estudio sobre la variación somaclonal en chile habanero ... · sistemas de regeneración de chile...
TRANSCRIPT

POSGRADO EN CIENCIAS Y BIOTECNOLOGÍA DE PLANTAS
Estudio sobre la variación somaclonal en chile Habanero (Capsicum chinense Jacq.) y su
aplicación en la obtención de líneas tolerantes al estrés hídrico
Tesis que presenta para obtener el grado de Doctor en Ciencias
M. en C. Jericó Jabín Bello Bello
CENTRO DE INVESTIGACIÓN CIENTÍFICA DE YUCATÁN, A.C.
Mérida, Yucatán, México. 2010

1-

t CENTRO DE INVESTIGACIÓN CIENTÍFICA DE YUCAT ÁN A.C. ~ ~ POSGRADO EN CIENCIAS Y BIOTECNOLOGIA DE PLANTAS
RECONOCIMIENTO
Por medio de la presente, hago constar que el trabajo de tesis titulado:
"Estudio sobre la variación somaclonal en chile habanero (Capsicum
chinense Jacq.) y su aplicación en la obtención de líneas tolerantes al
estrés hídrico", fue realizado en los laboratorios de la Unidad de
Bioquímica y Biología Molecular de Plantas del Centro de Investigación
Científica de Yucatán, A.C. bajo la dirección de la Dra. Nancy Santana
Buzzy y co- dirección de la Dra. Lourdes Iglesias Andreu, dentro de la
Opción Biología Experimental, perteneciente al Programa de Posgrado en
Ciencias y Biotecnología de Plantas de este Centro.
Atentamente,
Director Académico


DECLARACIÓN DE PROPIEDAD
Declaro que la información contenida en la sección de Materiales y
Métodos Experimentales, los Resultados y Discusión de este documento
provienen de las actividades de experimentación realizadas durante el
período que se me asignó para desarrollar mi trabajo de tesis, en los
laboratorios del Centro de Investigación Científica de Yucatán y que dicha
información le pertenece en términos de la ley de propiedad industrial, por
lo que no me reservo ningún derecho sobre ello.
Atentamente
M. en C. Jericó Jabín Bello Bello


CONTENIDO
AGRADECIMIENTOS DICATORIA LISTA DE TABLAS LISTA DE FIGURAS LISTA DE ABREVIATURAS RESUMEN ABSTRACT INTRODUCCIÓN BIBLIOGRAFIA
Capitulo l. Antecedentes
1.1 1.1 .1 1.1.2 1.1.3 1.1.4
1.2 1.2.1 1.2.2 1.2.3
1.3 1.3.1 1.3.2 1.3.3 1.3.4 1.3.5 1.3.6
1.4 1.4.1 1.4.2 1.4.3
1.5 1.5.1 1.5.2 1.5.3
1.6 1.6.1 1.6.2 1.6.3
1.7
EL CHILE HABANERO Origen y distribución Descripción taxonómica Importancia económica Problemática: El chile habanero y el cambio climático
ESTRÉS ABIÓTICO EN PLANTAS Mecanismos de respuesta Tolerancia a estrés hídrico El papel de la prolina ante el estrés hídrico
EL CULTIVO in vitro Cultivo de Tejidos Vegetales Control del desarrollo in vitro Morfogénesis in vitro Organogénesis Embriogénesis somática Regeneración in vitro de chile (Capsicum spp.)
VARIACIÓN SOMACLONAL Causas de la variación somaclonal El polietilenglicol (PEG) y la variación somaclonal Niveles de detección de la variación somática
MARCADORES MOLECULARES Tipos de marcadores moleculares Uso de los RAPO para el análisis de la variación somaclonal Uso de los ISSR para el análisis de la variación somaclonal
HIPÓTESIS Y OBJETIVOS Hipótesis Objetivo general Objetivos específicos
BIBLIOGRAFIA
Página
1 111 V
VIl IX XI
XIII 1 3
5
5 5 6 6 7
7 7 8 9
10 10 10 11 12 13 14
17 17 19 20
22 22 23 24
26 26 26 26
27


Capitulo JI. Materiales y Métodos 35
2.1 ASPECTOS GENERALES 37 2.1 .1 Material vegetal y condiciones de cultivo 37 2.2 INDUCCIÓN DE LOS PROCESOS DE MORFOGÉNESIS in vítro 37 2.2.1 Organogénesis directa 37 2.2.2 Embriogénesis somática directa 37 2.2.3 Embriogénesis somática indirecta 38 2.2.4 Formación de callo embriogénico 38 2.3 ANÁLISIS DE LA VARIACIÓN SOMACLONAL 38 2.3.1 Extracción de ADN 38 2.3.2 Selección y uso de cebadores RAPOs 39 2.3.3 Selección y uso de cebadores ISSRs 39 2.4 OBTENCIÓN DE UNA LINEA TOLERANTES A ESTRÉS 40
HfDRICO 2.4.1 Selección bajo presión con PEG sobre la embriogénesis somática 40
directa 2.4.2 Selección asistida por marcadores moleculares RAPOs 41 2.4.3 Determinación del contenido de prolina 42 2.5 BIBLIOGRAFfA 42
Capitulo 111. Resultados y Discusión 45
3.1 ANÁLISIS MOLECULAR DE LA VARIACIÓN SOMACLONAL 45 (ARTICULO SOMETIDO)
3.1.1 Abstract 45 3.1.2 lntroduction 46 3.1 .3 Materials and Methods 47 3.1.4 Results and Discussion 51 3.1.5 References 57 3.2 OBTENCIÓN DE UNA LINEA EMBRIOGÉNICA TOLERANTE A 60
ESTRÉS HfDRICO 3.2.1 Análisis molecular RAPO de las líneas embriogénicas 63 3.2.2 Determinación del contenido de prolina 65 3.3 PROTOCOLO PARA LA OBTENCIÓN DE LA lÍNEA 67
EMBRIOGÉNICA TOLERANTE A ESTRÉS HfDRICO 3.4 DISCUSIÓN GENERAL 68 3.5 BIBLIOGRAF[A 69
Capitulo IV. Conclusiones y Perspectivas 4.1 CONCLUSIONES 4.2 PERSPECTIVAS Anexos 1 2 3 4 5
Medio MS (Murashige & Skoog, 1962) Método de extracción de ADN de Dellaporta (1983) Protocolo de tratamiento de RNAsa Kit de cebadores RAPOs Kit de cebadores ISSRs
73 73 74 75 75 76 76 77 78


AGRADECIMIENTOS
Este trabajo se realizó en el Centro de Investigación Científica de Yucatán en la Unidad de Bioquímica y Biología Molecular de Plantas, bajo la dirección de la Dra. Nancy Santana Buzzy y co-direción de la Dra. Lourdes Iglesias Andrew (INBIOTECA), a quienes agradezco por su apoyo, amistad y compartir conmigo sus conocimientos y experiencia.s.
Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por la beca otorgada para realizar los estudios de doctorado (No. de becario: 200308).
A los miembros de mi comité tutoral y revisores de tesis : Dra. Nancy Santana Buzzy, Dra. Lourdes Iglesias Andreu, Dra. Guadalupe López Puc, Dra. Luisa Alhucema López Ochoa, Dr. Luis Alfonso Sáenz, Dr. Manuel Martínez Estévez y Dr. Javier O. Mijangos, por la revisión crítica con la que enriquecieron este escrito.
Este trabajo fue financiado a través de los proyecto: Fondos Mixtos-2008 (proyecto: 1 05699) y Ciencia Básica-2008 (proyecto: 1 00496).
A mi grupo de trabajo: Susana, Daniela, Eunice, Carlos Regla, Carlos Lecona por su amistad, apoyo y colaboración.
A los técnicos del laboratorio: Adriana Canto-Fiick y Eduardo Balam-Uc.
1

II

DEDICATORIA
A mi familia
Por su apoyo incondicional en la realización de mis metas y objetivos
III

IV

Tabla
1.1
1.2
1.3
2.1
2.2
3.1
3.2
3.3
LISTA DE TABLAS
Descripción taxonómica de chile Habanero.
Reportes de regeneración de plantas in vitro del género Capsicum. Concentración de PEG empleada para inducir la tolerancia al estrés hídrico en diferentes cultivos. Grados de sequía según el descenso del potencial hídrico del agua y contenido relativo de agua. Potencial hídrico del medio de cultivo.
RAPO and ISSR primers evaluated in genomic DNA of Habanero pepper (C. chinense). List of RAPO and ISSR primer used for detecting somaclonal variation in Habanero pepper (C. chinense). Effectiveness of RAPO e ISSR markers in detecting polymorphism of Habanero pepper somaclonal variation.
V
Pág.
5
15
19
40
41
49
50
52

VI

Figura
1.1
1.2
2.1
2.2
3.1 3.2
3.3
3.4
3.5 3.6
3.7
3.8
3.9
LISTA DE FIGURAS
Control de la diferenciación ejercido por la interacción entre las auxinas y las citocininas en los cultivos in vítro. Frecuencia de la variación somaclonal en relación al tipo de explante, vía de regeneración y tiempo de cultivo. Estrategia experimental l. Análisis molecular de la variación somaclonal. Estrategia experimental 11. Obtención de una línea embriogénica tolerante a estrés hídrico. Morphogenesis induction in Habanero pepper. Dendrograms based on the Jaccard's coefficient in Habanero pepper regeneration protocols by UPGMA cluster analysis. Electrophoresis pattern of randomly amplified polymorphic ONA (RAPO) and inter-simple sequence repeats (ISSR) markers obtained from regeneration protocols of Habanero pepper. Efecto del PEG sobre la selección bajo presión y germinación después de un tratamiento con GA3 en embriones somáticos chile Habanero. Embriones somáticos bajo presión de selección con PEG. Germinación in vítro de embriones somáticos después de un tratamiento con GA3 y previo tratamiento con 15% de PEG. Perfil de bandas obtenido a través de geles RAPO separados en agarosa al 2% bajo diferentes tratamientos con PEG. Efecto del PEG sobre del contenido de prolina en embriones somáticos de chile Habanero. Etapas en el desarrollo del protocolo para la obtención de una línea embriogénica tolerante a estrés hídrico.
VII
Pág.
10
17
35
36
51 53
55
60
61 62
64
65
67

VIII

ADN
2,4-D
GA3
AlA
A lB
ANA
A.C. BAP
BioMIN~ AICI3
NaCI
CTV
cv
2n
dNTPs
ES
spp.
i.a.
Kb
Mpa
IJM MS
n PAC
p. f. PCR
pH
RCV
Taq
TDZ
TBE
LISTA DE ABREVIATURAS
Ácido desoxirribonucleico
Ácido 2,4-diclorofenoxiacético
Ácido giberélico
Ácido indolacético
Ácido indolbutírico
Ácido naftalenacético
Antes de Cristo
6-bencilaminopurina o benciladenina
Biorreactor modular de inmersión temporal
Cloruro de aluminio
Cloruro de sodio
Cultivo de Tejidos Vegetales
Cultivar
Diploide
Deoxyribonucleotide triphosphates
Error estándar
Especies
Ingrediente activo
Kilo base
Mega pascales Micromolar (10-6M)
Medio de cultivo de Murashige y Skoog (1962)
Número de muestra
Paclobutrazol
Peso fresco
Polymerase chain reaction
Potencial de hidrógeno
Reguladores del crecimiento vegetal
Thermus aquaticus
Tidiazurón (regulador del crecimiento)
Tris/Borate/EDT A
IX

X

RESUMEN
En este estudio evaluamos por medio de marcadores moleculares RAPO (Randomly Amplified Polymorphic ONA) e ISSR {lnter Simple Sequence Repeats), la variación somaclonal inducida a través de la organogénesis, embriogénesis somática directa e indirecta y callo embriogénico. Un total de 62 bandas RAPO y 219 bandas ISSR fueron generadas a partir de 7 primers RAPO y 13 primers ISSR. Mediante el coeficiente de similitud de Jaccard y empleando el método de agrupamiento por pares no ponderados con media aritmética (UPGMA), se obtuvieron dos dendrogramas por cada marcador. Los resultados indican que, los sistemas de regeneración de chile Habanero evaluados generan alto grado de variación somaclonal, por lo tanto, estos sistemas son inapropiados para la micropropagación, conservación o transformación genética. Sin embargo, en un segundo enfoque y haciendo uso de la variación generada de estos sistemas, se procedió a obtener una línea embriogénica tolerante a estrés hídrico, para ello se ejerció una presión de selección empleado polietilenglicol (PEG) como agente osmótico bajo diferentes concentraciones (0, 5, 1 O, 15, 20 y 25%). Para la obtención de las líneas tolerantes a estrés hídrico se realizó una selección temprana asistida con marcadores moleculares RAPO y un análisis bioquímico por medio de la determinación del contenido de prolina en los diferentes tratamientos con PEG. El análisis molecular RAPO permitió detectar polimorfismo en aquellas líneas tratadas bajo estrés. Por otro lado, el análisis bioquímico mostró un incremento significativo en el contenido de prolina a medida que aumentó la presión de selección. En conclusión, la variación somaclonal con ayuda de marcadores moleculares y bioquímicos, constituye una herramienta útil que puede ser aplicada en el mejoramiento genético de chile Habanero para la obtención de líneas tolerantes al estrés por sequía.
XI

XII

ABSTRACT
In this study we evaluate somaclonal variation induced through organogenesis, direct and indirect somatic embryogenesis and embryogenic callus, using molecular markers RAPO (Randomly Amplified Polymorphic ONA) and ISSR (lnter Simple Sequence Repeats). A total of 62 RAPO bands and 219 ISSR bands were generated with 7 RAPO primers and 13 ISSR primers. Genetic similarity was calculated by Jaccard's coefficient method and the similarity matrix was subjected to cluster analysis by the unweighted pair-group method with arithmetic averages (UPGMA). Two dendrograms were obtained for each marker. Results show that, the Habanero pepper regeneration systems evaluated generate a high degree of somaclonal variation making them inappropriate for micropropagation, conservation or genetic transformation. However, in a second approach, making use of the variation generated in these systems, we proceeded to obtain an embryogenic line that was tolerant to hydric stress. to achieve this we applied selective pressure by employing polyethylenglicol (PEG) as an osmotic agent under different concentrations (0, 5,1 O, 15, 20 and 25%). In order to obtain hydric stress tolerant lines, PEG treatments of embryogenic lines were carried out with RAPO molecular markers and a biochemical analysis by determining the proline content. With the RAPO molecular analysis it was possible to detect polymorphism in the lines treated with PEG. The biochemical analysis showed a significant increase in the proline content in response to an increase in selective pressure. In conclusion, somaclonal variation, with the aid of molecular and biochemical markers, represents a useful tool to be applied in the genetic improvement of Habanero pepper in order to obtain genotypes tolerant to drought stress.
XIII


INTRODUCCIÓN
El chile Habanero (Capsicum chinense Jacq.) pertenece a la familia de las Solanáceas y es un cultivo de gran importancia económica en el mercado nacional e internacional. El alto nivel de pungencia de sus frutos define su uso comercial y lo hace ser muy codiciado en diferentes industrias (Tun-Dzul, 2001 ). En México el chile Habanero se cultiva principalmente en los Estados de Tabasco, Campeche, Quintana Roo y Yucatán (Latournerie et al. , 2006).
Las temperaturas extremas, las inundaciones y la sequía, son entre otros, algunos de los factores abióticos que en los últimos años han afectando a los cultivos debido al cambio climático (IPCC, 2001 ). En los posibles escenarios del cambio climático global, las predicciones apuntan a que el riesgo a la sequía se incrementará durante el siglo XXI, asociada con un régimen mas irregular de las precipitaciones y con temperaturas más extremas y en general mas cálidas (IPCC, 2006). En lo particular, el estrés por sequía constituye en la actualidad la principal causa de pérdidas en el rendimiento de los cultivos económicamente importantes (Vinocur & Altman, 2005). Orellana et al. (201 0), pronostican que la precipitación pluvial en la Península de Yucatán disminuirá en los próximos 12 años, mientras que la temperatura aumentará, como consecuencia del calentamiento global. Por estas razones y dada la importancia del cultivo de chile Habanero, es necesario contar con programas de mejoramiento genético enfocados a la obtención de cultivares mejores adaptados a las condiciones climáticas que la naturaleza les impone.
La variación generada a partir del cultivo in vitro ó "variación somaclonal " (Larkin & Scowcroft, 1981) puede ser aprovechada mediante la selección de regenerantes con características agronómicas deseables, como son la tolerancia a sequía, salinidad o resistentes a enfermedades (Evans, 1989; Bajaj et al., 1990). La ventaja que tienen estos sistemas de selección in vitro es que permiten la manipulación eficiente de un gran número de individuos en espacios muy reducidos, y disminuyen notablemente el tiempo requerido para desarrollar los programas de mejoramiento genético de plantas (Jain, 2001 ; Cardone et al., 2004). Para seleccionar líneas tolerantes a estrés abiótico se han empleado diferentes agentes selectivos estresantes (AICI3, NaCI, PEG, entre otros) en dependencia de la problemática de interés. Por sus propiedades, el PEG se ha usado como agente osmótico para seleccionar líneas tolerantes a estrés hídrico en diversas especies vegetales. Se ha aplicado en suspensiones celulares de tomate (Handa et al., 1983) y chile (Santos-Díaz & Ochoa-Aiejo, 1994). También ha sido empleado en medio semisólido durante el cultivo de embriones somáticos de sorgo (Duncan et al., 1995) y de arroz (Biswas et al., 2002), y más recientemente durante la proliferación de brotes en banano, tanto en medio líquido como en semisólido (Ebrahim et al., 2006).
1

Los marcadores morfológicos, bioquímicos y moleculares han sido de gran utilidad en los estudios de variación somaclonal. Los marcadores moleculares son particularmente muy útiles en estos estudios, ya que permiten detectar variaciones en las regiones cod ificantes y no codificantes del AON, además de que no son influenciados por el ambiente ni por los estadios fenológicos de las plantas (Valadez & Günter, 2000; Cardone et al., 2004). Los más utlilizados son los RFLP (Restriction Fragment Length Polymorphism), RAPO (Random Amplified Polymorphic ONA), AFLP (Amplified Fragment Polymorphism), SSR (Single Sequence Repeat ó Microsatélites) e ISSR {lnter Single Sequence Repeat ó intermicrosatélites). En particular, los RAPOs e ISSRs son marcadores multiloci que no requieren conocimiento previo del genoma, su ensayo es rápido, simple, de bajo costo y han sido utilizados para estudiar la variación somaclonal en diversas especies (Aihani & Wilkinson, 1998; Soniya et al., 2001; Hossain et al., 2003; Martins et al., 2004; Lakshmanan et al. , 2007). En este contexto, el objetivo del presente estudio fue evaluar mediante los marcadores moleculares (RAPO e ISSR) los niveles de variación somaclonal inducidos por tres vías de regeneración en chile Habanero (Capsicum chinense Jacq.) y seleccionar líneas tolerantes a estrés hídrico usando PEG como agente selectivo.
BIBLIOGRAFÍA
Alhani M. and Wilkinson M. {1998). lnter simple sequence repeat polymerase chain reaction for the detection of somaclonal variation. Plant Breeding 17 (6):573-575.
Bajaj, Y.P. (1990). Somaclonal variation-origin, induction, cryopreservation, and implications in plant breeding. In: Bajaj Y. P.(ed). Somaclonal variation in crop improvement. Springer-Verlag, Berlín, pp. 3-48.
Biswas, J., B. Chowdhury, A. Bhattacharya, and A. Mandal. (2002) . In vitro screening for increased drought tolerance in rice. In Vitro Cell. Oev. Bioi.-Piant, 38,525-530.
Cardone, S., S. Olmos y V. Echenique (2004). Variación somaclonal. In: Echenique V. , Rubinstein C., y Mroginski L. (eds). Biotecnología y Mejoramiento Vegetal. Ediciones INTA. 111 (1 ), 83-96.
Ouncan, R., R.M. Waskom and M. W. Nabors (1995). In vitro screening and field evaluation of tissue-culture-regenerated sorghum (Sorghum bicolor (L.) Moench) for soil stress tolerance. Euphytica, 85, 373-380.
Ebrahim, M.K., I.A. lbrahim, H.A. Emara and E. Komor (2006). lmpact of polyethylene g/yco!-induced water stress on growth and development of shoot tip cultures from different banana (Musa spp.) cu/tivars. Journal of Applied Horticultura, 8(1 ), 53-57.
Evans, O.A. (1989) . Somaclonal variation-genetic basis and breeding app/ications. Trends in Genetics, 5, 346-50.
2

Handa, A.K., R.A. Bressan, S. Handa and P.M. Hasegawa (1983). Clona/ Variation for Tolerance to Polyethy/ene G/yco/-lnduced Water Stress in Cu/tured Tomato Cells. Plant Physiol, 72, 645-653.
Hossain A. , K. Konisho, M. Minami and K. Nemoto (2003). Somac/onal variation of regenerated plants in chili pepper (Capsicum annuum L.). Euphytica, 130, 233-239.
IPCC (2006). Guidance notes for /ead authors of the IPCC fourth assessment report on addressing uncertainties. Appendix in Manning 2006. Adv Clim Change Res 2:13-21
IPCC (2001 ). Climate change 2001: the scientific basis. In: Houghton J., Y. Ding, J. Griggs, M. Noguer, P. Van der Linden, X. Dai, K. Maskell and C. Johnson(eds). Third Assessment Report of the lntergovernmental Panel on Climate Change. Cambridge University Press, Cambridge.
Jain, S.M. (2001 ). Tissue culture-derived variation in crop improvement. Euphytica, 118, 153-166.
Lakshmanan, V., S. Venkataramareddy and B. Neelwarne. (2007). Molecular analysis of genetic stability in /ong-term micropropagated shoots of banana using RAPO and /SSR markers. Electronic Journal of Biotechnology, 1 0(1 ),1-8.
Larkin, P.J. and S.C. Scowcroft. (1981). Somaclonal variation: a novel source of variability from cell culture for plant improvement. Theor Appl Genet, 60, 197-214.
Latournerie, L.M., M.E. Yupit, S.J. Tún, M.A. Pech y C.D. Lozada (2006). Compartimiento de materiales criollos de Chile Habanero ante el ataque de la virosis. Uuy u Táan. (Fundación Produce Yucatán, A.C.), 1, 15-17.
Martins, M., D. Sarmento and M. Oliveira (2004). Genetic stability of micropropagated a/mond plantlets, as assessed by RAPO and /SSR markers. Plant Cell Rep., 23, 492-496.
Orellana, R., C. Espadas, C. Conde y C. Gay (201 0). Atlas escenarios de cambio climático en la Península de Yucatán. CICY (ed). pp 133.
Santos-Díaz, M. and N. Ochoa-Aiejo (1994). PEG-tolerant cell clones of chili pepper: Growth, osmotic potentia/s and salute accumulation. Plant Cell, Tiss. Org. Cult. , 37,1-8.
Soniya, E., N. Banerjee and M. Das. (2001 ). Genetic ana/ysis of somaclona/ variation among callus-derived plants of tomato. Current Science, 80( 9), 1213-1215.
Tun-Dzul , J. (2001 ). Chile Habanero. Características y Tecnología de Producción, SAGARPA-INIFAP. Mocochá, Yucatán, México. pp 5-6.
Valadez, M.E. y K. Günter ( 2000). Huellas de ADN en Genomas de Plantas (Teoría y protocolos de laboratorio) . Editorial Mundi Prensa, Méx. pp. 147.
Vinocur, B. and A. Altman (2005). Recent advances in engineering plant to/erance to abiotic stress: achievements and limitations. Current Opinion in Biotechnology, 16, 123-132.
3

4

Capitulo 1
Antecedentes
1.1 EL CHILE HABANERO
1.1.1 Origen y distribución
El género Capsicum, al cual pertenecen todos los chiles cultivados, es originario de los trópicos de América. Las cinco especies domesticadas y sus parientes silvestres estuvieron confinadas en el Continente Americano en la época precolombina. Los primeros exploradores españoles y portugueses, encontraron los frutos tan pungentes que lo introdujeron rápidamente a Europa y Asia. El hallazgo de remanentes arqueológicos que datan de 7 000 años A.C. ha permitido determinar que las especies de este género se domesticaron en diferentes partes de América, principalmente en México {Tun-Dzul, 2001 ).
El centro de diversidad de las especies cultivadas de Capsicum incluye a México y Centroamérica. Existen centros secundarios en el centro y sureste de Europa, África, Asia y América Latina. Se indican como centros de origen de C. frutescecens L. y C. chinense Jacq. a Bolivia, Perú, sureste de Brasil, Los Andes y Colombia, aunque algunos tipos también se pueden encontrar en África y el sureste de Asia, ya que fueron introducidos por los portugueses en la época Colonial (Tun-Dzul, 2001 ).
El chile Habanero proviene de las tierras bajas de la cuenca Amazónica y de ahí se dispersó a Perú durante la época prehispánica. La distribución también se dirigió hacia la cuenca del Orinoco (ubicada actualmente en territorios de Colombia y Venezuela) hacia Guyana, Surinam, la Guyana Francesa y las Antillas del Caribe (González-Estrada et al., 2006)
1.1.2 Descripción taxonómica
La descripción taxonómica de chile Habanero se muestra en la Tabla 1.1
5

Tabla 1.1. Descripción taxonómica de chile Habanero de acuerdo al Código Internacional de Nomenclatura Botánica (CINB).
Reino Plantae
Subreino E m briophyta
División Angiosperma
Clase Dicotiledónea
Subclase Metachlam idea
Orden Tubiflorales
Familia Solanacea
Género Capsicum
Especie Capsicum chinense Jacquin
1.1.3 Importancia económica
En México el chile Habanero se cultiva principalmente en los Estados de Tabasco, Campeche, Quintana Roo y Yucatán, donde suele formar parte de ciertos platillos regionales. En Yucatán, el cultivo de chile Habanero es uno de los más importantes debido a su demanda local , nacional e internacional (Latournerie et al., 2006). Se estima que la superficie de cultivo en el Estado de Yucatán es de 423.3 hectáreas, con una producción de 3 390 toneladas que corresponden al 47.9% de la producción nacional, los rendim ientos varían entre 8 y 12 toneladas por hectárea (ton/ha), en suelos pedregosos al norte del estado, y de 1 O a 15 ton/ha en suelos mecanizados al sur (González-Estrada et al., 2006). La mayor superficie de cultivo se encuentra en la parte norte del estado y contribuye en más del 90% del volumen de la producción estatal, la mayor parte de la producción se comercial iza y se consume en fresco, sólo una pequeña parte se utiliza en la industria como materia prima para la elaboración de salsa picante (Tun-Dzul , 2001 ).
En el 2004, el consumo de chile Habanero para la venta en fresco y procesado alcanzó 1,372 toneladas según datos proporcionados por las principales compañías comercializadoras y procesadoras de Yucatán, lo cual representa un volumen de venta de 100 millones de pesos (Ledón, 2006}.
Los principales mercados nacionales donde se comercializa la producción de chile Habanero son las centrales de abastos del Distrito Federal, Guadalajara, Monterrey, San Luis Potosí, Morelia, Cancún, Puebla y Oxkutzcab. En los
6

mercados internacionales se comercializa en los Estados Unidos (California, Florida, Texas, Nueva York, Oregón, Nuevo México y Massachusset), Canadá y Cuba. En los mercados europeos se exportó a España (Madrid, Barcelona, Zaragoza y Valencia) e Inglaterra (Londres). En Asia, el principal mercado es Japón, país que ha iniciado una estrategia sumamente interesante y atractiva para establecer negocios relacionados con la industria de chile Habanero (Ledón, 2006).
1.1.4 Problemática: El chile Habanero y el cambio climático
Al cambio climático se le atribuye el continuo incremento en la temperatura de la superficie del mar en el Golfo de México, el aumento del nivel del mar, huracanes de mayor intensidad, así como los cambios en los ciclos de lluvia, todos ellos de carácter irreversible (González, 2007). El aumento de la sequía no sólo es el resultado de una menor precipitación anual, sino también de un patrón de distribución estacional diferente, con lluvias torrenciales e irregulares que tienden a aumentar durante el invierno, seguidas de largos períodos secos, que tienden a aumentar durante el verano (Rambal & Debussche, 1995; Reichstein et al., 2002). Además, la disponibilidad real de agua para las plantas disminuirá durante el siglo XXI debido al incremento de evapotranspiración como consecuencia del incremento de temperatura (IPCC, 2001 ). En lo que respecta al Estado de Yucatán, la Comisión Nacional del Agua (CONAGUA) en el 2008 determinó que 49 municipios del estado fueron afectados por una sequía atípica, durante los meses de julio y agosto. Por otro lado Orellana-Lanza (201 O) menciona que es probable que disminuya la precipitación pluvial en la Península de Yucatán en los próximos 12 años mientras que la temperatura aumentará, como consecuencia del calentamiento global.
Tomando en cuenta todos estos aspectos del cambio climático, y dada la importancia del cultivo de chile Habanero para la región, es importante desarrollar modelos de selección utilizando alternativas más eficientes para la . mejora genética de esta especie, a fin de contar con líneas tolerantes a estrés, en este caso a estrés hídrico.
1.2 ESTRÉS ABIÓTICO EN PLANTAS
1.2.1 Mecanismos de Respuesta
Levitt (1980) define al estrés ambiental como al cambio en las condiciones del medio que reducen o cambian desfavorablemente el crecimiento o desarrollo de las plantas. El estrés ambiental provoca en las plantas respuestas complejas y constituyen un problema fundamental para la agricultura, ya que influyen sobre la supervivencia y la productividad de los cultivos. Estas respuestas se manifiestan a
7

nivel celular, fisiológico y durante el desarrollo (Hsiao, 1973; Levitt, 1980) y son caracteres de variación continua, controlados por un importante número de genes aditivos y probablemente sinérgicos (Bohnert et al., 1995).
Los genes inducidos bajo condiciones de estrés abiótico, tales como las bajas temperaturas, la salinidad y la sequía, codifican proteínas que tienen funciones de señalización y protección celular, como por ejemplo: proteínas de canales iónicos que alteran el potencial del agua celular, enzimas requeridas para la síntesis de sustancias protectoras (azúcares, prolina y betaína), proteínas protectoras como las de tipo LEA (Late Embryogenesis Abundant), proteínas anticongelantes, chaperonas, proteasas y enzimas detoxificantes como la catalasa y la ascorbato peroxidasa (Vinocur & Altman, 2005).
La hipótesis de señalización actual postula que las señales ambientales son percibidas primero por receptores específicos, que con su activación inician o suprimen una cascada de transducción de señales intracelulares y, en muchos casos, activan factores de transcripción nucleares que inducen la expresión de grupos específicos de genes. Una forma común de señal de iniciación es la fosforilación de proteínas acoplada a receptores (Xiong & Zhu, 2001 ).
Los genes inducidos por estrés incluyen genes de respuesta temprana y genes de respuesta tardía. Los primeros son inducidos en pocos minutos y a menudo transitoriamente, su inducción no requiere de síntesis de nuevas proteínas, ya que estos componentes de señal ización serían constitutivos. En contraste, los genes de respuestas tardías son activados más lentamente (en horas) y su expresión es generalmente sostenida durante el tiempo en que el estrés se encuentra actuando. Los genes tempranos codifican factores de transcripción que activan a los genes de respuesta tardía , que constituyen la mayoría de los genes inducidos por estrés (Puebla & Del Viso, 2004)
1.2.2 Tolerancia a estrés hídrico
El estrés hídrico, en su sentido más amplio incluye tanto a la sequía como la salinidad. Estos dos tipos de estrés afectan aspectos en la fisiología de una planta y su metabolismo. El déficit hídrico actúa sobre las plantas produciendo dos tipos de respuestas: 1) evitar o prevenir la tensión del agua (mecanismos evitadores), y 2) activando mecanismos o adaptaciones que perm iten soportar o resistir el estrés (mecanismos tolerantes) (Levit, 1980). En las plantas, los mecanismos para afrontar esta situación han sido denominados estrategias, y son características de cada especie (Larcher, 1995). Otra estrategia, cuestionada por numerosos autores para enfocar los trabajos de resistencia al déficit hídrico es la de "escape" de la sequía, donde las plantas completan su ciclo vital antes de la llegada del estrés
8

hídrico, dentro de la estrategia cabe incluir especies que entran en una dormancia vegetativa parcial durante el período de sequía (Valladares & Pearcy, 1997).
Un aspecto de gran relevancia en la respuesta de los cultivos a sequía es el ajuste osmótico, es decir, el incremento neto en la cantidad de solutos osmóticamente activos en los tejidos. Ese ajuste osmótico se alcanza mediante la acumulación de iones inorgánicos, o a través de la síntesis de solutos orgánicos, entre los cuales la prol ina es uno de los osmolitos más ampliamente distribuidos en plantas superiores (Stewart, 197 4 ).
1.2.3 El papel de la prolina ante el estrés hídrico
La prolina es un aminoácido que se encuentra en pequeñas cantidades en las plantas cuando éstas crecen en condiciones óptimas. En condiciones de estrés, se ha encontrado que el contenido de prolina aumenta para actuar como un agente osmótico, protegiendo a la planta de la deshidratación (Stewart, 1974; Kuznetsov & Schevyakova, 1999). Se ha sugerido que la prolina actúa también como un estabil izador de proteínas y membranas, inductor de genes relacionados a estrés osmótico (lyer & Caplan, 1998), como fuente de carbono y nitrógeno fácilmente disponible en la rehidratación celular (Brugiére et al. 1999).
Los iones inorgánicos son determinantes en el ajuste osmótico y se acumulan preferencialmente en vacuolas, mientras que solutos orgánicos como la prolina, glicinabetaina, sacarosa, manitol y otros se conocen como "solutos compatibles" se acumulan principalmente en el citoplasma, sin causar inhibición de la actividad enzimática y además impiden la deshidratación del citosol por la reducción en el potencial hídrico derivada de la acumulación de sales en vacuolas (Stewart, 1974).
El ajuste osmótico mediado por el aumento en la síntesis de prolina en un mecanismo fisiológico cuya determinación puede realizarse en etapas tempranas del desarrollo (Stewart & Lee, 1974). El aumento en el contenido de prolina bajo estrés osmótico in vitro se ha llevado acabo en especies como Sapindus trifoliatus L. (Unnikrishnan et al., 1991 ), Capsicum annum L. (Santos-Díaz & Ochoa-Aiejo, 1994 ), Oryza sativa L. (Lutts et al., 1996; Sajid et al., 2007).
9

1.3 EL CULTIVO in vitro
1.3.1 Cultivo de Tejidos Vegetales
Pérez-Molphe-Balch et al. (1999), definieron al Cultivo de Tejidos Vegetales (CTV} como el conjunto de técnicas que permiten el establecimiento, mantenimiento y desarrollo de brotes y/o embriones somáticos a partir de cualquier célula, tejido u órgano de una planta, bajo condiciones artificiales, axénicas y controladas.
El CTV se ha usado como una alternativa para la propagación de plantas, además, es una herramienta invaluable para la resolución de problemas básicos y aplicados en la biología vegetal, ya que por una parte ofrece una serie de sistemas modelo ideales para la investigación fisiológica, bioquímica, genética y estructural, por otro lado tiene una aplicación práctica en la clonación, conservación y manipulación in vitro de cualquier material vegetal (Pérez-MolpheBalch et al., 1999}.
1.3.2 Control del desarrollo in vitro
A las primeras hormonas y reguladores sintéticos con efectos sobre crecim iento en plantas fueron identificados como: hormonas del crecimiento, reguladores del crecimiento, sustancias reguladoras del crecimiento o sustancias del crecimiento (Szweykowscy, 1993, citado por Jankiewicz, 2003). Dependiendo de su estructura química y su efecto fisiológico, los reguladores del crecimiento vegetal (RCV) se clasifican en 5 grupos básicos: auxinas, citocininas, giberelinas, ácido abscísico y etileno. Actualmente se han aislado y estudiado otras sustancias fuera de estos grupos, que son consideradas hormonas vegetales, por ejemplo: poliaminas, jasmonatos, ácido salicílico y los brasinoesteroides (Davies, 1995). Los RCV más utilizados en los cultivos in vitro son los pertenecientes a los grupos de las auxinas y de las citocininas, de los cuales se conoce que regulan en gran medida los procesos de crecimiento y desarrollo en el cultivo de tejidos vegetales . En menor grado se utilizan algunos reguladores no pertenecientes a estos grupos como las giberel inas y el ácido abscísico (Pérez-Molphe-Balch et al., 1999}.
Alterando ligeramente las concentraciones relativas de auxina y citocinina, los investigadores han podido modificar el desarrollo de las células indiferenciadas en el cultivo in vitro de muchas especies vegetales . Una concentración más o menos igual de estos RCV provoca que las células sigan indiferenciadas, formando masas celulares amorfas llamadas callos . Cuando la concentración de auxina es superior, el tejido indiferenciado forma raíces; con una concentración superior de citocinina se forman yemas. Un cuidadoso equilibrio de las dos hormonas puede inducir raíces y/o yemas (Fig. 1.1) (Pérez-Molphe-Balch et al., 1999}.
10

CONCENTRACION DEAUXINAS
CONCENTRACION DE CITOCININAS
ALTA
NULA
FORMACIÓN DE RAÍCES
EMBRIOGÉNESIS
TEJIDO CALLOSO
ORGANOGÉNESIS
PROLIFERACIÓN DE YEMAS
NULA
ALTA
Figura 1.1. Control de la diferenciación ejercido por la interacción entre las auxinas y las citocininas en los cultivos in vitro (Pérez-Molphe-Balch et al., 1999).
1.3.3 Morfogénesis in vitro
Existen tres vías diferentes para la regeneración de plantas in vitro:1) Brotación y proliferación de yemas, 2) Organogénesis, y 3) Embriogénesis somática. En el primer caso se parte de meristemos ya formados para la obtención a partir de ellos de una o varias plantas completas. En la organogénesis y la embriogénesis somática se parte de tejidos no meristemáticos y se requiere de un proceso de desdiferenciación y rediferenciación de los tejidos para dar lugar a embriones somáticos o brotes adventicios. En estos sistemas las nuevas plantas se originan a partir de estructuras multicelulares, mientras que, en los sistemas de embriogénesis somática y or,ganogénesis las nuevas plantas se originan al parecer a partir de una sola célula (Pérez-Molphe-Balch et al., 1999).
La brotación y proliferación de yemas es un proceso donde no ocurre la morfogénesis in vitro y con número limitado de brotes. La organogénesis a diferencia de la embriogénesis somática, puede llevarse en muchas especies vegetales de una forma relativamente sencilla. Pérez-Molphe-Balch et al. (1999) recomiendan que al trabajar con una especie por primera vez, deberán montarse experimentos que nos permitan desarrollar el mejor sistema para regeneración de órganos a partir del material vegetal.
11

1.3.4 Organogénesis
El término organogénesis aplicado al cultivo in vitro de tejidos vegetales se refiere a la formación de órganos de novo en los explantes cultivados. Entre los órganos que se pueden formar están las raíces o los llamados brotes adventicios, los cuales son estructuras muy similares a una yema y tienen la capacidad de originar una nueva planta después de elongarse y formar raíces . El fenómeno de la organogénesis in vitro se basa en la llamada totipotencial idad celular, es decir, cada célula posee toda la información genética necesaria para constituir una planta completa o desempeñar las funciones de cualquier órgano o tejido vegetal. Sin embargo, en condiciones normales este potencial no se expresa, y se requieren condiciones extraordinarias como las que pueden derivarse del cultivo in vitro para que una célula exprese su potencial y pueda originar un órgano o una planta completa Por otro lado, aún cuando en teoría cualquier célula viva que posea núcleo es totipotente, se ha visto que en la real idad no todos los tipos celulares son capaces de desencadenar el proceso de organogénesis y la probabilidad de que esto suceda decrece conforme se incrementa el grado de diferenciación de una célula (Pérez-Molphe-Balch et al., 1999).
La organogénesis puede ser directa, cuando tiene lugar en el explante original, o bien indirecta cuando primeramente se origina tejido calloso y luego se originan los órganos a partir de este. Aún cuando la organogénesis in vitro es un proceso muy utilizado en la actualidad, incluso con fines comerciales para la propagación de plantas, es muy poco lo que se sabe acerca de los fenómenos fisiológicos, bioquímicos y genéticos que la hacen posible. De manera general , se sabe que bajo las condiciones adecuadas, ciertas células dentro de un explante comienzan a dividirse cambiando su estado de diferenciación y dando lugar a regiones llamadas meristemoides. Los meristemoides están compuestos por células pequeñas, isodiamétricas, de pared celular delgada, varias vacuolas periféricas pequeñas y un núcleo grande en posición central , lo cual corresponde a las características de las células meristemáticas comunes. Cada uno de estos meristemoides es capaz de desarrollarse para dar lugar a uno o varios brotes adventicios. El punto crítico en este proceso es el momento en que una célula dentro del explante, usualmente parenquimatosa y vacuolada, comienza a dividirse para dar lugar al grupo de células que comprenden el meristemoide. A este proceso en concreto se le da el nombre de inducción. Existen dos teorías básicas acerca del proceso de inducción. La primera de ellas propone que en un momento dado, ciertas células del explante sufren un aislamiento físico y/o fisiológico con respecto al resto del tejido, acarreando la pérdida de los controles externos. Esto permite que las células aisladas expresen su potencial de desarrollo dividiéndose y formando los llamados meristemoides. La segunda teoría propone que son los cambios cuantitativos en la disponibilidad de nutrientes y reguladores del crecimiento en una célula en particular, lo que induce
12

a ésta a dividirse y formar el meristemoide (Hicks, 1994). En la mayoría de los casos, los brotes adventicios se originan a corta distancia del medio de cultivo, o bien del tejido vascular, lo cual perece indicar que los gradientes fisiológicos que se dan dentro del explante tienen mucho que ver con el desarrollo de los meristemoides. De acuerdo a los conocimientos actuales, esta segunda teoría pudiera ser la más acertada (Pér~z-Molphe-Balch et al., 1999).
1.3.5 Embriogénesis somática
Ciertas células, y bajo ciertas condiciones de cultivo in vitro, tienen la capacidad de formar embriones mediante un proceso muy similar a la embriogénesis cigótica. A este proceso se le denomina embriogénesis somática y es una de las pruebas más notables de la totipotencia celular. Los embriones somáticos tienen, al igual que los cigóticos, la capacidad de formar una nueva planta después de un proceso de germinación, con la diferencia de que la embriogénesis somática es un proceso asexual , por lo tanto, la nueva planta será exactamente igual a la donadora de la célula inicial. La embriogénesis somática no es un proceso que suceda únicamente en cultivos in vitro, ocurre naturalmente en algunas familias de plantas y se conoce como apomixis (Kiran y Thorpe, 1995).
Para que una célula vegetal lleve acabo el proceso de embriogénesis somática debe estar determinada o inducida. Por lo anterior, las células capaces de formar embriones somáticos reciben el nombre de células embriogénicas determinadas (CED). Este proceso de inducción aún no se conoce con exactitud pero se sabe que implica un cambio drástico en los patrones de expresión génica mediante el cual las células pasan del patrón normal a uno embriogénico (Pérez-MolpheBalch et al., 1999).
Existen dos tipos de embriogénesis somática in vitro, la directa y la indirecta. En la embriogénesis somática directa, los embriones aparecen directamente sobre el explante original. Por otro lado, en la embriogénesis somática indirecta, es indispensable obtener un tejido calloso o bien una suspensión celular embriogénica a partir de la cual se obtendrá la diferenciación de los embriones somáticos. Este es un proceso mucho más complejo que la embriogénesis somática directa. Se supone que la embriogénesis somática directa se da cuando en el explante original existen ya células proembriogénicas, por lo que no es necesario el proceso de inducción y sólo se requiere proporcionar al tejido las condiciones adecuadas para que suceda la diferenciación de los embriones somáticos. En la embriogénesis somática indirecta, el explante original no tiene células proembriogénicas, por lo que se requiere primero darle al tejido las condiciones para que ocurra la inducción y luego cambiarlo a otras que sean propicias para la diferenciación de los embriones. Las células de tejidos muy jóvenes, como por ejemplo los embriones cigóticos inmaduros son ya en una
13

buena proporción células proembriogénicas, de no ser así, son fácilmente inducibles para que lo sean. Por el contrario, las células más diferenciadas o de tejidos adultos muy difícilmente o casi nunca se llegan a convertir en proembriogénicas. Esto explica por qué la embriogénesis somática resulta sencilla en algunos tejidos y prácticamente imposible en otros (Pérez-MolpheBalch et al., 1999).
1.3.6 Regeneración de chile (Capsicum spp.J in vitro
En C. annuum L. se han publicado varios protocolos de regeneración basados en la inducción de organogénesis, en los que destacan trabajos realizados por Agrawal et al. (1989), Ochoa-Aiejo y !reta-Moreno (1990), Valera-Montero y Ochoa-Aiejo (1992), Binzel et al. (1996b), Buyukalaca y Mavituna (1996), Steinitz et al. (1999), Ramirez-Malagón y Ochoa-Aiejo (1996). Venkataiah et al. (2003) reportó la regeneración de C. annuum L. utilizando segmentos de hoja como explante con TDZ, obteniendo de 2-25 brotes por explante. Kumar et al., (2005) utilizando meristemos y hojas cotiledonarias, lograron la regeneración de brotes (20-25 por explante) en presencia de BAP, AlA, nitrato de plata y ácido benzoico como inhibidor del transporte de auxinas. Recientemente Ahman et al. (2006) obtuvieron la inducción de brotes múltiples a partir de segmentos de nudos en presencia de 1¡.JM de TDZ alcanzando un promedio de 14 brotes por explante.
En todos los trabajos publicados, la fuente de explante es muy variada (cotiledones, hipocótilos, ápices, segmentos de hoja, nudo y entrenudo), al igual que el tipo y concentraciones de reguladores de crecimiento. Por lo tanto, podemos mencionar que no existe un protocolo estable y eficiente de regeneración en C. annuum L. {Tabla 1.2) debido a la gran variación de genotipos y la recalcitrancia a la morfogénesis del género. La regeneración de C. annuum L. a través de embriogénesis somática ha sido reportada por Binzel et al. (1996a), Buyukalaca y Mavitura (1996), Steintz et al. (2003) y recientemente un protocolo de embriogénesis somática directa fue desarrollado por Khan et al. , {2006) utilizando meristemo apical y segmentos de tallo que fueron tratados con TDZ (0.5 IJM). En ninguno de estos reportes se ha logrado regenerar plantas completas .
El primer reporte de regeneración de plantas in vitro de chile Habanero se realizó por primera vez por Santana-Buzzy et al. (2005) obteniendo brotes múltiples a partir de nudos provenientes de plantas asépticas como fuente de explante, utilizando 3.4 IJM de tidiazurón {TDZ) en medio semisólido MS (Murashige y Skoog, 1962). Mas tarde, Santana-Buzzy et al. {2006) reportaron la producción de etileno durante el cultivo in vitro de chile Habanero. En este estudio se observó que la máxima concentración de este RCV ocurrió entre los 21-24 días de cultivo (0.12 iJL. L"\ donde los explantes mostraron una alta sensibilidad a este gas, con
14

efectos negativos sobre los brotes y desarrollo de las plantas de chile Habanero in vitro. Estos efectos desaparecen al mejorar el intercambio gaseoso manteniendo ventilados los recipientes de cultivo.
López-Puc G. et al. (2006) reportaron el primer sistema de regeneración de chile Habanero vía embriogénesis somática directa. En este protocolo se utilizó como fuente de explante segmentos de hipocótilos de chile Habanero cultivados en medio MS con 2,4-D (9.05 !JM). Se obtuvieron embriones directamente del explante y la eficiencia de este sistema una de las más altas del género Capsicum hasta la fecha (175±20, brotes por explante). Siguiendo esta misma línea, ZapataCastillo et al. (2007) reportaron un protocolo de regeneración vía embriogénesis somática indirecta, el primero en formar embriones a partir de células en suspensión en C. chinense. La composición del medio estuvo basada en las sales MS, suplementado con 2,4-D (9.05 !JM) durante la fase de inducción, sustituyendo esta auxina por TDZ (3.5 !JM) al momento de iniciar la formación de los embriones somáticos. En ambos protocolos de embriogénesis somática reportan que la mayor dificultad para el éxito de este proceso radica en la baja eficiencia en la etapa de conversión de los embriones a plantas completas, y a la alta frecuencia de embriones deformados que se observa en las poblaciones generadas.
Bello-Bello et al. (201 O) reportaron un nuevo sistema para la regeneración in vitro de chile Habanero vía organogénesis directa, donde a partir de nudos de 45-65 días de edad en medio semisólido MS con TDZ (3.4 !JM) + PAC (3.4 !JM) se logró aumentar el número de brotes por explante (25-30), superando al protocolo anterior (Santana-Buzzy et al., 2005). En este estudio se logró la elongación y enraizamiento de los brotes en medio semisólido y/o líquido (empleando el BioMIN~) compuesto por MS + GA3 (2 .77 !JM) + AgN03 (10 !JM) para la elongación y MS + AlA ( 1. 70 !JM) para el enraizamiento.
15

Tabla 1.2. Reportes de regeneración de plantas in vitro del género Capsicum. Especie RCV Explante Respuesta Referencia C. annuum L. AJA, BA Hipocótilos Callo, Agrawal et al. {1989)
cotiledones Organogénesis
C. annuum L. AIA,2iP Hipocótilos Organogénesis Ochoa-Aiejo e lreta-Moreno {1990)
C. annuum L. AlA, BAP Hipocótilo Organogénesis Valera-Montero y Ochoa-Aiejo {1992)
C. annuum L. 2,4-D, Ápices, Embriogénesis Binzel et al. (1996) TDZ embriones somática {AgN03) cigóticos
C. annuum L. 2,4-D, BA Embriones Embriogénesis Buyukalaca y Mavituna cigóticos Somática et al. (1 996) maduros
C. annuum L. 2,4-D Embriones Embriogénesis Steinitz et al. {2003) cigóticos somática
C. annuum L. A lB Hipocótilos Organogénesis Ramírez-Malagón y Ochoa-Aiejo {1996)
C. annuum L. TDZ Hipocótilos, Organogénesis Venkataiah et al. cotiledones, {2003) hoja
C. annuum L BAP,AIA, Meristemos y Organogénesis Kumar et al. {2005) {AgN03) hojas
C. chinense TDZ Nudos Organogénesis Santana-Buzzy et al. Jacq. {2005)
C. chinense TDZ Nudos Organogénesis Santana-Buzzy et al. Jacq. Desarrollo {2006)
C. chinense 2,4-D Hipocótilos Embriogénesis López-Puc et al. Jacq. somática {2006)
directa C. annuum L. TDZ Nudos Organogénesis Ahmad et al. {2006)
C. annuum L. TDZ, IBA hipocótilos Embriogénesis Khan et al. {2006) somática
C. chinense 2,4-D Hipocótilos Embriogénesis Zapata-Castillo et al. Jacq. TDZ somática {2007)
indirecta C. chinense TDZ, Nudos Organogénesis Bello-Bello et al. Jacg. PAC {201 O}
16

1.4 VARIACIÓN SOMACLONAL
1.4.1 Causas de la variación somaclonal
El cultivo in vitro per se puede llegar a ser muy estresante para las células vegetales y puede involucrar procesos mutagénicos durante el establecimiento del explante, la inducción de callo, la formación de embriones ó durante la formación de órganos. Por esta vía es posible obtener variación somaclonal de origen nuclear y/o citoplasmático, esta variación puede ser utilizada como una estrategia en el mejoramiento genético vegetal. El proceso denominado variación somaclonal, por Larkin y Scowcroft {1981}, involucra cambios genéticos en las plantas regeneradas a partir del cultivo de tejidos vegetales, y que dichos cambios son transmitidos a la progenie. Asimismo, cabe mencionar que los cambios epigenéticos son también considerados como una variación somaclonal. Esta forma de variación puede llegar a generar cambios estables y transmisibles que conducen a cambios en la expresión génica (Kaeppler et al., 2000; Cardone et al., 2004.).
Los mecanismos por los cuales ocurre la variación somaclonal no han sido completamente dilucidados, pero entre sus causas se mencionan alteraciones en el cariotipo, mutaciones puntuales, recombinación somática e intercambio de cromátidas hermanas, elementos genéticos móviles (transposones y retrotransposones) y alteración en los patrones de metilación del ADN . (Phillips et al., 1994; Kaeppler et al., 2000; Cardone et al., 2004). En general , las mutaciones pueden ser inducidas por medio de tratamientos específicos con agentes mutágenos físicos y químicos, o bien por medio del cultivo de tejidos (Predieri, 2001 ). La aparición de la variación somaclonal obtenida a partir del CTV depende de varios factores como: genotipo, nivel de ploidía, fuente de explante, tiempo de cultivo, la vía de regeneración y las condiciones y composición del medio de cultivo (Skirvin et al. , 1994; Duncan, 1997; Cardone et al., 2004}. Rani y Raina (2000} mencionan que una causa importante de variación somaclonal la conforman los reguladores de crecimiento como son el 2,4-D y el ANA, el cual a elevadas concentraciones causan aberraciones cromosomicas durante el cultivo in vitro. Arnold {2008} menciona que de forma general, el uso de 2,4-D y una prolongada fase de tejido calloso, son los responsables de inducir variación ya sea de tipo genética como epigenética durante el cultivo in vitro. De manera general, Novak (1991) menciona que entre los factores que causan la variación somaclonal son: tipo y grado de diferenciación del explante, vía de regeneración, nivel de ploidía del genotipo y tiempo de cultivo. Los explantes más diferenciados (embriones cigóticos y yemas axilares) presentan menos o nula variación genética, mientras que los menos diferenciados (células meristemáticas, cambium y tejido calloso) tienden a aumentar la frecuencia de variación. En cuanto a la vía de regeneración se refiere, la embriogénesis somática genera
17

menores alteraciones que la organogénesis, para ambos casos, la vanac1on aumenta cuando la vía regeneración se lleva acabo de forma indirecta. Las especies poliploides tienden a incrementar la frecuencia de variación. El tiempo de cultivo y número de subcultivos aumentan también la probabilidad de que ocurra mayor variación genética (ver Fig. 1.2.).
ZYGOTIC El\lllRYO
MERISTEMTIP EXPLANT
DIRECT MORPHOGENESIS
CALLUS CELL(PROTOPLASD
~ INDIRECT MORPHOGENESIS CALLUS
1t~~~ •• ~ ~ ~
•D .~ •· (\ . (/• .o
SOMATIC ORGANO· AXILLARY EMBRYO· GENESIS
IN VITRO BRANCHING GENESIS EMBRYO· GENE SIS GENESIS
GERMINATION
POLYSOMATIC SPECIES
NON-POLYSOMATIC SPECIES
TIME __ __. ...
SOMACLONAL VARATION
Figura 1.2. Frecuencia de la variación somaclonal en relación al tipo de explante, vía de regeneración y tiempo de cultivo (Novak, 1991).
La variación somaclonal resulta ser una herramienta que puede ser empleada en los programas de mejoramiento vegetal en cultivares de importancia económica mediante la selección in vitro de mutantes con características agronómicas deseables, entre los que se incluyen cierta tolerancia al estrés biótico o abiótico como son: plagas, enfermedades, sequía y salinidad, que pueden ser obtenidos in vitro en muy poco tiempo (Jain, 2001 ; Cardone et al. , 2004). Hasta el 2006, FAOIIAEA (Food and Agriculture Organization /lnternational Atomic Energy Agency) en su base de datos reportaron mas de 2500 variedades de plantas en más de 200 especies, obtenidas mediante técnicas de mutagénesis (Shu & Lagoda, 2007).
18

Para inducir la tolerancia al estrés abiótico se han empleado agentes estresantes como son el AICI3, NaCI y el polietilenglicol (PEG). Ben-Hayyim (1987) logró inducir la tolerancia al estrés salino y osmótico aplicando NaCI y PEG durante el cultivo in vitro en suspensiones celulares de naranja dulce (Citrus sinensis) y naranjo agrio (Citrus aurantium). En sorgo Duncan et al. (1995) trabajando con diferentes agentes estresantes (AICI3, NaCI y PEG), obtuvieron mediante selección in vitro líneas tolerantes al aluminio, salinidad y sequía. En trigo (Triticum aestivum L.) Benderradji et al. (2007) desarrollaron un sistema para la obtención de líneas tolerantes a salinidad mediante la aplicación de NaCI durante la formación de callo.
1.4.2 El polietilenglicol (PEG) y la variación somaclonal
El polietilenglicol (PEG) es un alcohol polimérico hidrofílico con una alta solubilidad en agua y baja toxicidad (Fontana et al., 2001 ). Actúa como un agente osmótico no penetrante que disminuye el potencial hídrico del medio de cultivo, cuyo efecto es producir un estrés hídrico en las células vegetales (Aimansouri et al., 2001 ).
Debido sus propiedades como agente osmótico, el PEG ha sido utilizado ampliamente en programas de mejoramiento genético mediante la selección bajo presión para la obtención de líneas tolerancia al estrés hídrico. Biswas et al. (2002) indica que al simular una sequía química bajo condiciones de cultivo in vitro, el PEG altera los patrones de respuesta durante la morfogénesis e inducción de tejido calloso. Este compuesto se ha empleado para la selección temprana de líneas tolerantes a estrés hídrico en especies de importancia económica durante el cultivo in vitro en suspensiones celulares de tomate (Handa et al., 1983), en células de chile (Santos-Díaz & Ochoa-Aiejo, 1994 ), durante el cultivo en medio semisólido de embriones somáticos de sorgo (Duncan et al., 1995), en los embriones somáticos de arroz en medio semisólido (Biswas et al., 2002), durante la proliferación de brotes en plantas de banano en medio líquido y semisólido (Ebrahim et al., 2006). El empleo de PEG durante la regeneración de plantas in vitro, posiblemente induce de novo ciertas características de tolerancia a sequía (Biswas et al. , 2002). Trabajando con Capsicum annum (Santos-Díaz & OchoaAiejo, 1994) aplicaron una presión de selección con 15% de PEG (PM-8000) y realizando 5 subcultivos cada 15 días con 20-25% de PEG, obtuvieron líneas celulares tolerantes a estrés hídrico pero sin llegar a regenerar plantas completas .
En la Tabla 1.3 se puede observar las concentración de PEG (PEG 6000 y PEG 8000) empleada para inducir la tolerancia al estrés hídrico en diferentes cultivos.
19

Tabla 1.3. Concentración de PEG empleada para inducir la tolerancia al estrés hídrico en diferentes cultivos.
Cultivo Concentración Referencia
Tomate (Lycopersicon escu/entum Mili.) Chile (Capsicum annuum L.) Sorgo (Sorghum bicolor (L.) Moench.) Trigo (Triticum durum Desf.) Arroz (Oryza sativa L.) Palma datilera (Phoenix dactylifera L.) Banano (Musa spp.)
dePEG 20-25%
(PEG 6000) 15%
(PEG 8000) 9-12%
(PEG 8000) 18%
(PEG 6000) 1%
(PEG 6000) 15%
(PEG 8000) 1.5%
(PEG 6000)
Handa et al. (1983)
Santos-Díaz y OchoaAiejo (1994) Duncan et al. (1995)
Almansouri et at. (2001)
Biswas et al. (2002)
AI-Khayri & AI-Bahrany (2004) Ebrahim et al, (2006)
Aunque los caracteres morfológicos (ej. hábito de crecim iento, morfología, color floral , etc.) son fáciles de evaluar, muchos aspectos de la variación somaclonal suceden sin manifestarse en cambios morfológicos evidentes. Una diferencia estructural en un producto génico puede no alterar su actividad biológica lo suficiente como para producir un fenotipo modificado. Por ello, la variación debe analizarse a diferentes niveles (Cardone et al., 2004).
1.4.3 Niveles de detección de la variación somática
La frecuencia de variación somaclonal puede detectarse con marcadores fenotípicos, bioquímicos y moleculares. La correlación de dichos marcadores con caracteres agronómicos es un requisito relevante para su implementación en los programas de mejoramiento genético (Cardone et al., 2004). A continuación se citan algunos ejemplos de diferentes tipos de marcadores utilizados en la evaluación de la variación somaclonal :
Marcadores morfológicos. El uso de este tipo de marcadores para detectar la variación somaclonal se basa en visualización del fenotipo de las plantas regeneradas in vitro. El empleo de descriptores morfológicos resulta útil para este tipo de estudio. Cassels et al. (1997) por medio de un analizador de imágenes visualizaron la diferenciación entre fenotipos normales y aberrantes de plantas regeneradas de Pe/argonium. Anu et al. (2004) lograron identificar la variación somaclonal en chile (Capsicum annuum L.) basandose un descriptor del IPGRI
20

(lnternational Plant Genetic Resources lnstitute) específico para Capsicum spp. Plantas de cacahuate regeneradas in vitro presentaron variaciones ex vitro en cuanto a su altura, tamaño de hojas, número y peso de semillas (Cardone et al. , 2004).
Marcadores fisiológicos. Estos estudios se basan en una alta presión de selección durante la etapa de regeneración celular mediante la adición de agentes selectivos en el medio de cultivo durante el establecimiento y la regeneración, estos agentes pueden ser: herbicidas, pHs extremos, metales pesados, concentraciones salinas elevadas o condiciones de estrés osmótico. Mercado et al. (2000} en tomate (Lycopersicon esculentum Mili.) evaluaron el efecto de diferentes concentraciones de NaCI durante la regeneración in vitro de esta especie, como resultado lograron obtener sOmaclones tolerantes a salinidad. Ebrahim et al. (2006} demostraron que al aplicar PEG al medio de cultivo se logran obtener somaclones tolerantes a sequía.
Análisis de cariotipo. Este estudio permite detectar cambios en el número y estructura de los cromosomas. En cuanto al número de cromosomas se pueden encontrar las aneuploidias y poliploidias. Por otro lado, las posibles alteraciones en la estructura de los cromosomas son: translocaciones, inversiones, deleciones y duplicaciones. Los cambios en el cariotipo constituyen una fuente importante de variación que muchas veces es subestimada cuando se realizan recuentos cromosómicos. Nutrí Ronchi et al. (1981) mediante un análisis citológico de cariotipo lograron detectar aneuploidias en el cultivo in vitro de dos especies de tabaco (Nicotiana spp.). Sin embargo, mediante esta misma técnica, Ahmad et al. (2006} lograron constatar la estabilidad genética en plantas regeneradas in vitro chile (Capsicum annuum L.). Existen otras técnicas que estiman cambios en el cariotipo con mayor eficiencia, como por ejemplo el bandeo cromosómico. Esta técnica fue aplicada a suspensiones celulares de Brachycome dichromosomatica (2n=4), donde permitió detectar rearreglos importantes en individuos donde el número cromosómico permaneció estable (Cardone et al., 2004).
Marcadores bioquímicos. Los marcadores bioquímicos se basan en las técnicas de electroforesis y métodos citohistológicos de coloración, lo que permite el estudio de isoenzimas, las cuales pueden ser utilizadas como marcadores para revelar la información contenida en un gen. La aplicación de métodos cuantitativos a esta información permite el cálculo de diversos parámetros utilizados en la determinación de la estructura genética y reproductiva de una población natural. Una limitante de esta técnica, es que los marcadores bioquímicos son regulados a través del desarrollo, es decir, solo serían expresados fenotípicamente en estados específicos del desarrollo y en órganos o tejido específico. Además, los caracteres morfológicos pueden tener efectos pleiotrópicos en características de interés. Por otra parte, los marcadores
21

bioquímicos no son tan útiles como los marcadores de ADN, debido al bajo nivel de polimorfismo y el limitado número de loci. Las isoenzimas, por lo tanto, pueden proveer información útil acerca de la variación generada in vitro. Sin embargo, se están analizando los productos de genes expresados cuya regulación puede estar en cierta medida afectada por factores ambientales o fisiológicos.
Mediante las técnicas antes mencionadas no es posible detectar diferentes tipos de mutaciones como son: mutaciones puntuales, cambios en los patrones de metilación del ADN y/o elementos genéticos móviles (transposones y retrotransposones). Por ello, los métodos de análisis por medio de marcadores moleculares pueden proporcionar mucho mas información, ya que las secuencias de ADN (hasta que no se demuestre lo contrario) son genéticamente las mismas en todas las células vivas de la planta, independientemente del estado fisiológico o de desarrollo de las mismas.
1.5 MARCADORES MOLECULARES
1.5.1 Tipos de marcadores moleculares
Un marcador molecular es un fragmento o secuencia de ADN que corresponde a una característica que puede ser detectada y su herencia monitoreada (Ferreira y Grattapaglia, 1998).
Este tipo de marcadores detecta vanac1ones directas a nivel de ADN y tiene ventajas tales como el hecho de ser dominantes o codominantes, de desarrollarse de manera estable, su amplia cobertura del genoma, y sobre todo de no estar influenciados por el ambiente ni al estado de desarrollo del organismo en estud io. Las propiedades antes mencionadas hacen que éstos sean extremadamente útiles en comparación con los análisis a nivel morfológico, fisiológico o de proteínas (del tipo isoenzimas) (Valadez & Günter, 2000; Cardone et al., 2004).
Para obtener marcadores de ADN se usan diferentes métodos, la mayoría de estos marcadores usan la hibridación o están basados en la Reacción en Cadena de la Pol imerasa o PCR (Polymerase Chain Reacton) y se pueden agrupar de manera convencional en tres categorías (Caetano-Anollés & Gresshoff, 1997; Valadez & Günter, 2000):
Técnicas basadas en la hibridación. Esta técnica tiene el propósito de explorar las variaciones en la longitud de los fragmentos de ADN, ocasionados por la digestión del genoma con alguna endonucleasa particular. Esta categoría incluye principalmente los RFLP (Restriction Fragment Length Polymorphism) y a los VNTR (Variable Number of Tandem Repeats), donde las sondas son hibridadas a membranas que contienen ADN fragmentado por enzimas de digestión.
22

Técnicas basadas en PCR que usan iniciadores arbitrarios. Tienen la característica de no necesitar información de la secuencia del genoma que se esta investigando. Esta tecnología utiliza secuencias de oligonucleótidos que inician la síntesis in vitro de fragmentos de ADN de longitudes variables (no mayores a 6 Kb.). Para esta tecnología, se pueden citar por ejemplo, los RAPO (Randomly Amplified Polymorphic DNA), AFLP (Amplified Fragment Length Polymorphism) y los ISSR (lnter Simple Sequence Repeat).
Técnicas basadas en PCR de secuencia específica. Para el uso de estos marcadores es un requisito necesario el conocimiento de la secuencia de las zonas flanqueadas para la obtención de iniciadores. Dentro de esta categoría cabe mencionar a los CAPS (Cieaved Ampl ified Polymorphic Sequence) y los microsatélite (SSR, Simple Sequence Repeats).
Los marcadores moleculares son ampliamente utilizados en genética humana, animal, microbiana y vegetal. Estos permiten evidenciar vanac1ones (polimorfismos) en la secuencia del ADN entre dos individuos sin importar que éstos modifiquen o no su fenotipo. En plantas son empleados en estudios evolutivos y de genética poblacional, caracterización y protección legal de germoplasma, mapeo de genes y en la detección de la variación somaclonal (Cardone et al., 2004). Actualmente, el empleo de marcadores de ADN permite aumentar la eficiencia en la selección de plantas y/o de variedades en los programas de mejoramiento genético. Esta metodología es conocida como selección temprana asistida por marcadores moleculares (Ribaut y Hoistong, 1998) y reduce notablemente el tiempo de selección durante la mejora genética de plantas (Carrera et al., 2004 ).
1.5.2 Uso de los RAPO para el análisis de la variación somaclonal
Esta técnica consiste en la amplificación de ADN genómico usando iniciadores de secuencia arbitraria de 1 O nucleótidos de longitud y una composición en G+C entre 50 y 80%. Los polimorfismos que detecta son generados por pequeños cambios o por rearreglos genómicos (inserción o deleciones), dichos cambios permite la estimación de variabilidad genética en individuos o poblaciones. Las características de los RAPOs son las siguientes: no es radioactiva, fácil de manejar, requiere pequeñas cantidades de ADN y puede detectar polimorfismos en secuencias altamente repetidas . Sin embargo, presenta las desventajas de ser de naturaleza dominante en loci no específicos y ser de baja reproducibilidad (Saliba-Colombani et al., 2000).
23

En el CTV, la técnica RAPO es suficientemente específica debido a que permite diferenciar entre líneas de plantas y suficientemente sensible para discriminar tejidos en diferentes estadios de cultivo y entre las plantas regeneradas, aún cuando entre ellas no se detecten anormalidades morfológicas (Brown et al., 1993). Así, el cultivo in vitro parece incrementar el número de sitios donde los iniciadores pueden anclarse para iniciar la amplificación del ADN, si se compara con el número de sitios en las plantas control; esto trae como consecuencia que el método es válido para detectar semejanzas y diferencias entre los somaclones, pero no para identificar en forma precisa cada uno de ellos, pues ninguno de los iniciadores utilizados produce patrones de amplificación únicos para cada línea (Banerjee et al., 1997}.
La variación somaclonal, ha sido detectada a partir del empleo de marcadores moleculares tipo RAPO en arroz (Banerjee et al., 1997}, en durazno (Hashmi et al., 1997}, en orquídea (Chen et al. , 1998}, en tomate (Rus-Kortekaas et al., 1994; Soniya et al., 2001 ), en banano (Giménez et al., 2001 ), en almendro (Martins et al., 2004}, entre otros. En chile (C. annuum L.), Hossain et al. (2003) empleando RAPO detectaron variación genética en plantas regeneradas vía organogénesis directa. De los 15 cebadores que utilizaron, tres resultaron polimórficos. Esta variación también fue detectada a través de marcadores morfológicos, observándose diferencias en cuanto al hábito de crecimiento, color del tallo, contenido de antocianinas y cambios en la coloración de la flor y del fruto. Esta variación generada in vitro puede ser aprovechada para el mejoramiento genético en C. annuum y en otras especies (Hossain et al., 2003)
La aparición de fragmentos únicos en los regenerantes podría indicar la presencia de loci hipervariables o de secuencias de genes con una gran susceptibilidad a la inducción de estrés durante el cultivo de tejidos . Estas secuencias pueden ser clonadas y utilizadas como marcadores para el estrés en cuestión, y pueden correlacionarse con la tolerancia bajo ciertas condiciones de crecimiento (Brown et al. , 1993).
1.5.3 Uso de los ISSR para el análisis de la variación somaclonal
Los marcadores inter-microsatélite (ISSR, lnter Simple Sequence Repeat) en una técnica basada en la PCR y han sido desarrollados a partir de las repeticiones microsatélites (con, di, tri, tetra o penta repeticiones de nucleótidos) mediante el anclaje (5' o 3'} de cebadores arbitrarios para dichas repeticiones . Estos marcadores ISSR poseen características que los hacen atractivos para la diferenciación varietal de diferentes especies de eucariontes (Lagercrantz et al., 1993; Zietkiewicz et al., 1994}.
24

Los marcadores ISSR, contrario a los marcadores microsatélites (SSR), son marcadores dominantes y no necesitan información previa de la secuencia que amplifican, mostrando ser más reproducibles que los RAPOs (Random amplified polymorphic ADN), además son considerados altamente polimórficos. Estas características los hacen un marcador idóneo para realizar estudios de diversidad genética, para establecer huella genética y como marcador de selección en el mejoramiento genético de plantas (Pradeep et al., 2002). Esta técnica ha sido usada en estudios de diversidad genética en arroz (Oryza sativa) (Biair et al., 1999; Virk et al., 2000), naranja trifoliada (Poncirus trifo/iata) (Fang et al., 1997), frijol (Phaseolus vulgaris) (Galvan et al., 2003), sorgo (Sorghum bicolor) , banano (Musa acuminata) (Godwin et al. 1997), trigo (Triticum spp.) (Nagaoka & Ogihara, 1997), papa (Solanum tuberosum) (Prevost & Wilkinson, 1999) y cebada silvestre (Hordeum vulgarae (Tanyolac, 2003).
Los ISSR pueden ser utilizados también como una herramienta para el estudio de la variación somaclonal. Monte-Corvo et al., (2001) lograron identificar variación entre somaclones de pera (Pyrus communís). Sin embargo, en almendro (Martins et al., 2004) y banano (Lakshmanan et al., 2007), no se logró detectar variación somaclonal mediante esta técnica.
25

1.6 HIPÓTESIS Y OBJETIVOS
1.6.1 Hipótesis
Si las vías de regeneración constituyen una fuente de vanac1on somaclonal durante el cultivo in vitro de chile Habanero, entonces esta variación puede ser utilizada como una herramienta para la obtención de líneas tolerantes al estrés hídrico mediante la selección bajo presión con PEG.
1.6.2 Objetivo general
Estudiar la variación somaclonal generada a partir de diferentes vías de regeneración in vitro de chile Habanero (Capsicum chinense Jacq.), y utilizar esta variación para seleccionar una línea embriogénica tolerante a estrés hídrico usando PEG como agente selectivo.
1.6.3 Objetivos específicos
Evaluar mediante marcadores moleculares RAPOs e ISSRs, la variación somaclonal inducida durante la organogénesis directa y la embriogénesis somática directa e indirecta de chile Habanero.
Establecer un modelo de selección bajo presión para la obtención de una línea tolerante a estrés hídrico.
Determinar la variación genética de las líneas embriogénicas bajo diferentes tratamientos con PEG mediante la técnica RAPO.
Estudiar el efecto del PEG sobre el contenido de prolina en los embriones somáticos de chile Habanero.
26

1. 7 BIBLIOGRAFÍA
Agrawal, S., N. Chandra and S.L. Kothari (1989). Plant regeneration in tissue cultures of pepper (Capsicum annuum L. cv 'MathaniaJ. Plant Cell Tissue Organ Cult., 16, 47-55.
Ahman, N., l. Siddique and M. Anís (2006). lmproved plant regeneration in Capsicum annum L. from nodal segments. Biología Plantarum, 50(4), 701-704.
AI-Khayri, J. and A. AI-Bahrany (2004). Growth, water content, and proline accumulation in drought-stressed callus of date palm. Biología Plantarum, 48(1 ), 105-108.
Almansouri , M, Kinet J. and S. Lutts (2001 ). Effect of salt and osmotic stresses on germination in durum wheat (Triticum durum Desf.). Plant and Soil, 231, 243-254.
Anu, A. , K. Babu and K. Peter (2004). Variations among somaclones and its seedling progeny in Capsicum annuum. Plant Cell , Tissue and Organ Culture, 76, 261-267.
Arnold , S.V. (2008). Somatic Embryogenesis. In: George E. , M. Hall and G. De Klerk G.(ed). Plant Propagation by Tissue Culture 3rd. Springer. Edition Chapter, 9, 335-354.
Banerjee, H., V. Chimote and S.K. Raina (1997). DNA polymorphism among rice somaclones. Biologia Plantarum , 40, 543-533.
Bello-Bello, J. J., A. Canto-Fiick, E. Balam-Uc, E. Gómez-Uc, M. L. Robert, L. G. lglesias-Andreu and N. Santana-Buzzy (2010). lmprovement of in vitro pro/iferation and elongation of Habanero pepper shoots (Capsicum chinense Jacq.) by Temporary lmmersion. Hortscience, 45(7), 1-6.
Benderradji, L., H. Bouzerzou1, A. Djekoun, N. Yekhlef and A. Benmahammed (2007). Effects of NaCI Stress on Callus Proliferation and Plant Regeneration from Mature Embryos of Bread Wheat (Triticum aestivum L.) Cultivars Mahon Demias and Hidhab. Plant Tissue Cult. & Biotech., 17(1 ), 19-27.
Ben-Hayyim, G. (1987). Relationship between Salt Tolerance and Resistance to Polyethylene Glycol-/nduced Water Stress in Cultured Citrus Cells. Plant Physiol., 85, 430-433.
Binzel, M. L., N. Sankhla, S. Joshi and D. Sankhla (1996). lnduction of direct somatic embryogenesis and plant regeneration in pepper (Capsicum annuum L.) . Plant Cell Reports, 15, 536-540.
Biswas, J. , B. Chowdhury, A. Bhattacharya and A. Mandal (2002). In vitro screening for increased drought tolerance in rice. In Vitro Cell . Dev. Bioi.-Piant. ,38, 525-530.
Blair, M.W., O. Panaud and S.R. McCough (1999). lnter simple sequence repeat (ISSR) amplification for analysis of microsatellite motif frequency and fingerprinting of rice (Oryza sativa L.) . Theor. Appl. Genet. , 98, 780-792.
Bohnert H.J., D.E. Nelson and R.G Jensen (1995). Adaptations to environmental stresses. Plant cell, 7, 1099-1111 .
27

Brown, P.T. , F.O. Lange. E. Franz and H. Lorz. (1993). Analysis of single protoplast and regenerated plants by PCR and RAPO technology. Mol. Gen. Genet., 237, 311-317.
Brugiere N., F. Dubois, A. Limami, M. Lelandais, Y. Roux, R. Sangwan and B. Hirel (1999). Glutamine synthetase in the phloem plays a major role in controlling proline production. Plant Cell , 11 , 1995-2011 . .
Buyukalaca, S. and F. Mavituna. (1996). Somatic embryogenesis and plant regeneration of papper in liquid media. Plant Cell , Tissue and Organ Culture, 46, 227-235.
Caetano-Anollés, G., and P. Gresshoff (1997). ONA Markers: Protocols, Aplications, and Overviews. Wiley-Liss (ed), lnc. USA pp. 353.
Cardone, S. , S. Olmos and V. Echenique. (2004). Variación somaclonal. In: Echenique V. , Rubinstein C. , y Mroginski L. (eds). Biotecnología y Mejoramiento Vegetal. Ediciones INTA., 111 (1), 83-96.
Carrera, A., G. Tranquilli y M. Helguera. (2004). Aplicaciones de /os marcadores moleculares. In: Echenique V., Rubinstein C., Mroginski L. (Eds). Biotecnología y Mejoramiento Vegetal. Ediciones INTA., IV (2), 149-160.
Cassels, A.C., J.T. Croke and B. M. Doyle. (1997) . Evaluation of lmage Analysis, Flow Cytometry, and RAPO Ana/ysis for the Assessment of Somac/onal Variation and lnduced Mutation in Tissue Culture-Oerived Pe/argonium Plants. Angew. Bot. , 71 , 125-130.
Chen, W., T. Chen., Y. Fu., and R. Hsieh. (1998). Studies on somaclonal variation in Phalaenopsis. Plant Cell Reports, 18, 7-13.
Davies, P.J. (1995). The Plant Hormones: Their Nature, Ocurrence, and Functions. In: Davies P.J. (Ed). Plant Hormones. Physiology, Biochemistry and Molecular Bio/ogy. Kluwer Academic Publisher, Dordrecht, pp 1-11.
Duncan, R.R. (1997) Tissue culture-induced variation and crop improvement. Adv. Agron ., 58, 201-240.
Duncan, R.R. , R.M. Waskom and M.W. Nabors (1995). In vitro screening and field evaluation of tissue-culture-regenerated sorghum (Sorghum bicolor (L.) Moench) for soil stress tolerance. Euphytica, 85, 373-380.
Ebrahim, M. K., I.A lbrahim, H.A. Emara and E. Komor (2006). lmpact of polyethy/ene glycol-induced water stress on growth and deve/opment of shoot tip cultures from different banana (Musa spp.) cu/tivars. Journal of Applied Horticultura, 8(1 ), 3-57.
Fang, D., M. Roose, R. Krueger and C. Federici (1997). Fingerprinting of trifoliate orange germplasm accessions with isozymes, RFLPs, and ínter-simple sequence repeat markers. Theor. Appl. Genet. , 95, 211-219.
Fontana, G. , M. Licciardi, S. Mansueto. , D. Schillaci and G. Giammona (2001 ). Amoxicillin-/oaded polyethylcyanoacrylate nanopartic/es: lnfluence of PEG coating on the particle size, drug release rafe and phagocytic uptake. Biomaterial, 22, 2857-2865.
28

Galvan, M., B. Bornet., P. Balatti and M. Branchard . (2003}. lnter simple sequence repeat (/SSR) markers as a too/ for the assessment of both genetic diversity and gene pool origin in common bean (Phaseolus vulgaris L.) . Euphytica, 132, 297-301 .
Giménez, C. and E. García. (2001 ). Somaclonal variation in Banana: Cytogenetic and molecular Characterization of the somaclonal variant CIEN BTA-03. In Vitro Cell . Dev. Bioi.-Piant, 37, 217-222
Godwin, l. D., E.A. Aitken, and L.W. Smith (1997). Application of ínter simple sequence (/SSR) markers to plant genetics. Electrophoresis, 18, 1524-1528.
González, A. (2007). En México, varios daños del calentamiento global ya son irreversibles, alerta el BM. La Jornada (http://www.jornada.unam.mx/2007/1 0/12/)
González-Estrada, T., L.C. Gutiérrez-Pacheco y F. Contreras-Martín. (2006).EI chile Habanero de Yucatán .Ciencia y Desarrollo, Mayo, 9-15.
Handa, A.K. , R.A. Bressan, S. Handa and P.M. Hasegawa. (1983). Clona! Variation for Tolerance to Polyethylene Glycol-lnduced Water Stress in Cultured Tomato Cells. Plant Physiol., 72, 645-653.
Hashmi G, R. Huettel, R. Meyer, L. Krusberg and F. Hammerschlag. (1997). RAPO analysis of somaclonal variants derived from embryo callus cultures of peach. Plant Cell Rep. , 16, 624-627.
Hicks, G.S. (1994). Shoot induction and organogenesis in vitro: A developmental perspective. In vitro Cell Dev. Biol. , 30, 10-15.
Hossain A. , K. Konisho, M. Minami and K. Nemoto (2003). Somaclonal variation of regenerated plants in chili pepper (Capsicum annuum L.). Euphytica, 130, 233-239.
Hsiao, T.C. (1973). Plant response to water stress. Annual Review of Plant Physiology, 24, 519-570.
IPCC (2001 ). Climate change 2001: the scientific basis. In: Houghton J., Y. Ding, J. Griggs, M. Noguer, P. Van der Linden, X. Dai, K. Maskell and C. Johnson(eds). Third Assessment Report of the lntergovernmental Panel on Climate Change. Cambridge University Press, Cambridge.
lyer, S., and A. Caplan (1998}. Products of proline catabolism can induce osmotically regulated genes. Plant Physiology, 116, 203-211 .
Jain, S.M. (2001 ). Tissue culture-derived variation in crop improvement. Euphytica, 118, 53-166.
Kaeppler, S. , H. Kaeppler and Y. Rhee. (2000}. Epigenetic aspects of somaclonal variation in plants. Plant Molecular Biology, 43, 179-188.
Kiran, K.S. and T.A. Thorpe. (1995}. Asexual embryogenesis in vascular plants in nature. In: Thorpe T. A. (ed.) In vitro embryogenesis in plants. Kluwer Academia Publishers. Dondrecht. , pp. 17-72.
Kuznetsov, V. and N., Schevyakova (1999). Proline under stress: biological role, metabolism and regulation. Russian J. Plant Physiol., 46, 274-287.
29

Kuznetsov, V. and N., Schevyakova (1999). Proline under stress: biological role, metabolism and regulation. Russian J. Plant Physiol. , 46, 274-287.
Lagercrantz, U., H. Ellegren and T. Kakanuga (1993) . The abundance of various polymorphic microsatellite motifs differs between plants and vertebrales. Nucleic Acids Res., 21, 1111-1115.
Lakshmanan, V., S. Venkataramareddy and B. Neelwarne (2007). Molecular analysis of genetic stability in long-term micropropagated shoots of banana using RAPO and ISSR markers. Electronic Journal of Biotechnology, 1 0(1 ), 1-8.
Larcher, W. (1995). Physiological plant ecology. Ecophysiology and stress physiology of functional groups. Springer-Verlag, Berlin-Heidelberg.
Larkin, P.J. and S.C. Scowcroft {1981 ). Somaclonal variation: a novel source of variability from ce// culture for plant improvement. Theor Appl Genet, 60, 197-214.
Latournerie, L.M., M. E. Yupit, S.J . Tún, M.A. Pech y C.D. Lazada. (2006). Compartimiento de materiales criollos de Chile habanero ante el ataque de la virosis. Uuy u Táan. (Fundación Produce Yucatán, A.C.), 1,15-17.
Ledón, V. J. (2006) . Importancia del chile habanero en Yucatán. Uuy u Táan. (Fundación Produce Yucatán, A.C.), pp. 4-5.
Levitt, J. (1980).Responses of plants to environmental stresses. Vol. 1 Chilling, freezing, and high temperature stress. London, New York, Toronto: Academic Press, pp 698
López-Puc, G., A. Canto-Fiick, F. Barreda-Pool, P. Zapata-Castillo, M. Montalvo-Peniche, F. Barahona-Pérez and N. Santana-Buzzy {2006). Oirect Somatic Embryogenesis: A Highly Efficient Protocol for In Vitro Regeneration of Habanero Pepper (Capsicum chinense Jacq.) HortScience, 41 (6), 1-7.
Lutts, S., J.M. Kinet and J. Bouharmont (1996). Effects of salt stress on growth, mineral nutrition and pro/ine accumulation in relation to osmotic adjustment in rice (Oryza saliva L.) cultivars differing in salinity resistance . Plant Growth Regul, 19, 207-218.
Martins, M., D. Sarmento and M. Oliveira (2004) . Genetic stability of micropropagated almond plantlets, as assessed by RAPO and ISSR markers. Plant Cell Rep. , 23, 492-496.
Mercado, J., M. Sancho-Carrascosa and S. Jiméenez-Bermúudez. {2000). Assessment of in vitro growth of apical stem sections and adventitious organogenesis to evaluate salinity tolerance in cultivated tomato. Plant Cell , Tissue and Organ Culture, 62, 101-106.
Monte-Corvo, L., L. Goulao and C. Oliveira {2001 ). ISSR Analysis of Cultivars of Pear and Suitability of Molecular Markers for Clone Oiscrimination. J. Amer. Soc. Hort. Sci., 126{5), 517-522.
Nagaoka, T. and Y. Ogihara {1997). Applicability of intersimplesequence repeat polymorphisms in wheat for use as ONA markers in comparison to RFLP and RAPO markers. Theor. Appl. Genet. , 94, 597-602.
30

Novak, F.J. (1991 ). Plant tissue culture techniques for mutation breeding: a training manual. fnternationaf Atomic Energy Agency, Vienna. p. 182.
Pérez-Mofphe-Bafch, E. , R. Ramírez-Malagón, H. Núñez-Palenius y N. Ochoa-Afejo. (1999). Introducción al Cultivo de Tejidos Vegetales. Universidad Autónoma de Aguascalientes (ed), Primera edición México. pp. 179.
Phiffips, R. L, S.M. Kaeppler and P. Olhoft (1994). Genetic instability of plant tissue cultures: breakdown of normal controls. Proc. Natl. Acad. Sci. USA, 91, 5222-5226.
Pradeep, R.M., N. Sarla and E.A Siddiq (2002). lnter simple sequence repeat (ISSR) po/ymorphism and its application in plant breeding. Euphytica, 128( 1 ), 9-17.
Predieri, S. (2001 ). Mutation induction and tissue culture in improving fruits. Plant Ce//, Tissue and Organ Culture, 64, 185-210.
Prevost, A. and M. Wilkinson. (1999). A new system of comparing PCR primers applied to ISSR fingerprinting of potato cultivars. Theor. Appl. Genet., 98, 107-112.
Puebla, A. y F. Del Viso (2004). Tolerancia a factores abióticos. In: Echenique V., Rubinstein C., y Mroginski L. (eds). Biotecnología y Mejoramiento Vegetal. Ediciones INTA, Vllfl (12), 1-11 .
Rambal, S. and G. Debussche (1995). Water balance of Mediterranean ecosystems under a changing climate. In: Moreno J. and Oechel W. (eds). Global change and Mediterranean-type ecosystems. Springer Verlag, New York. pp. 386-407
Ramírez-Malagon, R. and N. Ochoa-Aiejo (1996). An improved and reliab/e chili pepper (Capsicum annuum L.) plant regeneration method. Plant Cell Reports, 16(3-4), 226-231 .
Rani, V. and S. Raina. (2000). Genetic fidelity of organized meristem-derived micropropagated plants: a critica/ reappraisa/. In Vitro Cell Dev Biol - Plant. , 36, 319-330.
Reichstein, M., J. Tenhunen, O. Roupsard, J. Ourcival , S. Rambal, F. Miglietta, A. Peressotti, M. Pecchiari, G. Tirone and R. Valentini. ( 2002). Severe drought effects on ecosystem C02 and H20 f/uxes at three Mediterranean evergreen sites: revision of current hypotheses? Global Change Biology, 8, 999-1017.
Ribaut, J. M. and D.A. Hoisington (1998). Marker assisted selection: new tools and strategies. Trends Plant Sci., 1998, 3, 236-239
Roy, B. and A. Manda!. (2005). Towards development of Al-toxicity tolerant fines in indica rice by exploiting somac/onal variation. Euphytica 145, 221-227.
Rus-Kortekaas, W. , M.J. Smulders, P. Arens and B. Vosman (1994). Direct comparison of /evels of genetic variation in tomato detected by GACA-containing microsatellite probe and random amplified polymorphic DNA. Genome, 37, 375-381 .
31

Sajid Aqeel Ahmad M., F. Javed and M. Ashraf (2007). lso-osmotic effect of NaCI and PEG on growth, cations and free proline accumulation in callus tissue of two indica rice (Oryza sativa L.) genotypes. Plant Growth Regul, 53, 53-63.
Saliba-Colombani, V., M. Causse, L. Gervais and J. Philouze (2000). Efficiency of RFLP, RAPO and AFLP markers for the construction of an intraspecific map of the tomato genome. Genome, 43, 29-40.
Santana-Buzzy, N., A. Canto-Fiick, F. Barahona-Pérez., M. Montalvo-Peniche, P. Y. Zapata-Castillo, A. Solís-Ruiz, A. Zaldívar-Collí., O. Gutiérrez-Aionso and M. L. Miranda-Ham (2005). Regeneration of Habanero Pepper (Capsicum chinense Jacq.) Via Organogenesis. HortScience, 40(6), 829-1831 .
Santana-Buzzy, N., A. Canto-Fiick, L. G. lglesias-Andreu ., M. MontalvoPeniche, G. López-Puc and F. Barahona-Pérez {2006). lmprovement of In vitro Culturing of Habanero Pepper by lnhibition of Ethylene Effects. HortScience, 41 (2), 405-409.
Santos-Díaz, M. and M. Ochoa-Aiejo (1994). PEG-tolerant ce// clones of chili pepper: Growth, osmotic potentia/s and so/ate accumulation. Plant Cell, Tiss . Org. Cult. , 37,1-8.
Shu, Q . and P. Lagoda. (2007). Mutation Techniques for Gene Discovery and Crop /mprovement. Molecular Plant Breeding, 5(2), 193-195.
Skirvin, R.M., K.D. McPheeters and M. Norton {1994) Sources and frequency of somac/onal variation. HortSci., 29, 1232-1246.
Soniya, E., N. Banerjee and M. Das {2001 ). Genetic ana/ysis of somac/onal variation among cal/us-derived plants of tomato .Current Science, 80{ 9), 1213-1215.
Steinitz, B., D. Wolf, T. Matzevich-Josef andA. Zelcer (1999). Regeneration in vitro and genetic transformation of pepper (Capsicum spp.): The current state of art. Capsicum and Eggplant News Lett, 18, 8-15.
Stewart, G. and J. Lee {1974). The role of proline acumulation in ha/ophytes.Pianta,120, 279-289.
Szweykowscy, A.J . {1993). Diccionario de Botánica (en polaco). Wiedza Powszechna, Varsovia. En: Reguladores del crecimiento, desarrollo y resitencia en plantas (2004) Jankiewicz Leszek S. (ed). Vol.1 (Propiedades y acción) UACH. Edición Mundi-Prensa. México, D.F. pp. 16-99 y 242-247.
Tanyolac, B. (2003). lnter-simple sequence repeat (ISSR) and RAPO variation among wild barley (Hordeum vu/garae subsp. Spontaneum) populations from west Turkey. Genet. Resour. Crop Evol., 50, 611-614.
Tun-Dzul, J. (2001 ). Chile habanero. Características y Tecnología de Producción, SAGARPA-INIFAP. Mocochá, Yucatán, México. p .5-6.
Unnikrishnan, S.K., L. Prakash, P.C. Josekutty, P.N. Bhatt and A.R. Mehta. (1991 ). Effect of NaC/ Salinity on Somatic Embryo Deve/opment in Sapindus trifoliatus L. Journal of Experimental Botany, 42, 401-406.
Valadez, M. E. y K. Günter ( 2000). Huellas de ADN en Genomas de Plantas (Teoría y protocolos de laboratorio). Editorial Mundi Prensa, Méx. pp. 147.
32
1

Valera-Montero, L. , and N. Ochoa-Aiejo (1992). A novel approach for chili pepper (Capsicum annuum L.) plant regeneration: shoot induction in rooted hypocotyls. Plant Sci. , 84, 215-219.
Valladares, F. and R.W. Pearcy (1997). lnteractions between water stress, sun-shade acclimation, heat tolerance and photoinhibition in the sclerophy/1 Heteromeles arbutifolia. Plant Cell and Environment, 20, 25-36.
Venkataiah, P. , T. Christopher and K. Subhash (2003). Thidiazuron induced high frecuency adventitious shoot formation and plant regeneration in Capsicum annuum L. J. Plant Biotechnology, 5, 245-250.
Vinocur, B. and A. Altman (2005). Recent advances in engineering plant tolerance to abiotic stress: achievements and limitations. Current Opinion in Biotechnology, 16, 123-132.
Virk, P.S., J. Zhu, H.J. Newbury, G.J. Bryan, M.T. Jackson and B.V. FordLioyd (2000). Effectiveness of different classes of molecular markers for classifying and revealing variation in rioce (Oryza sativa) germplasm. Euphytica, 112, 275-284.
Xiong L. and J.K. Zhu (2001 ). Abiotic estrés signa/ transduction in p/ants: molecular and genetic perspectives. Physiol. Plant. , 112, 152-166.
Zapata-Castillo, Y. P., A. Canto-Fiick, G. López-Puc, A. Solís-Ruiz., F. Barahona-Pérez and N. Santana-Buzzy (2007). Somatic Embiogenesis in Habanero Pepper (C. chinense Jacq.) From Ce// Suspensions. HortScience, 42(2), 1-5.
Zietkiewicz, E., A. Rafalski and D. Labuda (1994). Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics, 20, 176-183.
33

34

Capitulo 11
Materiales y Métodos
En las Figuras 2.1 y 2.2 se muestran las estrategias experimentales de este proyecto.
ESTRATEGiA EXPERIMENTA 1
--------- ----------i Accesión: Naranja 1
• 1
Inducción de la morfogénesis 1
.. 1 1 1
Planta Organogénes is E m briogénesis Callo Embriogénesis jonante directa directa embriogénico indirecta
1 1 1 1
1 Extracción de ADN
1
¡ Selección de cebadores: 20 RAPOs y 20 ISSRs
Análisis molecular de la variación somaclonal (Método UPGMA e índice de similitud de Jaccard)
Figura 2.1 . Estrategia experimental l. Análisis molecular de la variación somac/onal.
35
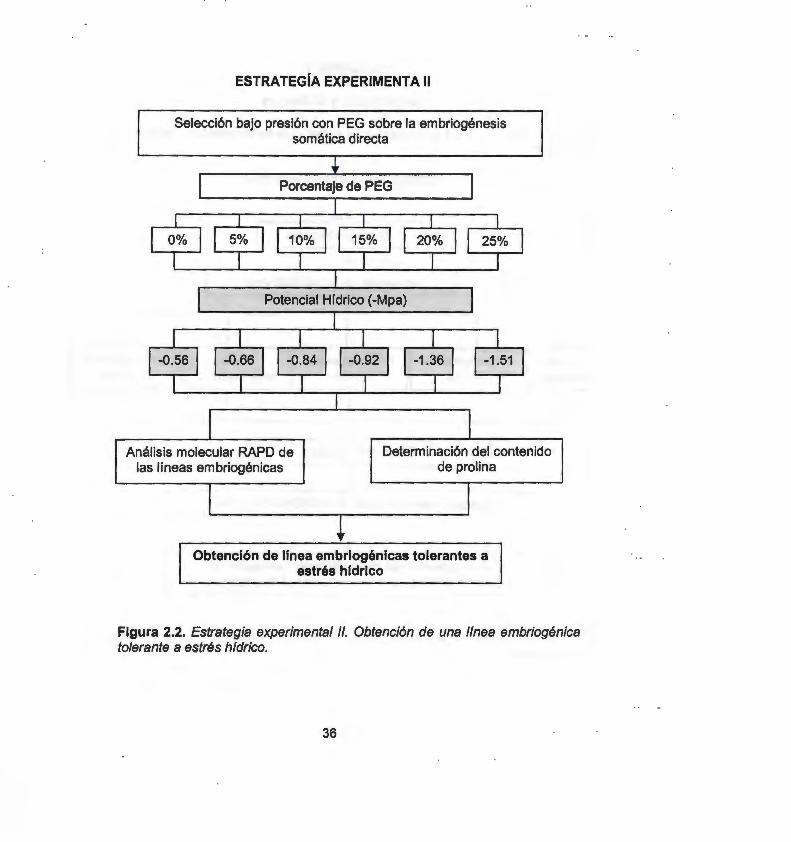
ESTRATEGÍA EXPERIMENTA 11
Selección bajo presión con PEG sobre la embriogénesis somática directa
+ 1
Porcentaje de PEG 1
1 1 1 1 1
1 0%
1 1 5% 1 1 1 O% 1 1 15% 1 1 20% 11 25% 1 1 1 1 1
1
1 Potencial Hídrico (-Mpa)
1
1
1 1 1 1
1 -0.56 1 1 -0.66 1 1 -0.84 1 1 -0.92 1 1 -1.36 1 1 -1 .51 1
1 1 1 1
1
1 1
Análisis molecular RAPD de Determinación del contenido las líneas embriogénicas de prolina
1 1
~ Obtención de línea embriogénicas tolerantes a
estrés hídrico
Figura 2.2. Estrategia experimental 11. Obtención de una línea embriogénica tolerante a estrés hídrico.
36

2.1 ASPECTOS GENERALES
2.1.1 Material vegetal y condiciones de cultivo. Se utilizaron semillas provenientes del banco de germoplasma registrado en el Centro de Investigación Científica de Yucatán (CICY) de la accesión de chile Habanero naranja (Capsicum chinense Jacq., accesión NSBm-04), estas fueron desinfectadas y germinadas in vitro de acuerdo a la metodología propuesta por Santana-Buzzy et al. (2005), en medio semisólido MS (Murashige y Skoog, 1962) suplementado con 1.15 1-1M de ácido giberélico (GA3). Una vez germinadas, fueron transferidas al mismo medio pero sin reguladores de crecimiento e incubadas en fotoperíodo para continuar su desarrollo. Posteriormente, se tomaron nudos e hipocótilos como fuente de explante para inducir los procesos e morfogénesis in vitro.
En todos los experimentos fue empleado el medio de cultivo MS (Anexo 1 ), 3% de sacarosa y como agente gelificante para el medio semisólido se usó gelrite al 0.22% (w/v), el pH fue siempre ajustado a 5.8 y los medios fueron esterilizados en autoclave a 1.5 Kg cm·2 de presión y 121 °C durante 15 minutos. Los explantes fueron incubados a una temperatura de 25±2°C, con fotoperiodo de 16 h luz y 8 h oscuridad a una intensidad lumínica de 40-50 1-1mol·m·2·s· .
2.2 INDUCCIÓN DE LOS PROCESOS DE MORFOGÉNESIS in vitro
2.2.1 Organogénesis directa
La organogénesis directa fue inducida siguiendo el protocolo propuesto por BelloBello et al. {201 0), utilizando nudos de 45-65 días de edad en medio MS + TDZ {3.4 1-1M) + PAC {3.4 1-1M), la elongación de lo brotes se llevó a cabo en medio MS+ GA3 {2.77 !JM) + AgN03 (10 IJM) y el enraizamiento en medio MS +AlA (1.70 IJM), todos los medios fueron en medio semisólido.
2.2.2 Embriogénesis somática directa
La embriogénesis somática directa fue inducida mediante el protocolo reportado por López-Puc et al. {2006) modificado por Avilés-Viñas (2007). Para ello, se utilizó como fuente de explante segmentos de hipocótilos incubados en medio semisólido MS + 2,4-D (9.05 1-1M), una vez formados los embriones, estos fueron transferidos a medió líquido MS a la mitad de 2,4-D (4.0 !JM) por 15 días, después fueron subcultivados a medio líquido con ABA {1 .89 !JM) para su maduración y para la germinación fueron transferidos a medio líquido con GA3 (1 .15 IJM).
37

2.2.3 Embriogénesis somática indirecta
El protocolo de embriogénesis somática indirecta se realizó de acuerdo a ZapataCastillo et al. (2007), en el cual se indujo la formación de callo a partir de hipocótilos en medio MS + 2,4-D (9.05 ¡.¡M). Una vez formado el callo, este es transferido a un sistema de de células en suspensión sustituyendo el 2,4-D por TDZ {3.5 ¡.¡M).
2.2.4 Formación de callo embriogénico
Para la formación de callo se siguió el mismo protocolo de embriogénesis somática indirecta de Zapata-Castillo et al. (2007), una vez formado el callo, este se trasfirió a un sistema de células en suspensión con 2,4-D a la mitad de su concentración (4.0 ¡.¡M) durante 2 subcultivos.
Una vez inducidos los procesos de morfogénesis in vitro, las muestras fueron maceradas con nitrógeno líquido y se mantuvieron a -80°C, posteriormente se liofilizaron para la extracción de ADN.
2.3 ANÁLISIS DE LA VARIACIÓN SOMACLONAL
2.3.1 Extracción de ADN
Para la extracción de ADN se utilizó el protocolo de Dellaporta (1983) (Anexo 2). En todos los casos se utilizaron 30 mg de tejido vegetal liofilizado. El ADN extraído fue resuspendido en 300 ¡.¡1 de agua bidestilada ultrapura estéril para su posterior tratamiento con RNAsa (Anexo 3). Finalmente, para determinar la concentración y pureza del ADN se realizaron lecturas de absorbancia de 260 y 280 nm en un espectrofotómetro marca Génesis TM 1 Ouv. Se visualizó la calidad de ADN mediante electroforesis en geles de agarosa al 1% usando como buffer TBE 1X y teñidos con bromuro de etidio. Para la corrida de ADN se utilizó una cámara de electroforesis submarino BioRad™ con una fuente de poder marca Consort™ modelo E844 para efectuar la separación a 90 volts durante 30 min. Se empleó un marcador de peso molecular de 1 kb plus (BioRad™) y se aplicaron 8 ¡.¡L de la solución de ADN resuspendido en los pocillos del gel con 2 ¡.¡L de buffer de carga 6X. Los geles fueron fotografiados con ayuda de un fotodocumentador (Universal Hood 11 Gel Doc marca BioRad™).
38

2.3.2. Selección y uso de cebadores RAPOs
Como primer paso se realizó un rastreo para identificar aquellos cebadores RAPO capaces de amplificar en genoma de chile Habanero. Para ello se empleó la técnica RAPO propuesta por Williams et al. (1990). Se evaluó un Kit de 20 cebadores BIOSELEC® (Anexo 4) para observar el patrón de bandeo que éstos pudieron amplificar mediante ensayos de ADN foliar de un pool de 5 individuos de la misma accesión. Aquellos cebadores que amplificaron fueron utilizados para analizar la variación somaclonal.
En el cóctel de reacción se preparó un volumen final de 30 IJL. La mezcla de reacción estuvo constituida por 3 IJL de buffer PCR 10X, 2 mM MgCI2, 100 1-1M de dNTPs, 5 pM de cebador, 2.5 unidades de Taq polimerasa y 300 ng de ADN, en todos los casos se emplearon reactivos de la marca lnvitrogen TM . Las reacciones de amplificación de PCR se llevaron a cabo en un termociclador My cycler BIORAD™ con el siguiente programa: 4 mina 94 oc; 40 ciclos de 1 min a 94 oc , 2 min a 32 ó 34 oc (según el cebador) y 1 min a 72oC; por ultimo se llevo acabo una extensión final de 72 oc por 7 min. Finalmente, los productos fueron visualizados mediante electroforesis en geles de agarosa al 2.5% usando como buffer TBE 1X teñidos con bromuro de etidio. Para estimar el tamaño de los productos se empleó un marcador de peso molecular de 1 kb-Pius (lnvitrogen™). Los geles fueron fotografiados con el fotodocumentador antes mencionado.
2.3.3. Selección y uso de cebadores ISSRs
Para los ISSRs se empleó la técnica propuesta por Zietkiewicz et al. (1994}. De forma similar a los RAPOs, se evaluó un kit de 20 cebadores (Anexo 5) y se seleccionaron aquellos cebadores que fueron capaces de amplificar en ADN genómico de la misma accesión de chile Habanero.
El coctel de reacción de los ISSRs se preparó a un volumen final de 30 IJL con las siguientes concentraciones: 3 IJL de buffer PCR 10X, 2 mM MgCI2, 200 IJM de dNTPs, 1 O 1JM de cebador, 2 unidades de Taq polimerasa y 100 ng de ADN, en todos los casos se emplearon reactivos de la marca lnvitrogen™. Las reacciones de amplificación de PCR se llevaron a cabo en un termociclador My cycler BIORAD™ con el siguiente programa: 4 min a 94 oc; 37 ciclos de 40 seg a 94 oc, 40 sega 35, 37, 40 6 45 oc (según el cebador) y 50 sega 72°C; por ultimo se llevo acabo una extensión final de 72 oc por 7 min. Posteriormente los productos fueron visualizados mediante electroforesis en geles poliacrilamida 6% usando como buffer TBE 1X. Para estimar el tamaño de los productos se empleó un marcador de peso molecular de 1 kb-Pius (lnvitrogen™).
39

Análisis de datos. El perfil de bandas fue analizado mediante una matriz binaria, asignando el número "1" para la presencia ó un "O" para la ausencia de una banda. Las distancia genéticas se llevaron acabo empleando el coeficiente de similitud de Jaccard (Jaccard, 1908}. Éste coeficiente fue usado para construir un dendograma mediante el método de agrupamiento por pares no ponderados con media aritmética (UPGMA, Unweighted Pair Group Method with Arithmetic Average) con ayuda del programa MVSP 3.0 (MultiVariate Statistical Package). Para poner a prueba el apoyo que tienen las ramas del árbol, se llevó acabo la prueba estadística del bootstrap con el programa FREETREE 7.0.
2.4 OBTENCIÓN DE UNA LÍNEA TOLERANTE AL ESTRÉS HÍDRICO
2.4.1 Selección bajo presión con PEG sobre la embriogénesis somática directa
En la Tabla 2.1 se muestra la escala propuesta por Hsiao (1973) para establecer el nivel de estrés hídrico o los grados de sequía a que pueden estar expuestas las plantas en la naturaleza y que será utilizada en este estudio.
Tabla 2.1. Grados de sequía según el descenso del potencial suelo y contenido relativo de agua (Hsiao, 1973).
hídrico del
Nivel de estrés Potencial hídrico del Contenido relativo de agua hídrico suelo (-MPa)
Suave Desciende de -0.5 a -0 .8 Se reduce entre 8 y 1 0%
Moderado Desciende de -1 .2 a -1 .5 Se reduce hasta un 20%
Severo Desciende > -1.5 Se reduce a valores > 20%
Para simular las condiciones de estrés hídrico in vitro , se adicionó polietilenglicol (PEG 8000 SIGMA ™) a diferentes concentraciones al medio de cultivo: O, 5, 1 O, 15, 20 y 25 %, que corresponden a un potencial hídrico de -0.56, -0.66, -0.84, -0.92, -1.36 y -1.51 Mpa, respectivamente. El sistema de regeneración utilizado fue el de Embriogénesis Somática Directa (ESD), por ser el más eficiente (mayor número de regenerantes o embriones por volumen de medio) y porque además, los análisis moleculares permitieron detectar un alto nivel de variación genética en sus regenerantes. En cada tratamiento fueron utilizados 50 explantes (hipocótilos), distribuidos a razón de 5 explantes por magenta. Una vez inducidos los embriones, en cada explante se obtuvieron en promedio 168 embriones en estadio globular, posteriormente estos fueron transferidos a medio líquido para
40

iniciar el proceso de selección bajo presión con PEG, los tratamientos estuvieron distribuidos de la siguiente manera: 5 explantes por matraz y cada matraz contenía 50 mi de medio de cultivo (se emplearon matraces de 250 mi). La tolerancia al estrés hídrico fue estimada mediante la dosis letal media (DL50}, y midiendo los porcentajes germinación en presencia de GA3 (1.15 1-1M) previo tratamiento con PEG.
El potencial hídrico del medio de cultivo se determinó con un medidor de potencial hídrico WP4 (Decagon-Devices, lnc.). En la Tabla 2.2 se observan la medida del potencial hídrico del medio de cultivo registrado en diferentes concentraciones de PEG (%).
Tabla 2.2. Potencial hídrico del medio de cultivo. Concentración de PEG Potencial hídrico en
(%) -MPa
o -0 .56 5 -0 .66 10 -0.84 15 -0.92 20 -1.36 25 -1.51
Análisis estadístico. Los datos recopilados fueron procesados utilizando el paquete estadístico SPSS 11.5 (para Windows). Se realizó un análisis de varianza (ANDEVA) y comparación de medias de acuerdo a Tukey (p =::0.05), cuando fue requerido.
2.4.2 Selección asistida por marcadores moleculares RAPOs
Con el objetivo de determinar si la respuesta de los embriones en condiciones de estrés hídrico (provocado por la presencia del PEG) es de naturaleza genética, se evaluaron los mismos cebadores RAPOs que fueron utilizados para evaluar la variación somaclonal (ver ANEXO 4). Para ello se seleccionaron los embriones somáticos más desarrollados, ya que fueron los que mostraron mayor capacidad de sobrevivencia a la presión de selección con PEG. Posteriormente se identificaron las bandas polimórficas como los posibles marcadores de ADN.
41

2.4.3 Determinación del contenido de prolina
A fin de determinar si la respuesta de los embriones en condiciones de estrés hídrico (provocado por la presencia del PEG) es de tipo fisiológica, se procedió a determinar el contenido de prolina como un posible marcador bioquímico. Para ello se seleccionaron los embriones somáticos que fueron capaces de sobrevivir bajo los diferentes tratamientos con PEG. La determinación del contenido de prolina se realizó siguiendo el método descrito por Bates et al. (1973}, basado en la reacción de la prolina con la ninhidrina), para lo cuál se maceró 250 mg de tejido embrionario fresco en un mortero, posteriormente se homogeneizó con 5 mi de ácido sulfosalicílico 3%. Los residuos fueron removidos con papel filtro, y del líquido libre de tejido foliar se tomó una alícuota de 1 mi al cual se le agregaron 1 mi de ácido acético glacial y 1 mi de ninhidrina (compuesto que al reaccionar con prolina a pH ácido forma un complejo coloreado). Esta mezcla se agitó en Vortex por 15 s y se dejó incubar a baño termorregulado por 1 h a 1 00°C. Se retiraron los tubos y se enfriaron en hielo rápidamente. Se añadió 2 mi de tolueno y se agitó por 30 s. La fase superior (tolueno + complejo coloreado) se retiró con pipeta. Finalmente, se leyó la absorbancia a 520 nm en un espectrofotómetro marca Génesis TM 1 Ouv. Los valores obtenidos se interpolaron en la curva de calibración hecha con estándar de L-prolina (SIGMA®).
2.5. BIBLIOGRAFÍA
Avilés-Viñas, S. (2007}. Papel del etileno durante la embriogénesis somática del chile habanero (Capsicum chinense Jacq.}.Tesis de maestría. CICY. Méx. 65 pp.
Buyukalaca, S. and F. Mavituna (1996). Somatic embryogenesis and p/ant regeneration of papper in liquid media. Plant Cell , Tissue and Organ Culture, 46, 227-235.
Dellaporta, S.L. , J. Wood and J.B. Hicks (1983). A plant DNA minipreparation: version 2. Plant Molecular Biology Reporter, 1, 19-22.
Hsiao, T.C. (1973). Plant response to water stress. Annual Review of Plant Physiology, 24, 519-570.
Kuznetsov, V. and N., Schevyakova (1999}. Proline under stress: biological role, metabolism and regulation. Russian J. Plant Physiol. , 46, 274-287.
López-Puc, G., A. Canto-Fiick, F. Barredo-Pool, P. Y. Zapata-Castillo, M. Montalvo-Peniche, F. Barahona-Pérez and N. Santana-Buzzy (2006}. Direct Somatic Embryogenesis : A Highly Efficient Protocol for In vitro Regeneration of Habanero Pepper (Capsicum chinense Jacq.). HortScience, 41 (6), 1-7.
Murashige, T. and F.A. Skoog (1962}. A revised medium for rapid growth and bioassays with tobacco cultures. Physiol. Plantarum, 15,473-497.
Santana-Buzzy, N., A. Canto-Fiick, F. Barahona-Pérez, M.C. MontalvoPeniche, P.Y. Zapata-Castillo, A. Solís-Ruíz, A. Zaldívar-Collí, O. Gutiérrez-Aionso
42

and M.L. Miranda-Ham (2005). Regeneration of Habanero Pepper (Capsicum chinense Jacq.) Vía Organogenesis. HortScience, 40(6), 829-1831.
Stewart, G. and J. Lee (1974). The role of proline acumulation in halophytes. Planta, 120, 279-289.
Williams, J.G., A.R. Kuubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey. (1990). DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. NucleicAcids Res. ,18, 6531-6535.
Zapata-Castillo, Y. P., A. Canto-Fiick, G. López-Puc, A. Solís-Ruiz, F. Barahona-Pérez and N. Santana-Buzzy. (2007). Somatic Embiogenesis in Habanero Pepper (C. chinense Jacq.) From Ce// Suspensions. HortScience, 42(2), 1-5.
Zietkiewicz, E., A. Rafalski and D. Labuda (1994). Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics, 20, 176-183.
43

44

In Vitro Cell Dev Bioi-Piant (Submitted Article)
Capitulo 111
Resultados y Discusión
3.1 ANÁLISIS MOLECULAR DE LA VARIACIÓN SOMACLONAL (ARTÍCULO SOMETIDO)
Somaclonal variation in Habanero pepper assessed by RAPO and ISSR markers
Jericó J. Bello-Bello and Nancy Santana-Buzz/
Unidad de Bioquímica y Biología Molecular de Plantas, Centro de Investigación Científica de Yucatán, Calle 43 # 130, Chuburná de Hidalgo, Mérida, Yucatán. CP. 97200, México
· ro whom reprint requests should be addressed: E-mail: [email protected]
3.1.1 Abstract
In order to evaluate the effects of in vitro culture on the genetic variation of Habanero pepper (Capsicum Chinense Jacq.) regeneration protocols, two different molecular markers were employed: random amplified polymorphic DNA (RAPD) and inter-simple sequence repeat (ISSR). A total of 62 RAPD and 219 ISSR clear and reproducible fragments were generated with 7 RAPD and 13 ISSR primers in direct organogenesis, direct and indirect somatic embryos and embryogenic callus system. Of the two different markers used, RAPD detected more polymorphism than ISSR (57.6% vs 27.5%, respectively). Two cluster analyses were performed to express - in the form of dendrograms - the relationships among different regeneration systems and the genetic variability detected. Genetic distance analysis indicates that our regeneration protocols are inappropriate for micropropagation, conservation or genetic transformation; however, they could be applicable for breeding. This is the first report on the use of molecular markers to evaluate genetic variation of in vitro regenerated plants of Habanero pepper using RAPD and ISSR.
Key words: Plant regeneration, organogenesis, somatic embryogenesis.
45

3.1.2 lntroduction
Larkin and Scowcroft ( 1981) coined the term somaclonal variation to cover genetic changes that occur in plants regenerated from cultured cells or tissues. The phenomenon of novel variation or somaclonal variation observed in tissue culture derived plants is well recognized. Regardless of the mechanism by which somaclonal variation is produced, evidence has accumulated demonstrating that such variation does produce useful variants (Evans& Sharp, 1983; Evans, 1989). Plant cell culture has often been hailed as one of the more significant potential adjuncts to plant improvement. This is usually seen in terms of the ability to apply cellular selection for recovering useful genetic variants, anther culture to speed the attainment of homozygosity, somatic hybridization for recombining genomes of sexually incompatible species and more recently the possibility of specific gene addition or modification by recombinant DNA techniques (Scowcroft, 1977; Thomas et al., 1979; Kado and Kleinhofs, 1980). These approaches have made only limited direct contribution to genotype improvement of commercial species to date. Often potentially valuable contributions of science emerge from unexpected quarters. Plant cell culture has provided a new and exciting option for obtaining increased genetic variability relatively rapidly and without sophisticated technology, generated by the use of a tissue culture cycle (Larkin & Scowcroft, 1981 ).
Severa! strategies are available for detecting genetic variation, including phenotypic identification and DNA analysis techniques. Phenotypic identification based on a description of the morphological and physiological traits can be used, although this method requires an extensive observation of the plants until maturity. Furthermore, some changes induced by in vitro culture cannot be observed because the structural difference in the gene product does not always alter its biological activity to such an extent that this change can also be observed in the phenotype. When this occurs, somaclonal variability can be evaluated by DNA analysis techniques. Various DNA markers have been developed, such as RFLP (restriction fragment length polymorphism), AFLP (amplified fragment length polymorphism), RAPO (random amplified polymorphic DNA), SSR (simple sequence repeats) and inter-simple sequence repeat (ISSR).
The occurrence of useful somaclonal variation has been demonstrated in many crop species, such as wheat (Hasim et al. , 1990), rice (Abe et al. , 1999), maize (Beckert et al., 1983), sugarcane (Larkin & Scowcroft, 1983), patato (Sibi, 1982; Austin & Cassells, 1983) and in tomato (Evans & Sharp, 1983). Somaclonal variations can be distinguished not only by their morphological traits (Maralappanavar et al., 2000) but also by DNA analysis (Chen et al., 1998). Nevertheless, critica! analysis of somaclonal variants in pepper in the literature is very scarce. The aim of this work was to evaluate somaclonal variation in
46

propagated plants of Habanero pepper by different regeneration via, using RAPOs and ISSR techniques.
3.1.3 Materials and Methods
Preparatíon of explants. Seeds of the accession NSNm-04 (orange type) of Habanero pepper (C. chinense) were first submerged in ethanol at 70% (v/v) for 5 min and subsequently in a solution of commercial chlorine (Cioralex, 6% active chlorine) diluted to 30% (v/v) for 15 min . The seeds were then rinsed three to four times with sterile distilled water and deposited in semisolid MS medium, supplemented with 1.15 J,JM giberelic acid (GA3) and incubated in darkness at 25 ± 2°C. U pon germination (1 O days), plantlets were transferred to the same MS medium without growth regulators. Photoperiod was 16 h light (40 to 50 ¡.Jmol· m-2
•
s-1) , at 25 ± 2°C.
Dírect organogenesís índuction. Nodes from 45 to 60 day-old plants were cultivated in MS semisolid medium supplemented with 3.4 J,JM TDZ + 3.4 J.JM PAC (paclobutrazol). The explants were incubated in photoperiod 16 h light ( 40 to 50 ¡.Jmol·m-2·s-1
), at 25 ± 2°C for two months, according to Bello-Bello et al. (2010). Once the shoots reached an adequate size ( ~ cm), leaf samples were taken for DNA extraction and subsequent molecular analyses.
Dírect somatic embryogenesis induction. Somatic embryogenesis was induced directly from hypocotyls segments taken from 15 day-old aseptic plants after in vítro germination of the seed. The hypocotyls explants were placed in MS semisolid medium supplemented with 2,4-dichlorophenoxyacetic acid (2,4-D; 9.05 J,JM) during one month, following the protocol of López-Puc et al. (2006). Photoperiod was 16 h light (40 to 50 J,Jmol·m-2·s-1
), at 25 ± 2°C.
lndirect somatíc embryogenesis ínductíon. For the induction of callus, hypocotyls were placed in MS semisolid medium with 9.05 J,JM (2,4-D) and incubation was carried out over a period of 30 d at 25 ±oc under continuous light (J,Jmol·m-2·s-1
).
Once the callus formed, it was transferred to a liquid medium maintaining the same composition as the initial medium but without Gelrite and with half the concentration of 2,4-D. The suspensions were established by inoculating 1 g of callus with 1 month of induction into 250 mi Erlenmeyer flasks containing 50 mi of liquid medium. The cultures were maintained in agitation at 100 rpm under the same light and temperatura conditions used in the callus induction stage. Subcultures were carried out every 14 d over a period of 2 months. lndirect somatic embryos were formed in liquid medium containing 3.4 J,JM thidiazuron (TDZ), according to Zapata-Castillo et al., 2007).
47

DNA extraction and PCR amplification
DNA extraction. Samples from embryogenic callus, somatic embryos and shoots provided plant tissue for ONA extraction to be used in molecular analyses. After liofilization, 30 mg of tissue were taken from each sample for the extraction of total genomic ONA. The protocol used for ONA extraction was Oellaporta (1983). The ONA concentration was determined using a Spectrometer (Genesis™ 10uv).
PCR amplifications. Twenty RAPOs (T-01 to T-20) primers, and twenty ISSR (IS-01 to IS-20) primers, were evaluated for ONA analysis (Table 3.1 ). ONA of Habanero pepper donor plants germinated in vitro was used as control. PCR amplification for both RAPOs and ISSR, was carried out in a Thermocycler (My cycler BIORAO™). RAPO amplification was performed according to Williams et al. (1990) in a total vol u me of 30 !JI containing 100 ng ONA template, 3 !JI of 1 O X polymerase chain reaction (PCR) buffer, 2 mM MgCI2, 200 !JM of each dNTP, 5 pM primer and 2.5 U Taq ONA polymerase (lnvitrogen™). ONA amplification was performed using the following cycling program : one time at 94 °C for 4 min; 40 cycles at 94 °C for 1 min, 32 or 34 °C (according primer) for 2 min, 72 °C for 1 min; and a final extension at 72 °C for 7 min. RAPO fragments were separated by electrophoresis on 2.5% agarose gels in 1X TBE. The gels were stained in ethidium bromide, visualized in a trans-illuminator (Universal Hood 11 Gel OocBioRad™) and photographed.
48

Table 3.1. RAPD and ISSR primers evaluated in genomic DNA of Habanero pepper (C. chinense) .
RAPD primer sequences (5'-3')
T-01 T-02 T-03 T-04 T-05 T-06 T-07 T-08 T-09 T-10
CATTCGAGCC GTCTCCGCAA CCAGCTTAGG CCGCCCAAAC TCTGTCGAGG CAC CTTTCCC AGCGAGGAAG GAACACTGGG CCCTACCGAC GTGCAACGTG
ISSR primer sequences (5'-3')
IS-01 IS-02 IS-03 IS-04 IS-05 IS-06 IS-07 IS-08 IS-09 IS-10
GTAGTAGATAGATA GTAGTAGATAGATARG GTAGTAGATAGATARY GACAGACAGACAGACA GACAGACAGACAGACARG GACAGACAGACAGACARY GACACGACACGACACGACAC ACTGACTGACTGACTG ACTGACTGACTGACTGRG ACTGACTGACTGACTGRY
R: A and G residues, Y: C and T residues.
T-11 T-12 T-13 T-14 T-15 T-16 T-17 T-18 T-19 T-20
IS-11 IS-12 IS-13 IS-14 IS-15 IS-16 IS-17 IS-18 IS-19 IS-20
AATGCCCCAG TGGCCCTCAC GGTIGTACCC CCCGCTACAC CTCCTGCCAA GAGCGTCGAA CCCAGCTGTG CCTAGTCGAG CACAGGCGGA CACAGGCGGA
GATAGATAGATA GTAGTAGATAGATARG YRGATAGATAGATA GACAGACAGACAGACA GACAGACAGACARG YRGACAGACAGACA GACACGACAC ACTGACTGACTG ACTGACTGACTG YRACTGACTGACTG
ISSR amplification was performed according to Zietkiewiez et al. (1994) in a total vol u me of 30 !JI containing 100 ng DNA template, 3 ¡JI of 1 O X polymerase chain reaction (PCR) buffer, 2 mM MgCI2 , 200 IJM of each dNTP, 1 O !JM primer and 2 U Taq DNA polymerase (lnvitrogen™). DNA amplification was performed using the following cycling program: one time at 94 °C for 4 min; 37 cycles at 94 °C for 40 s, 35, 37, 40 or 45 °C (according to primer) for 40 s, 72 °C for 50 s; and a final extension at 72 °C for 7 min. ISSR fragments were separated by electrophoresis on 6% polyacrylamide gels in 1X TAE and the band profiles were detected by silver nitrate staining. Primers generating multiple discrete bands that were completely identical between the two duplicates were selected as suitable for the present study (Table 3.2).
49

Table 3.2. List of RAPD and ISSR primer used for detecting somaclonal variation in Habanero pepper (C. chinense) .
Primer Sequence Tm*
No. of bands Range
*%P (5'-3') (OC) (pb)
RAPD
T-01 CATTCGAGCC 32 9 1000-2000 44.44 T-03 CCAGCTTAGG 32 8 300-2072 87.5 T-04 CCGCCCAAAC 34 13 300-1300 30.76 T-06 CACCTTTCCC 32 5 380-800 80 T-09 CCCTACCGAC 34 6 300-1200 50 T-10 GTGCAACGTG 32 10 150-680 50 T-19 CACAGGCGGA 34 11 200-1400 90.9
ISSR
IS-01 GTAGTAGATAGATA 40 22 180-2500 9 IS-02 GTAGTAGATAGATARG 45 14 140-3000 28 IS-03 GTA GTAGATAGATARY 45 17 200-2900 35.29 IS-11 GATAGATAGATA 35 27 160-2900 40.74 IS-12 GTAGTAGATAGATARG 37 16 120-2600 31.25 IS-13 YRGATAGATAGATA 37 19 160-2500 55.55 IS-14 GACAGACAGACAGACA 40 18 280-2800 16.66 IS-15 GACAGACAGACARG 45 12 150-2200 33.33 IS-16 YRGACAGACAGACA 40 5 220-800 o IS-17 GACACGACAC 35 25 120-1900 12 IS-18 ACTGACTGACTG 40 18 180-2000 27.77 IS-19 ACTGACTGACTG 45 11 250-2900 9 IS-20 YRACTGCTGACTG 40 15 220-2700 60 *Tm: Annealing temperature, %P: Percentage of polymorphism, R: A and G residues, Y: C and T residues.
Data analysis. RAPD and ISSR reproducible fragments were scored as present (1) or absent (0), and bands were entered in a computer file as a binary matrix. In the data analysis, we analyzed two binary matrices for each of the molecular markers. The matrices were then analyzed by MVSP 3.0 (MultiVariate Statistical Package). Similarity for qualitative data was computed using Jaccard"s coefficient method (Jaccard, 1908). The similarity matrixes were subjected to cluster analysis by the unweighted pair-group method with arithmetic averages (UPGMA) and the resulting clusters were expressed as dendrograms. We supported the resulting nades using 100 bootstrap replicates analyzed by FREETREE 7.0 program.
50

3.1.4 Results and Discussion
The results obtained during induction and development of in vitro morphogenesis in Habanero pepper, through direct organogenesis (Bello-Bello et al. , 201 0) , direct somatic embryogenesis (López-Puc et al., 2006) and indirect somatic embryogenesis (Zapata-Castillo et al., 2007), corroborated the reproducibility and efficiency of the protocols previously established for this specie (Fig . 3.1 a-d) .
Figure 3.1. Morphogenesis induction in Habanero pepper: a) Direct organogenesis, b) and e) Direct and indirect somatic embryogenesis and d) Embryogenic cal/us.
In arder to increase the confidence level of the fragments included in the matrices (for RAPO and SSR), we scored only those that were very conservative, and excluded weak bands or bands that were ambiguous for some genotypes. In RAPO analysis, 20 primers were used to amplify all the regeneration systems.
51

Seven of them showed reproducible and well-resolved bands, and these were analysed. These primers produced fragments ranging from 150 to 2072 bp in size. Of 62 fragments scored from these primers, 36 were polymorphic, including the different regeneration via examinad. The highest polymorphic bands were obtained by primer T-19 (%P=90.9) and the lowest by primer T-04 (%P=30.76}. RAPOs revealed a total of 42 amplified fragments, from ONA of the control treatment.
Amplification of the ISSR markers was also performed using 20 primers. However, only 13 primers showed reproducible and well-resolved bands. These primers produced fragments ranging from 120 to 3000 bp in size. All these fragments were judged to be reproducible and subsequently scored. The analysed primers produced 219 reproducible fragments for the different regeneration via evaluated, 64 of which were polymorphic. Only one primer (IS-16) revealed monomorphic fragments . The highest number of polymorphic and scorable bands was obtained by primer IS-20 (%P=60) and the lowest by primers IS-01 (%P=9), IS-19 (%P=9) y IS-18 (%P=12). ISSRs revealed a total of 197 amplified fragments from the donor plant ONA. A summary of the effectiveness of RAPO and ISSR markers is given in Table 3.3.
Table 3.3. Effectiveness of RAPO e ISSR markers in detecting ~ol;tmor~h ism of Habanero ~e~~er somaclonal variation.
RAPO ISSR
Number of primers screened 20 20
Number of primers used 7 13
Total bands scored 62 219
Polymorphic fragments scored 36 64
Average bands per primer 8.8 16.8
Range (pd) 150-2072 120-3000
Percentage of polymorphism 57.6 27.5
The results of the cluster analyses based on the RAPO and ISSR fragments (Fig. 2a and b, respectively) are representad in two dendrograms constructed from matrices in which both monomorphic and polymorphic bands were included. Contrasting results can be observad between both molecular markers. With the cluster analysis using Jaccard's similarity index, based on RAPO (Fig. 3.2a), it was possible to classify the regeneration processes evaluated into four groups: Group /, which included only the donor plant or control (TEST); Group 11, comprising only
52

the Direct Somatic Embryogenesis process (DSE); Group 111, with lndirect Somatic Embryogenesis (ISE) and Embryogenic Callus (CALL), and Group IV, including only Direct Organogenesis (ORG). From these results we can infer that direct organogenesis is the process in which we can expect a greater difference or genetic distance (0.54) between regenerated plants and the donar plant. This proved to be the least reliable regeneration system for clonal propagation, but the most recommendable for obtaining "variant" or "somaclonal" Habanero pepper plants. In contrast, direct somatic embryogenesis was found to be the process in which regenerated plants showed greater genetic similarity (0.73) with the donar plant. Of all the systems evaluated, this regeneration system would be the most adequate for a genetically reliable propagation of Habanero pepper. lndirect somatic embryogenesis and Embryogenic callus, however, were greater contributors to genetic variation (0.67 and 0.59, respectively) in regenerated plants. Thus, as with organogenesis, these systems would be recommendable in genetic improvement of the species.
UPGMA b UPGMA
ORG IV CALL a
IV
CALL ISE 111
ISE OSE 111 H r-
11 OSE 11 ORG '-----
1 TEST TEST 0.52 0.8 0.88 0.18 0.8. 0.92 0.16 0.8 0.8. 0.88 0.92 0.96
Jaccard's Coemclent Jaccard's Coemcient
Figure 3.2. Dendrograms based on the Jaccard's coefficient in Habanero pepper regeneration protocols by UPGMA cluster analysis. a) RAPO and b) ISSR markers in Direct Organogenesis (ORG), Direct Somatic Embryogenesis (DSE), lndirect Somatic Embryogenesis (/SE), Embryogenic Cal/us (CALL) and donar plants (TEST).
With ISSR, the regeneration systems were distributed in the following groups (Fig. 3.2b): Group 1, which included only the donar plant; Group 11, with Organogenesis (ORG) which, contrary to expectations, showed the greatest genetic similarity (0.87) to the donar plant (TEST); Group 11/, comprising Direct Somatic
53

Embryogenesis (DSE) and lndirect Somatic Embryogenesis {ISE); and Group IV, including only Embryogenic Callus (CALL), which presented the lowest genetic similarity {0. 72) to the donar plant. An analysis of the RAPD results obtained from callus, befare and after embryo formation, revealed 48 bands present in common and 6 bands absent in common in both, with 54 loci being shared between both processes (unreported data). Part of the variation observed in the ESI could be associated with the presence of 2,4-D, which was used in both processes. Figure 3.3a and b shows polymorphism through RAPD obtained using primers T-04 and T-19 to estimate somaclonal variation in each regeneration via evaluated for Habanero pepper. These results coincide with those reported by Novak (1991) regarding the relative tendency generally observed in the genetic variation generated in different in vitro regeneration systems. The somaclonal variation detected with the use of RAPD technique has also been reported in rice Oriza sativa L. (Banerjee et al., 1997), Phalaenopsis {Chen et al., 1998), Actinidia deliciosa A. (Palombi & Damiano, 2002), Capsicum annum L. (Hossain et al., 2003), Pyrus Pyraster B. (Palombi et al., 2007). Similarly, Hashmi et al. (1997) were able to find genetic variation in somatic embryos of peach (Prunus persica). In contrast, Isabel et al. (1993), using the same technique, found no evidence of variation during somatic embryogenesis in Picea mariana (Mili). With banana, Gimenez et al. {2001 ), using 6 primers, found that 86% of the bands were polymorphic and 14 were monomorphic, corroborating the genetic instability of the genus Musa.
With ISSR markers, direct organogenesis showed the higher genetic similarity {0.87), in comparison with the other regeneration systems evaluated. These results differ from reports by Novak (1991) on somaclonal variation of in vitro plants. Direct and indirect embryogenesis coincided in the level of variation {0.83), sharing 188 bands in common and 21 bands absent; however, they did differ in 1 O polymorphic bands. Analyses of Callus and ISE revealed 169 bands shared and 21 bands absent; a total of 29 polymorphic bands were detected in both processes, from which we can infer that part of the variation found in ISE may have originated in the callus phase. The presence of 2,4-D during the induction stage in both processes may have had some influence on the variation detected. Figure 3.3b and e show polymorphic primers {IS-11 and IS-18, respectively) evaluated in the different regeneration systems, using ISSR.
54

bp M 1 2 3 4 S N bp M 1 2 3 4 5 N
1500
1300 1100
1000 1000
900 900
800 800 700
700 800
600 500
500 400
400
300 300
200 200
100 100
bp M 1 2 3 4 5 N bp M 2 3 4 5 N
3000 2000
2500 1650
2000
1650
1000 850
1000
650 850
500
650 500
400
400
300
300
200
200
100 100
55

Figure 3.3. Electrophoresis pattern of randomly amplified polymorphic ONA (RAPO) and ínter-simple sequence repeats (ISSR) markers obtained from regeneration protocols of Habanero pepper. a) and b) RAPOs using primer T-04 and T-19, e) and d) ISSR using primer IS-11 and IS-18, in mother plant ~ane 1), direct organogenesis ~ane 2), direct somatic embryogenesis ~ane 3), indirect somatic embryogenesis ~ane 4) and cal/us system ~ane 5). N: negative control, M: molecular mass marker 1 kb plus ONA ladder.
Numerous reports refer to the presence of somaclonal variants in regenerated plants of many species, using different in vitro culture systems. Rus-Kortekaas et al. (1994}, working with Lycopersicon esculentum, observed physiological mutations in regenerated plants while studying the descendants obtained from callus. Albani and Wilkinson (1998} also reported genetic variation in shoots obtained from induced callus of Solanum tuberosum. In Capsicum annum L. Hossain et al. (2003), using RAPO, reported somaclonal variants in plants regenerated via direct organogenesis. Variations were also found in sorne morphological characters, such as growth habits, stem color, antocianine content and flower and fruit color. Similarly, Anu et al. (2004), working with the same species (C. annum) but using only morphological markers, observed variations in plant growth habit, flower position, fruit shape and color. Alhani and Wilkinson (1998} working with ISSR, detected somaclonal variation in induced callus of potato; while Leroy et al. (2000) obtained similar results in cauliflower callus. More recently, Thomas et al. (2006) reported up to 55% somaclonal variation in regenerated tea plants Camellia sinensis (L.), using direct somatic embryogenesis. The use of different molecular markers for evaluating the variation induced during in vitro culture has also been reported. More precisely, RAPOs and ISSR have been used in Camellia ssp. (Oevarumath et al., 2002), Prunus dulcis Mili (Martins et al. , 2004}, banano (Lakshmanan et al., 2007), where somaclonal variation has been detected in all regenerants. However, Martins et al. (2004) in Prunus dulcis, using the same markers (RAPOs and ISSR) to evaluate plants obtained from axillary bud proliferation, did not detect polymorphic bands, corroborating the genetic fidelity of this system for clonal micropropagation. Similar results have been reported by Lakshmanan et al. (2007) who also detected a high genetic homogeneity in banana plants obtained through in vitro culture of axillary buds.
Two hypotheses were formulated to explain these results; both of them are in agreement with the results obtained using these two types of molecular markers. We conclude that when the tissue culture technique is used, the analysis of somaclonal variability could require more than one ONA-based technique; in fact, the genetic variation present in different sources could interfere or combine with the more or less polymorphic ability, as our results show for ISSR and RAPO markers. This response could be associated with the type of molecular marker
56

used. Micropropagation, conservation or genetic transformation requires a high degree of genetic fidelity. However, somaclonal variation generates an unexpectedly rich source of genetic diversity in plants and has been used to introduce novel traits into a wide range of crops. In many cases it has been shown to be a potential tool for plant breeding when utilized to improve crop cultivars for agronomically useful characteristics, such as resistance to disease and environmental stress factors.
3.1.5 References
Abe, K., T. Otomo and S. Saito (1999). The use of somaclonal variation in rice breeding. l. Genetic variation in rice regenerant pedigree lines derived from a single callus. Report of the Kyushu Branch of the Crop Science Society of Japan, 56, 59-63.
Alhani, M.C. and M.J. Wilkinson (1998). lnter simple sequence repeat polymerase chain reaction for the detection of somaclonal variation. Plant Breed, 117, 573-575.
Anu, A. , K. Babu and K. Peter (2004). Variations among somaclones and its seedling progeny in Capsicum annuum. Plant Cell, Tissue and Organ Culture, 76, 261-267.
Austin, S. and A.C. Cassells (1983). Variation between plants regenerated from individual calli produced from separated patato stem cal/us cel/s. Plant Sci Lett, 31, 107-114.
Banerjee, H., V. Chimote and S. K. Raina (1997). DNA polymorphism among rice somaclones. Biología Plantarum, 40, 543-533.
Beckert, M., M. Pollacsek and M. Caenen (1983). Study of genetic variability in maize (Zea mays L.) after in vitro regeneration of plants from cal/us culture. Agronomie, 3, 9-18.
Bello-Bello, J. J., A. Canto-Fiick, E. Balam-Uc, E. Gómez-Uc, M. L. Robert, L. G. lglesias-Andreu and N. Santana-Buzzy (201 0). lmprovement of in vitro proliferation and elongation of Habanero pepper shoots (Capsicum chinense Jacq.) by Temporary lmmersion. Hortscience, 45(7), 1-6.
Chen, W.H., T.M. Chen, Y.M. Fu, R.M. Hsieh and W.S. Chen (1998). Studies on somaclonal variation in Phalaenopsis. Plant Cell Rep, 18, 7-13.
Dellaporta, S. L., J. Wood and J.B. Hicks (1983). A plant DNA minipreparation: Version 1/. Plant Molecular Biology Reporter, 1, 19-22.
Devarumath, R., S. Nandy, V. Rani, S. Marimuthu, N. Muraleedharan and S. Raina (2002). RAPO, ISSR and RFLP fingerprints as useful markers to evaluate genetic integrity of micropropagated plants of three diploid and triploid elite tea clones representing Camellia sinensis (China type) and C. assamica ssp. assamica (Assam-lndia type). Plant Cell Rep., 21, 166-173, 2002.
Evans, D.A. and W.R. Sharp (1983). Single gene mutations in tomato plants regenerated from tissue culture. Science, 221 , 949-951.
57

Evans, D.A. (1989). Somaclonal variation-genetic basis and breeding applications. Trends in Genetics, 5, 346-50.
Giménez, C. and E. García (2001 ). Somac/onal variation in Banana: Cytogenetic and molecular Characterization of the somaclonal variant CIEN BTA-03. In Vitro Cell . Dev. Bioi.-Piant, 37, 217-222.
Hashmi, G., R. Huettel, R. Meyer, L. Krusberg and F. Hammerschlag (1997) . RAPO analysis of somaclonal variants derived from embryo callus cultures of peach. Plant Cell Rep. , 16, 624-627.
Hasim, Z.N. , W.F. Cambell and J.G. Carman (1990). Morpho/ogical analysis of spring wheat (C/MMYT cultivar PC YT-10) somaclones. Plant Cell Tiss Organ Cult. , 20, 95-100.
Hossain, A., K. Konisho, M. Minami and K. Nemoto. (2003). Somaclonal variation of regenerated plants in chili pepper (Capsicum annuum L.). Euphytica, 130, 233-239.
Isabel, N., L. Tremblay, M. Michaud, F.M. Tremblay and J. Bousquet (1993). RAPOs as an aid to eva/uate the genetic integrity of somatic embryogenesisderived populations of Picea mariana (Mil/.) B.S.P. Theor. Appl. Genet., 86, 81-87.
Jaccard, P. (1908) Nouvel/es recherches sur la distribution flora/e . Bulletin Societe Vaudoise des Sciences Naturelles, 44, 223-270
Kado, C.l. and A. Kleinhofs (1980). Genetic modification of plant cells through uptake offoreign ONA. lntern. Rev. Cytol. Suppl., 11 , 47-80
Lakshmanan, V., S. Venkataramareddy and B. Neelwarne (2007). Molecular analysis of genetic stability in long-term micropropagated shoots of banana using RAPO and /SSR markers. Electronic Journal of Biotechnology, 10(1), 1-8.
Larkin, P.J . and W.R. Scowcroft (1983). Somaclonal variation and eyespot toxin tolerance in sugarcane. Plant Cell Tiss Organ Cult. , 2, 111-121 .
Larkin, P.J. and S.C. Scowcroft (1981). Somaclonal variation: a novel source of variability from ce// culture for plant improvement. Theor Appl Genet, 60, 197-214.
Leroy, X., K. Leon and M. Branchard (2000). Plant genomic instability detected by microsatelfite-primers. Electronic Journal of Biotechnology, 3(2), 1-9.
López-Puc, G., A. Canto-Fiick, F. Barredo-Pool, P. Zapata-Castillo, M. Montalvo-Peniche, F. Barahona-Pérez and N. Santana-Buzzy (2006). Oirect Somatic Embryogenesis: A Highly Efficient Protocol for In vitro Regeneration of Habanero Pepper (Capsicum chinense Jacq.) HortScience, 41 (6), 1-7.
Maralappanavar, M.S., M.S. Kuruvinashetti and C.C. Harti (2000). Regeneration, establishment and eva/uation of somaclones in Sorghum bicolor (L.) Moench. Euphytica, 115, 173-180.
Martins, M., D. Sarmento and M. Oliveira (2004) .Genetic stability of micropropagated a/mond plantlets, as assessed by RAPO and /SSR markers. Plant Cell Rep., 23,492-496
Novak, F.J. (1991 ). In vitro mutation system for crop improvement. In: Plant Mutation Breeding for Crop lmprovement, IAEA, Vienna. Vol 2, pp. 327-342.
58

Palombi, M., B. Lombardo and E. Caboni (2007). In vitro regeneration of wild pear (Pyrus pyraster Burgsd) clones tolerant to Fe-chlorosis and somaclonal variation analysis by RAPO markers. Plant Cell Rep., 26, 489-496.
Palombi, M. and V. Damiano (2002). Comparison between RAPO and SSR molecular markers in detecting genetic variation in kiwifruit (Actinidia deliciosa A. Chev). Plant Cell Rep., 20, 1061-1066.
Rus-Kortekaas, W., M.J . Smulders, P. Arens and B. Vosman (1994). Direct comparison of levels of genetic variation in tomato detected by GACA-containing microsatellite probe and random amplified polymorphic DNA. Genome, 37, 375-381.
Scowcroft, W.R. (1977) Somatic ce// genetics and plant improvement. Adv. Agron ., 29, 39-81
Sibi, M. (1982). Heritable epigenetic variations from in vitro tissue culture of Lycopersicon esculentum (var. Monalbo). In: E.D. Earle & Y. Demarly (Eds.), Variability in Plants Regenerated from Tissue Culture, Praeger Press, New York. pp. 228-244.
Thomas, E., P.J. King and l. Potrykus (1979). lmprovement of crop plants vía single ce lis in vitro - an assessment. Zeitschrtfljur Pflanzenzuchtigen. 82, 1-30
Thomas, J., D. Vijayan, S.D. Joshi, S.J. Lopez, and R.R. Kumar (2006). Genetic integrity of somaclonal variants in tea (Camellia sinensis (L.) O Kuntze) as revealed by ínter simple sequence repeats. Journal of Biotechnology, 123: 149-154
Williams, J.G., A.R. Kuubelik, K.J . Livak, J.A. Rafalski and S.V. Tingey. (1990). ONA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res ., 18, 6531-6535.
Zapata-Castillo, Y. P., A. Canto-Fiick, G. López-Puc, A. Solís-Ruiz. , F. Barahona-Pérez and N. Santana-Buzzy. (2007). Somatic Embiogenesis in Habanero Pepper (C. chinense Jacq.) From Ce// Suspensions. HortScience, 42(2), 1-5.
Zietkiewicz, E., A Rafalski and D. Labuda. (1994). Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics, 20, 176-183.
59
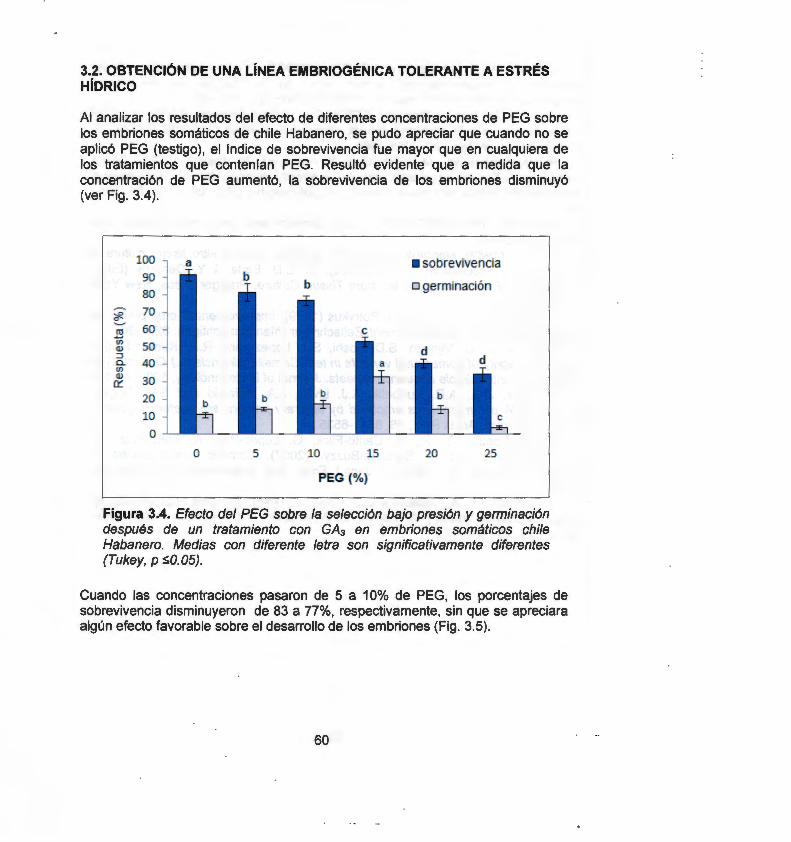
3.2. OBTENCIÓN DE UNA LÍNEA EMBRIOGÉNICA TOLERANTE A ESTRÉS HÍDRICO
Al analizar los resultados del efecto de diferentes concentraciones de PEG sobre los embriones somáticos de chile Habanero, se pudo apreciar que cuando no se aplicó PEG (testigo) , el índice de sobrevivencia fue mayor que en cualquiera de los tratamientos que contenían PEG. Resultó evidente que a medida que la concentración de PEG aumentó, la sobrevivencia de los embriones disminuyó (ver Fig. 3.4 ).
100 a 90
80 ~ 70 '# ~
60 2 Ul 50 IV :J ·C.. 40 VJ IV 30 0:::
20
10
o o 5 10 15
PEG (%)
• sobrevivencia
o germinación
20 25
Figura 3.4. Efecto del PEG sobre la selección bajo presión y germinación después de un tratamiento con GA3 en embriones somáticos chile Habanero. Medias con diferente letra son significativamente diferentes (Tukey, p :50. 05).
Cuando las concentraciones pasaron de 5 a 1 0% de PEG, los porcentajes de sobrevivencia disminuyeron de 83 a 77%, respectivamente, sin que se apreciara algún efecto favorable sobre el desarrollo de los embriones (Fig. 3.5).
60

Figura 3.5. Embriones somáticos bajo presión de selección con PEG. Apariencia de /os explantes y embriones en diferentes estadios de desarrollo con diferentes concentraciones de PEG (O, 5, 1 O 15, 20 y 25%), a /os 15 días de cultivo.
61

Cuando los embriones fueron expuestos a 15% de PEG, se observó una drástica disminución en el porcentaje de sobrevivencia (53%). Sin embargo, se vió favorecido el desarrollo de los embriones, logrando su germinación después de un tratamiento con GA3 , como se puede observar en la Figura 3.6. Los índices más bajos de sobrevivencia se observaron cuando los embriones fueron expuestos a concentraciones de 20 y 25% de PEG, en estos tratamientos se inhibió tanto el desarrollo como la germinación de los embriones, con una sobrevivencia fue menor al 50%. Probablemente esto pudiera estar relacionado con el elevado potencial osmótico generado en ambos medios de cultivo (-1 .33 y-1.51 MPa., respectivamente) , lo que pudiera estar limitando la toma de agua y nutrientes por parte de los embriones.
Figura 3.6 Germinación in vitro de embriones somáticos después de un tratamiento con GA3 y previo tratamiento con 15% de PEG.
El efecto positivo del PEG sobre el desarrollo de los embriones somáticos se ha reportado también en pino (Ramarosandratana et al., 2001 ; Maruyama et al., 2007) y palma datilera (Aikhateeb, 2006). Al aumentar la osmolaridad de los medios de cultivo, esta provoca un efecto de estrés osmótico, similar a lo que ocurre durante la desecación de los embriones, este efecto tiene una implicación directa sobre la maduración y germinación en los embriones somáticos (Attree and Fowke, 1993; Alkhateeb, 2006).
La selección bajo presión in vitro ha sido empleada para la obtención de líneas tolerantes a estrés hídrico en numerosas especies. En tomate (Handa et al., 1983) reportaron una línea celular tolerante a estrés hídrico mediante la selección bajo presión con 20 ó 25% de PEG (PM-6000). Santos-Díaz & Ochoa-Aiejo (1994) trabajando con células en suspensión de chile (C. annum) aplicaron una presión de selección con 15% de PEG (PM-8000), en una segunda etapa las líneas aparentemente tolerantes fueron tratadas nuevamente con 20-25% de PEG. Sin
62

embargo, mediante estos protocolos sólo es posible trabajar con células en suspensión, lo cual limita la obtención de posibles genotipos tolerantes a estrés hídrico. Por otro lado, en sorgo (Duncan et al., 1995) obtuvieron una línea embriogénica tolerante a estrés hídrico mediante la aplicación de 9 a 12% de PEG (PM- 8000). En arroz (Biswas et al., 2002) obtuvieron líneas embriogénicas tolerantes a estrés hídrico mediante la aplicación de 5 a 10% de PEG (PM-6000). En banano (Ebrahim et al., 2006) con 1.5% de PEG (PM-6000) lograron obtener plantas tolerantes a estrés hídrico. En el género Capsicum, este es el primer reporte de obtención de una línea embriogénica tolerante a estrés hídrico (línea: NSBm-04-PEG), obtenida mediante la aplicación de 15% de PEG, el cual se registró un potencial hídrico de -0.92 Mpa, pasando de un estrés suave a moderado según los grados de sequía propuestos por Hsiao (1973), a este potencial hídrico aproximadamente el 50% de una población embriones somático muere, lo que constituye la dosis letal media (DL50). Sin embargo, a este nivel de estrés hídrico observamos mejor desarrollo y germinación por parte de los embriones somáticos.
3.2.1 . Análisis molecular RAPO de las líneas embriogénicas
El análisis molecular realizado en las líneas embriogénicas a partir 7 cebadores RAPOs reveló un total de 36 /oci entre los diferentes tratamientos con PEG. En la línea embriogénica usada como testigo (0% de PEG), se obtuvieron 34 bandas presentes. Por otro lado, sólo fue posible obtener bandas polimórficas en los cebadores T-03, T-04, T-10 y T-19. Con el cebador T-03 se observo la ausencia de 1 banda de 1500 pb en los tratamientos con mayor estrés (20 y 25% de PEG), este mismo fenómeno sucedió con en el cebador T-19 al perder una banda de 2600 pb en los tratamientos con 10, 15 20 y 25% de PEG. Con el cebador T-04 se generó un nuevo fragmento de 1200 pb en los tratamientos de 1 O, 15, 20 y 25% de PEG (Fig. 3.7a). De manera similar, en el cebador T-10 se encontró una nueva banda de 1800 pb en todas las líneas embriogénicas tratadas con PEG (Fig. 3.7b). El polimorfismo obtenido por la ausencia o presencia de nuevos fragmentos en los tratamientos bajo presión de selección con PEG, posiblemente podría estar ligado a genes involucrados a la tolerancia al estrés hídrico. Carrera et al. , (2004) mencionan que si un marcador resulta estar genéticamente ligado a un gen que controla un carácter de interés agronómico, la selección de este marcador resulta en la selección indirecta del gen de interés. Aplicado al mejoramiento genético vegetal, este hecho constituye el fundamento del proceso de selección de genotipos de interés agronómico por medio de marcadores moleculares.
63
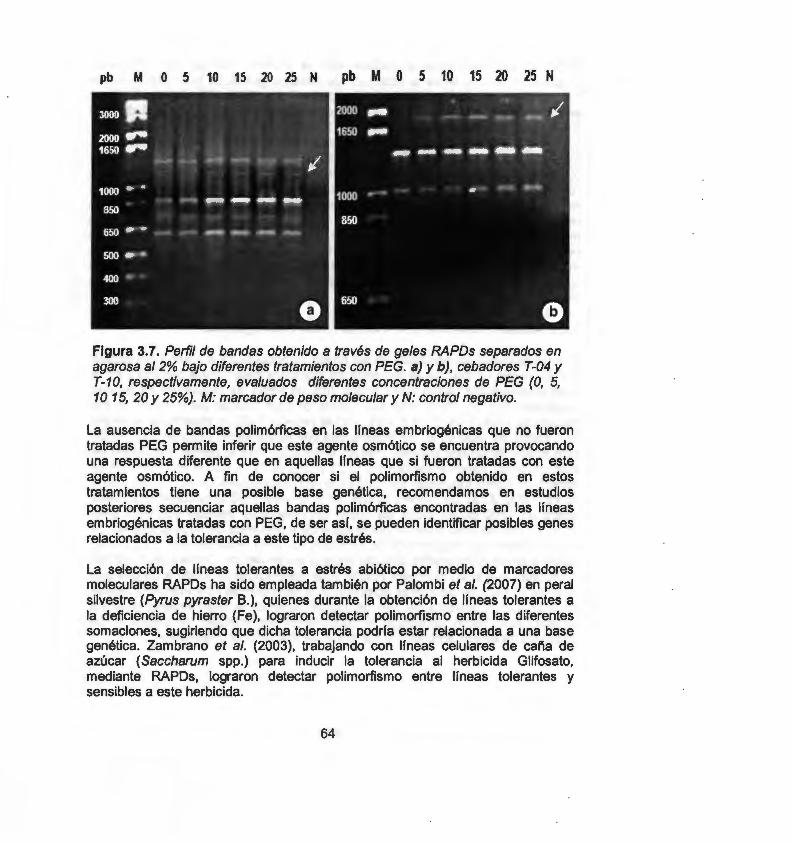
pb M O 5 1'0 15 20 25 N ,pb M O 5 10 1i5 20 25 N
Figura 3.7. Perfil de bandas obtenido a través de geles RAPOs separados en agarosa al 2% bajo diferentes tratamientos con PEG. a) y b), cebadores T-04 y T-10, respectivamente, evaluados diferentes concentraciones de PEG (0, 5, 1 O 15, 20 y 25%). M: marcador de peso molecular y N: control negativo.
La ausencia de bandas polimórficas en las líneas embriogénicas que no fueron tratadas PEG permite inferir que este agente osmótico se encuentra provocando una respuesta diferente que en aquellas líneas que si fueron tratadas con este agente osmótico. A fin de conocer si el polimorfismo obtenido en estos tratamientos tiene una posible base genética, recomendamos en estudios posteriores secuenciar aquellas bandas polimórficas encontradas en las líneas embriogénicas tratadas con PEG, de ser así, se pueden identificar posibles genes relacionados a la tolerancia a este tipo de estrés.
La selección de líneas tolerantes a estrés abiótico por medio de marcadores moleculares RAPOs ha sido empleada también por Palombi et al. (2007) en peral silvestre (Pyrus pyraster B.), quienes durante la obtención de líneas tolerantes a la deficiencia de hierro (Fe), lograron detectar polimorfismo entre las diferentes somaclones, sugiriendo que dicha tolerancia podría estar relacionada a una base genética. Zambrano et al. (2003), trabajando con líneas celulares de caña de azúcar (Saccharum spp.) para inducir la tolerancia al herbicida Glifosato, mediante RAPOs, lograron detectar polimorfismo entre líneas tolerantes y sensibles a este herbicida.
64
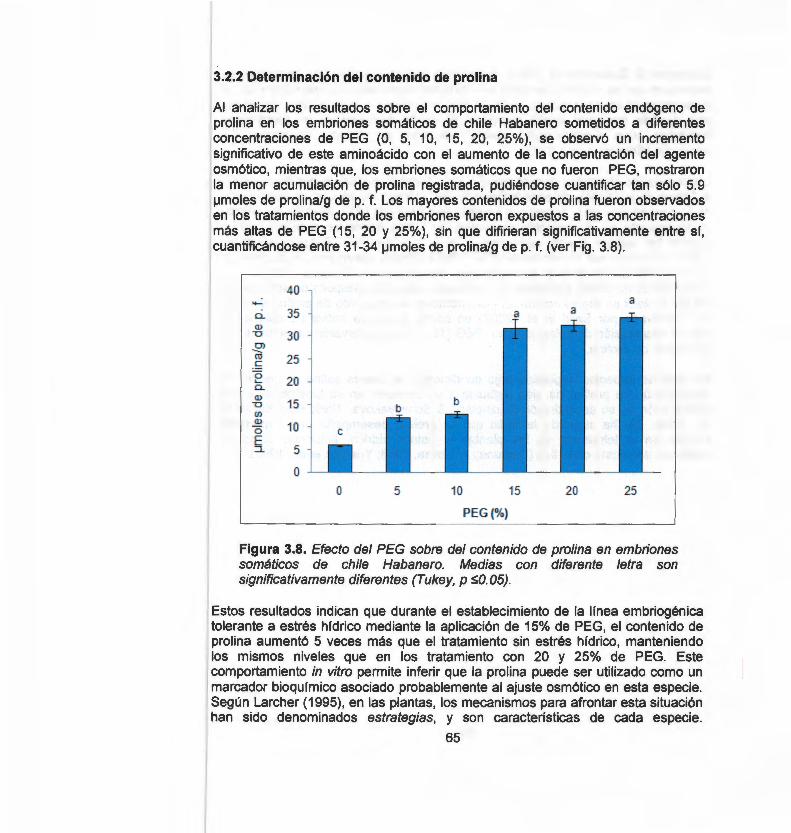
3.2.2 Determinación del contenido de prolina
Al analizar los resultados sobre el comportamiento del contenido endógeno de prolina en los embriones somáticos de chile Habanero sometidos a diferentes concentraciones de PEG (0, 5, 1 O, 15, 20, 25%), se observó un incremento significativo de este aminoácido con el aumento de la concentración del agente osmótico, mientras que, los embriones somáticos que no fueron PEG, mostraron la menor acumulación de prolina registrada, pudiéndose cuantificar tan sólo 5.9 ¡Jmoles de prolina/g de p. f. Los mayores contenidos de prolina fueron observados en los tratamientos donde los embriones fueron expuestos a las concentraciones más altas de PEG (15, 20 y 25%), sin que difirieran significativamente entre sí, cuantificándose entre 31-34 ¡Jmoles de prolina/g de p. f. (ver Fig. 3.8).
40 ,._ a o_ 35 Q)
-o 30 O) -ro 25
-~ o 20 ..... o_ Q)
15 -o (/)
..!l:! ~o o E :l. 5
o o 5 10 1!5 20 25
PEG {%)
Figura 3.8. Efecto del PEG sobre del contenido de prefina en embriones somáticos de chile Habanero. Medias con diferente letra son significativamente diferentes (Tukey, p sO. OS).
Estos resultados indican que durante el establecimiento de la línea embriogénica tolerante a estrés hídrico mediante la aplicación de 15% de PEG, el contenido de prolina aumentó 5 veces más que el tratamiento sin estrés hídrico, manteniendo los mismos niveles que en los tratamiento con 20 y 25% de PEG. Este comportamiento in vitro permite inferir que la prolina puede ser utilizado como un marcador bioquímico asociado probablemente al ajuste osmótico en esta especie. Según Larcher (1995), en las plantas, los mecanismos para afrontar esta situación han sido denominados estrategias, y son características de cada especie.
65

Kuznetsov & Schevyakova (1999) mencionan que la acumulación de prolina se incrementa en las plantas afectadas por condiciones de sequía o salinidad y que el incremento de este compuesto confiere cierta tolerancia a ambos tipos de estrés, esto sucede mediante ajuste osmótico. El ajuste osmótico es un mecanismo fisiológico cuya determinación puede realizarse en etapas tempranas del desarrollo (Stewart & Lee, 197 4 ).
Los resultados obtenidos en este estudio, corroboran lo planteado por Santos-Díaz & Ochoa-Aiejo (1994), quienes trabajando con cultivos de células en suspensión de Capsicum annum L. sometidos a estrés osmótico con PEG, también reportaron un incremento en el contenido de prolina en líneas tolerantes a estrés hídrico cuando los cultivos fueron expuestos a 20 y 25% de PEG. Estudios similares fueron realizados por Unnikrishnan et al. (1991) quienes observaron un incremento en el contenido de prolina en embriones somáticos de Sapindus trifoliatus L. , provenientes de líneas tolerantes a la salinidad, obtenidas después de someter las células a NaCI en medio líquido. El incremento en el contenido de prolina también fue observado por Sajid et al. (2007) en callos de Oryza sativa L., quienes al ejercer una presión de selección con PEG (10 y 18%) observaron un aumento en la síntesis de prolina.
En muchas especies vegetales bajo condiciones de estrés salino y sequía, la acumulación de prolina ha sido atribuida a un aumento en su biosíntesis y una disminución en su degradación (Kuznetsov & Schevyakova, 1999; Kavi Kishor et al. , 2005). Se ha sugerido también que la prolina desempeña una importante función en la tolerancia de las plantas al estrés hídrico, actuando como un mediador del ajuste osmótico (Delauney & Verma, 1993; Yoshiba et al., 1997).
66

3.3 PROTOCOLO PARA LA OBTENCIÓN DE LA LÍNEA EMBRIOGÉNICA NSBM-04-PEG, TOLERANTE A ESTRÉS HÍDRICO
Línea: NSBm-04-PEG
e --+
Figura 3.9. Etapas en el desarrollo del protocolo para la obtención de una línea embriogénica tolerante a estrés hídrico: A) asepsia de semillas; B) germinación in vitro de semillas (MS+1 . 15 pM de AG3) ; C) selección de explantes (hipocótilos de 15 días de edad); D) inducción de la embriogénesis a partir de hipocótilos en medio semisólido (MS+9.05 pM de 2,4-0 + 2 gL1 de Gelrite™) por 30 días en luz continua; E) embriones somáticos (estadio globular y corazón) sometidos a 15% de PEG, en medio líquido en agitación (100 rpm en luz continua); F) subcultivo a los 14 días, ídem condiciones al medio anterior (desarrollo de los embriones somáticos a estadías torpedo y cotiledonar); G) línea embriogénica NSBm-04-PEG tolerante a estrés hídrico (germinación de los embriones de esta línea en medio líquido MS+1.15 pM de AG3) .
67

3.4 DISCUSIÓN GENERAL
En los últimos años, los estudios de vanac1on somaclonal han aumentado considerablemente para evaluar la frecuencia de variación genética generada en los sistemas de micropropagación in vitro (Giménez et al. , 2001; Devarumath et al. , 2002; Hossain et al., 2003; Martins et al., 2004; Guo et al., 2006; Lakshmanan et al., 2007) y para la generación de genotipos con nuevas características agronómicas (Mercado et al., 2000; Biswas et al. , 2002; Roy & Mandal, 2005; Venkataiah et al. , 2005; Patnaik et al., 2006).
En C. annum existen reportes donde se ha reconocido una alta frecuencia de variación genética durante el cultivo in vitro (Hossain et al., 2003; Anu et al., 2004). En C. chinense, éste constituye el primer reporte donde se evalúa la variación in vitro generada por diferentes vías de regeneración empleando dos marcadores moleculares. En este estudio, utilizando RAPOs e ISSRs se pudo detectar bandas polimórficas en los perfiles de ADN, arrojando mediante el análisis de similitud genética, alto nivel de variación en todos los sistemas de regeneración que fueron analizados. Según Evans {1989), esta variación somaclonal puede convertirse en un recurso importante para programas de mejoramiento genético de plantas de interés. En este sentido, aprovechando que la variación genética generada fue alta en todos los sistemas de regeneración de chile, la elección del sistema para la selección bajo presión fue en base a la eficiencia de cada sistema. Por ello, la selección de líneas tolerantes a estrés hídrico se hizo utilizando el protocolo de "embriogénesis somática directa de alta eficiencia en medio líquido", reportado por Avilés-Viñas (2007), debido a que consideramos que este sistema es altamente eficiente y reproducible, además este protocolo, a diferencia de la organogénesis directa, permite trabajar con un mayor número de individuos por explante, aumentando así la probabilidad de obtener mayor índice de sobrevivencia bajo condiciones de estrés.
Como resultado, estamos reportando para C. Chinense el primer protocolo de selección bajo presión para la obtención de líneas tolerantes a estrés hídrico, utilizando PEG como agente selectivo (ver Fig. 3.9). La selección bajo presión in vitro con PEG es una técnica que ha sido también empleada para la obtención de líneas tolerantes a estrés hídrico diferentes especies (Handa et al. , 1983; SantosDíaz & Ochoa-Aiejo, 1994; Duncan et al., 1995; Biswas et al., 2002; Ebrahim et al., 2006). En Capsicum annum (Santos-Díaz & Ochoa-Aiejo, 1994), lograron seleccionar líneas celulares tolerantes a estrés hídrico al emplear PEG como agente selectivo. Sin embargo, mediante este protocolo sólo es posible trabajar con cultivos celulares, lo cual limita la obtención de posibles individuos tolerantes a este estrés.
68

El polimorfismo encontrado en aquellas líneas tratadas con PEG permite inferir que este agente osmótico provocó una respuesta genética o epigenéticamente diferente respecto a aquellas líneas que no fueron estresadas con este agente osmótico. La presencia de nuevas bandas en nuestra línea seleccionada a estrés hídrico sugiere que probablemente estas bandas estén relacionadas con la tolerancia a dicho estrés. La selección de líneas tolerantes a estrés por medio de marcadores moleculares ha sido también reportada por otros autores (Zambrano et a/., 2003; Palombi et al., 2007; Rzepka-Pievnes et a/., 2007).
La respuesta en cuanto a la acumulación de prolina es compleja y en algunos casos, más que una causa de tolerancia a sequía, podría ser un sensor del grado de estrés al que está sometida la planta (Kuznetsov & Schevyakova, 1999). Para el caso de C. chinense, no descartamos la posibilidad de que este osmolito actúe como mediador en el ajuste osmótico. La tolerancia al estrés osmótico in vito mediante el aumento en el contenido de prolina ha sido reportado también en Sapindus trifolíatus L. (Unnikrishnan et a/., 1991 ), Capsicum annum L. (SantosDíaz & Ochoa-Aiejo, 1994 ), Oryza sativa L. (Lutts et al., 1996; Sajid et al., 2007). Stewart & Lee (1974) mencionan que el ajuste osmótico mediado por el aumento en la síntesis de prolina en un mecanismo fisiológico cuya determinación puede realizarse en etapas tempranas del desarrollo. En tabaco se han producido plantas transgénicas que sobreproducen prolina, mejorando así su tolerancia al estrés osmótico (Kishor, 1995).
Aunque los embriones somáticos de la línea NSBm-04-PEG germinan, la conversión en planta sigue siendo una problemática para la especie durante el cultivo in vitro.
3.5 BIBLIOGRAFÍA
Alkhateeb, A. (2006). Somatic Embryogenesis in Date Palm (Phoenix dactylífera L.) cv. Sukary in Response to Sucrose and Polyethy/ene G/yco/. Biotecnology, 5(4), 466-470.
Anu, A., K. Babu and K. Peter (2004). Variations among somaclones and its seedlíng progeny in Capsicum annuum. Plant Cell, Tissue and Organ Culture, 76, 261-267.
Attree, S.M. and L.C Fowke (1993). Embryogeny of gymnosperms: advances in synthetic seed technology of conifers. Plant Cell Tissue Organ Cult. , 35,1-35.
Avilés-Viñas, S. (2007). Papel del etileno durante la embriogénesis somática del chile Habanero (Capsicum chinense Jacq.).Tesis de maestría. CICY. Méx. pp 65.
Biswas, J., B. Chowdhury, A. Bhattacharya, and A. Manda!. (2002). In vitro screening for increased drought tolerance in rice. In Vitro Cell. Dev. Bioi.-Piant, 38,525-530.
69

Brar, S.M. and D.S. Brar (1998). Somaclonal variation: mechanism and applications in crop improvement. In: Jain, SM and D.S. Brar (eds). Variation and induced mutations in crop improvement, Dordrecht, Kluwer. p. 15-37.
Buyukalaca, S. and F. Mavituna {1996). Somatic embryogenesis and plant regeneration of papper in /iquid media. Plant Cell, Tissue and Organ Culture, 46, 227-235.
Delauney, A.J ., and O.P. Verma. (1993). Proline biosynthesis and osmoregulation in plants. Plant Journal, 4, 215-223.
Devarumath, R. , S. Nandy, V. Rani, S. Marimuthu, N. Muraleedharan and S. Raina {2002). RAPO, /SSR and RFLP fingerprints as useful markers to evaluate gene tic integrity of micropropagated plants of three diploid and triploid elite tea clones representing Camellia sinensis (China type) and C. assamica ssp. assamica (Assam-lndia type). Plant Cell Rep., 21 , 166-173, 2002.
Duncan, R. , R.M. Waskom and M.W. Nabors (1995). In vitro screening and field evaluation of tissue-cu/ture-regenerated sorghum (Sorghum bicolor (L.) Moench) for soil stress tolerance. Euphytica, 85, 373-380.
Duncan, R.R. {1997). Tissue cu/ture-induced variation and crop improvement. Adv. Agron. , 58, 201-240.
Ebrahim, M. K. , l. A. lbrahim, H. A. Emara and E. Komor. (2006). !mpact of po/yethylene glycol-induced water stress on growth and development of shoot tip cultures from different banana (Musa spp.) cultivars. Journal of Applied Horticultura, 8(1 ), 53-57.
Evans, D.A. (1989). Somac!onal variation-genetic basis and breeding applications. Trends in Genetics, 5, 346-50.
Giménez, C. and E. García. (2001 ). Somaclonal variation in Banana: Cytogenetic and molecular Characterization of the somaclonal variant CIEN BTA-03. In Vitro Cell. Dev. Bioi.-Piant, 37, 217-222.
Guo W., Y. Li, L. Gong, F. Li, Y. Dong, and B. Liu (2006). Efficient micropropagation of Robinia ambigua var. idahoensis (ldaho Locust) and detection of genomic variation by ISSR markers. Plant Cell , Tissue and Organ Culture, 84, 343-351 .
Handa, A.K., R.A. Bressan, S. Handa and P.M. Hasegawa {1983). Clona! Variation for To!erance to Po/yethylene G/yco!-lnduced Water Stress in Cultured Tomato Cells. Plant Physiol., 72, 645-653.
Hossain, A., K. Konisho, M. Minami and K. Nemoto. (2003). Somac/onal variation of regenerated plants in chili pepper (Capsicum annuum L.). Euphytica, 130, 233-239.
Kavi Kishor, P.B., S. Sangam, R.N. Amrutha, P. Sri Laxmi, K.R. Naidu, K.R., Rao, Sreenath Rao, K.J. Reddy, P. Theriappan and N. Screenivasulu (2005). Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: its implications in plant growth and abiotic stress tolerance. Current Science, 88{3), 424-438
70

Kishor, K., Z. Hong, G. Miao, C. Hu and D. Verma (1995). Overexpression of Pyrrolíne 5-Carboxy/ate Syntetase increases proline production and confer osmotolerance in transgenic plants. Plant Physiol. 108: 1387-1394.
Kuznetsov, V. and N., Schevyakova (1999). Proline under stress: biological role, metabolism and regu/ation. Russian J. Plant Physiol., 46, 274-287.
Lakshmanan, V., S. Venkataramareddy and B. Neelwarne. (2007). Molecular ana/ysis of genetic stabi!íty in /ong-term micropropagated shoots of banana using RAPO and /SSR markers. Electronic Journal of Biotechnology, 1 O( 1 },1-8.
Larcher, W. (1995). Physiological plant ecology. Ecophysiology and stress physio/ogy of functíonal groups. Springer-Verlag, Berlín-Heidelberg.
López-Puc G., A. Canto-Fiick, F. Barreda-Pool, P. Zapata-Castillo, M. Montalvo-Peniche, F. Barahona-Pérez and N. Santana-Buzzy (2006). Oírect Somatic Embryogenesis: A Highly Efficient Protocol for In vitro Regeneration of Habanero Pepper (Capsicum chinense Jacq.) HortScience, 41 (6), 1-7.
Lutts, S., J.M. Kinet and J. Bouharmont (1996). Effects of salt stress on growth, mineral nutrition and prolíne accumulation in relation to osmotic adjustment in rice (Oryza sativa L.) cu/tivars differíng in salinity resistance. Plant Growth Regul, 19, 207-218.
Martins, M., D. Sarmento and M. Oliveira (2004} .Genetíc stability of micropropagated almond plantlets, as assessed by RAPO and /SSR markers. Plant Cell Rep., 23,492-496
Maruyama, E., Y. Hosoi and K. lshii (2007). Somatic embryogenesis and p/ant regenerationin yakutanegoyou, Pinus armandíí Franch. var. amamiana (Koidz.) Hatusima, an endemic and endangered species in Japan. In Vitro Ceii.Dev.Bioi.-Piant., 43, 28-34.
Mercado, J., M. Sancho-Carrascosa and S. Jiméenez-Bermúudez. (2000). Assessment of in vitro growth of apica/ stem sections and adventitious organogenesis to evaluate sa/inity to/erance in cultivated tomato. Plant Cell, Tissue and Organ Culture, 62, 101-106.
Palombi, M., B. Lombardo and E. Caboni (2007). In vitro regeneration of wild pear (Pyrus pyraster Burgsd) clones tolerant to Fe-chlorosis and somaclonal variation ana!ysis by RAPO markers. Plant Cell Rep., 26, 489-496.
Patnaik A. , Chaudhary D., and G.J. Rao ( 2006), Genetic improvement of long grain aroma tic rices through mutatíon approach. Plant M u t. Rep., 1 ( 1 ), 7-1 O
Ramarosandratana, A., L. Harvengt, A. Bouvet, R. Galvayrac and M. Paques (2001 ). Effects of carbohydrate source PEG and gellan fum concentration on embryona/-suspensor mass (ESM) prolíferatíon and maturation of maritime pine somatic embryos. In vitro Cell. Dev. Biol. Plant., 32, 29-34.
Roy, B. and A. Manda!. (2005). Towards development of Al-toxicity toleran! fines in indica rice by exploiting somac!onal variation. Euphytica 145, 221-227.
Rzepka-Pievnes D., D. Kulpa, M. Smolik and M. Glówka (2007). Somaclonal variation in tomato L. pennellí and L. peruvianum f. g!andu/osum characterized in
71

respect to salt tolerance. Journal of Food, Agriculture & Environment.5 (2), 194-201
Sahijram, L., H.J. Soneyi and K.T. Bollamma (2003). Analyzing somaclonal variation in micropropagated bananas (Musa spp.) . In Vitro cellular and Developmental Biology Plant. 39. 551-556.
Sajid Aqeel Ahmad M., F. Javed and M. Ashraf (2007). lso-osmotic effect of NaCI and PEG on growth, cations and free proline accumulation in callus tissue of two indica rice (Oryza saliva L.) genotypes. Plant Growth Regul , 53, 53-63.
Santos-Díaz M. and N. Ochoa-Aiejo (1994). PEG-tolerant ce/1 clones of chili pepper: Growth, osmotic potentials and solote accumulation. Plant Cell, Tiss. Org. Cult., 37, 1-8.
Stewart, G. and J. Lee (1974). The role of proline acumulation in halophytes. Planta, 120, 279-289.
Unnikrishnan, S.K., L. Prakash, P.C. Josekutty, P.N . Bhatt and A.R. Mehta. (1991 ). Effect of NaCI Salinity on Somatic Embryo Development in Sapindus trifoliatus L. Journal of Experimental Botany, 42, 401-406.
Venkataiah, P. , C. Thamidala and K. Subhash (2005). Selection of atrazineresistant plants by in vitro mutagenesis in pepper (Capsicum annuum). Plant Cell , Tissue and Organ Culture, 83, 75-82.
Yoshiba, Y.T., K. Kiyosue, K.Y. Nakashima,Yamaguchi-Shinozaki, K., K. Shinozaki.(1997). Regulation of /evels of proline as an osmolyte in plants under water stress. Plant Ce/1 and Physiology, 38, 1095-1102.
Zambrano, A. , J. Demey and V. González (2003). In Vitro Selection of a Glyphosate-Tolerant Sugarcane Cellular Une. Plant Molecular Biology Reporter, 21 , 365-373.
72

Capitulo IV
Conclusiones y Perspectivas
4.1 CONCLUSIONES
Los marcadores moleculares RAPOs fueron más polimórficos que los ISSRs; sin embargo, ambos permitieron detectar variación somaclonal en los diferentes sistemas de regeneración de chile Habanero estudiados.
La adición de PEG hasta el 15%, al medio de cultivo, tuvo un efecto favorable sobre el desarrollo de los embriones somáticos hacia estadios más avanzados de desarrollo.
Se estableció por primera vez en chile Habanero un protocolo para la obtención una línea embriogénica tolerante a estrés hídrico (Línea: NSBm-04-PEG).
El polimorfismo obtenido mediante marcadores RAPOs en la línea embriogénica tolerante a estrés hídrico, sugiere que el efecto del estrés provocado por el PEG provocó posibles cambios genéticos o epigenéticos, respecto al tratamiento sin PEG.
El incremento en la concentración de PEG en el medio de cultivo promueve un aumento significativo en el contenido endógeno de prolina en los embriones somáticos de chile Habanero, lo que permite inferir que este osmolito puede ser considerado como un marcador bioquímico asociado a la tolerancia a condiciones de estrés hídrico.
73

4.2 PERSPECTIVAS
La denominación de origen otorgada a chile Habanero representa un valor agregado al cultivo de esta especie en la península Yucatán. Por ello, es necesario contar con programas de mejoramiento genético enfocados a la obtención de cultivares mejores adaptados a las condiciones que el cambio climático impone. La variación somaclonal es una herramienta que puede ser aprovechada no solo para selecionar nuevos genotipos tolerantes a estrés hídrico, si no para inducir tolerancia a otros tipos de estrés abiótico.
El presente trabajo representa una importante contribución biotecnológica al género Capsicum y a la especie C. Chinense, y son la base para futuras aplicaciones de nuestros sistemas de regeneración in vitro con fines de mejora genética. Sin embargo, en nuestro estudio sobre la variación somaclonal mediante RAPO e ISSR, recomendamos evaluar otros tipos de marcadores moleculares o bien, realizar estudios sobre posibles cambios epigenéticos en nuestras vías regeneración in vitro. Para el caso especifico de la embriogénesis somática, las anormalidades observadas representan un problema que a la fecha no se ha podido resolver, por lo que recomendamos continuar con los estudios bioquímicos y moleculares que conlleven a lograr una mayor comprensión de este fenómeno que afecta a muchas especies vegetales.
Por otro lado, a fin de profundizar en el conocimiento del polimorfismo revelado en los diferentes sistemas de regeneración in vitro de esta especie, sería importante estudiar la base genética de esta amplia variación observada. Para ello, una de las recomendaciones a corto plazo sería seleccionar y secuenciar bandas polimórficas, especialmente encontradas en las líneas embriogénicas sometidas a estrés, y posteriormente identificar posibles genes relacionados a su tolerancia. De manera similar, se lograría una mayor comprensión del papel de la prolina en la adaptación de los embriones somáticos a condiciones de estrés hídrico, si se consideran realizar estudios adicionales en los que se incluyan cuantificar la acumulación de otros osmolitos compatibles y/o de iones salinos.
74

ANEXOS
Anexo 1. Medio MS (Murashige & Skoog, 1962). Compuesto Macroelementos Nitrato de amonio Nitrato de potasio Cloruro de calcio dihidratado Sulfato de magnesio heptahidratado Fosfato de potasio Sal disódica de etilendiaminotetracetato Sulfato ferroso heptahidratado Microelementos Sulfato de manganeso monohidratado Ácido bórico Yoduro de potasio Molibdato de sodio dihidratado Cloruro de cobalto hexahidratado Sulfato cúprico pentahidratado Sulfato de zinc heptahidratado Suplementos Tia mina Piridoxina Ácido nicotínico Glicina Mionositol Sacarosa
75
Formula
NH4N03 KN03 CaCI2·2H20 MgS04·7H20 KH2P04 Na2EDTA-2H20 FeS04·7H20
MnS04·H20 H3803 Kl Na2Mo04·2H20 CuCI2·6H20 CuS04·6H20 ZnS04·7H20
mg. L'
1,650 1,900 440 370 170 37.3 27.8
16.9 6.2 0.83 0.25 0.025 0.025 8.6
0.10 0.5 0.5 2.0 100 30,000

Anexo 2. Método de extracción de ADN de Dellaporta (1983).
1. Pulverizar con nitrógeno líquido tejido foliar joven recién cortado. 2. En un tubo eppendorf de 1,5 mi colocar material vegetal pulverizado hasta la marca de 500 IJI y resuspender con 750 IJI de buffer CTAB {2X), (2% de CTAB, 100 mM de tris base pH 8,0, 10 mM de EDTA y 0,7 M de NAC1 agregar agua hasta 500 mi y autoclavar). Adicionar 30 IJI de 2-13 mercaptoetanol. Agitar suavemente e incubar por 30 minutos a 65 °C. 3. Agregar 300 IJI de acetato de potasio {3 M, pH 4,8), mezclar suavemente e incubar en hielo por 15 m in. Centrifugar 1 O m in a 14 000 rpm y transferir el sobrenadante a otro tubo. 4. Adicionar 500 IJI de cloroformo: alcohol isoamílico {24:1 ), mezclar suavemente por inversión del tubo y centrifugar 10 mina 14 000 rpm. 5. Cuidadosamente pipetear la fase superior en tubo con 500 IJI de lsopropanol frío para precipitar el ADN . Incubar en hielo durante 60 m in. 6. Centrifugar en 5 min a 14 000 rpm y descartar el sobrenadante. 7. Lavar el pellet de ADN dos veces con 500 IJI de Etanol al 70% frío . Descartar el Etanol y permitir que el ADN se seque al aire libre por 30m in. 8. Resuspender el ADN en 30 a 50 IJI de buffer TE (1 O mM de Tris-HC1, 1 mM de EDTA pH 8,4) o agua estéril y almacenar a -20 °C.
Anexo 3. Protocolo de tratamiento de RNAsa.
1. Secar la pastilla Resuspender en 300 IJL de agua bidestila ultrapura 3.Agregar 3 IJL de RNAsa (10 mg/ mi) 4. Incubar durante 30 min. A 37 °C. S.Precipitar con 1 V de acetato de sodio 3M y 1000 IJL de isopropanol 6.1ncubar 1 ha -80 °C ó tode ADN 2. da la noche a -20 °C ?.Centrifugar a 1300 rpm 20 mina 4 °C 8.Lavar la pastilla 2 veces con 500 IJL de etanol absoluto 9.Secar la pastilla y resuspender en agua bidestila ultrapura
76

Anexo 4. Kit de cebadores RAPOs marca BIOSELEC®. Nombre del
Secuencia Temperatura de
cebador. fusión (T m) Tubek-01 5'-CAT TCG AGC C-3' 32 o e Tubek-02 5'-GTC TCC GCA A-3' 32 o e Tubek-03 5'-CCA GCT TAG G-3' 32 o e Tubek-04 5'-CCG CCC AAA C-3' 34 o e Tubek-05 5'-TCT GTC GAG G-3' 32 o e Tubek-06 5'-CAC CTT TCC C-3' 32 o e Tubek-07 5'-AGC GAG GAA G-3' 32 oc Tubek-08 5'-GAA CAC TGG G-3' 32 o e Tubek-09 5'-CCC TAC CGA C-3' 34 o e Tubek-10 5'-GTG CAA CGT G-3' 32 o e Tubek-11 5'-AAT GCC CCA G-3' 32 o e Tubek-12 5'-TGG CCC TCA C-3' 34 o e Tubek-13 5'-GGT TGT ACC C-3' 32 o e Tubek-14 5'-CCC GCT ACA C-3' 34 o e Tubek-15 5'-CTC CTG CCA A-3' 32 o e Tubek-16 5'-GAG CGT CGA A-3' 32 o e Tubek-17 5' -CCC AGC TGT G-3' 34 o e Tubek-18 5'-CCT AGT CGA G-3' 32 o e Tubek-19 5'-CAC AGG CGG A-3' 34 o e Tubek-20 5'-CAC AGG CGG A-3' 34 o e
77
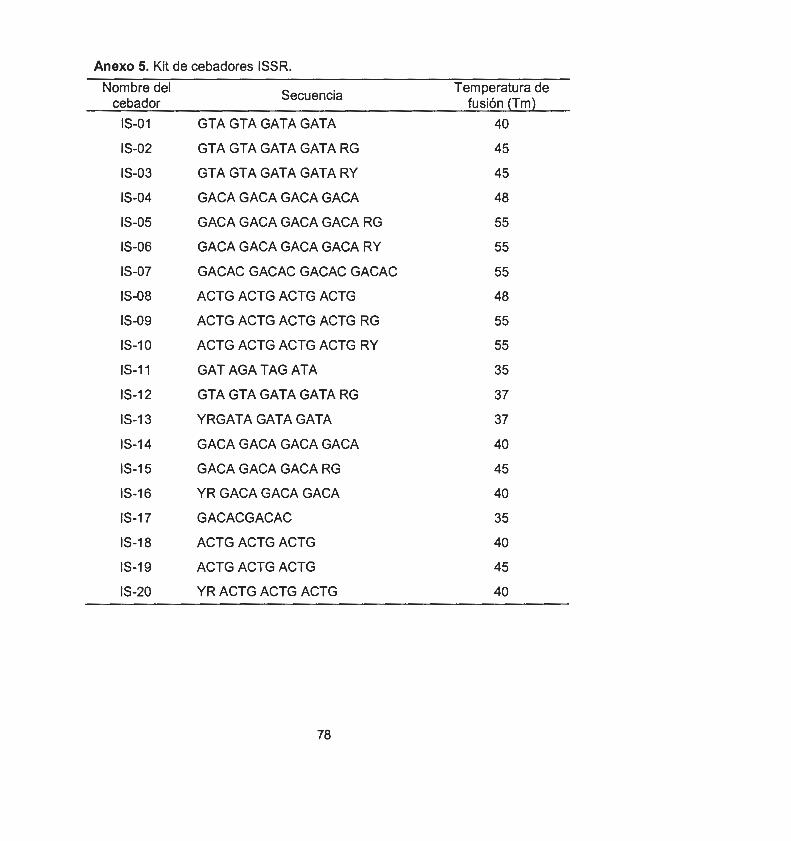
Anexo 5. Kit de cebadores ISSR.
Nombre del Secuencia
Temperatura de cebador fusión (Tm)
IS-01 GTA GTA GATA GATA 40
IS-02 GTA GTA GATA GATA RG 45
IS-03 GTA GTA GATA GATA RY 45
IS-04 GACA GACA GACA GACA 48
IS-05 GACA GACA GACA GACA RG 55
IS-06 GACA GACA GACA GACA RY 55
IS-07 GACAC GACAC GACAC GACAC 55
IS-08 ACTG ACTG ACTG ACTG 48
IS-09 ACTG ACTG ACTG ACTG RG 55
IS-10 ACTG ACTG ACTG ACTG RY 55
IS-11 GAT AGA TAG ATA 35
IS-12 GTA GTA GATA GATA RG 37
IS-13 YRGATA GATA GATA 37
IS-14 GACA GACA GACA GACA 40
IS-15 GACA GACA GACA RG 45
IS-16 YR GACA GACA GACA 40
IS-17 GACACGACAC 35
IS-18 ACTG ACTG ACTG 40
IS-19 ACTG ACTG ACTG 45
IS-20 YR ACTG ACTG ACTG 40
78