dr. en tao wang hu departamento de microbiología...
TRANSCRIPT

INFORME FINAL TÉCNICO DEL PROYECTO DE 20050014
“Diversidad genética de las bacterias endofíticas de la leguminosa
Conzattia multiflora
Dr. En Tao Wang Hu
Departamento de Microbiología
Escuela Nacional de Ciencias Biológicas
15 del marzo de 2006

2
Resumen
Para investigar la diversidad de bacterias endofíticas de La leguminosa Conzattia multiflora
que es un árbol nativo de México. Estas plantas no forman nódulos radicales con los rhizobia,
pero tienen nódulos en su tallo. Se encontraron bacterias endofíticas en estos nódulos. Estas
bacterias son entrobacterias de los géneros Enterobacter, Erwinia, Klebsiella, Pantoea y
Salmonella. No hay genes de fijación de nitrógeno ni de nodulación encontrados en estas
bacterias y la inoculación de ellas no causa la formación de nódulos. En el trabajo presente,
se aisló el DNA metagenómico bacteriano de tejidos de nódulos de C. multiflora, el cual se
usó en PCR del genes de 16S rRNA, nifH, y nodA. No fue detectado el gen nifH por PCR,
indicando que las bacterias endofíticas del Conzattia multiflora no fijan nitrógeno. Se obtuvo
producto de nodA, pero se necesitan más estudios, como hibridación y secuenciación, para
confirmar que estos productos son gen de nodulacón. Se obtuvo una biblioteca del gen de
16S rRNA por medio de clonación, ya se encontraron diferentes clonas, cuales representen
bacterias distintas por el análisis de RFLP de EcoRI. Se harán más trabajos para seleccionar,
caracterizar por RFLP y secuenciar las clonas positivas. Las secuencias obtenidas se van a
comparar con las bacterias aisladas de Conzattia multiflora y con las reportadas en las
publicaciones, para identificar su diversidad por las relaciones de filogenía.

3
NTRODUCCIÓN
Microorganismos endofíticos
La asociación de los microorganismos endofíticos con su hospedero puede ser mutualista y
llegar hasta el umbral de un microorganismo patógeno en estado de latencia. (Strobel y Long,
1998). Pero generalmente los microorganismos endofíticos comprenden a los hongos y
bacterias que viven sin causar daño en el interior de células y tejidos de plantas superiores
durante una parte considerable de su ciclo de vida y pueden localizarse en espacios
intracelulares, intercelulares o tejido vascular (Quispel, 1992; Reinhold-Hurek y col., 1998).
Entonces, los microorganismos asociados a las plantas se pueden clasificar en tres tipos
basándose en su efecto en el organismo huésped:
A. Simbióticos: Son aquellos microorganismos que establecen relaciones que conllevan
beneficios tanto para ellos como para el organismo huésped.
B. Fitopatógenos: Son aquellos microorganismos que establecen relaciones que causan
daño al organismo huésped.
C. Endofíticos: Son aquellos microorganismos que establecen relaciones que no causan
ningún daño en el organismo huésped. (Hallmann y col. 1997)
Con base en estos conceptos, los microorganismos endofíticos no incluyen microorganismos
patógenos ni simbióticos. La existencia de los microorganismos endofíticos ha sido
confirmada desde hace más de 100 años. El primer reporte sobre microorganismos
endofíticos fue por Freeman (1904). Aunque la investigación sobre los microbios endofíticos
se desarrolló lentamente comparando con las de fitopatógenos y simbióticos, ya se tienen
conocimientos básicos sobre la diversidad e impacto en las plantas, productos metabólicos y
sus usos potenciales de estos microbios.
Interacción entre los microorganismos endofíticos y las plantas
La asociación entre los microorganismos endofíticos y las plantas es mutualista pues, los

4
microbios solos o ambas partes pueden obtener beneficios. Se ha considerado que las
bacterias endofíticas pueden tener ventajas competitivas incluso sobre las bacterias
rizosféricas (que viven en la superficie de las raíces) ya que la disponibilidad de nutrientes es
mayor en el interior de las plantas y el número de microorganismos endofíticos es menor que
el de rizosféricos (James, 2000). Por otro lado también los microorganismos obtienen
ventajas tales como condiciones óptimas de pH y temperatura, variadas fuentes de carbono y
nitrógeno, además de que la planta puede proporcionar un ambiente anoxigénico el cual
puede ser apropiado para el desarrollo óptimo de unas bacterias.
Aunque en muchos casos no conocemos los efectos que causan las bacterias endofíticas en la
planta, la existencia de estas bacterias puede ser vital y las plantas sin bacterias endofíticas
pueden llegar a morir gradualmente en algunos casos. Por lo que las bacterias endofíticas son
de gran importancia para la conservación de bosques temperados y selvas tropicales debido a
que confiere a sus plantas ventajas competitivas sobre otras especies vegetales. Se han
encontrado beneficios tales como:
A. Fijación de nitrógeno: Esta dada por bacterias tales como los géneros
Gluconoacetobacter, Acetobacter, Azospirillum y Azoarcus entre otros. Los cuales
pueden vivir en la raíz y en los tallos (Muñoz Rojas y col. 2005). Estas bacterias
fijadoras de nitrógeno han sido aisladas o detectadas en varias plantas, como café,
papas y maíz (Caballero-Mellado y col., 2004; Estrada-De Los Santos 2001;
Jimenez-Salgado y col. 1997; Reis y col., 2004; Tapia-Hernández y col. 2000), y
unas plantas herbáceas salvajes (Reinhold–Hurek y Hurek, 1998). Se ha estimado que
estas bacterias pueden ofrecer la mayor parte de nutrientes nitrogenados a las plantas
en campos no fertilizados.
B. Producción de fitohormonas: Se tiene información de bacterias endofíticas que son
capaces de producir fitohormonas tales como gibelerina (Bottini y col., 2004) y ácido
indol-acético (Fuentes-Ramírez y col., 1993). Este efecto ha sido usado a explicar
porque la inoculación de bacterias puede aumentar la producción de biomasa de
plantas.
C. Resistencia a los ambientes adversos: Esta puede darse por la existencia de

5
mecanismos como la supresión o sobreexpresión de hormonas ya antes mencionado,
que hacen a la planta adquirir ventajas para desarrollarse mejor en un ambiente
determinado. Los ejemplos son la resistencia a climas secos y a metales pesados
pueden revisarse en el trabajo de Newman y Reynolds (2005).
D. Resistencia a fitopatógenos: Muchos microorganismos pueden secretar sustancias
que actúan como agentes antibióticos y bioinsecticidas tal es el caso de muchas
especies de hongos endofíticos y algunas bacterias (Tabla 1) las cuales tienen esta
propiedad. La inhibición de las bacterias endofíticas, tal como Pseudomonas y
Bacillus a los fitopatógenos se han reportado en plantas de papas (Sessitsch y col.,
2004).
Los microorganismos endofíticos también tienen importancias comerciales. Basándose en
sus impactos a las plantas y diversos productos metabólicos, los microorganismos
endofíticos se pueden usar de diferentes maneras:
A. Inóculos como biofertilizantes sobre la base de sus capacidades de fijación de
nitrógeno (Paredes-Cardona y col., 1988), y de producción de fitohormonas (Bottini
y col., 2004).
B. Inóculo como biocontrol de insectos y fitopatógenos.
C. Inoculo para bioremediación (Newman y col., 2005).
D. Producción de antibióticos, anticancerígenos, como un diterpeno que tiene alta
capacidad de anticancerígena, el cual, extraído de diversas especies, ha formado un
producto comercial de billones de dólares (Strobel y Daisy, 2003).
E. Otras sustancias incluyendo enzimas, polisacáridos, etc. Un producto importante es la
goma natural xantana, proveniente de los polisacáridos de la bacteria fitopatógena
Xanthomonas campestris (García-Ochoa y col., 2000). Este producto se usa en
extracción de petróleo y en otras industrias.
Generalmente, otras bacterias o microorganismos también se utilizan para estos fines, pero
las bacterias endofíticas tienen algunas ventajas:

6
A. Como inóculo de biofertilizantes o biocontrol, las bacterias endofíticas tienen efectos
más estables que otras bacterias, debido a que su hábitat es localizado dentro las
plantas donde impera un ambiente protegido para las bacterias.
B. Viviendo dentro las plantas, las bacterias endofíticas desarrollan rutas específicas de
metabolismo, que ofrecen la posibilidad de producir metabolitos diferentes a otras
bacterias del mismo ambiente. Han sido encontrados antibióticos nuevos (Tabla 1).
Tabla 1. Algunos productos naturales aislados de microorganismos endofíticos (Strobel y
col., 2003)
Microorganismo
Huésped Producto
Bacterias Serratia marscesens R. penicillata Oocydin A (antifúngico Pseudomonas viridiflava
Pastos Ecomycina (antifúngico)
Streptomyces Raíces de plantas de patatas
Estreptomicina (antibiótico)
Pseudomonas Plantas diversas Pseudomycina (antifúngico) Hongos
Criptosporocis quercina
Tripterigeum wilfordii Cryptocin, Criptocandin, (antifúngicos)
Cytonamea sp.
Plantas diversas
Inhibidores proteicos de Cytomelovirus (antivirales)
Muscodor albus Cinnamomum zeylanicum
(Compuestos volátiles de actividad antfúngica)
Muscodor roseus Cinnamomum zeylanicum
Antibióticos volátiles
Glicocadium sp. Plantas diversas Antibióticos volátiles T. andreanae T. brevifolia Paclitaxel (anticancerígeno) Seimatoantlerium tepoiense
Maguireothamus speciosus
Paclitaxel (anticancerígeno)
P. microspora Terminalia morobensis
Pestacin, Isopestacin, (antioxidantes)
Nodulisporum sp. Bontia daphnoides Ácido nodulispórico, (insecticida) Pseudomassaria sp. Selva lluviosa africana Naftalina (insecticida) Fusarium subglutinans T. wilfordii Subglutinol A y B (inmunorepresores)

7
Biodiversidad de las bacterias endófiticas
Con los datos mencionados arriba, podemos ver que los microorganismos endofíticos tienen
diferentes huéspedes, diferentes capacidades de metabolismo, y varios efectos en plantas.
Para conocer y explorar estos recursos biológicos, se necesita a estudiar los microorganismos
mismos, incluyendo que tipo de microorganismos estén dentro de las plantas, y que se
reproduzcan dentro de las mismas. La diversidad de los microbios endofíticos es la base para
entender sus diversas capacidades de producir compuestos, vivir en diversos ambientes,
asociarse a diferentes plantas, etc.
En la tierra existe una gran diversidad de bacterias endofíticas. Existen alrededor de 300,000
especies de plantas superiores en el planeta y cada planta una de estas contiene a su vez, una
o más especies bacterianas dentro de ella (Strobel y col., 2004). Por lo que la diversidad de
bacterias endofíticas se puede estimar en valores superiores, sugiriendo una extensa
diversidad de endofítas, la posibilidad de encontrar nuevas especies bacterianas y encontrar
nuevos compuestos útiles. Algunas bacterias endofíticas han sido reportadas en los últimos 5
años como especies nuevas, tal como Samsonia erythrinae (Sutra y col., 2001), Candidatus
Burkholderia kirkii y Burkholderia nigropunctata (Van Oevelen y col. 2002; 2004).
De los miles de ecosistemas del planeta, las selvas tropicales son los ecosistemas con mayor
diversidad biológica, cubren el 1.44 % de la superficie de la tierra, pero contienen más del
60% de su biodiversidad (Mittermeier y col. 1999). La elevada cantidad de plantas
endémicas hace posible encontrar gran cantidad de endofítas.
Han sido encontrados hongos y bacterias en plantas tales como los géneros bacterianos
Pseudomonas, Enterobacter, Bacillus, Erwina y Xanthomonas (Muñoz Rojas y col., 2005).
Además de géneros fúngicos tales como Cryptosporiopsis, Muscodor, Fusarium, etc.
(Strobel y col., 2003). Estas bacterias representan solo una pequeña parte de la gran
diversidad que pueden tener estos microorganismos. Debemos decir también que muchos de
estos microorganismos no han podido ser aislados por métodos convencionales, por lo que
las plantas pueden ser una gran fuente de microorganismos nuevos que valgan la pena ser

8
estudiados, tanto por su gran potencial para obtener productos naturales como para dilucidar
la relación que existe entre estos y el organismo hospedero.
Para investigar la diversidad de bacterias endofíticas, se han usado dos estrategias: (1) aislar
los microorganismos con diferentes medios de cultivo y (2) con métodos independientes de
cultivo, como amplificación por PCR de los genes de 16S rRNA u otros, como clonación y
secuenciación. El aislamiento es una estrategia básica e importante porque con este método
se pueden tener cepas las cuales se usan para investigaciones avanzas y para aplicaciones.
Pero la desventaja de este método es que muchas o la mayoría de las bacterias no crecen en
los medios de cultivo. Entonces con los métodos de cultivo, no podemos encontrar la
diversidad real de las bacterias endofíticas en una planta. Con los métodos independientes de
cultivo, se tiene la capacidad a encontrar todas las bacterias que existen en una planta, pero
no se puede tener las cepas para otras investigaciones. En muchos trabajos publicados sobre
las bacterias endofíticas, se han usado las dos estrategias para tener las ventajas de ambos
métodos.
Conzattia multiflora
Existe una gran variedad y riqueza en la flora de nuestro país, por lo que es posible encontrar
muchas especies vegetales que tienen interés económico, tales como plantas medicinales,
alimenticias, madereras y bioremediación etc. Algunas investigaciones han sido realizadas
en México sobre las bacterias endofíticas fijadoras de nitrógeno de plantas de café, de caña
de azúcar, y de pino (Caballero-Mellado y col., 2004; Estrada-De Los Santos 2001;
Jiménez-Salgado y col. 1997; Reis y col., 2004; Tapia-Hernández y col. 2000), y sobre
Rhizobium etli endofítico de maíz (Gutiérrez-Zamora y Martínez-Romero, 2001). Varias
especies bacterianas nuevas han sido reportadas en estas investigaciones, indicando que hay
una gran diversidad de las bacterias endofíticas asociadas a las plantas mexicanas.
La leguminosa arbórea Conzattia multiflora es nativa en México, la cual crece en diversos
estados del país. No tiene ningún valor comercial. Por eso, es una planta no atrapa
investigaciones científicas. Esa planta es un miembro del familia Caesalpinieae de la

9
subfamilia Caesalpinioideae. Esta familia incluye 47 géneros y la mayoría no forman
nódulos, casi la mitad de ellos no han sido reportados, incluyendo Conzattia (Allen & Allen,
1981), aunque sus flores se utilizan para comida y como ornamentación. Puede encontrarse
en los climas semiárido y de bosque lluvioso, crece junto a cactos arborescentes y con plantas
del tipo Ipomea, Brusela, Fouqueira, Ceiba y Hura, principalmente en la región de la cuesta
del pacífico, en los valles secos de la cuenca del Río Santiago y los estados de Morelos,
Puebla, Oaxaca, Sinaloa, Colima, Jalisco, Michoacán etc. Crece solo a alturas entre 400 y
1400 m sobre el nivel del mar. Como características principales podemos citar que es un
árbol de temporada pues florea entre junio y julio y da fruto de octubre a enero, crece hasta
alturas de 10 a 20 m. Su tronco puede medir de 30 a 50 cm de diámetro, la corteza es lisa y de
color café rojizo, con pequeñas ramas que tienen de 20 a 50 cm de longitud las cuales
contienen de 10 a 15 pares de hojuelas de hojas pinadas. Las hojas pinadas tienen de 5 a 10
mm de longitud, son apiculadas y redondeadas en ambos extremos. Da muchas flores con
pedículos de 5 a 10 mm de longitud, sépalos sin vellosidades de 5 mm de longitud y pétalos
elípticos y ovalados de 7 a 10 mm de largo. (Mc Vaugh, 1984.)
Llama la atención su capacidad de formación de nódulos (Fig. 1) en el tronco del árbol, los
cuales, difieren de los tres tipos de nódulos comúnmente encontrados (Tabla 2, Fig. 2) en
otras especies de plantas, los cuales son todos causados por infección de bacterias. En las
células del los nódulos de C. multiflora, también encontramos muchas bacterias (Fig. 1). Por
lo que vale la pena el estudio de estos nódulos de la planta, tanto por su posible potencial de
conocer bacterias nuevas como por la posibilidad de conocer sus mecanismos de nodulación.
La nodulación de leguminosas ha sido investigada extensivamente y se tienen muchos
conocimientos sobre la diversidad de las bacterias contenidas en los nódulos radicales entre
los que destacan los géneros de Rhizobium y Frankia, existen algunas diferencias entre el
tipo de nódulos de cada planta aunque básicamente tienen la misma función. La diferencia
estriba principalmente en su estructura ya que en el caso de los nódulos actinorrízicos se
forman lóbulos definidos, mientras que en las leguminosas se hallan formas de glóbulos y de
lóbulos definidos. También existen diferencias en cuanto a la estructura y función con
respecto a las agallas de las hojas y en forma de corona, ya que estas son resultado de

10
procesos patógenos. A continuación se da una breve explicación sobre cada tipo de nódulo.
Nódulos radicales. Los nódulos radicales son organelos simbióticos y son los que están
formados por bacterias fijadoras de nitrógeno como los géneros Rhizobium y Frankia los
cuales se localizan preferentemente en las raíces de plantas leguminosas o actinorrízicas. En
unas leguminosas, los nódulos también se forman en el tallo, como en Sesbania rostrata (Fig.
2). Los nódulos de leguminosas han sido estudiados intensivamente. Estos nódulos tienen
dos tipos: forma determinada e indeterminada, las cuales se determinan por las plantas.
Las leguminosas de fríjol, chicharo, cacahuate etc. tienen nódulos determinados, con forma
de pelotita. Las de trébol, haba, y alfalfa etc. tienen nódulos indeterminados los cuales son
lóbulos. Estos nódulos contienen leghemoglobina, una proteína responsable de transportar
oxigeno a las bacterias para producir energía. Las bacterias que causan la formación de
nódulo viven dentro las células y reducen nitrógeno gaseoso a amoniaco el cual se usa por la
planta en la síntesis de proteínas. Hasta la fecha, han sido descritas más de 40 especies
bacterianas que inducen la formación de los nódulos en las leguminosas. La asociación entre
estas bacterias y las leguminosas es especifica: cada especie de leguminosa forma nódulos
con ciertos rhizobia, y cada especie de rhizobia tiene un rango limite de huéspedes en las
leguminosas (Wang y col., 2005.)
Figura 2. Diferentes tipos de nódulos de plantas: A) Agallas en corona causada por Agrobacterium en el tallo
de tabaco; B). Nódulos de tallo inducidos por Azorhizobium en Sesbania rostrata; C) Nódulos radicales
inducidos por Frankia en Alnus glutinosa; (Madigan y col. 2004) D) Agallas de las hojas causadas por
Phylobacterium en Pavetta zimmermanniana (Knösel DH. 1984).
Figura 1. Fotos demostrando los nódulos en
tronco de Conzattia multiflora y las bacterias
endofíticas en los nódulos. A. Nódulos en tronco;
B. Nódulos aislados del tronco; C. Bacterias
endofíticas en las células del nódulo de C.
multiflora (Wang y col. Dato no publicado).
D
Fotos eliminados
Fotos eliminados
11
Agallas de corona (crown galls) (Fig. 2). Los tumores en las plantas se dan por bacterias
patógenas en algunas especies vegetales tales como los géneros Brassica, Bryophyllum,
Lilium, Lycopersicon, y Nicotiana. La formación de estas agallas de corona causa perdida de
biomasa o mata gradualmente la planta. Las agallas de corona son causadas principalmente
por especies de Agrobacterium, como A. tumenfaciens y A. rhizogenes, atacan
principalmente a plantas dicotiledóneas. Existen 2 tipos de tumores en forma de corona, los
que se llaman teratomas son tumores con forma de hoja o excrecencias continuas en la raíz, el
otro tipo de tumores no muestran una diferenciación remarcable. Las cepas fitopatógenas de
Agrobacterium contienen un plásmido Ti (de tumor inducción) o Ri (de raíz pilosa
inducción). La infección de Agrobacterium no es específica y normalmente entra por las
heridas. Dentro las células infectadas, un fragmento (T-DNA,) del plásmido Ti se inserta en
el cromosoma de la planta por hibridación. Este fragmento es responsable de producir
fitohormonas que estimulan la división de las células. Este fragmento también causa la
producción de opinas, (octopina y nopalina, dos derivados de la arginina), los cuales se usan
como fuente de carbono únicamente por la cepa de Agrobacterium. De este modo se asegura
una fuente alimenticia a las cepas de Agrobacterium mientras viven en las células de las
plantas (Sengbusch, 2005).
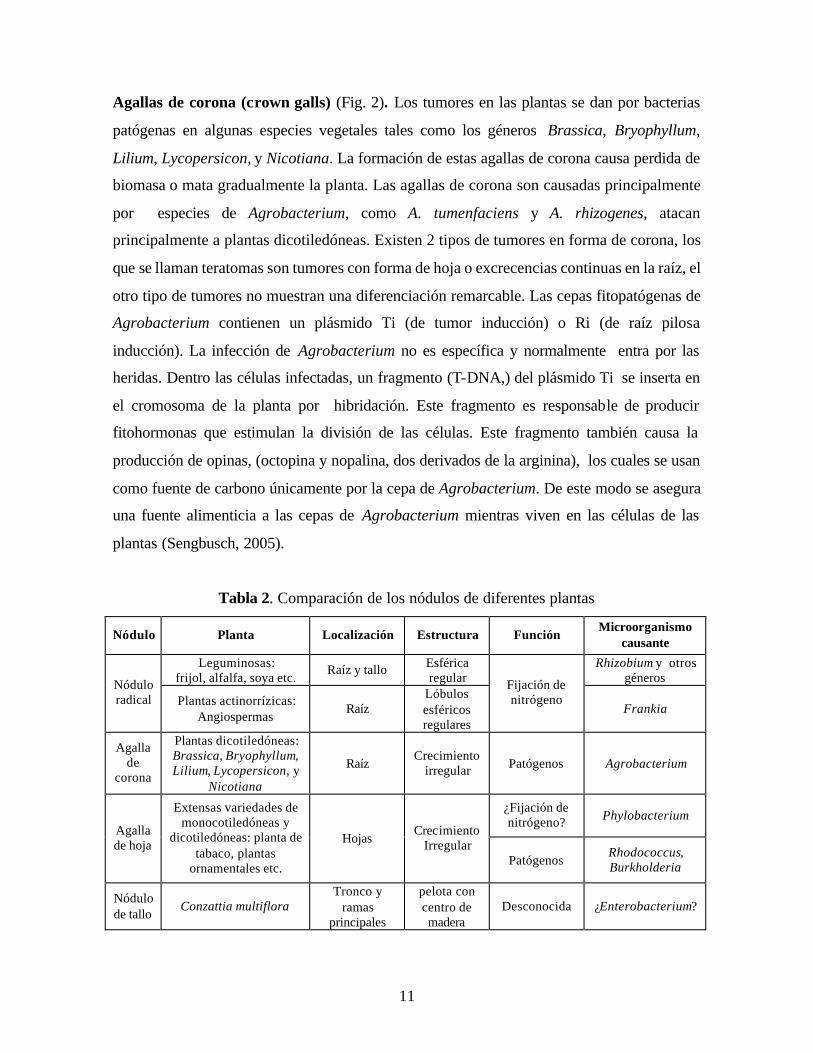
Tabla 2. Comparación de los nódulos de diferentes plantas
Nódulo Planta Localización Estructura Función Microorganismo
causante
Leguminosas: frijol, alfalfa, soya etc.
Raíz y tallo Esférica regular
Rhizobium y otros géneros Nódulo
radical Plantas actinorrízicas: Angiospermas
Raíz Lóbulos esféricos regulares
Fijación de nitrógeno
Frankia
Agalla de
corona
Plantas dicotiledóneas: Brassica, Bryophyllum, Lilium, Lycopersicon, y
Nicotiana
Raíz Crecimiento
irregular Patógenos Agrobacterium
¿Fijación de nitrógeno? Phylobacterium
Agalla de hoja
Extensas variedades de monocotiledóneas y
dicotiledóneas: planta de tabaco, plantas
ornamentales etc.
Hojas Crecimiento
Irregular Patógenos
Rhodococcus, Burkholderia
Nódulo de tallo
Conzattia multiflora Tronco y
ramas principales
pelota con centro de madera
Desconocida ¿Enterobacterium?

12
Agallas de las hojas (leafy galls) (Fig. 2). Las bacterias Gram positivas del género
Rhodococcus y las Gram negativas Phylobacterium y Burkholderia causan malformaciones
en las partes aéreas de las plantas, propiamente en las hojas formando tumores en los
meristemas axilares de estas.
La naturaleza y posición de las malformaciones depende de la especie y el método de
infección, la más grave se caracteriza por disparos múltiples y disparos primordiales, los que
se originan por la existencia de tejidos meristemáticos. La patogenicidad está dada por un
plásmido lineal que acarrea un locus de gran virulencia. (Vereecke y col. 2002)
La formación de agallas por el género Phylobacterium aún no está clara, y es posible que las
gallas de hojas formadas por esta bacteria tengan capacidad de fijación de nitrógeno.
Sobre las agallas de las hojas ocasionadas por el género Burkhodelia, solo se han encontrado
estas bacterias por métodos de biología molecular.
Comparados con los nódulos mencionados aquí, los nódulos de Conzzatia multiflora son de
un tipo nuevo por la presencia de una “pelota” de madera y por la presencia de
enterobacterias en los tejidos del nódulo, aunque aún no se confirma si estas bacterias tienen
relación con la formación del nódulo.
En la literatura no existen trabajos acerca de la nodulación y las bacterias endofíticas en
Conzattia multiflora (Allen y Allen, 1981). En un trabajo “Aislamiento y caracterización de
las bacterias endofíticas de la leguminosa arbórea Conzattia multiflora” realizado por el
equipo del profesor En Tao Wang Hu, se estudiaron los nódulos en esta leguminosa por
medio de técnicas de microscopia electrónica, aislamiento de microorganismos en medio
semi-sólido, análisis con técnicas de PCR-RFLP (PCR amplificación-fragmentos de
restricción de tamaño polimórfico) y secuenciación de los genes 16S rRNA obteniéndose las
siguientes conclusiones:
- Estos nódulos no son parecidos a los anteriormente reportados en la literatura.
- Se encontraron muchas bacterias dentro de estos.

13
- De los mismos se han aislado bacterias de géneros Enterobacter, Salmonella,
Erwinia, Klebsiella, y Pantoea, (Fig. 3) y se confirma que son endófitas.
- No se han encontrado los genes de nodulación y no se ha relacionado la nodulación
de Conzattia con los aislados bacterianos.
Consecuentemente, estudios de diversidad de bacterias endofiticas por métodos moleculares
son necesarios para confirmar si algunas bacterias tienen relación con la formación de los
nódulos de Conzattia, porque muchos microorganismos endofiticos no son cultivables.
Grimontella senegalensis
Pantoea stewartii Pantoea ananatis
Co9945 Pantoea agglomerans
Co9927 C09926
Co9941 Erwinia psidii
Co9929 Erwinia amylovora
Erwinia rhapotici Co9936
Salmonella enterica bv. typhimurium Salmonella enterica bv. typhi
Salmonella enterica bv. typhi Salmonella bongori
Co9901 Co9937
Co9902 Enterobacter homaechei
Enterobacter cloacae Co9935
Klebsiella pneuminiae Klebsiella oxytoca
Figura. 3. Árbol simplificado de filogenia que muestra las relaciones entre los aislados de los nódulos de Conzattia y las bacterias relevantes. El árbol se construye con las secuencias del gen 16S rRNA. Las cepas presentadas en letras negras son aisladas de los nódulos de Conzattia multiflora (Wang y col. Dato no publicado).

14
El objetivo general del trabajo fue a determinar mediante métodos de biología molecular la
diversidad de las bacterias endofíticas en los nódulos de la leguminosa arbórea Conzattia
multiflora. Los objetivos específicos fueron a determinar si estos nódulos son causados por
bacterias; determinar si existen bacterias fijadoras de nitrógeno en estos nódulos y
determinar que otro tipo de bacterias existe además de las ya encontradas.
JUSTIFICACIÓN
La interacción entre microbios y plantas es una rama de las ciencias biológicas. Sobre la base
del efecto de microbios en las plantas se clasifican en tres tipos las relaciones entre ellos:
simbiosis, patogénesis y endófitas. Extensas investigaciones han sido realizadas sobre
simbiosis y patogénesis y muchos conocimientos importantes han sido obtenidos incluyendo
el registro de nuevos microorganismos.
En años recientes se han aumentado los estudios sobre microorganismos endofíticos, en estos
estudios ya encontraron gran cantidad de diversas bacterias endofiticas. La importancia del
estudio de estos microorganismos radica en que se tiene una gran posibilidad de conocer
bacterias nuevas y de potencial biotecnológico para diferentes industrias como la
farmacéutica y la alimenticia, además de su importancia agrícola y en los campos de la
medicina y la biotecnología. También ofrecen pistas sobre la relación que guardan estos
microorganismos con su entorno para tener una mejor comprensión sobre como se
desenvuelven en el ambiente y que papel juegan en este.
En México existe una gran diversidad de ambientes con una flora muy rica y variada, con
respecto a la flora puede decirse que es un recurso importante para hacer investigaciones
básicas y aplicadas, incluyendo el estudio de las bacterias endofiticas.
La Conzattia multiflora es una leguminosa nativa en México y es una planta casi no
estudiada. Nos interesa esta planta porque forma nódulos en su tronco que son diferentes a
los nódulos o tumores conocidos. Las bacterias aisladas de estos nódulos son enterobacterias,
pero no se ha confirmado que estas causen la formación del nódulo. Por lo que es importante

15
realizar un estudio para conocer los microorganismos presentes dentro los nódulos, para
saber la diversidad de bacterias endofíticas de la planta.
El estudio de las bacterias endófitas de C. multiflora tiene posibilidad de reportar un nuevo
tipo de asociación de bacteria- leguminosa, además de la posibilidad de encontrar nuevos
tipos de bacterias.
MATERIALES Y MÉTODOS
Para realizar este estudio se están llevando a cabo los pasos que se detallan a continuación:
Muestreo de nódulos
Las muestras de nódulos se toman de distintos árboles de C. multiflora escogidos por sus
características de la sierra de Huautla en el estado de Morelos, estos se cortan y almacenan en
hielo, al llegar al laboratorio se almacenan en refrigeración a 4º C, para extraer el ADN de las
bacterias posteriormente. Se realiza el muestreo en junio cuanto las plantas están en la fase de
crecimiento vegetal y tienen un gran número de bacterias endofíticas.
Extracción de DNA metagenómico (Holme y col. 1996)
• Cosechar 10 g de nódulos mediante cortes con bisturí.
• Desinfectarlos superficialmente mediante lavados con etanol al 70% durante 5 min,
solución de hipoclorito de sodio (2% de cloruros disponible) 5 min, etanol al 70% 30
s y lavar dos veces con agua destilada estéril (Vincent 1970).
• Macerar en regulador de fosfatos salino (PBS) pH 8, hasta obtener partículas muy
pequeñas y extraer la mayor cantidad posible de bacterias.
• Agitar 2 h a 120 rpm y temperatura ambiente.
• Transferir 1.5 mL de la suspensión a un tubo Eppendorf nuevo y centrifugar 30 s a
12,000 x g, repetir este paso hasta terminar con la suspensión.
• La pastilla resultante se disuelve en 500 µL de regulador de TE. (tris – HCl 10 mM,
EDTA 1 mM, pH 8)
• Agregar 50 µL de lysosima e incubar a 37º C de 30 a 60 min.

16
• Adicionar 70 µL de SDS al 10%, 6 µL de proteinasa K (10 mg/mL) e incubar en baño
maría a 65º C 10 min.
• Adicionar 100 µL de solución 5 M de NaCl y poner 10 s en el vortex.
• Adicionar 80 µL de CTAB/NaCl, poner en el vortex 10 s e incubar en baño maría a
65º C 10 min.
• Adicionar un volumen de cloroformo alcohol isoamílico, (24:1) poner en vortex 10 s
y centrifugar a 12,000 x g 10 min.
• Pasar el sobrenadante a un nuevo tubo Eppendorf y desechar el precipitado.
• Adicionar 0.6 volúmenes de isopropanol a -20º C y reposar 30 min.
• Centrifugar 10 min a 12,000 x g y desechar el sobrenadante.
• Adicionar 800 µL de etanol 70% a -20º C, poner 10 s en el vortex y centrifugar 5 min
a 12,000 x g.
• Desechar el sobrenadante y secar al aire 10 min.
• Adicionar de 30 a 100 µL de agua desionizada, bidestilada o miliQ.
• Almacenar a 4 ó -20º C.
Para comprobar que se obtuvieron aislados de DNA se utiliza la electroforesis en gel de
agarosa tal como se describe a continuación:
• Mezclar de 5 a 10 µL de muestra con 2 µL de regulador de carga en un gel de agarosa
al 1% a 100 V durante 40 min.
• Revelar el gel en una cantidad apropiada de agua destilada adicionada con 0.5 µg/mL
de bromuro de etidio.
• Detectar el DNA por medio de luz ultravioleta en un transiluminador.
Amplificación del gen 16S rRNA. (Van Berkum y col. 1996)
Colocar en un microtubo estéril de 200 µL, en una cama de hielo: (mezcla de reacción para 2
tubos)
• 75 µL de agua miliQ.
• 10 µL de regulador de PCR 10 x

17
• 3 µL de MgCl2 50 mM
• 2 µL de dNTP (10 mM de cada uno de dTTP, dGTP, dCTP y dATP).
• 6 µL de iniciadores específicos fD1 (3 µL) y rD1 (3 µL) hasta la concentración final
de 12 pmol cada uno.
• 2 µL de DNA Taq polimerasa hasta una concentración final de 10 U por 100 µL.
• Colocar 49 µL de la mezcla de reacción en cada tubo de PCR y adicionar 1 µL (0.4-4
ng) de DNA extraído, para tener un volumen total de 50 µL por tubo.
• Colocar un testigo negativo con 1 µL de agua miliQ en vez del DNA extraído.
La amplificación se lleva a cabo en un termociclador mediante un paso inicial de
desnaturalización de 95º C durante 3 min, seguido por 35 ciclos de 94º C por 1 min, 57º C por
1 min y 72º C por 2 min, seguidos por un paso final de extensión de 72º C por 3 minutos,
llevando la muestra a una temperatura final de 10º C.
Para verificar el éxito de la amplificación se lleva a cabo una electroforesis en gel de agarosa
al 1% 40 min a 100 V con marcador de 100 pb para verificar el tamaño de los amplificados.
Una vez obtenidos los amplificados se lleva a cabo un paso de purificación con el sistema de
purificación rápida de amplificados de PCR , de Marligen, sigiendo las indicaciones del
fabricante
Amplificación de los genes nifH y nodA.(Eardly y col. 1985), (Haukka y col. 1997)
Para llevar a cabo la amplificación se prepara la mezcla de reacción de la misma manera que
para el gen 16S rRNA, pero utilizando los iniciadores específicos nodA1 y nodA2, para el
gen nodA y nifH1 y nifH2 para el gen nifH bajo las siguientes condiciones: nifH: un paso
inicial de desnaturalización de 94° C por 2 min, seguido por 35 ciclos de 94° C por 30 s, 62°
C por 30 s, y 72° C por 1 min, seguidos por un paso final de extensión de 72° C por 5 min,
llevando la muestra a temperatura final de 10° C. Para el gen nodA: un paso inicial de
desnaturalización de 94° C por 5 min, seguido por 35 ciclos de 94° C por 45 s, 49° C por 1
min y 72° C por 2 min, seguidos por un paso final de extensión de 72° C por 5 min, llevando
la muestra a una temperatura final de 10° C.

18
Clonación de los amplificados.
Los amplificados se clonan utilizando el plásmido pCR2.1 - TOPO de 3.9 Kpb como vector
para transformar la cepa TOP10 de E. coli (químicamente competente) empleando el kit de
clonación TOPO- TA de Invitrogen de acuerdo a las indicaciones del fabricante, las clonas
obtenidas se inoculan en el medio de Luria Bertani con 50 µg/mL de Kanamicina y se realiza
la extracción de plásmidos por el método de lisis alcalina según el procedimiento reportado
en el Manual de Clonación Molecular de Sambrook y Russell.
Caracterización de las clonas por análisis de fragmentos de restricción de tamaño
polimórfico.
Una vez que se haya realizado la extracción de los plásmidos estos se digieren con enzimas
de restricción como a continuación se describe:
• 10 µL de muestra (4.4 ng)
• 2 µL de regulador de enzimas de restricción 10 x
• 0.5 µL de enzima (5 U/µL)
• 7.5 µL de agua miliQ
Se utilizan las enzimas MspI, HindI, HhaI, y XbaI, pues son de corte frecuente ya que cortan
fragmentos de 4 pb y ofrecen buenos result ados para este trabajo, se corren en gel de agarosa
al 3% a 100 V durante 90 min, para verificar los patrones de RFLP.
Secuenciación y análisis filogenético.
Se lleva a cabo la secuenciación de las clonas que tengan diferentes patrones de RFLP para
después realizar la construcción de un árbol filogenético utilizando el programa Blast para
buscar las bacterias más relacionadas en la base de dato. Las secuencias obtenidas en el
trabajo, las de bacterias más relacionadas y las de aislados del Conzattia multiflora (Wang y
col. Dato no publicado) se van a usar en análisis por el programa Clustal W para producir un
árbol que demuestre la diversidad de estas bacterias y ayude la identificación de ellas.
RESULTADOS

19
Muestreo de nódulos: Se llevaron a cabo dos muestreos, uno en julio, cuando las bacterias
están en fase vegetativa (Fig. 4) y otro en octubre, cuando el árbol comienza a dar fruto.
Obteniéndose en cada uno de unas 10 a 15 muestras de nódulos de diferentes árboles de la
región, el criterio para escogerlos fue tamaño y forma del nódulo. Las muestras obtenidas en
julio resultaron mejores para el trabajo ya que presentaban mayor cantidad de DNA, mientras
que las obtenidas en octubre casi no lo presentaron.
Extracción de DNA total: Se realizaron bastantes ensayos con el método de aislamiento de
Holme combinado con el proceso de Vincent hasta obtener aislados de calidad aceptable.
Después del aislamiento fue necesario llevar a cabo un paso de purificación con el kit
comercial de purificación de DNA de Promega, siguiendo las indicaciones del fabricante, ya
que contenían impurezas. De los resultados podemos decir que la cantidad obtenida es poca
debido al proceso de purificación pero si este no se lleva a cabo no es posible obtener
amplificados (Fig. 5).
Amplificación de los genes 16S rRNA, nifH y nodA: En la amplificación del gen 16S se
obtuvieron fragmentos del tamaño de 1500 bp, el cual corresponde al gen de 16S rRNA (Fig.
6).
Para la amplificación de los genes de nodulación y fijación de nitrógeno se amplificó el DNA
obtenido con dos testigos positivos de cepas de Rhizobium caracterizadas como buenas
productoras de nódulos y fijadoras de nitrógeno por el equipo de trabajo del Dr. En Tao
Figura 4. Flor de Conzattia multiflora y su hábitat, las fotografías fueron tomadas en junio, cuando la
planta florea y las bacterias del nódulo se encuentran en fase de crecimiento vegetativo.
Figura 5. Fotografía que muestra el DNA metagenómico obtenido a partir de los aislados de nódulos de Conzattia multiflora . (De arriba hacia abajo: Pozos 1, 2 y 3: DNA metagenómico.) La muestra se procesó por electroforesis en gel de agarosa al 1%, durante 40 min a 100 V y se tiñó con bromuro de etidio (0.5 µg/ml).
1
2
3
Fotos eliminados
Foto eliminado

20
Wang Hu las cuales fueron las cepas Le39 y S182. No hubo indicios de que el gen de fijación
de nitrógeno se halle presente en las muestras de DNA obtenido, ya que no presenta ninguna
banda al analizar los amplificados por electroforesis en gel de agarosa, mientras la banda
correspondiente nifH se encontró en las cepas testigo (Fig. 7).
Sobre de los resultados de la amplificación del gen de nodulación podemos decir que se
obtuvieron indicios de que este podría estar presente en la muestra por el análisis en gel de
agarosa, por lo cual se decidió llevar a cabo una hibridación, en condiciones de baja
astringencia, del amplificado con una sonda marcada con enzima Klenow hecha a partir de el
amplificado de la cepa Le39. La hibridación fue realizada transfiriendo el DNA a una
membrana de nylon por el método de Southern de acuerdo a las indicaciones del manual de
clonación molecular de Sambrook y Russell y revelando con digoxigenina, utilizando el kit
de Roche y siguiendo las indicaciones del fabricante, para comprobar si existía homología
entre los amplificados, pero los resultados de la prueba no han sido concluyentes, por lo que
se seguirá intentando la hibridación (Fig. 8) .
Figura 7. Fotografía que mu estra los amplificados del los genes nifH. Pozo M: marcador de 100 pb. Pozo 1: cepa Le39. Pozo 2: cepa S182. Pozo 3: muestra problema. La muestra se proceso por electroforesis en gel de agarosa al 1%, durante 40 min a 100 V, y se tiñó con bromuro de etidio (0.5 µg/mL)
Figura 8. Fotografía que muestra el amplificado del gen de nodA. Pozo M: marcador de 100 pb; B: blanco; Pozo 1: amplificado de la cepa Le39; Pozo 2: amplificado de la cepa S182; Pozo 3: amplificado muestra problema. La muestra se proceso por electroforesis en gel de agarosa al 1%, durante 40 min a 100 V, y se tiñó con bromuro de etidio (0.5 µg/mL).
1 2 3
3 2 1
M
nodA
nifH
1500 bp
M 1 2 3 Figura 6. Fotografía que muestra los amplificados del gen 16S rRNA, obtenidos a partir del DNA metagenómico. (De arriba hacia abajo: Pozo 1: marcador de 100 pb; Pozos 2 y 3: Gen 16S rRNA amplificado; pozo 4: control negativo) la muestra se proceso por electroforesis en gel de agarosa al 1%, durante 40 min a 100 V, y se tiñó con bromuro de etidio (0.5 µg/mL)
M
M B
Foto eliminado
Foto eliminado

21
Clonación de los amplificados: Una vez obtenidos los amplificados se llevo a cabo el paso
de transformación de los fragmentos obtenidos en el vector pCR 2.1 TOPO, para después
transformar la cepa de E. coli TOP10 por métodos químicos siguiendo las indicaciones del
fabricante. Las cepas transformadas se sembraron en medio sólido de Luria Bertani
adicionado con IPTG, X–gal y kanamicina como marcador de resistencia a las
concentraciones recomendadas por el fabricante. Se picaron mil colonias blancas en sendas
cajas con medio sólido de LB con kanamicina y en este momento se está realizando la
extracción de plásmidos para posteriormente agrupar las clonas por sus patrones de RFLP y
realizar el análisis filogenético (Figura 9). En los primero 20 clonas, se encontraron 5 clonas
positivas, cuales tienen 2 patrones de restricción por EcoRI: 2 clonas solo presentaron una
banda, indicando que ellos no tienen el sitio de EcoRI en el gen de 16S rRNA; tres tienen 2
bandas, demostrando que hay un sitio para la dicha enzima.
DISCUSIÓN DE RESULTADOS Aislamiento y purificación de DNA metagenómico. Para realizar la extracción de DNA
metagenómico se realizaron diversos ensayos para determinar cual era el mejor método para
obtener buenos resultados, se probaron dos métodos reportados en la literatura: El de Araujo
y col (2002) y el de Holme, combinado con Vincent, después de varias pruebas se llego a la
conclusión de que lo mejor para obtener lisados bacterianos era una combinación entre los
métodos de Vincent y Araujo, pues la obtención del lisado es un paso crítico en la obtención
de DNA metagenómico. Para la obtención de DNA es mejor el método de Holme, pues
utiliza rompimiento de células con lisozima y proteinasa K, lo cual es mas eficiente que el
rompimiento mecánico del método de Araujo. Se hicieron pruebas con muestras obtenidas
en diferentes tiempos, las primeras en el mes de junio y las segundas en el mes de octubre,
Figura 9: Fotografía que muestra las primeras 20 clonas analizadas mediante el método de lisis alcalina. Se observan un total de 5 clonas positivas. Los fragmentos de inserto y del vector se indican con de la muestra fue procesada por electroforesis en gel de agarosa al 1% durante 90 min a 100 V y se tiñó con bromuro de etidio (0.5 µg/mL) Vector
inserto
Foto eliminado

22
siendo las mejores para nuestro trabajo las obtenidas en junio, ya que la cantidad de ADN
extraída en las muestras de octubre fue muy poca y de muy mala calidad. Después de la
extracción se llevaron a cabo varios ensayos de amplificación por PCR, los cuales fueron
infructuosos, por lo que se sometió a la muestra a un paso extra de purificación para poder
obtener amplificados, para lo cual se utilizó un el kit comercial de purificación de promega,
que se basa en elusiones de DNA en microcolumnas para su purificación. Con lo cual fue
posible obtener muestras amplificables.
La ausencia de nifH. Anteriormente, se encontró que las bacterias endofíticas aisladas de
Conzattia multiflora produjeron muy baja concentración de C2H4 bajo la condición de aire
con C2H2, cual es una reacción especifica a la enzima de nitrogenasa, la enzima de fijación de
nitrógeno. Pero no se detectó el gene de fijación de nitrógeno (nifH) por PCR. En el presente
trabajo, tampoco encontramos nifH en el DNA metagenómico, demostrando que no existen
bacterias fijadoras de nitrógeno en esta planta, porque los iniciadores usados son universales
para bacterias. En este caso, es posib le que C2H4 fue un producto metabólico de las bacterias
endofíticas.
El producto de nodA. Con respecto a la capacidad de nodulación, aún es necesario realizar
más estudios para poder afirmar con certeza si las bacterias dentro del nódulo son
responsables de la formación de este. Se han logrado obtener amplificados con iniciadores
específicos para los genes nodA y nodB los cuales codifican para la síntesis de factores de
nodulación (oligopolisacáridos) pero no podemos decir que el amplificado sea el gen de
nodulación por lo que se han llevado a cabo pruebas de hibridación para comprobar si existe
homología entre nuestro amplificado y un gen de nodulación testigo, pero hasta ahora los
resultados no son concluyentes, por lo que se seguirá tratando de hibridar para
posteriormente clonar y secuenciar el amplificado.
Selección de clonas positivas. Han sido analizadas hasta el momento veinte clonas de un
total de mil, encontrándose cinco clonas positivas, con las cuales se están llevando a cabo
estudios de RFLP para comprobar sus patrones de restricción y posteriormente secuenciar las
clonas representativas. El resultado de RFLP del enzima EcoRI, estas cinco clonas

23
representaron mínimo dos diferentes bacterias, indicando que se incluyen diferentes
bacterias en la biblioteca del gen de16S rRNA.
CONCLUSIONES Y PROSPECTOS
A partir de los resultados del trabajo experimental hasta ahora podemos concluir que:
1. En las muestras aisladas de DNA metagenómico no está presente el gen de fijación
de nitrógeno, por lo que las bacterias dentro del nódulo no tienen esta capacidad.
2. Se obtengan productos de PCR para el gen nodA. Es necesario realizar más estudios
para afirmar con certeza la existencia de este gen en las bacterias dentro del nódulo.
3. Se obtenga una biblioteca del gen de 16S rRNA, cual incluyen diferentes bacterias.
Se va a continuar con la selección y caracterización de las clonas para llevar a cabo la
secuenciación y análisis de diversidad de las bacterias endofíticas del árbol.
BIBLIOGRAFÍA
Allen, O. N. y Allen, E.K. 1981. The Leguminosae: a Source Book of Characteristics, Uses,
and Nodulation. Madison, WI: University of Wisconsin Press.
Araujo, L. W, Marcon, J, Maccheroni, W. Jr, van Elsas, J. D, van Vuurde, J. W. L. y
Azevedo, J. L. 2002. Diversity of endophytic bacterial populations and their interaction
with Xylela fastidiosa in citrus plant. Appl. Environ. Microbiol. 68: 4906 – 4914.
Bottini R, Cassan F, Piccoli P. 2004. Gibberellin production by bacteria and its involvement
in plant growth promotion and yield increase. Appl Microbiol Biotechnol. 65:497-503.
Caballero-Mellado J, Martinez-Aguilar L, Paredes-Valdez G, Santos PE. 2004 Burkholderia
unamae sp. nov., an N2-fixing rhizospheric and endophytic species. Int J Syst Evol

24
Microbiol. 54:1165-72.
Earldy, B. D, Hannaway, D. B, Bottomley, P. J,. 1985. Characterization of rhizobia from
ineffective alfalfa nodules: ability to nodulate bean plants [Phaseolus vulgaris (L.)
savi] Appl. Environ. Microbiol; 50: 1422 -1427
Estrada-De Los Santos P, Bustillos-Cristales R, Caballero-Mellado J. 2001. Burkholderia, a
genus rich in plant-associated nitrogen fixers with wide environmental and geographic
distribution. Appl Environ Microbiol. 67:2790-8.
Fuentes y col. 1993. Capítulo 13: Gluconobacter diazotrophicus, modelo de bacteria
endófita. FEMS Microbiol. Ecol. 29:117-128.
Fuentes-Ramirez LE, Bustillos-Cristales R, Tapia-Hernandez A, Jimenez-Salgado T, Wang
ET, Martinez-Romero E, Caballero-Mellado J. 2001. Novel nitrogen-fixing acetic acid
bacteria, Gluconacetobacter johannae sp. nov. and Gluconacetobacter azotocaptans
sp. nov., associated with coffee plants. Int J Syst Evol Microbiol. 51:1305-14.
Freeman, E M, 1904. Philos Trans. R. Soc. Lond. (Biol.), 196:1.
Garcia-Ochoa F, Santos VE, Casas JA, Gomez E. 2000. Xanthan gum: production, recovery,
and properties. Biotechnol Adv. 18:549-79.
Gutierrez-Zamora ML, Martinez-Romero E. 2001. Natural endophytic association between
Rhizobium etli and maize (Zea mays L.). J Biotechnol. 91:117-26.
Hallman, J., Quadt – Hallmann, A., Mahaffe, W. F and Kloepper, J. W. 1997. Bacterial
endophytes in agriculturals corps. Canadian Journal of Microbiology 43: 895 – 814.
Haukka K, Linndström K, Young J. P. W., 1997. Three phylogenetic groups of nodA and
nifH genes in Sinorhizobium and Mesorhizobium isolates from leguminous trees

25
growing in Africa and Latin America. Appl. Environ. Microbiol. 64: (419 – 426)
Holme, I. B. and K. K. Petersen. 1996. Callus induction and plant regeneration from different
explant types of Myscantus x ogiformis Honda “Giganteus”. Pl. Cell. Tiss. Org. Cult.
45: 43 – 52.
James, E. K. 2000. Nitrogen fixation in endophytic and associative symbiosis. Field Crops
Research 65: 197 – 209.
Jimenez-Salgado T, Fuentes-Ramirez LE, Tapia-Hernandez A, Mascarua-Esparza MA,
Martinez-Romero E, Caballero-Mellado J. 1997. Coffea arabica L., a new host plant
for Acetobacter diazotrophicus, and isolation of other nitrogen-fixing acetobacteria.
Appl Environ Microbiol. 63:3676-83.
Knösel DH. 1984. Genus IV. Phylobacterium (ex Knösel 1962) nom. rev. Phyllobacterium
Knösel (1962, 96). In: Bergey's Manual of Systematic Bacteriology, vol. 1. (Krieg, N. R. and
Holt, J. G., Eds.), pp. 234-254. The Williams & Wilkins Co., Baltimore.
Madigan MT, Martinko JM, Parker J. 2004. Brock. Biología de los Microorganismos.
Décima edición. pp. 6675-684. España
Mc Vaugh, R. 1984. Flora Novo - Galiciana: A descriptive account of the vascular plants of
western México, Vol 5: Leguminosae. Ann Arbor The University of Michigan Press.
pp 60 – 63.
Mittermeier, R. A., N. Meyers, P. R. Gil, and C. G. Mittermeier. 1999. Hotspots: Earth’ s
biologically richest and most endangered ecoregions. Topan Printing Co., Tokyo.

26
Muñoz, R. J., Caballero – Mellado, J. 2005. Microbios, CIFN Cápitulo 13: Gluconobacter
diazhotrophicus, modelo de bacteria endófita.
Newman LA, Reynolds CM. 2005. Bacteria and phytoremediation: new uses for endophytic
bacteria in plants. Trends Biotechnol. 23:6-9.
Paredes-Cardona E, Carcano-Montiel M, Mascarua-Esparza MA, Caballero-Mellado J.
1988. [Response of maize to inoculation with Azospirillum brasilense]. Rev Latinoam
Microbiol. 30:351-5.
Quispel A. 1992. A search for signals in endophytic microorganism, pp. 471–491. In Verma,
D. P. S. (ed.), Molecular signals in plant–microbe communications, CRC press, Boca
Ratón.
Reinhold–Hurek B, Hurek T. 1998. Life in grasses: diazotrophic endophytes. Trends
Microbiol. 6: 139 – 144.
Reis VM, Estrada-de los Santos P, Tenorio-Salgado S, Vogel J, Stoffels M, Guyon S,
Mavingui P, Baldani VL, Schmid M, Baldani JI, Balandreau J, Hartmann A,
Caballero-Mellado J. 2004. Burkholderia tropica sp. nov., a novel nitrogen-fixing,
plant-associated bacterium. Int J Syst Evol Microbiol. 54:2155-62.
Sambrook, J. Russell, D, W. 2001 Molecular Cloning a laboratory manual. Vol. 1. Chapter 1:
Plasmids and their usefulness in molecular cloning. Cold Spring Harbor Laboratory
Press. pp. 1.32 – 1.34.
Sengbusch VP. 2005. Interactions of plants and Bacteria–Genetic Engineering.
www.botanyonline.com.kl
Sessitsch A, Reiter B, Berg G. 2004. Endophytic bacterial communities of field-grown
potato plants and their plant-growth-promoting and antagonistic abilities. Can J

27
Microbiol. 50:239-49.
Strobel G, Daisy B. 2003. Bioprospecting for microbial endophytes and their natural
products. Microbiol. Molec. Biol. Rev. 67: 491–502.
Strobel GA, Long DM. 1998. Endophytic microbes embody pharmaceutical potential. ASM
News 64: 263–268.
Tapia-Hernandez A, Bustillos-Cristales MR, Jimenez-Salgado T, Caballero-Mellado J,
Fuentes-Ramirez LE. 2000. Natural endophytic occurrence of Acetobacter
diazotrophicus in pineapple plants. Microbiol Ecol. 39:49-55.
Temmerman W, Vereecke D, Dreesen R, Van Montagu M, Holsters M, Goethals K. 2000.
Leafy gall formation is controlled by fasR, an AraC-type regulatory gene in
Rhodococcus fascians. J Bacteriol. 182:5832-5840.
Van Berkum P., Beyene D., Eardy B. D. 1996. Phylogenetic relationship among Rhizobium
species nodulating the common bean (Phaseolus vulgaris). Int J Bacteriol.
46:240–244.
Van Oevelen S, De Wachter R, Vandamme P, Robbrecht E, Prinsen E. 2002. Identification
of the bacterila endosymbionts in leaf galls of Psychotria (Rubiaceae, angiosperms)
and proposal of 'Candidatus Burkholderia kirkii' sp. nov. Int J Syst Evol Microbiol.
52:2023-2027.
Van Oevelen S, De Wachter R, Vandamme P, Robbrecht E, Prinsen E. 2004. 'Candidatus
Burkholderia calva' and 'Candidatus Burkholderia nigropunctata' as leaf gall
endosymbionts of African Psychotria. Int J Syst Evol Microbiol. 54:2237-2239.
Vereecke D, Corneils K, Temmerman W, Jaziri M, Van Montagu M, Holsters M, Goethals

28
K. 2002. Chromosomal locus that affects pathogenicity of Rhodococcus fascians. J
Bacteriol. 184: 1112–1120.
Vincent, J. M. 1970. A Manual for the Practical Study of the Root – Nodule Bacteria.
Blackwell Scientific Publications, Oxford.
Wang ET, Martínez-Romero J, López, LI, 2001. 12: Rhizobium y su destacada simbiosis con
plantas. En Martínez-Romero (ed.) Microbios en línea. UNAM.
Weidern S, Arnold W, Pühler A, 1996. Diversity of uncultured microorganism associated
whit the seagrass Halopita stipulacea estimated by restriction fragment length
polymorphism analysis of PCR amplified 16S rRNA genes. Appl Environ Microbiol.
62: 766–771.
Weisburg WG, Barns SM, PelletierDA, Lane D. 1991. 16S Ribosomal DNA amplification
for phylogenetic study. J Bacteriol. 173:697–703.