enhancement of diepoxin ζ production in liquid culture of endophytic fungus berkleasmium sp. dzf12...
TRANSCRIPT

ORIGINAL PAPER
Enhancement of diepoxin f production in liquid cultureof endophytic fungus Berkleasmium sp. Dzf12 by polysaccharidesfrom its host plant Dioscorea zingiberensis
Yan Li • Peiqin Li • Yan Mou • Jianglin Zhao •
Tijiang Shan • Chunbang Ding • Ligang Zhou
Received: 4 September 2011 / Accepted: 1 November 2011 / Published online: 8 November 2011
� Springer Science+Business Media B.V. 2011
Abstract This study is the first report of the enhancement
of diepoxin f production in liquid culture of the endophytic
fungus Berkleasmium sp. Dzf12 by the polysaccha-
rides from its host plant Dioscorea zingiberensis which
serve as elicitors. Three polysaccharides, namely water-
extracted polysaccharide (WEP), sodium hydroxide-
extracted polysaccharide and acid-extracted polysaccharide
were sequentially prepared from the rhizomes of D. zing-
iberensis. Among them, WEP was found to be the most
effective elicitor to enhance diepoxin f production. When
WEP was added to the medium at 400 mg l-1 on day 3 of
culture, the maximal diepoxin f yield (intracellular diep-
oxin f in mycelia plus extracellular diepoxin f in medium)
of 350.76 mg l-1 on day 15 was achieved, which was
about 2.69-fold in comparison with that (130.43 mg l-1) of
the control.
Keywords Diepoxin f � Endophytic fungus
Berkleasmium sp. Dzf12 � Polysaccharide � Dioscorea
zingiberensis
Introduction
Spirobisnaphthalenes are a rapidly growing group of
naphthoquinone derivatives with interesting structures and
various biological activities such as antitumor, antibacterial,
antifungal, antileishmanial, enzyme-inhibitory, and other
properties to display their potential applications in agri-
culture, medicine and food industry (Krohn 2003; Cai et al.
2010; Zhou et al. 2010). The fungal metabolite diepoxin f(also namly palmarumycin C13, Sch 53514 and cladospi-
rone bisepoxide), a member of the spirobisnaphthalene
group, has been isolated from fungal species such as fungus
LL-07F725 associated with a tree growing in Panama
(Schlingmann et al. 1993), Nattrassia mangiferae (Chu
et al. 1994), Coniothyrium sp. (Krohn et al. 1994), Clado-
sporium sp. (Petersen et al. 1994), and endophytic fungus
Berkleasmium sp. Dzf12 (Cai et al. 2009). Diepoxin f was
examined to have strong antitumor activity in phospholi-
pase D (PLD) assay (IC50, 0.2 lM) (Schlingmann et al.
1993), against HT 1080 human fibro-sarcoma in the inva-
sion assay (IC50, 0.37 lM) (Chu et al. 1994), as well as
strong antibacterial activity (IC50, 5.0–12.5 lg ml-1) (Cai
et al. 2009). These tremendous discoveries about diepoxin fhave received a particular attention (Krohn et al. 1997;
Powis et al. 2006; Cai et al. 2010).
In our previous study, five spirobisnaphthalenes were
successfully obtained from the endophytic fungus Berk-
leasmium sp. Dzf12 associated with the healthy rhizomes of
a medicinal plant Dioscorea zingiberensis C. H. Wright,
and diepoxin f was found to be the most abundant and
exhibit obvious antimicrobial activity among them (Cai
et al. 2009; Zhang et al. 2010; Zhao et al. 2011a, b). In order
to speed up its application, increased diepoxin f yield in
fermentation culture of Berkleasmium sp. Dzf12 is required.
Many strategies (i.e., medium optimization, elicitation by
using polysaccharides and oligosaccharides, as well as two-
phase culture) to enhance the production of bioactive
compounds in either microorganism or plant cultures have
been well developed. Among them, elicitation was regarded
Y. Li � C. Ding
College of Biology and Science, Sichuan Agricultural
University, Yaan 625014, China
Y. Li � P. Li � Y. Mou � J. Zhao � T. Shan � L. Zhou (&)
Department of Plant Pathology, College of Agronomy and
Biotechnology, China Agricultural University,
Beijing 100193, China
e-mail: [email protected]
123
World J Microbiol Biotechnol (2012) 28:1407–1413
DOI 10.1007/s11274-011-0940-y

as a convenient and effective approach (Zhou and Wu 2006;
Zhao et al. 2010; Xu et al. 2009a, 2011).
Carbohydrates (i.e., polysaccharides and oligosaccha-
rides) have been widely regarded as the preferable elicitors
for overproduction of secondary metabolites in plant cell
cultures (Dornenburg and Knorr 1994; Zhou et al. 2003,
2007; Zhang et al. 2009). However, their use for enhancing
production of secondary metabolites in fungal cultures has
been rarely reported. The oligosaccharides prepared from
sodium alginate by partial acid hydrolysis were found to
cause significant increase of penicillin G production in
cultures of Penicillium chrysogenum (Ariyo et al. 1997).
Further investigation showed that the stimulation of peni-
cillin G production in cultures of P. chrysogenum by both
alginate and alginate-derived oligosaccharides was derived
from an increase in the transcriptional activity of the
penicillin biosynthesis genes (i.e., pcbAB, pcbC, and
penDE) (Liu et al. 2001). In our previous study, an obvious
stimulatory effect on diepoxin f accumulation in liquid
culture of Berkleasmium sp. Dzf12 was observed, when the
yeast polysaccharide fraction was applied to the medium at
0.5 g l-1 on day 3 of culture (Zhao et al. 2011a). To the
best of our knowledge, there are no previous reports on the
effects of host plant polysaccharide elicitors on secondary
metabolite accumulation of its endophytic fungus. The
purpose of this study was to investigate the enhancement of
diepoxin f production in liquid culture of endophytic fun-
gus Berkleasmium sp. Dzf12 by polysaccharides from its
host plant D. zingiberensis.
Materials and methods
Endophytic fungus and culture conditions
The endophytic fungus Berkleasmium sp. Dzf12 (GenBank
accession number EF543255) was isolated from healthy
rhizomes of the medicinal plant D. zingiberensis C.
H. Wright (Dioscoreaceae) in our previous study (Cai et al.
2009; Zhang et al. 2010). The living culture has been
deposited in the China General Microbiological Culture
Collection Center (CGMCC) under the number CGMCC
2476. It was also maintained on potato dextrose agar
(PDA) slants at 25�C, and in 40% glycerol at -70�C at the
Herbarium of the College of Agronomy and Biotechnol-
ogy, China Agricultural University. Liquid culture experi-
ments were carried out in 150-ml Erlenmeyer flasks, each
of which was filled with 30 ml of liquid medium, and
maintained on a rotary shaker at 150 rev min-1 and 25�C.
The liquid medium was composed of (l-1): 40 g glucose,
10 g peptone, 1.0 g KH2PO4, 0.5 g MgSO4�7H2O, 0.05 g
FeSO4�7H2O, pH 6.5, a formulation based on Sabouraud
broth medium (Zhao et al. 2011a). The inoculum for the
shake-flask culture was prepared by shaking incubation of
the mycelia from the solid stock culture in potato dextrose
broth for 4 days, and 0.9 ml of mycelial broth was inocu-
lated to each flask (3.0%, v/v).
Preparation of polysaccharides
The polysaccharides obtained from the rhizomes of
D. zingiberensis were prepared according to the method of
Lazaridou et al. (2008) with some modifications. Briefly,
the dried and powdered rhizomes (800 g) of D. zingiber-
ensis were refluxed three times with 95% ethanol at 60�C to
remove lipids. The defatted powder was obtained by drying
in air, and then extracted three times with hot water at 80�C
for 2 h with the ratio of the material (g) to water (ml) as 1:15
(w/v), which was then filtered through a muslin cloth. The
filtrate was collected and concentrated to a certain volume
under vacuum at 60�C, and mixed with three volumes of
95% ethanol, then kept at 4�C overnight. The precipitate
was collected by centrifugation, washed with ethanol first,
and then with diethyl ether. The polysaccharide obtained
(45.60 g) was named as crude water-extracted polysac-
charide (WEP). The residue not containing WEP was fur-
ther extracted with 1 M sodium hydroxide (NaOH) solution
at room temperature for 24 h. The remaining steps were the
same as the treatments of WEP. The obtained polysaccha-
ride (34.80 g) was designated as crude sodium hydroxide-
extracted polysaccharide (SEP). The residue not containing
WEP and SEP was further extracted with 1 M HCl solution
at room temperature for 24 h. After ethanol precipitation,
the resulting polysaccharide (33.36 g) was named as crude
acid-extracted polysaccharide (AEP). Each crude polysac-
charide was treated with the Sevag reagent (n-butanol-
chloroform = 1:4, v/v) for deproteination (Xu et al. 2009b),
and then treated with 3% H2O2 for decoloration to obtain
the pure polysaccharide fractions of WEP, SEP and AEP,
which were freeze-dried and stored in a desiccator at room
temperature. The carbohydrate content of each polysac-
charide as well as glucose consumption in medium was
measured by the method of anthrone-sulfuric acid spectro-
photography (Zhao et al. 2010), which involved sulfuric
acid hydrolysis of the sample in the presence of anthrone
reagent at 100�C. The absorbance at 620 nm was measured
and calibrated to carbohydrate content using glucose as a
reference. The carbohydrate content of WEP, SEP and AEP
was determined as 81, 72 and 76%, respectively.
Application of polysaccharides
Stock solutions (100 mg carbohydrate equivalent per mil-
liliter) were prepared by dissolving each polysaccharide
(i.e., WEP, SEP and AEP) in distilled water, and the pH
was adjusted to 6.5. The solutions were sterilized by
1408 World J Microbiol Biotechnol (2012) 28:1407–1413
123

filtering through a microfilter (0.45 lm), diluted with
sterile water into different concentrations, and then stored
at 4�C before use. Each polysaccharide was applied to the
medium with final concentrations of 100, 400 and 800 mg
carbohydrate equivalent per liter in the medium, respec-
tively. WEP, which had been screened to be the most
effective polysaccharide (data shown in Table 1), was
added to the liquid medium of Berkleasmium sp. Dzf12 on
days 0, 3, 6, 9 and 12 in combination with its final con-
centrations of 100, 200, 400, 600 and 800 mg l-l in the
medium, respectively. In order to study the kinetics of the
mycelial growth and diepoxin f accumulation in liquid
culture of Berkleasmium sp. Dzf12, WEP was treated at
400 mg l-1 on day 3 of culture. The mycelial suspension
cultures were harvested every 2 days after addition of
WEP.
Measurement of biomass and diepoxin f content
The mycelia of Berkleasmium sp. Dzf12 were separated
from the liquid medium by filtration under vacuum and
rinsed thoroughly with distilled water, and then dried at
50–55�C in an oven to constant dry weight (dw). Diepoxin
f extraction and determination was carried out as previ-
ously described (Zhao et al. 2011a, b). In brief, diepoxin fwas extracted from the dry mycelial powder (10 mg ml-1)
with methanol-chloroform (9:1, v/v) in an ultrasonic bath
(three times, 60 min each). After removal of the solid by
filtration, the filtrate was evaporated to dryness and re-
dissolved in 1 ml of methanol. For analysis of diepoxin fquantity in the culture broth, the sample was obtained by
evaporating 5 ml of the culture broth to dryness, extracting
with 5 ml methanol-chloroform (9:1, v/v) in an ultrasonic
bath (three times, 60 min each), then evaporating the liquid
extract to dryness, and re-dissolving in 1 ml of methanol.
Diepoxin f content was analysed by HPLC (Shimadzu,
Japan), which consisted of two LC-20AT solvent delivery
units, an SIL-20A autosampler, an SPD-M20A photodi-
ode array detector, and CBM-20Alite system controller.
The reversed-phase Agilent TC-C18 column (250 mm 9
4.6 mm i.d., particle size 5 lm) was used for separation by
using a mobile phase of methanol-H2O (50:50, v/v) at a
flow rate of 1 ml min-1. The temperature was maintained
at 40�C, and UV detection at 226 nm. The sample injection
volume was 10 ll. The LCsolution multi-PDA workstation
was employed to acquire and process chromatographic
data. The diepoxin f was detected and quantified with the
standard obtained from our previous study, which was
identified according to its physicochemical and spectro-
metric data (Cai et al. 2010; Krohn et al. 1994). According
to the method of Zhu et al. (2008), mycelial growth against
glucose consumption was calculated as: (maximum
mycelia biomass—initial mycelia biomass)/(initial glucose
concentration—residual glucose concentration when the
maximal mycelia biomass obtained). Diepoxin f yield
against glucose consumption was calculated as: (maximum
total diepoxin f yield—initial total diepoxin f yield)/(initial
glucose concentration—residual glucose concentration
when the maximal total diepoxin f yield obtained).
Statistical analysis
All experiments were repeated three times, and the results
were represented by their mean values and the standard
deviations (SD). The data were submitted to analysis of
variance (one-way ANOVA) to detect significant differ-
ences by PROC ANOVA of SAS version 8.2. The term
significant has been used to denote the differences for
which P B 0.05.
Table 1 Effects of the polysaccharides WEP, SEP and AEP on mycelial growth and diepoxin f production in liquid culture of Berkleasmium sp.
Dzf12
Treatment Polysaccharide
concentration (mg l-1)
Mycelial biomass
(g l-1)
Diepoxin f content in
mycelia (mg g-1)
Diepoxin f yield in
medium (mg l-1)
Total diepoxin f yield
(mg l-1)
Control 0 6.97 ± 0.32 d 3.45 ± 0.18 e 101.45 ± 3.10 g 125.50 ± 5.17 g
WEP 100 9.18 ± 0.38 b 4.05 ± 0.20 d 127.46 ± 4.12 f 164.64 ± 6.37 e
400 11.70 ± 0.35 a 7.05 ± 0.34 a 236.61 ± 3.57 a 319.10 ± 9.74 a
800 11.41 ± 0.11 a 5.17 ± 0.23 c 184.42 ± 6.07 c 243.41 ± 6.07 b
SEP 100 7.69 ± 0.24 cd 4.25 ± 0.21 d 202.30 ± 4.95 b 234.98 ± 8.12 bc
400 8.05 ± 0.20 c 5.19 ± 0.29 c 172.52 ± 3.63 d 214.30 ± 8.63 d
800 7.82 ± 0.35 cd 3.80 ± 0.25 de 104.72 ± 5.06 g 134.44 ± 7.47 f
AEP 100 7.25 ± 0.36 d 3.88 ± 0.14 de 107.30 ± 3.33 g 135.43 ± 5.17 f
400 8.84 ± 0.48 b 6.21 ± 0.28 b 175.68 ± 6.96 cd 230.58 ± 9.17 c
800 7.15 ± 0.13 d 3.96 ± 0.13 d 140.79 ± 5.36 e 169.10 ± 7.04 e
The polysaccharides were applied to the medium on day 3. The period of culture was in 13 days. The values represented M ± SD (n = 3).
Different letters indicated significant differences among the treatments in each column at P = 0.05 level
World J Microbiol Biotechnol (2012) 28:1407–1413 1409
123

Results
Effects of the polysaccharides WEP, SEP and AEP
on mycelial growth and diepoxin f production
The effects of three polysaccharides (i.e., WEP, SEP and
AEP) on mycelial growth and diepoxin f production in
liquid culture of Berkeasmium sp. Dzf12 are shown in
Table 1. All three polysaccharide enhanced mycelia
growth and diepoxin f production at concentration ranges
of 100, 400 and 800 mg l-1. The appropriate concentra-
tions for WEP, SEP and AEP to notably stimulate mycelia
growth were all at 400 mg l-1. Correspondingly, appro-
priate concentration for WEP, SEP and AEP to obviously
increase diepoxin f accumulation was at 400, 100 and
400 mg l-1, respectively. Among three polysaccharides,
WEP was the most effective elicitor to stimulate mycelia
growth and diepoxin f production. Treated with WEP at
400 mg l-1, the total diepoxin f yield (intracellular diep-
oxin f in mycelia plus extracellular diepoxin f in medium)
of Berkeasmium sp. Dzf12 was increased to 319.10 mg l-1,
about 2.54-fold in comparison with that (125.50 mg l-1) of
the control. Therefore, WEP was selected for further
studying its enhancement effects.
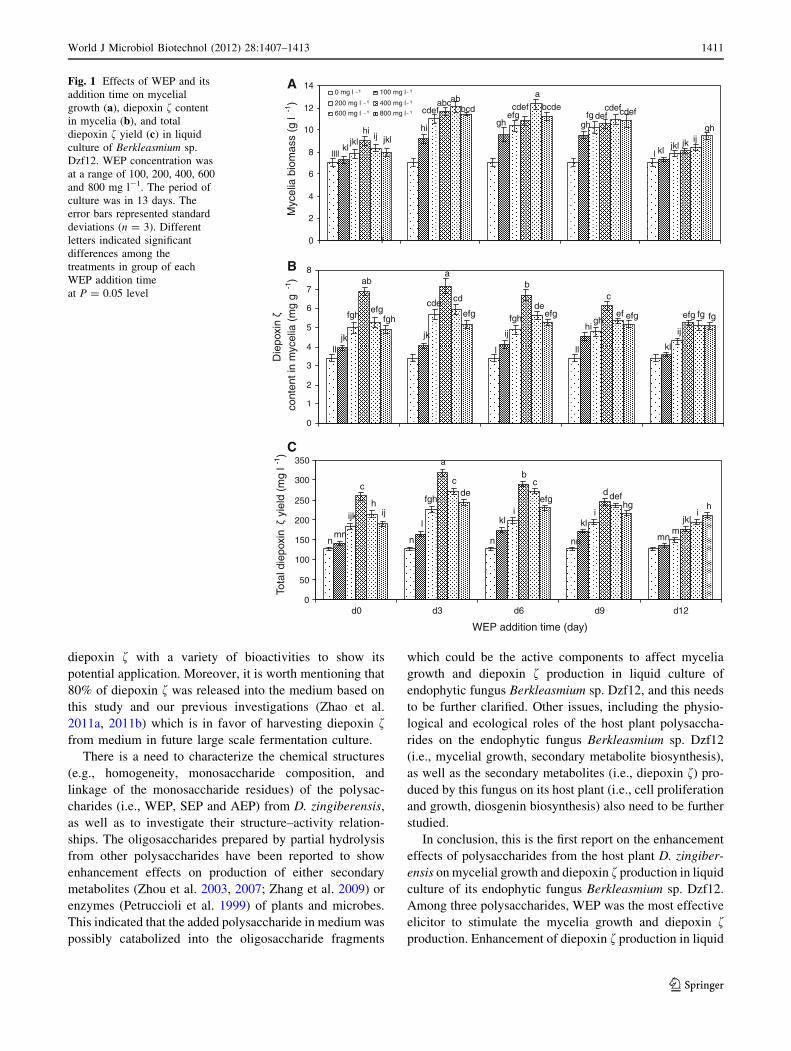
Effects of WEP and its addition time on mycelia growth
and diepoxin f production
WEP at 100, 200, 400, 600 and 800 mg l-1 was added to
liquid medium of Berkleasmium sp. Dzf12 on days 0, 3, 6,
9 and 12, respectively. Effects of WEP on mycelial growth,
diepoxin f content in mycelia, and diepoxin f yield of
Berkleasmium sp. Dzf12 liquid cultures are shown in
Fig. 1. When the cultures were fed with 600 mg of WEP
l-1 on day 6, the mycelia biomass was increased 1.75-fold
in comparison with that of the control (12.33 g l-1 versus
7.06 g l-1) (Fig. 1a). The diepoxin f production was also
effectively enhanced by WEP. With 400 mg of WEP l-1
fed on day 3, the highest content (7.17 mg g-1) of diepoxin
f in mycelia was obtained, which was 2.11-fold, compared
to that (3.40 mg g-1) of the control (Fig. 1b), and the total
diepoxin f yield (intracellular diepoxin f in mycelia plus
extracellular diepoxin f in medium) was as much as
318.41 mg l-1, about 2.50-fold in comparison with that
(127.42 mg l-1) of the control (Fig. 1c).
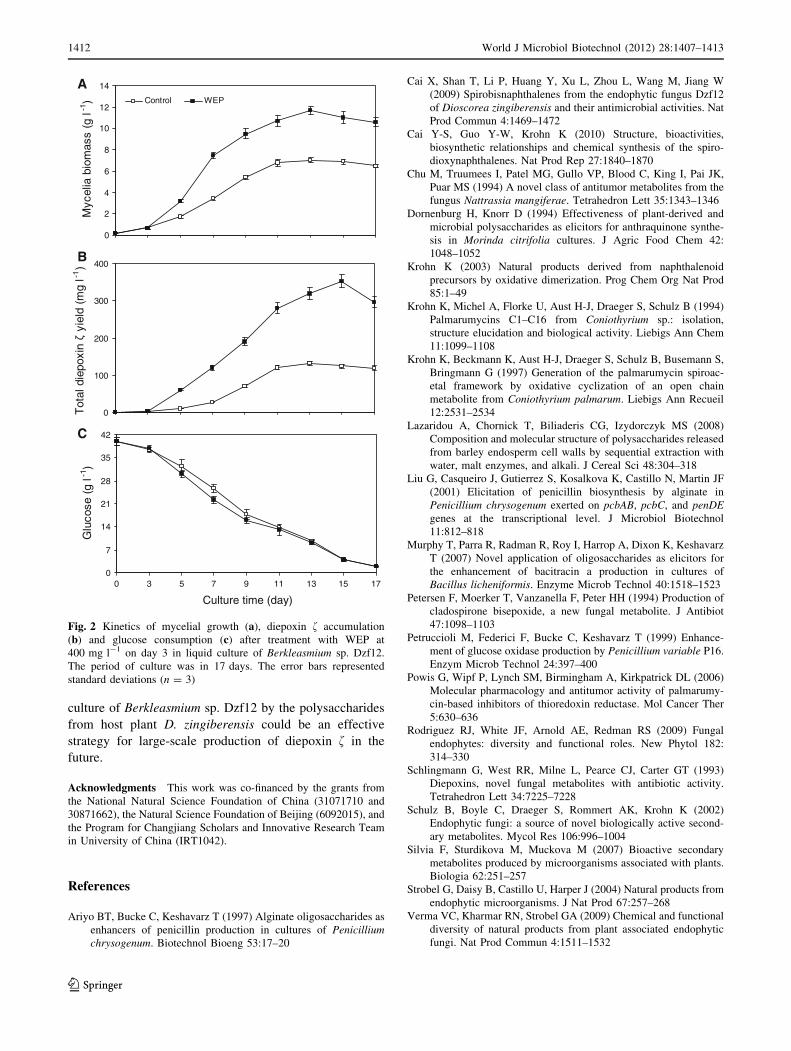
Kinetics of the mycelia growth, diepoxin faccumulation and glucose consumption after treatment
with WEP
Based on the above results (shown in Fig. 1), kinetic studies
of mycelial growth, diepoxin f accumulation and glucose
consumption after feeding with WEP at 400 mg l-1 on day
3 in liquid culture of Berkleasmium sp. Dzf12 are shown in
Fig. 2. Mycelial dry weight and diepoxin f yield of Berk-
leasmium sp. Dzf12 treated with WEP were always higher
than those of the control during the days after elicitation.
The enhancing effects of WEP on mycelial growth and
diepoxin f production of Berkleasmium sp. Dzf12 could be
observed on day 5, and then followed a steady increase. The
highest mycelial biomass was 11.66 g l-1 obtained on day
13, about 1.66-fold in comparison with that (7.04 g l-1) of
the control (Fig. 2a), and total diepoxin f yield
(350.76 mg l-1) reached its maximum on day 15 of culture,
about 2.69-fold in comparison with that (130.43 mg l-1) of
the control (Fig. 2b). The glucose consumption pattern of
the cultures treated with WEP was quite similar to the
control (Fig. 2c). For the WEP-treated cultures, the glucose
was consumed at a little higher rate on day 5, and the trend
maintained till day 11 in comparison with the control. The
mycelia growth against glucose consumption was
0.23 g g-1 for the control, and 0.37 g g-1 for the WEP-
treated cultures, and the corresponding total diepoxin fyield on glucose consumption was 4.32 mg g-1 (for the
control) and 9.71 mg g-1 (for the WEP treated cultures). It
concluded that the increase in both mycelial growth and
diepoxin f yield was due to the elicitation of WEP, which
was added to the cultures in trace amounts (mg l-1) and
assumed not to be used as carbon source (Ariyo et al. 1997;
Murphy et al. 2007).
Discussion
Plant endophytic fungi are the fungal microorganisms that
live asymptomatically within plant tissues (Zhang et al.
2006; Rodriguez et al. 2009). During the long period of co-
evolution, a friendly relationship has developed between
each endophyte and its host plant (Zhao et al. 2011c). The
host plant can supply plenteous nutriment and a safe hab-
itation for the survival of its endophytes. On the other hand,
the endophytes would produce a number of bioactive
constituents for helping their host plants to resist external
biotic and abiotic stresses, and benefiting for the host
growth (Silvia et al. 2007). The results from the present
study show that polysaccharides produced by D. zingiber-
ensis notably stimulated the mycelia growth and diepoxin fproduction of its endophytic fungus Berkleasmium sp.
Dzf12. It indicated that there should be a close relationship
between them which needs further investigation. Further-
more, plant endophytic fungi have been regarded as
important sources of natural bioactive products, and have
been proved to be useful in human medicine, agriculture
and food industry (Schulz et al. 2002; Strobel et al. 2004;
Verma et al. 2009; Zhou et al. 2010; Zhao et al. 2011c).
Berkleasmium sp. Dzf12 is a high yield strain for producing
1410 World J Microbiol Biotechnol (2012) 28:1407–1413
123
diepoxin f with a variety of bioactivities to show its
potential application. Moreover, it is worth mentioning that
80% of diepoxin f was released into the medium based on
this study and our previous investigations (Zhao et al.
2011a, 2011b) which is in favor of harvesting diepoxin ffrom medium in future large scale fermentation culture.
There is a need to characterize the chemical structures
(e.g., homogeneity, monosaccharide composition, and
linkage of the monosaccharide residues) of the polysac-
charides (i.e., WEP, SEP and AEP) from D. zingiberensis,
as well as to investigate their structure–activity relation-
ships. The oligosaccharides prepared by partial hydrolysis
from other polysaccharides have been reported to show
enhancement effects on production of either secondary
metabolites (Zhou et al. 2003, 2007; Zhang et al. 2009) or
enzymes (Petruccioli et al. 1999) of plants and microbes.
This indicated that the added polysaccharide in medium was
possibly catabolized into the oligosaccharide fragments
which could be the active components to affect mycelia
growth and diepoxin f production in liquid culture of
endophytic fungus Berkleasmium sp. Dzf12, and this needs
to be further clarified. Other issues, including the physio-
logical and ecological roles of the host plant polysaccha-
rides on the endophytic fungus Berkleasmium sp. Dzf12
(i.e., mycelial growth, secondary metabolite biosynthesis),
as well as the secondary metabolites (i.e., diepoxin f) pro-
duced by this fungus on its host plant (i.e., cell proliferation
and growth, diosgenin biosynthesis) also need to be further
studied.
In conclusion, this is the first report on the enhancement
effects of polysaccharides from the host plant D. zingiber-
ensis on mycelial growth and diepoxin f production in liquid
culture of its endophytic fungus Berkleasmium sp. Dzf12.
Among three polysaccharides, WEP was the most effective
elicitor to stimulate the mycelia growth and diepoxin fproduction. Enhancement of diepoxin f production in liquid
0
2
4
6
8
10
12
14
Myc
elia
bio
mas
s (g
l -1
)
0 mg l ¹ 100 mg l ¹
200 mg l ¹ 400 mg l ¹
600 mg l ¹ 800 mg l ¹
a
kl
l
llllkl
high gh
ljkl
cdef fg
jkl
hiij jkl
abc
def
jk
cdefbcdebcd
ij
cdefcdefab
gh
efg
0
1
2
3
4
5
6
7
8D
iepo
xin
con
tent
in m
ycel
ia (
mg
g -1
)
efg
ab
jk
efgefgfgh
ab
c
fgefefg
cdde
efgcde
fgfgh
ij
ghijjk
fghhi
klll l ll
0
50
100
150
200
250
300
350
d0 d3 d6 d9 d12
WEP addition time (day)
Tota
l die
poxi
n y
ield
(m
g l -1
)
mn
ijjkl
i
cc
h
ab
d
i
defefgfgh
c
hgde
hijk i
ml kl kl
mnn n n nn
B
A
C
Fig. 1 Effects of WEP and its
addition time on mycelial
growth (a), diepoxin f content
in mycelia (b), and total
diepoxin f yield (c) in liquid
culture of Berkleasmium sp.
Dzf12. WEP concentration was
at a range of 100, 200, 400, 600
and 800 mg l-1. The period of
culture was in 13 days. The
error bars represented standard
deviations (n = 3). Different
letters indicated significant
differences among the
treatments in group of each
WEP addition time
at P = 0.05 level
World J Microbiol Biotechnol (2012) 28:1407–1413 1411
123
culture of Berkleasmium sp. Dzf12 by the polysaccharides
from host plant D. zingiberensis could be an effective
strategy for large-scale production of diepoxin f in the
future.
Acknowledgments This work was co-financed by the grants from
the National Natural Science Foundation of China (31071710 and
30871662), the Natural Science Foundation of Beijing (6092015), and
the Program for Changjiang Scholars and Innovative Research Team
in University of China (IRT1042).
References
Ariyo BT, Bucke C, Keshavarz T (1997) Alginate oligosaccharides as
enhancers of penicillin production in cultures of Penicilliumchrysogenum. Biotechnol Bioeng 53:17–20
Cai X, Shan T, Li P, Huang Y, Xu L, Zhou L, Wang M, Jiang W
(2009) Spirobisnaphthalenes from the endophytic fungus Dzf12
of Dioscorea zingiberensis and their antimicrobial activities. Nat
Prod Commun 4:1469–1472
Cai Y-S, Guo Y-W, Krohn K (2010) Structure, bioactivities,
biosynthetic relationships and chemical synthesis of the spiro-
dioxynaphthalenes. Nat Prod Rep 27:1840–1870
Chu M, Truumees I, Patel MG, Gullo VP, Blood C, King I, Pai JK,
Puar MS (1994) A novel class of antitumor metabolites from the
fungus Nattrassia mangiferae. Tetrahedron Lett 35:1343–1346
Dornenburg H, Knorr D (1994) Effectiveness of plant-derived and
microbial polysaccharides as elicitors for anthraquinone synthe-
sis in Morinda citrifolia cultures. J Agric Food Chem 42:
1048–1052
Krohn K (2003) Natural products derived from naphthalenoid
precursors by oxidative dimerization. Prog Chem Org Nat Prod
85:1–49
Krohn K, Michel A, Florke U, Aust H-J, Draeger S, Schulz B (1994)
Palmarumycins C1–C16 from Coniothyrium sp.: isolation,
structure elucidation and biological activity. Liebigs Ann Chem
11:1099–1108
Krohn K, Beckmann K, Aust H-J, Draeger S, Schulz B, Busemann S,
Bringmann G (1997) Generation of the palmarumycin spiroac-
etal framework by oxidative cyclization of an open chain
metabolite from Coniothyrium palmarum. Liebigs Ann Recueil
12:2531–2534
Lazaridou A, Chornick T, Biliaderis CG, Izydorczyk MS (2008)
Composition and molecular structure of polysaccharides released
from barley endosperm cell walls by sequential extraction with
water, malt enzymes, and alkali. J Cereal Sci 48:304–318
Liu G, Casqueiro J, Gutierrez S, Kosalkova K, Castillo N, Martin JF
(2001) Elicitation of penicillin biosynthesis by alginate in
Penicillium chrysogenum exerted on pcbAB, pcbC, and penDEgenes at the transcriptional level. J Microbiol Biotechnol
11:812–818
Murphy T, Parra R, Radman R, Roy I, Harrop A, Dixon K, Keshavarz
T (2007) Novel application of oligosaccharides as elicitors for
the enhancement of bacitracin a production in cultures of
Bacillus licheniformis. Enzyme Microb Technol 40:1518–1523
Petersen F, Moerker T, Vanzanella F, Peter HH (1994) Production of
cladospirone bisepoxide, a new fungal metabolite. J Antibiot
47:1098–1103
Petruccioli M, Federici F, Bucke C, Keshavarz T (1999) Enhance-
ment of glucose oxidase production by Penicillium variable P16.
Enzym Microb Technol 24:397–400
Powis G, Wipf P, Lynch SM, Birmingham A, Kirkpatrick DL (2006)
Molecular pharmacology and antitumor activity of palmarumy-
cin-based inhibitors of thioredoxin reductase. Mol Cancer Ther
5:630–636
Rodriguez RJ, White JF, Arnold AE, Redman RS (2009) Fungal
endophytes: diversity and functional roles. New Phytol 182:
314–330
Schlingmann G, West RR, Milne L, Pearce CJ, Carter GT (1993)
Diepoxins, novel fungal metabolites with antibiotic activity.
Tetrahedron Lett 34:7225–7228
Schulz B, Boyle C, Draeger S, Rommert AK, Krohn K (2002)
Endophytic fungi: a source of novel biologically active second-
ary metabolites. Mycol Res 106:996–1004
Silvia F, Sturdikova M, Muckova M (2007) Bioactive secondary
metabolites produced by microorganisms associated with plants.
Biologia 62:251–257
Strobel G, Daisy B, Castillo U, Harper J (2004) Natural products from
endophytic microorganisms. J Nat Prod 67:257–268
Verma VC, Kharmar RN, Strobel GA (2009) Chemical and functional
diversity of natural products from plant associated endophytic
fungi. Nat Prod Commun 4:1511–1532
0
2
4
6
8
10
12
14M
ycel
ia b
iom
ass
(g l
-1) Control WEP
0
100
200
300
400
Tot
al d
iepo
xin
yie
ld (
mg
l -1 )
0
7
14
21
28
35
42
0 3 5 7 9 11 13 15 17
Culture time (day)
Glu
cose
(g
l -1 )
A
C
B
Fig. 2 Kinetics of mycelial growth (a), diepoxin f accumulation
(b) and glucose consumption (c) after treatment with WEP at
400 mg l-1 on day 3 in liquid culture of Berkleasmium sp. Dzf12.
The period of culture was in 17 days. The error bars represented
standard deviations (n = 3)
1412 World J Microbiol Biotechnol (2012) 28:1407–1413
123

Xu L, Liu Y, Zhou L, Wu J (2009a) Enhanced beauvericin production
with in situ adsorption in mycelial liquid culture of Fusariumredolens Dzf2. Process Biochem 44:1063–1067
Xu W, Zhang F, Luo Y, Ma L, Kou X, Huang K (2009b) Antioxidant
activity of a water-soluble polysaccharide purified from Pteri-dium aquilinum. Carbohyd Res 344:217–222
Xu L, Liu Y, Zhou L, Wu J (2011) Modeling of Fusarium redolensDzf2 mycelial growth kinetics and optimal fed-batch fermenta-
tion for beauvericin production. J Ind Microbiol Biotechnol
38:1187–1192
Zhang HW, Song YC, Tan RX (2006) Biology and chemistry of
endophytes. Nat Prod Rep 23:753–771
Zhang R, Li P, Xu L, Chen Y, Sui P, Zhou L, Li J (2009)
Enhancement of diosgenin production in Diocorea zingiberensiscell culture by oligosaccharide elicitor from its endophytic
fungus Fusarium oxysporum Dzf17. Nat Prod Commun 4:
1459–1462
Zhang R, Li P, Zhao J, Yin C, Zhou L (2010) Endophytic fungi from
Dioscorea zingiberensis and their effects on the growth and
diosgenin production of the host plant cultures. Nat Prod Res
Dev 22:11–15
Zhao J, Zhou L, Wu J (2010) Effects of biotic and abiotic elicitors on
cell growth and tanshinone accumulation in Salvia miltiorrhizacell cultures. Appl Microbiol Biotechnol 87:137–144
Zhao J, Zheng B, Li Y, Shan T, Mou Y, Lu S, Li P, Zhou L (2011a)
Enhancement of diepoxin f production by yeast extract and its
fractions in liquid culture of Berkleasmium-like endophytic
fungus Dzf12 from Dioscorea zingiberensis. Molecules 16:
847–856
Zhao J, Li Y, Shan T, Mou Y, Zhou L (2011b) Enhancement of
diepoxin f production with in situ resin adsorption in mycelial
liquid culture of the endophytic fungus Berkleasmium sp. Dzf12
from Dioscorea zingiberensis. World J Microbiol Biotechnol.
doi:10.1007/s11274-011-0750-2
Zhao J, Shan T, Mou Y, Zhou L (2011c) Plant-derived bioactive
compounds produced by endophytic fungi. Mini-Rev Med Chem
11:159–168
Zhou L, Wu J (2006) Development and application of medicinal plant
tissue cultures for production of drugs and herbal medicinals in
China. Nat Prod Rep 23:789–810
Zhou L, Yang C, Li J, Wang S, Wu J (2003) Heptasaccharide and
octasaccharide isolated from Paris polyphylla var. yunnanensisand their plant growth-regulatory activity. Plant Sci 165:571–575
Zhou L, Cao X, Zhang R, Peng Y, Zhao S, Wu J (2007) Stimulation
of saponin production in Panax ginseng hairy roots by two
oligosaccharides from Paris polyphylla var. yunnanensis. Bio-
technol Lett 29:631–634
Zhou L, Zhao J, Shan T, Cai X, Peng Y (2010) Spirobisnaphthalenes
from fungi and their biological activities. Mini-Rev Med Chem
10:977–989
Zhu L-W, Zhong J–J, Tang Y-J (2008) Significance of fungal elicitors
on the production of ganoderic acid and Ganoderma polysac-
charides by the submerged culture of medicinal mushroom
Ganoderma lucidum. Process Biochem 43:1359–1370
World J Microbiol Biotechnol (2012) 28:1407–1413 1413
123