diversidade de anuros e lagartos em fisionomias de cerrado na … · 2006-10-03 · (campo sujo e...
TRANSCRIPT

Maria Tereza Chiarioni Thomé
Diversidade de anuros e lagartos em
fisionomias de Cerrado na região de
Itirapina, Sudeste do Brasil.
São Paulo
2006

ii
Maria Tereza Chiarioni Thomé
Diversidade de anuros e lagartos em
fisionomias de Cerrado na região de
Itirapina, Sudeste do Brasil.
Dissertação apresentada ao Instituto
de Biociências da Universidade de
São Paulo, para a obtenção de Título
de Mestre em Ciências, na Área de
Ecologia.
Orientador: Márcio R. C. Martins
São Paulo
2006

iii
Thomé, Maria Tereza Chiarioni.
Diversidade de anuros e lagartos em fisionomias de
Cerrado na região de Itirapina, Sudeste do Brasil. 59 pp.
Dissertação (Mestrado) - Instituto de Biociências da
Universidade de São Paulo. Departamento de Ecologia.
1. herpetofauna 2. Cerrado. Universidade de São
Paulo. Instituto de Biociências. Depto. de Ecologia.
Comissão Julgadora:
________________________ _______________________
Prof(a). Dr(a). Prof(a). Dr(a).
______________________
Prof. Dr. Márcio R.C. Martins
Orientador

iv
Capa: Cercosaura schreibersii e Physalaemus cuvieri.
Fotos: Ricardo Sawaya (disponível em http://eco.ib.usp.br/labvert/SiteItirapina/iti.htm).

v
Aos meus pais,
pelo apoio
e incentivo constante.

vi
Agradecimentos
Ao Professor Márcio Martins pela orientação e Cinthia A. Brasileiro pela co-
orientação informal.
À CAPES pela bolsa concedida. À FAPESP (Projeto Temático 00/12339-2)
e CNPq (processo 470621/2003-6) pelo apoio financeiro.
À Fundação O Boticário de Proteção à Natureza não só por patrocinar meu
projeto, mas pela paciência e fé no meu trabalho. Ao Idea Wild pelos
equipamentos. Ao Depto. de Ecologia do Instituto de Biociências da Universidade
de São Paulo pelo apoio logístico.
Ao IBAMA pelas licenças de coleta (02027.010426/99-21 e
02010.000987/04-19), ao Instituto Florestal e Fazenda São José S.A. pelo acesso
aos fragmentos.
À Denise Zanchetta e, especialmente, Gilson Gianez (ambos da Estação
Ecológica de Itirapina) pelo apoio logístico (leia-se desatolar a Brasa no meio da
noite, entre outras).
À Cinthia A. Brasileiro e Mara C. Kiefer por cederem parte dos dados de
coleta. Ao Cristiano Nogueira pela identificação dos lagartos e incentivo. À Ripasa
Celulose e Papel S.A. pelos dados climáticos da região. Ao Professor Roberto
Shimizo pela sugestão de como analisá-los.
Ao Tiago Castellar, Hilton Oyamaguchi, Roberta Bruscagim, Kika Braga,
Tessa, Noni, Zé Mário e tantos outros que ajudaram nos trabalhos de campo
empurrando a Brasa, plantando balde ou simplesmente dividindo o que tinha pra

vii
dividir em Itirapina. À Dna. Isabel por sempre reservar um cantinho no alojamento,
mesmo se fosse amontoando os outros. À Brasa, guerreira capenga e
desdentada. Sem você o que seria de mim...
Aos todos os colegas do LABVERT pela companhia, ajuda e paciência.
Beto, Robertinha, Hirotôn, Renato... À Cinthia pelo companheirismo e prontidão
em ajudar nos aspectos malucos da vida (dita) “acadêmica”.
Ao Tiago pelo amor e cumplicidade.

viii
Índice
I. Introdução...........................................................................................1
II. Objetivos............................................................................................5
III. Material e método
1. Área de estudo.............................................................................6
2. Delineamento amostral...............................................................12
3. Coleta de dados..........................................................................12
4. Análise de dados........................................................................15
IV. Resultados
1. Descrição da diversidade de anuros e lagartos em diferentes
fisionomias na região de Itirapina...............................................19
2. Estrutura da vegetação e composição de espécies nas
fisionomias..................................................................................30
V. Discussão........................................................................................34
VI. Referências Bibliográficas..............................................................41
VII. Resumo/ Abstract..........................................................................51

1
I. Introdução
A partir da segunda metade do século XX, diversas teorias foram
desenvolvidas para explicar a ocorrência de áreas de grande diversidade no
mundo (PIANKA, 1994; BEGON et al., 2005). Com o surgimento da disciplina
de ecologia de comunidades, levantou-se a possibilidade de que interações
ecológicas pudessem afetar o número de espécies que co-existem em um
mesmo local (RICKLEFS & SCHLUTER, 1993). Processos históricos de
ampla escala foram abandonados por serem considerados fracos e lentos,
dando espaço ao determinismo local de fatores ecológicos como o
responsável pelos padrões observados (RICKLEFS, 2004).
Dentre os fatores ecológicos evidenciados nestas teorias, receberam
maior atenção os mecanismos pelos quais espécies evitariam competição
(ZOBEL, 1997). A produtividade primária e a complexidade estrutural do
hábitat embasaram teorias ecológicas consagradas (RICKLEFS &
SCHLUTER, 1993; GASTON, 2000), segundo as quais hábitats como
florestas tropicais seriam ricos em espécies porque a maior complexidade
estrutural e produtividade resultariam em maior oferta e variedade de micro
hábitats e presas, permitindo especializações de nicho (PIANKA, 1994) e
evitando a competição.
Mais recentemente, teorias foram propostas evidenciando processos
regionais, como migração, dispersão, colonização e extinção, como
modeladores da diversidade local em tempo ecológico (TAYLOR et al, 1990,
ZOBEL, 1997). Como estes processos dependem de uma matriz de espécies

2
disponíveis, estas teorias relacionam-se com as origens da diversidade em
tempo evolutivo, o que promoveu a retomada da importância dos fatores de
ampla escala (RICKLEFS, 1987; RICKLEFS & SCHLUTER, 1993;
RICKLEFS, 2004) ligados à biogeografia histórica (ZOBEL, 1992). A
interpretação mais direta das teorias baseadas em efeitos regionais assume
o número de espécies que ocorre em um hábitat como uma função da
disponibilidade deste hábitat em uma região (ZOBEL, 1997). Atualmente,
estudos sobre esse assunto consideram tanto a disponibilidade regional de
espécies (ZOBEL, 1997; CALEY & SCHLUTER, 1997; MOUQUET &
LOREAU, 2002) quanto os fatores ecológicos tradicionais (ver SHURIM &
ALLEN, 2001, para uma revisão sobre o assunto).
O Cerrado é a maior e mais rica savana no mundo (SILVA & BATES,
2002) e constitui um dos componentes de vegetação terrestre mais
importantes do continente sul-americano, ocorrendo ao longo de mais de 20°
de latitude (RATTER et al., 1997). É também um dos biomas brasileiros mais
ameaçados (SILVA & BATES, 2002) e faz parte da lista mundial dos
ecossistemas com maior diversidade e perda de hábitat, os hotspots
(MYERS et al 2000). Atualmente, 20% de sua vegetação original se encontra
relativamente bem preservada (MITTERMEYER et al., 2000), mas apenas
1,2% está protegida em unidades de conservação. Apesar do crescimento do
interesse por parte da comunidade científica nas últimas décadas, a fauna do
Cerrado ainda é pouco conhecida e o número de estudos básicos que
possam fornecer subsídios para a conservação, como os inventários
faunísticos, é ainda muito pequeno (CAVALCANTI & JOLY, 2002).

3
Até recentemente, acreditava-se que a herpetofauna do Cerrado fosse
descaracterizada e depauperada, constituída em sua maior parte por
espécies generalistas oriundas dos biomas vizinhos (VANZOLINI, 1976;
WEBB, 1978; VITT, 1991; SILVA & SITES, 1995). Hoje, é consenso entre
autores que a diversidade real da herpetofauna do Cerrado havia sido
subestimada; diversos endemismos são conhecidos (30% dos anfíbios, 20%
dos répteis; MYERS, 2000) e o número de espécies que ocorrem no bioma
vem crescendo consideravelmente nos últimos anos (e.g. NOGUEIRA,
2006). Segundo HEYER (1988) e COLLI (1998), a razão deste equívoco
deve-se a amostragens insatisfatórias; conforme novas localidades foram
amostradas, espécies foram e ainda são descritas freqüentemente (COLLI et
al., 2002; DINIZ et al, 2005).
Para explicar esta alta diversidade e endemismo, alguns autores
sugeriram como causa a estratificação horizontal do bioma. No Cerrado
existem diversos tipos vegetacionais (fisionomias) que representam hábitats
diferentes para a herpetofauna (COLLI et al, 2002). Estes hábitats variam de
florestas a campos em um gradiente de complexidade estrutural (EITEN,
1972; COUTINHO, 1978), e se distribuem na paisagem em mosaicos
complexos de fisionomias com características próprias, potencializando a
especialização das linhagens aí presentes.
Esta estratificação horizontal, associada à megadiversidade da
herpetofauna, confere ao bioma condições propícias para o estudo dos
fatores condicionantes da diversidade. Fisionomias fechadas são
estruturalmente mais complexas (COUTINHO, 1978) e produtivas do que
fisionomias abertas (VOURLITIS et al., 2001; RATANA et al., 2005); se

4
fatores locais tiverem maior relevância para a determinação da riqueza de
espécies, deve-se esperar uma maior riqueza nestas fisionomias mais
complexas e produtivas. Por outro lado, de uma forma geral, as fisionomias
mais abertas são predominantes nos Cerrados (EITEN, 1972; RATTER et al,
1997); portanto, se houver preponderância de fatores regionais na
determinação da riqueza de espécies, a maior riqueza deve ser encontrada
nestas fisionomias. Do ponto de vista da paisagem, manchas de fisionomias
raras (neste caso as fisionomias mais fechadas) podem não ser suficientes
para manter metapopulações de espécies especialistas nestes habitats raros,
ou seja, não haveria manchas que se comportariam como fontes de novos
indivíduos para colonização (ver HOLT, 1993).
Na região de Itirapina, estado de São Paulo, tanto ocorrem fisionomias
de Cerrado fechado (cerrado típico e cerradão) quanto fisionomias abertas
(campo sujo e campo cerrado), sendo que, na escala regional, as últimas são
predominantes. Este estudo compara a diversidade de anuros e lagartos em
diferentes fisionomias da região, averiguando quais fatores explicam melhor
a distribuição da diversidade encontrada.

5
II. Objetivos
Os objetivos gerais do presente trabalho foram descrever e comparar
a diversidade das taxocenoses de anuros e lagartos nas fisionomias de
campo sujo, campo cerrado, cerrado típico (cerrado sensu stricto) e cerradão
da região de Itirapina, testando a hipótese de que a diversidade é mais alta
em ambientes estruturalmente mais complexos e produtivos. Para tanto, os
objetivos específicos foram:
• Descrever como a diversidade de anuros e lagartos se distribui
espacialmente nas fisionomias de campo sujo, campo cerrado,
cerrado típico e cerradão, na região de Itirapina;
• Descrever a variação estrutural da vegetação nestas fisionomias e sua
relação com a composição das taxocenoses,
• Discutir os possíveis fatores condicionantes da diversidade de anuros
e lagartos da região.

6
III. Materiais e Métodos
1. Áreas de estudo
O estudo foi realizado na região de Itirapina, São Paulo, Brasil, em
fragmentos preservados pertencentes aos municípios de Itirapina e Brotas. A
região (fig. 1) encontra-se na porção Sul do domínio do Cerrado (RATTER et
al, 1997) em uma bacia sedimentar de 730-770 m de altitude, caracterizada
por solos predominantemente arenosos (VICENTE et al., 2005), cobertos por
fisionomias abertas de Cerrado. Em locais onde os solos são argilosos,
pequenos enclaves naturais de vegetação mais densa podem ser
encontrados, como matas de galeria nas áreas ripárias, cerrados típicos e
cerradões nos interflúvios. Como os solos argilosos são escassos nos
interflúvios da região, áreas de cerrados típicos e cerradões são mais raras,
prevalecendo as fisionomias de campo sujo e campo cerrado.
O campo sujo (fig. 2 A) é uma formação caracterizada por extensos
campos de gramíneas com alguns arbustos e pequenas palmeiras (Syagrus
sp e Attalea sp) dispersos. O campo cerrado (fig. 2 B), por sua vez,
apresenta árvores esparsas, além dos estratos arbustivo e herbáceo com
predomínio de gramíneas. Já o cerrado típico (fig. 2 C) possui uma cobertura
arbóreo-arbustiva mais densa, sendo o estrato herbáceo menos
desenvolvido devido ao sombreamento proporcionado pelos outros estratos.
No cerradão (fig. 2 D), o estrato herbáceo é praticamente ausente; as árvores
são mais altas e em maior densidade, formando um dossel fechado.
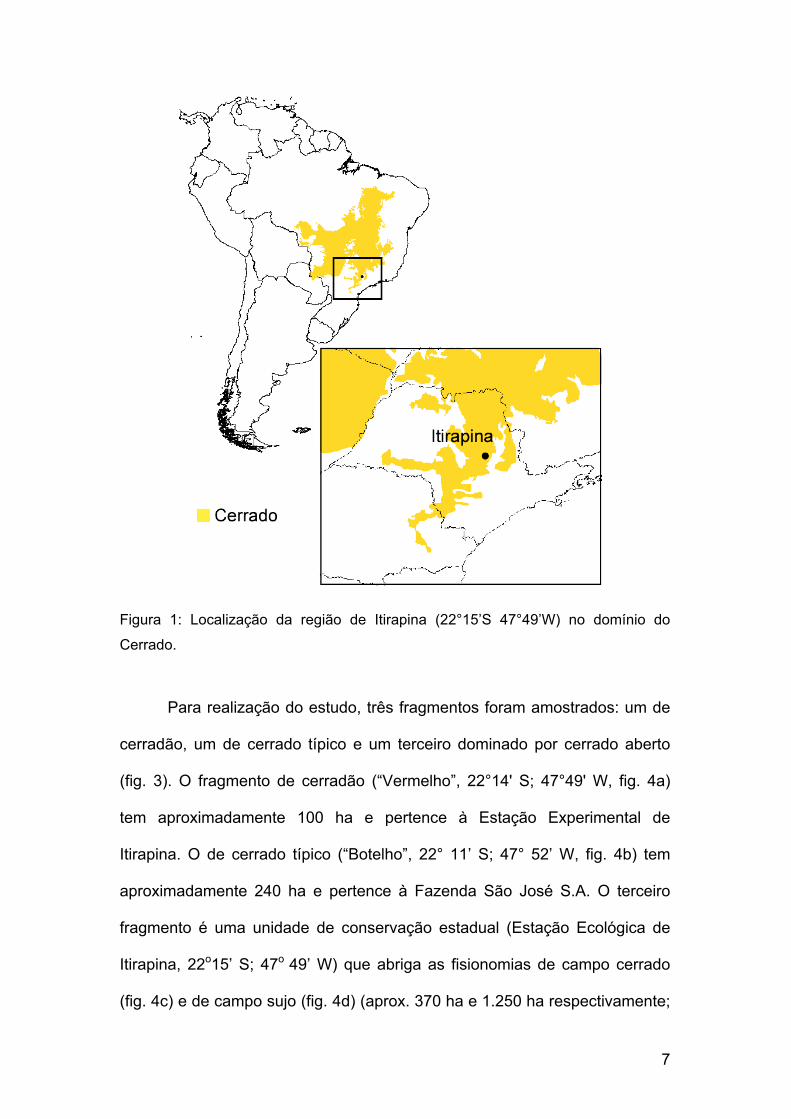
7
Figura 1: Localização da região de Itirapina (22°15’S 47°49’W) no domínio do
Cerrado.
Para realização do estudo, três fragmentos foram amostrados: um de
cerradão, um de cerrado típico e um terceiro dominado por cerrado aberto
(fig. 3). O fragmento de cerradão (“Vermelho”, 22°14' S; 47°49' W, fig. 4a)
tem aproximadamente 100 ha e pertence à Estação Experimental de
Itirapina. O de cerrado típico (“Botelho”, 22° 11’ S; 47° 52’ W, fig. 4b) tem
aproximadamente 240 ha e pertence à Fazenda São José S.A. O terceiro
fragmento é uma unidade de conservação estadual (Estação Ecológica de
Itirapina, 22o15’ S; 47o 49’ W) que abriga as fisionomias de campo cerrado
(fig. 4c) e de campo sujo (fig. 4d) (aprox. 370 ha e 1.250 ha respectivamente;

8
HÜLLE, 2006) amostradas neste estudo. A matriz entre os três fragmentos é
formada atualmente por pequenas áreas urbanizadas, uma represa,
pastagens e monoculturas extensas (cana-de-açúcar, Pinus e Eucaliptus).
Figura 2: Fisionomias de Cerrado, amostradas na região de Itirapina. A, campo sujo;
B, campo cerrado; C, cerrado típico e D; cerradão. A e B retiradas de BRASILEIRO
(2004).
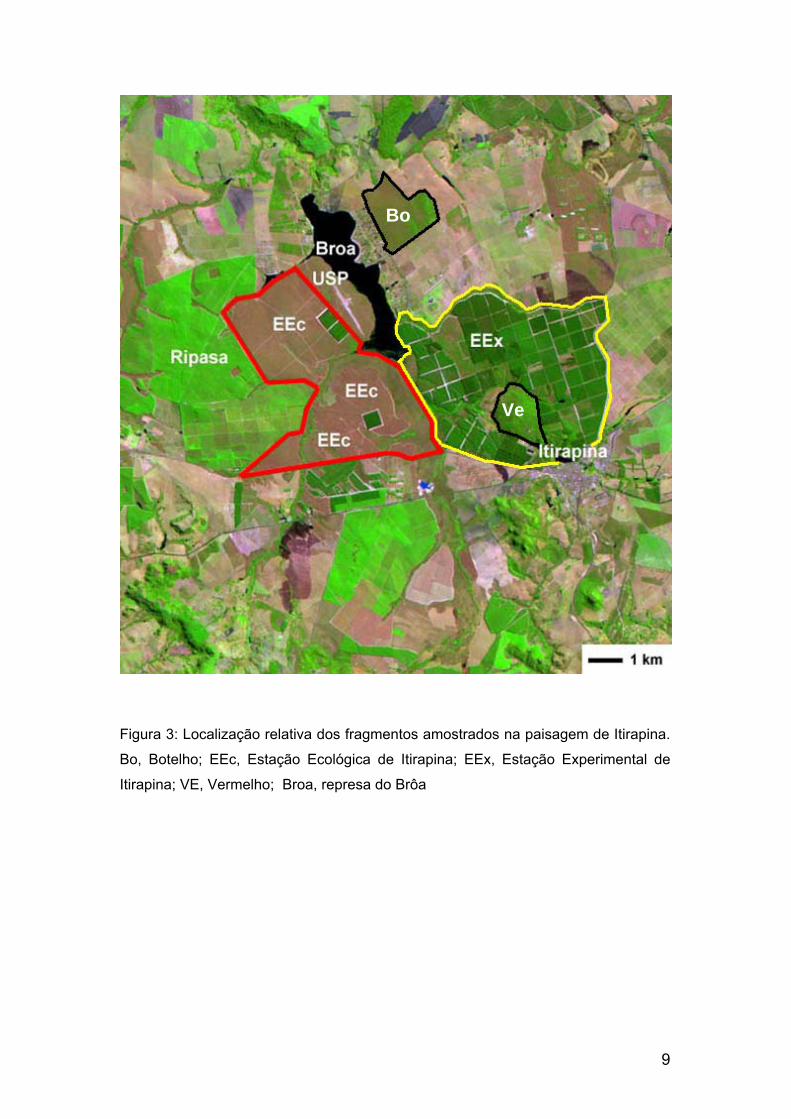
9
Figura 3: Localização relativa dos fragmentos amostrados na paisagem de Itirapina.
Bo, Botelho; EEc, Estação Ecológica de Itirapina; EEx, Estação Experimental de
Itirapina; VE, Vermelho; Broa, represa do Brôa
Bo
Ve

10
Figura 4: Vegetação dos fragmentos amostrados. (a), cerradão no Vermelho; (b),
cerrado típico no Botelho; (c), campo cerrado e (d), campo sujo, ambos na Estação
Ecológica de Itirapina. Fotos (c) e (d) cedidas por Márcio Martins.
a b
c d

11
Como outras regiões do Cerrado (KLINK & MACHADO, 2005), a
região de Itirapina apresenta clima mesotérmico com sazonalidade
acentuada. Devido à concentração das chuvas em seis meses seguidos, o
clima ao longo do ano pode ser tipicamente reconhecido em apenas duas
estações; uma estação seca bem pronunciada de temperaturas mais
amenas, de abril a setembro, e uma estação chuvosa de outubro até março,
com temperaturas mais elevadas (fig. 5). De julho de 2000 a junho de 2005,
as médias mensais de temperatura máxima chegaram a 34,5 °C durante as
estações chuvosas, enquanto que o menor valor de temperatura mínima
média chegou a 7,9 °C, registrado durante a estação seca. A pluviosidade
total mensal atingiu 433 mm durante a estação chuvosa, sendo que durante
as estações secas houve meses sem qualquer registro de chuva.
2000 a 2005
0
100
200
300
400
jul ago set out nov dez jan fev mar abr mai jun
pluv
iosi
dade
(mm
)
5
15
25
temperatura (°C
) m
in max
Figura 5: Pluviosidade total mensal (barras) e médias mensais das temperaturas
mínimas (linha azul) e máximas (linha vermelha) para região de Itirapina. Cada mês
representa a média dos valores mensais registrados entre julho de 2000 e junho de
2005. A linha tracejada delimita os meses da estação chuvosa.

12
2. Delineamento amostral
Para a amostragem dos anuros e lagartos nas fisionomias, foram
feitas coletas durante duas estações chuvosas: 2001/2002 e 2004/2005
(estações “1” e “2” respectivamente). Os dados das coletas referentes à
estação chuvosa 1 foram obtidos em BRASILEIRO (2004) (anuros) e cedidos
por M. C. Kiefer (lagartos, dados não publicados). Os dados da estação
chuvosa 2 foram coletados pela presente autora, de forma padronizada,
semelhante àquela utilizada pelas autoras anteriores.
Durante a estação chuvosa 1 foram amostradas as fisionomias
campestres, enquanto as fisionomias de cerrado típico e cerradão foram
amostradas durante a estação 2. Devido à padronização nas coletas de
dados, o esforço amostral foi o mesmo nas quatro fisionomias, permitindo
comparações diretas de diversidade. Entretanto, como as coletas não foram
concomitantes em todas as fisionomias, o delineamento causou um viés
temporal, já que comparações de diversidade entre estações chuvosas
diferentes podem ser influenciadas por possíveis flutuações populacionais.
Este problema é uma limitação deste trabalho que será discutida
posteriormente.
3. Coleta de dados
As amostragens foram feitas com armadilhas de interceptação e
queda (sensu GREENBERG et al. 1994, CECHIN & MARTINS, 2000),
distribuídas igualmente entre fisionomias. As armadilhas eram compostas por

13
baldes de 100 L enterrados, interligados por cercas de tela plástica com 50
cm de altura, formando linhas de armadilhas. Cada linha tinha 45 m de
comprimento e quatro baldes, um a cada 15 m, intercalados pelas cercas-
guia (fig. 6a). As linhas foram dispostas em pares, posicionados em
sobreposição a um eixo imaginário e distantes 100 m uma da outra (fig. 6b).
Cada par de linhas foi considerado um ponto amostral. Em cada fisionomia
foram instalados três pontos amostrais, respeitando-se uma distância mínima
de 500 m entre pontos, sendo estes considerados unidades independentes
(réplicas). Os animais capturados foram marcados por amputação de
artelhos e soltos no mesmo local da coleta. Recapturas foram
desconsideradas nas análises. Devido às dificuldades na identificação entre
jovens de Physalaemus cuvieri e P. centralis, ambas espécies são
consideradas conjuntamente como “Physalaemus spp. gr. cuvieri”.
As amostragens ocorreram em viagens a campo (coletas) quinzenais,
realizadas sempre entre a segunda quinzena de outubro e a primeira
quinzena de março (tabela I). Dez coletas foram feitas por estação chuvosa,
cada uma com duração de cinco dias consecutivos, totalizando 100 dias de
coleta, 50 dias em campo sujo e em campo cerrado na estação chuvosa 1 e
50 dias em cerrado típico e em cerradão na estação chuvosa 2.
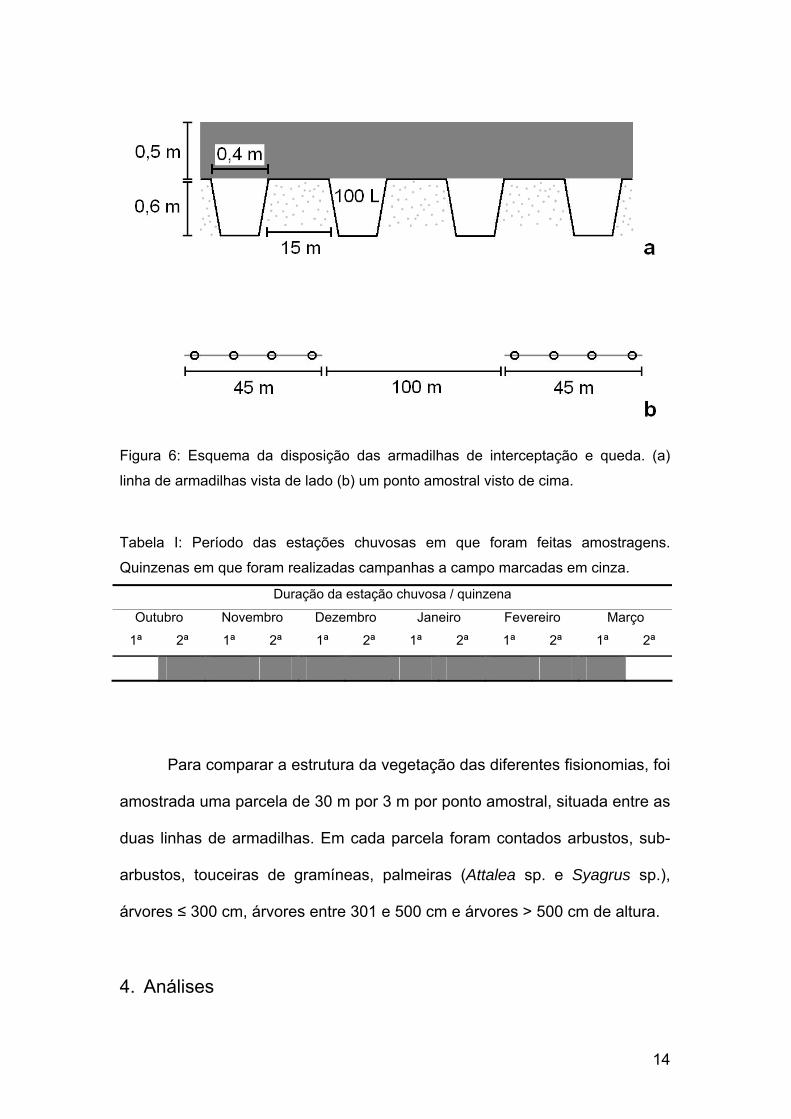
14
Figura 6: Esquema da disposição das armadilhas de interceptação e queda. (a)
linha de armadilhas vista de lado (b) um ponto amostral visto de cima.
Tabela I: Período das estações chuvosas em que foram feitas amostragens.
Quinzenas em que foram realizadas campanhas a campo marcadas em cinza.
Duração da estação chuvosa / quinzena
Outubro Novembro Dezembro Janeiro Fevereiro Março
1ª 2ª 1ª 2ª 1ª 2ª 1ª 2ª 1ª 2ª 1ª 2ª
Para comparar a estrutura da vegetação das diferentes fisionomias, foi
amostrada uma parcela de 30 m por 3 m por ponto amostral, situada entre as
duas linhas de armadilhas. Em cada parcela foram contados arbustos, sub-
arbustos, touceiras de gramíneas, palmeiras (Attalea sp. e Syagrus sp.),
árvores ≤ 300 cm, árvores entre 301 e 500 cm e árvores > 500 cm de altura.
4. Análises

15
4.1. Esforço amostral
Para avaliar o esforço amostral de captura foram feitas curvas de
rarefação de espécies (SANDERS, 1968; SIMBERLOFF, 1978; GOTELLI &
GRAVES, 1996) com 1.000 aleatorizações no programa EstimateS
(COLWELL, 2005). As curvas foram feitas por amostra e escalonadas pelo
número de indivíduos (GOTELLI & COLWELL, 2001), separadamente para
cada fisionomia. O número de capturas registrado por ponto amostral
(réplica) a cada duas campanhas foi considerado uma amostra.
4.2. Descrição da diversidade
Tradicionalmente, herpetofaunas tropicais têm sido comparadas em
termos de riqueza, densidade, dominância e equitatividade (HOFER &
BERSIER, 2001). Para comparar riqueza, foram utilizadas as curvas de
rarefação de espécies, riqueza estimada e riqueza rarefeita por abundância
(GOTELLI & GRAVES, 1996). Estimativas de riqueza, bem como as curvas
de rarefação de espécies (ver 4.1. Esforço amostral) foram feitas com 1.000
aleatorizações no programa EstimateS (COLLWEL, 2005). O estimador
escolhido foi “Chao 1” por seu desempenho em amostras com número alto
de espécies com apenas uma captura (“Singletons”, ver COLWELL &
CODDINGTON, 1994). A riqueza rarefeita por abundância foi calculada sobre
o total de capturas a partir da menor amostra, no programa EcoSim 7.00
(GOTTELI & ENTSMINGER, 2004), com 1.000 aleatorizações.

16
Análises envolvendo densidade de indivíduos não foram realizadas, já
que o método de captura escolhido não permite inferências sobre sua área
de abrangência. Por outro lado, devido à padronização do esforço amostral,
as abundâncias absolutas ou abundâncias médias por amostra encontradas
nas diferentes fisionomias puderam ser comparadas diretamente. No caso
das abundâncias médias, o total de capturas em duas campanhas por
fisionomia foi considerado uma amostra.
O grau de dominância foi expresso em termos de porcentagem da
espécie mais abundante. Para a equitatividade, foram seguidas as
recomendações de GOTELLI & GRAVES (1996), com o uso da probabilidade
de encontro interespecífico (PEI) de HULBERT (1971), obtida com o
programa EcoSim 7.00 (GOTTELI & ENTSMINGER, 2004), com 1.000
aleatorizações. Como na riqueza rarefeita por abundância, a PEI foi
calculada sobre o total de capturas por espécie.
Para comparar riqueza e abundância absoluta entre fisionomias,
primeiramente foi avaliada a efetividade dos pontos amostrais como réplicas.
Para isso, foram feitas análises de variância de Kruskal-Wallis, onde a
riqueza e a abundância registradas em duas campanhas por ponto amostral
(replica) foram consideradas uma amostra. Quando Ho foi rejeitada, testes de
Dunn foram realizados a posteriori (ZAR, 1996). Todos os testes foram
realizados com nível de significância 0,05 no programa Bioestat (AYRES et
al. 1998).
Uma vez testadas as réplicas, foram feitas também análises de
variância de Kruskall-Wallis para comparar riqueza e abundância absoluta
entre as fisionomias e individualmente para as espécies mais abundantes.

17
Nestas análises, as capturas de cada ponto amostral (réplica) foram
consideradas conjuntamente para cada fisionomia, sendo então considerada
uma amostra a riqueza ou a abundância registrada em duas campanhas por
fisionomia. Quando Ho foi rejeitada, testes de Dunn foram realizados a
posteriori (ZAR, 1996).
Para investigar alterações do clima entre as estações chuvosas foram
obtidos dados diários das temperaturas máxima e mínima e de pluviosidade
para a região. Para uma avaliação mais robusta, os períodos considerados
incluíram não só as estações chuvosas 1 e 2, mas também as respectivas
estações secas anteriores. Para avaliar se estes períodos se comportaram
de forma semelhante, foram feitas correlações de Spearman entre dados
climáticos quinzenais (médias de temperatura e pluviosidade total). Se a
correlação foi significativa, para avaliar se e como os períodos diferiram nos
valores da variável em questão foram feitas regressões lineares e
comparados seus respectivos coeficientes de inclinação da reta.
4.3. Composição de espécies e estrutura da vegetação
Para averiguar quais fisionomias tiveram as composições de espécies
mais semelhantes entre si, foram feitas análises de agrupamento (“UPGMA”)
utilizando o índice de similaridade modificado de Morisita-Horn (ver
MAGURRAN, 1988). Nestas análises foram consideradas as abundâncias
das espécies por ponto amostral. As abundâncias foram transformadas em
seu logaritmo natural. Para verificar as diferenças na estrutura da vegetação
das fisionomias, foi feita uma análise dos componentes principais (ACP) a

18
partir dos dados obtidos nas parcelas de vegetação. Os dados brutos das
variáveis foram padronizados (ver MINGOTI, 2005). Todas estas análises
foram feitas no programa MVSP (KOVACH, 2000).

19
IV. Resultados
1. Descrição da diversidade de anuros e lagartos entre
fisionomias na região de Itirapina
1.1. Esforço amostral
Foi capturado um total de 1.437 anuros e 169 lagartos, sendo treze
espécies de anuros pertencentes a três famílias, e doze espécies de lagartos
pertencentes a cinco famílias (tabela II). As curvas de rarefação de espécies
de anuros (fig. 7) não estabilizaram, indicando que provavelmente nem todas
as espécies foram capturadas nestas fisionomias. Nas fisionomias de campo
sujo e campo cerrado, as curvas ultrapassaram o ponto de inflexão, visto que
os desvios-padrão tendem a diminuir no final. Já em cerrado típico e
cerradão, não foram capturados indivíduos suficientes para que este ponto
fosse atingido. Das curvas de rarefação de espécies de lagartos (fig. 7)
apenas a do cerradão estabilizou; provavelmente um viés derivado do
número extremamente baixo de capturas (tabela II). Nas outras fisionomias,
o ponto de inflexão foi atingido apenas no campo sujo, enquanto que no
campo cerrado e cerrado típico, as curvas para lagartos permaneceram em
ascensão.
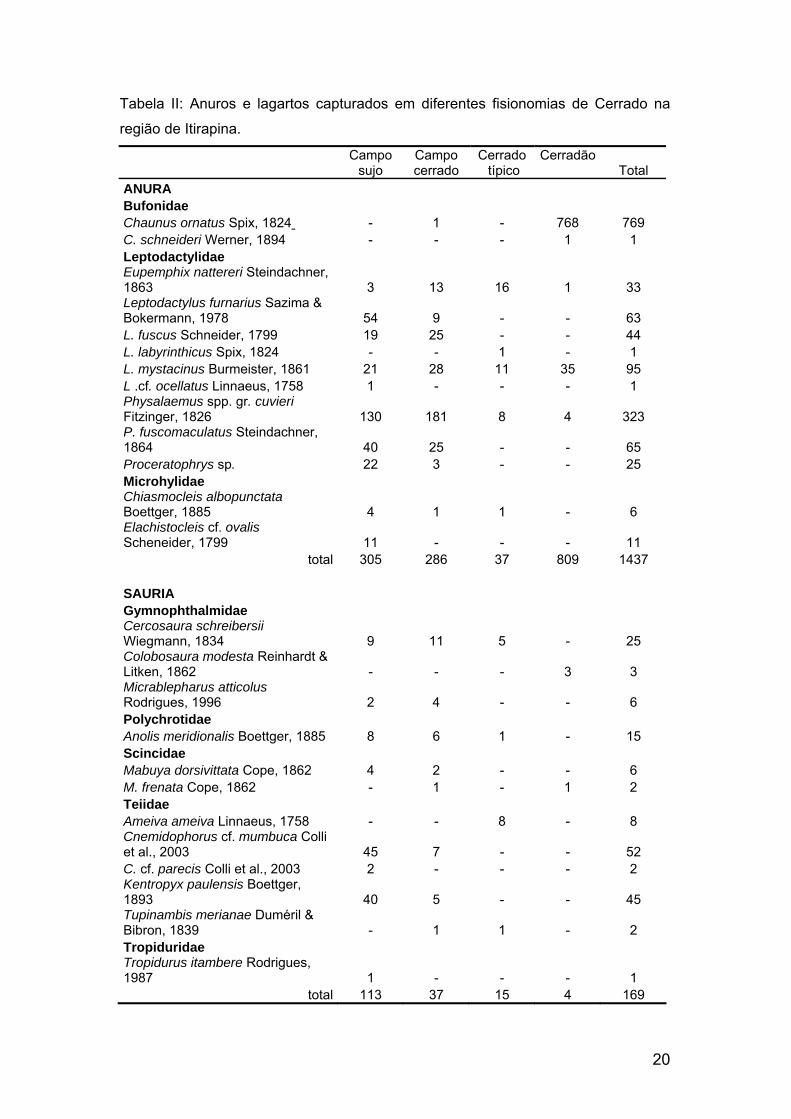
20
Tabela II: Anuros e lagartos capturados em diferentes fisionomias de Cerrado na
região de Itirapina.
Campo
sujo Campo cerrado
Cerrado típico
Cerradão Total
ANURA Bufonidae Chaunus ornatus Spix, 1824 - 1 - 768 769 C. schneideri Werner, 1894 - - - 1 1 Leptodactylidae Eupemphix nattereri Steindachner, 1863 3 13 16 1 33 Leptodactylus furnarius Sazima & Bokermann, 1978 54 9 - - 63 L. fuscus Schneider, 1799 19 25 - - 44 L. labyrinthicus Spix, 1824 - - 1 - 1 L. mystacinus Burmeister, 1861 21 28 11 35 95 L .cf. ocellatus Linnaeus, 1758 1 - - - 1 Physalaemus spp. gr. cuvieri Fitzinger, 1826 130 181 8 4 323 P. fuscomaculatus Steindachner, 1864 40 25 - - 65 Proceratophrys sp. 22 3 - - 25 Microhylidae Chiasmocleis albopunctata Boettger, 1885 4 1 1 - 6 Elachistocleis cf. ovalis Scheneider, 1799 11 - - - 11
total 305 286 37 809 1437 SAURIA Gymnophthalmidae Cercosaura schreibersii Wiegmann, 1834 9 11 5 - 25 Colobosaura modesta Reinhardt & Litken, 1862 - - - 3 3 Micrablepharus atticolus Rodrigues, 1996 2 4 - - 6 Polychrotidae Anolis meridionalis Boettger, 1885 8 6 1 - 15 Scincidae Mabuya dorsivittata Cope, 1862 4 2 - - 6 M. frenata Cope, 1862 - 1 - 1 2 Teiidae Ameiva ameiva Linnaeus, 1758 - - 8 - 8 Cnemidophorus cf. mumbuca Colli et al., 2003 45 7 - - 52 C. cf. parecis Colli et al., 2003 2 - - - 2 Kentropyx paulensis Boettger, 1893 40 5 - - 45 Tupinambis merianae Duméril & Bibron, 1839 - 1 1 - 2 Tropiduridae Tropidurus itambere Rodrigues, 1987 1 - - - 1
total 113 37 15 4 169

21
anuros
0
3
6
9
0 100 200 300indivíduos
espé
cies CS
0
3
6
9
0 100 200indivíduos
espé
cies
CC
0
3
6
9
0 10 20 30indivíduos
espé
cies SS
0
3
6
9
0 200 400 600 800indivíduos
espé
cies
CE
lagartos
0
3
6
9
0 5 10 15indivíduos
espé
cies
0
3
6
9
0 50 100indivíduos
espé
cies
0
3
6
9
0 10 20 30indivíduos
espé
cies
CC
0
3
6
9
0 1 2 3 4indivíduos
espé
cies
CE
SS
CS
Figura 7: Curvas de rarefação de espécies de anuros e lagartos em fisionomias de
Cerrado na região de Itirapina. CS, campo sujo; CC, campo cerrado; CT, cerrado
típico e CE, cerradão. Barras indicam o desvio padrão

22
1.2. Diversidade de anuros e lagartos
A riqueza de anuros e lagartos foi mais alta nas fisionomias abertas do
que nas fisionomias fechadas (tabela III). Das treze espécies de anuros, onze
foram capturadas em campo sujo e campo cerrado (tabela II). Das doze
espécies de lagartos, também onze foram capturadas nestas fisionomias
(tabela II). Análises de variância evidenciam diferenças significativas de
riqueza entre as fisionomias, tanto para anuros quanto para lagartos (tabela
IV). Testes a posteriori para anuros indicam que a riqueza diferiu entre
campo sujo e cerrado típico e entre campo sujo e cerradão (tabela V). Para
lagartos, a riqueza no campo sujo também diferiu da riqueza no cerradão
(tabela V).
Os descritores de riqueza indicam que as fisionomias abertas foram
mais ricas do que as fechadas (tabela III). Para anuros, os maiores valores
de riqueza absoluta, rarefeita e de riqueza média por amostra foram
registrados no campo sujo seguido pelo campo cerrado, enquanto as
riquezas estimadas apresentaram valores iguais para estas fisionomias. O
cerrado típico e o cerradão apresentaram valores mais baixos e parecidos
entre si para todas as riquezas (tabela III). Houve sobreposição nos
intervalos de confiança das riquezas rarefeita e estimada de todas as
fisionomias, com exceção da riqueza rarefeita do cerradão (tabela III).
Para lagartos, a riqueza média por amostra também foi mais alta no
campo sujo seguido do campo cerrado (tabela III). Entretanto, as riquezas
absoluta e estimada apresentaram valores extremamente próximos nestas

23
fisionomias, enquanto que as riquezas rarefeitas apresentaram valores sem
nenhum padrão (o que provavelmente se deve ao número baixo de capturas
pelo qual as amostras foram rarefeitas). Em todos os descritores de riqueza o
cerrado típico se apresentou mais pobre do que as fisionomias abertas, e
mais rico do que o cerradão (tabela III). Também houve sobreposição nos
intervalos de confiança (tabela III), exceto para a riqueza estimada no
cerradão.
Análises de variância também evidenciaram diferenças significativas
na abundância de anuros e de lagartos entre as fisionomias (tabela IV).
Testes a posteriori para anuros indicam que a abundância diferiu entre
cerradão e cerrado típico (tabela V). Para lagartos, a abundância diferiu entre
campo sujo e cerrado típico e entre campo sujo e cerradão (tabela V).
O cerradão foi a fisionomia com maior abundância de anuros (tabela
III), tanto absoluta quanto abundância média por amostra, seguido pelo
campo sujo, campo cerrado e por último o cerrado típico. Para lagartos, a
abundância seguiu o mesmo padrão da riqueza, com valores altos para as
fisionomias abertas, decrescendo no sentido das fisionomias fechadas
(tabela III). Entretanto, o padrão da abundância encontrado para anuros foi
extremamente influenciado pelo número de capturas de Chaunus ornatus. Ao
se retirar esta espécie das amostras, ainda as fisionomias apresentam
abundâncias significativamente diferentes (H = 14,31; p < 0,01), e o campo
sujo se torna a fisionomia de maior abundância, seguido do campo cerrado
(tabela III). Nos testes a posteriori, o campo sujo diferiu do cerrado típico e do
cerradão e o campo cerrado diferiu do cerrado típico.

24
Não foram encontradas diferenças significativas na riqueza ou
abundância por amostra de anuros entre pontos amostrais (tabela VI),
mostrando que estes se comportaram como réplicas verdadeiras em todas
as fisionomias. O mesmo ocorreu para a riqueza de lagartos; para a
abundância, houve variações entre as réplicas das fisionomias campestres.
No campo sujo esta variação foi marginalmente significativa, enquanto no
campo cerrado foi significativa (tabela IV). Devido às variações serem
apenas na abundância, nenhum dos pontos amostrais foi desconsiderado
nas análises anteriores.
Tabela III: Descritores de diversidade de anuros e lagartos em fisionomias de
Cerrado na região de Itirapina. Aabs, abundância absoluta; μA, abundância média
por amostra; Sabs riqueza observada; μS, riqueza média por amostra; Sr riqueza
rarefeita por abundância; Se riqueza estimada com o estimador Chao 1; C.I.
intervalos de confiança em 95%; PIE probabilidade de encontro interespecífico.
Campo sujo Campo cerrado Cerrado típico Cerradão
Anuros
Aabs 305 286 37 809
μA 61 57,2 7,4 161,8
Sabs 10 9 5 5
μS 7 5,8 2,6 2,8
Sr (C,I, 95%) 7,6 (6 - 9) 6,1 (4 - 8) - 2,07 (1 - 3)
Se (C,I, 95%) 10 10 (9,1 - 22,9) 6 (5,2 – 28,1) 6 (5 – 18,5)
PIE 0,76 0,57 0,69 0,10
Lagartos
Aabs 111 37 15 4
μA 22,2 7,4 3 0,8
Sabs 8 8 4 2
μS 5,4 4,4 1,6 0,6
Sr (C,I, 95%) 2,6 (1 - 4) 3,18 (2 - 4) 2,35 (1 – 4) -
Se (C,I, 95%) 8,25 (8 - 12,7) 10 (8,2 - 30,1) 5 (4,1 - 17,3) 2 (2 – 3,4)
PIE 0,71 0,84 0,64 0,50

25
Tabela IV: Análises de variância de Kruskal-Wallis para riqueza e abundância entre
fisionomias. Grau de liberdade 3. Resultados significativos estão em negrito.
Anuros Lagartos
H P H P
Riqueza 13,37 < 0,01 14,07 < 0,01 Abundância 11,1 0,01 16,1 < 0,01
Tabela V: Testes à posteriori de Dunn das análises da tabela VI. Testes
significativos marcados por X.
Anuros Lagartos
Riqueza Abundância Riqueza Abundância
Campo sujo e campo cerrado
Campo sujo e cerrado típico X X
Campo sujo e cerradão X X X Campo cerrado e cerrado típico
Campo cerrado e cerradão X
Cerrado típico e cerradão X
Tabela VI: Análises de variância de Kruskal-Wallis para teste de réplicas. Grau de
liberdade 2. Resultados significativos estão em negrito.
Campo sujo Campo cerrado Cerrado típico Cerradão
Riqueza
Anuros H 0,08 0,29 2,59 1,85
p 0,96 0,86 0,27 0,24
Lagartos H 0,5 4,38 1,12 1,52
p 0,78 0,11 0,56 0,46
Abundânci
a

26
Anuros H 0,32 3,33 3,44 0,74
p 0,85 0,19 0,18 0,69
Lagartos H 6,43 7,27 1,12 1,52
p 0,04 0,02 0,56 0,47
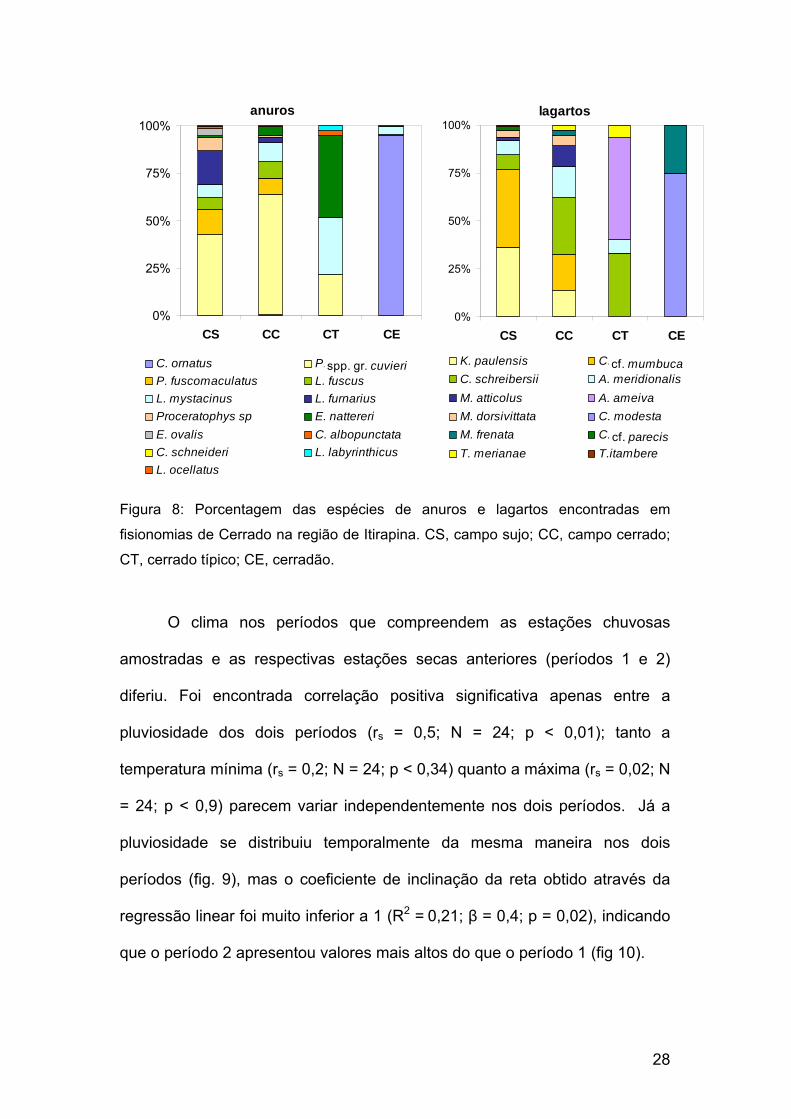
As espécies dominantes entre os anuros foram as mesmas nas
fisionomias abertas; Physalaemus spp. gr. cuvieri predominaram tanto em
campo sujo (42%) quanto em campo cerrado (63%). Em cerrado típico e
cerradão as espécies dominantes foram Eupemphix nattereri (43%) e
Chaunus ornatus (95%), respectivamente (fig. 8). A probabilidade de
encontro interespecífico (tabela IV) foi semelhante nas fisionomias abertas e
no cerrado típico. No cerradão, a dominância extrema resultou em uma
probabilidade de encontro interespecífico muito baixa.
Entre lagartos a espécie dominante nunca foi a mesma:
Cnemidophorus cf. mumbuca (40%) no campo sujo, Cercosaura schreibersii
(30%) no campo cerrado, Ameiva ameiva (53%) no cerrado típico e
Colobosaura modesta (75%) no cerradão (fig. 8). A probabilidade de
encontro interespecífico (tabela IV) permaneceu ligeiramente mais alta nas
fisionomias abertas, seguida do cerrado típico e cerradão.
A maioria das espécies de anuros e lagartos apresentou distribuição
diferenciada entre as fisionomias. Desconsiderando as espécies raras
(menos de 15 capturas; tabela II), entre anuros, Physalaemus spp. gr. cuvieri
e Proceratophrys sp. foram significativamente mais abundantes nas
fisionomias abertas (tabela VII). Eupemphix nattereri, Leptodactylus furnarius,
L. fuscus e Physalaemus fuscomaculatus não apresentaram variação de
abundância significativa e detectável a posteriori (tabela VII), embora
aparentemente sejam mais abundantes nas fisionomias abertas. Chaunus

27
ornatus foi mais abundante em cerradão e apenas L. mystacinus parece
ocorrer igualmente entre as fisionomias (tabela VII).
Entre os lagartos, Kentropyx paulensis e Cnemidophorus cf. mumbuca
ocorreram apenas nas fisionomias campestres e preferencialmente em
campo sujo (tabela VII). Cercosaura schreibersii e Anolis meridionalis não
apresentaram variações significativas de abundância entre fisionomias
(tabela VII) e ambas ocorreram também em cerrado típico (embora A.
meridionalis tenha apenas um registro para esta fisionomia).
Tabela VII: Análises de variância de Kruskal-Wallis para abundância das espécies
mais comuns nas fisionomias. Negrito indica significância. CS, campo sujo; CC,
campo cerrado; CT, cerrado típico; CE, cerradão e NS, testes a posteriori
inconclusivos.
H P Testes a posteriori
Anuros
Chaunus ornatus 10,93 0,01 CE>CT CE>CC
Eupemphix nattereri 7,98 0,04 NS
Leptodactylus furnarius 7,73 0,05
L. fuscus 7,07 0,07
L. mystacinus 3,29 0,34
Physalaemus spp. gr. cuvieri 14,73 < 0,01 CC>CT CC>CE
P. fuscomaculatus 9,22 0,02 NS
Proceratophys sp 12,17 < 0,01 CS>CT CS>CE
Lagartos
Cercosaura schreibersii 6,34 0,1
Anolis meridionalis 6,32 0,09
Cnemidophorus cf. mumbuca 16 < 0,01 CS>CT CS>CE
Kentropyx paulensis 11,16 0,01 CS>CT CS>CE
28
anuros
0%
25%
50%
75%
100%
CS CC CT CE
C. ornatus P. spp.gr. cuvieriP. fuscomaculatus L. fuscusL. mystacinus L. furnariusProceratophys sp E. nattereriE. ovalis C. albopunctataC. schneideri L. labyrinthicusL. ocellatus
lagartos
0%
25%
50%
75%
100%
CS CC CT CE
K. paulensis C.cf.mumbucaC. schreibersii A. meridionalisM. atticolus A. ameivaM. dorsivittata C. modestaM. frenata C. cf. parecisT. merianae T.itambere
spp. gr. cuvieri cf. mumbuca
cf. parecis
Figura 8: Porcentagem das espécies de anuros e lagartos encontradas em
fisionomias de Cerrado na região de Itirapina. CS, campo sujo; CC, campo cerrado;
CT, cerrado típico; CE, cerradão.
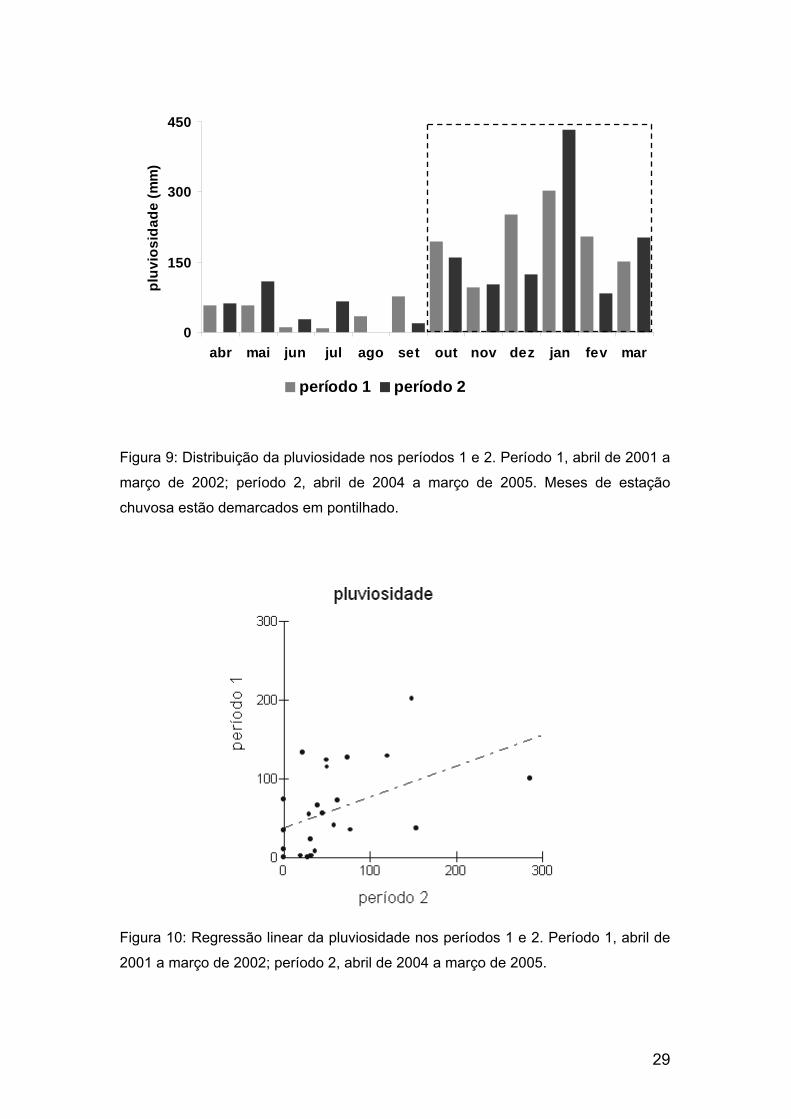
O clima nos períodos que compreendem as estações chuvosas
amostradas e as respectivas estações secas anteriores (períodos 1 e 2)
diferiu. Foi encontrada correlação positiva significativa apenas entre a
pluviosidade dos dois períodos (rs = 0,5; N = 24; p < 0,01); tanto a
temperatura mínima (rs = 0,2; N = 24; p < 0,34) quanto a máxima (rs = 0,02; N
= 24; p < 0,9) parecem variar independentemente nos dois períodos. Já a
pluviosidade se distribuiu temporalmente da mesma maneira nos dois
períodos (fig. 9), mas o coeficiente de inclinação da reta obtido através da
regressão linear foi muito inferior a 1 (R2 = 0,21; β = 0,4; p = 0,02), indicando
que o período 2 apresentou valores mais altos do que o período 1 (fig 10).
29
0
150
300
450
abr mai jun jul ago set out nov dez jan fev mar
pluv
iosi
dade
(mm
)
período 1 período 2
Figura 9: Distribuição da pluviosidade nos períodos 1 e 2. Período 1, abril de 2001 a
março de 2002; período 2, abril de 2004 a março de 2005. Meses de estação
chuvosa estão demarcados em pontilhado.
Figura 10: Regressão linear da pluviosidade nos períodos 1 e 2. Período 1, abril de
2001 a março de 2002; período 2, abril de 2004 a março de 2005.

30
2. Estrutura da vegetação e composição de espécies nas
fisionomias
Na análise de componentes principais (ACP, fig. 11) realizada com as
variáveis obtidas nas parcelas de vegetação, o cerradão e o cerrado típico
formaram grupos definidos e isolados, enquanto as fisionomias campestres
formaram grupos parcialmente sobrepostos entre si. O cerradão foi a
fisionomia mais homogênea.
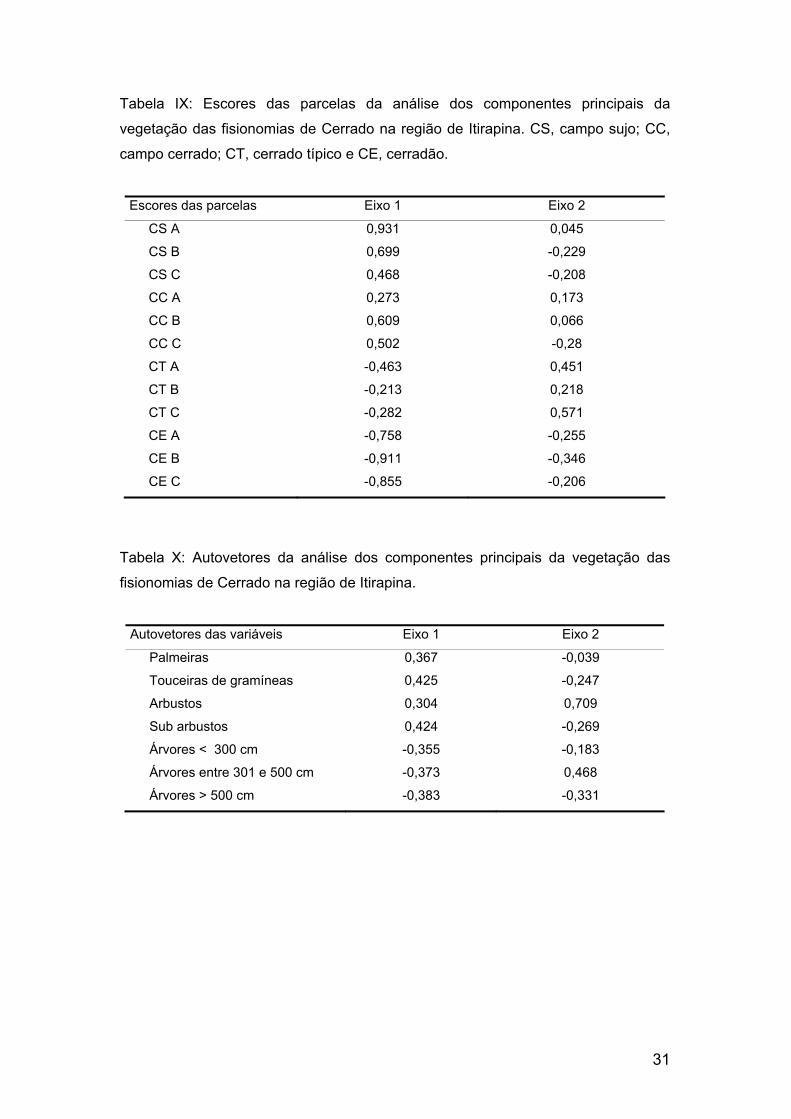
O eixo 1 explicou 67,8% da variância (tabela VIII). As parcelas das
fisionomias campestres foram ordenadas principalmente pelo número de
palmeiras, touceiras e subarbustos (tabela IX) no sentido dos valores
positivos (tabela X). Densidade de árvores ≤ 300 cm e > 500 cm foram as
variáveis principais (tabela IX) na ordenação das parcelas do cerradão e do
cerrado típico, no sentido dos valores negativos (tabela X). Já o eixo 2
ordenou principalmente as parcelas do cerrado típico pelo número de
arbustos e árvores entre 301 e 500 cm de altura (tabela IX) no sentido dos
valores positivos, e as do cerradão pelo número de árvores com mais de 500
cm de altura no sentido dos valores negativos, explicando mais 14,5% da
variância (tabela VIII).
Tabela VIII: Autovalores e porcentagem de variância explicada para os eixos 1 e 2
da análise dos componentes principais da vegetação das fisionomias de Cerrado na
região de Itirapina.
ACP Eixo 1 Eixo 2
Autovalores 4,75 1,01
Porcentagem (%) 67,81 14,50
31
Tabela IX: Escores das parcelas da análise dos componentes principais da
vegetação das fisionomias de Cerrado na região de Itirapina. CS, campo sujo; CC,
campo cerrado; CT, cerrado típico e CE, cerradão.
Escores das parcelas Eixo 1 Eixo 2
CS A 0,931 0,045
CS B 0,699 -0,229
CS C 0,468 -0,208
CC A 0,273 0,173
CC B 0,609 0,066
CC C 0,502 -0,28
CT A -0,463 0,451
CT B -0,213 0,218
CT C -0,282 0,571
CE A -0,758 -0,255
CE B -0,911 -0,346
CE C -0,855 -0,206
Tabela X: Autovetores da análise dos componentes principais da vegetação das
fisionomias de Cerrado na região de Itirapina.
Autovetores das variáveis Eixo 1 Eixo 2
Palmeiras 0,367 -0,039
Touceiras de gramíneas 0,425 -0,247
Arbustos 0,304 0,709
Sub arbustos 0,424 -0,269
Árvores < 300 cm -0,355 -0,183
Árvores entre 301 e 500 cm -0,373 0,468
Árvores > 500 cm -0,383 -0,331

32
-1
0
1
-1 0 1
campo sujocampo cerradocerrado típicocerradão
arb
301 - 500
> 500
< 300
palmtou
sub
Figura 11: Diagrama “biplot” da análise dos componentes principais da estrutura da
vegetação das fisionomias. Arb, arbustos; palm, palmeiras; tou, touceiras de
gramíneas; sub, subarbustos; < 300, árvores com alturas menores ou iguais a 300
cm; 301-500, árvores entre 301 e 500 cm de altura; > 500, árvores com mais de 500
cm de altura.
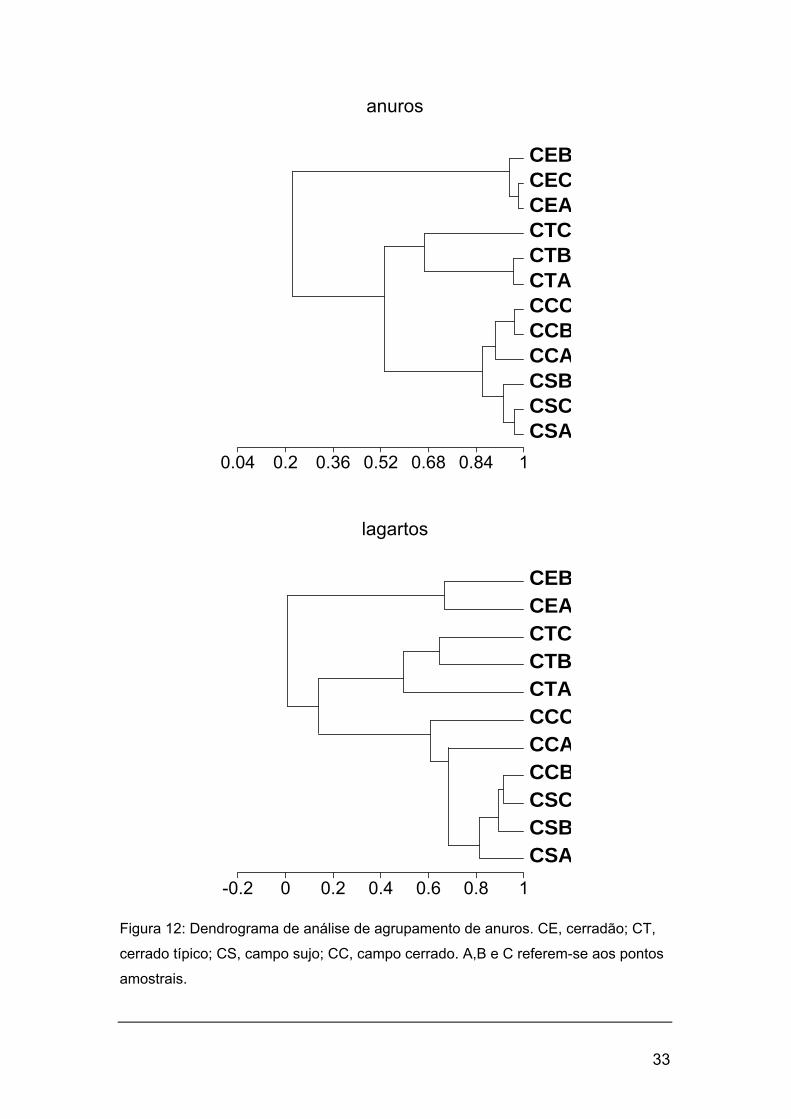
As análises de agrupamento (fig. 12) formaram quatro grupos coesos,
concordantes para ambas taxocenoses. O cerradão representa o grupo mais
isolado. O cerrado típico e as fisionomias campestres formaram um segundo
grupo de fisionomias savânicas, por sua vez subdividido em dois: um
contendo o cerrado típico e outro as fisionomias campestres. As fisionomias
campestres não formaram um grupo coeso segundo suas faunas de lagartos,
mas sim segundo suas faunas de anuros (fig 12).
33
anuros
CSACSCCSBCCACCBCCCCTACTBCTCCEACECCEB
0.04 0.2 0.36 0.52 0.68 0.84 1
lagartos
CSACSBCSCCCBCCACCCCTACTBCTCCEACEB
-0.2 0 0.2 0.4 0.6 0.8 1
Figura 12: Dendrograma de análise de agrupamento de anuros. CE, cerradão; CT,
cerrado típico; CS, campo sujo; CC, campo cerrado. A,B e C referem-se aos pontos
amostrais.

34
V. Discussão
De um modo geral, a diversidade de anuros e lagartos na região de
Itirapina variou nas diferentes fisionomias de forma semelhante para ambas
taxocenoses: as fisionomias abertas de campo sujo e campo cerrado foram
as mais ricas, seguidas pelo cerrado típico e, por último, o cerradão. As
abundâncias dos anuros não apresentaram o mesmo padrão geral, o
cerradão apresentou maior abundância devido ao número elevado de
capturas de Chaunus ornatus. Esta discrepância provavelmente se deu pelo
recrutamento de jovens desta espécie (observação pessoal); ao
desconsiderá-la o padrão de abundância segue o de riqueza, com valores
que crescem no sentido das fisionomias abertas.
Embora escassos, os dados da literatura de diferentes regiões
sugerem que os cerrados abertos e savânicos tendem a apresentar riquezas
e abundâncias mais altas do que formações florestais no domínio do
Cerrado. Na região do Rio Manso (MT), STRÜSSMANN (2000) encontrou
riquezas e abundâncias de anuros e de lagartos maiores nas fisionomias
abertas de cerrado do que nas matas de galeria, matas ciliares e cerradões.
O mesmo ocorreu na região de Brasília (DF) para anuros (BRANDÃO &
ARAÚJO, 2001 e BRANDÃO & ARAÚJO, 1998 apud COLLI et al 2002) e
lagartos (NOGUEIRA et al, 2005). Ainda para lagartos, VALDUJO (2003)
encontrou maior riqueza nas fisionomias campestres e savânicas no Parque
Nacional das Emas (GO), assim como NOGUEIRA (2006) em mais nove
outras localidades do bioma.

35
Além da riqueza e da abundância, as composições das taxocenoses
de anuros e lagartos em Itirapina diferiram entre fisionomias de forma
parecida. Padrões concordantes na estrutura da vegetação e composição de
espécies sugerem que o hábitat exerceu grande influência na distribuição dos
anuros e dos lagartos, principalmente nas fisionomias fechadas de cerradão
e de cerrado típico. A maior semelhança na composição de espécies do
cerrado típico com fisionomias abertas e não com o cerradão provavelmente
resulta de maior tolerância de espécies oriundas das fisionomias campestres
em ocupar esta fisionomia; o cerradão foi uma fisionomia aparentemente
pouco habitável para a maioria das espécies savânicas. Segundo VALDUJO
(2003), a falta de radiação solar direta no nível do solo torna este hábitat
pouco favorável para muitas espécies de lagartos.
Por outro lado, fisionomias abertas não apresentaram padrões de
composição concordantes para ambas taxocenoses. Para lagartos, a
sobreposição na composição de espécies sugere que estas fisionomias
representam uma única unidade contínua de hábitat. Para os anuros, a
formação de composições individuais no campo sujo e no campo cerrado
parece sugerir uma fauna com maior especialização de hábitat. Esta
discrepância entre taxocenoses coloca em questão se a sobreposição da
estrutura da vegetação entre o campo sujo e o campo cerrado não é
decorrente de escolha imprópria de variáveis para a análise. Ou então, se a
distribuição das espécies de anuros nestas fisionomias pode ter outras
razões que não a estrutura da vegetação.
De qualquer maneira, a influência do hábitat sobre a herpetofauna do
Cerrado já havia sido observada anteriormente. Na região de Brasília,

36
NOGUEIRA et al. (2005) encontraram diferentes conjuntos de espécies de
lagartos em hábitats semelhantes aos de Itirapina. As composições das
espécies apresentaram as mesmas relações de semelhança nas duas
localidades; florestas de galeria se segregaram das áreas de Cerrado aberto
(assim como o cerradão em Itirapina) e áreas de cerrado aberto, por sua vez,
se dividiram em cerrado típico e áreas de campos (NOGUEIRA et al. 2005).
Segundo estes dois estudos, a herpetofauna do Cerrado parece estar
subdivida em taxocenoses florestais e savânicas, sendo as últimas divididas
em campestres e de cerrado típico.
Algumas das espécies encontradas freqüentemente em Itirapina já
haviam sido mencionadas por outros autores em hábitats muito parecidos,
porém em outras localidades de Cerrado. Entre anuros, Physalaemus
centralis, P. cuvieri, P. fuscomaculatus, Eupemphix nattereri, Leptodactylus
furnarius e L. fuscus parecem habitar preferencialmente fisionomias
campestres e cerrado típico (STRÜSSMANN, 2000; BRANDÃO & ARAÚJO,
2001; BASTOS et al, 2003). Entre os lagartos, Anolis meridionalis,
Cnemidophorus cf. mumbuca e Kentropyx paulensis também parecem ser
espécies típicas de formações campestres, sendo C. cf. mumbuca e K.
paulensis encontradas em abundância em locais de solo arenoso (COLLI et
al., 2002; COLLI et al., 2003; VALDUJO, 2003; STRÜSMANN, 2003;
NOGUEIRA et al., 2005; NOGUEIRA, 2006). Das espécies de anuros e
lagartos mais comuns em cerrado típico e cerradão, Colobosaura modesta
parece ocorrer exclusivamente nestas formações (STRUSSMANN, 2000;
VALDUJO, 2003; NOGUEIRA, 2006) enquanto Chaunus ornatus é uma das
espécies mais abundantes em áreas perturbadas na Mata Atlântica (DIXO,

37
2000; 2005), raramente encontrada em levantamentos de fauna do Cerrado.
Estes registros corroboram as interpretações de COLLI et al. (2002) e
NOGUEIRA et al. (2005) de que a herpetofauna do Cerrado não é formada
por espécies generalistas, mas distribuída de forma não aleatória entre
hábitats (NOGUEIRA, 2006). A repetição das mesmas espécies em hábitats
semelhantes em localidades diferentes parece indicar que esta dimensão do
nicho permanece muito conservada mesmo ao longo de grandes distâncias.
A existência de relações entre a fauna e a estruturação do hábitat é
cada vez mais clara no Cerrado. Entretanto, a concentração da diversidade
nas fisionomias abertas em Itirapina e em outras localidades contrariou a
hipótese de que a estruturação do hábitat e a produtividade seriam agentes
promotores da biodiversidade no Cerrado, assim como supostamente o são
nos biomas florestais (PIANKA, 1994; BEGON et al., 2005). Ao invés disso,
suporta a hipótese alternativa de que a diversidade nas fisionomias é
condicionada por suas disponibilidades, havendo predomínio dos processos
regionais sobre processos locais. Aparentemente, na paisagem da região de
Itirapina, as manchas de fisionomias mais fechadas não são suficientes para
manter, na paisagem, metapopulações de espécies especialistas nestes
habitats. Assim, a disponibilidade de área e/ ou o isolamento das fisionomias
fechadas da paisagem da região de Itirapina pode ter comprometido o
surgimento e/ ou a permanência de outras espécies de anfíbios e lagartos
especialistas nestes habitats.
Para lagartos, isso já havia sido apontado por NOGUEIRA (2006), que
atribuiu a baixa diversidade encontrada na Serra da Bodoquena (NOGUEIRA
2006) e em Lavrado (por VITT, 1996) ao domínio de formações florestais em

38
detrimento das formações abertas nessas regiões. Em Itirapina, outros
estudos corroboram esta tendência, como é o caso da riqueza dos
mamíferos de médio e grande porte (HÜLLE, 2006), mais alta nas
fisionomias campestres. No Cerrado, diferentes regiões apresentam
diferentes proporções de fisionomias abertas e fechadas (RATTER et al,
1997, OLIVEIRA-FILHO & RATTER, 2002). Áreas grandes de vegetação
densa, como florestas e cerradões (24% do bioma; SILVA & BATES, 2002),
podem ter sido capazes de produzir e manter uma fauna própria. A
investigação destas áreas pode fornecer resultados mais conclusivos sobre
este assunto.
A restrição de algumas espécies a ambientes específicos parece ter
tido papel fundamental na distribuição da herpetofauna do Cerrado; padrões
de uso de hábitat por lagartos podem ser considerados evidência de
associação entre a história da paisagem e conservação de seus nichos
(NOGUEIRA, 2006). Durante flutuações climáticas drásticas, a inércia
filogenética provavelmente impõe limitações à distribuição das populações.
Quando a vegetação de área aberta se expandiu durante os ciclos mais frios
do Quaternário (PRANCE, 1982), linhagens de área aberta podem ter sido
favorecidas, enquanto linhagens de formações fechadas podem ter se
isolado naturalmente, com eventuais extinções locais. Considerando estes
eventos, a disponibilidade regional das fisionomias parece ser determinante
na composição de espécies (ver GAINSBURY & COLLI, 2003).
A inércia filogenética parece ter contribuído no passado para modelar
comunidades em diferentes localidades de forma similar, tendo como produto
um padrão repetido no uso do hábitat, mais um indicativo da predominância

39
dos fatores históricos regionais sobre os locais (BROOKS & McLENNAN,
1991). Atualmente, a inércia filogenética continua uma força relevante,
dificultando ou impedindo a colonização das fisionomias fechadas pela
herpetofauna de área aberta (e vice-versa); hábitats abertos e florestados
podem atuar como barreiras mútuas para os lagartos (NOGUEIRA, 2006),
limitando a sobreposição de fauna.
Existe ainda a possibilidade de que a baixa riqueza nos fragmentos de
Cerrado denso em Itirapina seja oriunda de extinções locais modernas
causadas pelo homem, e não por processos históricos. Entretanto, os poucos
estudos sobre a fragmentação no Cerrado não indicaram quaisquer efeitos
de área sobre a riqueza de lagartos (COLLI, 2003; GAINSBURY & COLLI,
2003), embora estes estudos tenham sido feitos em escala muito maior.
Além disso, as amostragens nas fisionomias abertas e fechadas não foram
concomitantes e este resultado pode ter sido enviesado pela variação no
regime de chuvas nos dois períodos (ver COLLI, 1991; VITT & COLLI, 1994;
COLLI et al., 1997; MESQUITA & COLLI, 2003; GIARETTA & MENIN, 2004;
BRASILEIRO et al., 2005).
Atualmente, cresce a idéia de que mecanismos locais e regionais
interagem em um contínuo de tempo e espaço para produzir variações
geográficas na estrutura de comunidades (RICKLEFS, 2004). Entretanto, a
contribuição individual dos processos locais e regionais como determinantes
da diversidade local ainda é pouco compreendida (RICKLEFS, 1987;
CORNELL & LAWTON, 1992; GASTON, 2000), já que estes processos
atuam em diferentes escalas temporais e espaciais (HILLEBRAND &
BLENKCNER, 2002). Apesar do atual estado de conservação do Cerrado,

40
características como área extensa (SILVA & BATES, 2002), antigüidade
(RATTER et al., 1996) e heterogeneidade de ambientes (EITEN, 1972)
conferem a este bioma grande potencial para estudo de padrões de
diversidade em escala local, regional e continental. A abundância, riqueza
(COLLI et al, 2002) e endemismo (MYERS et al, 2000) dos anuros e lagartos
no Cerrado, associados à disponibilidade de métodos de captura
padronizados e de fácil aplicação, podem fazer desses grupos organismos-
modelo para este tipo de estudo.

41
VI. Referências Bibliográficas
AYRES, M.; M. AYRES JR.; D. L. AYRES & A. S. DOS SANTOS. 1998.
BioEstat: aplicações estatísticas nas áreas das ciências biológicas e
médicas. Publicações Avulsas Mamirauá. 193p. Acompanha CD-ROM
para PC compatível com Windows 95. Manaus: Sociedade Civil
Mamirauá.
BASTOS, R.P; J.A.O. MOTTA, L.P. LIMA & L.D.A. GUIMARÃES. 2003.
Anfíbios da Floresta nacional de Silvânia, estado de Goiás. Editora
Stylo, Goiânia.
BEGON, M.; J. L. HARPER & C. R. TOWSEND 2005. Ecology: Individuals,
Populations, and Communities. Blackwell Science Ltd, Oxford.
BRANDÃO, R. A. & A. F. B. ARAUJO. 1998. A herpetofauna da Estação
Ecológica de Águas Emendadas. Em J. S. MARINHO-FILHO, ed.,
Vertebrados da Estação Ecológica de Águas Emendadas: História
Natural e Ecologia de um Fragmento de Cerrado do Brasil Central.
Instituto de Ecologia e Meio Ambiente do Distrito Federal, Brasília.
BRANDÃO, R.A & A.F.B. ARAÚJO. 2001. A herpetofauna associada às
matas de galeria no Distrito Federal. Em RIBEIRO, J.F.; C.E.L.
FONSECA & J.C. SOUSA-SILVA. Cerrado: caracterização e
recuperação das matas de galeria. Embrapa, Planaltina.
BRASILEIRO, C. A. 2004. Diversidade de Anfíbios Anuros em Área de
Cerrado no Estado de São Paulo. Tese de Doutorado. IB- Universidade
de São Paulo, SP, Brasil.

42
BRASILEIRO, C. A.; R. J. SAWAYA; M. C. KIEFER & M. MARTINS. 2005.
Amphibians of an open Cerrado fragment in Southeastern Brazil. Biota
Neotropica:http://www.biotaneotropica.org.br/v5n2/pt/abstract?article+BN004050220
05
BROOKS, D. R. & D. A. McLENNAN. 1991. Phylogeny, Ecology and
Behavior. The University of Chicago Press, Chicago and London.
CALEY, J. C. & D. SCHLUTER. 1997. The relationship between local and
regional diversity. Ecology, 78: 70-80.
CAVALCANTI, R. B. & C. A. JOLY. 2002. Biodiversity and conservation
priorities in the Cerrados. Em: OLIVEIRA, P.S. & R.J MARQUIS (eds.).
The Cerrados of Brazil: Ecology and Natural History of a Neotropical
Savanna. Columbia University Press, Irvington.
CECHIN, S. Z.& M. MARTINS 2000. Eficiência de armadilhas de queda
(pitfall traps) em amostragens de anfíbios e répteis no Brasil. Revista
Brasileira de Zoologia, 17: 729-740.
COLLI, G. R. 1991. Reproductive ecology of Ameiva ameiva in the Cerrado of
central Brazil. Copeia, 1991: 1002-1012.
COLLI, G. R. 1998. Biogeografia e conservação da herpetofauna no Cerrado,
Pantanal e Savanas Amazônicas: Relatório final do grupo temático
Herpetofauna. Em: Workshop Ações Prioritárias para Conservação
da Biodiversidade do Cerrado e Pantanal. Ministério do Meio
Ambiente, FUNATURA, Conservation International, Fundação
Biodiversitas, Universidade de Brasília. Web:
www.bdt.org.br/workshop/cerrado/br.

43
COLLI, G. R. 2003. Estrutura de taxocenoses de lagartos em fragmentos
naturais e antrópicos de Cerrado. Em: CLAUDINO-SALES (Ed):
Ecossistemas Brasileiros: Manejo e Conservação. Expressão Gráfica
e Editora, Fortaleza.
COLLI, G. R., A. K. PÉRES JR. & M. G. ZATZ. 1997. Foraging mode and
reproductive seasonality in tropical lizards. Journal of Herpetology, 31:
490-499.
COLLI, G. R.; R. P. BASTOS & F. B. ARAUJO. 2002. The character and
dynamics of the Cerrado herpetofauna. Em: OLIVEIRA, P.S. & R. J.
MARQUIS. (eds.) The Cerrados of Brazil: Ecology and Natural History
of a Neotropical Savanna. Columbia University Press, Columbia.
COLLI, G. R; J. P. CADWELL, G. C. COSTA, A. M. GAINSBURY, A. A.
GARDA, D. O. MESQUITA, C. M. M. R. FILHO, A. H. B. SOARES, V. N.
SILVA, P. H. VALDUJO, G. H. C. VIEIRA, L. J. VITT, F. P. WERNECK,
H. C. WIEDERHECKER & M. ZATS. 2003. A new species of
Cnemidophorus (Squamata: Teiidae) from the Cerrado biome in central
Brazil. Occasional Papers of the Oklahoma Museum of Natural
History 14: 1-14.
COLWELL, R. K. 2005. EstimateS: Statistical estimation of species
richness and shared species from samples. Version 7.5. User's Guide
and application published at: http://purl.oclc.org/estimates.
COLWELL, R. K. & J. A. CODDINGTON. 1994. Estimating terrestrial
biodiversity through extrapolation. Philosophical Transactions of the
Royal Society (Series B), 345:101-118.

44
CORNELL, H. V. & J. H. LAWTON. 1992. Species interactions, local and
regional processes, and limits to the richness of ecological communities:
a theoretical perspective. Journal of Animal Ecology, 61:1-12.
COUTINHO, L. M. 1978. O conceito de Cerrado. Revista Brasileira de
Botânica, 1: 17-23.
DINIZ-FILHO, J. A. F.; R. P. BASTOS; T. F. V. L. RANGEL; L. M. BINI; P.
CARVALHO & R. J. SILVA. 2005. Macroecological correlates and spatial
patterns of anuran description dates in the Brazilian Cerrado. Global
Ecology and Biogeography, 14: 469–477
DIXO, M. B. O. 2001. Efeito da fragmentação da floresta sobre a comunidade
de sapos e lagartos de serapilheira no sul da Bahia. Dissertação de
mestrado. IB- Universidade de São Paulo, SP, Brasil.
DIXO, M. B. O. 2005 . Diversidade de sapos e lagartos de serrapilheira numa
paisagem fragmentada do Planalto Atlântico de São Paulo. Tese de
Doutorado. IB- Universidade de São Paulo, SP, Brasil.
EITEN, G. 1972. The Cerrado vegetation of Brazil. Botanical Review,
38:201-341.
GOTTELI, N. & R. K. COLWELL. 2001. Quantifying biodiversity: Procedures
and pitfalls in the measurement and comparison of species richness.
Ecology Letters, 4: 379-391.
GAINSBURY, A. M. & G. R. COLLI 2003. Lizard assemblages from natural
Cerrado enclaves in southwestern Amazonia: The role of stochastic
extinctions and isolation. Biotropica, 35: 503 – 519.
GASTON, K. J. 2000. Global patterns in biodiversity. Nature, 405: 220-227.

45
GIARETTA, A. A. & M. MENIN. 2004. Reproduction, phenology, and mortality
sources of Physalaemus (Anura:Leptodactylidae). Journal of Natural
History, 38: 1711-1722.
GOTTELI, N. J. & G. L. ENTSMINGER. 2004. EcoSim: Null models
software for ecology. Version 7. Acquired Intelligence Inc. & Kesey-
Bear. Jericho, VT 05465. http://garyentsminger.com/ecosim/index.htm.
GOTELLI, N. J. & G. R. GRAVES. 1996. Null Models in Ecology.
Smithsonian Institution Press, Washington, DC.
GREENBERG, C. H.; NEARY, D.G. & L. D. HARRIS,1994. Comparison of
herpetofaunal sampling effectiveness of pitfall, single-ended, and double-
ended funnel traps used with drift fences. Journal of Herpetology,
28:319-324.
HEYER, W.R. 1988. On frog distribution patterns east of the Andes. Em:
Vanzolini, P. E. & W. R. Heyer (eds.) Proceedings of a Workshop on
Neotropical Distribution Patterns. Academia Brasileira de Ciências, Rio
de Janeiro.
HILLEBRAND, H. & T. BLENCKNER. 2002. Regional and local impact on
species diversity – from pattern to processes. Oecologia, 132:479–491
HOFER, U. & L. F. BERSIER. 2001. Herpetofaunal diversity and abundance
in tropical upland forests of Cameroon and Panama. Biotropica, 33: 142-
152.
HOLT, R. D.1993. Ecology at the mesoscale: The influence of regional
processes on local communities. Em: Ricklefs, R. E. & Schluter, D.
(Eds.). 1993. Species Diversity in Ecological Communities: Historical
and Geographical Perspectives. Univ. Chicago Press, Chicago, Illinois.

46
HULBERT, S. H. 1971. The nonconcept of species diversity: a critique and
alternative parameters. Ecology, 52: 577-585.
HÜLLE, N.L. 2006. Mamíferos de médio e grande porte num remanescente
de Cerrado no sudeste do Brasil (Itirapina, SP). Dissertação de mestrado,
IB- Universidade de São Paulo, SP, Brasil.
KLINK, C. A. & R. B. MACHADO 2005. Conservation of the brazilian Cerrado.
Conservation Biology, 19: 707-713.
KOVACH W. L. 2000. MVSP - A multi-variate statistical package for
Windows, version 3.1. Penthraeth: Kovach Computing Services.
MAGURRAN, A. E. 1988. Ecological diversity and its measurement.
London: Croom Helm Ltd.
MESQUITA, D. O. & G. R. COLLI. 2003. The ecology of Cnemidophorus
ocellifer (Squamata, Teiidae) in a neotropical savanna. Journal of
Herpetology, 37: 498-509.
MINGOTI, S. A. 2005. Análise de dados através de métodos de
estatística multivariada: Uma abordagem aplicada. Editora UFMG,
Belo Horizonte.
MITTERMEYER, R. A.; N. MYERS & C. G. MITTERMEYER. 2000.
Hotspots: Earth’s Biologically Richest and Most Endangered
Terrestrial Ecoregions. CEMEX, Mexico.
MOUQUET, N. & M. LOREAU. 2002. Coexistence in metacommunities: the
regional similarity hypothesis. The American Naturalist, 159: 420-426.
MYERS, N.; R. A. MITTERMEYER; C. G. MITTERMEYER; G. A. B
FONSECA & J. KENT. 2000. Biodiversity hotspots for conservation
priorities. Nature, 403: 853-858.

47
NOGUEIRA, C. 2006. Diversidade e padrões de distribuição da fauna de
lagartos no Cerrado. Tese de doutorado, IB- Universidade de São Paulo,
SP, Brasil.
NOGUEIRA, C.; P. H. VALDUJO & F.G. R. FRANÇA. 2005. Habitat variation
and lizard diversity in a Cerrado area of Central Brazil. Studies on
Neotropical Fauna and Environment, 40: 105 – 112.
OLIVEIRA-FILHO, A. T. & J. A. RATTER. 2002. Vegetation physiognomies
and woody flora of the Cerrado Biome. Em: OLIVEIRA, P. S. & R. J.
MARQUIS. (eds.) The Cerrados of Brazil: Ecology and Natural History
of a Neotropical Savanna. Columbia University Press, Irvington.
PIANKA, E. R. 1994. Evolutionary Ecology. HarperCollins College
Publishers.
PRANCE, G.T. 1982. A review of the phytogeographic evidence for
pleistocene climate changes in the neotropics . Annals of the Missouri
Botanical Garden 69: 594-624.
RATANA, P.; A. HUETE & L. FERREIRA 2005. Analysis of Cerrado
Physiognomies and Conversion in the MODIS Seasonal-Temporal
Domain. Earth Interactions, 9, 1-22.
RATTER, J. A.; J. F. RIBEIRO, & S. BRIDGEWATER 1997. The brazilian
Cerrado vegetation and Threats to its biodiversity. Annals of Botany, 80:
223-230.
RICKLEFS, R. E. 1987. Community diversity: relative roles of local and
regional processes. Science, 235:167–171.
RICKLEFS, R. E. 2004. A comprehensive framework for global patterns in
biodiversity. Ecology Letters, 7:1-15.

48
RICKLEFS, R. E. & D. SCHLUTER (Eds.). 1993. Species Diversity in
Ecological Communities: Historical and Geographical Perspectives.
Univ. Chicago Press, Chicago.
SANDERS, H. L. 1968. Marine benthic diversity: a comparative study.
American Naturalist, 102: 243-282.
SHURIN, J. B. & E. G. ALLEN. 2001. Effect of competition, predation, and
dispersal on species richness at local and regional scales. The American
Naturalist, 158: 624-637.
SILVA J. M. C. & J. M. BATES. 2002. Biogeographic patterns and
conservation in the South American Cerrado: A tropical savanna hotspot.
BioScience, 52:225-233.
SILVA Jr,, N. J. & J. W. SITES Jr. 1995. Patterns of of diversity of neotropical
squamate reptile species with emphasis on the Brazilian Amazon and the
conservation potential of indigenous reserves. Conservation Biology,
Malden, 9:873-901.
SIMBERLOFF, D. 1978. Use of rarefaction and related methods in ecology.
Em: Dickson K. L., Cairns J. Jr & Livingston R. J., (eds) Biological Data
in Water Pollution Assessment: Quantitative and Statistical
Analyses. American Society for Testing and Materials, Philadelphia.
STRUSMANN, C. 2000. Herpetofauna. Em Alho, C. J. R (ed). Fauna
Silvestre da região do Rio Manso- MT. Edições Ibama, Brasília.
TAYLOR, D. R.; L. W. ARSSEN & C. LOEHLE. 1990. On the relationship
between r/K selection and environmental carrying capacity: new habitat
templet for plant life history strategies. Oikos, 58: 239-250.

49
VALDUJO, P.H. 2003. Distribuição da comunidade de lagartos em um
gradiente de vegetação no cerrado e suas implicações para a
conservação. Dissertação de mestrado. IB- Universidade de Brasília, DF,
Brasil.
VANZOLINI P. E. 1976. On the lizards of a Cerrado – Caatinga contact,
evolutionary and zoogeographical implications (Sauria). Papéis Avulsos
de Zoologia, 29: 111–119.
VICENTE, L. E., C. R. SOUZA FILHO & A. PERES FILHO. 2005.
Mapeamento de formações arenosas em fragmentos de Cerrado
utilizando dados e produtos do sensor ASTER. Anais XII Simpósio
Brasileiro de Sensoriamento Remoto, Goiânia, Brasil, 16-21 abril
2005, INPE,.
VITT, L. J. 1991. An introduction to the ecology of Cerrado lizards. Journal of
Herpetology, 25: 79-90.
VITT L.J. 1996. Biodiversity of Amazonian Lizards. Em GIBSON, A.C. (ed),
Neotropical Biodiversity and Conservation. Occasional Publication of
the Mildred E. Mathias Botanical Garden 1, Los Angeles.
VITT, L. J. & G. R. COLLI. 1994. The geographical ecology of a neotropical
lizard: Ameiva ameiva (Teiidae) in Brazil. Canadian Journal of Zoology,
72(11): 1986-2008.
VOURLITIS, G. L.; N. P. FILHO; M. M. S. HAYASHI; J. S. NOGUEIRA; F. T.
CASEIRO & J. H. CAMPELO Jr. 2001. Seasonal variations in the net
ecosystem CO2 exchange of a mature Amazonian transitional tropical
forest (cerradão). Functional Ecology, 15: 388-395.

50
WEBB, S. D. 1978. A history of savanna vertebrates in the new world: Part II,
South America and the great interchange. Annual Review of Ecology
and Systematics, 9:393-426.
ZAR, J. H.1996. Biostatistical Analysis. Third ed. Prentice-Hall.
ZOBEL, M. 1992. Plant species coexistence: The role of historical,
evolutionary, and ecological factors. Oikos, 65: 314-320.
ZOBEL, M. 1997. The relative role of species pools in determining plant
species richness: an alternative explanation of species coexistence?
Trends in Ecology and Evolution, 12: 266-269.

51
VII. Resumo/ Abstract
Diversas teorias foram propostas para explicar a ocorrência de áreas
de grande diversidade no mundo. O Cerrado é a savana tropical mais rica,
apresentando herpetofauna diversa e endêmica distribuída em diferentes
formações vegetais. Estas formações variam de florestas a campos e
representam hábitats distintos para a herpetofauna. Na região de Itirapina,
porção sul do domínio do Cerrado, a diversidade de anuros e lagartos foi
comparada entre fisionomias abertas e fechadas para testar se a riqueza
destes animais é maior em ambientes mais produtivos e estruturalmente
mais complexos. Para isso foram feitas amostragens em duas estações
chuvosas com uso de armadilhas de queda. As fisionomias abertas foram
mais diversas do que fisionomias fechadas, uma tendência observada em
outras regiões. Neste estudo, foi encontrada relação entre estrutura da
vegetação e composição de espécies, indicando que o hábitat exerce grande
influência da distribuição das espécies. Ainda, o padrão de distribuição das
espécies mais abundantes foi concordante com a literatura de diferentes
regiões, sugerindo conservação de nicho por longas distâncias. Entretanto, a
hipótese de maior riqueza em ambientes mais produtivos e estruturalmente
mais complexos foi rejeitada. A hipótese alternativa implica em um efeito
regional causado por baixa disponibilidade de fisionomias fechadas em
relação às abertas. Estes resultados evidenciam fatores históricos e
regionais como determinantes nos padrões de diversidade no Cerrado.

52
Several theories have been proposed to explain areas of high diversity
worldwide. The Cerrado is the most diverse tropical savanna, with a diverse,
endemic herpetofauna distributed in different types of vegetation. The cerrado
vegetational types vary from forest to grasslands and represent distinct
habitats for the herpetofauna. In the region of Itirapina, southeastern extent of
the Cerrado domain, the diversity of frogs and lizards among these distinct
physiognomies was compared to test the hypothesis that the richness of
these animals is higher in more productive and more structurally complex
habitats. Sampling occurred over two rainy seasons by pitfall trapping. Open
formations were more diverse than closed ones. Literature information from
other regions suggests that there is a tendency of open forms to be richer.
The correlation between species composition and vegetation structure
indicates that habitat plays an important role in determining species
distributions. The distribution pattern of the most abundant species was
corroborated by the literature from different regions, suggesting niche
conservatism over long geographic distances. However, the hypothesis of
higher richness in more productive and more structurally complex habitats
was not corroborated. The proposed alternative hypothesis implies a regional
effect caused by low availability of closed formations in relation to open ones.
These results underscore the importance of historical and regional factors on
Cerrado diversity patterns.