
UNIVERSIDADE FEDERAL DO PARÁ
INSTITUTO DE ESTUDOS COSTEIROS
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA AMBIENTAL
MESTRADO EM ECOLOGIA DE ECOSSISTEMAS
COSTEIROS E ESTUARINOS
TAYANA MARIA CABRAL FERREIRA
EFEITO DE BORDAS NATURAIS E ARTIFICIAIS NA ASSEMBLÉIA DE FORMIGAS
DOS MANGUEZAIS
BRAGANÇA
2013

UNIVERSIDADE FEDERAL DO PARÁ
INSTITUTO DE ESTUDOS COSTEIROS
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA AMBIENTAL
MESTRADO EM ECOLOGIA DE ECOSSISTEMAS
COSTEIROS E ESTUARINOS
TAYANA MARIA CABRAL FERREIRA
EFEITO DE BORDAS NATURAIS E ARTIFICIAIS NA ASSEMBLÉIA DE FORMIGAS
DOS MANGUEZAIS
Dissertação de Mestrado apresentada ao Programa de Pós-
Graduação em Biologia Ambiental, do Instituto de Estudos
Costeiros, UFPA, como requisito final necessário à obtenção
do grau de Mestre em Biologia Ambiental, com ênfase em
Ecologia de Ecossistemas Costeiros e Estuarinos.
ORIENTADOR: Prof. Dr. Marcus Emmanuel B. Fernandes
CO-ORIENTADORA: Profª. Drª. Ana Yoshi Harada
BRAGANÇA
2013

TAYANA MARIA CABRAL FERREIRA
EFEITO DE BORDAS NATURAIS E ARTIFICIAIS NA ASSEMBLÉIA DE FORMIGAS
DOS MANGUEZAIS
Este trabalho será para obtenção do Grau de Mestre em Biologia Ambiental pelo
Programa de Pós-Graduação em Biologia Ambiental (Mestrado em Biologia de Ecossistemas
Costeiros e Estuarinos).
Aprovado em:
Banca Examinadora
________________________________________________
Prof. Dr. Marcus Emanuel B. Fernandes (Orientador) - UFPA
________________________________________________
Profª. Drª. Ana Yoshi Harada (Co-orientadora)- MPEG
________________________________________________
Profª. Drª. Rita de Cássia Oliveira dos Santos (Titular) - UFPA
________________________________________________
Profª. Drª. Marlucia Bonifácio Martins (Titular) - MPEG
________________________________________________
Prof. Dr. César França Braga (Titular) - UFPA

Dedico este trabalho aos meus pais,
José Maria e Maria das Graças,
Como sinal de gratidão pela confiança em mim depositada.

AGRADECIMENTOS
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), pela concessão da
bolsa;
À Universidade Federal do Pará (UFPA) e ao Museu Paraense Emílio Goeldi (MPEG);
Ao Programa de Pós-graduação em Biologia Ambiental (PPBA);
Ao Dr. Marcus Fernandes (Marquinho) e à Drª Ana Harada pela orientação;
Aos meus pais, José Maria e Graça, e meus irmãos Tiago, Tássia e Tomás, bem como à toda a
minha família pelo apoio e incentivo;
Ao meu companheiro Arthur Raiol;
Aos amigos-irmãos descobertos em Bragança: Lenita Sousa, Kelle Cunha, Mariana Santos,
Adriano Martins, Rosivan Pereira, Ilmarina Menezes, Dani Arruda, Cinthya Arruda, Dênis
Domingues, Kelli Garboza, e a todos que colaboraram para meu crescimento pessoal;
Aos ajudantes de campo: Milene, Diomara, Cássio, Thiely, Camila, Romário, Clarissa e Neto.

SUMÁRIO
CAPÍTULO 1 - Introdução Geral Página
Biologia das formigas 2
Formigas em manguezais 3
Formigas como bioindicadores 4
Fragmentação de habitats 5
Contexto da fragmentação dos manguezais em Bragança, Pará 7
Referências Bibliográficas
8
CAPÍTULO 2 – Artigo: “Efeito de bordas naturais e artificiais na assembléia de
formigas dos manguezais”
13
Resumo 14
Abstract 15
Introdução 16
Métodos 17
Área de estudo 17
Metodologia de coleta 19
Análise dos dados 20
Resultados 22
Assembléia de formigas de manguezal 22
Distribuição vertical de formigas 27
Comparação entre os sítios 31
Variação na assembléia de formigas no sentido borda-centro do manguezal 31
Discussão 34

Literatura citada 38
ANEXO 1: Regras para submissão do CAPÍTULO 2 42
Conclusão 44

1
CAPÍTULO 1
INTRODUÇÃO GERAL

2
INTRODUÇÃO GERAL
Biologia das formigas
As formigas constituem um grupo dominante de insetos himenópteros, exercendo papel-
chave na ecologia dos mais diversos ecossistemas terrestres (HOLLDOBLER; WILSON, 1990;
FOWLER, 1995). Elas constituem uma única família taxonômica, Formicidae, que apresenta uma
história de sucesso evolutivo desde o período Cretáceo, há aproximadamente 100 milhões de
anos atrás (WILSON, 1987). Segundo o catálogo online gerenciado por Barry Bolton (AntCat
2014), as formigas são atualmente classificadas em 21 subfamílias, 308 gêneros e 12.928
espécies. Para a região Neotropical são registradas 8 subfamílias, 119 gêneros e 3100 espécies
(FERNÁNDEZ, 2003).
O comportamento eussocial apresentado por estes insetos (com exceção de algumas
poucas formas parasitas) exerce papel crucial no sucesso do grupo, e pode ser caracterizado pelo
cuidado cooperativo da colônia, pela sobreposição de gerações de operárias e pelo
desenvolvimento de um avançado sistema de castas, que envolve a divisão de trabalhos dentro e
fora do ninho (WILSON, 2005).
A literatura acerca das formigas descreve casos de simbiose, tanto facultativa quanto
obrigatória, com mais de 465 espécies de plantas, pertencentes a 52 famílias (JOLIVET, 1996;
BRONSTEIN, 1998); com diversas espécies de artrópodes (DELABIE, 2001; HOLLDOBLER;
WILSON 1990), e com inúmeras espécies de fungos e microrganismos (BOURSAUX-EUDE;
GROSS 2000; D'ETTORRE et al., 2002).
As formigas podem utilizar uma variedade de ambientes para nidificação, desde o solo de
desertos (ANDERSEN; CLAY, 1996) e de florestas úmidas (WILKIE et al., 2007), embaixo de
pedras (BRIAN; BRIAN, 1951), em galhos na serapilheira (CARVALHO; VASCONCELOS,
2002); cavidades existentes nas plantas (TEPE et al., 2009;) e na copa das árvores (FLOREN;
LINSENMAIR, 2005).
Nestes microhabitats, utilizam uma ampla gama de itens alimentares (onivoria). Existem
espécies com dietas características, tal como as formigas cortadeiras que alimentam-se de fungo
cultivado no interior do ninho subterrâneo; as formigas que alimentam-se de exsudatos vegetais,
seja via nectários extraflorais ou simbiose com hemípteros e larvas de lepidópteros (YOO;

3
HOLWAY, 2011); e ainda aquelas predadoras especialistas, com dietas taxonomicamente
limitadas, como o gênero Strumigenys, predador de colêmbolos (CREIGHTON, 1937).
Formigas em manguezais
Nos ecossistemas terrestres, a maior riqueza de formigas é registrada no solo, devido este
ambiente apresentar maior complexidade estrutural e estabilidade microclimática. Em
ecossistemas transicionais, como o manguezal, o solo representa uma limitação de sítios para
nidificação das formigas, favorecendo o estilo de vida arbóreo. Desta forma, ao forragear
constantemente sobre a vegetação de mangue, as formigas podem eventualmente proteger a
planta hospedeira contra herbívoros, exercendo um efeito positivo no desempenho reprodutivo
vegetal (OFFENBERG et al., 2006a).
Cannicci et al (2008) revisaram o efeito da fauna na estrutura da vegetação e no
funcionamento do manguezal, e destacaram a importância numérica e ecológica das formigas.
Segundo os autores, os trabalhos publicados sobre formigas em manguezais investigam a
composição e a distribuição espacial de espécies (WETTERER; O’HARA, 2002), bem como
seus comportamentos de nidificação e adaptações fisiológicas (NIELSEN, 1997). Além disso,
diversos estudos demonstram que a acentuada territorialidade apresentada pelas formigas
arborícolas no manguezal pode afastar importantes herbívoros (ADAMS, 1994), embora o custo
de manutenção da colônia pela árvore seja elevado (OFFENBERG et al., 2006b).
Vale ressaltar que a mirmecofauna de manguezal é composta por espécies de florestas de
terra firme, bem como por espécies urbanas dispersas através de atividades antrópicas sobre o
manguezal (DELABIE et al., 2006). Estas espécies cuja ocorrência e dispersão estão
necessariamente sujeitas à atuação humana são chamadas de sinantrópicas, e sua incidência pode
ser um indicativo do grau de perturbação do manguezal, em decorrência das atividades humanas
desenvolvidas em seu entorno.

4
Formigas como bioindicadores
Eventos de perturbação ambiental geram diferentes respostas na estrutura da comunidade
de formigas, e podem refletir mudanças em grupos de organismos animais e vegetais
(ANDERSEN; SPARLING, 1997; WATT et al., 2002). Em bordas de habitats criadas após
fragmentação, a alta insolação influencia positivamente a produção de plantas pioneiras, recurso
que promove aumento populacional de formigas-cortadeiras, e consequentemente da herbivoria,
na “zona de efeito da borda” (URBAS et al., 2007). Por outro lado, especialistas de floresta como
as formigas de correição, são suscetíveis à fragmentação florestal e evitam pequenos fragmentos,
devido seu hábito nômade requerer ampla área de vida (SUAREZ et al., 1998). Em áreas de
manguezal sujeitas a um gradiente de impacto antrópico no sudeste da Bahia, Delabie et al
(2006) revelaram que a riqueza de espécies foi negativamente correlacionada com o nível de
impacto humano. Segundo os autores, a ocupação humana nos manguezais e nas suas periferias
tende a depauperar a comunidade de formigas arborícolas, induzindo à uniformidade
composicional.
Outros exemplos da utilização de formigas como bioindicadores foram sumarizados por
Underwood e Fisher (2006), os quais detectaram cinco áreas de pesquisa em que as formigas
podem gerar valiosas informações para planos de manejo, baseados no monitoramento ambiental:
i) detecção de tendências entre espécies ameaçadas e aquelas em risco de extinção,
exemplificado no estudo da simbiose entre formigas do gênero Myrmica com
borboletas Maculinea. O estado ameaçado de conservação em que se encontram
as borboletas atenta para a necessidade prioritária de conservação das áreas em
que estas ocorrem (ALS et al., 2004; ELMES; THOMAS, 1992);
ii) detecção de tendências entre espécies-chave, tal como as formigas cortadeiras, as
quais comumente ocupam bordas recentemente criadas em florestas
fragmentadas, o que pode acarretar uma mudança no recrutamento e
consequentemente alterar a assembléia de plantas numa zona de influência de
60-120m² ao redor do ninho (CORRÊA et al., 2010);
iii) avaliação das ações de manejo da paisagem, tais como a silvicultura
(VASCONCELOS et al., 2000), pastagens (DIAS et al., 2008), agricultura

5
(ANDERSEN et al., 2002), mineração (MAJER, 1983) e fogo (IZHAKI et al.,
2003);
iv) acesso às mudanças no ecossistema a longo prazo, monitorando grupos-chave de
formigas e sua relação com a restauração das propriedades físico-químicas do
solo após queimadas (OLIVEIRA; FRANKLIN, 1993), atividade mineradora
(MOUTINHO et al., 2005) e fragmentação florestal (VASCONCELOS, 1998;
VASCONCELOS et al., 2006).
v) detecção da presença de espécies invasoras, as quais são conhecidas na literatura
por impactar de forma negativa a diversidade de formigas nativas, além de
reduzir as populações de insetos e vertebrados e alterar a dispersão de sementes
nas plantas (STORZ; TSCHINKEL, 2004; WARD; HARRIS, 2005;
HOLWAY, 2005). Estas formigas formam um grupo distinto de pelo menos
150 espécies com perfil sinantrópico, disseminadas em novos habitats
principalmente através de atividades humanas (tramp ants) (MCGLYNN,
1999).
Fragmentação de habitats
A fragmentação de habitats é considerada um processo natural, haja vista que resulta de
mudanças ambientais como, por exemplo, as glaciações, queimadas e erupções vulcânicas, as
quais geram descontinuidades na paisagem. Entretanto, a modificação da paisagem natural por
intervenção humana intensifica este processo, promovendo destruição e mudança na estrutura dos
habitats. Segundo Viana e Tabanez (1996) um fragmento florestal pode ser definido como
qualquer área de vegetação natural contínua interrompido por barreiras naturais (lagos, tipos de
solo, formações vegetais ou rochosas) ou barreiras antrópicas (estradas, hidrelétricas, culturas
agrícolas, pecuária, ocupações rurais ou urbanas).
A construção de rodovias, ferrovias e trilhas em áreas florestais é apontada como uma das
formas mais frequentes de criação de bordas artificiais de habitats, resultando na reconfiguração
da paisagem natural (ANDREWS, 1990). Os efeitos ecológicos de uma rodovia estendem-se
sobre os componentes bióticos e abióticos do sistema, e podem ser visualizadas como mudanças
na dinâmica de populações de plantas e animais, alteração do fluxo de materiais na paisagem,

6
introdução de elementos exóticos e mudanças na disponibilidade de água, luz e nutrientes
(FORMAN; ALEXANDER, 1998).
A fragmentação de hábitats por rodovias também promove condições ambientais
diferenciadas daquelas experimentadas no interior da área fragmentada, produzindo mudanças
bióticas e abióticas locais, denominadas genericamente de “efeitos de borda”. Segundo Murcia
(1995), estes efeitos podem ser mensurados até 150 m no interior da área fragmentada, e podem
ser classificados em três tipos: i) efeitos abióticos, os quais são resultantes de alterações na
luminosidade, temperatura do ar e umidade, por exemplo, próximo à borda do fragmento; ii)
efeitos bióticos diretos, visualizados como mudanças na estrutura e composição da fauna e flora
em função das alterações físicas ocorridas na borda do fragmento e iii) efeitos bióticos indiretos,
que representam as mudanças nas interações entre espécies como, por exemplo, a predação,
competição, dispersão e herbivoria.
A crescente fragmentação dos habitats via a criação de bordas artificiais tem alterado a
composição de espécies de formigas em habitats naturais, por meio da introdução acidental ou
não de espécies sinantrópicas, as quais possuem eficientes estratégias de estabelecimento
(GRAHAM et al., 2004, FAHRIG, 2003). A disparidade numérica alcançada por algumas
espécies sinantrópicas introduzidas determina sua influência sobre os outros componentes da
biota, conferindo então à espécie status de invasora (HOLWAY et al., 2002, TSUTSUI;
SUAREZ, 2003; SUAREZ; TSUTSUI, 2008).
Muitas destas espécies invasoras nidificam de forma efêmera e tem uma estrutura colonial
variável tanto temporal quanto espacialmente, permitindo mover rapidamente suas colônias em
resposta á variações ambientais. Podem ainda nidificar em materiais que são transportados pelo
homem, tal como lixo, solo ou vasos de plantas, dispersando-se assim para grandes distâncias
(SUAREZ et al., 2001; SUAREZ et al., 2005). Sua reprodução ocorre geralmente por construção,
onde as rainhas inseminadas partem do ninho oficial e fundam subunidades, vinculadas entre si e
com ausência de agressividade entre seus membros.

7
Contexto da fragmentação dos manguezais em Bragança, Pará
O extenso ecossistema de manguezal existente na região sofreu um severo processo de
fragmentação, por meio da construção da Rodovia PA-458, a qual foi iniciada em 1974 e
concluída em 1983. Esta rodovia secciona aproximadamente 26 km de floresta de mangue, e foi
projetada com o objetivo de ligar a cidade de Bragança à praia de Ajuruteua, incentivando o
turismo na região e facilitando o escoamento do pescado proveniente da Vila dos Pescadores.
A execução deste projeto ocasionou a remoção de 86 ha de floresta de mangue, obstrução
de canais de maré, compactação do solo e degradação florestal, além da criação de um lago
artificial. Estudos locais revelaram que esta rodovia impactou negativamente os componentes da
fauna bentônica (NASCIMENTO, 2005), além de ocasionar eventos constantes de atropelamento
da fauna associada às florestas de mangue (PEREIRA et al., 2006).
Partindo do pressuposto que a modificação estrutural da paisagem em função de
atividades humanas resulta em mudanças na composição e riqueza de espécies de formigas, este
estudo buscou avaliar se a presença da rodovia PA-458 afeta a mirmecofauna associada ao
manguezal, promovendo a ocorrência de espécies de formigas sinantrópicas, e tomando-as como
uma medida do grau de impacto ambiental.
Inicialmente, a mirmecofauna foi inventariada em um sítio de manguezal próximo à
rodovia, considerando esta como uma barreira artificial de habitat. Ao mesmo tempo,
amostramos a mirmecofauna em um sítio de manguezal situado na borda de um canal de maré,
considerando este como uma barreira natural de habitat. Foi feita uma comparação da
composição de espécies entre os sítios de coleta, partindo da teoria de que a incidência de
formigas sinantrópicas está mais relacionada à borda artificial, apresentando ainda maior
diversidade quanto mais próximo da rodovia.

8
REFERÊNCIAS BIBLIOGRÁFICAS
ADAMS, E. S. Territory defense by the ant Azteca trigona: maintenance of an arboreal ant
mosaic. Oecologia v. 97, p. 202-208. 1994.
ALS, T. D. et al. The evolution of alternative parasitic life histories in Large Blue butterflies.
Nature v. 432, p. 386–390. 2004.
ANDERSEN, A. N.; CLAY, R. E. The Ant Fauna of Danggali Conservation Park in Semi-arid
South Australia: a Comparison with Wyperfeld (Vic.) and Cape Arid (W.A.) National Parks.
Australian Journal of Entomology v. 35, p. 289-295. 1996.
ANDERSEN, A. N.; SPARLING, G. P. Ants as Indicators of Restoration Success: Relationship
with Soil Microbial Biomass in the Australian Seasonal Tropics. Restoration Ecology v. 5, p.
109–114. 1997.
ANDERSEN, A. N. et al. Using ants as bioindicators in land management: simplifying
assessment of ant community responses. Journal of Applied Ecology v. 39, p. 8–17. 2002.
Science
ANDREWS, A. Fragmentation of habitat by roads and utility corridors: A Review. Australian
Zoologist v. 26, p. 130-141. 1990.
BOLTON, B. AntCat: An Online Catalog of the Ants of the World. Disponível em:
<http://www.antcat.org>. Acesso em: 18 março 2014.
BOURSAUX-EUDE, C.; GROSS, R. New insights into symbiotic associations between ants and
bacteria. Research in microbiology v. 151, p. 513-519. 2000.
BRIAN, M. V.; BRIAN, A. D. Insolation and ant populations in the west of Scotland.
Transactions of the Royal Entomological Society of London v. 102, p. 303-330. 1951.
BRONSTEIN, J. L. The Contribution of Ant-Plant Protection studies to our understanding of
Mutualism. Biotropica v. 30, p. 150-161. 1998.
CANNICCI, S. et al. Faunal impact on vegetation structure and ecosystem function in mangrove
forests: A review. Aquatic Botany v. 89, p. 186–200. 2008.
CARVALHO, K. S.; VASCONCELOS, H. L. Comunidade de formigas que nidificam em
pequenos galhos da serrapilheira em floresta da Amazônia Central, Brasil. Revista Brasileira de
Entomologia v. 46, p. 115-121. 2002.
CORRÊA, M. M. et al. How leaf-cutting ants impact forests: drastic nest effects on light
environment and plant assemblages. Oecologia v. 162, p. 103–115. 2010.

9
CREIGHTON,W. S. Notes on the habits of Strumigenys. Psyche v. 44, p. 97-109. 1937.
DELABIE, J. H. C. Trophobiosis Between Formicidae and Hemiptera (Sternorrhyncha and
Auchenorrhyncha): an Overview. Neotropical Entomology v. 30, p. 501-516. 2001.
DELABIE, J. H. C. et al. As Formigas como Indicadores Biológicos do Impacto Humano em
Manguezais da Costa Sudeste da Bahia. Neotropical Entomology v. 35, p. 602-615. 2006.
DIAS, N. S. et al. Interação de fragmentos florestais com agroecossistemas adjacentes de café e
pastagem: respostas das comunidades de formigas (Hymenoptera, Formicidae). Iheringia, Série
Zoologia, Porto Alegre, v. 98, p. 136-142. 2008.
D’ETTORRE, P. et al. The role of the symbiotic fungus in the digestive metabolism of two
species of fungus-growing ants. Journal of Comparative Physiology. B v. 172, p. 169-176.
2002.
ELMES, G. W.; THOMAS, J. A. Complexity of species conservation in managed habitats:
interaction between Maculinea butterflies and their ant hosts. Biodiversity and Conservation v.
1, p. 155–169. 1992.
FAHRIG, L. Effects of Habitat Fragmentation on Biodiversity. Annual Reviews in Ecology,
Evolution and Systematics v. 34, p. 487–515. 2003.
FERNÁNDEZ, F. Introducción a las Hormigas de la región Neotropical. Instituto de
Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá, Colombia. 2003. 398 p.
FOWLER, H. G. Diversity estimates: Ant communities and the rare ant species (Hymenoptera:
Formicidae) in fauna of a sub-tropical island. Revista de Matemática e Estatística v. 13, p. 29-
38. 1995.
FORMAN, R. T. T.; ALEXANDER, L. E. Roads and their major ecological effects. Annual
Reviews in Ecology and Systematic v. 29, p. 207–31. 1998.
FLOREN, A.; LINSENMAIR, K. E. The Importance of Primary Tropical Rain Forest For
Species Diversity: An Investigation Using Arboreal Ants as an example. Ecosystems v. 8, p.
559–567. 2005.
GRAHAM, J. H. et al. Habitat disturbance and the diversity and abundance of ants (Formicidae)
in the Southeastern Fall-Line Sandhills. Journal of Insect Science v. 4, p. 30-45. 2004.
HOLWAY, D. A. et al. The Causes and Consequences of Ant Invasions. Annual Review in
Ecology and Systematics v. 33, p. 181–233. 2002.
HOLWAY, D. A. Edge effects of an invasive species across a natural ecological boundary.
Biological Conservation v. 121, p. 561–567. 2005.

10
HÖLLDOLBLER, B.; WILSON, E. O. The Ants. Belknap Press, Cambridge, Massachusetts.
1990. 733 p.
JOLIVET, P. Ants and plants, an example of coevolution. Backhuys Publishers, Leiden. 1996.
303 pp.
IZHAKI , I.; LEVEY, D. J.; SILVA,W. R. Effects of prescribed fire on an ant community in
Florida pine savanna. Ecological Entomology v. 28, p. 439–448. 2003.
MAJER, J. D. Ants: Bio-indicators of minesite rehabilitation, land-use, and land conservation.
Environmental Management v. 7, p. 375-383. 1983.
MCGLYNN, T. P. The worldwide transfer of ants: Geografical distribution and ecological
invasions. Journal of Biogeography v. 23, p. 535-548. 1999.
MOUTINHO, P.; NEPSTAD, D. C.; DAVIDSON, E. A. Influence of Leaf-Cutting Ant Nests on
Secondary Forest Growth Soil Properties in Amazonia. Ecology v. 84, p. 1265-1276. 2005.
MURCIA, C. Edge effects in fragmented forests: implications for conservation. Trends in
Ecology and Evolution v. 10, p. 58-62. 1995.
NASCIMENTO, J. R. O efeito da construção da rodovia PA-458 sobre a macrofauna
bentônica dos manguezais no município de Bragança- Pará. Trabalho de Conclusão de Curso
(Licenciatura em Ciências Biológicas). Universidade Federal do Pará. Campus de Bragança.
Bragança. 2005.
NIELSEN, M. G. Nesting biology of the mangrove mud‐nesting ant Polyrhachis sokoolva Forel
(Hymenoptera: Formicidae) in northern Australia. Insects Sociaux v. 44, p. 14‐21. 1997.
OFFENBERG, J.; MACINTOSH, D. J.; NIELSEN, M. G. Indirect ant-protection against crab
herbivory: damage-induced susceptibility to crab grazing may lead to its reduction on ant-
colonized trees. Functional Ecology v. 20, p. 52–57. 2006a.
OFFENBERG, J.; NIELSEN, M. G.; MACINTOSH, D. J.; AKSORNKOAE, S.; HAVANON, S.
Weaver ants increase premature loss of leaves used for nest construction in Rhizophora trees.
Biotropica v. 38, p. 782–785. 2006b.
OLIVEIRA, E. P.; FRANKLIN, E. Efeito do fogo sobre a mesofauna do solo: recolonização em
áreas queimadas. Pesquisa Agropecuária Brasileira, Brasília, 28 (3): 357-369. 1993.
PEREIRA, A. P. F. G.; ANDRADE, F. A. G.; FERNANDES, M. E. B. Dois anos de
monitoramento dos atropelamentos de mamíferos na rodovia PA-458, Bragança, Pará. Boletim
do Museu Paraense Emílio Goeldi, Ciências Naturais, v. 1, p. 77-83. 2006.
SOTRZ, S. R.; TSCHINKEL, W. R. Distribution, spread, and ecological associations of the
introduced ant Pheidole obscurithorax in the southeastern United States. Journal of Insect
Science v. 4, 12 p. 2004.

11
SUAREZ, A. V.; BOLGER, D. T.; CASE, T. J. Effects of Fragmentation and Invasion on Native
Ant Communities in Coastal Southern California. Ecology v. 79, p. 2041–2056. 1998.
SUAREZ, A. V.; HOLWAY, D. A.; CASE, T. J. Patterns of spread in biological invasions
dominated by long-distance jump dispersal: insights from Argentine ants. Proceedings of the
National Academy of Sciences of the United States of America v. 98, p. 1095–1100. 2001.
SUAREZ, A. V.; HOLWAY, D. A.; WARD, P. S. The role of opportunity in the unintentional
introduction of nonnative ants. Proceedings of the National Academy of Sciences of the
United States of America v. 102, p. 17032–17035. 2005.
SUAREZ, A. V.; TSUTSUI, N. D. The evolutionary consequences of biological invasions.
Molecular Ecology v. 17, p. 351–360. 2008.
TEPE, E. J.; KELLEY, W. A.; RODRIGUEZ-CASTAÑEDA, G.; DYER, L. A. Characterizing
the cauline domatia of two newly discovered Ecuadorian ant plants in Piper: an example of
convergent evolution. Journal of Insect Science v. 9, 9 p. 2009.
TSUTSUI, N. D.; SUAREZ, A. V. The Colony Structure and population biology of Invasive
ants. Conservation Biology v. 17, p. 48–58. 2003.
UNDERWOOD, E. C.; FISHER, B. L. The role of ants in conservation monitoring: If, when, and
how. Biological Conservation v. 132, p. 166 –182. 2006.
URBAS, P. et al. Cutting More from Cut Forests: Edge Effects on Foraging and Herbivory of
Leaf-Cutting Ants in Brazil. Biotropica v. 39, p. 489–495. 2007.
VASCONCELOS, H. L. Resposta das formigas à fragmentação florestal. Série Técnica IPEF,
INPA, v. 12 n. 32, p. 95-98. 1998.
VASCONCELOS, H. L.; VILHENA, J. M. S.; CALIRI, G. J. A. Responses of ants to selective
logging of a central Amazonian forest. Journal of Applied Ecology v. 37, p. 508–514. 2000.
VASCONCELOS, H. L. et al. M. Long-term effects of forest fragmentation on Amazonian ant
communities. Journal of Biogeography v. 33, p. 1348–1356. 2006.
VIANA , V. M.; TABANEZ , A. A. J. Biology and conservation of forest fragments in the
Brazilian Atlantic moist forest. In: Forest patches in tropical landscapes. Washington: Island
Press, 1996. p. 151-167.
WARD, D. F.; HARRIS, R. J. Invasibility of native habitats by Argentine ants, Linepithema
humile, in New Zealand . New Zealand Journal of Ecology v. 29, p. 215-219. 2005.
WATT, A. D.; STORK, N. E.; BOLTON, B. The diversity and abundance of ants in relation to
forest disturbance and plantation establishment in southern Cameroon. Journal of Applied
Ecology v. 39, p. 18–30. 2002.

12
WETTERER, J. K.; O’HARA, B. C. Ants (Hymenoptera: Formicidae) of the Dry Tortugas, the
outermost Florida Keys. Florida Entomologist v. 85, p. 303–307. 2002.
WILKIE, K. T.; MERTL, A. L.; TRANIELLO, J. F. Biodiversity below ground: probing the
subterranean ant fauna of Amazonia. Naturwissenschaften v. 94, p. 725–731. 2007.
WILSON, E. O. Causes of Ecological Success: The Case of the Ants. Journal of Animal
Ecology v. 56, p. 1-9. 1987.
WILSON, E. O.; HÖLLDOLBLER, B. Eusociality: Origin and consequences. PNAS v. 102, p.
13367–13371. 2005.
YOO, H. S .; HOLWAY D. A. Context-dependence in an ant–aphid mutualism: direct effects of
tending intensity on aphid performance. Ecological Entomology v. 36, p. 450–458. 2011.

13
CAPÍTULO 2
EFEITO DE BORDAS NATURAIS E ARTIFICIAIS NA ASSEMBLÉIA DE FORMIGAS
DOS MANGUEZAIS

14
RESUMO
Estudos recentes indicam que a fragmentação de hábitats e a criação de bordas artificiais tem
promovido mudanças na diversidade de formigas, favorecendo a introdução de espécies
sinantrópicas e alterando as relações interespecíficas. Neste cenário, nós examinamos a influência
de bordas artificial e natural, apontando as diferenças na riqueza e na frequência de ocorrência da
assembléia de formigas associada aos manguezais, com ênfase nas espécies sinantrópicas. Nós
amostramos a mirmecofauna ao longo de transecções de 180 m, com intervalos de 30 m, a partir
da borda para dentro do manguezal. Utilizamos dois tipos de armadilhas atrativas, destinadas a
capturar formigas na copa, tronco e superfície do solo, além de coletas manuais. O inventário
registrou 45 espécies, 13 delas comuns às duas áreas. A composição de espécies de formigas foi
similar entre os estratos da floresta, em ambos os tipos de borda. A riqueza não foi influenciada
pelo tipo ou distância da borda, enquanto a frequência de ocorrência de formigas foi
significativamente maior no sítio com borda artificial. Nove espécies de formigas sinantrópicas
foram registradas (20% do total) e representadas regularmente ao longo das transecções,
tornando-se elementos dominantes em todos os estratos florestais amostrados. Nossos resultados
indicam que a fragmentação ocasionada pela construção de rodovia favorece a ocorrência de
espécies de formigas sinantrópicas em áreas de manguezal.
Palavras-chave: Riqueza de espécies; estratificação florestal; gradiente de distância;
fragmentação; zona costeira; Brasil.

15
ABSTRACT
Recent studies indicate that habitat fragmentation and the formation of artificial edges have
promoted changes in ant diversity, favoring the introduction of synanthropic species and altering
interspecific relations. In this scenario, we examined the influence of natural and artificial edges,
pointing out the differences in richness and frequency of occurrence of the assemblage of ants
associated with mangroves, with emphasis on synanthropic species. We sampled the ant fauna
along transects of 180 m, at intervals of 30 m, from the edge into the mangrove. We use two
types of attractive traps designed to capture ants in the canopy, trunk and soil surface, and hand
collecting. The survey recorded 45 species, 13 of them common to both areas, besides manual
collection. The inventory recorded 45 species, 13 of them common to both areas. The
composition of ant species was similar among the strata of mangrove forest at both study sites.
Richness was not influenced by the type of edge or distance from the edge, while the frequency
of occurrence of ants was significantly affected, being recorded higher values at the site with
artificial edge. Nine species of synanthropic ants were recorded (20% of total) and represented
regularly along the transects, becoming dominant elements in all forest strata sampled. Our
results indicate that the fragmentation caused by highway construction influences positively the
occurrence of synanthropic species of ants in mangrove areas.
Key words: Species richness; Forest stratification; distance gradient; fragmentation; coastal zone;
Brazil.

16
INTRODUÇÃO
FORMIGAS (HYMENOPTERA: FORMICIDAE) CONSTITUEM UM GRUPO DE INSETOS COM NOTÁVEL
BIOMASSA NOS DIVERSOS ECOSSISTEMAS TERRESTRES, exercendo funções ecológicas de grande
importância. Servem como agentes de movimentação do solo, redistribuição de nutrientes e
distúrbios em pequena escala (Folgarait 1998). Apresentam plasticidade alimentar e atuam em
uma série de interações com plantas e outros insetos (Bronstein 1998, Philpott & Armbrecht
2006). Seu sucesso ecológico está relacionado ao fato de ser o primeiro grupo predador social a
nidificar e forragear sobre o solo e também sobre a vegetação, ampliando significativamente sua
dominância nestes hábitats (Wilson 1987).
Devido à elevada abundância, ampla distribuição geográfica, facilidade de coleta e
identificação, as formigas tem sido frequentemente utilizadas como bioindicadores do nível de
saúde ambiental, pois são capazes de responder rapidamente às mudanças estressantes no
ambiente (Andersen & Sparling 1997, Lobry de Bruyn 1999). Os distúrbios de origem
antropogênica, tais como o desmatamento visando à expansão da fronteira agrícola e a
desenfreada construção de rodovias em hábitats nativos, promovem mudanças na estrutura e
complexidade da paisagem, alteram as relações interespecíficas, além de facilitar a dispersão de
espécies nos hábitats perturbados (Fagan et al. 1999). Espécies cuja ocorrência e dispersão estão
necessariamente sujeitas à atuação humana são chamadas de espécies sinantrópicas, e uma vez
introduzidas, tais espécies podem exercer forte impacto sobre os outros componentes da biota.
Diversos estudos correlacionam positivamente o nível de integridade ambiental com a
diversidade de espécies de formigas, enfatizando que os eventos envolvidos na fragmentação de
uma paisagem são responsáveis por promover a perda e/ou substituição da diversidade nativa
(Dunn 2004, Silva et al. 2007). A presença de rodovias é uma característica essencial ao
deslocamento humano, atuando ecologicamente como uma barreira ao movimento de algumas
espécies, enquanto facilita a dispersão de outras, sendo apontada como a principal causa do
aumento da suscetibilidade dos hábitats ao fenômeno da invasão biológica (Pimm & Raven 2000,
Trombulak & Frissell 2000, Holway & Suarez 2006).
Em florestas de terra firme, por exemplo, diversos trabalhos relataram elevada densidade
de organismos exóticos e invasores associada às margens de rodovias, tais como formigas-de-
fogo (Forys et al. 2002) e plantas de sub-bosque (Watkins et al. 2003), revelando que as
diferenças na composição e na abundância destes grupos são consequência da criação de feições

17
antrópicas em hábitats naturais. Outros grupos de formigas, como as de correição, por exemplo,
tendem a evitar as bordas, podendo até desaparecer se o fragmento remanescente for muito
pequeno (Zheng & Chen 2000).
A influência de rodovias tem aumentado também em florestas alagáveis, como os
manguezais, intensificando a sobre- exploração dos seus recursos naturais e a diminuição de sua
área em função de atividades antrópicas (Luther & Greenberg 2009). Considerando a importância
ecológica das formigas e sua aplicabilidade em estudos de bioindicação, o presente estudo
investigou o efeito do grau de perturbação sobre a assembléia de formigas associada ao
manguezal, relacionado à influência de uma rodovia (borda criada artificialmente), que corta uma
floresta de mangue. Inicialmente, nós examinamos a composição da mirmecofauna ao longo do
estrato vertical da floresta de mangue, no intuito de descrever a distribuição espacial de formigas
nos microhábitats disponíveis (copa, tronco, raízes e superfície do solo), e identificar as espécies
mais importantes em cada estrato florestal. Posteriormente, testamos as seguintes hipóteses: i)
devido à instabilidade do solo no manguezal, é na copa das árvores que se concentra a maior
riqueza de espécies e frequência de ocorrência de formigas, ii) a assembléia de formigas
apresenta maior riqueza e frequência de ocorrência em borda criada artificialmente do que em
borda natural e iii) os valores de riqueza e frequência de ocorrência de formigas são mais altos
em direção à borda do que no interior da floresta. Ao mesmo tempo, nós testamos essas hipóteses
considerando as espécies de formigas sinantrópicas.
MÉTODOS
ÁREA DE ESTUDO.− O presente estudo foi realizado na Reserva Extrativista Marinha Caeté-
Taperaçu, localizada na cidade de Bragança, PA, Brasil (46°50’46°30’W e 0°45’1°07’S) (Fig. 1).
As áreas de coleta estão inseridas na península de Ajuruteua, cuja diversidade de ambientes inclui
as florestas de mangue, as quais dominam a maior parte da cobertura vegetal. A paisagem dos
manguezais dessa região é composta por quatro espécies arbóreas: Rhizophora mangle L.,
Avicennia germinans (L.) Stearn, A. schaueriana Stapf e Leechman ex Moldenke e Laguncularia
racemosa (L.) Gaertn. f. (Menezes et al. 2008).
As coletas foram conduzidas no período de abril a dezembro de 2012, abrangendo a
transição dos períodos pluviométricos da região. O clima da região é quente e úmido, a

18
temperatura média do ar no período estudado foi de 27°C (variação anual de 23 a 32°C). A
umidade relativa do ar alcançou cerca de 85% (variação anual de 77 a 91%). A região apresenta
um período chuvoso (>200 mm) de janeiro a junho, com média anual de precipitação em torno de
2597.2 mm (Instituto de Meteorologia, Estação Meteorológica de Tracuateua, 30 km sudoeste da
área de estudo).
O extenso ecossistema de manguezal existente na região sofreu um processo de
fragmentação, decorrente da construção da rodovia estadual PA-458, no período de 1974-1983,
com o objetivo de ligar a cidade de Bragança à praia de Ajuruteua para facilitar o turismo e
escoamento da produção pesqueira local (Lara et al. 2002). A rodovia tem 36 km de extensão,
dos quais 20 km cruzam as florestas de mangue, onde foram produzidos impactos negativos, tais
como a obstrução de canais-de-maré, mudanças na estrutura da vegetação e na diversidade da
fauna bentônica (Beasley et al. 2010), além de propiciar eventos constantes de atropelamento da
fauna associada ás florestas de mangue (Pereira et al. 2006). A criação da rodovia também
impulsionou a ocupação desordenada na praia de Ajuruteua, modificando a dinâmica das dunas
costeiras, e ocasionando problemas de caráter socioambiental (Pereira et al. 2007).
Para a realização das coletas selecionamos dois sítios de trabalho localizados em área de
manguezal, os quais apresentam diferentes tipos de bordas. O Sítio 1 (00°59’42’’S,
046°43’20’’W) localiza-se às margens da rodovia PA-458 (borda artificial) e foi escolhido por
sua proximidade com as florestas de terra firme, com as áreas urbana e rural de Bragança (< 20
km), consideradas fontes de espécies sinantrópicas, e também com uma área de manguezal que
foi degradada pela construção da rodovia e atualmente se encontra em processo de regeneração
(Fig. 1A). Neste sítio, observa-se intensa exploração do caranguejo-uçá (Ucides cordatus), além
da retirada seletiva de espécies arbóreas para produção de carvão e artefatos de pesca (Glaser
2003). O Sítio 2 (00°83’55°S, 046°63’83’’W) está localizado às margens de um canal-de-maré,
conhecido localmente por Furo Grande, e foi selecionado para ilustrar as condições da fauna de
formigas associada à presença de borda natural (Fig. 1B). Esta área de manguezal está localizada
ao norte da península de Ajuruteua e sofre inundação diária, de acordo com o regime semidiurno
da maré. A fim de minimizar a influência da rodovia (borda artificial) neste sítio, as amostras
foram tomadas a partir de 1 km da rodovia. Em ambos os sítios predominam as espécies arbóreas
A. germinans e R. mangle.

19
FIGURA 1: Mapa da área de estudo, destacando os sítios de trabalho na península de
Ajuruteua, Pará, Brasil: Sítio 1– bordas artificiais (A) e Sítio 2 – bordas naturais (B).
METODOLOGIA DE COLETA. –No intuito de examinar os efeitos gerados por diferentes tipos
de borda sobre a composição da fauna de formigas, sorteamos em cada sítio de trabalho uma
transecção, no sentido da borda para o interior da floresta de mangue. Em cada transecção,
demarcamos sete pontos amostrais, a cada intervalo de 30 m, partindo de uma distância de 0 até
180 m no interior do manguezal. No total, foram feitas quatro campanhas de coleta, em intervalo
trimestral.
Em cada ponto amostral, selecionamos aleatoriamente uma árvore para a disposição de
armadilhas atrativas de queda, destinadas a analisar a estratificação vertical da fauna de formigas,
considerando os estratos da copa e o tronco das árvores de mangue. Foram instaladas duas
armadilhas na copa, enquanto outras duas armadilhas foram atadas ao tronco da árvore, a uma
altura de no máximo dois metros do solo. Estas armadilhas foram descritas por Oliveira Santos et
al. (2009), e funcionalmente atuam atraindo formigas até um copo descartável que contém isca

20
atrativa, e está fixado ao fundo de um recipiente PET (10x10 cm) contendo uma solução
conservante de álcool com algumas gotas de detergente. Usamos dois tipos de isca atrativa por
estrato, sendo uma de base proteica (sardinha macerada em óleo vegetal) e a outra rica em
carboidratos (solução saturada de açúcar), buscando melhor representar a diversidade de formigas
através da variedade de recursos alimentares oferecidos. Os espécimes que caíram nas armadilhas
foram coletados 48 horas após a instalação, sendo devidamente acondicionados em sacos
plásticos etiquetados. Cada armadilha foi contabilizada como uma unidade amostral, totalizando
224 unidades amostrais para este método (112 para cada sítio de coleta).
Sorteamos outras quatro transecções, contendo também sete pontos amostrais a cada
intervalo de 30 m, onde foi aplicado o método de armadilhas atrativas de captura manual. Estas
armadilhas consistem de copos plásticos, com capacidade de 30 ml, com um suporte de metal
para fixar a armadilha na árvore. Em cada ponto amostral colocamos quatro armadilhas por
árvore, sendo as duas primeiras fixadas em fissuras, galhos, ou ainda sobre as raízes-escora de R.
mangle, enquanto as outras duas foram dispostas próximo ao solo. No interior destas armadilhas
continha iscas atrativas de sardinha e iscas atrativas açucaradas. Os espécimes atraídos para as
iscas foram coletados 01 30 h após a disposição, e acondicionados em sacos contendo acetato de
etila. Este método também totalizou 224 unidades amostrais (112 em cada sítio de coleta).
Realizamos também busca ativa em troncos caídos, no solo, ou onde houvesse indícios de
nidificação de formigas, sendo que dados provenientes deste método de coleta foram utilizados
apenas de modo a complementar o inventário da mirmecofauna associada aos manguezais da área
de estudo.
Os espécimes coletados foram acondicionados em frascos com álcool 70% e
transportados para a Coleção de Zoologia de Bragança (CZB), na Universidade Federal do Pará
(UFPA), Campus Bragança, onde foram conservados para triagem e identificação. Utilizamos a
chave taxonômica para subfamílias e gêneros de formigas, proposta por Fernández e Sharkey
(2003), bem como 57% das espécies coletadas foram identificadas ao nível de espécie, com ajuda
de taxonomista, no Museu Paraense Emílio Goeldi, em Belém, PA.
ANÁLISE DOS DADOS.− Nós caracterizamos inicialmente a estrutura da assembléia de
formigas associada ao manguezal utilizando os parâmetros ecológicos de riqueza de espécies e
frequência de ocorrência (número de registros). A média destes valores foi posteriormente

21
comparada entre os sítios de coleta, por meio de Teste t. Para verificar a eficiência amostral em
acessar a riqueza total da assembléia de formigas, projetamos curvas de acumulação de espécies
(Curva do coletor) por cada tipo de método amostral padronizado, excluindo, portanto, os dados
da busca ativa.
Investigamos possíveis diferenças na composição da assembléia de formigas através de
Análises de Similaridade (ANOSIM), considerando os fatores 1) estratos da floresta de mangue,
2) sítios de trabalho e 3) distância da borda. Estas análises foram baseadas em uma matriz de
similaridade entre as 448 unidades amostrais, calculada com base no índice de similaridade de
Bray Curtis, o qual é menos afetado pelo número de espécies raras nas amostras (Clarke &
Warwick 2001). Para a construção da matriz, usamos dados de presença e ausência das espécies,
de forma a evitar resultados superestimados, os quais são gerados quando se usa os dados de
abundância absoluta.
Utilizamos a Análise de Percentagem de Similaridade (SIMPER) para calcular o
percentual de contribuição de cada espécie, e indicar quais espécies mais contribuíram para a
diferença observada na assembléia de formigas. ANOSIM e SIMPER foram conduzidas
utilizando o pacote estatístico Primer versão 6.0 (Clarke & Gorley 2006), sendo considerados
significativos os resultados com P< 0,05.
Diferenças nas médias dos parâmetros ecológicos de riqueza e frequência de ocorrência
de formigas ao longo das transecções foram avaliadas por meio de Análises de Variância
(ANOVA- um fator), com teste post-hoc de Tukey. Representamos esquematicamente a
frequência das espécies ao longo das transecções como círculos, sendo o tamanho do círculo
proporcional à frequência da espécie. Desta forma, buscamos evidenciar possíveis gradientes de
distribuição das espécies no sentido borda-centro do manguezal, o que pode sugerir a ocorrência
de efeitos de borda.
Nós também avaliamos os resultados através da Análise de Correspondência (CA),
utilizando a frequência relativa das espécies em cada estrato arbóreo, com o intuito de representar
de forma esquemática a importância da espécie nos diferentes microhábitats disponíveis em cada
sítio de trabalho, enfatizando a ocorrência das formigas sinantrópicas. As Análises de Variância
(ANOVA) e Análises de Correspondência foram conduzidas no pacote estatístico STATISTICA
versão 8.0 (StatSoft 2001).

22
RESULTADOS
ASSEMBLÉIA DE FORMIGAS DE MANGUEZAL._Foi coletado um total de 2784 espécimes,
obtidos em 292 registros. A Tabela 1 apresenta o computo geral de espécies de formigas
registradas para a área de manguezal estudada, sendo 45 espécies, distribuídas em 16 gêneros e
cinco subfamílias. Quatro táxons foram os mais frequentes neste estudo: Camponotus brettesi
(106 registros), Azteca chartifex (50 registros), Camponotus atriceps (42 registros), Camponotus
sexguttatus (21 registros), representando 75% do total dos registros (Tabela 1). Nove espécies
com identificação já confirmada são consideradas sinantrópicas (20% do total de espécies)
(Tabela 1). Do total de espécies de formigas, 14 foram coletadas exclusivamente através de busca
ativa e, portanto, as análises foram executadas considerando as 31 espécies restantes.
De uma forma geral, a subfamília Myrmicinae foi a mais representativa quanto à riqueza
de espécies (n=23), seguida por Formicinae (n=11), Dolichoderinae (n=5), Pseudomyrmicinae
(n=4) e Ponerinae (n=2), sendo esta última amostrada apenas através de busca ativa.

23
TABELA 1. Frequência das espécies de formigas registradas em dois sítios de manguezal ao longo da península de Ajuruteua, Pará,
Brasil. A=Armadilha atrativa suspensa, B=Armadilha atrativa de captura manual e C=Coleta manual.
Subfamília Sítio 1 Sítio 2
Espécie Característica A
(n=112)
B
(n=112) C
A
(n=112)
B
(n=112) C
Formicinae
Camponotus brettesi Forel,
1899
Arborícola, Comum em manguezais 42 22 20 23 19 22
Camponotus atriceps (F.
Smith, 1858)
Arborícola, Sinantrópica 32 1 1 8 1 2
Camponotus cingulatus Mayr,
1862
Arborícola, Sinantrópica - - - 2 - -
Camponotus sexguttatus
(Fabricius, 1793)
Arborícola, Sinantrópica 3 1 5 11 6 3
Camponotus sp.1 Arborícola - - - 2 - -
Camponotus sp.2 Arborícola - - - 1 - -
Camponotus sp.3 Arborícola - - - 4 - -
Camponotus sp.4 Arborícola - - 1 - - -
Paratrechina sp.1 Nidifica oportunisticamente no solo, liteira e
madeira apodrecida - - - 4 - -

24
Paratrechina sp.2 Nidifica oportunisticamente no solo, liteira e
madeira apodrecida 1 2 2 - - -
Paratrechina sp.3 Nidifica oportunisticamente no solo, liteira e
madeira apodrecida - - 1 - - -
Myrmicinae
Wasmannia lutzi Forel, 1908 Generalista quanto à nidificação - - 1 1 - 1
Pheidole mamore Mann, 1916 Nidifica em madeira apodrecida, é típica de
florestas de terra firme e planícies alagáveis ‘ - 4 - - -
Pheidole gigas Wilson, 2003 Arborícola 1 2 - - - -
Pheidole grupo fallax Arborícola 1 - - - - -
Pheidole nr angusta Arborícola - - - - - 1
Pheidole nr. machutela Generalista quanto à nidificação - - - 1 - 1
Pheidole sp.1 Arborícola - - 2 - - -
Pheidole sp.2 Arborícola 2 3 6 - - -
Pheidole sp.3 Arborícola - - - 2 - -
Nesomyrmex sp.1 Arborícola 1 - - - - -
Nesomyrmex sp.2 Arborícola - - - - - 2
Solenopsis pusillignis Trager,
1991
Generalista quanto à nidificação - - 5 - - 9
Solenopsis sp.1 Generalista quanto à nidificação - - - 3 - -
Solenopsis sp.2 Generalista quanto à nidificação - - - 1 - -
Solenopsis sp.3 Generalista quanto à nidificação 2 - - - - -

25
Solenopsis sp.4 Generalista quanto à nidificação - - 1 - - -
Solenopsis sp.5 Generalista quanto à nidificação - - 1 - - -
Crematogaster erecta Mayr,
1866
Arborícola, Sinantrópica - - - 6 2 16
Cephalotes cordiventris
(Santschi, 1931)
Arborícola - 1 8 - - 6
Cephalotes atratus (Linnaeus,
1758)
Arborícola 4 1 8 - - -
Cephalotes pallens (Klug,
1804)
Arborícola, comum em vegetação de capoeira
e bordas de hábitats. 1 - - - - -
Cephalotes maculatus (F.
Smith, 1876)
Arborícola, Sinantrópica - - 1 - - -
Strumigenys elongata Roger,
1863
Generalista quanto à nidificação - - 1 - - -
Dolichoderinae
Azteca chartifex Emery, 1896 Arborícola, típica de florestas úmidas e
vegetação na borda de hábitats 24 23 24 3 - 8
Linepithema sp.1 Generalista quanto à nidificação - - 1 1 - 1
Linepithema sp.2 Generalista quanto à nidificação - - - - - 1
Dolichoderus lutosus F.
Smith, 1858
Generalista quanto à nidificação,
Sinantrópica 3 - 2 5 - 1
Tapinoma melanocephalum Generalista quanto à nidificação, 1 2 - - - -
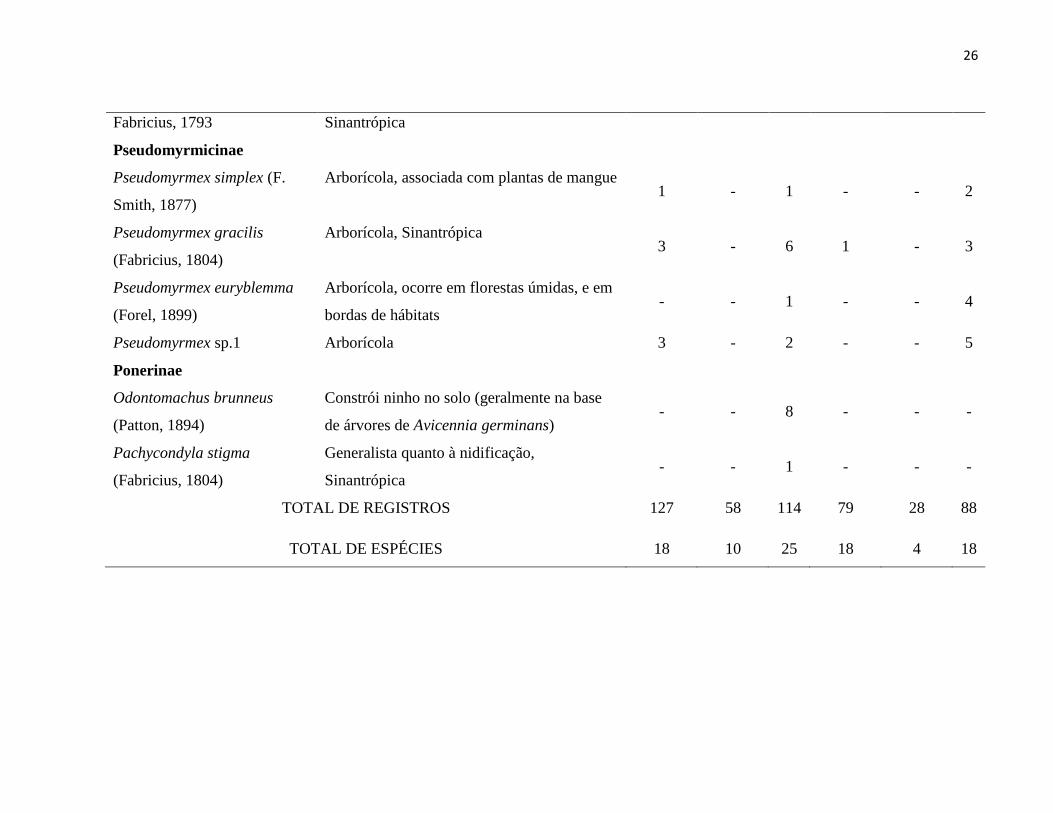
26
Fabricius, 1793 Sinantrópica
Pseudomyrmicinae
Pseudomyrmex simplex (F.
Smith, 1877)
Arborícola, associada com plantas de mangue 1 - 1 - - 2
Pseudomyrmex gracilis
(Fabricius, 1804)
Arborícola, Sinantrópica 3 - 6 1 - 3
Pseudomyrmex euryblemma
(Forel, 1899)
Arborícola, ocorre em florestas úmidas, e em
bordas de hábitats - - 1 - - 4
Pseudomyrmex sp.1 Arborícola 3 - 2 - - 5
Ponerinae
Odontomachus brunneus
(Patton, 1894)
Constrói ninho no solo (geralmente na base
de árvores de Avicennia germinans) - - 8 - - -
Pachycondyla stigma
(Fabricius, 1804)
Generalista quanto à nidificação,
Sinantrópica - - 1 - - -
TOTAL DE REGISTROS 127 58 114 79 28 88
TOTAL DE ESPÉCIES 18 10 25 18 4 18

27
O maior tempo de exposição das armadilhas atrativas suspensas influenciou positivamente
na riqueza de espécies capturada por este método, em ambos os sítios. O número acumulativo de
espécies sofreu incremento com a realização de novas coletas, demonstrando que a diversidade
de formigas nestes manguezais é elevada e, pode revelar-se ainda maior, se ampliarmos a
dimensão da área sob estudo e o número de métodos amostrais empregados.
FIGURA 2. Curva de Acumulação de espécies, considerando o número de espécies
de formigas capturadas nas 112 unidades amostrais contabilizadas para cada método
de coleta, em cada sítio de manguezal, na península de Ajuruteua, Pará, Brasil.
DISTRIBUIÇÃO VERTICAL DE FORMIGAS._ Aproximadamente 58% das espécies de
formigas identificadas apresentaram hábito arborícola, 11% nidificaram em madeira apodrecida,
na liteira ou diretamente no solo e 31% foram classificadas como generalistas quanto à
nidificação. As espécies com maior amplitude de distribuição vertical no Sítio 1 foram C. brettesi
e A. chartifex, enquanto no Sítio 2, C. brettesi e C. sexguttatus foram as únicas espécies que
ocorreram em todos os estratos arbóreos.
A análise da estratificação vertical da fauna de formigas, baseada apenas no método das
armadilhas atrativas suspensas, revelou que em ambos os sítios de trabalho a riqueza e a

28
frequência de ocorrência de formigas foram maiores no tronco do que na copa das árvores de
mangue (Tabela 2). Ao comparar estes resultados com aqueles obtidos pelo método das
armadilhas atrativas de captura manual, observamos que a riqueza de espécies foi baixa em
ambos os sítios, sendo que no Sítio 1 a frequência de formigas foi maior nas amostras tomadas na
superfície do solo (n= 36), enquanto no Sítio 2 este parâmetro foi maior no tronco das árvores de
mangue (n=21, Tabela 2).
A composição de formigas ao longo dos estratos da floresta de mangue foi similar entre
os sítios (ANOSIM, R= 0,022, P= 0,001). No Sítio 1, a superfície do solo e o tronco
apresentaram mais espécies em comum (n=8), enquanto que no Sítio 2 a copa e o tronco
compartilham mais espécies (n=8). De acordo com a rotina SIMPER, três táxons foram os mais
representativos nos estratos arbóreos do Sítio 1 (C. brettesi, C. atriceps, A. chartifex), enquanto
no Sítio 2, quatro espécies predominaram nas amostras (C. brettesi, C. sexguttatus, C. atriceps e
Crematogaster erecta (Tabela 2).
A Análise de Correspondência confirmou a importância numérica das espécies
sinantrópicas em todos os estratos arbóreos, sendo que no Sítio 1 as espécies D. lutosus e P.
gracilis apresentam elevada frequência na copa das árvores, enquanto C. atriceps, C sexguttatus,
Paratrechina sp.2 e T. melanocephalum foram frequentes nos estratos arbóreos inferiores (Fig.
3A). Quando analisamos o Sítio 2, D. lutosus, C. atriceps, e Paratrechina sp.1 foram as
principais espécies registradas na copa, ao passo que C. cingulatus, P. gracilis, Crematogaster
erecta e C. sexguttatus as espécies mais frequentes no tronco das árvores deste sítio (Fig. 3B).

29
TABELA 2. Valores para os parâmetros ecológicos da mirmecofauna em cada estrato da floresta de mangue, mostrando também os
resultados das análises de similaridade (ANOSIM, R= valor do teste de similaridade e P= nível de significância), estrato de acordo com
o método amostral utilizado (A= Armadilha atrativa suspensa e B= Armadilha atrativa de captura manual), na península de Ajuruteua,
Pará, Brasil.
Sítio Método Estrato Principais espécies Riqueza Frequência ANOSIM
R P
Km 15
A
Tronco Camponotus brettesi, Camponotus atriceps, Azteca chartifex,
Pheidole mamore 14 69
0.009 0.7
Copa C. brettesi, C. atriceps, A. chartifex, Pseudomyrmex sp.1,
Dolichoderus lutosus, Pseudomyrmex gracilis 11 58
B
Solo C. brettesi, A. chartifex, Pheidole sp.2, Pheidole gigas 9 36
0.002 0.32
Tronco A. chartifex, C. brettesi 4 22
Furo
Grande
A
Tronco C. brettesi, C. sexguttatus, Crematogaster erecta, Camponotus
atriceps 15 45
0.003 0.22
Copa C. brettesi, C. atriceps, Paratrechina sp.1, D. lutosus 11 34
B Solo C. brettesi, C. sexguttatus 2 7
0.032 0.005
Tronco C. brettesi, C. sexguttatus, Crematogaster erecta 4 21

30
ASI
ASS
AMI
AMS
C b
C a
C sPa 2
P g
P f
P m
P 2
N 1
S 3
C c
C a
C p
A c
D l
T m
Ps s
Ps g
Ps 1
-1.5 -1.0 -0.5 0.0 0.5 1.0 1.5
Eixo 1
-1.0
-0.8
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4E
ixo
2
ASI
ASS
AMI
AMS
C b
C a
C sPa 2
P g
P f
P m
P 2
N 1
S 3
C c
C a
C p
A c
D l
T m
Ps s
Ps g
Ps 1
A
ASI
ASS
AMI
AMS
C b C a
C c
C s
C 1
C 2
C 3
Pa 1
W l
P ma
P 3
S 1
S 2
C e
A c
L 1
D l
Ps g
-1.0 -0.8 -0.6 -0.4 -0.2 0.0 0.2 0.4 0.6 0.8 1.0 1.2
Eixo 1
-2.0
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0
Eix
o 2
ASI
ASS
AMI
AMS
C b C a
C c
C s
C 1
C 2
C 3
Pa 1
W l
P ma
P 3
S 1
S 2
C e
A c
L 1
D l
Ps g
B
FIGURA 3. Análises de Correspondência baseadas na frequência de ocorrência das espécies de formigas em relação aos estratos
arbóreos, considerando os dois tipos de bordas (borda artificial - Sítio 1, rodovia PA-458 (A) e borda natural – Sítio 2, canal-de-
maré (B), na península de Ajuruteua, PA, Brasil. Métodos (ASS= Armadilha Suspensa Superior; ASI= Armadilha Suspensa
Inferior; AMS= Armadilha Manual Superior; AMI= Armadilha Manual Inferior). Espécies de formigas (Camponotus brettesi= C b;
C. atriceps= C a; C. sexguttatus= C s; C. cingulatus= C c; C. sp.1= C 1; C. sp.2= C 2; C. sp.3= C 3; Paratrechina sp.1= Pa 1; P.
sp.2= Pa 2; Pheidole gigas= P g; P. grupo falax= P f; P. mamore= P m; P. nr. machutela= P ma; P. sp.2= P 2; P. sp.3= P 3;
Nesomyrmex sp.1= N 1; Solenopsis sp.1= S 1; S. sp.2 = S 2; S. sp.3= S 3; Crematogaster erecta= C e; Cephalotes cordiventris=
Ce c; C. atratus= Ce a; C. pallens= Ce p; Azteca chartifex= A c; Linepithema sp.1= L 1; Dolichoderus lutosus= D l; Tapinoma
melanocephalum= T m; Pseudomyrmex simplex= Ps s; P.gracilis= Ps g; P. sp.1= Ps 1).

31
COMPARAÇÃO ENTRE OS SÍTIOS._ A riqueza de espécies foi similar entre os sítios de
coleta (Sítio 1= 19 spp., Sítio 2= 18 spp.). Por outro lado, o Sítio 1 apresentou valores
significativamente maiores de frequência de ocorrência (t=2.68, gl=12, P= 0.01), sendo
obtidos 185 registros de formigas no Sítio 1 (2103 indivíduos) e 107 registros no Sítio 2 (681
indivíduos).
A coleta manual acrescentou 13 táxons à lista de espécies do Sítio 1 e 8 táxons ao Sítio
2. Os dois sítios compartilharam treze espécies, que estão representadas em quatro
subfamílias. Das 45 espécies inventariadas, 19 foram registradas apenas no Sítio 1, enquanto
13 ocorreram exclusivamente no Sítio 2. Das nove espécies sinantrópicas registradas neste
estudo, quatro ocorreram em ambos os sítios de coleta, três exclusivamente no Sítio 1 e duas
apenas no Sítio 2.
Assim como a riqueza, a composição de espécies de formigas também foi similar entre
os sítios de coleta (ANOSIM, R= 0,036, P = 0,001), apresentando-se mais heterogênea nas
amostras do Sítio 1, onde a espécie mais frequente foi Camponotus brettesi (60% dos
registros), seguida por Azteca chartifex (29%) e Camponotus atriceps (10%). Já no Sítio 2, C.
brettesi também é a espécie mais representativa, contribuindo com 78% do total de registros,
seguida por Camponotus sexguttatus (15%).
VARIAÇÃO NA ASSEMBLÉIA DE FORMIGAS NO SENTIDO BORDA-CENTRO DO
MANGUEZAL._ A riqueza de espécies de formigas não variou significativamente entre os
pontos amostrais em nenhum dos sítios estudados (ANOVA, F= 1.14, gl=1, P= 0.3), sendo o
valor máximo registrado apartir dos 120 m de distância da borda em ambos os sítios de coleta
(Fig. 4). Diferente da riqueza, a frequência de ocorrência de espécies apresentou variação
significativa ao longo das transecções, sendo geralmente maior quanto mais próximo à borda
(ANOVA, F=7.23, gl= 1, P=0.01) (Fig. 4).

32
FIGURA 4. Análise comparativa dos parâmetros ecológicos (riqueza e frequência de
ocorrência) da fauna de formigas em diferentes pontos amostrais, dispostos no sentido borda-
centro do manguezal, na península de Ajuruteua, Pará, Brasil.
A composição de espécies apresentou pequena variação entre os pontos amostrais
(ANOSIM, Sítio 1: R= 0.018, P= 0.02 e Sítio 2: R= 0.012, P= 0.02), sendo que as espécies A.
chartifex, C. brettesi e C. atriceps ocorreram ao longo de todas as transecções no Sítio 1,
enquanto no Sítio 2 C. brettesi, C. sexguttatus e C. atriceps foram as espécies mais
frequentemente registradas nas transecções, nesta ordem de importância.
No Sítio 1, algumas espécies tiveram sua ocorrência restrita até os 60 m de distância
da rodovia (Pheidole sp.2, Nesomyrmex sp.1, Solenopsis sp.3 e Pseudomyrmex simplex),
enquanto outras ocorreram a no mínimo 90 m da rodovia (Pheidole gigas, P. gr. fallax,
Pseudomyrmex sp.1, Cephalotes cordiventris, C. atratus e C. pallens) (Fig. 5A). No Sítio 2,
três espécies foram registradas somente nas amostras tomadas na borda do canal de maré
(Pheidole nr. machutela, Solenopsis sp.2 e Linepithema sp.1), enquanto outras quatro
espécies foram registros exclusivos para o interior florestal (Camponotus sp.1, C. sp.2,
Paratrechina sp.1, Solenopsis sp.1) (Fig. 5B). A representação esquemática da distribuição de
formigas ao longo das transecções nos permite ainda inferir que muitas das espécies
sinantrópicas se distribuem desde a borda até o interior da floresta de mangue, levando em
consideração também o fato de que as amostras do Sítio 2 foram tomadas no mínimo a um
quilômetro da rodovia.
0
2
4
6
8
10
12
Riq
ueza
de
esp
écie
s
0 30 60 90 120 150 180
Sítio 1
Sítio 2
Distância da borda
0
5
10
15
20
25
30
35
40
Fre
quência
de
ocorr
ência
0 30 60 90 120 150 180
Distância da borda
Sítio 1
Sítio 2

33
FIGURA 5. Representação esquemática da frequência de ocorrência das
espécies de formigas ao longo das transecções amostradas no Sítio 1 – bordas
artificiais (A) e Sítio 2 – bordas naturais (B), na península de Ajuruteua, Pará,
Brasil.

34
DISCUSSÃO
Os resultados do presente estudo mostram que a riqueza da mirmecofauna dos
manguezais da península de Ajuruteua, em Bragança (n=45), está dentro do espectro de
variação dos valores já registrados para os outros manguezais ao redor do mundo, como é o
caso das florestas de mangue do México com o registro de 37 espécies de formigas (Dejean et
al. 2003) e da Austrália com 64 táxons (Dangerfield et al. 2003). No caso dos manguezais
brasileiros, 22 espécies de formigas foram registradas na região sul, em Santa Catarina
(Cortes–Lopes et al. 1996), ao passo que no nordeste, 64 espécies foram associadas aos
manguezais da Bahia (Delabie et al. 2006). Tais variações nos valores de riqueza certamente
são provenientes do viés exercido pelas variações nos métodos de amostragem populacional,
no esforço amostral empregado, bem como no direcionamento da amostragem para os
diferentes hábitats existentes nos manguezais.
Considerando os valores de riqueza da mirmecofauna para as florestas de terra firme,
segundo a compilação de Agosti et al. (2000), o número de espécies de formigas encontrado
somente em amostragens no solo variou de 106 (Amazonas, Brasil) a 471 espécies
(Madagascar, África), o que representa em torno de duas e dez vezes, respectivamente, a
riqueza da fauna de formigas encontrada nos manguezais do presente estudo (n=45). Por outro
lado, no caso de estudos que amostraram apenas a copa das árvores os valores variaram desde
21 espécies, para a Costa Rica (Longino & Nadkarni 1990) até 52 táxons para o estudo
realizado no Amazonas, Brasil (Harada & Adis 1998). Estes valores de riqueza são maiores
do que aqueles encontrados nas copas dos manguezais do presente estudo (n=18), onde, em
geral, a menor complexidade dos bosques de mangue, que é evidenciada através do número
de estratos da floresta e da composição das espécies vegetais, proporciona menos
microhábitats para nidificação e forrageio de formigas.
É notório que a diversidade de formigas nos manguezais tende a ser muito menor do
que nas florestas de terra firme. Isto certamente decorre da baixa disponibilidade para o uso
do solo nos manguezais por esse grupo de insetos, devido às inundações diárias pelas marés
(Cannicci et al. 2008). Ao contrário, nas florestas de terra firme, o solo possui
comparativamente um número de espécies de formigas aproximadamente oito vezes maior do
que àquele registrado na copa das suas árvores, considerando os estudos de Agosti et al.
(2000) para o solo, e de Longino e Nadkarni (1990) e Harada e Adis (1998) para a copa das
árvores. Além do mais, é importante ressaltar que no presente estudo o solo da área de
manguezal próximo à rodovia parece mais favorável, tanto para a nidificação quanto para o

35
forrageio de formigas, haja vista que esse sítio é inundado apenas durante as marés de sizígia.
Tais condições são diferentes daquelas na área próximo ao canal-de-maré, onde o solo é
diariamente inundado pela maré, dificultando a nidificação e limitando o tempo de forrageio
das formigas sobre o solo. Essa hipótese é corroborada pelo fato de que o sítio próximo à
rodovia apresenta aproximadamente cinco vezes mais espécies no solo do que àquele próximo
ao canal-de-maré (Tabela 2).
Adicionalmente, o menor número de espécies que compõe a mirmecofauna registrada
na copa das árvores dos manguezais pode ser reflexo da prevalência de espécies consideradas
dominantes, pertencentes aos gêneros Camponotus e Azteca. Espécies destes dois gêneros são
reconhecidas em outros trabalhos por seu comportamento altamente agressivo e territorial,
controlando desta forma os recursos disponíveis, bem como a riqueza de espécies
consideradas subordinadas (Adams 1994, Parr 2008). De fato, a defesa da copa das árvores
pelas formigas dominantes é maximizada principalmente em locais estratégicos, como a base
do fuste, impedindo ou limitando o acesso de outras espécies de formigas (Hölldobler &
Wilson 1990). Da mesma forma, a similaridade na composição de espécies entre a copa e o
tronco é também um exemplo da dominância destas espécies, já que ao descer para forragear
sobre o tronco, as formigas frequentemente reforçam seus sinais químicos de defesa
territorial. Contudo, a fauna arborícola de formigas pode ainda sofrer homogeneização como
resultado de distúrbios antropogênicos, resultando em mudanças na riqueza e na dinâmica das
relações entre espécies e, consequentemente, na promoção de efeitos diretos sobre a dinâmica
do sistema como um todo (Floren & Linsenmair 2001).
A riqueza e a composição da fauna de formigas podem ser diretamente afetadas
dependendo da metodologia aplicada para o seu inventário (Gotelli et al. 2011). A maior
eficiência das armadilhas atrativas suspensas em determinar os parâmetros ecológicos da
mirmecofauna, por exemplo, deve-se certamente ao princípio funcional do método, uma vez
que os espécimes atraídos caem na solução conservante, limitando o recrutamento de mais
operárias para aquele recurso alimentar, e permitindo inclusive a aproximação de outras
espécies de formigas (Oliveira-Santos et al. 2009). Por outro lado, alguns fatores relativos às
armadilhas atrativas de captura manual, como o recrutamento em massa para as iscas, o curto
tempo de exposição em campo e os diferentes períodos de forrageio entre as espécies de
formigas podem subestimar os valores de riqueza de espécies, sendo a utilização deste método
justificada por sua rapidez e praticidade (King & Porter 2005).
Nossos achados mostraram que o tipo de borda não influenciou na riqueza de espécies
de formigas registradas para o manguezal como um todo, mas interferiu na frequência destas

36
espécies, refutando a segunda hipótese proposta de que tanto a riqueza quanto a frequência de
espécies seriam maiores no sítio próximo à rodovia. Da mesma forma, quando consideramos a
distância da borda em cada sítio de trabalho, percebemos que a riqueza de espécies registrada
novamente não foi afetada, enquanto a frequência de formigas variou significativamente ao
longo das transecções, sendo refutada também a terceira hipótese de que ambos os parâmetros
seriam maiores quanto mais próximo da rodovia. Os resultados mostraram ainda que a
frequência das espécies de formiga apresentou valores mais altos no sítio com borda artificial.
Esse fato parece estar relacionado com as características deste último sítio, tais como: a
colonização por espécies de gramíneas, acúmulo de serapilheira, além de sua proximidade às
áreas urbana e de pastagem. Tais características favorecem a composição de espécies
oportunistas e generalistas, como aquelas pertencentes às subfamílias Myrmicinae e
Dolichoderinae, consideradas por Andersen e Sparling (1997) como típicas de ambientes
degradados.
Os resultados mostraram que duas das quatro espécies mais frequentes na península de
Ajuruteua, C. atriceps e C. sexguttatus, são reputadas como sinantrópicas, sendo a primeira
reconhecida por sua íntima associação com os ambientes urbanos e considerada como uma
peste de residências (Campos 2011), enquanto a outra é frequentemente associada aos
ambientes perturbados, áreas abertas e ambientes submetidos a efeitos de borda (Wild 2007).
Adicionalmente, é relevante enfatizar que, no presente estudo, as espécies sinantrópicas
apresentaram-se distribuídas de forma similar em ambos os sítios de trabalho e ao longo de
suas transecções, o que indica não haver influência nem do tipo nem da proximidade da
borda, respectivamente, refutando a hipótese proposta de que estas espécies estariam mais
associadas à borda artificial, apresentando maior riqueza e frequência quanto mais próximo à
rodovia.
É importante ressaltar que os impactos antrópicos (ex. construção de rodovias,
assentamentos humanos) não parecem ser os únicos meios de dispersão das espécies
sinantrópicas nas áreas de manguezal. Sucessivos eventos de reprodução, realizada através de
voos nupciais, os quais segundo Simberloff e Wilson (1969) representam o principal meio de
colonização de insetos em ilhas de manguezal, além do transporte pelas marés de fragmentos
vegetais contendo colônias de formigas, como observado em manguezais na Bahia por
Delabie et al. (2006), são também fenômenos responsáveis pela dispersão destas espécies.
Por fim, sabe-se que a fragmentação florestal representa uma das principais ameaças à
biodiversidade, na medida em que promove a extinção e substituição, tanto dos hábitats
quanto das espécies (Vasconcelos et al. 2006). O ecossistema de manguezal, por exemplo,

37
tem sido alvo frequente desse processo, tornando-se cada vez mais exposto às atividades
humanas (Wade et al. 2003). Nossos resultados indicam que a incidência de espécies de
formigas sinantrópicas, na área de estudo, pode ser influência do processo de fragmentação
ocasionado pela construção da rodovia PA-458, representando uma consequência da
intervenção antrópica em ambientes naturais. Adicionalmente, acreditamos que amostragens
em áreas de manguezal em uma escala maior, isto é, mais distanciadas das feições antrópicas,
podem revelar composições de assembléias de formigas mais intrínsecas ao manguezal, o que
acentuaria a influência de bordas artificiais como fonte de espécies sinantrópicas nesse
ambiente.

38
LITERATURA CITADA
ADAMS, E. S. 1994. Territory defense by the ant Azteca trigona: maintenance of an arboreal
ant mosaic. Oecologia 97 : 202-208.
AGOSTI, D., J. D. MAJER, L. ALONSO, AND E. T. SCHULTZ. 2000. Sampling Ground-dwelling
Ants: Case studies from the World’s Rain forests. Curtin University School of
Environmental Biology Bulletin. Perth, Australia. 75 p.
ANDERSEN, A.N., AND G. P. SPARLING. 1997. Ants as Indicators of Restoration Success:
Relationship with Soil Microbial Biomass in the Australian Seasonal Tropics.
Restoration Ecology 5: 109–11.
BEASLEY, C. R., M. E. B. FERNANDES, E. A. G. FIGUEIRA, D. S. SAMPAIO, K. R. MELO, AND R.
S. BARROS. 2010. Mangrove Infauna and Sessile Epifauna. In SAINT-PAUL, U. AND
H. SCHNEIDER (Eds.). Mangrove Dynamics and Management in North Brazil, pp.
109-120. Springer Berlin Heidelberg.
BRONSTEIN, J. L. 1998. The Contribution of Ant-Plant Protection Studies to Our
Understanding of Mutualism. Biotropica 30: 150-161.
CAMPOS, A. E. C. 2011. Emerging urban pests and vector-borne diseases in Brazil. In Dhang,
P. Urban Pest Management: An Environmental Perspective, pp. 19-31. CABI,
Cambridge, UK.
CANNICCI, S., D. BURROWS, S. FRATINI, T. J. SMITH, J. OFFENBERG, AND F. DAHDOUH-
GUEBAS. 2008. Faunal impact on vegetation structure and ecosystem function in
mangrove forests: A review. Aquatic Botany 89: 186–200.
CLARKE, K. R., AND R. M., WARWICK. 2001. Change in marine communities: an approach to
statistical analysis and interpretation. PRIMER-E, Plymouth. UK.
CLARKE, K. R., AND R. N., GORLEY. 2006. PRIMER v. 6: User Manual/ Tutorial. PRIMER-E
Plymouth. UK.
CORTES-LOPES, B., R. AUGLARDAS SANTOS, AND R. A. DOES SANNTUS. 1996. Aspects of the
ecology of ants (Hymenoptera: Formicidae) on the mangrove vegetation of Rio
Ratones, Santa Catarina Island, SC, Brazil. Boletin de Entomologia Venezuelana 11:
123‐133.
DANGERFIELD, J. M., A. J. PIK, D.BRITTON, A.HOLMES, M. I.OLIVER, D. BRISCOE, AND A. J.
BEATTIE. 2003. Patterns of Invertebrate Biodiversity across a natural edge. Austral
Ecology 28: 227–236.

39
DEJEAN, A., S. DUROU, I. OLMSTED, R. R. SNELLING, AND J. ORIVEL. 2003. Nest site selection
by ants in a flooded Mexican mangrove, with special reference to the epiphytic
orchid Myrmecophila christinae. Journal of Tropical Ecology 19: 325–331.
DELABIE, J. H. C., V. R. L. PAIM, I. C. NASCIMENTO, S. CAMPIOLO, AND C. S. F. MARIANO.
2006. As Formigas como Indicadores Biológicos do Impacto Humano em
Manguezais da Costa Sudeste da Bahia. Neotropical Entomology 35: 602-615.
DUNN, R. R. 2004. Managing the tropical landscape: a comparison of the effects of logging
and forest conversion to agriculture on ants, birds, and Lepidoptera. Forest Ecology
and Management 191: 215–224.
FERNÁNDEZ, F., AND M. J. SHARKEY. 2003.Introducción a los Hymenoptera de la región
Neotropical. Sociedad Colombiana de Entomología y Universidad Nacional de
Colombia, Bogotá.
FAGAN, W. F., R. S. CANTRELL, AND C. COSNER. 1999. How Habitat Edges Change Species
Interactions. The American Naturalist 153 : 165-182.
FOLGARAIT, P. J. 1998. Ant biodiversity and its relationship to ecosystem functioning: a
review. Biodiversity and Conservation 7: 1221-1244.
FORYS, E. A., C. R. ALLEN, AND D. P. WOJCIK. 2002. Influence of the proximity and amount
of human development and roads on the occurrence of the red imported fire ant in the
lower Florida Keys. Biological Conservation 108: 27–33.
FLOREN, A., AND K. E., LINSENMAIR. 2001. The influence of anthropogenic disturbances on
the structure of arboreal arthropod communities. Plant Ecology 153: 153–167.
GLASER, M. 2003. Interrelations between mangrove ecosystem, local economy and social
sustainability in Caeté Estuary, North Brazil. Wetlands Ecology and
Management 11: 265-272.
GOTELLI, N. J., A. M. ELLISON, R. R. DUNN, AND N. J. SANDERS. 2011. Counting ants
(Hymenoptera: Formicidae): biodiversity sampling and statistical analysis for
myrmecologists. Myrmecological News 15: 13-19.
HARADA, A. Y., AND J. ADIS. 1998. Ants obtained from trees of a “jacareúba” (Calophyllum
brasiliense) forest plantation in Central Amazonia by canopy fogging: first results.
Acta Amazonica 28: 309-318.
HOLWAY, D. A., AND A. V. SUAREZ. 2006. Homogenization of ant communities in
Mediterranean California: The effects of urbanization and invasion. Biological
Conservation 127: 319 –326.

40
HÖLLDOBLER, B. E., AND O. WILSON. 1990. The ants. Harvard University Press, Cambridge,
UK.
KING, J. R., AND S. D. PORTER. 2005. Evaluation of Sampling Methods and Species Richness
Estimators for Ants in Upland Ecosystems in Florida. Environ. Entomol. 34: 1566-
1578.
LARA, R., C. SZLAFSZTEIN, M. COHEN, U. BERGER, AND M. GLASER. 2002. Implications of
mangrove dynamics for private land use in Bragança, North Brazil: a case
study. Journal of Coastal Conservation 8: 97-102.
LOBRY DE BRUYN, L. A. 1999. Ants as bioindicators of soil function in rural environments.
Agriculture, Ecosystems and Environment 74: 425–441.
LONGINO, J., AND N. M. NADKARNI. 1990. Comparison of Ground and Canopy Leaf Litter
Ants (Hymenoptera: Formicidae) in a Neotropical montane Forest. Psyche 97: 81-93.
LUTHER, D. A., AND R. GREENBERG. 2009. Mangroves: A Global Perspective on the Evolution
and Conservation of their Terrestrial Vertebrates. BioScience 59: 602–612.
MENEZES, M., U. BERGER, AND M. WORBES. 2008. Mangrove vegetation in Amazonia: a
review of studies from the coast of Pará and Maranhão States, north Brazil. Acta
Amazonica 38: 403-420.
OLIVEIRA-SANTOS, L. G. R., R. D. LOYOLA, AND A. B. VARGAS. 2009. Armadilhas de dossel:
uma técnica para amostrar formigas no estrato vertical de florestas. Neotropical
Entomology 38: 691-694.
PARR, C. L. 2008. Dominant ants can control assemblage species richness in a South African
savanna. Journal of Animal Ecology 77: 1191–1198.
PEREIRA, A. P. F. G., F. A. G. ANDRADE, AND M. E. B. FERNANDES. 2006. Dois anos de
monitoramento dos atropelamentos de mamíferos na rodovia PA-458, Bragança,
Pará. Boletim do Museu Paraense Emílio Goeldi Ciências Naturais 1: 77-83.
PEREIRA, L. C. C., D. O. GUIMARÃES, M. J. S. RIBEIRO, R. M. COSTA, AND P. W. M. SOUZA
FILHO. 2007. Use and Occupation in Bragança Littoral, Brazilian Amazon. Journal of
Coastal Research 50: 1116 – 1120.
PHILPOTT, S. M., AND I. ARMBRECHT. 2006. Biodiversity in tropical agroforests and the
ecological role of ants and ant diversity in predatory function. Ecological
Entomology 31: 369–377.
PIMM, S. L., AND P. RAVEN. 2000. Extinction by numbers. Nature 403: 843–845.

41
SILVA, R. R., R. S. M. FEITOSA, AND F. EBERHARDT. 2007. Reduced ant diversity along a
habitat regeneration gradient in the southern Brazilian Atlantic Forest. Forest
Ecology and Management 240: 61–69.
SIMBERLOFF, D. S., AND E. O. WILSON. 1969. Experimental Zoogeography of Islands: The
Colonization of Empty Islands. Ecology 50: 278-296.
StatSoft. 2001. Statistica: Date Analysis Software System. Versão 6.0. StatSoft, USA.
TROMBULAK, S. C., AND C. A. FRISSELL. 2000. Review of Ecological Effects of Roads on
Terrestrial and Aquatic Communities. Conservation Biology 14: 18-30.
VASCONCELOS, H. L., J. M. S. VILHENA, W. E. MAGNUSSON, AND A. L. K. M. ALBERNAZ.
2006. Long-term effects of forest fragmentation on Amazonian ant communities.
Journal of Biogeography. 33: 1348–1356.
WADE, T. G., K. H. RIITTERS, J. D. WICKHAM, AND K. B. JONES. 2003. Distribution and causes
of global forest fragmentation. Conservation Ecology 7: 7 p.
WATKINS, R. Z., J. CHEN, J. PICKENS, AND K. D. BROSOFSKE. 2003. Effects of Forest Roads on
Understory Plants in a Managed Hardwood Landscape. Conservation Biology 17:
411–419.
WILD, A.L. 2007. A catalogue of the ants of Paraguay (Hymenoptera: Formicidae). Zootaxa
1622: 1-55.
WILSON, E. O. 1987. Causes of Ecological Success: The Case of the Ants. Journal of Animal
Ecology 56: 1-9.
ZHENG, D. AND J. CHEN. 2000. Edge effects in fragmented landscapes: a generic model for
delineating area of edge influences (D-AEI). Ecological Modelling 132:175–190.

42
ANEXO 1
BIOTROPICA – JOURNAL OF THE ASSOCIATION FOR TROPICAL BIOLOGY AND
CONSERVATION
Checklist for preparation of manuscripts and illustrations
Online submission and review of manuscripts is mandatory effective 01 January 2005.
I. General Instructions
Manuscripts may be submitted in the following categories, based on these suggested word
limits:
Paper (up to 5000 words); Insights (up to 2000 words); Review (up to 8000 words),
Commentary (up to 2000 words). Word counts exclude title page, abstract(s), literature cited,
tables, figures, or appendices.
Use 8.5" x 11" page size (letter size). Double space everything, including tables, figure
legends, abstract, and literature cited.
Use a 1" margin on all sides. Align left. Avoid hyphens or dashes at ends of lines; do not
divide a word at the end of a line.
Use standard 12 point type (Times New Roman).
Indent all but the first paragraph of each section.
Use italics instead of underline throughout. Italicize non-English words such as e.g., i.e., et
al., cf., ca, n.b., post-hoc, and sensu (the exceptions being ‘vs.’ and ‘etc.’).
Include page number in the centre of all pages. Do use line numbering starting on each page.
Cite each figure and table in the text. Tables and figures must be numbered in the order in
which they are cited in the text.
Use these abbreviations: yr (singular & plural), mo, wk, d, h, min, sec, diam, km, cm, mm, ha,
kg, g, L, g/m². For units, avoid use of negative numbers as superscripts: use the notation /m²
rather than m-².
Write out other abbreviations the first time they are used in the text; abbreviate thereafter: "El
Niño Southern Oscillation (ENSO) . . ."
Numbers: Write out one to ten unless a measurement ( e.g. , four trees, 6 mm, 35 sites, 7 yr,
10 × 5 m, m, ± SE) or in combination with other numbers (e.g. , 5 bees and 12 wasps). Use a
comma as a separator in numbers with more than four digits ( i.e., 1000, but 10,000); use
decima l points as in 0.13; 21°C (no spaces); use dashes to indicate a set location of a given
size (e.g., 1-ha plot).
Spell out ‘percent’ except when used in parentheses (20%) and for 95% CI.
Statistical abbreviations: Use italics for P, N, t, F, R, r, G, U, N, χ2 (italics, superscripts non-
italics); but use roman for: df, SD, SE, SEM, CI, two-way ANOVA, ns.
Dates: 10 December 1997; Times: 0930 h, 2130 h Latitude and Longitude are expressed as:
10°34′ 21 ″ N, 14°26 ′ 12″ W Above sea level is expressed as: asl Regions: SE Asia, UK (no
periods), but note that U.S.A. includes periods. Geographical place names should use the
English spelling in the text (Zuric h, Florence, Brazil), but authors may use their preferred
spelling when listing their affiliation (Zürich, Firenze, Brasil).
Lists in the text should follow the style: … : (1)… ; (2)…; and (3)…, as in, “The aims of the
study were to: (1) evaluate pollination success in Medusagyne oppositifolia; (2) quantify gene
flow between populations; and (3) score seed set.”
Each reference cited in text must be listed in the Literature Cited section, and vice versa.
Double check for consistency, spelling and details of publication, including city and country
of publisher.
Literature citations in the text are as follows:
One author: Yaz (1992) or (Yaz 1992)

43
Two authors: Yaz and Ramirez (1992); (Yaz & Ramirez 1992)
Three or more authors: Yaz et al . (1992), but include ALL authors in the literature cited
section.
Cite unpublished materials or papers not in press as (J. Yaz, pers. obs.) or (J. Yaz, unpubl.
data).
Initials and last name must be provided. ‘In prep’ or ‘submitted’ are NOT acceptable, and we
encourage authors not to use ‘pers. obs.’ or ‘ unpubl. data’ unless absolutely necessary.
Personal communications are cited as (K. A. Liston, pers. comm.).
Use commas (Yaz & Taz 1981, Ramirez 1983) to separate citations, BUT use semicolon for
different types of citations (Fig. 4; Table 2) or with multiple dates per author (Yaz et al.
1982a, b; Taz 1990, 1991). Order references by year, then alphabetical (Azy 1980, Yaz 1980,
Azy 1985).
For the review purpose, submit the entire manuscript, with Tables, Figure legends and Figures
embedded at the end of the manuscript text, as a Microsoft Word for Windows document
(*.doc), or equivalent for Mac or Linux. Do NOT submit papers as pdf files.
II- Text
No heading for Introduction. First line or phrase of Introduction should be S MALL CAPS.
Main headings are METHODS, RESULTS, and DISCUSSION: All CAPITALS and Bold.
Flush left, one line.
Second level headings: SMALL CAPS, flush left, Capitalize first letter, begin sentence with
em-dash, same line (e.g., I NVENTORY TECHNIQUE.—The ant inventory…).
References to figures are in the form of ‘Fig. 1’, and tables as ‘Table 1’. Reference to online
Supporting Information is as ‘Fig. S1’ or ‘Table S1’.
III- Literature Cited
(Continue page numbering and double spacing)
No ‘in prep.’ or ‘submitted’ titles are acceptable; cite only articles published or ‘in press’. ‘In
press’ citations must be accepted for publication. Include journal or publisher.
Insert a period and space after each initial of an author's name; example: YAZ, A. B., AND
B. AZY. 1980.
Authors Names: use SMALL CAPS .
Every reference should spell out author names as described above. BIOTROPICA no longer
uses ‘em-dashes’ (—) to substitute previously mentioned authors.
Double-space. Hanging indent of 0.5 inch.
Leave a space between volume and page numbers and do not include issue numbers. 27: 3–12
Article in books, use: AZY, B. 1982. Title of book chapter. In G. Yaz (Ed.). Book title, pp.
24–36. Blackwell Publications, Oxford, UK.
Dissertations, use: ‘PhD Dissertation’ and ‘MSc Dissertation’.

44
CONCLUSÃO
O número de espécies registrado para o manguezal da península de Ajuruteua (n=45)
apresenta um valor próximo àqueles encontrados em outras áreas de manguezal, tanto no
Brasil quanto em outras regiões do mundo. No entanto, essa riqueza é bastante inferior
quando comparada com sistemas de terra firme, onde a fauna de formiga utiliza o chão da
floresta, chegando a alcançar neste ambiente mais de dez vezes o número de espécies
encontrado nos manguezais da Amazônia brasileira.
A assembléia de formigas associada ao manguezal é predominantemente arborícola,
sendo a copa das árvores caracterizada por espécies dominantes, as quais podem ter
influenciado a baixa riqueza da mirmecofauna encontrada nesse estrato da floresta.
As bordas artificiais apresentam riqueza de espécies de formigas similar às bordas
naturais, sendo que a ocorrência de formigas nas iscas foi maior próximo ás bordas, tanto
artificial quanto natural. Espécies oportunistas e generalistas estão mais relacionadas às
bordas do tipo artificial.
Espécies de formigas sinantrópicas representaram uma importante fração da
assembléia de formigas nos manguezais estudados, registrando elevada ocorrência em todos
os estratos da floresta, e sendo aqui avaliado como um efeito da presença de atividades
antrópicas, como a construção da rodovia PA-458.







