
KARAKTERISASI SENYAWA BIOAKTIF ISOFLAVON DAN UJI AKTIVITAS ANTIOKSIDAN DARI EKSTRAK
TEMPE BERBAHAN BAKU BUNCIS (Phaseolus vulgaris) DAN KECIPIR (Psophocarpus tetragonolobus)
TESIS Disusun untuk memenuhi sebagian persyaratan
guna memperoleh gelar Magister Sains Program Studi Biosains
oleh
SRI WAHYUNI
S 900208023
PROGRAM PASCA SARJANA BIOSAINS UNIVERSITAS SEBELAS MARET
SURAKARTA 2010

ii
KARAKTERISASI SENYAWA BIOAKTIF ISOFLAVON DAN UJI AKTIVITAS ANTIOKSIDAN DARI EKSTRAK TEMPE
BERBAHAN BAKU BUNCIS (Phaseolus vulgaris) DAN KECIPIR (Psophocarpus tetragonolobus)
Disusun oleh
Nama : Sri Wahyuni
Nim : S 900208023
Telah disetujui oleh Tim Pembimbing
Dosen Pembimbing Nama Tanda tangan Tanggal
Pembimbing I Prof. Ir. Sri Handajani, M.S.,Ph.D .......................... ........ 2010 NIP: 19470729 197612 2 001
Pembimbing II Dr. Artini Pangastuti, M.Si .......................... ......... 2010 NIP: 19750531 200003 2 001
Mengetahui
Ketua Program Studi Biosains
Dr. Sugiyarto, M.Si NIP. 19670430 199203 1.002

iii
KARAKTERISASI SENYAWA BIOAKTIF ISOFLAVON DAN UJI AKTIVITAS ANTIOKSIDAN DARI EKSTRAK TEMPE
BERBAHAN BAKU BUNCIS (Phaseolus vulgaris ) DAN KECIPIR (Psophocarpus tetragonolobus)
TESIS
Oleh Sri Wahyuni
S 900208023 Telah dipertahankan di depan penguji
Dinyatakan telah memenuhi syarat pada tanggal, 25 Januari 2010
Jabatan Nama Tanda tangan Tanggal
Ketua Dr. Sugiyarto, M.Si ..................... .......... 2010 NIP. 19670430 199203 1 002 Sekretaris Dr. Edwi Mahajoeno, M.Si ..................... .......... 2010 NIP. 19601025 199702 1 001
Anggota Prof. Ir. Sri Handajani, M.S.,Ph.D ..……………. ...…… 2010 Penguji NIP. 19470729 197612 2 001 Dr. Artini Pangastuti, M.Si ……………… ……… 2010 NIP.19750531 200003 2 001
Mengetahui
Direktur Program Pascasarjana UNS Ketua Program Studi Biosains
Prof.Drs. Suranto,M.Sc.,PhD Dr. Sugiyarto, M.Si NIP.19570820 198503 1 004 NIP.19670430 199203 1 002

iv
PERNYATAAN ORISINALITAS TESIS
Saya menyatakan dengan sebenar-benarnya bahwa :
1. Tesis dengan judul : “Karakterisasi senyawa bioaktif isoflavon dan uji
aktivitas antioksidan dari ekstrak tempe berbahan baku Buncis
(Phaseolus vulgaris) dan Kecipir (Psophocarpus tetragonolobus)” ini
merupakan sebagian karya penelitian dari Prof. Ir. Sri Handayani, M.S. dan Sri
Retno Dwi Ariani, M.Si dan tidak terdapat karya ilmiah yang pernah diajukan
oleh orang lain untuk memperoleh gelar akademik serta tidak terdapat karya
atau pendapat yang pernah ditulis atau diterbitkan oleh orang lain, kecuali
yang secara tertulis dikutip dalam naskah ini dan disebutkan dalam sumber
kutipan dan daftar pustaka. Apabila ternyata di dalam naskah tesis ini dapat
dibuktikan terdapat unsur-unsur jiplakan, maka saya bersedia Tesis beserta
gelar MAGISTER saya dibatalkan, serta diproses sesuai dengan peraturan
perundang-undangan yang berlaku (UU No. 20 Tahun 2003, pasal 25 ayat 2
dan pasal 70).
Surakarta, Januari 2010
Mahasiswa
Sri Wahyuni S 900208023

v
KARAKTERISASI SENYAWA BIOAKTIF ISOFLAVON DAN UJI AKTIVITAS ANTIOKSIDAN DARI EKSTRAK TEMPE BERBAHAN BAKU BUNCIS (Phaseolus vulgaris) DAN KECIPIR (Psopocarpus tetragonolobus )
Sri Wahyuni, Sri Handajani, Artini Pangastuti
Program Studi Magister Biosains, PPS-UNS Surakarta
ABSTRAK
Biji buncis dan kecipir yang tergolong leguminoceae berpotensi sebagai sumber antioksidan alami. Penelitian ini bertujuan mengetahui kandungan isoflavon aglikon dan membandingkan aktivitas antioksidan pada buncis, kecipir, kedelai kuning dengan antioksidan alami maupun antioksidan BHT (Butyl Hidroksi Toluena).
Metode untuk ekstraksi isoflavon dengan maserasi, identifikasi isoflavon dengan HPLC, uji aktivitas antioksidan dengan DPPH. Untuk mengetahui perbedaan antar perlakuan fermentasi dan penghitungan aktivitas antioksidan dianalisis dengan General Linear model-Univariete, untuk membandingkan tingkat aktivitas antioksidannya dianalisis dengan Compare Means-One Way Annova, menggunakan program SPSS version 15.
Hasil penelitian menunjukkan bahwa biji kedelai kuning, buncis dan kecipir mentah memiliki total isoflavon masing-masing sebanyak (0,18%), (0,15%), dan (0,21%). Pada kedelai kuning mentah tidak ditemukan isoflavon factor-2, tetapi buncis dan kecipir mentah ditemukan sebanyak (0,006%) dan (0,001%). Pada hasil fermentasi buncis, kecipir, kedelai kuning ditemukan jenis isoflavon factor-2, daidzein, glisitein, genistein. Hasil uji aktivitas antioksidan, berurutan dari tinggi ke rendah adalah tempe kecipir 0-hr (85,19%), tempe kedelai 3-hr (81,43%), BHT (81,15%), α-tokoferol (76,41%), vitamin C (75,62%), tempe buncis 0-hr (52,95%) dan β-karoten (43,25%). Aktivitas antioksidan pada tempe kecipir 0-hr signifikan lebih besar dari kedelai kuning fermentasi 3-hr maupun BHT, berarti kecipir berpotensi sebagai sumber antioksidan yang prospektif untuk digunakan sebagai pengganti kedelai kuning dan BHT. Aktifitas antioksidan pada buncis signifikan lebih rendah dari antioksidan BHT, α-tokoferol, vitamin C tetapi masih lebih tinggi dari antioksidan β-karoten.
Kata kunci : Buncis, Kecipir, Fermentasi, Isoflavon, Aktivitas antioksidan

vi
THE CHARACTERIZATION OF THE BIOACTIVE COMPOUNDS OF ISOFLAVONE AND THE TEST OF ANTIOXIDANT ACTIVITY OF EXTRACT
THE OF TEMPEH MADE OF POLE BEAN (Phaseolus vulgaris) AND WINGED BEAN (Psopocarpus tetragonolobus)
Sri Wahyuni, Sri Handajani, Artini Pangastuti Program Study of Biosains, Post Graduate Program, Sebelas Maret
University Surakarta
ABSTRACT
Pole bean and Winged bean belong to leguminoceae which have a potential as natural antioksidan resource. The aims of the research at finding out the content of the isoflavone aglikon compounds and comparing the antioxidant activity of the pole bean, winged bean, and yellow soybean with both the natural antioxidant and BHT antioxidant.
The methods used to extract the isoflavone compounds were maceration, identification of isoflavone by using HPLC, and test of antioxidant activity was done with DPPH. To know the difference between fermentation treatment and the measurement of antioksidan activity which was analized by General Linear Model-Univariete, to compare the level of antioxidant activity which was analized by Compare Means-One Way Annova using the program of SPSS version 15. The results of the research show that the whole soybean, pole bean, and winged bean seed had the total contents of the isoflavone compounds of 0.18%, 0.15%, and 0.21% respectively. The isoflavone of factor-2 was not found in the yellow soybean, but it was found in the pole bean (0.006%) and in the winged bean (0.001%). In the fermentation result of pole bean, winged bean and yellow soybean was I found kind of the isoflavone compounds of factor-2, daidzein, glisitein and genistein. The results of the antioxidant activity test show that the antioxidant activity from the highest to the lowest was owned by winged bean tempeh in the 0-day fermentation (85.19%), soybean tempeh in the 3-day fermentation (81.43%), BHT (81.15%), α-tocoferol (76.41%), vitamin C (75.62%), pole bean tempeh in the 0-day fermentation (52.95%), and ß-carotene (43.25%). Antioxidant activity in the winged bean tempeh in the 0-day fermentation was higher than and really different from the yellow soybean in the 3-day fermentation and BHT, meaning that the winged bean had the potential to be the antioxidant resource which was prospective to be used as a substitute for the soybean and BHT. The antioxidant activity of the pole bean was lower than that of BHT, α-tocoferol, and vitamin C, but was higher than that of ß-carotene. Keywords: Pole bean, winged bean, fermentation, isoflavone, antioxidant activity

vii
PERSEMBAHAN
Karya ilmiah ini kupersembahkan kepada Keluarga besarku, Suamiku, dan anakku yang tercinta
“Harta yang paling menguntungkan ialah sabar
Teman yang paling setia adalah amal
Pengawal pribadi yang paling waspada adalah diam
Bahasa yang paling manis adalah senyum
dan ibadah yang paling indah tentunya khusyuk”
(Harun Yahya)

viii
KATA PENGANTAR
Segala puji bagi Allah Tuhan semesta alam, tiada daya dan upaya kecuali
kekuatan dari Allah SWT. Berkat limpahan rahmat dan hidayah-Nya, penulis
dapat menyelesaikan tugas akhir berupa tesis dengan judul “Karakterisasi
Senyawa Bioaktif Isoflavon dan Uji Aktivitas Antioksidan dari Ekstrak Etanol
Tempe Berbahan Baku Buncis (Phaseolus vulgaris) dan Kecipir (Psopocarpus
tetragonolobus)”, merupakan bagian dari penelitian yang dilaksanakan oleh
Prof. Dr. Ir. Sri Handajani, MS. dan Dra. Retno Dwi Ariani, M.Si.
Di dalam tulisan ini, disajikan pokok-pokok bahasan yang meliputi
ekstraksi isoflavon dari biji buncis dan kecipir beserta produk fermentasinya,
identifikasi isoflavon aglikon (faktor-2, daidzein, glisitein, genistein) dan uji
aktivitas antioksidan.
Nilai penting penelitian ini adalah mengetahui kandungan isoflavon dan
aktivitas antioksidan pada buncis dan kecipir dengan variasi lama waktu
fermentasi (0, 1, 2, 3, 4 hari) serta mengetahui tingkat aktivitas antioksidatifnya
bila dibandingkan dengan ekstrak etanol dari kedelai dan produk fermentasinya
serta beberapa antioksidan alami (α-tokoferol, β-karoten, vitamin C) maupun
antioksidan sintetis BHT. Dari hasil penelitian ditemukan bahwa kecipir memiliki
tingkat aktivitas antioksidan yang lebih tinggi dan signifikan bila dibanding
dengan antioksidan sintetis BHT, sehingga kecipir dapat digunakan sebagai
sumber antioksidan alami yang bagus dan aman.
Pengembangan penelitian ini ke arah pembuatan senyawa antioksidan
untuk pengawet alami dari bahan yang murah dan melimpah serta dapat
dimanfaatkan sebagai food suplemen yang memiliki banyak khasiat pendukung
kesehatan.

ix
Penulis menyadari dengan sepenuh hati adanya kekurangan dan
keterbatasan, oleh karena itu penulis mengharapkan kritik dan saran yang
membangun agar tulisan ini bermanfaat bagi yang membutuhkan.
Surakarta, 30 Desember 2009
Penulis

x
UCAPAN TERIMA KASIH
Assalamu’alaikum Warahmatullahi wabarakatuh.
Segala puji bagi Allah Tuhan semesta alam, tiada daya dan upaya kecuali
kekuatan dari Allah SWT, atas berkah dan inayah-Nya maka penulis dapat
menyelesaikan tesis dengan judul “Karakterisasi Senyawa Bioaktif Isoflavon
dan Uji Aktivitas Antioksidan dari Ekstrak Tempe Berbahan Baku Buncis
(Phaseolus vulgaris) dan Kecipir (Psophocarpus tetragonolobus)”.
Pada kesempatan ini, penulis tidak lupa mengucapkan terima kasih kepada :
1. Direktur Program Pascasarjana Universitas Sebelas Maret Surakarta, Bapak
Prof. Suranto, MSc.,Ph.D sebagai Pembimbing Akademik yang telah
memberikan semua fasilitas dan dorongan semangat selama penulis
mengikuti pendidikan di Program Pascasarjana, Prodi Biosains UNS
Surakarta.
2. Prof. Ir. Sri Handajani, M.S.,Ph.D selaku Pembimbing I yang senantiasa
memberikan dorongan moril dan fasilitas serta pengarahan dan bimbingan
selama menyelesaikan tesis ini.
3. Dr. Artini Pangastuti, M.Si selaku Pembimbing II yang selalu memotivasi
dan memberi arahan serta bimbingan pada saat menulis dan menyelesaikan
tesis.
4. Dra. Retno Dwi Ariani, M.Si atas semua bimbingan dan pengarahannya
dalam pelaksanaan penelitian dan penulisan tesis.
5. Dr. Sugiyarto, M.Si, selaku dosen penguji yang dengan kesabarannya
memberikan bimbingan dan dorongan moril sehingga penulis dapat
menyelesaikan penulisan dan ujian tesis dengan lancar.

xi
6. Dr. Edwi Mahajoeno, M.Si atas semua masukan dan bimbingan serta
arahannya selama ujian komprehensif sehingga penulisan tesis ini menjadi
lebih berkualitas.
7. Ketua Laboratorium Pusat MIPA UNS beserta jajarannya, terima kasih atas
dukungan dan kerjasamanya sehingga penulis dapat melaksanakan
penelitian dengan lancar.
8. Ketua Laboratorium Kimia, FKIP UNS beserta jajarannya yang telah member
kan fasilitas sehingga penelitian ini dapat terlaksana.
9. Bapak Poyo, Laboran di Laboratorium Kimia organik Fakultas MIPA UGM
yang telah membantu, memberi fasilitas dan mendukung pelaksanaan
penelitian ini, semoga dibalas oleh Alloh SWT.
10. Saudari Wiji Hastuti, Yani, Yuli di FKIP UNS yang telah sangat membantu
terlaksananya penelitian ini.
11. Saudari Rini, Heni, Yurina yang merupakan teman seperjuangan dalam
suka dan duka, terima kasih atas kerjasama dan dukungannya selama tesis
ini diawali dan diakhiri sampai dinyatakan lulus.
12. Sadara M. Rosyid beserta seluruh jajaran staf administrasi Program
Pascasarjana UNS Surakarta yang dengan tulus telah membantu
memperlancar sarana administrasi penulis selama belajar di Program S2
Biosains.
13. Mbah Wagiyem, perajin dan pembuat tempe yang telah membimbing dan
membantu kami dalam proses pembuatan tempe sampai didapatkan tempe
yang memenuhi syarat untuk penelitian.
14. Anakku Tia dan Ida yang telah membantu kelancaran penulisa tesis ini.

xii
15. Suamiku, cinta dan kasih sayang serta motivasimu telah menyemangati aku
dalam menyelesaikan studi di Prodi Biosains, UNS Surakarta.
16. Semua pihak yang tidak dapat penulis sebutkan satu per satu, yang telah
memberikan bantuan baik moril maupun materiil sehingga penulis dapat
menyelesaikan studi S2.
Penulis tidak dapat membalas semua kebaikan dan segala bantuan yang
telah diberikan, semoga Alloh SWT mencatat amal kebaikan saudara/i dan
membalas semua dengan kasih sayang-Nya yang lebih besar lagi.
Wassalamu’alaikum Warahmatullahi wa barakatuh.
Surakarta, Desember 2009
Penulis
Sri Wahyuni

xiii
DAFTAR ISI
HALAMAN JUDUL .......................................................................................... i
HALAMAN PENGESAHAN PEMBIMBING...................................................... ii
HALAMAN PENGESAHAN PENGUJI ............................................................. iii
PERNYATAAN ORISINILITAS TESIS............................................................. iv
ABSTRAK ....................................................................................................... v
ABSTRACT ..................................................................................................... vi
HALAMAN PERSEMBAHAN........................................................................... vii
KATA PENGANTAR........................................................................................ viii
UCAPAN TERIMA KASIH ............................................................................... x
DAFTAR ISI…………………………………………………………………………..xiii
DAFTAR TABEL……………………………………………………………………..xvi
DAFTAR GAMBAR ........................................................................................ xvii
DAFTAR LAMPIRAN....................................................................................... xix
BAB I. PENDAHULUAN ................................................................................ 1
A. Latar Belakang .................................................................................... 1
B. Perumusan Masalah ........................................................................... 4
C. Tujuan Penelitian ................................................................................ 5
D. Manfaat Penelitian ............................................................................... 6
BAB II. TINJAUAN PUSTAKA ........................................................................ 7
A. Tinjauan Pustaka ................................................................................ 7
1. Leguminoceae .............................................................................. 7
a. Kecipir ( Psophocarpus tetragonolobus ) ................................. 8
b. Buncis ( Phaseolus vulgaris ) .................................................. 10
2. Tempe ........................................................................................... 12

xiv
a. Tempe Kedelai ........................................................................ 13
b. Tempe Non Kedelai ................................................................. 13
c. Proses Pembuatan Tempe ...................................................... 14
1. Perendaman ...................................................................... 14
2. Pencucian .......................................................................... 15
3. Pengupasan ...................................................................... 16
4. Pemasakan ....................................................................... 16
5. Penirisan dan Pendinginan ................................................ 16
6. Penambahan Inokulum ...................................................... 17
7. Pemeraman ...................................................................... 17
d. Proses Fermentasi .................................................................. 18
e. Rhizopus oligosporus .............................................................. 19
3. Isoflavon ....................................................................................... 20
3.1. Isoflavon Pada Kedelai ......................................................... 22
3.2. Isoflavon Pada Tempe Kedelai ............................................. 23
3.3. Metabolisme Isoflavon Pada Proses Pengolahan Kedelai
Menjadi Tempe ..................................................................... 26
3.4. Manfaat Senyawa Isoflavon Pada Tempe Kedelai ................ 28
4. Antioksidan ................................................................................... 29
4.1. Pengertian Antioksidan ......................................................... 29
4.2. Antioksidan Sintetik .............................................................. 32
4.3. Antioksidan Alami.................................................................. 32
4.3.a. Antioksidan Pada Kedelai ............................................ 33
4.3.b. Antioksidan Pada Tempe Kedelai ................................ 34
B. Kerangka Berpikir ................................................................................ 34
C. Hipotesis ............................................................................................. 35
BAB III. METODOLOGI PENELITIAN ............................................................. 36
A. Waktu dan Tempat Penelitian ............................................................. 36
B. Alat dan Bahan ................................................................................... 36
C. Prosedur Penelitian ............................................................................. 37
1. Pembuatan Tempe Kedelai ........................................................... 37
a. Pemilihan Biji Kedelai .............................................................. 37

xv
b. Perendaman ............................................................................ 38
c. Pengupasan ............................................................................ 38
d. Pengukusan ............................................................................ 38
e. Penambahan Inokulum ............................................................ 38
f. Pengemasan ........................................................................... 39
g. Fermentasi .............................................................................. 39
2. Pembuatan Tempe Berbahan Baku Biji Buncis dan Kecipir .......... 39
3. Mengekstraksi Isoflavon dengan Metode Maserasi ....................... 40
4. Identifikasi Isoflavon ...................................................................... 40
5. Uji Aktivitas Antioksidan ................................................................ 41
a. Pembuatan Larutan DPPH ........................................................ 41
b. Pembuatan Larutan Sampel ...................................................... 42
c. Pengukuran Kadar Antioksidan ................................................. 42
D. Teknik Analisa Data ............................................................................ 43
BAB IV. HASIL DAN PEMBAHASAN ............................................................. 44
A. Hasil Fermentasi Kedelai, Buncis dan Kecipir ..................................... 44
B. Hasil Ekstraksi Senyawa Isoflavon ...................................................... 46
C. Hasil Identifikasi Isoflavon dengan menggunakan Metode
HPLC .................................................................................................. 50
D. Hasil Uji Aktivitas Antioksidan ............................................................. 60
BAB V. PENUTUP ......................................................................................... 67
A. Kesimpulan ......................................................................................... 67
B. Saran ................................................................................................. 68
DAFTAR PUSTAKA ....................................................................................... 69

xvi
DAFTAR TABEL
Tabel 1. Komposisi Asam Amino Biji Kecipir ................................................... 10
Tabel 2. Struktur Daidzin, Genistin, Glisitin...................................................... 23
Tabel 3. Potensi Pemanfaatan Senyawa Isoflavonoid ..................................... 34
Tabel 4. Konsentrasi Isoflavon Pada Legume dan Produk Tempenya dengan
Variasi Lama Waktu Fermentasi (g/100 g sampel)............................. 53
Tabel 5. Kandungan Isoflavon Total (g) dan Aktivitas Antioksidan (%)
pada Kedelai Kuning, Buncis, Kecipir, dengan Variasi
Lama Fermentasi (hr) ......................................................................... 61
Tabel 6. Perbandingan Aktivitas Antioksidan Alami,Sintesis dan Legume (%). 65

xvii
DAFTAR GAMBAR
Gambar 1. Tanaman Kecipir ........................................................................... 9
Gambar 2. Polong Kecipir muda ..................................................................... 9
Gambar 3. Biji Kecipir ..................................................................................... 9
Gambar 4. Tanaman Buncis ........................................................................... 12
Gambar 5. Polong Buncis muda...................................................................... 12
Gambar 6. Biji Buncis...................................................................................... 12
Gambar 7. Rhizopus sp................................................................................... 20
Gambar 8. Hubungan antara metabolit primer dan metabolit sekunder........... 22
Gambar 9. Struktur dan sifat kimia Daidzein ................................................... 25
Gambar 10.Struktur dan sifat kimia Genistein.................................................. 25
Gambar 11. Struktur dan sifat kimia Glisitein................................................... 25
Gambar 12. Struktur dan sifat kimia Faktor- 2 ................................................. 26
Gambar 13.Reaksi Hidrolisis Glikosida Isoflavon menjadi Aglikon Isoflavon.... 27
Gambar 14. Massa Hasil Ekstraksi Biji Kedelai Kuning dan Produk
Fermentasinya ........................................................................... 48
Gambar 15. Massa Hasil Ekstraksi Biji Buncis dan Produk Fermentasinya ..... 48
Gambar 16. Massa Hasil Ekstraksi Biji Kecipir dan Produk Fermentasinya ..... 48
Gambar 17. Grafik Kandungan Isoflavon Faktor -2 pada Kedelai,
Buncis dan Kecipir ..................................................................... 57
Gambar 18. Grafik Kandungan Isoflavon Daidzein pada Kedelai,
Buncis dan Kecipir ..................................................................... 57
Gambar 19. Grafik Kandungan Isoflavon Glisitein pada Kedelai,
Buncis dan Kecipir ...................................................................... 57

xviii
Gambar 20. Grafik Kandungan Isoflavon Genistein pada Kedelai,
Buncis dan Kecipir ..................................................................... 57
Gambar 21. Grafik Perbandingan Aktivitas Antioksidan Alami, Sintesis dan
legume ........................................................................................ 64

xix
DAFTAR LAMPIRAN
Lampiran
1. Mekanisme Kerja Pembuatan Tempe Kedelai ..................................... 74
2. Mekanisme Pembuatan Tempe berbahan Buncis dan Kecipir ............ 75
3. Mekanisme Ekstraksi Isoflavon dengan Metode Maserasi .................. 76
4. Identifikasi Isoflavon dengan Metode HPLC......................................... 77
5. a. Pembuatan Larutan DPPH............................................................... 78
b. Pembuatan Larutan Sampel dan Uji Aktivitas Antioksidannya.......... 79
6. Hasil Pengukuran dan Perhitungan besarnya Aktivitas Antioksidan
dari kedelai, buncis,, kecipir, antioksidan alami (α-tokoferol, β-karoten,
serta vitamin C) dan antioksidan sintetis BHT...................................... 80
7. Hasil Pengamatan Biji Kedelai Kuning dan produk Tempenya............. 81
8. Hasil Pengamatan Biji Buncis dan poduk Tempenya ........................... 82
9. Hasil Pengamatan Biji Kecipir dan produk Tempenya.......................... 83
10. Hasil Ekstraksi Senyawa Isoflavon dari Kedelai, Buncis, Kecipir beserta
produk Tempenya................................................................................ 84
11. Analisis Statistik SPSS dua faktor........................................................ 85
12. Aktivitas Antioksidan (%) pada Kedelai, Buncis, dan Kecipir dengan
Variasi Lama Waktu Fermentasi .......................................................... 88
13. Perbandingan Aktivitas Antioksidan alami, Sintetik, dan Legume........ 88
14. Surat Pernyataan dan Publikasi Tesis.................................................. 89
15. Biodata Mahasiswa.............................................................................. 90

xx
BAB I
PENDAHULUAN
A. Latar Belakang Penelitian
Tempe kedelai dikonsumsi oleh berbagai lapisan masyarakat, berbagai
umur dan berbagai status ekonomi serta sering dijumpai sebagai makanan dalam
menu sehari-hari, baik sebagai lauk pauk maupun sebagai makanan camilan.
Bahan baku untuk pembuatan tempe kedelai masih sangat tergantung
dari pasokan impor, sehingga harganya sering fluktuatif dan menjadi mahal. Hasil
penelitian Handajani et al., (1991), menyatakan bahwa Indonesia mempunyai
banyak jenis legume yang beberapa diantaranya belum dimanfaatkan secara
optimal. Salah satu jenis legume yang cocok dibudidayakan di Indonesia dan
dapat berfungsi sebagai bahan pangan tetapi produk olahannya masih jarang
dikonsumsi yaitu buncis (Phaseolus vulgaris) dan kecipir (Psophocarpus
tetragonolobus). Dalam rangka pengembangan senyawa antioksidan alami
khususnya isoflavon, maka perlu dilakukan penelitian tentang optimasi produksi
senyawa antioksidan dari buncis dan kecipir dengan produk tempenya serta
karakterisasi kandungan isoflavonnya. Buncis dan kecipir sebagai alternatif
obyek penelitian sumber isoflavon, karena isoflavon merupakan metabolit
sekunder yang banyak disintesis oleh tanaman, terutama dari kelompok
leguminoceae (Anderson, 1997). Oleh karena itu, buncis (P. vulgaris) dan kecipir
(P. tetragonolobus) yang merupakan dua spesies dari familia leguminoceae,
dimungkinkan juga mengandung senyawa bioaktif isoflavon seperti yang
dijumpai pada kedelai.
Selama ini tempe kedelai yang dikonsumsi oleh masyarakat adalah
tempe hasil fermentasi kedelai selama 36 – 48 jam. Lama waktu fermentasi

xxi
tersebut merupakan lama waktu fermentasi kedelai untuk menghasilkan tempe
optimum dari sisi cita rasa untuk dikonsumsi, tetapi lama waktu fermentasi
optimum untuk menghasilkan ekstrak antioksidan khususnya senyawa isoflavon
yang maksimum, belum diketahui. Dalam rangka pengembangan produk tempe
tersier, misalnya produk isoflavon dari tempe untuk pengawet alami, maka perlu
dilakukan penelitian tentang optimasi produksi isoflavon. Dalam penelitian ini
akan difokuskan pada optimasi produksi senyawa antioksidan khususnya
senyawa bioaktif isoflavon dengan variasi lama waktu fermentasi pada biji
kedelai dan produk tempenya, maupun pada biji buncis dan biji kecipir beserta
produk tempenya.
Penelitian ini menggunakan sampel tempe berbahan baku buncis dan
kecipir, karena buncis (P.vulgaris) dan kecipir (P. tetragonolobus) tergolong
spesies dari famili leguminoceae seperti halnya kedelai, dan sudah diketahui
dengan positif bahwa tempe yang berbahan baku kedelai ternyata mengandung
senyawa bioaktif isoflavon aglikon yaitu faktor-2, daidzein, glisitein dan genistein
( Pawiroharsono, 1995).
Menurut hasil penelitian (Gyorgy et al., 1964 dalam Ariani 2009)
dilaporkan bahwa isoflavon faktor-2 hanya dijumpai pada kedelai rendam yang
difermentasi oleh Rhizopus sp. Isoflavon yang ditemukan dalam biji dorman
adalah daidzin, glisitin, dan genistin yang tergolong bentuk isoflavon glikosida.
Senyawa faktor-2 tidak dijumpai pada kedelai yang tidak difermentasi
oleh kapang (Laksani, 2003), oleh karena itu agar diperoleh senyawa faktor-2
harus membuat biji-biji legum menjadi tempe, melalui proses fermentasi oleh
kapang jenis Rhizopus sp. Faktor-2 mempunyai aktivitas antioksidan dan
antihemolisis yang lebih baik dari daidzein dan genistein. Selain itu, telah

xxii
ditemukan bahwa senyawa isoflavon faktor-2 lebih aktif 10 kali lipat dari senyawa
karboksi kroman (Pawiroharsono, 1995).
Studi epidemiologi di Jepang menemukan bahwa konsumsi isoflavon
bermanfaat mengurangi konsentrasi kolesterol serum pada hiperkolesterolemia,
sehingga dapat menurunkan insiden kanker payudara (Aldercreutsz, 1998).
Isoflavon juga memiliki khasiat farmakologi, sifat fisiologis aktif dari senyawa
isoflavon antara lain antifungi, antioksidan, antihemolisis dan antikanker.
Isoflavon adalah salah satu jenis fitoestrogen, merupakan suatu zat yang
memiliki khasiat mirip estrogen dan berasal dari tumbuhan. Pentingnya manfaat
isoflavon bagi kesehatan manusia, mendorong peneliti untuk melakukan studi
kandungan isoflavon pada tempe yang berbahan baku buncis (P. vulgaris) dan
kecipir (P. tetragonolobus) yang banyak terdapat dilingkungan sekitar kita. Salah
satu aktivitas fisiologis yang menonjol dari isoflavon daidzein, genistein, glisitein
dan faktor-2 adalah aktivitas antioksidan.
Pada umumnya, antioksidan yang digunakan sebagai pengawet pada
bahan makanan adalah antioksidan sintetik seperti Butylated Hydroxyanisole
(BHA), Butylated Hydroxytoluene (BHT), Propyl Gallat (PG) dan Etylene Diamine
Tetra Acetic Acid (EDTA). Pemanfaatan zat antioksidatif sintetik dapat
menimbulkan gangguan kesehatan bagi konsumen antara lain gangguan fungsi
hati, paru, mukosa usus dan keracunan (Chang et al.,1977 dalam Suryo dan
Tohari, 1995). Untuk itu perlu dicari alternatif lain untuk mengatasi permasalahan
tersebut. Salah satu cara adalah dengan mengganti pemanfaatan antioksidan
sintetik dengan antioksidan alami. Mengingat adanya kandungan isoflavon
dalam kedelai yang dapat berfungsi sebagai antioksidan, maka tempe berbahan
baku dari leguminoceae dapat direferensikan sebagai bahan baku sumber

xxiii
antioksidan alami. Dengan demikian isoflavon dari tempe berbahan baku legume
selain berkhasiat sebagai antioksidan juga mempunyai khasiat farmakologis
seperti yang sudah diuraikan diatas.
Untuk memperoleh zat antioksidan alami, dapat dilakukan dengan cara
ekstraksi menggunakan pelarut organik seperti, heksana, benzena, etil eter,
kloroform, etanol atau metanol. Metanol 90 % merupakan pelarut optimum untuk
mengekstrak isoflavon dari kedelai, namun penggunaanya untuk skala komersial
masih perlu dikaji lebih lanjut karena bersifat toksik. Penelitian dengan
menggunakan pelarut etanol 70% untuk ekstraksi diharapkan dapat mengganti
metanol untuk menghasilkan ekstrak antioksidan alami secara komersial, karena
kepolaran etanol mendekati metanol dan relatif tidak beracun (Kudou et al.,
1991, Susanto et al.,1998 dalam Ariani, 2009). Untuk selanjutnya pada
penelitian ini juga akan difokuskan pada ekstraksi dengan menggunakan pelarut
etanol 70%.
Berdasarkan uraian diatas, maka dilakukan penelitian tentang
karakterisasi senyawa isoflavon dan uji aktivitas antioksidan dari ekstrak tempe
berbahan baku buncis (Phaseolus vulgaris) dan kecipir (Psophocarpus
tetragonolobus).
B. Perumusan Masalah
Dari uraian latar belakang di atas, permasalahan dapat dirumuskan
sebagai berikut :
1. Isoflavon jenis apa sajakah yang terkandung dalam biji buncis dan biji
kecipir beserta produk fermentasinya yang dibuat dengan lama waktu
fermentasi 0,1, 2, 3, dan 4 hari.

xxiv
2. Berapa lama waktu fermentasi yang optimum untuk menghasilkan ekstrak
etanol tempe buncis dan kecipir, yang mengandung isoflavon aglikon
(Faktor-2, daidzein, glisitein, genistein) dengan aktivitas antioksidan
maksimum ?
3. Bagaimana potensi biji buncis dan kecipir serta produk fermentasinya
sebagai sumber antioksidan alami khususnya isoflavon, bila dibandingkan
dengan ekstrak etanol dari kedelai dan produk fermentasinya serta
beberapa antioksidan alami ( α-tokoferol, β-karoten, vitamin C ) maupun
antioksidan sintetis (Butyl Hidroksi Toluena / BHT) dengan metode DPPH.
C. Tujuan Penelitian
Adapun tujuan dari penelitian yang dilakukan adalah :
1. Mengetahui jenis Isoflavon aglikon yang terkandung dalam biji buncis dan
kecipir beserta produk fermentasinya berdasarkan variasi lama waktu
fermentasi (0, 1, 2, 3, dan 4 hari ).
2. Mengetahui lama waktu fermentasi yang optimum untuk menghasilkan
ekstrak etanol tempe buncis dan kecipir yang mengandung isoflavon aglikon,
dengan aktivitas antioksidan yang maksimum.
3. Mengetahui potensi biji buncis dan kecipir serta produk fermentasinya
sebagai sumber antioksidan alami khususnya isoflavon bila dibandingkan
dengan ekstrak etanol dari kedelai dan produk fermentasinya serta beberapa
antioksidan alami (α-tokoferol, β-karoten, vitamin C) maupun antioksidan
sintetis Butylat Hidroksitoluena (BHT) dengan metode DPPH.

xxv
D. Manfaat Penelitian
Adapun manfaat dari penelitian yang dilakukan adalah :
a. Dapat memberikan informasi pada masyarakat mengenai kandungan
isoflavon faktor-2, daidzein, glisitein, genistein, serta aktivitas antioksidatif
dari biji buncis dan kecipir beserta produk fermentasinya
b. Mengetahui sejauh mana manfaat biji buncis dan kecipir beserta produk
fermentasinya sebagai sumber antioksidan alami khususnya senyawa
bioaktif isoflavon.
c. Diharapkan dapat dijadikan referensi bagi penelitian selanjutnya, untuk
mencari dan menentukan macam senyawa yang berguna sebagai
antioksidan (selain senyawa isoflavon), yang terkandung di dalam biji
buncis dan biji kecipir.

xxvi
BAB II
TINJAUAN PUSTAKA
A. Tinjauan Pustaka
A.1. Leguminoceae
Legume (kacang polong) adalah jenis tanaman dikotil dan merupakan
tanaman setahun berumur pendek yang memiliki bunga kupu-kupu, tumbuh
merambat sebagai tanaman semak. Di lingkungan kita, legume sebagian besar
dibudidayakan untuk dimanfaatkan sebagai sayuran (misalnya buncis dan
kecipir) atau untuk diambil bijinya (misalnya kedelai, kacang hijau, kacang tanah).
Koro merupakan salah satu jenis legum yang dapat tumbuh di tanah yang kurang
subur dan kering. Selain untuk dimanfaatkan bijinya, tujuan penanaman koro
adalah sebagai tanaman pelindung dan pupuk hijau (Kanetro dan Hastuti, 2006).
Koro-koroan merupakan salah satu jenis legum lokal yang memiliki beragam
varietas dan biasa digunakan sebagai bahan baku pengganti kedelai dalam
pembuatan tempe. Kandungan gizi koro tidak kalah dengan kedelai yaitu
karbohidrat dan protein yang cukup tinggi serta kandungan lemak yang rendah.
Dalam Kanetro dan Hastuti (2006) disebutkan bahwa senyawa antigizi yang
sering terdapat pada legum antara lain enzim lipoksigenase, tripsin inhibitor,
asam fitat, oligosakarida, senyawa glikosida dan sianida. Pengolahan koro pada
umunya diawali dengan perendaman untuk menghilangkan sianida, karena kadar
sianida pada koro reltif tinggi. Setelah perendaman, biasanya diikuti dengan
pemasakan dan pengupasan kulit, karena kandungan karbohidarat yang tinggi
menyebabkan koro memiliki tekstur yang keras sehingga pemasakan dilakukan
agar teksturnya menjadi lunak (Handajani dan Atmaka, 1993).

xxvii
Selain bersifat sebagai senyawa antinutrisi, fitat memiliki peranan dalam
kesehatan yang dianggap positif yaitu sebagai antioksidan yang dapat
menangkal adanya radikal bebas maupun senyawa non radikal yang dapat
menimbulkan oksidasi pada biomolekuler seperti protein, karbohidrat dan lipida
(Anonim, 2007).
Anggota famili leguminoceae (polongan) diantaranya adalah :
a. Kecipir atau Winged Beans (Psophocarpus tetragonolobus )
Kedudukan tanaman kecipir dalam klasifikasi ilmiah ( taksonomi tumbuhan)
sebagai berikut:
Kingdom : Plantae
Divisio : Magnoliophyta
Classis : Magnoliopsida
Ordo : Fabales
Famili : Fabaceae
Sub famili : Faboideae
Genus : Psophocarpus
Species : Psophocarpus tetragonolobus
Sumber : (Heyne, 1987 ).
Tanaman kecipir tumbuh subur didaerah tropis basah dengan curah hujan
lebih besar dari 1600 m di atas permukaan laut. Tanah-tanah yang baik untuk
kecipir adalah tanah dengan bahan organik yang rendah, berpasir atau lempung.
Tanaman kecipir tahan terhadap kekeringan. Kecipir disebut juga kacang botol
(Malaysia), sirahu avarai (Thailand), dragon bean (Vietnam), jaat (Sunda) dan
sigarillas (Tagalog) termasuk tanaman setahun, mempunyai keunggulan
dibandingkan dengan kedelai, karena seluruh bagian tanaman dapat

xxviii
dimanfaatkan. Daun, buah dan umbi kecipir memiliki kadar protein yang tinggi,
bahkan rata-rata melampaui kadar protein biji-bijian atau umbi-umbian lainnya
sepert beras, ubi jalar, ketela pohon dan kentang.
Dibawah ini disajikan gambar tanaman, buah, dan biji kecipir yang biasa
dijumpai dalam kehidupan sehari-hari.
Gambar 1. Tanaman Kecipir
Gambar 2. Polong Kecipir muda
Gambar 3. Biji-biji Kecipir
Buah kecipir berbentuk persegi empat siku yang masing-masing segi
bersayap dan bergelombang. Panjang polong antara 6-20 cm dan bijinya putih,
kuning hitam, sawo matang atau coklat muda. Berat bijinya rata-rata 30-64 gr
tiap 100 butir (Handajani, 1992 ).
Biji tanaman kecipir mengandung protein, lemak yang cukup tinggi, setara
dengan kedelai (Salunkhe and Kadam, 1990). Kadar mineral dalam bentuk zat
kapur, fosfor dan zat besinya juga lebih unggul. Selain itu, biji kecipir juga
mengandung sejumlah asam amino esensial dan asam lemak esensial, setara
dengan kedelai atau kacang tanah (Handayani, 1993 ). Komposisi asam amino

xxix
dari biji kecipir hampir sama dengan biji kedelai. Secara lengkap kandungan
asam amino yang menyusun protein dalam kecipir dapat dilihat pada tabel
berikut :
Tabel 1. Komposisi asam amino biji kecipir (%) Jenis Asam
Amino Kandungan ( % ) Jenis Asam Amino Kandungan ( % )
Alanin
Arginin
Asam aspartat
Asam glutamat
Glisin
Histidin
Prolin
Serin
Tryptophan
296
283
751
1080
268
176
449
360
104
Tirosin
Isoleusin
Leusin
Lisin
Methionin
Fenil alanin
Threonin
Valin
Sistein
281
263
506
488
58
321
294
265
54
Sumber : (Haryoto, 1996).
Biji kecipir belum dimanfaatkan secara optimal, karena memiliki bau langu
yang kuat dan kulit biji yang keras (Kanetro, 2001). Kulit biji yang keras
menyebabkan daya serap air sangat rendah sehingga membutuhkan waktu yang
lama dalam pengolahannya.
b. Buncis ( Phaseolus vulgaris )
Buncis adalah sayuran dari genus Phaseolus yang paling dikenal.
Umumnya sistem perakaran berbagai jenis buncis tidak ekstensif, percabangan
lateralnya dangkal. Daun buncis beranak daun tiga dan menyirip. Bunga
berukuran besar, berwarna putih, merah jambu atau ungu. Bunganya sempurna
memiliki 10 benangsari, 9 diantaranya menyatu berbentuk tabung yang

xxx
melingkupi batang, buah panjang. Bunga menyerbuk sendiri dan umumnya
jarang terjadi persilangan .
Kedudukan tanaman buncis dalam klasifikasi ilmiah (taksonomi tumbuhan)
sebagai berikut:
Kingdom : Plantae
Divisio : Magnoliophyta
Classis : Magnoliopsida
Ordo : Fabales
Famili : Fabaceae
Sub famili : Faboideae
Genus : Phaseolus
Species : Phaseolus vulgaris
Sumber: ( Heyne, 1987 )
Panjang polong berkisar 8 hingga 20 cm atau lebih, dengan lebar 1 cm.
Ketika biji telah matang sempurna, polong akan membelah terbuka. Sebagian
polong buncis tidak berbulu, sedikit diantaranya berbulu halus. Buncis tidak
memiliki kelopak daun yang persisten sebagaimana yang dimiliki kapri (Heyne,
1987 ).
Sebagian besar kultivar buncis berbiji 3 hingga 5. Tipe buncis bijian atau
buncis segar cenderung berbiji banyak. Panjang biji berkisar 5 mm hingga 20 mm
dan bobot biji tunggal kultivar berkisar dari 0,15 hingga lebih dari 0,80 gram. Biji
dapat berbentuk bundar, ovoid, lonjong dan seperti ginjal. Warna kulit biji sangat
khas menurut kultivarnya, dan dapat muncul dalam berbagai macam warna dan
kombinasi warna (Rubatzky and Yamaguchi, 1998).

xxxi
Dibawah ini disajikan gambar tanaman, buah polong muda, dan biji buncis yang
sering dijumpai dilingkungan sekitar kita.
Gambar 4. Tanaman Buncis
Gambar 5. Polong Buncis
Gambar 6. Biji Buncis
A.2. Tempe
Tempe secara luas dikenal sebagai makanan khas Indonesia, dikonsumsi
oleh semua lapisan masyarakat dan sangat digemari oleh masyarakat Jawa.
Ada berbagai macam tempe di Indonesia seperti misalnya tempe gembus dibuat
dari ampas tahu, tempe lamtoro dibuat dari biji lamtoro, tempe benguk dibuat dari
biji koro benguk, tempe bongkrek dibuat dari ampas kelapa, tempe gude dibuat

xxxii
dari kacang gude dan tempe kedelai dibuat dari kedelai. Dari beberapa jenis
tempe tersebut yang paling banyak digemari masyarakat adalah tempe kedelai.
Tempe dibuat dengan proses fermentasi kedelai dengan kapang jenis
Rhizopus sp. Tempe bermutu tinggi bila kacang terlekat dengan jalinan miselium
putih. Jika proses fermentasi dibiarkan terlalu lama, spora hitam terbentuk di
permukaan. Spora tersebut tidak berbahaya namun mempengaruhi kenampakan
dan penerimaan konsumen.
2.a. Tempe Kedelai
Tempe kedelai tergolong sebagai makanan hasil fermentasi oleh jamur
jenis Rhizopus sp. dan merupakan produk fermentasi yang amat dikenal oleh
masyarakat. Tempe kedelai berpotensi untuk digunakan melawan radikal bebas,
sehingga dapat menghambat proses penuaan dan mencegah terjadinya penyakit
degeneratif (aterosklerosis, jantung koroner, diabetes melitus, kanker). Selain itu
tempe juga mengandung zat antibakteri penyebab diare, penurun kolesterol
darah, hipertensi, dan penyakit degeneratif lainnya (Astuti , 1995 ).
Komposisi gizi tempe baik kadar protein, lemak, dan karbohidratnya tidak
banyak berubah dibandingkan dengan kedelai. Namun, karena adanya enzim
pencerna yang dihasilkan oleh kapang tempe, maka protein, lemak, dan
karbohidrat pada tempe menjadi lebih mudah dicerna di dalam tubuh
dibandingkan yang terdapat dalam kedelai.
2.b. Tempe Non Kedelai
Selain tempe berbahan dasar kacang kedelai, terdapat pula berbagai
jenis makanan berbahan bukan kedelai yang juga disebut tempe. Terdapat dua
golongan besar tempe menurut bahan dasarnya, yaitu tempe berbahan dasar
legume dan tempe berbahan dasar non-legume .

xxxiii
Dalam pembuatan tempe buncis dan kecipir, biji direndam dalam air
bersih dan dilakukan penggantian air rendaman setiap 8 jam sekali (9 kali diganti
airnya selama 3 x 24 jam) perendaman, agar kandungan HCN dapat dihilangkan.
Setelah selesai perendaman, kemudian kulit biji dikupas, dan direbus dengan
air selama satu jam, kemudian ditiriskan dan didinginkan. Proses selanjutnya,
termasuk jenis ragi yang digunakan relatif sama dengan pembuatan tempe
kedelai. Tempe berbahan dasar non-legume mencakup tempe bongkrek dari
bungkil kacang atau ampas kelapa, yang terkenal didaerah Banyumas, dan
tempe jamur merang terbuat dari jamur merang (Astawan, 2003).
2.c. Proses Pembuatan Tempe
1. Perendaman
Perendaman merupakan tahap awal dan penting dalam pembuatan
tempe secara tradisional. Menurut Kasmidjo (1990), perendaman bertujuan
memberikan kesempatan kepada biji untuk menyerap air (hidrasi) sehingga biji
menjadi lebih lunak dan dapat menurunkan pH, karena terjadi pengasaman yang
disebabkan oleh proses fermentasi oleh bakteri dan mikroorganisme lain yang
berada dalam air rendaman.
Pada kacang-kacangan, secara umum dilakukan perendaman sebelum
proses pengolahan. Perendaman ini berfungsi untuk melunakkan biji,
mengurangi bau langu dari biji yang diolah serta mereduksi lendir dan kotoran
yang menempel pada keping biji (Atikoh dan Supriyanti, 1997).
Pengasaman terjadi karena pertumbuhan bakteri penghasil asam laktat
diantaranya Lactobacillus sp. Pertumbuhan bakteri yang baik ditandai oleh bau
kecut dan busa pada permukaan rendaman kedelai. Bila pertumbuhan bakteri
kurang, sehingga pH yang diperlukan tidak tercapai, rendaman kedelai perlu

xxxiv
ditambah bahan pengasaman. Asam laktat merupakan pilihan pertama sebagai
bahan pengasaman. Jika tidak mungkin memperoleh asam laktat, asam cuka
dapat digunakan (Suwahyono, 1989 )
Penggunaan bahan pengasaman menjadi keharusan pada cara cepat
pembuatan tempe, yang cocok sebagai cara industri. Pada cara cepat
perendaman cukup 2-3 jam. Pada cara tradisional perendaman berlangsung
20-30 jam karena bakteri perlu waktu panjang untuk tumbuh dan menghasilkan
asam.
Menurut Kasmidjo (1990), bakteri pembentuk asam yang banyak tumbuh
pada proses perendaman kedelai sebagian besar adalah golongan termobakteri
yang mempunyai suhu optimum 40 – 45 C, antara lain Bacillus sp, Lactobacillus
sp, Streptococcus thermopilus dan Enterobacteriaceae. Sehingga dengan lebih
banyaknya bakteri pembentuk asam yang tumbuh pada suhu tersebut, maka
semakin banyak asam laktat yang terbentuk. Besarnya pH berkorelasi dengan
kandungan asam laktat, semakin tinggi kandungan asam laktat maka semakin
rendah nilai pH nya.
2. Pencucian
Pencucian ini dimaksudkan untuk menghilangkan kotoran dan
mengurangi mikroorganisme lain yang tumbuh selama perendaman, juga untuk
membuang kelebihan asam dan lendir yang terproduksi. Pencucian harus
dilakukan sampai keping biji kedelai tidak licin lagi oleh lendir dan kedelai tidak
terlalu asam. Lendir dan bakteri apabila tidak tercuci bersih akan mengganggu
pertumbuhan kapang Rhizopus sp. dan dapat menyebabkan kegagalan produksi
(Atikoh dan Supriyanti, 1997 )

xxxv
3. Pengupasan
Setelah proses hidrasi, dilakukan pengupasan yaitu melepaskan kulit ari
dari keping biji kedelai. Proses ini harus dilakukan agar terjadi penetrasi asam
dan miselium kapang ke dalam keping biji. Miselium kapang tidak dapat
menembus lapisan kulit ari kedelai karena zat tanduk (kitin) yang terkandung
dalam kulit, sehingga bila kulit tidak terlepas dari keping kedelai, produk tempe
yang dihasilkan tidak atau kurang kompak.
4. Pemasakan
Proses pemasakan dapat dilakukan dengan cara merebus atau
mengukus. Proses pemasakan dilakukan selain untuk melunakkan kedelai, juga
sebagai proses sterilisasi untuk mematikan bakteri-bakteri yang tumbuh selama
proses perendaman. Keuntungan melalui proses ini adalah tempe akan lebih
tahan lama, tidak berasa asam dan tidak pernah berlendir (Susanto et al., 1998).
Menurut Suwaryono dan Ismeini (1988) proses pengukusan dilakukan
setelah air mendidih. Pada pengukusan, kerusakan biji terjadi lebih lambat
karena biji tidak berinteraksi secara langsung dengan air panas, namun melalui
uap air panas sehingga pada proses ini suhu yang digunakan dibawah atau
sama dengan 100 o C.
5. Penirisan dan Pendinginan
Proses penirisan dilakukan, untuk menghilangkan air yang menempel
pada biji kedelai. Menurut Kasmidjo (1990), tidak adanya air pada biji kedelai
akan menghambat pertumbuhan bakteri dan mempercepat pertumbuhan
kapang. Sedangkan proses pendinginan dilakukan setelah pemasakan, untuk
menghilangkan air yang menempel pada keping biji dan juga untuk mengkondisi
suhu agar sesuai untuk pertumbuhan kapang.

xxxvi
6. Penambahan Inokulum
Secara tradisisonal inokulum tempe disebut ragi tempe atau bibit tempe.
Ragi adalah sumber mikrobia yang digunakan dalam proses fermentasi. Bibit
tempe sebenarnya spora kapang. Jumlah spora pada satu unit ukuran inokulum,
gram atau sendok, dapat diubah dengan memperbanyak bahan pembawa
berupa campuran spora kapang dengan tepung beras, tepung singkong atau
tepung terigu. Jumlah bahan pembawa yang lebih baik memudahkan
penyebaran spora yang merata, lebih menjamin pertumbuhan yang baik.
Inokulum yang dibuat dalam keadaan terbuka ke lingkungan
memungkinkan untuk dimasuki mikrooraganisme lain, seperti bakteri, dan
khamir. Mikroorganisme tersebut dapat menimbulkan perubahan-perubahan
disaat proses fermentasi sehingga dihasilkan zat-zat berbeda (Judoamidjojo
et al.,1992).
Perubahan yang terjadi selama proses fermentasi ditimbulkan oleh enzim-enzim
yang dihasilkan kapang yaitu pengurai protein, karbohidrat, dan asam lemak.
Inokulum merupakan bahan yang paling penting pada pembuatan tempe, karena
sebagai pembawa kapang yang akan melakukan proses fermentasi. Setiap jenis
kapang mempunyai kemampuan yang berbeda dalam hal pertumbuhan,
pembentukan enzim dan senyawa-senyawa yang bermanfaat bagi kesehatan.
7. Pemeraman
Keping-keping biji kedelai yang sudah dicampur dengan inokulum
diperam dalam kemasan pada suhu kamar (29ºC-31ºC). Kemasan diperlukan
karena kapang hanya memerlukan sedikit oksigen untuk tumbuh. Kemasan juga
berguna untuk mengkondisikan suhu agar selalu sesuai untuk pertumbuhan
kapang, bahan pengemas menggunakan daun pisang.

xxxvii
d. Proses Fermentasi
Ariani dan Hastuti (2009) melaporkan, selama fermentasi terjadi
perubahan bau dan rasa karena adanya aktivitas enzim. Selama fermentasi,
miselia jamur yang berwarna putih akan menyelubungi permukaan tempe, dan
mengeluarkan enzim protease, lipase dan amilase ke lingkungan sekitarnya.
Enzim-enzim tersebut dapat memecah komponen dalam bahan yaitu protein,
lemak dan karbohidrat menjadi bahan yang lebih sederhana. Aktivitas
mikroorganisme didalam proses pembuatan tempe secara tradisional, dapat
dibedakan menjadi 2 tahapan proses yaitu :
1) Proses Fermentasi Awal (Fermentasi I)
Proses perendaman selama 12 jam dilakukan terhadap biji-biji yang
telah disortir dan kemudian dikupas kulitnya, kemudian direndam kembali selama
12 jam, pada suhu kamar (27-300C), dengan menggunakan air tanah (sumur).
Pada proses ini terjadi proses fermentasi awal oleh bakteri pembentuk asam-
asam organik. Tujuan utama proses ini adalah untuk pengasaman kedelai. Untuk
maksud pengasaman ini, maka pada proses perendaman dilakukan inokulasi
bakteri pembentuk asam yaitu dengan menambahkan air kedalam rendaman dari
proses perendaman sebelumnya, sehingga tahapan ini disebut proses
fermentasi awal (fermentasi I). Pada biji buncis dan kecipir proses
perendamannya lebih lama yaitu 3 x24 jam, karena kulit biji kecipir itu keras
sedangkan buncis sangat berwarna biru air rendamannya sehingga untuk
membeningkan air rendaman diperlukan waktu yang lebih lama.
2) Proses Fermentasi Utama (Fermentasi II)
Mikroorganisme yang berperan utama didalam pembuatan tempe adalah
kapang Rhizopus sp. Aktivitas fisiologis kapang pada proses fermentasi tempe

xxxviii
dimulai sejak diinokulasinya inokulum (ragi tempe) pada bahan yang telah siap
difermentasikan yaitu biji kedelai, biji buncis dan kecipir masak yang telah dikuliti
dan ditiriskan. Spora kapang tersebut mulai tumbuh berkecambah dengan
membentuk benang-benang hifa yang makin memanjang membalut dan
menembus biji kedelai. Apabila benang-benang tersebut telah sedemikian padat,
maka terbentuklah tempe yang kompak, putih dan dengan aroma khas tempe.
Secara keseluruhan tahapan ini disebut sebagai proses fermentasi II, dan
berlangsung disaat pemeraman.
e. Rhizopus sp. / kapang tempe
Mikroorganisme yang berperan utama didalam pembuatan tempe, salah
satunya adalah kapang Rhizopus sp. Sebagai contoh inokulum dalam bentuk
tepung dan diproduksi skala pabrik oleh Puslitbang Kimia Terapan-LIPI Bandung,
menggunakan Rhizopus oligosporus (Lindajati, 1985). Didalam klasifikasi,
kapang ini digolongkan ke dalam genus Rhizopus, familia Mucoraceae, ordo
Mucorales, subklas Zygomicotina, dan klasis zygomycetes (Hesseltine, 1985).
Kapang yang tergolong dalam genus Rhizopus dicirikan berupa sel vegetatif
yang berupa benang dan disebut hifa/misellium yang membentuk stolon-stolon
(semacam ruas/buku) yang dilengkapi dengan rhizoid (mirip akar) yang tumbuh
bercabang-cabang masuk kedalam subtrat. Pada tempat tumbuhnya rhizoid,
terdapat sporangiospora yang tumbuh mengarah keudara (berlawanan arah
dengan rhizoid), dan dari tempat inilah terbentuk spora. Dibawah ini disajikan
gambar Rhizopus sp.

xxxix
Gambar 7. Rhizopus sp.
Rhizopus sp. sebagai kapang pemeran utama yang telah terbukti dapat
memfermentasikan kedelai dan membentuk tempe secara sempurna. Waktu
yang dibutuhkan sampai terbentuk tempe secara sempurna memerlukan waktu
24-36 jam.
Selama proses fermentasi berlangsung, kedelai berubah menjadi tempe
dan perubahan tersebut pada dasarnya dapat dibedakan sebagai perubahan
secara fisik dan secara kimia. Perubahan sifat fisik tempe dibandingkan dengan
kedelai antara lain, bertekstur kompak, warna putih dengan aroma khas tempe.
Perubahan secara kimia ditandai dengan terjadinya hidrolisa senyawa-senyawa
komplek (protein, karbohidrat, lemak, ikatan glikosida) menjadi senyawa yang
lebih sederhana dan mudah dicerna (Astuti, 1995).
A.3. Isoflavon
Flavonoid merupakan kelompok fenol dengan sebuah cincin aromatik
dan satu atau lebih gugus hidroksil yang tersebar di alam. Senyawa fenol
cenderung larut dalam air karena paling sering dijumpai bergabung dengan gula

xl
berupa glikosida dan biasanya terdapat dalam rongga sel. Flavonoid merupakan
kelompok molekul organik yang tersebar di hampir seluruh bagian tanaman.
Kurang lebih dua ribu jenis golongan flavonoid tersebar di alam (Goldberg,1996).
Penyebaran jenis flavonoid terbanyak dijumpai pada angiospermeae
(tumbuhan berbiji tertutup), dan flavonoid mempunyai potensi sebagai
antioksidan (Goldberg, 1996). Isoflavon adalah salah satu senyawa yang
termasuk dalam golongan flavonoid dan merupakan salah satu hasil metabolit
sekunder. Metabolit sekunder merupakan senyawa-senyawa yang terdapat pada
spesies tertentu dan sangat khas untuk setiap spesies. Metabolit sekunder
berperan untuk kelangsungan hidup suatu spesies dalam perjuangan
menghadapi keadaan darurat dan untuk bertahan hidup (Judoamidjojo et al.,
1992). Senyawa metabolit sekunder terbentuk pada saat tidak ada pertumbuhan
sel yang dikarenakan keterbatasan nutrien zat gizi dalam medium sehingga
merangsang dihasilkannya enzim-enzim yang berperan dalam pembentukan
metabolit sekunder dengan memanfaatkan metabolit primer untuk
mempertahankan kelangsungan hidupnya. Polisakarida, protein, lemak dan
asam nukleat merupakan penyusun utama dari makhluk hidup, karena itu disebut
metabolit primer. Proses metabolisme merupakan keseluruhan proses sintetis
dan perombakan zat-zat yang dilakukan oleh makhluk hidup untuk kelangsungan
hidupnya. Metabolit primer dari semua organisme sama meskipun sangat
berbeda genetiknya. Dibawah ini disajikan skema hubungan antara metabolit
primer dan metabolit sekunder yang disintesis oleh tumbuhan :

xli
Metabolisme Primer Metabolit Sentral Metabolit Sekunder
Gambar 8. Hubungan antara metabolit primer dan metabolit sekunder, (Muhanifa, 2000 dalam Ariani dan Hastuti, 2009)
3.1. Isoflavon Pada Kedelai
Mengingat berbagai potensi kedelai sebagai sumber gizi dan senyawa
aktif serta prospeknya untuk dikembangkannya produk-produk baru, kedelai
banyak disebut sebagai “The golden bean, the miracle bean, food for the future”.
Isoflavon yang dominan pada kedelai terdapat dalam bentuk glikosida,
sedangkan yang dominan pada produk kedelai yang sudah mengalami
fermentasi adalah aglukan (Coward et al., 1993). Bentuk senyawa glikosida
dipertahankan oleh tanaman sebagai bentuk inaktif sehingga disimpan oleh
tanaman dalam keadaan yang stabil.
Polisakarida Glukosa Glikosida
Polisakarida Pentosa Tetrosa
Asam amino aromatik
Fenilpropanoid
Alkaloid
Protein
Tritosa Asam amino
alifatik Poliketida
Asam asetat Lemak
Asam mevalonat
Terpena Karotenoid
Asam Nukleat Siklus Krebs Asam - asam Tetrapirol

xlii
Bentuk aktif glikosida adalah aglikon, yang dihasilkan dari pelepasan glukosa
dari ikatan glikosida (Anderson and Carner, 1997).
Isoflavon pada kedelai terdapat dalam empat bentuk, yaitu dalam bentuk
aglikon (Faktor-2, daidzein, genistein, dan glisitein); dalam bentuk glikosida
(daidzin, genistin dan glisitin); dalam bentuk asetilglikosida dan malonilglikosida
(Wang and Murphy, 1994).
Struktur kimia senyawa isoflavon glikosida (daidzin, genistin dan glisitin) terlihat
pada tabel 2. dihalaman berikutnya :
Tabel 2. Struktur Daidzin, Genistin dan Glisitin
Nama Senyawa Struktur
Genistin O
OH
OH
O
O
O
OH
HOH
HOH
H
CH2OH
H
Glisitin
OH
O
H3CO
OO
OH
HOH
H
H
OH
H
CH 2OH
O
Daidzin
3.2. Isoflavon Pada Tempe Kedelai
Kedelai mengalami berbagai perubahan pada proses pembuatan untuk
dijadikan tempe, baik melalui proses fisik maupun proses enzimatik yang
CH2OH
H H
O H H
OH
O H
O
O
O H
O
O H

xliii
dikarenakan oleh aktivitas mikroorganisme. Keterlibatan mikroorganisme pada
proses pembuatan tempe terutama terjadi pada proses perendaman,
dikarenakan oleh bakteri-bakteri pembentuk asam dan proses fermentasi oleh
kapang khususnya Rhizopus oligosporus. Sebagai akibat perubahan-perubahan
tersebut tempe menjadi lebih enak, lebih bergizi, dan lebih mudah dicerna. Salah
satu faktor penting dalam perubahan tersebut adalah terbebasnya senyawa-
senyawa isoflavon dalam bentuk bebas (aglikon), dan teristemewa hadirnya
Faktor-2 yang terdapat pada tempe tetapi tidak terdapat pada kedelai, ternyata
berpotensi tinggi (dibanding dengan jenis isoflavon yang lainnya) sebagai
antioksidan (Gyorgy et al., 1964).
Faktor-2 (6,7,4' tri-hidroksi isoflavon) merupakan senyawa yang sangat
menarik perhatian, karena senyawa ini terbentuk selama proses fermentasi oleh
aktivitas mikroorganisme. Senyawa ini mula-mula ditemukan oleh Gyorgy (1964)
pada ekstrak tepung tempe, yang merupakan senyawa konjugat/terikat dengan
senyawa karbohidrat melalui ikatan glikosida.
Selama proses pengolahan, baik melaui fermentasi maupun proses non-
fermentasi, senyawa isoflavon dapat mengalami biokonversi, terutama melalui
proses hidrolisis sehingga dapat diperoleh senyawa isoflavon bebas yang
disebut aglikon yang lebih tinggi aktivitasnya. Senyawa aglikon tersebut adalah
genistein, daidzein, glisitein dan faktor-2 (Purwoko et al., 2001).
Struktur dan sifat kimia daidzein, genistein, glisitein dan faktor-2 ditampilkan pada
gambar 9, 10, 11 dan 12 dihalaman berikut:
xliv
OH
OH
OOH
O
Genistein
OH
OH O
H3CO
O
Glisitein
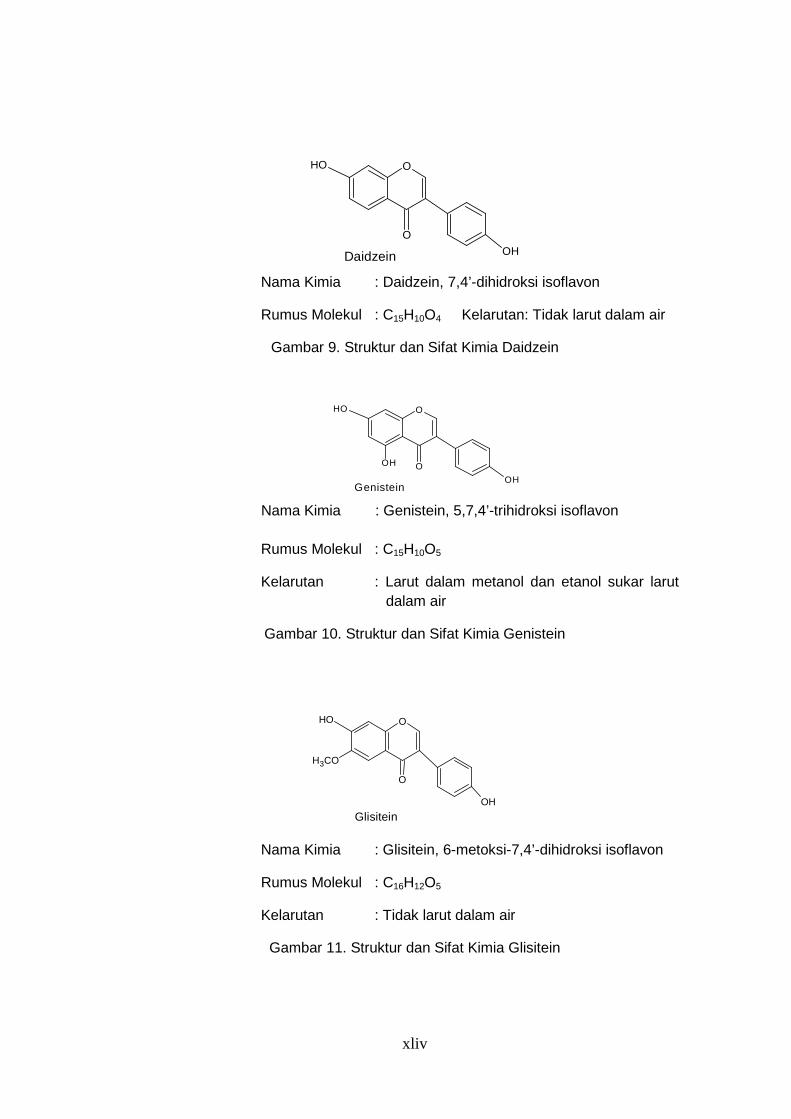
Nama Kimia : Daidzein, 7,4’-dihidroksi isoflavon
Rumus Molekul : C15H10O4 Kelarutan: Tidak larut dalam air
Gambar 9. Struktur dan Sifat Kimia Daidzein
Nama Kimia : Genistein, 5,7,4’-trihidroksi isoflavon
Rumus Molekul : C15H10O5
Kelarutan : Larut dalam metanol dan etanol sukar larut dalam air
Gambar 10. Struktur dan Sifat Kimia Genistein
Nama Kimia : Glisitein, 6-metoksi-7,4’-dihidroksi isoflavon
Rumus Molekul : C16H12O5
Kelarutan : Tidak larut dalam air
Gambar 11. Struktur dan Sifat Kimia Glisitein
OH
OOH
O
Daidzein

xlv
OH
OH
OH
O
O
Faktor II
Nama Kimia : Faktor-2 , 6,7,4’-trihidroksi isoflavon
Rumus Molekul : C15H10O5
Kelarutan : Tidak larut dalam air
Gambar 12. Struktur dan Sifat Kimia faktor-2
3.3.Metabolisme Isoflavon pada Proses Pengolahan Kedelai menjadi Tempe
Senyawa isoflavon merupakan salah satu komponen yang juga
mengalami metabolisme. Senyawa isoflavon ini pada kedelai berbentuk senyawa
konjugat dengan senyawa gula melalui ikatan -O- glikosidik. Selama proses
fermentasi, akan berlangsung proses hidrolisis, sehingga dibebaskan senyawa
gula dan isoflavon aglukan. Senyawa isoflavon aglukan ini dapat mengalami
transformasi lebih lanjut membentuk senyawa transforman baru. Hasil
transformasi lebih lanjut dari senyawa aglukan ini justru menghasilkan senyawa-
senyawa yang mempunyai aktivitas biologi lebih tinggi. Selama proses
pengolahan, baik melalui fermentasi maupun proses non-fermentasi, senyawa
isoflavon dapat mengalami biokonversi, terutama melalui proses hidrolisis
sehingga dapat diperoleh senyawa isoflavon bebas yang disebut aglukan yang
lebih tinggi aktivitasnya. Hal ini terlihat pada faktor-2, yang mempunyai aktivitas
antioksidan dan antihemolisis lebih baik dari daidzein dan genistein. Selain itu,
telah ditemukan bahwa senyawa isoflavon faktor-2 lebih aktif 10 kali lipat dari
senyawa karboksikroman (Pawiroharsono, 1995).
Faktor-2 dipandang sebagai senyawa yang sangat prospektif sebagai
senyawa antioksidan (10 kali aktivitas dari vitamin A atau karboksi kroman dan

xlvi
sekitar 3 kali dari senyawa isoflavon aglukan lainnya pada tempe) serta
berkhasiat antihemolisis. Reaksi hidrolisis glikosida isoflavon menjadi aglikon
isoflavon ditampilkan pada gambar 13 dibawah ini :
Gambar 13. Reaksi Hidrolisis Glikosida Isoflavon menjadi Aglikon Isoflavon
Purwoko et al., (1993) dalam Restuhadi (2001), menyatakan bahwa 99%
isoflavon glikosida yang terdapat pada biji kedelai, selama proses perendaman,
dapat terhidrolisis menjadi aglikon isoflavon dan glukosa. Pada proses fermentasi
kedelai rendam dengan kapang Rhizopus oligosporus, daidzein dan genistein
dapat mengalami proses hidroksilasi sehingga menjadi senyawa yang memilik
O
O H O H
O
O
O
O H
H O H
H O H
H H
Genistin
O H O H
O O H
O
Genistein
+
H H
O H O H
O H
O H
H
2OH
Glukosa
b - glukosidase
+ H 2O
CH2OH
CH20H
H H
O H H
O H
O H
O O
O H
O
O H
Daidzin
O H
O O H
O
Daidzein
+ H H
O H O H
O H
O
O H
H
H
OH
Glukosa
b - Glukosidase
+ H2 O
O H
O
H 3 CO
O O
O H
H O H
H
H
O H
H
CH2OH
O
Glisitin
O H
O H O
H 3 CO O
H H
O H O H
O H
O
O H
H
H
CH2 OH
Glisitein Glukosa
+
b - glukosidase
+ H 2O
OH
CH
CH2

xlvii
aktivitas fisiologis tinggi dan disebut faktor-2. Senyawa isoflavon faktor-2 ini tidak
dijumpai pada kedelai yang tidak difermentasi (Trilaksani, 2003).
3.4. Manfaat Senyawa Isoflavon dari Tempe Kedelai
Isoflavon pada tempe dapat mencegah aktivitas sel menjadi sel kanker,
tetapi juga dapat memperbaiki metabolisme hormon steroid, menurunkan
kolesterol dan trigleserida, serta melindungi sel-sel hati dari paparan senyawa
beracun. Selain itu isoflavon juga dapat berfungsi untuk memperlancar sirkulasi
darah. Isoflavon mempunyai beberapa efek posifif, diantaranya adalah
antiadrenalin, yang membuat jantung bekerja lebih santai, di samping
antiperadangan serta mencegah ketidak teraturan denyut jantung. Khususnya
isoflavon pada tempe yang aktif sebagai antioksidan, yaitu factor-2 terbukti
berpotensi sebagai anti-kontriksi pembuluh darah (konsentrasi 5 µg/ml) dan juga
berpotensi menghambat pembentukan LDL (low density lipoprotein). Dengan
demikian, isoflavon dapat mengurangi terjadinya arteriosclerosis pada pembuluh
darah (Zilliken, 1987). Isoflavon mempunyai struktur kimia hamper sama dengan
estrogen sehingga disebut fitoestrogen (Pakasi, 2000 dalam Iswandari, 2006)
Hasil olahan kedelai lain seperti minyak kedelai, juga dapat menangkal
kolesterol. Menurut Zilliken (1987), faktor-2 merupakan senyawa isoflavon yang
paling besar pengaruhnya. Karena itulah isoflavon menumbuhkan harapan cerah
pada pencegahan dan penurunan kejadian penyakit jantung. Manfaat senyawa
Isoflavon dibidang kesehatan diantaranya adalah sebagai antikanker/antitumor,
antivirus, antikolesterol, mencegah jantung koroner, mencegah osteoporosis dan
membantu terapi hormon estrogen (Pawiroharsono, 1998).

xlviii
A.4. ANTIOKSIDAN
4.1. Pengertian Tentang Antioksidan
Antioksidan didefinisikan sebagai senyawa yang dapat menunda,
memperlambat, dan mencegah proses oksidasi. Dalam arti khusus, antioksidan
adalah zat yang dapat menunda atau mencegah terjadinya reaksi oksidasi
radikal bebas dalam oksidasi lipid, protein dan karbohidrat, dalam proses
metabolisme yang berlangsung dalam tubuh (Winarsi, 2004). Antioksidan bekerja
dengan cara mendonorkan satu elektronnya kepada senyawa yang bersifat
oksidan sehingga aktivitas senyawa oksidan tersebut bisa dihambat.
Keseimbangan oksidan dan antioksidan sangat penting, karena berkaitan
dengan berfungsinya sistem imunitas tubuh. Defisiensi antioksidan yang berupa
vitamin C, vitamin E, Se, Zn dan glutation, dalam derajat ringan hingga berat,
sangat berpengaruh terhadap respon imunitas (Meydani, et al., 1995)
Radikal bebas adalah suatu molekul yang mempunyai satu elektron atau
lebih yang tidak berpasangan pada orbital luarnya. Adanya elektron yang tidak
berpasangan, menyebabkan senyawa tersebut sangat reaktif mencari pasangan,
dengan cara menyerang dan mengikat elektron dari molekul yang berada
disekitarnya, ia sangat reaktif dan merusak jaringan (Soeatmaji, 1998).
Radikal bebas merupakan molekul yang kehilangan elektron, sehingga
molekul tersebut menjadi tidak stabil dan selalu berusaha mengambil elektron
dari molekul sel lain. Radikal bebas dapat dihasilkan dari hasil metabolisme
tubuh dan faktor eksternal seperti asap rokok, hasil penyinaran UV, zat kimiawi
dalam makanan dan polutan lain. Penyakit yang disebabkan oleh radikal bebas
bersifat kronis, yaitu dibutuhkan waktu bertahun-tahun untuk penyakit tersebut
menjadi nyata. Radikal bebas sangat reaktif karena kehilangan satu atau lebih

xlix
elektron yang bermuatan listrik, dan untuk mengembalikan keseimbangannya
maka radikal bebas berusaha mendapatkan elektron dari molekul lain atau
melepas elektron yang tidak berpasangan tersebut.
Radikal bebas dalam jumlah berlebih di dalam tubuh sangat berbahaya
karena menyebabkan kerusakan sel, asam nukleat, protein dan jaringan lemak.
Radikal bebas terbentuk di dalam tubuh akibat produk sampingan proses
metabolisme ataupun karena tubuh terpapar radikal bebas melalui pernapasan.
Di dalam tubuh terdapat mekanisme antioksidan atau antiradikal bebas secara
endogenik. Tetapi bila jumlah radikal bebas dalam tubuh berlebih maka
dibutuhkan antioksidan yang berasal dari sumber alami atau sintetik dari luar
tubuh. Senyawa antioksidan ini akan menyerahkan satu atau lebih elektronnya
kepada radikal bebas sehingga dapat menghentikan kerusakan yang disebabkan
oleh radikal bebas.
Antioksidan dapat berasal dari dalam tubuh dan luar tubuh. Didalam
tubuh kita memiliki sistem enzym antioksidan yang bekerja secara simultan
memetabolisme radikal bebas sehingga tidak meninggalkan kerusakan pada
jaringan (Hodgson and Levi, 2000). Sementara itu jenis antioksidan yang lainnya
berasal dari luar tubuh, yaitu yang berasal dari makanan, atau komponen bahan
makanan (fitokimia) seperti fenol, atau alkaloid (Schultze et al., 1984).
Menurut Winarsi (2007), secara umum antioksidan dikelompokkan
menjadi dua, yaitu antioksidan enzimatis dan non-enzimatis. Antioksidan
enzimatis misalnya enzim superoksida dismutase (SOD), katalase dan glutation
peroksidase. Antioksidan non-enzimatis meliputi antioksidan larut dalam lemak
(tokoferol, karotenoid, flavonoid, quinon dan bilirubin) dan antioksidan larut dalam
air (vitamin C, asam urat, protein pengikat logam dan protein pengikat heme).

l
Senyawa antioksidan memiliki peran yang sangat penting dalam
kesehatan. Berbagai bukti ilmiah menunjukkan bahwa senyawa antioksidan
mengurangi resiko terhadap penyakit kronis, seperti kanker dan penyakit jantung
koroner (Amrun et al., 2007). Reaksi oksidasi lemak yang terjadi pada makanan
atau bahan makanan berlemak dapat dihambat dengan pemberian zat
antioksidan. Pada umumnya zat antioksidan yang digunakan adalah zat
antioksidan sintetik seperti Butylated Hydroxyanisole (BHA), Butylated
Hydroxytoluene (BHT), Propyl Gallat (PG) dan Etylene Diamine Tetra Acetic Acid
(EDTA). Sementara itu penggunaan zat antioksidan sintetik tertentu misalnya
BHT dapat menimbulkan akibat buruk terhadap kesehatan konsumen seperti
gangguan fungsi hati, paru, mukosa usus dan keracunan. Salah satu usaha
untuk mengatasi masalah tersebut adalah mengganti zat antioksidan sintetik
dengan zat antioksidan alami. Zat antioksidan alami dapat diperoleh dari ekstrak
bagian-bagian tanaman tertentu terutama yang banyak mengandung senyawa-
senyawa flavonoid yang tersusun dari gugus-gugus fenol (Suryo dan Tohari,
1995).
Perkembangan pengetahuan menunjukkan adanya hubungan antara
kimiawi radikal dengan keterlibatannya pada proses biologi normal ataupun pada
beberapa penyakit yang dihubungkan dengan ketuaan. Antioksidan merupakan
senyawa yang mampu menghambat oksidasi molekul lain. Tubuh tidak
mempunyai sistem pertahanan antioksidatif yang berlebihan, sehingga jika terjadi
paparan radikal bebas yang berlebihan, tubuh membutuhkan antioksidan
eksogen. Kekhawatiran terhadap efek samping penggunaan antioksidan sintetik,
maka antioksidan alami menjadi alternatip yang terpilih. Antioksidan alami
mampu melindungi tubuh terhadap kerusakan yang disebabkan oleh oksigen

li
reaktif, menghambat penyakit degeneratif serta mampu menghambat peroksidasi
lipid pada makanan. Beberapa tahun terkhir terjadi peningkatan minat untuk
mendapatkan antioksidan alami.
Jenis penggolongan antioksidan yang lain adalah berdasarkan sumber
diperolehnya senyawa tersebut. Penggolongan ini ada dua yaitu antioksidan
sintetik dan antioksidan alami.
4.2. Antioksidan sintetik
Antioksidan sintetik efektif dalam mencegah ketengikan pada minyak dan
bahan pangan berlemak (Purwoko 2001 dalam Meyri, 2003). Contoh antioksidan
sintetik adalah BHA, BHT, propil galat dan EDTA. Menurut Chang et al. (1977),
penggunaan BHT pada tikus percobaan dapat menyebabkan kerusakan organ
tubuh seperti paru-paru dan organ pencernaan. oleh karena itu penggunaan food
additive (bahan tambahan pada makanan) lebih baik dibatasi.
Penggunaan antioksidan tidak boleh berlebihan karena aktivitas
antioksidan akan hilang pada konsentrasi yang tinggi dan mungkin akan menjadi
prooksidan. Penggunaan antioksidan berlebihan akan menyebabkan senyawa
lebih bersifat sebagai akselerator daripada inhibitor dalam oksidasi lemak.
Dalam keadaan berlebih, antioksidan akan meningkatkan dekomposisi oksidasi
lemak dan pembentukan produk radikal.
4.3. Antioksidan Alami
Merupakan senyawa antioksidan yang diisolasi dari sumber alami dan
ditambahkan ke bahan makanan. Kebanyakan senyawa antioksidan yang
diisolasi dari sumber alami adalah berasal dari tumbuhan. Isolasi antioksidan
alami telah dilakukan dari tumbuhan yang dapat dimakan, tetapi tidak selalu dari
bagian yang dapat dimakan.

lii
4.3.a. Antioksidan Pada Kedelai
Salah satu senyawa dari tumbuhan yang memiliki kemampuan
bioaktivitas sebagai antioksidan adalah isoflavon. Isoflavon sebagai hasil
metabolit sekunder, banyak disintesa oleh tanaman angiospermae dan banyak
terdapat pada kelompok leguminoceae, khususnya pada tanaman kedelai. Pada
tanaman kedelai, kandungan isoflavon yang lebih tinggi terdapat pada biji kedelai
bagian hipokotil dan kotiledon (Anderson, 1997). Pawiroharsono (1995)
melaporkan bahwa kandungan isoflavon pada kedelai berkisar 2-4 mg/g.
Senyawa isoflavon pada kedelai, umumnya berupa senyawa kompleks
atau konjugasi dengan senyawa gula melalui ikatan glikosida. Jenis senyawa
tersebut diantaranya adalah genistin, daidzin, dan glisitin. Bentuk senyawa
demikiaan ini mempunyai aktivitas fisiologis kecil, apabila melalui proses
pengolahan (baik melalui proses fermentasi maupun non fermentasi), senyawa
isoflavon dapat mengalami transformasi terutama melalui proses
hidrolisis,sehingga dapat diperoleh senyawa isoflavon bebas yang disebut
aglikon yang lebih tinggi aktivitas fisiologisnya.
Selain isoflavon, asam fitat yang terkandung pada kelompok tanaman
leguminoceae, juga memiliki efek antioksidan. Asam fitat merupakan bentuk
penyimpanan fosfor terbesar pada tanaman sereal dan leguminosa.
Menurut Nuraida dan Yasni (1998) dinyatakan bahwa produk olahan
kedelai tanpa fermentasi, masih mengandung asam fitat tetapi fermentasi
Rhizopus pada kedelai dapat menghilangkan asam fitat, sehingga tempe tidak
mengandung asam fitat.

liii
4.3.b. Antioksidan Pada Tempe Kedelai
Dalam tempe kedelai terdapat empat jenis isoflavon yang mempunyai
khasiat antioksidan, yaitu daidzein, glisitein, dan genistein serta faktor-2. Selain
itu, isoflavon juga berpotensi untuk dimanfaatkan sebagai obat. Pada tabel
berikut, disajikan beberapa potensi pemanfaatan isoflavon yang diperoleh dari
hasil penelitian.
Tabel 3. Potensi Pemanfaatan Senyawa Isoflavonoid
No Isoflavon Bioaktivitas Referensi 1 Daidzein,Genistein Antioksidan, Antikanker Gyorgy et al.(1964) Glisitein, Faktor-2 Antioksidan, Anrikanker Kramer, et al (1984)
2 Faktor-2 Antioksidan, Antiinflamasi Zilliken (1987) Antikontriksi, Antikanker
3 Faktor – 2 Antiheimolitic Murata, Jha (1985) 4 Glisitein Antialergi Jha ( 1985 )
5 Daidzein,Genistein Estrogenik Oilis (1962 )
Snyder dan Kwon ( 1987)
Sumber : Pawiro Harsono ( 1998 )
B. Kerangka Berpikir
Senyawa isoflavon banyak ditemukan pada semua jenis tanaman, dan
dari berbagai hasil penelitian melaporkan bahwa tanaman leguminoceae /kacang
polong mempunyai kandungan isoflavon yang lebih tinggi bila dibandingkan
dengan jenis tanaman bukan leguminoceae. Buncis dan Kecipir merupakan dua
spesies dari keluarga leguminoceae, diduga bahwa buncis dan kecipir juga
memiliki kandungan isoflavon yang tinggi. Peneliti menggunakan kedelai kuning
sebagai kontrol pembanding, karena kedelai sudah diketahui dari berbagai
penelitian ilmiah sebagai sumber isoflavon yang dikenal oleh masyarakat luas.
Seberapa banyakkah kandungan isoflavon yang dimiliki oleh buncis dan kecipir,
serta bagaimanakah kemampuan aktivitas antioksidatifnya akan diketahui dari

liv
hasil penelitian ini. Lebih tinggi ataukah lebih rendah kandungan isoflavonnya
bila dibanding dengan senyawa antioksidan yang sudah dikenal oleh masyarakat
seperti vitamin C, alfatokoferol, betakaroten maupun BHT.
C.Hipotesis
1. Biji buncis dan kecipir hasil fermentasi 0, 1, 2, 3, 4 hari, mengandung senyawa
isoflavon yang bervariasi.
2. Kandungan isoflavon yang tinggi, memiliki kemampuan sebagai antioksidan
yang bagus (optimum).

lv
BAB III
METODOLOGI PENELITIAN
A. Waktu dan Tempat Penelitian
Penelitian tentang Karakterisasi Senyawa Bioaktif Isoflavon dan Uji
Aktivitas Antioksidan dari Ekstrak Tempe Berbahan Baku Buncis dan Kecipir,
dilakukan dengan metode eksperimental laboratorium yang berlangsung dari
bulan Februari 2009 sampai bulan Juli 2009.
Penelitian ini dilakukan di:
a. Laboratorium Program Kimia FKIP UNS
b. Sub Laboratorium Biologi Pusat MIPA UNS
c. Laboratorium Kimia Organik Fakultas MIPA UGM
B. Alat dan Bahan Penelitian
Alat-alat yang digunakan dalam penelitian ini adalah :
a. Alat untuk uji HPLC ( Perkin Elmer LC 295)
b. Alat untuk uji Aktivitas Antioksidan ( Spektrofotometer UV – VIS )
c. Blender (National)
d. Pipet mikro ( Master Pet )
e. Spatula
f. Gelas Erlenmeyer, Bekerglas, Petridish, Corong kaca, gelas arloji,
pengaduk, dan botol-botol vial.
Bahan-bahan yang digunakan dalam penelitian ini terdiri dari :
a. Kedelai kuning dari Madura, buncis dan kecipir dari Ndarjo Wonogiri
b. Etanol 95 % , Metanol p.a ( Merck )
c. Standar Genistein ( Sigma Chemical Co )
d. Standart Daidzein ( Sigma Chemical Co )

lvi
e. Standar Glisitein ( Sigma Chemical Co )
f. Standar Faktor-2 ( Sigma Chemical Co )
g. DPPH, BHT, Betakaroten, Alfatokoferol, Vitamin C (Sigma Chemical Co)
h. Metanol gradient grade for liquid chromatography ( Merck )
i. Aluminium foil, kertas saring, dan akuades
C. Prosedure Penelitian
1. Pembuatan Tempe Kedelai
Tempe kedelai dibuat dari biji kedelai sebanyak 0,5 kg dan diinokulasi
dengan 0,5 gram inokulum tempe. Sebelum diinokulasi, biji kedelai disortir agar
didapatkan kedelai dengan kualitas yang bagus. Biji kedelai kemudian direndam
selama 24 jam, kemudian kulit biji dikupas secara manual dengan menggunakan
tangan. Proses perendaman ini selain untuk menghilangkan bau langu pada
kedelai juga untuk melunakkan biji, sehingga diperoleh kedelai kupas yang
bersih dari kulit. Kemudian kedelai kupas dikukus 30 menit dan untuk selanjutnya
diproses lebih lanjut dengan cara difermentasikan dengan ragi tempe dan
menghasilkan tempe kedelai. Tahapan dalam pembuatan tempe kedelai
dijelaskan dalam uraian berikut :
a. Pemilihan Biji Kedelai
Biji kedelai yang akan dibuat tempe perlu dipilih agar diperoleh bahan
dasar tempe yang berkualitas baik, sehingga tempe yang dihasilkan juga baik.
Biji kedelai yang digunakan adalah biji kedelai yang bentuknya utuh, bernas,
berwarna kuning bersih dan tidak terdapat cacat pada seluruh permukaannya
seperti bekas hama, lubang-lubang, dan berkerut kulitnya.

lvii
b. Perendaman
Biji kedelai yang telah dipilih selanjutnya direndam dalam air biasa
selama 24 jam. Air diganti setiap 6 jam sekali untuk menghindari bau yang tidak
enak selama proses perendaman. Perendaman merupakan salah satu cara
dalam persiapan bahan, yaitu menyiapkan kedelai kupas sebelum
difermentasikan.
c. Pengupasan
Selama perendaman, biji kedelai dikupas kulitnya menggunakan tangan
untuk menjamin kebersihan dan kedelai tidak rusak. Biji kedelai yang telah
dihilangkan kulit arinya (kedelai kupas), selanjutnya dikukus.
d. Pengukusan
Biji kedelai yang telah dikupas selanjutnya dikukus menggunakan
dandang tertutup selama 30 menit. Pengukusan merupakan salah satu cara
persiapan bahan, sebelum difermentasi menjadi tempe. Proses pengukusan ini
juga merupakan proses untuk menambah kadar air pada biji kedelai, sehingga
kelembabannya memenuhi syarat untuk pertumbuhan jamur Rhizopus
oligosporus. Hasil dari pengukusan ini adalah kedelai kukus yang sedikit lunak
dan lebih besar ukuran bijinya.
e. Penambahan inokulum (ragi tempe)
Setelah sampel dalam keadaan tidak terlalu basah, ditaburi ragi atau
inokulum sebanyak 0,5 gram untuk 500 gram sampel kedelai. Inokulum yang
digunakan adalah Rhizopus sp. produk LIPI dengan merk RAPRIMA
Inokulum tempe yang digunakan berupa serbuk halus berwarna putih
kekuningan dan tidak berbau. Sebelum ditambahkan inokulum, kedelai hasil
pengukusan diangin-anginkan terlebih dahulu untuk mengurangi kadar air

lviii
berlebih pada kedelai. Pencampuran dilakukan dengan diaduk perlahan-lahan
secara merata agar jamur tumbuh pada permukaan biji-biji kedelai secara
optimum, sehingga diperoleh tempe yang teksturnya kompak dan merata.
f. Pengemasan
Kedelai yang telah diinokulasi selanjutnya dikemas menggunakan daun
pisang. Pengemasan bertujuan untuk mencegah kontaminasi bahan lain atau
mikrobia yang dapat mengganggu pertumbuhan jamur pada proses fermentasi
tempe.
g. Fermentasi
Kedelai yang telah diinokulasi dan dikemas, selanjutnya difermentasi
selama 0, 1, 2, 3 dan 4 hari. Fermentasi 0 hari merupakan kedelai kukus yang
setelah diinokulasi dengan ragi namun tidak difermentasikan lebih lanjut.
Fermentasi bertujuan memberi kesempatan pada jamur untuk melanjutkan
pertumbuhan. Mekanisme kerja pembuatan tempe kedelai dapat dilihat pada
lampiran 1.
2. Pembuatan Tempe berbahan biji Buncis dan biji Kecipir
Tempe dari bahan buncis dan kecipir, dibuat dari biji buncis dan biji kecipir
sebanyak masing-masing 0,5 kg dan diinokulasi dengan 0,5 gram inokulum
tempe. Sebelum diinokulasi, biji buncis dan biji kecipir disortir agar didapatkan biji
dengan kualitas yang bagus. Biji buncis maupun biji kecipir kemudian direndam
selama 3x24 jam, sambil dikupas kulitnya dan diganti air perendamnya sampai
tidak berwarna biru (air perendamnya bening). Tujuan dari perendaman adalah
untuk menghilangkan bau langu dan menghilangkan zat sianida yang terdapat
pada biji buncis dan kecipir. Setelah selesai perendaman dan pengupasan akan
dihasilkan biji buncis dan biji kecipir kupas, kemudian dikukus 60 menit, sehingga

lix
dihasilkan biji kupas yang sudah masak sekaligus relatif steril, untuk selanjutnya
diproses lebih lanjut secara fermentasi dengan variasi lama fermentasi (0, 1, 2, 3,
dan 4 hari) dan akhirnya dihasilkan tempe. Fermentasi 0 hari adalah biji legum
buncis dan kecipir yang sudah di inokulasi dengan ragi, tetapi tidak
difermentasikan lebih lanjut. Bagan alir pembuatan tempe berbahan buncis dan
kecipir ada pada lampiran 2.
3. Mengekstraksi Isoflavon dengan Metode Maserasi
Untuk mengisolasi isoflavon, dapat diuraikan sebagai berikut :
Sebanyak 100 g sampel diblender hingga terbentuk bubur, kemudian
dimaserasi dalam 250 ml etanol 70 % selama 24 jam, kemudian disaring dan
filtratnya ditampung. Residu ditambah dengan 100 ml etanol 70 %, kemudian
dimaserasi selama 24 jam, kemudian disaring dan filtratnya ditampung. Residu
kedua ditambah dengan 100 ml etanol 70 %. Filtrat hasil maserasi kemudian
dipekatkan dengan rotary vacuum evaporator hingga diperoleh ekstrak kental.
Ekstrak kental di oven selama 30 menit dengan suhu 50oC sehingga diperoleh
massa hasil ekstraksi, kemudian diidentifikasi isoflavonnya dengan metode
HPLC. Mekanisme Isolasi Isoflavon dengan metode maserasi dapat dilihat pada
lampiran 3.
4. Identifikasi Isoflavon
Adapun mekanisme kerja untuk mengidentifikasi isoflavon dari hasil
ekstraksi dengan etanol dari bahan baku legum, dapat diuraikan sebagai berikut :
Identifikasi isoflavon dengan menggunakan metode HPLC dilakukan
dengan pengkondisian instrumen HPLC dan pembuatan larutan sampel. Larutan
sampel dibuat dengan mengambil 1 mg massa hasil ekstraksi lalu masing-
masing dilarutkan dalam etanol 10 mL. Larutan kemudian disentrifuge lalu

lx
diambil 20 µL dengan alat injeksi. Selanjutnya sampel diinjeksikan ke dalam
HPLC setelah pengkondisian HPLC selesai. Menganalisa kromatogram HPLC
dengan menggunakan pembanding kromatogram isoflavon standar yang terdiri
dari daidzein, genistein, glisitein dan faktor-2. Adapun kondisi HPLC adalah
sebagai berikut:
a. Panjang Kolom : 10 cm
b. Jenis Kolom : Lichrosper (R) 100 RP-18 (non polar)
c. Fase Gerak : metanol : asam asetat 0,02 ( 57,5% ; 42,5%)
d. Volume Injeksi : 20 µL
e. Detektor : sinar UV pada panjang gelombang 265 nm
f. Suhu Oven : suhu kamar
Mekanisme kerja untuk mengidentifikasi Isoflavon dengan metode HPLC dapat
dilihat pada lampiran 4.
5. Uji Aktivitas Antioksidan
a. Pembuatan larutan DPPH dengan cara menimbang kristal sebanyak
7,88 mg DPPH dan dilarutkan dalam metanol 100 mL sehingga diperoleh
konsentrasi 0,2 mM sebagai larutan kontrol. Pengukuran absorbansi larutan
DPPH dilakukan dengan memipet 600 µL pelarut (metanol) ke dalam kuvet dan
ditambahkan larutan DPPH sampai volume 3 mL, kemudian ditutup dan dikocok
sampai homogen warnanya. Selanjutnya membuat spektra sinar tampak pada
panjang gelombang (λ) 400-600 nm dan mencatat absorbannya pada puncak
panjang gelombang 517nm sebagai absorban kontrol.
Untuk menganalisis adanya senyawa antioksidan dari hasil ekstraksi
kedelai kuning Madura, buncis dan kecipir Wonogiri menggunakan metode
DPPH. Prosedur yang dikerjakan untuk uji aktivitas antioksidan meliputi :

lxi
b. Pembuatan Larutan Sampel, dengan cara menimbang ekstrak
sebanyak 2 mg dan melarutkan ke dalam etanol 4 mL untuk membuat larutan uji
dengan konsentrasi 100 ppm. Kemudian pengukuran antioksidan bahan uji
digunakan metode yang sama, dimana 600 µL pelarut diganti dengan 600 µL
larutan uji (sampel). Selanjutnya membuat spektra sinar tampak pada panjang
gelombang (λ) 400-600 nm dan mencatat absorbannya pada puncak panjang
gelombang mendekati 517nm sebagai absorban sampel.
c. Pengukuran Kadar Antioksidan
Aktivitas antiradikal dihitung dengan metode DPPH dimana sampel
direaksikan dengan larutan DPPH. Aktivitas antiradikal diperlihatkan pada sistem
yang warnanya berubah dari ungu menjadi kekuningan.
Perubahan warna larutan menunjukkan aktivitas penangkapan radikal
bebas DPPH dan dapat diukur dengan perbedaan absorbansi yang dihasilkan
pada sampel dibandingkan dengan kontrol. Aktivitas antiradikal dinyatakan
dalam bentuk persen penangkapan radikal DPPH dan dihitung dengan
persamaan ( Yen dan Chen, 1995 dalam Ariani dan Hastuti, 2009).
absorbansi sampel % aktivitas antioksidan = ( 1 - ) x 100% absorbansi kontrol
Nilai 0% berarti tidak mempunyai aktivitas antiradikal bebas atau
antioksidan, sedangkan nilai 100% berarti peredaman total dan pengujian perlu
dilanjutkan dengan pengenceran larutan uji untuk melihat batas konsentrasi
aktivitasnya. Mekanisme pembuatan larutan DPPH dan pembuatan larutan
sampel serta uji aktivitas antioksidannya, dapat dilihat pada lampiran 5a. dan 5b.

lxii
E. Teknik Analisa Data
Isoflavon yang diperoleh dari hasil ekstraksi, dianalisa jenis-jenis
isoflavonnya dengan metode HPLC dengan menggunakan standar isoflavon
genistein, daidzein, glisitein dan faktor-2 sebagai pembanding.
Instrumen HPLC akan memberikan puncak-puncak kromatogram untuk
masing-masing jenis isoflavon. Untuk mengetahui jenis isoflavon dalam sampel,
dilakukan dengan membandingkan tr (waktu retensi) dari sampel dengan tr (waktu
retensi) standar yang telah diketahui. Dengan membandingkan luas puncak
kromatogram sampel dengan luas puncak kromatogram standar, akan diketahui
kadar setiap jenis isoflavon dalam sampel.
Untuk mengetahui perbedaan antar perlakuan fermentasi pada kedelai,
buncis, dan kecipir dengan pengukuran dan penghitungan aktivitas
antioksidannya, dianalisa dengan menggunakan program SPSS version 15,
analisa data berupa General Linear Model-Univariete. Untuk membandingkan
aktivitas antioksidan pada legum (kedelai, buncis,kecipir) dengan antioksidan
alami (α-tokoferol, β-karoten, dan vitamin C) serta BHT, dianalisa dengan
Compare Means – One Way Annova menggunakan program SPSS version 15.

lxiii
BAB IV
HASIL DAN PEMBAHASAN
Dalam bab ini, hasil yang di peroleh dari penelitian yang dilakukan akan di
tabulasikan dan disertai dengan pembahasan. Urutan hasil penelitian dan
pembahasannya akan disusun sebagai berikut :
A. Hasil Fermentasi Kedelai, Buncis dan Kecipir
Dari fermentasi yang dilakukan pada biji-biji leguminoceae (kedelai,
buncis, dan kecipir) dapat diamati bahwa pertumbuhan miselium kapang yang
paling optimum untuk ketiga jenis legum adalah fermentasi hari ke 2 dan ke 3.
Pada fermentasi hari pertama, pertumbuhan miselium belum banyak dan belum
merata, sedangkan fermentasi hari keempat sudah over fermented sehingga
kelihatan warnanya berubah dari putih menjadi kuning kecoklatan, kepadatannya
menurun dan disertai munculnya aroma amonia (sengak).
Dari hasil fermentasi hari ke 2 dan ke 3, tekstur tempe padat dan
pertumbuhan miseliumnya paling banyak dan tebal, diduga kandungan senyawa
isoflavonnya paling optimum (terutama faktor-2), bila hasil fermentasinya paling
optimum diharapkan kandungan isoflavonnya juga optimum, menurut hasil
penelitian (Gyorgy et al., 1964) dilaporkan bahwa isofdlavon faktor-2 hanya
dijumpai pada kedelai rendam dan hasil fermentasi oleh kapang Rhizopus sp .
Fermentasi hari ke 0, merupakan biji legume yang telah mengalami
proses perendaman sampai pengukusan dan penambahan inokulum, namun
tidak difermentasikan lebih lanjut (berarti tidak muncul miselium)..
Pada dasarnya, fermentasi hari ke 0 sebenarnya sudah terjadi proses fermentasi
awal yang berlangsung di saat perendaman biji, sehingga dimungkinkan telah
terjadi perubahan melalui proses hidrolisis senyawa glikosida isoflavon menjadi

lxiv
aglikon isoflavon. Pada proses perendaman juga terjadi proses pembentukan
senyawa asam organik yang dilakukan oleh bakteri pembentuk asam, seperti
Lactobacillus sp., Streptococcus thermopillus yang ditandai munculnya lendir dan
busa serta aroma kecut/asam pada air rendamannya (Kasmidjo, 1990).
Fermentasi hari ke 1, sudah muncul miselium berupa serabut-serabut
halus tetapi belum tebal dan perlekatan miselium dengan biji-biji belum merata.
Selama fermentasi tempe, hifa Rhizopus sp. menembus biji-biji yang keras dan
mengambil makanan dari dalam biji tersebut untuk pertumbuhannya. Mula-mula
hifa hanya menyebabkan lunaknya biji dengan dorongan mekanis akibat
pertumbuhannya, selanjutnya akan diikuti dengan berlangsungnya proses
perombakan secara enzimatis. Perubahan kimiawi dalam biji terjadi oleh aktivitas
enzim ekstraseluler yang dilepaskan oleh ujung-ujung miselia, antara lain enzim
lipase, fitase, proteolitik dan, enzim β-glukosidase (Kasmidjo, 1990; Suwaryono
dan Ismeini, 1998).
Fermentasi hari ke 4, miselium mulai berkurang yang ditandai dengan
perubahan warna dari putih menjadi kuning kecoklatan serta kepadatannya
berkurang, walaupun produk fermentasinya bila di iris tidak remuk (pecah),
berarti masih ada sisa-sisa miselium kapang di dalam tempe yang melekat pada
biji-biji, diperkirakan kandungan isoflavonnya juga mulai berkurang karena
berlangsung juga proses penguraian oleh bakteri yang ditandai oleh munculnya
bau busuk dan aroma amoniak. Aktivitas enzim lipase akan mendegradasi lemak
dalam kedelai menjadi asam-asam lemak yang akan digunakan sebagai sumber
energi untuk pertumbuhan Rhizopus sp, sedangkan aktivitas oleh proteolitik
dapat merombak protein kedelai menjadi asam amino sebagai sumber nitrogen,
kemudian asam amino ini akan masuk kedalam metabolisme intra sel dan

lxv
akhirnya melepaskan amoniak keluar dari sel. Akumulasi amoniak yang
dilepaskan selama fermentasi tempe dapat meningkatkan nilai pH.
Seiring dengan telah terurainya senyawa yang terkandung dalam kedelai secara
keseluruhan oleh aktivitas berbagai enzim yang dihasilkan oleh Rhizopus, maka
aktivitas pertumbuhan pun mulai berkurang. Nilai pH yang semakin meningkat
akan menghambat pertumbuhan Rhizopus, karena sudah tidak cocok dengan
lingkungannya. Peningkatan nilai pH dan penurunan aktivitas pertumbuhan
Rhizopus akan membuka kesempatan bagi bakteri penyebab kebusukan untuk
tumbuh dan berkembang biak, karena bakteri pembusuk pada tempe dapat
hidup pada nilai pH 6-10, diantaranya adalah Bacillus sp. (Suwahyono,1989;
Kasmidjo, 1990). Aktivitas enzim protease yang dihasilkan oleh bakteri
pembusuk akan semakin melunakkan tempe dan menghasilkan banyak
akumulasi amoniak dan menimbulkan bau busuk dan sengak. Untuk lebih
jelasnya, karakterisasi biji legume mentah dan hasil fermentasinya dapat dilihat
pada lampiran 7, 8, dan 9.
B. Hasil Ekstraksi Senyawa Isoflavon
Ekstraksi isoflavon dilakukan dengan cara maserasi menggunakan
pelarut etanol 70%. Etanol 70% diketahui mampu mengekstrak isoflavon secara
optimal (Kudou et al., 1991). Etanol merupakan salah satu pelarut optimum
untuk mengekstrak isoflavon dari kedelai, tetapi penggunaanya untuk skala
komersial masih dikaji lebih lanjut karena bersifat toksik (Susanto et al.,1998).
Dalam penelitian ini menggunakan pelarut etanol karena selain kepolarannya
mendekati metanol, etanol juga relatif tidak beracun. Proses penyiapan bahan
adalah dengan memotong tempe kedelai hasil fermentasi 1, 2, 3 dan 4 hari
dalam ukuran yang tipis, kemudian diblender hingga berbentuk bubur tempe.

lxvi
Khusus untuk tempe fermentasi 0 hari, langsung diblender setelah penambahan
inokulum. Pembuatan bubur tempe bertujuan untuk memperkecil ukuran partikel
sehingga diharapkan senyawa isoflavon yang terekstrak akan semakin banyak
karena interaksi antara pelarut dengan bahan yang akan diekstrak semakin
tinggi.
Bahan mentah dimaserasi dalam pelarut etanol selama 24 jam,
selanjutnya filtrat dipisahkan dari residu untuk diproses lebih lanjut menjadi
ekstrak yang murni. Pada penelitian ini, maserasi dilakukan dalam pelarut etanol
70%. Etanol merupakan pelarut yang sesuai untuk mengekstrak senyawa-
senyawa organik (Susanto, 1998). Dari proses maserasi dan penyaringan
diperoleh hasil berupa filtrat, kemudian filtrat tersebut diuapkan dengan rotary
vacum evaporator pada suhu 500 C sampai didapatkan ekstrak yang pekat atau
hampir semua etanol teruapkan. Pada saat merotari dengan rotary vacum
evaporator menggunakan suhu 500 C, dikarenakan etanol yang digunakan untuk
maserasi memiliki titik didih 800 C, sehingga untuk menguapkan etanol secara
perlahan-lahan dipanaskan secara tidak langsung (labu ekstrak berputar
dipermukaan air dengan suhu dibawah 800 C) agar senyawa-senyawa dalam
hasil ekstraksi tidak mengalami kerusakan tetapi etanolnya dapat menguap.
Isoflavon diketahui memiliki titik didih diatas 2000C (Ariani dan Hastuti, 2009)
sehingga pemanasan dengan suhu 500C tentu tidak akan merusakkan senyawa
isoflavon yang terkandung dalam ekstrak. Ekstrak tersebut selanjutnya disimpan
dalam oven dengan suhu 500 C untuk menguapkan pelarut yang masih tersisa.
Hasilnya adalah massa hasil ekstraksi seperti yang tercantum dalam gambar
grafik berikut ini :

lxvii
Gambar 14. massa hasil ekstraksi biji kedelai kuning dan produk fermentasinya
Gambar 15. massa hasil ekstraksi biji buncis dan produk fermentasinya
Gambar 16. massa hasil ekstraksi biji kecipir dan produk fermentasinya

lxviii
Dari gambar diatas dapat ditunjukkan, bahwa ekstraksi yang
menghasilkan massa ekstraks dari masing-masing legume (kedelai, buncis dan
kecipir) mentah beserta produk fermentasinya ternyata bervariasi, walaupun
ketiganya tergolong kelompok leguminoceae, Massa hasil ekstraksi yang
diperoleh dari biji buncis dan kecipir mentah ternyata lebih banyak daripada
produk fermentasinya kecuali pada kedelai. Massa hasil ekstraksi dari biji
mentah, terbanyak berasal dari biji kecipir yaitu (3,910 g), kemudian biji kedelai
kuning madura (3,422 g) dan paling sedikit diperoleh dari biji buncis sebanyak
2,550 g dari 100 gram sampel yang di ekstraksi. Massa hasil ekstraksi yang
diperoleh dari produk fermentasi, jumlah tertinggi pada tempe kedelai dan tempe
buncis 4hari, masing-masing (5,192 g) dan (1,241 g), sedangkan tempe kecipir
menghasilkan ekstrak tertinggi pada fermentasi 3hari yaitu sebanyak 3,310 g
dari setiap 100 g sampel.
Hasil ekstraksi yang diperoleh dari kedelai, buncis dan kecipir beserta
produk tempenya ternyata bervariasi, dimungkinkan karena varietasnya, sifat
keras atau lunaknya biji, kandungan zat yang ada dalam biji dan kepadatan
komponen zat yang ada didalam biji-biji tersebut, juga berbeda walaupun
termasuk dalam famili yang sama. Hasil pertanian selalu dipengaruhi oleh faktor-
faktor lingkungan seperti tanah tempat tumbuhnya, musim tanamnya, cara
pengelolaan tanaman dan pengelolaan hasil panennya. Senyawa yang
terekstraksi oleh etanol tentulah senyawa-senyawa yang kepolarannya sama
dengan etanol, diantaranya adalah protein, lemak, alkaloid dan yang akan
diambil dari sampel penelitian ini adalah isoflavon. Hasil Ekstraksi dari kedelai
kuning, buncis, kecipir, beserta produk tempenya dapat dilihat pada lampiran 10.

lxix
Massa hasil ekstraksi yang diperoleh dari biji buncis dan kecipir mentah
berwarna hitam pekat, karena masih banyak mengandung senyawa sianida
sedangkan massa hasil ekstraksi dari biji kedelai mentah berwarna coklat tua ,
kemungkinan kandungan sianidanya relatif lebih sedikit dan akan hilang pada
saat dilakukan perendaman dan pengukusan. Proses perendaman dalam air
(apalagi air panas) dan perlakuan fermentasi dapat menurunkan kandungan
sianida dan asam fitat, karena terjadi hidrolisis asam fitat menjadi inositol dan
asam fosfat oleh enzim fitase yang diaktifkan selama perendaman dan
fermentasi. Hasil penelitian Samson et al., 1987; Koswara, 1995; Pramita 2008),
melaporkan adanya Lactobacillus casei, Streptococcus jaecium, Klebsiella
pneumonia, Enterobacter cloaceae, Bacillus brevis dan Bacillus pumilus dalam
air rendaman kedelai. Dilaporkan juga bahwa bakteri jenis Bacillus sp
mempunyai aktivitas enzim fitase, yang dapat menguraikan asam fitat menjadi
inositol dan asam fosfat, sedangkan senyawa HCN dalam biji dapat terhidrolisis
disaat perendaman menjadi ion H+ dan CN - .
C.Hasil Identifikasi Isoflavon dengan menggunakan Metode HPLC (High Performance Liquid Chromatography)
Analisis dengan HPLC bertujuan untuk mengidentifikasi keberadaan
senyawa isoflavon faktor-2, daidzein, glisitein dan genistein dalam sampel biji
mentah, tempe kedelai, tempe buncis dan tempe kecipir pada variasi lama waktu
fermentasi ( 0, 1, 2, 3, dan 4 hari ). Seperti metode kromatografi yang lain,
analisis HPLC dilakukan dengan membandingkan waktu retensi dari senyawa
isoflavon standar dengan waktu retensi dari masing-masing sampel.
Adanya puncak-puncak yang memiliki waktu retensi relatif sama
dengan senyawa isoflavon daidzein, glisitein, genistein dan faktor-2 standar
menunjukkan bahwa dalam sampel tersebut terdapat kandungan isoflavon

lxx
daidzein, glisitein, genistein dan faktor-2. Penentuan waktu retensi senyawa
daidzein, glisitein, genistein maupun faktor-2 standar, dilakukan pada hari yang
sama dengan penentuan waktu retensi dari masing-masing sampel, agar
diperoleh kondisi yang sama.
Analisis kuantitatif senyawa isoflavon dilakukan dengan cara menghitung
luas kromatogram. Konsentrasi senyawa isoflavon faktor-2, daidzein, glisitein
dan genistein dapat diketahui, dengan mengalikan % luas masing-masing
senyawa isoflavon dalam kromatogram dengan massa hasil ekstraksi yang
dihasilkan.
Identifikasi isoflavon yang dilakukan pada massa hasil ekstraksi biji
kedelai kuning mentah, tidak ditemukan isoflavon faktor-2 tetapi ditemukan
daidzein, glisitein, dan genistein, sedangkan dari hasil fermentasinya ditemukan
faktor-2, daidzein, glisitein dan genistein. Identifikasi isoflavon pada massa hasil
ekstraksi biji buncis mentah, ditemukan faktor-2, daidzein dan glisitein,
sedangkan dari hasil fermentasinya ditemukan faktor-2, daidzein, glisitein, dan
genistein. Identifikasi isoflavon pada kecipir mentah ditemukan faktor-2, daidzein,
glisitein dan genistein, sedangkan pada hasil fermentasi kecipir diketahui bahwa
isoflavon faktor-2, daidzein, glisitein, dan genistein ditemukan pada hasil
fermentasi 0, 2, 3, dan 4 hari, sedangkan pada hasil fermentasi 1 hari tidak
ditemukan genistein. Menurut hasil penelitian (Trilaksani, 2003) dilaporkan
bahwa isoflavon faktor-2 tidak diketemukan pada biji kedelai yang tidak
difermentasi, tetapi dari hasil identifikasi penulis di dapatkan bahwa faktor-2
dapat ditemukan pada biji yang belum difermentasi oleh kapang Rhizopus sp.
tetapi jumlahnya sangat sedikit (biji kecipir: 0,001g/100 g sampel dan biji
buncis:0,006 g/100 g sampel).

lxxi
Walaupun belum mengalami fermentasi yang utama (menggunakan
inokulum Rhizopus sp), tetapi bila biji itu mengandung air disaat dorman
dimungkinkan ada proses hidrolisis dari senyawa glikosida menjadi aglikon yang
dilakukan oleh enzim β-glikosidase dalam biji. Dalam kondisi dorman, biji
mengandung senyawa glikosida (daidzin, genistin, dan glisitin) dapat terhidrolisis
menjadi senyawa isoflavon dalam bentuk aglikon (daidzein, genistein, glisitein,
dan faktor-2) serta glukosa oleh enzim ß-glikosidase yang ada dalam biji mentah.
Hidrolisis mudah terjadi karena faktor air dan suasana asam, bila waktu
penyimpanan biji ada kelembaban dalam waktu yang relatif lama (misalnya
disimpan dalam plastik atau disimpan di tempat tertutup) berarti ada faktor air /
kelembaban yang dapat mendorong bekerjanya enzim. Selain oleh faktor air,
keasaman (pH) 5, merupakan pH optimum bagi enzim ß-glikosidase untuk
beraktivitas (Iswandari, 2006).
Genistein (bentuk aglikon) pada biji buncis mentah tidak ditemukan,
kemungkinan memang tidak terkandung dalam bijinya (yang ada dalam bentuk
glikosida), tetapi pada hasil fermentasinya dapat ditemukan, diduga karena
adanya aktivitas kapang Rhizopus. Dalam proses perendaman dan fermentasi
dengan Rhizopus akan dihasilkan enzim ß-glikosidase yang berperan mengubah
isoflavon dari bentuk glikosida menjadi bentuk aglikonnya (Koswara,1995). Pada
proses perendaman dan fermentasi terjadi pembebasan senyawa aglikon
dengan pola yang identik yaitu terlepasnya glukosa dan aglikon isoflavon dari
ikatan glikosida, namun demikian pembebasan senyawa aglikon selama proses
fermentasi tempe lebih besar bila dibandingkan dengan proses perendaman
(Pawiroharsono,1995). Untuk mengetahui konsentrasi kandungan isoflavon,
dilakukan perhitungan dengan cara mengalikan besarnya luas % pada
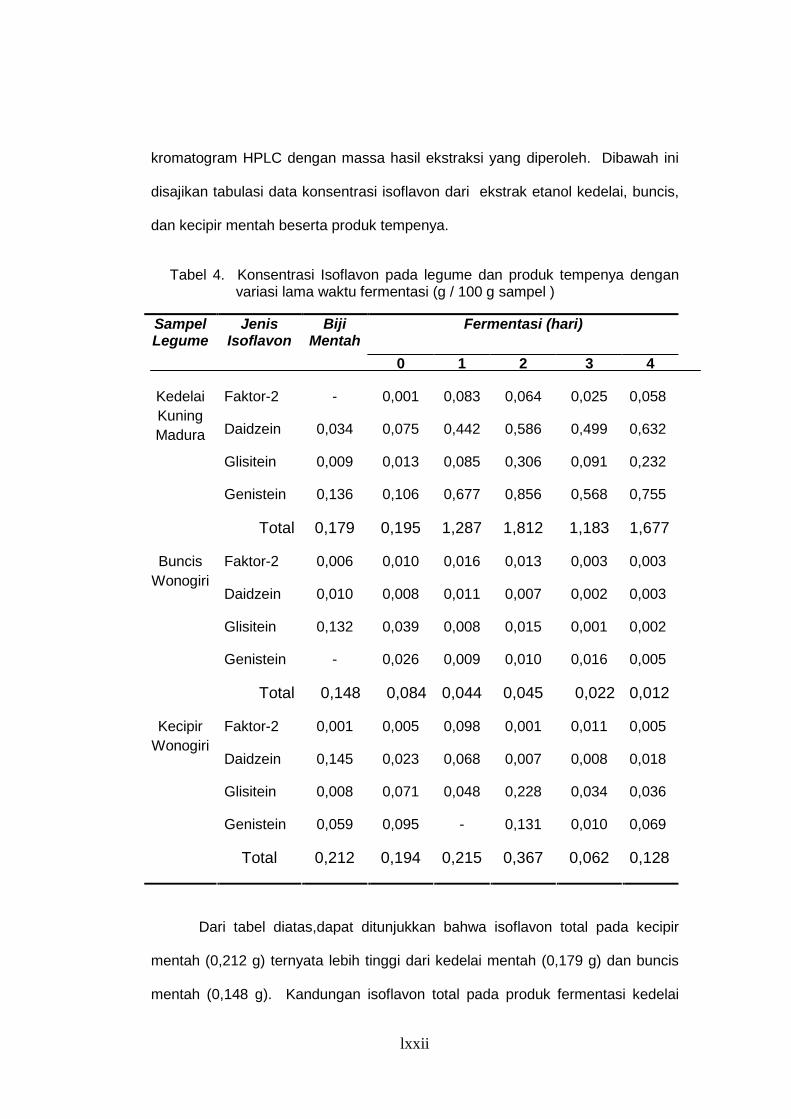
lxxii
kromatogram HPLC dengan massa hasil ekstraksi yang diperoleh. Dibawah ini
disajikan tabulasi data konsentrasi isoflavon dari ekstrak etanol kedelai, buncis,
dan kecipir mentah beserta produk tempenya.
Tabel 4. Konsentrasi Isoflavon pada legume dan produk tempenya dengan variasi lama waktu fermentasi (g / 100 g sampel )
Fermentasi (hari) Sampel Legume
Jenis Isoflavon
Biji Mentah
0 1 2 3 4
Faktor-2 - 0,001 0,083 0,064 0,025 0,058
Daidzein 0,034 0,075 0,442 0,586 0,499 0,632
Glisitein 0,009 0,013 0,085 0,306 0,091 0,232
Genistein 0,136 0,106 0,677 0,856 0,568 0,755
Kedelai Kuning Madura
Total 0,179 0,195 1,287 1,812 1,183 1,677
Faktor-2 0,006 0,010 0,016 0,013 0,003 0,003
Daidzein 0,010 0,008 0,011 0,007 0,002 0,003
Glisitein 0,132 0,039 0,008 0,015 0,001 0,002
Genistein - 0,026 0,009 0,010 0,016 0,005
Buncis Wonogiri
Total 0,148 0,084 0,044 0,045 0,022 0,012
Faktor-2 0,001 0,005 0,098 0,001 0,011 0,005
Daidzein 0,145 0,023 0,068 0,007 0,008 0,018
Glisitein 0,008 0,071 0,048 0,228 0,034 0,036
Genistein 0,059 0,095 - 0,131 0,010 0,069
Kecipir Wonogiri
Total 0,212 0,194 0,215 0,367 0,062 0,128
Dari tabel diatas,dapat ditunjukkan bahwa isoflavon total pada kecipir
mentah (0,212 g) ternyata lebih tinggi dari kedelai mentah (0,179 g) dan buncis
mentah (0,148 g). Kandungan isoflavon total pada produk fermentasi kedelai

lxxiii
dan kecipir yang tertinggi, diperoleh dari fermentasi 2 hari masing-masing
(1,812g) dan (0,367g), sedangkan untuk produk fermentasi buncis yang tertinggi
diperoleh dari fermentasil 0 hari (0,084 g).
Dari 100 g sampel yang digunakan, kandungan isoflavon Faktor-2 yang
terbanyak pada kedelai, buncis dan kecipir diperoleh dari fermentasi 1 hari, dan
kadar yang dimiliki oleh kecipir (0,098g) sedikit lebih tinggi dari kedelai (0,083g),
dan yang paling rendah dijumpai pada buncis (0,016g). Isoflavon Daidzein yang
terbanyak pada buncis dan kecipir diperoleh dari fermentasi 1 hari, sedangkan
pada kedelai kuning diperoleh dari fermentasi 4 hari. Jumlah daidzein pada
kedelai (0,632g) jauh lebih banyak dari yang dimiliki oleh kecipir (0,068g) dan
buncis (0,011g). Isoflavon Glisitein dan Genistein tertinggi pada kedelai dan
kecipir, diperoleh dari fermentasi 2 hari, sedangkan pada buncis diperoleh dari
hasil fermentasi 0 hari. Kandungan glisitein pada kedelai (0,036g) lebih tinggi
dari kecipir (0,228g) dan buncis (0,039g) dari 100 gr sampel. Kandungan
genistein pada kedelai (0,856 g), lebih tinggi dari kecipir (0,131 g) dan buncis
(0,026 g) dari 100 g sampel, ini berarti pola kecenderungan kandungan isoflavon
aglikon pada kedelai dan kecipir hampir sama, hal ini sesuai dengan laporan
penelitian Handajani et al., 1983 bahwa kedelai dan kecipir memiliki kandungan
senyawa yang hampir sama untuk protein, lemak, karbohidrat, mineral-mineral
serta asam aminonya.
Dari uraian diatas dapat disimpulkan bahwa kandungan jenis isoflavon
pada tempe buncis dan tempe kecipir sama dengan tempe kedelai kuning,
terutama hasil fermentasi 0,1,2,3 dan 4 hari, namun kadar isoflavonnya berbeda.
Dengan demikian buncis dan kecipir dapat dimanfaatkan sebagai pengganti
kedelai yang merupakan sumber isoflavon berkhasiat antioksidan.

lxxiv
Perendaman untuk buncis dan kecipir lebih lama, sehingga pada
fermentasi 1 hari sudah ditemukan isoflavon aglikon walaupun aktivitas dan
populasi Rhizopus sp belum optimum, diduga aglikon isoflavon sudah terbentuk
disaat perendaman. Untuk kedelai ditemukan isoflavon aglikon tertinggi pada
fermentasi 4 hari yaitu dalam kondisi over fermented dan sudah terjadi proses
pembusukan oleh bakteri-bakteri pembusuk. Dengan demikian, isoflavon aglikon
yang ditemukan pada biji dan hasil fermentasinya, dikarenakan oleh aktivitas
mikoorganisme disaat perendaman dan pembusukan, serta aktivitas kapang
Rhizopus sp, disaat proses pembentukan tempe.
Dari uraian diatas dapat disimpulkan bahwa kandungan isoflavon total
pada biji kecipir mentah ternyata lebih tinggi dari biji kedelai mentah , dan yang
paling sedikit ditemukan pada biji buncis mentah untuk setiap 100 g sampel
yang digunakan. Yang dimaksud dengan fermentasi hari ke-0 dalam penelitian
ini adalah biji yang sudah direndam selama 3x24 jam, kemudian dipanaskan
(dikukus), selanjutnya ditiriskan dan didinginkan, barulah diberi inokulum.
Walaupun Rhizopus belum beraktivitas, tetapi disaat perendaman sudah terjadi
fermentasi awal yang dilakukan oleh bakteri dari golongan termobakteri seperti
Lactobacillus sp, Streptococcus thermopillus dan Enterobacteriaceae yang
merupakan bakteri pembentuk asam yang banyak tumbuh pada proses
perendaman (Kasmidjo,1990), padahal perendaman untuk biji buncis dilakukan
selama 3x24 jam, berarti isoflavon aglikon yang terbentuk diduga berasal dari
proses hidrolisis yang dibantu oleh bakteri yang tumbuh dalam air rendaman.
Dari data yang diperoleh dapat diartikan bahwa kedelai kuning dan kecipir
produk fermentasi, ternyata menghasilkan jenis isoflavon yang sama, tetapi
kwantitasnya berbeda. Dalam penelitian ini menggunakan pembanding legume

lxxv
kedelai, karena sudah diketahui adanya kandungan isoflavon didalam biji kedelai
dan produk fermentasinya. Beberapa penelitian melaporkan bahwa kandungan
isoflavon pada kacang-kacangan dipengaruhi oleh varietas, waktu panen dan
lokasi penanaman (Mazur et al.,1998), waktu tanam (Aussenac et al.,1998), dan
kondisi iklim (Tsukamoto et al., 1995). Kondisi pertumbuhan, varietas, lokasi, dan
waktu tanam membedakan jumlah senyawa isoflavon (Harbone, 1996).
Senyawa isoflavon merupakan senyawa metabolit sekunder yang banyak
disintesa oleh tanaman angiospermae (tumbuhan berbiji tertutup). Oleh karena
itu, tanaman berbiji merupakan sumber senyawa isoflavon alami disekitar kita.
Dari berbagai tanaman, kandungan isoflavon yang lebih tinggi terdapat pada
kelompok tanaman leguminoceae dan tidak terdapat pada organisme seperti
bakteri, alga, jamur dan lumut (Markham, 1998). Senyawa metabolit sekunder
terbentuk pada saat tidak ada pertumbuhan sel yang dikarenakan keterbatasan
nutrient zat gizi dalam medium sehingga merangsang dihasilkannya enzim-enzim
yang berperan dalam pembentukan metabolit sekunder dengan memanfaatkan
metabolit primer untuk mempertahankan kelangsungan hidupnya. Polisakarida,
protein, lemak dan asam nukleat merupakan penyusun utama dari makhluk
hidup, karena itu disebut metabolit primer. Banyaknya kandungan jenis isoflavon
pada kedelai kuning, buncis dan kecipir mentah dan produk fermentasi 0, 1, 2, 3,
dan 4 hari disajikan pada gambar 16, 17, 18, 19 berikut ini:

lxxvi
Gambar 17. Kandungan Isoflavon Faktor-2 pada kedelai kuning, buncis, kecipir.
Gambar 18.Kandungan Isoflavon Daidzein pada kedelai kuning , buncis, kecipir.
Gambar 19. Kandungan Isoflavon Glisitein pada kedelai kuning, buncis, kecipir.
Gambar 20. Kandungan Isoflavon Genistein pada kedelai kuning, buncis, kecipir

lxxvii
Kandungan isoflavon faktor-2 pada kedelai, buncis, dan kecipir hasil
fermentasi memiliki kecenderungan hampir sama (kadar tertinggi ditunjukkan dari
tempe hasil fermentasi 1 hari), tetapi jumlah faktor-2 dari ketiga legum tersebut
secara keseluruhan lebih sedikit bila dibandingkan dengan kandungan daidzein,
glisitein dan genistein (Gambar 17 ). Pada kedelai kuning, kandungan isoflavon
faktor-2 akan menurun secara landai dan naik kembali pada fermentasi 4 hari.
Pada kecipir, terjadi penurunan kandungan isoflavon faktor-2 secara tajam pada
fermentasi 2 hari, sedangkan pada buncis ada kenaikan dan penurunan relatif
sedikit sehingga garis grafiknya tampak landai. Kadar isoflavon faktor-2 pada
ketiga legume relatif sedikit dan naik turun dengan pola yang berbeda karena
tempe yang dibuat menggunakan cara tradisional serta penyimpanan dialam
terbuka dan tidak menggunakan inkubator sehingga kontaminasi mikroflora
sangat mungkin terjadi dengan kontaminan yang bervariasi.
Kandungan daidzein pada kedelai hasil fermentasi ternyata jauh lebih
tinggi bila dibandingkan dengan buncis dan kecipir. Kadar tertinggi pada kedelai
diperoleh pada hasil fermentasi 4 hari (Gambar 18). Kadar daidzein yang rendah
pada buncis dan kecipir dapat terjadi diduga karena banyaknya senyawa yang
terbuang dari buncis dan kecipir disaat proses perendaman (direndam selama
3x24 jam dengan penggantian air setiap 8 jam). Proses perendaman yang lama
dimungkinkan terjadi reaksi hidrolisis, yang dilakukan oleh mikroorganisme yang
ada diair rendaman dengan dihasilkannya enzim β-glikosidase, sehingga akan
terbentuk isoflavon aglikon. Penggantian air rendaman sebanyak 9 kali,
memungkinkan terbuangnya banyak senyawa hasil hidrolisis, akibatnya
kandungan isoflavon pada buncis dan kecipir hasil fermentasi lebih sedikit bila
dibanding kedelai kuning (lama perendaman kedelai kuning 1x24 jam).

lxxviii
Kandungan glisitein dan genistein pada kedelai dan kecipir jauh lebih
tinggi bila dibandingkan dengan buncis, tetapi jumlah terbanyaknya memiliki pola
yang sama yaitu pada hasil fermentasi 2 hari. Pada buncis mentah ternyata
mengandung glisitein yang lebih tinggi daripada hasil fermentasinya, tetapi
isoflavon genistein pada buncis mentah tidak ditemukan (Gambar 19 dan 20),
diduga senyawa yang ada pada biji mentah masih dalam bentuk glikosida yaitu
glisitin. Dari hasil perbandingan ke-empat gambar diatas dapat disimpulkan
bahwa kadar isoflavon aglikon (faktor-2, daidzein, genistein dan glisitein) pada
buncis dan kecipir lebih rendah bila dibandingkan dengan kedelai kuning,
walaupun ketiganya termasuk dalam famili yang sama. Karena berbeda spesies,
sehingga masing-masing bijinya memiliki karakter dan kandungan senyawa
isoflavon yang tidak sama (Harbone,1996). Selain berbeda spesies, perendaman
yang lebih lama pada biji buncis dan kecipir diduga mengakibatkan hilangnya
senyawa hasil hidrolisis terutama disaat penggantian air rendaman.
Isoflavon faktor-2 pada kedelai kuning tanpa fermentasi tidak ditemukan,
hal itu sesuai dengan hasil penelitian Gyorgy et al., 1964; Trilaksani, 2003; Ariani
dan Hastuti, 2009), tetapi dari hasil penelitian penulis didapatkan isoflavon
faktor-2 dari biji buncis dan kecipir tanpa fermentasi, walaupun kadarnya sangat
sedikit (kecipir : 0,001g dan buncis : 0,006 g) dari100 gram sampel. Hal tersebut
dapat terjadi diduga ada proses pembentukan isoflavon faktor-2 disaat biji
dorman dalam penyimpanan. Bila penyimpanan biji dalam tempat yang tertutup
atau berada dalam plastik, maka akan muncul kelembaban dan memungkinkan
tumbuhnya mikroorganisme yang akan mendorong terjadnyai proses hidrolisisi,
dengan menghasilkan enzim β-glikosidase untuk memecah glikosida dalam biji
menjadi senyawa aglikon.

lxxix
D. Hasil Uji aktivitas Antioksidan
Uji aktivitas antioksidatif dilakukan dengan metode DPPH melalui
pengukuran absorbansi menggunakan spektrofotometer UV-Vis. Uji aktivitas
antioksidan dari penelitian ini terdiri dari biji mentah kedelai kuning beserta
produk tempenya, biji buncis mentah beserta produk tempenya serta biji kecipir
mentah dan produk tempenya.
Metode yang dipilih untuk pengujian aktivitas antioksidan adalah
metode DPPH karena sederhana, mudah, cepat dan peka serta hanya
memerlukan sedikit sampel. Metode aktivitas antiradikal bebas DPPH (2,2 difenil
1 picril hidrazil) merupakan metode terpilih untuk menapis aktivitas antioksidan
bahan alam (Molyneux, 2004; Luo et al., 2002; Santosa et al., 1998 dalam Amrun
dan Umayah, 2007). Senyawa antioksidan akan bereaksi dengan radikal DPPH
melalui mekanisme donasi atom hidrogen dan menyebabkan terjadinya
peluruhan warna DPPH dari ungu ke warna kuning. Perhitungan persentase (%)
peredaman DPPH oleh massa hasil ekstraksi biji kedelai kuning, buncis, dan
kecipir beserta produk fermentasinya ditampilkan pada lampiran 9.
Hasil identifikasi isoflavon dan uji aktivitas antioksidan dari sampel biji kedelai,
buncis dan kecipir beserta hasil fermentasi 0, 1, 2, 3 dan 4 hari terangkum dalam
tabel dibawah ini.
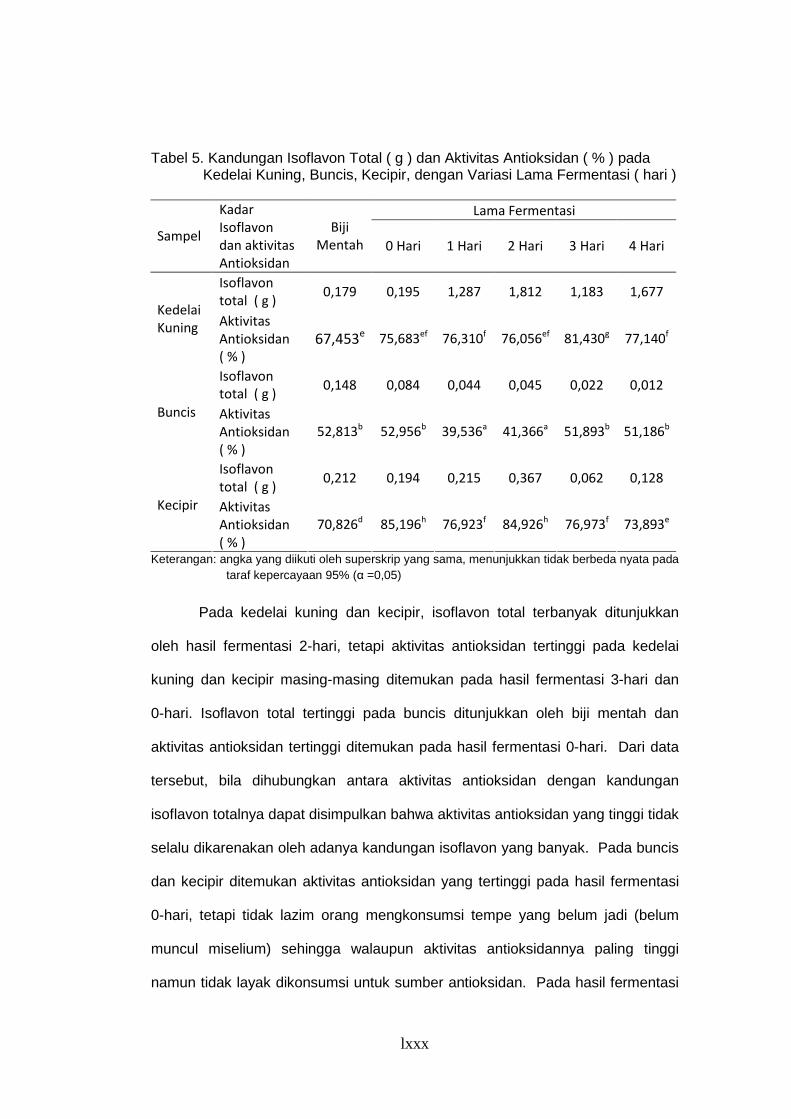
lxxx
Tabel 5. Kandungan Isoflavon Total ( g ) dan Aktivitas Antioksidan ( % ) pada Kedelai Kuning, Buncis, Kecipir, dengan Variasi Lama Fermentasi ( hari )
Lama Fermentasi
Sampel
Kadar Isoflavon dan aktivitas Antioksidan
Biji Mentah 0 Hari 1 Hari 2 Hari 3 Hari 4 Hari
Isoflavon total ( g ) 0,179 0,195 1,287 1,812 1,183 1,677
Kedelai Kuning Aktivitas
Antioksidan ( % )
67,453e 75,683ef 76,310f 76,056ef 81,430g 77,140f
Isoflavon total ( g ) 0,148 0,084 0,044 0,045 0,022 0,012
Buncis Aktivitas Antioksidan ( % )
52,813b 52,956b 39,536a 41,366a 51,893b 51,186b
Isoflavon total ( g ) 0,212 0,194 0,215 0,367 0,062 0,128
Kecipir Aktivitas Antioksidan ( % )
70,826d 85,196h 76,923f 84,926h 76,973f 73,893e
Keterangan: angka yang diikuti oleh superskrip yang sama, menunjukkan tidak berbeda nyata pada taraf kepercayaan 95% (α =0,05)
Pada kedelai kuning dan kecipir, isoflavon total terbanyak ditunjukkan
oleh hasil fermentasi 2-hari, tetapi aktivitas antioksidan tertinggi pada kedelai
kuning dan kecipir masing-masing ditemukan pada hasil fermentasi 3-hari dan
0-hari. Isoflavon total tertinggi pada buncis ditunjukkan oleh biji mentah dan
aktivitas antioksidan tertinggi ditemukan pada hasil fermentasi 0-hari. Dari data
tersebut, bila dihubungkan antara aktivitas antioksidan dengan kandungan
isoflavon totalnya dapat disimpulkan bahwa aktivitas antioksidan yang tinggi tidak
selalu dikarenakan oleh adanya kandungan isoflavon yang banyak. Pada buncis
dan kecipir ditemukan aktivitas antioksidan yang tertinggi pada hasil fermentasi
0-hari, tetapi tidak lazim orang mengkonsumsi tempe yang belum jadi (belum
muncul miselium) sehingga walaupun aktivitas antioksidannya paling tinggi
namun tidak layak dikonsumsi untuk sumber antioksidan. Pada hasil fermentasi

lxxxi
kecipir 2-hari diketahui aktivitas antioksidannya juga tinggi (84,926%), sehingga
tempe hasil fermentasi 2-hari dapat dikonsumsi oleh manusia sebagai sumber
antioksidan.
Dari analisis statistik Program SPSS, dapatlah disimpulkan bahwa
aktivitas antioksidatif pada kedelai dan kecipir hasil fermentasi, ternyata lebih
tinggi dari kedelai dan kecipir tanpa fermentasi (mentah). Dari tabel diatas dapat
diketahui juga bahwa, besarnya aktivitas antioksidan pada kedelai hasil
fermentasi 1-hari tidak berbeda nyata dengan fermentasi 4-hari, hasil fermentasi
0-hari tidak berbeda nyata dengan fermentasi 2-hari, tetapi berbeda nyata
dengan hasil fermentasi 3-hari. Besarnya aktivitas antioksidan pada buncis, hasil
fermentasi 0-hari, 3-hari, 4-hari tidak berbeda nyata dengan buncis tanpa
fermentasi. Pada kecipir tanpa fermentasi berbeda nyata dengan hasil
fermentasinya, tetapi hasil fermentasi 0-hari dan 2-hari serta fermentasi 1-hari
dan 3-hari tidak signifikan. Aktivitas antioksidatif pada tempe buncis yang paling
tinggi adalah hasil fermentasi 0-hari (±52,96%), kemudian 3-hari (±51,89%),
4-hari (±51,19%), 2-hari (±41,37%) dan yang terendah adalah hasil fermentasi
1-hari yaitu sebesar (±39,54%) ; sedangkan aktivitas antioksidatif untuk tempe
kecipir yang paling tinggi adalah hasil fermentasi 0-hari yaitu (±85,20); kemudian
2-hari (±84,93%); 3-hari (±76,97%); 1-hari (±76,92%) dan yang paling rendah
adalah hasil fermentasi 4-hari (±73,89%).
Dari hasil penelitian yang di uraikan diatas, dapatlah ditarik kesimpulan
bahwa tingkat aktivitas antioksidatif dari biji kecipir mentah dan hasil
fermentasinya ternyata tergolong tinggi, yaitu berkisar antara ±73,89 % sampai
dengan ±85,20%. Untuk kedelai kuning, tingkat aktivitas antioksidatifnya berkisar
antara ±68,64% sampai dengan ± 81,43 %, berarti lebih rendah dari kecipir dan

lxxxii
produk fermentasinya. Tingkat aktivitas antioksidatif dari biji buncis dan produk
fermentasinya ternyata lebih rendah (±39,54% s/d ±52,96% ) bila dibanding
dengan kedelai kuning. Pada kecipir mentah maupun hasil fermentasinya
memiliki kandungan isoflavon yang relatif rendah bila dibandingkan dengan
kedelai kuning, tetapi aktivitas antioksidannya tinggi. Ini dapat diartikan bahwa,
walaupun senyawa isoflavon pada kecipir mempunyai kemampuan sebagai
antioksidan, tetapi senyawa antioksidan itu tidak hanya isoflavon, dimungkinkan
di dalam kecipir terdapat senyawa-senyawa flavonoid dan alkaloid lain (selain
isoflavon) tetapi juga memiliki kemampuan sebagai antioksidan, bahkan telah
lama diyakini oleh sebagian masyarakat kita bahwa kecipir (baik biji maupun
buahnya) memiliki khasiat obat. Dari uraian diatas dapat disimpulkan bahwa,
aktivitas antioksidan yang tinggi tidak selalu dikarenakan oleh kadar isoflavon
yang tinggi, dan total isoflavon terbanyak tidak secara otomatis memiliki aktivitas
antioksidan yang tinggi.
Proses yang terjadi selama pengolahan biji legume menjadi tempe
adalah adanya reaksi hidrolisis yang berlangsung disaat perendaman biji bahan
baku, proses fermentasi Rhizopus disaat inokulum bekerja pada substrat, proses
pembusukan oleh bakteri yang terjadi pada tempe over fermented, hasil
fermentasi 4 hari.
Pada tempe buncis dan kecipir, aktivitas antioksidan yang maksimum
diperoleh dari hasil fermentasi 0-hari, dimungkinkan karena perendaman pada
biji buncis dan kecipir dilakukan selama 3 x 24 jam sehingga pada perendaman
yang lama tersebut akan terjadi proses hidrolisis yang lebih besar bila dibanding
dengan kedelai kuning (lama perendaman hanya 24 jam). Enzim β-glikosidase
yang dihasilkan oleh mikroorganisme disaat proses perendaman, akan

lxxxiii
melakukan pemecahan senyawa glikosida dalam biji menjadi aglikon dan
glukosa dan dapat berlangsung lebih dominan.
Kecipir hasil fermentasi 0hari diperoleh kadar isoflavon sebanyak
±0,194g/100g sampel dan kadar isoflavon tertinggi diperoleh setelah berlangsung
proses fermentasi 2 hari ±0,367 g/100 g sampel, ini berarti selama Rhizopus
bekerja pada substrat akan dihasilkan enzim β-glikosidase yang akan memecah
glikosida menjadi aglikonnya sehingga akan menambah jumlah senyawa
isoflavon aglikon. Pada hasil fermentasi 4-hari sudah mulai terjadi pembusukan
pada tempe yang ditandai dengan munculnya aroma busuk dan amonia yang
menyengat, dikarenakan adanya aktivitas mikroorganisme.
Aktivitas antioksidatif dari biji kedelai kuning, buncis, dan kecipir bila
dibandingkan dengan aktivitas antioksidan α-tokoferol, ß-karoten, vitamin C,
maupun BHT (Butyl Hidroksitoluena) yang merupakan antioksidan sintetis, dapat
dicermati pada gambar grafik berikut ini :
Gambar 21. Perbandingan aktivitas antioksidan alami, sintetis dan legume

lxxxiv
Bila dibandingkan dengan antioksidan alami (α-tokoferol, ß-karoten dan
vitamin C) ternyata aktivitas antioksidan yang dimiliki oleh kecipir dan produk
tempenya jauh lebih tinggi. Aktivitas antioksidatif β-karoten sebesar ± 43,25%;
vitamin C (± 75,62%), α-tokoferol (76,41 %), BHT (81,15%) dan kedelai
(81,43%), sedangkan kecipir fermentasi 0 hari tingkat antioksidatifnya mencapai
(± 85,19%). Selain itu, aktivitas antioksidatif pada kecipir hasil fermentasi 2 hari
(± 84,93%), juga lebih tinggi dari pengawet sintetis BHT. Dengan demikian,
kecipir dan hasil fermentasinya dapat digunakan sebagai sumber antioksidan
alami yang prospektif, karena keberadaannya melimpah di Indonesia. Aktivitas
antioksidatif pada buncis fermentasi 0 hari sebesar ± 52,95%, berarti masih lebih
tinggi dari β-karoten tetapi lebih rendah dari antioksidan BHT, vitamin C dan
α-tokoferol. Dengan demikian, buncis dan produk tempenya tidak dapat
digunakan sebagai pengganti BHT. Untuk mengetahui perbandingan aktivitas
antioksidan pada buncis, kecipir, dan kedelai dengan antioksidan alami yang
sudah ada (α-tokoferol, vitamin C dan β-karoten) serta antioksidan sintetis dapat
disajikan dalam tabel dibawah ini:
Tabel 6. Perbandingan Aktivitas Antioksidan Alami, Sintetis dan Legum (%)
Sampel Aktivitas Antioksidan
Betakaroten 43,2533 a
Tempe buncis (0 hari) 52,9567 b
Vitamin C 75,6200 c
Alfatokoferol 76,4100 c
BHT 81,1567 d
Tempe kedelai kuning (3 hari) 81,4300 d
Tempe kecipir (0 hari) 85,1967 e
Keterangan: Angka yang diikuti oleh superskrip yang sama, tidak menunjukkan beda nyata pada taraf kepercayaan 95% (α : 0,05)

lxxxv
Aktivitas antioksidan dari kecipir bila dibandingkan dengan antioksidan
alami (vitamin C, ß-karoten, α-tokoferol), dan BHT ternyata menunjukkan
perbedaan yang signifikan dan lebih besar, sedangkan aktivitas antioksidatif
pada buncis walaupun lebih rendah secara nyata dengan BHT tetapi lebih tinggi
bila dibanding dengan β-karoten, sehingga legum Kecipir dapat digunakan
sebagai pengganti BHT tetapi legum Buncis tidak dapat digunakan sebagai
pengganti BHT.
Penggunaan pengawet sintetis BHT dalam skala luas dimasyarakat dapat
menimbulkan dampak buruk bagi kesehatan, seperti gangguan fungsi hati, paru-
paru, mukosa usus bahkan bersifat meracuni (Suryo dan Tohari, 1995), oleh
karena itulah maka legume kecipir sangatlah potensial untuk dimanfaatkan
sebagai sumber antioksidan alami yang dapat digunakan untuk pengawet alami.
Kedelai digunakan sebagai pembanding, dengan pertimbangan bahwa
kedelai dan produk tempenya secara paten dan positif telah diketahui memiliki
manfaat sebagai antioksidan alami dan sumber isoflavon yang sudah tidak
diragukan, yang diperoleh dari beberapa hasil penelitian (Astuti, 1995 dan
Pawiroharsono, 1995). Buncis sebagai jenis legume , walaupun kemampuan
antioksidannya rendah tetapi masih dapat dimanfaatkan sebagai sumber
isoflavon yang berkhasiat, sekaligus sebagai bahan pangan sumber protein dan
mineral yang potensial bagi pemenuhan gizi masyarakat. Hasil pengukuran dan
perhitungan besarnya aktivitas antioksidan dari kedelai, buncis, kecipir dan
antioksidan alami (α-tokoferol, β-karoten, vitamin C) serta antioksidan sintetik
BHT dapat dilihat pada lampiran 6.

lxxxvi
BAB V
PENUTUP
A. Kesimpulan
Dari hasil penelitian yang telah dilakukan, dapat disusun kesimpulan
sebagai berikut :
1. Kecipir, Buncis dan produk fermentasinya memiliki potensi sebagai sumber
antioksidan alami yang prospektif dan mengandung senyawa isoflavon
faktor-2, daidzein, glisitein dan genistein. Total isoflavon terbanyak pada
kedelai dan kecipir ditemukan pada hasil fermentasi 2-hari, sedangkan
pada buncis ditemukan pada hasil fermentasi 0-hari
2. Lama waktu fermentasi yang optimum untuk menghasilkan ekstrak yang
mengandung isoflavon dengan aktivitas antioksidan maksimum pada
tempe kedelai kuning ditemukan pada hasil fermentasi 3-hari (sebesar
81,43 %) dan tempe koro buncis serta koro kecipir ditemukan pada hasil
fermentasi 0-hari, masing-masing 52,95 % dan 85,19 %.
3. Aktivitas antioksidan yang dimiliki oleh kecipir dapat digunakan sebagai
pengganti BHT, kedelai kuning, α-tokoferol, β-karoten maupun vitamin C,
sedangkan aktivitas antioksidan pada buncis hanya dapat menggantikan
β-karoten .

lxxxvii
B.Saran
Dari hasil penelitian yang diperoleh, penulis memberikan saran sebagai
berikut :
1. Perlu dilakukan penelitian secara in vivo mengenai manfaat senyawa
bioaktif yang terdapat dalam kecipir, maupun penelitian lanjutan untuk
mengetahui senyawa-senyawa lain (selain isoflavon) yang terkandung
dalam kecipir dan berpotensi sebagai antioksidan.
2. Senyawa bioaktif isoflavon dari buncis dan kecipir dapat dimanfaatkan
sebagai food suplement dan sebagai pengawet alami.
3. Biji kecipir dan produk fermentasinya dengan Rhizopus sp. dapat
disosialisasikan kepada masyarakat untuk dimanfaatkan sebagai sumber
antioksidan alami.

lxxxviii
DAFTAR PUSTAKA
Aldercreutz H. 1998. Epidemiology of phytoestrogens. Baillieres Clin. Endocrinol. Metab. 12: 605-623
Amrun,H; Umiyah dan Umayah E.U. 2007. Uji Aktivitas Antioksidan Ekstrak Metanol Beberapa Varian Buah Kenitu (Chrysopylum cainito L) dari Daerah Jember.. Jurusan Biologi Universitas Jember.
Anderson JJB, Carner SC. 1997. The effect of phytoestrogens on bone. Nutr.Res. 17: 1617-1623.
Ariani,S.R.D. dan Hastuti W. 2009. Analisis Isoflavon dan Uji Aktivitas Antioksidan pada Tempe dengan Variasi Lama Waktu Fermentasi dan Metode Ekstraksi. FKIP UNS Surakarta.
Astawan M. 2004. Sehat Bersama Aneka Serat Pangan Alami. Tiga Serangkai.
Solo
Astuti, Mary. 1995. Tempe dan Antioksidan Prospek Pencegahan Penyakit Degeneratif. Yayasan Tempe Indonesia.
Atikoh dan Supriyanti.1997. Perlakuan Perendaman, Pengukusan, Prebusan serta Kombinasinya terhadap Kandungan asam Fitat dan anti Kemotripsin pada Kacang Tholo dan Gude. Skripsi S1. UGM. Yogyakarta.
Aussenac T, Lacombe S, Dayde J. 1998. Quantification of isoflavones by capillary zone electrophoresis in soybean seeds : effects of variety and environment. Am. J. Clin. Nutr. (68):1480-1485.
Barz, W. and Papendorf. 1991. Metabolism of isoflavones and formation of factor-2 by tempeh producing microorganism.Tempeh Workshop, Cologne. 20 May 1991.
Barz, W., Heskamp, Klus,K.,Rehms, H and Steinkamp,R. Recent Aspect of Protein, Phytate and Isoflavone Metabolism by Microorganisms Isolated from Tempe Fermentation. Tempe Workshop. Jakarta. 15 February 1993.
Chang, S.S., Bostric-Matijasevic, O.A.L. Hsieh and C.L. Huang, 1977. Natural Antioxidants from Rosemary and Sage. J.Food Sci.42:574
Coward L, Barnes NC, Setchell KDR, Barnes S. 1993. Genistein, daidzein, and their ß-glycoside conjugates: antitumor isoflavones in soybean food from American and Asian diets. J. Agric. Food Chem. 41: 1967.

lxxxix
Dian Sri Pramita. 2008. Pengaruh Teknik Pemanasan Terhadap Kadar Asam Fitat dan Aktivitas Antioksidan Koro Benguk (Mucuna pruriens), Koro Glinding (Phaseolus lunatus), dan Koro Pedang (Canavalia ensiformis ). Skripsi. Fakultas Pertanian UNS. Surakarta.
Fujimaki. 1968. Fundamental Investigation of Proteolytic Enzim Aplication to Soybean Protein Inrelation Flavour. Tokyo University. Tokyo.
Goldberg I. 1996. Functional Foods : Designer foods, pharmafoods, nutraceuticals. London : Chapman & Halll, Inc.
Gyorgy P, Murata K, Ikehata H. 1964. Antioxidant isolated from fermented soybeans (tempeh). Nature, 203 (4947), 870-871.
Handajani S dan Bukle. 1991. Characteristic of Winged Bean (Psopocarpus tetragonolobus) seed. PhD Thesis, Universitas of New South Wales, Kensington, Australia.
Handajani S dan Windi Atmaka. 1993. Analisa Sifat Phisis-khemis Beberapa Biji Kacang-kacangan, Kekerasan, Kualitas Tanak, Protein, dan Kandungan Mineralnya. Lembaga Penelitian UNS. Surakarta.
Handajani S, Supriyono, Triharjanto, Marwanti S, Astuti D, Pujiasmanto B. 1996. Pengembangan Budidaya dan Pengolahan Hasil Kacang-kacangan Sebagai Usaha Produktif Wanita Di Lahan Kering Daerah Tangkapan Hujan Waduk Kedung Ombo. Lembaga Penelitian UNS. Surakarta.
Harborne JB. 1996. The Flavonoid : Advances in research since 1986. London : Chapman & Hall, Inc.
Hery Winarsi. 2007. Antioksidan Alami & Radikal Bebas. Kanisius, Yogyakarta.
Hesseltine, C.W. 1985. Genus Rhizopus and Tempeh Microorganisms. Asian Symposium Non-Salted Soybean Fermentation. Tsukuba, Japan.
Heyne K. 1987. Tumbuhan Berguna Indonesia (Jilid 3). Badan Penelitian dan Pengembangan Kehutanan. Departemen Kehutanan, Jakarta : Yayasan Sarana Wana Jaya. Terjemahan dari : De Nuttige Planten Van Indonesie.
Hidayati WB. 2003. Peran Isoflavon Untuk Kesehatan Reproduksi Wanita. Cermin Dunia Kedokteran 139 : 49-50.
Hodgson, E and P.E.Levi. 2000. A Textbook of Modern Toxicology. Elsevier, New York.
Iswandari R. 2006. Studi Kandungan Isoflavon Pada Kacang Hijau. IPB. Bogor.

xc
Judoamidjojo M, Darwis AA, Gumbira. 1992. Teknologi Fermentasi. Jakarta : Rajawali Press.
Kanetro B, Hastuti S. 2006. Ragam Produk Olahan Kacang-Kacangan. Universitas Wangsa Manggala Press. Yogyakarta.
Kasmidjo, R.B. 1990. Tempe: Mikrobiologi dan Biokimia, Pengolahan Serta Pemanfaatannya. Pusat Antar Universitas Pangan dan Gizi. UGM Yogyakarta.
Koswara, S. 2006. Isoflavon, Senyawa Multi Manfaat Dalam Kedelai. Ebookpangan.com, Bogor.
Kudou.S, Y.Fleury, D.Welti, D.Magnolato, K.Kitamura and K.Okubo. 1991. Malonyl Isoflavone Glycosides in Soybeans Seed (Glycine max Merril). Agric. Biol. Chem. 55: 2227-2233.
Lindajati, T. 1985. Large Scale Tempe Inoculum Production. Asian Symposium on Non salted Soybeans Fermentation, Tsukuba, Japan, July 14-16, 1985.
Markham KR. 1988. Cara Mengidentifikasi Flavonoid. Bandung : Penerbit ITB.
Mazur WM, Duke JA, Wahala K, Rasku S, Adlercreutz H. 1998. Isoflavonoids and lignans in legumes : nutritional and health aspects in humans. J Nutr Biochem 9: 193-200.
Messina MJ. 1991. The role of soy products in reducing risk of cancer. Journal NCL. 83(3) : 541-546.
Meydani, S.N, D. Wu, Santos, Hayek. 1995. Antioxidants and Immune Response in Aged Persons. Clinical Nutrition. 62 (6):1462-1476.
Meyri Sulasmi. 2003. Aktifitas Antioksidatif Ekstrak Tempe Hasil Fermentasi Rhizopus oryzae Terhadap Oksidasi Minyak Kedelai. Skripsi. FMIPA. UNS Surakarta.
Nuraida, L. dan S. Yasni. 1998. Kajian Gizi Produk Olahan Kedelai. Prosiding Seminar Pengembangan Pengolahan dan Penggunaan Kedelai selain Tempe. Kerjasama Pusat Studi Pangan dan Gizi-IPB dengan American Soybean Association. Bogor.
Pawiroharsono S. 1995. Metabolisme Isoflavon dan Faktor 2 (6,7,4’ trihidroksi isoflavon) pada Proses Pembuatan Tempe. Makalah Simposium Nasional Pengembangan Tempe dalam Industri Pangan Modern, Yogyakarta, 15-16 April.

xci
Pawiroharsono, S.1998. Benarkah Tempe Sebagai Anti Kanker. Jurnal Kedokteran dan Farmasi MEDIKA. 12: 815-817
Pratt, D.E. and B.J.F. Hudson. 1990. Natural Antioxidans not Exploited Comercially. Didalam: B.J.F. Hudson, editor. Food Antioxidants. Elsevier Applied Science. London.
Purwoko, T; S.Pawiroharsono dan I.Ginandjar. 2001. Biotransformasi Isoflavon oleh Rhizopus oryzae UICC 524. BioSMART, 3(2), 36-39.
Restuhadi, F. 2001. Studi Pendahuluan Biokonversi Isoflavon Pada Proses Fermentasi Kedelai Menggunakan Rhizopus sp. Tesis. Bandung. Magister Kimia ITB.
Rubatzky V.E, Yamaguchi M.1998. Sayuran Dunia : Prinsip, Produksi dan Gizi Jilid II. (diterjemahkan oleh Catur Herison). ITB Bandung.
Salunkhe D. K., and Kadam S.S. 1990. Handbook of world food legumes : Nutritional chemistry, processing tecknology, and utilization. Vol.1. CRC Press, Boca Raton, FL.
Schultze, J.E. 1984. R. Hansel and V.E. Tayler. Rational Phytotherapy A Physician’s Guide to Herbal Medicine. Springer Verlag, Heidelberg.
Setchell KDR, Aedin C. 1999. Dietary Isoflavones : biological effects and relevance to human health. J. Nutr. 129: 767-785
Setchell KDR. 2001. Bioavailability of pure isoflavones in healthy humans and analysis of commercial soy isoflavone supplements. J.of Nutrition. 131: 362-1375.
Shahidi,F. and Naczk. 1995. Food Phenolic. Technomic pub.Co. Inc. Lancester-
Bas.
Soeatmaji, D.W.1998. Peran Stress Oksidatif dalam Patogenesis Angiopati Mikro dan Makro DM. Medica.5 (24) : 318-325
Somaatmojo et al., 1985. Kedelai. Badan Penelitian dan Pengembangan Tanaman Pangan. Bogor.
Sri Kumalaningsih, 2006. Sumber, Manfaat, dan Penyajian Antioksidan Alami. Trubus Agrisarana. Jakarta.
Suryo dan Imam Tohari. 1995. Aktivitas Antioksidan Buah Jambu Mete dan Penerapannya pada Abon. Biosains. 1(7): 50-61

xcii
Susanto T, B Zubaidah, S.B.Wijanarko.. 1998. Studi Tentang Aktivitas Antioksidan pada Tempe Terhadap Lama Fermentasi, Jenis Pelarut dan Ketahanan Terhadap Proses Pemanasan. Fakultas Teknologi Pertanian Universitas Brawijaya Malang.
Suwahyono,U. 1989. Ringkasan Makalah Kongres Nasional V. Perhimpunan Mikrobiologi Indonesia. Yogyakarta. 4-6 Desember. 107-267.
Suwaryono,O dan Ismeini,Y.1988. Fermentasi Bahan Makanan Tradisional. PAU Pangan Gizi..22-25.
Taher A. 2003. Peran fitoestrogen kedelai sebagai antioksidan dalam penanggulangan aterosklerosis (tesis). Bogor : Program Pascasarjana, Institut Pertanian Bogor.
Trilaksani, W. 2003. Antioksidan : Jenis, Sumber, Mekanisme Kerja dan Peran terhadap Kesehatan. Bogor : Institut Pertanian Bogor.
Tsukamoto C, Shimada S, Igita K, Kudou S, Kokubun M, Okubo K, Kitamura K. 1995. Factors affecting isoflavones content in soybean seeds : changes in isoflavones, saponins, and compotition of fatty acids at different temperatures during seed development. J. Agric. Food Chem. 43:1184-1192.
Wang H, Murphy PA. 1994. Isoflavon compotition of American and Japanese soybeans in lowa : effects of variety, crop year, and location. J. Agric. Food Chem. 42 : 1674-1677.
Ziliken, F.I. 1987. Production of Novel Isoflavons. Material Meeting. BMBF, Bonn, Germany.
Zuheid, Noor.1989. Senyawa Antigizi (skripsi). Fakultas Teknologi Pertanian Universitas Gadjah Mada. Yogyakarta.







