caveolina-1 inhibe la proliferacion y promueve la … · ii universidad de chile facultad de...
TRANSCRIPT

UNIVERSIDAD DE CHILE
FACULTAD DE MEDICINA ESCUELA DE POSTGRADO
CAVEOLINA-1 INHIBE LA PROLIFERACION Y PROMUEVE LA MUERTE CELULAR MEDIANTE LA
REGULACION NEGATIVA DE LA CICLOOXIGENASA-2 Y SURVIVINA
DIEGO ALEJANDRO RODRIGUEZ GONZALEZ
TESIS PARA OPTAR AL GRADO DE DOCTOR EN CIENCIAS BIOMEDICAS
Directores de Tesis: Prof. Dr. Andrew Quest
Prof. Dra. Lisette Leyton
Santiago-Chile
2009

ii
UNIVERSIDAD DE CHILE FACULTAD DE MEDICINA ESCUELA DE POSTGRADO
INFORME DE APROBACION TESIS DE DOCTORADO EN CIENCIAS BIOMEDICAS
Se informa a la Comisión de Grados Académicos de la Facultad de Medicina, que la Tesis de Doctorado en Ciencias Biomédicas presentada por el candidato
DIEGO ALEJANDRO RODRIGUEZ GONZALEZ
ha sido aprobada por la Comisión Informante de Tesis como requisito para optar al Grado de Doctor en Ciencias Biomédicas en Examen de Defensa de Tesis rendido el día 7 de Abril de 2009.
Prof. Dr. Andrew Quest
Prof. Dra. Lisette Leyton
DIRECTORES DE TESIS Dpto. de Biología Celular y Molecular, Instituto de Ciencias Biomédicas,
Facultad de Medicina, Universidad de Chile
COMISION INFORMANTE DE TESIS
PROF. DR. MARIO GALINDO PROF. DR. SERGIO LAVANDERO PROF. DR. MIGUEL BRONFMAN PROF. DR. ENRIQUE CASTELLON Presidente Comisión de Examen

iii
DEDICATORIA
Agradezco con mucho cariño a mi gran familia, en especial a mis padres, Jorge y
Jacqueline y hermanas, Lorena y Constanza, gracias a los valores y cariño que me han
dado son un pilar fundamental en mi formación como persona y profesional, por esto
les dedico la obtención de este grado académico. También se los dedico con mucho
cariño a mis padrinos, Emilio y Clara y mis primos-hermanos Cristian y Carolina. Quiero
agradecer con mucho cariño a mi Mamaria y los otros tatas que están descansando
(Tatalalo, Tatamanuel y Mamaminga), sobrinos Tomas, Isidora y Pau-mama. A la
familia Barría (Tíos y primos), Claudio Sharkawi, tía Lucy y Chin también les dedico
esta tesis por su cariño y apoyo.
Quiero agradecer en especial al Dr.Andrew Quest y Dra. Lisette Leyton de quienes he
recibido una gran guía profesional, científico y además han sido grandes personas y
amigos. Muchas gracias a los integrantes y amigos que he hecho en el laboratorio:
Loro, Alejandra, Manuel, Jorge, Claudia, Lillian, Enrique, Lorena, Sergio, Hery, Nicolás,
Natalia, Carlos, Rodrigo, Adam y los que ya han tomado otros caminos como: Ana
María, Daniel, Nati, Cristian, Denisse, Freddy, Joan, gracias a todos por los gratos
momentos vividos en el laboratorio.
Otro especial agradecimiento a mis grandes amigos Andrés y Koke que me han
apoyado en las buenas y en las malas, gracias Julio por tu gran apoyo en el laboratorio
y por ser un gran amigo. A mis amigos desde el pregrado GES, Vicente y Álvaro les
dedico este grado por su apoyo incondicional, también les dedico este logro a Gonzalo
y Mauricio que gracias al lab he conocido a estos grandes amigos. Gracias a mis
amigos de siempre, Canito, Alejandro, Mauricio (Lukas), Marcelito, Niel, Daniel,
Miguelito (Q.E.P.D), Araceli, Denise y Jazmín.
Finalmente, quiero agradecer al Dr. Jay Heinecke y Charles Holmes por su valiosa
cooperación, apoyo y los buenos momentos vivídos durante la realización de mi
estadía en Seattle.
Nuevamente, muchas gracias a todos los compañeros y amigos con quienes compartí
estos años inolvidables.

iv
AGRADECIMIENTOS
Esta Tesis de Doctorado se desarrolló en el laboratorio de Comunicaciones
Celulares, Centro FONDAP de Estudios Moleculares de la Célula (CEMC), Instituto de
Ciencias Biomédicas, Facultad de Medicina, Universidad de Chile y fue posible gracias
a los siguientes proyectos y becas:
Financiamiento otorgado a Andrew F. G. Quest .
• Proyecto FONDAP 15010006
Financiamiento otorgado a Lisette Leyton C.
• Proyecto Fondecyt 1040390 y 1070699
Financiamiento otorgado a Julio Tapia P.
• Proyecto Fondecyt 3050037 y 11070116
Financiamiento otorgado a Diego Rodríguez G.
1. Beca de post-grado MECESUP, período 2004-2007.
2. Beca de post-grado FONDAP, periodo 2007-2008.
3. Beca para estadías cortas en el extranjero, Departamento de Postgrado y
Postitulo, Universidad de Chile, Santiago, Chile. Actividad realizada entre Marzo-
Junio 2008. “Entrenamiento en la detección de proteínas por espectrometría de
masas MALDI-TOF/TOF en complejos multiproteicos mediados por caveolina-1:
Papel de E-cadherina en la supresión de la expresión de COX-2 por
caveolina-1”. Laboratorio de Metabolismo, Endocrinología y Nutrición.
Departamento de Medicina, Universidad de Washington, Seattle, Washington,
USA.

v
Parte de la investigación derivada de este trabajo o bien relacionada con éste, ha sido o será publicada en los siguientes artículos:
Manuscritos publicados:
1. Rodriguez, D.A., Tapia, J.C., Fernández, J.G.R., Torres. V.A., Muñoz, N., Leyton, L.,
Galleguillos, D, Quest, A.F.G. Caveolin-1-mediated suppression of cyclooxygenase-2 via
a β-catenin-Tcf/Lef-dependent transcriptional mechanism reduced PGE2 production and
survivin expression. Mol Biol Cell. 20(8): 2297-2310, 2009 2. Torres, V.A., Tapia, J.C., Rodriguez, D.A., Lladser, A., Leyton, L., Quest, A.F.G.
E-cadherin is required for caveolin-1-mediated down-regulation of the inhibitor of
apoptosis protein survivin via reduced β-catenin/Tcf-Lef-dependent transcription. Mol
Cell Biol. 27(21): 7703-7717, 2007
3. Tapia, J.C., Torres, V.A., Rodriguez, D.A., Leyton, L., Quest, A. F.G. Casein kinase 2
(CK2) increases survivin expression via enhanced β-catenin-T cell factor / lymphoid
enhancer binding factor-dependent transcription. Proc Natl Acad Sci USA. 103
(41):15079-84, 2006
4. Torres. V.A., Tapia, J.C., Rodriguez, D.A., Parraga, M., Lisboa, P., Montoya, M.,
Leyton, L., Quest, A. F.G. Caveolin-1 down-regulates the expression of survivin via a
transcriptional mechanism involving the β-catenin-Tcf/Lef pathway. J Cell Sci. 119
(9):1812-23, 2006
Manuscritos en preparación:
1. Rodriguez, D.A., Sanhueza, C., Lobos, L., Lladser, A., Torres. V.A., Leyton, L., Quest,
A.F.G. Caveolin-1-enhanced degradation of survivin via the proteasome pathway is
blocked by PGE2. En preparación.
2. Tapia J.C., Rodriguez, D.A., Ponce D.P., Fernández J.G., Cataldo L.R., and Quest
A.F.G. Protein Kinase CK2-dependent Up-regulation of Cyclooxygenase-2 Enhances
Viability and Resistance to Apoptosis in Cancer Cells. En preparación.

1
INDICE GENERAL INDICE GENERAL................................................................................................................. 1 INDICE DE FIGURAS ............................................................................................................ 5 INDICE DE TABLAS.............................................................................................................. 6 ABREVIATURAS ................................................................................................................... 7 RESUMEN.............................................................................................................................. 8 SUMMARY ........................................................................................................................... 11 I. INTRODUCCION............................................................................................................... 14 1. Cáncer: antecedentes generales .................................................................................. 14 2. Caveolina-1 en la fisiología celular............................................................................... 15
2.1. Rol de caveolas y caveolina-1 .................................................................................. 15
2.2. Caveolina-1 en la transformación oncogénica y cáncer........................................... 16
2.3. Estructura, dominios y señalización intracelular de caveolina-1.............................. 19
3. Vías de señalización mediada por β-catenina y su papel en el cáncer .................... 21
4. Relación entre COX-2, survivina y caveolina-1: papel en carcinogénesis .............. 23
4.1. Rol de COX-2 en cáncer de colon. ........................................................................... 23
4.2. Survivina en cáncer de colon y su regulación negativa por la expresión de
caveolina-1 a través de la vía β-catenina-Tcf/Lef............................................................ 27
4.3. PGE2 activa la vía de β-catenina-Tcf/Lef.................................................................. 29
II. HIPOTESIS DE TRABAJO.............................................................................................. 31 III. OBJETIVO GENERAL.................................................................................................... 31 IV. OBJETIVOS ESPECIFICOS .......................................................................................... 31 V. METODOLOGIA .............................................................................................................. 32 1. Materiales ........................................................................................................................ 32 2. Anticuerpos..................................................................................................................... 33 3. Cultivo celular y transfecciones ................................................................................... 33
3.1. Cultivo líneas celulares............................................................................................. 33
3.2. Plasmidios................................................................................................................. 34
3.3. Expresión ectópica de caveolina-1, COX-2 y plasmidios reporteros ....................... 36
Páginas

2
3.3.1. Expresión ectópica de caveolina-1 y/o COX-2 en células HEK293T.................... 37
3.3.2. Expresión de caveolina-1 y/o COX-2 en células de cáncer .................................. 37
3.3.3. Expresión de caveolina-1 y E-cadherina en células de cáncer de colon
HT29(US). ........................................................................................................................ 38
4. Análisis de la expresión de survivina, COX-2 y caveolina-1 ..................................... 39 4.1. Cuantificación del mRNA de COX-2 y survivina por RT-PCR semicuantitativo y
PCR en tiempo real.......................................................................................................... 39
4.2. Reacción de transcripción inversa (RT) ................................................................... 39
4.3. Reacción de RT-PCR semicuantitativo .................................................................... 40
4.4. Reacción de PCR en tiempo real ............................................................................. 41
5. Western blot e inmunoprecipitación ............................................................................ 42 5.1. Extracción de proteínas, SDS-PAGE y Western blot ............................................... 42
5.2. Inmunoprecipitación de caveolina-1 con proteína A-sefarosa ................................. 43
5.3. Preparación de anticuerpos inmovilizados en esferas magnéticas ......................... 44
5.4. Análisis de inmunoprecipitados de caveolina-1 por espectrometría de masas ....... 45
6. Ensayos de reporteros................................................................................................... 46 6.1. Medición de la actividad transcripcional de Tcf/Lef.................................................. 46
6.2. Medición de la actividad transcripcional del promotor de survivina ......................... 47
6.3. Medición de la actividad transcripcional del promotor de COX-2 ............................ 47
7. Ensayos de viabilidad y proliferación.......................................................................... 47 7.1. Ensayo de exclusión con Azul de tripan................................................................... 47
7.2. Ensayo de proliferación MTS®................................................................................. 48
8. Inmunofluorescencia indirecta-microscopia confocal............................................... 49
9. Cuantificación de PGE2.................................................................................................. 50 10. Incubación de células con PGE2................................................................................. 51 11. Análisis estadístico ...................................................................................................... 52
VI. RESULTADOS................................................................................................................ 53 1. SECCION 1: Objetivo específico 1: Analizar el efecto de la expresión ectópica de caveolina-1 sobre la regulación transcripcional de COX-2 ................................................................ 53
1.1. Cuantificar la expresión de COX-2 en células humanas de cáncer de colon
[DLD-1, HT29(ATCC) y HT29 (US)], de mama (ZR75) y de origen embrionario
(HEK293T) en presencia y ausencia de caveolina-1 ..................................................... 53

3
1.2. Cuantificar la actividad transcripcional de β-catenina-Tcf/Lef (TOP/FOP-FLASH)
y de COX-2 (Promotor de COX-2) .................................................................................. 60
2. SECCION 1: Objetivo específico 2: Estudiar el mecanismo de regulación transcripcional de survivina por la expresión de COX-2 y su nexo con la regulación negativa en la proliferación celular por caveolina-1................................................................................ 64
2.1 Determinar el efecto de la sobreexpresión de COX-2 sobre la actividad
transcripcional de β-catenina-Tcf/Lef y sobre la expresión de survivina........................ 64
2.2. Determinar el efecto de PGE2 sobre la actividad transcripcional de β-catenina-
Tcf/Lef y sobre la expresión de survivina ....................................................................... 68
2.3. Determinar el efecto de la PGE2 o bien la sobreexpresión de COX-2 sobre la
expresión de survivina y la proliferación en células que expresan caveolina-1............. 71
2.4. Investigar el efecto de la inhibición de COX-2 sobre la proliferación en células
de cáncer de colon [HT29(ATCC) y DLD-1] que expresan caveolina-1......................... 77
2.5. Estudiar el efecto de PGE2 en la localización nuclear de β-catenina y sobre el
secuestro de β-catenina a la membrana plasmática por caveolina-1 en células de
cáncer de colon [HT29(ATCC)]....................................................................................... 82
3. SECCIÓN 2: Objetivo especifico 3: Analizar el efecto de la expresión ectópica de caveolina-1 sobre la regulación post-traduccional de survivina....................................................... 87
3.1. Estudio de los complejos multiproteicos que conforma caveolina-1 en células de
cáncer de colon [HT29(ATCC) y HT29(US)] .................................................................. 87
3.1.1. Preparación de células y control de la expresión de caveolina-1 ........................ 88
3.1.2. Inmunoprecipitación de caveolina-1 ..................................................................... 89
3.1.3. Análisis de muestras por MALDI-TOF .................................................................. 90
3.1.4. Análisis de muestras por LTQ (Cromatografía capilar de fase reversa acoplado a
masas) ............................................................................................................................. 90
3.2. Analizar el efecto de la expresión ectópica de caveolina-1 sobre la regulación
post-traduccional de survivina ........................................................................................ 91
Resumen de resultados................................................................................................ 99
VII. DISCUSION ................................................................................................................. 100 1. Caveolina-1 disminuye la expresión de COX-2 por un mecanismo
transcripcional dependiente de β-catenina-Tcf/Lef .................................................. 100

4
1.1. Inhibición de la expresión de COX-2 en células humanas de cáncer de colon,
de mama y de origen embrionario ................................................................................ 100
1.2. La inhibición de la expresión de COX-2 por caveolina-1 requiere la presencia
de E-cadherina.............................................................................................................. 101
1.3. La inhibición de la expresión de COX-2 por caveolina-1 ocurre por la represión
de vía de β-catenina-Tcf/Lef......................................................................................... 102
1.4. Mecanismos alternativos inhibitorios de caveolina-1............................................. 104
2. La sobreexpresión de COX-2 o la incubación con PGE2 estimulan la actividad
transcripcional de β-catenina-Tcf/Lef aumentando la expresión de survivina y
COX-2................................................................................................................................. 106
2.1. Efecto de COX-2 sobre la transcripción dependiente de β-catenina-Tcf/Lef ........ 106
2.2. Efecto de PGE2 y COX-2 sobre la regulación negativa de caveolina-1 en la
expresión de survivina .................................................................................................. 109
3. La inhibición específica de COX-1 o COX-2 potencia la disminución de la proliferación inducida por caveolina-1 .......................................................................... 114
4. La presencia de caveolina-1 disminuye la vida media de survivina....................... 117
VIII. CONCLUSION Y PROYECCIONES .......................................................................... 119 IX. REFERENCIAS ............................................................................................................ 121

5
INDICE DE FIGURAS
Figura 1. Eventos secuenciales que conllevan al desarrollo del cáncer.............................................................. 14
Figura 2. Caveolina-1 se encuentra disminuida en muestras tumorales de colon humano y en líneas
celulares de cáncer de colon y su re-expresión en células HT29 disminuye su tumorigenicidad in vivo............. 18
Figura 3. Estructura y dominios de caveolina-1. ................................................................................................. 20
Figura 4. Vía de señalización mediada por β-catenina. ...................................................................................... 24
Figura 5. PGE2 activa la vía de β-catenina-Tcf/Lef. ............................................................................................ 29
Figura 6. Sistema de reporteros TOP/FOP-FLASH y reporteros basados en el promotor de survivina y de
COX-2................................................................................................................................................................. 36
Figura 7 Esquema resumen de los tratamientos con PGE2 ................................................................................ 51
Figura 8. Niveles basales de caveolina-1, COX-2 y survivina de líneas celulares humanas de cáncer de
colon [HT29(ATCC), DLD-1], de mama (ZR75) y de origen embrionario (HEK293T) ......................................... 54
Figura 9. Caveolina-1 reduce los niveles de mRNA y proteína de COX-2 en células de cáncer de mama
(ZR75) y colon [HT29(ATCC), DLD1]. ................................................................................................................ 55
Figura 10. Caveolina-1 inhibe la expresión de COX-2 en células HEK293T. ...................................................... 57
Figura 11. La reducción de los niveles de mRNA de COX-2 requiere la presencia de E-cadherina en
células HT29(US). .............................................................................................................................................. 59
Figura 12. La inhibición de la quinasa GSK-3β aumenta los niveles de mRNA y proteicos de COX-2 y
survivina. ............................................................................................................................................................ 61
Figura 13. Caveolina-1 inhibe la expresión de COX-2 por la supresión de la actividad transcripcional
dependiente de β-catenina-Tcf/Lef en células HEK293T. ................................................................................... 62
Figura 14. Caveolina-1 inhibe la actividad transcripcional dependiente de β-catenina-Tcf/Lef en células de
cáncer................................................................................................................................................................. 63
Figura 15. Caracterización del vector de expresión de COX-2 (pOSML-COX-2) ................................................ 64
Figura 16. La sobreexpresión de COX-2 aumenta la expresión de survivina y β-catenina en células
HEK293T ............................................................................................................................................................ 65
Figura 17. La sobreexpresión de COX-2 en células HEK293T aumenta la actividad de los reporteros de
β-catenina-Tcf/Lef, survivina y COX-2 ................................................................................................................ 66
Figura 18. La sobreexpresión de COX-2 en células HEK293T aumenta la producción y liberación de
PGE2................................................................................................................................................................... 67
Figura 19. PGE2 aumenta la expresión de survivina y COX-2 ............................................................................ 69
Figura 20. La incubación de células HEK293T con PGE2 aumenta la actividad de reporteros de
β-catenina-Tcf/Lef, survivina y COX-2 ................................................................................................................ 69
Figura 21. PGE2 aumenta los niveles proteicos de survivina y COX-2 y se correlaciona con un aumento
en la proliferación de células HEK293T .............................................................................................................. 70
Figura 22. La expresión de caveolina-1 disminuye la producción de PGE2 ........................................................ 71
Figura 23. PGE2 revierte la inhibición de la expresión de COX-2 y survivina por caveolina-1 en líneas
celulares de cáncer............................................................................................................................................. 73
Figura 24. La sobreexpresión de COX-2 previene la inhibición de la expresión de survivina por
caveolina-1 en células DLD-1 ............................................................................................................................. 74

6
Figura 25. La sobreexpresión de COX-2 en células DLD-1 previene la inhibición de la expresión de
survivina, la producción de PGE2 y la proliferación causada por caveolina-1 ..................................................... 75
Figura 26. La sobreexpresión de COX-2 o la incubación con PGE2 previene la inhibición de la
proliferación causada por la sobreexpresión de caveolina-1............................................................................... 76
Figura 27. Efecto de PGE2 e inhibidores de COX-1 y COX-2 en células DLD-1 ................................................. 78
Figura 28. PGE2 previene la inhibición de la proliferación por la expresión de caveolina-1 ................................ 78
Figura 29. El efecto de caveolina-1 sobre la transcripción de survivina y producción de PGE2 en células
DLD-1 se ve potenciado por la inhibición de COX-2........................................................................................... 80
Figura 30. La inhibición de la proliferación por la inhibición de COX-2 es potenciada en células que
expresan caveolina-1.......................................................................................................................................... 81
Figura 31. PGE2 inhibe la co inmunoprecipitación de �-catenina con caveolina 1 en células HT29(ATCC) ...... 83
Figura 32. PGE2 inhibe la co-inmunoprecipitación de caveolina-1 con β-catenina.............................................. 83
Figura 33. PGE2 aumenta la localización nuclear de β-catenina......................................................................... 84
Figura 34. PGE2 aumenta la localización nuclear de β-catenina e inhibe la capacidad de caveolina-1 para
secuestrar β-catenina a la membrana plasmática............................................................................................... 85
Figura 35. Expresión de caveolina-1 en células de cáncer de colon [HT29(US) y HT29(ATCC)] ....................... 88
Figura 36. Inmunoprecipitación de caveolina-1 en células de cáncer de colon [HT29(US) y HT29(ATCC)] ....... 89
Figura 37. Análisis de muestras por MALDI-TOF ............................................................................................... 96
Figura 38. Caveolina-1 disminuye la vida media de survivina en células DLD-1................................................. 97
Figura 39. Caveolina-1 disminuye la vida media de survivina en células HT29(US)........................................... 98
Figura 40. Caveolina-1 disminuye la vida media de survivina en células B16F10 .............................................. 98
Figura 41. Modelo de inhibición de la expresión de COX-2 por caveolina-1 ....................................................... 104
Figura 42. Mecanismos alternativos inhibitorios de caveolina-1 ......................................................................... 106
Figura 43. β-catenina se ubica en la membrana plasmática sólo en su forma desfosforilada............................. 112
Figura 44. La presencia de caveolina-1 podria promover la degradacion de survivina via el proteasoma .......... 118
Figura 45. Resumen del trabajo del laboratorio en la identificación de blancos de caveolina-1 en su papel
como supresor de tumores y de aquellos mecanismos que impiden que caveolina-1 cumpla este papel .......... 120
INDICE DE TABLAS
Tabla 1. Comparación y características de las líneas celulares versus célula normal ........................................ 34
Tabla 2. Oligonucleótidos y Tm usados para la obtención de cDNAs de COX-2, survivina, caveolina-1
y β-actina ............................................................................................................................................................ 40
Tabla 3. Análisis de muestras por LTQ (inmunoprecipitados HT29(ATCC)) ....................................................... 92
Tabla 4. Análisis de muestras por LTQ (inmunoprecipitados HT29(US))............................................................ 94

7
ABREVIATURAS AMV: transcriptasa inversa de AMV
APC: Poliposis Coli Adenomatosa
BAL: benzamidina-antipaína-leupeptina
BCA: ácido bicincoínico
BSA: seroalbúmina de bovino
CHX: cicloheximida COX-2: ciclooxigenasa-2
DABCO: diazobiciclooctano
DMSO: dimetilsulfóxido
dNTP: desoxinucleótido trifosfato
E-cadherina: caderina epitelial
EDTA: ácido etilendiaminotetracético
FBS: suero fetal bovino
FITC: isotiocianato de fluoresceína
GFP: proteína fluorescente verde
GSK-3β: glicógeno sintetasa kinasa 3β
HRPO: peroxidasa de rabanito
IAP: inhibidor de la apoptosis
IFI: inmunofluorescencia indirecta
IgG: inmunoglobulina G
iNOS: sintetasa de óxido nítrico inducible
IPTG: isopropil-β-D-tiogalactósido
LiCl: Cloruro de litio MALDI-TOF: Matrix-Assisted Laser Desorption/Ionization-Time Of Flight RNAm: ácido ribonucleico mensajero
MTS®: 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium)
ONPG: 2-nitrofenil-β-D-galactopiranósido
PBS: tampón fosfato salino
PI3K: fosfatidil inositol 3-kinasa
PMSF: fenil metil sulfonil fluoruro
RIPA: tampón de radioinmunoprecipitación
RT-PCR: transcripción inversa-reacción en cadena de la polimerasa
PAGE: electroforesis en gel de poliacrilamida
PGE2: Prostaglandina E2
SDS: dodecil sulfato de sodio
TBE: sitios de unión a Tcf/Lef
Tcf/Lef: factor de células T / factor de células linfoides
VEGF: factor de crecimiento vascular endotelial
Wnt-1: Wingless-type MMTV integration site family member 1

8
RESUMEN El cáncer es una de las mayores causas de muerte a nivel mundial. Múltiples
cambios a nivel molecular se han asociado con la génesis de esta enfermedad, los cuales se ubican dentro de dos categorías, estas alteraciones atribuidas a oncogenes (ganancia de función) o a supresores de tumores (pérdida de función). Un ejemplo de gen supresor de tumores es el gen de caveolina-1, ya que su expresión es eliminada en muchos tipos de células de cáncer y su re-expresión, dependiendo del contexto y tipo celular, inhibe las características asociadas con el fenotipo transformado. Caveolina-1 es una proteína integral de membrana de 21-24 kDa y constituye el principal componente proteico de las caveolas o “pequeñas cuevas” que son invaginaciones vesiculares de la membrana plasmática de 50-100 nm de tamaño. Caveolina-1 pertenece a una familia de proteínas que comprende tres isoformas diferentes (caveolina-1, 2 y 3). La caveolina-1 y 2 son usualmente co-expresadas en diferentes tejidos y células, mientras que la expresión de caveolina-3 se limita principalmente a células musculares.
En diferentes tipos de cáncer, genes cuya transcripción es normalmente inducible, frecuentemente se vuelve constitutiva, como es el caso de la enzima ciclooxigenasa-2 (COX-2), que cataliza la síntesis de prostaglandinas (PGs) a partir del ácido araquidónico (AA). Particularmente, en cáncer de colon, PGE2, aumenta la actividad transcripcional de β-catenina que en asociación con los factores de transcripción Tcf/Lef (Tcf, T cell factor; Lef, Lymphoid enhancer factor) promueven la expresión de genes tales como: ciclina D1 y survivina. Survivina, es un miembro de la familia de proteínas inhibidoras de la apoptosis (IAPs) y está normalmente ausente en los tejidos diferenciados adultos, pero se sobreexpresa en células cancerosas y su expresión correlaciona bien con una tasa proliferativa aumentada y el mal pronóstico para los pacientes.
Se ha descrito previamente que caveolina-1 co-localiza en la membrana plasmática con E-cadherina y β-catenina, dado que co-inmunoprecipita con estas proteínas. Adicionalmente, la expresión de caveolina-1 reprime la actividad transcripcional dependiente de β-catenina-Tcf/Lef, inducida por la proteína secretada Wnt o por la sobreexpresión de β-catenina. En concordancia con estas observaciones, nuestro laboratorio demostró que la sobreexpresión de caveolina-1 en células HEK293T, ZR75, DLD-1 y HT29(ATCC), reprime la expresión de survivina, por un mecanismo transcripcional que involucra la vía de β-catenina-Tcf/Lef, disminuye la proliferación y promueve la muerte celular. Además, dicha inhibición requiere de la presencia de E-cadherina, dado que células de origen metastático HT29(US), que carecen de E-cadherina no responden a dicha regulación, pero lo hacen al re-expresar esta proteína.
Alternativamente, demostramos previamente que caveolina-1 regula blancos río abajo por un mecanismo de regulación post-transcripcional mediado por la degradación proteasomal, como es el caso de la isoforma sintasa inducible del oxido nítrico (iNOs).

9
El trabajo descrito aquí se focalizó en la identificación de mecanismos por el cual caveolina-1 regula la actividad/expresión de COX-2, también estudiamos como alteraciones en la expresión de COX-2 se relaciona con posibles cambios en la expresión de survivina y la proliferación celular. En estos estudios se analizaron tres líneas celulares de cáncer de colon [DLD-1; HT29(ATCC) y HT29(US)] y una línea celular de cáncer mamario (ZR-75) que expresan establemente caveolina-1 (inducción por IPTG). También se usó células de origen embrionario (HEK293T) transfectadas de manera transitoria con caveolina-1. Los resultados indicaron que la presencia de caveolina-1 disminuye los niveles de mRNA y proteicos de COX-2 así como también, la liberación de PGE2. Células HT29(US) que sobreexpresan caveolina-1 no revelaron cambios en los niveles de mRNA de COX-2, pero si se restablece este tipo de regulación cuando se co-expresó E-cadherina junto con caveolina-1. También, en células HEK293T, DLD-1 y ZR75 la presencia de caveolina-1 disminuyó la actividad transcripcional β-catenina-Tcf/Lef, así como la actividad reportera específica de COX-2. La inhibición observada por la presencia de caveolina-1 fue revertida en células incubadas con los inhibidores de la quinasa GSK-3β. Estos resultados indican que caveolina-1 inhibe la expresión de COX-2 por un mecanismo de regulación transcripcional, similar al que fue previamente descrito para survivina.
Adicionalmente, investigamos si la expresión ectópica de COX-2 o la adición de PGE2 al cultivo celular, aumenta la expresión de survivina y si dicho efecto es suficiente para sobrellevar la inhibición de survivina por caveolina-1. Resultados obtenidos por análisis de RT-PCR, Western blot y ensayos de reporteros, revelaron que la sobreexpresión de COX-2 aumenta la expresión de survivina, activa la vía de β-catenina-Tcf/Lef y específicamente aumenta la actividad reportera de plasmidios que contienen elementos de respuesta de la región promotora de COX-2 y survivina. Estos eventos fueron acompañados por una mayor liberación de PGE2. Adicionalmente, PGE2 incrementó los niveles de mRNA y proteicos de survivina y COX-2, así como la actividad de vectores reporteros previamente mencionados, indicando que COX-2 promovería su propia expresión por un mecanismo de retroalimentación positivo.
Estos experimentos revelaron también que la adición de PGE2 previene la inhibición de la expresión de survivina y la proliferación debido a la presencia de caveolina-1, así como en presencia del inhibidor específico de COX-2. Además, la inhibición de COX-2 disminuyó los niveles de PGE2, la actividad transcripcional de β-catenina-Tcf/Lef, los niveles proteicos de survivina y la proliferación celular. En células de cáncer de colon, la inhibición de COX-1 reflejó un efecto similar pero menos pronunciado. Como fue observado en células HEK293T, transfecciones transitorias adicionales de COX-2 en células DLD-1 que expresan o no caveolina-1, sobrellevó los efectos inhibitorios observados por la presencia de caveolina-1, incluyendo la reducción de la expresión de survivina, reducida actividad reportera de β-catenina-Tcf/Lef y del reportero que contiene el promotor de survivina, así como también redujo la producción de PGE2 y la proliferación

10
celular. Además, la presencia de PGE2, en células HT29(ATCC) que expresan caveolina-1, aumenta los niveles nucleares de β-catenina, disminuye la co-localización y la co-inmunoprecipitación de β-catenina con caveolina-1. En conjunto, estos resultados sugieren que el tratamiento con PGE2 previene la formación eficiente del complejo caveolina-1/β-catenina/E-cadherina en la membrana plasmática y por ende permite la translocación nuclear de β-catenina. Finalmente, mediante espectrometría de masas (MALDI-TOF), se comparó proteínas que componen los complejos multiproteicos asociados a caveolina-1 en células HT29(ATCC) y HT29(US) que expresan o no caveolina-1. Los resultados revelaron diferencias sustanciales en la composición de los complejos, algunas de las cuales fueron proteínas involucradas en la degradación proteasomal. En concordancia, se observó que la presencia de caveolina-1 disminuyó sustancialmente la vida media de survivina, sugiriéndose que ocurre a través de un mecanismo post-transcripcional mediado por la degradación vía proteasomal. Interesantemente, dicho efecto fue independiente de la presencia de E-cadherina.
En resumen, este trabajo demuestra que caveolina-1 inhibe la expresión de COX-2 de una manera similar a lo descrito para ciclina D1 y survivina. Además, se identificó un mecanismo de retroalimentación positivo entre COX-2/PGE2 y la expresión de survivina. Este estudio también muestra cómo caveolina-1 o COX-2, inhiben o activan la transcripción dependiente de β-catenina-Tcf/Lef. Por un lado, PGE2 un producto derivado de la actividad enzimática de COX-2 se une a receptores EP2 incrementando la actividad transcripcional de β-catenina-Tcf/Lef, promoviendo la expresión de genes proliferativos como cox-2 y survivina. Por otra parte, caveolina-1 secuestra β-catenina a la membrana plasmática en un complejo multiproteico con E-cadherina, inhibiendo la transcripción de survivina y COX-2 la cual es dependiente de β-catenina-Tcf/Lef. Además, según la evidencia presentada se predice que la producción de PGE2 por alguna célula vecina reducirá la capacidad de caveolina-1 para inhibir la transcripción dependiente de β-catenina-Tcf/Lef y, por ende, su función como supresor de tumores en la misma célula o en células vecinas. Por el contrario, incrementos en la expresión de caveolina-1 podrían modular características de células por la reducción en la producción de PGE2 sugiriendo que alteraciones en la expresión de caveolina-1 en células epiteliales y del estroma podrían ser importantes en el desarrollo tumoral.
Finalmente, este estudio sugiere que caveolina-1 de manera simultánea promueve la degradación de survivina por un mecanismo de regulación post-transcripcional. Por lo tanto, el impacto de caveolina-1 sobre survivina ocurre a diferentes niveles al parecer por mecanismos interconectados. Dada la importancia atribuida a la expresión de survivina en células de cáncer, estas observaciones representan un avance sustancial en el entendimiento de la función de caveolina-1 como supresora de tumores.

11
SUMMARY Cancer is a leading cause of death worldwide. Multiple changes at the molecular
level are associated with the genesis of this disease, which fall into two general categories, referred to as “gain-of-function” or “loss-of-function” alterations in oncogenes or tumor suppressors, respectively. One example of a tumor suppressor that has attracted considerable attention in recent years is caveolin-1, which is considered as such, since its expression is reduced in many types of cancer cells and re-expression, depending on the context and cell type, has been shown to inhibit characteristics associated with the transformed phenotype. Caveolin-1 is an integral membrane protein of 21-24 kDa and constitutes the main component of caveolae or small caves that are vesicular invaginations of the plasma membrane of 50-100 nm in size. Caveolin-1 belongs to a family of proteins comprised of three different isoforms (caveolin-1, 2 and 3). Caveolin-1 and -2 are usually co-expressed in different tissues and cells, while caveolin-3 expression is limited mainly to muscle cells.
In many types of cancer, genes whose transcription is usually inducible, frequently
become constitutively expressed, as is the case for cyclooxygenase-2 (COX-2), an enzyme that catalyzes prostaglandin synthesis (PGs) from arachidonic acid (AA). Particularly, in colon cancer, Prostaglandin E2 (PGE2), increases β-catenin dependent transcriptional activity in association with the transcription factors Tcf/Lef (Tcf, T cell factor; Lef, Lymphoid enhancer factor) and thereby promotes the expression of genes, such as cyclin D1 and survivin. Survivin, an inhibitor of apoptosis protein (IAP) family member, is usually absent in the normal, differentiated tissue, but is overexpressed in cancer cells, where its expression correlates with augmented proliferation and poor patient prognosis.
Caveolin-1 has previously been shown to co-localize at the plasma membrane with
E-cadherin and β-catenin, as well as co-immunoprecipitate with these two proteins. Additionally, caveolin-1 expression precludes β-catenin-Tcf/Lef-dependent transcriptional activity, induced by soluble Wnt or the overexpression of β-catenin. In agreement with these observations, our laboratory demonstrated that caveolin-1 overexpression in HEK293T, ZR75 and DLD-1 cells, repressed survivin expression by a transcriptional mechanism that involves the β-catenin-Tcf/Lef pathway, decreased cell proliferation and promoted cell death. Moreover, such inhibition required the presence of E-cadherin, since metastatic cells HT29(US) cells lacking E-cadherin did not down-regulate survivin but restored this ability upon re-expression of E-cadherin. Alternatively, we have previously shown that caveolin-1 regulates downstream targets by post-transcriptional mechanisms mediated by proteasomal degradation, as is the case for the inducible isoform of nitric oxide synthase (iNOS).

12
The work described here focused on identifying mechanisms by which caveolin-1 regulates the expression/activity of COX-2, as well as studying how alterations in COX-2 expression relate to changes in survivin and cell proliferation. These studies analyzed three colon [DLD-1; HT29(ATCC) and HT29(US)] and a breast (ZR-75) cancer cell line that stably express caveolin-1 (induction by IPTG) and also human embryonic kidney cells (HEK293T) that transiently expressed caveolin-1. The experiments indicated that caveolin-1 presence decreased the mRNA and protein levels of COX-2, as well as the liberation of PGE2. HT29(US) cells that overexpressed caveolin-1 did not reduce COX-2 mRNA levels, but this mode of regulation was restored upon co-expression of E-cadherin with caveolin-1. Also, in HEK293T, DLD-1 and ZR75 cells the presence of caveolin-1 decreased generic β-catenin-Tcf/Lef-dependent, as well as COX-2-specific reporter activity. Suppression by caveolin-1 was reverted in cells incubated with inhibitors of the kinase GSK-3β. These results indicate that caveolin-1 inhibits the expression of COX-2 by a transcriptional mechanism, similar to that previously described for survivin.
Additionally, we investigated whether ectopic expression of COX-2 or addition of
PGE2 to the culture medium augmented survivin expression and whether this sufficed to overcome the observed inhibition of survivin expression by caveolin-1 presence. Results obtained by RT-PCR, Western blot analysis and using reporter assays, revealed that COX-2 overexpression increased survivin expression, activated the β-catenin-Tcf/Lef pathway and specifically increased the activity of reporter constructs that contain responsive elements of the promoter regions of cox-2 and survivin. These events were accompanied by enhanced PGE2 liberation. Additionally, PGE2 was shown to increase the mRNA and protein levels of survivin and COX-2, as well as the activity of the afore-mentioned luciferase reporters, indicating that COX-2 could promote its own expression via a positive feedback loop.
These experiments also revealed that the addition of PGE2 precluded diminished
survivin expression and proliferation observed in the presence of caveolin-1, as well as in the presence of a specific inhibitor for COX-2. Furthermore, COX-2 inhibition decreased PGE2 levels, β-catenin-Tcf/Lef reporter activity, survivin protein levels and cell proliferation. Inhibition of COX-1 had similar but less pronounced effects in colon cancer cells.
As for HEK293T cells, additional transient transfection of DLD-1 cells expressing
caveolin-1 with COX-2, overcame limitations imposed on these cells by the presence of caveolin-1, including reduced survivin mRNA and protein levels, reduced generic β-catenin-Tcf/Lef and survivin reporter activities, as well as reduced PGE2 production and cell proliferation. Furthermore, the presence of PGE2, in HT29(ATCC) cells that express caveolin-1, increased nuclear β-catenin levels, precluded caveolin-1-dependent sequestration of β-catenin to the plasma membrane and decreased the

13
co-immunoprecipitation of β-catenin with caveolin-1. Taken together, these results indicate that PGE2 treatment precludes efficient formation of caveolin-1/β-catenin/E-cadherin complexes at the plasma membrane and in doing so permits nuclear translocation of β-catenin.
Finally, by mass spectrometry (MALDI-TOF) different proteins present in the
multiprotein complex associated with caveolin-1 were compared in HT29(ATCC) and HT29(US) cells expressing or not caveolin-1. The results revealed substantial differences in complex composition, some of which were proteins involved in proteasomal degradation. Accordingly, caveolin-1 was found to promote the degradation of survivin by a post-transcriptional mechanism, since the half life of survivin protein decreased substantially in cells expressing caveolin-1, although this effect did not require E-cadherin presence.
In summary, we demonstrate have that caveolin-1 down-regulates COX-2 in a
manner similar to that described for cyclin D1 and survivin. Furthermore, a feed-forward amplification loop between COX-2/PGE2 and survivin expression was identified. This study reveals how caveolin-1 or COX-2, inhibit or activate, respectively, β-catenin-Tcf/Lef-dependent transcription. On the one hand, PGE2, produced as a consequence of COX-2 activity and released from cells, is suggested to bind to EP2 receptors, increase β-catenin-Tcf/Lef activity and promote the expression of growth-enhancing genes, such as cox-2 and survivin. On the other hand, caveolin-1 sequesters β-catenin to the plasma membrane in a complex with E-cadherin and inhibits β-catenin-Tcf/Lef-dependent transcription of survivin and COX-2. Thus, augmented PGE2 production by any given cell is predicted to reduce the ability of caveolin-1 to inhibit β-catenin-Tcf/Lef-dependent transcription and function as a tumor suppressor in the same cell, as well as in neighboring cells. On the contrary, increments in the expression of caveolin-1 may modulate cell features by reducing PGE2 production suggesting that alterations in caveolin-1 expression in both epithelial and stroma cells are likely to be important in tumor development.
Finally, some preliminary results obtained in this study suggested that caveolin-1
also promoted the degradation of survivin by a post-transcriptional mechanism. Thus, caveolin-1 is able to impact on survivin presence in cells at several different levels that appear interconnected. Given the importance attributed to survivin expression in cancer cells, these observations are likely to represent a substantial advance in our understanding of caveolin-1 function as a tumor suppressor.

14
I. INTRODUCCION 1. Cáncer: antecedentes generales
El cáncer es una de las principales causas de muerte en Chile y en el mundo.
El desarrollo del cáncer esta dado por la transformación de células de un tejido que
es consecuencia directa de alteraciones genéticas y epigenéticas (Ponder, 2001).
En general, genes cuyas mutaciones originan la transformación celular se
denominan oncogenes y otros genes que la impiden, son supresores de tumores.
Actualmente, los procedimientos terapéuticos, farmacológicos y quirúrgicos clásicos
han logrado importantes avances que permiten tratar una enfermedad asociada a
un mal pronóstico. Sin embargo, la eficacia de las terapias convencionales depende
de muchos factores y su éxito se basa principalmente en el diagnóstico temprano.
Por otra parte, los tumores son diversos y heterogéneos, pero las células
cancerosas comparten características como la capacidad de proliferar sin control.
Aberraciones en la regulación de vías de señalización que controlan la proliferación
celular y sobrevida son esenciales para el desarrollo de todos los tumores. En
consecuencia de lo anterior, muchas vías de señalización se vuelven
constitutivamente activas, promoviendo un potencial replicativo ilimitado, evasión a
la apoptosis, insensibilidad frente a señales antiproliferativas, invasión tisular y
metástasis (Hanahan y Weinberg, 2000) (Figura 1).
Insensibilidad a señales antiproliferativas
Insensibilidad a señales antiproliferativas
Independencia a señales de crecimiento
Independencia a señales de crecimiento
Replicación ilimitada
Replicación ilimitada
Replicación ilimitada
Aumento angiogénesis
Aumento angiogénesis
Evadir la apoptosisEvadir la apoptosis
Invasión tisular y metástasis
Invasión tisular y metástasis
Figura 1. Eventos secuenciales que conllevan al desarrollo del cáncer. El siguiente modelo ilustra 6 alteraciones esenciales denominadas eventos principales o “capacidades adquiridas” que explican el desarrollo del cáncer. Dependiendo del tipo de cáncer estos eventos varían en su presencia, cronología y su jerarquía.

15
El punto central es identificar los cambios que ocurren en diversas vías de
señalización en células tumorales y cómo estos cambios podrían explotarse tanto
en el diagnóstico como la terapia del cáncer.
2. Caveolina-1 en la fisiología celular 2.1. Rol de caveolas y caveolina-1
Las caveolas o “pequeñas cuevas” son invaginaciones vesiculares de la
membrana plasmática de 50-100 nm de tamaño y fueron descritas inicialmente por
Yamada en 1955 (Yamada, 1955). Las caveolas han sido consideradas como un
subgrupo dentro de la categoría de los microdominios de la membrana plasmática,
denominados “balsas lipídicas” (Lipid rafts) cuya composición lipídica es rica en
esfingolípidos (esfingomielina y ceramídas) y colesterol. La composición lipídica se
asocia con un mayor grado de organización de los constituyentes lipídicos y con
propiedades fisicoquímicas específicas, tales como una menor densidad e
insolubilidad en detergentes no iónicos, como por ejemplo Tritón X-100, a bajas
temperaturas (Anderson, 1998; Quest et al., 2008).
Posteriormente, Rothberg y colaboradores (1992) describieron a caveolina-1 ó
VIP21 como la primera proteína marcadora de las caveolas (Rothberg et al., 1992;
Murata et al., 1995). Caveolina-1 es una proteína integral de membrana de 21-24
kDa y constituye el principal componente proteico de las caveolas, las que están
presentes en muchos tipos celulares, tales como células endoteliales, adipositos,
fibroblastos, células de músculo liso y estriado (Kogo y Fujimoto, 2000; Liu et al.,
2002). Caveolina-1 pertenece a una familia de proteínas que comprende tres
isoformas diferentes (Caveolina-1, 2 y 3) (Smart et al., 1999).

16
La caveolina-1 y 2 son usualmente co-expresadas en diferentes tejidos y
células, mientras que la expresión de caveolina-3 se limita principalmente a células
musculares (Okamoto et al., 1998). De caveolina-1 se han descrito dos variantes,
caveolina-1α (residuos 1-178) y una más pequeña, caveolina-1β (residuos 32-178),
la que es generada desde el mRNA completo debido a la iniciación alternativa o por
la transcripción de una variante de corte y empalme (Kogo y Fujimoto, 2000; Kogo
et al., 2004). Interesantemente, caveolina-1β se encuentra presente en la parte baja
de las caveolas y en el borde principal de las células migratorias (Scherer et al.,
1995), sugiriendo que caveolina-1α y 1β pueden cumplir distintas funciones. Esto
se confirmó analizando por ejemplo el rol de caveolina-1α y 1β en el desarrollo del
pez cebra (Fang et al., 2006). Además, se ha descrito diferentes variantes de
caveolina-2, sin embargo, no se ha caracterizado su relevancia (Razani et al., 2002;
van Deurs et al., 2003).
2.2. Caveolina-1 en la transformación oncogénica y cáncer
Se ha descrito el cáncer como una enfermedad multifactorial que involucra
pérdida de la capacidad de la célula para responder de manera apropiada frente a
los distintos estímulos de su microambiente. Durante el proceso de tumorigénesis,
diferentes mecanismos contribuyen al desarrollo de un “fenotipo adquirido” el cual
explica las características autónomas de células transformadas (Hanahan y
Weinberg, 2000). Los cambios moleculares que dan cuenta de dicho fenómeno se
explican por alteraciones en la regulación por mecanismos epigenéticos o bien por
mutaciones genéticas de genes esenciales en el control de la homeostasis celular.
En vista de que estos cambios involucran una “ganancia” o “pérdida” de función, las
moléculas involucradas se han clasificado como oncogenes o genes supresores de

17
tumores. Interesantemente, caveolina-1 pertenece a un grupo selecto de proteínas
que poseen un rol dual, es decir dependiendo del contexto celular caveolina-1
puede prevenir o promover el desarrollo tumoral.
Existe clara evidencia que apoya el rol de caveolina-1 en prevenir el desarrollo
tumoral, por ejemplo, se ha descrito que los niveles de mRNA y proteína de
caveolina-1 se encuentran muy disminuídos en fibroblastos NIH3T3 transformados
con diferentes oncogenes (v-Abl, Bcr-Abl, H-Ras (G12V)) (Koleske et al., 1995).
Además, la disminución de la expresión de caveolina-1 mediante el uso de
oligonucleótidos de antisentido es suficiente para promover la transformación de
células NIH3T3 (Galbiati et al., 1998) y la re-expresión de caveolina-1 en células
NIH3T3 transformadas, que carecen de caveolina-1, es suficiente para revertir o
prevenir la transformación celular (Engelman et al., 1997). A su vez, en estudios in
vivo se demostró que ratones nulos para caveolina-1 tratados con carcinógenos
químicos en la piel desarrollan tumores epidermales, con mayor frecuencia que
ratones normales (Capozza et al., 2003). Estos ratones carentes de caveolina-1
tampoco tienen caveolas en los tejidos no musculares, confirmando la necesidad de
la presencia esta proteína en la biogénesis de las caveolas. Específicamente,
nuestro laboratorio demostró que la expresión de caveolina-1 se encuentra
disminuida en muestras tumorales de colon humano y en diferentes líneas celulares
de cáncer de colon (Figura 2A) y que la re-expresión de caveolina-1 en líneas
celulares de adenocarcinoma de colon humano HT29 disminuye la capacidad de
estas células para formar tumores en ratones inmunodeprimidos (Figura 2B,
recopilado desde Bender et al., 2000; Bender et al., 2002).

18
En conjunto, estas evidencias apoyan la idea que caveolina-1 se comporta
como un supresor de tumores, ya que en un gran número de tumores humanos se
han encontrado bajos niveles de expresión de caveolina-1 (Quest et al., 2008) y se
han corroborado los efectos negativos de la re-expresión de caveolina-1 en
diferentes líneas tumorales de colon (Bender et al., 2000), pulmón (Wikman et al.,
2002) y mama (Lee et al., 1998).
Sin embargo, hay otras evidencias indican que caveolina-1 también puede
promover características patológicas como metástasis y multiresistencia a drogas.
Específicamente, en células de cáncer de mama (MCF7) la expresión de
caveolina-1 es marcadamente elevada (Lavie et al., 1998) y caveolina-1 promueve
la sobrevida previniendo la muerte celular por “anoikis” haciendo a las células
independientes de su anclaje (Fiucci et al., 2002), de tal modo que la
norm
al
tumor
Cav-1
Cav-1
actin
norm
al
tumor
Cav-1
Cav-1
actin
A B
Figura 2. Caveolina-1 se encuentra disminuida en muestras tumorales de colon humano y en líneas celulares de cáncer de colon y su re-expresión en células HT29 disminuye su tumorigenicidad in vivo. (A) La expresión de caveolina-1 se encuentra disminuida en muestras tumorales de colon humano (panel superior) así como en diferentes líneas de adenocarcinoma de colon (panel inferior). (B) La re-expresión de caveolina-1 en células de adenocarcinoma de colon humano HT29 causa una disminución en la incidencia y tamaño de tumores formados por estas células en ratones inmunodeprimidos. La foto muestra un ensayo representativo de formación de tumores por células HT29. Flecha verde, tumor formado por células transfectadas con el plasmidio pLacIOP sin caveolina-1 (HT29(US)/M1); flecha negra, ausencia de formación de tumores por células HT29 transfectadas con el plasmidio pLacIOP-caveolina-1 (clon C14 ó C16).

19
sobreexpresión de caveolina-1 se asocia a la promoción de metástasis y a un mal
pronóstico para los pacientes (Garcia et al., 2007).
Sin embargo, este papel dual depende del contexto celular. Esta tesis se ha
enfocado al estudio de mecanismos por los cuales caveolina-1 actúa como un
supresor de tumores.
2.3. Estructura, dominios y señalización intracelular de caveolina-1
Estudios mediante mutagénesis sitio-dirigida han identificado diversas
regiones en la secuencia aminoacídica de caveolina-1 que explican la mayoría de
sus funciones. Se ha definido que los extremos amino y carboxilo terminal distales
están orientados hacia el citoplasma, lo que les permite interactuar con diversas
proteínas citosólicas (Figura 3A) (Williams y Lisanti, 2005).
La caveolina-1 posee un dominio central hidrofóbico anclado a la membrana
plasmática (residuos 102-134), un dominio aminoterminal implicado en la formación
de oligómeros (residuos1-101) y en su dominio carboxiloterminal, los residuos
135-178 funcionan como puente entre los distintos oligómeros (Okamoto et al.,
1998; Smart et al., 1999).
Además, en el extremo aminoterminal, entre los residuos 82-101, se ubica el
dominio de andamiaje de caveolina-1 o CSD (Caveolin-1 Scaffolding Domain), el
que se une a moléculas que presentan “dominios de unión a caveolina-1” o CBD
(Caveolin-1 Binding Domain) (Figura 3B) (Okamoto et al., 1998; Smart et al., 1999;
Cohen et al., 2004).

20
Este último fue definido como un segmento que muestra la siguiente
secuencia: φxφx4φ o φx4φx2φ, donde “φ” es un aminoácido aromático (W, F o Y) y “x”
cualquier otro aminoácido (Couet et al., 1997). Por otra parte, la región que
comprende los residuos 98-132 contiene una secuencia similar al dominio WW o
WWP (Sudol et al., 1995).
Este dominio corresponde a una secuencia de aproximadamente 40
aminoácidos, muy conservada en una gran variedad de proteínas reguladoras,
citoesqueléticas y de señalización y su nombre deriva de que está principalmente
compuesto por dos triptófanos separados por 20-23 aminoácidos y una prolina
carboxiloterminal. Posee un plegamiento caracterizado por una hoja β estable de
tres hebras y sus ligandos contienen secuencia ricas en prolina, fosfoserina o
fosfotreonina, dentro de una secuencia de consenso “PPXY” (Einbond y Sudol,
1996).
N
C
N
C
Medio
N
C
N
C
N
C
N
C
MedioMembrana plasmática
Colesterol y esfingomielina
Monómero de caveolina-1Secuencia de inserción en membrana (residuos 102-134)Secuencia de unión a membranaDominio de andamio de caveolina (residuos 82-101)Grupos palmitoilo (133, 143, 156)Secuencia de oligomerización (residuos 61-161)
CAVEOLINA-1
A B
Figura 3. Estructura y dominios de caveolina-1. (A) El esquema muestra caveolina-1 insertada en microdominios de la membrana plasmática ricos en esfingolípidos y colesterol (en amarillo y rojo). Dichas estructuras de membrana, denominadas caveolas, pueden variar en su morfología desde ensamblajes planares hasta las típicas invaginaciones de la membrana de 50-100 nm, cuya formación depende de la presencia de caveolina-1 (Smart et al., 1999) y una proteína denominada cavina (Hill et al., 2008). (B) Caveolina-1 presenta en su estructura una serie de dominios necesarios para la inserción a membrana, oligomerización e interacción con otras proteínas (Cohen et al., 2004).

21
Se ha descrito un dominio tipo-WW en caveolina-3 que se une con la
secuencia 32PPAY35 del extremo carboxiloterminal de β-distroglicano, bloqueando la
interacción de distrofina con β-distroglicano en células musculares y promoviendo
de tal manera su degradación por el proteasoma (Sotgia et al., 2000; Ilsley et al.,
2002). Además, se ha visto que caveolina-1 es capaz de promover la degradación
de proteínas como iNOS vía el proteasoma, posiblemente a través de la interacción
con la región 101-135 de caveolina-1 donde existe un dominio putativo tipo WW
(Felley-Bosco et al., 2000; Felley-Bosco et al., 2002)
Existe una amplia variedad de proteínas reguladas por su interacción con
caveolina-1 (Cohen et al., 2004; Quest et al., 2004; Williams y Lisanti, 2005; Quest
et al., 2008). Por ejemplo, se ha determinado que muchas de las proteínas que
interactúan con caveolina-1, lo hacen a través de uno o más CBD presentes en su
estructura y se ha observado que, generalmente la unión al dominio de andamiaje
de caveolina-1 inhibe su actividad (Couet et al., 1997). Así, se ha propuesto dos
mecanismos de inhibición donde: (i) caveolina-1 interactúa con un CBD que es
parte del sitio activo de la proteína blanco o (ii) caveolina-1 interactúa con el CBD de
la proteína en su conformación inactiva.
3. Vías de señalización mediada por β-catenina y su papel en el cáncer
De las muchas vías de señalización relacionadas con el cáncer, existe una
particularmente importante para los objetivos de esta tesis, la vía de Wnt, que está
alterada en células de cáncer de colon y tiene a β-catenina como proteína efectora
clave (Gumbiner, 1995).
β-catenina fue inicialmente identificada por su asociación con moléculas de
adhesión como E-cadherina (Potter et al., 1999). No obstante, β-catenina también

22
posee una función transactivadora de la transcripción de genes, en su mayoría,
involucrados en proliferación y sobrevivencia celular (Eastman y Grosschedl, 1999;
Waterman, 2004).
La actividad transcripcional de β-catenina está mediada por su interacción con
factores de transcripción de la familia Tcf/Lef (Tcf, T cell factor; Lef, Lymphoid
enhancer factor) (ver figura 4) (Waterman, 2004). β-catenina provee los dominios
necesarios para el reclutamiento de las proteínas involucradas en la activación
transcripcional, mientras que los factores Tcf/Lef son proteínas de unión al DNA
secuencia-específicos (Eastman y Grosschedl, 1999). Por otro lado, se ha
observado que la proteína APC (adenomatous polyposis coli) interactúa con
β-catenina, en un complejo multiproteico con axina y con la proteína quinasa
GSK-3β regulando su degradación en el citosol. En este complejo, la quinasa
GSK-3β fosforila a β-catenina promoviendo su ubiquitinación y posterior
degradación por el proteasoma (Aberle et al., 1997; Salomon et al., 1997).
Asimismo, la inhibición del proteasoma o la sobreexpresión de Wnt-1 evita la
ubiquitinación de β-catenina y en consecuencia no es degradada por el proteasoma
(Aberle et al., 1997). A su vez, la sobreexpresión de β-catenina es suficiente para la
activación transcripcional de los factores Tcf/Lef (Hsu et al., 1998).
Cabe destacar que en células de cáncer de colon esta vía se encuentra
alterada debido a mutaciones tanto en la proteína APC como en β-catenina,
observándose como resultado final que la degradación de β-catenina no ocurre.
Por el contrario, se acumula en el citoplasma y luego transloca al núcleo
donde, en asociación con los factores Tcf/Lef (Rubinfeld et al., 1993; Rubinfeld et
al., 1996), promueve la expresión de sus genes blancos entre los cuales se
encuentra ciclina D1, c-myc, VEGF y mas recientemente descritos, cox-2 y survivina

23
(ver figura 4) (Haertel-Wiesmann et al., 2000; Giles et al., 2003; Logan y Nusse,
2004; He et al., 2005).
En un estudio realizado por Galbiati et al. (2000), se demostró que caveolina-1
co-localiza con E-cadherina, β-catenina y γ-catenina en la membrana plasmática,
además, co-inmunoprecipita con estas proteínas en extractos de membranas
enriquecidas en caveolas. Adicionalmente, caveolina-1 reprime eficientemente la
actividad transcripcional Tcf/Lef dependiente de β-catenina, inducida por la proteína
secretable Wnt o bien por la sobreexpresión de β-catenina.
En este estudio se demuestra también, que la re-expresión de caveolina-1 en
células que no la expresan, promueve el reclutamiento de β-catenina hacia la
membrana plasmática (Galbiati et al., 2000). Sin embargo, en estos experimentos
no se entrega información sobre la naturaleza de la interacción entre β-catenina y
caveolina-1. Los resultados indicaron más bien que no existe una interacción directa
entre ambas proteínas (Galbiati et al., 2000). A pesar de esta limitación, los
resultados sugirieron que la presencia de caveolina-1 podría reducir la
disponibilidad de β-catenina en el citosol y por lo tanto su capacidad de promover la
transcripción.
4. Relación entre COX-2, survivina y caveolina-1: papel en carcinogénesis
4.1. Rol de COX-2 en cáncer de colon
Las ciclooxigenasas (COXs) catalizan la conversión del ácido araquidónico a
PGH2, el que es procesado a PGE2 (Prostaglandina E2), PGI2, PGD2 o tromboxano
A2 por sintasas específicas (Smith et al., 2000; Wang et al., 2007).

24
Dos genes designados como cox-1 y cox-2 codifican para las respectivas
COXs (Kanaoka et al., 2007). Ambas isoenzimas exhiben propiedades enzimáticas
similares (William et al., 1996). Sin embargo, la expresión de COX-1 es constitutiva
en la mayoría de los tejidos, mientras que la expresión de COX-2 es inducida por
factores de crecimiento, mediadores de la inflamación y carcinógenos (Telliez et al.,
2006; Kanaoka et al., 2007).
Así, esta expresión inducida de COX-2 se transforma en constitutiva en varios
tipos de cáncer humano, incluyendo los de colon, mama, páncreas, pulmón y piel
(Sano et al., 1995; Tucker et al., 1999; Soslow et al., 2000). De éstos, el más
estudiado en relación a COX-2 es el cáncer colorrectal (Sinicrope y Gill, 2004;
Telliez et al., 2006; Liao et al., 2007).
Figura 4: Vía de señalización mediada por β-catenina. 1) Las proteínas α-catenina y β-catenina se asocian con moléculas de adhesión, como E-cadherina y participan con el citoesqueleto en funciones de adhesión celular. 2) En el citoplasma, β-catenina interactúa con las proteínas APC, axina y la quinasa GSK-3β. La quinasa GSK-3β fosforila a las proteínas APC, axina y β-catenina. La fosforilación de β-catenina por la quinasa GSK-3β presente en este complejo multiprotéico, produce la ubiquitinización y posterior degradación de β-catenina
GSK3βPP
1
4
2
3
GSK3βPP
11
44
22
33
3) Por otra parte, β-catenina se transloca al núcleo en asociación con los factores de transcripción Tcf/Lef promoviendo la expresión de genes involucrados en la proliferación celular, como por ejemplo: cox-2, survivina y ciclina D1. 4) La unión de Wnt a su receptor, a través de diversas vías de señalización, inactiva a la quinasa GSK-3β y por lo tanto, β-catenina aumenta sus niveles citoplasmáticos, debido a que no es degradada. En cáncer de colon, mutaciones de la proteína APC, aumentan los niveles citoplasmáticos de β-catenina y en consecuencia, células que poseen la proteína APC mutada poseen una elevada actividad transcripcional Tcf/Lef dependiente de β-catenina.
E-cadherina
ciclina D1 cox-2 survivina
Degradación proteasomal

25
En cáncer de colon los niveles de mRNA y proteicos de COX-2 se encuentran
significativamente elevados (DuBois et al., 1996; Kutchera et al., 1996). Los niveles
elevados del mRNA de COX-2 se deben a mecanismos transcripcionales y/o post-
transcripcionales. Por ejemplo, a nivel de regulación post-transcripcional se ha visto
que β-catenina se une a elementos ricos en AU ubicados en el 3´-UTR del mRNA
de COX-2 y de esta forma permite su estabilización (Dixon, 2004).
En general, se piensa que las proteínas COXs una vez expresadas, están
constitutivamente activas asumiendo que todos los cofactores requeridos están
presentes (Telliez et al., 2006; Wang et al., 2007). Además, los niveles elevados de
COX-2 se han asociado directamente con una producción aumentada de PGE2,
proliferación celular elevada y tumorigénesis (Hla et al., 1999; Tong et al., 2005). En
células humanas de cáncer colorectal, una expresión elevada de COX-2 está
asociada con el progreso del cáncer (Kanaoka et al., 2007) y conlleva a cambios
fenotípicos que promueven la metástasis (Tsujii et al., 1997).
Se piensa que una expresión elevada de COX-2 promueve la formación de
tumores, lo que es una consecuencia de las múltiples acciones de COX-2 en la
dinámica celular y que incluyen principalmente: i) promover la proliferación celular
(Tsujii et al., 1997; Sheng et al., 2001; Pai et al., 2003); ii) inhibir la apoptosis a
través de la generación de un producto anti-apoptótico, PGE2, la que aumenta los
niveles de BCL-2 (Sheng et al., 1998) y también a través de la remoción de un
sustrato pro-apoptótico, el ácido araquidónico (Cao y Prescott, 2002) el que debido
a debido a niveles elevados de ceramidas induce la muerte celular (Chan et al.,
1998; Ilsley et al., 2005); iii) inhibir la vigilancia inmune, ya que varias
prostaglandinas, principalmente PGE2, tienen efectos inmunosupresores (Tsujii et
al., 1997; Pockaj et al., 2004) y iv) aumentar la movilidad celular y adhesión, puesto

26
que la activación constitutiva de COX-2 conduce a una expresión aumentada de
metaloproteinasa-2 y reduce la expresión de E-cadherina, lo que promueve la
invasión celular y metástasis (Tsujii y DuBois, 1995; Tsujii et al., 1997).
En un principio, se determinó indirectamente la participación de COX-2 en
cáncer y mediante estudios epidemiológicos se demostró una reducción de un
40-50% en la incidencia de cáncer colorectal en personas con tratamiento de
drogas antiinflamatorias no esteroidales o NSAID (Chell et al., 2005; Cha y DuBois,
2007), así como también en modelos experimentales animales de formación de
tumores en el colon (Kawamori et al., 1998; Reddy et al., 2000; Rao et al., 2002).
El uso de NSAIDs específicos para COX-2 promueve la apoptosis en células
tumorales (Jendrossek et al., 2003; Swamy et al., 2004) y por lo tanto, han revelado
efectos benéficos para la prevención del desarrollo tumoral (Warner y Mitchell,
2004). Actualmente, algunos NSAIDs específicos para COX-2 que se han
desarrollado son: lumiracoxib, rofecoxib, etoricoxib y celecoxib (Warner y Mitchell,
2004; Cha y DuBois, 2007).
Sin embargo, aún no es claro si el mecanismo por el cual estos fármacos
median su efecto antitumoral sea sólo a través de la inhibición de COX-2, ya que el
uso de dimetil-celecoxib, una molécula que no inhibe la actividad catalítica de
COX-2, posee propiedades antitumorales, independiente de la actividad de COX-2
(Schonthal et al., 2008).

27
4.2. Survivina en cáncer de colon y su regulación negativa por la expresión de
caveolina-1 a través de la vía β-catenina-Tcf/Lef
Survivina, es una proteína pequeña de 16,4 kDa que pertenece a la familia de
las proteínas inhibidoras de la apoptosis (IAP) (Verhagen et al., 2001), posee un
papel anti-apoptótico posiblemente a través de la inhibición de varios tipos de
caspasas (Shin et al., 2001). Sin embargo, existen otros mecanismos mas
aceptados que explican su capacidad de funcional como una proteína anti-
apoptótica (Altieri, 2008) y su característica principal es que se expresa en etapas
tempranas del desarrollo, estando prácticamente ausente en la mayoría de los
tejidos diferenciados adultos. Sin embargo, se expresa fuertemente en un gran
número de tipos de cáncer humanos (Ambrosini et al., 1997; Velculescu et al.,
1999). De hecho, survivina fue identificada como uno de los cuatro transcritos más
expresados en tumores humanos, no así en células normales del mismo tejido
(Velculescu et al., 1999).
En un estudio in vivo, la mutación de survivina (T34A) suprimió el crecimiento
tumoral de novo y redujo el crecimiento de tumores establecidos (Grossman et al.,
2001). También la expresión de survivina ha sido inhibida con ribozimas (Pennati et
al., 2004) y oligonucleótidos de antisentido, demostrando la función anti-apoptótica
de esta proteína (Chen et al., 2000). La función anti-apoptótica de survivina parece
ser un prerrequisito para la sobrevivencia tumoral, lo que explicaría la alta
prevalencia de esta proteína en distintos tipos de tumores humanos, entre ellos el
carcinoma de colon (Kim et al., 2003). Específicamente en cáncer de colon, la
sobreexpresión de survivina se debe en parte a una elevada actividad
transcripcional de β-catenina-Tcf/Lef, ya que el promotor de survivina contiene tres
sitios de unión TBE (Tcf/Lef-binding elements) (Kim et al., 2003). A su vez, se ha

28
descrito la co-localización de COX-2/β-catenina/survivina en muestras tumorales de
cáncer de colon, sugiriéndose un posible efecto regulatorio de COX-2/β-catenina
sobre la expresión de survivina (Mori et al., 2007).
Por otra parte, y en concordancia con el papel supresor de tumores de
caveolina-1 (Quest et al., 2004; Williams y Lisanti, 2005; Quest et al., 2008), la
sobreexpresión de ésta en células HEK293T, ZR75 y DLD-1, reprime la expresión
de survivina, por un mecanismo transcripcional que involucra la vía de β-catenina-
Tcf/Lef, disminuye la proliferación y promueve la muerte celular (Torres et al., 2006).
Además, en un estudio reciente de nuestro laboratorio se demostró también que la
regulación negativa de survivina a través de la vía de β-catenina-Tcf/Lef por
caveolina-1 requiere la presencia de E-cadherina en células HT29(US) (Torres et
al., 2007). Dicha inhibición de la expresión de survivina requiere el reclutamiento de
β-catenina a la membrana plasmática, en donde forma un complejo multiproteico
con E-cadherina (Torres et al., 2007).
Importantemente, los niveles proteicos de survivina son también regulados por
el proteasoma (Zhao et al., 2000; Vong et al., 2005) y el tratamiento con NSAIDs
específicos para COX-2, promueven la degradación de survivina por este mismo
camino (Chiou y Mandayam, 2007; Gaiser et al., 2008). En concordancia, existe
también la evidencia que la unión de caveolina-1 a la iNOS, por medio de su
dominio tipo-WW putativo, promueve la degradación de esta enzima por el
proteasoma (Felley-Bosco et al., 2000; Felley-Bosco et al., 2002). Esto sumado a
que la presencia de caveolina-1 reprime la expresión de survivina, por un
mecanismo transcripcional (Torres et al., 2006), surge la interrogante si, de manera
adicional, caveolina-1 podría promover también la degradación de survivina vía el
proteasoma.

29
4.3. PGE2 activa la vía deβ-catenina-Tcf/Lef
Como fue mencionado anteriormente, PGE2 es un producto generado a través
de la actividad enzimática de COX-2 que aumenta el crecimiento de células de
cáncer de colon (Shao et al., 2003) y también la viabilidad (Sheng et al., 1998; Shao
et al., 2003). Interesantemente, la unión de PGE2 al receptor EP2, inhibe la
regulación negativa de β-catenina por la quinasa GSK-3β.
Un mecanismo descrito por Castellone y colaboradores (2005) demuestra que
la PGE2 activa el receptor EP2, que se ubica en la membrana plasmática asociado
con proteínas heterotrímericas Gαs. La activación por PGE2 promueve la unión de la
proteína axina con Gαs, mientras que los heterodímeros Gβγ activan la vía de
PI3K-Akt, la cual fosforila e inactiva la quinasa GSK-3β. En consecuencia, la
quinasa GSK-3β es desplazada del complejo axina/APC/β-catenina y por ende
aumentan los niveles proteicos de β-catenina, así como también la actividad
transcripcional de β-catenina-Tcf/Lef (Figura 5) (Castellone et al., 2005; Shao et al.,
2005).
Figura 5: PGE2 activa la vía de β-catenina-Tcf/Lef. La sobreexpresión de COX-2 en cáncer de colon conlleva a una mayor producción de PGE2. A su vez, PGE2 activa al receptor EP2 promoviendo la unión de la proteína axina con la subunidad Gαs, desplazando la quinasa GSK-3β del complejo. Concomitantemente, los heterodimeros Gβγ activan la vía de PI3K-PDK1-Akt y Akt fosforila e inactiva la quinasa GSK-3β. En resumen, la quinasa GSK-3β es desplazada del complejo axina/ APC/β-catenina y por ende aumentan los niveles proteicos de β-catenina, así como también la actividad transcripcional de β-catenina-Tcf/Lef, aumentando la expresión de genes que promueven el crecimiento y desarrollo tumoral (Tomado de Castellone et al., 2005).

30
Adicionalmente, se ha descrito que en células de cáncer de pulmón la
expresión de survivina depende de la actividad de COX-2 (Krysan et al., 2004a;
Krysan et al., 2004b) y que la expresión de COX-2 es crítica para la resistencia a la
apoptosis dado que la PGE2 disminuye la degradación de survivina por el
proteasoma (Krysan et al., 2003; Krysan et al., 2004c).
Además, se ha visto que PGE2 aumenta la expresión de survivina en células
dendríticas las cuales se hacen más resistentes a estímulos apoptóticos comparado
con células controles (Baratelli et al., 2005). Considerando que survivina es
regulada por la vía de β-catenina-Tcf/Lef y que también podría ser positivamente
regulada por PGE2, en esta tesis se investigó el efecto de PGE2 en células humanas
embrionarias de riñón (HEK293T), de cáncer de colon (DLD-1, HT29) y de mama
(ZR75).
En resumen, la evidencia presentada sugiere que: 1) La expresión alterada de
COX-2 actúa en la carcinogénesis de células del colon y que la deficiencia de la
actividad enzimática de COX-2 a través del uso de inhibidores específicos puede
inhibir la formación de tumores o causar eventualmente su regresión. 2) COX-2 a
través de la generación de PGE2, aumenta la actividad de β-catenina-Tcf/Lef y
posiblemente aumenta la expresión de survivina. 3) Caveolina-1 podría inhibir la
expresión de COX-2 por un mecanismo transcripcional dependiente de β-catenina-
Tcf/Lef. 4) Caveolina-1 podría, al reducir el nivel de expresión de COX-2, también
reducir la liberación de PGE2 y a su vez PGE2 promovería la estimulación de la
transcripción de COX-2 como la de survivina a través de un circuito de
retroalimentación positivo. 5) Caveolina-1 podría modular los niveles de survivina a
través de un mecanismo post-traduccional, promoviendo la degradación de la
proteína. Estos antecedentes han llevado a la formulación de la hipótesis de trabajo.

31
II. HIPOTESIS
“Caveolina-1 reduce la proliferación y sobrevivencia celular al disminuir la
expresión de ciclooxigenasa-2 y survivina por un mecanismo transcripcional y/o
post-traduccional”
Para desarrollar esta hipótesis de trabajo se ha formulado un objetivo general
y tres objetivos específicos.
III. OBJETIVO GENERAL
Estudiar la regulación transcripcional y post-traduccional de caveolina-1 sobre
la ciclooxigenasa-2 (COX-2) ver cómo este proceso afecta la regulación de survivina.
Finalmente, se evaluará el papel de estos procesos en la modulación de la
proliferación.
IV. OBJETIVOS ESPECIFICOS:
1.- Analizar el efecto de la expresión ectópica de caveolina-1 sobre la regulación
transcripcional de COX-2.
2.- Estudiar el mecanismo de regulación transcripcional de survivina por la expresión
de COX-2 y su nexo con la regulación negativa en la proliferación celular por
caveolina-1.
3.- Analizar el efecto de la expresión ectópica de caveolina-1 sobre la regulación
post-traduccional de survivina.

32
V. METODOLOGIA
1. Materiales
Sustrato de quimioluminiscencia EZ-ECL (Biological Indrustries, Kibbutz Beit
Haemek, Israel), reactivo BCA (Pierce, Rockford, IL, USA), reactivo de transfección
Superfect (#301305) y kit de extracción de DNA Midi Kit (#12143) (Qiagen,
Valencia, CA, USA), reactivo TriZOL® (Invitrogen, Life Technologies, Carlsbad, CA,
USA), antibiótico higromicina y Prostaglandina E2 (PGE2) (Calbiochem, La Jolla,
CA, USA), reactivo IPTG (Isopropyl β-D-1-thiogalactopyranoside) (USBiological,
Swampscott, MA, USA). Todos los medios de cultivo y antibióticos usados
obtenidos de Gibco BRL (Paisley, Scotland, UK). Suero fetal bovino (FBS)
(HyClone, Logan, Utah, USA). Kit para ensayos de proliferación MTS® (Promega,
Madison, WI, USA). Para los ensayos de actividad luciferasa, los reactivos luciferina
y 2 nitrofenil-β-D-galactopiranósido (ONPG) adquiridos de USBiological
(Swampscott, MA, USA) y Boehringer Mannheim (Mannheim, Alemania),
respectivamente.
Respecto a los inhibidores utilizados, para la quinasa GSK-3β se utilizó el
inhibidor específico, SB-216763 de Sigma (St. Louis, MO, USA) y para inhibir
específicamente COX-1 ó COX-2, se utilizaron los inhibidores FR122047 y SC-791
(Calbiochem, La Jolla, CA, USA). La proteína A-sefarosa usada en ensayos de
inmunoprecipitación y todos los reactivos usados para inmunofluorescencia y
Western blots se adquirieron de Sigma (St. Louis, MO, USA) o de la mejor calidad
disponible.

33
2. Anticuerpos
Se utilizó anticuerpos primarios policlonales anti-caveolina-1 (#C3237,
Sigma, St. Louis, MO, USA) y (#610060, Transduction Laboratories, Lexington, KY,
USA). Los anticuerpos monoclonales anti-caveolina-1 (#C13620), anti-COX-2
(#610204), anti-β-catenina (#610154) y anti-E-cadherina (#610181) fueron
obtenidos de Transduction Laboratories (Lexington, KY, USA). El anticuerpo
monoclonal anti-GAPDH (Ambion, Austin, TX USA) y los anticuerpos policlonales
anti-survivina humana (#AF886) y anti-actina (#A5060) eran de RD Systems
(Minneapolis, MN, USA) y Sigma (St. Louis, MO, USA), respectivamente. Los
anticuerpos secundarios (caprinos) acoplados a peroxidasa de rabanito (HRPO),
anti-IgG de conejo (#1706515) y anti-IgG de ratón (#A4416) eran de Bio-Rad
Laboratories (Hercules, CA, USA) y Sigma, respectivamente. Los anticuerpos
secundarios (caprinos) anti-IgG de ratón conjugado a Cy3TM (#115-165-068) y
anti-IgG de conejo conjugado a FITC (#111-0950144) se obtuvieron de Jackson
ImmunoResearch Laboratories (West Grove, PA, USA).
3. Cultivo celular y transfecciones
3.1 Cultivo líneas celulares
La línea celular derivada de epitelio renal humano embrionario, HEK293T
(ATCC, CRL-11268), la línea celular de cáncer mama (ZR75) y células de
adenocarcinoma de colon humano DLD-1, HT29 (ATCC, HTB-38) [HT29(ATCC)] y
HT29(US) se cultivaron en medio DMEM rico en glucosa suplementado con FBS al
10% y antibióticos (10.000 U/ml de penicilina y estreptomicina 10 µg/ml) a 37ºC y
CO2 al 5%. Las células HT29(US) fueron seleccionadas desde HT29(ATCC) por su
mayor potencial metastásico a través de repetidos pasajes en ratones

34
inmunosuprimidos, en donde las células fueron inyectadas dorsalmente y luego
recuperadas de los respectivos pulmones metastáticos (Bender et al., 2000). Todas
las líneas celulares fueron sub-cultivadas cada 3-5 días por tripsinización de 2 a 5
min, permitiendo entre cada pasaje un máximo de 80% de confluencia.
El uso de cada línea celular y todas las transfectantes derivadas de estas
células fueron utilizadas entre 15-45 pasajes (ciclos de sub-cultivo). La tabla 1
muestra un resumen de las principales características de las líneas celulares
usadas comparado con células normales.
Tabla 1: Comparación y características de las líneas celulares versus una célula normal.
3.2 Plasmidios.
Los plasmidios pLacIOP fueron descritos previamente en (Bender et al.,
2000). Brevemente, el plasmidio pLacIOP-caveolina-1 expresa caveolina-1 inducida
por IPTG y permite la selección de células debido a que incorpora resistencia a
higromicina. El plasmidio pEGFP-caveolina-1, se generó subclonando el cDNA
completo de caveolina-1 el que fue extraído de pGEX2T (Felley-Bosco et al., 2000)
y situado en el marco de lectura del vector pEGFP-C1 (Clontech, Palo Alto, CA,
USA) mediante el uso de las enzimas BglII-KpnI como fuera descrito previamente
(Torres et al., 2006).

35
El plasmidio pOSML-COX-2 que codifica para la COX-2 silvestre fue
gentilmente donado por el Dr. Tim Hla (Center for Vascular Biology, University of
Connecticut Health Center, Farmington). El plasmidio reportero pTOP-FLASH
(Figura 6) contiene el promotor de c-Jun con tres sitios de unión Tcf/Lef y el
plasmidio control pFOP-FLASH contiene los tres sitios de unión a Tcf/Lef mutados.
Estos plasmidios fueron descritos previamente (van de Wetering et al., 1997) y
fueron generosamente donados por el Dr. Hans Clevers (Hubrecht Laboratory,
Uppsalalaan, The Netherlands). Los plasmidios reporteros para cuantificar la
actividad transcripcional del promotor de survivina (pLuc-1710 y pLuc-420-3M)
fueron generosamente donados por el Dr. Dario Altieri y están descritos
previamente por su grupo de trabajo (Kim et al., 2003). Este plasmidio pLuc-1710
corresponde a un reportero con los 3 TBEs y en el caso de pLuc-420-3M se eliminó
un TBE y otro TBE se encuentra mutado (ver detalle de plasmidios en figura 6).
El plasmidio de expresión de la enzima β-galactosidasa (pON) fue generosamente
donado por el Dr. Sergio Lavandero (Universidad de Chile, Santiago, Chile) y fue
utilizado para la normalización de los experimentos con reporteros. Los plasmidios
reporteros para medir la actividad transcripcional del promotor de cox-2,
pGL3-COX-2 (contiene el promotor completo con un sitio de unión a Tcf/Lef) o
pGL3 (contiene solo el gen de la luciferasa) fueron donados por el Dr. Ying Zhu
(Laboratory of Virology, Wuhan University, Wuhan, China).
En esta tesis todos los plasmidios fueron purificados mediante el sistema
comercial de extracción de DNA Midi Kit de Qiagen (Valencia, CA, USA).

36
3.3. Expresión ectópica de caveolina-1, COX-2 y plasmidios reporteros
Las diferentes líneas celulares usadas en este estudio se transfectaron de
acuerdo a las instrucciones entregadas por el fabricante de un dendrímero
fracturado, llamado Superfect® (Qiagen). El protocolo se ajustó para realizar
transfecciones con diluciones de 1-4 μg de cada plasmidio en 100 μl de medio
DMEM sin suero. Luego, se agregó 10 μl del reactivo Superfect® y se agitó
suavemente con una micropipeta.
Se dejó la mezcla durante 15-20 min para dar paso a la formación del
complejo dendrímero-DNA y posteriormente este complejo fue diluido por la adición
de 600 μl de DMEM suplementado con suero fetal bovino al 10% y antibióticos.
Figura 6. Sistema de reporteros TOP/FOP-FLASH y reporteros basados en el promotor de survivina y de COX-2. (A) Sistema reportero a Tcf/Lef. Se esquematiza el plasmidio reportero para Tcf/Lef, que codifica para el gen de la luciferasa precedido por un promotor con 3 sitios de unión a Tcf/Lef en tándem (pTOP-FLASH). El plasmidio control (pFOP-FLASH) contiene mutaciones puntuales en los sitios de unión a Tcf/Lef. La secuencia consenso de unión para Tcf/Lef es esquematizada en el panel superior (pTOP-FLASH) y los sitios mutados son indicados en rojo (pFOP-FLASH). (B) Sistema de reporteros basadas en el promotor de survivina. Se esquematiza el promotor del gen de survivina humano, que contiene en su secuencia 3 sitios consenso de unión a Tcf/Lef (TBE). Las 2 construcciones de plasmidios reporteros (actividad luciferasa) usados para determinar la actividad transcripcional del promotor de survivina fueron: pLuc-1710 y pLuc-420-3M. El plasmidio pLuc1710 corresponde a un reportero con los 3 TBEs. En el caso de pLuc-420-3M se eliminó un TBE y el otro TBE, responsable de la activación, se encuentra mutado (Kim et al., 2003). (C) Sistema de reporteros basadas en el promotor de cox-2. Se esquematiza el promotor completo del gen de cox-2, el cual posee un sitio TBE. El plasmidio pGL3-COX-2 tiene el promotor y bajo este control expresa el gen de la luciferasa. En cambio, el plasmidio pGL3 (control vacío) no contiene al promotor, pero si el gen de la luciferasa.
A
TBE LucTBETBE
TBE LucTBETBE
TBE LucTBETBE
TBE LucTBETBE
TBE LucTBE Luc
TOP-FLASH
FOP-FLASH
p-GL3-COX-2
p-GL3
B C

37
La mezcla resultante se agitó suavemente con una micropipeta y se agregó
sobre las células previamente lavadas con PBS. La incubación de células con el
complejo varió de entre 2-3 h para células HEK293T y 4-5 h para el resto de las
líneas celulares (ZR75, DLD-1 y HT29). Cumplido el tiempo de incubación, en
células HEK293T se reemplazó la mezcla por medio de cultivo completo (2 ml) o
bien para células de cáncer sólo fue adicionado 1 ml del medio de cultivo completo
conteniendo IPTG 2 mM (concentración final 1 mM) y según el experimento las
células se cultivaron durante 24 ó 48 h.
3.3.1. Expresión ectópica de caveolina-1 y/o COX-2 en células HEK293T
Se sembró 5x105 células en placas de 6 pocillos y se crecieron por 24 h,
alcanzando un 40-60% de confluencia. Luego, las células fueron transfectadas o
bien fueron co-transfectadas con 2 μg (excepto donde se indica) de cada plasmidio.
Para la expresión ectópica de caveolina-1 se utilizó dos tipos vectores diferentes:
pLacIOP, pLacIOP-caveolina-1 o bien pEGFP-C1, pEGFP-caveolina-1 (genera una
proteína de fusión GFP-caveolina-1) y para la expresión de COX-2, pOSML-COX-2
(0-4 μg). Finalmente, 24 h post-transfección, se obtuvo extractos según cada
procedimiento experimental llevando a cabo el análisis de reporteros (ver sección 6
de metodología), Western blot, RT-PCR o bien ensayos de inmunoprecipitación.
3.3.2. Expresión de caveolina-1 y/o COX-2 en células de cáncer
La transfección y obtención de clones a partir de las líneas celulares de
adenocarcinoma de colon humano DLD-1 y la sub-línea celular metastásica
HT29(US) ha sido descrita previamente en nuestro laboratorio (Bender et al., 2000),
siendo la expresión de caveolina-1 de manera inducible por IPTG.

38
En esta tesis, se utilizaron los clones derivados de éstas, DLD-1/M1 (control
mock, pLacIOP), DLD1/C4 (pLacIOP-caveolina-1), HT29(US)/M1 (control mock,
pLacIOP) HT29(US)/C14 (pLacIOP-caveolina-1) (ver tabla 1). En todos los
experimentos se indujo la expresión de caveolina-1 agregando IPTG 1 mM al medio
de cultivo durante 24 h. La expresión de COX-2 y análisis de reporteros en células
DLD-1, se llevó a cabo utilizado el mismo protocolo descrito previamente, con la
salvedad que las células DLD-1 M1/C4 fueron co-transfectadas con 2 μg del vector
pOSML-COX-2 con pTOP-FLASH o pFOP-FLASH incubando con el complejo
dendrímero-DNA durante 5 h. Luego, se adicionó 1 ml del medio de cultivo
completo con IPTG 2 mM (concentración final 1 mM) incubándose durante 24 h
hasta posterior ensayos de la actividad luciferasa (ver más adelante), Western blot
o RT-PCR. La línea celular de adenocarcinoma de colon humano HT29 de
American Type Cell Culture, HT29(ATCC) y la línea celular de cáncer de mama
humano ZR75 fueron previamente transfectadas con los plasmidios pLacIOP o
pLacIOP-caveolina-1 como fue descrito por (Torres et al., 2006). En esencia, se
utilizó 5 μg de cada vector para 5x106 células en placas de 100 mm, luego fueron
seleccionadas por resistencia a higromicina, caracterizadas y usadas en estudios
previos y para este trabajo de tesis.
3.3.3. Expresión de caveolina-1 y E-cadherina en células de cáncer de colon
HT29(US)
La sub-línea celular metastásica de adenocarcinoma de colon humano
HT29(US) fue utilizada para re-expresar las proteínas E-cadherina (E-cad) y
caveolina-1 (estudio descrito por Torres et al., 2007). En dicho trabajo se
co-transfectó células HT29(US) con el plasmidio pBATEM2 (expresión de

39
E-cadherina) con los plasmidios pLacIOP o pLacIOP-caveolina-1. Después de 24 h
de transfección, las células fueron seleccionadas por su resistencia a higromicina
750 μM y luego fueron caracterizadas para la expresión de caveolina-1,
E-cadherina y survivina. En esta tesis, se utilizó las mismas células (HT29(US)
cav-1/E-cad (-) o HT29(US) cav-1/E-cad (+)) para evaluar la expresión de COX-2
mediante RT-PCR y PCR en tiempo real.
4. Análisis de la expresión de survivina, COX-2 y caveolina-1
4.1. Cuantificación del mRNA de COX-2 y survivina por RT-PCR
semicuantitativo y PCR en tiempo real
Una vez cumplido el tiempo de cultivo para cada experimento se procedió a
aislar el RNA usando el reactivo comercial Trizol (Invitrogen, Carlsbad, CA, USA)
según el protocolo del proveedor. Se utilizó 1 ml del reactivo para cada placa de
3,5 cm de diámetro. La concentración de RNA se determinó midiendo la
absorbancia a 260 nm y se confirmó su integridad mediante su migración
electroforética en geles de agarosa al 2% con bromuro de etidio.
4.2. Reacción de transcripción inversa (RT)
Se realizó una reacción de transcripción inversa en un volumen total de
20 µl: RNA total 1 μg, 2 μl de partidores al azar (50 μM) y agua nanopura hasta
completar un volumen de 11,2 μl. Luego se incubó a 70ºC durante 5 min y
posteriormente en hielo 15 min. Cumplida dicha incubación se agregó la siguiente
mezcla: dNTPs 0,4 µM, tampón (Tris HCl 50 mM pH 8,3, KCl 75 mM, MgCl2 3 mM,
DTT 10 mM) y transcriptasa inversa de AMV 10 U/µl, todo de Promega (Madison,

40
WI, USA). Finalmente, las muestras se incubaron 2 h a 42ºC y luego la enzima se
inactivó durante 5 min a 95ºC.
4.3. Reacción de RT-PCR semicuantitativo
Los cDNAs para COX-2 survivina, caveolina-1 y actina fueron amplificados
usando los siguientes partidores descritos en la tabla 2. Todos los productos de
reacción fueron analizados después de 25-30 ciclos de amplificación. Cada ciclo
estuvo dado por pasos consecutivos de 1 minuto a 94ºC, 55-60ºC (Tm usado
dependiendo de los partidores usados) y 72ºC (ver tabla 2). Los productos de PCR
fueron analizados en geles de agarosa al 2%. Los niveles de survivina y COX-2
fueron normalizados por la expresión de actina. Los valores numéricos indicados
entre paneles, fueron obtenidos por análisis densitométrico de cada banda en el gel
y expresado relativo a células control (100%).
Tabla 2. Oligonucleótidos y Tm usados para la obtención de cDNAs de COX-2, survivina, caveolina-1 y β-actina.
Partidor Secuencia 5’→3’ gen Tm sentido TTCAAATGAGATTGTGGGAAAATTGCT
antisentido AGATCATCTCTGCCTGAGTATCTT cox-2 55ºC
sentido CCGACGTTGCCCCCTGC
antisentido TCGATGGCACGGCGCAC survivina 60ºC
sentido GGGCAACATCTAGAAGCCCAACAA
antisentido CTGATGCACTGAATTCCAATCAGGAA caveolina-1 55ºC
sentido AAATCGTGCGTGACATTAAGC
antisentido CCGATCCACACGGAGTACTT β-actina 60ºC

41
4.4. Reacción de PCR en tiempo real
Los resultados obtenidos por RT-PCR semicuantitativo se confirmaron
mediante análisis por PCR en tiempo real. El procedimiento consistió en realizar
curvas estándar de COX-2 y survivina con productos de PCR diluidos desde 1/50
hasta 1/100000, de tal modo que la amplificación de una dilución de las muestras
(cDNAs) pueda ser interpolada en dichas curvas. Para ello se utilizó cDNAs de
cada experimento por línea celular y con el fin de establecer la mejor dilución, los
cDNAs fueron diluidos desde 1/10 hasta 1/100, siendo la dilución más adecuada la
de 1/50. Los niveles de COX-2 y survivina fueron normalizados respecto la
expresión del RNA 18S (partidor sentido 5’-TCAAGAACGAAAGTCGGAGG-3` y
partidor antisentido 5’-GGACATCTAAGGGCATCACA-3`) en dilución de 1/50 e
interpolada en su respectiva curva estándar. La amplificación de cada producto
PCR se realizó en un volumen de 25 μl de cada mezcla de reacción: 12,5 μl de
brilliant SYBR green quantitative PCR (Stratagene, La Jolla, CA), 0,5 μl de partidor
sentido (10 μM), 0,5 μl de partidor antisentido (10 μM), 6,5 μl de agua nanopura y
5 μl de cada dilución de cDNAs.
La reacción de PCR en tiempo real se llevó a cabo utilizando un equipo
Chromo4 real-time PCR detection system kit (Bio-Rad Laboratories) de acuerdo a
los protocolos sugeridos por el fabricante. La expresión relativa de cada gen fue
calculada usando el método de 2ΔΔCT (Livak y Schmittgen, 2001). Los datos fueron
expresados relativizando a un 100 % con los valores obtenidos en células control
(mock o sin tratamiento).

42
5. Western blot e inmunoprecipitación
5.1. Extracción de Proteínas, SDS-PAGE y Western blot
Una vez cumplido cada procedimiento experimental, las distintas líneas
celulares fueron utilizadas para el análisis de proteínas mediante Western blot. La
obtención de extractos celulares se realizó por lisis con un tampón RIPA (Tris-HCl
100 mM pH 7,4; Nonidet P-40 al 2%; SDS al 0,2%; NaCl 300 mM) más los
inhibidores de proteasas BAL (concentración final de benzamidina 11 µg/ml;
antipaína 2,2 µg/ml; leupeptina 5,5 µg/ml) y PMSF (fenil metil sulfonil fluoruro)
2 mM. En el caso de células adherentes (líneas de cáncer) estas fueron lavadas
con PBS frío y luego lisadas con 50-150 μl del tampón de lisis, seguido por
incubación en hielo durante 15 min y luego por dos ciclos de sonicación.
Los extractos fueron centrifugados a 15.000xg por 5 min a 4ºC y de la
fracción sobrenadante se cuantificó la concentración de proteínas usando el
método del BCA según las indicaciones del fabricante (Pierce). Las proteínas
fueron hervidas por 5 min en tampón de carga 2X (glicerol 8,7 %; 2 mercaptoetanol
5 %; SDS 5 %; Tris 62,5 mM pH 6,8), separadas posteriormente mediante
electroforesis en geles de poliacrilamida (SDS-PAGE) al 12 % (cargando 35-80 μg
de proteínas por carril) y transferidas a membranas de nitrocelulosa. Las
membranas fueron bloqueadas en PBS/Tween al 0,1% con leche al 5% (tampón de
bloqueo) o bien gelatina al 2% y después incubadas durante 1 h con diluciones de
los anticuerpos en solución de bloqueo: anti-caveolina-1 policlonal (1:3000) o
anti-caveolina-1 monoclonal (1:500), anti-survivina policlonal (1:2500), anti-
β-catenina monoclonal (1:2000), anti-E-cadherina monoclonal (1:3000), anti-COX-2
policlonal (1:500), anti-COX-2 monoclonal (1:350), anti-actina policlonal (1:5000) o
anti-GFP policlonal (1:5000).

43
Posteriormente, las membranas se lavaron 3 veces con PBS/Tween al 0,1%
y después incubadas con los anticuerpos acoplados a HRPO anti-IgG de conejo
(1:5000), anti-IgG de cabra (1:10000) o anti-IgG de ratón (1:4000) durante 2 h.
Finalmente, se agregó el sustrato de quimioluminiscencia EZ-ECL y la señal fue
detectada por el uso de films de rayos X o mediante el uso de un sistema de
quimioluminiscencia modelo Discovery 12iC de Ultralum (Claremont, CA USA).
5.2. Inmunoprecipitación de caveolina-1 con proteína A-sefarosa
Las células se homogenizaron en tampón de lisis (Tris 10mM pH 8,0; NaCl
150 mM; EDTA 5 mM, Tritón X-100 1% o bien n-octyl-beta-D-glucopyranoside
(OBG) 1 % más los inhibidores de proteasas BAL (benzamidina 11 µg/ml; antipaína
2,2 µg/ml; leupeptina 5,5 µg/ml) y fenil metil sulfonil fluoruro (PMSF) 2 mM,
incubando en hielo durante 20 min. Los extractos se centrifugaron a 15.000xg por 5
min a 4ºC y finalmente la fracción sobrenadante fue usada para determinar la
concentración de proteínas y para los ensayos de inmunoprecipitación.
La conjugación de los anticuerpos policlonales anti-caveolina-1 a esferas de
proteína A-sefarosa, se llevó a cabo por la incubación de 15 μl de una mezcla 1:1
en volumen de proteína A sefarosa/PBS con 2-4 μg de anticuerpo anti-caveolina-1
policlonal o una IgG irrelevante (anti-GAPDH policlonal), todo en un volumen total
de 500 μl de tampón de lisis a 4ºC con agitación orbital durante 1 h. Las esferas
con anticuerpos acoplados se lavaron 3 veces con PBS y bloquearon con PBS/BSA
al 2 % durante 1 h a 4ºC con agitación orbital. Después de 3 lavados adicionales
con PBS, las esferas acopladas con anticuerpos fueron incubadas con 500 μg de
cada extracto celular durante 1 h a 4ºC.

44
Finalmente, la suspensión se centrifugado a 15.000xg y el pellet, luego de 3
lavados con PBS, se resuspendió en tampón de carga para SDS-PAGE y analizado
mediante Western Blot. El sobrenadante y el extracto inicial (input) también se
analizaron. La cantidad de β-catenina co-inmunoprecipitada con caveolina-1 se
determinó por densitometría y fue expresada con respecto a su control de carga y a
la cantidad de caveolina-1 inmunoprecipitada.
5.3. Preparación de anticuerpos inmovilizados en esferas magnéticas
La conjugación de las esferas magnéticas con los anticuerpos
anti-caveolina-1 ocurre a través de una unión covalente y dichas esferas permiten
su separación una vez que son expuestas a un magneto. Es importante destacar
que para lograr una unión covalente entre las esferas magnéticas y anticuerpos,
este último debe estar resuspendido sólo en PBS, ya que la existencia de BSA u
otra proteína interfiere en la unión del anticuerpo con las esferas. En esencia, 25 ó
50 μg del anticuerpo policlonal anti-caveolina-1 resuspendido en PBS (Sigma,
#C3237) o bien una IgG irrelevante (anti-GAPDH policlonal) se incubaron con las
esferas magnéticas (Dynabeads M-280 Tosylactivated) de Invitrogen (Carlsbad,
CA) a una concentración final de 1x109 (esferas/ml) durante toda la noche a 37ºC
en una solución de borato 0,1 M, pH 9,5. El porcentaje de unión anticuerpo-esferas
fue calculado por la diferencia de absorbancia (280 nm) entre la solución inicial del
anticuerpo y el sobrenadante después de la incubación.

45
5.4. Análisis de inmunoprecipitados de caveolina-1 por espectrometría de
masas
Se incubó 500 μg de los extractos proteicos con las esferas magnéticas
(2x108) unidas a una IgG irrelevante (anti-GAPDH) durante 1 h a 4ºC (para eliminar
la unión no específica de proteínas), luego se separó el sobrenadante de las
esferas y se incubó el nuevo sobrenadante toda la noche a 4ºC con las esferas
unidas al anticuerpo anti-caveolina-1 en agitación orbital. Posteriormente, las
esferas se lavaron cuatro veces con PBS/OBG (1%). Las proteínas fueron eluídas
por temperatura (15 min a 90ºC) en una solución de NH4HCO3, 50 mM, pH 8,0.
Para controlar la inmunoprecipitación algunas muestras se chequearon por Western
blot. Luego, los eluidos provenientes de inmunoprecipitaciones realizadas en
paralelo fueron digeridos con tripsina toda la noche a 37ºC (NH4HCO3 50 mM;
Tripsina de porcino 0,1 μg/ml; CaCl2 1 mM) y posteriormente se detuvo la reacción
por adición de ácido trifluoro acético (TFA) 0,5 % (pH <2) e incubación durante
45 min a 37ºC. La solución resultante se centrifugó 10 min a 13000 rpm a 4ºC.
Finalmente, el contenido de sal en los sobrenadantes se eliminó mediante el
uso de minicolumnas de afinidad C18 ZIP TIP de Millipore (Billerica, MA, USA) y los
péptidos fueron analizados en un espectrómetro de masas MALDI-TOF/TOF
(Matrix-Assisted Laser Desorption/Ionization-Time Of Flight) o analizados en un
equipo de cromatografía capilar de fase reversa con ionizador de electrospray
acoplado a un espectrómetro de masas (MS/MS) con trampa de iones (LTQ,
Thermo Electron, Waltham, MA, USA) (ver metodología descrita en (Vaisar et al.,
2007)).

46
6. Ensayos de reporteros
6.1. Medición de la Actividad Transcripcional de Tcf/Lef
Se utilizó el sistema de actividad reportera luciferasa pTOP-FLASH y pFOP-
FLASH (vector control, sitios de unión a Tcf/Lef mutados) descrito previamente (van
de Wetering et al., 1997). Luego de 24 h de transfección o co-transfección, se
realizó la lisis de las células con el tampón de lisis (KH2PO4 0,1M, pH 7,9; DTT
2,5 mM; Tritón X-100 0,5 %) y los extractos se incubaron en hielo durante 15 min,
seguido de una centrifugación a 15.000xg durante 5 min a 4ºC.
Posteriormente, se realizó diluciones seriadas de los sobrenadantes (en
tampón de lisis) sobre placas blancas de 96 pocillos, luego se agregó 50 μl de
tampón KTME (Tris HCl 100 mM pH 7,8; MgSO4 10 mM; EDTA 2 mM) conteniendo
los sustratos luciferina 0,073 mM y ATP 5,5 µM y la actividad luciferasa se
cuantificó en un luminómetro. Las mediciones fueron normalizadas respecto a la
razón de las cuentas obtenidas con los vectores pTOP-FLASH/pFOP-FLASH.
Adicionalmente, la transfección de los vectores reporteros fue controlada por
la co-transfección con el vector pON (codifica para la β-galactosidasa) y los datos
se normalizaron por la actividad β-galactosidasa determinada en cada extracto.
La actividad β-galactosidasa fue determinada por la metabolización del reactivo
2-nitrofenil-β-D-galactopiranósido (ONPG) a 37ºC, siguiendo la absorbancia a
410 nm (lector de ELISA VERSA Max, Molecular Devices). La normalización de
datos con β-galactosidasa no reveló diferencias comparado sólo con la razón de los
vectores reporteros.
Por esto, para la mayoría de los experimentos, los resultados se expresaron como
la razón de las cuentas de cada vector reportero (excepción donde se indica). Los
valores obtenidos se expresaron con respecto a células control (100%).

47
6.2. Medición de la actividad transcripcional del promotor de survivina
Se utilizó el sistema de construcciones plasmidiales descrita previamente
(Figura 6). Los vectores pLuc-1710, que contiene los tres sitios de unión a Tcf/Lef
en la región promotora del gen de survivina o bien el pLuc-420-3M, que contiene
uno de los dos sitios de unión a Tcf/Lef proximales mutados (Kim et al., 2003),
fueron co-transfectados con pOSML-COX-2 o bien transfectados seguidos al
tratamiento con PGE2. La cuantificación de la actividad luciferasa y expresión de
datos fue realizada como se ha indicado anteriormente.
6.3. Medición de la actividad transcripcional del promotor de cox-2
La actividad reportera del promotor de cox-2 fue determinada por la
transfección de células con 2 μg de los plasmidios pGL3-COX-2 (contiene el
promotor de COX-2) o bien pGL3 (vector control). Células de cáncer (DLD-1 y
ZR75) o células HEK293T que expresan o no caveolina-1 se transfectaron con los
vectores reporteros descritos previamente (Figura 6) e incubadas según cada caso
en presencia de los inhibidores de la quinasa GSK-3β (LiCl, 1 mM o SB-216763,
20 μM) y 24 h post-transfección la actividad luciferasa fue cuantificada y los valores
se expresaron considerando los de células control como un 100 %.
7. Ensayos de viabilidad y proliferación
7.1. Ensayo de exclusión con Azul de tripan
Después de cada experimento, las células se recolectaron mecánicamente
(HEK293T) o por tripsinización (líneas celulares de cáncer), resuspendidas en PBS,

48
teñidas con azul de tripan por dilución 1:1 y observadas al microscopio óptico
invertido (XDS 1B) para cuantificar el porcentaje de células viables.
7.2. Ensayo de proliferación MTS®
Células HEK293T co-transfectadas con pOSML-COX-2 y pLacIOP o
pLacIOP-caveolina-1 o bien incubadas con PGE2, fueron sub-cultivadas en placas
de 96 pocillos durante 24 h a razón de 1x104 células por pocillo. La proliferación
celular se evaluó usando el kit de proliferación MTS®. Se agregó 20 μl de una
mezcla 20:1 de MTS:PMS (phenazine methosulfate; PMS) a cada pocillo, se incubó
esta mezcla durante 1-2 h a 37ºC y la reacción fue detenida por adición de 10 μl de
SDS al 10%. Finalmente, se determinó la reducción del compuesto MTS en
formazán por medición de la absorbancia a 490 nm (lector de ELISA VERSA Max,
Molecular Devices).
En el caso de experimentos de líneas celulares de cáncer de colon DLD-1 y
HT29(ATCC), transfectadas establemente con pLacIOP o pLacIOP-caveolina-1,
éstas se deprivaron de suero durante 4 h e incubadas 3 h en ausencia o presencia
de PGE2 (20 μM) en medio sin suero. Posteriormente, las células fueron sub-
cultivadas por 24 h adicionales en placas de 96 pocillos, sólo en presencia de IPTG
1 mM o en conjunto con el inhibidor de COX-1 (FR122047, 10 μM) o bien con el
inhibidor de COX-2 (SC-791, 20 μM). En experimentos adicionales, se incubó
células DLD-1 transfectadas establemente con pLacIOP o pLacIOP-caveolina-1 con
cantidades crecientes del inhibidor de COX-2 (0, 1, 5 y 20 μM).
En ambos casos la proliferación celular se determinó como fue descrito al
principio. Para todos los casos los valores están expresados como el porcentaje de

49
actividad residual con respecto a células control (100%), obtenidos de un promedio
de 3 experimentos independientes (promedio ± error estándar).
8. Inmunofluorescencia indirecta-microscopía confocal
Células HT29(ATCC) (2x104), establemente transfectadas con los plasmidios
pLacIOP o pLacIOP-caveolina-1, se crecieron por 24 h en presencia de IPTG 1 mM
sobre cubreobjetos. Posteriormente, las células se deprivaron de suero por 4 h e
incubaron 3 h con PGE2 (20 μM) en medio sin suero, alcanzando un 60% de
confluencia final. Luego de dos lavados con PBS, éstas fueron fijadas durante
20 min con paraformaldehído al 4%, en un tampón PIPES 100 mM pH 6,8,
conteniendo KOH 0,04 M, EGTA 2 mM y MgCl2 2 mM. Subsecuentemente, las
células se lavaron tres veces con un tampón Tris HCl 50 mM pH 7,6, NaCl 0,15 N y
azida de sodio 0,1% (tampón universal). Las células fueron permeabilizadas por
10 min con Tritón X-100 0,1% diluido en tampón universal, luego lavadas dos veces
con tampón universal y bloqueadas durante 30 min con BSA al 2% en el mismo
tampón. Después de bloquear, las células se incubaron con diluciones de los
anticuerpos primarios policlonal anti-caveolina-1 (1:100) y monoclonal
anti-β-catenina (1:100), seguido de los anticuerpos secundarios anti-IgG de conejo
acoplado a FITC (1:200) o anti-IgG de ratón acoplado a Cy3 (1:200) en BSA
2%/tampón universal. Finalmente, los cubreobjetos se lavaron 3 veces con tampón
universal, una vez con agua nanopura, estilados y después se montaron sobre
portaobjetos, usando 5 μl de solución de montaje Mowiol/DABCO (Tris 0,13 M
pH 8,5; Mowiol 13,3%; glicerol 30%; y DABCO 2,5%). Las muestras se visualizaron
con un microscopio confocal Carl Zeiss Axiovert-135 (LSM Microsystems),
utilizando láser de 488 nm y 543 nm (FONDAP, CEMC, Universidad de Chile).

50
La superposición y generación de un pseudocolor, así como la
reconstrucción de imágenes en 3D de las diferentes secciones ópticas obtenidas
por microscopía confocal fue generada la usando un software Imaris (Bitplane AG,
Zuerich, Switzerland). Utilizando el mismo software, se realizó un corte virtual a
5 μm del cubreobjeto de las imágenes reconstruidas, obteniendo así una mejor
visualización del interior del núcleo.
9. Cuantificación de PGE2.
Se recolectó el medio de cultivo (sobrenadantes) proveniente de líneas de
cáncer de colon [HT29(ATCC) y DLD-1] y de mama (ZR75) establemente
transfectadas con los plasmidios pLacIOP o pLacIOP-caveolina-1 en presencia o
ausencia de inhibidores de COX-1 (10 μM) y COX-2 (20 μM). Los sobrenadantes
fueron inmediatamente centrifugados y almacenados a -80ºC hasta ser analizados.
También, sobrenadantes provenientes de células HEK293T o células DLD-1
transfectadas con el vector pOSML-COX-2 (0-4 μg) se recolectaron del mismo
modo. La PGE2 presente en los sobrenadantes se cuantificó usando las
instrucciones de un sistema comercial “PGE2 EIA monoclonal” de Cayman
Chemical Company (Ann Arbor, MI, USA). Este ensayo consiste en la competencia
entre la PGE2 (presente en sobrenadantes) con un conjugado de PGE2-
acetilcolinesterasa (trazador de PGE2), por la unión al anticuerpo monoclonal
anti-PGE2. Para el cálculo de los resultados se realizó una curva de calibración
lineal (gráfico alternativo sugerido por el fabricante) desde la curva de calibración
con el estándar de PGE2 (proveniente del kit) y a partir de la regresión lineal se
interpolaron los datos para cada muestra.

51
Los resultados obtenidos provenientes de las distintas muestras se expresaron
como ng/ml. Los niveles basales de PGE2 se definieron usando medio de cultivo
como control.
10. Incubación de células con PGE2
En esta tesis se utilizaron concentraciones de PGE2 y deprivación de sueros
de acuerdo a lo descrito en la literatura (Shao et al., 2005; Liu et al., 2008). Los
experimentos realizados con PGE2 se han focalizado en al menos tres tipos de
estrategias que son: (1) Inducir la actividad transcripcional dependiente de
β-catenina-Tcf/Lef; (2) Revertir la inhibición de la expresión de survivina y de COX-2
por caveolina-1 y (3) Prevenir la disminución de la proliferación de células
embrionarias y tumorales que expresan caveolina-1. La figura 7 muestra un
esquema que resume los diferentes tratamientos empleados con PGE2.
HEK293T
Siembra
Deprivación de suero PGE2 (5 μM)24 h 12 h 4 h
-RT-PCR / PCR en tiempo real-Western blot
Siembra
Transfección ectópica PGE2 (5 μM)24 h 12 h 4 h
-Análisis de reporteros
Deprivación de suero24 h
Siembra
Deprivación de suero PGE2 (5 μM)
24 h 12 h 16 h
-MTS®
HEK293T
Siembra
Deprivación de suero PGE2 (5 μM)24 h 12 h 4 h
-RT-PCR / PCR en tiempo real-Western blot
Siembra
Transfección ectópica PGE2 (5 μM)24 h 12 h 4 h
-Análisis de reporteros
Deprivación de suero24 h
Siembra
Deprivación de suero PGE2 (5 μM)
24 h 12 h 16 h
-MTS®
A
(Figuras 18, 20)
(Figura 19)
(Figuras 20, 25)
Figura 7. Esquema resumen de los tratamientos con PGE2. (A) Células HEK293T (1x106) se crecieron durante 24 h y según corresponde se transfectaron con los diferentes vectores reporteros y posteriormente se deprivaron de suero durante 12 h seguido de la incubación con PGE2 (5 mM) por 4 h o bien 16 h para los ensayos de proliferación.

52
11. Análisis estadístico.
Donde se estimó pertinente, los resultados se compararon usando prueba t
no pareada de al menos 3 experimentos independientes y en los casos de
comparaciones múltiples se usó un test de ANOVA no paramétrico de Kruskal-
Wallis con un post-test (Dunn's Multiple Comparison Test). Un valor de p menor a
0,05 fue considerado significativo.
ZR75 / DLD-1 / HT29(ATCC)
Siembra
Deprivación de suero PGE2 (20 μM)24 h 4 h 3 h
-RT-PCR / PCR en tiempo real
-Western blot / IPP
Siembra
PGE2 (20 μM)24 h 3 h 24 h
-Análisis de proliferación
4 h
-Inmunofluorescencia
DLD-1 / HT29(ATCC)
Deprivación de suero
Reposición de sueros /
IPTG / Inhibidores
COX-1/2
ZR75 / DLD-1 / HT29(ATCC)
Siembra
Deprivación de suero PGE2 (20 μM)24 h 4 h 3 h
-RT-PCR / PCR en tiempo real
-Western blot / IPP
Siembra
PGE2 (20 μM)24 h 3 h 24 h
-Análisis de proliferación
4 h
-Inmunofluorescencia
DLD-1 / HT29(ATCC)
Deprivación de suero
Reposición de sueros /
IPTG / Inhibidores
COX-1/2
IPTG (1 mM)
IPTG (1 mM)
(Figuras 22, 30-33)
(Figuras 26, 27)
B
C
Figura 7. Esquema resumen de los tratamientos con PGE2. (B) Células ZR75, DLD-1 y HT29(ATCC) se crecieron por 24 h, luego se deprivaron de suero por 4 h seguido del tratamiento con PGE2 (20 μM) durante 3 h para finalmente realizar experimentos de RT-PCR / PCR en tiempo real, Western blot / inmunoprecipitación o inmunofluorescencia. (C) Células DLD-1 y HT29(ATCC) se crecieron por 24 h, deprivadas de suero 4 h seguido de la incubación con PGE2 (20 μM). Finalmente se repuso por medio con sueros con IPTG en combinación con los inhibidores específicos de COX-1/2. Transcurridas 24 h se cuantificó la proliferación celular.

53
VI. RESULTADOS
Los resultados obtenidos de este trabajo de tesis han sido organizados de
acuerdo a los objetivos específicos planteados. Por lo tanto, se presentan 2 secciones
de resultados: la sección I agrupa a los objetivos específicos Nº 1 y 2 y la sección II
contiene al objetivo específico Nº 3.
SECCION I
Objetivo específico 1
1. Analizar el efecto de la expresión ectópica de caveolina-1 sobre la regulación transcripcional de COX-2
1.1. Cuantificar la expresión de COX-2 en células humanas de cáncer de colon [DLD-1, HT29(ATCC) y HT29 (US)], de mama (ZR75) y de origen embrionario (HEK293T) en presencia y ausencia de caveolina-1
Los resultados de este objetivo se obtuvieron usando cuatro líneas celulares
humanas: cáncer de mama (ZR75), adenocarcinoma de colon (DLD-1, HT-29) y células
de riñón de origen embrionario humano (HEK293T). Dicha elección se basó en un
estudio previo de nuestro laboratorio, el cual demostró en estas células que caveolina-1
inhibe la expresión de survivina a través de la vía de β-catenina-Tcf/Lef (Torres et al.,
2006). Sobre esta base, se utilizó los mismos modelos celulares para investigar si la
expresión de caveolina-1 podría regular negativamente la expresión de COX-2, ya que
COX-2 es un blanco transcripcional de la vía β-catenina-Tcf/Lef (Haertel-Wiesmann et
al., 2000). Además, estas células son buenos candidatos por su muy baja expresión
endógena de caveolina-1, sumado a su expresión de COX-2 y survivina (Figura 8).

54
Para abordar nuestra propuesta, se realizó experimentos de RT-PCR, PCR en
tiempo real y Western blot. Los resultados de RT-PCR y Western blot para COX-2 en
células de cáncer de mama (ZR75) y de colon [HT29(ATCC), DLD-1] que expresan
caveolina-1 de manera ectópica e inducible por IPTG, revelaron que la presencia de
caveolina-1 disminuye significativamente los niveles de mRNA y niveles proteicos de
COX-2 (Figura 9A-C).
Para corroborar que células de cáncer que expresan caveolina-1 presentan una
reducción en los niveles de mRNA de COX-2 se utilizó PCR en tiempo real. Los
resultados de la figura 9D muestran una disminución del mRNA de COX-2 cercana al
50% en las tres líneas celulares estudiadas.
Figura 8. Niveles basales de caveolina-1, COX-2 y survivina de líneas celulares humanas de cáncer de colon [HT29(ATCC), DLD-1], de mama (ZR75) y de origen embrionario (HEK293T). Células HT29(ATCC), DLD-1, ZR75 y HEK293T transfectadas con el vector vacío pLacIOP, se sembraron en placas de 60 mm y se crecieron durante 24 h. Los extractos proteicos de cada línea celular se separaron mediante SDS-PAGE (50 µg/carril) y fueron analizados por Western blot (WB) con anticuerpos específicos para COX-2, survivina, caveolina-1 o actina. Los números entre paneles indican un análisis densitométrico de COX-2, survivina y caveolina-1, comparados con los niveles detectados para cada proteína en la línea HT29(ATCC). La cantidad comparativa de COX-2 fue: DLD-1, 0,5; ZR75, 0,3 y HEK293T, 0,1. La cantidad comparativa de survivina fue: DLD-1, 0,7; ZR75, 0,1 y HEK293T, 1,1 y la cantidad comparativa de caveolina-1 fue: DLD-1, 0,6; ZR75, 0 y HEK293T, 0. Western blot representativo de dos experimentos independientes.
COX-2
survivina
caveolina-1
actina
HT29
(ATC
C)DL
D-1
ZR75
HEK2
93T
1,0 0,5 0,3 0,1
1,0 0,7 0,1 1,1
1,0 0,6 0 0
COX-2
survivina
caveolina-1
actina
HT29
(ATC
C)DL
D-1
ZR75
HEK2
93T
1,0 0,5 0,3 0,1
1,0 0,7 0,1 1,1
1,0 0,6 0 0
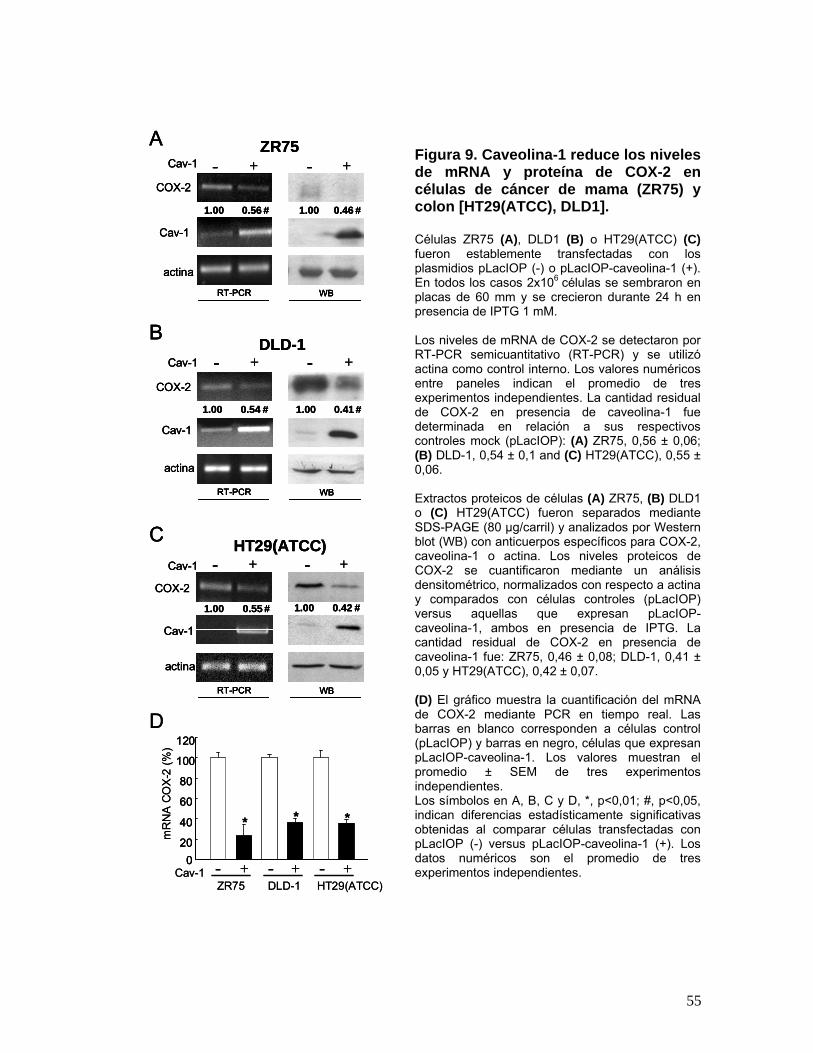
55
D
A
COX-2
C
B
HT29(ATCC)+-
COX-2
Cav-1
actina
+-Cav-1
#1.00 0.42#1.00 0.55
RT-PCR WB
ZR75
Cav-1
actina
Cav-1 +- +-
#1.00 0.56 #1.00 0.46
RT-PCR WB
DLD-1
COX-2
Cav-1
actina
+- +-Cav-1
#1.00 0.41#1.00 0.54
RT-PCR WB
+-ZR75
Cav-1
mR
NA
CO
X-2
(%)
+-DLD-1
+-HT29(ATCC)
020
40
60
80
100
120
* * *
D
A
COX-2
C
B
HT29(ATCC)+-
COX-2
Cav-1
actina
+-Cav-1
#1.00 0.42#1.00 0.55
RT-PCR WB
ZR75
Cav-1
actina
Cav-1 +- +-
#1.00 0.56 #1.00 0.46
RT-PCR WB
DLD-1
COX-2
Cav-1
actina
+- +-Cav-1
#1.00 0.41#1.00 0.54
RT-PCR WB
A
COX-2
C
B
HT29(ATCC)+-
COX-2
Cav-1
actina
+-Cav-1
#1.00 0.42#1.00 0.55
RT-PCRRT-PCR WBWB
ZR75
Cav-1
actina
Cav-1 +- +-
#1.00 0.56 #1.00 0.46
RT-PCR WBRT-PCR WB
DLD-1
COX-2
Cav-1
actina
+- +-Cav-1
#1.00 0.41#1.00 0.54
RT-PCR WBRT-PCR WB
+-ZR75
Cav-1
mR
NA
CO
X-2
(%)
+-DLD-1
+-HT29(ATCC)
020
40
60
80
100
120
020
40
60
80
100
120
* * *
Figura 9. Caveolina-1 reduce los niveles de mRNA y proteína de COX-2 en células de cáncer de mama (ZR75) y colon [HT29(ATCC), DLD1]. Células ZR75 (A), DLD1 (B) o HT29(ATCC) (C) fueron establemente transfectadas con los plasmidios pLacIOP (-) o pLacIOP-caveolina-1 (+). En todos los casos 2x106 células se sembraron en placas de 60 mm y se crecieron durante 24 h en presencia de IPTG 1 mM. Los niveles de mRNA de COX-2 se detectaron por RT-PCR semicuantitativo (RT-PCR) y se utilizó actina como control interno. Los valores numéricos entre paneles indican el promedio de tres experimentos independientes. La cantidad residual de COX-2 en presencia de caveolina-1 fue determinada en relación a sus respectivos controles mock (pLacIOP): (A) ZR75, 0,56 ± 0,06; (B) DLD-1, 0,54 ± 0,1 and (C) HT29(ATCC), 0,55 ± 0,06. Extractos proteicos de células (A) ZR75, (B) DLD1 o (C) HT29(ATCC) fueron separados mediante SDS-PAGE (80 µg/carril) y analizados por Western blot (WB) con anticuerpos específicos para COX-2, caveolina-1 o actina. Los niveles proteicos de COX-2 se cuantificaron mediante un análisis densitométrico, normalizados con respecto a actina y comparados con células controles (pLacIOP) versus aquellas que expresan pLacIOP-caveolina-1, ambos en presencia de IPTG. La cantidad residual de COX-2 en presencia de caveolina-1 fue: ZR75, 0,46 ± 0,08; DLD-1, 0,41 ± 0,05 y HT29(ATCC), 0,42 ± 0,07. (D) El gráfico muestra la cuantificación del mRNA de COX-2 mediante PCR en tiempo real. Las barras en blanco corresponden a células control (pLacIOP) y barras en negro, células que expresan pLacIOP-caveolina-1. Los valores muestran el promedio ± SEM de tres experimentos independientes. Los símbolos en A, B, C y D, *, p<0,01; #, p<0,05, indican diferencias estadísticamente significativas obtenidas al comparar células transfectadas con pLacIOP (-) versus pLacIOP-caveolina-1 (+). Los datos numéricos son el promedio de tres experimentos independientes.

56
En un trabajo previo de nuestro laboratorio, se utilizó el modelo de células
embrionarias humanas HEK293T debido a que permiten realizar transfecciones
transientes eficientes, expresan survivina, caveolina-1 está casi ausente y su
re-expresión demostró inhibir la expresión de survivina (Torres et al., 2006). En base a
esto, decidimos utilizar este modelo, ya que estas células expresan COX-2 y la
sobreexpresión de caveolina-1 podría modular los niveles de COX-2 por la misma vía
descrita para survivina.
Por lo tanto, la estrategia experimental consistió en utilizar dos sistemas de
expresión de caveolina-1 en células HEK293T. Con uno se expresó caveolina-1 como
proteína de fusión a GFP (transfección con vectores pEGFP-C1 o pEGFP-caveolina-1).
Este sistema tiene la ventaja de que las células transfectadas son visualizadas por la
fluorescencia verde del GFP. La desventaja, es que caveolina-1 aparece como proteína
de fusión con GFP y esto podría cambiar la conformación de caveolina-1 y alterar así su
función. Por esto, se usó un segundo sistema de expresión, basado en el vector
pLacIOP, el cual permite la expresión de caveolina-1 de manera inducible por IPTG.
Células HEK293T (1x106) transfectadas con cada vector se lisaron 24 h post-
transfección para preparar extractos los que se analizaron para evaluar el contenido de
mRNA y de proteína para COX-2. Los resultados muestran que la presencia de
caveolina-1 o GFP-caveolina-1 disminuye significativamente los niveles basales del
mRNA de COX-2, cuantificados mediante RT-PCR (Figura 10A).
Estos resultados también se corroboraron por PCR en tiempo real (Figura 10B).
En concordancia, ensayos de Western blot revelaron una disminución significativa en la
cantidad del contenido proteico de COX-2, también con el uso de ambos vectores
pLacIOP-caveolina-1 (inducible por IPTG) y pEGFP-caveolina-1 (proteína de fusión con
GFP) (Figura 10C).

57
pLacIOPpEGFP-C1
Cav-1
COX-2
actin
- + - +
1.00 0.35 1.00 0.55# #
0
20
40
60
80
100
120
140
#
CO
X-2
mR
NA
(%)
#
pEGFP-C1+-Cav-1 +- -
pLacIOP
0
20
40
60
80
100
120
140
0
20
40
60
80
100
120
140
#
CO
X-2
mR
NA
(%)
#
pEGFP-C1+-Cav-1 +- -
pLacIOP
1.00 0.58 1.00 0.61#
pLacIOPpEGFP-C1
Cav-1 - + - + (C+)
COX-2
Cav-1
GFP
actin
#1.00 0.58 1.00 0.61#
pLacIOPpEGFP-C1
Cav-1 - + - + (C+)
COX-2
Cav-1
GFP
actin
#
A
B
C
Figura 10. Caveolina-1 inhibe la expresión de COX-2 en células HEK293T. Células HEK293T (1x106) transfectadas con los plasmidios pEGFP-C1, pEGFP-caveolina-1, pLacIOP o pLacIOP-caveolina-1 (2 μg en cada caso). Post-transfección, las células fueron incubadas en ausencia (pEGFP-C1, pEGFP-caveolina-1) o presencia de IPTG (pLacIOP, pLacIOP-caveolina-1) por 24 h. (A) Los niveles de mRNA de COX-2 se detectaron por RT-PCR semicuantitativo y se utilizó actina como control interno. Los valores numéricos entre paneles indican el promedio de tres experimentos independientes. La cantidad residual de COX-2 en presencia de caveolina-1 fue determinada en relación a sus respectivos controles mock (pEGFP-C1 o pLacIOP): 0,35 ± 0,07 (pEGFP-caveolina-1) y 0,55 ± 0,07 (pLacIOP-caveolina-1), respectivamente. (B) Células HEK293T transfectadas del mismo modo descrito anteriormente, luego el mRNA de COX-2 fue cuantificado mediante PCR en tiempo real. Los datos corresponden a tres experimentos independientes y fueron normalizados a un 100% con aquellos valores obtenidos con células no transfectadas. Se utilizó como control interno el mRNA ribosomal 18S. (C) La expresión de cada proteína fue determinada por Western blot, cargando 80 μg de proteínas por carril, usando anticuerpos específicos anti-COX-2, anti-caveolina 1, anti-GFP o anti-actina. Como control positivo (C+) de COX-2, células HEK293T fueron transfectadas con 2 μg de un vector que expresa COX-2 (pOSML-COX-2). Los niveles proteicos de COX-2 fueron normalizados con respecto a actina y cuantificados por densitometría. La cantidad residual de COX-2 fue: 0,58 ± 0,12 (pEGFP-caveolina-1) y 0,61 ± 0,10 (pLacIOP-caveolina-1). El símbolo #, p<0,05, indica las diferencias estadísticamente significativas se obtuvieron de tres experimentos independientes utilizando células transfectadas con pEGFP-C1 y pEGFP-caveolina-1, o también con pLacIOP y pLacIOP-caveolina-1.

58
La cuantificación mediante un análisis densitométrico para tres experimentos
independientes demostró que la presencia de caveolina-1 sola o como proteína de
fusión a GFP causa una reducción de un 40-50% en el contenido de COX-2. Sin
embargo, en las células HEK293T los niveles basales de COX-2 son relativamente
bajos respecto a células de cáncer de colon. Por lo tanto, para identificar la banda
correspondiente para COX-2 se utilizó como control positivo (C+) células HEK293T
transfectadas con un vector de expresión de COX 2 (pOSML-COX-2).
En un estudio reciente de nuestro Laboratorio se demostró que la regulación
negativa de survivina a través de la vía de β-catenina-Tcf/Lef por caveolina-1 requiere la
presencia de E-cadherina en células HT29(US) (Torres et al., 2007). Por este motivo,
investigamos si lo mismo ocurre para COX-2 y para ello utilizamos células HT29(US)
que poseen niveles muy bajos de E-cadherina y expresan o no caveolina-1 inducible
con IPTG 1 mM. Los resultados revelaron que no existen diferencias significativas en
los niveles del mRNA de COX-2 (Figura 11A). En cambio, cuando células HT29(US)
que expresan o no caveolina-1 fueron adicionalmente transfectadas con el plasmidio
pBATEM2 (vector de expresión de E-cadherina), si se observó una reducción
significativa en los niveles del mRNA de COX-2 (Figura 11B). Estos resultados también
se corroboraron por PCR en tiempo real (Figura 11C). Finalmente, nuestros datos
sugieren que la presencia de caveolina-1 disminuye los niveles de mRNA de COX-2 en
células que expresan E-cadherina y no en células que carecen de E-cadherina.

59
Estos resultados sugieren que la re-expresión de caveolina-1, en líneas celulares de cáncer y de origen embrionario, promueve la regulación negativa de la expresión de COX-2 por un mecanismo transcripcional que depende de la presencia de E-cadherina.
Figura 11. La reducción de los niveles de mRNA de COX-2 requiere la presencia de E-cadherina en células HT29(US). Células HT29(US) (E-cadherina, (-)) establemente transfectadas con pLacIOP (-) o pLacIOP-caveolina-1 (+) se incubaron en ausencia o presencia de IPTG 1 mM. (A) Los niveles de mRNA de COX-2 y actina fueron analizados por RT-PCR semi-cuantitativo. La expresión de actina se utilizó como control interno. Los datos numéricos entre paneles, fueron obtenidos mediante análisis densitométrico y corresponden al promedio de tres experimentos independientes (promedio ± S.E.M) cada uno de ellos normalizados con actina. (B) Células HT29(US) fueron co-transfectadas con 2 μg de los plasmidios: pBATEM2 (E-cadherina, (+)) y pLacIOP (-) o pLacIOP-caveolina-1 (+) y posteriormente se crecieron por 24 h en ausencia o presencia de IPTG 1 mM. Los niveles de mRNA de COX-2 fueron analizados del mismo modo descrito en (A). Los datos numéricos provenientes de tres experimentos independientes (promedio ± SEM) se normalizaron con actina. Los niveles residuales de mRNA de COX-2 en células que expresan E-cadherina y caveolina-1 fueron 0,65 ± 0,14 y son estadísticamente significativos entre células co-transfectadas con pLacIOP/pBATEM2 versus pLacIOP-caveolina-1/pBATEM2 (#, p<0.05). (C) Análisis de los niveles de mRNA de COX-2 mediante PCR cuantitativo en tiempo real (ver metodología) de células HT29(US) co-transfectadas como en (B). Las barras en blanco corresponden a células transfectadas con pLacIOP (-) y barras en negro aquellas transfectadas con pLacIOP-caveolina-1. Los datos numéricos son el promedio dos experimentos independientes realizados por triplicado y normalizados por la expresión del RNA 18S ribosomal.
IPTG
+-Cav-1 +-COX-2
β-actina
1,0 0,9 1,1 0,9
E-Cadherina (-)
IPTG
+-Cav-1 +-COX-2
β-actina
1,0 0,9 1,1 0,9
E-Cadherina (-)
IPTG
+-Cav-1 +-
β-actina
COX-2
1,0 0,8 1,1 0,65#
E-Cadherina (+)
IPTG
+-Cav-1 +-
β-actina
COX-2
1,0 0,8 1,1 0,65#
E-Cadherina (+)
Cav-1 +- +-0
20
40
60
80
100
120
140
Cav-1 +- +-
mR
NA
CO
X-2
(%)
E-CadherinaCav-1 +- +-0
20
40
60
80
100
120
140
Cav-1 +- +-
mR
NA
CO
X-2
(%)
E-Cadherina
A
B
C

60
1.2. Cuantificar la actividad transcripcional de β-catenina-Tcf/Lef (TOP/FOP-
FLASH) y de COX-2 (Promotor de cox-2)
Inicialmente, se realizaron experimentos de dosis respuesta en células HEK293T
con un inhibidor específico (SB-216763) de la quinasa GSK-3β y uno menos especifico
como litio, ya que se sabe que la inhibición de dicha enzima incrementa la actividad
transcripcional de β-catenina-Tcf/Lef y por ende aumenta también la expresión de
blancos transcripcionales, tales como cox-2 y survivina. Con esta estrategia
controlamos de manera indirecta la inhibición de la quinasa GSK-3β, ya que nuestro
laboratorio no posee las herramientas para cuantificar la actividad quinasa in vitro.
Los resultados revelaron que la inhibición de la quinasa GSK-3β aumenta los niveles de
mRNA (Figura 12A) y proteicos (Figura 12B) de COX-2 y survivina.
La actividad transcripcional fue cuantificada en células que expresan o no
caveolina-1. Para ello se cuantificó mediante ensayos de transfección con vectores de
expresión reporteros para β-catenina-Tcf/Lef, TOP-FLASH y FOP-FLASH (ver mas
detalle en figura 6). Para evaluar la actividad transcripcional de COX-2, se utilizó el
promotor para el gen de cox-2 humana que ha sido clonado por el grupo de Dr. Ying
Zhu (Laboratory of Virology, Wuhan University, Wuhan, China), quien nos donó
generosamente los plasmidios reporteros para determinar la actividad transcripcional
del promotor de cox-2 pGL3-COX-2 (contiene un sitio de unión a Tcf/Lef) o pGL3
(contiene sólo el gen de la luciferasa).
La presencia de caveolina-1 en células HEK293T causó una disminución
significativa en la actividad transcripcional del plasmidio reportero TOP-FLASH lo que
se reflejó también con menores niveles del mRNA de COX-2 (Figura 13A).

61
Este efecto en la actividad reportera fue significativamente revertido cuando
células HEK293T se incubaron con inhibidores de la quinasa GSK-3β (Figura 13A).
Al mismo tiempo, se evaluó los niveles proteicos de COX-2, survivina y β-catenina los
que también reflejan la modulación de la actividad transcripcional de β-catenina-Tcf/Lef
por caveolina-1 (Figura 13B).
Para demostrar específicamente que caveolina-1 reduce la expresión de COX-2
por la misma vía se utilizó los plasmidios reporteros para COX-2 descritos previamente
en metodología. Los resultados revelaron que caveolina-1 disminuye significativamente
la actividad reportera de cox-2 y en este caso, el efecto de caveolina-1 también fue
inhibido con el uso de ambos inhibidores de la quinasa GSK-3β (Litio, 20 mM o
SB-216763, 10 μM) (Figura 13C).
Figura 12. La inhibición de la quinasa GSK-3β aumenta los niveles de mRNA y proteicos de COX-2 y survivina. Dosis respuesta del inhibidor específico de la quinasa GSK-3β (SB-216763). Células HEK293T fueron incubadas 24 h en presencia de concentraciones crecientes del inhibidor SB-216763 (0, 10 y 20 μM). Adicionalmente, células HEK293T se incubaron con un inhibidor menos específico, LiCl (20 mM). (A) Una vez incubadas las células en presencia de los inhibidores se extrajo el RNA total y se llevó a cabo experimentos de RT-PCR semicuantitativo utilizando partidores específicos para COX-2, survivina y actina. (B) Extractos proteicos (80 μg/carril) fueron separados por SDS-PAGE y analizados mediante Western blot usando anticuerpos específicos anti-COX-2, anti-survivina o anti-actina. En ambos casos, se muestra un resultado representativo de dos experimentos independientes. Los datos numéricos indicados entre paneles fueron obtenidos mediante un análisis densitométrico donde se utilizó actina para su normalización.
A
SB-216763 (μM)
COX -2
survivina
actina
0 10 20 Li+(20 mM)
1.00 1.6 1.90 2.4
1.0
B
2.1 2.3 3.6
SB-216763 (μM)0 10 20 Li+(20 mM)
COX -2
survivina
actina
A
SB-216763 (μM)
COX -2
survivina
actina
0 10 20 Li+(20 mM)
1.00 1.6 1.90 2.4
1.0
B
2.1 2.3 3.6
SB-216763 (μM)0 10 20 Li+(20 mM)
COX -2
survivina
actina

62
Decidimos investigar si caveolina-1 específicamente disminuye la transcripción de
COX-2 en células de cáncer de mama (ZR75) y de colon (DLD-1) transfectadas
establemente con pLacIOP o pLacIOP-caveolina-1. Para ello las células fueron
adicionalmente transfectadas con los plasmidios reporteros pGL3 o pGL3-COX-2.
Figura 13. Caveolina-1 inhibe la expresión de COX-2 por la supresión de la actividad transcripcional dependiente de β-catenina-Tcf/Lef en células HEK293T. Células HEK293T (1x106) se co-transfectaron con los vectores reporteros pTOP-FLASH o pFOP-FLASH y pLacIOP o pLacIOP-caveolina-1 (2 μg por cada vector). Una vez transfectadas, las células fueron incubadas 24 h con IPTG 1 mM. Adicionalmente, células HEK293T transfectadas con pLacIOP-caveolina-1 fueron cultivadas en presencia de litio (20 mM) o SB-216763 (10 μM). (A) Transcurridas 24 h, se cosechó las células y se preparó extractos para ensayos de reportero y RT-PCR semicuantitativo. Las barras indican la actividad luciferasa normalizada por la razón entre pTOP-FLASH/pFOP-FLASH (ver metodología) respecto a células transfectadas con pLacIOP. El panel inferior muestra los niveles de mRNA de COX-2 y actina analizados mediante RT-PCR semicuantitativo. (B) Extractos proteicos (80 μg/carril) fueron separados por SDS-PAGE y analizados mediante Western blot usando anticuerpos específicos anti-COX-2, anti-β-catenina, anti-survivina, anti-caveolina-1 o anti-actina. (C) Células HEK293T co-transfectadas con los vectores reporteros pGL3-COX-2 o pGL3 y pLacIOP o pLacIOP-caveolina-1 (2 μg por cada vector). Una vez transfectadas, las células se incubaron con inhibidores de la quinasa GSK-3β, como fue descrito anteriormente. La actividad luciferasa fue normalizada por cada condición calculando la razón entre la actividad pGL3-COX-2/pGL3. Los datos de A y C representan tres experimentos independientes (promedio ± SEM). Se indican diferencias estadísticamente significativas (#, p<0,05; *, p<0,01).
LitioSB-216763
- +Cav-1-- -
-+ ++ -
+-
COX-2
actina
caveolina-1
Survivina
β-catenina
0
100
200
300
400
LitioSB-216763
- +Cav-1-- -
-+ ++ -
+-
Rep
orte
ro C
OX
-2 (%
)
**
#
*
#
*A
ctiv
idad
luci
fera
sa(%
)
COX-2
actina
LitioSB-216763
Cav-1 - +-- -
-+ ++ -
+-
TOP/FOP
0
50100150200
250300
LitioSB-216763
- +Cav-1-- -
-+ ++ -
+-
COX-2
actina
caveolina-1
Survivina
β-catenina
0
100
200
300
400
LitioSB-216763
- +Cav-1-- -
-+ ++ -
+-
Rep
orte
ro C
OX
-2 (%
)
**
#
*
#
*A
ctiv
idad
luci
fera
sa(%
)
COX-2
actina
LitioSB-216763
Cav-1 - +-- -
-+ ++ -
+-
TOP/FOP
0
50100150200
250300
A
B
C
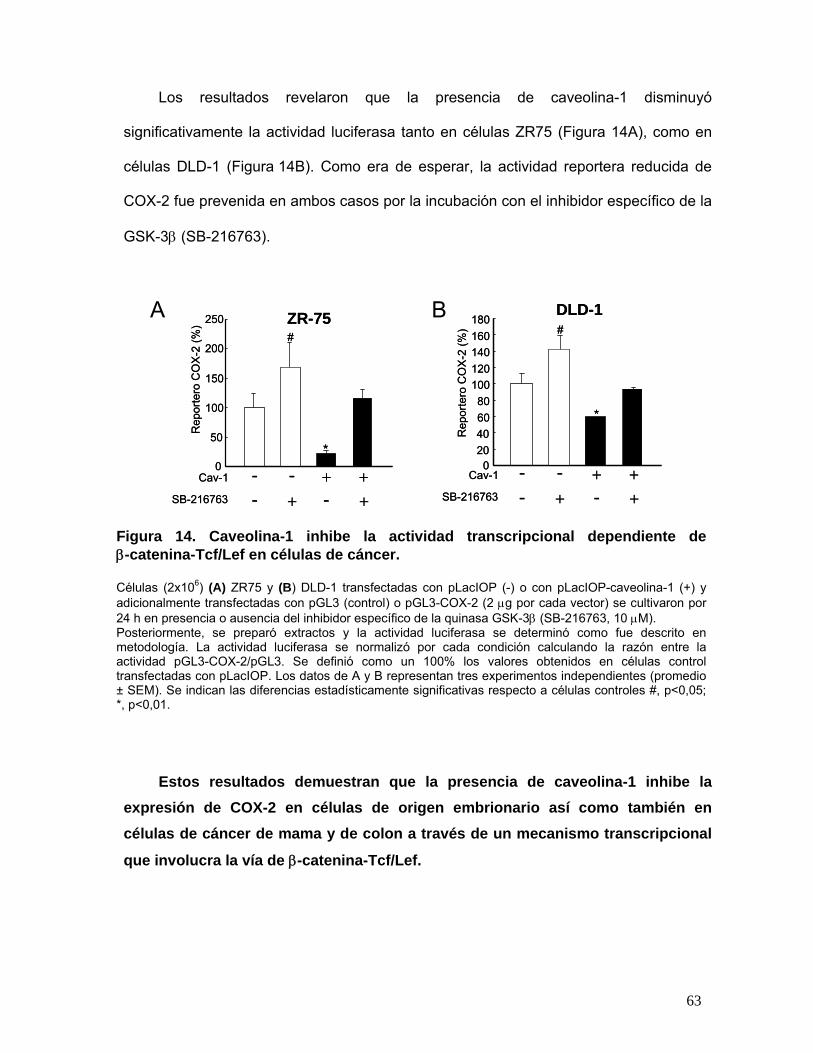
63
Los resultados revelaron que la presencia de caveolina-1 disminuyó
significativamente la actividad luciferasa tanto en células ZR75 (Figura 14A), como en
células DLD-1 (Figura 14B). Como era de esperar, la actividad reportera reducida de
COX-2 fue prevenida en ambos casos por la incubación con el inhibidor específico de la
GSK-3β (SB-216763).
Estos resultados demuestran que la presencia de caveolina-1 inhibe la
expresión de COX-2 en células de origen embrionario así como también en células de cáncer de mama y de colon a través de un mecanismo transcripcional
que involucra la vía de β-catenina-Tcf/Lef.
Figura 14. Caveolina-1 inhibe la actividad transcripcional dependiente de β-catenina-Tcf/Lef en células de cáncer. Células (2x106) (A) ZR75 y (B) DLD-1 transfectadas con pLacIOP (-) o con pLacIOP-caveolina-1 (+) y adicionalmente transfectadas con pGL3 (control) o pGL3-COX-2 (2 μg por cada vector) se cultivaron por 24 h en presencia o ausencia del inhibidor específico de la quinasa GSK-3β (SB-216763, 10 μM). Posteriormente, se preparó extractos y la actividad luciferasa se determinó como fue descrito en metodología. La actividad luciferasa se normalizó por cada condición calculando la razón entre la actividad pGL3-COX-2/pGL3. Se definió como un 100% los valores obtenidos en células control transfectadas con pLacIOP. Los datos de A y B representan tres experimentos independientes (promedio ± SEM). Se indican las diferencias estadísticamente significativas respecto a células controles #, p<0,05; *, p<0,01.
ZR-75
Cav-1 +- +-SB-216763 - + - +
Rep
orte
ro C
OX
-2 (%
)
#
*0
50
100
150
200
250 ZR-75
Cav-1 +- +-SB-216763 - + - +
Rep
orte
ro C
OX
-2 (%
)
#
*0
50
100
150
200
250DLD-1
Cav-1 +- +-SB-216763 - + - +
Rep
orte
ro C
OX
-2 (%
)
*
#
020406080
100120140160180
DLD-1
Cav-1 +- +-SB-216763 - + - +
Rep
orte
ro C
OX
-2 (%
)
*
#
020406080
100120140160180A B

64
A B
C D
Objetivo específico 2 2. Estudiar el mecanismo de regulación transcripcional de survivina por la expresión de COX-2 y su nexo con la regulación negativa en la proliferación celular por caveolina-1 2.1. Determinar el efecto de la sobreexpresión de COX-2 sobre la actividad
transcripcional de β-catenina-Tcf/Lef y sobre la expresión de survivina
Se sabe que survivina es regulada directamente por la vía de β-catenina-Tcf/Lef
(Zhang et al., 2001; Torres et al., 2006) y además, existe evidencia que la PGE2, un
producto derivado de la actividad enzimática de COX-2, promueve la estabilización de
survivina y así incrementa la resistencia apoptótica de células tumorales de pulmón
(Krysan et al., 2003; Krysan et al., 2004b). Por este motivo, nos enfocamos en
investigar el efecto de la sobreexpresión de COX-2 sobre la actividad transcripcional de
β-catenina-Tcf/Lef y la expresión de survivina en células HEK293T, que también
expresan los receptores de PGE2 (Hoshino et al., 2007).
Figura 15. Caracterización del vector de expresión de COX-2 (pOSML-COX-2). (A) Mapa del
vector pOSML-COX-2. Los cuadros indican sitios de corte por la enzima NotI, la cual permite liberar el inserto de aprox. 1,9 kb. (B) Gel de agarosa al 1% teñido con bromuro de etidio. Digestión enzimática del vector para COX-2, el orden de carga es el siguiente: 1.-Std 1 kb; 2.-vector digerido por NotI; 3.-vector intacto; 4.-amplificación de COX-2 por PCR (amplicón de 305 pb).
(C) RT-PCR de células HEK293T transfectadas con 0 ó 2 μg del vector pOSML-COX-2, como control de carga se utilizó actina. (D) Ensayo de Western blot con anticuerpos específicos para COX-2 y actina como control de carga, de izquierda a derecha se muestra cantidades crecientes (0, 0,5, 1, 2 y 4 μg) del vector pOSML-COX-2 utilizado para la expresión ectópica de COX-2 en células HEK293T.

65
Para sobreexpresar COX-2 se utilizó un vector que expresa la enzima silvestre
(pOSML-COX-2). La figura 15A-D muestra la caracterización y sobreexpresión del
vector en células HEK293T. En la figura 16A se muestra el efecto de la sobreexpresión
de COX-2 sobre survivina analizado por RT-PCR. Este reveló incrementos en los
niveles del mRNA de survivina en la medida que se aumentó la cantidad del vector
pOSML-COX-2. También se detectó aumentos en los niveles proteicos de COX-2,
β-catenina y survivina por análisis de Western blot (Figura 16B).
Figura 16. La sobreexpresión de COX-2 aumenta la expresión de survivina y β-catenina en células HEK293T. Células HEK293T (1x106) transfectadas con cantidades crecientes (0, 0,5, 1, 2 y 4 μg) del vector que codifica para la COX-2 (pOSML-COX-2), 24 h post-transfección se cosechó las células y se procedió a la extracción de RNA total o bien se preparó extractos de proteínas para Western blot. (A) Cuantificación de mRNA para COX-2 y survivina mediante RT-PCR semicuantitativo utilizando partidores específicos. Se utilizó actina como control de carga. Los datos numéricos entre paneles corresponden al promedio de tres experimentos independientes. Los valores relativos (promedio ± SEM) de mRNA para survivina, comparado con células control sin transfectar fueron: 2,0 ± 0,07, 2,1 ± 0,05 y 2,8 ± 0,1, con 1, 2 y 4 μg del vector pOSML-COX-2, respectivamente. (B) Análisis de Western blot de células HEK293T transfectadas con el vector pOSML-COX-2, descrito en metodología. El contenido de proteínas totales (80 μg/ carril) fue separado mediante SDS-PAGE y posteriormente analizado por inmunoblot usando anticuerpos específicos anti-COX-2, anti-β-catenina, anti-survivina o anti-actina. Los datos numéricos fueron obtenidos mediante un análisis densitométrico. Los valores indicados entre paneles fueron normalizados contra actina y corresponden al promedio de tres experimentos independientes (promedio ± SEM). Los valores relativos para β-catenina comparado con células control fueron: 2,6 ± 0,04, 2,7 ± 0,03 y 2,4 ± 0,08, con 1, 2 y 4 μg de pOSML-COX-2, respectivamente. A su vez, los valores relativos para survivina fueron: 2,5 ± 0,04, 4,2 ± 0,08 y 4,6 ± 0,08, con 1, 2 y 4 μg de pOSML-COX-2, respectivamente. Las diferencias estadísticamente significativas encontradas en (A) y (B) respecto células controles se indican como #, p<0,05; *, p<0,01.
###
***
0 0,5 1,0 2,0 4,0pOSML-COX-2 (μg)
COX-2
actina
survivina1,0 1,3 1,4 1,9 2,9
1,0 1,9 2,0 2,1 2,8 ###
***
0 0,5 1,0 2,0 4,0pOSML-COX-2 (μg)
COX-2
actina
survivina1,0 1,3 1,4 1,9 2,9
1,0 1,9 2,0 2,1 2,8
###
# ##
***
COX-2
β-catenina
actina
survivina
0 0,5 1,0 2,0 4,0pOSML-COX-2 (μg)
1,0 1,7 3,4 3,5 4,4
1,0 1,6 2,5 4,2 4,6
1,0 1,4 2,6 2,7 2,4
###
# ##
***
COX-2
β-catenina
actina
survivina
0 0,5 1,0 2,0 4,0pOSML-COX-2 (μg)
1,0 1,7 3,4 3,5 4,4
1,0 1,6 2,5 4,2 4,6
1,0 1,4 2,6 2,7 2,4
A B
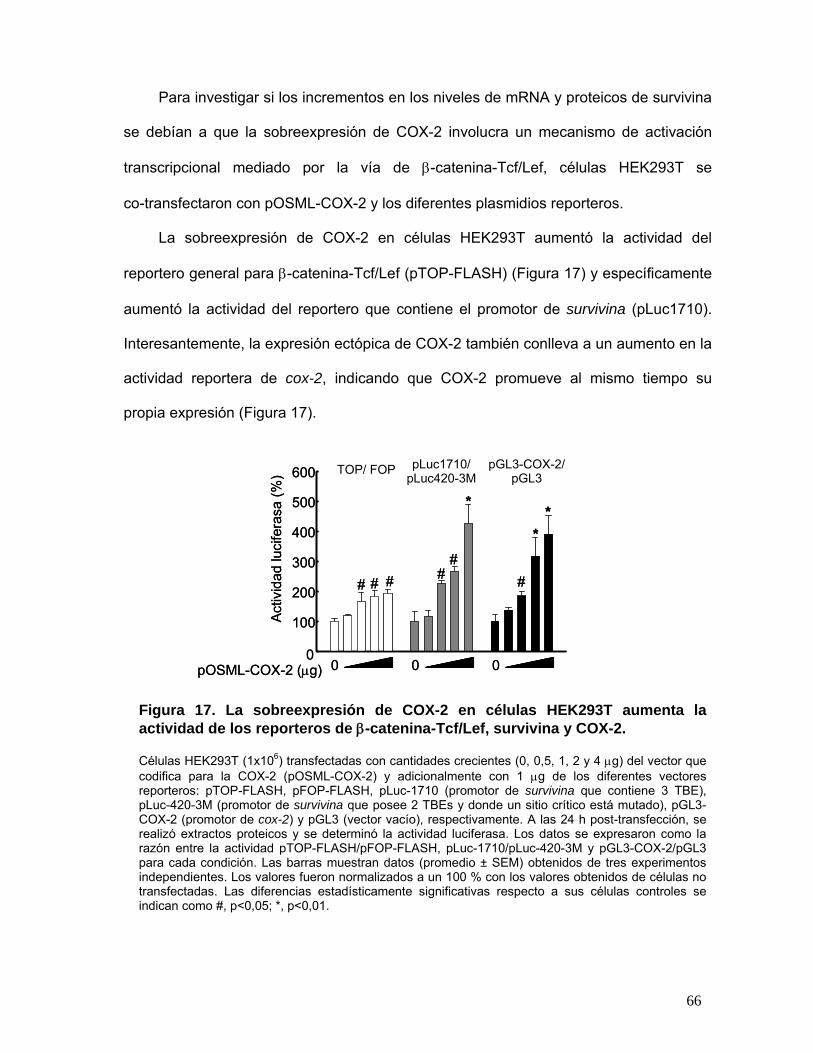
66
Para investigar si los incrementos en los niveles de mRNA y proteicos de survivina
se debían a que la sobreexpresión de COX-2 involucra un mecanismo de activación
transcripcional mediado por la vía de β-catenina-Tcf/Lef, células HEK293T se
co-transfectaron con pOSML-COX-2 y los diferentes plasmidios reporteros.
La sobreexpresión de COX-2 en células HEK293T aumentó la actividad del
reportero general para β-catenina-Tcf/Lef (pTOP-FLASH) (Figura 17) y específicamente
aumentó la actividad del reportero que contiene el promotor de survivina (pLuc1710).
Interesantemente, la expresión ectópica de COX-2 también conlleva a un aumento en la
actividad reportera de cox-2, indicando que COX-2 promueve al mismo tiempo su
propia expresión (Figura 17).
Figura 17. La sobreexpresión de COX-2 en células HEK293T aumenta la actividad de los reporteros de β-catenina-Tcf/Lef, survivina y COX-2. Células HEK293T (1x106) transfectadas con cantidades crecientes (0, 0,5, 1, 2 y 4 μg) del vector que codifica para la COX-2 (pOSML-COX-2) y adicionalmente con 1 μg de los diferentes vectores reporteros: pTOP-FLASH, pFOP-FLASH, pLuc-1710 (promotor de survivina que contiene 3 TBE), pLuc-420-3M (promotor de survivina que posee 2 TBEs y donde un sitio crítico está mutado), pGL3-COX-2 (promotor de cox-2) y pGL3 (vector vacío), respectivamente. A las 24 h post-transfección, se realizó extractos proteicos y se determinó la actividad luciferasa. Los datos se expresaron como la razón entre la actividad pTOP-FLASH/pFOP-FLASH, pLuc-1710/pLuc-420-3M y pGL3-COX-2/pGL3 para cada condición. Las barras muestran datos (promedio ± SEM) obtenidos de tres experimentos independientes. Los valores fueron normalizados a un 100 % con los valores obtenidos de células no transfectadas. Las diferencias estadísticamente significativas respecto a sus células controles se indican como #, p<0,05; *, p<0,01.
Act
ivid
ad lu
cife
rasa
(%)
pOSML-COX-2 (μg) 0 0 00
100
200
300
400
500
600
**
*
## ## # #
Act
ivid
ad lu
cife
rasa
(%)
pOSML-COX-2 (μg) 0 0 00
100
200
300
400
500
600
**
*
## ## # #
pLuc1710/ pLuc420-3M
pGL3-COX-2/ pGL3
TOP/ FOP
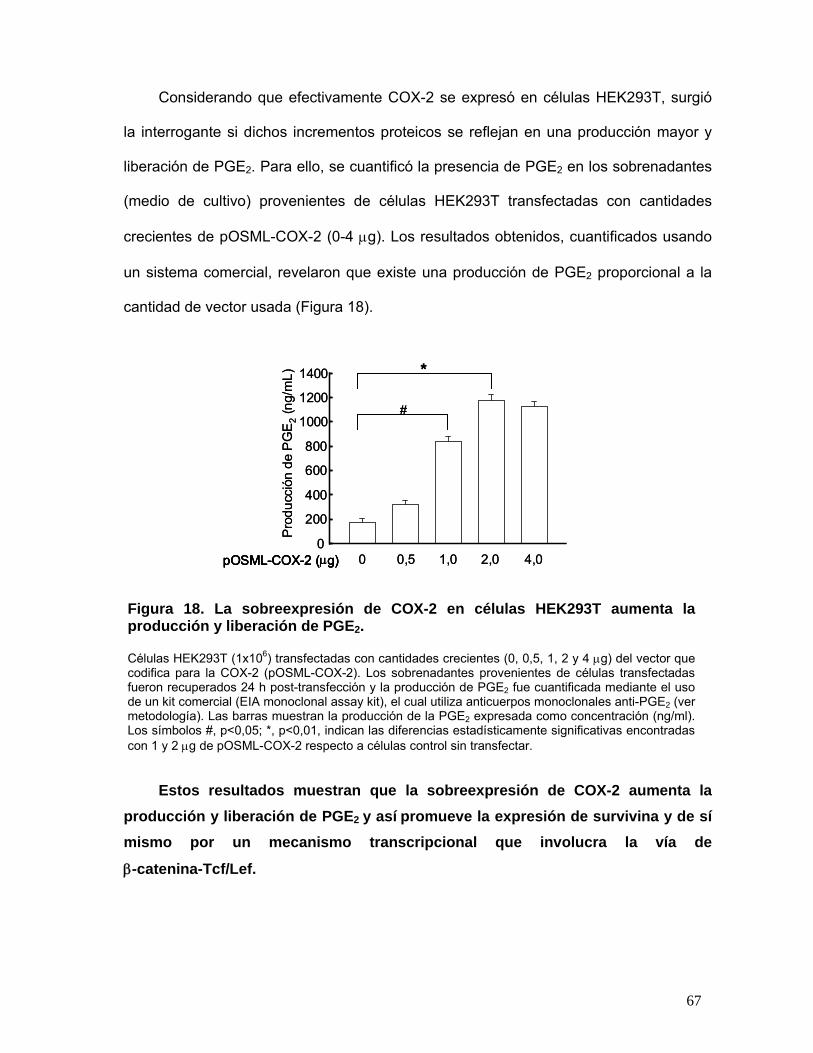
67
Considerando que efectivamente COX-2 se expresó en células HEK293T, surgió
la interrogante si dichos incrementos proteicos se reflejan en una producción mayor y
liberación de PGE2. Para ello, se cuantificó la presencia de PGE2 en los sobrenadantes
(medio de cultivo) provenientes de células HEK293T transfectadas con cantidades
crecientes de pOSML-COX-2 (0-4 μg). Los resultados obtenidos, cuantificados usando
un sistema comercial, revelaron que existe una producción de PGE2 proporcional a la
cantidad de vector usada (Figura 18).
Estos resultados muestran que la sobreexpresión de COX-2 aumenta la producción y liberación de PGE2 y así promueve la expresión de survivina y de sí mismo por un mecanismo transcripcional que involucra la vía de
β-catenina-Tcf/Lef.
0
200
400
600
800
1000
1200
1400
pOSML-COX-2 (μg)
Pro
ducc
ión
de P
GE
2(n
g/m
L)
#
*
pOSML-COX-2 (μg) 0 0,5 1,0 2,0 4,00
200
400
600
800
1000
1200
1400
pOSML-COX-2 (μg)
Pro
ducc
ión
de P
GE
2(n
g/m
L)
#
*
pOSML-COX-2 (μg) 0 0,5 1,0 2,0 4,0
Figura 18. La sobreexpresión de COX-2 en células HEK293T aumenta la producción y liberación de PGE2. Células HEK293T (1x106) transfectadas con cantidades crecientes (0, 0,5, 1, 2 y 4 μg) del vector que codifica para la COX-2 (pOSML-COX-2). Los sobrenadantes provenientes de células transfectadas fueron recuperados 24 h post-transfección y la producción de PGE2 fue cuantificada mediante el uso de un kit comercial (EIA monoclonal assay kit), el cual utiliza anticuerpos monoclonales anti-PGE2 (ver metodología). Las barras muestran la producción de la PGE2 expresada como concentración (ng/ml). Los símbolos #, p<0,05; *, p<0,01, indican las diferencias estadísticamente significativas encontradas con 1 y 2 μg de pOSML-COX-2 respecto a células control sin transfectar.

68
2.2. Determinar el efecto de PGE2 sobre la actividad transcripcional de β-catenina-
Tcf/Lef y sobre la expresión de survivina
Los resultados obtenidos mediante transfecciones con COX-2 nos sugirió que
parte de los efectos de COX-2 sobre la actividad transcripcional de β-catenina-Tcf/Lef y
por ende sobre la expresión de survivina, podrían estar mediados por una mayor
producción y liberación de PGE2, ya que se ha descrito que PGE2 puede activar la vía
de β-catenina-Tcf/Lef por la estimulación del receptor EP2 (Castellone et al., 2005;
Castellone et al., 2006). Para confirmar esta interpretación, se decidió investigar si sólo
la adición de PGE2 al medio de cultivo era suficiente para aumentar la expresión de
survivina por el mecanismo que involucra una activación transcripcional como fuera
observado al sobreexpresar COX-2.
Para ello, células HEK293T se deprivaron de suero e incubaron en presencia o
ausencia de PGE2. La presencia PGE2 aumentó significativamente los niveles de mRNA
de COX-2 y survivina medido por RT-PCR (Figura 19A) y corroborados por PCR en
tiempo real para COX-2 (Figura 19B) y survivina (Figura 19C). Posteriormente, se
investigó si dichos aumentos de mRNA de COX-2 y survivina se deberían a un aumento
transcripcional. Células HEK293T fueron transfectadas con los diferentes plasmidios
reporteros TOP-FLASH (β-catenina-Tcf/Lef) y específicamente aquellos que contienen
los promotores de COX-2 y survivina (Figura 20).
Los resultados revelaron que la incubación con PGE2 aumenta significativamente
la actividad de los reporteros, confirmando la hipótesis de que COX-2 promueve su
propia expresión a través de su actividad enzimática, efecto que aparentemente está
mediado por una mayor producción y liberación de PGE2 (Figura 20).

69
Figura 20. La incubación de células HEK293T con PGE2 aumenta la actividad de reporteros de β-catenina-Tcf/Lef, survivina y COX-2. Células HEK293T (1x106) fueron transfectadas con 1 μg de los diferentes vectores reporteros: pTOP-FLASH, pFOP-FLASH, pLuc-1710, pLuc-420-3M, pGL3-COX-2 o pGL3 (vector vacío). Posteriormente, las células se deprivaron de suero durante 12 h e incubaron 4 h con PGE2 (5 μM) en medio sin suero. A las 24 h post-transfección se realizó extractos proteicos y se determinó la actividad luciferasa. Los datos fueron expresados por cada condición como la razón entre la actividad pTOP-FLASH/pFOP-FLASH, pLuc-1710/pLuc-420-3M y pGL3-COX-2/pGL3. Las barras en blanco muestran células sin tratar y las barras en negro aquellas incubadas con PGE2 (5 μM). Los resultados corresponden al promedio ± SEM de tres experimentos independientes. Los valores se normalizaron con respecto a los valores obtenidos de células control (100%). Las diferencias estadísticamente significativas respecto células controles se indican como *, p<0,01; #, p<0,05.
PGE2 (5 μM) +-COX-2
survivina
actina
1,00 1,40
1,00 2,00
#
#
PGE2 (5 μM) +-COX-2
survivina
actina
1,00 1,40
1,00 2,00
#
#
+-
*
0
100
200
300
400
500
600
700
+-
*
0
100
200
300
400
500
600
700
#
0
100200
300
400
500
600
700
+-
#
0
100200
300
400
500
600
700
+-PGE2 (5 μM)
mR
NA
sur
vivi
na (%
)
PGE2 (5 μM)
A B C
Figura 19. PGE2 aumenta la expresión de survivina y COX-2. Células HEK293T se deprivaron de suero durante 12 h y posteriormente fueron incubadas 4 h con PGE2 (5 μM) en medio sin suero. Cumplido el tratamiento, se aisló el RNA total (ver metodología) y los niveles de mRNA de COX-2 y survivina fueron determinados mediante (A) RT-PCR semicuantitativo usando actina como control o mediante PCR en tiempo real (B y C). (A) Los niveles relativos de mRNA estimados mediante análisis densitométrico fueron: 2,0 ± 0,03 para survivina y 1,4 ± 0,06 para COX-2. (B y C) Cuantificación del mRNA de COX-2 y survivina mediante PCR en tiempo real (ver protocolo en metodología). La obtención de las muestras se llevó a cabo del mismo modo que en (A). Se utilizó RNA ribosomal 18S como control interno de carga. Los resultados corresponden al promedio ± SEM de tres experimentos independientes. Los valores se normalizaron con respecto a los valores obtenidos de células control (100%). Las diferencias estadísticamente significativas se obtuvieron comparando células tratadas con células control sólo incubadas en ausencia de suero y se indican como *, p<0.01; #, p<0.05.
mR
NA
CO
X-2
(%)
0
50
100
150
200
250
300
- + - + - +
#*
#
- + - + - +
#*
#
PGE2 (5 μM)
Activ
idad
luci
fera
sa(%
)
0
50
100
150
200
250
300
- + - + - +
#*
#
- + - + - +
#*
#
PGE2 (5 μM)
Activ
idad
luci
fera
sa(%
)
pLuc1710/ pLuc420-3M
pGL3-COX-2/ pGL3 TOP/ FOP

70
Posteriormente, se investigó si dichos incrementos a nivel transcripcional de
COX-2 y survivina por PGE2 se reflejan en mayores niveles proteicos de COX-2,
β-catenina y survivina. Para ello, células HEK293T fueron tratadas con PGE2 y
posteriormente se obtuvo extractos proteicos. Los resultados confirman que la
presencia de PGE2 aumenta significativamente los niveles proteicos de estas tres
proteínas (Figura 21A). Además, se investigó si tal incremento en los niveles de
survivina y COX-2 se correlacionan con incrementos en la proliferación, ya que se ha
descrito previamente que PGE2 promueve el crecimiento y viabilidad en células de
cáncer de colon (Shao et al., 2003). Nuestros datos revelaron que la adición de PGE2
aumenta significativamente la proliferación de células HEK293T (Figura 21B).
Figura 21. PGE2 aumenta los niveles proteicos de survivina y COX-2 y se correlaciona con un aumento en la proliferación de células HEK293T. Células HEK293T se deprivaron de suero durante 12 h y posteriormente incubaron 4 h con PGE2 (5 μM) en medio sin suero. (A) Extractos proteicos obtenidos después del tratamiento (80 μg/ carril) fueron separados mediante SDS-PAGE y posteriormente analizados por inmunoblot usando anticuerpos específicos anti-COX-2, anti-β-catenina, anti-survivina o anti-actina. Los datos numéricos se obtuvieron mediante un análisis densitométrico de las bandas. Los valores indicados entre paneles se normalizaron contra actina y corresponden al promedio de tres experimentos independientes. Los valores relativos (promedio ± SEM) para COX-2, β-catenina y survivina comparado con células control fueron: 1,55 ± 0,03, 1,35 ± 0,05 y 1,74 ± 0,04, respectivamente. (B) La proliferación celular fue cuantificada mediante ensayos de MTS en células HEK293T cultivadas 12 h en ausencia de suero y luego incubadas 16 h en medio sin suero con PGE2 (5 μM). Las barras en blanco corresponden a células control sin PGE2 y las barras en negro aquellas incubadas en presencia de PGE2 (5 μM). Los valores corresponden a tres experimentos independientes (promedio ± SEM). Las diferencias estadísticamente significativas se obtuvieron comparando células tratadas con células control sólo incubadas en ausencia de suero (#, p<0.05).
+-COX-2
β-catenina
survivina
actina
PGE2 (5 μM)
1.00 1.55
1.00 1.35
1.00 1.74
#
#
#
+-COX-2
β-catenina
survivina
actina
PGE2 (5 μM)
1.00 1.55
1.00 1.35
1.00 1.74
#
#
#
Pro
lifer
ació
n(%
)
+-PGE2 (5 μM)
#
0
100
200
300
400
Pro
lifer
ació
n(%
)
+-PGE2 (5 μM)
#
0
100
200
300
400
A B

71
En resumen, la adición de PGE2 en células HEK293T promueve la expresión de survivina y COX-2 y aumenta la proliferación por un mecanismo que involucra
la activación de la vía de β-catenina-Tcf/Lef.
2.3. Determinar el efecto de la PGE2 o bien la sobreexpresión de COX-2 sobre la
expresión de survivina y la proliferación en células que expresan caveolina-1
Los resultados previos de nuestros experimentos demostraron que la presencia de
caveolina-1 disminuye significativamente los niveles de mRNA y proteicos de COX-2
(Figuras 9 y 10). Por lo tanto, nos preguntamos si tal disminución en la expresión de
COX-2, conlleva a menores niveles de PGE2. Como se esperaba, los niveles de PGE2
cuantificados en los sobrenadantes de células de embrionarias humanas (HEK293T),
cáncer de mama (ZR75) y de colon [DLD-1; HT29(ATCC)] que expresan ectópicamente
caveolina-1 presentaron niveles reducidos de PGE2 comparado con células control
transfectadas con el vector vacío (Figura 22A-D).
Posteriormente, los sobrenadantes (medio de cultivo) se recolectaron y los niveles de PGE2 se cuantificaron mediante un ensayo comercial “PGE2 EIA monoclonal kit” (ver metodología). Los datos muestran el promedio de tres experimentos independientes (promedio ± SEM). Las diferencias estadísticamente significativas respecto a células control (placIOP, (-)) se indican como #, p<0.05.
DLD-1
+-Cav-1
#
0200400600800
1000120014001600
ZR75
+-Cav-10
200
400
600
800
1000
1200 HT29(ATCC)
+-Cav-1
#
0500
10001500200025003000
HEK293T
+-Cav-1
Pro
ducc
ión
de P
GE
2(n
g/m
L)
0
50100
150
200
250
300
#
Prod
ucci
ón d
e PG
E2
(ng/
mL)
Pro
ducc
ión
de P
GE
2(n
g/m
L)
Pro
ducc
ión
de P
GE
2(n
g/m
L)
DLD-1
+-Cav-1
#
0200400600800
1000120014001600
ZR75
+-Cav-10
200
400
600
800
1000
1200 HT29(ATCC)
+-Cav-1
#
0500
10001500200025003000
HEK293T
+-Cav-1
Pro
ducc
ión
de P
GE
2(n
g/m
L)
0
50100
150
200
250
300
#
Prod
ucci
ón d
e PG
E2
(ng/
mL)
Pro
ducc
ión
de P
GE
2(n
g/m
L)
Pro
ducc
ión
de P
GE
2(n
g/m
L)
A C
B D
Figura 22. La expresión de caveolina-1 disminuye la producción de PGE2. Inmunodetección y cuantificación de la producción de PGE2. Células HEK293T (A), células de cáncer (B) ZR75, (C) DLD1 o (D) HT29(ATCC) transfectadas con los plasmidios pLacIOP (-), barras blancas o pLacIOP-caveolina-1 (+), barras negras, fueron cultivadas en placas de 60 mm (2x106) y crecidas en todos los casos por 24 h en presencia de IPTG 1 mM.

72
Estudios previos de nuestro laboratorio establecieron que la re-expresión de
caveolina-1 en células de cáncer de mama (ZR75) y de colon [DLD-1; HT29(ATCC)]
reduce significativamente la expresión de survivina (Torres et al., 2006). Además, en
esta tesis se ha demostrado que PGE2 aumenta la expresión de survivina, entonces se
investigó si la presencia de PGE2 revierte la regulación negativa de caveolina-1 sobre la
expresión de COX-2 y de survivina en células de cáncer de mama (ZR75) y de colon
[DLD-1; HT29(ATCC)].
En las tres líneas celulares estudiadas, la adición de PGE2 restauró los niveles de
mRNA de COX-2 y survivina que fueron cuantificados por RT-PCR semicuantitativo
(Figura 23A). En el caso de COX-2, la inhibición de los niveles de mRNA por la
presencia de caveolina-1 y la posterior reversión causada por la adición de PGE2 fue
también corroborada por PCR en tiempo real (Figura 23B). Además, se observó el
mismo efecto al examinar los niveles proteicos de COX-2 y survivina mediante
experimentos de Western blot. Adicionalmente, se observó en todas las líneas celulares
un aumento en la cantidad proteica de β-catenina al agregar PGE2, también en
presencia de caveolina-1 (Figura 23C).
Con el fin de revertir la regulación negativa de caveolina-1, células de cáncer de
colon DLD-1 transfectadas establemente con pLacIOP (M1) o pLacIOP-caveolina-1
(C4) fueron adicionalmente transfectadas en forma transitoria con el plasmidio
pOSML-COX-2 (2 μg).

73
HT29(ATCC)DLD-1
PGE2
+-Cav-1 +-- - + +
+ +
- - + +
- -
survivina
COX-2
Cav-1
actina
1,0 0,6 1,8 0,9
1,0 0,7 0,9 1,3
ZR75+ +
- - + +
- -
1,0 0,7 1,4 1,6
1,0 0,7 0,8 0,9 #
# 1,0 0,4 2,5 0,8
1,0 0,6 1,9 1,4 #
#
HT29(ATCC)DLD-1
PGE2
+-Cav-1 +-- - + +
+ +
- - + +
- -
survivina
COX-2
Cav-1
actina
1,0 0,6 1,8 0,9
1,0 0,7 0,9 1,3
ZR75+ +
- - + +
- -
1,0 0,7 1,4 1,6
1,0 0,7 0,8 0,9 #
# 1,0 0,4 2,5 0,8
1,0 0,6 1,9 1,4 #
#1,0 0,4 2,5 0,8
1,0 0,6 1,9 1,4 #
#
HT29(ATCC)
PGE2
+-Cav-1 +-- - + +
CO
X-2
mR
NA
(%)
050
100150200250300350
050
100150200250300350
PGE2
+-Cav-1 +-- - + +
CO
X-2
mR
NA
(%)
*DLD-1
CO
X-2
mR
NA
(%)
0200400
1200
PGE2
+Cav-1 +
- - + +
- -
*
CO
X-2
mR
NA
(%)
0200400
1200
0200400
1200
PGE2
+Cav-1 +
- - + +
- -
DLD-1ZR75
CO
X-2
mR
NA
(%)
PGE2
+Cav-1 +
- - + +
- -
#
CO
X-2
mR
NA
(%)
0
50100150200250
0
50100150200250
PGE2
+Cav-1 +
- - + +
- -
#ZR75 HT29(ATCC)
PGE2
+-Cav-1 +-- - + +
CO
X-2
mR
NA
(%)
050
100150200250300350
050
100150200250300350
PGE2
+-Cav-1 +-- - + +
CO
X-2
mR
NA
(%)
*HT29(ATCC)
PGE2
+-Cav-1 +-- - + +
CO
X-2
mR
NA
(%)
050
100150200250300350
050
100150200250300350
PGE2
+-Cav-1 +-- - + +
CO
X-2
mR
NA
(%)
*DLD-1
CO
X-2
mR
NA
(%)
0200400
1200
PGE2
+Cav-1 +
- - + +
- -
*
CO
X-2
mR
NA
(%)
0200400
1200
0200400
1200
PGE2
+Cav-1 +
- - + +
- -
DLD-1DLD-1
CO
X-2
mR
NA
(%)
0200400
1200
0200400
1200
PGE2
+Cav-1 +
- - + +
- -
*
CO
X-2
mR
NA
(%)
0200400
1200
0200400
1200
PGE2
+Cav-1 +
- - + +
- -
DLD-1ZR75
CO
X-2
mR
NA
(%)
PGE2
+Cav-1 +
- - + +
- -
#
CO
X-2
mR
NA
(%)
0
50100150200250
0
50100150200250
PGE2
+Cav-1 +
- - + +
- -
#ZR75ZR75
CO
X-2
mR
NA
(%)
PGE2
+Cav-1 +
- - + +
- -
#
CO
X-2
mR
NA
(%)
0
50100150200250
0
50100150200250
PGE2
+Cav-1 +
- - + +
- -
#ZR75
PGE2
+-Cav-1 +-- - + +
1,0 0,9 1,8 2,1
1,0 0,4 0,9 1,1
1.0 0,2 2,1 1,6
survivina
COX-2
Cav-1
actina
β-catenina
#
#
HT29(ATCC)+ +
- - + +
- -
1,0 0,7 1,3 1,2
1,0 0,6 1,1 1,1
1,0 1,3 1,7 1,9
#
#
DLD-1+ +
- - + +
- -
1,0 0,5 0,9 0,8
1,0 0,8 3,6 2,9
1,0 1,4 1,7 1,6
#
ZR75
PGE2
+-Cav-1 +-- - + +
1,0 0,9 1,8 2,1
1,0 0,4 0,9 1,1
1.0 0,2 2,1 1,6
survivina
COX-2
Cav-1
actina
β-catenina
#
#
HT29(ATCC)+ +
- - + +
- -
1,0 0,7 1,3 1,2
1,0 0,6 1,1 1,1
1,0 1,3 1,7 1,9
#
#
DLD-1+ +
- - + +
- -
1,0 0,7 1,3 1,2
1,0 0,6 1,1 1,1
1,0 1,3 1,7 1,9
#
#
DLD-1+ +
- - + +
- -
1,0 0,5 0,9 0,8
1,0 0,8 3,6 2,9
1,0 1,4 1,7 1,6
#
ZR75+ +
- - + +
- -
1,0 0,5 0,9 0,8
1,0 0,8 3,6 2,9
1,0 1,4 1,7 1,6
#
ZR75
A
B
C
Figura 23. PGE2 revierte la inhibición de la expresión de COX-2 y survivina por caveolina-1 en líneas celulares de cáncer. Las líneas celulares de cáncer de mama (ZR75) y de colon [DLD-1; HT29(ATCC)] transfectadas establemente con pLacIOP (-) o pLacIOP-caveolina-1 (+) fueron incubadas 24 h en presencia de IPTG 1 mM o alternativamente con IPTG 1 mM y 4 h con PGE2 20 μM en medio sin suero. (A) RT-PCR semicuantitativo del mRNA de COX-2, survivina, caveolina-1 y actina. La amplificación de actina fue usada como control interno. Los datos numéricos entre paneles, obtenidos mediante análisis densitométrico, corresponden al promedio de tres experimentos independientes normalizados con actina. (B) Análisis del mRNA de COX-2 mediante PCR en tiempo real de células ZR75, DLD-1 y HT29(ATCC) tratadas como fue descrito al inicio. Las barras en blanco representan células control (pLacIOP, (-)) y barras en negro, células que expresan caveolina-1 (pLacIOP-caveolina-1, (+)). Como control de carga se utilizó el RNA ribosomal 18S. Los datos corresponden al promedio de tres experimentos independientes (promedio ± SEM), expresando como un 100% a células control (pLacIOP, (-)) sin PGE2. (C) Western blot de las mismas muestras descritas anteriormente. El contenido de proteínas totales (80 μg/ carril) fue separado mediante SDS-PAGE y posteriormente analizado por inmunoblot usando anticuerpos específicos anti-COX-2, anti-β-catenina, anti-survivina o anti-actina. Los datos numéricos se normalizaron contra actina y corresponden al promedio de tres experimentos independientes. Se indican las diferencias estadísticamente significativas encontradas en (A-C) y corresponden a la comparación de células transfectadas con pLacIOP-caveolina-1 cultivadas en ausencia o presencia de PGE2 (20 μM) (*,p<0,01; #, p<0.05).

74
Interesantemente, la sobreexpresión de COX-2 previene la inhibición de survivina
por caveolina-1, ya que la transfección con pOSML-COX-2 (2 μg) incrementó los niveles
de mRNA (Figura 24A), la actividad transcripcional de β-catenina-Tcf/Lef (Figura 24B) y
específicamente del promotor de survivina (Figura 24C).
Para investigar si dicho aumento transcripcional se correlaciona con mayores
niveles proteicos de COX-2 y por ende mayor producción y liberación de PGE2, se
realizó experimentos de Western blot para analizar COX-2 y se cuantificó los niveles de
PGE2 presentes en los sobrenadantes de células DLD-1 transfectadas con
pOSML-COX-2 y pLacIOP (M1) o pLacIOP-caveolina-1 (C4).
La figura 25A muestra que la transfección ectópica de células DLD-1 con
pOSML-COX-2 (2 μg) revierte la inhibición de survivina y restablece la producción de
PGE2 incluso en aquellas células que expresan caveolina-1 (Figura 25B).
Figura 24. La sobreexpresión de COX-2 previene la inhibición de la expresión de survivina por caveolina-1 en células DLD-1. Células DLD-1 transfectadas establemente con pLacIOP (M1) o pLacIOP-caveolina-1 (C4) fueron adicionalmente transfectadas con 2 μg del vector que codifica para COX-2 (pOSML-COX-2) y cultivadas 24 h en presencia de IPTG 1 mM. (A) RT-PCR semicuantitativo del mRNA de COX-2, survivina y actina. La amplificación de actina fue usada como control interno. Células DLD-1 (M1) y (C4) adicionalmente co-transfectadas con 1 μg de los diferentes vectores reporteros: pTOP-FLASH y pFOP-FLASH (B), pLuc-1710 y pLuc-420-3M (C), respectivamente. Los datos numéricos se normalizaron con el control de cada experimento (100%) y corresponden al promedio de tres experimentos independientes (promedio ± SEM). Se indican las diferencias estadísticamente significativas encontradas en (B y C) y corresponden a compareción entre células transfectadas con pLacIOP-caveolina-1 versus aquellas co-transfectadas con pLacIOP-caveolina-1 y pOSML-COX-2 (2 μg) (#, p<0.05).
A B C RT-PCR
++
actina
+- -Cav-1pOSML-COX-2 - - +
survivina1.0 0.4 1.6 1.7
COX-21.0 0.4 12.4 14.1
RT-PCR
++
actina
+- -Cav-1pOSML-COX-2 - - +
survivina1.0 0.4 1.6 1.7
COX-21.0 0.4 12.4 14.1
++
actina
+- -Cav-1pOSML-COX-2 - - +
survivina1.0 0.4 1.6 1.7
COX-21.0 0.4 12.4 14.1
+- +-Cav-1pOSML-COX-2 - - ++
*
050
100150200250300350
Act
ivid
ad lu
cife
rasa
(%)
pLuc1710/ pLuc420-3M
+- +-Cav-1pOSML-COX-2 - - ++
*
050
100150200250300350
Act
ivid
ad lu
cife
rasa
(%)
pLuc1710/ pLuc420-3M
Act
ivid
adlu
cife
rasa
(%)
+- +-Cav-1pOSML-COX-2 - - ++
#
020406080
100120140160
TOP/FOP
Act
ivid
adlu
cife
rasa
(%)
+- +-Cav-1pOSML-COX-2 - - ++
#
020406080
100120140160
Act
ivid
adlu
cife
rasa
(%)
+- +-Cav-1pOSML-COX-2 - - ++
#
020406080
100120140160
TOP/FOP

75
Por otra parte, utilizando la misma estrategia experimental se evaluó los efectos
de la sobreexpresión de COX-2 sobre la proliferación de células DLD-1 que expresan o
no caveolina-1. Nuestros resultados muestran que la sobreexpresión de COX-2
previene la inhibición de la proliferación causada por la expresión de caveolina-1
(Figura 25C).
Figura 25. La sobreexpresión de COX-2 en células DLD-1 previene la inhibición de la expresión de survivina, la producción de PGE2 y la proliferación causada por caveolina-1. Células transfectadas establemente con pLacIOP o pLacIOP-caveolina-1 se transfectaron adicionalmente con 2 μg del vector que codifica para la COX-2 (pOSML-COX-2) y cultivaron 24 h en presencia de IPTG 1 mM. (A) Análisis de Western blot de los lisados tratados según descrito anteriormente. El contenido de proteínas totales (50 μg/ carril) fue separado mediante SDS-PAGE y posteriormente analizado por inmunoblot usando anticuerpos específicos anti-COX-2, anti-β-catenina, anti-survivina y anti-actina. Los datos numéricos entre paneles fueron obtenidos mediante análisis densitométrico y corresponden al promedio de dos experimentos independientes normalizados con actina. (B) Inmunodetección y cuantificación de la producción de PGE2. A las 24 h post-transfección, los sobrenadantes (medio de cultivo) se recolectaron para cuantificar los niveles de PGE2 mediante un ensayo comercial (ver mas detalle en metodología). Las barras blancas muestran los niveles de PGE2 provenientes de células DLD-1 control, en cambio las barras negras muestran los niveles de PGE2 en células DLD-1 que expresan caveolina-1. Los valores corresponden a tres experimentos independientes (promedio ± SEM). Las diferencias estadísticamente significativas fueron obtenidas comparando células control o aquellas que expresan caveolina-1, ambas en ausencia y presencia de PGE2 como se indica (*,p<0,01; #, p<0.05). (C) Análisis de la proliferación celular mediante ensayos de MTS en células DLD-1 co-transfectadas como fue descrito mas arriba. Post-transfección, las células (1x104) fueron sembradas en placas de 96 pocillos y cultivadas 24 h en presencia de IPTG 1 mM. Los datos numéricos fueron normalizados con respecto a células control (100%) y corresponden al promedio de tres experimentos independientes (promedio ± SEM). El valor de P (p=0.054) indica la diferencia entre células que expresan pLacIOP-caveolina-1 versus aquellas que expresan pLacIOP-caveolina-1/pOSML-COX-2.
++
18.0
COX-2
Cav-1
actina
+- -Cav-1
WBpOSML-COX-2 (2 μg) - - +
survivina1.0 0.3 22.0
1.0 0.5 1.7 2.0
++
18.0
COX-2
Cav-1
actina
+- -Cav-1
WBpOSML-COX-2 (2 μg) - - +
survivina1.0 0.3 22.0
1.0 0.5 1.7 2.0
A
B
C
Pro
ducc
ion
de P
GE
2(n
g/m
L)
+- +-Cav-1pOSML-COX-2 (2 μg) - - ++
#
*
0200400600800
100012001400
Pro
ducc
ion
de P
GE
2(n
g/m
L)
+- +-Cav-1pOSML-COX-2 (2 μg) - - ++
#
*
0200400600800
100012001400
+- +-Cav-1pOSML-COX-2 (2 μg) - - ++
Pro
lifer
ació
n (%
)
p=0.054
020406080
100120140
+- +-Cav-1pOSML-COX-2 (2 μg) - - ++
Pro
lifer
ació
n (%
)
p=0.054
020406080
100120140
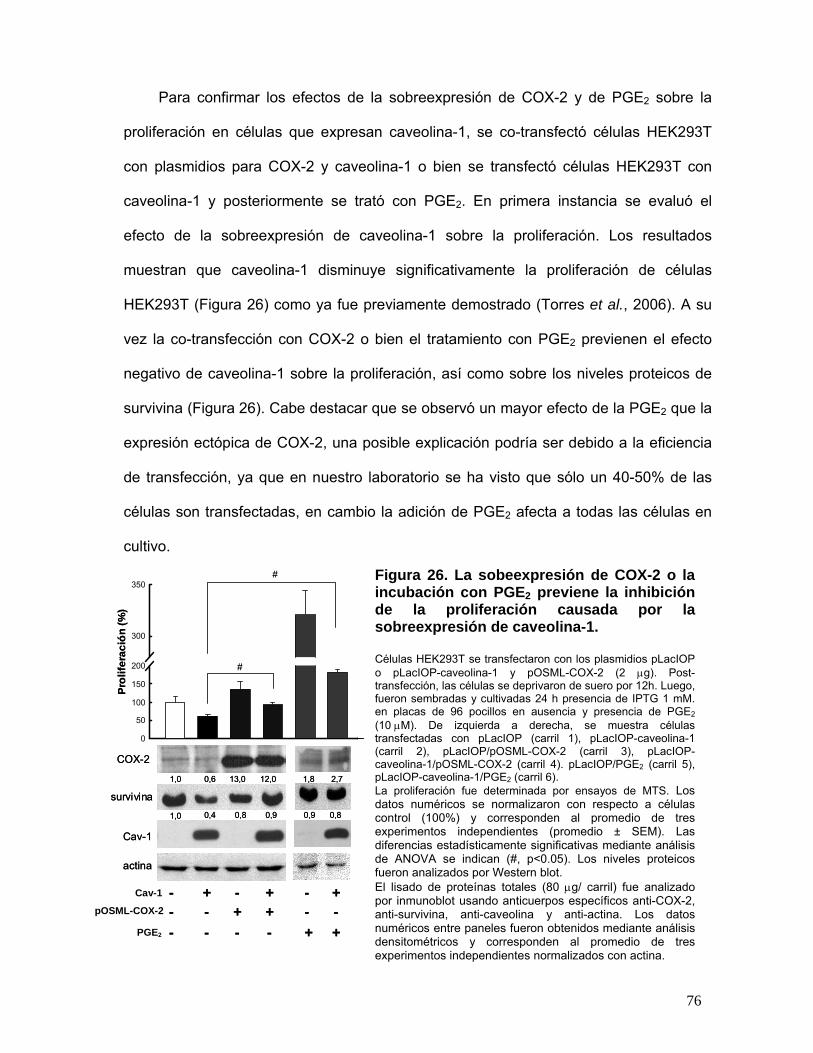
76
Para confirmar los efectos de la sobreexpresión de COX-2 y de PGE2 sobre la
proliferación en células que expresan caveolina-1, se co-transfectó células HEK293T
con plasmidios para COX-2 y caveolina-1 o bien se transfectó células HEK293T con
caveolina-1 y posteriormente se trató con PGE2. En primera instancia se evaluó el
efecto de la sobreexpresión de caveolina-1 sobre la proliferación. Los resultados
muestran que caveolina-1 disminuye significativamente la proliferación de células
HEK293T (Figura 26) como ya fue previamente demostrado (Torres et al., 2006). A su
vez la co-transfección con COX-2 o bien el tratamiento con PGE2 previenen el efecto
negativo de caveolina-1 sobre la proliferación, así como sobre los niveles proteicos de
survivina (Figura 26). Cabe destacar que se observó un mayor efecto de la PGE2 que la
expresión ectópica de COX-2, una posible explicación podría ser debido a la eficiencia
de transfección, ya que en nuestro laboratorio se ha visto que sólo un 40-50% de las
células son transfectadas, en cambio la adición de PGE2 afecta a todas las células en
cultivo.
0
50
100
150
200
300
350
Prol
ifera
ción
(%)
COX-2
PGE2
---
+--
+-
-+-
+ - +- -+ +
COX-2
survivina
Cav-1
actina
#
#
1,0
1,0
0,6
0,4
13,0 12,0
0,8 0,9 0,9 0,8
2,71,8
Cav-1
0
50
100
150
200
300
350
Prol
ifera
ción
(%)
COX-2
PGE2
---
+--
+-
-+-
+ - +- -+ +
COX-2
survivina
Cav-1
actina
#
#
1,0
1,0
0,6
0,4
13,0 12,0
0,8 0,9 0,9 0,8
2,71,8
Cav-1
Figura 26. La sobeexpresión de COX-2 o la incubación con PGE2 previene la inhibición de la proliferación causada por la sobreexpresión de caveolina-1. Células HEK293T se transfectaron con los plasmidios pLacIOP o pLacIOP-caveolina-1 y pOSML-COX-2 (2 μg). Post-transfección, las células se deprivaron de suero por 12h. Luego, fueron sembradas y cultivadas 24 h presencia de IPTG 1 mM. en placas de 96 pocillos en ausencia y presencia de PGE2 (10 μM). De izquierda a derecha, se muestra células transfectadas con pLacIOP (carril 1), pLacIOP-caveolina-1 (carril 2), pLacIOP/pOSML-COX-2 (carril 3), pLacIOP-caveolina-1/pOSML-COX-2 (carril 4). pLacIOP/PGE2 (carril 5), pLacIOP-caveolina-1/PGE2 (carril 6). La proliferación fue determinada por ensayos de MTS. Los datos numéricos se normalizaron con respecto a células control (100%) y corresponden al promedio de tres experimentos independientes (promedio ± SEM). Las diferencias estadísticamente significativas mediante análisis de ANOVA se indican (#, p<0.05). Los niveles proteicos fueron analizados por Western blot. El lisado de proteínas totales (80 μg/ carril) fue analizado por inmunoblot usando anticuerpos específicos anti-COX-2, anti-survivina, anti-caveolina y anti-actina. Los datos numéricos entre paneles fueron obtenidos mediante análisis densitométricos y corresponden al promedio de tres experimentos independientes normalizados con actina.
pOSML-COX-2
PGE2
Cav-1

77
Estos resultados confirman la hipótesis que la reversión de los efectos
negativos de caveolina-1 causada por la sobreexpresión de COX-2 sobre la
actividad transcripcional de β-catenina-Tcf/Lef son mediados en parte por PGE2.
2.4. Investigar el efecto de la inhibición de COX-2 sobre la proliferación en células
de cáncer de colon [HT29(ATCC) y DLD-1] que expresan caveolina-1
Diversos estudios enfocados en cáncer de colon han demostrado que la
administración de analgésicos anti-inflamatorios no esteroidales disminuye o previene el
desarrollo tumoral, tanto en modelos animales (DuBois et al., 1996), como en humanos
(Hull, 2005; Chan et al., 2007). Con estos antecedentes, sumado al papel de
caveolina-1 como supresor de tumores (Quest et al., 2008) investigamos en nuestro
modelo, el efecto de la inhibición selectiva de la COX-2 en células que expresan
caveolina-1.
Para ello, células DLD-1 y HT29(ATCC) que expresan o no caveolina-1 fueron
incubadas en ausencia o presencia del inhibidor específico de COX-1
(FR122047, 10 μM) o COX-2 (SC-971, 20 μM). La proliferación fue evaluada por
ensayos de exclusión con azul de tripan y mediante ensayos de MTS.
Los resultados muestran que caveolina-1 disminuye la proliferación en ambas
líneas celulares (Figura 27; Figura 28A y 28B; comparar barras 1 y 2) y este efecto fue
potenciado por la incubación con el inhibidor especifico de COX-2, pero no con el
inhibidor de COX-1 (Figura 28A y 28B; comparar barras 2, 6 y 10). Cabe destacar, que
la adición de PGE2 previene la reducción observada en la proliferación debido a la
presencia de caveolina-1 o la presencia de los inhibidores de COX-1 y COX-2, o bien la
combinación de ambas condiciones (Figura 2 y Figura 28A y 28B; comparar barras
4, 8 y 12).

78
Figura 28. PGE2 previene la inhibición de la proliferación por la expresión de caveolina-1. Células (A) DLD-1 o (B) HT29(ATCC) transfectadas establemente con pLacIOP (-) o pLacIOP-caveolina-1 (+) fueron deprivadas de suero durante 4 h e incubadas 3 h en ausencia o presencia de PGE2 (20 μM) en medio sin suero. Posteriormente, las células fueron sub-cultivadas por 24 h adicionales en placas de 96 pocillos, en medio completo con IPTG 1 mM o en conjunto con el inhibidor de COX-1 (10 μM) o bien con el inhibidor de COX-2 (20 μM). La proliferación celular fue determinada por el ensayo de MTS®. Los valores indican el porcentaje de actividad residual con respecto a células control (100%), obtenido de un promedio de tres experimentos independientes (promedio ± SEM) Las diferencias estadísticamente significativas encontradas en (A) y (B) se indican como #, p<0.05.
A B HT29(ATCC)
Cav-1PGE2
+-- -
+- +- +- +- +-++ - - ++ - - ++
60
80
100
120
140
Pro
lifer
ació
n(%
)
#
#
#
Inhibidor de COX-1 Inhibidor de COX-2
#
HT29(ATCC)
Cav-1PGE2
+-- -
+- +- +- +- +-++ - - ++ - - ++
60
80
100
120
140
Pro
lifer
ació
n(%
)
#
##
#
Inhibidor de COX-1 Inhibidor de COX-2
#
DLD-1
Cav-1PGE2
+-- -
+- +- +- +- +-++ - - ++ - - ++
60
80
100
120
140#
#
#
Pro
lifer
ació
n(%
)
Inhibidor de COX-1 Inhibidor de COX-2
#
DLD-1
Cav-1PGE2
+-- -
+- +- +- +- +-++ - - ++ - - ++
60
80
100
120
140#
#
#
Pro
lifer
ació
n(%
)
Inhibidor de COX-1 Inhibidor de COX-2
##
Figura 27. Efecto de PGE2 e inhibidores de COX-1 y COX-2 en células DLD-1 Células DLD-1 transfectadas establemente con pLacIOP (-) o pLacIOP-caveolina-1 (+) fueron deprivadas de suero durante 4 h e incubadas 24 h o 48 h en ausencia o presencia de PGE2 (20 μM) en medio completo con IPTG (1 mM). Adicionalmente, bajo el mismo tratamiento, las células se crecieron en presencia de los inhibidores específicos de COX-1 (10 μM) o de COX-2 (20 μM). La proliferación se determinó por conteo de células con azul de tripan. Las barras en blanco corresponden al tratamiento durante 24 h y en negro aquellas tratadas por 48 h. El gráfico representa un experimento independiente realizado por triplicado.
0
1e+5
2e+5
3e+5
4e+5
5e+5
6e+5
7e+5
Cav-1PGE2
Inhibidor de COX-2
+-- -
+-- -
+- +-+ + + +
+-- -
+-- -
+- +-+ + + +
+-- -
+-- -
+- +-+ + + +
Inhibidor de COX-1
24 h 48 h
0
1e+5
2e+5
3e+5
4e+5
5e+5
6e+5
7e+5
Cav-1PGE2
Inhibidor de COX-2
+-- -
+-- -
+- +-+ + + +
+-- -
+-- -
+- +-+ + + +
+-- -
+-- -
+- +-+ + + +
Inhibidor de COX-1
24 h 48 h
0
100
200
300
400
500
600
700
Núm
ero
de c
élul
as (
x103 )

79
En vista a los resultados anteriores, investigamos cual es la concentración minima
requerida del inhibidor de COX-2 (SC-791) para inhibir la actividad de COX-2 y por
ende la actividad transcripcional de β-catenina-Tcf/Lef, y así como también, su efecto
sobre la proliferación. Al mismo tiempo se investigó la contribución de la COX-1 sobre la
activación de dicha vía, para ello se utilizó un inhibidor específico (FR122047). Nuestra
estrategia experimental consistió en utilizar células DLD-1 transfectadas establemente
con pLacIOP (M1) y con pLacIOP-caveolina-1 (C4) e incubarlas con concentraciones
crecientes del inhibidor SC-971 (0, 1, 5 y 20 μM) o bien con el inhibidor FR122047
(10 μM). Posteriormente, se evaluó la producción de PGE2 (cuantificación desde
medios de cultivo) y para estudiar si menores niveles de PGE2 también inciden sobre la
expresión de survivina, se determinó la actividad reportera de β-catenina-Tcf/Lef y se
examinó los niveles proteicos de β-catenina y de survivina.
Los resultados revelaron que efectivamente el inhibidor de COX-2 (SC-791) tuvo
un efecto dependiente de la dosis, ya que al cuantificar la producción de PGE2 se
observó que a concentraciones de 5 μM disminuye los niveles de PGE2 y aun más a
20 μM. Interesantemente, en células que expresan caveolina-1, las mismas
concentraciones del inhibidor llevaron a niveles menores de PGE2 comparado con
células control (Figura 29A). La inhibición de la COX-1 reveló una disminución en la
producción de PGE2 sugiriendo que ambas enzimas contribuyen en la producción de
PGE2. Sin embargo, las diferencias observadas con el inhibidor de COX-1 no fueron
significativas. Así también, se observó el mismo efecto a nivel transcripcional, el cual se
tradujo en una menor actividad transcripcional de β-catenina-Tcf/Lef (Figura 29B). Esto
último se reflejó en una mayor reducción en los niveles proteicos de survivina de células
que expresan caveolina-1 comparado con células control incubadas con los inhibidores
(Figura 29C).
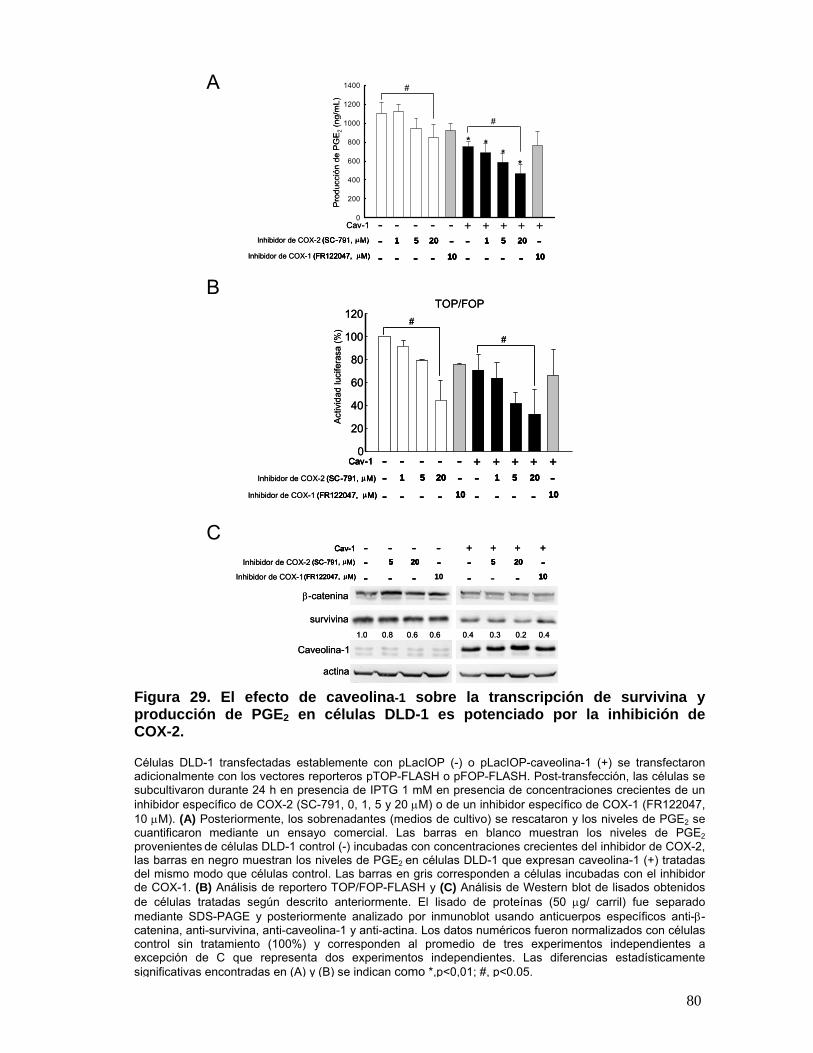
80
Figura 29. El efecto de caveolina-1 sobre la transcripción de survivina y producción de PGE2 en células DLD-1 es potenciado por la inhibición de COX-2. Células DLD-1 transfectadas establemente con pLacIOP (-) o pLacIOP-caveolina-1 (+) se transfectaron adicionalmente con los vectores reporteros pTOP-FLASH o pFOP-FLASH. Post-transfección, las células se subcultivaron durante 24 h en presencia de IPTG 1 mM en presencia de concentraciones crecientes de un inhibidor específico de COX-2 (SC-791, 0, 1, 5 y 20 μM) o de un inhibidor específico de COX-1 (FR122047, 10 μM). (A) Posteriormente, los sobrenadantes (medios de cultivo) se rescataron y los niveles de PGE2 se cuantificaron mediante un ensayo comercial. Las barras en blanco muestran los niveles de PGE2 provenientes de células DLD-1 control (-) incubadas con concentraciones crecientes del inhibidor de COX-2, las barras en negro muestran los niveles de PGE2 en células DLD-1 que expresan caveolina-1 (+) tratadas del mismo modo que células control. Las barras en gris corresponden a células incubadas con el inhibidor de COX-1. (B) Análisis de reportero TOP/FOP-FLASH y (C) Análisis de Western blot de lisados obtenidos de células tratadas según descrito anteriormente. El lisado de proteínas (50 μg/ carril) fue separado mediante SDS-PAGE y posteriormente analizado por inmunoblot usando anticuerpos específicos anti-β-catenina, anti-survivina, anti-caveolina-1 y anti-actina. Los datos numéricos fueron normalizados con células control sin tratamiento (100%) y corresponden al promedio de tres experimentos independientes a excepción de C que representa dos experimentos independientes. Las diferencias estadísticamente significativas encontradas en (A) y (B) se indican como *,p<0,01; #, p<0.05.
B
C
β-catenina
survivina
Caveolina-1
actina
-+- - - -
5 20 -+ + +
- 5 20 -- - - 10 - - - 10-
+- - - -5 20 -
+ + +- 5 20 -
Cav-1
(SC-791, μM)
(FR122047, μM)
Cav-1
(SC-791, μM)
(FR122047, μM) - - - - - 10
Inhibidor de COX-2
Inhibidor de COX-1
1.0 0.8 0.6 0.6 0.4 0.3 0.2 0.4
β-catenina
survivina
Caveolina-1
actina
-+- - - -
5 20 -+ + +
- 5 20 -- - - 10 - - - 10-
+- - - -5 20 -
+ + +- 5 20 -
Cav-1
(SC-791, μM)
(FR122047, μM)
Cav-1
(SC-791, μM)
(FR122047, μM) - - - - - 10
Inhibidor de COX-2
Inhibidor de COX-1
1.0 0.8 0.6 0.6 0.4 0.3 0.2 0.4
#
#
0
20
40
60
80
100
120
Cav-1
-+-
1- - - -
5 20 -+ + + +
- 1 5 20 -- - - - 10 - - - - 10
Act
ivid
ad lu
cife
rasa
(%)
Cav-1
-+-
1- - - -
5 20 -+ + + +
- 1 5 20 -- - - - 10 - - - - 10
TOP/FOP
(SC-791, μM)
(FR122047, μM)
Inhibidor de COX-2 (SC-791, μM)
(FR122047, μM)Inhibidor de COX-1
#
#
0
20
40
60
80
100
120
Cav-1
-+-
1- - - -
5 20 -+ + + +
- 1 5 20 -- - - - 10 - - - - 10
Act
ivid
ad lu
cife
rasa
(%)
Cav-1
-+-
1- - - -
5 20 -+ + + +
- 1 5 20 -- - - - 10 - - - - 10
TOP/FOP
(SC-791, μM)
(FR122047, μM)
Inhibidor de COX-2 (SC-791, μM)
(FR122047, μM)Inhibidor de COX-1
A
0
200
400
600
800
1000
1200
1400
Prod
ucci
ón d
e P
GE 2
(ng/
mL)
Cav-1
(SC-791, μM) -+-
1- - - -
5 20 -+ + + +
- 1 5 20 -(FR122047, μM) - - - - 10 - - - - 10
Cav-1
Inhibidor de COX-2 (SC-791, μM) -+-
1- - - -
5 20 -+ + + +
- 1 5 20 -(FR122047, μM) - - - - 10 - - - - 10
#
#
* **
*
Inhibidor de COX-1
0
200
400
600
800
1000
1200
1400
Prod
ucci
ón d
e P
GE 2
(ng/
mL)
Cav-1
(SC-791, μM) -+-
1- - - -
5 20 -+ + + +
- 1 5 20 -(FR122047, μM) - - - - 10 - - - - 10
Cav-1
Inhibidor de COX-2 (SC-791, μM) -+-
1- - - -
5 20 -+ + + +
- 1 5 20 -(FR122047, μM) - - - - 10 - - - - 10
#
#
* **
*
Inhibidor de COX-1

81
Por otra parte, los experimentos de proliferación concuerdan muy bien con lo
observado previamente, es decir, el aumento de la cantidad del inhibidor de COX-2
(SC-791; 0, 1, 5 y 20 μM) disminuyó levemente la proliferación de células control, pero
aún más en aquellas que expresan caveolina-1, siendo efectiva una concentración de
5 μM para reducir significativamente la proliferación (Figura 30). Interesantemente, el
inhibidor específico de la COX-1 también disminuye la proliferación en células que
expresan caveolina-1. Sin embargo, como fue descrito anteriormente para la producción
de PGE2 (Figura 29A), las diferencias no alcanzaron significancia estadística.
En resumen, nuestros hallazgos muestran que la inhibición selectiva de
COX-2 disminuye la producción de PGE2, la actividad transcripcional de
β-catenina-Tcf/Lef y la proliferación celular. Este efecto se ve potenciado en
células que expresan caveolina-1.
Figura 30. La inhibición de la proliferación por la inhibición de COX-2 se potencia en células que expresan caveolina-1. Células DLD-1 transfectadas establemente con pLacIOP (-) o pLacIOP-caveolina-1 (+) se incubaron 24 h en presencia de IPTG 1 mM. Adicionalmente, las células fueron sub-cultivadas en presencia de IPTG 1 mM durante 24 h en placas de 96 pocillos y con concentraciones crecientes de un inhibidor específico de COX-2 (SC-791, 0, 1, 5 y 20 μM) o de un inhibidor específico de COX-1 (FR122047, 10 μM). La proliferación celular fue determinada por el ensayo de MTS®. Los valores indican el porcentaje de actividad residual con respecto a células control (100%), obtenido de un promedio de 3 experimentos independientes (promedio ± SEM). Las diferencias estadísticamente significativas se indican como #, p<0.05.
Cav-1
(SC-791, μM) -+-
1- - - -
5 20 -+ + + +
- 1 5 20 -(FR122047, μM) - - - - 10 - - - - 10
Prol
ifera
ción
(%)
Cav-1
(SC-791, μM) -+-
1- - - -
5 20 -+ + + +
- 1 5 20 -(FR122047, μM) - - - - - - - - 10
##
0
20
40
60
80
100
120
Inhibidor de COX-2
Inhibidor de COX-1
Cav-1
(SC-791, μM) -+-
1- - - -
5 20 -+ + + +
- 1 5 20 -(FR122047, μM) - - - - 10 - - - - 10
Prol
ifera
ción
(%)
Cav-1
(SC-791, μM) -+-
1- - - -
5 20 -+ + + +
- 1 5 20 -(FR122047, μM) - - - - - - - - 10
##
0
20
40
60
80
100
120
Inhibidor de COX-2
Inhibidor de COX-1

82
2.5. Estudiar el efecto de PGE2 en la localización nuclear de β-catenina y sobre el
secuestro de β-catenina a la membrana plasmática por caveolina-1 en células de
cáncer de colon [HT29(ATCC)]
En un estudio reciente (Castellone et al., 2005) se describió que la adición de
PGE2 promueve la translocación y acumulación nuclear de β-catenina en células
DLD-1. Nuestros estudios previos demuestran que la re-expresión de caveolina-1 en
células HT29(ATCC) induce el secuestro de β-catenina a la membrana plasmática
(Torres et al., 2007). Por lo tanto, decidimos investigar si PGE2 podría modular la
distribución de β-catenina en células HT29(ATCC) en presencia y ausencia de
caveolina-1. En primera instancia, los resultados obtenidos en esta tesis confirman que
caveolina-1 co-inmunoprecipita con β-catenina en células HT29(ATCC) (Figura 31A)
como fue previamente demostrado (Torres et al., 2007). Interesantemente, la adición de
PGE2 (20 μM) disminuyó la cantidad de β-catenina que co-inmunoprecipita con
caveolina-1 (Figura 31A) en aproximadamente un 70% (Figura 31C). Adicionalmente, la
figura 31B muestra la cantidad de β-catenina presente en sobrenadante inicial (input) y
la figura 31D muestra la inmunoprecipitación de GAPDH (eliminar la unión no específica
de proteínas) realizada previamente a la inmunoprecipitación de caveolina-1 (ver
metodología).
Para confirmar estos resultados, bajo el mismo diseño experimental se
inmunoprecipitó β-catenina y se cuantificó la cantidad de caveolina-1 que
co-inmunoprecipita. Como se esperaba, se observó el mismo efecto inhibitorio de PGE2,
es decir, también disminuyó la co-inmunoprecipitación de caveolina-1 con β-catenina en
alrededor de 70% consenso

83
Figura 31. PGE2 inhibe la co-inmunoprecipitación de β-catenina con caveolina-1 en células HT29(ATCC). Células HT29(ATCC) establemente transfectadas con los plasmidios pLacIOP (-) o pLacIOP-caveolina-1 (+), fueron cultivadas por 24 h en presencia de IPTG 1 mM. Posteriormente, las células se deprivaron de suero por 4 h e incubaron 3 h con PGE2 (20 μM) en medio sin suero. Luego se preparó extractos proteicos para ensayos de inmunoprecipitación. (A) Inmunoprecipitación de caveolina-1(anticuerpo anti-caveolina-1 policlonal) y Western blot del inmunoprecipitado total con los anticuerpos anti-β-catenina y anti-caveolina-1 (ver metodología). (B) Se muestra un Western blot del control de carga para β-catenina y caveolina-1 (C) La cantidad de β-catenina co-inmunoprecipitada se determinó por densitometría y fue expresada con respecto a su control de carga y a la cantidad de caveolina-1 inmunoprecipitada. Las barras muestran la eficiencia de co-inmunoprecipitación de β-catenina en células que expresan caveolina-1 en ausencia y presencia de PGE2 (20 μM). Los datos representan el promedio de 4 experimentos independientes. Las diferencias estadísticamente significativas se indica (#, p<0.05) (D) Inmunoprecipitación de GAPDH. Se eliminó la unión no específica de proteínas mediante la inmunoprecipitación de GADPH. Los sobrenadantes resultantes fueron usados para inmunoprecipitar caveolina-1 como se muestra en A (ver metodología).
02468
101214161820
PGE2
Cav-1 + ++-
Eficiencia de Inmunoprecipitación
02468
101214161820
PGE2
Cav-1 + ++-
Eficiencia de Inmunoprecipitación
A IPP:Caveolina-1
PGE2
+-Cav-1 +-- - + +
--
IgG
β-catenina
Cav-1
IPP:Caveolina-1
PGE2
+-Cav-1 +-- - + +
--
IgG
β-catenina
Cav-1
B
C D PGE2
+-Cav-1 +-- - + +
IP: GAPDH
INPUT: GAPDH
GAPDH
GAPDH
PGE2
+-Cav-1 +-- - + +
IP: GAPDH
INPUT: GAPDH
GAPDH
GAPDH
PGE2
+-Cav-1 + -- - + +
β-catenina
Cav-1
PGE2
+-Cav-1 + -- - + +
β-catenina
Cav-1
INPUT
# IP
P β-
cat/
inpu
t β-c
at /
IPP
cave
olin
a-1
(%)
Figura 32. PGE2 inhibe la co-inmunoprecipitación de caveolina-1 con β-catenina. Células HT29(ATCC) establemente transfectadas con los plasmidios pLacIOP (-) o pLacIOP-caveolina-1 (+), se cultivaron por 24 h en presencia de IPTG 1 mM. Posteriormente, las células fueron deprivadas de suero por 4 h e incubadas 3 h con PGE2 (20 μM) en medio sin suero. Luego se preparó extractos proteicos para ensayos de inmunoprecipitación. (A) Inmunoprecipitación de β-catenina y Western blot del inmunoprecipitado total con los anticuerpos anti-caveolina-1 y anti-β-catenina. (B) Se muestra un Western blot del control de carga para β-catenina y caveolina-1(input). (C) La cantidad de caveolina-1 se determinó por densitometría y fue expresada con respecto a su control de carga y a la cantidad de β-catenina inmunoprecipitada. Las barras muestran la eficiencia de co-inmunoprecipitación de caveolina-1 con β-catenina en células que expresan caveolina-1 en ausencia y presencia de PGE2 (20 μM). La figura muestra los resultados de un experimento.
A B CIPP:β-catenina
PGE2
+-Cav-1 +-- - + +
--
IgG
β-catenina
Cav-1
IPP:β-catenina
PGE2
+-Cav-1 +-- - + +
--
IgG
β-catenina
Cav-1
PGE2
+-Cav-1 +-- - + +
INPUT (control de carga)
β-catenina
Cav-1
PGE2
+-Cav-1 +-- - + +
INPUT (control de carga)
β-catenina
Cav-1
PGE2
Cav-1 + ++-
IPP
Cav
-1/ i
nput
Cav
-1
/ IP
P β
-cat
enin
a (%
)
0
1
23
4
5
6
PGE2
Cav-1 + ++-
IPP
Cav
-1/ i
nput
Cav
-1
/ IP
P β
-cat
enin
a (%
)
0
1
23
4
5
6

84
Estos hallazgos también fueron corroborados por microscopía confocal en células
HT29(ATCC) que expresan o no caveolina-1. Previamente, mediante el uso de
microscopía confocal, nuestro laboratorio reportó que el secuestro de β-catenina a la
membrana plasmática, debido a la expresión de caveolina-1, se refleja en menores
niveles nucleares de β-catenina (Torres et al., 2007). Como fue previsto, la adición de
PGE2 indujo la acumulación nuclear de β-catenina en células que expresan caveolina-1
(+) y también en aquellas que no la expresan (-) (Figura 33).
Figura 33. PGE2 aumenta la localización nuclear de β-catenina. Células HT29(ATCC) establemente transfectadas con los plasmidios pLacIOP (-) o pLacIOP-caveolina-1 (+), se cultivaron por 24 h en presencia de IPTG 1 mM sobre cubreobjetos. Posteriormente, las células se deprivaron de suero por 4 h e incubaron 3 h con PGE2 (20 μM) en medio sin suero. Luego, fueron procesadas para el análisis de β-catenina (rojo) y de contraste de fase por inmunofluorescencia y microscopía confocal. La barra de magnificación es equivalente a 10 μm. La sobreposición de imágenes se generó usando un software Imaris (Bitplane, Zuerich, Switzerland). Se muestra un resultado representativo de 3 experimentos independientes.
Cav-1 - + - +PGE2
β-ca
teni
naC
ontra
ste
de fa
seS
obre
posi
ción
Cav-1 - + - +PGE2
β-ca
teni
naC
ontra
ste
de fa
seS
obre
posi
ción

85
Figura 34. PGE2 aumenta la localización nuclear de β-catenina e inhibe la capacidad de caveolina-1 para secuestrar β-catenina a la membrana plasmática Células HT29(ATCC) establemente transfectadas con los plasmidios pLacIOP (-) o pLacIOP-caveolina-1 (+), se cultivaron por 24 h en presencia de IPTG 1 mM sobre cubreobjetos. Posteriormente, las células se deprivaron de suero por 4 h e incubaron 3 h con PGE2 (20 μM) en medio sin suero. (A) Luego, fueron procesadas para el análisis de caveolina-1 (verde) y β-catenina (rojo) por inmunofluorescencia indirecta y microscopía confocal. La sobreposición de imágenes y la generación de la imagen 3D (con corte virtual a 5 μm del cubreobjeto para mejor visualización nuclear) de las diferentes secciones ópticas obtenidas por microscopía confocal fue generada usando el software Imaris (Bitplane, Zuerich, Switzerland) (ver mas detalle en metodología). (B) El panel inferior muestra la sobreposición de imágenes generadas con el software Imaris para estudiar co-localización, en donde un tercer pseudo color blanco ilustra la co-localización no azaroza de pixeles rojos y verdes. Resultado representativo de 3 experimentos independientes. Barra = 10μm.
cave
olin
a-1
β-ca
teni
naso
brep
osic
ión
Cav-1 - + - +PGE2
sobr
epos
ició
n 3D
+ co
rte h
oriz
onta
lca
veol
ina-
1β-
cate
nina
sobr
epos
ició
n
Cav-1 - + - +PGE2
Cav-1 - + - +PGE2
sobr
epos
ició
n 3D
+ co
rte h
oriz
onta
l
A
B Cav-1 - + - +
Sobr
epos
ició
n
PGE2Cav-1 - + - +
Sobr
epos
ició
n
PGE2PGE2

86
Además, en células HT29ATCC que expresan caveolina-1, PGE2 redujo la
co-localización de caveolina-1/β-catenina en la membrana plasmática, promoviendo la
acumulación nuclear de β-catenina (Figura 34). Estos resultados sugieren que el
tratamiento con PGE2 evita la eficiente formación del complejo caveolina-1/β-catenina
en la membrana plasmática y por ende permite la translocación nuclear de β-catenina.
En resumen, en este objetivo específico 2 se encontró que la sobreexpresión
de COX-2 o bien la adición de PGE2 aumentaron la actividad transcripcional de
β-catenina-Tcf/Lef promoviendo la expresión de survivina. Además, COX-2
promueve en parte su propia expresión por esta vía.
Interesantemente, la inhibición de la expresión de survivina causada por
caveolina-1 fue prevenida por la adición de PGE2 o bien por la sobreexpresión de
COX-2 a través de un mecanismo que involucra inhibición de la formación de los
complejos multiproteicos entre β-catenina/caveolina-1. Esto disminuye por ende,
el reclutamiento de β-catenina a la membrana plasmática por caveolina-1.

87
Sección II
Objetivo específico 3
3. Analizar el efecto de la expresión ectópica de caveolina-1 sobre la regulación
post-traduccional de survivina.
3.1. Estudio de los complejos multiproteicos que forma caveolina-1 en células de
cáncer de colon [HT29(ATCC) y HT29(US)]
Estudios de nuestro laboratorio han demostrado que la re-expresión de
caveolina-1 en células de cáncer de colon HT29(ATCC) regula negativamente la
expresión de survivina mediante la vía de β-catenina-Tcf/Lef (Torres et al., 2006).
Asimismo, este proceso requiere la presencia de E-cadherina, ya que en células
HT29(US) que no la expresan, no existe regulación transcripcional de survivina (Torres
et al., 2007) ni de COX-2 (Figura 11A y 11C) y sólo al re-expresar E-cadherina
[HT29(US)/E-cad/Cav-1], se regula negativamente la expresión de survivina (Torres et
al., 2007) y de COX-2 (Figura 11A y 11C). Sin embargo, estudios preliminares
sugirieron que en células HT29(US) que expresan caveolina-1 y no E-cadherina se
promueve la degradación proteasomal de survivina, lo cual indicaría la existencia de un
mecanismo de regulación post-traduccional.
Para identificar qué proteínas forman los complejos moleculares que dan cuenta
de dicha regulación post-traduccional de survivina, se utilizó como modelo, células
HT29(ATCC) que expresan E-cadherina y células HT29(US), que no la expresan.
Establecimos una colaboración con el Dr. Jay W. Heinecke de la Escuela de Medicina
de la Universidad de Washington (Seattle, WA), cuyo laboratorio estudia la composición
de complejos macromoleculares mediante espectrometría de masas
(MALDI-TOF/TOF).

88
De esta manera, se cultivó células de cáncer de colon HT29(ATCC) y HT29(US)
transfectadas establemente con pLacIOP o pLacIOP-caveolina-1. Luego de cultivarlas
24 h en presencia de IPTG 1 mM, caveolina-1 se inmunoprecipitó y se chequeó su
presencia por SDS-PAGE e inmunoblot. De los inmunoprecipitados se preparó
extractos digeridos por tripsina para después realizar el análisis de las muestras por
espectrometría de masas MALDI-TOF/TOF y por cromatografía capilar de fase reversa
con ionizador de electrospray acoplado a un espectrómetro de masas.
3.1.1. Preparación de células y control de la expresión de caveolina-1
Células HT29(US) y HT29 (ATCC) que expresan caveolina-1 se seleccionaron dos
semanas con higromicina 500 μg/ml y la expresión de caveolina-1 se indujo por
incubación con IPTG 1 mM. La presencia de caveolina-1 en los extractos celulares fue
confirmada por Western blot utilizando GAPDH como control de carga. En ambos casos
se expresa caveolina-1 (Figura 35), con la salvedad que el nivel basal de expresión de
caveolina-1 en células HT29(US) transfectadas con pLacIOP (Cav-1, -) es mayor que lo
esperado según resultados previos del laboratorio.
Figura 35. Expresión de caveolina-1 en células de cáncer de colon [HT29(US) y HT29(ATCC)]. Células HT29 (US) y HT29 (ATCC) transfectadas establemente con pLacIOP (-) o pLacIOP-caveolina-1 (+), fueron cultivadas en placas de 100 mm durante 24 h en presencia de IPTG 1 mM (inducción expresión de caveolina-1). Los extractos de proteínas totales fueron separados mediante SDS-PAGE (50 μg proteínas por carril) y analizados por Western blot usando anticuerpos específicos anti-caveolina-1 y anti-GAPDH, este último como control de carga. Se muestra un resultado representativo de tres experimentos independientes.
HT29(US)
Cav-1 - + - +
caveolina-1
GAPDH
HT29(ATCC)HT29(US)
Cav-1 - + - +
GAPDH
HT29(US)
Cav-1 - + - +
caveolina-1
GAPDH
HT29(ATCC)HT29(US)
Cav-1 - + - +
GAPDHGAPDH
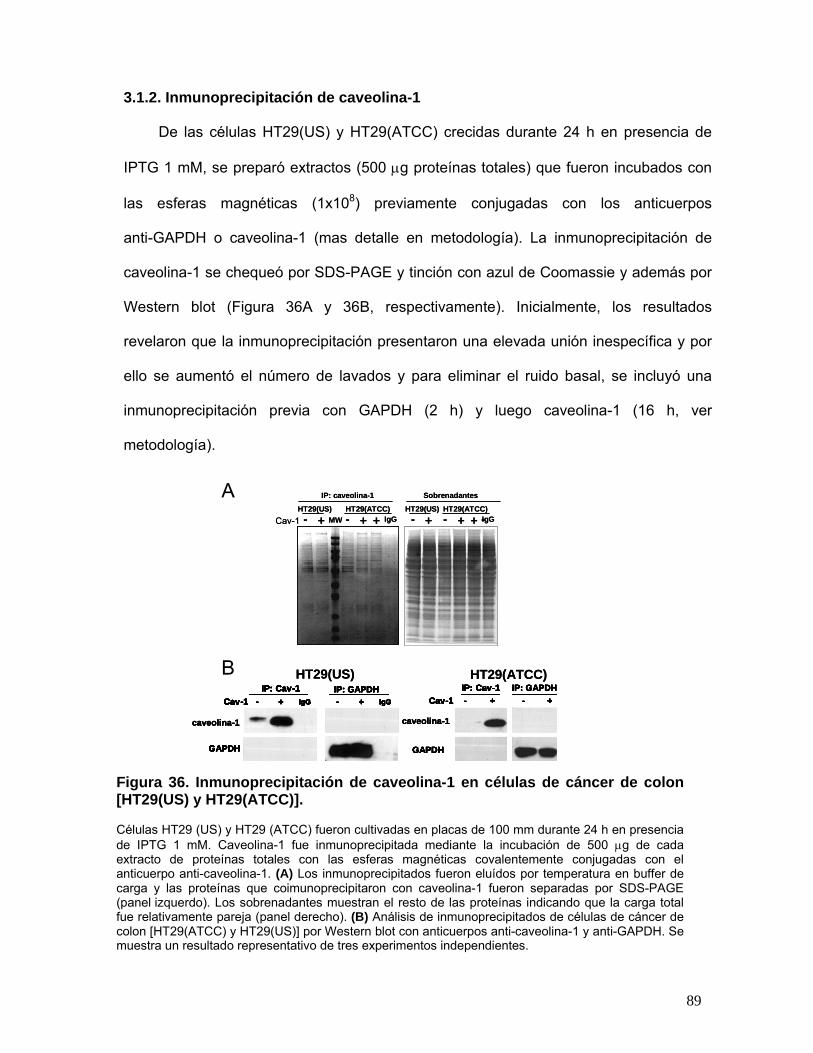
89
3.1.2. Inmunoprecipitación de caveolina-1
De las células HT29(US) y HT29(ATCC) crecidas durante 24 h en presencia de
IPTG 1 mM, se preparó extractos (500 μg proteínas totales) que fueron incubados con
las esferas magnéticas (1x108) previamente conjugadas con los anticuerpos
anti-GAPDH o caveolina-1 (mas detalle en metodología). La inmunoprecipitación de
caveolina-1 se chequeó por SDS-PAGE y tinción con azul de Coomassie y además por
Western blot (Figura 36A y 36B, respectivamente). Inicialmente, los resultados
revelaron que la inmunoprecipitación presentaron una elevada unión inespecífica y por
ello se aumentó el número de lavados y para eliminar el ruido basal, se incluyó una
inmunoprecipitación previa con GAPDH (2 h) y luego caveolina-1 (16 h, ver
metodología).
IP: caveolina-1
Cav-1 - + - + + -MWHT29(US) HT29(ATCC)
IgG
Sobrenadantes
- + - + +HT29(US) HT29(ATCC)
IgG
IP: caveolina-1
Cav-1 - + - + + -MWHT29(US) HT29(ATCC)
IgG
Sobrenadantes
- + - + +HT29(US) HT29(ATCC)
IgG
A
B
Figura 36. Inmunoprecipitación de caveolina-1 en células de cáncer de colon [HT29(US) y HT29(ATCC)]. Células HT29 (US) y HT29 (ATCC) fueron cultivadas en placas de 100 mm durante 24 h en presencia de IPTG 1 mM. Caveolina-1 fue inmunoprecipitada mediante la incubación de 500 μg de cada extracto de proteínas totales con las esferas magnéticas covalentemente conjugadas con el anticuerpo anti-caveolina-1. (A) Los inmunoprecipitados fueron eluídos por temperatura en buffer de carga y las proteínas que coimunoprecipitaron con caveolina-1 fueron separadas por SDS-PAGE (panel izquerdo). Los sobrenadantes muestran el resto de las proteínas indicando que la carga total fue relativamente pareja (panel derecho). (B) Análisis de inmunoprecipitados de células de cáncer de colon [HT29(ATCC) y HT29(US)] por Western blot con anticuerpos anti-caveolina-1 y anti-GAPDH. Se muestra un resultado representativo de tres experimentos independientes.
IP: Cav-1 IP: GAPDH
GAPDH
-Cav-1 + - +
HT29(ATCC)HT29(US)
+ IgG
IP: Cav-1 IP: GAPDH
GAPDH
-Cav-1 + IgG-+ IgG
IP: Cav-1 IP: GAPDH
GAPDH
-Cav-1 + IgG-IP: Cav-1 IP: GAPDH
GAPDH
-Cav-1 + - +
caveolina-1 caveolina-1
IP: Cav-1 IP: GAPDH
GAPDH
-Cav-1 + - ++ IgG
IP: Cav-1 IP: GAPDH
GAPDH
-Cav-1 + IgG-+ IgG
IP: Cav-1 IP: GAPDH
GAPDH
-Cav-1 + IgG-IP: Cav-1
GAPDH
-Cav-1 + - +
caveolina-1 caveolina-1
IP: Cav-1 IP: GAPDH
GAPDH
-Cav-1 + - +
HT29(ATCC)HT29(US)
+ IgG
IP: Cav-1 IP: GAPDH
GAPDH
-Cav-1 + IgG-+ IgG
IP: Cav-1 IP: GAPDH
GAPDH
-Cav-1 + IgG-IP: Cav-1 IP: GAPDH
GAPDH
-Cav-1 + - +
caveolina-1 caveolina-1
IP: Cav-1 IP: GAPDH
GAPDH
-Cav-1 + - ++ IgG
IP: Cav-1 IP: GAPDH
GAPDH
-Cav-1 + IgG-+ IgG
IP: Cav-1 IP: GAPDH
GAPDH
-Cav-1 + IgG-IP: Cav-1
GAPDH
-Cav-1 + - +
caveolina-1 caveolina-1

90
3.1.3. Análisis de muestras por MALDI-TOF
Una vez confirmada la inmunoprecipitación de caveolina-1, las proteínas fueron
digeridas con tripsina y concentradas por mini-columnas C18ZipTip. Los eluidos
correspondientes para cada línea celular fueron analizados en un espectrómetro de
masas MALDI-TOF/TOF. Según se observa en la figura 37, sólo fue posible detectar la
presencia de GAPDH y no caveolina-1. El mismo ensayo se realizó dos veces con un
control positivo como BSA. Sin embargo, la abundancia de caveolina-1 en los
inmunoprecipitados no fue suficiente para ser detectada mediante este equipo. Para
ello se utilizó otro espectrómetro de masas acoplado a cromatografía capilar de fase
reversa.
3.1.4. Análisis de muestras por LTQ (Cromatografía capilar de fase reversa
acoplado a masas)
Células HT29 (US) y HT29 (ATCC) que expresan o no caveolina-1 se trataron del
mismo modo descrito anteriormente. Una vez realizada la inmunoprecipitación de
caveolina-1 las muestras se eluyeron por temperatura (90ºC, 15 min), digeridas con
tripsina y concentradas por centrifugación al vacío (speedvac). Posteriormente, las
muestras se resuspendieron en 20 μL con una solución de formaldehído 0,1% y
acetonitrilo 5%. El equipo fue programado para una gradiente de una hora, es decir de
menor a mayor polaridad (solución A; 5 % acetonitrilo / ácido trifluoroacético 0,1% y
solución B; 50 % acetonitrilo / ácido trifluoroacético 0,1%). Finalmente, los péptidos
separados se detectaron con el espectrómetro de masas (MS/MS).
El análisis de datos y búsqueda del espectro MS/MS se llevó a cabo utilizando la
base de datos humana “International Protein Index (IPI)” v3.01
(http://www.ebi.ac.uk/IPI/IPIhelp.html) mediante el programa SEQUEST (Thermo

91
Electron). Se consideró posibles modificaciones como oxidación de péptidos en
metionina y un corte incompleto por tripsina. Los resultados de la búsqueda fueron
validados mediante el uso de otro motor de búsqueda “PeptideProphet y ProteinProphet
(http://tools.proteomecenter.org/TPP.php). La selección de datos fue realizada usando
una probabilidad mayor que 0,9 para péptidos y valores mayores que 0,95 para
proteínas como fue previamente descrito (Vaisar et al., 2007).
Los resultados que se presentan en las tablas 3 y 4 confirman la presencia de
caveolina-1 en ambas líneas celulares HT29(US) y HT29(ATCC) que expresan
caveolina-1 de manera inducible por IPTG. El análisis de las proteínas que co-
inmunoprecipitaron con caveolina-1 (Tabla 3 y 4) reveló la presencia de proteínas
involucradas en la degradación proteasomal (Ligasas de ubiquitina), lo cual nos llevó a
proponer que caveolina-1 prodría estar promoviendo la degradación de survivina (Ver
esquema en figura 44). Además, en células HT29(ATCC) se encontró proteínas
activadoras de fosfatasas de tirosina (Liprin alpha-2/4) (Tabla 3) (Serra-Pages et al.,
1998). En cambio, en inmunoprecipitados de células HT29(US), se observó un patrón
muy distinto de las proteínas que co-inmunoprecipitaron con caveolina-1 (Tabla 4) y no
fue detectada la presencia de estas proteínas reguladoras de fosfatasas de tirosina
(Liprin alpha 4) como fue observado en células HT29(ATCC), sugiriendo que su
presencia podría depender de la expresión de E-cadherina.
3.2. Analizar el efecto de la expresión ectópica de caveolina-1 sobre la regulación
post-traduccional de survivina
En esta tesis se propuso investigar si caveolina-1 podría regular negativamente
la expresión de survivina a través de un mecanismo de regulación post-traduccional, el
cual involucra la ubiquitinación y degradación vía el proteasoma, ya que se ha descrito

92
que survivina es degradada por esta vía (Zhao et al., 2000) y el tratamiento con NSAIDs
potencia su degradación (Chiou y Mandayam, 2007).
Tabla 3. Análisis de muestras por LTQ (inmunoprecipitados HT29(ATCC))
HT29(ATCC) pLacIOP
HT29(ATCC) pLacIOP-caveolina-1
similar to GOLGA8A protein Glycerophosphodiester phosphodiesterase domain-containing protein
MAD1 mitotic arrest deficient-like 1 Protein kinase C epsilon type Isoform 5 of Hypoxia-inducible factor 3 alpha Regulator of G-protein signaling 12 Isoform 3 of EF-hand calcium-binding domain-containing protein 5
Heat shock protein beta-1
Putative p150 Non-specific lipid-transfer protein similar to RE32881p Transcriptional repressor NF-X1 Isoform 1 of Ankyrin repeat domain-containing protein 41 CTA-126B4.3 Gastric cancer ant... Isoform 1 of Transmembrane protease, serine 13 Ral guanine nucleotide exchange factor RalGPS1A Isoform 2 of Integrator complex subunit 7 MAX interactor 1 similar to LOC642852 protein Alanine aminotransferase 2 Dishevelled 1 isoform a variant (Fragment) Nuclear transcription factor NFX2 Small subunit processome component 20 homolog Nuclear transcription factor, X-box binding 1 Isoform 1 of Sialoadhesin precursor WW domain-binding protein 11 Probable ribosome biogenesis protein RLP24 Junctophilin-2 Coagulation factor X precursor Cytospin-A Isoform 1 of GRAM domain-containing protein 4 BAI1-associated protein 3 Isoform 2 of Serine/threonine-protein kinase TAO2 C9orf86 Isoform 3 of Putative GTP-binding protein Parf Isoform 2 of Supervillin Genetic suppressor element 1 Isoform 1 of Latent-transforming growth factor beta-binding protein 3 precursor
Ankyrin-1
t-SNARE domain containing 1 ALB protein Ras-related protein Rab-40C Sterol carrier protein 2 Msx2-interacting protein Schlafen family member 12 Isoform 1 of MARVEL domain-containing protein 3 Regulator of G-protein signalling 12 Isoform 3 of LisH domain-containing protein ARMC9 Peroxisomal proliferator-activated receptor A-interacting
complex 285 kDa protein ETS domain-containing transcription factor ERF Ral GEF with PH domain and SH3 binding motif 1 Isoform 1 of Protein kinase C theta type USP5 Isoform Short of Ubiquitin carboxyl-terminal hy... Gamma-tubulin complex component 2 FH1/FH2 domain-containing protein 3 Synaptotagmin-4 IGHG1 protein Isoform 2 of Peptidyl-prolyl cis-trans isomerase-like 2 ATP7B Isoform 3 of Copper-transporting ATPase 2 Isoform 1 of Fragile X mental retardation 1 protein ZNF56 32 kDa protein DNA-repair protein complementing XP-C cells Adenylate cyclase type 10 Isoform 1 of Deoxyribonuclease-2-beta precursor Serine palmitoyltransferase 3 Epoxide hydrolase 2 Nuclear transcription factor, X-box binding 1 Repetin Uncharacterized protein DKC1 Cytoplasmic dynein 1 heavy chain 1 E3 ubiquitin-protein ligase UBR2 Isoform 2 of Transmembrane channel-like protein 5 GW112 protein echinoderm microtubule associated protein like 5 HNRPUL1 Isoform 2 of Heterogeneous nuclear
ribonucleoprotein U-like protein 1 similar to proline-rich proteoglycan 2 UBAP2L Isoform 2 of Ubiquitin-associated protein 2... CXXC-type zinc finger protein 6 Ubiquitin carboxyl-terminal hydrolase 5 Isoform 1 of Protein phosphatase Slingshot homolog 2 UBC;UBB;RPS27A ubiquitin and ribosomal protein S27a
pr... PR domain containing 1, with ZNF domain isoform 2 UBAP2L Isoform 3 of Ubiquitin-associated protein 2-like Serine/threonine-protein kinase RIO2 UBR2 Isoform 4 of E3 ubiquitin-protein ligase UBR2 Isoform 5 of Vasohibin-2 HNRPUL1 Isoform 3 of Heterogeneous nuclear
ribonucleoprotein U-like protein 1 EOSINOPHIL LYSOPHOSPHOLIPASE UBR2 Isoform 3 of E3 ubiquitin-protein ligase UBR2

93
Putative collagen homolog protein-a Ubiquitin A-52 residue ribosomal protein fusion product 1 Ubiquitin-associated protein 2-like Ubiquitin associated protein 2-like
Ubiquitin B Ubiquitin C splice variant UBC;UBB;RPS27A 30 kDa protein UBC;UBB;RPS27A 16 kDa protein UBC;UBB;RPS27A 25 kDa protein UBC;UBB;RPS27A Ribosomal protein S27a UBC;UBB;RPS27A 39 kDa protein UBC;UBB;RPS27A 12 kDa protein UBC protein UBC;UBB;RPS27A 19 kDa protein UBC;UBB;RPS27A 18 kDa protein Ubiquitin C splice variant UBC;UBB;RPS27A 79 kDa protein Protein tyrosine phosphatase, receptor type, f polypeptide (PTPRF), interacting protein (Liprin), alpha 4 PPFIA2 protein Liprin alpha4 Liprin-alpha-3 PPFIA3 Isoform 2 of Liprin-alpha-3 Protein tyrosine phosphatase, receptor type, f polypeptide (PTPRF), interacting protein (Liprin), alpha 4 PPFIA4 Isoform 2 of Liprin-alpha-4 PPFIA2 Liprin-alpha-2
Q03135|CAV1_HUMAN
Tabla 3. Análisis de muestras por LTQ (inmunoprecipitados HT29(ATCC)) Células HT29(ATCC) establemente transfectadas con los plasmidios pLacIOP (-) o pLacIOP-caveolina-1 (+), se cultivaron por 24 h en presencia de IPTG 1 mM. Luego se preparó extractos proteicos para ensayos de inmunoprecipitación de caveolina-1. La inmunoprecipitación de caveolina-1 se llevó a cabo en el laboratorio del Dr. Jay W. Heinecke de la Escuela de Medicina de la Universidad de Washington (Seattle, WA). Para ello se utilizó anticuerpos policlonales anti-caveolina-1 inmovilizados de manera covalente con esferas magnéticas. Posteriormente, se eluyó las proteínas que co-inmunoprecipitan con caveolina-1 y se digirieron con tripsina. Los péptidos resultantes se analizaron en un equipo de cromatografía capilar de fase reversa con ionizador de electrospray acoplado a un espectrómetro de masas (MS/MS) con trampa de iones (LTQ, Thermo Electron, Waltham, MA, USA) (ver metodología y métodos descritos en (Vaisar et al., 2007)). El análisis de datos y búsqueda del espectro MS/MS se llevó a cabo utilizando la base de datos humana “International Protein Index (IPI)” v3.01 (http://www.ebi.ac.uk/IPI/IPIhelp.html) mediante el programa SEQUEST (Thermo Electron). Se consideró posibles modificaciones como oxidación de péptidos en metionina y un corte incompleto por tripsina. Los resultados de la búsqueda fueron validados mediante el uso de otro motor de búsqueda “PeptideProphet y ProteinProphet (http://tools.proteomecenter.org/TPP.php). La selección de datos fue realizada usando una probabilidad mayor que 0,9 para péptidos y valores mayores que 0,95 para proteínas. La tabla muestra los resultados obtenidos de un experimento.
94
Tabla 4. Análisis de muestras por LTQ (inmunoprecipitados HT29(US))
HT29(US) pLacIOP
HT29(US) pLacIOP-caveolina-1 Desmocollin 1b Protein beta-actin-like Caspase-14 precursor Small subunit processome component 20 homolog Calmodulin-like protein 5 Gene_Symbol=UBAP2L NICE-4 protein ALB Uncharacterized protein ALB Fatty acid-binding protein, epidermal Filaggrin Frizzled-8 precursor Dermcidin precursor Desmoplakin Cystatin-A Lysozyme C precursor Uncharacterized protein ENSP00000382160 Dynein heavy chain 12, axonemal Desmocollin-1 precursor Heat shock protein beta-1 ALB protein Desmoglein-1 precursor Uncharacterized protein ENSP00000300992 Gene_Symbol=UBAP2L Isoform 2 of Ubiquitin-associated
protein 2... Uncharacterized protein ENSP00000368744 Polycystic kidney disease 2-like 1 protein Profilaggrin Alpha-2-glycoprotein 1, zinc-binding Desmocollin 1a SYNPO2 synaptopodin 2 Hornerin UBAP2L Isoform 3 of Ubiquitin-associated protein 2-like Hemoglobin subunit alpha Thioredoxin ANXA2 annexin A2 isoform 1 DSP Isoform DPII of Desmoplakin - CDNA FLJ46113 fis, clone TESTI20... Tau-tubulin kinase CDNA FLJ45296 fis, clone BRHIP3003340, moderately similar to Actin, alpha skeletal muscle 2
Hemoglobin subunit epsilon
Annexin A2 Glyceraldehyde-3-phosphate dehydrogenase ANKRD26-like family C member 1A PKD2L1 Isoform 2 of Polycystic kidney disease 2-like 1
protein ACTA1 Uncharacterized protein ACTA1 PKD2L1 Isoform 3 of Polycystic kidney disease 2-like 1
protein FLG Profilaggrin Splice Isoform 4 of Polycystic kidney disease 2-like 1
protein PRB3 protein Arginase-1 RPS27A protein PRO0899 Hemoglobin subunit beta LOC729440 hypothetical protein LOC729440 Profilaggrin Polycystic kidney disease 2 related protein LOC653781 similar to Prostate, ovary, testis expressed protein on chromosome 2
ARG1 Isoform 3 of Arginase-1
ANKRD26-like family C member 1B Corneodesmosin precursor LOC653269 similar to Prostate, ovary, testis expressed protein on chromosome 2 isoform 2
Ifapsoriasin
FLG 115 kDa protein ARG1 Isoform 2 of Arginase-1 FLG Profilaggrin LOC387934 similar to Fatty acid-binding protein,
epidermal PRB3 protein Q03135|CAV1_HUMAN LOC730839 similar to Hornerin Fructose-bisphosphate aldolase A Pseudogene candidate Pyruvate kinase isozymes M1/M2 PSPHL;ACTG1 22 kDa protein Ubiquitin-associated protein 2-like PSPHL;ACTG1 ACTG1 protein Thioredoxin PSPHL;ACTG1 16 kDa protein Junction plakoglobin ANXA2 24 kDa protein Kappa-actin ANXA2 8 kDa protein Pyruvate kinase ACTA2 protein PKM2 27 kDa protein Dermcidin isoform 2 Tax_Id=9606 52 kDa protein Alpha 2 globin variant CDSN Corneodesmosin precursor FLG PREDICTED: filaggrin variant HBD;HBB 15 kDa protein UBA52 11 kDa protein HBE1 15 kDa protein ALB 56 kDa protein Synaptopodin-2 LOC730839 similar to hornerin LOC642956 similar to Fatty acid-binding protein,
epidermal Lipocalin-1 precursor SYNPO2 similar to synaptopodin 2 isoform 3 Putative lipocalin 1-like protein 1 precursor SYNPO2 similar to synaptopodin 2 isoform 2 Probable cation-transporting ATPase 13A5 HBE1 17 kDa protein CDNA FLJ36874 fis, clone ASTRO2018169, weakly PKD2L1 88 kDa protein

95
similar to Oocyte-specific protein P100 CSTA 7 kDa protein HBE1 17 kDa protein Protein PAT1 homolog 1 LOC648187 similar to filaggrin 2 OK/KNS-CL.5 PKM2 26 kDa protein Q03135|CAV1_HUMAN JUP 30 kDa protein
GAPDH Glyceraldehyde 3-phosphate dehydrogenase Putative uncharacterized protein DKFZp686P24273 JUP 32 kDa protein HBD 11 kDa protein Hemoglobin Lepore-Baltimore HBB 19 kDa protein Pyruvate kinase ALDOAP2 Similar to Fructose-bisphosphate aldolase (EC 4.1.2.13) A Uncharacterized protein HBE1 Uncharacterized protein AZGP1 Uncharacterized protein ENSP00000381161 ALB 23 kDa protein IPI:IPI00884436.1|H-INV:HIT000393097 Tax_Id=9606 Gene_Symbol=- Similar to Beta-hemoglobin LOC100129958 similar to hCG1643231 LOC100129958 similar to hCG1643231 LOC100129958 similar to hCG1643231 Heat shock 27kDa protein 1 Protein S100-A7 UBC;UBB;RPS27A ubiquitin and ribosomal protein S27a pr... Ubiquitin A-52 residue ribosomal protein fusion product 1 Ubiquitin C splice variant UBC;UBB;RPS27A Ribosomal protein S27a UBC;UBB;RPS27A 39 kDa protein Ubiquitin B UBC;UBB;RPS27A 44 kDa protein UBC;UBB;RPS27A 30 kDa protein UBC;UBB;RPS27A 16 kDa protein UBC;UBB;RPS27A 25 kDa protein UBC;UBB;RPS27A 18 kDa protein UBC;UBB;RPS27A 21 kDa protein UBC protein UBC;UBB;RPS27A 17 kDa protein UBC;UBB;RPS27A 18 kDa protein UBC;UBB;RPS27A 19 kDa protein UBC;UBB;RPS27A CDNA FLJ32377 fis, clone SKMUS100... UBC;UBB;RPS27A ubiquitin C Ubiquitin C splice variant UBC;UBB;RPS27A 79 kDa protein UBC;UBB;RPS27A 12 kDa protein
Tabla 4. Análisis de muestras por LTQ (inmunoprecipitados HT29(US)) Células HT29(US) establemente transfectadas con los plasmidios pLacIOP (-) o pLacIOP-caveolina-1 (+), se cultivaron por 24 h en presencia de IPTG 1 mM. Luego se preparó extractos proteicos para ensayos de inmunoprecipitación de caveolina-1 y se procedió a la identificación de proteínas y posterior análisis del mismo modo descrito en la leyenda de la tabla 3. El listado de proteínas encontradas corresponde a un experimento.

96
HT29(ATCC) pLacIOP-Cav-1
HT29(ATCC) pLacIOPIP: Cav-1 GAPDH
- + - +IP: Cav-1 GAPDH
- + - +
HT29(ATCC) pLacIOP-Cav-1
HT29(ATCC) pLacIOPIP: Cav-1 GAPDH
- + - +IP: Cav-1 GAPDH
- + - +
A
B
C
Figura 37. Análisis de muestras por MALDI-TOF. Extractos proteicos de células HT29(ATCC) y HT29(US) preparados bajo las mismas condiciones experimentales descritas en métodos. Los inmunoprecipitados eluídos por temperatura y las proteínas que co-imunoprecipitaron con caveolina-1 se concentraron por mini columnas C18ZipTip. Posteriormente, las muestras se colocaron en una placa especial para el equipo de MALDI-TOF/TOF. Análisis de inmunoprecipitados con esferas magnéticas conjugadas con anticuerpos anti-GAPDH (A) o anti-caveolina-1 (B) en células HT29(ATCC) transfectadas establemente con el vector vacío (pLacIOP) o caveolina-1 (pLacIOP-caveolina-1). (C) Análisis de inmunoprecipitados con esferas magnéticas conjugadas con anticuerpos anti-caveolina-1 de células HT29(US) transfectadas establemente con el vector vacío (pLacIOP) o caveolina-1 (pLacIOP-caveolina-1). Las tablas indican el nombre de las proteínas encontradas y el “score” o “índice de confianza” expresado como porcentajes (%). Los valores de scores mayor al 65 % fueron considerados como valores confiables para la proteína encontrada (resultados corresponden a tres experimentos independientes)
HT29(US) pLacIOp
HT29(US) pLacIOP-caveolina-1
HT29(US) pLacIOp
HT29(US) pLacIOP-caveolina-1
+ IgGIP: Cav-1
Cav-1
-Cav-1 + IgGIP: Cav-1
Cav-1
-Cav-1
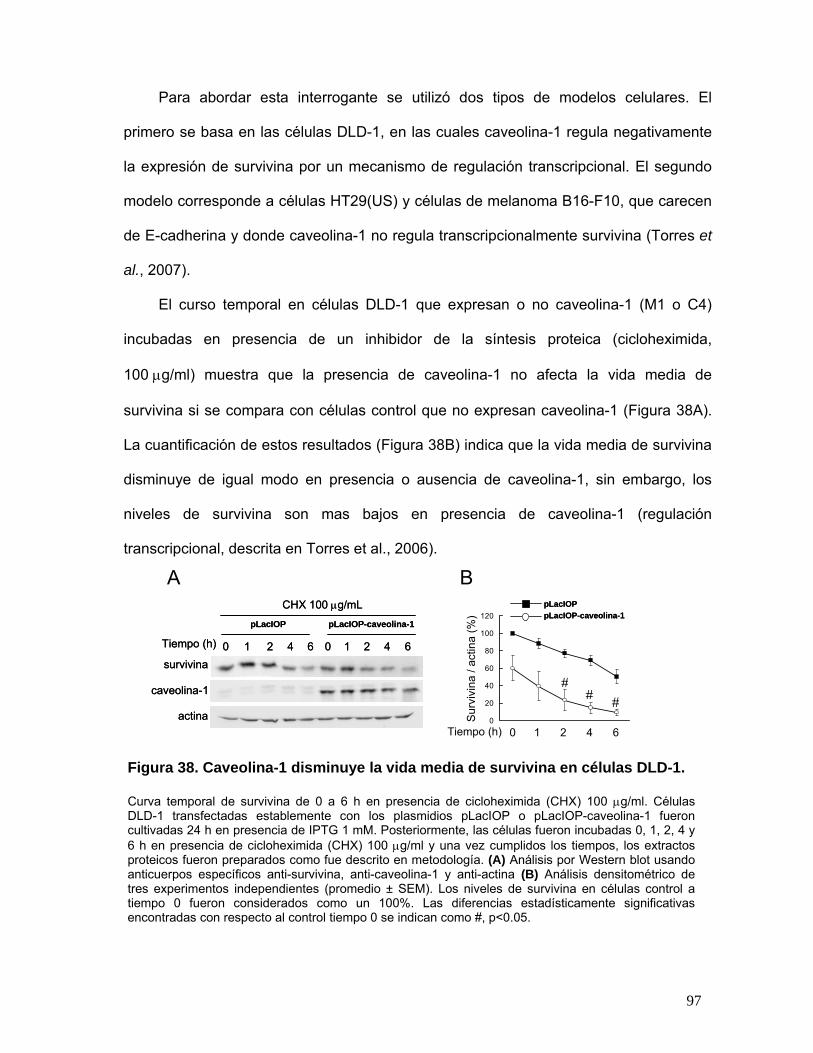
97
0
20
40
60
80
100
120
Para abordar esta interrogante se utilizó dos tipos de modelos celulares. El
primero se basa en las células DLD-1, en las cuales caveolina-1 regula negativamente
la expresión de survivina por un mecanismo de regulación transcripcional. El segundo
modelo corresponde a células HT29(US) y células de melanoma B16-F10, que carecen
de E-cadherina y donde caveolina-1 no regula transcripcionalmente survivina (Torres et
al., 2007).
El curso temporal en células DLD-1 que expresan o no caveolina-1 (M1 o C4)
incubadas en presencia de un inhibidor de la síntesis proteica (cicloheximida,
100 μg/ml) muestra que la presencia de caveolina-1 no afecta la vida media de
survivina si se compara con células control que no expresan caveolina-1 (Figura 38A).
La cuantificación de estos resultados (Figura 38B) indica que la vida media de survivina
disminuye de igual modo en presencia o ausencia de caveolina-1, sin embargo, los
niveles de survivina son mas bajos en presencia de caveolina-1 (regulación
transcripcional, descrita en Torres et al., 2006).
Figura 38. Caveolina-1 disminuye la vida media de survivina en células DLD-1. Curva temporal de survivina de 0 a 6 h en presencia de cicloheximida (CHX) 100 μg/ml. Células DLD-1 transfectadas establemente con los plasmidios pLacIOP o pLacIOP-caveolina-1 fueron cultivadas 24 h en presencia de IPTG 1 mM. Posteriormente, las células fueron incubadas 0, 1, 2, 4 y 6 h en presencia de cicloheximida (CHX) 100 μg/ml y una vez cumplidos los tiempos, los extractos proteicos fueron preparados como fue descrito en metodología. (A) Análisis por Western blot usando anticuerpos específicos anti-survivina, anti-caveolina-1 y anti-actina (B) Análisis densitométrico de tres experimentos independientes (promedio ± SEM). Los niveles de survivina en células control a tiempo 0 fueron considerados como un 100%. Las diferencias estadísticamente significativas encontradas con respecto al control tiempo 0 se indican como #, p<0.05.
CHX 100 μg/mL
0 1 2 4 6 0 1 2 4 6Tiempo (h)
survivina
caveolina-1
actina
pLacIOP pLacIOP-caveolina-1
CHX 100 μg/mL
0 1 2 4 6 0 1 2 4 6Tiempo (h)
survivina
caveolina-1
actina
pLacIOP pLacIOP-caveolina-1
A
Tiempo (h)
Sur
vivi
na /
actin
a (%
)
B
0 1 2 4 6
pLacIOP-caveolina-1pLacIOPpLacIOP-caveolina-1pLacIOP
## #

98
Las diferencias encontradas en células HT29(US), fueron diferentes a lo
observado en células DLD-1. A tiempo cero no se observó diferencias en los niveles
proteicos de survivina en ausencia o presencia de caveolina-1. En cambio, el curso
temporal con CHX reveló que células que expresan caveolina-1 poseen menores
niveles proteicos de survivina comparado con células control, sugiriendo que la vida
media de survivina disminuye desde 4h a 1h (Figura 39A y B).
Figura 39. Caveolina-1 disminuye la vida media de survivina en células HT29(US). Curva temporal de survivina de 0 a 6 h en presencia de cicloheximida (CHX) 100 μg/ml. Células HT29(US) transfectadas establemente con los plasmidios pLacIOP o pLacIOP-caveolina-1 fueron cultivadas 24 h en presencia de IPTG 1 mM. Posteriormente, incubadas 0, 1, 2, 4 y 6 h en presencia de cicloheximida (CHX) 100 μg/ml y una vez cumplidos los tiempos, los extractos proteicos fueron preparados como fue descrito en metodología. (A) Análisis por Western blot usando anticuerpos específicos anti-survivina, anti-caveolina-1 y anti-actina (B) Análisis densitométrico de tres experimentos independientes (promedio ± SEM). Los niveles de survivina en células control sin caveolina-1 a tiempo 0 fueron considerados como un 100%.
CHX 100 μg/mL
0 1 2 4 6 0 1 2 4 6Tiempo (h)survivina
actina
pLacIOP pLacIOP-caveolina-1
caveolina-1
CHX 100 μg/mL
0 1 2 4 6 0 1 2 4 6Tiempo (h)survivina
actina
pLacIOP pLacIOP-caveolina-1
caveolina-1
A B
Tiempo (h) S
urvi
vina
/ ac
tina
(%)
0 1 2 4 6
pLacIOP-caveolina-1pLacIOPpLacIOP-caveolina-1pLacIOP
Tiempo (h)
Sur
vivi
na /
actin
a (%
)
0 1 2 4
pLacIOP-caveolina-1pLacIOPpLacIOP-caveolina-1pLacIOP
Figura 40. Caveolina-1 no disminuye la vida media de survivina en células B16F10. Curva temporal de survivina de 0 a 4 h en presencia de cicloheximida (CHX) 100 μg/ml. Células B16F10 transfectadas establemente con los plasmidios pLacIOP o pLacIOP-caveolina-1 fueron cultivadas 24 h en presencia de IPTG 1 mM. Posteriormente, incubadas 0, 1, 2 y 4 h en presencia de cicloheximida (CHX) 100 μg/ml y una vez cumplidos los tiempos, los extractos proteicos fueron preparados como fue descrito en metodología. (A) Análisis por Western blot usando anticuerpos específicos anti-survivina, anti-caveolina-1 y anti-actina (B) Análisis densitométrico de tres experimentos independientes (promedio ± SEM). Los niveles de survivina en células control sin caveolina-1 a tiempo 0 fueron considerados como un 100%.
A B
0
20
40
60
80
100
120
0 20 40 60 80
100 120
#
#
CHX 100 μg/mL
0 1 2 4Tiempo (h)
survivina
caveolina-1
actina
pLacIOP
0 1 2 4
pLacIOP-caveolina-1
CHX 100 μg/mL
0 1 2 4Tiempo (h)
survivina
caveolina-1
actina
pLacIOP
0 1 2 4
pLacIOP-caveolina-1

99
Sin embargo, nuestros experimentos en células B16F10 (carecen de una
regulación transcripcional de survivina por caveolina-1) muestran que la presencia de
caveolina-1 no disminuye la vida media de survivina en estas células (figura 40A y B).
Estos resultados sugieren que posiblemente células B16F10 que expresan caveolina-
1 requieren la presencia de otra(s) proteínas(s) para promover la degradación de
survivina por un mecanismo de regulación post-transcripcional.
Resumen de resultados más relevantes
I.- Caveolina-1 inhibe la expresión de COX-2 por un mecanismo transcripcional
dependiente de la vía de β-catenina-Tcf/Lef.
II.- La sobreexpresión de COX-2 o la adición de PGE2 por un mecanismo de
retroalimentación positivo aumenta la expresión de survivina y así mismo de COX-2.
III.- PGE2 disminuye la presencia de β-catenina en la membrana plasmática e
inhibe la formación del complejo multiproteico entre caveolina-1 y β-catenina.
IV.-La incubación de células con PGE2 o la sobreexpresión de COX-2 previenen la
disminución de la proliferación provocada por la presencia de caveolina-1.
V.- Los datos sugieren que caveolina-1 promueve la degradación de survivina por
un mecanismo de regulación post-traduccional en celulas HT29(US).

100
VII. DISCUSION
1. Caveolina-1 disminuye la expresión de COX-2 por un mecanismo
transcripcional dependiente de β-catenina-Tcf/Lef
1.1. Inhibición de la expresión de COX-2 en células humanas de cáncer de colon,
de mama y de origen embrionario
El rol de caveolina-1 de funcionar como un supresor de tumores se ha atribuido
en parte a su capacidad de funcionar como un regulador negativo de la transcripción
dependiente de β-catenina-Tcf/Lef (Quest et al., 2008). Ejemplo de genes blancos que
son regulados negativamente debido a la represión de la transcripción dependiente de
β-catenina-Tcf/Lef por caveolina-1 (Galbiati et al., 2000) incluye ciclina D1 (Hulit et al.,
2000) y survivina (Torres et al., 2006). Asociado con estos cambios, se ha observado
una menor proliferación celular y una susceptibilidad incrementada a la muerte por
apoptosis (Torres et al., 2006; Torres et al., 2007).
En el objetivo específico 1 de esta tesis se investigó si la expresión de cox-2, otro
gen regulado por la vía de β-catenina-Tcf/Lef (Araki et al., 2003), es afectada por la
expresión ectópica de caveolina-1. Nuestros hallazgos muestran que la presencia de
caveolina-1 reduce los niveles de mRNA y proteicos de COX-2 en células de cáncer de
mama (ZR75) y de colon [HT29(ATCC); DLD-1] (Figura 9). También, una inhibición
similar se observó en células embrionarias humanas de riñón (HEK293T) (Figura 10), a
pesar de que estas células expresan niveles muy bajos de COX-2 (Figura 8). La
relevancia de este hallazgo sugiere que caveolina-1 reprime efectivamente la expresión
de COX-2 por un mecanismo transcripcional dada la reducción en los niveles de mRNA.
Sin embargo, aparentemente esta regulación podría ser exclusivamente dependiente
del contexto celular estudiado, ya que como se discutirá más adelante, este efecto
ocurre en células donde la presencia de caveolina-1 disminuye la actividad

101
transcripcional de β-catenina-Tcf/Lef. Experimentos adicionales in vivo serían de gran
aporte para confirmar estos hallazgos (análisis de células ex-tumor). También es
importante destacar otros mecanismos adicionales que puedan explicar la reducción en
los niveles del mRNA de COX-2. Por ejemplo, en células de cáncer de colon, que la
proteína HuR se une a la región 3`-UTR del mRNA de cox-2 aumentando su vida media
y favoreciendo la expresión constitutiva de COX-2 (Cok et al., 2003; Doller et al., 2008).
En nuestros estudios no se exploró la posibilidad que la presencia de caveolina-1
pueda afectar la vida media del mRNA de COX-2.
1.2. La inhibición de la expresión de COX-2 por caveolina-1 requiere la presencia
de E-cadherina
En un estudio reciente de nuestro laboratorio, se demostró el requerimiento de
E-cadherina en la supresión eficiente de la transcripción dependiente de
β-catenina-Tcf/Lef y la expresión de survivina. Este fenómeno fue corroborado en
células HT29(US) que carecen de E-cadherina, mediate la expresión ectópica de esta
proteína (Torres et al., 2007). Similarmente, en esta tesis se observó en células
HT29(US) que los niveles de mRNA de COX-2 no fueron afectados por la expresión
ectópica de caveolina-1 (Figura 11A) y que la co-expresión de E-cadherina/caveolina-1
restaura la inhibición de la expresión de COX-2 como fue observado en las otras líneas
celulares (Figura 11B-C). Por lo tanto, estos hallazgos refuerzan la idea que la pérdida
de la expresión de E-cadherina durante el desarrollo tumoral representa un paso
importante en la generación de un microambiente celular permisivo donde la capacidad
de caveolina-1 de ejercer su rol como supresor de tumores y/o inhibición de la
metástasis se ve severamente deteriorada (Quest et al., 2004; Quest et al., 2008).

102
Por ejemplo, estímulos adicionales del microambiente como el tratamiento
crónico con EGF, disminuye la expresión de caveolina-1, E-cadherina y potencia la
invasión celular y por ende la metástasis (Lu et al., 2003). Concomitantemente, se ha
descrito en células de epitelio mamario (HC11) el corte proteolítico de E-cadherina
inducido por receptores de estrógenos (ER). Específicamente, en ausencia de un
receptor ERβ, el receptor ERα promueve el corte del dominio extracelular de
E-cadherina (“shedding”) y a su vez, el dominio restante de E-cadherina es endocitado
en un proceso dependiente de caveolas (Helguero et al., 2008). Esto conlleva a la
pérdida en la interacción de E-cadherina con β-catenina en la membrana plasmática,
la translocación de β-catenina al núcleo, aumentando la actividad transcripcional de
β-catenina-Tcf/Lef (Helguero et al., 2008). Un mecanismo similar podría estar
ocurriendo en nuestro modelo de trabajo, donde la asociación de
E-cadherina/caveolina-1/β-catenina en la membrana plasmática regula la actividad
transcripcional de β-catenina-Tcf/Lef. Sin embargo, nuestros estudios no han
clarificado si eventualmente en las líneas celulares utilizadas, que poseen bajos
niveles proteicos de E-cadherina (HT29(US) y melanoma B16F10), existe un corte
proteolítico de E-cadherina. Futuros estudios apuntarían a resolver si existe este
mecanismo en células de cáncer que poseen niveles indetectables de E-cadherina.
1.3. La inhibición de la expresión de COX-2 por caveolina-1 ocurre por la
represión de la vía de β-catenina-Tcf/Lef
La ausencia de caveolina-1 se asocia con la activación de la vía de
β-catenina-Tcf/Lef (Reportero TOP/FOP) y específicamente con la activación del gen de
cox-2. Como se mencionó anteriormente, estudios recientes han mostrado que COX-2
es un blanco transcripcional de β-catenina-Tcf/Lef (Haertel-Wiesmann et al., 2000), ya

103
que contiene un sitio TBE en su promotor (Araki et al., 2003). En concordancia, en esta
tesis se muestra que la expresión de caveolina-1 en células embrionarias (HEK293T)
disminuye la actividad transcripcional de β-catenina-Tcf/Lef y disminuye
significativamente la actividad del reportero de COX-2 (promotor de cox-2) (Figuras 13 y
14), asi mismo también los niveles basales de mRNA y proteicos de COX-2 (Figuras 9,
10 y 13).
Para confirmar que la inhibición de COX-2 por caveolina-1 ocurre por esta vía, se
utilizó como control un inhibidor específico de la quinasa GSK-3β (SB-216763), ya que,
se ha descrito que la inhibición de la quinasa GSK-3β aumenta la actividad
transcripcional de β-catenina-Tcf/Lef e incrementa la expresión de COX-2 (Norvell et al.,
2004; Thiel et al., 2006). En nuestro modelo, el uso del inhibidor específico aumentó la
expresión de COX-2 (Figura 12A-B) e importantemente, revirtió significativamente la
inhibición de la actividad del reportero de COX-2 causada por la presencia de
caveolina-1 en células HEK293T, ZR75 y DLD-1 (Figuras 13 y 14) demostrando que la
regulación de β-catenina es crucial en la represión o activación transcripcional.
Por otra parte, se ha demostrado la co-localización e interacción de COX-2 con
caveolina-1 en fibroblastos humanos estimulados con un éster de forbol (PMA) (Liou et
al., 2001). Además, mediante experimentos in vitro, se observó que la proteína de
fusión, glutatión-S-transferasa-caveolina-1 se une directamente con COX-2 sin afectar
la actividad de COX-2, sugiriendo que caveolina-1 no regula COX-2 por este
mecanismo post-traduccional (Liou et al., 2001). Sin embargo, otros tipos de regulación
post-traducionales mediados por caveolina-1, tales como una aumentada degradación
por el proteasoma (Felley-Bosco et al., 2000; Felley-Bosco et al., 2002) o bien una
desestabilización del mRNA de COX-2 (Cok et al., 2003) no pueden ser descartados en
este estudio.
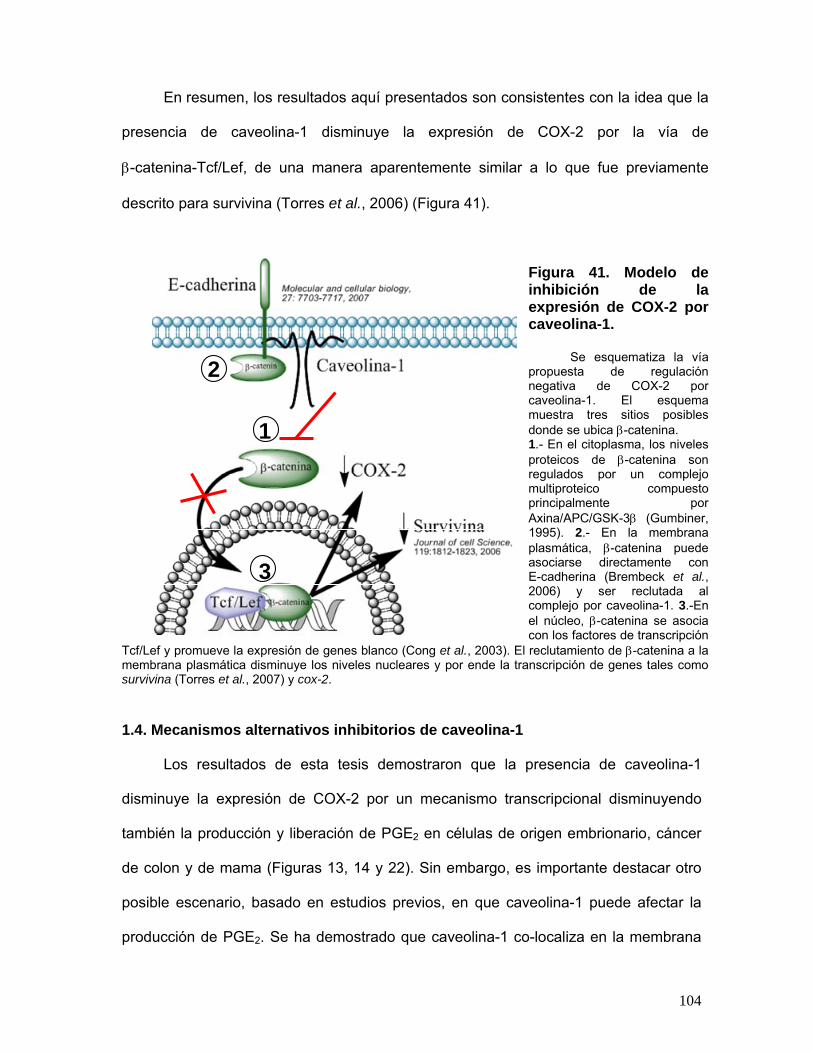
104
2
3
1
En resumen, los resultados aquí presentados son consistentes con la idea que la
presencia de caveolina-1 disminuye la expresión de COX-2 por la vía de
β-catenina-Tcf/Lef, de una manera aparentemente similar a lo que fue previamente
descrito para survivina (Torres et al., 2006) (Figura 41).
Figura 41. Modelo de inhibición de la expresión de COX-2 por caveolina-1.
Se esquematiza la vía
propuesta de regulación negativa de COX-2 por caveolina-1. El esquema muestra tres sitios posibles donde se ubica β-catenina. 1.- En el citoplasma, los niveles proteicos de β-catenina son regulados por un complejo multiproteico compuesto principalmente por Axina/APC/GSK-3β (Gumbiner, 1995). 2.- En la membrana plasmática, β-catenina puede asociarse directamente con E-cadherina (Brembeck et al., 2006) y ser reclutada al complejo por caveolina-1. 3.-En el núcleo, β-catenina se asocia con los factores de transcripción
Tcf/Lef y promueve la expresión de genes blanco (Cong et al., 2003). El reclutamiento de β-catenina a la membrana plasmática disminuye los niveles nucleares y por ende la transcripción de genes tales como survivina (Torres et al., 2007) y cox-2. 1.4. Mecanismos alternativos inhibitorios de caveolina-1
Los resultados de esta tesis demostraron que la presencia de caveolina-1
disminuye la expresión de COX-2 por un mecanismo transcripcional disminuyendo
también la producción y liberación de PGE2 en células de origen embrionario, cáncer
de colon y de mama (Figuras 13, 14 y 22). Sin embargo, es importante destacar otro
posible escenario, basado en estudios previos, en que caveolina-1 puede afectar la
producción de PGE2. Se ha demostrado que caveolina-1 co-localiza en la membrana

105
plasmática con la fosfolipasa A2 (PLA2) (Graziani et al., 2004) y con COX-2 (Liou et
al., 2001; Perrone et al., 2007), específicamente en caveolas (Graziani et al., 2004;
Perrone et al., 2007). También que caveolina-1 co-inmunoprecipita y co-fracciona con
ambas proteínas (Liou et al., 2001; Gaudreault et al., 2004). Interesantemente,
caveolina-1 a través de su dominio “Caveolin-1 Scaffolding Domain (CSD)” inhibe la
actividad enzimática de la PLA2 disminuyendo la producción del ácido araquidónico
(Gaudreault et al., 2004). Sin embargo, no afecta la actividad enzimática de COX-2
(Liou et al., 2001). Por lo tanto, en este complejo entre caveolina-1/PLA2/COX-2,
COX-2 permanecerá activa pero tendrá menos disponibilidad de ácido araquidónico
(inhibición de la PLA2 por caveolina-1) para producir PGH2, el cual es un precursor de
las enzimas específicas que producen los diferentes eicosanoides, entre ellos la PGE2
(Park et al., 2006).
La figura 42 sugiere un mecanismo inhibitorio dual de caveolina-1 sobre la
producción de PGE2. El primero se relaciona con la inhibición a nivel transcripcional de
la expresión de COX-2 por caveolina-1 y el segundo, propone como hipótesis
alternativa, que caveolina-1 al co-localizar con PLA2 y COX-2 en la membrana
plasmática podría disminuir la disponibilidad de ácido araquidónico debido a la
inhibición de la actividad enzimática de la PLA2 (dominio CDS de caveolina-1), por
ende, COX-2 tiene una menor disponibilidad del precursor ácido araquidónico
disminuyendo la producción de PGE2, la actividad transcripcional de β-catenina-Tcf/Lef
y activación transcripcional de survivina y cox-2 (ver más adelante). Sin embargo, no
se puede descartar la posibilidad que caveolina-1 interfiera en el traspaso concertado
de precursores entre ambas enzimas debido a su interacción directa con PLA2 y
COX-2, o bien que interfiera en el transporte de la PGE2 desde el citosol al exterior de
la célula (se discutirá mas adelante).

106
Figura 42. Mecanismos alternativos inhibitorios de caveolina-1.
Se esquematiza un mecanismo inhibitorio dual de caveolina-1 sobre la producción de PGE2. En A se muestra la inhibición a nivel transcripcional de la expresión de COX-2 por caveolina-1 (Objetivos específicos 1 y 2). En B se propone alterna-tivamente que caveolina-1 al co-localizar con fosfolipasa A2 (PLA2) y COX-2 en la membrana plasmática podría disminuir la disponibilidad de ácido araquidónico (AA) debido a la inhibición de la actividad enzimática de la PLA2 (dominio CDS de caveolina-1), por ende, COX-2 tiene una menor disponibilidad del pre-cursor ácido araquidónico
disminuyendo la producción de PGE2 y esta disminución conllevaría a una reducida actividad transcripcional de β-catenina-Tcf/Lef (Castellone et al., 2005) y activación transcripcional de survivina y cox-2. 2. La sobreexpresión de COX-2 o la incubación con PGE2 estimulan la actividad
transcripcional de β-catenina-Tcf/Lef aumentando la expresión de survivina y
COX-2
2.1. Efecto de COX-2 sobre la transcripción dependiente de β-catenina-Tcf/Lef
La expresión constitutiva de COX-2 se ha asociado con angiogénesis, invasión
celular, resistencia a la apoptosis y promoción de la proliferación de células cancerosas
(Kanaoka et al., 2007). Se han estudiado ampliamente los mecanismos moleculares
involucrados en el desarrollo del cáncer, en particular, en el cáncer de colon, la
aplicación de nuevas terapias y búsqueda de blancos terapéuticos eficaces, como por
ejemplo COX-2, sigue siendo un tema de continua investigación dada su gran
complejidad. En este contexto, el estudio de pacientes que padecen de cáncer de colon
A
B

107
tratados con NSAIDs ha revelado efectos benéficos en la prevención del cáncer de
colon (Herendeen y Lindley, 2003), específicamente en pacientes que se les administró
celecoxib (Arber et al., 2006) o bien aspirina (Chan et al., 2007).
Sin embargo, muchos de los mecanismos y vías involucradas de manera
simultánea requieren de un mayor entendimiento. En base a ello, este estudio se
focalizó en primera instancia en el papel de caveolina-1 como supresor de tumores,
dado los antecedentes previamente presentados y por otra parte en el papel de COX-2
en promover la proliferación y resistencia a la muerte celular.
Como fuera mencionado, muchos estudios han demostrado que COX-2 se
encuentra sobreexpresada en cáncer de colon (Tsujii et al., 1997; Hasegawa et al.,
2001). Para simular en parte los efectos de la sobreexpresión de COX-2 se utilizó una
línea celular de origen embrionario HEK293T que expresa niveles basales bajos de
COX-2 (Figura 8), pero que posee la vía de wnt/β-catenina/Tcf/Lef funcional. Los
experimentos de sobreexpresión de COX-2 revelaron un interesante hallazgo, ya que
en la medida que se sobreexpresó COX-2 aumentó la expresión de survivina
(Figura 26).
Survivina, como se mencionó anteriormente es un gen que se sobreexpresa en la
mayoría de los distintos tipos de cáncer (Kim et al., 2003) y se sigue investigado y
utilizado como un posible blanco terapéutico (Ambrosini et al., 1997; Grossman et al.,
2001; Altieri, 2003). En el modelo celular (HEK293T) investigamos adicionalmente
cómo COX-2 podría modular la actividad transcripcional de β-catenina-Tcf/Lef, ya que
se ha demostrado previamente que PGE2 activa esta vía (Shao et al., 2005), a través
de la inhibición de la quinasa GSK-3β (Lim et al., 2008). Nuestros hallazgos revelaron
que la sobreexpresión de COX-2 incrementa la actividad del reportero de survivina y el
promotor de cox-2, indicando un mecanismo de retroalimentación positivo (Figura 16).

108
Específicamente, nuestros datos muestran que este proceso podría estar
mediado por una mayor producción de PGE2, ya que la sobreexpresión de COX-2 se
correlacionó con una mayor producción y liberación de PGE2 (Figura 18).
Además, PGE2 como ya se mencionó, es un producto importante derivado de la
actividad enzimática de COX-2 que incrementa los niveles proteicos de β-catenina a
través de la inhibición de la quinasa GSK-3β (Lim et al., 2008), aumenta la actividad
transcripcional (Shao et al., 2005) y promueve el crecimiento de células tumorales de
cáncer de colon (Shao et al., 2003; Hawcroft et al., 2007). Con esta amplia variedad de
efectos de PGE2, no sorprende que actualmente se estén desarrollando inhibidores
específicos para los receptores de prostaglandinas (Robertson et al., 2008; Steinert et
al., 2009) con el fin de evitar los efectos adversos de un exceso de PGE2.
En concordancia, sólo la adición de PGE2 produjo el mismo efecto, es decir,
aumenta la expresión de survivina y COX-2, lo cual se tradujo también en una mayor
proliferación celular (Figura 21B), sugiriéndose que dichos efectos son mediados por un
mecanismo de activación transcripcional (Figuras 19, 20 y 21).
Sin embargo, estudios realizados por Krissan y cols. (2003) muestran que PGE2
aumenta los niveles proteicos de survivina debido a una menor degradación por el
proteasoma, lo cual indicaría otro mecanismo alternativo que podría también ser
funcional en nuestro modelo de trabajo. Cabe destacar que PGE2 ejerce su acción, en
parte, a nivel de la membrana plasmática donde se encuentran situados los receptores
de prostaglandinas EP1-4 (Schuster, 2002), no descartándose su acción a nivel
intracelular.
Una vez activados los receptores de PGEs, éstos pueden activar diversas vías
de transducción de señales (Regan, 2003), específicamente el receptor EP2 activa la
vía de PI3K/Akt que termina por inhibir la quinasa GSK-3β y por ende aumentan los

109
niveles proteicos de β-catenina (Figuras 5 y 41) explicándose la mayor actividad
transcripcional encontrada en este estudio (Figura 20) y previamente descrito
(Castellone et al., 2005). Sin embargo, queda por dilucidar otras vías que puedan estar
simultáneamente activas y conduzcan a una menor actividad del proteasoma y por
ende una menor degradación de survivina como fuera descrito (Krysan et al., 2004c).
Por otra parte, también en células DLD-1, la sobreexpresión de COX-2 incrementó la
actividad transcripcional de β-catenina-Tcf/Lef y específicamente el reportero
conteniendo el promotor de survivina. Esto se tradujo en mayores niveles de mRNA y
proteína de survivina, así como también, mayor producción y liberación de PGE2
(Figuras 24 y 25).
Cabe destacar que estas células poseen niveles basales de COX-2 cuatro veces
más alto que células HEK2932T (Figura 8) indicando que COX-2 se encuentra
sobreexpresada. Pero aun así, la expresión ectópica de COX-2 ejerce una fuerte
activación de los promotores de survivina y cox-2, sugiriendo que el sustrato ácido
araquidónico no sería un factor limitante para la producción de PGE2. Estos datos
apoyarían el mecanismo inhibitorio de caveolina-1, vía secuestro de β-catenina
(Figura 42, ruta A) y no la vía de inhibición de PLA2 (Figura 42, ruta B).
2.2. Efecto de PGE2 y COX-2 sobre la regulación negativa de caveolina-1 en la
expresión de survivina
Como fue previamente mostrado, la presencia de caveolina-1 disminuye la
expresión de survivina en un escenario que depende de la expresión de E-cadherina
(Figuras 9-11) y de este modo ocurre el reclutamiento de β-catenina a la membrana
plasmática (Torres et al., 2007) (Figuras 33 y 34). En este estudio se confirmó dicho
fenómeno e interesantemente, la incubación con PGE2 en células que expresan o no

110
caveolina-1 disminuyó la co-inmunoprecipitación de caveolina-1 con β-catenina,
experimento que fue controlado al realizar la inmunoprecipitación inversa, es decir se
inmunoprecipitó β-catenina y se cuantificó los niveles proteicos de caveolina-1
(Figuras 31 y 32). A su vez, mediante inmunofluorescencia indirecta se observó una
disminución de β-catenina en la membrana plasmática en aquellas células que
expresan caveolina-1 (Figuras 33 y 34), fenómeno que corrobora lo observado por
inmunoprecipitaciones.
Respecto, a los hallazgos con PGE2 y su efecto inhibitorio en la formación de
los complejos caveolina-1/β-catenina resta aún clarificar si dicha inhibición sólo es
producto de la señalización debido a la unión de la PGE2 al receptor EP2, o bien el
receptor EP4 también podría contribuir, dado que la señalización por el receptor EP4
también puede inhibir la quinasa GSK3β (Hull et al., 2004). Para descartar o incluir
esta posibilidad, se podrían realizar experimentos de pérdida o ganancia de función
utilizando células de cáncer de colon HT29(ATCC) que se sabe expresan los
receptores EP1-4 (Chell et al., 2006). En experimentos de pérdida de función, el uso
de inhibidores específicos de los receptores EP1-4 (Cherukuri et al., 2007; Birkenmeier
et al., 2008), o bien el uso de shRNA para inhibir la expresión de estos receptores
(Robertson et al., 2008), permitiría explorar un posible efecto intracelular de la PGE2 y
por otra parte, en experimentos de ganancia de función, el uso de agonistas selectivos
de cada receptor EP1-4 (Xiao et al., 2007) otorgaría información valiosa sobre la
contribución de cada receptor en la señalización que involucra una activación de la
actividad transcripcional de β-catenina-Tcf/Lef y su efecto inhibitorio en la formación de
los complejos entre caveolina-1/β-catenina.
Cabe destacar que la PGE2 es inactivada (catabolismo intracelular) en el citosol
y llevada a su forma cetónica a través de un proceso mediado por la enzima

111
15-hidroxiprostaglandina deshidrogenasa (15-PGDH) (Ensor y Tai, 1995). Sin
embargo, esta enzima no se expresa en células de cáncer de colon y tampoco en
células HT29 (Backlund et al., 2005), lo cual explicaría en parte el efecto sostenido de
PGE2 considerando que es una molécula con una vida media corta (Bygdeman, 2003).
PGE2 es transportada o bien pasa a través de la membrana vía
transportadores específicos, PGT, que son transportadores que permiten su entrada y
así permiten su remoción del medio extracelular. Por otra parte, existen
transportadores que permiten su salida, por ejemplo la proteína 4 asociada a la
multiresistencia a drogas (MRP-4). Interesantemente, la expresión de las proteínas
PGT se encuentran disminuidas mientras que la expresión de las proteínas MRP-4
está elevada en cáncer de colon y células HT29 (Holla et al., 2008), estos
antecedentes concuerdan con nuestros estudios, donde la adición exógena de PGE2
no sería internalizada para su catabolismo y favorecería la señalización a través de
sus receptores.
PGE2 podría también ejercer un efecto citosólico sobre la formación del complejo
entre β-catenina/caveolina-1. Respecto a la formación de estos complejos
multiproteicos es importante señalar que en experimentos de inmunoprecipitación de
caveolina-1 y posterior análisis de los complejos por Western blot o espectrometría de
masas, una limitante importante fue la baja abundancia relativa de las proteínas que
forman dichos complejos. Caveolina-1 como proteína de andamio puede unir diversas
proteínas que se asocian a membrana y caveolas o también puede unir proteínas
presentes en retículo-aparato de Golgi. Dada esta complejidad resultó difícil identificar
las diferentes proteínas asociadas a caveolina-1 en la membrana plasmática que
podrían formar parte del complejo β-catenina/caveolina-1. Nuestros datos obtenidos en
la estadía de investigación en el laboratorio del Dr. Jay Heinecke entregaron

112
información valiosa en la identificación de las proteínas que conforman los complejos
multiproteicos con caveolina-1.
Un hallazgo interesante, fue la identificación de proteínas de andamio de
fosfatasas de tirosina (Liprin alpha 2 y 4) (Tabla 3), dado que estas proteínas se
detectaron sólo en células HT29(ATCC) que expresan caveolina-1. Se ha descrito que
la ubicación de β-catenina en la célula es dependiente de los sitios donde sea
fosforilada, es decir si es fosforilada en la tirosina 654 es destinada a su degradación
vía el proteasoma o bien si es fosforilada en la tirosina 142 se transloca al núcleo
donde promueve la actividad transcripcional (Brembeck et al., 2006). Sin embargo, para
que β-catenina pueda asociarse con E-cadherina en la membrana plasmática requiere
estar desfosforilada (Brembeck et al., 2006), para lo cual es importante la presencia de
fosfatasas de tirosina (Figura 43).
Figura 43. β-catenina se ubica en la membrana plasmática sólo en su forma desfosforilada.
El esquema modificado desde
(Brembeck et al., 2006) muestra la función dual de β-catenina en la interacción célula-célula y en la actividad transcripcional. En la membrana plasmática, β-catenina en su forma desfosforilada, participa en la interacción célula-célula al unirse con E-cadherina y α-catenina. En el citoplasma, los niveles de β-catenina son regulados por su degradación mediada por un complejo multiproteico compuesto por las proteínas APC, GSK-3β y axina. La formación de este complejo requiere la fosforilación de β-catenina por la quinasa c-src en el residuo Y654 cercano al carboxilo-terminal de β-catenina. En cambio, la fosforilación de β-catenina en su residuo Y142 (cercano al amino-terminal) por quinasas como Fer o Fyn, promueve la translocación nuclear de
β-catenina, su asociación con los factores de trascripción Tcf/Lef y la subsecuente expresión de genes blanco como cox-2 y survivina.

113
Una de las proteínas fosfatasas que ejercen dicha acción es una proteína tirosina
fosfatasa (PTPRF) que al ser sobreexpresada disminuye la actividad transcripcional de
β-catenina-Tcf/Lef de manera dependiente de E-cadherina (Yan et al., 2006).
Interesantemente, esta proteína fosfatasa PTPRF, puede a su vez interactuar con estas
proteínas liprinas y quizás formar de esta manera parte de este complejo multiproteico
que mantendría a β-catenina desfosforilada. En consecuencia, β-catenina puede
asociarse con E-cadherina en la membrana plasmática y formar el complejo
multiproteico con caveolina-1. Nuestro estudio, por lo tanto sugiere, que la presencia de
caveolina-1 facilitaría la localización de esta proteína reguladora de fosfatasa en la
membrana plasmática, de tal modo que se estaría promoviendo la desfosforilación de
β-catenina y así su reclutamiento a la membrana plasmática reprimiendo la actividad
transcripcional de β-catenina-Tcf/Lef en un proceso dependiente de E-cadherina.
Por otra parte, la presencia de PGE2 y su unión al receptor EP2 promueve la
acumulación citoplasmática de β-catenina debido a la inhibición de la quinasa GSK-3β
de tal modo que aumenta la translocación nuclear de β-catenina y así la actividad
transcripcional de β-catenina-Tcf/Lef. Sin embargo, en este estudio a pesar que PGE2
disminuyó la formación del complejo entre β-catenina/caveolina-1, disminuyendo la
presencia de β-catenina en la membrana plasmática, no se identificó el mecanismo
específico, especulándose que PGE2 podría modular negativamente la actividad de las
fosfatasas o bien PGE2 podría activar la actividad enzimática de las tirosina quinasas
Fer o Fyn, ya que se ha descrito que éstas, al fosforilar a β-catenina en el residuo de
tirosina 142, disminuye la asociación de β-catenina con α-catenina en la membrana
plasmática (Piedra et al., 2003).
Otro mecanismo alternativo de regulación positiva en la actividad transcripcional
de β-catenina-Tcf/Lef, lo establece el rol de la caseína quinasa 2 (CK2). Esta

114
serina/treonina quinasa participa en la señalización de Wnt, ya que se ha sugerido que
la CK2 formaría un complejo multiproteico con la proteína Dvl (dishevelled) evitando la
degradación de β-catenina y por ende promoviendo la transcripción dependiente de
β-catenina-Tcf/Lef (Song et al., 2000). En concordancia, estudios previos de nuestro
laboratorio demostraron que la sobreexpresión de CK2 en células HEK293T aumenta la
actividad transcripcional de β-catenina-Tcf/Lef y la expresión de survivina y la inhibición
específica de la CK2 disminuye dicho efecto (Tapia et al., 2006).
Estos antecedentes nos llevan a especular en nuestro modelo de trabajo que la
CK2 podría evitar la formación del complejo multiproteico entre β-catenina/
E-cadherina/caveolina-1 en un proceso en que CK2 indirectamente promueva la
fosforilación de β-catenina en el residuo Y142 (cercano al N-terminal), aumentando la
translocación nuclear de β-catenina, su asociación con los factores de transcripción
Tcf/Lef y la subsecuente expresión de genes blanco (ver figura 45). Dicho proceso se
especula ocurrir en paralelo al efecto observado por una mayor expresión de COX-2 y
la presencia de PGE2.
3. La inhibición específica de COX-1 o COX-2 potencia la disminución de la
proliferación inducida por caveolina-1
En este trabajo de tesis se investigó si los niveles proteicos reducidos de COX-2
en las distintas líneas celulares de cáncer de colon [HT29(ATCC) y DLD-1], de mama
(ZR75) y de origen embrionario (HEK2893T) que expresan caveolina-1 se correlacionan
con niveles menores de PGE2. Efectivamente, la cuantificación reveló niveles menores
de PGE2 en los medios de cultivo y además, los distintos niveles de PGE2 se
correlacionan con los distintos niveles proteicos de COX-2 en cada línea celular, es
decir ordenadas de mayor a menor los niveles proteicos de COX-2 son: HT29 > DLD-1

115
> ZR75 > HEK293T (Figura 8), en concordancia los niveles de PGE2 se ordenan del
mismo modo. Sin embargo, a pesar de que la presencia de caveolina-1 disminuyó
significativamente los niveles de PGE2, no se observó un bloqueo completo en la
producción de PGE2, lo cual nos llevó a pensar la posible contribución de la COX-1, ya
que por ejemplo, células de cáncer de mama ZR75 no revelaron una disminución
significativa en la producción de PGE2 y células de cáncer de colon disminuyeron su
producción en aproximadamente un 40-50% (Figura 22). Para resolver esta
interrogante se incubó células DLD-1 en presencia de inhibidores específicos de las
enzimas COX-1 y COX-2. La incubación de células DLD-1 que expresan o no
caveolina-1 con cantidades crecientes del inhibidor de COX-2 (SC-971; 0, 1, 5 y
20 μM), siendo concentraciones superiores al IC50 4 nM (determinada in Vitro) (Blomme
et al., 2003) y lo descrito en cultivo celular concuerda a lo utilizado en este estudio
(Furst et al., 2005). Los resultados revelaron que no existe un bloqueo completo en la
producción de PGE2, así mismo también se observó con el inhibidor de COX-1, pero en
menor grado y no significativo.
Estos datos nos llevaron a especular al menos tres escenarios distintos que
permitan explicar lo observado. Primero, se podría pensar que las células poseen de
algún modo un reservorio vesicular de PGE2 de tal modo que los efectos, ya sea por si
solo de caveolina-1 o bien ambos (presencia de caveolina-1 mas inhibidores) no logran
reducir completamente la presencia de PGE2, debido a que sigue siendo liberada desde
reservorios intracelulares. Segundo, una elevada vida media de su precursor PGH2
permitiría mantener una producción de PGs independiente de una menor producción de
PGH2 a partir del ácido araquidónico y tercero, podría existir otra prostaglandina en
mayor abundancia que pueda interferir en nuestras mediciones.

116
En células ZR75 nuestros datos fueron contradictorios respecto a las otras líneas
celulares usadas, debido a que en ausencia y presencia de caveolina-1, si bien
caveolina-1 disminuyó la producción de PGE2 los cambios no fueron significativos,
además la adición exógena de PGE2 sólo aumentó la expresión de COX-2 y no
survivina. Sería necesario hacer experimentos adicionales, como por ejemplo,
cuantificar la PGE2 y realizar ensayos de proliferación en presencia de los inhibidores
específicos de COX-1 y COX-2, así como evaluar la expresión de los receptores de
PGE2 (EP1-4) y el efecto de agonistas y antagonistas específicos, de tal modo que se
pueda explicar por que éstas células presentan una respuesta diferencial comparado
con las líneas de cáncer de colon.
Por otra parte, los resultados obtenidos en células DLD-1, utilizando por
separado ambos inhibidores reveló que caveolina-1 potencia la inhibición específica de
COX-1 o COX-2 disminuyendo la producción de PGE2, actividad transcripcional de
β-catenina-Tcf/Lef y expresión de survivina, efecto que se tradujo en una disminución
significativa en la proliferación celular.
Estos datos confirman el papel de survivina en la proliferación, ya que
previamente nuestro grupo demostró que la sobreexpresión de survivina previene la
muerte celular por apoptosis inducida por un inhibidor específico de la CK2α (Tapia et
al., 2006). En concordancia, la disminución de la proliferación inducida por la presencia
de caveolina y en conjunto con los inhibidores de COX-1/2 fue prevenida cuando
células DLD-1 o bien HT29(ATCC) fueron incubadas con PGE2 (Figuras 27 y 28), de la
cual previamente se demostró que es un buen inductor de la expresión de survivina y
por ende de la proliferación (Figuras 19 y 21).

117
4. La presencia de caveolina-1 disminuye la vida media de survivina
Como fue previamente mencionado la presencia de caveolina-1 promueve la
degradación de la sintasa inducible del oxido nítrico (iNOs) vía el proteasoma (Felley-
Bosco et al., 2000; Felley-Bosco et al., 2002).
Estos antecedentes, sumado a los hallazgos por espectrometría de masas en
donde se encontró la co-inmunoprecipitación de proteínas ligasas de ubiquitina con
caveolina-1 en células HT29(ATCC) y en HT29(US), apoyan la idea que podría existir
un mecanismo anexo de regulación post-transcripcional de survivina, en el cual
caveolina-1 promovería su degradación vía el proteasoma (Figura 44). Sin embargo, los
datos aquí presentados son preliminares, ya que sólo se ha visto que la presencia de
caveolina-1 disminuye la vida media de survivina en células HT29(US) (Figura 40),
quedando aún por investigar el efecto de los inhibidores del proteasoma y el menor o
mayor grado de ubiquitinación de survivina en células que expresan o no caveolina-1.
También es interesante mencionar que este efecto de caveolina-1 sobre
survivina es independiente de la presencia de E-cadherina sugiriendo que posiblemente
existe un pool de survivina y caveolina-1 que podría estar conformando un complejo
multiproteico de tal modo que caveolina-1 promueva así la ubiquitinación y por ende
degradación de survivina, ya que resultados preliminares no mostrados (Datos del
laboratorio) revelaron que una pequeña cantidad de survivina co-inmunoprecipita con
caveolina-1. Sin embargo, survivina no fue detectada mediante espectrometría de
masas, posiblemente debido a la baja abundancia de survivina en los complejos
multiproteicos.
Otro punto importante de mencionar, es que se debería investigar el posible rol
de PGE2 en la modulación de la degradación de survivina en células que expresen o no
caveolina-1, ya que como fue previamente descrito, PGE2 previene la degradación de

118
survivina por el proteasoma (Krysan et al., 2003; Krysan et al., 2004c). Una predicción
muy posible es que en células que expresan caveolina-1, survivina posee un mayor
grado de ubiquitinación y se degrada más rápidamente. Al incubar estas células con
PGE2 se prevendría tal efecto sobre survivina.
Figura 44. La presencia de caveolina-1 podría promover la degradación de survivina vía el proteasoma. Se ha descrito en la literatura que los niveles proteicos de survivina son regulados a nivel post-traduccional mediante su degradación vía el proteasoma (Zhao et al., 2000). El siguiente esquema propone que la presencia de caveolina-1 promueve la degradación de survivina vía el proteasoma, ya que proteínas que participan en esta vía co-inmunoprecipitan con caveolina-1 en células de cáncer de colon (ver tablas 3 y 4). Adicionalmente, estudio previos del laboratorio demostraron que caveolina-1 promueve la degradación de la sintasa inducible del oxido nítrico en células de cáncer de colon (Felley-Bosco et al., 2000; Felley-Bosco et al., 2002).
Caveolina-1
Degradación de survivina vía el proteosoma
survivina
Caveolina-1
Degradación de survivina vía el proteosoma
survivina
proteasoma

119
VIII. CONCLUSION Y PROYECCIONES
En conclusión, nuestros principales hallazgos permiten formular un modelo de
trabajo en donde caveolina-1 inhibe la expresión de COX-2 por un mecanismo
transcripcional que involucra la regulación negativa de la vía de β-catenina-
Tcf/Lef. Adicionalmente, dicha inhibición puede ser sobrellevada por un mecanismo de
retroalimentación positivo de COX-2/PGE2 el cual interfiere en el reclutamiento de
β-catenina a la membrana plasmática por caveolina-1 promoviendo la expresión de
survivina y la proliferación celular (Figura 45). Por lo tanto, nuestros estudios indican
que caveolina-1 puede funcionar como un supresor de tumores, basándose en la idea
que su presencia disminuye la proliferación de una célula tumoral y retarda el
crecimiento tumoral en ratones, efecto que es sustentado por el hecho que se observó
una disminución de dos genes (cox-2 y survivina) que promueven la proliferación
tumoral y que se encuentran sobreexpresados en la gran mayoría de tumores
humanos.
Sin embargo, queda destacar que los efectos inhibitorios de caveolina-1 pueden
ser sobrellevados bajo algunas circunstancias como por ejemplo, la sobreexpresión de
COX-2, la presencia de un estímulo como PGE2 o posiblemente también por una
elevada expresión de la quinasa CK2, en todos estos casos se genera un potente
estímulo en la activación de la vía de β-catenina-tcf/Lef.
Finalmente, el esquema resume las principales vías estudiadas en esta tesis y
otros trabajos realizados en el laboratorio identificando por un lado, blancos río abajo
de caveolina-1 que explican su papel como supresor de tumores y por otro lado,
mecanismos que impiden el cumplimiento de este papel.

120
`
A
B
C
D
E
F
Figura 45. Resumen del trabajo del laboratorio en la identificación de blancos de caveolina-1 en su papel como supresor de tumores y de aquellos mecanismos que impiden que caveolina-1 cumpla este papel. (A) La presencia de caveolina-1 disminuye la expresión de survivina, por un mecanismo que involucra la actividad transcripcional dependiente de β-catenina-Tcf/Lef (Torres et al., 2006). La expresión de COX-2 también es regulada por este mecanismo (Rodriguez et al., 2009). (B) COX-2 promueve la síntesis de prostaglandina E2 (PGE2). En este estudio se ha mostrado que una mayor producción de PGE2 estimula la actividad transcripcional dependiente de β-catenina-Tcf/Lef y específicamente aumenta la expresión de survivina y de COX-2, proceso que permite un circuito de retroalimentación positivo (Rodriguez et al., 2009). (C) Caveolina-1 recluta a β-catenina a la membrana plasmática en un proceso dependiente de E-cadherina (Torres et al., 2007). La adición de PGE2 disminuye la formación de estos complejos aumentando la actividad transcripcional de β-catenina-Tcf/Lef y por ende previendo la represión por caveolina-1 de COX-2 y survivina y a su vez la proliferación celular. (D) La disminución de la expresión de survivina y de COX-2 por la presencia caveolina-1 de conlleva a una baja en la proliferación de células de cáncer. (E) A través de un mecanismo de regulación post-traduccional, posiblemente caveolina-1 estaría involucrada en promover la degradación de survivina por el proteasoma. (F) Experimentos de co-inmunoprecipitación sugieren que caveolina-1 en la formación de complejos multiproteicos con β-catenina/E-cadherina y posiblemente una fosfatasa PTPRF con una proteína activadora (Liprina), lo cual apoya la idea que una vez reclutada la β-catenina a la membrana plasmática se mantendría desfosforilada por dichas fosfatasa. (G) Estudios previos de nuestro laboratorio demostraron que la sobreexpresión de la caseína quinasa CK2 aumenta la actividad transcripcional de β-catenina-Tcf/Lef y por consiguiente la expresión de survivina (Tapia et al., 2006). Sin embargo, resta investigar si dicha regulación también se refleja en una mayor expresión de COX-2 e interesantemente, si la presencia de CK2 afectaría la formación de los complejos multiproteicos entre β-catenina/E-cadherina, contraponiéndose al efecto supresor de tumor de caveolina-1.
¿?
G CK2
Proc. Nat. Acad. Sci., 103(41), 15079-84, 2006
P

121
IX. REFERENCIAS Aberle, H., Bauer, A., Stappert, J., Kispert, A., and Kemler, R. (1997). beta-catenin is a target for the ubiquitin-proteasome pathway. Embo J 16, 3797-3804. Altieri, D.C. (2003). Validating survivin as a cancer therapeutic target. Nat Rev Cancer 3, 46-54. Altieri, D.C. (2008). New wirings in the survivin networks. Oncogene 27, 6276-6284. Ambrosini, G., Adida, C., and Altieri, D.C. (1997). A novel anti-apoptosis gene, survivin, expressed in cancer and lymphoma. Nat Med 3, 917-921. Anderson, R.G. (1998). The caveolae membrane system. Annu Rev Biochem 67, 199-225. Araki, Y., Okamura, S., Hussain, S.P., Nagashima, M., He, P., Shiseki, M., Miura, K., and Harris, C.C. (2003). Regulation of cyclooxygenase-2 expression by the Wnt and ras pathways. Cancer Res 63, 728-734. Arber, N., Eagle, C.J., Spicak, J., Racz, I., Dite, P., Hajer, J., Zavoral, M., Lechuga, M.J., Gerletti, P., Tang, J., Rosenstein, R.B., Macdonald, K., Bhadra, P., Fowler, R., Wittes, J., Zauber, A.G., Solomon, S.D., and Levin, B. (2006). Celecoxib for the prevention of colorectal adenomatous polyps. N Engl J Med 355, 885-895. Backlund, M.G., Mann, J.R., Holla, V.R., Buchanan, F.G., Tai, H.H., Musiek, E.S., Milne, G.L., Katkuri, S., and DuBois, R.N. (2005). 15-Hydroxyprostaglandin dehydrogenase is down-regulated in colorectal cancer. J Biol Chem 280, 3217-3223. Baratelli, F., Krysan, K., Heuze-Vourc'h, N., Zhu, L., Escuadro, B., Sharma, S., Reckamp, K., Dohadwala, M., and Dubinett, S.M. (2005). PGE2 confers survivin-dependent apoptosis resistance in human monocyte-derived dendritic cells. J Leukoc Biol 78, 555-564. Bender, F.C., Reymond, M.A., Bron, C., and Quest, A.F. (2000). Caveolin-1 levels are down-regulated in human colon tumors, and ectopic expression of caveolin-1 in colon carcinoma cell lines reduces cell tumorigenicity. Cancer Res 60, 5870-5878. Birkenmeier, K., Janke, I., Schunck, W.H., Trimpert, C., Krieg, T., Landsberger, M., Volker, U., Felix, S.B., and Staudt, A. (2008). Prostaglandin receptors mediate effects of substances released from ischaemic rat hearts on non-ischaemic cardiomyocytes. Eur J Clin Invest 38, 902-909. Blomme, E.A., Chinn, K.S., Hardy, M.M., Casler, J.J., Kim, S.H., Opsahl, A.C., Hall, W.A., Trajkovic, D., Khan, K.N., and Tripp, C.S. (2003). Selective cyclooxygenase-2 inhibition does not affect the healing of cutaneous full-thickness incisional wounds in SKH-1 mice. Br J Dermatol 148, 211-223. Brembeck, F.H., Rosario, M., and Birchmeier, W. (2006). Balancing cell adhesion and Wnt signaling, the key role of beta-catenin. Curr Opin Genet Dev 16, 51-59. Bygdeman, M. (2003). Pharmacokinetics of prostaglandins. Best Pract Res Clin Obstet Gynaecol 17, 707-716. Cao, Y., and Prescott, S.M. (2002). Many actions of cyclooxygenase-2 in cellular dynamics and in cancer. J Cell Physiol 190, 279-286. Capozza, F., Williams, T.M., Schubert, W., McClain, S., Bouzahzah, B., Sotgia, F., and Lisanti, M.P. (2003). Absence of caveolin-1 sensitizes mouse skin to carcinogen-induced epidermal hyperplasia and tumor formation. Am J Pathol 162, 2029-2039. Castellone, M.D., Teramoto, H., and Gutkind, J.S. (2006). Cyclooxygenase-2 and colorectal cancer chemoprevention: the beta-catenin connection. Cancer Res 66, 11085-11088.

122
Castellone, M.D., Teramoto, H., Williams, B.O., Druey, K.M., and Gutkind, J.S. (2005). Prostaglandin E2 promotes colon cancer cell growth through a Gs-axin-beta-catenin signaling axis. Science 310, 1504-1510. Cohen, A.W., Hnasko, R., Schubert, W., and Lisanti, M.P. (2004). Role of caveolae and caveolins in health and disease. Physiol Rev 84, 1341-1379. Cok, S.J., Acton, S.J., and Morrison, A.R. (2003). The proximal region of the 3'-untranslated region of cyclooxygenase-2 is recognized by a multimeric protein complex containing HuR, TIA-1, TIAR, and the heterogeneous nuclear ribonucleoprotein U. J Biol Chem 278, 36157-36162. Cong, F., Schweizer, L., Chamorro, M., and Varmus, H. (2003). Requirement for a nuclear function of beta-catenin in Wnt signaling. Mol Cell Biol 23, 8462-8470. Couet, J., Li, S., Okamoto, T., Ikezu, T., and Lisanti, M.P. (1997). Identification of peptide and protein ligands for the caveolin-scaffolding domain. Implications for the interaction of caveolin with caveolae-associated proteins. J Biol Chem 272, 6525-6533. Cha, Y.I., and DuBois, R.N. (2007). NSAIDs and cancer prevention: targets downstream of COX-2. Annu Rev Med 58, 239-252. Chan, A.T., Ogino, S., and Fuchs, C.S. (2007). Aspirin and the risk of colorectal cancer in relation to the expression of COX-2. N Engl J Med 356, 2131-2142. Chan, T.A., Morin, P.J., Vogelstein, B., and Kinzler, K.W. (1998). Mechanisms underlying nonsteroidal antiinflammatory drug-mediated apoptosis. Proc Natl Acad Sci U S A 95, 681-686. Chell, S., Patsos, H.A., Qualtrough, D., AM, H.Z., Hicks, D.J., Kaidi, A., Witherden, I.R., Williams, A.C., and Paraskeva, C. (2005). Prospects in NSAID-derived chemoprevention of colorectal cancer. Biochem Soc Trans 33, 667-671. Chell, S.D., Witherden, I.R., Dobson, R.R., Moorghen, M., Herman, A.A., Qualtrough, D., Williams, A.C., and Paraskeva, C. (2006). Increased EP4 receptor expression in colorectal cancer progression promotes cell growth and anchorage independence. Cancer Res 66, 3106-3113. Chen, J., Wu, W., Tahir, S.K., Kroeger, P.E., Rosenberg, S.H., Cowsert, L.M., Bennett, F., Krajewski, S., Krajewska, M., Welsh, K., Reed, J.C., and Ng, S.C. (2000). Down-regulation of survivin by antisense oligonucleotides increases apoptosis, inhibits cytokinesis and anchorage-independent growth. Neoplasia 2, 235-241. Cherukuri, D.P., Chen, X.B., Goulet, A.C., Young, R.N., Han, Y., Heimark, R.L., Regan, J.W., Meuillet, E., and Nelson, M.A. (2007). The EP4 receptor antagonist, L-161,982, blocks prostaglandin E2-induced signal transduction and cell proliferation in HCA-7 colon cancer cells. Exp Cell Res 313, 2969-2979. Chiou, S.K., and Mandayam, S. (2007). NSAIDs enhance proteasomic degradation of survivin, a mechanism of gastric epithelial cell injury and apoptosis. Biochem Pharmacol 74, 1485-1495. Dixon, D.A. (2004). Dysregulated post-transcriptional control of COX-2 gene expression in cancer. Curr Pharm Des 10, 635-646. Doller, A., Akool el, S., Huwiler, A., Muller, R., Radeke, H.H., Pfeilschifter, J., and Eberhardt, W. (2008). Posttranslational modification of the AU-rich element binding protein HuR by protein kinase Cdelta elicits angiotensin II-induced stabilization and nuclear export of cyclooxygenase 2 mRNA. Mol Cell Biol 28, 2608-2625. DuBois, R.N., Radhika, A., Reddy, B.S., and Entingh, A.J. (1996). Increased cyclooxygenase-2 levels in carcinogen-induced rat colonic tumors. Gastroenterology 110, 1259-1262. Eastman, Q., and Grosschedl, R. (1999). Regulation of LEF-1/TCF transcription factors by Wnt and other signals. Curr Opin Cell Biol 11, 233-240.

123
Einbond, A., and Sudol, M. (1996). Towards prediction of cognate complexes between the WW domain and proline-rich ligands. FEBS Lett 384, 1-8. Engelman, J.A., Wykoff, C.C., Yasuhara, S., Song, K.S., Okamoto, T., and Lisanti, M.P. (1997). Recombinant expression of caveolin-1 in oncogenically transformed cells abrogates anchorage-independent growth. J Biol Chem 272, 16374-16381. Ensor, C.M., and Tai, H.H. (1995). 15-Hydroxyprostaglandin dehydrogenase. J Lipid Mediat Cell Signal 12, 313-319. Fang, P.K., Solomon, K.R., Zhuang, L., Qi, M., McKee, M., Freeman, M.R., and Yelick, P.C. (2006). Caveolin-1alpha and -1beta perform nonredundant roles in early vertebrate development. Am J Pathol 169, 2209-2222. Felley-Bosco, E., Bender, F., and Quest, A.F. (2002). Caveolin-1-mediated post-transcriptional regulation of inducible nitric oxide synthase in human colon carcinoma cells. Biol Res 35, 169-176. Felley-Bosco, E., Bender, F.C., Courjault-Gautier, F., Bron, C., and Quest, A.F. (2000). Caveolin-1 down-regulates inducible nitric oxide synthase via the proteasome pathway in human colon carcinoma cells. Proc Natl Acad Sci U S A 97, 14334-14339. Fiucci, G., Ravid, D., Reich, R., and Liscovitch, M. (2002). Caveolin-1 inhibits anchorage-independent growth, anoikis and invasiveness in MCF-7 human breast cancer cells. Oncogene 21, 2365-2375. Furst, S.M., Komocsar, W.J., Khan, K.N., White, K.L., Jr., Peachee, V.L., and Mennear, J.H. (2005). Screening New Drugs for Immunotoxic Potential: I. Assessment of the Effects of Conventional Nonsteroidal Anti-Inflammatory Drugs and Selective COX-2 Inhibitors on In Vitro and In Vivo Phagocytic Activity. J Immunotoxicol 1, 149-158. Gaiser, T., Becker, M.R., Habel, A., Reuss, D.E., Ehemann, V., Rami, A., and Siegelin, M.D. (2008). TRAIL-mediated apoptosis in malignant glioma cells is augmented by celecoxib through proteasomal degradation of survivin. Neurosci Lett 442, 109-113. Galbiati, F., Volonte, D., Brown, A.M., Weinstein, D.E., Ben-Ze'ev, A., Pestell, R.G., and Lisanti, M.P. (2000). Caveolin-1 expression inhibits Wnt/beta-catenin/Lef-1 signaling by recruiting beta-catenin to caveolae membrane domains. J Biol Chem 275, 23368-23377. Galbiati, F., Volonte, D., Engelman, J.A., Watanabe, G., Burk, R., Pestell, R.G., and Lisanti, M.P. (1998). Targeted downregulation of caveolin-1 is sufficient to drive cell transformation and hyperactivate the p42/44 MAP kinase cascade. Embo J 17, 6633-6648. Garcia, S., Dales, J.P., Charafe-Jauffret, E., Carpentier-Meunier, S., Andrac-Meyer, L., Jacquemier, J., Andonian, C., Lavaut, M.N., Allasia, C., Bonnier, P., and Charpin, C. (2007). Poor prognosis in breast carcinomas correlates with increased expression of targetable CD146 and c-Met and with proteomic basal-like phenotype. Hum Pathol 38, 830-841. Gaudreault, S.B., Chabot, C., Gratton, J.P., and Poirier, J. (2004). The caveolin scaffolding domain modifies 2-amino-3-hydroxy-5-methyl-4-isoxazole propionate receptor binding properties by inhibiting phospholipase A2 activity. J Biol Chem 279, 356-362. Giles, R.H., van Es, J.H., and Clevers, H. (2003). Caught up in a Wnt storm: Wnt signaling in cancer. Biochim Biophys Acta 1653, 1-24. Graziani, A., Bricko, V., Carmignani, M., Graier, W.F., and Groschner, K. (2004). Cholesterol- and caveolin-rich membrane domains are essential for phospholipase A2-dependent EDHF formation. Cardiovasc Res 64, 234-242. Grossman, D., Kim, P.J., Schechner, J.S., and Altieri, D.C. (2001). Inhibition of melanoma tumor growth in vivo by survivin targeting. Proc Natl Acad Sci U S A 98, 635-640.

124
Gumbiner, B.M. (1995). Signal transduction of beta-catenin. Curr Opin Cell Biol 7, 634-640. Haertel-Wiesmann, M., Liang, Y., Fantl, W.J., and Williams, L.T. (2000). Regulation of cyclooxygenase-2 and periostin by Wnt-3 in mouse mammary epithelial cells. J Biol Chem 275, 32046-32051. Hanahan, D., and Weinberg, R.A. (2000). The hallmarks of cancer. Cell 100, 57-70. Hasegawa, K., Ichikawa, W., Fujita, T., Ohno, R., Okusa, T., Yoshinaga, K., and Sugihara, K. (2001). Expression of cyclooxygenase-2 (COX-2) mRNA in human colorectal adenomas. Eur J Cancer 37, 1469-1474. Hawcroft, G., Ko, C.W., and Hull, M.A. (2007). Prostaglandin E2-EP4 receptor signalling promotes tumorigenic behaviour of HT-29 human colorectal cancer cells. Oncogene 26, 3006-3019. He, B., Reguart, N., You, L., Mazieres, J., Xu, Z., Lee, A.Y., Mikami, I., McCormick, F., and Jablons, D.M. (2005). Blockade of Wnt-1 signaling induces apoptosis in human colorectal cancer cells containing downstream mutations. Oncogene 24, 3054-3058. Helguero, L.A., Lindberg, K., Gardmo, C., Schwend, T., Gustafsson, J.A., and Haldosen, L.A. (2008). Different roles of estrogen receptors alpha and beta in the regulation of E-cadherin protein levels in a mouse mammary epithelial cell line. Cancer Res 68, 8695-8704. Herendeen, J.M., and Lindley, C. (2003). Use of NSAIDs for the chemoprevention of colorectal cancer. Ann Pharmacother 37, 1664-1674. Hill, M.M., Bastiani, M., Luetterforst, R., Kirkham, M., Kirkham, A., Nixon, S.J., Walser, P., Abankwa, D., Oorschot, V.M., Martin, S., Hancock, J.F., and Parton, R.G. (2008). PTRF-Cavin, a conserved cytoplasmic protein required for caveola formation and function. Cell 132, 113-124. Hla, T., Bishop-Bailey, D., Liu, C.H., Schaefers, H.J., and Trifan, O.C. (1999). Cyclooxygenase-1 and -2 isoenzymes. Int J Biochem Cell Biol 31, 551-557. Holla, V.R., Backlund, M.G., Yang, P., Newman, R.A., and DuBois, R.N. (2008). Regulation of prostaglandin transporters in colorectal neoplasia. Cancer Prev Res (Phila Pa) 1, 93-99. Hoshino, T., Nakaya, T., Homan, T., Tanaka, K., Sugimoto, Y., Araki, W., Narita, M., Narumiya, S., Suzuki, T., and Mizushima, T. (2007). Involvement of prostaglandin E2 in production of amyloid-beta peptides both in vitro and in vivo. J Biol Chem 282, 32676-32688. Hsu, S.C., Galceran, J., and Grosschedl, R. (1998). Modulation of transcriptional regulation by LEF-1 in response to Wnt-1 signaling and association with beta-catenin. Mol Cell Biol 18, 4807-4818. Hulit, J., Bash, T., Fu, M., Galbiati, F., Albanese, C., Sage, D.R., Schlegel, A., Zhurinsky, J., Shtutman, M., Ben-Ze'ev, A., Lisanti, M.P., and Pestell, R.G. (2000). The cyclin D1 gene is transcriptionally repressed by caveolin-1. J Biol Chem 275, 21203-21209. Hull, M.A. (2005). Cyclooxygenase-2: how good is it as a target for cancer chemoprevention? Eur J Cancer 41, 1854-1863. Hull, M.A., Ko, S.C., and Hawcroft, G. (2004). Prostaglandin EP receptors: targets for treatment and prevention of colorectal cancer? Mol Cancer Ther 3, 1031-1039. Ilsley, J.L., Sudol, M., and Winder, S.J. (2002). The WW domain: linking cell signalling to the membrane cytoskeleton. Cell Signal 14, 183-189. Ilsley, J.N., Nakanishi, M., Flynn, C., Belinsky, G.S., De Guise, S., Adib, J.N., Dobrowsky, R.T., Bonventre, J.V., and Rosenberg, D.W. (2005). Cytoplasmic phospholipase A2 deletion enhances colon tumorigenesis. Cancer Res 65, 2636-2643.

125
Jendrossek, V., Handrick, R., and Belka, C. (2003). Celecoxib activates a novel mitochondrial apoptosis signaling pathway. Faseb J 17, 1547-1549. Kanaoka, S., Takai, T., and Yoshida, K. (2007). Cyclooxygenase-2 and tumor biology. Adv Clin Chem 43, 59-78. Kawamori, T., Rao, C.V., Seibert, K., and Reddy, B.S. (1998). Chemopreventive activity of celecoxib, a specific cyclooxygenase-2 inhibitor, against colon carcinogenesis. Cancer Res 58, 409-412. Kim, P.J., Plescia, J., Clevers, H., Fearon, E.R., and Altieri, D.C. (2003). Survivin and molecular pathogenesis of colorectal cancer. The Lancet 362, 205-209. Kogo, H., Aiba, T., and Fujimoto, T. (2004). Cell type-specific occurrence of caveolin-1alpha and -1beta in the lung caused by expression of distinct mRNAs. J Biol Chem 279, 25574-25581. Kogo, H., and Fujimoto, T. (2000). Caveolin-1 isoforms are encoded by distinct mRNAs. Identification Of mouse caveolin-1 mRNA variants caused by alternative transcription initiation and splicing. FEBS Lett 465, 119-123. Koleske, A.J., Baltimore, D., and Lisanti, M.P. (1995). Reduction of caveolin and caveolae in oncogenically transformed cells. Proc Natl Acad Sci U S A 92, 1381-1385. Krysan, K., Dalwadi, H., Sharma, S., Pold, M., and Dubinett, S. (2004a). Cyclooxygenase 2-dependent expression of survivin is critical for apoptosis resistance in non-small cell lung cancer. Cancer Res 64, 6359-6362. Krysan, K., Dohadwala, M., Luo, J., Lin, Y., Zhu, L., Heuze-Vourc'h, N., Goodglick, L., Merchant, F., Seligson, D., Pold, M., Strieter, R., Sharma, S., and Dubinett, S. (2004b). Cyclooxygenase-2-dependent expression of survivin in non-small cell lung cancer. Chest 125, 140S. Krysan, K., Merchant, F.H., Zhu, L., Dohadwala, M., Luo, J., Lin, Y., Heuze-Vourc'h, N., Pold, M., Seligson, D., Chia, D., Goodglick, L., Wang, H., Strieter, R., Sharma, S., and Dubinett, S. (2003). COX-2-dependent stabilization of survivin in non-small cell lung cancer. FASEB J, 03-0369fje. Krysan, K., Merchant, F.H., Zhu, L., Dohadwala, M., Luo, J., Lin, Y., Heuze-Vourc'h, N., Pold, M., Seligson, D., Chia, D., Goodglick, L., Wang, H., Strieter, R., Sharma, S., and Dubinett, S. (2004c). COX-2-dependent stabilization of survivin in non-small cell lung cancer. Faseb J 18, 206-208. Kutchera, W., Jones, D.A., Matsunami, N., Groden, J., McIntyre, T.M., Zimmerman, G.A., White, R.L., and Prescott, S.M. (1996). Prostaglandin H synthase 2 is expressed abnormally in human colon cancer: evidence for a transcriptional effect. Proc Natl Acad Sci U S A 93, 4816-4820. Lavie, Y., Fiucci, G., and Liscovitch, M. (1998). Up-regulation of caveolae and caveolar constituents in multidrug-resistant cancer cells. J Biol Chem 273, 32380-32383. Lee, S.W., Reimer, C.L., Oh, P., Campbell, D.B., and Schnitzer, J.E. (1998). Tumor cell growth inhibition by caveolin re-expression in human breast cancer cells. Oncogene 16, 1391-1397. Liao, Z., Mason, K.A., and Milas, L. (2007). Cyclo-oxygenase-2 and its inhibition in cancer: is there a role? Drugs 67, 821-845. Lim, K., Han, C., Xu, L., Isse, K., Demetris, A.J., and Wu, T. (2008). Cyclooxygenase-2-derived prostaglandin E2 activates beta-catenin in human cholangiocarcinoma cells: evidence for inhibition of these signaling pathways by omega 3 polyunsaturated fatty acids. Cancer Res 68, 553-560. Liou, J.Y., Deng, W.G., Gilroy, D.W., Shyue, S.K., and Wu, K.K. (2001). Colocalization and interaction of cyclooxygenase-2 with caveolin-1 in human fibroblasts. J Biol Chem 276, 34975-34982. Liu, P., Rudick, M., and Anderson, R.G. (2002). Multiple functions of caveolin-1. J Biol Chem 277, 41295-41298.

126
Liu, Y., Borchert, G.L., Surazynski, A., and Phang, J.M. (2008). Proline oxidase, a p53-induced gene, targets COX-2/PGE2 signaling to induce apoptosis and inhibit tumor growth in colorectal cancers. Oncogene 27, 6729-6737. Livak, H.J., and Schmittgen, T.D. (2001). Analysis of Relative Gene Expression Data Using Real- Time Quantitative PCR and the 2-DDCT Method. Methods 25, 402-408. Logan, C.Y., and Nusse, R. (2004). The Wnt signaling pathway in development and disease. Annu Rev Cell Dev Biol 20, 781-810. Lu, Z., Ghosh, S., Wang, Z., and Hunter, T. (2003). Downregulation of caveolin-1 function by EGF leads to the loss of E-cadherin, increased transcriptional activity of beta-catenin, and enhanced tumor cell invasion. Cancer Cell 4, 499-515. Mori, F., Piro, F.R., Della Rocca, C., Mesiti, G., Giampaoli, S., Silvestre, G., and Lazzaro, D. (2007). Survivin and Cyclooxygenase-2 are co-expressed in human and mouse colon carcinoma and in terminally differentiated colonocytes. Histol Histopathol 22, 61-77. Murata, M., Peranen, J., Schreiner, R., Wieland, F., Kurzchalia, T.V., and Simons, K. (1995). VIP21/caveolin is a cholesterol-binding protein. Proc Natl Acad Sci U S A 92, 10339-10343. Norvell, S.M., Alvarez, M., Bidwell, J.P., and Pavalko, F.M. (2004). Fluid shear stress induces beta-catenin signaling in osteoblasts. Calcif Tissue Int 75, 396-404. Okamoto, T., Schlegel, A., Scherer, P.E., and Lisanti, M.P. (1998). Caveolins, a family of scaffolding proteins for organizing "preassembled signaling complexes" at the plasma membrane. J Biol Chem 273, 5419-5422. Pai, R., Nakamura, T., Moon, W.S., and Tarnawski, A.S. (2003). Prostaglandins promote colon cancer cell invasion; signaling by cross-talk between two distinct growth factor receptors. Faseb J 17, 1640-1647. Park, J.Y., Pillinger, M.H., and Abramson, S.B. (2006). Prostaglandin E2 synthesis and secretion: the role of PGE2 synthases. Clin Immunol 119, 229-240. Pennati, M., Binda, M., De Cesare, M., Pratesi, G., Folini, M., Citti, L., Daidone, M.G., Zunino, F., and Zaffaroni, N. (2004). Ribozyme-mediated down-regulation of survivin expression sensitizes human melanoma cells to topotecan in vitro and in vivo. Carcinogenesis 25, 1129-1136. Perrone, G., Zagami, M., Altomare, V., Battista, C., Morini, S., and Rabitti, C. (2007). COX-2 localization within plasma membrane caveolae-like structures in human lobular intraepithelial neoplasia of the breast. Virchows Arch 451, 1039-1045. Piedra, J., Miravet, S., Castano, J., Palmer, H.G., Heisterkamp, N., Garcia de Herreros, A., and Dunach, M. (2003). p120 Catenin-associated Fer and Fyn tyrosine kinases regulate beta-catenin Tyr-142 phosphorylation and beta-catenin-alpha-catenin Interaction. Mol Cell Biol 23, 2287-2297. Pockaj, B.A., Basu, G.D., Pathangey, L.B., Gray, R.J., Hernandez, J.L., Gendler, S.J., and Mukherjee, P. (2004). Reduced T-cell and dendritic cell function is related to cyclooxygenase-2 overexpression and prostaglandin E2 secretion in patients with breast cancer. Ann Surg Oncol 11, 328-339. Ponder, B.A. (2001). Cancer genetics. Nature 411, 336-341. Potter, E., Bergwitz, C., and Brabant, G. (1999). The cadherin-catenin system: implications for growth and differentiation of endocrine tissues. Endocr Rev 20, 207-239. Quest, A., Gutierrez-Pajares, J.L., and Torres, V. (2008). Caveolin-1: an ambiguous partner in cell signaling and cancer. J Cell Mol Med 12, 1130 - 1150.

127
Quest, A.F., Leyton, L., and Parraga, M. (2004). Caveolins, caveolae, and lipid rafts in cellular transport, signaling, and disease. Biochem Cell Biol 82, 129-144. Rao, C.V., Indranie, C., Simi, B., Manning, P.T., Connor, J.R., and Reddy, B.S. (2002). Chemopreventive properties of a selective inducible nitric oxide synthase inhibitor in colon carcinogenesis, administered alone or in combination with celecoxib, a selective cyclooxygenase-2 inhibitor. Cancer Res 62, 165-170. Razani, B., Woodman, S.E., and Lisanti, M.P. (2002). Caveolae: from cell biology to animal physiology. Pharmacol Rev 54, 431-467. Reddy, B.S., Hirose, Y., Lubet, R., Steele, V., Kelloff, G., Paulson, S., Seibert, K., and Rao, C.V. (2000). Chemoprevention of colon cancer by specific cyclooxygenase-2 inhibitor, celecoxib, administered during different stages of carcinogenesis. Cancer Res 60, 293-297. Regan, J.W. (2003). EP2 and EP4 prostanoid receptor signaling. Life Sci 74, 143-153. Robertson, F.M., Simeone, A.M., Mazumdar, A., Shah, A.H., McMurray, J.S., Ghosh, S., and Cristofanilli, M. (2008). Molecular and pharmacological blockade of the EP4 receptor selectively inhibits both proliferation and invasion of human inflammatory breast cancer cells. J Exp Ther Oncol 7, 299-312. Rodriguez, D.A., Tapia, J.C., Fernandez, J.G., Torres, V.A., Munoz, N., Galleguillos, D., Leyton, L., and Quest, A.F. (2009). Caveolin-1-mediated Suppression of Cyclooxygenase-2 via a {beta}-catenin-Tcf/Lef-dependent Transcriptional Mechanism Reduced PGE2 Production and Survivin Expression. Mol Biol Cell 20, 2297-2310. Rothberg, K.G., Heuser, J.E., Donzell, W.C., Ying, Y.S., Glenney, J.R., and Anderson, R.G. (1992). Caveolin, a protein component of caveolae membrane coats. Cell 68, 673-682. Rubinfeld, B., Albert, I., Porfiri, E., Fiol, C., Munemitsu, S., and Polakis, P. (1996). Binding of GSK3beta to the APC-beta-catenin complex and regulation of complex assembly. Science 272, 1023-1026. Rubinfeld, B., Souza, B., Albert, I., Muller, O., Chamberlain, S.H., Masiarz, F.R., Munemitsu, S., and Polakis, P. (1993). Association of the APC gene product with beta-catenin. Science 262, 1731-1734. Salomon, D., Sacco, P.A., Roy, S.G., Simcha, I., Johnson, K.R., Wheelock, M.J., and Ben-Ze'ev, A. (1997). Regulation of beta-catenin levels and localization by overexpression of plakoglobin and inhibition of the ubiquitin-proteasome system. J Cell Biol 139, 1325-1335. Sano, H., Kawahito, Y., Wilder, R.L., Hashiramoto, A., Mukai, S., Asai, K., Kimura, S., Kato, H., Kondo, M., and Hla, T. (1995). Expression of cyclooxygenase-1 and -2 in human colorectal cancer. Cancer Res 55, 3785-3789. Scherer, P.E., Tang, Z., Chun, M., Sargiacomo, M., Lodish, H.F., and Lisanti, M.P. (1995). Caveolin isoforms differ in their N-terminal protein sequence and subcellular distribution. Identification and epitope mapping of an isoform-specific monoclonal antibody probe. J Biol Chem 270, 16395-16401. Schonthal, A.H., Chen, T.C., Hofman, F.M., Louie, S.G., and Petasis, N.A. (2008). Celecoxib analogs that lack COX-2 inhibitory function: preclinical development of novel anticancer drugs. Expert Opin Investig Drugs 17, 197-208. Schuster, V.L. (2002). Prostaglandin transport. Prostaglandins Other Lipid Mediat 68-69, 633-647. Serra-Pages, C., Medley, Q.G., Tang, M., Hart, A., and Streuli, M. (1998). Liprins, a family of LAR transmembrane protein-tyrosine phosphatase-interacting proteins. J Biol Chem 273, 15611-15620. Shao, J., Jung, C., Liu, C., and Sheng, H. (2005). Prostaglandin E2 Stimulates the beta-catenin/T cell factor-dependent transcription in colon cancer. J Biol Chem 280, 26565-26572.

128
Shao, J., Lee, S.B., Guo, H., Evers, B.M., and Sheng, H. (2003). Prostaglandin E2 stimulates the growth of colon cancer cells via induction of amphiregulin. Cancer Res 63, 5218-5223. Sheng, H., Shao, J., Morrow, J.D., Beauchamp, R.D., and DuBois, R.N. (1998). Modulation of apoptosis and Bcl-2 expression by prostaglandin E2 in human colon cancer cells. Cancer Res 58, 362-366. Sheng, H., Shao, J., Washington, M.K., and DuBois, R.N. (2001). Prostaglandin E2 increases growth and motility of colorectal carcinoma cells. J Biol Chem 276, 18075-18081. Shin, S., Sung, B.J., Cho, Y.S., Kim, H.J., Ha, N.C., Hwang, J.I., Chung, C.W., Jung, Y.K., and Oh, B.H. (2001). An anti-apoptotic protein human survivin is a direct inhibitor of caspase-3 and -7. Biochemistry 40, 1117-1123. Sinicrope, F.A., and Gill, S. (2004). Role of cyclooxygenase-2 in colorectal cancer. Cancer Metastasis Rev 23, 63-75. Smart, E.J., Graf, G.A., McNiven, M.A., Sessa, W.C., Engelman, J.A., Scherer, P.E., Okamoto, T., and Lisanti, M.P. (1999). Caveolins, liquid-ordered domains, and signal transduction. Mol Cell Biol 19, 7289-7304. Smith, W.L., DeWitt, D.L., and Garavito, R.M. (2000). Cyclooxygenases: structural, cellular, and molecular biology. Annu Rev Biochem 69, 145-182. Song, D.H., Sussman, D.J., and Seldin, D.C. (2000). Endogenous protein kinase CK2 participates in Wnt signaling in mammary epithelial cells. J Biol Chem 275, 23790-23797. Soslow, R.A., Dannenberg, A.J., Rush, D., Woerner, B.M., Khan, K.N., Masferrer, J., and Koki, A.T. (2000). COX-2 is expressed in human pulmonary, colonic, and mammary tumors. Cancer 89, 2637-2645. Sotgia, F., Lee, J.K., Das, K., Bedford, M., Petrucci, T.C., Macioce, P., Sargiacomo, M., Bricarelli, F.D., Minetti, C., Sudol, M., and Lisanti, M.P. (2000). Caveolin-3 directly interacts with the C-terminal tail of beta -dystroglycan. Identification of a central WW-like domain within caveolin family members. J Biol Chem 275, 38048-38058. Steinert, D., Kuper, C., Bartels, H., Beck, F.X., and Neuhofer, W. (2009). PGE2 potentiates tonicity-induced COX-2 expression in renal medullary cells in a positive feedback loop involving EP2-cAMP-PKA signaling. Am J Physiol Cell Physiol 296, C75-87. Sudol, M., Chen, H.I., Bougeret, C., Einbond, A., and Bork, P. (1995). Characterization of a novel protein-binding module--the WW domain. FEBS Lett 369, 67-71. Swamy, M.V., Cooma, I., Patlolla, J.M., Simi, B., Reddy, B.S., and Rao, C.V. (2004). Modulation of cyclooxygenase-2 activities by the combined action of celecoxib and decosahexaenoic acid: novel strategies for colon cancer prevention and treatment. Mol Cancer Ther 3, 215-221. Tapia, J.C., Torres, V.A., Rodriguez, D.A., Leyton, L., and Quest, A.F. (2006). Casein kinase 2 (CK2) increases survivin expression via enhanced beta-catenin-T cell factor/lymphoid enhancer binding factor-dependent transcription. Proc Natl Acad Sci U S A 103, 15079-15084. Telliez, A., Furman, C., Pommery, N., and Henichart, J.P. (2006). Mechanisms leading to COX-2 expression and COX-2 induced tumorigenesis: topical therapeutic strategies targeting COX-2 expression and activity. Anticancer Agents Med Chem 6, 187-208. Thiel, A., Heinonen, M., Rintahaka, J., Hallikainen, T., Hemmes, A., Dixon, D.A., Haglund, C., and Ristimaki, A. (2006). Expression of cyclooxygenase-2 is regulated by glycogen synthase kinase-3beta in gastric cancer cells. J Biol Chem 281, 4564-4569.

129
Tong, X., Yin, L., Joshi, S., Rosenberg, D.W., and Giardina, C. (2005). Cyclooxygenase-2 regulation in colon cancer cells: modulation of RNA polymerase II elongation by histone deacetylase inhibitors. J Biol Chem 280, 15503-15509. Torres, V.A., Tapia, J.C., Rodriguez, D.A., Lladser, A., Arredondo, C., Leyton, L., and Quest, A.F. (2007). E-Cadherin Is Required for Caveolin-1-Mediated Down-Regulation of the Inhibitor of Apoptosis Protein Survivin via Reduced {beta}-Catenin-Tcf/Lef-Dependent Transcription. Mol Cell Biol 27, 7703-7717. Torres, V.A., Tapia, J.C., Rodriguez, D.A., Parraga, M., Lisboa, P., Montoya, M., Leyton, L., and Quest, A.F. (2006). Caveolin-1 controls cell proliferation and cell death by suppressing expression of the inhibitor of apoptosis protein survivin. J Cell Sci 119, 1812-1823. Tsujii, M., and DuBois, R.N. (1995). Alterations in cellular adhesion and apoptosis in epithelial cells overexpressing prostaglandin endoperoxide synthase 2. Cell 83, 493-501. Tsujii, M., Kawano, S., and DuBois, R.N. (1997). Cyclooxygenase-2 expression in human colon cancer cells increases metastatic potential. Proc Natl Acad Sci U S A 94, 3336-3340. Tucker, O.N., Dannenberg, A.J., Yang, E.K., Zhang, F., Teng, L., Daly, J.M., Soslow, R.A., Masferrer, J.L., Woerner, B.M., Koki, A.T., and Fahey, T.J., 3rd. (1999). Cyclooxygenase-2 expression is up-regulated in human pancreatic cancer. Cancer Res 59, 987-990. Vaisar, T., Pennathur, S., Green, P.S., Gharib, S.A., Hoofnagle, A.N., Cheung, M.C., Byun, J., Vuletic, S., Kassim, S., Singh, P., Chea, H., Knopp, R.H., Brunzell, J., Geary, R., Chait, A., Zhao, X.Q., Elkon, K., Marcovina, S., Ridker, P., Oram, J.F., and Heinecke, J.W. (2007). Shotgun proteomics implicates protease inhibition and complement activation in the antiinflammatory properties of HDL. J Clin Invest 117, 746-756. van de Wetering, M., Cavallo, R., Dooijes, D., van Beest, M., van Es, J., Loureiro, J., Ypma, A., Hursh, D., Jones, T., Bejsovec, A., Peifer, M., Mortin, M., and Clevers, H. (1997). Armadillo coactivates transcription driven by the product of the Drosophila segment polarity gene dTCF. Cell 88, 789-799. van Deurs, B., Roepstorff, K., Hommelgaard, A.M., and Sandvig, K. (2003). Caveolae: anchored, multifunctional platforms in the lipid ocean. Trends Cell Biol 13, 92-100. Velculescu, V.E., Madden, S.L., Zhang, L., Lash, A.E., Yu, J., Rago, C., Lal, A., Wang, C.J., Beaudry, G.A., Ciriello, K.M., Cook, B.P., Dufault, M.R., Ferguson, A.T., Gao, Y., He, T.C., Hermeking, H., Hiraldo, S.K., Hwang, P.M., Lopez, M.A., Luderer, H.F., Mathews, B., Petroziello, J.M., Polyak, K., Zawel, L., Kinzler, K.W., and et al. (1999). Analysis of human transcriptomes. Nat Genet 23, 387-388. Verhagen, A.M., Coulson, E.J., and Vaux, D.L. (2001). Inhibitor of apoptosis proteins and their relatives: IAPs and other BIRPs. Genome Biol 2, REVIEWS3009. Vong, Q.P., Cao, K., Li, H.Y., Iglesias, P.A., and Zheng, Y. (2005). Chromosome alignment and segregation regulated by ubiquitination of survivin. Science 310, 1499-1504. Wang, M.T., Honn, K.V., and Nie, D. (2007). Cyclooxygenases, prostanoids, and tumor progression. Cancer Metastasis Rev 26, 525-534. Warner, T.D., and Mitchell, J.A. (2004). Cyclooxygenases: new forms, new inhibitors, and lessons from the clinic. Faseb J 18, 790-804. Waterman, M.L. (2004). Lymphoid enhancer factor/T cell factor expression in colorectal cancer. Cancer Metastasis Rev 23, 41-52.

130
Wikman, H., Kettunen, E., Seppanen, J.K., Karjalainen, A., Hollmen, J., Anttila, S., and Knuutila, S. (2002). Identification of differentially expressed genes in pulmonary adenocarcinoma by using cDNA array. Oncogene 21, 5804-5813. Williams, T.M., and Lisanti, M.P. (2005). Caveolin-1 in oncogenic transformation, cancer, and metastasis. Am J Physiol Cell Physiol 288, C494-506. Xiao, Y., Araldi, G.L., Zhao, Z., Brugger, N., Karra, S., Fischer, D., and Palmer, E. (2007). Discovery of novel prostaglandin analogs of PGE2 as potent and selective EP2 and EP4 receptor agonists. Bioorg Med Chem Lett 17, 4323-4327. Yamada, E. (1955). The fine structure of the gall bladder epithelium of the mouse. J Biophys Biochem Cytol 1, 445-458. Yan, H.X., Yang, W., Zhang, R., Chen, L., Tang, L., Zhai, B., Liu, S.Q., Cao, H.F., Man, X.B., Wu, H.P., Wu, M.C., and Wang, H.Y. (2006). Protein-tyrosine phosphatase PCP-2 inhibits beta-catenin signaling and increases E-cadherin-dependent cell adhesion. J Biol Chem 281, 15423-15433. Zhang, T., Otevrel, T., Gao, Z., Gao, Z., Ehrlich, S.M., Fields, J.Z., and Boman, B.M. (2001). Evidence that APC regulates survivin expression: a possible mechanism contributing to the stem cell origin of colon cancer. Cancer Res 61, 8664-8667. Zhao, J., Tenev, T., Martins, L.M., Downward, J., and Lemoine, N.R. (2000). The ubiquitin-proteasome pathway regulates survivin degradation in a cell cycle-dependent manner. J Cell Sci 113 Pt 23, 4363-4371.