAUS M Todos los sitios PARQ, AERO, TORT, CAÑO
41 Hylocichla mustelina HYMU M PARQ, TORT CAÑO
42 Turdus grayi TUGR R CCC, PARQ CCC, PARQ, TORT, CAÑO
43 Vermivora peregrina VEPE M CCC CCC, AERO
44 Dendroica petechia DPET M CCC, AERO, CAÑO CCC, AERO, CAÑO
45 Dendroica pensylvanica DEPE M AERO Todos los sitios
46 Dendroica magnolia DEMA M CCC, PARQ, TORT AERO
47 Dendroica castanea DECA M AERO, TORT CCC, AERO
48 Mniotilta varia MNVA M CAÑO CCC
49 Setophaga ruticilla SERU M CCC, AERO CCC, AERO
50 Protonotaria citrea PRCI M Todos los sitios CCC, AERO, CAÑO
51 Seiurus noveboracensis SENO M Todos los sitios AERO, CAÑO
52 Oporornis formosus OPFO M CCC, AERO, CAÑO CAÑO
53 Habia fuscicauda HAFU R TORT TORT
54 Piranga rubra PIRU M CCC, AERO, TORT CCC, AERO 94

CUADRO 2. (Continuación)
Nºa Nombre Científico Código Estatusb Sitio de Detecciónc
Captura con Redes Búsqueda de Aves
55 Thraupis palmarum TAPA R CCC, AERO CCC, AERO
56 Euphonia gouldi EUGO R TORT, CAÑO PARQ, TORT, CAÑO
57 Sporophila americana SPAU R Todos los sitios CCC, PARQ, AERO, CAÑO
58 Arremonops conirostris ARCO R CCC CCC, PARQ, CAÑO
59 Quiscalus mexicanus QUME R CCC CCC
60 Icterus dominicensis ICDO R CCC CCC
61 Icterus galbula ICGA M CCC PARQ
a Los códigos de las especies corresponden a los de la Figura 9.
b R = Residentes, M = Migratorias.
c CCC = Corporación para la Conservación del Caribe; PARQ = Parque Nacional Tortuguero; AERO: Aeropuerto Barra de Tortuguero; TORT = Tortuga Lodge; CAÑO = Estación Biológica Caño Palma.
d Incluye a Empidonax alnorum y E. trailli
95


CUADRO 3. Parámetros de regresión del modelo de Clench para cinco sitios de estudio en el Caribe norte de Costa Rica.
Captura con Redes Conteos de Búsqueda de Aves
Sitio a
R2
a b
b b
Asíntota
(a/b)
Porcentaje de Especies Capturadas
Esfuerzo para capturar el 90% de las Especies (h-r) c
R2
a
b
Asíntota
(a/b)
Porcentaje de Especies Observadas
Esfuerzo para contar el 90% de las
Especies (Conteos)
CCC 0.96 0.04 2.8-4 143 38 32,142 0.96 5.72 0.07 82 68 129 PARQ 0.96 0.13 2.6-3 50 60 3,461 0.96 10.58 0.19 56 55 47 AERO 0.94 0.09 5.6-4 d 161 22 16,071 0.94 4.31 7.4-4 e 5824 1 12,162 TORT 0.96 0.06 8.5-4 70 41 10,588 0.94 2.56 2.8-3 f 914 4 3,214 CAÑO 0.98 0.15 3.2-3 47 70 2,812 0.94 3.71 -0.01-3 g -371 13 900
a CCC = Corporación para la Conservación del Caribe; PARQ = Parque Nacional Tortuguero; AERO: Aeropuerto Barra de Tortuguero; TORT = Tortuga Lodge; CAÑO = Estación Biológica Caño Palma.
b Los parámetros (a y b) son estadísticamente significativos (P<0.05) si no se indica lo contrario.
c h-r = horas-red.
d P=0.07
e P=0.98
f P=0.69
g P=0.36
96

LISTA DE TÍTULOS DE FIGURAS
Figura
1. Área de estudio con la ubicación de los sitios muestreados.
2. Curva de acumulación de especies para especies de aves capturadas basada en el
modelo de Clench.
3. Curva de acumulación de especies para especies de aves observadas basada en el
modelo de Clench.
4. Asociación de la tasa de detección de especies de aves residentes entre métodos, en
cinco sitios del Caribe norte de Costa Rica.
5. Asociación de las tasas medias de detección de especies de aves migratorias entre
métodos, en cinco sitios del Caribe norte de Costa Rica.
6. Tasa media de detección de especies en cinco sitios por los métodos de captura con
redes y conteos de búsqueda de aves (intervalos de confianza son +/- intervalos de
Tukey).
7. Porcentaje medio de especies migratorias detectadas por medio de captura con redes y
conteos de búsqueda de aves en cinco sitios del Caribe norte de Costa Rica (intervalos
de confianza son +/- intervalos de Tukey).
8. Número total de especies por familia detectadas por los métodos de captura con redes y
conteos de búsqueda de aves.
9. Relación entre la proporción de días en que las especies fueron detectadas (PDD) por los
métodos de captura con redes y búsqueda de aves. Los datos fueron estandarizados y
luego transformados logarítmicamente (base 10). Ver la Tabla 2 para los códigos de
las especies.
97

’
Fig. 2; 98
Tortuguero
Cerro Tortuguero Boca de las lagunas de Tortuguero

.
0
5
10
15
20
25
30
35
40
45
50
55
1 3 5 7 9 11 13 15 17 19 21 23 25 27
Hora-Red
Núm
ero Acumulado de Especies Capturadas
CCC
PARQ
AERO
TORT
CAÑO
0 150 300 450 600 750 1000 1150 1300 1450 1600 1750 1900 2050 2200
Fig. 2; 99

Fig. 3; 100
0
5
10
15
20
25
30
35
40
45
50
55
1 3 5 7 9 11 13 15 17 19 21 23 25 27
Conteos de Búsqueda de Área/día (40 min esfuerzo-persona)
Núm
ero Acumulado de Especies Observadas
CCC
PARQ
AERO
TORT
CAÑO
Conteos de Búsqueda de Aves/día (40 min esfuerzo-persona)

r = 0.133 n = 71 P = 0.27
Fig. 4; 101
0.4
0.6
0.8
1.0
1.2
1.4
-0.7 -0.6 -0.5 -0.4 -0.3 -0.2 -0.1 0.0 0.1 0.2
Log(Tasa Media de Detección de Especies Residentes en Conteos)
Log
(Tasa Media de Detección
de
Especies Residentes Capturadas)
r = 0.133 n = 71 P = 0.27

0.0
0.2
0.4
0.6
0.8
1.0
1.2
-1.0 -0.9 -0.8 -0.7 -0.6 -0.5 -0.4 -0.3 -0.2 -0.1 0.0 0.1 0.2
Log(Tasa Media de Detección de Especies Migratorias en Conteos)
Log(Tasa Media de Detección de
Especies Migratorias Capturadas)
r = 0.499 n = 71 P = 0.002
Fig. 5; 102

Método: F=870; gl=1,132; P<0.0001 Sitio: F= 9.14; gl=4,132; P<0.0001 Interacción: F=1.20; gl=4,132; P=0.312
Fig. 6; 103
Conteos de Búsqueda de Aves
Captura con Redes

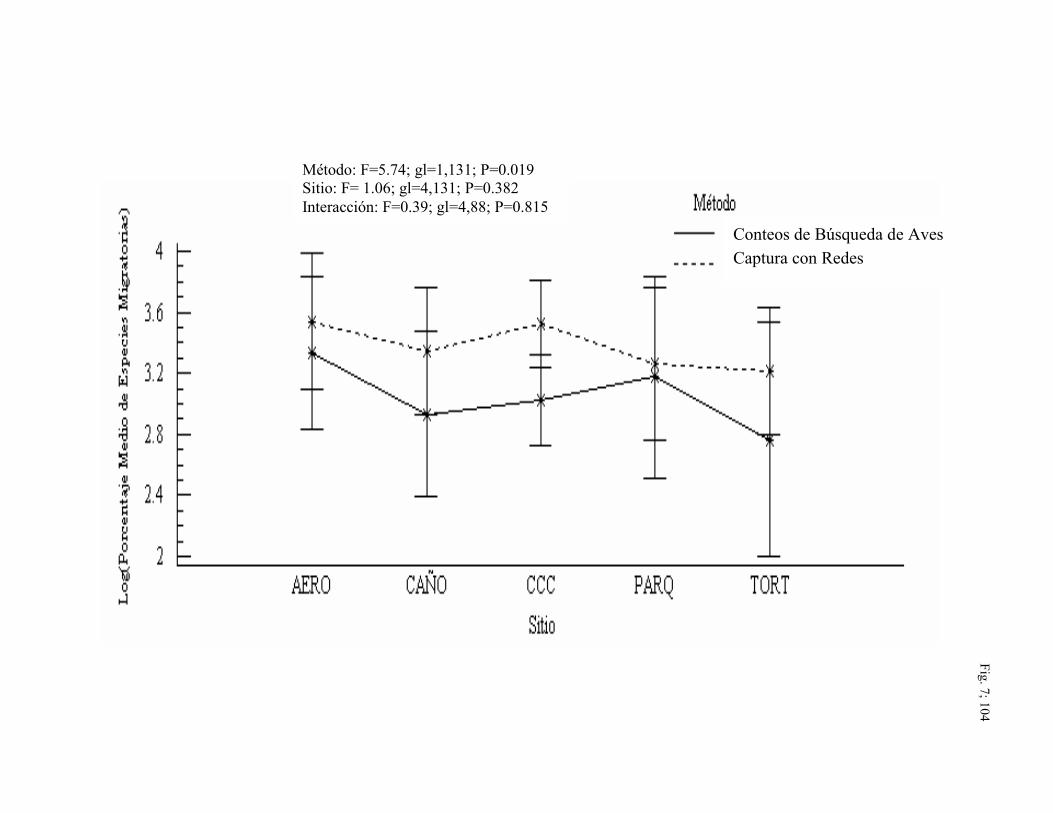
Método: F=5.74; gl=1,131; P=0.019 Sitio: F= 1.06; gl=4,131; P=0.382 Interacción: F=0.39; gl=4,88; P=0.815
Fig. 7; 104
Conteos de Búsqueda de Aves
Captura con Redes

0
2
4
6
8
10
12
14
16
18
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
Familia
Núm
ero de Especies
Captura con Redes
Conteos de Búsqueda de Área
12. Dendrocolaptidae13. Thampnophilidae14. Tyrannidae15. Cotingidae16. Pipridae17. Vireonidae18. Hirudinidae19. Troglodytidae20. Sylviidae21. Turdidae22. Mimidae
1. Cathartidae2. Accipitridae3. Columbidae4. Psittacidae5. Cuculidae6. Trochilidae7. Trogonidae8. Alcedinidae9. Bucconidae10. Ramphastidae11. Picidae
23. Parulidae24. Thraupidae25. Emberizidae26. Icteridae
X2=18.82 gl=25 P=0.806
Fig. 8; 105
Conteos de Búsqueda de Aves
Captura con Redes

AMTZ
MACA
SPAU GLAE
HYDE
TOCI
PISU
MYSI
MEPI
THNI
COCA EUGO
TUGR
VIOL
COVI
EMSP
PRCI
SENO
CAUS THCO
OPFO
MYEX, TAPU
y = a + bx y = -0.00065 +/- 0.478x R2 = 0.21 F1,59 = 16.17 P = 0.002
DEPE, THTH
PHSU
EMFL
Fig. 9; 106
LESO
CAFU
ANPR
PIME
PTTO
SERU
TRAE
DPET
DRLI, VEPE
TRRU PIRU HELE
TYME, QUME
VIFL, ICDO, ICGA
MNVA, ARCO
ELFL, THPA
PHLO
MICR
HYMU HAFU THPA
DECA
DECE
AMAM
GYLE, DEMA
ATSP
LECA
APÉNDICE A. Nombres científicos de todas las especies capturadas en redes de niebla y/o observadas durante conteos de búsqueda de aves, estatus, sitios de detección y tipo de hábitat.
Nº Nombre Científico Estatus a Método Hábitat b Sitio c
1
CATHARTIDAE Coragyps atratus
R
conteos
I, NI
CCC, AERO, TORT, CAÑO
2 Cathartes aura R conteos I, NI CCC, CAÑO
3
ACCIPITRIDAE ButeogTodosus anthracinus
R
conteos
I
CCC
4
COLUMBIDAE Columba flavirostris
R
conteos
I
CCC
5 Columba cayennensis R ambos I CCC
6 Leptotila cassini R ambos I, NI PARQ, AERO,TORT, CAÑO
7
PSITTACIDAE Aratinga nana
R
conteos
I
AERO
8 Ara ambigua R conteos NI TORT
9 Pionus senilis R conteos I, NI AERO, TORT
10 Amazona autumnalis R conteos I AERO
11 Amazona farinosa R conteos NI TORT
12
CUCULIDAE Piaya cayana
R
conteos
NI
CAÑO
13
TROCHILIDAE Glaucis aenea
R
ambos
I, NI
Todos los sitios
14 Phaethornis superciliosus R ambos I, NI Todos los sitios
15 Phaethornis longuemareus R ambos I, NI CCC, CAÑO
16 Anthracothorax prevostii R ambos I, NI CCC, PARQ, AERO
17 Thalurania colombica R ambos I, NI CCC, AERO, TORT, CAÑO
18 Hylocharis eliciae R captura I CCC
19 Amazilia amabilis R ambos I, NI CCC, CAÑO 107

APÉNDICE A. (Continuación)
Nº Nombre Científico Estatus a Método Hábitat b Sitio c
20 Amazilia tzacatl R ambos I, NI Todos los sitios
21 Heliothryx barroti R captura NI CAÑO
22
TROGONIDAE Trogon rufus
R
ambos
NI
PARQ, TORT
23 Trogon massena R ambos I, NI CCC, PARQ, AERO, CAÑO
24
ALCEDINIDAE Ceryle torquata
R
conteos
I
AERO
25 Chloroceryle americana R conteos I AERO
26 Chloroceryle aenea R captura NI TORT, CAÑO
27
BUCCONIDAE Monasa morphoeus
R
conteos
NI
PARQ, TORT
28
RAMPHASTIDAE Pteroglossus torquatus
R
ambos
I, NI
PARQ, AERO, TORT, CAÑO
29 Ramphastos sulfuratus R conteos I, NI PARQ, AERO, TORT, CAÑO
30 Ramphastos swainsonii R conteos NI TORT, CAÑO
31
PICIDAE Dryocopus lineatus
R
ambos
I, NI
CCC, TORT, CAÑO
32
DENDROCOLAPTIDAE Dendrocolaptes certhia*
R
ambos
I, NI
PARQ, AERO, TORT, CAÑO
33 Xiphorhynchus erythropygius R conteos I CCC
34 Lepidocolaptes souleyetii R ambos I CCC, AERO
35
THAMNOPHILIDAE Thamnophilus doliatus
R
captura
I
CCC
36 Thamnophilus atrinucha R ambos NI PARQ, TORT, CAÑO
37 Myrmotherula axillaris R captura NI CAÑO
108

APÉNDICE A. (Continuación)
Nº Nombre Científico Estatus a Método Hábitat b Sitio c
38 Cercomacra tyrannina R captura I CCC
39 Myrmeciza exsul R ambos NI PARQ, TORT, CAÑO
40 Gymnopithys leucaspis R ambos NI PARQ, CAÑO
41
TYRANNIDAE Elaenia flavogaster
R
ambos
I, NI
CCC, PARQ
42 Mionectes oleagineus R captura NI PARQ
43 Myiornis atricapillus R conteos I, NI CCC, PARQ, TORT, CAÑO
44 Todirostrum cinereum R ambos I, NI Todos los sitios
45 Todirostrum nigriceps R captura I AERO
46 Rhynchocyclus brevirostris R captura NI CAÑO
47 Contopus virens M ambos I, NI Todos los sitios
48 Empidonax flaviventris M ambos I, NI CCC, PARQ, CAÑO
49 Empidonax sp. e M ambos I, NI CCC, PARQ, AERO, CAÑO
50 Attila spadiceus R ambos I, NI PARQ, AERO, TORT, CAÑO
51 Myiarchus crinitus M ambos I, NI CCC, AERO, TORT
52 Pitangus sulphuratus R ambos I, NI Todos los sitios
53 Megarynchus pitangua R ambos I, NI CCC, PARQ, AERO, TORT
54 Myiozetetes similis R ambos I, NI Todos los sitios
55 Tyrannus melancholicus R ambos I, NI CCC, PARQ, AERO
56 Tyrannus tyrannus M conteos I, NI CCC, AERO, CAÑO
57
COTINGIDAE Querula purpurata
R
conteos
NI
TORT
58
PIPRIDAE Manacus candei
R
ambos
I, NI
Todos los sitios 109
APÉNDICE A. (Continuación)
Nº Nombre Científico Estatus a Método Hábitat b Sitio c
59 Pipra mentalis R ambos I, NI Todos los sitios
60
VIREONIDAE Vireo flavifrons
M
conteos
I
CCC, AERO
61 Vireo olivaceus M ambos I, NI CCC, PARQ, AERO, CAÑO
62 Vireo flavoviridis M ambos I CCC
63 Hylophilus decurtatus R ambos I, NI Todos los sitios
64
HIRUDINIDAE Hirudindo rustica
R
conteos
I
CCC
65
TROGLODYTIDAE Thryothorus nigricapillus
R
ambos
I, NI
CCC, AERO, TORT, CAÑO
66 Thryothorus thoracicus R ambos NI TORT
67 Troglodytes aedon R ambos I, NI CCC, PARQ, AERO
68 Henicorhina leucosticta R ambos NI TORT, CAÑO
69
SILVIIDAE Polioptila plumbea
R
conteos
I
CCC
70
TURDIDAE Catharus fuscescens
M
ambos
I, NI
CCC, PARQ, AERO, TORT
71 Catharus minimus M captura I, NI Todos los sitios
72 Catharus ustulatus M ambos I, NI Todos los sitios
73 Hylocichla mustelina M ambos NI PARQ, TORT, CAÑO
74 Turdus grayi R ambos I, NI CCC, PARQ, TORT, CAÑO
75
MIMIDAE Dumetella carolinensis
M
captura
I
CCC, AERO
110

APÉNDICE A. (Continuación)
Nº Nombre Científico Estatus a Método Hábitat b Sitio c
76
PARULIDAE Vermivora peregrina
M
ambos
I
CCC, AERO
77 Dendroica petechia M ambos I, NI CCC, AERO, CAÑO
78 Dendroica pensylvanica M ambos I, NI Todos los sitios
79 Dendroica magnolia M ambos I, NI CCC, PARQ, AERO, TORT
80 Dendroica fusca M conteos I, NI AERO, CAÑO
81 Dendroica castanea M ambos I, NI CCC, AERO, TORT
82 Dendroica cerulea M conteos I CCC
83 Mniotilta varia M ambos I, NI CCC, CAÑO
84 Setophaga ruticilla M ambos I CCC, AERO
85 Protonotaria citrea M ambos I, NI Todos los sitios
86 Helmitheros vermivorus M captura I, NI CCC, AERO, TORT
87 Seiurus aurocapillus M captura I CCC
88 Seiurus noveboracensis M ambos I, NI Todos los sitios
89 Seiurus motacilla M captura I, NI CCC, CAÑO
90 Oporornis formosus M ambos I, NI CCC, AERO, CAÑO
91 Oporornis philadelphia M captura I CCC
92 Geothlypis semiflava R captura I CCC
93 Wilsonia citrina M captura I, NI CCC, PARQ
94 Wilsonia canadensis M captura I CCC, AERO
95
THRAUPIDAE Habia fuscicauda
R
ambos
NI
TORT
111

APÉNDICE A. (Continuación)
Nº Nombre Científico Estatus a Método Hábitat b Sitio c
96 Piranga rubra M ambos I, NI CCC, AERO, TORT
97 Piranga olivacea M conteos I CCC, AERO
98 Thraupis episcopus R conteos I, NI CCC, PARQ, AERO, CAÑO
99 Thraupis palmarum R ambos I CCC, AERO
100 Euphonia gouldi R ambos I, NI PARQ, TORT, CAÑO
101 Tangara larvata R conteos NI CAÑO
102
EMBERIZIDAE Sporophila americana
R
ambos
I, NI
Todos los sitios
103 Oryzoborus funereus R captura I, NI CCC, PARQ, TORT, CAÑO
104 Arremonops conirostris R ambos I, NI CCC, PARQ, CAÑO
105 Saltator maximus R captura I CCC
106 Pheucticus ludovicianus M conteos I CCC
107
ICTERIDAE Quiscalus mexicanus
R
ambos
I
CCC
108 Icterus dominicensis R ambos I CCC
109 Icterus galbula M ambos I, NI CCC, PARQ
110 Psaracollus montezuma R conteos I, NI CCC, AERO, TORT, CAÑO
a R = Residentes; M = Migratorias. b I = Intervenido; NI = No intervenido.
c CCC = Corporación para la Conservación del Caribe; PARQ = Parque Nacional Tortuguero; AERO: Aeropuerto Barra de Tortuguero; TORT = Tortuga Lodge; CAÑO = Estación Biológica Caño Palma.
d Incluye a Empidonax alnorum y E. trailli.
112

SÍNTESIS
En la presente tesis se realizó una caracterización de la comunidad de aves asociada a
cinco sitios diferentes en los alrededores de Tortuguero, en el Caribe norte de Costa Rica
durante los meses de agosto hasta diciembre del 2002. Medidas de abundancia,
composición y riqueza de especies residentes y migratorias fueron tomadas y comparadas
entre sitios intervenidos y no intervenidos, con la finalidad de sugerir estrategias
apropiadas de manejo. Para el muestreo de aves se utilizaron las técnicas de captura con
redes de niebla y conteos de búsqueda de aves y se estimó el número de frutos presentes
para las aves en cuatro de los sitios. Se registraron un total de 128 especies de aves
distribuidas en 32 familias, de las cuales el 28.1% fueron especies migratorias. Las
especies residentes tienden a evitar más los hábitats intervenidos que las especies
migratorias. El mantenimiento de hábitats primarios debería ser una prioridad debido al
mayor tiempo que implica su formación y porque muchas especies de aves, especialmente
residentes son dependientes de este tipo de hábitats. Sin embargo, los esfuerzos de
conservación dirigidos a maximizar la riqueza de especies de aves deberían incluir la
preservación de los sitios intervenidos ya que proveen hábitats importantes para las
especies de aves, especialmente para migratorias.
Por otro lado se compararon los resultados obtenidos por las dos técnicas de muestreo
empleadas. Para las comparaciones entre métodos se consideraron 110 especies
distribuidas en 26 familias en los cinco sitios, de las cuales 61 e fueron comunes a ambos
métodos. La tasa promedio de detección de especies fue mayor con los conteos de
búsqueda de aves que con capturas, sin embargo las capturas tuvieron un mayor porcentaje
de especies migratorias. Para el total de especies registradas por ambos métodos, la
abundancia relativa mostró una correlación positiva significativa y para el 90% de las
especies los estimados de abundancia relativa fueron precisos para ambos métodos. Es
necesario reconocer los sesgos asociados a las técnicas de muestreo de aves e interpretar
cuidadosamente los resultados antes de tomar decisiones de manejo. A pesar de que el
método de búsqueda de aves resulta más eficiente en cuanto a tiempo y costos, debería ser
113

SÍNTESIS (continuación)
utilizado en combinación con otros métodos para obtener una relativamente más completa
representación de la comunidad de aves en los trópicos.
114

Apéndice A. Instrucciones para autores para la revista Wilson Bulletin.
Instructions to Authors
Be advised that manuscripts that do not conform to these guidelines will not be
accepted for publication.
I. General Instructions
Prepare manuscripts on 8.5 X 11 inch paper (if your standard-sized paper is A4 (210 X 297 mm), please leave at least a 40-mm margin at bottom so that manuscript can be copied on 8.5 X 11 inch paper without lines being cut off at the button). Double-space throughout, including tables, figure legends, and literature cited. Leave at least a 1-inch (25-mm) margin on all sides. Do not hyphenate words at ends of lines. Place be sure and submit four copies of the manuscript to: Robert C. Beason, Editor, Biology Dept., State Univ. of New York, 1 College Circle, Geneseo, NY 14454-1401. If possible, use italic type instead of underlining words to be italicized. Only the following Latin terms should be italicized: in vivo, in vitro, in utero, in situ, ad libitum, a priori, a posteriori. All others Latin terms ( except scientific names) should be left unitalicized. Cite each figure and table in the text. Tables and figures must be sequenced in the order cited. Use “ Figure” only outside of parentheses: otherwise, use “Fig.” if singular, “Figs.” If plural (e.g., Fig.2; Figs. 2 and 3; Figs. 3-6). To cite figures from another work, write figure or fig. in lowercase (e.g., figure 2 in Smith (1980); Smith 1980: fig. 2; Jones 1987: table 5). All measurements are to be given in SI units. Use continental dating (e.g., 28 September 1992), the 24-hour clock (e.g., 08:00 and 23:00), and standard time (not daylight savings time). Specify that it is Standard Time (e.g., EST for Eastern Standard Time) at first reference to time of day. English proper names of bird species that occur in North America should follow the 6th edition of AOU’s Check-list of North American Birds and its supplements. Names for other species of birds should follow an appropiate standard. English proper names of birds should be capitalized, but not the English names of other organisms. Use the following abbreviations: s (second), min (minute), h (hour); designate temperature as C. Do not abbreviate day, week, month, or year. User-defined abbreviations should be written out full words the first time the term used in text. Abbreviate thereafter: “Second-year (SY) birds ... We found SY birds in large numbers. “ Use the following statistical abbreviations: ANOVA, SD, SE, df, CV, ns, n, P, r, F, G, t-test, U-test. Other statistical abbreviations, in general, conform to sixth edition of Scientific Style and Format: The CBE Manual for Authors, Editors, and Publishers (1994, Cambridge Univ. Press, Cambridge, UK). Write out numbers one to nine unless a measurement, but use numerals for larger numbers (e.g., three birds. 6 mm, 12 days, 2 min). If number is in a series with at least one number being 10 or more, then use all numerals (e.g., 6 males and 13 females). Use 1,000 not
115

1000, 0.01 not .01, and 50% instead 50 percent. All gene or amino acid sequences must be deposited in Genbank or an equivalent repository, and the accession number(s) reported in the Methods. Each reference cited in text must be listed in Literature Cited section and vice versa. Please make final check when revised manuscript is complete. Cite literature in text as follows: 1. One author: Able (1989) or (Able 1989). 2. Two authors: Able and Baker (1989) or (Able and Baker 1989). 3. Three or more authors: Able and coworkers (1989) or (Able et al. 1989). In Literature
Cited section, give names of all authors. 4. Manuscripts that are accepted for publication but not yet published: Able (in press) or
Able (1998) if date known. 5. Unpublished materials: (Able, unpubl, data); (Able, pers, obs.); or (Able, pers. comm.). 6. 0Within parentheses: (Charley 1980; Able 1983,1990; Able and Baker 1984); (Baker
1989, Able 1992, Charley 1996); (Able 1988a, b, c). Assemble manuscript in following order: (1) title page; (2) abstract; (3) text; (4) acknowledgments; (5) literature cited; (6) figure
legends; (7) figures; (8) tables. II. Title Page (number as page 1, and present items in following order) rrh, Author’s names (in italics) · RUNNING HEAD (36 characters or less, all caps). Running head not needed for Short Communications. Title in capital letters for long articles, or caps and lowercase for Short Communications; centered. Author names in the same form as title; centered. Author address at time research was carried out. Current addresses if different should be indicated. All addresses should be numbered footnotes. Name, current address, telephone number and E-mail address of corresponding author. III. Text (page 2, etc.) Follow the instructions in section I. Do not repeat information given on title page. The following are typical main headings: Abstract, Methods, Results, Discussion, Acknowledgments and Literature Cited (no heading for Introduction). Short Communications need not be subdivided into sections, but must include an abstract. 0Up to three levels of headings can be used: (1) caps, centered; (2) caps and small caps, flush left; (3) italics, indented, with period and followed by a dash. Keep headers to a minimum. In long article, use header type 1 first, and type 3 next, then type 2 (if needed).
116

IV. Literature Cited (continue page numbering) Verify all entries against original sources, especially journal titles, volume and pages numbers, accents, diacritical marks, and spelling in languages other than English. Capitalize all nouns in German. Cite references in alphabetical order by first author’s surname and then his/her first and middle initials. References by a single author precede multiauthored works by same first author, regardless of date. List multiple authors by first author’s name (surname and then initials), second author’s name, etc. List works by the same author(s) in chronological order, beginning with earliest date of publication. If author has two works in same year, place in alphabetical order by first significant word in title; these works ahould be lettered consecutively (e.g., 1991ª, 1991b). “In press” citations must have been accepted for publication, with the name of journal or publisher included. Do not write author names in uppercase. Use the “small caps” command or “normal” case (e.g. Hendricks, D.P.) Insert a period and space after each initial of an author’s name. Journal titles should be abbreviated using the standard of the National Information Standards Organization (ANSI-NISO 1985) such as listed in the Serials Sources for the BIOSIS Data Base. Citations should follow formats below : Papers: Stutchbury, B. J. M. 1997. Effects of female cowbird removal on reproductive success of Hooded Warblers. Wilson Bull. 109: 74-81. Woodrey, M. S. and C. R. Chandler. 1997. Age-related timing of migration: geographic and interspecific patterns. Wilson Bull. 109: 52-67. Gerhardt, R. P., D. M. Gerhardt, and M. A. Vasquez. 1997. Siblicide in Swallow-tailed Kites. Wilson Bull. 109: 112-120. Willis, E. O. and Y. Oniki. 1978. Birds and army ants. Annu. Rev. Ecol. Syst. 9:243-263. Fahrig, L.and G Merriam. 1994. Conservation of fragmented populations. Conserv. Biol. 8:50-59. Dissertations, books, chapters: American Ornithologists’ Union. 1983. Check-list of North American birds, 6th ed. American Ornithologists’ Union, Washington, D.C. Freeman, S. 1991. Molecular systematics and morphological evolution in the blackbirds. Ph.D. diss., Univ. of Washington, Seattle. Gaunt, A. S. 1988. Interaction of syringeral structure and airflow in avian phonation. Pp. 915-924 in Acta XIX Congressus Internationalist Ornithologici (H. Ouellet , Ed.). Ottawa, Ontario, 1986. National Museum of Natural Sciences, Ottawa. Lack, D. 1954. The natural regulation of animal numbers. Oxford Univ. Press, London, UK. Lowther, P. E. 1993. Brown-heated Cowbird (Molothrus ater). In The birds of North
117

America, no. 47 (A. Poole and F. Gill, Eds.). Academy of Natural Sciences, Philadelphia, and American Ornithologist’s Union, Washington, D. C. Kear, J. 1970. The adaptive radiation of parental care in waterfowl. Pp. 357-392 in Social behavior in birds and mammals (J. H. Crook, Ed.). Academic Press, London, UK. Walsberg, G. E. 1983. Avian ecological energetics. Pp. 161-120 in Avian biology, vol. 7 (D. S. Farner, J. R. King, and K. C. Parkes, Eds.). Academic Press, New York. V. Tables (continue page numbering) Each table must start on separated sheet and be double-spaced throughout (header, table body, footnotes). Each table should have a legend above it and be intelligible without recourse to the text. Indent caption and begin with TABLE and Arabic numeral followed by a period. Capitalize first word of title; all other words should be lowercase except proper nouns. Include a period at end of title. Indicated footnotes by lowercase superscript letters (a, b, c, etc.). Do not use vertical lines in tables. Include horizontal lines above and below boxhead, and at end of table. Follow details of style used in The Wilson Bulletin for headings in boxhead. VI. Figures
A. Legends (continue page numbering) Start with “FIG.” Indent and double-space legends. Type legends in paragraph form separate from the figures themselves. Indicate “exotic symbols” (lines, dots, triangles, etc.) or refer to them by name in figure legend. Be aware that only a few symbols can be represented in the legend. B. Preparation Routine illustrations are black-and-white halftones (photographs), drawings, or graphs. Consult editor about color for frontispiece. Write author(s), name(s), and figure number(s) in pencil on back of each original figure or plate and each review copy. Figures in the Wilson Bulletin are virtually identical to those submitted (little degradation occurs, but flaws will show). Thus, illustrations should be prepared to professional standards. Drawings should be on good-quality paper and allow for about 20% reduction. Do not submit originals larger than 22 X 28 cm in size, unless impractical to do otherwise. Illustrations should be prepared for one- or two-column width, keeping in mind dimensions of a page in The Wilson Bulletin. Where possible, group several illustrations as panels in a single figure. Figures may be submitted with the manuscript on computer disk in eps or tiff format. Helvetica or a similar san serif typeface is recommended for figures. Symbols may be added to figures with press-on symbols and letters, but make sure they will not peel off. Handwritten or typed symbols are unacceptable. Copies of halftone figures and places must be of good quality for manuscript copies.
118

VII. What and Where to Submit
For initial submission, send four copies of original manuscript and illustrations to Editor. Initially, do not send original figures. For accepted revisions, send one copy along with the original figures and a word processor version on diskette [ WordPerfect or Word preferred, but identify sofware and platform ( Mac or IBM ) ]. The cover letter with initial submission should include a statement indicating that the manuscript reports on original research not published elsewhere and that it is submitted exclusively to The Wilson Bulletin. The letter should include any special instructions and any special instructions and any adress change during the next several months, as well as a daytime phone number and, if possible, FAX and E-mail information for the corresponding author. ( If this information was not included with the original submission letter, it must be provided in the letter with the revised maniscript ). Names of possible reviewers may be included. VIII. Proofs, reprints, and Chargers
Authors will receive page proofs for approval, along with the original typescript. Both must be returned within 48 h to the Editorial Office to avoid publication delays. A reprint order form will be sent with proofs; reprints are billed following the printer’s current cost schedule. Because changes in proofs are expensive, authors should not expect to make major modifications in their work at this stage. Excessive alterations in proofs will be charged to the author at the rate of US$3 per reset line. Authors should keep the Editor informed of adresses changes, so proofs will not be delayed. Authors are required to pay pages charges for pages in excess of the 20 page limit. The WOS request that authors, if possible, bear part or all of the cost of publishing their papers when grant, institutional, or personal funds are available for the purpose. Current cost per printed page are US$100; a minimum contribution of US$50 is recommended, but the WOS is pleased to receive any payment considered appropriate. Authors who do not have access to publication funds may request a waiver of this payment. If you have questions, contact the Editor at <[email protected]> .
119

INSTRUCTIONS TO AUTHORS
The Journal of Field Ornithology is the quarterly of the Association of Field Ornithologists. The Journal welcomes original articles that emphasize the descriptive or experimental study of birds in their natural habitats. Articles describing techniques or methodological refinements must have relevance beyond a single species or study system. Contributions are encouraged from throughout the world, but must de written in English. The Journal is especially interested in studies conducted in the Neotropics and those involving participation by nonprofessional ornithologists. SUBMISSION
Articles should de submitted in quadruplicate to the Editor: Charles R. Brown, Department of Biological Science, University of Tulsa, Tulsa, Oklahoma 74104-3189 USA. A cover letter should include the tittle of the paper, a statement that the paper (as a whole or in part) has not be published or submitted for publication elsewhere, and the name, mailing address, and email address for the subsequent nine months of the corresponding author. Authors are encouraged to suggest the name of 4-5 potential reviewers (although the Editor may choose reviewers other than those suggested). The Journal as yet does not accept electronic submissions.
FORMAT
Prepare manuscripts carefully with
attention to all details. Manuscripts that depart from these guidelines will be returned without review. Print the paper double-spaced throughout (including the title page, tables and figure legends); use the same font (no smaller than 12 point) throughout the manuscript. Manuscripts should be printed on one side of good quality bond paper that measures 21.5 X 28 cm (8.5 X 11 in). Margins should be at least 2,.5 cm (1 in) on all sides of the page. Do not use italic or bold type; underline text that should appear in italic (e.g., scientific names). Place the first author’s last name and the page number (starting with the abstract on page 2 and continuing throughout the Literature Cited) in the upper right corner of each page. Authors should use recent issues of the Journal as a general guide in preparing their manuscripts. Title page. In the upper left corner
provide the author’s name (e.g., R.T. Smith; R.T. Smith and P. R. Jones; or E.F. Hunt et al.) as a left running head and, below this, a short title as a right running head. In the upper right provide the name and address of the author to receive proofs. Centered below these, provide the full title (double-spaced) and the name of all authors and their addresses at the time the research was conducted. Each author’s current address, if different, should be given as a numbered footnote at the bottom of the title page. The corresponding author should be indicated by providing his/her email address in a footnote. Use a recent issue of the Journal for correct formatting and style of author and address listings. Abstract. The second page that does not
exceed 5% of the length of the paper. The abstract should explain the purpose of the study, describe the principal findings, and
Apéndice B. Instrucciones para autores para la revista Journal of Field Ornithology.
120

state the main conclusions. Many readers will rely heavily on the abstract so it should be as descriptive as possible. Void uninformative sentences such as “ the significance of these results is discussed. ” Generally, quantitative results (e.g., sample sizes, P-values) are unnecessary in Abstracts and be summarized in narrative fashion. Below the Abstract, provide 5-6 keywords that describe the subject of the paper; these need not duplicate words in the title. The editorial staff will articles accepted for publication. Authors are welcome to submit their own Spanish translations. Text. Begin the text on page 3.
The English and scientific names of a species should be given the first time it is mentioned. The scientific name should be underlined; do not use italic type. Bird names should follow the AOU Check-list of North American Birds (1998) and supplements or the appropriate equivalent unless departures are explained and defined. The first letter of common names of bird species should be capitalized. Use metric units. Measurement unit abbreviations should be those given in the fifth edition of the CBE style manual, 5th edition (1983). Use the 24-hour clock (05:00 and 17:00) and “continental” dating (10 March 1992). Define all symbols, abbreviations, and acronyms but minimize their use. Probability values should be written as P = 0.025; give exact values when possible even for nonsignificant results (P=0.67, not P>0.05 or NS). Sample size is abbreviated N. Use the following abbreviations: second, s; minute, min; hour, h; month, mo; week, wk; year, yr. Statistic (e.g., X2
, F34,56). Test
statistics should be given with all P-values. Acknowledgments. Institutional
affiliations are not allowed for persons thanked in acknowledgments. Literature Cited. List literature citations
alphabetically by the first author’s last name in a Literature Cited section following the Acknowledgments. Literature Cited entries (in a style conforming to that in the latest issue of the Journal) should be carefully double-checked against citations in the text. Journal and publisher names are spelled out in their entirety. Text citations should be in the author-year format (LeConte 1995; Edwards and Sutton 1994, 1996; Darwin et al. 1997); multiple text citations should be in sequential order by year of publication. If you cite or quote critical material directly from longer works, indicate the pertinent pages (e.g., Smith 1994: 23-24). Unpublished papers should not be cited unless a copy has been deposited in the Van Tyne Library at Museum of Zoology, University of Michigan, where they may be obtained for use by other researchers. Do not cite manuscripts that are in preparation or review and avoid citation of “gray” literature such as technical reports by governmental agencies that are often difficult for other researchers to find. Regularly published serial publications containing chapters by multiple authors, such as Current Ornithology, Farner and King’s Avian Biology, and Studies in Avian Biology should be cited as journal articles. Accounts from the Birds of North America series should be cited using the style for book chapters. Example of citation style: Journal article
Hoogland, J. L., and P.W. Sherman. 1976.
Advantages and disadvantages of Bank Swallow (Riparia riparia) coloniality. Ecological Monographs 46: 33-58.
121

Book
Sharpe, R. S., W. R. Silcock, and J.G.
Jorgensen. 2001. Birds of Nebraska: their distribution and temporal occurrence. University of Nebraska Press, Lincoln, NE.
Book chapter
Rogers, C. A., R. J. Robertson, and B.
J. Stutchbury. 1991. Patterns and effects of parasitism by Protocalliphora sialia on Tree Swallow nestlings. In: Bird-parasite interactions: ecology evolution and behaviour (J. E. Loye and M. Zuk, eds.), pp.123-139. Oxford University Press, Oxford, UK.
Thesis or dissertation
Brown, C. R. 1985. The costs and
benefits of coloniality in the Cliff Swallow. Ph.D. dissertation. Princeton University, Princeton, NJ.
Tables. Each table should be
printed double-spaced throughout on a separate page. Place the tables after the Literature Cited. Tables should be numbered sequentially and include a concise and informative title. Do not use additional sentences after the table’s title; material necessary to clarify the table should be presented as footnotes to the table. Tables should supplement, not duplicate, material in the text or figures. Compose tables carefully and use all stylistic conventions (e.g., abbreviations) as the text. Tables
should be prepared with the size of the journal’s pages and its double-column format in mind. Avoid wide, shallow tables. Figures. Submit figures as glossy 13 X
18 cm (5 X 7 in) black-and-white prints or laser-printed originals. Lightly pencil the author’s name and the figure number on the back of each figure. Figures should be uncluttered, but convey a maximum amount of information; they should not duplicate material in the text or tables. When preparing figures use “thin” rather than “fat” letters so that reduction of the figure will not close the spaces in letters such as “e” or “p”, and do not use boldface font in figures. Use high-quality computer graphics and lettering on a laser printer; for laser-printed graphs, submit original printed versions on each manuscript copy rather than photocopies. All lettering on the figure should conform to the same stylistic conventions as the text of the manuscript, including abbreviations. Original drawings should be large enough to permit reduction to the size they will appear in print. Figures should be prepared with the size of the journal’s pages and double-column format in mind. Type (double-space) figure legends consecutively on one page. Do not print keys to dot symbols or shading types on the figure; define these in the legend. Do not use four-letter Bird Banding Laboratory species abbreviations (e.g., CLSW) in figures or anywhere in the manuscript. Authors are encouraged to follow the suggestions of Kroodsma (2000, Auk 117: 1081-1083) in preparing figure legends and titles of tables, in which the main point of the figure or table is clearly illustrated in the legend or title. Figures and tables should be designed to convey information when standing alone; extensive cross-reference of the to the text (e.g., “see Methods”) is unacceptable.
122

Editorial Assistance Program
The Association of Field Ornithologist offers a free service assisting authors of ornithological articles who are not natives speakers of English. The goal of the Editorial Assistance Program (EAP) is to enable and encourage Latin American and other ornithologist to publish their work in widely read international journals. This is not a translation service, however. Manuscript must be written in English (even if flawed), and a AFO volunteer will work with the authors to refine the writing into idiomatic English appropriate for scientific publication. It is often useful for the English version to be accompanied by one in the authors’ native language.
It is important to realize that scientific content will not generally be addressed through this program, rather only suggestions for improving clarity and grammar will be provided. While submission of appropriate articles to the AFO’s own Journal of Field Ornithology is encouraged, it is not required for this program. If interested in helping authors with manuscripts as part of this program, please contact the EAP Coordinator. All inquiries from authors should de directed to Daniel M. Brooks, EAP Coordinator, Department of Vertebrate Zoology, Houston Museum of Natural Science, One Herman Circle Drive, Houston, Texas 77030-1799 USA, (phone 713-639-4776), email dbrooks@ hmns.org. Electronic submission of manuscripts to the EAP Coordinator via an email is strongly encouraged.
123