copertina suppl. sv804 copertina suppl....
TRANSCRIPT

Supp
lem
ento
LA SETTIMANAeterinariaVsettimanale d’informazione professionale per il veterinario • supplemento al n. 804 - settimana dal 25 al 31 ottobre 2012
Poste Italiane Spa - Spedizione in abbonamento postale - D.L. 353/2003 (conv. In L. 27/02/2004 n. 46) art. 1, comma 1, DCB Brescia - In caso di mancato recapito ritornare al mittente che si impegna a pagare la corrispondente tassa DCB - Brescia
Co
ntie
ne I
.P.
Copertina suppl. SV804_Copertina suppl. SV804 31/10/12 11.13 Pagina 1

Sommario
Il complessodell’infertilitàestiva nellascrofa
22
Patologie ovariche-uterine e gravidanza
13
Metabolismo e fertilità nellavacca da latte
2
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 20121
EditoreLe Point Vétérinaire Italie s.r.l.Edizioni Veterinarie e AgrozootecnicheVia Medardo Rosso, 11 - 20159 MilanoTel. 02 608 52 31 r.a.; fax 02 668 28 66
■ Direttore responsabile:Gabriele Lanzarotti
■ Redazione (e-mail: [email protected]):
Maria Rosa Cama(Tel. 02 60 85 23 41, e-mail: [email protected])Monica Vajna de Pava(Tel. 02 60 85 23 29, e-mail: [email protected])
■ A cura diChiara Avanzini, Alessandro Fantini, Claudio Mazzoni,Chiara Piancastelli e Valeria Raffi
■ Grafica:Patrizia Zagni
■ Pubblicità (Tel. 02 60 85 23 20)e-mail: [email protected]: Riccardo Cini (responsabile)
■ Assistente pubblicità (Tel. 02 60 85 23 20)e-mail: [email protected]: Lucia Medas (annunci economici)
■ AbbonamentiGestione e rinnovo abbonamenti:Donatella Sgaria - Tel. 02 60 85 23 32 e-mail: [email protected]
Registrazione Tribunale di Milanon. 541 del 27-11-1993Iscr. Reg. Naz. della Stampan. 4820 del 16/02/1995
Stampa:A.G.F. Italia - Peschiera Borromeo (Mi)
Associazione Nazionale Editoria Specializzata
CONFINDUSTRIA
Questa rivista le è stata inviata tramite abbonamento.L’informativa sul trattamento dei dati personali è consultabile
sul sito www.pointvet.it
LA SETTIMANAeterinariaV
Fertilità e patologie in bovini e suini
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 1

l declino della fertilità della Frisona, alle-vata in ogni parte del mondo, è ormai sot-to gli occhi di tutti e abbondantemente do-cumentato dai dati pubblicati negli ultimianni. Che ci sia una correlazione fenotipica
tra la “sindrome della sub-fertilità nella vacca dalatte” (SSFVL) e l’incremento produttivo di que-sta razza è altrettanto evidente, anche se l’osser-vazione delle performance produttive delle stalleal top della produzione o delle singole bovine spes-so dimostrano il contrario. Molti sono i quesitiche, in questi anni, sia la comunità scientifica sia itecnici si sono posti. È sufficiente inserire negli indici genetici, attraver-so i quali si selezionano le razze da latte, alcuni ca-ratteri funzionali che direttamente o indirettamentepossano ridurre l’interparto e quindi accorciarel’intervallo parto-concepimento? In animali sele-zionati a produrre più latte possibile con percen-tuali e chilogrammi di proteina e grasso semprepiù elevati, quale aspetto del metabolismo è statoalterato per rendere sempre potenzialmente menofertili le Frisone? Prendendo atto della necessitàeconomica della vacca da latte di produrre semprepiù latte e caseina, le tecniche d’allevamento, nu-trizione e gestione sanitaria sono ancora adatte orappresentano il vero fattore limitante? A tuttequeste domande probabilmente non esiste unarisposta univoca.
◗ La genetica In molti Paesi, e già da tempo, troviamo tra gli in-dici genetici anche i cosiddetti caratteri funziona-li come la fertilità. Li troviamo nel TPI statunitense,nell’LPI canadese e comunque in tutti i Paesi delVecchio Continente dove sono maggiormente pre-senti le Frisone da latte. Il peso della fertilità in al-cuni indici è rilevante, anche se i caratteri ripro-duttivi hanno in genere una bassa, se non bassis-sima, ereditabilità. In Italia, attualmente, il PFTnon prevede la selezione di caratteri riferibili al-l’attività riproduttiva, anche se nel nuovo indicequesto verrà a breve previsto. Ad aggravare la si-tuazione è stato il fatto che prima dell’adozionedel PFT in Italia l’indice di selezione precedente,ILQM, prevedeva solo la selezione per caratteriproduttivi e morfologici. Questa particolarità, percerti versi, ha distratto la revisione delle tecniched’allevamento, nutrizione e gestione sanitaria at-tribuendo i fallimenti riproduttivi a una geneticanon favorevole e all’eccessiva voglia di produrre
degli allevatori italiani. Produrre molto latte, allaconcentrazione di proteina e grasso più alta pos-sibile, è una condizione ineludibile per animalida reddito. Incrementi nelle dinamiche dei costi edel tenore di vita della popolazione rendono nonpraticabili ripensamenti riproduttivi verso il bas-so. Vista la situazione, è quindi necessario com-prendere come la selezione operata sulla vacca dalatte abbia modificato gli assetti ormonali e meta-bolici e dove essi agiscono e interferiscono su unottimale andamento riproduttivo. La domestica-zione dei ruminanti da latte è avvenuta diverse mi-gliaia di anni fa e contestualmente a questo è ini-ziato il processo di selezione artificiale. È facile pen-sare che già allora venissero premiati gli animaliche producevano più latte, allevandone le figlie,oppure ricavando da animali particolarmente pre-giati i maschi da riproduzione. Questo tipo di se-lezione, definito “massale”, è ancora in uso nei pic-coli ruminanti e comunque in molte specie mino-ri. Gli animali, in natura, in genere si riproduco-no, ossia diventano fertili, nei periodi dell’anno chepotranno garantire quella sufficiente quantità dicibo in grado di assicurare alla madre un’adegua-ta produzione di latte, così da poter accompagna-re il prodotto del concepimento fino allo svezza-mento. Questa programmazione delle nascite pre-vede pertanto quel tipico andamento dei cicli estra-li definito mono-estrale o poli-estrale stagionale,ossia il sincronizzare l’attività riproduttiva con lemaggiori disponibilità di cibo per la madre e quin-di per il nascituro. Essendosi tutti gli animali, uo-mo compreso, selezionati in condizioni di quasipermanente carenza di cibo, o meglio, di quasi maiillimitata disponibilità di esso per 365 giorni al-l’anno, i mammiferi hanno sviluppato, per sele-zione naturale, dei meccanismi di gestione delle li-mitate risorse alimentari molto sofisticati. La se-lezione naturale non ha mai dovuto risolvere il pro-blema di una disponibilità illimitata di cibo per pe-riodi molto lunghi. L’uomo primitivo che alleva-va ruminanti da latte aveva la necessità, per il pro-prio sostentamento, di disporre di latte possibil-mente tutti i giorni e in quantità costante, anchese le tecniche di conservazione del latte si sono evo-lute di pari passo con la domesticazione degli ani-mali. Per avere una produzione di latte costantenell’anno era necessario premiare, selezionandoli,quegli animali che avevano cicli estrali meno con-centrati ma più diluiti e che quindi potessero par-torire in ogni periodo dell’anno e produrre per-tanto latte più costantemente. Già allora, ma ben
Metabolismo e fertilitànella vacca da latte
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 20122
Alessandro FantiniFacoltà di Medicina veterinaria– Università di Perugia I
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 2

descritte da Darwin nel suo “L’origine della spe-cie”, si era osservato che i ruminanti da latte co-stantemente alimentati con cibo di alto valore nu-tritivo risultavano più fertili e meno condizionatidalla stagione. In particolare nella vacca da latte,in un momento di difficile datazione, si è arrivatiad avere animali non più poli-estrali stagionali ben-sì annuali, ossia con un susseguirsi regolare, ogni21 giorni, di cicli estrali potenzialmente fertili. Per-tanto, in un momento dato, è nata la vacca da lat-te moderna, ossia un animale fertile tutto l’anno eche quindi può partorire regolarmente e, in virtùdella selezione artificiale, sempre più produttivo.Le allora modeste produzioni di latte erano suffi-cientemente soddisfatte da alimentazioni semplicibasate essenzialmente sui foraggi, verdi o essicca-ti che fossero. L’implacabile e inevitabile selezio-ne artificiale, ossia la programmazione degli ac-coppiamenti, rendeva gli animali sempre più pro-duttivi aumentandone costantemente i fabbisogninutritivi. Il passaggio dalla selezione artificiale“massale” a tecniche più sofisticate, come l’ani-mal model, ha esasperato i fabbisogni nutritivi de-gli animali rendendo sempre meno sufficienti i nu-trienti nella prima fase di lattazione per produrrelatte di qualità e avviare una corretta attività ri-produttiva. Già agli albori della zootecnia era no-to che il ruminante da latte produce di più quan-do è “fresco”, cioè tanto più è vicino al parto, aven-do il picco di lattazione tra il secondo e terzo me-se di lattazione. Un allevamento di bovine da lat-te sarà tanto più produttivo tanto più mungerà ani-mali “ freschi”. Per realizzare questa condizioneideale in allevamento gli animali devono rimane-re gravidi più presto possibile per tenere la curvadi lattazione più alta possibile. La curva di latta-zione ha un andamento tipico nella vacca da lat-te, ma anche negli altri ruminanti da latte, ed è sta-ta ben descritta da Wood nel 1967.La motivazione, ricavabile dalla biologia evoluti-va, della forma e dell’andamento di questa curva èlegata alla necessità di fornire a un nascituro, so-stanzialmente monogastrico, nutrimento e idrata-zione attraverso il latte almeno per i primi mesi divita. Per incoraggiare il vitello a interessarsi adalimenti diversi dal latte materno e permettere allamadre di disporre di risorse nutritive per avviareuna nuova gravidanza, la natura ha previsto unaprogressiva riduzione di latte dal picco produtti-vo in avanti per poi scendere più o meno rapida-mente, una volta instaurata la nuova gravidanza.Questo per dirottare le sempre scarse risorse nu-tritive disponibili per la lattazione alla gravidanza.Questi principi di fisiologia animale possono, dal-la selezione artificiale, essere modulati ma mai stra-volti, a meno che si possa ricorrere alle tecniche del-la bio-ingegneria. Questo concetto, seppur banale,deve essere ben compreso se si vuole affrontare inmaniera sostanziale la sub-fertilità della vacca dalatte. Da questa introduzione sono scaturite duechiavi di lettura importanti. La prima è che nellaprima fase di lattazione, in virtù della selezione ge-
netica, la produzione di latte è superiore alla quan-tità di nutrienti che la bovina è in grado di ingeri-re. Espressione palese di questa condizione fisiolo-gica è l’inevitabile dimagrimento nei primi due me-si di lattazione. Altra considerazione è che per pre-cise esigenze di redditività, e solo di essa, la bovinadovrebbe rimanere gravida quando massima è lasua produzione di latte o appena dopo, condizio-ne fisiologica spesso di difficile realizzazione. Piùalto è il potenziale genetico degli animali, mag-giore sarà la difficoltà di far riprodurre le bovineper avere un interparto entro l’anno. A confonde-re le idee, a far cadere paradigmi a volte molto su-perficiali e a dare grandi speranze ai genetisti, ai ve-terinari e agli zootecnici, sono i dati riproduttivi deisingoli soggetti e di alcuni allevamenti di vacche dalatte la cui produttività e potenziale genetico nonsono affatto correlate negativamente con le per-formance riproduttive.
◗ I nuovi assetti ormonali e metabolici
La domanda fondamentale da porsi, la cui rispo-sta può dare le chiavi di lettura per ispirare la mo-difica dei paradigmi zootecnici, veterinari e ma-nageriali, evidentemente non più adatti a gestire lemoderne bovine d’allevamento, è: cosa ha modi-ficato la selezione genetica nelle bovine per ren-derle così produttive? Nel 1980, Bauman e Curriedel Dipartimento di Animal science della CornellUniversity enfatizzarono fortemente il concetto dicome sia controllato il metabolismo durante la lat-tazione e la gravidanza ribadendo come esistanodue tipi di regolazione: una omeostatica e unaomeoressica [1]. Per regolazione omeostatica si in-tende il mantenimento dell’equilibrio in condizio-ne ambientali interne costanti. Per regolazioneomeoressica si intende, invece, la variazione or-chestrata dalle priorità degli stati fisiologici dei va-ri tessuti. La ripartizione dei nutrienti durante lagravidanza e la lattazione è un tipico esempio diregolazione omeoressica del metabolismo. A gui-dare questa ripartizione dei nutrienti e soprattut-to a dare le priorità alle varie funzioni metaboli-che intervengono gli ormoni. In animali di alto po-tenziale genetico la selezione rende alcuni caratte-ri produttivi come la produzione di latte, grassoe proteina sempre più prioritari rispetto ad altriseppur importanti funzioni metaboliche. La sele-zione genetica amplifica, per così dire, l’attitudinematerna della bovina, dotandola di una capacitàpotenziale di produrre latte molto elevata e ren-dendo questa funzione metabolica sempre più prio-ritaria rispetto alle altre. La selezione genetica, per-tanto, per produrre più latte agisce modificando,a vantaggio dell’allevatore, l’assetto ormonale de-gli animali. Durante il primo terzo di lattazione laproduzione di latte delle bovine supera, spesso digran lunga, la possibilità di ingerire quella cor-retta quantità di nutrienti che impedisce loro di an-dare in deficit energetico e proteico. Limiti fisio-
Fertilità e patologie in bovini e suini
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 20124
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 4

logici nella capacità di ingestione o nell’eccessivaconcentrazione d’energia e proteine non consen-tono di evitare questo status metabolico. Perchésuccede questo? Appunto perché la selezione ge-netica amplifica una caratteristica già a suo tem-po fissata dalla selezione naturale, di per sé moltopiù lenta, ma più efficace. Esistono nell’organismodei mammiferi funzioni vitali essenziali, ossia chesi riducono solo quando i meccanismi omeostati-ci sono “saltati”, riducibili, ossia modulabili in fun-zione della disponibilità di nutrienti e anche dialtri fattori ambientali, e sospendibili, ossia atti-vabili solo quando si ha una piena disponibilità dinutrienti sia sul breve che sul lungo periodo. Ve-diamone le principali. Si ritiene, oggi, che siano essenziali per la vacca dalatte: il metabolismo basale, l’attività neurale, lacircolazione sanguigna e la produzione di latte del-la bovina non gravida. Riducibili sono invece: ter-moregolazione, locomozione e crescita. Sono poi considerate non essenziali e quindi ese-guite in piena disponibilità di nutrienti e più in ge-nerale di condizioni ambientali favorevoli, la ri-produzione e lo stoccaggio energetico di grasso,glicogeno e proteine labili. L’essenzialità delle va-rie funzioni metaboliche e l’ordine con il quale inutrienti vengono ripartiti è di fatto l’essenza stes-sa della selezione genetica artificiale. Nel noto mo-dello nutrizionale CNCPS, utilizzato per studiaree applicare la nutrizione della vacca da latte ormaisu buona parte della Terra, nella sezione di calco-lo dei fabbisogni nutritivi essi vengono ripartiti inmantenimento, gravidanza, lattazione, crescita eriserve corporee. Il criterio di ripartizione dei nu-trienti varia in funzione dello stato fisiologico prin-cipale in cui si trova l’animale: gravidanza o lat-tazione. In un futuro, speriamo non troppo lon-tano, il CNCPS potrebbe stabilire il livello dellepriorità in funzione del livello genetico degli ani-mali. Nella fase di lattazione, e specificatamentenel primo terzo, dove la gravidanza non si è an-cora instaurata, la produzione di latte degli ani-mali di alto potenziale genetico sani è in generemolto elevata e prescinde dall’apporto nutritivo,in quanto conseguita a scapito delle riserve ener-getiche e proteiche accumulate durante il ciclo pro-duttivo precedente. Un adeguato apporto nutriti-vo ha la sola finalità di mitigare gli effetti negatividel deficit energetico e proteico e funge da fatto-re preventivo di alcune patologie metaboliche co-me la lipidosi epatica e la chetosi. Durante la gra-vidanza, che per buona parte del tempo si so-vrappone a una lattazione in corso di riduzione,funzioni come il ripristino delle riserve corporeeritornano a essere prioritarie perché legate alla pos-sibilità di produrre più latte possibile e per la mag-gior parte del tempo nella lattazione successiva.Questa regolazione omeoressica, continuamentemodulata dalla selezione genetica, ha come baseun pool di ormoni che ne rappresentano i veri at-tori. Molti sono gli ormoni coinvolti. Nel 1988,Bonczek e altri ricercatori dell’Università del Mis-
souri hanno eseguito un interessante esperimen-to che aveva la finalità di determinare come or-moni e metaboliti variano nel sangue in virtù del-la selezione genetica. Le bovine sono state suddi-vise in due gruppi e uno di essi è stato fecondatocon tori del 1964 (LMG) e l’altro con tori più re-centi (HMG). La produzione delle figlie di questidue gruppi è risultata pari, rispettivamente (305giorni), a 7.401 e 9.878 kg. Nel gruppo degli ani-mali LMG il livello di insulina e somatotropinamedio registrato dal 42° al 161° giorno dopo ilparto è stato, rispettivamente, di 22,5 µU/ml e 3,7ng/ml con un rapporto di 6,1. Nel gruppo HMG,invece, si è registrato un livello di insulina di 18,8µU/ml e di somatotropina di 4,5 ng/ml con un rap-porto pari a 4,2 [2]. Lavori successivi a quelli diBonczek, tuttavia piuttosto esigui, hanno via viaconfermato cosa significa a livello ormonale unabovina HMG. In un lavoro successivo si è evi-denziato che l’assetto ormonale caratteristico del-le bovine a più alto potenziale genetico sia tale,in lattazione che in asciutta, con una netta diffe-renza tra concentrazione di somatotropina e in-sulina. La differenze nella concentrazione ematicadi prolattina e tiroxina non sembrerebbero così si-gnificative. Differenze significative sono invece sta-te riscontrate nei NEFA e nel BHBA. Le bovine adalto potenziale genetico, probabilmente in virtù diuna concentrazione ematica di insulina tenden-zialmente più bassa, hanno una maggiore necessi-tà di ricorrere alle riserve lipidiche (NEFA), sotto-ponendo il fegato a un’intensa attività di esterifi-cazione e beta-ossidazione che però tende ad ar-restarsi e produrre più corpi chetonici (BHBA) del-le bovine a basso potenziale genetico. In un suc-cessivo lavoro di Gong e Webb (vedere figura 1a pag. 6) si è riscontrato il medesimo andamentodell’insulina, della somatotropina e dei BHBA [3].In un lavoro di Weber del 2007 dal titolo “Effet-ti della selezione genetica sulla produzione di so-matotropina (ST), IGF-1 e lattogeno placentare”si sono approfonditi questi aspetti. L’autore del-l’esperimento ha suddiviso gli animali per poten-ziale genetico, per numero di parti e per fase fi-siologica, lattazione o gestazione. Gli animali inlattazione, sia le primipare sia le pluripare, hannopresentato e confermato i dati di altri autori per lebovine HMG, cioè un incremento del livello ema-tico di somatotropina di circa il 40%, una ridu-zione di circa il 10% di IGF-1 e un significativo in-cremento del livello plasmatico dei NEFA [4].Analoga situazione è stata riscontrata per l’ST nel-le bovine in gestazione. Le variazioni di lattoge-no placentare (PL) non sono state ritenute signi-ficative. Esaminando l’andamento della ST e del-l’IGF-1 si è notato che nei soggetti HMG la ST au-menta e rimane elevata dal parto in avanti. Nellebovine ad alto potenziale genetico l’IGF-1 è ten-denzialmente più bassa nei primi 3 mesi dopo ilparto. Non direttamente legati alla ripartizione deimetaboliti, ma comunque coinvolti nelle modifi-cazioni ormonali direttamente o indirettamente in-
Metabolismo e fertilità nella bovina da latte
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 20125
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 5

dotte dalla selezione genetica, sono i lavori di Veer-kamp del 2003 e quello di Wiltbank del 2006.Veerkamp verificò il livello di progesterone ema-tico su latte prelevato due volte a settimana tra il35° giorno di lattazione e la fecondazione di 673primipare ad alto e basso potenziale genetico [5].Le osservazioni di Veerkamp conducono al fattoche il livello di progesterone delle primipare dialto e basso potenziale genetico erano, prima del1994, significativamente più alte delle bovine ana-loghe dopo il 1994. Wiltbank (vedere figura 2) in-dividua una serie di meccanismi fisiologici a spie-gazione della sub-fertilità della vacca da latte, di-versi ma comunque complementari ai dati sin quimostrati. Le bovine HMG in virtù della loro ele-vata capacità di ingestione presentano un accre-sciuto flusso di sangue su tutto l’apparato dige-rente e soprattutto a livello epatico. Questo de-termina un più elevato metabolismo degli estro-geni e del progesterone, riducendone la disponi-bilità ematica con una conseguente riduzione deltasso di concepimento e del comportamento estra-le e un aumento delle ovulazioni multiple e delleperdite embrionali [6].
◗ L’azione degli ormoni I pochi ma significativi esperimenti inequivocabil-mente riconducono gli effetti della selezione gene-tica a un aumento della concentrazione ematica diormone somatotropo, a una riduzione dell’insuli-na e dell’IGF-system e a una sostanziale stabilitàdegli altri ormoni, con la sola eccezione del pro-gesterone. Lucy, in una comunicazione del 2006(vedere figura 3) apparsa su Cattle Pratice fa al-
cune considerazioni provocatorie constatando al-cune analogie tra l’assetto ormonale e metabolicodella vacca nel post partum e il diabete umano ti-po 1 (da carenza di insulina) e tipo 2 da insulino-resistenza [7], individuate da Bereket et al., nel 1999[8], e da Radcliff et al., nel 2003 [9, 10]. L’insuli-na plasmatica risulta bassa nel diabete tipo 1 e nel-la vacca nel post partum. La sensibilità all’insulinarisulta bassa sia nelle due forme di diabete che nel-la bovina nel post partum. Il glucosio ematico ri-sulta basso nella bovina dopo il parto e alto nelledue forme di diabete. Il GH ematico è elevato neldiabete tipo uno e nella vacca post partum, men-tre l’IGF-1 e i recettori epatici del GH (GHR) ri-sultano bassi sia nel diabete da carenza di insuli-na che nella vacca dopo il parto. La selezione ge-netica per la produttività premia, e quindi agevo-la nella riproduzione, le bovine che da un lato ri-escono a ingerire più nutrienti possibile e nelle qua-li la mammella, organo insensibile all’azione del-l’insulina, capta la maggiore quantità possibile diglucosio, per trasformarlo in lattosio e quindi inlatte. Esasperando il concetto, per ragioni di mi-glior comprensione, la vacca ad alto potenziale ge-netico è quel soggetto la cui mammella è più “pre-potente” di quella di bovine con minor potenzialegenetico nella captazione dei nutrienti ematici. Du-rante il continuo processo di selezione artificiale,premiando il GH o la somatotropina, si sono inqualche modo favoriti animali con un livello di in-sulina più basso, creando quel quadro ormonalee metabolico tipico del paziente diabetico o predi-sposto ad esso. GH elevato, insulina bassa, ten-denza spiccata all’inizio della lattazione alla lipo-mobilizzazione e alla chetosi sono caratteristiche
Fertilità e patologie in bovini e suini
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 20126
Figura 1.
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 6
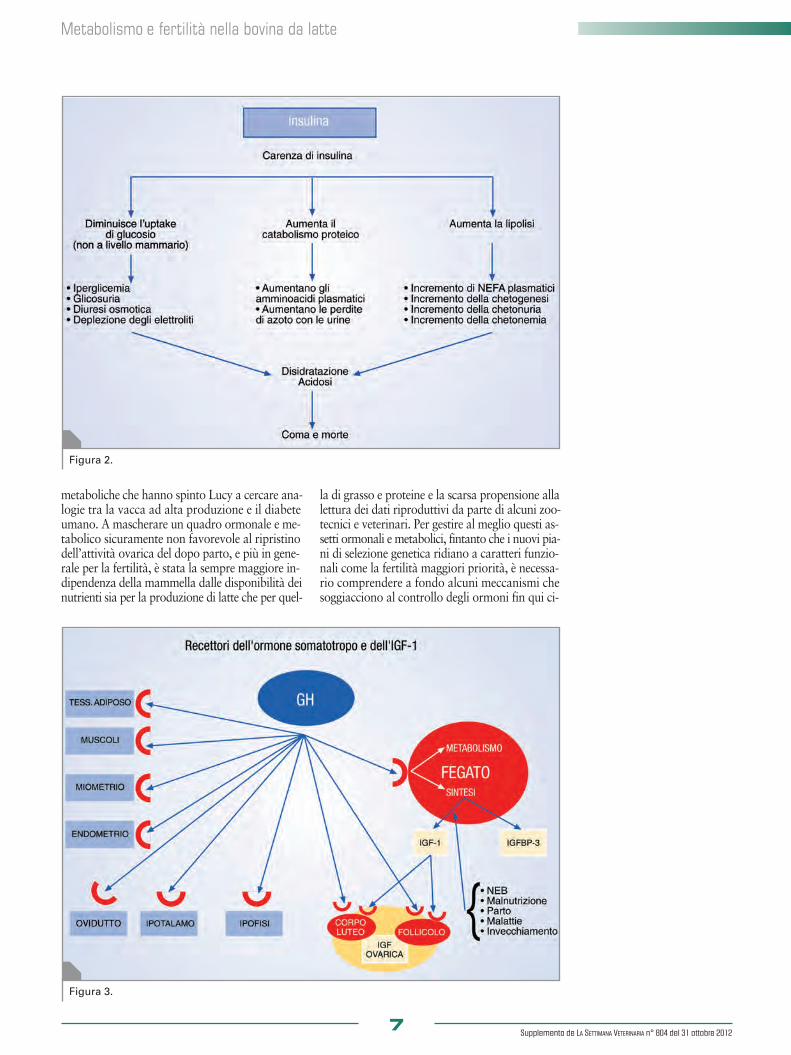
metaboliche che hanno spinto Lucy a cercare ana-logie tra la vacca ad alta produzione e il diabeteumano. A mascherare un quadro ormonale e me-tabolico sicuramente non favorevole al ripristinodell’attività ovarica del dopo parto, e più in gene-rale per la fertilità, è stata la sempre maggiore in-dipendenza della mammella dalle disponibilità deinutrienti sia per la produzione di latte che per quel-
la di grasso e proteine e la scarsa propensione allalettura dei dati riproduttivi da parte di alcuni zoo-tecnici e veterinari. Per gestire al meglio questi as-setti ormonali e metabolici, fintanto che i nuovi pia-ni di selezione genetica ridiano a caratteri funzio-nali come la fertilità maggiori priorità, è necessa-rio comprendere a fondo alcuni meccanismi chesoggiacciono al controllo degli ormoni fin qui ci-
Metabolismo e fertilità nella bovina da latte
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 20127
Figura 2.
Figura 3.
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 7

tati. Il “team” ormonale che direttamente e indi-rettamente è coinvolto nella fertilità della vacca dalatte è estremamente complesso e non definitiva-mente chiarito. I momenti fondamentali sono rap-presentati dalla ripresa dell’attività ovarica dopo ilparto, dalla crescita delle coorti follicolari e dal-l’emersione del follicolo dominante, dalla deiscen-za del follicolo di Graff, dalla possibilità di so-pravvivenza dell’embrione prima dell’adesione al-la mucosa uterina e dalle fasi successive dello svi-luppo embrionale. Esiste una forte connessione trail metabolismo dell’animale e l’attività riprodutti-va. La bovina da latte deve riprodursi, ossia in-staurare una nuova gravidanza, quando il bilancioenergetico e proteico è ancora negativo per cui labovina nel prendere “ la decisione di riprodursi”ha la necessità di effettuare una ricognizione a bre-ve, medio e lungo termine del suo stato metaboli-co per vedere se ci sono le condizioni per consen-tire il ripristino di una gravidanza, di portarla a ter-
mine e, una volta nato il vitello, di poter assicura-re quella produzione di latte necessaria al sosten-tamento prima dello svezzamento. Per poter ef-fettuare questa complessa “ricognizione metaboli-ca” viene coinvolto l’insieme del complesso ormo-nale. Tra gli ormoni maggiormente coinvolti tro-viamo l’insulina (vedere figura 4). L’insulina è clas-sificata come ormone metabolico e più specifica-tamente anabolico, ossia coinvolto nella gestionedello stoccaggio e della sintesi di alcune molecoleorganiche. L’insulina è un ormone proteico sinte-tizzato dalle cellule beta delle isole di Langerhansdel pancreas. Il suo ruolo fondamentale è quello dilegarsi al recettore esterno della membrana cellu-lare favorendo la penetrazione del glucosio nei tes-suti dove questi recettori sono presenti. Una voltache il glucosio è penetrato nelle cellule viene fosfo-rilato a glucosio-6-fosfato dalla glucochinasi e quin-di in esse stoccato per essere utilizzato per la pro-duzione di energia (ATP). Il suo diretto antagoni-sta è il glucagone definito ormone catabolico, pro-dotto dalle cellule alfa delle isole di Langerhans.L’insulina e il glucagone sono coinvolti direttamentenel sistema che garantisce l’omeostasi del glucosio,ossia la sua concentrazione costante nel sangue. Se-condo le indicazioni contenute in “Clinical bio-chemistry of domestic animals” di Kaneko et al.,tale valore normale è compreso tra 2,50 e 4,16mmol/l oppure tra 45 e 75 mg/dl. È necessario sot-tolineare che l’incessante pressione selettiva sullavacca da latte, con l’obiettivo di incrementare laproduzione di latte e la concentrazione sia di gras-so che proteine, tende a incidere su questi valori ri-
Fertilità e patologie in bovini e suini
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 20128
Figura 4.
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 8

tenuti normali in una fase intermedia della latta-zione. Quando la concentrazione di glucosio ema-tico tende a innalzarsi, l’insulina favorisce l’uptakedi glucosio nei tessuti dove sono presenti recettoriper questo ormone. Di converso, quando per unforte utilizzo dei tessuti o per scarso approvvigio-namento alimentare la glicemia tende ad abbas-sarsi, interviene il glucagone, essenzialmente a li-vello delle cellule epatiche, attivando la degrada-zione del glucosio in esse stoccato come glicogeno(glicolisi). In realtà la secrezione dell’insulina è sti-molata anche dal mannosio e dagli aminoacidi eparticolarmente dalla leucina, per la quale esiste unrecettore specifico nelle cellule beta. Esiste poi un’a-zione di stimolo nella secrezione esercitata dai cor-pi chetonici, dagli acidi grassi e da parte di alcuniormoni gastro-intestinali come la gastrina, la pan-cretina e la pancreozimina. Va ricordato che l’e-mivita dell’insulina in circolo è di soli 5-10 minu-ti. L’insulina agisce anche sulle cellule nervose, maè bene ricordare che il sistema di trasporto in essepresente è limitato. Altro aspetto, che è bene ri-cordare come premessa nella comprensione delleproblematiche riproduttive della vacca da latte, èche le cellule dell’epitelio alveolare mammario, incui avviene la sintesi dei costituenti del latte e quel-la del latte stesso, sono indipendenti dall’insulinanel meccanismo di captazione del glucosio emati-co per la sintesi del lattosio e quindi del latte. È necessario anche approfondire l’effetto dell’in-sulina sui vari tessuti. A livello del tessuto adipo-so un ormone anabolico come l’insulina stimolala sintesi degli acidi grassi, del glicerolo fosfato,l’uptake degli aminoacidi e del potassio e infineinibisce la sensibilità ormonale della lipasi. Nel fe-gato stimola la sintesi del glicogeno, ossia lo stoc-caggio del glucosio, l’ossidazione dello stesso e lasintesi dei lipidi e delle proteine. La stimolazionedell’insulina a livello degli epatociti determina unariduzione della produzione di corpi chetonici (che-togenesi). A livello muscolare, oltre a stimolarela sintesi proteica, di acidi grassi, del glicerolo fo-sfato e ovviamente l’ingresso di glucosio nelle cel-lule muscolari, riduce il catabolismo degli ami-noacidi chetogenetici. L’insulina favorisce l’upta-ke muscolare dei corpi chetonici e dello ione po-tassio. In senso generale e sull’intero organismol’insulina stimola la crescita cellulare e il quozien-te respiratorio. È chiaro dunque che l’insulina, es-sendo secreta quando la glicemia e alcuni ami-noacidi nel sangue sono elevati, ha la possibilità dimisurare il bilancio energetico dell’intero animalee di informare i vari sensori metabolici in esso pre-senti. L’azione dell’insulina può essere divisa in ra-pida, intermedia e ritardata. L’azione rapida, os-sia che si realizza in pochi secondi, è quella di au-mentare il trasporto di glucosio, aminoacidi e io-ni potassio nelle cellule insulino-sensibili. L’azione intermedia si espleta nell’ordine di minutied è relativa alla stimolazione della sintesi protei-ca, all’inibizione e degradazione delle proteine eall’attivazione della glicogeno-sintetasi. L’azione
ritardata, che si espleta nell’arco di alcune ore, èrelativa all’incremento della sintesi di mRNA perla lipogenesi e di altri enzimi. Sia il glucosio, comealtri nutrienti, sia direttamente l’insulina agisconoa livello di sistema nervoso centrale per “infor-marlo” sullo status energetico dell’intero organi-smo. Nella sezione caudale dell’encefalo esistonoa livello dei neuroni NPY-CA dei recettori per l’in-sulina. Questi neuroni sono in connessione conquelli ipotalamici che secernono il GnRH e quin-di in grado di modulare la secrezione di LH e FSHe anche il comportamento estrale. Anche a livello ovarico, e più specificatamente alivello di follicolo dominante, esistono recettori perl’insulina. L’insulina è in grado di stimolare la pro-duzione di progesterone. Come spesso accade, percomprendere il ruolo di un ormone si studiano leanomalie del metabolismo e le patologie conse-guenti in caso di carenza. La ridotta secrezionedi insulina si identifica con il diabete, per cui a fron-te di uno stimolo alla sua secrezione derivante daun innalzamento della glicemia non si verifca un’a-deguata risposta da parte delle cellule beta delleisole di Langerhans pancreatiche. Tale risposta ina-deguata causerà la sintomatologia descritta nel dia-bete mellito o tipo 1. Nel caso della bovina, a cau-sa della carenza di insulina ed essendo il tessutomammario da essa indipendente, la produzione dilatte non calerà, ma anzi ne verrà avvantaggiata equesto può aver fatto sottovalutare il problema. Ènoto che una somministrazione farmacologica diinsulina provoca nella vacca da latte una bruscacaduta della produzione. Aumentando il glucosioin circolo ne aumenterà la disponibilità per la sin-tesi di lattosio e quindi di latte. Noteremo, ac-canto all’iperglicemia, glicosuria, diuresi osmoti-ca e conseguentemente perdita di elettroliti. Man-cando glucosio ai tessuti verrà incentivato il ca-tabolismo degli aminoacidi da destinare alla glu-coneogenesi. Ci sarà un conseguente aumentodi aminoacidi plasmatici con incrementata per-dita di azoto nelle urine. Nella necessità di averea disposizione energia, la carenza di insulina in-duce la lipomobilizzazione con il conseguente in-cremento di NEFA plasmatici e l’inevitabile con-seguente chetonuria, chetogenesi e chetonemia. Ilprotrarsi e l’aggravarsi di questa condizione puòportare ad acidosi metabolica, disidratazione e, al-la fine, alla morte. Abbiamo visto come la selezione genetica per laproduzione di latte abbia favorito un assetto or-monale che premia una maggiore secrezione di GHa scapito di quella di insulina, offrendo un quadrometabolico degli animali ad alto potenziale gene-tico che ricorda quello del paziente diabetico. Esi-ste anche la possibilità che a causa del persisteredi valori elevati di glicemia possa maturare un qua-dro biochimico riconducibile al diabete tipo 2, os-sia da insulino-resistenza, caratterizzato da unacorretta secrezione pancreatica di insulina ma dauna refrattarietà dei recettori, troppo sollecitati neltempo e nelle quantità. Dai dati attualmente dis-
Fertilità e patologie in bovini e suini
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201210
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 10

ponibili sembrerebbe quindi che la superiorità ge-netica di una bovina da latte si traduca nel disporredi una maggiore concentrazione ematica di ormonesomatotropo (GH) (vedere figura 5) in tutte lefasi del ciclo produttivo, dalla nascita al primo par-to e, soprattutto, presupponga la presenza di unmaggior numero di recettori sugli organi bersaglio.Il GH aumenta l’efficienza alimentare, ossia la ca-pacità di trasformare gli alimenti ingeriti in latte,stimola l’anabolismo proteico, incrementa la ri-tenzione dell’azoto e sopprime la capacità del-l’insulina di stimolare l’uptake del glucosio. Tro-viamo i recettori del GH su molti organi e tessuticome quello adiposo, muscolare, nell’utero e sul-l’ovidutto, e in particolare a livello epatico, ovari-co e anche ipotalamico e ipofisario. I recettori epa-tici di GH sono molto importanti perché correla-ti con la produzione dell’IGF-1 con il quale esistecomunque un feedback negativo. Nel pre-partoaumenta la concentrazione di GH, mentre aumentaquella di IGF-1. Nel parto e nel post partum av-viene il contrario. Situazioni come il bilancio ener-getico negativo, malattie, invecchiamento ecc., ini-biscono la sintesi epatica di IGF-1. Questo ormo-ne stimola la risposta del follicolo all’LH. Esisteuna correlazione tra IGF-1 follicolare e plasmati-co. Questo ormone stimola, inoltre, la prolifera-zione e la capacità steroidogenica della teca e del-la granulosa. Un altro effetto noto e strettamente correlato al-la produzione di latte è la correlazione positiva esi-stente tra GH e afflusso di sangue alla mammella.Un animale geneticamente selezionato per pro-durre latte avrà un maggior afflusso di sangue equindi di nutrienti alla mammella. L’uso della so-matotropina sintetica bST, autorizzata negli Usanella vacca da latte, con lo scopo di aumentare laproduttività degli animali agendo sulla persisten-za della curva di lattazione, ha permesso di ap-profondire il meccanismo d’azione di questo or-mone. In uno studio fatto da Barman nel 1987 ven-gono confrontate alcune situazioni metaboliche divacche geneticamente superiori con quelle deri-vanti dall’uso del bST riscontrando interessantianalogie. Sia la bovina ad alto potenziale genetico(HMG) che quella trattata con bST (bST-T) han-no un incremento di ingestione che purtroppo au-menta solo dopo alcune settimane dal picco di lat-tazione, quindi come nelle vacche di basso po-tenziale genetico. Sia nelle bovine HMG che inquelle bST-T si osserva un ricorso più massiccioalle riserve corporee di grassi per garantire un’a-deguata copertura energetica all’incremento pro-duttivo del post partum o del periodo successivoalla somministrazione della bST. In ambedue leclassi di bovine si osserva un aumento della quan-tità di tessuto secretorio mammario. Non è notal’attività diretta dell’ormone sulle cellule dell’al-veolo mammario. Sia negli animali HMG che inquelli bST-T è necessario un incremento delle ca-pacità manageriali per garantire adeguate perfor-mance riproduttive. Da questo semplice confron-
to viene ulteriormente confermato come la supe-riorità genetica si traduce in un diverso assetto or-monale che tende a premiare il GH a scapito del-l’insulina. La secrezione di GH può comunque es-sere influenzata da fattori metabolici e ormonali.Lo stress paradossalmente stimola la produzionedi GH. L’ipoglicemia, il digiuno, una bassa con-centrazione di NEFA, l’arginina, la leucina e il dia-bete stimolano la produzione di GH, così come al-cuni ormoni quali il GNRH, gli estrogeni, il glu-cagone e bassi livelli di IGF-1. L’ipotiroidismo eun alto livello di glucocorticoidi inibiscono il GH,come anche l’obesità. Abbiamo visto come l’asse-ipotalamo ipofisario ha la necessità di “conosce-re” la disponibilità di nutrienti, sia direttamenteche indirettamente, attraverso l’assetto ormonale.Abbiamo accennato all’inizio che la “decisione diriprodursi” deve essere presa disponendo di in-formazioni sulla situazione dei nutrienti a brevetermine, ma anche sul medio-lungo periodo. Perquesto e per altre motivazioni esiste un ormone se-creto dal tessuto adiposo denominato leptina (ve-dere figura 6). La sintesi di questo ormone è di-rettamente proporzionale alla quantità di riservelipidiche presenti e più specificatamente alle di-mensioni degli adipociti che costituiscono il tes-suto adiposo. Più gli adipociti sono grandi, mag-giore sarà la concentrazione ematica di leptina.La sintesi della leptina è inibita da una minoredisponibilità di nutrienti per il tessuto adiposo,dall’ipo-insulinemia, dalla presenza dei glucocor-ticoidi e da un’aumentata concentrazione di GHe catecolamine. La leptina dovrebbe e il condizio-nale è d’obbligo ridurre l’appetito, il catabolismoproteico e aumentare i depositi energetici. Nel-l’ambito della fisiologia comparata, un soggettoumano obeso dovrebbe veder ridurre sensibilmenteil proprio appetito e quindi non ingrassare più deldovuto. Esistono dei meccanismi complessi e al-cuni non ben noti e non ben regolati da un’evo-luzione naturale, che forse non aveva mai dovu-to affrontare una situazione di illimitata disponi-bilità di cibo nel tempo e nella quantità. Anchein questo caso il “baco” evolutivo potrebbe chia-marsi leptino-resistenza, ossia quella condizionepatologica derivante da una continua e incessan-te stimolazione dei recettori leptinici dei tessuti alpunto di renderli resistenti allo stimolo stesso. Stes-sa condizione che abbiamo prima descritto per l’in-sulino-resistenza. Una bovina da latte ha continuadisponibilità di cibo di alta concentrazione ener-getica e proteica, cosa che in natura si può verifi-care solo in alcune stagioni dell’anno e comunqueper periodi molto limitati. La leptina, a causa deinumerosi recettori presenti sui tessuti coinvolti nel-l’attività riproduttiva può avere un ruolo nella fer-tilità. Troviamo recettori a livello ovarico, ipofi-sario e sopratutto a livello ipotalamico nella se-zione caudale dell’encefalo, dove troviamo i re-cettori dell’insulina e dove la leptina esercita la stes-sa funzione. Il GH inibisce l’azione di stimolo deiglucorticoidi e dell’insulina nella secrezione di lep-
Metabolismo e fertilità nella bovina da latte
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201211
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 11

tina mentre l’IGF-1 avrebbe un feedback positi-vo con questo ormone. L’insulina stimola diretta-mente la produzione di leptina. Il complesso in-sulina-leptina è in grado, pertanto, di fornire al si-stema nervoso centrale un quadro sulla situazio-ne nutrizionale di breve e medio periodo che, uni-tamente alle informazioni derivanti dal fotoperio-do possono dare il “consenso” o meno alla bovi-na di riprodursi. La selezione genetica artificiale,cioè guidata dall’uomo, ha dato un vantaggio ri-produttivo a quegli animali la cui produzione dilatte, grasso e proteine era superiore alla mediadella popolazione. Per far questo, ossia per ga-rantire un maggior apporto di nutrienti alla mam-mella e dare ad essa sempre più priorità rispettoalle altre funzioni metaboliche, si è dovuto inces-santemente modificare l’assetto ormonale e meta-bolico degli animali, molto spesso inconsapevol-mente.
◗ Conclusioni La selezione, in sostanza, agisce premiando lecaratteristiche o meglio l’attitudine materna del-le bovine. L’attività riproduttiva, ossia l’instau-rarsi di una nuova gravidanza, non ha prioritàmetaboliche o meglio ne ha di più in quei Pae-si dove negli indici genetici sono stati aggiunticaratteri funzionali come la fertilità. Ad aggra-vare la situazione è la necessità assoluta che lanuova gravidanza si instauri quando la bovinaè ancora in deficit energetico e proteico ossia nel
primo terzo della lattazione, per avere nell’alle-vamento quel regolare alternarsi di parti e asciu-gamenti, funzionale a tenere entro 160 i giornimedi di lattazione dell’allevamento. Per ottene-re questo straordinario incremento della pro-duttività della Frisona si è dovuto consapevol-mente o inconsapevolmente riassettare l’equili-brio ormonale e conseguentemente metabolico.Sembrerebbe ormai chiaro che l’animale di al-to potenziale genetico ha una maggiore con-centrazione di GH e dei suoi recettori rispettoad animali di minor potenziale. Conseguente-mente a questo, e forse mascherata dalla suascarsa influenza sulla mammella, si è ridotta laconcentrazione ematica di insulina, creando ne-gli animali un quadro metabolico, molto evi-dente all’inizio lattazione, riconducibile al dia-bete tipo-1. Animali cioè che dimagriscono mol-to velocemente nelle prime fasi di lattazione perpoi ingrassare altrettanto rapidamente alla finedi essa specialmente se non gravidi. Si tratta divacche con un livello tendenzialmente più ele-vato di NEFA e BHBA nel sangue di quelle dibasso livello genetico. Probabilmente, nell’ag-gravamento di questo quadro derivante da un’a-zione su un assetto ormonale modificato, po-trebbero giocare un ruolo l’instaurarsi o la piùfacile predisposizione all’insulino e leptino-re-sistenza, patologie derivanti dalla mancanza diun’adeguata protezione evolutiva, in quantodi difficile realizzazione nell’ambito della solaselezione naturale.
Fertilità e patologie in bovini e suini
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201212
PER SAPERNE DI PIÙ
• Bauman D.E., Currie W.B.: Partitioning of nutrients du-ring pregnancy and lactation: a review of mechanismsinvolving homeostasis and homeorhesis. J. Dairy Sci.1980; 63(9):1514-1529.
• Bonczek R.R., Young C.W., Wheaton J.E., Miller K.P.:Responses of somatotropin, insulin, prolactin, and thyro-xine to selection for milk yield in holsteins. J. Dairy Sci.1988; 71(9):2470-2479.
• Gong J.G., Lee W.J., Garnsworthy P.C., Webb R.: Effectof dietary-induced increases in circulating insulin con-centrations during the early postpartum period on re-productive function in dairy cows. Reproduction 2002;123(3):419-427.
• Weber W.J., Wallace C.R., Hansen L.B., Chester-JonesH., Crooker B.A.: Effects of genetic selection for milk yieldon somatotropin, insulin-like growth factor-I, and pla-cental lactogen in Holstein cows. J. Dairy Sci. 2007;90(7):3314-3325.
• Veerkamp R.F., Beerda B., van der Lende T.:Effects ofgenetic selection for milk yield on energy balance, le-vels of hormones, and metabolites in lactating cattle,and possible links to reduced fertility. Livest. Prod. Sci.2003; 83(2-3):257-275.
• Wiltbank M., Lopez H., Sartori R., Sangsritavong S., G-men A.: Changes in reproductive physiology of lactatingdairy cows due to elevated steroid metabolism. Therio-genology 2006; 65(1):17-29.
• Lucy M.: Mechanisms linking growth hormone, insu-lin and reproduction: lessons from the postpartum dairycow. Cattle Pract. 2006; 14(1):23-28.
• Bereket A., Lang C.H., Wilson T.A.: Alterations in thegrowth hormone-insulin-like growth factor axis in insu-lin dependent diabetes mellitus. Horm. Metab Res. 1999;31(2-3):172-181.
• Radcliff R.P., McCormack B.L., Crooker B.A., LucyM.C.: Plasma hormones and expression of growthhormone receptor and insulin-like growth factor-ImRNA in hepatic tissue of periparturient dairy cows.J. Dairy Sci. 2003; 86(12):3920-3926.
• Radcliff R.P., McCormack B.L., Crooker B.A., LucyM.C.: Growth hormone (GH) binding and expressionof GH receptor 1A mRNA in hepatic tissue of peri-parturient dairy cows. J. Dairy Sci. 2003; 86(12):3933-3940.
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 12

ffinché si instauri una gravidanza esi completi la gestazione fino ad ar-rivare al parto, è necessario che leovaie siano esenti da patologie, per-ché ciò può compromettere una cor-
retta ovulazione. Inoltre, è fondamentale la qua-lità dell’oocita e un corpo luteo in grado di pro-durre adeguate quantità di progesterone, alme-no inizialmente. Altrettanto importante è la ca-pacità dell’utero di nutrire adeguatamente l’em-brione, almeno fino alla placentazione, che av-viene intorno al 35° giorno di gestazione.
◗ Le infezioni dell’utero Per infezioni dell’utero si intendono tutti i proces-si infettivi che possono interessare l’endometrio e/ol’intero l’utero. Possono anche essere definite comeinfezioni non specifiche, che riducono l’efficienzariproduttiva e, quindi, la redditività. Le infezioniuterine possono essere classificate in base al tipo(endometrite o metrite), all’epoca di insorgenza eal decorso (acuto, subacuto o cronico). La piome-tra rappresenta invece un caso a sé stante.
Infezioni uterine: alcuni termini• Periodo post partum: corrisponde all’inter-vallo temporale tra il parto e la completa in-voluzione uterina. Tale periodo è ulteriormen-te suddiviso in:- puerperio (fase puerperale): l’arco temporale com-preso tra il parto e il momento in cui l’ipofisi tor-na ad essere sensibile al GnRH. La durata di que-sto periodo è generalmente di 7-14 giorni;- fase intermedia: rappresenta l’intervallo fra lafine del puerperio e la prima ovulazione, chepuò verificarsi in tempi molto variabili;- periodo post ovulatorio: l’arco temporale trala prima ovulazione e la completa involuzioneuterina, ossia quando il diametro delle cornauterine e della cervice è inferiore a 40 mm e lecaruncole sono di nuovo epitelizzate. • Endometrite: qualsiasi processo infiammato-rio a carico dell’endometrio.• Metrite: processo infiammatorio a carico ditutti gli strati della parete uterina.Durante il periodo puerperale, si registra la ri-duzione più marcata del volume dell’utero: a se-condamento avvenuto, l’utero pesa circa 10 kg,per arrivare a 2 kg in circa dieci giorni. L’involuzione uterina inizia già 2 giorni dopo ilparto e, dopo 5 giorni, le caruncole uterine ca-
dono in necrosi e liquefazione; alla fine del pe-riodo puerperale (15 giorni), inizia la rigenera-zione dell’epitelio. In questa fase, la produzio-ne di prostaglandine (che contribuiscono all’in-voluzione uterina) è massima, per ridursi poiprogressivamente, fino a raggiungere i livelli nor-mali 20 giorni dopo il parto.Circa il 90% delle bovine sviluppa un’endome-trite moderata e fisiologica, sostenuta dai bat-teri ambientali penetrati durante il parto.La dissoluzione delle caruncole, che avviene nei pri-mi 10 giorni dopo il parto, produce un materialedi colore variabile, dal biancastro al rossastro, fi-no al marrone scuro, detto “lochi”. Nelle prime 48ore dopo il parto, questo materiale è prodotto innotevole quantità (2 l) e, in seguito, si riduce finoad arrivare a 1 l al quinto giorno post partum e a500 ml all’ottavo. Solo pochi millilitri caratteriz-zano invece la fase finale del puerperio.Un odore intenso e fetido dei lochi è espressio-ne di infiammazione uterina; in questo caso, èragionevole sospettare la presenza di una me-trite puerperale. Il colore, al contrario dell’o-dore, non è legato a patologie endometriali.Nella fase intermedia, o meglio alla terza setti-mana, la mucosa uterina è del tutto rigeneratae la bovina, nella fase successiva, potrebbe ini-ziare una nuova gravidanza.Le vacche sane riducono o eliminano comple-tamente i germi patogeni che invadono l’uterodopo il parto. In questa fase, è possibile riscon-tare sia metrite sia endometrite. Le metriti diquesto periodo sono definite croniche.• Piometra: grave processo infiammatorio a ca-rattere purulento che può insorgere nel periodopost ovulatorio. Per la presenza del corpo luteoe, quindi, data la chiusura della cervice, il pusnon può raggiungere l’esterno e si accumula nellume uterino.L’utero è un ambiente normalmente sterile ma, inconcomitanza del parto, la cervice si apre, con-sentendo la penetrazione dei batteri ambientali(cute, feci). Normalmente i batteri sono elimina-ti, ma almeno il 20% delle bovine non è in gradodi effettuare il risanamento dell’ambiente uterino,con conseguente sviluppo di metrite. Quest’ulti-ma, nel 10-15% degli allevamenti, può persisteresotto forma di endometrite. Il dato allarmante èche le bovine in cui l’endometrite va incontro a ri-soluzione hanno un tasso di concepimento infe-riore del 20% rispetto alle bovine che non si so-no mai ammalate, mentre il 3% diventa sterile.
Patologie ovariche-uterine e gravidanza
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201213
Alessandro FantiniFacoltà di Medicina veterinaria– Università di PerugiaA
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 13

◗ Gli agenti patogeni Nel post partum, il lume uterino consente la cre-scita di una notevole varietà di batteri, sia anae-robi sia aerobi. Molti dei batteri opportunistiche troviamo in utero sono rimossi e, per alcu-ne settimane dopo il parto, si osserva l’alternarsidi condizioni di risanamento e infezione.Le specie batteriche che possono essere isolatedal lume uterino sono più di 30 e sono classi-ficate in patogene, potenzialmente patogene eopportuniste.Al gruppo delle specie patogene, così definite inquanto associate a lesioni uterine, appartengo-no Escherichia coli, Arcanobacterium pyogenes,Fusobacterium necrophorum e Prevotella spp.Con la definizione “potenziali patogeni” si in-tendono tutti i batteri presenti in utero, non ne-cessariamente associati a patologie.Infine, gli opportunisti sono rappresentati daspecie batteriche non patogene, che colonizza-no l’utero dopo il parto.Interessante è notare l’azione sinergica che eser-citano E. coli, A. pyogenes e Prevotella sull’in-sorgenza dell’endometrite, sulla sua gravità e sulsuo decorso.Dopo il parto, il primo batterio ad essere indivi-duato a livello uterino è E. coli, fin dai primissi-mi giorni e in percentuale dominante. E. coli ren-de l’utero particolarmente suscettibile all’infezio-ne da A. pyogenes, con conseguente insorgenzadi gravi endometriti. A. pyogenes, a sua volta, pro-duce un fattore di crescita per F. necrophorus.Man mano che la bovina si allontana dal parto,gli isolamenti batterici si riducono e la presenzadi A. pyogenes, 28-35 giorni dopo il parto, è as-sociata a un ridotto tasso di concepimento.L’inoculazione di A. pyogenes ed E. coli evocauna risposta anticorpale che aiuta l’utero a con-tenere la durata e la gravità dell’infezione, an-che se il ruolo dell’immunità specifica rimaneancora non ben definito.Nelle fasi iniziali, il meccanismo difensivo più im-portante è quello cellulo-mediato, anche controbatteri non opsonizzati dagli anticorpi. La fago-citosi è un’attività che compete fondamentalmenteai polimorfonucleati (PMN) e agli altri leucociti.I cambiamenti dell’assetto ormonale, tipici del pe-riparto, possono interferire con l’efficacia del-l’immunità innata della vacca da latte. La soma-totropina induce la produzione, da parte delle cel-lule epatiche, di somatomedina (IGF-1) e, per que-sto motivo, la sua concentrazione nel peripartoè bassa. Tale assetto ormonale può essere ulte-riormente compromesso dalla presenza di corpichetonici ematici. L’IGF-1 e le proteine ad essoassociate influenzano la sopravvivenza cellulare,l’apoptosi, e la funzionalità dei PMN. Nel peri-parto, la presenza di NEFA e BHBA altera la ca-pacità di migrazione, la fagocitosi e l’abilità neldistruggere il batterio fagocitato; nei primi tre gior-ni dopo il parto, una concentrazione di BHBA >1
mmol/l predispone la bovina alle mastiti clinichee alla metrite puerperale.In questa fase, un meccanismo difensivo moltoimportante è la contrattilità del miometrio, checonsente la totale ed efficace espulsione dei lochie, quindi, della flora batterica, patogena e non,presente nell’utero. Tuttavia, è contraddittoria lamancanza di correlazione tra il calcio ionizzatoplasmatico e la contrattilità uterina.La presenza in utero della massa batterica è di-rettamente correlata con un aumento della con-centrazione di endotossine. Queste ultime, de-rivanti essenzialmente dalla parete cellulare deibatteri Gram-, sono la conseguenza dell’inten-sa proliferazione e morte batterica e, per la lo-ro natura chimica, sono definite lipopolisacca-ridi (LPS). Gli LPS batterici sono rappresentatidalla porzione glicolipidica (lipide A) della pa-rete interna dei batteri Gram-negativi. Gli LPSpossono diffondere dall’utero, così come da al-tri distretti (mammella, rumine e intestino), nelsangue periferico e nel fluido follicolare e rag-giungere l’ipotalamo. Le tossine LPS stimolanola produzione di citochine, come l’IL-1 e il TNF,le prostaglandine, l’ACTH e il cortisolo.In presenza di elevate concentrazioni di LPS, siriduce la concentrazione ematica di estrogeni,LH e progesterone, ma non quella di FSH. Que-sto “riassetto” ormonale è dovuto all’azione di-retta delle citochine e dei glucocorticoidi a livel-lo ipotalamico, dove si osserva un aumento del-la concentrazione di citochine, prostaglandine,oppiodi, CRH e vasopressina. Tale condizioneinibisce la secrezione di GnRH e conseguente-mente la secrezione ipofisaria di LH. La presen-za di LPS nel liquido follicolare inibisce la pro-duzione di estrogeni e, a livello di corpo luteo,quella di progesterone. Quindi, l’infezione bat-terica uterina può sopprimere la secrezione diLH, la steroidogenesi follicolare, con conseguenteaumento della lunghezza della fase luteinica, elo sviluppo di follicoli e di corpi lutei di minoridimensioni. Queste interferenze sullo sviluppodel follicolo e del corpo luteo si ripercuotono alungo, concorrendo negativamente sulla ripresadell’attività ovarica dopo il parto e sulla conse-guente qualità dell’oocita (vedere figura 1). Quan-do la presenza di batteri, più o meno patogeni,supera la capacità del sistema immunitario di te-nere sotto controllo la situazione, insorgono leforme cliniche classificate come metriti puerpe-rali, endometriti e piometre.
◗ La metrite puerperale La metrite puerperale è una complicazione batte-rica della fase puerperale; pertanto, si verifica nel-le prime due settimane dopo il parto, tipicamen-te tra il 4° e il 10° giorno, ed è caratterizzata daun essudato fetido e maleodorante di colore ros-so-bruno, contenente brandelli necrotici. La pa-rete uterina appare inizialmente sottile, poi ede-
Fertilità e patologie in bovini e suini
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201214
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 14

matosa. La metrite puerperale può essere asso-ciata a segni sistemici, come sonnolenza, prostra-zione e febbre (>39,5 °C), e la sua incidenza ne-gli allevamenti può variare dal 2,2 al 37,3%.I processi infiammatori e degenerativi, unita-mente all’infezione batterica, possono interes-sare non solo l’endometrio, ma anche la sub-mucosa e gli strati più profondi della parete ute-rina, causando danni che dall’epitelio ghiando-lare possono estendersi a tutto l’organo. I bat-teri evidenziati in corso di metrite puerperale so-no A. pyogenes (33-83%), E. coli (67-85%) eGram- anaerobi, come F. necrophorus, Bacte-roides spp. e Prevotella spp. (49-67%). In ca-so di metrite puerperale, nell’essudato patolo-gico, si rinvengono frequentemente streptococ-chi non emolitici e Mannheimia haemolytica.La ridotta contrattilità uterina e l’alterata fun-zionalità dei neutrofili, in termini di migrazio-ne, fagocitosi e scoppio respiratorio, condizio-nano l’insorgenza e la gravità della metrite puer-perale della vacca da latte. La presenza dei bat-teri e dei fattori condizionanti, come il ristagnodei lochi, una pregressa distocia e la ritenzioneplacentare, determinano una massiccia libera-zione di LPS, che stimolano, per chemiotassi, lamigrazione dei leucociti dal sangue verso il lu-me dell’utero. Questa azione primaria parte es-senzialmente dalla liberazione, da parte dei ma-crofagi, di istamina e citochine pro-infiamma-torie (IL-1, IL-6 e TNFα) e dall’attivazione dienzimi come la fosfolipasi A2, la COX-2 e la 5-lipossigenasi. Questo gruppo di enzimi pro-muove la sintesi e la secrezione di alcune pro-staglandine, come la PGF2α e la PGE2. La pri-ma di questi eicosanoidi ha proprietà luteoliti-che e immunostimolanti, nonché la capacità di
stimolare la contrattilità uterina. Nelle prime 2-3 settimane dopo il parto, aumenta il livello pla-smatico di PGF2α e di PGFM inattivo. La pre-senza di PGFM coincide con la rapida regres-sione dell’utero.L’altra prostaglandina, la PGE2, prodotta dal-l’endometrio e dalle cellule dello stroma sottola stimolazione delle endotossine, ha attività im-munosoppressiva e luteotropica. Le citochinee alcune delle altre sostanze prodotte dai ma-crofagi attivati raggiungono il sistema nervosocentrale, alterando la produzione e la secrezio-ne di neurotrasmettitori e ormoni come il cor-tisolo, le catecolamine, la prolattina, il GH,l’IGF-1, l’insulina, il glucagone, la leptina, gliormoni tiroidei e l’LH. Contemporaneamente,si osserva anche una maggiore produzione diproteine della fase acuta, soprattutto l’α1-AG el’aptoglobina (HP). Queste profonde modifica-zioni inducono febbre, letargia, anoressia e cam-biamenti nel metabolismo energetico, proteicoe minerale, con perdita di peso e aggravamentodel bilancio energetico negativo (NEBAL). Neicasi molto gravi, si può verificare ipotensione,tachicardia e diarrea.La fase in cui può insorgere una metrite puer-perale coincide con il reclutamento della primacorte follicolare e l’emergenza e la selezione delprimo follicolo dominante. A causa della ridot-ta produzione di LH, estrogeni, insulina e IGF-1, la ripresa dell’attività ovarica dopo il partorisulta compromessa. La diagnosi di metrite puerperale è relativamen-te semplice. Il sintomo primario è uno scolo va-ginale acquoso, particolarmente fetido e di colo-re rosso-bruno, rilevabile nelle bovine in puer-perio. Nelle forme molto gravi, lo scolo appare
Patologie ovariche-uterine e gravidanza
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201215
Figura 1. Possibili mediatori della soppressione endotossica della concentrazione di LH in circolo.
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 15

alterato già dal 3° giorno dopo il parto, mentre inquelle più leggere dal 6° al 10° giorno. L’utero del-le bovine con metrite puerperale appare edema-toso e assottigliato. I sintomi generali, in partico-lare la febbre, non sempre sono presenti; infatti,è bene sottolineare che, per questa infezione ute-rina, l’ipertermia non è patognomonica.
◗ Endometrite clinica e piometra
Per endometrite clinica si intende la presenza dimateriale muco-purulento e purulento nell’ute-ro di bovine che si trovano nella fase successi-va al puerperio, ossia 14 giorni dopo il parto.L’incidenza è molto variabile (7,5-61,6%). L’es-sudato patologico che caratterizza l’endometri-te clinica può essere mucopurulento: costituitoper il 50% da pus e per il 50% da muco, o pu-rulento quando la concentrazione di pus supe-ra il 50%. A causa dell’apertura della cervice,l’essudato patologico esce dall’utero e invade illume vaginale. L’infezione è in genere sostenu-ta da A. pyogenes e batteri Gram- anaerobi, de-rivanti da metriti puerperali persistenti o recentie da successive contaminazioni esterne. In ca-so d’endometrite, l’isolamento di E. coli è raro.Questa patologia è caratterizzata dal coinvol-gimento del solo endometrio, e solo raramentel’infezione si estende allo strato spongiforme sot-tostante. Si osserva un cambiamento degene-rativo dell’epitelio superficiale, con congestio-ne vascolare ed edema dello stroma.A causa dell’infezione, un potente stimolo che-miotattico richiama dal sangue i PMN, i linfocitie le plasmacellule, con un’imponente produzionelocale di TNFα, leucotrieni ed eicosanoidi (PGE2e PGF2α). Dato lo scarso assorbimento di cito-chine da parte dell’utero, l’endometrite clinica nonè associata a nessuna sintomatologia generale enon si registra alcun calo della produzione di lat-te. Si osserva iperemia e congestione a carico del-l’endometrio, mentre le cellule superficiali des-quamano e necrotizzano. Pertanto, l’essudato pa-tologico è costituito dalle cellule immunitarie e dalmateriale necrotico proveniente dalla distruzionedell’epitelio superficiale.Nella forma cronica, il tessuto cicatriziale puòsostituire il tessuto funzionale dell’endome-trio, con conseguente fibrosi perighiandolare,degenerazione cistica e atrofia delle ghiando-le uterine.La piometra è sostanzialmente un’endometriteclinica, che insorge nelle bovine in cui è già av-venuta la ripresa dell’attività ovarica. Infatti, lapresenza del corpo luteo, con la conseguenteproduzione di progesterone, determina la chiu-sura della cervice e, quindi, l’accumulo del ma-teriale muco-purulento nel lume uterino.La diagnosi di endometrite clinica è relativa-mente semplice, anche se i criteri di valutazio-ne sono spesso molto soggettivi. Essa si basa sul-
la presenza di essudato patologico a livello va-ginale e sulla valutazione delle dimensioni del-l’utero, tenendo presente che, a 3-5 settimane,non è ancora completamente involuto.L’endometrite clinica e la piometra non sono ca-ratterizzate né da sintomi generali, né da calodella produzione lattea. Inoltre, la sola palpa-zione transrettale, se non associata a un esamecon il vaginoscopio, non consente di diagnosti-care con precisione questo gruppo di patologie.LeBlanc, in un importante studio effettuato su1.865 bovine in 27 allevamenti, ha riscontratoun’incidenza dell’endometrite clinica pari al16,9% negli animali visitati tra il 20° e il 33°giorno post partum. LeBlanc ha pertanto defi-nito l’endometrite clinica come la patologia ute-rina che, 21 giorni dopo il parto e quindi do-po il puerperio, determina la presenza di un es-sudato muco-purulento o purulento, a volte ma-leodorante, in vagina, unitamente a un diame-tro della cervice >7,5 cm[1]. In ogni caso, la pre-senza di un essudato vaginale oltre il 26° gior-no dopo il parto è considerata patologica.Un’alternativa al vaginoscopio è lo strumento mes-so a punto da G. Reddle (Nuova Zelanda), deno-minato “metricheck”. Secondo l’autore, prima del2004, anno di presentazione dello strumento, lapercentuale d’endometrite clinica riscontrata in4.554 vacche di 15 allevamenti, era del 10%.La diagnosi di piometra è relativamente ancorapiù semplice e si basa sulla palpazione transret-tale o sull’ultrasonografia. Questi esami con-sentono di individuare un aumento delle di-mensioni dell’utero e la presenza d’essudato nellume uterino, contemporaneamente alla presen-za di un corpo luteo e di una cervice chiusa.
◗ Endometrite subclinica L’endometrite subclinica è un’infiammazionecronica e inapparente dell’endometrio, caratte-rizzata da un’elevata infiltrazione di PMN. Ta-le patologia insorge al completamento istologi-co dell’involuzione uterina, ossia 8 settimanedopo il parto. La sintomatologia clinica è ine-sistente, in quanto non sono presenti sintomi ge-nerali o alterazioni dell’essudato uterino.La bovina con endometrite sub-clinica non è fer-tile. In genere, per diagnosticare le patologie ute-rine, si ricorre alla palpazione transrettale e al-l’ultrasonografia. In questo caso, l’unica pos-sibilità diagnostica è data dalla valutazione quan-titativa dei PMN in utero (citologia endome-triale, EC). La tecnica impiegata è il cytobrush,adattata ai grossi animali. I valori normali di-pendono dalla distanza dal parto.Il sospetto d’endometrite subclinica insorge quan-do si riscontra più del 18% di PMN, a 20-33 gior-ni dopo il parto, mentre l’EC è considerato posi-tivo con PMN >10%, a 34-47 giorni dopo il par-to. In ogni caso, l’utero della bovina è sano quan-do i PMN sono <5%. Le positività in allevamen-
Fertilità e patologie in bovini e suini
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201216
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 16

to possono essere molto elevate. Secondo Gilbert,è possibile riscontrare prevalenze del 53%, convariazioni dal 37 al 74%3.
◗ Principi di terapia e prevenzione
Nella bovina da latte la prevenzione gioca unruolo fondamentale nella riduzione dell’inci-denza del complesso metrite-endometrite. Fon-damentale è la riduzione dell’incidenza di de-terminate condizioni, come la ritenzione pla-centare (RFM), le ipocalcemie post partum, ledistocie e la gemellarità. Per quanto riguarda leprime due patologie, è importante la gestionedella preparazione al parto, al fine di evitare glistress derivanti dal sovraffollamento, e garanti-re un apporto corretto di nutrienti come pro-teina, energia e minerali. Altrettanto importan-te è assicurare un apporto adeguato di principiattivi antiossidanti come la vitamina E, il sele-nio, il rame, lo zinco e il manganese.In asciutta, la prevenzione delle ipocalcemie postpartum aiuta in modo determinante la contrat-tilità uterina e, quindi, la possibilità di un ra-pido svuotamento dei lochi e la riduzione delleinfezioni uterine.Le distocie e la gemellarità possono essere pre-venute quasi solo esclusivamente attraverso l’at-tenta scelta dei tori, sempre ricordando che la pre-disposizione alla gemellarità è caratteristica deglianimali di alto potenziale genetico, per l’aumen-to dell’incidenza della doppia ovulazione. Inoltre,per la prevenzione delle infezioni uterine è decisi-va l’igiene degli ambienti in cui le bovine partori-scono e delle attrezzature impiegate. Pertanto, igie-ne e precocità di intervento giocano un ruolo fon-damentale nel controllare l’insorgenza e nel fa-vorire la guarigione delle patologie uterine.Consigliabili sono le visite ginecologiche nel postpartum, tramite palpazione transrettale e im-piego del vaginoscopio, che dovrebbero essereidealmente realizzate a 1-3, 6-10, 14-21, 28-35giorni dopo il parto. Nei grandi allevamenti, ingenere, il veterinario concentra i suoi interven-ti in una settimana.Un altro fattore d’estrema importanza, è l’ado-zione di uno schema diagnostico che consentel’individuazione dell’endometrite subclinica a40-60 giorni dopo il parto. Per mitigare gli ef-fetti predisponenti e negativi della RFM, delledistocie e delle ipocalcemie è utile, dopo il par-to, la stimolazione della contrattilità uterina tra-mite la somministrazione di prostaglandine e os-sitocine. L’uso di questi principi attivi aiuta aprevenire il ristagno uterino dei lochi e, quindi,le infezioni conseguenti.È particolarmente importante combattere la pre-senza E. coli a livello uterino nel periodo postpartum, in quanto è in grado di favorire, dap-prima la presenza di A. pyogenes e, successi-vamente, di Gram- anaerobi.
L’antibiotico tradizionalmente utilizzato percombattere le infezione uterine è l’ossitetraci-clina (OTC), sebbene la sua somministrazionepresenti alcuni svantaggi; innanzitutto, questamolecola esercita un’azione irritante a livellouterino e, secondariamente, il suo frequente usonegli anni ha portato a un aumento della MIC,tanto che è necessario somministrarne 3-4 g peralmeno 3-5 giorni.Per la terapia antibiotica sistemica o locale, so-no utili le cefalosporine di terza o quarta gene-razione, in quanto caratterizzate da un breve tem-po di sospensione e MIC ridotta. Il trattamentosistemico con antibiotici è giustificato solo in pre-senza di metriti puerperali, quando il processo in-fettivo interessa tutto l’utero, preferibilmente inassociazione con antinfiammatori non steroideie, se necessario, un’adeguata terapia elettrolitica.Per il trattamento delle endometriti subcliniche èconsigliabile la cefapirina, associata a una pro-staglandina per via generale, già 20-30 giorni do-po il parto, mentre, in caso di metriti puerperali,sono stati ottenuti risultati più che soddisfacenticon il ceftiofur, una cefalosporina attiva contro iprincipali patogeni uterini.Molto importante è l’uso delle prostaglandine, nonsolo per l’effetto luteolitico che esercitano in casodi piometra ma, più in generale, per la loro azio-ne in caso di endometrite. In un lavoro del 2004,Melendez et al. hanno verificato l’effetto di unadoppia inoculazione di prostaglandine, a distanzadi 8 ore, 8 giorni dopo il parto, valutando la con-centrazione ematica di proteine nella fase acuta,come espressione dell’andamento dell’infiamma-zione uterina. I ricercatori hanno riscontrato uneffetto positivo del trattamento sulle performan-ces riproduttive degli animali trattati rispetto aglianimali di controllo [3].
◗ Le cisti ovariche Le cisti ovariche bovine (CO) o la degenerazio-ne cistica delle ovaie è una condizione patolo-gica risultante dalla mancata ovulazione dei fol-licoli pre-ovulatori. Le cisti possono insorgerein qualsiasi stadio della lattazione, anche se l’in-cidenza più alta si riscontra 40-150 giorni do-po il parto, nelle bovine adulte e nei mesi in-vernali. La produttività dei singoli soggetti sem-brerebbe rappresentare un fattore di rischio, an-che se questa ipotesi non trova un consenso una-nime. Secondo quanto riportato dagli studi dis-ponibili in letteratura, l’incidenza media delleCO varia dal 9 al 25%.Molti sono i fattori che alterano la comunica-zione ormonale dell’asse ipotalamo-ipofisi-ovaio,o meglio, che interferiscono sul feedback posi-tivo tra estradiolo follicolare e secrezione diGnRH. Tra i principali fattori predisponenti, sidevono annoverare i disordini metabolici, lostress, le infezioni uterine, le zoppie e una dis-creta predisposizione genetica.
Fertilità e patologie in bovini e suini
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201218
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 18

Dopo il parto, la bovina subisce profonde mo-dificazioni ormonali, caratterizzate da una ri-dotta produzione di insulina e IGF-1 e da un ca-lo, appena dopo il parto, della concentrazionedi estrogeni. Contemporaneamente, la bovinaacquisisce un inevitabile NEBAL, per lo squili-brio tra produzione e capacità di ingerire unaproporzionale quantità di nutrienti (vedere fi-gura 2).Tra i fattori causali delle CO, sembrerebbe ave-re un ruolo l’insulino-resistenza, derivante dauna continua sovrastimolazione, da parte del-l’alimento, della secrezione di insulina; questacondizione caratterizza quasi tutto il ciclo pro-duttivo della vacca da latte e, soprattutto, il pri-mo terzo della lattazione.Altri fattori causali sono la carenza di β-caro-tene, l’eccesso di selenio, alcune micotossine, lapresenza di sostanze estrogeno-simili, come al-cuni inquinanti ambientali, o i fitormoni pre-senti nelle leguminose. Inoltre, non deve esseretrascurato il ruolo rivestito dallo stress ossida-tivo. Dopo il parto e dopo l’espulsione degli in-vogli fetali, a causa della brusca caduta dellaconcentrazione di estrogeni circolanti, si assistea un aumento della produzione ipofisaria di FSHche, a sua volta, stimola la crescita follicolare,fin dalla prima settimana dopo il parto.Alle ondate follicolari segue la selezione del pri-mo follicolo dominante, in genere caratteriz-zato da una ridotta produzione di estrogeni; ilprimo follicolo non sarà quindi in grado di in-durre un’adeguata attività pulsatile per la pro-duzione ipofisaria di LH e, di conseguenza, dif-ficilmente potrà arrivare a piena maturazione ealla conseguente ovulazione. In questa fase, lascarsa secrezione di IGF-1 non consente la pie-na funzionalità del follicolo. Pertanto, la ridot-ta produzione di LH e IGF-1 dopo il parto, con-dizione fisiologica fortemente influenzata dal-
l’entità del NEBAL, non consente ai follicoli del-le prime ondate follicolari di giungere all’ovu-lazione. Inoltre, in questa fase molto precoce, ineuroni che rilasciano GnRH non esprimonoappieno i recettori per l’estradiolo. In condizioninormali, ma nella realtà poco riscontrabili, ilfeedback positivo estrogeni follicolari-neuroniipotalamici secernenti GnRH si osserva dopodue settimane e, sebbene l’ovulazione avven-ga, solo il 50% dei soggetti manifesta un com-portamento estrale. In genere, la prima ovula-zione avviene a livello dell’ovaio controlateraleal corno uterino che ha accolto il feto durantela precedente gravidanza.Al completamento dell’involuzione uterina, quin-di, oltre il 30° giorno post partum, la frequen-za dei cicli estrali e le ovulazioni tornano nor-mali, a meno che non intervengano situazionimetaboliche e/o patologiche. È bene ricordare,per meglio inquadrare le degenerazioni cistichedei follicoli, che la fase di sviluppo follicolare,in cui il follicolo possiede un diametro inferio-re a 3 mm, è indipendente dalle gonadotropi-ne ipofisarie, mentre il suo sviluppo dipende dal-l’FSH quando il diametro è pari a 3-10 mm. Successivamente, fino alla fase preovulatoria, lacrescita dei follicoli e la loro capacità di produrreormoni steroidei è condizionata dall’IGF-1 e dal-l’LH. In particolare, un’adeguata concentrazio-ne ematica di IGF-1 stimola sia la steroidogene-si follicolare sia la crescita stessa del follicolo,aumentando la sensibilità del follicolo dominanteall’FSH e all’LH. La maggior parte dei follicoliestrogenici e ovulatori emerge dopo il nadir delNEBAL. A questo punto, è fondamentale defi-nire cosa si intende per cisti ovarica.Per CO si intende un follicolo con un diametrosuperiore a 25 mm, che persiste sull’ovaio peroltre 10 giorni, in assenza di corpi lutei, o folli-coli multipli con un diametro >15 mm, che per-
Patologie ovariche-uterine e gravidanza
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201219
Figura 2. Modificazioni ormonali dopo il parto.
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 19

sistono per almeno 7 giorni. In ogni caso, qual-siasi follicolo con un diametro maggiore di 17mm può essere potenzialmente una CO. Le COnon sono strutture statiche, in quanto possonopersistere, luteinizzare o diventare atresiche.Le bovine con cisti hanno ondate follicolari chepossono portare all’ovulazione o alla forma-zione di altre cisti. La vacca cistica può esibire ninfomania o cicliestrali irregolari, ma l’anestro è il segno clini-co più comune.Uno dei problemi maggiori della vacca da latteHMG è la ripresa dell’attività ovarica dopo il par-to e, quindi, è importante differenziare le CO dal-l’anestro post partum: in questo periodo, la nu-trizione gioca un ruolo fondamentale. La visitaginecologica evidenzia la presenza di un singolofollicolo non estrogenico, di diametro <18 mm, edi pochi follicoli decisamente più piccoli, di dia-metro <8 mm. Lo scarso apporto di nutrienti, omeglio, la mancanza d’equilibrio nell’apporto dinutrienti alla base del NEBAL, si evidenzia conun basso BCS. Questo quadro metabolico è as-sociato a una scarsa secrezione di FSH (aspettomolto controverso) e a un’insufficiente produzionedi IGF-1 e LH. Le ovaie sono quindi piccole e losviluppo follicolare minimo, con conseguente ri-dotta produzione d’estradiolo; ne deriva l’assen-za d’ovulazione e del comportamento estrale.Dato che l’IGF-1 e il colesterolo, precursore degliormoni steroidei, sono prodotti dal fegato, la pie-na funzionalità di questo organo è di enorme im-portanza. Le bovine con lipidosi epatica, anche dimedia entità, riducono la produzione di coleste-rolo e IGF-1, con ripercussioni assolutamente ne-gative sullo sviluppo follicolare e sulla capacità d’o-vulare ed esibire un corretto comportamento estra-le. A differenza dell’anestro post partum, le COpossono formarsi in qualsiasi stadio della lattazio-ne, in seguito all’alterazione del feedback positivotra estradiolo e GnRH. In questa fase, lo stress puòrappresentare un fattore di rischio molto impor-
tante. Oltre allo stress comportamentale, un ruolosicuramente non trascurabile deve essere ricono-sciuto alle LPS prodotte dai Gram- presenti a livellodi utero, mammella e apparato gastroenterico.La presenza di LPS stimola la produzione di di-verse sostanze, come le interleuchine (TNF e IL-1), gli ormoni glucocorticoidi e le prostaglan-dine. Le interleuchine agiscono a livello ipota-lamico, inibendo la secrezione di GnRH e, con-seguentemente, di LH. Inoltre, le LPS possonointerferire direttamente a livello ipofisario sul-la secrezione di LH.Nella diagnostica delle cisti ovariche è impor-tante differenziare le cisti luteiniche dalle cistifollicoliniche (vedere foto 1 e 2), non tanto peri risvolti terapeutici, ma soprattutto profilatti-ci. La sensibilità e la specificità dell’esplorazio-ne rettale (ER) è molto bassa, mentre la dia-gnostica ecografica (USG) è più efficace per lecisti luteiniche rispetto alle cisti follicoliniche.L’USG, associata al dosaggio del progestero-ne, ha un’alta specificità e sensibilità per le cistiluteiniche, in quanto producono questo ormo-ne in quantità superiori alle cisti follicolari e in-feriori rispetto al corpo luteo.Con l’ER si può escludere la presenza del cor-po luteo e valutare la tonicità dell’utero, aspet-to particolarmente significativo ai fini della dia-gnosi di CO. Inoltre, l’USG è un valido aiutoper confermare l’assenza di un corpo luteo e de-terminare la taglia del follicolo, nonché il gra-do di luteinizzazione.La misurazione del livello di progesterone ser-ve per determinare il grado di luteinizzazionedella cisti.Un’altra diagnosi differenziale di estrema im-portanza è quella tra cisti luteiniche e corpo lu-teo. La cisti luteinica è una patologia ovaricaderivante dai follicoli che, pur non avendo ovu-lato, vanno incontro a luteinizzazone e presen-tano una cavità interna in concomitanza con losviluppo del corpo luteo. All’esplorazione ret-
Fertilità e patologie in bovini e suini
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201220
Le pareti dell'utero e delle corna uterine sono composte da tre strati: sierosa (strato ester-no), strato muscolare (strato intermedio) e mucosa (stato interno). La componente mu-scolare, o miometrio, è costituita da due strati di muscolatura liscia, uno esterno longi-tudinale e uno interno trasversale. Tra i due strati, si trovano i vasi e le cellule nervose.Queste ultime sono stimolate dagli estrogeni e dall'ossitocina, in grado di conferire toni-cità e stimolare la contrattilità uterina.Vi è poi l’endometrio, struttura anatomica costituita da un epitelio ghiandolare e una mu-cosa sovrastante il miometrio. Nella mucosa uterina, è possibile individuare tre compo-nenti: l'epitelio superficiale, il corion e le ghiandole uterine. Sotto lo strato superficialetroviamo il corion, una struttura costituita da fibre collagene con cellule reticolari, all'in-terno della quale sono ospitati linfociti, neutrofili e plasmacellule.Le ghiandole uterine si trovano tra gli spazi intercotiledonali, come invaginazioni dell'e-pitelio, e la loro funzione è secernere abbondanti quantità di muco durante il calore, uti-le alla nutrizione, al transito e alla capacitazione degli spermatozoi. Inoltre, durante lafase luteinica, le ghiandole uterine secernono un muco idoneo alla nutrizione dell'em-brione, prima del suo attecchimento.
Richiami anatomici
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 20

tale, il corpo luteo presenta una linea di de-marcazione e una netta distorsione sulla lineadell’ovaio. L’esplorazione rettale ha una bassaspecificità e sensibilità per individuare la lutei-nizzazione del follicolo, ma il problema è ri-solvibile con l’adozione della USG.Molto più importante della diagnosi differenzia-le tra cisti follicoliniche e luteiniche è la distinzio-ne tra cisti ovariche e follicolo preovulatorio (ve-dere foto 3). La diagnosi differenziale si basa es-senzialmente sul numero e la taglia dei follicoli e,soprattutto, sull’assenza di tonicità uterina. Per de-finire alla ER le CO, si parla di follicoli multiplipiù grandi del normale, associati a un utero flac-cido. Le bovine, nella fase di proestro hanno giàun utero tonico e turgido a causa dell’ossitocinaprodotta a seguito della luteolisi del corpo luteodel ciclo precedente. È comunque bene sottolineareche le CO sono diagnosticate anche dopo il “fal-limento” dell’ovulazione e che, in questo periodo,l’utero risulta flaccido e poco tonico.L’USG è un valido aiuto per la determinazio-ne del numero dei follicoli e la loro taglia. Per-tanto, per cisti ovarica, si intende la presenza,a livello ovarico, di follicoli multipli con un dia-metro di 18-20 mm, l’assenza di corpi lutei as-sociati, la presenza di scarsa tonicità uterina ecalori assenti o molto irregolari. La caratteri-stica distintiva è la persistenza; quindi, è indi-spensabile la conferma diagnostica dopo 7-10giorni.
◗ Principi di terapia Molte CO, che si sviluppano nell’immediatopost partum, regrediscono e il 20% circa dellebovine riprende spontaneamente la normale ci-clicità follicolare dopo un episodio di CO. Se-condo alcuni autori, non è consigliabile inter-venire farmacologicamente o meccanicamantesu cisti diagnosticate prima del 50° giorno postpartum, anche se questa opinione non trova unconsenso unanime. La rottura manuale delle COè una pratica sconsigliabile, in quanto può es-sere causa di emorragie e determinare la for-mazione di aderenze. La terapia si basa sullasomministrazione di sostanze in grado di sti-molare il rilascio di un’adeguata quantità di LHe di indurre la lisi del corpo luteo.I principi attivi maggiormente impiegati sono ilGnRH, le prostaglandine, il progesterone e l’HCG.Il GnRH stimola il rilascio di LH ipofisario e, per-tanto, la rottura della CO e la luteinizzazione del-le cellule della teca interna, con conseguente sin-tesi di progesterone (P4). Frequente anche l’ado-zione di una dose unica di PGF2α ad attività lu-teolitica, seguita da GnRH, dopo 7-14 giorni.Complementare a queste tecniche è l’uso di pro-gesterone somministrato con Prid (Progesteronereleasing intravaginal device) e rimosso dopo unasettimana dall’impianto, per indurre un bloccodell’asse ipotalamo-ipofisi-ovaie e la successivamassiccia liberazione di LH.
Patologie ovariche-uterine e gravidanza
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201221
Foto 1. Cisti follicolinica. Foto 2. Cisti luteinica.
Foto 3. Utero con ovaie cistiche.
PER SAPERNE DI PIÙ
• LeBlanc S.J., Duffield T.F., Leslie K.E., Ba-teman K.G., Keefe G.P., Walton J.S., John-son W.H. (2002): Defining and diagnosingpostpartum clinical endometritis and its im-pact on reproductive performance in dairycows. J. Dairy Sci.; 85(9):2223-2236.
• Gilbert R.O., Shin S.T., Guard C.L., Erb H.N.,Frajblat M. (2005): Prevalence of endometritis
and its effects on reproductive performance ofdairy cows. Theriogenology; 64(9):1879-1888.
• Melendez P. McHale J., Bartolome. J,Archbald L.F., Donovan G.A. (2004): Uterineinvolution and fertility of holstein cows sub-sequent to early postpartum PGF2alpha treat-ment for acute puerperal metritis. J. Dairy Sci.;87(10):3238-3246.
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 21

ome sottolineato da diversi autori, il70% dei fallimenti riproduttivi all’in-terno delle aziende suinicole è dovu-to a cause di origine non infettiva e,nella maggioranza dei casi, questi si
concentrano nel periodo estivo-autunnale (Muir-head M.R., et al., 1997). Gli aborti propriamente detti sono quelli che av-vengono oltre il 35° giorno di gestazione e che pre-vedono l’espulsione di un feto già ossificato. Per leinterruzioni di gravidanza antecedenti al 34° gior-no, è preferibile invece parlare di riassorbimenti,che si evidenziano con l’espulsione da parte dellascrofa di piccole placente che al loro interno cela-no un embrione spesso visibile (vedere foto 1). Anche se in entrambi i casi si tratta di un conce-pimento senza mantenimento della gravidanza,da un punto di vista zootecnico questa differenzarisulta molto importante. Un riassorbimento av-venuto senza l’intervento di sintomi clinici impu-tabili ad agenti patogeni generalmente non pre-clude il successo nell’inseminazione seguente. Nel caso di un aborto intercorso senza l’inter-vento presunto di patogeni, risulta sempre diprimaria importanza verificare esattamente ilperiodo della gestazione in cui esso si è manife-
stato. Mentre per un aborto tardivo è preferi-bile far saltare l’imminente estro della scrofa,nel caso di un aborto precoce (alcuni giorni do-po l’ossificazione dei feti) vi sono maggiori pro-babilità di successo all’inseminazione successi-va. Ovviamente tutto ciò non sempre viene con-fermato dalla pratica, ma si verifica nella mag-gioranza dei casi e si spiega col permettere al-l’utero di recuperare le sue fisiologiche dimen-sioni al momento della fecondazione. I riassorbimenti non sempre sono ben visibili inazienda, soprattutto per la presenza dei grigliati,ma è possibile comunque sostenere che rappre-sentino la causa principale dei ritorni fuori ciclofra i 25 e i 38 giorni post inseminazione. Lo stru-mento migliore per confermare una diagnosi diriassorbimento risulta certamente l’ecografo che,tra i 20 e i 28 giorni dalla fecondazione, permet-te di individuare quadri inconfondibili.
◗ La sindrome dell’infertilitàstagionale (SIS)
Il convincimento maturato nel corso degli an-ni è quello di un aumento dei quadri ecogra-fici da riassorbimento durante i mesi estivi
Il complesso dell’infertilitàestiva nella scrofa
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201222
Claudio Mazzoni1, Valeria Raffi2, Chiara Avanzini3, Chiara Piancastelli21Veterinario libero professionista. Suivet - Reggio Emilia2Veterinario libero professionista.Suivet - Parma3Veterinario libero professionista. Suivet - Cremona
C
Foto 1. Riassorbimento. Placenta al cui interno risulta visibile un embrione.
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 22

e autunnali, particolarmente da luglio a ot-tobre, a cui corrisponde un amento dei ritornifuori ciclo. In realtà nulla di nuovo poiché,a un’indagine approfondita, questi riassor-bimenti, uniti ai ritorni fuori ciclo, sono laprova che ci troviamo nel contesto della sin-drome dell’infertilità stagionale (SIS), un tem-po nota come “ipofertilità estiva”.La SIS può essere suddivisa in complesso del-l’infertilità estiva (SIC) e sindrome degli abor-ti autunnali (AAS) in base ai sintomi e al pe-riodo di comparsa (Levis D.G., 2001).La SIC inizia solitamente verso la fine del me-se di giugno con la comparsa dei primi forti ri-alzi di temperatura, per esaurirsi verso la metàdi settembre; qui lascia il campo alla sindromedegli aborti autunnali (AAS) che, pur colpendoil trimestre settembre-ottobre-novembre con lesue notevoli fluttuazioni di temperature, vieneconsiderata parte integrante della SIS (vedere ta-bella 1 – modificata - Martineau, 2003). Le aziende affette da questa sindrome de-nunciano una diminuzione della messa in par-to compresa fra il 5 e il 20% (Mechler D.W.2002).
◗ Complesso dell’infertilitàestiva (SIC)
I cambiamenti climatici tipici della stagione esti-va sono poco compatibili con la sfera riprodut-tiva delle scrofe che, derivando dal suino selvati-co Europeo, hanno ereditato una certa predi-sposizione all’anestro stagionale nel trimestre lu-glio-agosto-settembre (Mauget, 1982). Tra le cau-se di questa situazione certamente gioca un ruo-lo fondamentale una lattazione a cui va incontrola scrofa, che in natura è protratta per oltre tremesi e a cui fa seguito uno svezzamento gradua-le nel tempo. Anche una disponibilità altalenan-te di alimenti con scarso valore nutritivo e la strut-tura sociale del suino selvatico, che limita forte-mente i contatti con il maschio al di fuori dellastagione degli accoppiamenti, risultano esserecause predisponenti all’anestro stagionale. L’avvento delle pratiche più moderne di ma-nagement quali lo svezzamento precoce, ladisponibilità di alimenti con elevato valorenutritivo e un appropriato contatto con il ver-ro hanno permesso di superare, nella stra-grande maggioranza dei casi, quei paletti chela natura aveva posto da tempi immemora-bili. Tuttavia non per tutte le scrofe vale que-
sto discorso, per alcune infatti, definite “scro-fe suscettibili”, le interazioni fra l’alimentazio-ne e la stabulazione sono aspetti insuperabili chescarsamente conciliano l’impianto di una gra-vidanza o il suo mantenimento.Alla base di tutti i concetti sopra indicati ci so-no complessi meccanismi ormonali regolati al-l’origine dall’alimentazione e in particolare dal-l’apporto di energia e nutrienti durante il pe-riodo della lattazione. Questa problematica vie-ne prevalentemente registrata nel periodo esti-vo, ove l’assunzione di alimento da parte dellescrofe in sala parto viene fortemente compro-messa dalle alte temperature raggiunte. Le conseguenze provocate dall’ipofertilità esti-va sono maggiormente evidenti nelle scrofette enelle primipare le quali, oltre a dover produrregli ormoni per l’attività riproduttiva, devono nel-lo stesso tempo continuare a secernere anche quel-li indispensabili per la propria crescita. Questofatto si ripercuote sulla sfera riproduttiva stessadelle scrofette e delle primipare in quanto l’or-mone della crescita è in grado di esprimere unaazione inibitoria su quelli preposti alla riprodu-zione. Inoltre, le scrofette, avendo una capacitàgastrica decisamente più contenuta rispetto allescrofe adulte, hanno un ostacolo in più da su-perare relativamente ai volumi di ingestione. Èpossibile sostenere quindi che, negli animali gio-vani, i meccanismi di crescita corporea e di ve-nuta in calore interferiscano in maniera piutto-sto negativa uno nei confronti dell’altro. Le manifestazioni cliniche dell’infertilità estivasono rappresentate da pubertà ritardata (Pelto-niemi O.A.T. et al. 1999), allungamento del-l’intervallo svezzamento-calore e da una ridu-zione della messa in parto (Ashworth C. 2006).Risulta importante sottolineare che gli effetti sulpost-svezzamento sono maggiormente visibilisulle primipare. Per quanto riguarda le scrofepluripare, l’evento più importante è rappresen-tato dall’incapacità di mantenere la gravidanzaassociato a un aumento di ritorni in ciclo e fuo-ri ciclo, soprattutto se paragonato agli altri me-si dell’anno. Da segnalare anche un incrementodelle pseudogravidanze (Kirkwood, 2003).Generalmente i casi di riassorbimento avven-gono senza una sintomatologia apparente, per-tanto non vengono evidenziati segni clinici qua-li febbre o anoressia ma si assiste solitamentealla perdita della gravidanza seguita da un ri-torno in calore fuori ciclo che risulta normal-mente fertile.
Il complesso dell’infertilità estiva nella scrofa
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201223
Tab. 1. Distribuzione del complesso dell’infertilità estiva (con le alte temperature) e della sindromedegli aborti autunnali (con le fluttuazioni delle temperature) nel corso dell’anno (Martineau, 2003).
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 23
È bene ricordare che i ritorni in ciclo sono quel-li fra i 18 e i 24 giorni (oltre che quelli fra i 39e i 44) dall’inseminazione, mentre vengono con-siderati ritorni fuori ciclo quelli che si presen-tano fra i 25 e 38 giorni post inseminazione.Poiché la problematica della SIC risulta essereben nota e radicata nelle nostre scrofaie, sareb-be molto utile tenere sempre aggiornati i dati ri-produttivi delle scrofe e possedere una sorta di“storia aziendale” per poter confrontare la fer-tilità nei diversi periodi dell’anno.Un esempio molto semplice potrebbe essere l’u-tilizzo di una tabella come quella riportata diseguito (vedere tabella 2).
◗ Caso pratico: l’ipofertilitàestiva (SIC) in azienda
Attraverso l’utilizzo della tabella precedente-mente esposta, e al fine di riportare un esempiopratico di quello che può accadere a causa del-la SIC, si sono voluti usare i dati di un alleva-mento a ciclo aperto composto da 600 scrofe si-to nella pianura padana rilevati durante l’an-no 2006 (vedere tabella 3).Da questa tabella sono state volutamente esclu-se altre problematiche riproduttive dell’aziendache hanno inciso sulla fertilità complessiva (zop-pie, scrofe in anestro e metriti). Sono stati indi-cati fra parentesi i ritorni, in ciclo e fuori ciclo,relativi al solo mese di gennaio e già compresinei rispettivi totali, conseguenti a una brutalecircolazione del virus della PRRS, caratterizza-ta da una tempesta di aborti e mortalità nellescrofe, iniziata già nel mese di dicembre 2005 econclusasi verso la fine di gennaio 2006. Da una veloce osservazione dei dati della tabella,è possibile subito notare che sui primi 9 mesidell’anno considerati, il 43,5% (57/131) deiritorni totali è avvenuto nel solo trimestre lu-glio-settembre. Tuttavia, il dato più interessan-te sembra essere quello relativo ai ritorni fuoriciclo dello stesso periodo che rappresentano il54,8% (34/62) dei totali e che sono il problemaprincipale causato dalla SIC nel caso di questa
azienda. Nel caso qui riportato, se dai dati ven-gono tolti i ritorni in ciclo e fuori ciclo causatidal focolaio di PRRS evidenziato a gennaio (30in tutto), è possibile avere una reale idea del dan-no economico causato dalla SIC. Ovviamenteprendere in esame un solo anno dello storicoaziendale non può essere considerato come suf-ficiente per confermare la problematica da SIC,pertanto sarebbe molto utile possedere un’a-namnesi remota inerente alla fertilità. Nel gra-fico 1 e nella tabella 4 (vedere a pag. 26) sonoriportate le percentuali di fertilità registrate al-l’ecografia effettuata entro i 35 giorni dall’in-seminazione (anni 2001-2006).Sono stati volutamente evidenziati in arancionei mesi nei quali la percentuale di fertilità è sce-sa sotto l’80%, valore che viene considerato co-me soglia fra il dato positivo (> 80%) e quellonegativo (< 80%). È facile notare la concen-trazione di aree in arancione nei mesi estivo-au-tunnali, con un’incidenza di 7 mesi sui 18 tota-li (38%) nel solo trimestre luglio-agosto-set-tembre. Inoltre, è da sottolineare una generaletendenza al ribasso della fertilità in questi me-si rispetto ai mesi invernali e primaverili.Per ovviare alle problematiche indotte dallaSIC, già a partire dal 2005 e negli anni suc-cessivi, l’azienda ha messo in atto alcune mi-sure preventive. Il punto di partenza nel 2005è stato quello di migliorare il comfort am-bientale degli animali in gestazione tramite l’im-piego di refrigeratori (tipo cooling) uniforme-mente disposti in tutti i settori della gestazio-ne. Nel 2006, oltre alla riduzione di un 10%del parco scrofe, una nuova e razionale dis-tribuzione del personale in azienda e l’arrivodi un nuovo tecnico in gestazione ha permes-so di proseguire in una graduale ricerca dellasoluzione al problema. È da sottolineare chetre delle otto sale parto erano munite da diversianni di refrigeratori che, anche se non perfet-tamente funzionanti, avevano dato il loro con-tributo; infatti, per il dato della mortalità sul-le scrofe in sala parto, imputabile al cosiddet-to “colpo di caldo”, è stata registrata in que-
Fertilità e patologie in bovini e suini
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201224
MESI FECONDAZIONI TOTALI RITORNI IN CICLO (18-24 GIORNI) RITORNI FUORI CICLO (25-38 GIORNI)
Gennaio-GiugnoLuglio-SettembreTotali
Tab. 2.
MESI FECONDAZIONI TOTALI RITORNI IN CICLO (18-24 GIORNI) RITORNI FUORI CICLO (25-38 GIORNI)
Gennaio-Giugno 2006 915 46 (15) 28 (15)Luglio-Settembre 450 23 34Totali 1365 69 62
Tab. 3.
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 24
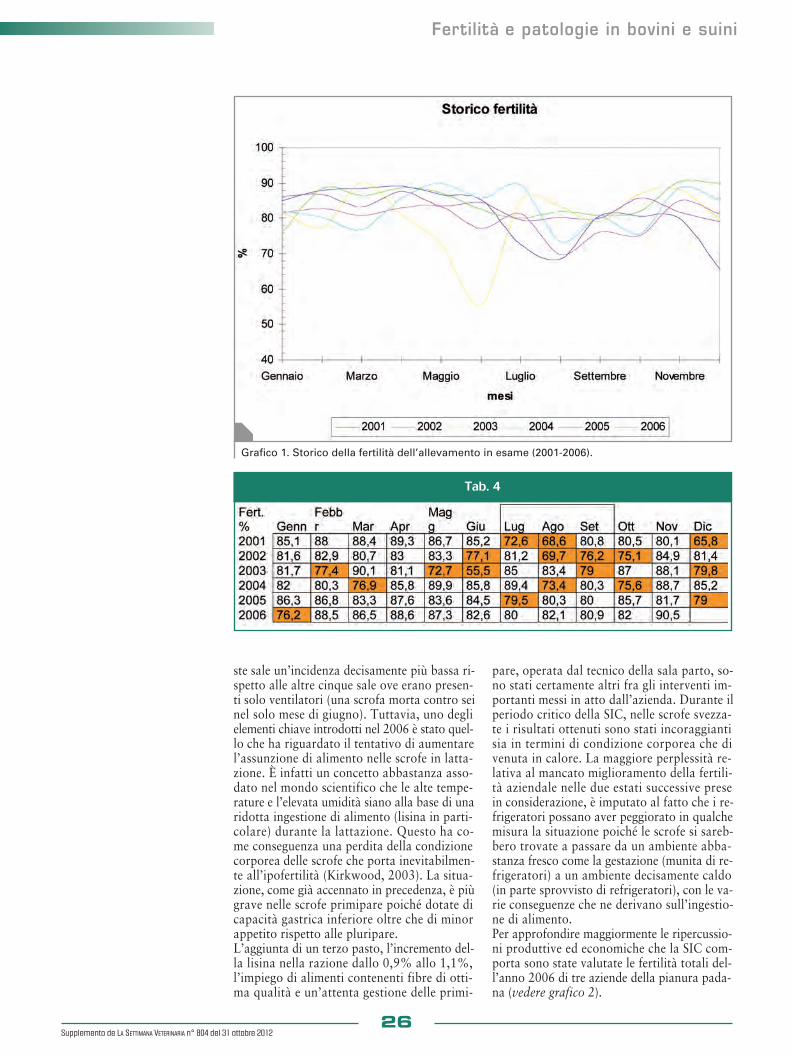
ste sale un’incidenza decisamente più bassa ri-spetto alle altre cinque sale ove erano presen-ti solo ventilatori (una scrofa morta contro seinel solo mese di giugno). Tuttavia, uno deglielementi chiave introdotti nel 2006 è stato quel-lo che ha riguardato il tentativo di aumentarel’assunzione di alimento nelle scrofe in latta-zione. È infatti un concetto abbastanza asso-dato nel mondo scientifico che le alte tempe-rature e l’elevata umidità siano alla base di unaridotta ingestione di alimento (lisina in parti-colare) durante la lattazione. Questo ha co-me conseguenza una perdita della condizionecorporea delle scrofe che porta inevitabilmen-te all’ipofertilità (Kirkwood, 2003). La situa-zione, come già accennato in precedenza, è piùgrave nelle scrofe primipare poiché dotate dicapacità gastrica inferiore oltre che di minorappetito rispetto alle pluripare.L’aggiunta di un terzo pasto, l’incremento del-la lisina nella razione dallo 0,9% allo 1,1%,l’impiego di alimenti contenenti fibre di otti-ma qualità e un’attenta gestione delle primi-
pare, operata dal tecnico della sala parto, so-no stati certamente altri fra gli interventi im-portanti messi in atto dall’azienda. Durante ilperiodo critico della SIC, nelle scrofe svezza-te i risultati ottenuti sono stati incoraggiantisia in termini di condizione corporea che divenuta in calore. La maggiore perplessità re-lativa al mancato miglioramento della fertili-tà aziendale nelle due estati successive presein considerazione, è imputato al fatto che i re-frigeratori possano aver peggiorato in qualchemisura la situazione poiché le scrofe si sareb-bero trovate a passare da un ambiente abba-stanza fresco come la gestazione (munita di re-frigeratori) a un ambiente decisamente caldo(in parte sprovvisto di refrigeratori), con le va-rie conseguenze che ne derivano sull’ingestio-ne di alimento.Per approfondire maggiormente le ripercussio-ni produttive ed economiche che la SIC com-porta sono state valutate le fertilità totali del-l’anno 2006 di tre aziende della pianura pada-na (vedere grafico 2).
Fertilità e patologie in bovini e suini
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201226
Grafico 1. Storico della fertilità dell’allevamento in esame (2001-2006).
Tab. 4
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 26
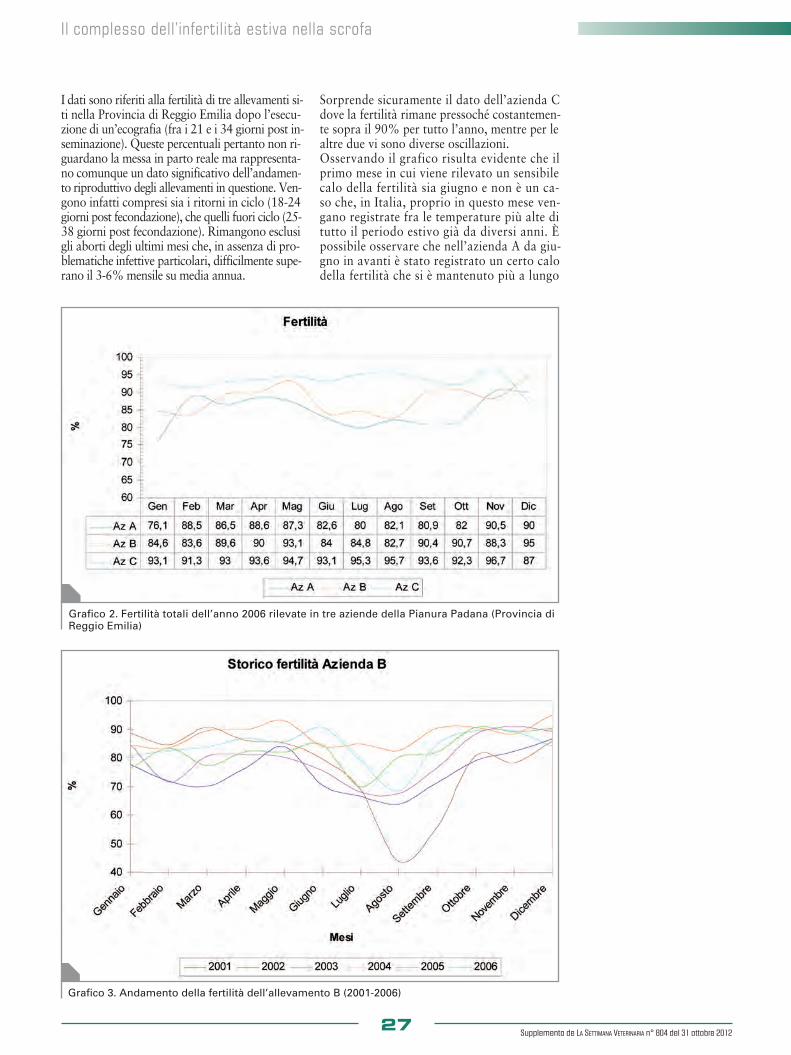
I dati sono riferiti alla fertilità di tre allevamenti si-ti nella Provincia di Reggio Emilia dopo l’esecu-zione di un’ecografia (fra i 21 e i 34 giorni post in-seminazione). Queste percentuali pertanto non ri-guardano la messa in parto reale ma rappresenta-no comunque un dato significativo dell’andamen-to riproduttivo degli allevamenti in questione. Ven-gono infatti compresi sia i ritorni in ciclo (18-24giorni post fecondazione), che quelli fuori ciclo (25-38 giorni post fecondazione). Rimangono esclusigli aborti degli ultimi mesi che, in assenza di pro-blematiche infettive particolari, difficilmente supe-rano il 3-6% mensile su media annua.
Sorprende sicuramente il dato dell’azienda Cdove la fertilità rimane pressoché costantemen-te sopra il 90% per tutto l’anno, mentre per lealtre due vi sono diverse oscillazioni. Osservando il grafico risulta evidente che ilprimo mese in cui viene rilevato un sensibilecalo della fertilità sia giugno e non è un ca-so che, in Italia, proprio in questo mese ven-gano registrate fra le temperature più alte ditutto il periodo estivo già da diversi anni. Èpossibile osservare che nell’azienda A da giu-gno in avanti è stato registrato un certo calodella fertilità che si è mantenuto più a lungo
Il complesso dell’infertilità estiva nella scrofa
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201227
Grafico 2. Fertilità totali dell’anno 2006 rilevate in tre aziende della Pianura Padana (Provincia diReggio Emilia)
Grafico 3. Andamento della fertilità dell’allevamento B (2001-2006)
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 27
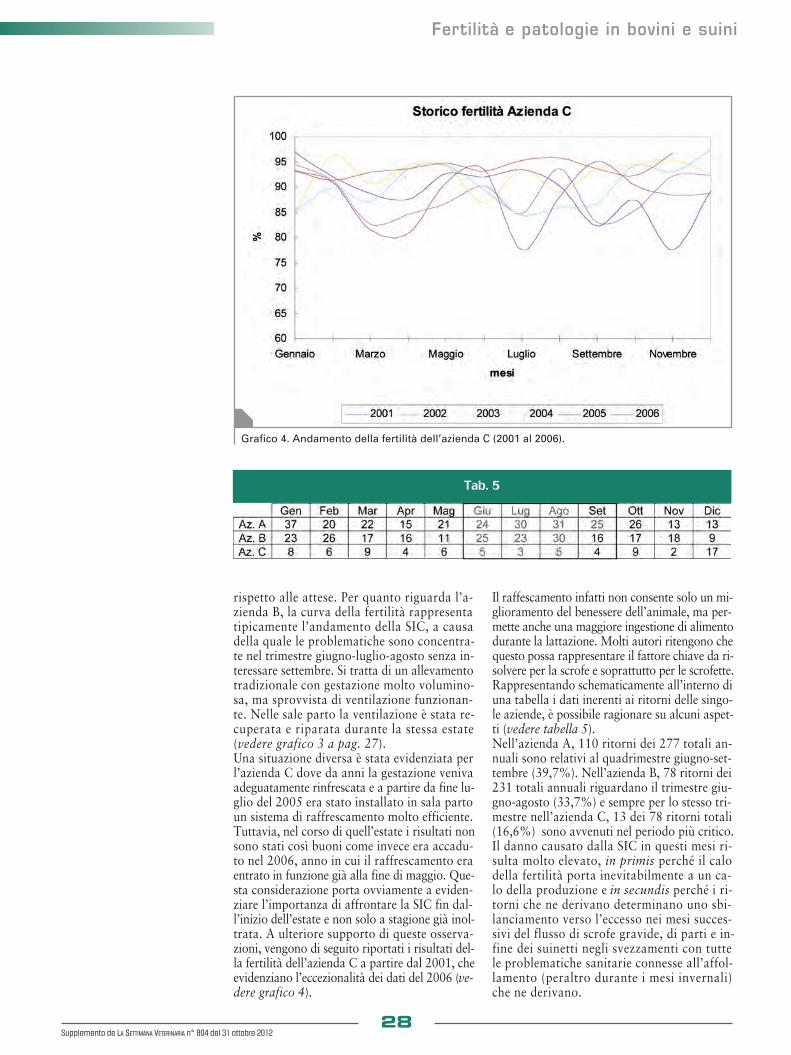
rispetto alle attese. Per quanto riguarda l’a-zienda B, la curva della fertilità rappresentatipicamente l’andamento della SIC, a causadella quale le problematiche sono concentra-te nel trimestre giugno-luglio-agosto senza in-teressare settembre. Si tratta di un allevamentotradizionale con gestazione molto volumino-sa, ma sprovvista di ventilazione funzionan-te. Nelle sale parto la ventilazione è stata re-cuperata e riparata durante la stessa estate(vedere grafico 3 a pag. 27).Una situazione diversa è stata evidenziata perl’azienda C dove da anni la gestazione venivaadeguatamente rinfrescata e a partire da fine lu-glio del 2005 era stato installato in sala partoun sistema di raffrescamento molto efficiente.Tuttavia, nel corso di quell’estate i risultati nonsono stati così buoni come invece era accadu-to nel 2006, anno in cui il raffrescamento eraentrato in funzione già alla fine di maggio. Que-sta considerazione porta ovviamente a eviden-ziare l’importanza di affrontare la SIC fin dal-l’inizio dell’estate e non solo a stagione già inol-trata. A ulteriore supporto di queste osserva-zioni, vengono di seguito riportati i risultati del-la fertilità dell’azienda C a partire dal 2001, cheevidenziano l’eccezionalità dei dati del 2006 (ve-dere grafico 4).
Il raffescamento infatti non consente solo un mi-glioramento del benessere dell’animale, ma per-mette anche una maggiore ingestione di alimentodurante la lattazione. Molti autori ritengono chequesto possa rappresentare il fattore chiave da ri-solvere per la scrofe e soprattutto per le scrofette. Rappresentando schematicamente all’interno diuna tabella i dati inerenti ai ritorni delle singo-le aziende, è possibile ragionare su alcuni aspet-ti (vedere tabella 5). Nell’azienda A, 110 ritorni dei 277 totali an-nuali sono relativi al quadrimestre giugno-set-tembre (39,7%). Nell’azienda B, 78 ritorni dei231 totali annuali riguardano il trimestre giu-gno-agosto (33,7%) e sempre per lo stesso tri-mestre nell’azienda C, 13 dei 78 ritorni totali(16,6%) sono avvenuti nel periodo più critico. Il danno causato dalla SIC in questi mesi ri-sulta molto elevato, in primis perché il calodella fertilità porta inevitabilmente a un ca-lo della produzione e in secundis perché i ri-torni che ne derivano determinano uno sbi-lanciamento verso l’eccesso nei mesi succes-sivi del flusso di scrofe gravide, di parti e in-fine dei suinetti negli svezzamenti con tuttele problematiche sanitarie connesse all’affol-lamento (peraltro durante i mesi invernali)che ne derivano.
Fertilità e patologie in bovini e suini
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201228
Grafico 4. Andamento della fertilità dell’azienda C (2001 al 2006).
Tab. 5
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 28

◗ Sindrome degli abortiautunnali (AAS)
La sindrome degli aborti autunnali (AAS) rap-presenta la parte terminale dell’insieme di tur-be riproduttive che vengono raggruppate nelcomplesso dell’infertilità stagionale, di cui laAAS è parte integrante. Nello specifico, con ladicitura sindrome degli aborti autunnali si in-tendono quelle interruzioni di gravidanza, mol-to spesso senza apparente spiegazione, tipichedella stagione autunnale. Come causa scate-nante di questi aborti non vengono coinvoltiagenti patogeni, ma da più parti sono chiamatiin causa fattori relativi al metabolismo del-l’animale e fattori manageriali. Molto spessogli aborti decorrono senza una sintomatolo-gia clinica di riferimento (febbre e anoressia)tanto che vengono individuati tardivamente(aumento delle “vuote al parto”). La problematica degli aborti autunnali è spessoinspiegabile. Può manifestarsi all’interno di unallevamento durante un anno per poi non es-sere evidenziata nei due anni successivi e infi-ne ripresentarsi nei tre anni seguenti.All’interno dello stesso gruppo di aziende è pos-sibile osservarne alcune che, pur avendo la stes-sa genetica, lo stesso sistema di alimentazione,lo stesso mangime, i medesimi sistemi di con-trollo ambientale e design strutturale non han-no mai evidenziato questo problema rispetto adaltre colpite annualmente.Lo stadio della gestazione in cui si eviden-ziano gli aborti è piuttosto variabile, spa-ziando dal 35° al 110° giorno. Tuttavia, lamaggioranza dei casi è individuabile fra le5 e le 7 settimane post inseminazione. Comeconsolidato anche nella pratica, è evidenteuna certa stagionalità di questi aborti, in par-ticolare da settembre a novembre. È pertan-to possibile ipotizzare che queste scrofe sia-no state fecondate da giugno, ovvero all’i-nizio del complesso dell’infertilità estiva(Martineau, 2003).Una possibile spiegazione a questi eventi può es-sere fornita dalla “teoria dell’LH”. L’LH (ormone luteinizzante) viene prodotto dal-l’ipofisi e possiede una fondamentale azione diprotezione del corpo luteo (CL). Il CL è unastruttura dell’ovaio che compare dopo l’ovula-zione ed è preposto alla produzione del proge-sterone (P4), ormone che, in caso di avvenutoconcepimento, è deputato al mantenimento del-la gravidanza. A differenza di altre specie dimammiferi, l’unica fonte rilevante di progeste-rone per la scrofa gravida è il CL e per questomotivo è facilmente intuibile che qualsiasi even-to in grado di influenzare negativamente la con-centrazione di LH circolante nel siero ematicodella scrofa potrà interferire sull’andamento del-la gestazione (Kirkwood, 2002). Se i livelli diLH circolante si abbassano troppo, il CL sarà
poco protetto e maggiormente esposto a fatto-ri diversi in grado di determinarne la rottura (li-si del corpo luteo) con conseguente crollo delprogesterone circolante e relativa perdita dellagravidanza.La AAS ha ancora oggi numerose caratteristi-che non del tutto chiarite. La fisiopatologia diquesto processo, collegata alla lisi del CL, è damettere in relazione a un evento stressante e nontutti gli autori sono concordi sulla teoria del-l’LH. La parola “stress” in questo caso permettedi capire che alcune scrofe sono più suscettibi-li di altre a questo processo (Martineau, 2003).Il concetto di suscettibilità individuale, già evi-denziato nella SIC, anche in questo caso vienesottolineato per la sua importanza e vuole ri-badire la predisposizione individuale di alcunescrofe verso fattori, definiti stressanti, in gra-do di indurre turbe riproduttive.I “fattori stressanti” sono decisamente molti (ve-dere figura 1 a pag. 30). Alcuni studi hanno evidenziato livelli più bassi diLH e progesterone proprio in concomitanza del-l’estate e dell’autunno; le alte temperature sonopoi responsabili della riduzione dei livelli emati-ci di LH. Inoltre, i bassi livelli energetici forniticon l’alimento durante la prima fase della ge-stazione, allo scopo di ridurre i riassorbimenti fe-tali, sono in grado, a loro volta, di inibire in ma-niera consistente la secrezione di LH. In questocontesto si inseriscono le variazioni di tempera-tura giorno/notte (caldo/freddo) e l’accorciamentodelle ore di luce della giornata (tipici dell’autun-no) che vengono da molti associate all’AAS. Perquanto riguarda gli sbalzi di temperatura è da os-servare che si verificano proprio in concomitan-za di un periodo in cui sia l’LH che il progeste-rone vengono secreti già in quantità ridotte, quin-di l’interazione fra alta temperatura e i bassi li-velli di energia assunti con l’alimento possonodare esiti più gravi in autunno che non nelle al-tre stagioni dell’anno dove il freddo non inibiscel’LH. Per quanto riguarda la riduzione delle oredi luce della giornata (spesso a partire da set-tembre), viene chiamato in causa un altro ormo-ne: la melatonina. Questa viene secreta dall’epi-fisi e la sua produzione viene incrementata conl’accorciarsi delle ore di luce. Anche le restrizio-ni alimentari causano un incremento della mela-tonina circolante e ciò risulta evidente in estatema non in inverno. Su questo argomento gli au-tori non sembrano essere in accordo anche se lamaggior parte di essi è giunto alla conclusioneche la melatonina possa essere un fattore in gra-do di contribuire alla patogenesi della AAS.In ogni caso, da qualunque parte venga osser-vata la sindrome SIC-AAS, il dato ricorrentein tutti i lavori riguarda lo scarso apporto ener-getico causato dalla limitata ingestione di ali-mento soprattutto in sala parto, ma anche in ge-stazione, a causa dell’eccessivo caldo in estate.Non è inoltre da sottovalutare l’incremento dei
Il complesso dell’infertilità estiva nella scrofa
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201229
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 29

fabbisogni energetici causati dall’affanno a cuivanno in contro le scrofe, nel tentativo di com-pensare l’eccesso di calore, così come impor-tante è il ruolo dello squilibrio termico gior-no/notte in autunno, specialmente durante la fa-se di inizio gestazione e specialmente in gabbia. In conclusione, e alla luce delle osservazionisinora fatte, si potrebbe ipotizzare per la SIS unapatogenesi da “accumulo”, nella quale, rag-giunto gradualmente e superato il “livello dicompensazione ambientale” della scrofa, le “rot-
ture” di alcuni soggetti si manifestano con lacomparsa dei disordini riproduttivi conosciuti. Verrebbe così a prendere corpo la proposta diKirkwood, il quale suggerisce l’esistenza di sot-topopolazioni di scrofe particolarmente sensi-bili allo stress termico. Più precisamente, sarebbel’integrità dei corpi lutei di queste scrofe ad es-sere intaccata, secondo modalità ancora nonchiarite, con l’effetto finale dell’interruzione del-la gravidanza (Kirkwood R. 2002, 2003). Que-sta genesi endocrina concorda con il quadro cli-nico, che spesso vede le scrofe in corso di rias-sorbimento/aborto normotermiche e con appe-tito conservato, rendendo poco percorribile l’i-potesi infettiva così spesso evocata (Levis D.G.2001, Martineau G.P. 2003).I meccanismi fisiologici alla base della SIC tut-tavia non sono ancora stati perfettamente chia-riti e ancora molto deve essere studiato sul ruo-lo svolto da certi ormoni e sulle loro intera-zioni. Certamente questa problematica si ma-nifesta su quelle “scrofe suscettibili” che rap-presentano una minoranza, ma che, quandoraggiungono i numeri che spesso osserviamonelle nostre realtà della pianura, non possonoessere trascurate. Quadri clinici come la SIS, che rientrano nelgrande capitolo della patologia multifattoria-le condizionata, saranno diagnosticabili piùfacilmente incrociando i dati produttivi d’al-levamento con i sintomi raccolti all’esame cli-nico, e potranno essere contrastati efficace-mente solo con un approccio di tipo preven-tivo, che metta in risalto, azienda per azienda,i vari fattori di rischio presenti (Madec F. etal. 2002).
Fertilità e patologie in bovini e suini
Supplemento de LA SETTIMANA VETERINARIA n° 804 del 31 ottobre 201230
PER SAPERNE DI PIÙ
• Martineau G.P. (2003) “Non infectious abor-tions in saws”, 10th International Symposiumon Pig Reproduction and Artificial Insemina-tion, Rome 5-7 May.
• Levis D.G. (2001) “What’s New with Seaso-nal Infertility?”, George A. Young Swine Con-ference 2001, University of Nebraska, 28-64.
• Mauget R (1982) “Seasonality of reproduc-tion in the wild boar”. Control of Pig Repro-duction. Cole DJA, Foxcroft GR (Eds). Butter-worths, London. pp. 509-526.
• Peltoniemi O.A.T., Love R.J., Heinonen M.,Tuovinen V., Saloniemi H. (1999) “Seasonaland management effects on fertility of the saw:a descriptive study”, An Reprod Sci, 55, 47-61.
• Kirkwood R. (2003) “Understanding and ma-naging seasonal infertility”, Allen D. LemanSwine Conference 2003, University of Minne-sota, 164-168.
• Ashworth C. (2006) “Reproduction”, in Kyria-zakis I., Whittemore C.T. “Whittemore’s scien-ce and practice of pig production”, 3th ed., Ox-ford, Blackwell Publishing, 104-147.
• Kirkwood R. (2002) “Seasonal infertility-cur-rent understandings”, Swine Disease Confe-rence for Swine Practitioners 2002, Iowa Sta-te University, 118-121.
• Madec F., Buddle J.R. (2002) “A reflectionon the strategies for tackling multifactorialdisease problems in pig, with specific referenceto post-weaningenteric disorders”, Proc. of the17° IPVS Congress, Ames, vol. 1, 113-120.
• Mechler D.W. (2002) “Seasonal infertility:early planning steps to minimize production ef-fects”, Swine Disease Conference for Swine Prac-titioners 2002, Iowa State University, 131-136.
• Muirhead M.R., Alexander T.J.L. (1997) “Ma-naging Pig Health and the Treatment of Dis-ease”, 1a ed., 5 M Enterprises Ltd, Uk.
Figura 1. Fattori stressanti che possono predisporre alla AAS.
01_30_Suppl SV804_01_30_Suppl SV804 31/10/12 11.12 Pagina 30