genotipizacija slovenskih bolnikov z revmatoidnim … · 2020. 1. 30. · magnezijev klorid nacl...
TRANSCRIPT

Magistrsko delo
GENOTIPIZACIJA SLOVENSKIH BOLNIKOV Z REVMATOIDNIM ARTRITISOM ZA DNA
POLIMORFIZME PREDHODNO POVEZANE Z BOLEZNIJO V ASOCIACIJSKIH ŠTUDIJAH V CELOTNEM
GENOMU
Avgust, 2016 Doroteja Gselman

Doroteja Gselman
Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane
z boleznijo v asociacijskih študijah v celotnem genomu
Magistrsko delo
Maribor, 2016

Genotipizacija slovenskih bolnikov z revmatoidnim
artritisom za DNA polimorfizme predhodno povezane
z boleznijo v asociacijskih študijah v celotnem
genomu
Magistrsko delo študijskega programa II. stopnje
Študent: Doroteja Gselman
Študijski program: magistrski študijski program II. stopnje Kemijska
tehnika
Predvideni strokovni naslov: magistrica inženirka kemijske tehnike
Mentor: red. prof. dr. Uroš Potočnik
Komentor: prim. prof. dr. Artur Pahor, dr. med.
Maribor, 2016


Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
I
Kazalo
Kazalo .......................................................................................................................................... I Izjava......................................................................................................................................... III Zahvala ..................................................................................................................................... IV
Povzetek ..................................................................................................................................... V Abstract ..................................................................................................................................... VI Seznam tabel ............................................................................................................................ VII Seznam slik ............................................................................................................................ VIII Uporabljeni simboli in kratice .................................................................................................. IX
1 Uvod .................................................................................................................................... 1
1.1 Opredelitev problema ................................................................................................... 1
1.2 Namen, hipoteze in cilji ............................................................................................... 1 2 Revmatoidni artritis ............................................................................................................. 3
2.1 Splošno o bolezni ......................................................................................................... 3 2.2 Patogeneza in patologija revmatoidnega artritisa ........................................................ 5
2.3 Klinična slika ............................................................................................................... 6 2.3.1 Prizadetost sklepov ............................................................................................... 7 2.3.2 Prizadetost drugih organov ................................................................................... 8
2.3.3 Laboratorijski testi ................................................................................................ 8 2.4 Diagnoza ...................................................................................................................... 9
2.5 Genetske študije pri RA ............................................................................................. 10 2.5.1 Študije genetske vezave ...................................................................................... 10 2.5.2 Asociacijske študije ............................................................................................ 10
2.6 Genetika RA ............................................................................................................... 13
2.7 Zdravljenje ................................................................................................................. 14 2.8 Farmakološko zdravljenje .......................................................................................... 15
2.8.1 Zdravila z nesteroidnimi protivnetnimi učinkovinami (NSAIDs) in analgetiki . 15
2.8.2 Glukokortikoidi................................................................................................... 16 2.8.3 Imunomodulirajoče protirevmatične učinkovine (DMARDs) ........................... 16
2.8.4 Biološka DMARDs ............................................................................................. 16 2.9 Genotipizacija ............................................................................................................ 17 2.10 Opis SNP-ov uporabljenih v nalogi ........................................................................... 17
2.10.1 rs1571878 (CCR6) .............................................................................................. 17 2.10.2 rs7574865 (STAT4) ............................................................................................. 18
2.10.3 rs909685 (SYNGR1)............................................................................................ 19 2.10.4 rs2476601 (PTPN22) .......................................................................................... 19
2.10.5 rs3087243 (CTLA4) ............................................................................................ 20 3 Eksperimentalni del ........................................................................................................... 21
3.1 Vzorci DNK ............................................................................................................... 21 3.2 Materiali ..................................................................................................................... 21
3.2.1 Osnovne kemikalije ............................................................................................ 21
3.2.2 CEPH vzorci ....................................................................................................... 22 3.2.3 Raztopine in pufri ............................................................................................... 22 3.2.4 Laboratorijska oprema ........................................................................................ 23
3.3 Laboratorijske metode ............................................................................................... 23 3.3.1 Izolacija DNK, RNK in proteinov ...................................................................... 23 3.3.2 Merjenje koncentracij in redčenje DNK ............................................................. 25

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
II
3.3.3 Verižna reakcija z encimom polimerazo (PCR) ................................................. 26 3.3.4 Priprava 2 % agaroznega gela in elektroforeza .................................................. 29
3.3.5 Polimorfizem dolžin restrikcijskih fragmentov (RFLP) ..................................... 29 3.3.6 Metoda HRM ...................................................................................................... 31
3.4 Računalniški programi ............................................................................................... 34 3.4.1 Bioinformatska orodja ........................................................................................ 34 3.4.2 Primer3 ............................................................................................................... 34
3.4.3 SNP cutter ........................................................................................................... 34 3.4.4 GeneRunner 3.05 ................................................................................................ 34 3.4.5 Program SPSS 20.0 ............................................................................................. 37
4 Rezultati ............................................................................................................................. 38 4.1 Izbor polimorfizmov .................................................................................................. 38
4.2 Optimizacija pogojev za RFLP test ........................................................................... 38 4.3 Optimizacija pogojev za HRM test ............................................................................ 42
4.4 Asociacijska analiza izbranih SNP pri bolnikih z RA ............................................... 44 4.4.1 Rezultati RFLP ................................................................................................... 44 4.4.2 Rezultati HRM .................................................................................................... 45 4.4.3 Primerjava rezultatov dobljenih iz genotipizacij ................................................ 45
4.4.4 Rezultati statističnih analiz ................................................................................. 46 5 Diskusija ............................................................................................................................ 51
6 Zaključek ........................................................................................................................... 53 7 Literatura ........................................................................................................................... 54 8 Življenjepis ........................................................................................................................ 59

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
III
Izjava
Izjavljam, da sem magistrsko delo izdelala sama, prispevki drugih so posebej označeni.
Pregledala sem literaturo s področja magistrskega dela po naslednjih geslih:
Vir: PubMed (http://www.ncbi.nlm.nih.gov/pubmed)
Gesla: Število referenc
rheumatoid arthritis 111464
rheumatoid arthritis IN genetics 11457
rheumatoid arthritis IN single nucleotide polymorphism 1443
rheumatoid arthritis IN GWAS 389
rheumatoid arthritis IN treatment IN methotrexate 6181
PCR IN RFLP IN HRM 32
Vir: COBISS/OPAC (http://www.cobiss.si/scripts/cobiss?ukaz=getid, COBIB.SI)
Gesla: Število referenc
revmatoidni artritis IN zdravljenje 78
Skupno število pregledanih člankov: 112
Skupno število pregledanih knjig: 6
Maribor, avgust 2016 Doroteja Gselman

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
IV
Zahvala
Zahvaljujem se mentorju, red. prof. dr. Uroš Potočniku, za vodenje
in svetovanje pri nastanku magistrskega dela. Prav tako se
zahvaljujem somentorju prim. prof. dr. Artur Pahor, dr. med. za
pomoč in pregled magistrskega dela.
Zahvala gre tudi dr. Katji Repnik in Staši Jurgec za pomoč pri delu v
laboratoriju.
Zahvaljujem se tudi svoji družini za vso podporo in spodbujanje pri
študiju.

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
V
Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
Povzetek
Revmatoidni artritis je kronična avtoimunska bolezen s kompleksno genetsko etiolologijo.
Prizadene vse sklepne dele telesa. Študije so pokazale da poleg okoljskih dejavnikov k razvoju
bolezni pomemben delež prispevajo tudi genetski dejavniki. Ocenjen prispevek dednega
dejavnika je 60 %. V patogenezo bolezni je vpleteno veliko število polimorfizmov posameznega
nukleotida (SNP).
Namen magistrskega dela je ugotoviti ali so SNP-eji, ki so bili najbolj statistično značilni v
asociacijskih študijah na celotnem genomu (GWAS), statistično značilni tudi za slovenske
bolnike z RA. V raziskavi smo izbrali rs1571878 (CCR6), rs7574865 (STAT4), rs909685
(SYNGR1), rs2476601 (PTPN22) in rs3087243 (CTLA4). Izvedli smo genotipizacijo s PCR-
RFLP metodo in HRM metodo na 276 vzorcih bolnikov in 276 vzorcih zdravih kontrol. Rezultate
smo analizirali s statistično analizo.
Statistična analiza je pokazala povezavo med RA in SNP-jema rs2476601 (PTPN22) (p = 1,3 x
10-5) in rs3087243 (CTLA4) (p = 0,002). Alel A za SNP rs2476601 zvišuje tveganje za nastanek
RA. Za SNP rs3087243 ima alel G zaščitno vlogo proti nastanku RA.
Ključne besede: revmatoidni artritis, asociacijske študije na celotnem genomu, verižna reakcija
s polimerazo, polimorfizem dolžin celotnega genoma, genotipizacija
UDK: 575.22:616.72-002(043.2)

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
VI
Genotyping of Slovenian patients with rheumatoid arthritis for DNA polymorphisms previously associated with the disease in genome wide association studies
Abstract
Rheumatoid arthritis is a chronic an autoimmune disease with a complex genetic etiololoy. It
affects all joints of the body. Studies have shown that in addition to the environmental factors
genetic factors also contribute an important proportion to the development of the disease. The
estimated contribution of hereditary factors is 60%. However, it is known that a substantial
number of single nucleotid polymorphisms (SNP) are involved in the pathogenesis of the disease.
The aim of the master thesis was to find out whether are the most strongly associated SNP-s in
the genome wide association studies (GWAS) also associated with the slovenian patients with
RA. In this study, we have chosen rs1571878 (CCR6), rs7574865 (STAT4), rs909685 (SYNGR1),
rs2476601 (PTPN22) and rs3087243 (CTLA4). We performed genotypization using PCR-RFLP
method and HRM method in 276 samples of patients and 276 samples of healthy controls. Results
have been analyzed with the statistical analysis.
In the assignment we have confirmed associations between SNPs rs2476601 (PTPN22) (p = 1,3
x 10-5) and rs3087243 (CTLA4) (p = 0,002). Allele A for SNP-rs2476601 increases the risk of
developing RA. For SNP-rs3087243, allel G has a protective role in the development of RA.
Key words: rheumatoid arthritis, Genome Wide Association Studies, Polymerase Chain
Reaction, Restriction Fragment Lenght Polymorphism, genotyping
UDK: 575.22:616.72-002(043.2)

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
VII
Seznam tabel
Tabela 2-1: Klasifikacijska merila RA glede na simptome. ..................................................... 9
Tabela 3-1: Osnovne kemikalije in njihovi proizvajalci......................................................... 21
Tabela 3-2: Izbrani začetni oligonukleotidi (primerji). .......................................................... 22
Tabela 3-3: Laboratorijska oprema in njeni proizvajalci ........................................................ 23
Tabela 3-4: Prikaz protokala za PCR reakcijo. ....................................................................... 28
Tabela 3-5: Izbrani protokol, ki je uporabljen v termociklizatorju. ....................................... 29
Tabela 3-6: Parametri uporabljeni pri RFLP metodi. ............................................................. 31
Tabela 3-7: Protokol za pripravo HRM reakcijske mešanice. ................................................ 34
Tabela 4-1: Kandidatni polimorfizmi posameznega nukleotida (SNP). ................................. 38
Tabela 4-2: Protokol za PCR reakcijo. ................................................................................... 42
Tabela 4-3: Protokol za RFLP reakcijo. ................................................................................. 42
Tabela 4-4: Optimalni pogoji za HRM test. ........................................................................... 43
Tabela 4-5: Ponovno optimizirani optimalni pogoji za HRM. ............................................... 43
Tabela 4-6: Primerjava izračunih frekvenc s frekvencami dobljenih iz projekta HapMap. ... 45
Tabela 4-7: p-vrednosti, razmerja obetov in intervali zaupanja za RA bolnike in kontrole za
CCR6. ..................................................................................................................................... 47
Tabela 4-8: p-vrednosti, razmerja obetov in intervali zaupanja za RA bolnike in kontrole za
SYNGR1. ................................................................................................................................. 48
Tabela 4-9: p-vrednosti, razmerja obetov in intervali zaupanja za RA bolnike in kontrole za
PTPN22. ................................................................................................................................. 49
Tabela 4-10: p-vrednosti, razmerja obetov in intervali zaupanja za RA bolnike in kontrole za
CTLA4. .................................................................................................................................... 50
Tabela 5-1: Primerjava študij za SNP rs2476601. .................................................................. 51

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
VIII
Seznam slik
Slika 2-1: Prikaz simetrične prizadetosti sklepov (rdeča barva) in drugih organov (oči, srce,
pljuča, ledvice) (rumena barva) pri RA [8]. ............................................................................. 4
Slika 2-2: Imunopatogeneza RA [3]. ........................................................................................ 6
Slika 2-3: Prikaz zdravega in revmatoidnega sklepa [3]. ......................................................... 7
Slika 2-4: Vezavno ravnovesje in vezavno neravnovesje [18]. .............................................. 12
Slika 2-5: Položaj gena CCR6 [62]. ........................................................................................ 18
Slika 2-6: Položaj gena STAT4. [63] ....................................................................................... 18
Slika 2-7: Položaj gena SYNGR1 [72]. ................................................................................... 19
Slika 2-8: Položaj gena PTPN22 [73]. .................................................................................... 19
Slika 2-9: Položaj gena CTLA4 [75]. ...................................................................................... 20
Slika 3-1: Mešanica krvi in pufra PBS nanešenega na Ficol. ................................................. 24
Slika 3-2: Krvni vzorec, ki je po centrifugiranje razdeljen na štiri faze. ................................ 24
Slika 3-3: Prikaz postopka PCR reakcije [79]. ....................................................................... 27
Slika 3-4: Prikaz eksponentne rasti tarčne DNK [81]............................................................. 28
Slika 3-5: Primer razreza treh restrikcijskih encimov [83]. .................................................... 30
Slika 3-6: Razklop dvojne vijačnice DNK v enoverižno DNK [86]. ..................................... 32
Slika 3-7: Normalinizirana talilna krivulja. ............................................................................ 33
Slika 3-8: Normaliniziran diferencialni graf. .......................................................................... 33
Slika 3-9: Prikaz uporabe GeneRunner. ................................................................................. 36
Slika 4-1: Optimizacija optimalne temperature pripenjanja začetnih oligonukeotidov za rs909685
(SYNGR1). .............................................................................................................................. 39
Slika 4-2: Optimizacija koncentracij začetnih oligonukleotidov za rs909685 (SYNGR1). .... 40
Slika 4-3: Optimizacija volumna restrikcijskega encima za rs909685 (SYNGR1). ................ 41
Slika 4-4: Shematski in slikovni prikaz detekcije polimorfizma rs909685 (SYNGR1) na
agaroznem gelu. ...................................................................................................................... 44
Slika 4-5: Normalizirana in obdelana talilna krivulja HRM testa. ......................................... 45

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
IX
Uporabljeni simboli in kratice
Simboli c množinska koncentracija (M; mM)
f število obratov (vrt./min; g)
m masa (g)
t čas (h; min; s)
T temperatura (°C)
U napetost (V)
V volumen (L; mL; µL)
Grški simboli
masna koncentracija (g/L)
Δ /
λ valovna dolžina
Kratice
A adenin
anti-CCP protitelesa proti cikličnemu citruliranemu peptidu
bp bazni pari
C citozin
CI interval zaupanja
DNK deoksiribonukleinska kislina
dH2O destilirana voda
dNTP deoksiribonukleozid trifosfat
EDTA etilendiamin tetraocetna kislina
G gvanin
GWAS asociacijske študije na celotnem genomu
HCl klorovodikova kislina
HRM analiza talilne krivulje visoke ločljivosti
LD vezavno neravnovesje
LOD logaritem obetov
MgCl2 magnezijev klorid
NaCl natrijev klorid
NaOH natrijev hidroksid
OR razmerje obetov
p p-vrednost signifikance
PCR verižna reakcija s polimerazo
RA revmatoidni artritis

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
X
RE restrikcijski encim
RFLP polimorfizem dolžin restrikcijskih fragmentov
SNP polimorfizem posameznega nukleotida
T timin
TNF-α faktor tumorske nekroze alfa
TRIS 2-amino 2-hidroksimetil-1,3-propandiol
U enota

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
1
1 Uvod
1.1 Opredelitev problema
Revmatoidni artritis (RA) je kronična, vnetna bolezen, za katero zboli približno 1 %
prebivalstva. Ta bolezen prizadene vse sklepe in tudi notranje organe. Ženske za RA zbolevajo
od tri- do štirikrat pogosteje kot moški. RA se v nosečnosti umiri. Bolezen je posledica
kombinacije genetskih faktorjev in faktorjev okolja. Genetski faktorji predstavljajo približno
65 % dovzetnosti za bolezen [1].
Zdravljenje RA je individualno prilagojeno vsakemu bolniku. Poteka po smernicah, ki
predvidevajo, da je potrebno vedno upoštevati bolnikovo specifično stanje in položaj:
življenjski slog, starost, spol, kako dolgo RA že traja in stopnjo prizadetosi zaradi bolezni.
Glede na aktivnostno skalo bolezni (DAS28, ang. disease activity score) ločimo:
- remisijo,
- manjšo,
- zmerno,
- hudo aktivnost bolezni.
Za zdravljenje RA so na voljo zdravila iz različnih skupin:
- NSAIDS (nesteroidni antirevmatiki),
- glukokortikoidi,
- imunomodulatorji: metotreksat, leflunomid, sulfasalazin ipd. in
- biološka zdravila.
Prvi izbor, ki mu rečejo tudi zlati standard zdravljenja je običajno metotreksat. Zdravniki ga
predpisujejo za lajšanje bolečin in osnovnega vnetja.
Asociacijske študije celotnega genoma (GWA, ang. za genome wide association) so z RA
povezale že več kot 100 lokusov na genomu. GWA študije temeljijo na primerjavi frekvence
bolezenskega alela med kontrolami in bolniki, pri čemer genotipiziramo na tisoče
polimorfizmov posameznega nukleotida (SNP, ang. za single nucleotide polymorphism) v
celotnem genomu na skupini posameznikov z boleznijo in zdravih posameznikih. Kljub temu,
pa velik delež dejavnika dednosti ostaja neznanka. Ker se frekvence polimorfizmov SNP med
populacijami razlikujejo, je pomembno, da asociacijske študije za odkrite povezave
ponavljamo na različnih populacijah. SNP-ji, ki so do sedaj pokazali najmočnejšo povezavo z
RA v genih PTPN22, STAT4, TNFAIP3, PADI4 in CCR6, ki so prav tako biološko zelo
pomembni v patogenezi RA.
1.2 Namen, hipoteze in cilji
Namen magistrskega dela je ugotoviti ali so najmočneje povezani SNP-ji v GWA študijah
povezani tudi s slovenskimi bolniki z RA.
Hipoteza magistrskega dela je, da so izbrani polimorfizmi SNP povezani z RA v slovenski
populaciji.

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
2
V delu smo naredili obsežen pregled do sedaj objavljenih GWA študij na drugih populacijah
ter izbrali 5 SNP-jev. Izvedli smo genotipizacijo na vzorcih bolnikov in vzorcih zdravih
kontrol. Genotipizacijo smo izvedli s PCR-RFLP in HRM metodo. Povezavo analiziranih
SNP-jem smo nato statistično ovrednotili.

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
3
2 Revmatoidni artritis
Poznamo pet glavnih skupin revmatičnih bolezni [2]:
1. Zunaj-sklepni revmatizem
2. vnetne revmatične bolezni (kosti, sklepi, mišice):
a) revmatoidni artritis,
b) spondiloartritis
3. osteoartroza
4. sistemske vezivno-tkivne bolezni, ki lahko prizadenejo tako rekoč sleherni organ v
našem telesu
5. s kristali povzročeni artritisi (metabolni revmatizem).
Revmatoidni artritis (RA) je avtoimunska, kompleksna bolezen. Ta bolezen prizadene vse
sklepe in tudi obsklepne dele. Za RA lahko zboli vsak, najpogosteje pa ženske. Čeprav vzrok
za RA ni znan [3], se domneva, da je povezan s patogenimi povzročitelji ali alergeni novejšega
sveta [4].
2.1 Splošno o bolezni
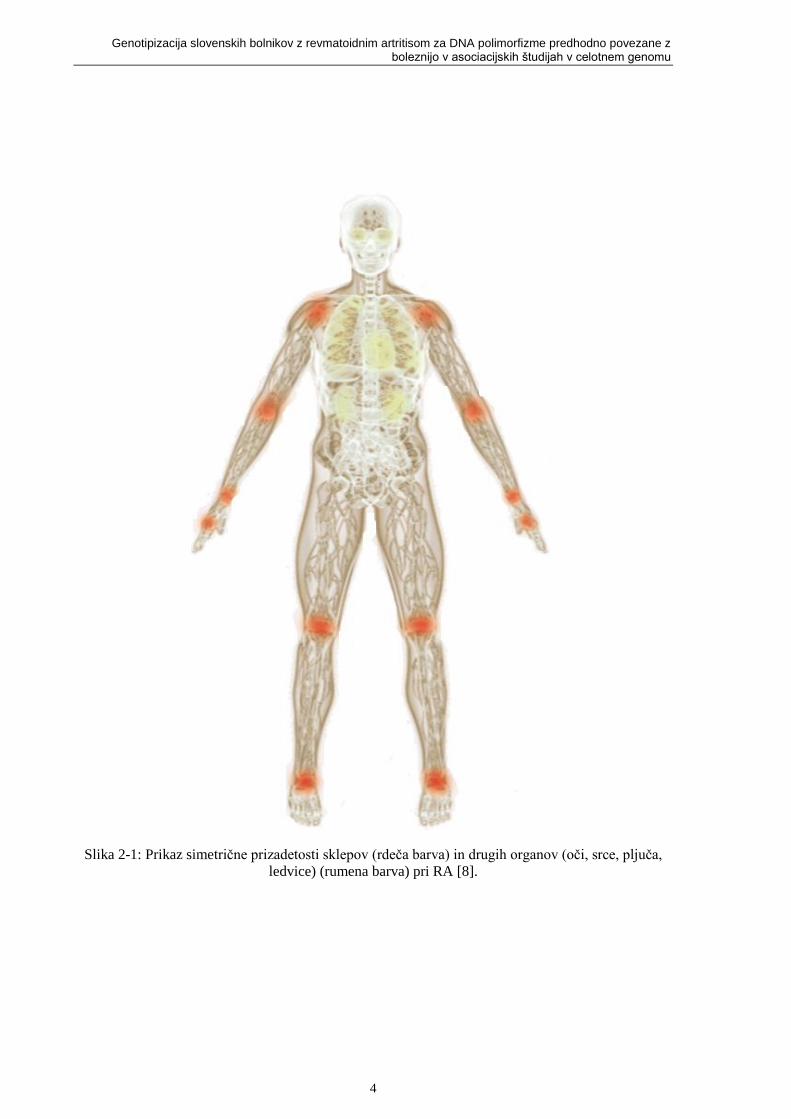
Bolezen ni omejena le na sklepe, saj lahko prizadene skoraj vse organske sisteme (koža, ožilje,
srce, pljuča, oči in mišice) (slika 2-1). RA je razširjen po vsem svetu in prizadene ljudi vseh
ras in etničnih skupin, čeprav se zdi, da sta incidenca in pogostost manjša v ruralni sub-Saharni
Afriki in Karibih [5]. Razširjenost bolezni je približno 1 % in se lahko pojavi v vsaki starosti,
kar pomeni, da lahko zbolijo tako otroci kot tudi starejši. Najpogosteje pa zbolijo ljudje med
30. in 55. letom starosti. Dvakrat do trikrat pogosteje zbolevajo ženske kot moški [4].
Prevalenca se s starostjo poveča, razlika med spoloma pa se zmanjša [5]. V Sloveniji je okrog
20.000 bolnikov s to boleznijo.
Vzročnih dejavnikov za nastanek RA je več. Najpogosteje se bolezen pojavlja pri genetsko
dovzetnih ljudeh. Druge dejavnike, kot so okužbe, poškodbe, hormonske spremembe, zunanji
dejavniki, ki lahko sprožijo RA, še proučujejo. Pomembno vlogo pri sklepnem vnetju ima
imunski sistem. Le-ta ščiti organizem pred bakterijami, virusi in drugimi tujimi snovmi. Pri
RA imunski sistem napade lastna tkiva in sklepe ter druge organe [6].
Če se bolezen ne zdravi ustrezno, lahko privede do nastanka nepopravljivih okvar sklepov in
invalidnosti, posredno pa vodi do povečane umrljivosti. Bolniki z RA živijo 3 do 12 let manj
kot splošna populacija. Vzrok za hitrejšo umrljivost teh bolnikov so pogosto kardiovaskularne
bolezni, katerih incidenca je pri le-teh večja. Ustrezno oz. hitro zdravljenje bolezni ob uporabi
vseh razpoložljivih zdravil (tudi bioloških), lahko zavre napredovanje RA in podaljša življenje
teh bolnikov [7].
Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
4
Slika 2-1: Prikaz simetrične prizadetosti sklepov (rdeča barva) in drugih organov (oči, srce, pljuča,
ledvice) (rumena barva) pri RA [8].

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
5
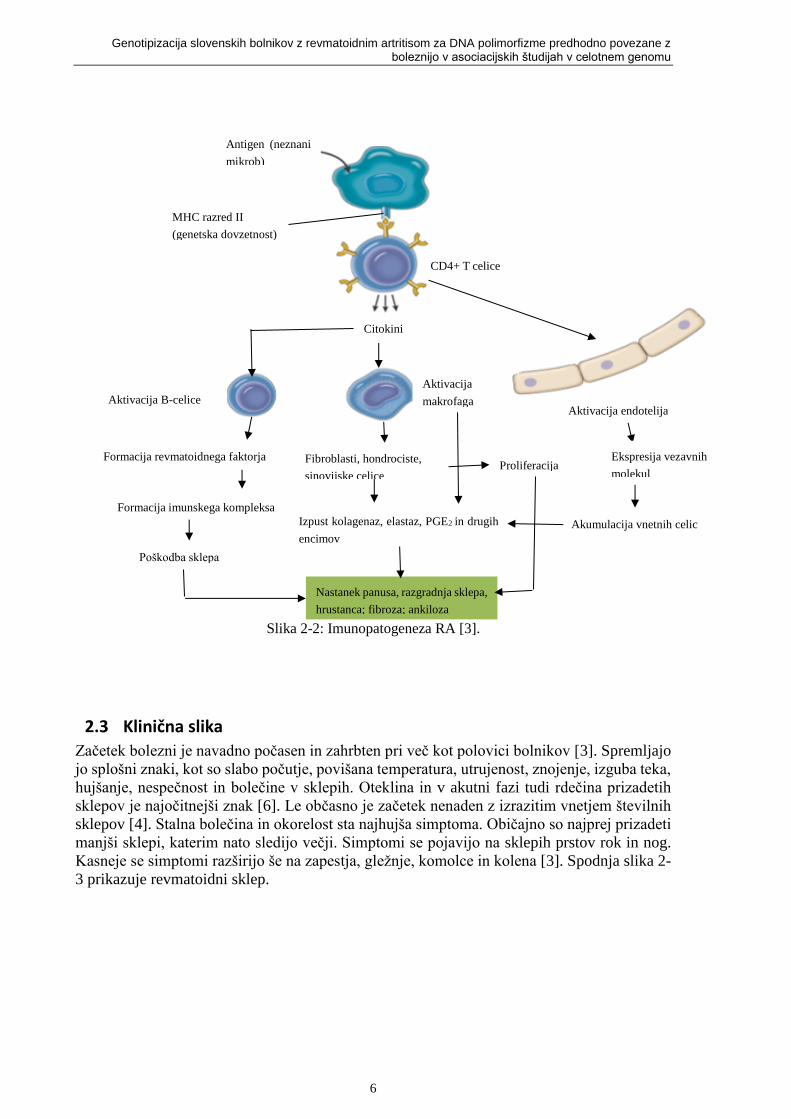
2.2 Patogeneza in patologija revmatoidnega artritisa RA je avtoimunska bolezen, ki jo sproži izpostavljanje genetsko dovzetnega gostitelja
neznanemu antigenu. Ponavljajoča avtoimunska reakcija z aktivacijo T celic pomagalk in
drugih limfocitov ter lokalno sproščanje vnetnih mediatorjev in citokinov je proces, ki uniči
sklep [3]. Postopek patogeneze prikazuje slika 2-2.
Antigen, ki sproži bolezensko dogajanje še ni znan. Bilo je veliko zanimanja za raziskovanje
mikrobnih antigenov kot začetnih povzročiteljev RA. Vendar za zdaj še niso našli trdnih
dokazov za identifikacijo mikroorganizma kot etiološkega agensa pri RA [3]. Antigen pride v
sinovijsko ovojnico. V sinovijski ovojnici le-tega predelajo antigen predstavitvene celice, ki
so lahko sinoviociti A (makrofagom podobne celice), makrofagi ali dentritične celice. Vse
predstavitvene celice izražajo antigene HLA-DR na svojih celičnih ovojnicah [4].
Med citokini, ki jih izločajo aktivirane antigen predstavitvene celice, sta najpomembnejša
interlevkin 1 (IL-1) in tumorje nekrotizirajoči faktor alfa (TNF-alfa) [3], [4]. Omenjena
citokina sta provnetna mediatorja, ki vzpodbujata različne celice (T celice pomagalke,
sinovijske celice, hondrocite, fibroblaste in endotelijske celice). Prav tako vplivata tudi na
razvoj simptomov pri RA (izguba apetita, hujšanje in povišana telesna temperatura). Pri RA
sta pomembna tudi limfokina interlevkin 2 (IL-2) in interferon gama (INF-gama). IL-2
spodbuja rast limfocitov T in B in diferenciacijo, ter aktivira T celice pomagalke. IFN-gama
aktivira makrofage in na antigen predstavitvenih celicah povečuje izražanje gena HLA-DR.
Sinovijske celice, hondrociti in fibroblasti pod vplivom provnetnih citokinov proliferirajo in
izločajo prostaglandin E2 in proteolitične encime [4].
Adhezijske molekule, ki jih izražajo aktivirane endotelijske celice na svoji površini,
privabljajo limfocite T in druge provnetne celice. Le-te nato migrirajo v sinovijsko tkivo.
Sinovijske, epitelijsko urejene celice nabreknejo in sinovija se zadebeli. Tvori se granulacijsko
tkivo, ki mu rečemo panus [4]. Ta se vrašča v sklepni hrustanec in ga razgrajuje. Po razgradnji
hrustanca se panus začne vraščati na kosti in tvori fibrozno ankilozo, ki okosteni [3].
Makrofagi in aktivirane endotelijske celice sintetizirajo levkotrien B4, ki deluje na granulocite
močno kemotaktično. Vzporedno citokini spodbujajo še proliferacijo limfocitov B in tvorbo
protiteles. Imunski kompleksi, ki jih protitelesa tvorijo z antigenom, se sproščajo v sklep. Tam
se vežejo v komplement, ki jih nato fagocitirajo polimorfonuklearni levkociti. Med fagocitozo
se tvorijo in sproščajo kisikovi prosti radikali, prostaglandini in različne proteaze, ki
povzročajo tkivno okvaro [4].
Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
6
Slika 2-2: Imunopatogeneza RA [3].
2.3 Klinična slika
Začetek bolezni je navadno počasen in zahrbten pri več kot polovici bolnikov [3]. Spremljajo
jo splošni znaki, kot so slabo počutje, povišana temperatura, utrujenost, znojenje, izguba teka,
hujšanje, nespečnost in bolečine v sklepih. Oteklina in v akutni fazi tudi rdečina prizadetih
sklepov je najočitnejši znak [6]. Le občasno je začetek nenaden z izrazitim vnetjem številnih
sklepov [4]. Stalna bolečina in okorelost sta najhujša simptoma. Običajno so najprej prizadeti
manjši sklepi, katerim nato sledijo večji. Simptomi se pojavijo na sklepih prstov rok in nog.
Kasneje se simptomi razširijo še na zapestja, gležnje, komolce in kolena [3]. Spodnja slika 2-
3 prikazuje revmatoidni sklep.
Antigen (neznani
mikrob)
MHC razred II
(genetska dovzetnost)
CD4+ T celice
Citokini
Aktivacija B-celice
Aktivacija
makrofaga Aktivacija endotelija
Formacija revmatoidnega faktorja
Formacija imunskega kompleksa
Fibroblasti, hondrociste,
sinovijske celice
Izpust kolagenaz, elastaz, PGE2 in drugih
encimov
Poškodba sklepa
Nastanek panusa, razgradnja sklepa,
hrustanca; fibroza; ankiloza
Ekspresija vezavnih
molekul Proliferacija
Akumulacija vnetnih celic

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
7
Slika 2-3: Prikaz zdravega in revmatoidnega sklepa [3].
2.3.1 Prizadetost sklepov
Sinovitis pri RA simetrično zajame različne sklepe, najpogosteje metakarpofalangealne in
proksimalne interfalangealne sklepe rok in/ali zapestja ter metatarzofalangealne sklepe stopal.
Prizadeti so lahko tudi vsi veliki periferni sklepi in mali sinovijski sklepi hrbtenice, posebno
v vratnem delu (redko na začetku bolezni). Bolečina v sklepih je pri RA stalna, bolj intenzivna
je zgodaj zjutraj. Prizadeti sklepi so otekli, v kronični fazi tudi toplejši od okolice in včasih
pordeli. Zaradi bolečine in otekline so sklepi tudi omejeno gibljivi. Okorelost prizadetih
sklepov je pomemben simptom RA in lahko traja več ur. Najizrazitejša je zjutraj in po daljšem
počitku ter se pogosto ujema z aktivnostjo bolezni [4].
Pri napredovani, slabo zdravljeni ali nezdravljeni bolezni se razvijejo deformacije sklepov,
najpogosteje ulnarna deviacija in deformacije prstov rok v obliki labodjega vratu ter flektorne
kontrakture komolcev in kolen [4].
Pri RA je pogost sinovitis upogibalnih in izteznih kitnih ovojnic na rokah. Najpogosteje ga
vidimo nad zapestji in na hrbtišču rok, kjer se pojavi kot podkožna vretenasta elastična in
boleča zatrdlina. V kronični fazi RA so prisotne tudi sinovijske ciste in sinovijske hernije, ki
nastanejo zaradi povečanega pritiska v vnetno spremenjenih sklepih. Najpogostejša je
Bakerjeva cista v kolenski jami, ki nastane zaradi raztezanja središčnih kitnih burz
gastroknemične in semitendinozne mišice [4].
Dentridska
celica

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
8
2.3.2 Prizadetost drugih organov
Zunajsklepno prizadetost se opaža pri bolnikih s hujšo obliko bolezni. Domnevajo, da se
razvije zaradi vpliva imunskih kompleksov, ki sprožijo vaskulitis in zaradi vpliva rastnih
faktorjev. Oblike izvensklepnih prizadetosti so lahko naslednje [4]:
- Revmatoidni vozliči nastanejo predvsem pri serološko pozitivnih bolnikih (pri 20 do
25 %). Nahajajo se podkožno, na mehanično izpostavljenih mestih: iztezne strani
komolcev, podlakti, na malih sklepih rok, nad kolenoma, redkeje tudi v notranjih
organih. Histološka slika revmatoidnega vozliča prikazuje v sredini prisotna
fibrinoidna nekroza veziva, katero obdaja plast palisadno razporejenih fibroblastov, ki
so obkroženi z granulacijskim tkivom, ta pa vsebuje številne skupke mononuklearnih
celic ob žilah.
- Kožni vaskulitis se pojavlja pr hujši obliki RA in prizadane oba spola enako. Pojavlja
se kot pikčaste krvavitve, tipna purpura, drobni infarkti ob nohtih, kožne razjede,
gangrena. Razjede na koži se pogosto zagnojijo in so prisotne na hrbtišču stopal in
okoli gležnjev.
- Vaskulitis notranjih organov je možen na srcu, pljučih, črevesju, ledvicah, jetrih,
trebušni slinavki, bezgavkah, modih.
- Revmatična pljuča so najpogostejša komplikacija revmatoidnega artritisa. Prizadeta
sta lahko pljučni parenhim z ožiljem in plevra, kar se kaže kot kronična fibrozirajoča
pljučnica, revmatoidni vozliči, difuzni fibrozirajoči alveolitis ali plevritis. Omenjene
spremembe pljuč se navadno razvijejo pri dalj časa trajajočem RA. Redko kot prvi in
izolirani pojav bolezni.
- Pri spremembah na srcu je najpogostejši perikarditis (1 do 10 % bolnikov), ki se
navadno pojavi zgodaj, ko je RA še zelo aktivna. Redko se pojavlja endokarditis,
revmatoidni vozliči, miokarditis in koronaritis.
- Spremembe na očeh so pri RA pogoste. Pojavi se lahko episkleritis, skleritis,
prizadetost veznice in roženice v sklopu sindroma suhih oči. Pojavijo se lahko depoziti
zlata v roženici kot posledica zdravljenja RA z zlatom, spremembe mrežnice po
jemanju antimalarikov in siva mrena po zdravljenju s kortikosteroidi.
- Nevrološka prizadetost. Utesnitveni sindromi, difuzna senzorična nevropatija, multipli
mononevritis, vratna mielopatija (pri atlantoaksialni dislokaciji ali subluksaciji).
- Amiloidoza prizadane 5–15 % bolnikov in je najpogostejša v ledvicah, redkeje na srcu,
jetrih, vranici ali prebavilih.
2.3.3 Laboratorijski testi
Avtoimunske bolezni kot je RA so pogosto karakterizirane s prisotnostjo protiteles.
Revmatoidni faktor ni specifičen za RA in je lahko prisoten tako pri bolnikih z drugimi bolezni
(npr. hepatitis C) kot tudi pri zdravih ljudeh. Protitelesa proti cikličnemu citruliranemu peptidu
(anti-CCP) so visoko specifična za RA in lahko imajo vlogo pri patogenezi bolezni [7]. Poleg
diagnostične vrednosti, ki jo imajo, so tudi dober prognostični dejavnik. Približno 50 do 80 %
bolnikov z RA ima v krvi revmatoidni faktor, anti-CCP ali oboje, takrat govorimo o serološko
pozitivnem RA. IgM proti revmatoidnemu faktorju določamo z Waaler-Rosejevo reakcijo in
lateks testom, z encimsko imunskim testom (ELISA) pa poleg IgM določamo še IgA in IgG
proti revmatoidnemu faktorju. Sedimentacija eritrocitov je navadno pospešena in v sorazmerju
z aktivnostjo bolezni. Koncentracija reaktivnega proteina C v serumu je običajna. Pogosto se
pojavlja normocitna ali mikrocitna anemija [4]. S hitrostjo sedimentacije eritrocitov in
koncentracije reaktivnega proteina C v serumu lahko sledimo napredku bolezni in odzivu na
zdravila [7]. Sočasna prisotnost anti-CCP in IgM proti revmatoidnemu faktorju namreč kaže
na bolj neugoden potek bolezni z zelo zgodnjim pojavom erozij [4].

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
9
2.4 Diagnoza Diagnozo RA postavi osebni zdravnik, ki jo nato potrdi revmatolog. Bolezen diagnosticiramo
na podlagi bolnikovih težav, telesnega pregleda, s krvnimi preiskavami, rentgenskim
slikanjem, ultrazvokom, ipd. Pri postavitvi diagnoze so bila v pomoč klasifikacijska merila iz
leta 1987, ki jih prikazuje tabela 2-1. Vendar so bila merila iz leta 1987 primernejša za
postavitev diagnoze pri bolnikih z že razvito obliko bolezni. Za zgodnjo odkrivanje
nediferenciranih oblik artritisa pa imajo nizko občutljivost [9]. Sodobno zdravljenje narekuje
hitro diagnozo, zato so strokovnjaki Ameriškega revmatološkega združenja in Evropske lige
proti revmatizmu leta 2010 izdelali nova klasifikacijska merila (Tabela 2-1) [10].
Tabela 2-1: Klasifikacijska merila RA glede na simptome [11].
Kriteriji Ameriškega
revmatološkega združenja leta
1987
Kriteriji Ameriškega
revmatološkega združenja leta
2010
1. Jutranja otrdelost sklepov vsaj 1 uro
pred popolnim izboljšanjem
2. Artritis treh ali več sklepov
3. Artritis na sklepih v roki
4. Simetrični artritis
5. Revmatoidni noduli
6. Pozitiven revmatoidni faktor
7. Radiografske spremembe na rokah
ali zapestjih (erozija ali pomanjkanje
kalcija)
1. Prizadetost sklepov (0-5 točk))
- Eden srednji ali velik sklep (0 točk)
- Dva do deset srednjih ali velikih
sklepov (1 točka)
- Ena do tri malih sklepov (2 točki)
- Štiri do deset sklepov (veliki sklepi
niso šteti)(3 točke)
- Več kot deset sklepov (vsaj en mali
sklep)(5 točk)
2. Serologija (0-3 točke)
- Negativen revmatoidni faktor in
negativen proti-citrulinirani
protein/protitelesa (0 točk)
- Nizko pozitiven revmatoidni faktor ali
nizko pozitiven proti citrulinirani
protein (2 točki)
- Visoko pozitiven revmatoidni faktor
ali visoko pozitiven proti citrulinirani
protein (3 točke)
3. Akutni parametri vnetja (0-1 točka)
- Reaktivn protein C ni zvišan in
sedimentacija eritrocitov ni pospešena
(0 točk)
- Zvišan reaktivni protein C ali
pospešena sedimentacija eritrocitov (1
točka)
4. Trajanje simptomov (0-1 točka)
- Manj kot šest tednov (0 točk)
- Šest tednov ali več (1 točka)
Pri bolniku z razvito klinično sliko serološko pozitivnega RA pravilne diagnoze običajno ni
težko določiti. Težje je ločiti začetne znake RA od drugih sorodnih bolezni. Virusne okužbe
potekajo pogosto s sklepnimi bolečinami in celo otekanjem sklepov. Nekaterim vrstam
bakterijskih okužb lahko sledijo artritisi. Diferencialno diagnostično nam v teh primerih
pomagajo natančna anamneza o okužbi ter pozitivni laboratorijski testi na povzročitelja
okužbe.

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
10
2.5 Genetske študije pri RA Najpomembnejši pristopi, ki se uporabljajo za odkrivanje genetskih lokusov, povezanih z RA
so [12]:
- študije genetske vezave,
- asociacijske študije:
- asociacijske študije kandidatnih genov
- asociacijske študije celotnega genoma.
Uporaba študij genetske vezave in študij kandidatnih genov za odkrivanje genetske
dovzetnosti ni dala nobenih dokončnih odgovorov glede etiologije RA [13]. Asociacijske
študije celotnega genoma so se izkazale za najbolj uspešne [12].
2.5.1 Študije genetske vezave
Pretekla desetletja so bile analize genetske vezave vodilne pri genetskih študijah bolezni.
Študije genetske vezave testirajo kosegregacijo genetskih markerjev in fenotipov bolezni z
uporabo družinskih podatkov. Signifikantni rezultat genetske vezave pokaže, da sta marker in
susceptibilni gen genetsko povezana. Analize vezave so bile zelo uspešne pri mnogih redkih
monogenskih bolezni.
Slabost študije analize genetske vezave je v tem, da potrebuje veliko obolelih svojcev [15].
Omejen uspeh analize genetske vezave pri kompleksnih boleznih je zaradi nezadostne moči
in resolucij za odkrivanje variant [12]. Alternativna metoda za genetske študije vezave so
asociacijske študije.
Cilj analize genetske vezave je določiti nahajanje gena ali genov, ki so odgovorni za nastanek
bolezni. Poznamo dve vrsti analize genetske vezave: parametrično in neparametrično. Če je
dovolj informacij glede parametrov (dednost in DNK številnih članov iste družine), uporabimo
parametrično študijo genetske vezave. Če pa je genetski model neznan, se uporabi
neparametrična študija. Parametrična študija genetske vezave je močna strategija za mapiranje
genov z Mendlovim tipom dedovanja. Rezultat te študije je izražen z LOD vrednostjo
(logaritem obetov). LOD vrednost je funkcija rekombinantnega količnika (ϴ). Rekombinantni
količnik poda verjetnost rekombinacije med dvema markerjema. Dva lokusa sta povezama
takrat, kadar je rekombinantni količnik manj kot 0,5. Če je LOD vrednost 3 (kar ponazarja
verjetnost 1000:1) ali več je to dokaz statistično signifikantne vezave. Negativna LOD
vrednost dokazuje odsotnost vezave [16].
2.5.2 Asociacijske študije
Namen genetskih asociacijskih študij je odkriti povezavo med enim ali več genetskimi
polimorfizmi in lastnostmi, ki so lahko kvantitativno značilne ali diskretni atributi ali bolezni.
Asociacijska študija se razlikuje od vezavne študije po tem, da je isti alel (ali aleli) povezan z
lastnostmi na podoben način skozi celotno populacijo, medtem, ko so pri analizi vezave
različni aleli asociirani z lastnostjo v različnih družinah. Asociacijske študije lahko
obravnavamo kot študije vezave, saj imamo ljudje skupne prednike, kjer je povečana družina
širša populacija. Asociacijske študije delujejo na kratke razdalje v genomu. Z asociacijskimi
študijami lahko odkrijemo majhne učinke, ki jih imajo geni, ki so prisotni pri običajnih
kompleksnih boleznih. Toda pregledati moramo veliko več markerjev kot pri vezavni. To

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
11
spoznanje je, skupaj z identifikacijo velikega števila polimorfizmov posameznega nukleotida
(SNP) v genomu in zmanjšanja stroškov genotipizacije, privedlo do pomembnosti
asociacijskih študij v genetski epidemiologiji [17].
Z asociacijskimi študijami skušamo najti povezavo med polimorfizmi SNP in boleznijo pri
posameznikih, ki niso v sorodu. Asociacijo izvedemo tako, da primerjamo frekvence alelov
med skupino bolnikov in kontrolno skupino zdravih posameznikov, t.j. študija
primeri:kontrole.
Asociacijske študije so deljene na posredne in neposredne študije. Pri posrednih asociacijskih
študijah je kandidatni gen znan. Asociacija je direktna. Metoda uporablja kavzalno mutacijo
določenega gena pri analizi asociacije bolezenskega fenotipa. Pri neposredni metodi
kandidatni gen ni znan in je vezan na genske markerje. Vezavno neravnovesje (LD – linkage
disequilibrium) je oblika neposredne metode [17].
Vezavno neravnovesje (LD – linkage disequilibrium)
Kartiranje genov za kompleksne motnje in drugih genskih značilnosti z indirektno metodo je
odvisno od asociacije na stopnji populacije, med naključnimi variacijami in bližnjimi markerji
[17].
LD opisuje zvezo med aleli, ki so na različnih lokusih. Vezavno neravnovesje med aleloma je
takrat, ko asociacija med dvema aleloma v dveh lokusih ni naključna. Če pa je asociacija med
aleloma v dveh lokusih naključna, takrat govorimo o vezavnem ravnovesju (LE-linkage
equilibrium) [16].
Za merjenje LD se uporablja veliko različnih parametrov, med njimi tudi Lewontinov D', ki
se imenuje tudi »asociacijska verjetnost« [7] in indeks r2 [8]. Lewontinov D' je pomemben
parameter za identifikacijo regij, v katerih je bilo malo rekombinacij in kjer je možnost za
kartiranje kavzalnih lokusov z indirektnimi asociacijskimi študijami. [6] Kadar je vrednost D'
= 0, takrat govorimo o popolnem vezavnem ravnovesju. D' = 1 ponazarja popolno LD, kjer ni
rekombinacije med dvema markerjema pri populaciji [8]. Indeks r2 nam pokaže moč
neposrednih študij. Kljub temu, da so loci v popolnem neravnovesju (D' = 1), lahko indeks r2
variira. Vrednosti indeksa r2 so povezane s frekvencami alelov in s pozicijo korespodenčnih
mutacij [17].
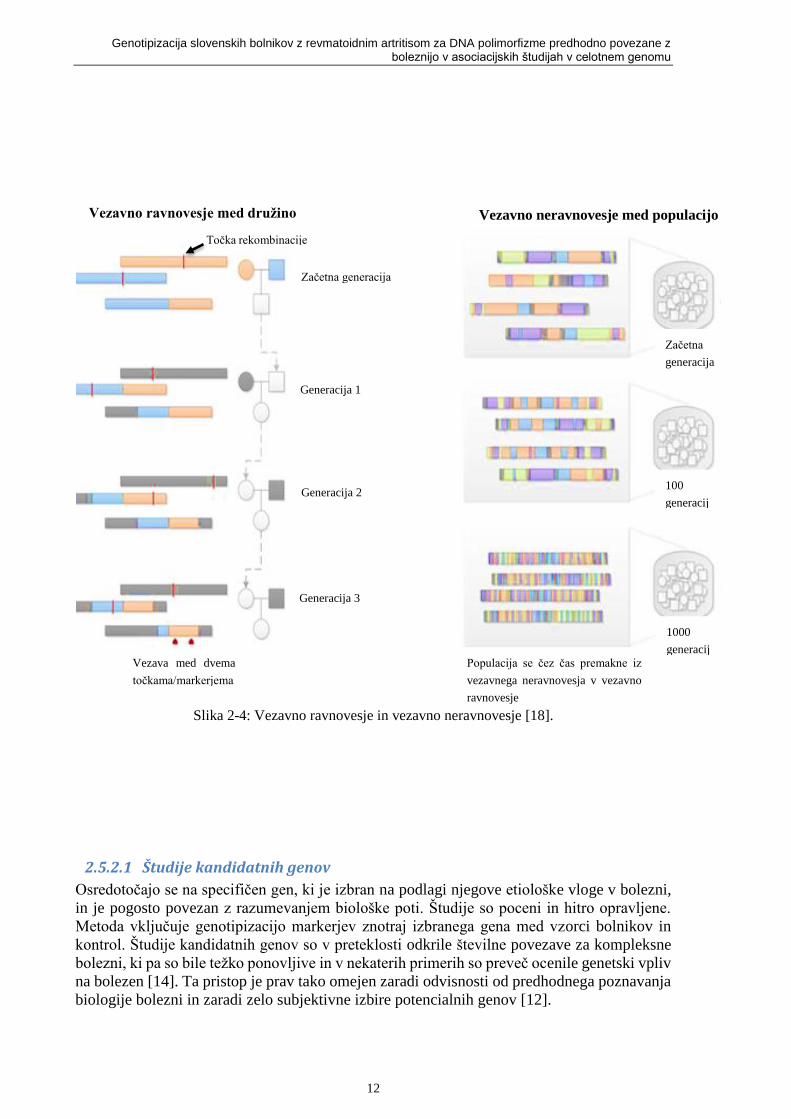
Slika 2-4 prikazuje dva originalna kromosoma (eden je modri, drugi oranžen). Rekombinacije
v družini iz generacije v generacijo ločujejo kromosomske segmente. Učinek se skozi
generacijo še ojača in na populaciji fiksne velikosti doživlja naključno parjenje. Ponavljajoča
naključna rekombinacija bo ločila segmente strnjenega kromosoma (vsebuje povezane alele),
dokler ne bodo vsi aleli v populaciji v vezavnem ravnovesju ali pa bodo neodvisni. Vezavo
med markerji pri populaciji imenujemo vezavno neravnovesje [18].
Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
12
Slika 2-4: Vezavno ravnovesje in vezavno neravnovesje [18].
2.5.2.1 Študije kandidatnih genov
Osredotočajo se na specifičen gen, ki je izbran na podlagi njegove etiološke vloge v bolezni,
in je pogosto povezan z razumevanjem biološke poti. Študije so poceni in hitro opravljene.
Metoda vključuje genotipizacijo markerjev znotraj izbranega gena med vzorci bolnikov in
kontrol. Študije kandidatnih genov so v preteklosti odkrile številne povezave za kompleksne
bolezni, ki pa so bile težko ponovljive in v nekaterih primerih so preveč ocenile genetski vpliv
na bolezen [14]. Ta pristop je prav tako omejen zaradi odvisnosti od predhodnega poznavanja
biologije bolezni in zaradi zelo subjektivne izbire potencialnih genov [12].
Vezavno ravnovesje med družino
Točka rekombinacije
Začetna generacija
Generacija 1
Generacija 2
Generacija 3
Vezava med dvema
točkama/markerjema
Vezavno neravnovesje med populacijo
Začetna
generacija
100
generacij
1000
generacij Populacija se čez čas premakne iz
vezavnega neravnovesja v vezavno
ravnovesje

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
13
2.5.2.2 Asociacijske študije na celotnem genomu
Asociacijske študije na celotnem genomu (ang. Genome Wide Association Studies, GWAS)
sistematično preiščejo celoten genom z namenom odkrivanja SNP-jev, povezanih z boleznijo.
Zadnjih nekaj let so GWAS pripomogle pri razumevanju in odkrivanju polimorfizmov,
povezanih s kompleksnimi bolezni in tako predstavljajo zanesljiv in na široko uporabljen
pristop [12]. Uspeh GWA študij se lahko pripiše trem glavnim dejavnikom v humani genetiki:
(1) Zaključek projekta Humanega genoma leta 2001 [19], katerega cilj je bil razviti
haplotipsko strukturo humanega genoma, ki bo opisal skupne vzorce genskih variacij
človeškega genoma in zaključek HapMap projekta leta 2003 [20].
(2) Razvoj zanesljivih in cenovno ugodnih platform za genotipizacijo [21].
(3) Oblikovanje konzorcijev za proučevanje velikega števila kompleksnih bolezni [21].
GWA študije genotipizirajo veliko število SNP-jev v celotnem genomu na veliki skupini
bolnikov (primeri) in zdravih posameznikov (kontrole). Nična hipoteza GWA študije pravi,
da med danim SNP-jem in nastankom bolezni ni povezave. Hipoteza se testira s primerjavo
alelnih frekvenc ali s frekvencami genotipov med primeri in kontrolami. Nična hipoteza ni
potrjena takrat, ko je nivo signifikance p = 5 x 10-8 in je genski marker zaznal prisotnost
vzročne variante [22]. Napredek v tehnologiji, predvsem pri razvoju DNK mikročipov in nižji
stroški, so omogočili razpon GWA študij. Označevalci, ki jih za posamezno bolezen
ugotovimo, so pogosto v vezavnem neravnotežju [12].
Uspeh in popularnost te strategije je prikazana v spletnem katalogu, ki vsebuje 812 objavljenih
GWA študij za 476 različnih kompleksnih bolezni od marca 2011 in še vedno raste [12].
2.6 Genetika RA
Prve dokaze za genetsko dovzetnost pri RA so dokazale prav študije dvojčkov in družinske
študije. Študije dvojčkov so pokazale vlogo genetskih faktorjev za RA, s konkordanco od 12-
15 % med enojajčnimi dvojčki in 2-4 % med dvojajčnimi dvojčki [23], [24], [25]. Družinske
študije in študije dvojčkov so pokazale velik vpliv genetske komponenten na razvoj RA,
katerih dednost je ocenjena med 50-60 % [24], [26].
Najbolj pomemben genetski faktor za RA je HLA lokus [22]. Študije so pokazale, da so
številni HLA-DRB1 aleli povezani z RA in da so različni aleli povezani z različnimi etničnimi
populacijami [27], [28].
HLA-DRB1 aleli, ki so znotraj glavnega histokompatibilnega kompleksa (MHC), so
odgovorni za imunski odziv. Peptidi, produkti teh genov, so na površinah celic in so odgovorni
za vezavo HLA-antigena in posredovanje le tega limfocitom T. Vsi HLA-DRB1 aleli, ki so
povezani s pojavom RA, imajo skupno regijo podobnih zaporedij. Ti aleli se imenujejo tudi
skupni epitopi (SE) [25], [29]. Domneva se, da so prav SE odgovorni pri patogenezi RA, ki
določijo kateri patogeni peptid bo predstavljen limfocitom T. Natančen mehanizem tega
procesa še ni poznan [25].
HLA lokus je najbolj močan genetski faktor in predstavlja okoli 30 % genetske dovzetnosti za
RA [30].
Preteklo je skoraj 30 let, preden so odkrili genetske povezave zunaj HLA lokusa. Najprej sta
bila odkrita gena PADI4 in PTPN22. Prvi gen, povezan z RA, je bil PADI4 in je bil odkrit pri

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
14
japonski populaciji na kromosomu 1p36 leta 2003 [31]. PADI4 spada v gensko družino
peptidil arginin deaminaz (PADI4). Ta gen kodira PAD encim, kateri je potreben za
posttranslacijsko modifikacijo arginina v citrulin. Asociacijo so uspešno ponovili še na drugih
azijskih populacijah. Niso je pa mogli ponoviti na populaciji evropejcev [32], [33]. Po odkritju
gena PADI4 so v povezavi z RA odkrili še gen PTPN22 leta 2004 pri evropejski populaciji
[34]. Po letu 2004 se je odkrivanje novih genetskih faktorjev za RA pospešilo zaradi napredka
pri genotipizaciji. Z asociacijsko študijo kandidatnega gena so odkrili CTLA4 gen [35],
kateremu je sledilo odkritje TRAF1/C5 regije [36]. Povezava gena STAT4 z RA je bila odkrita
s kombinacijo asociacijske študije kandidatnih genov in študije genetske vezave [37].
Številne GWA študije so bile izvedene na svetovni populaciji in so odkrile številne z RA
lokuse, vključno s PADI4, PTPN22 [38], TNFAIP3 [39], TRAF1/C5 [40], REL [41] in CCR6
[42]. REL predstavlja dober primer RA lokusa, ki je bil odkrit z neodvisno GWA študijo [41]
in ga niso odkrili s predhodno asociacijsko študijo [43]. Vendar pa vsaki od teh študij
primanjkuje statistične moči za odkritje lokusov, ki imajo zmerno velikost [22]. To omejitev
so premostili z meta-analizami. Leta 2008 so odkrili dodatni lokus z meta-analizo različnih
GWAS, ki se nahaja v bližini gena CD40 [44]. Z meta-analizami GWA študij evropejskih in
azijskih populacij so povečali število lokusov [45], [46]. Nedavno nazaj so izvedli večetnično
metastatistično GWA študijo, v kateri je sodelovalo 25 študijskih skupin iz celega sveta in
skupaj več kot 100,000 preiskovancev (29,880 primerov RA in 73,758 kontrol) azijskega in
evropejskega porekla. To je bila največja meta-analiza za imunske bolezni GWA študije, ki
so bile kadarkoli izvedene. V tej študiji so potrdili 101 genetskih lokus povezan z nastankom
RA [47].
Imunočip olajšuje gosto genotipizacijo in kartiranje 186 genetskih lokusov, vključno z že
potrjenimi avtoimunskimi lokusi in drugimi aleli. Z uporabo imunočipa, so v tej študiji
identificirali 14 genetskih lokusov za RA. Zanimivo je, da so SNP na genu PADI4, ki je
nedvomno povezan z RA v azijski populaciji v prejšnjih študijah, prav tako povezali tudi z
RA bolniki evropejskega porekla [48]. Tega jim v prejšnjih študijah ni uspelo. Prav tako so v
študiji leta 2013 [49] s pomočjo imunočipa pri Korejcih in Evropejcih identificirali dodatnih
8 novih genetskih lokusov za RA. Med njimi je tudi SNP rs909685.
Čeprav so GWA študije močno orodje za identifikacijo z boleznijo povezanih lokusov, ostaja
velik delež dejavnika dednosti neznanka. Pri RA odkritih 101 lokusov, ki so jih odkrili zunaj
HLA regije, razložijo samo 5,5 % in 4,7 % tveganja za nastanek bolezni pri evropejcih in
azijcih [47]. Analiza GWAS podatkov z uporabo Bayesian pristopa sklepanja je ocenila, da
100 do 1000 lokusov vsebuje skupne vzročne variante, ki skupaj z HLA-DRB1 in GWAS
identificiranih lokusov razložijo le približno 30 % tveganja za RA. To je približno polovica
dedovanja [50]. Preostali genetsi dejavnik veganja lahko razložimo z vplivom redkih variant,
katere frekvenca alela je < 1 % [51]. Redke variante pa se lahko analizirajo le s tehnologijo
sekvenciranja, ki zagotovijo večjo statistično moč.
2.7 Zdravljenje
V večini primerov je RA neozdravljiva bolezen. Le 5 % bolnikov doseže spontano remisijo
bolezni [52]. Obstoječa zdravljenja zato niso usmerjena k odstranjevanju vzrokov, temveč
zgolj blažijo vnetne reakcije in skušajo upočasniti napredovanje bolezni. Za uspešno terapijo

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
15
RA je bilo ključno spoznanje, da so erozije kosti in deformacije sklepov ireverzibilne, zato je
zgodnja diagnoza in čim hitrejši pričetek zdravljenja bistvenega pomena. Le na ta način lahko
preprečimo trajne poškodbe in uničenje sklepov. Cilj antirevmatskega zdravljenja je dvojen in
zajema remisijo simptomov bolezni (zmanjšanje bolečine v sklepih in zatekanje,
preprečevanje deformacij) ter spremembo poteka bolezni. Zdravila z nesteroidnimi
protivnetnimi učinkovinami (ang. non-steroidal anti-inflammatory drugs ali NSAIDs) se
uporabljajo za odpravljanje simptomov RA, kot so vnetje in bolečine v sklepih. Zdravljenje z
NSAIDs je simptomatsko, zaradi česar ni nobenega učinka na dolgoročne posledice bolezni.
Potek RA spreminjamo z imunomodulirajočimi protirevmatičnimi učinkovinami /zdravili
(ang. Disease modifying antirheumatic drugs ali DMARDs) in kortikosteroidi, katerih učinek
je večplasten [53].
Zaporedje ukrepov pri zdravljenju RA je naslednje [54]:
1. zgodnja uporaba DMARD (praviloma metotreksata),
2. redni pregledi in sistematično spremljanje terapevtskih učinkov,
3. hitro stopnjevanje terapije (višanje odmerkov in krajšanje odmernega intervala),
4. uvedba kombinirane terapije,
5. uvedba zdravljenja z biološkimi zdravili, če bolnik ni odziven na DMARD,
6. zamenjava biološkega zdravila, če bolnik ni odziven na prvega ali če pride do
znižane učinkovitosti prvega,
7. uvedba pomožnega zdravljenja s kortikosteroidi (praviloma časovno omejena).
Operativno zdravljenje je potrebno, kadar nastanejo nepopravljive okvare sklepov in
obsklepnih struktur. Dolgoročni rezultati so dobri. Ponovitev operativnega posega je
potrebna samo pri 4 do 13 % bolnikov [8].
Pri zdravljenju RA je pomembna tudi redna telesna aktivnost [55]. Le-ta mora biti prilagojena
potrebam in zmožnostim vsakega posameznika, zaradi česar so v proces zdravljenja vključeni
tudi fizioterapevti in delovni terapevti, ter psihologi, psihiatri in socialni delavci.
2.8 Farmakološko zdravljenje Kot je že omenjeno v poglavju 2.7, se zdravila, ki se uporabljajo pri zdravljenju RA, delijo v
naslednje skupine:
- zdravila z nesteroidnimi protivnetnimi učinkovinami (NSAIDS) in analgetiki,
- glukokortikoidi,
- imunomodulirajoče protirevmatične učinkovine (DMARDS)
- biološka imunomodulirajoča zdravila
2.8.1 Zdravila z nesteroidnimi protivnetnimi učinkovinami (NSAIDs) in analgetiki
NSAIDS delujejo zaviralno na encime iz skupine ciklooksigenaz (COX). Ti encimi so v telesu
odgovorni za sintezo prostaglandinov, ki imajo funkcijo mediatorjev vnetja, povzročijo dvig
telesne temperature in povečajo vzdraženost živčnih končičev. Zaviralci COX pa delujejo
protivnetno, protibolečinsko in protivročinsko. NSAIDS delimo na neselektivne in selektivne
(COX-2) zaviralce. Selektivni zaviralci COX-2 imajo manj neželenih učinkov na prebavila
[6]. V primerjavi s klasičnimi neselektivnimi NSAID pa povzročajo več neželenih učinkov na
srčno-žilnem sistemu [56]. V zadnjih letih se je izkazalo, da so vsi NSAIDS, razen naproksena
in deloma ibuprofena, odgovorni za neželene učinke na srčno-žilnem sistemu [57].

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
16
2.8.2 Glukokortikoidi
Kortikosteroidi se uporabljajo zaradi pleotropnih protivnetnih učinkov. Zaradi hitrega
delovanja hitro umirijo vnetje. Včasih je bila njihova uporaba precej razširjena. Danes pa se
jih uporablja s previdnostjo, saj ob dolgotrajnem zdravljenju povzročajo neželene stranske
učinke (osteoporoza, arterijska hipertenzija, steroidni diabetes, itn.) [55].
Zdravljenje RA z glukokortikoidi v kombinaciji z DMARD upočasni nastanek erozivnih
sprememb [58]. Najpogosteje se uporablja metilprednizolon.
Kombinacija NSAIDS in kortikosteroidov ni priporočljiva.
2.8.3 Imunomodulirajoče protirevmatične učinkovine (DMARDs)
DMARDs v kombinaciji z glukokortikoidi uporabljamo za vzdrževanje remisije. Ta zdravila
na različne načine zavirajo imunski sistem in so pri zdravljenju RA precej spremenila
prognoze te bolezni. Predstavljajo terapijo prvega izbora in jih je potrebno začeti uporabljati
v najkrajšem možnem času po postavitvi diagnoze. Zmanjšajo aktivnost bolezni, omejijo
okvaro sklepov in preprečujejo invalidnost [52].
Iz skupine DMARDs se uporabljajo metotreksat, leflunomid in sulfalsalazin. Druga zdravila
iz te skupine (antimalariki, azatioprin, ciklofosfamid, parenteralno zlato in ciklosporin A) se
zaradi učinkovitejših in varnejših alternativ pri zdravljenju RA le redko uporabljajo [52].
V skupino DMARDs se uvršča tudi novo zdravilo tofacitinib, ki zavira encim JAK3 kinazo.
Ameriška agencija za zdravila in prehrano (ang. FDA) je njegovo uporabo za zdravljenje RA
udobrila, medtem ko Evropska agencija za zdravila njegove uporabe ni udobrila. Trenutno je
namenjem za zdravljenje tistih bolnikov z RA, pri katerih zdravljenje s klasičnimi in
biološkimi NSAIDs ni bilo uspešno [52].
Metotreksat (MTX) je visoko učinkovit in varen, zato velja za zdravilo prvega izbora [59].
Glavni mehanizem delovanje je zaviranje encima dihidro folat reduktaza, ki je potreben v
sintezi DNK.
Najpogostejši neželeni učinki so slabost, bruhanje, driska, hematološki odkloni in toksičen
vpliv na jetra [60].
Zdravilo drugega izbora je leflunomid, na katerega preidemo ob neučinkovitosti ali
nezaželenih stranskih učinkov MTX.
Leflunomid uporabljamo v monoterapiji ali v kombinaciji z MTX. Zavira delovanje encima
dihidroorotatna dehidrogenaza in s tem zavira sintezo pirimidinskih nukleotidov in posledično
razrast limfocitov T.
Najpogostejši neželeni učinki so hujšanje, izpadanje las, kožne spremembe, driska, arterijska
hipertenzija, hematološki odkloni in toksični vpliv na jetra.
Sulfalsalazin se kot monoterapija pri zdravljenju RA le redko uporablja. Pogosteje se
uporablja v kombinaciji z MTX. Čeprav mehanizem delovanja ni povsem pojasnjen, se je
pokazalo zaviranje izločanja citokinov, izločanje makrofagov in apoptoze limfocitov T.
Najpogostejši neželeni učinki so slabost, bruhanje, zvišana telesna temperatura, kožni
izpuščaj, fotosenzitivnost in glavobol.
2.8.4 Biološka DMARDs
Biološka DMARDs se uporabljajo ob odpovedi ali neželenih učinkih standardnih zdravil. Na
različne načine zavirajo delovanje specifičnih citokinov ali celic. Tako zmanjšajo vnetje in
upočasnijo napredovanje bolezni. Zdravljenje z biološkimi zdravili poveča tveganje za okužbe

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
17
in aktivacijo latentne tuberkuloze ali virusnega hepatitisa. Ob zdravljenju je nujen reden
laboratorijski nadzor.
Zaviralci TNF-alfa. TNF-alfa je eden izmed glavnih provnetnih citokinov. Zaviranje
njegovega delovanja ima protivnetni učinek. V to skupino uvrščamo monoklinska protitelesa
(adalimumab, certolizumab pegol, infliksimab, golimumab) in TNF-alfa receptor, vezan na Fc
del IgG1-etanercept, ki specifično vežejo TNF-alfa in preprečijo vezavo na njegove receptorje.
Uporabljajo se v kombinaciji z MTX ali leflunomidom.
Tocilizumab je monoklinsko protitelo, ki je usmerjeno proti receptorju za IL-6. IL-6 je
provnetni citokin s širokim spektrom delovanja. Je edino biološko zrdavilo, ki je pokazalo
podobno učinkovitost v monoterapiji kot v kombinaciji z MTX pri zdravljenju RA [59].
Kontraindikacija za zdravljenje s tocilizumabom je divertikuloza črevesja.
Rituksimab je monoklinsko protitelo, ki je usmerjeno proti antigenu CD20. Antigen CD20 se
nahaja na površini nezrelih limfocitov B. Rituksimab po vezavi povzroči apoptozo tarčne
celice. Uporablja se v kombinaciji z MTX ali leflunomidom.
Anakinra je rekombinantna beljakovina, ki deluje kot antagonist receptorja IL-1. Receptor
IL-1 spodbuja vnetje z indukcijo IL-6 in COX. Pri zdravljenju RA se redko uporablja.
2.9 Genotipizacija Genotip je genska zasnova organizma. Nanaša se lahko na kompleten genom ali na posamezen
odsek DNK, ki je lahko gen ali pa posamezen SNP. Genotip SNP-ja je tako lahko:
- ˝wild type˝, ki je homozigot za normalen alel,
- homozigot za mutiran alel in
- heterozigot, ki vsebuje oba alela.
Genotipizacija je proces ugotavljanja genotipa posameznika. Pri vsakem posamezniku se
lahko ugotavlja genotip enega ali večih genov/SNP-jev. SNP markerji se uporabljajo pri
genotipizaciji bolezni ali določanju pripadnosti določeni etnični skupini. Pojavljanje SNP-jev
je povezano s pojavljanjem določenega gena. Pri določanju verjetnosti za razvoj kompleksnih
bolezni ugotavljamo, kakšen genotip ima za določene markerje posameznik [61]. Za gensko
tipizacijo polimorfizmov se uporabljajo različne metode, npr. verižna reakcija s polimerazo
(PCR), polimorfizem dolžin restrikcijskih fragmentov (RFLP), analiza talilne krivulje visoke
ločljivosti (HRM), itd.
2.10 Opis SNP-ov uporabljenih v nalogi
2.10.1 rs1571878 (CCR6)
CCR6 ali kemokinski (C-C) receptor 6 se nahaja na dolgi ročici q kromosoma 6 na položaju
27 (6q27). Položaj gena prikazuje slika 2-5.
Gen kodira člana iz družine beta kemokinskih receptorjev. CCR6 je izražen iz nedozorelih
dendritskih celic in spominskih T celic. Ligand teh receptorjev je makrofag vnetni protein 3
alfa (MIP-3 alfa). Ta receptor je pomemben za B linijsko dozorelost in diferenciacijo antigen
B celice, ter regulacijo migracije dentritskih in T celic med vnetnim in imunskim odzivom
[62].
CCR6 povezujejo z RA, Chronovo boleznijo in Basedovo boleznijo [22].

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
18
Slika 2-5: Položaj gena CCR6 [62].
2.10.2 rs7574865 (STAT4)
Signalni pretvornik in aktivator transkripcije 4 oz. STAT4 spada v družino genov, ki vsebujejo
SH2 domene. Gen je lociran na daljši ročici kromosoma 2 med položajema 32.2 in 32.3
(2q32.2-q32.3), kar prikazuje slika 2-6 [63].
STAT4 je član družine sedmih transkripcijskih faktorjev, kar pomeni, da se veže na specifično
mesto na DNK in pomaga kontrolirati aktivnost določenih genov. STAT4 aktivirajo proteine
imunskega sistema (citokine), kateri pomagajo pri boju z okužbo. Ko je gen aktiviran, ta zviša
aktivnost genov, ki pomagajo T celicam dozoreti v specializirane T celice. Specializirane T-
celice se imenujejo Th celice, ki proizvedejo specifične citokine in stimulirajo druge imunske
celice za odstranitev patogena iz celice [64].
STAT4 je povezan s številnimi avtoimunskimi boleznimi, kot so RA, sistemski lupus
eritematozus [65], diabetes I [66] in Sjögren sindrom [67].
Slika 2-6: Položaj gena STAT4. [63]

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
19
2.10.3 rs909685 (SYNGR1)
Synaptogyrin 1 je protein in je pri ljudeh kodiran z genom SYNGR1 [68]. Lociran je na daljši
ročici kromosoma 22 na položaju 13.1 (22q13.1). Lego gena SYNGR1 prikazuje slika 2-7.
Sestavljen je iz šestih eksonov. Gen kodira sestavni del membrane proteina povezanega s
presinaptičnim veziklom v nevronski celici. Točna funkcija proteina ni znana, vendar študije
podobnega proteina murina kažejo na funkcijo v sinaptični plastičnosti brez zahteve po
sinaptičnem prenosu [69]. SYNGR1 gen povezujejo s shizofrenijo [70], [71] in bipolarno
motnjo [71].
SNP rs909685 se nahaja na genu SYNGR1. Z GWA študijo [49] so ga povezali s pojavom RA.
Slika 2-7: Položaj gena SYNGR1 [72].
2.10.4 rs2476601 (PTPN22)
Gen PTPN22 je lociran na krajši ročici (p) kromosoma 1, položaj 13.2 (1p13.2). Položaj
PTPN22 prikazuje slika 2-8. Sestavlja ga 24 eksonov [73].
PTPN22 kodira nereceptorski protein tirozin fosfatazo 22 (LYP), ki je izražen v limfnih tkivih.
Mutacija rs2476601, ki se nahaja na genu PTPN22, spremeni aminokislinsko zaporedje na
mestu 620 iz arginina v triptofan. Fizična povezava med LYP in proteina tirozin kinaza CSK
(ang. C terminal SRC kinase) zavira LYP aktivnost. SNP rs 2476601 zmoti vezavo LYP na
CSK in sproži odzivnost B in T celičnih receptorjev [1]. Le-ta SNP je povezan s pojavom več
kot 20 različnih avtoimunskih bolezni v evropski populaciji, ki vključuje RA, diabetes tipa 1,
Graves-ovo bolezen in multiplo sklerozo [74].
Slika 2-8: Položaj gena PTPN22 [73].

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
20
2.10.5 rs3087243 (CTLA4)
Citotoksik T-limfocit-povezan protein 4 (CTLA4) je gen, ki kodira protein. Nahaja se na daljši
roki kromosoma 2, položaj 33.3 (2q33.3). Položaj CTLA4 vidimo na sliki 2-9.
Gen je član super družine imunoglobulinov in kodira protein za transmisijo zaviralnih signalov
do T celic. Protein vsebuje V domeno, transmembransko domeno in citoplazmitski rep.
Mutacije tega gena so povezali z inzulin-odvisnim diabetesom, Graves-ovo boleznijo,
Hašimoto tiroidizmom, celiakijo, sistematičnim lupos eritematozisom, RA in drugimi
avtoimunskimi boleznimi [75].
Slika 2-9: Položaj gena CTLA4 [75].

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
21
3 Eksperimentalni del
3.1 Vzorci DNK Laboratorijske raziskave so bile izvedene na 276 vzorcih DNK, kateri so bili predhodno
izolirani iz periferne krvi, natančneje iz krvnih limfocitov iz vzorcev bolnikov diagnosticiranih
z RA ter na 276 vzorcih DNK, kateri so bili predhodno izolirani iz vzorcev zdravih
posameznikov.
3.2 Materiali
3.2.1 Osnovne kemikalije
Pri naši nalogi smo uporabili kemikalije, ki so predstavljene v spodnji tabeli 3-1.
Tabela 3-1: Osnovne kemikalije in njihovi proizvajalci.
Kemikalija Proizvajalec
Agaroza SeaKem
dNTP-ji Sigma Aldrich
EDTA Sigma Aldrich
Etanol absolut Merck
Etidijev bromid Sigma Aldrich
Ficoll-Paque Plus GE Healthcare
Izopropanol Sigma Aldrich
Kloroform Sigma Aldrich
Ksilen cianol Sigma Aldrich
Light Cycler Master Mix Roche
Magnezijev klorid (HRM) Roche
Magnezijev klorid (PCR) Fermentas
PBS pufer Sigma Aldrich
PCR pufer Sigma Aldrich
Pufri za restrikcijske encime Fermentas
Restrikcijski encimi Fermentas
Saharoza Merck
TAQ polimeraza Sigma Aldrich
TRI reagent Sigma Aldrich
TRIS baza Calbiochem
Začetni oligonukleotidi Sigma Aldrich

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
22
Za RFLP in HRM smo uporabljali začetne oligonukleotide, ki so navedeni v tabeli 3-2.
Začetne oligonukleotide smo ročno oblikovali s pomočjo spletnega programa Primer3
(http://bioinfo.ut.ee/primer3-0.4.0/).
Tabela 3-2: Izbrani začetni oligonukleotidi (primerji).
Gen SNP ID Oznaka Zaporedje 5´-3´
CCR6 rs1571878 CCR6_rs1571878hrm(1) CTCAAGTGATCCACCCACCT
CCR6_rs1571878hrm(2) AGCCTGGGAACCCCTATCTA
SYNGR1 rs909685 SYNGR1_rs909685(1) ATCTGTGCCAGTCTCCTGCT
SYNGR1_rs909685(2) GGCCCAGCAGATTCAATAAA
STAT4 rs7574865 STAT4_rs7574865hrm(1) TGGAAACTGAAGGGATTGCT
STAT4_rs7574865hrm(2) CCTCATGCCCACAGTTGAC
PTPN22 rs2476601 PTPN22_rs2476601(1) CCAGCTTCCTCAACCACAAT
PTPN22_rs2476601(2) GGATAGCAACTGCTCCAAGG
CTLA4 rs3087243 CTLA4_rs3087243(1) CACCACTATTTGGGATATACC
CTLA4_rs3087243(2) AGGTCTATATTTCAGGAAGGC
3.2.2 CEPH vzorci
Pri eksperimentalnem delu smo pri genotipizacijah kot referenčne vzorce uporabili CEPH
vzorce. Fundacija Jean Dausset – Centre d'Etude du Polymorphisme Humain (CEPH) oziroma
Center za študijo človeških polimorfizmov vsebuje podatkovno bazo genotipov za genetske
markerje. Le-ti so bili genotipizirani na CEPH referenčnih družinah. Iz rakastih celic in celic
posameznikov so v fundaciji razvili nesmrtne celične linije, ki so vir DNK za genetske študije
po vsem svetu [76].
3.2.3 Raztopine in pufri
Vse raztopine in pufri v našem delu so bile pripravljene s posebno čisto destilirano vodo. Vodo
je destilirala naprava TKA PacificUP-UPW water purification system.
3.2.3.1 TBE pufer
TBE pufer smo pripravili kot založno raztopino z 10 x koncentracijo. Sestava 10 x TBE pufra
je iz:
- c = 445 mM borove kisline,
- c = 445 mM TRIS pufra in
- c = 5 mM EDTA.
Za pripravo V = 1 L 10 x TBE pufra smo zatehtali m = 55 g borove kisline, m = 108 g TRIS
baze in m = 3,72 EDTA. Vse skupaj smo raztopili v V = 1 L vode.
Za pripravo 1 x pufra smo V = 100 mL 10 x TBE pufra dodali v V = 900 mL destilirane vode.

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
23
3.2.3.2 PBS pufer
Pri pripravi PBS pufra smo uporabili PBS v obliki tablet. V V = 200 mL destilirane smo
raztopili 1 tableto PBS. Raztopino smo nato sterilizirali v avtoklavu s programom za
sterilizacijo raztopin.
3.2.3.3 Nanašalni pufer za gelsko elektroforezo
Nanašalni pufer oziroma barvilo za gelsko elektroforezo smo pripravili tako, da smo zatehtali
m = 40 g saharoze in m = 0,25 g ksilen cianola. Vse skupaj smo raztopili v V = 100 mL 1 x
TBE pufra.
3.2.4 Laboratorijska oprema
V nalogi smo uporabili laboratorijsko opremo, ki je navedena v tabeli 3-3.
Tabela 3-3: Laboratorijska oprema in njeni proizvajalci
Oprema Proizvajalec
Centrifuge Biosan CMC-3000
Eppendorf centrifuge 5415R
Ciklični termostat Biometra T1 Thermocycler
Biometra Professional basic gradient
PCR v realnem času Light Cycler LC480
Mikropipete Finnpipette 10 µL, 100µL, 1000 µL
Mikrovalovna pečica Gorenje
Spektrofotometer BioTek Synergy2 Plate Reader
UV transiluminator Gel imaging system GBOX
3.3 Laboratorijske metode Poleg pregleda znanstvenih člankov smo v nalogi uporabili tudi različne laboratorijske metode
in tehnike, ki se uporabljajo pri molekularni biologiji. Uporabili smo naslednje laboratorijske
metode:
Izolacija DNK, RNK in proteinov
Merjenje koncentracij in redčenje DNK
Verižna reakcija z encimom polimeraza (PCR)
Polimorfizem dolžin restrikcijskih fragmentov (RFLP)
PCR v realnem času (qPCR) in metoda analize talilne krivulje visoke ločljivosti (HRM)
3.3.1 Izolacija DNK, RNK in proteinov
3.3.1.1 Zbiranje limfocitov
Z izolacijo limfocitov smo pričeli s pripravo vzorca. Kri z V = 12 mL, ki je bila odvzeta v
epruveto z EDTA smo prenesli v novo sterilno V = 50 mL centrifugirko ter dodali V = 12 mL
pufra PBS (skupen volumen 24 mL). Raztopino smo dobro premešali. V novo sterilno V = 50
mL centrifugirko smo nalili V = 12 mL Ficola. Na Ficol smo previdno in počasi ob steni

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
24
nalivali celotno suspenzijo krvi in PBS-a. Paziti smo morali, da se raztopina ni pomešala s
Ficolom, kot prikazuje spodnja slika 3-1.
Slika 3-1: Mešanica krvi in pufra PBS nanešenega na Ficol.
Centrifugirko smo pri f = 2500 rpm, t = 20 min in T = 18 °C centrifugirali. Po centrifugiranju
smo dobili vzorec, razdeljen na štiri faze, ki so vidne na sliki 3-2: eritrocite, Ficol, limfocite
in krvno plazmo.
Slika 3-2: Krvni vzorec, ki je po centrifugiranje razdeljen na štiri faze.
Krvno plazmo smo prenesli v sterilno V= 50 mL centrifugirko in jo shranili na T = - 20 °C.
Drugo fazo z limfociti smo s pipeto prenesli v novo V = 50 mL sterilno centrifugirko, katerim
smo nato dodali 3 volumne pufra PBS. Raztopino smo nato centrifugirali pri f = 2800 rpm in
t = 10 min. Po koncu centrifugiranja smo supernatant odsesali. Limfocitom smo dodali V = 10
mL PBS-a, zvorteksirali in ponovno centrifugirali pri f = 2800 rpm in t = 10 min. Supernatant
smo nato s pomočjo pipete v celoti odsesali in limfocite, ki so ostali, shranili v zmrzovalnik
pri T = 70 °C do njihove nadalnje uporabe.
Prav tako smo odstrabili Ficol nad plastjo eritrocitov in eritrocite shranili na – 20 °C.
3.3.1.2 Izolacija RNA s TRI reagentom
Zbrane limfocite smo odmrznili na sobni temperaturi in jim dodali V = 1500 µL TRI reagenta.
Celice smo razbili s pipetiranjem tako, da ni bilo več vidnih večjih koščkov. Vse skupaj smo
nato iz centrifugirk prenesli v V = 2 mL mikrocentrifugirke in inkubirali t = 5 min na sobni
temperaturi. Mešanici smo dodali V = 300 mL kloroforma, zaprli mikrocentrifugirke in
Kri pomešana s
pufrom PBS
Ficol
Krvna plazma
Limfociti
Ficol
Eritrociti

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
25
vorteksirali t = 15 s, da je raztopina postala motno bela. Vzorce smo inkubirali na sobni
temperaturi t = 2 -15 min. Po inkubaciji smo mešanico centrifugirali pri f = 12 000 x g (rcf), t
= 15 min pri T = 4 °C, da se loči v tri faze:
- spodnja rdeča – proteini,
- vmesna bela – DNK,
- zgornja prozorna – vodna raztopina RNK.
Vodno fazo smo prenesli v svežo V = 1,5 mL mikrocentrifugirko, preostanek pa smo uporabili
za izolacijo DNK in proteinov, kateri postopek je opisan v nadaljevanju tega dela. Vodni fazi
smo dodali V = 700 µL izopropanola in premešali z obračanjem mikrocentrifugirke. Vzorec
smo inkubirali na sobni temperaturi t = 5 – 10 min. Po inkubaciji smo vzorec centrifugirali pri
f = 12 000 x g (rcf), t = 10 min in T 4 °C. Na dnu mikrocentrifugirke nastane viden pelet.
Supernatant smo odlili in dodali V = 1500 µL 75 % etanola. Mešanico smo dobro premešali z
obračanjem in nato ponovno centrifugirali na f = 12 000 x g (rcf), t = 5 min pri T = 4 °C.
Supernatant smo odlili in pelet, ki je ostal na dnu mikrocentrifugirke smo posušili na zraku t
= 5 – 10 min. Vzorcu smo dodali V = 60 µL RNase free water in ga shranili pri T = - 70 °C.
3.3.1.3 Izolacija DNK
Po odstranitvi vodne faze za izolacijo RNK, smo DNK precipitirali iz vmesne faze in
organeske faze tako, da smo dodali V = 450 µL 100 % etanola. Mikrocentrifugirko smo nato
zvorteksirali in pustili stati na sobni temperaturi t = 2 – 3 min. Nato smo mešanico
centrifugirali pri f = 5 000 x g (rcf), t = 5 min in T = 4 °C. Po centrifugiranju smo odstranili
supernatant in ga shranili pri T = 4 °C za izolacijo proteinov. Pelet DNK smo dvakrat sprali v
V = 1500 µL, c = 0,1 M natrijevega citrata/10 % etanolu in dobro zvorteksirali. Med vsakim
spiranjem smo pustili pelet DNK stati z občasnim mešanjem t = 30 min. Sledilo je
centrifugiranje pri f = 5000 x g (rcf) t = 5 min pri T = 4 °C. Supernatant smo z odlivanjem
zavrzili. Pelet DNK smo resuspendirali v V= 1,0 mL 75 % etanola in pustili stati t = 10 – 20
min pri sobni temperaturi. Vzorce smo lahko na tej stopnji že shranili na – 20 °C. Lahko pa
smo jih ponovno centrifugirali pri f = 5000 x g (rcf), t = 5 min in T = 4 °C in supernatant smo
po koncu centrifugiranja ponovno zavrgli z odlivanjem. Pelet DNK smo nato posušili na zraku
in ga raztopili v V = 400 µL vode (ponavljajoče pipetiranje, uporabi konice s filtrom).
Raztopljeno DNK smo shranili na T = 4 °C v hladilnik.
3.3.1.4 Izolacija proteinov
Proteine smo iz fenol-etanolnega supernatanta precipitirali tako, da smo dodali V = 2250 µL
izopropanola. Vzorce smo pustili stati najmanj t = 10 min pri sobni temperaturi in jih nato
centrifugirali pri f = 12 000 x g (rcp), t = 10 min in T = 4 °C. Supernatant, kot zgornjo fazo
smo zavrgli, pelet pa smo sprali v V = 1 mL, c = 0,3 M gvanidijevega hidroklorida/95 %
etanolu. Spiranje smo ponovili dvakrat. Med vsakim spiranjem pa smo mešanico pustili stati
t = 20 min pri sobni temperaturi in nato centrifugirali pri f = 7 600 x g (rcf), t = 5 min pri 4 °C.
Nastali supernatant smo zavrgli. Po treh spiranjih smo dodali V = 1 mL 100 % etanola in
premešali z vorteksiranjem. Do nadaljne uporabe smo proteine shranili v 100 % etanolu na T
= - 20 °C.
3.3.2 Merjenje koncentracij in redčenje DNK
Koncentracija DNK je za metodo HRM zelo pomembna. Izoliranim vzorcem DNK smo
izmerili koncentracije s spektrofotometrom BioTek Synergy2 Plate Reader. DNK smo nato
razredčili do koncentracije c = 15 ng/µL. Za PCR-RFLP in HRM smo uporabili 2 µL tako
razredčene koncentracije (30 ng).

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
26
Volumen vzorca in vode za izračun želene koncentracije vzorca smo izračunali po enačbah
3.1 in 3.2:
𝑐 ∗ 𝑣 = 𝑐1 ∗ 𝑣1 + 𝑐2 ∗ 𝑣2 (3.1)
𝑣 = 𝑣1 + 𝑣2 (3.2)
Kjer je
c = končna koncentracija (15 ng/µL)
v = končni volumen
c1 = koncentracija raztopine DNK
v1 = volumen raztopine DNK, ki ga moramo dodati, da dobimo željen končni volumen
c2 = koncentracija DNK v vodi, ki je enaka 0
v2 = volumen vode, ki ga moramo dodati, da dobimo želeni končni volumen z želeno
koncentracijo
3.3.3 Verižna reakcija z encimom polimerazo (PCR)
Verižna reakcija s polimerazo (ang. PCR za Polymerase Chain Reaction) je metoda, ki
omogoča kopiranje odsekov DNK s pomočjo termično obstojnega encima DNK polimeraze.
Metodo je leta 1983 zasnoval ameriški biokemik Kary Mullis in je zanjo tudi dobil Nobelovo
nagrado za kemijo leta 1993. Je enostavna, hitra in cenovno ugodnejša od drugih metod, ki se
uporabljajo za pomnoževanje DNK. PCR metoda posnema in vivo pomnoževanje DNK.
Uporabljajo jo pri diagnostičnih postopkih molekularne medicine in forenzični znanosti [77],
[78].
Za PCR reakcijo potrebujmo naslednje osnovne komponente:
- dve kratki molekuli DNK, ki sta komplementarni začetnemu in končnemu delu
segmenta, ki ga želimo kopirati,
- dva sintetična začetna oligonukleotida, dolga med 18 in 25 nukleotidov, ki sta
komplementarna sosednjima zaporedjema,
- termostabilno DNK polimerazo (Taq DNK polimeraza), ki kopira regije,
- vse štiri deoksiribonukleozid trifosfate oz. dNTP (dATP, dGTP, dCTP, dTTP), za
gradnjo nove DNK,
- MgCl2,
- pufer, ki poskrbi za primerno kemijsko okolje v katerem encim DNK polimeraza
optimalno deluje.
PCR se izvaja v cikličnem termostatu. To je naprava, ki segreva in hladi reakcijske posodice
do določene temperature, ki je potrebna za vsak korak reakcije. PCR vsebuje serije od 20 do
35 ciklov. Vsak cikel je sestavljen iz treh stopenj, ki jih predstavlja slika 3-3:
1. Denaturacija. Matrično DNK segrejemo do T = 93 – 95 °C za t = 15 – 30 s. Tako
razbijemo vodikove vezi in ločimo verigi matrične DNK. Dobimo enoverižno DNK.
2. Pripenjanje začetnih oligonukleotidov. Temperaturo znižamo do T = 37 – 55°C za t =
15 – 30 s, kar omogoči pripenjanje začetnih oligonukleotidov (primer-jev) na ustrezne
komplementarne regije. Za uspešno pripenjanje oligonukleotidov in preprečitev
ponovnega prileganja matričnih verig DNK se v reakcijsko mešanico doda prebitek
začetnih nukleotidov [78]. Prav tako pa je ključnega pomena izbira temperature.
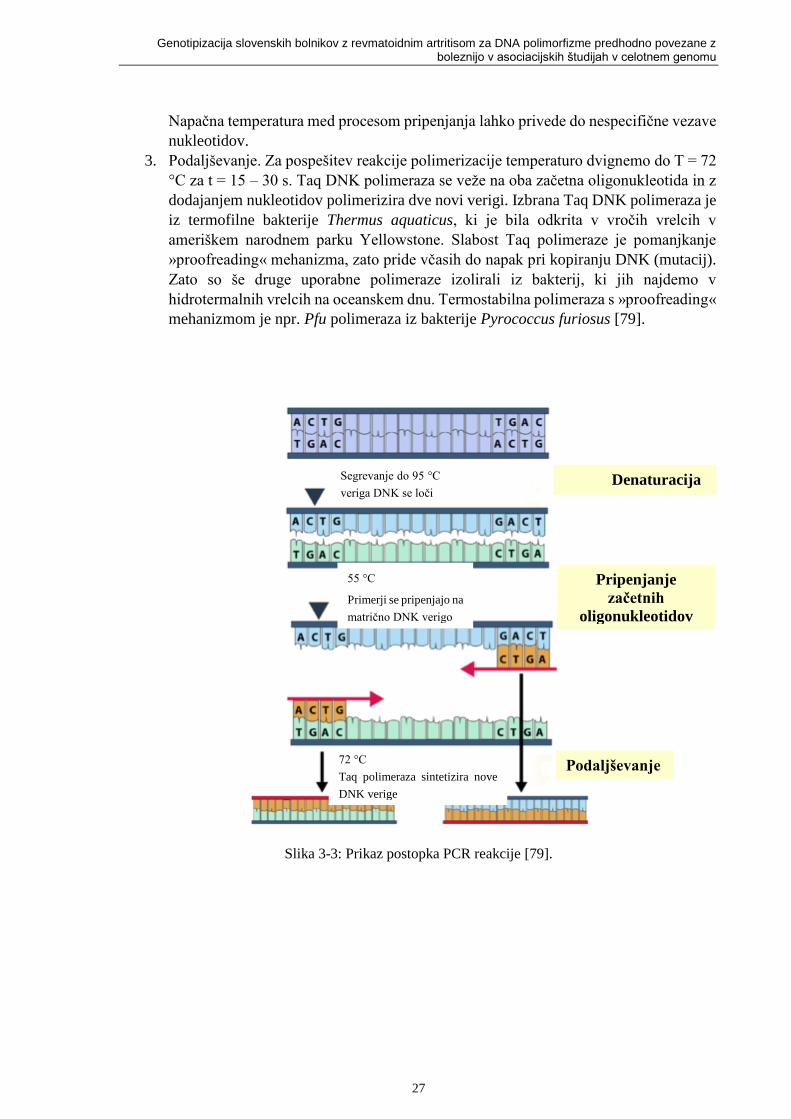
Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
27
Napačna temperatura med procesom pripenjanja lahko privede do nespecifične vezave
nukleotidov.
3. Podaljševanje. Za pospešitev reakcije polimerizacije temperaturo dvignemo do T = 72
°C za t = 15 – 30 s. Taq DNK polimeraza se veže na oba začetna oligonukleotida in z
dodajanjem nukleotidov polimerizira dve novi verigi. Izbrana Taq DNK polimeraza je
iz termofilne bakterije Thermus aquaticus, ki je bila odkrita v vročih vrelcih v
ameriškem narodnem parku Yellowstone. Slabost Taq polimeraze je pomanjkanje
»proofreading« mehanizma, zato pride včasih do napak pri kopiranju DNK (mutacij).
Zato so še druge uporabne polimeraze izolirali iz bakterij, ki jih najdemo v
hidrotermalnih vrelcih na oceanskem dnu. Termostabilna polimeraza s »proofreading«
mehanizmom je npr. Pfu polimeraza iz bakterije Pyrococcus furiosus [79].
Slika 3-3: Prikaz postopka PCR reakcije [79].
Denaturacija Segrevanje do 95 °C
veriga DNK se loči
Pripenjanje
začetnih
oligonukleotidov
55 °C
Primerji se pripenjajo na
matrično DNK verigo
Podaljševanje 72 °C
Taq polimeraza sintetizira nove
DNK verige

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
28
Koncentracija tarčne DNK narašča eksponentno (Slika 3-4), saj vsaka na novo sintetizirana
veriga služi kot matrica. Po 20 ponovitvah reakcije dobimo iz 1 kopije DNK teoretično več
kot 1 milijon kopij [80].
Slika 3-4: Prikaz eksponentne rasti tarčne DNK [81].
Števila ciklov ni smiselno povečevati v neskončnost, saj običajno pri 35 do 45 ciklov pride do
t.i. efekta PCR platoja. Pri efektu platoja novi produkti ne nastajajo več. Vzrok za efekt platoja
je prebitek pridelkov PCR, ki izpodrinejo pri prilepljanju primerje in se kompetitivno vežejo
nazaj na drugo verigo DNK [77].
PCR produkte preverimo in analiziramo z gelsko elektroforezo. S pomočjo velikostnega
markerja ocenimo velikost nastalih pridelkov. Za detekcijo mutacij na DNK sekvenci
uporabimo metodo verižna reakcija s polimerazo, polimorfizem dolžin restrikcijskih
fragmentov (RFLP), metodo analize talilne krivulje visoke ločljivosti (HRM), TaqMan
metodo genotipiziacije [82].
Poznamo več vrst PCR metod, npr. »touch down« PCR, »hot start« PCR, »nested« PCR, Alu-
PCR, inverzni CPR, Real time PCR, itd. [83].
Reakcije smo izvedli po protokolu, ki je naveden v tabeli 3-4.
Tabela 3-4: Prikaz protokala za PCR reakcijo.
Reagenti Volumen za 1 vozrec (µL)
DNK 2
H2O a
pufer + MgCl2 1
dNTP-ji 0,2
primer 1 b
primer 2 b
TAQ polimeraza 0,05
tarča
Teoretično,
neskočna
eksponenčna
amplifikacija Matrična DNK
1 cikel
2 cikel
3 cikel 4 cikel
4 kopije 8 kopij 16 kopij 32 kopij

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
29
Parametra v tabeli 3-4 označena z a in b sta odvisna od vrste para začetnih oligonukleotidov.
S procesom optimizacije smo določili parameter b oziroma njegov volumen. Volumen a smo
izračunali naknadno.
Vse reagente smo dodali v mikrocentrifugirko, jih premešali na vorteksu in jih na kratko
centrifugirali. Mešanico smo nanesli na stripe ali ploščice, na katere smo predhodno
odpipetirali V = 2 µL DNK vzorca. Ploščico smo nato dali v termociklizator, kjer smo izbrali
ustrezen protokol glede na začetne oligonukleotide.
Najpogosteje izbrani protokol prikazuje tabela 3-5.
Tabela 3-5: Izbrani protokol, ki je uporabljen v termociklizatorju.
Začetna denaturacija T = 94 – 96 °C, t = 5 min
Denaturacija T = 94 – 96 °C, t = 30 – 60 s
Pripenjanje začetnih oligonukleotidov T = X, t = 20 – 30 s
Podaljševanje T = 72 °C, t = 30 s
Število ciklov 35 – 40 ciklov
Končno podaljševanje T = 72 °C, t = 7 min
Parameter X smo določili z optimizacijo.
3.3.4 Priprava 2 % agaroznega gela in elektroforeza
Agarozni gel smo pripravili tako, da smo v erlenmajerici zatehtali m = 0,8 g agaroze in dodali
40 mL 1 x TBE pufra. Mešanico smo segrevali v mikrovalovni pečici tako dolgo, dokler se
agaroza ni popolnoma raztopila. Nato smo dodali V = 3 µL etidijevega bromida, ki se vgrajuje
med bazne pare DNK in fluorescira pod UV svetlobo ( λ = 365 nm). Raztopino smo nalili na
nosilec z glavnički, katerih glavna naloga je narediti žepke v gel. Po strditvi gela smo
glavničke odstranili in gel potopili v banjico za elektroforezo, napolnjeno z 1 x TBE pufrom.
V = 10 µL PCR produkta smo dodali V = 5 µL barvila (ksilencianol) in vse skupaj nanesli na
dno žepka. Na gel smo tudi dali velikostni marker, tj. mešanica več DNK produktov znanih
dolžin, ki služijo kot standard, da lahko ocenimo velikost nastalih PCR produktov. Banjico z
gelom priključimo na konstantno napetost U = 150 V in pustimo teči t = 10 – 15 min. Molekula
DNK ima negative naboj, zato bo v elektroforeznem sistemu potovala skozi agarozni gel v
smeri pozitivno nabite elektrode. Večje molekule DNK so težje in potujejo počasneje v
primerjavi z manjšimi.
3.3.5 Polimorfizem dolžin restrikcijskih fragmentov (RFLP)
Pojem polimorfizem dolžin restrikcijskih fragmentov (ang. RFLP za restriction fragment
lenght polymorphism) se nanaša na različno dolge fragmente DNK, ki nastanejo po razrezu
DNK z restrikcijskim encimom (RE) ter na laboratorijsko metodo, ki se uporablja za ločevanje
in analizo teh fragmentov. Je ena izmed prvih metod, ki so jih uporabili v forenziki. V kliničnih
študijah pa se uporablja na področjih študije DNK polimorfizmov in študije mutacij tumorjev
[82].
V preteklosti se je za tipizacijo RFLP-jev uporabljala hibridizacija (Southern blotting) in z
radioaktivnimi sondami. Hibridizacijo je zamenjala PCR metoda saj je preprostejša, hitra in
ne zahteva velikih količin DNK. S PCR metodo pomnožimo željen odsek DNK, ki vsebuje

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
30
SNP, ki ga preiskujemo. PCR produktom se doda RE, ki specifično reže produkt. Fragmente
različnih velikosti ločimo z gelsko elektroforezo in na podlagi velikosti fragmentov določimo
genotip [82].
3.3.5.1 Restrikcijski encimi
Restrikcijski encimi, restrikcijske endonukleaze ali restriktaze so najbolj specifični bakterijski
encimi, ki režejo dvoverižne DNK molekule. Restrikcijske encime proizvajajo bakterije, ki z
njimi razgrajujejo tujo DNK bakteriofagov. RE prepoznajo specifično zaporedje nukleotidov
v DNK molekuli. Znotraj prepoznavnega zaporedja RE režejo DNK molekulo na fragmente
točno definirane dolžine, ti. restrikcijske fragmente. Uporabna vrednost RE je v tem, da so
sposobni rezati DNK molekulo na natančen in ponovljiv način [83].
Odkritje RE je švicarju Werner Arberju prineslo Nobelovo nagrado leta 1978. Delijo se na tri
glavne skupine:
- vrsta I,
- vrsta II in
- vrsta III.
Najbolj se uporablja vrsta II saj sta drugi dve skupini precej bolj kompleksni [84].
Izoliranih in preiskanih je bilo že več sto RE. Za raziskave jih uporabljajo približno 200. Vsaka
vrsta bakterij proizvede različne RE, ki prepoznajo različno zaporedje v DNK. Imena RE so
sestavljena iz treh črk bakterije iz katerih so bili izolirani, oznako seva in rimske številke, ki
označuje, kateri po vrsti je bil encim odkrit. Prvi odkriti RE, poimenovani Eco RI, so izolirali
iz seva R bakterij E. coli [83].
Večina RE prepoznajo specifična štiri, šest ali osem baznih parov dolga zaporedja nukleotidov
v DNK. Encimi se vežejo na določeno mesto v DNK in režejo dvoverižno DNK molekulo.
Prepoznano zaporedje nukleotidov, ki ga RE reže se imenuje mesto restrikcije. Številni RE
režejo prepoznavno zaporedje simetrično (na istem mestu na obeh verigah DNK). Nekateri
encimi pa režejo prepoznavna zaporedja asimetrično (pri različnih nukleotidih), da dobimo
DNK fragmente, ki imajo kratke enoverižne konce. Te konce imenujemo »lepljivi konci« [83].
Na sliki 3-5 je prikazan razrez DNK s tremi različnimi encimi. Restrikcijska encima EcoRI in
HindIII režeta DNK asimetrično, medtem ko AluI reže DNK simetrično.
Slika 3-5: Primer razreza treh restrikcijskih encimov [83].
EcoRI
(Escherichia coli)
HindIII
(Haemophilus influenzae)
AluI
(Arthrobacter luteus)
Tarčna sekvenca
in mesto razreza;
»lepljivi konci«
Tarčna sekvenca
in mesto razreza;
»topi konci«
Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
31
PCR produktu z V = 10 µL smo dodali V = 20 µL reakcijske mešanice. Reakcijska mešanica
za RFLP vsebuje vodo, restrikcijski encim in restrikcijski pufer. Reakcijsko mešanico dobro
premešamo in dodamo PCR produktu. Vse skupaj nato premešamo na vorteksu in
centrifugiramo. Nato inkubiramo na T = 37 °C za t = 15 min.
Parametri, ki smo jih uporabili za restrikcijsko mešanico, so prikazani v tabeli 3-6.
Tabela 3-6: Parametri uporabljeni pri RFLP metodi.
Sestavina Količina za 1 vzorec (µL)
PCR produkt 10
Restrikcijski encim a
10 x pufer za restrikcijski encim 3
dH2O b
Parameter a smo določili z optimizacijo restrikcijskega encima. Parameter b smo izračunali
naknadno.
3.3.6 Metoda HRM
Analiza talilne krivulje visoke ločljivosti (ang. HRM za High Resolution Melting) je preprosto
orodje, ki hitro in učinkovito odkrije genetske spremembe (SNP, mutacije, metilacije) in ne
potrebuje označenih sond in procesiranja ali separacije po PCR (High Resolution Melting
Applications for Clinical Laboratory Medicine). HRM je bila predstavljena leta 2003 v
sodelovanju med Univerzo v Utahu (University of Utah) in podjetjem (Idaho Technology, Inc
(Salt Lake City, UT)). Zaradi preprostosti in hitrosti se metoda vedno bolj uporablja v kliničnih
laboratorijih [84].
HRM metoda se nanaša na taljenje DNK v prisotnosti nasičenih DNK barvil. V preteklosti je
bila toplotna talitev dvojne vijačnice DNK nadzorovana z UV absorbanco (hiperkromični
efekt). Analiza je zahtevala µg količine DNK in veliko časa, saj se je temperatura višala za
0,1-1,0 °C na minuto. Sodobna DNK analiza taljenja uporablja fluorescenco in potrebuje le
ng količine DNK. Analiza taljenja DNK z fluorescenco je postala popularna leta 1997 z
napredkom PCR v realnem času in prihodom inštrumenta LightCycler®. Majhne količine
vzorca so pripomogle k boljši kontroli temperature in hitremu taljenju za 0,1- 1,0 °C na
sekundo. Fluorescentno barvilo iz leta 1997 za real-time podaljševanje in taljenje pomnoženih
produktov je bilo SYBR® Green I [84]. Visoko specifična barvila, kot so SYTO® 9,
EvaGreen® in LCGreen® I nam dajejo boljše rezultate, saj bolj svetijo in jih lahko
uporabljamo pri višjih koncentracijah v primerjavi s tradicionalnimi barvili, kot je SYBR®
Green I [85]. LCGreen® I ima v primerjavi s SYBR® Green I sposobnost zaznavanja
heterodupleksov med samo talilno analizo. Prav tako ne inhibira PCR produktov, zato ga lahko
uporabljamo pri koncentracijah nasičenja in s tem zmanjšamo možnost za nastanek
nespecifičnih produktov. SYBR® Green I pa PCR produkt inhibira, zato ga uporabljamo pri
nizkih koncentracijah. Zaradi nizkih koncentracij ne dosežemo nasičenja, kar lahko privede
do prerazporejanja molekul barvila znotraj DNK med taljenjem.
Za HRM potrebujemo reakcijsko mešanico, ki vsebuje barvilo in DNK. Prvi korak je PCR, pri
katerem se pomnožijo zaporedja okoli izbranega polimorfizma SNP. Pomnoženi regiji rečemo
amplikon, PCR produkti pa so navadno krajši (100 do 159 bp). Drugi korak je HRM analiza
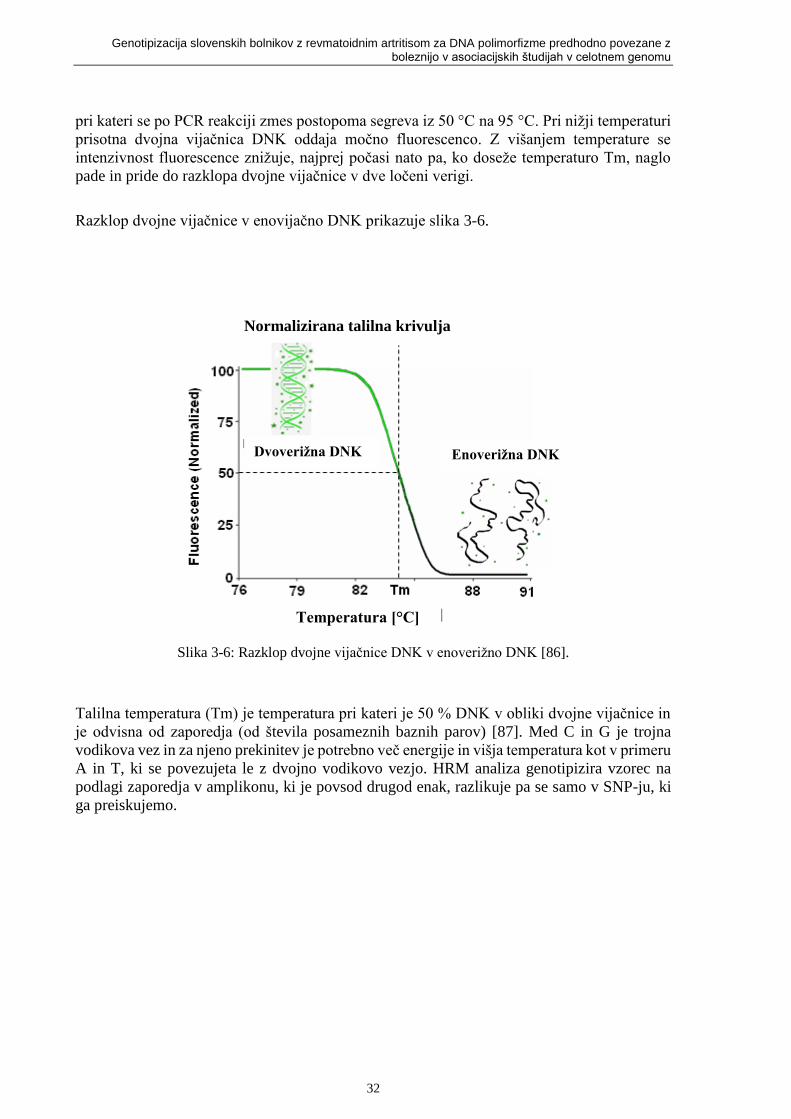
Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
32
pri kateri se po PCR reakciji zmes postopoma segreva iz 50 °C na 95 °C. Pri nižji temperaturi
prisotna dvojna vijačnica DNK oddaja močno fluorescenco. Z višanjem temperature se
intenzivnost fluorescence znižuje, najprej počasi nato pa, ko doseže temperaturo Tm, naglo
pade in pride do razklopa dvojne vijačnice v dve ločeni verigi.
Razklop dvojne vijačnice v enovijačno DNK prikazuje slika 3-6.
Slika 3-6: Razklop dvojne vijačnice DNK v enoverižno DNK [86].
Talilna temperatura (Tm) je temperatura pri kateri je 50 % DNK v obliki dvojne vijačnice in
je odvisna od zaporedja (od števila posameznih baznih parov) [87]. Med C in G je trojna
vodikova vez in za njeno prekinitev je potrebno več energije in višja temperatura kot v primeru
A in T, ki se povezujeta le z dvojno vodikovo vezjo. HRM analiza genotipizira vzorec na
podlagi zaporedja v amplikonu, ki je povsod drugod enak, razlikuje pa se samo v SNP-ju, ki
ga preiskujemo.
Normalizirana talilna krivulja
Temperatura [°C]
Enoverižna DNK Dvoverižna DNK
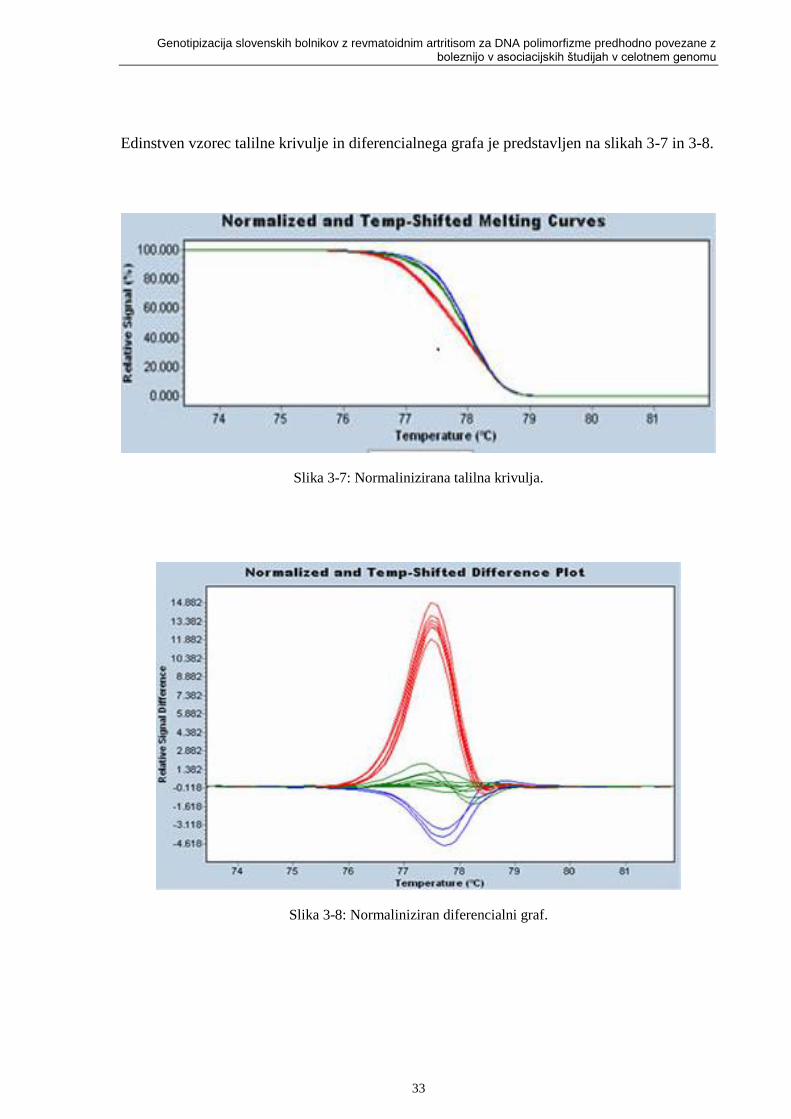
Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
33
Edinstven vzorec talilne krivulje in diferencialnega grafa je predstavljen na slikah 3-7 in 3-8.
Slika 3-7: Normalinizirana talilna krivulja.
Slika 3-8: Normaliniziran diferencialni graf.

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
34
Podobno kot pri PCR se tudi za HRM pripravi reakcijska mešanica. Reakcijsko mešanico smo
dodali V = 2 µL DNK vzorcem.
HRM reakcijsko mešanico smo naredili po protokolu, ki je prikazan v tabeli 3-7.
Tabela 3-7: Protokol za pripravo HRM reakcijske mešanice.
Reagent Volumen za 1 vzorec (µL)
DNK 2
PCR-grade H2O a
HRM Master Mix 5
MgCl2 b
Primer 1 c
Primer 2 c
Vrednosti a in b so bile določene po optmizaciji HRM. Parameter a smo izračunali naknadno.
Reakcije smo izvedli v Light Cycler LC480.
3.4 Računalniški programi Za načrtovanje eksperimentov in pri analizi podatkov smo si pomagali z različnimi
računalniškimi programi in bioinformatskimi orodji.
3.4.1 Bioinformatska orodja
Bioinformatika, uporaba računalniških tehnik za analizo informacij, povezane z
biomolekulami v velikem obsegu, je uveljavljena kot disciplina v molekularni biologiji in
zajema širok spekter področij od strukturne biologije, genomike do študij genske ekspresije
[88], [89], [90]. Bioinformatika se ukvarja s študijo metod za shranjevanje, pridobivanje in
analizo bioloških podatkov [91].
3.4.2 Primer3
Primer3 je spletni računalniški program, ki ga najdemo na spletni strani
http://bioinfo.ut.ee/primer3-0.4.0/. S Primer3 načrtujemo in analiziramo primerje za PCR in
PCR v realnem času. Program lahko zazna primerje za reakcije sekvenciranja in lahko načrtuje
hibridizacijske probe za oligonukleotide. Spletno orodje vsebuje pomembne aplikacije kot so
detekcija primerja, kloniranje, sekvenciranje in Primer seznam.
S Primer3 smo načrtovali začetne oligonukleotide za HRM.
3.4.3 SNP cutter
SNP cutter je prav tako spletno računalniško orodje za načrtovanje začetnih oligonukleotidov.
Najdemo ga na spletni strani http://bioapp.psych.uic.edu/SNP_cutter.htm. S tem programom
smo načrtovali začetne oligonukleotide za RFLP reakcijo.
3.4.4 GeneRunner 3.05
GeneRunner je preprost program za analizo v molekularni biologiji, katerega smo uporabili
za analizo sekvenc polimorfizmov SNP za RFLP.

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
35
V program lahko vstaviš sekvenco DNK ali proteinov. GeneRunner lahko izvede standardne
manipulacije sekvenčnih podatkov [92]:
- iskanje DNK sekvence za mesto restrikcije,
- iskanje sekvence proteinov za restikcijsko mesto peptidaz,
- translacija DNK v sekvenco proteinov in iskanje ORF,
- PCR in oblikovanje sekvenc začetnih oligonukleotidov,
- multi sekvenčno poravnanje.
Na spletnih straneh ameriškega Nacionalnega centra za biotehnološke informacije (NCBI)
smo v podatkovni bazi dbSN. poiskali sekvenco nukleotidnega tarčnega DNK fragmenta. V
program smo nato vnesli sekvenco prvega in sekvenco drugega alela. Dobili smo vse
restrikcijske encime, ki režejo eno in drugo sekvenco. Izbrali smo najbolj primeren
restrikcijski encim in ga naročili.
Slika 3-9 prikazuje primer uporabe programa GeneRunner. Zgoraj je prikazana sekvenca z
alelom A, katero nam restrikcijski encim NlaIII razreže na dva fragmenta. Spodaj pa je
prikazana homologna sekvenca z alelom T, ki je encim NlaIII ne reže. Zaradi razlik v velikosti
fragmentov lahko po končani elektroforezi na agaroznem gelu ločimo med posameznimi
genotipi.

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
36
Slika 3-9: Prikaz uporabe GeneRunner.
Alel A
Mesto
razreza z
restrikcijskim
encimom
NlaIII
Alel T

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
37
3.4.5 Program SPSS 20.0
Za statistično obdelavo smo uporabljali programski paket SPSS, pridobljeni z raziskavami. S
programom SPSS lahko opravljamo standardne statistične analize, kot so: opisno statistične
metode, korelacijske in regresijske metode, splošno linearno modeliranje, ipd. Z omenjenim
programskim orodjem smo analizirali dobljene rezultate genotipizacij. Za asociacijsko analizo
smo izvajali χ2 (hi-kvadrat) test t.j. Fisherjev natančni test. Fisherjev test se uporablja za
primerjavo frekvenc in za analizo variance. Za vsak SNP smo dobili p-vrednosti (statistično
signifikantne vrednosti p˂0,05), intervale zaupanja (confidence interval-CI95 %) in razmerja
obetov (odds ratio-OR). OR nam pove kakšen doprinos ima SNP, ki ga preiskujemo, za
nastanek določene bolezni. V našem primeru je to revmatoidi artritis.

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
38
4 Rezultati
4.1 Izbor polimorfizmov Kandidatne polimorfizme posameznega nukleotida (SNP), ki so pokazali povezavo z RA, smo
izbrali s sistematičnim pregledom GWA študij, ki so bile izvedene v zadnjih letih. Iz teh
asociacijskih študij smo izpisali vse polimorfizme posameznega nukleotida (SNP), ki
sodelujejo pri razvoju RA. V izbor smo uvrstili SNP-je, ki so bili največkrat replicirani in
imajo p < 5 x 10-8. Izbrali smo tudi gene, ki imajo p < 5 x 10-6 in še ni ustreznih replikacijskih
študij.
Po pregledu vse literature smo izbrali pet SNP-jev: rs1571878 (CCR6), rs7574865 (STAT4),
rs909685 (SYNGR1), rs2476601 (PTPN22) in rs3087243 (CTLA4). Izbrane SNP-je smo
predstavili v tabeli 4-1, v kateri so prikazani izbrani SNP-ji, citogenetska lokacija in
sprememba.
Tabela 4-1: Kandidatni polimorfizmi posameznega nukleotida (SNP).
SNP GEN Citogenetska
lokacija
Sprememba
rs1571878 CCR6 6q27 C/T
rs7574865 STAT4 2q32 G/T
rs909685 SINGR1 22q13.1 A/T
rs2476601 PTPN22 1p13 A/G
rs3087243 CTLA4 2q33 A/G
4.2 Optimizacija pogojev za RFLP test Za RFLP test smo uporabili polimorfizme SNP (rs909685 (SYNGR1), rs3087243 (CTLA4) in
rs2476601 (PTPN22). Začetne oligonukleotide (primerje) smo poiskali s pomočjo spletnega
programa SNPcutter-ja. Optimizacijo optimalnih pogojev smo izvedli samo za rs909685
(SYNGR1), ker so bili pogoji za rs3087243 (CTLA4) in rs2476601 (PTPN22) že optimizirani.
Najprej smo optimizirali optimalno temperaturo začetnih oligonukleotidov, pri kateri se le-ti
pripenjajo.
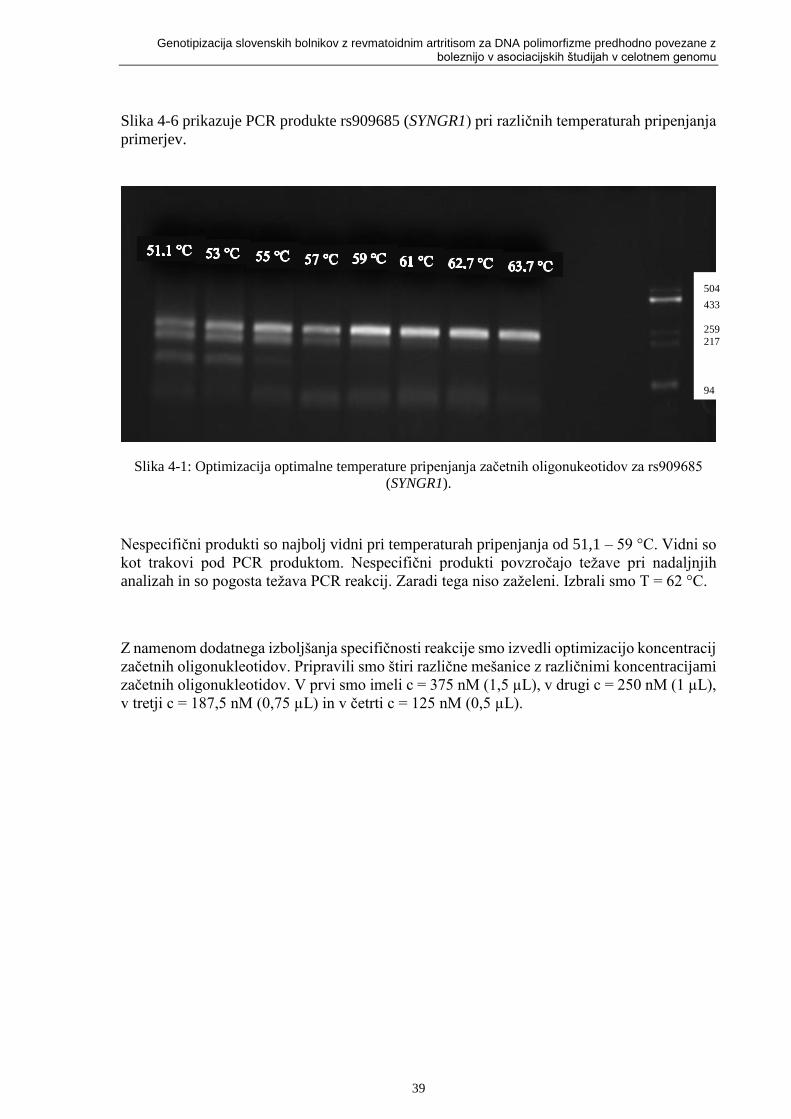
Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
39
Slika 4-6 prikazuje PCR produkte rs909685 (SYNGR1) pri različnih temperaturah pripenjanja
primerjev.
Slika 4-1: Optimizacija optimalne temperature pripenjanja začetnih oligonukeotidov za rs909685
(SYNGR1).
Nespecifični produkti so najbolj vidni pri temperaturah pripenjanja od 51,1 – 59 °C. Vidni so
kot trakovi pod PCR produktom. Nespecifični produkti povzročajo težave pri nadaljnjih
analizah in so pogosta težava PCR reakcij. Zaradi tega niso zaželeni. Izbrali smo T = 62 °C.
Z namenom dodatnega izboljšanja specifičnosti reakcije smo izvedli optimizacijo koncentracij
začetnih oligonukleotidov. Pripravili smo štiri različne mešanice z različnimi koncentracijami
začetnih oligonukleotidov. V prvi smo imeli c = 375 nM (1,5 µL), v drugi c = 250 nM (1 µL),
v tretji c = 187,5 nM (0,75 µL) in v četrti c = 125 nM (0,5 µL).
504
433
259
217
94

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
40
Na sliki 4-7 so vidni PCR produkti rs909685 (SYNGR1) pri različnih koncentracijah primerjev.
Slika 4-2: Optimizacija koncentracij začetnih oligonukleotidov za rs909685 (SYNGR1).
PCR produkti so zelo dobro vidni pri vseh volumnih primerjev, zato smo izbrali najmanjši
volumen, tj. V = 0,5 µL. Optimizacija MgCl2 ni bila potrebna, ker začetni oligonukleotidi niso
tvorili nobenih nespecifičnih produktov.
Za rs909685 (SYNGR1) smo morali še optimizirati količino restrikcijskega encima NlaIII, ki
ga potrebujemo pri RFLP reakciji. Produktom PCR reakcije smo dodali različne količine
restrikcijskega encima in izbrali optimalno količino pri kateri se izbrana sekvenca DNK
najlepše reže.
1,5 µL 1 µL
0,75 µL 0,5 µL
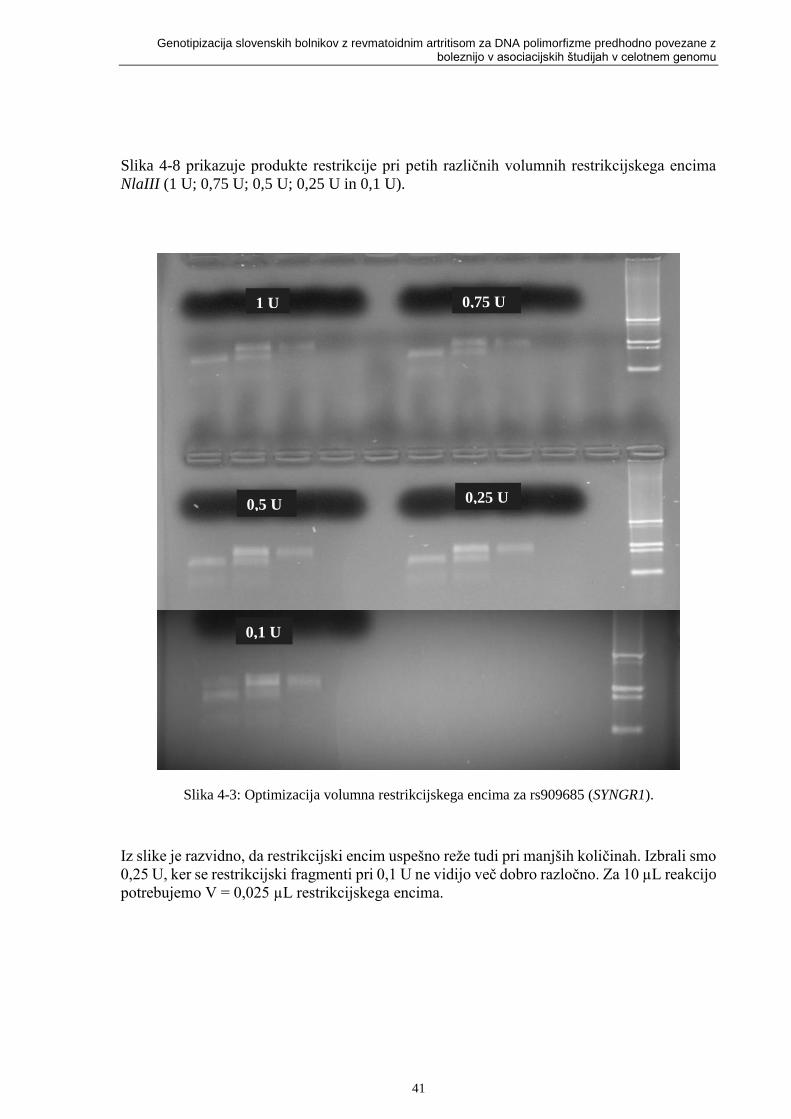
Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
41
Slika 4-8 prikazuje produkte restrikcije pri petih različnih volumnih restrikcijskega encima
NlaIII (1 U; 0,75 U; 0,5 U; 0,25 U in 0,1 U).
Slika 4-3: Optimizacija volumna restrikcijskega encima za rs909685 (SYNGR1).
Iz slike je razvidno, da restrikcijski encim uspešno reže tudi pri manjših količinah. Izbrali smo
0,25 U, ker se restrikcijski fragmenti pri 0,1 U ne vidijo več dobro razločno. Za 10 µL reakcijo
potrebujemo V = 0,025 µL restrikcijskega encima.
1 U 0,75 U
0,5 U 0,25 U
0,1 U
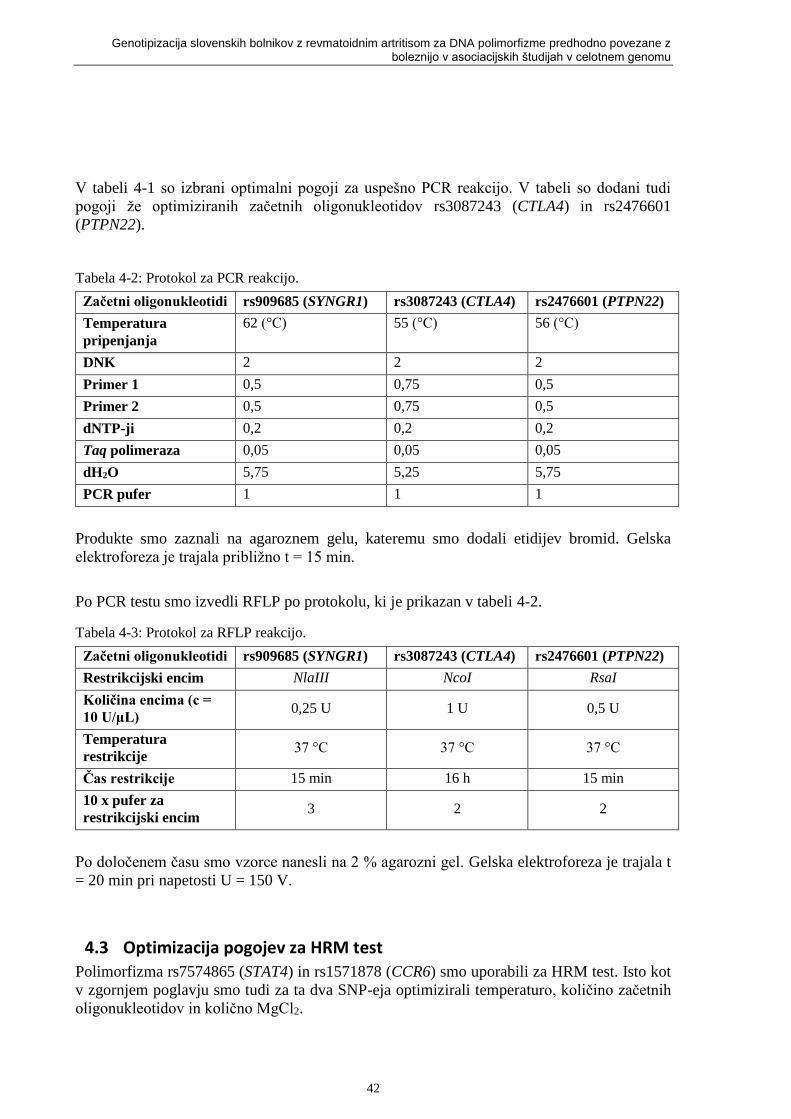
Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
42
V tabeli 4-1 so izbrani optimalni pogoji za uspešno PCR reakcijo. V tabeli so dodani tudi
pogoji že optimiziranih začetnih oligonukleotidov rs3087243 (CTLA4) in rs2476601
(PTPN22).
Tabela 4-2: Protokol za PCR reakcijo.
Začetni oligonukleotidi rs909685 (SYNGR1) rs3087243 (CTLA4) rs2476601 (PTPN22)
Temperatura
pripenjanja
62 (°C) 55 (°C) 56 (°C)
DNK 2 2 2
Primer 1 0,5 0,75 0,5
Primer 2 0,5 0,75 0,5
dNTP-ji 0,2 0,2 0,2
Taq polimeraza 0,05 0,05 0,05
dH2O 5,75 5,25 5,75
PCR pufer 1 1 1
Produkte smo zaznali na agaroznem gelu, kateremu smo dodali etidijev bromid. Gelska
elektroforeza je trajala približno t = 15 min.
Po PCR testu smo izvedli RFLP po protokolu, ki je prikazan v tabeli 4-2.
Tabela 4-3: Protokol za RFLP reakcijo.
Začetni oligonukleotidi rs909685 (SYNGR1) rs3087243 (CTLA4) rs2476601 (PTPN22)
Restrikcijski encim NlaIII NcoI RsaI
Količina encima (c =
10 U/µL) 0,25 U 1 U 0,5 U
Temperatura
restrikcije 37 °C 37 °C 37 °C
Čas restrikcije 15 min 16 h 15 min
10 x pufer za
restrikcijski encim 3 2 2
Po določenem času smo vzorce nanesli na 2 % agarozni gel. Gelska elektroforeza je trajala t
= 20 min pri napetosti U = 150 V.
4.3 Optimizacija pogojev za HRM test Polimorfizma rs7574865 (STAT4) in rs1571878 (CCR6) smo uporabili za HRM test. Isto kot
v zgornjem poglavju smo tudi za ta dva SNP-eja optimizirali temperaturo, količino začetnih
oligonukleotidov in količno MgCl2.
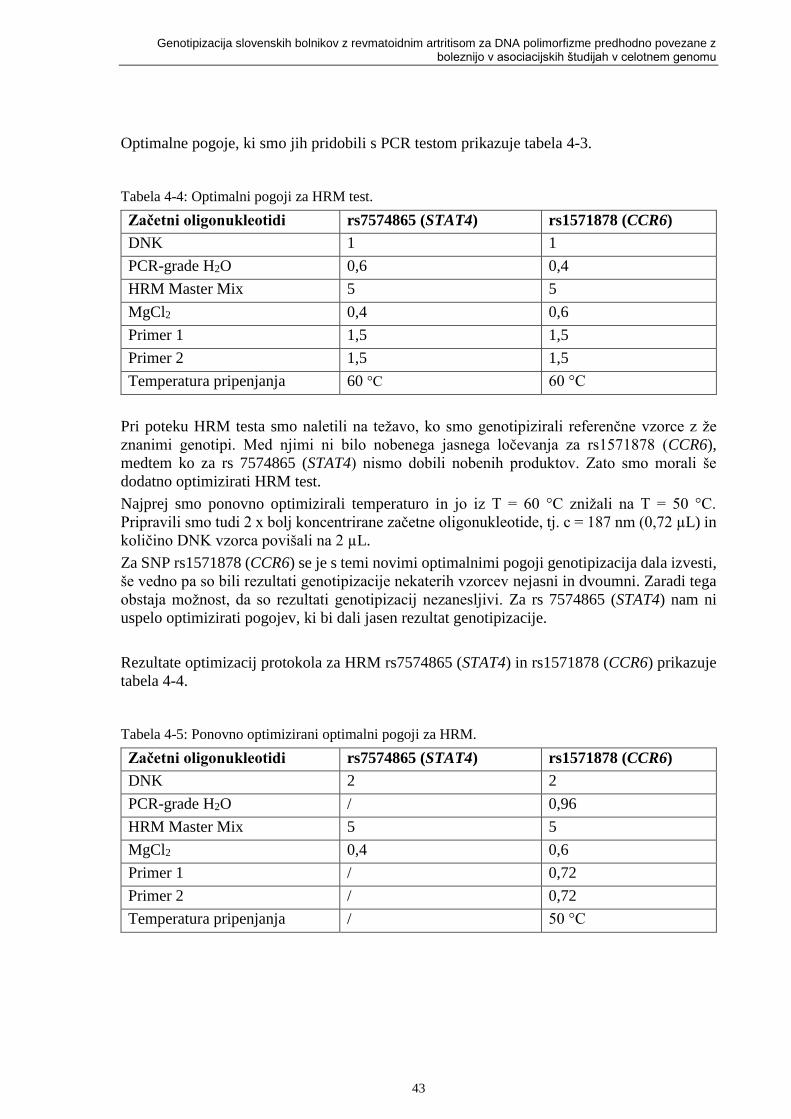
Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
43
Optimalne pogoje, ki smo jih pridobili s PCR testom prikazuje tabela 4-3.
Tabela 4-4: Optimalni pogoji za HRM test.
Začetni oligonukleotidi rs7574865 (STAT4) rs1571878 (CCR6)
DNK 1 1
PCR-grade H2O 0,6 0,4
HRM Master Mix 5 5
MgCl2 0,4 0,6
Primer 1 1,5 1,5
Primer 2 1,5 1,5
Temperatura pripenjanja 60 °C 60 °C
Pri poteku HRM testa smo naletili na težavo, ko smo genotipizirali referenčne vzorce z že
znanimi genotipi. Med njimi ni bilo nobenega jasnega ločevanja za rs1571878 (CCR6),
medtem ko za rs 7574865 (STAT4) nismo dobili nobenih produktov. Zato smo morali še
dodatno optimizirati HRM test.
Najprej smo ponovno optimizirali temperaturo in jo iz T = 60 °C znižali na T = 50 °C.
Pripravili smo tudi 2 x bolj koncentrirane začetne oligonukleotide, tj. c = 187 nm (0,72 µL) in
količino DNK vzorca povišali na 2 µL.
Za SNP rs1571878 (CCR6) se je s temi novimi optimalnimi pogoji genotipizacija dala izvesti,
še vedno pa so bili rezultati genotipizacije nekaterih vzorcev nejasni in dvoumni. Zaradi tega
obstaja možnost, da so rezultati genotipizacij nezanesljivi. Za rs 7574865 (STAT4) nam ni
uspelo optimizirati pogojev, ki bi dali jasen rezultat genotipizacije.
Rezultate optimizacij protokola za HRM rs7574865 (STAT4) in rs1571878 (CCR6) prikazuje
tabela 4-4.
Tabela 4-5: Ponovno optimizirani optimalni pogoji za HRM.
Začetni oligonukleotidi rs7574865 (STAT4) rs1571878 (CCR6)
DNK 2 2
PCR-grade H2O / 0,96
HRM Master Mix 5 5
MgCl2 0,4 0,6
Primer 1 / 0,72
Primer 2 / 0,72
Temperatura pripenjanja / 50 °C

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
44
4.4 Asociacijska analiza izbranih SNP pri bolnikih z RA Izvedli smo genotipizacijo z metodo PCR-RFLP in z HRM metodo na različnem številu
vzorcev bolnikov in zdravih kontrol. Genotipizirane vzorce smo analizirali in jih med seboj
primerjali. Primerjali smo genotipe bolnikov in genotipe zdravih kontrol.
4.4.1 Rezultati RFLP
Po končani optimizaciji smo izvedli genotipizacijo na RFLP in HRM testu. Genotipizirali smo
vzorce bolnikov z RA in vzorce kontrol.
Rezultati RFLP testa so bili vidni na agaroznih gelih z uporabo referenčnih CEPH vzorcev.
Te vzorce znanih genotipov smo predhodno poiskali v podatkovni bazi.
Slika 4-9 prikazuje shematski prikaz detekcije izbranega polimorfizma po restrikciji na
agaroznem gelu.
Slika 4-4: Shematski in slikovni prikaz detekcije polimorfizma rs909685 (SYNGR1) na agaroznem
gelu.
rs909685
Velikost
produkta
237
172
65
A/A A/T T/T

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
45
4.4.2 Rezultati HRM
Slika 4-10 prikazuje rezultate HRM testa za rs1571878 (CCR6).
Slika 4-5: Normalizirana in obdelana talilna krivulja HRM testa.
4.4.3 Primerjava rezultatov dobljenih iz genotipizacij
Genotipizirali smo izbrane SNP-je pri bolnikih in zdravih posameznikih in primerjali dobljene
frekvence genotipov posameznega SNP-ja in frekvence alelov s frekvencami iz podatkovne
baze projekta HapMap. Naše izračunane vrednosti smo primerjali s frekvencami izmerjenimi
na populaciji Evropejcev (CEU), katere smo našli na spletni strani NCBI [93], [94], [95], [96].
Primerjave prikazuje spodnja tabela 4-5.
Tabela 4-6: Primerjava izračunih frekvenc s frekvencami dobljenih iz projekta HapMap.
CCR6
(rs1571878)
SYNGR1
(rs909685)
PTPN22
(rs2476601)
Izmerjeno HapMap Izmerjeno HapMap Izmerjeno HapMap
Frekvenca
genotipov
CC: 0,211
CT: 0,463
TT: 0,325
CC: 0,183
CT: 0,450
TT: 0,367
AA: 0,188
AT: 0,437
TT: 0,376
AA: 0,133
AT: 0,450
TT: 0,417
AA: 0,011
AG: 0,144
GG: 0,844
AA: 0,009
AG: 0,216
GG: 0,775
Frekvenca
alelov
C: 0,443
T: 0,556
T: 0,592
C: 0,408
A: 0,406
T: 0,594
A: 0,358
T: 0,642
A: 0,083
G: 0,917
A: 0,117
G: 0,883
CTLA4
(rs3087243)
Izmerjeno HapMap
Frekvenca
genotipov
AA: 0,216
AG: 0,511
GG: 0,272
AA: 0,204
AG: 0,513
GG: 0,283

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
46
Frekvenca
alelov
A: 0,472
G: 0,528
A: 0,460
G: 0,540
Med izračunanimi frekvancami in frekvencami genotipov iz podatkovne baze je opazno
odstopanje. To razhajanje je lahko posledica različnih populacij saj smo pri našem delu
uporabljali vzorce slovenske populacije, pri projektu HapMap pa so bili uporabljeni vzorci
evropske populacije.
Največje razlike so vidne pri SNP rs1571878 (CCR6), kar lahko pripišemo manj zanesljivi
genotipizaciji ali pa kot smo že omenili, različni populaciji.
4.4.4 Rezultati statističnih analiz
Povezave med posameznim SNP-jem in boleznijo smo ugotavljali s statistično analizo. Izvedli
smo jo z uporabo pridobljenih podatkov za genotipe izbranih SNP-jev za 276 zdravih
posameznikov in 276 bolnikov z RA.
Tabele 4-7, 4-8, 4-9 in 4-10 predstavljajo statistične analize. Primerjali smo frekvence
genotipov in alelov med bolniki in kontrolno skupino (zdravi posamezniki). Za primerjavo
genotipskih frekvenc smo uporabili recesivni model, nato pa še dominantni model glede na
prvi alel.
S p-vrednostjo smo preverjali statistično signifikantnost. Izračunana p-vrednost je morala biti
p˂0,05, da bi povezavo ovrednotili kot statistično signifikantno.
Pri naši analizi je do razlik v signifikanci prišlo le pri dveh SNP-jih. Pri SNP-eju rs2476601
(Tabela 4-9) smo ugotovili, da je frekvenca genotipa A/A signifikantno višja (p = 1,3 x 10-5)
pri bolnikih (3,1 %) v primerjavi s kontrolami (1,1 %). Alelna frekvenca za alel A je prav tako
višja pri bolnikih (17,5 %). P-vrednost je bila p = 1,1 x 10-5.
Predvidevamo lahko, da alel A zvišuje tveganje za razvoj RA.
SNP rs3087243 prikazuje tabela 4-10. Frekvenca genotipa G/G je signifikantno nižja (p =
0,002) pri bolnikih (25,6 %) v primerjavi s kontrolami (29,5 %). Prav tako je alelna frekvenca
za alel G nižja pri bolnikih (44,5 %). P-vrednost je 0,028.
Sklepamo lahko, da alel G zmanjšuje tveganje za nastanek RA.

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
47
Tabela 4-7: p-vrednosti, razmerja obetov in intervali zaupanja za RA bolnike in kontrole za CCR6.
CCR6
rs1571878
RA
(n= 247)
Kontrole
(n= 246)
TT 38,0 % 32,5 %
CT 42,5 % 46,3 %
CC 19,8 % 21,1 %
T 0,589 0,557
C 0,411 0,443
Statistična analiza
Primerjava genotipov
CC proti CT + TT
p vrednost 0,739
OR 0,923
95 % CI 0,644-1,511
CC + CT proti TT
p vrednost 0,228
OR 0,786
95 % CI 0,596-1,430
Primerjava alelov
p vrednost 0,334
OR 0,877
95 % CI 0,681-1,129
TT = homozigot za alel T, CT = heterozigot, CC = homozigot za alel C, CC proti CT + TT je
dominantni model za alel T, CC + CT proti TT je dominantni model za alel C
n = število oseb z določenim genotipom, p = vrednost statistično značilne povezave, OR =
razmerje obetov z 95 % intervalom zaupanja
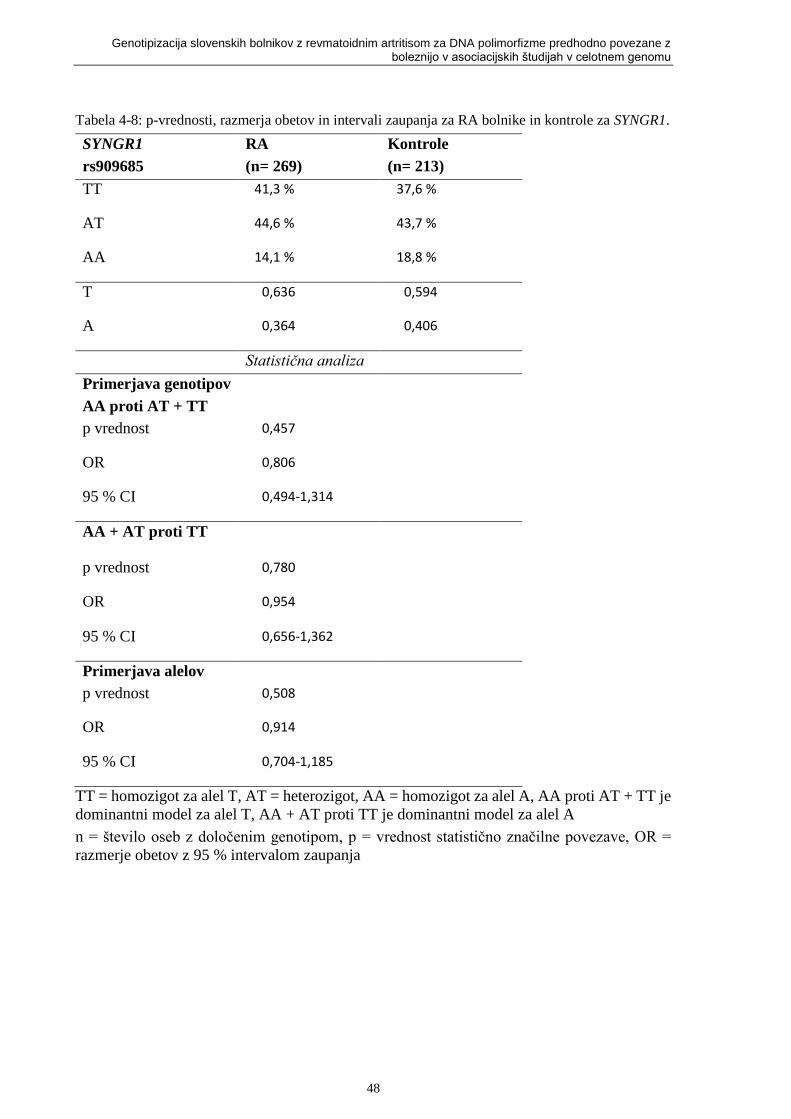
Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
48
Tabela 4-8: p-vrednosti, razmerja obetov in intervali zaupanja za RA bolnike in kontrole za SYNGR1.
SYNGR1
rs909685
RA
(n= 269)
Kontrole
(n= 213)
TT 41,3 % 37,6 %
AT 44,6 % 43,7 %
AA 14,1 % 18,8 %
T 0,636 0,594
A 0,364 0,406
Statistična analiza
Primerjava genotipov
AA proti AT + TT
p vrednost 0,457
OR 0,806
95 % CI 0,494-1,314
AA + AT proti TT
p vrednost 0,780
OR 0,954
95 % CI 0,656-1,362
Primerjava alelov
p vrednost 0,508
OR 0,914
95 % CI 0,704-1,185
TT = homozigot za alel T, AT = heterozigot, AA = homozigot za alel A, AA proti AT + TT je
dominantni model za alel T, AA + AT proti TT je dominantni model za alel A
n = število oseb z določenim genotipom, p = vrednost statistično značilne povezave, OR =
razmerje obetov z 95 % intervalom zaupanja

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
49
Tabela 4-9: p-vrednosti, razmerja obetov in intervali zaupanja za RA bolnike in kontrole za PTPN22.
PTPN22
rs2476601
RA
(n= 254)
Kontrole
(n= 270)
AA 3,1 % 1,1 %
AG 28,7 % 14,4 %
GG 68,1 % 84,4 %
A 0,175 0,083
G 0,825 0,917
Statistična analiza
Primerjava genotipov
AA proti AG + GG
p vrednost 1,3 x 10-5
OR 2,542
95 % CI 1,667-3,875
AA + AG proti GG
p vrednost 0,132
OR 2,894
95 % CI 0,759-11,033
Primerjava alelov
p vrednost 1,1 x 10-5
OR 2,337
95 % CI 1,596-3,421
AA = homozigot za alel A, AG = heterozigot, GG = homozigot za alel G, AA proti AG + GG
je dominantni model za alel G, AA + AG proti GG je dominantni model za alel A
n = število oseb z določenim genotipom, p = vrednost statistično značilne povezave, OR =
razmerje obetov z 95 % intervalom zaupanja
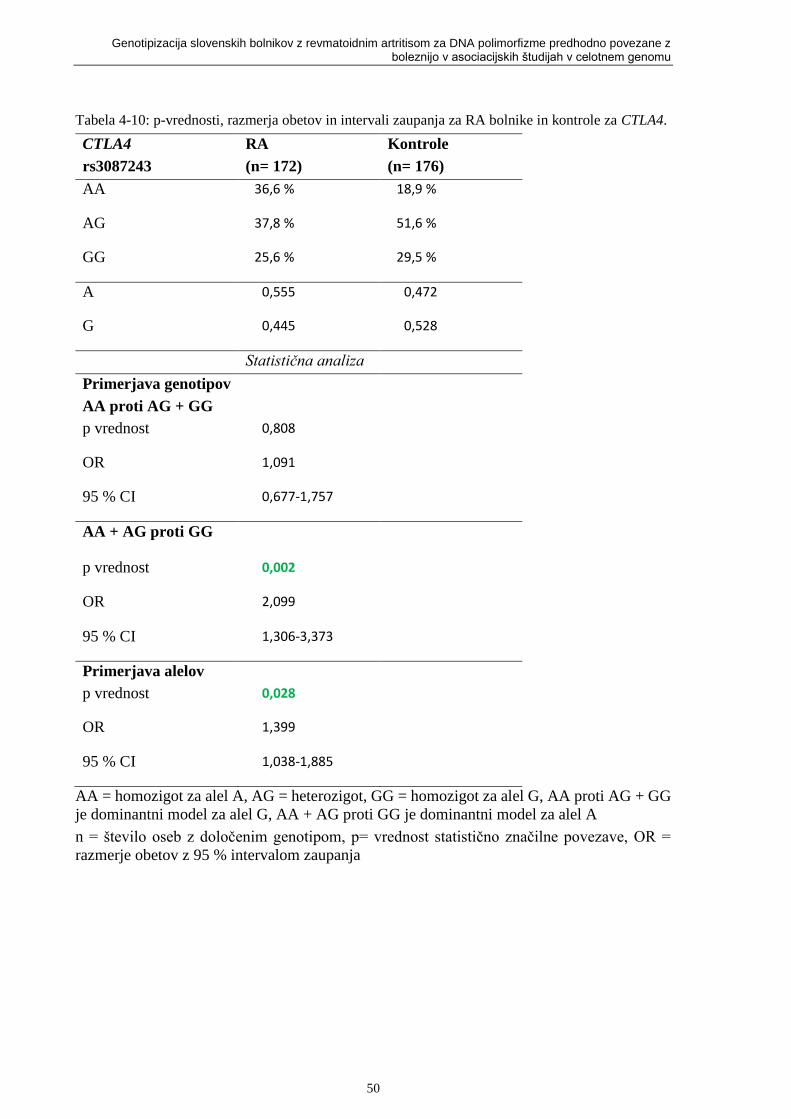
Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
50
Tabela 4-10: p-vrednosti, razmerja obetov in intervali zaupanja za RA bolnike in kontrole za CTLA4.
CTLA4
rs3087243
RA
(n= 172)
Kontrole
(n= 176)
AA 36,6 % 18,9 %
AG 37,8 % 51,6 %
GG 25,6 % 29,5 %
A 0,555 0,472
G 0,445 0,528
Statistična analiza
Primerjava genotipov
AA proti AG + GG
p vrednost 0,808
OR 1,091
95 % CI 0,677-1,757
AA + AG proti GG
p vrednost 0,002
OR 2,099
95 % CI 1,306-3,373
Primerjava alelov
p vrednost 0,028
OR 1,399
95 % CI 1,038-1,885
AA = homozigot za alel A, AG = heterozigot, GG = homozigot za alel G, AA proti AG + GG
je dominantni model za alel G, AA + AG proti GG je dominantni model za alel A
n = število oseb z določenim genotipom, p= vrednost statistično značilne povezave, OR =
razmerje obetov z 95 % intervalom zaupanja

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
51
5 Diskusija
V nalogi smo s povečanim tveganjem za nastanek RA v slovenski populaciji povezali dva
SNP-ja, rs2476601 (PTPN22) in rs3087243 (CTLA4), medtem ko za SNP-ja rs1571878
(CCR6) in rs909685 (SYNGR1) statistične povezave nismo dokazali.
Odkar so leta 2004 prvič odkrili povezavo SNP-ja rs2476601 (PTPN22) z RA [34], so številne
študije pri različnih populacijah, vključno s francosko, britansko, finsko, švedsko, nemško,
špansko in kanadsko, potrdile to povezavo. Ugotovili so, da alel A povečuje tveganje za
nastanek bolezni [46], [97].
V naši študiji smo za SNP rs2476601 (PTPN22) uspešno genotipizirali 254 vzorcev bolnikov
z RA in 270 vzorcev zdravih posameznikov in potrdili statistično značilno povezavo.
Ugotovili smo, da je frekvenca alela A višja pri bolnikih (17,5 %) v primerjavi z zdravimi
posamezniki (8,3 %, p = 1,1 x 10-5). Iz tega sklepamo da alel A zvišuje tveganje za razvoj RA
tudi v slovenski populaciji.
Primerjali smo rezultate naše študije s študijo, ki so jo izvedli na britanski populaciji [98].
Rezultate obeh študij smo predstavili v tabeli 5-1.
Kot je razvidno iz tabele 5-1, so za SNP rs2476601 na genu PTPN22 potrdili statistično
značilno povezavo z RA na britanski populaciji (p = 4,87 x 10-33) [98]. Želeli smo preveriti,
če bi dobili podoben rezultat z enakim številom vzorcev kontrol in RA bolnikov, kot v tuji
študiji, saj je p-vrednost v primerjavi z našo visoka. Izvedli smo ponovno statistično analizo
in dobili, da je p = 3,096 x 10-30.
Tabela 5-1: Primerjava študij za SNP rs2476601.
Število kontrol
(n)
Število RA
bolnikov (n)
p-vrednost
SLOVENSKA
ŠTUDIJA 270 254 p = 1,3 x 10-5
SLOVENSKA
ŠTUDIJA
(povečano
število kontrol
in RA
bolnikov)
2 334 1 361 p = 3,096 x 10-30
BRITANSKA
ŠTUDIJA [98] 2 334 1 361 p = 4,87 x 10-33

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
52
Za SNP rs3087243 na genu CTLA4 so različne replikacijske študije pokazale spodbudne,
vendar neponovljive rezultate v povezavi z RA. Študija na severno ameriški populaciji je
potrdila asociacijo SNP-ja rs3087243 (CTLA4) [41], toda študija, opravljena na švedski
populaciji [35] asociacije SNP-ja rs3087243 z RA ni dokazala. Povezavo med CTLA4 in RA
pa so potrdili pri evropski in azijski populaciji [46]. Plenge s sod. [35] je dokazal, da razlike
med študijami lahko nastanejo zaradi nezadostne statistične moči v nekaterih študijah.
Za SNP rs3087243 (CTLA4) smo uspešno genotipizirali 172 RA bolnikov in 176 zdravih
posameznikov.
Asociacijska študija na evropski populaciji [44] je pokazala, da ima alel A SNP-eja rs3087243
(CTLA4) zaščitno vlogo pred RA in zmanjšuje tveganje za nastanek RA. Alel A ima nižjo
frekvenco pri bolnikih (39 %) v primerjavi s kontrolami (44 %).
V naši študiji je frekvenca genotipa G/G pri bolnikih (25,6 %) nižja v primerjavi s kontrolami
(29,5 %). Sklepamo lahko, da ima G/G genotip za slovensko populacijo zaščitno vlogo (p =
0,002), kar pomeni, da imajo nosilci tega genotipa zmanjšano tveganje za nastanek bolezni.
Gen CCR6 je bil povezan z RA v različnih etničnih populacijah. Vendar je nekaj replikacijskih
študij poročalo o nasprotujočih rezultatih asociacije CCR6 z RA [99]. Nedavno so tri GWA
študije povezale CCR6 z RA [42], [45], [100]. Najmočnejša povezava med CCR6 in RA se je
pokazala na japonski populaciji [99].
Študije, ki so preučevale povezavo med genom CCR6 in RA, so v večini analizirale SNP-je
rs3093024, rs3099023 in rs1854853. Povezavo s SNP-jem rs1571878 na genu CCR6 so do
sedaj opravile dve GWA študiji [46], [47].
Okada s sod. je v GWA študiji [47], ki jo je opravil na japonski in evropski populaciji, potrdil
povezavo med SNP-jem rs1571878 (CCR6) in RA (p = 5,9 x 10-7).
V slovenski populaciji smo genotipizirali rs1571878 (CCR6) pri 247 RA bolnikih in 246
zdravih posameznikih. V naši študiji nismo našli povezave med RA in SNP-jem rs1571878
(CCR6). V prihodnjih raziskavah bo potrebno povečati število vzorcev saj je lahko na naš
rezultat vplivalo premalo število vzorcev, ki niso dosegli zadostne statistične moči.
V GWA študiji [47] so potrdili povezavo med SNP-jem rs909685 (SYNGR1) in RA. Študijo
so opravili na azijski in evropski populaciji pri 19 243 bolnikih in 61 565 kontrolah. Na azijski
populaciji so dobili vrednost statistično značilne povezave p = 2,0 x 10-7, na evropski
populaciji pa p = 6,4 x 10-12. V naši študiji nismo našli povezave med SNP-jem rs909685
(SYNGR1) in RA. Na to je lahko vplivalo premalo število vzorcev, ki ni doseglo zadostne
statistične moči in bo v prihodnjih raziskavah število vzorcev potrebno povečati.

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
53
6 Zaključek
Revmatoidni artritis je avtoimunska bolezen, ki prizadene vse sklepne dele telesa. Na nastanek
in patogenezo RA vplivajo dejavniki okolja in genetika. Na področju genetike RA niso
pojasnjeni vsi dejavniki, ki vplivajo na tveganje za razvoj RA. Odkar ugotavljajo vpliv
posameznih genov na razvoj bolezni so odkrili že veliko SNP-jev, ki v določeni meri povečajo
tveganje za nastanek RA.
V raziskavi smo izbrali pet SNP-jev in preverajli njihovo povezanost z RA pri slovenski
populaciji.
Statistična analiza je pokazala povezavo med RA in SNP-jem rs2476601 na genu PTPN22 za
genotipski model AA proti AG + GG (p = 1,3 x 10-5) in alelni model (p = 1,1 x 10-5). Za SNP
rs3087243 na genu CTLA4 smo potrdili statistično značilno povezavo za genotipski model AA
+ AG proti GG (p = 0,002) kot tudi za alelni model (p = 0,028). V primeru SNP-jev rs1571878
(CCR6) in rs909685 (SYNGR1) statistično značilne povezave nismo potrdili.
Z odkrivanjem in določanjem novih SNP-jev ter genov, ki povečujejo dovzetnost za razvoj
RA, omogočimo lažjo napoved bolezni in lažje načrtovanje zdravljenja. Zdravila za popolno
ozdravitev RA ni. Obstajajo pa standardna in biološka zdravila, s katerimi se pri bolnikih
lajšajo bolečine in simptomi. Z nadaljnjim odkrivanjem novih SNP-jev ter genov pri RA bi
pripomogli k razvoju zdravil, katera bi omogočila učinkovitejše zdravljenje in izboljšala
kakovost življenja bolnikov z RA.

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
54
7 Literatura
[1] Viatte S., Plant D., Raychaudhuri S. Genetics and epigenetics of rheumatoid arthritis.
Nat Rev Rheumatol., 9 (3), 141-153, 2014.
[2] Ješe R., Tomšič M. Vnetne revmatične bolezni. Farmacevtski vestnik, 64 (4), 267-271,
2013.
[3] Kumar V., Abbas A. K., Fausto N. Robbins and Cotran Pathologic Basis of Disease,
8th edition. Philadelphia: Elsevier Saunders, 2010.
[4] Kocjančič A., Mrevlje F., Štajer D. Interna medicina, tretja izdaja. Ljubljana: Littera
picta, 2005.
[5] Fauci A. S. et al. Harrison's principles of internal medicine, 17th edition. New York:
McGraw-Hill, Medical Publishing Division, 2008.
[6] Klemenc-Ketiš Z., Tušek-Bunc K. Navodila za bolnike: knjiga 2, Bolezni in poškodbe.
Ljubljana: Zavod za razvoj družinske medicine, 2009.
[7] Wasserman A. M. Diagnosis and Management of Rheumatoid Arthritis. American
Academy of Family Physicians, 84 (11) 1245-1252, 2011.
[8] Sotlar Južnič M., Kralj T. Viva, priloga skrb zase: Zdravljenje do cilja, revmatoidni
artritis, 2013. http://www.revmatiki.si/wp-content/uploads/2013/06/Priloga-Revma.pdf
(dostop 2. 4. 2015)
[9] Banal F, Dougados M, Combescure C, Gossec L. Sensitivity and specificity of the
American College of Rheumatology 1987 criteria for the diagnosis of rheumatoid arthritis
according to disease duration: a systematic literature review and meta-analysis. Ann Rheum
Dis, 68 (7), 1184-1191, 2009.
[10] Combe B, Landewe R, Lukas C, Bolosiu HD, Breedveld F, Dougados M, et al. EULAR
recommendations for the management of early arthritis: report of a task force of the European
Standing Committee for International Clinical Studies Including Therapeutics (ESCISIT).
Ann Rheum Dis, 66 (1), 34-45, 2007.
[11] Gupta S., Kumar S. Ethiopathogenesis and Investigation Tools: A Shadow of
Rheumatoid Arthritis. International Journal of Pharmaceutical Studies and Research, 2 (2), 63-
78, 2011.
[12] McAllister K., Eyre S., Orozco G. Genetic of rheumatoid arthritis: GWAS and beyond.
Open Access Rheumatology: Research Reviews, 3 (1), 31-46, 2011.
[13] Risch N., Merikangas K. The future of genetic studies of complex human diseases.
Science, 273 (5281), 1516-1517, 1996.
[14] Zhu M., Zhao S. Candidate gene identification approach: progress and challenges. Int
J Biol Sci., 3 (7), 420-427, 2007.
[15] Lengauer T. Bioinformatics–From Genomes to Therapies: Volume 3: Molecular
Function. Nemčija: Wiley-VCH, 2007.
[16] Sahebi L., Dastgiri S., Ansarin K., Sahebi R., Mohammadi S. A. Study Design in
Genetic Epidemiology. ISRN Genetics, 1-8, 2013.
[17] Cordell H. J., Clayton D. G. Genetic association studies. Lancet, 366, 1121-1131,
2005.
[18] Bush W. S., Moore J. H. Chapter 11: Genome-Wide Association Studies. PLoS
Comput Biol., 8 (12), e1002822, 2012.

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
55
[19] Lander E. S., Linton L. M., Birren B., et al. Initial sequencing and analysis of the
human genome. Nature, 409 (6822), 860-921, 2001.
[20] International HapMap Consortium. A haplotype map of the human genome. Nature,
437 (7063), 1299-1320, 2005.
[21] Lee J.K. Statistical Bioinformatics: A Guide For Biomedical and Life Science
Reasearchers. ZDA: John Wiley and Sons, 2010.
[22] Kochi Y., Suzuki A., Yamamoto K. Genetic basis of rheumatoid arthritis: A current
review. Biochemical and Biophysical Research Communications, 452, 254-262, 2014.
[23] Seldin M. F., Amos C. I., Ward R., Gregersen P. K. The Genetics Revolution and the
Assault on Rheumatoid Arthritis. Arthritis & Rheumatism, 42 (6), 1071-1079, 1999.
[24] Bax M., van Heemst J., Hulzinga T. W. J., Toes R. E. M. Genetics of Rheumatoid
Arthritis: what have we learned? Immunogenetics, 63, 459-466, 2011.
[25] McInnes I. B., Schnett G. Mechanisms of Disease: The Pathogenesis of Rheumatoid
Arthritis. The New England Journal of Medicine, 365 (23), 2205-2219, 2011.
[26] MacKay K., Eyre S., et al. Whole – Genome Linkage Analysis of Rheumatoid Arthritis
Susceptibility Loci in 252 Affected Sibling Pairs in the United Kingdom. Arthritis &
Rheumatism, 46 (3), 632-639, 2002.
[27] Wakitani S, Murata N, Toda Y et al. The relationship between HLA-DRB1 alleles and
disease subsets of rheumatoid arthritis in Japanese. Br J Rheumatol 36; 630-636, 1997.
[28] Wordsworth B. P., Lanchbury J. S., Sakkas L. I., Welsh K. I., Panayi G. S., Bell J. I.
HLA-DR4 subtype frequencies in rheumatoid arthritis indicate that DRB1 is the major
susceptibility locus within the HLA class II region. Proc Natl Acad Sci U S A, 86 (24), 10049-
10053, 1989.
[29] Gregersen PK, Shen M, Song QL et al. Molecular diversity of HLA-DR4 haplotypes.
Proc Natl Acad Sci U S A 83 (8), 2642-2646, 1986.
[30] Deighton C. M., Walker D. J., Griffiths I.D., Roberts D. F. The contribution of HLA
to rheumatoid arthritis. Clin Genet, 36 (3), 178-182, 1989.
[31] Suzuki A., Yamada R., Chang X. et al. Functional haplotypes of PADI4, encoding
citrullinating enzyme peptidylarginine deiminase 4, are associated with rheumatoid arthritis.
Nat Genet, 34 (4); 395-402, 2003.
[32] Burr M. L., Naseem H., Hinks A. et al. PADI4 genotype is not associated with
rheumatoid arthritis in a large UK Caucasian population. Ann Rheum Dis, 69; 666-670, 2010.
[33] Lee Y. H., Rho Y. H., Choi S. J., Ji J. D., Song G. G. PADI4 polymorphisms and
rheumatoid arthritis susceptibility: a meta-analysis. Rheumatol Int, 27 (9), 827-833, 2007.
[34] Begovich A. B., Carlton V. E., Honigberg L. A. et al. A missense single-nucleotide
polymorphism in a gene encoding a protein tyrosine phosphatase (PTPN22) is associated with
rheumatoid arthritis. Am J Hum Genet, 75 (2), 330-337, 2004.
[35] Plenge R. M., Padyukov L., Remmers E. F. et al. Replication of putative candidate-
gene associations with rheumatoid arthritis in >4,000 samples from North America and
Sweden: association of susceptibility with PTPN22, CTLA4, and PADI4. Am J Hum Genet,
77 (6), 1044-1060, 2005.
[36] Kurreeman F. A., Padyukov L., Marques R. B. et al. A candidate gene approach
identifies the TRAF1/C5 region as a risk factor for rheumatoid arthritis. PLoS Med, 4
(9),1515-1524, 2007.
[37] Remmers E. F., Plenge R. M., Lee A. T. et al. STAT4 and the risk of rheumatoid
arthritis and systemic lupus erythematosus. N Engl J Med, 357 (10), 977-986, 2007.

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
56
[38] The Wellcome Trust Case Control Consortium. Genome-wide association study of
14,000 cases of seven common diseases and 3,000 shared controls. Nature, 447, 661-678,
2007.
[39] Thomson W., Bardon A., Ke X., Eyre S., et al. Rheumatoid arthritis association at
6q23. Nat Genet., 39 (12), 1431-1433, 2007.
[40] Plenge R. M., Seielstad M., Padyukov L. et al. TRAF1-C5 as a risk locus for
rheumatoid arthritis--a genomewide study. N Engl J Med, 357 (12), 1199-1209, 2007.
[41] Gregersen P. K., Amos C. I., Lee A. T., Lu Y., et al. REL, encoding a member of the
NF-kappaB family of transcription factors, is a newly defined risk locus for rheumatoid
arthritis. Nat Genet., 41 (7), 820-823, 2009.
[42] Kochi Y., Okada Y., Suzuki A., Ikari K., et al. A regulatory variant in CCR6 is
associated with rheumatoid arthritis susceptibility. Nat Genet., 42 (6), 515-519, 2010.
[43] Ballina J. A., Canete J., et al. Genome-wide association study of rheumatoid arthritis
in the Spanish population: KLF12 as a risk locus for rheumatoid arthrits susceptibility.
Arthritis Rheum., 58 (8), 2275-2286, 2008.
[44] Raychaudhuri S., Remmers E. F., Lee A. T. et al. Common variants at CD40 and other
loci confer risk of rheumatoid arthritis. Nat Genet, 40 (10), 1216-1223, 2008.
[45] Stahl E. A., Raychaudhuri S., Remmers E. F., Eyre S., et al. Genome-wide association
study meta-analysis identifies seven new rheumatoid arthritis risk loci. Nat Genet., 42 (6),
508-514, 2010.
[46] Okada Y., Terao C., Ikari K., Kochi Y., et al. Meta-analysis identifies nine new loci
associated with rheumatoid arthritis in the Japanese population. Nat Genet., 44 (5), 511-516,
2012.
[47] Okada Y., Wu D., Trynka G., et al. Genetics of rheumatoid arthritis contributes to
biology and drug discovery. Nature, 506 (7488), 376-381, 2014.
[48] Eyre S., Bowes J., Diogo D., Lee A., et al. High-density genetic mapping identifies
new susceptibility loci for rheumatoid arthritis. Nat Genet., 44 (12), 1336-1340, 2012.
[49] Kim K., Bang S. Y., Lee H.-S., Cho S.-K., et al. High density genotyping of immune
loci in Koreans and Europeans identifies eight new rheumatoid arthritis risk loci. Ann Rheum
Dis., 74 (3), 1-17, 2015.
[50] Stahl E. A., Wegmann D., Trynka G. Gutierrez-Achury J., et al. Bayesian inference
analyses of the polygenic architecture of rheumatoid arthritis. Nat Genet., 44 (5), 483-489,
2012.
[51] Kiezun A., Garimella K., Do R., Stitziel N. O., et al. Exome sequencing and the genetic
basis of complex traits. Nat Genet., 44 (6), 623-630, 2012.
Zdravljenje
[52] Smolen J. S., Landewe R., Breedveld F. C., Buch M., Burmester G., Dougados M., et
al. EULAR recommendations for the management of rheumatoid arthritis with synthetic and
biological disease-modifying antirheumatic drugs. Ann Rheum Dis., 73 (3), 492-509, 2014.
[53] Ipavec M., Simonišek U., Pibernik N., Saje A. Revmatoidni artritis: zbornik predavanj.
Zdravstveni TIM, 1-52, 2011.
[54] van Vollenhoven R. F. Treatment of rheumatoid arthritis: state of the art 2009. Nat Rev
Rheumatol., 5 (10), 531-541, 2009.
[55] Ostrovršnik J., Tomšič M. Sodobno zdravljenje revmatoidnega artritisa. Rehabilitacija,
8 (1), 157-162, 2014.

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
57
[56] Bennett J. S., Daugherty A., Herrington D., Greenland P., Roberts H., Taubert K. A.
The use of nonsteroidal antiinflammatory drugs (NSAIDs): a science advisory from the
American Heart Association. Circulation, 111 (13), 1713-1716, 2005.
[57] Coxib, traditional NTC, Bhala N., Emberson J., Merhi A., et al. Vascular and upper
gastrointestinal effects of non-steroidal anti-inflammatory drugs: meta-analyses of individual
participant data from randomised trials. Lancet, 382 (9894), 769-779, 2013.
[58] Kirwan J. R., Bijlsma J. W., Boers M., Shea B. J. Effects of glucocorticoids on
radiological progression in rheumatoid arthritis. Cochrane Database Syst Rev., 1, CD006356,
2007.
[59] Jones G., Sebba A., Gu J., Lowenstein M. B., Calvo A., Gomez-Reino J. J., et al.
Comparison of tocilizumab monotherapy versus methotrexate monotherapy in patients with
moderate to severe rheumatoid arthritis: the AMBITION study. Ann Rheum Dis, 69 (1), 88-
96, 2010.
[60] Holt K. A Review of Pathophysiology and Medicinal Treatment Options of
Rheumatoid Arthritis. PharmCon, 1-21, 2011.
[61] http://www.geneplanet.si/genetika/slovarcek-genetskih-pojmov/genotipizacija/
(dostop 25. 5. 2015)
[62] http://ghr.nlm.nih.gov/gene/CCR6 (dostop 3. 6. 2015)
[63] http://ghr.nlm.nih.gov/gene/STAT4 (dostop 3. 6. 2015)
[64] Wurster A. L, Tanaka T., Grusby M. J. The biology of Stat4 and Stat6. Oncogene, 19
(21), 2577-2584, 2000.
[65] Kobayashi S., Ikari K., Kaneko K., et al. Association of STAT4 with susceptibility to
rheumatoid arthritis and systemic lupus erythematosus in the Japanese population. Arthritis
Rheum, 58 (7), 1940-1946, 2008.
[66] Zervou M. I., Mamoulakis D., Panierakis C., Boumpas D. T., Goulielmos G. N.
STAT4: a risk factor for type 1 diabetes? Hum Immunol, 69 (10), 647-650, 2008.
[67] Zheng J., Yin J., Huang R., Petersen F., Yu X. Meta-analysis reveals an association of
STAT4 polymorphisms with systemic autoimmune disorders and anti-dsDNA antibody. Hum
Immunol, 74 (8), 986-992, 2013.
[68] Kedra D., Pan H. Q., Seroussi E., Fransson I., et al. Characterization of the human
synaptogyrin gene family. Hum Genet, 103 (2), 131-141, 1998.
[69] http://www.ncbi.nlm.nih.gov/gene/9145 (dostop 3. 6. 2015)
[70] Latropoulos P., Gardella R., Valsecchi P., Magri C., et al. Association study and
mutational screening of SYNGR1 as a candidate susceptibility gene for schizophrenia.
Psychiatr Genet, 19 (5), 237-243, 2009.
[71] Verma R., Kubendran S., Das S. K., Jain S., et al. SYNGR1 is associated with
schizophrenia and bipolar disorder in southern India. J Hum Genet., 50 (12), 635-640, 2005.
[72] http://www.genecards.org/cgi-bin/carddisp.pl?gene=SYNGR1 (dostop 3. 6. 2015)
[73] http://www.genecards.org/cgi-bin/carddisp.pl?gene=PTPN22 (dostop 3. 6. 2015)
[74] Zheng J., Ibrahim S., Petersen F., Yu X. Meta-analysis reveals an association of
PTPN22 C1858T with autoimmune diseases, which depends on the localization of the affected
tissue. Genes Immun, 13 (8), 641-652, 2012.
[75] http://www.genecards.org/cgi-bin/carddisp.pl?gene=CTLA4 (dostop 3. 6. 2015)
[76] Foundation Jean Dausset. CEPH. http://www.cephb.fr/en/cephdb/ (dostop 7. 6. 2015)

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
58
[77] Lo Y. M. D., Chiu R. W. K., Chan K. C A. Clinical Applications of PCR, second
edition. New Jersey: Humana Press Inc., 2006.
[78] Boyer R. Temelji biokemije. Ljubljana: Študentska založba, 2005.
[79] Strachan T., Read A. P. Human molecular genetics 3, 3. edition. New York: Garland
Publishing, 2004.
[80] http://scienceinfoworld.blogspot.com/2012/11/polymerase-chain-reaction-pcr.htm
(dostop 25. 5. 2015)
[81] http://openwetware.org/wiki/CH391L/S12/PCR_and_advanced_PCR_techniques
(dostop 25. 5. 2015)
[82] Gormus U., Selvi N., Yaylim-Eraltan I. PCR-RFLP and Real-Time PCR Techniques
in Molecular Cancer Investigations. InTech, 2012.
[83] Hartl D. L., Jones E. W. Genetics: Analysis of Genes and Genomes, fifth edition. USA:
2001.
[84] https://dna.utah.edu/Hi-Res/TOP_Hi-Res%20Melting.html (dostop 25. 5. 2015)
[85] Applied biosystems. A Guide to High Resolution Melting (HRM) Analysis. USA:
2010.
[86] http://www.gene-quantification.de/hrm-dyes.html (dostop 7. 6. 2015)
[87] Erali M., Voelkerding K. V., Wittwe C. T. High Resolution Melting Applications for
Clinical Laboratory Medicine. Exp Mol Pathol., 85 (1), 50-58, 2008.
[88] Lascombe N. M., Greenbaum D., Gerstein M, et al. What is bioinformatics? A
proposed Definition and Overview of the Field. Method Inform Med, 40, 346-358, 2001.
[89] Sharma N., Kumar D., Investigating HMM in Protein Profile Analysis. UNIASCIT, 2
(1), 141-146, 2012.
[90] Mohammed J. Machine Learning in Bioinformatics. International Journal of Advanced
Research, 2 (12), 545-549, 2014.
[91] http://www.uniassignment.com/essay-samples/biology/bioinformatics-is-a-sub-field-
biology-essay.php (dostop 1. 6. 2015)
[92] http://primerssr.blogspot.com/2011/04/generunner.html (dostop 1. 6. 2015)
[93] http://www.ncbi.nlm.nih.gov/projects/SNP/snp_ref.cgi?rs=1571878 (dostop 1. 6.
2015)
[94] http://www.ncbi.nlm.nih.gov/projects/SNP/snp_ref.cgi?rs=909685 (dostop 1. 6. 2015)
[95] http://www.ncbi.nlm.nih.gov/projects/SNP/snp_ref.cgi?rs=2476601 (dostop 1. 6.
2015)
[96] http://www.ncbi.nlm.nih.gov/projects/SNP/snp_ref.cgi?rs=3087243 (dostop 1. 6.
2015)
[97] Fodil M., Benzaoui A., et al. Association of PTPN22 (rs2476601) and STAT4
(rs7574865) polymorphisms with Rheumatoid Arthritis in the Western Algerian population.
Acta Reumatol port., 40 (1), 56-62, 2015.
[98] Orozco G., Viatte S., Bowes J. Novel Rheumatoid Athritis Susceptibility Locus at
22g12 Identified in an Extended UK Genome Wide Association Study. Arthritis Rheumatol.,
66 (1), 24-30, 2014.
[99] Cheng P., Zhang Y., Huang H., et al. Association between CCR6 and rheumatoid
arthritis: a meta-analysis. Int J Clin Exp Med., 8 (4), 5388-5396, 2015.
[100] Jiang L., Yin J., Ye L., et al. Novel risk loci for rheumatoid arthritis in han chinese and
congruence with risk variants in europeans. Arthritis Rheumatol.;66 (5), 1121–1132, 2014

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
59
8 Življenjepis
OSEBNI PODATKI Gselman Doroteja
Na hribu 26, 2319 Poljčane (Slovenija)
+386 31228607
DELOVNE IZKUŠNJE
IZOBRAŽEVANJE IN USPOSABLJANJE
ŽELENO PODROČJE DELA Kemijski tehnolog, živilski tehnolog
24.3. 2014–v teku Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu (v okviru magistrskega dela)
Fakulteta za kemijo in kemjsko tehnologijo Maribor, Maribor (Slovenija)
Izolacija DNA iz vzorcev krvi, genotipizacija izbranih polimorfizov SNP s tehniko PCR-RFLP in HRM in statistična analiza s programom SPSS.
15.7. 2014–1.9. 2014 Priprava in analiza vzorcev (v okviru praktičnega usposabljanja)
Steklarna Rogaška d.d., Rogaška Slatina (Slovenija)
Analiza trdote hladilnih vod, priprava surovin (sejalna analiza, nasipna teža), določanje gostote stekla, določitev števila bakterij, pH in motnosti v vodah za hlajenje brusov in pomoč pri določanju izluževanja Pb iz steklenih površin.
18.1. 2012–27.6. 2012 Vpliv superkritičnega ogljikovega dioksida (SC CO2) na aktivnost zamreženih encimskih agregatov (CLEAs) iz celulaze (v okviru dela za diplomo)
Fakulteta za kemijo in kemijsko tehnologijo Maribor, Maribor (Slovenija)
Sinteza zamreženih encimskih agregatov (CLEAs) iz celulaze, izpostavljanje CLEAs superkritičnemu ogljikovemu dioksidu in merjenje aktivnosti CLEAs s pomočjo UV-Vis spektrofotometrom.
1.10. 2012–v teku Magistrica inženirka kemijske tehnike Raven 7 EOK
Fakulteta za kemijo in kemijsko tehnologijo, Maribor (Slovenija)
- Bioseparacijska tehnika in kataliza
- Industrijska mikrobiologija
- Procesne surovine za bio in prehrambeno industrijo
- Osnove tehnologije živilskih izdelkov
- Genomika v biomedicinski tehnologiji
- Encimske tehnologije
2009–2012 Diplomirana inženirka kemijske tehnologije (UN) Raven 6 EOK

Genotipizacija slovenskih bolnikov z revmatoidnim artritisom za DNA polimorfizme predhodno povezane z boleznijo v asociacijskih študijah v celotnem genomu
60
KOMPETENCE
Fakulteta za kemijo in kemijsko tehnologijo Maribor, Maribor (Slovenija)
- Splošna in anorganska kemija
- Analizna kemija
- Organska kemija
- Biokemijska tehnika-biotehnologija
- Biokemija in molekularna biologija
2004–2008 Gimnazijski maturant Raven 5 EOK
Prva gimnazija Maribor, Maribor (Slovenija)
Materni jezik slovenščina
Drugi jeziki RAZUMEVANJE GOVORJENJE PISNO SPOROČANJE
Slušno razumevanje Bralno razumevanje Govorno
sporazumevanje Govorno sporočanje
angleščina C1 C1 C1 C1 C1
nemščina A1 A1 A1 A1 A1
španščina B1 B1 A2 A2 A2
Stopnja: A1 in A2: Osnovni uporabnik - B1 in B2: Samostojni uporabnik - C1 in C2: Usposobljeni uporabnik Skupni evropski jezikovni okvir
Komunikacijske kompetence Komunikacijske veščine sem pridobila predvsem med delom v laboratoriju (med študijem), kjer smo bili razdeljeni v skupine.
Digitalna pismenost - Dobro poznavanje MS Office (Word, Excel, PowerPoint),
- delo v SPSS,
- elektronska pošta,
- spletni brskalniki.
Druge kompetence Vajena sem javnega nastopanja, se hitro učim, sposobna sem samostojnega dela kot tudi dela v skupini.
Vozniško dovoljenje B