induces insulinomas in hamsters
TRANSCRIPT

Vol. 32, No. 3JOURNAL OF VIROLOGY, Dec. 1979, p. 934-9420022-538X/79/12-0934/09$02.00/0
Viable Deletion Mutant of Human Papovavirus BK ThatInduces Insulinomas in Hamsters
SUMIE WATANABE,l KUNITO YOSHIIKE,l AKEMI NOZAWA,2 YASUHITO YUASA,' ANDSEIJIRO UCHIDA2
Department of Enteroviruses, National Institute ofHealth, Kamiosaki, Shinagawa-ku, Tokyo 141,' andDepartment of Viral Infection, Institute ofMedical Science, University of Tokyo, Tokyo 108,2 Japan
Received for publication 1 June 1979
A plaque morphology mutant (pm-522) of human papovavirus BK, which wasrescued from a human papovavirus BK-induced hamster pineocytoma, was char-acterized and compared with a cloned wild-type virus (wt-501). Mutant pm-522formed turbid plaques and grew more slowly than wt-501 in human embryonickidney (HEK) cells. The immunofluorescence assay revealed that more HEKcells underwent abortive infection withpm-522 than with wt-501. Whereas wt-501induced brain tumors and osteosarcomas, but no insulinomas, in hamsters, pm-522 induced brain tumors and insulinomas. The DNA of pm-522 was found byelectrophoresis and electron microscopy to have a deletion (85 ± 15 base pairs)and an insertion (40 ± 10 base pairs) between map coordinates 0.708 and 0.725from the endonuclease EcoRI cleavage site. These results demonstrate thepresence of a viable deletion human papovarivus BK mutant capable of inducinginsulinomas in hamsters.
Human papovavirus BK (BKV) was originallyisolated from the urine of a renal allograft recip-ient on immunosuppressive therapy (9). Thegenome ofBKV is a closed circular duplex DNAmolecule approximately 5,000 base pairs longand is considerably similar to the genome ofsimian virus 40 (SV40) in both nucleotide se-quence homology and genetic organization (5, 6,11, 12, 15, 16, 27, 28).Like SV40, BKV induces tumors in hamsters
(1-3, 7, 14, 18, 22, 24). We found that hamsters,inoculated intracerebrally soon after birth witha family of BKV samples originating from asingle stock of Gardner's original strain, devel-oped choroid plexus papillomas, malignant in-sulinomas, and osteosarcomas at high frequen-cies, each differing among the virus samples (21).For instance, the samples could be classified byincidence of insulinoma into two groups, thepotent and poor inducers of that tumor. Fromthe relationship between those samples in thefamily line, we concluded that there must existBKV mutants differing in capacity to inducetumors of the specific types, especially insuli-noma.
In the present study we have characterized anaturally occurring plaque morphology mutant(pm-522), which had been rescued from a BKV-induced hamster pineocytoma (21), comparingit with a cloned wild type (wt-501). Mutantpm-522 was found to be a viable deletion mutantcapable of inducing insulinomas in hamsters,
whereas the cloned wild type lacked that capac-ity. These results strongly support the hypoth-esis that the polyoncogenic capacity of BKV isaccounted for by the presence of BKV mutants.
MATERIALS AND METHODSCells. Human embryonic kidney (HEK) cell cul-
tures were used at passage 4 for propagation and assayof plaque-forming and antigen-forming activities ofBKV. The preparation of cell cultures from kidneysand conditions for cultivation were as described pre-viously (21, 22).
Viruses and their purification. Two clones ofBKV (wt-501 and pm-522) and one uncloned BKVstock of Gardner's original strain were used. The pas-sage history of their parental stocks and their mutualrelationship in the passage lines were described pre-viously (21). The purification procedure of viruses wasas described previously (22), except that the concen-tration of sodium deoxycholate was lowered to 0.5%.
(i) Wild-type clone (wt-501). From unclonedBKV stock 308 (purified sample P12) that inducedinsulinoma at the highest incidence (21), wt-501 wascloned through one limiting-dilution passage, followedby two successive plaque isolations. The original stockwas prepared by infecting HEK cells with the lastisolate at an input multiplicity of infection (MOI) ofless than lo-4 PFU/cell. Working stocks 152 (purifiedsample P17) and 252 (purified sample P35) were ob-tained from cell cultures infected with the originalstock (5 x 10' PFU/cell) and stock 152 (5 x 10-5PFU/cell), respectively.
(ii) Plaque morphology mutant (pm-522). BKVhad been rescued by the cell fusion technique, afterprolonged cultivation of the heterokaryon cultures,
934
on April 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

BKV MUT'ANT' IND)UCING HAMSTElR INSULINOMAS 935
from cell line Pc13 originating from a hamster pineo-cytoma. This tumor had been induced by virus sampleP3, which had also induced insulinomas at a highincidence (21). Mutant pm-522 was cloned by threesuccessive plaque isolations from a rescued virus stock.The original stock was prepared in the same way asdone for wt-501. Working stocks 154 (purified sampleP24) and 155 (purified sample P36) were prepared byinfection with the original stock at an MOI of 5 x 10-4PFU/cell.
(iii) Uncloned virus. Uncloned BKV stock 412(purified sample P23) was prepared by infection ofcells with purified P3 at an input MOI of 0.03 PFU/cell. Sample P23 was used for comparison in the testof tumorigenicity.
Infectivity and antigen-forming activities. In-fectivity was determined by the plaque method asdescribed previously (23), with the following modifi-cations: the adsorption period was for 2 h at 36°C;1.5% agar medium was composed of Eagle minimumessential medium containing 0.1% yeast extract, 0.25%lactalbumin hydrolysate, 1% bovine serum, and 5%heat-inactivated horse serum; and neutral red wasadded on day 13. V- and T-antigen-forming activitieswere measured by the indirect immunofluorescent as-say essentially as described previously (21, 22). Coverslip HEK cultures (1.2 x 3.2 cm2) in small bottles (1.4x 3.8 cm2) were prepared from the ceL's stored at-80°C at passage 3 so as to form monolayers 24 hlater. Monolayer cultures were washed with Dulbeccophosphate-buffered saline, inoculated with 0.7 ml (theeffective dose of inoculum was assumed to be 0.07 ml)of appropriately diluted virus, incubated at 36°C for 6h for virus adsorption, washed twice with phosphate-buffered saline, and then cultivated in the mediumcontaining anti-BKV guinea pig serum. At 4 days afterinfection, at which time the number of positive cellsreaches the maximum, the infected cultures were fixedin precooled (-20°C) acetone for 10 min, incubatedfor 45 min at 360C with either hamster serum againstSV40 or BKV T-antigen or hamster serum againstBKV V-antigen, and stained with fluorescent rabbitanti-hamster globulin (Miles Laboratories, Inc.) for 30min at 36°C. Stained cells were counted under a NikonUV microscope equipped with an FITC interferencefilter. Cell-infecting units were calculated from thenumber of positives, the dilution factor, and the effec-tive dose of inoculum (26).
Tumorigenicity. Syrian golden hamsters main-tained in the Department of Veterinary Science, Na-tional Institute of Health, Tokyo (22), were used.Newborn hamsters, fewer than 24 h old, were eachinoculated with 0.02 ml of purified BKV into themidportion of the right cerebral hemisphere and ob-served for 8 months. For comparison, some animalsreceived a sample prepared from uninfected HEK cellsin the same way as virus purification. Hamsters wereautopsied when they showed abnormality, and tumorswere examined histopathologically. T-antigen was ex-amined with tumor imprints by the indirect immuno-fluorescent test (22).
Preparation of viral DNA. BVK DNA was ex-tracted from purified virions and separated into twofractions, supercoiled closed circular DNA (DNA I)and nicked open circular DNA (DNA II), as describedpreviously (30).
Digestion of DNA with restriction endonucle-ases. Endonucleases HindIII (cleaving BKV DNA atfour sites) and EcoRI (cleaving BKV DNA once) werepurchased from Boehringer Mannheim Corp. andMiles Laboratories, Inc., respectively. DNA I was di-gested according to the protocols accompanying theenzymes. EcoRI-cleaved linear molecules were puri-fied as described previously (30).
Electrophoresis. Agarose gel electrophoresis wasperformed as described by Sugden et al. (20). Ethidiumbromide (0.5 ,ug/ml) was included in the running Tris-acetate buffer and 1.4% agarose slab gels. After run-ning, DNA bands were visualized on a short-wave-length UV transilluminator (Ultra-Violet Products)and photographed through a Nikon R60 filter withNEOPAN 400 135 film.
Electron microscopy. The formamide techniqueof Davis et al. (4) was essentially followed. BKV DNAII was mixed with human wart virus DNA II, whichwas used as an internal reference (29), for a lengthdetermination of the whole BKV genome. Micro-graphs from a limited area (a single-mesh hole) wereused for a determination of the length distributionpattern (29). The conditions for the heteroduplexmethod were as described previously (30), except gridswere filmed with carbon instead of Parlodion. Thecalculated effective temperatures for reannealing andmounting were Tm - 29°C and Tm - 20°C, respec-tively. Contour lengths of DNA were measured onprojected images with a Numonics digital length cal-culator at a final magnification of x200,000. Single-stranded DNA was longer by 10% than the correspond-ing double-stranded DNA. Correction was made ac-cordingly.
RESULTSGrowth and yield. Virus rescued from a
BKV-induced hamster pineocytoma and itsclonal derivative pm-522 formed turbid plaqueson HEK cells (Fig. 1). The formation of turbidplaques appears to indicate that some infectedcells stay alive for awhile within a plaque formedby pm-522. Plaques formed by pm-522 began toappear slightly later than those formed by wild-type virus (wt-501), but the numbers of plaquesformed by the two viruses reached the maximumat about day 20, at which time the diameter ofpm-522 plaques (1 to 4 mm) was about half thatof wt-501 plaques (2 to 7 mm). Plaque formationby the two viruses followed one-hit kinetics.
In a low-multiplicity infection, pm-522 multi-plied more slowly than wt-501. When HEK cellswere infected with the original stock of wt-501at an MOI of 5 x 10-5 PFU/cell, they showedthe first sign of cytopathic effects on day 15postinfection and complete cytopathic effects onday 26. HEK cells, infected with the pm-522original stock under the same conditions, startedto show cytopathic effects on day 21 and werecompletely destroyed on day 33. The yields ofthe resulting wt-501 and pm-522 stocks were 1.2x 109 and 4 x 108 PFU/150-cm2 glass cultureflask, respectively.
Voi,. 32>, 1979
on April 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

936 WAT'ANABE El' AL.
Although the yield of pm-522 in terms of in-fectivity was one-third that of wt-501, the yieldsof the two viruses in terms of physical particleswere approximately the same. As shown in Ta-ble 1, the calculated yield for two purified wt-501 samples (P17 and P35) and two purifiedpm-522 samples (P24 and P36) was about 4 x 1012virions per 150-cm2 culture. The specific infec-tivity (PFU per virions) of the pm-522 sampleswas less than half that of wt-501.Antigen formation in HEK cells. Using the
indirect immunofluorescent assay, we comparedT- and V-antigen formations between wt-501and pm-522 in low-multiplicity infections ofHEK cells. As shown in Fig. 2, there were twoclasses of T-antigen-positive cells; T-fluorescentgranules were densely distributed in nuclei inone type and sparsely distributed in the other.Although there was some variation in the degreeof fluorescence within each type, the two typeswere easily distinguishable at a magnification of
pm522 ~d.w t 501C
FIG. 1. Plaque morphologies of BKVpm-522 and
wt-501. The viruses were each plaqued on a mono-
layer ofHEK cells for 19 days at 360C. The plaques
were visualized by neutral red staining of the cells
on day 13.
10 x 20 as strongly and weakly fluorescent nu-
clei, respectively, except for an occasional occur-
rence of an intermediate type. The proportionof sparsely stained T-antigen positives was muchlarger in the infection withpm-522 than with wt-501. As the numbers of T- and V-antigen-posi-tive cells did not increase much after day 4, a
reading was made on that day for titration ofantigen-forming activities.
Figure 3 shows examples of a dose-responserelationship. T- and V-antigen formations in in-fections with wt-501 and pm-522 showed one-hit
FIG. 2. Immunofluorescent staining of HEK cellsinfected with pm-522 with sera from BKV tumor-bearing hamsters and fluorescein-labeled anti-ham-ster immunogloblin. HEK cells on a cover slip were
infected with pm-522 (for demonstration, at an MOIof 3 PFU/cell) and processed for immunofluorescentmicroscopy on day 4 after infection. One denselystained (D) and six sparsely stained (S) T-antigen-positive cells are shown here. x1,200.
TABLE 1. Yields and specific activities ofpurified BKV samples
Virus Yield per 150-cm2 culture Sp act (ClUb/105 virions)
Virions T-antigen- Dense-T-anti- V-antigen-Clone Sample rl1oI2 PFU x 10" forming activi- gen-forrning forming activi-
ties activities ties
wt-501 P17 4.9 2.7 2.0 1.4 1.3P35 3.7 1.7 2.1 1.2 1.0
pm-522 P24 4.1 1.0 3.0 0.6 0.5P36 4.3 0.8 2.2 0.5 0.3
a Estimated from the following: 1 optical density unit at 258 nm = 6.4 X 1012 virions.b CIU, Cell-infecting unit (26).
,J. V! XRolI
on April 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
BKV MUT'ANT' INI)UCING HAMST'ER INSULINOMAS 937
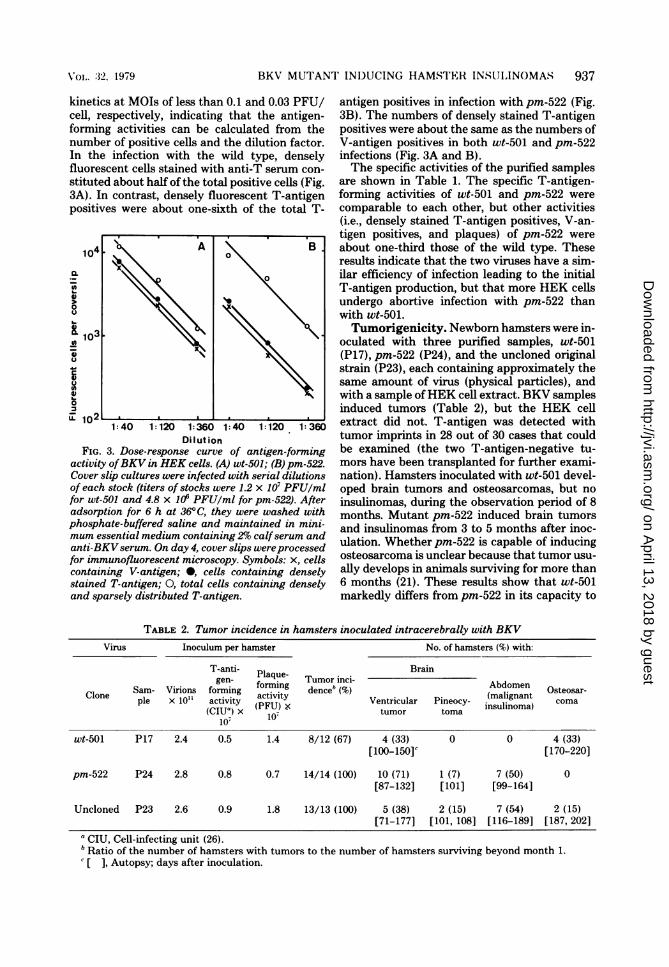
kinetics at MOIs of less than 0.1 and 0.03 PFU/cell, respectively, indicating that the antigen-forming activities can be calculated from thenumber of positive cells and the dilution factor.In the infection with the wild type, denselyfluorescent cells stained with anti-T serum con-
stituted about half of the total positive cells (Fig.3A). In contrast, densely fluorescent T-antigenpositives were about one-sixth of the total T-
104. A B
0~~~~~~~~~~
ui11A~~~~~~~~S
0
IL 1021 . I1:40 1:120 1:360 1:40 1:120 1:360
DilIutionFIG. 3. Dose-response curve of antigen-forming
activity ofBKV in HEK cells. (A) wt-501; (B) pm-522.Cover slip cultures were infected with serial dilutionsof each stock (titers of stocks were 1.2 x 107 PFU/mlfor wt-501 and 4.8 x 10' PFU/ml for pm-522). Afteradsorption for 6 h at 36°C, they were washed withphosphate-buffered saline and maintained in mini-mum essential medium containing 29,o calfserum andanti-BKVserum. On day 4, cover slips wereprocessedfor immunofluorescent microscopy. Symbols: x, cellscontaining V-antigen; *, cells containing denselystained T-antigen; O, total cells containing denselyand sparsely distributed T-antigen.
antigen positives in infection with pm-522 (Fig.3B). The numbers of densely stained T-antigenpositives were about the same as the numbers ofV-antigen positives in both wt-501 and pm-522infections (Fig. 3A and B).The specific activities of the purified samples
are shown in Table 1. The specific T-antigen-forming activities of wt-501 and pm-522 were
comparable to each other, but other activities(i.e., densely stained T-antigen positives, V-an-tigen positives, and plaques) of pm-522 were
about one-third those of the wild type. Theseresults indicate that the two viruses have a sim-ilar efficiency of infection leading to the initialT-antigen production, but that more HEK cellsundergo abortive infection with pm-522 thanwith wt-501.Tumorigenicity. Newborn hamsters were in-
oculated with three purified samples, wt-501(P17), pm-522 (P24), and the uncloned originalstrain (P23), each containing approximately thesame amount of virus (physical particles), andwith a sample ofHEK cell extract. BKV samplesinduced tumors (Table 2), but the HEK cellextract did not. T-antigen was detected withtumor imprints in 28 out of 30 cases that couldbe examined (the two T-antigen-negative tu-mors have been transplanted for further exami-nation). Hamsters inoculated with wt-501 devel-oped brain tumors and osteosarcomas, but no
insulinomas, during the observation period of 8months. Mutant pm-522 induced brain tumorsand insulinomas from 3 to 5 months after inoc-ulation. Whether pm-522 is capable of inducingosteosarcoma is unclear because that tumor usu-
ally develops in animals surviving for more than6 months (21). These results show that wt-501markedly differs from pm-522 in its capacity to
TABLE 2. Tumor incidence in hamsters inoculated intracerebrally with BKVVirus Inoculum per hamster No. of hamsters (%) with:
T-anti- Brain
Sam- Virions forming orminueg Tumor ici- Abdomen Osteosar-Clone Sam Viin fomnrm.acingt dece (malignantCleple x ll activity activity Ventricular Pineocy- inslignoa) coma(CIU") x (PFU- tumor toma nsulinoma)
10'
wt-501 P17 2.4 0.5 1.4 8/12 (67) 4 (33) 0 0 4 (33)[100-150]c [170-220]
pm-522 P24 2.8 0.8 0.7 14/14 (100) 10 (71) 1 (7) 7 (50) 0[87-132] [101] [99-164]
Uncloned P23 2.6 0.9 1.8 13/13 (100) 5 (38) 2 (15) 7 (54) 2 (15)[71-177] [101, 108] [116-189] [187, 202]
aCIU, Cell-infecting unit (26).b Ratio of the number of hamsters with tumors to the number of hamsters surviving beyond month 1.' [ ], Autopsy; days after inoculation.
Voi,. :32, 1979
on April 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

938 WAT'ANABE El AL.
induce insulinomas. Hamsters inoculated withuncloned virus P23 developed brain tumors, in-sulinomas, and osteosarcomas. The types of tu-mors induced by P23 and the time when theywere detected resembled those obtained withthe similar uncloned samples (21). It should benoted that pm-522 induced tumors earlier thanwt-501 and uncloned sample P23 did. During thefirst 4 months of the 8-month observation pe-riod, 11 out of 14 hamsters inoculated with pm-522 developed tumors, and we found 14 out of18 tumors observed throughout this test. Duringthe same 4-month period, wt-501 and P23 in-duced 2 out of 8 and 5 out of 16 tumors found inthe test of 8 months, respectively.Homogeneity of viral DNA. When DNA I
or EcoRI-cleaved linear DNA of pm-522 was
subjected to agarose gel electrophoresis, a singleDNA band was observed which moved slightlyfaster than the corresponding wild-type DNA(data not shown). A slight difference in DNAmass was confirmed by a length determinationof DNA II, relative to human wart virus DNA.When the mean length of 75 wt-501 moleculeswas assumed to be 1.000 ± 0.013 (standard de-viation [SD]), the mean for 65pm-522 moleculeswas 0.990 ± 0.022 (SD) and reference humanwart virus DNA had a mean length of 1.507,with SDs of 0.024 and 0.021, respectively, for thetwo determinations (25 molecules). The lengthdistribution patterns of wt-501 and pm-522DNAs showed that, as indicated by their smallSDs, their DNAs are as homogeneous in size asnondefective SV40 DNA (29). The difference inDNA mass between the two DNAs was esti-mated to be about 50 base pairs.Location of mutation on DNA map. DNA
I of wt-501 and DNA I ofpm-522 were digestedwith HindIlI and subjected to electrophoresison a 1.4% agarose gel (Fig. 4). As did Howley etal. (10), we found that the wild type generatedfour fragments, A, B, C, and D. In contrast, theC fragment of pm-522 migrated faster than thecorresponding fragment of the wild type, justbehind the D fragment. This result indicatesthat pm-522 DNA contains a small deletionwithin the C fragment defined by EcoRI map
positions 0.62 to 0.725 (6) or 0.615 to 0.72 (8). Ifthe C and D fragments are 525 and 475 basepairs long, respectively (8), the deletion ofpm-522 is about, but slightly fewer than, 50 basepairs.We used the electron microscope heterodu-
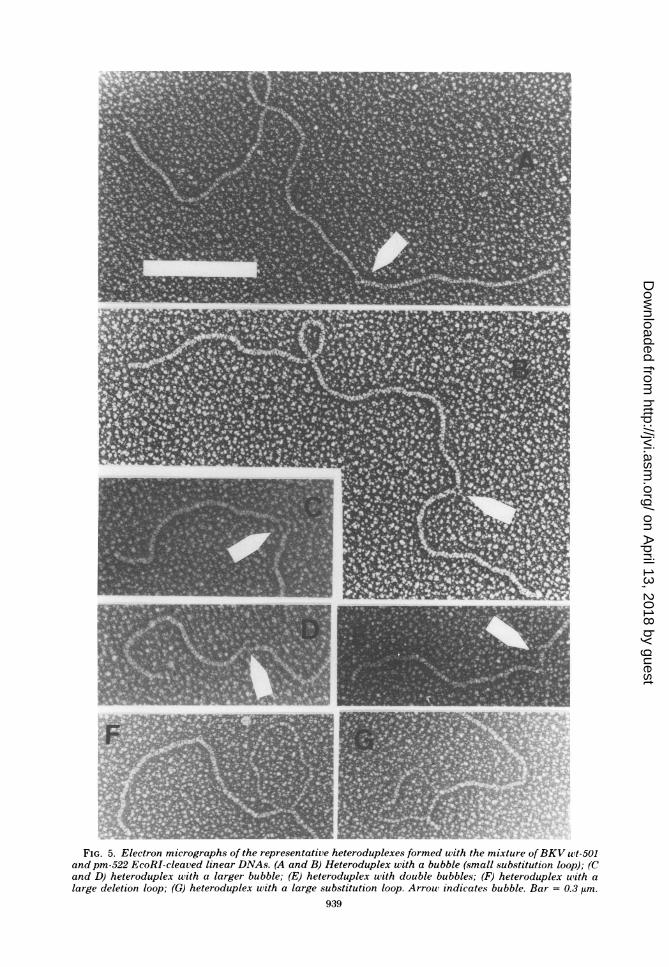
plex method in an attempt to map the deletionof pm-522 more precisely. The mixture (1:1) ofwt-501 and pm-522 EcoRI-cleaved linear DNAswas denatured and annealed for heteroduplexformation. For comparison, each DNA alone wassubjected to the same procedure. While the grids
.J. ViR0o1.
were being scanned, the clean homoduplexes(double stranded linear) were counted, and theheteroduplexes (molecules with single-strandedsegments) were photographed and, later, meas-ured and interpreted. The results are summa-rized in Table 3, and the representative hetero-duplexes are shown in Fig. 5. Heteroduplexeswere found most frequently with the mixture. Of45 heteroduplexes, 42 had bubbles (small substi-tution loops) at a seemingly fixed position fromone end as shown in Fig. 5A to E. The remaining
FIG. 4. Cleavage ofBKVwt-501 andpm-522 DNAs
with endonuclease HindIIIL DNA (0.4 gig) was di-
gested with HindIII (12.5 U) and subjected to electro-
phoresis on a 1.4% agarose slab gel for 4 h at V.
After running, DNA-ethidium bromide bands were
visualized and photographed on a short-wavelengthUV transilluminator.
TABLE 3. Proportion of heteroduplexes
Determination fMixture(t1 wt-501 pm-522and pm-522 alone alone
Clean homoduplex 103 (70)" 378 (95) 463 (92)Clean heteroduplex 45 (30) 20 (5) 40 (8)With bubble' 42 0 1With larger loop' 3 20 39
aNumbers within parentheses indicate percentages.As shown in Fig. 5A to E.
'As shown in Fig. 5F and G.
on April 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
F *.1
F. rc,
f)t
FIG. 5. Electron micrographs of the representative heteroduplexes formed with the mixture ofBKV uwt-501and pm-522 EcoRI-cleaved linear DNAs. (A and B) Heteroduplex with a bubble (small substitution loop); (Cand D) heteroduplex with a larger bubble; (E) heteroduplex with double bubbles; (F) heteroduplex with alarge deletion loop; (G) heteroduplex with a large substitution loop. Arrow indicates bubble. Bar = 0.3 ,im.
939
on April 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

940 WATIANABE El' AL.
three heteroduplexes had a large deletion orsubstitution loop (Fig. 5F and G). The hetero-duplexes found with wt-501 or pm-522 alonewere mostly those with larger loops, as shown inFig. 5F and G. As the heteroduplexes with abubble were found almost exclusively with themixture of wt-501 and pm-522, they were consid-ered to be hybrid molecules of the two DNAs.The proportion of these heteroduplexes (30%)was smaller than that expected theoretically(50%) of a 1:1 mixture. In practice, the smallbubbles apparently collapsed to form tiny nod-ules in some molecules. If found, those were,excluded from measurement, and some of themwould have been easily overlooked during scan-ning.The position and size of bubbles were deter-
mined from a length measurement of each seg-ment. Of 42 heteroduplexes that were consideredto be hybrids between wt-501 andpm-522, 2 witha larger bubble (Fig. 5C and D) and 2 withdouble bubbles (gig. 5E) were not included inthe calculation. The mean fractional length ofeach segment in 38 molecules with a bubble (Fig.5A and B) was as follows: the longer double-stranded segment, 0.708 ± 0.006 (SD); theshorter double-stranded segment, 0.275 + 0.005(SD); the longer strand of bubble, 0.017 ± 0.003(SD); and the shorter strand of bubble, 0.008 +0.002 (SD). As the deletion ofpm-522 was withinHindIII C fragment, it was located between mapcoordinates 0.708 and 0.725 from the EcoRIcleavage site. The size of the deletion estimatedhere was 85 ± 15 base pairs. The DNA of pm-522 was found to have an insertion (equivalentto the shorter strand of bubble) of 40 ± 10 basepairs at the site of deletion.Table 3 also shows that DNAs of wt-501 and
pm-522 are homogeneous with respect to basesequences. The proportion of heteroduplexes toclean homoduplexes found with the two DNAswas similar to that found with various nondefec-tive SV40 DNA samples prepared by low-mul-tiplicity infection (K. Yoshiike, unpublished ob-servations).
DISCUSSIONIn the present study we found that BKV mu-
tant pm-522 is a viable virus having a smalldeletion and an insertion at about 0.72 map unit.The nature of the inserted sequence is not clearyet. If the genomic organization of BKV is iden-tical with that of SV40, the deletion-insertion ofpm-522 lies in the leader segment for the latemRNA (17, 25). Several SV40 mutants havingsmall deletions in a similar region have beenisolated or constructed by the biochemicalmethod; they were reported to grow somewhat
more slowly and poorly and to form smallerplaques in monkey cells than did wild-type SV40(13, 19). Some of them were shown to havedecreased 19S late mRNA synthesis (relative to16S RNA) in the productive infection (25). Thepresent BKV mutant pm-522 formed smallerplaques and grew more poorly than the wildtype. Although the yield ofpm-522 expressed asinfectivity was about one-third that of the wildtype, there was no difference in yield expressedas physical particles between the two viruses.Whether pm-522 mutation affects mRNA syn-thesis remains to be determined.
Abortive infection of HEK cells appears to becharacteristic of BKV pm-522. The formation ofturbid plaques suggests that productive infectionis not completed in some infected cells withinplaques. The high ratio of T-antigen positives(sparse and dense) to V-antigen positives ordensely fluorescent T-antigen positives (Fig. 3and Table 1) also shows the abortive infectionwith pm-522. These results can be interpreted toindicate either (i) that in a low-multiplicity in-fection, the efficiency of pm-522 infection of asingle HEK cell leading to the production ofvirions is about one-third that of the wild type(Figure 3 may indicate that one out of six T-antigen positive cells, each infected singly witha pm-522 virus, produces virions in the sameamount as does the wild type, whereas one outof two cells does so in the wild-type infection.Which of the infected cells can complete theproductive infection is determined by chance.The cells that have survived the initial abortiveinfection will be reinfected later. Eventually, theentire monolayer culture will be destroyed,yielding a comparable amount of virus particlesper culture between pm-522 and wt-501.) or (ii)that pm-522, even grown at a low MOI, alwayscontains a large amount of defective viruses(Figure 3 may indicate that five out of six pm-522 particles are defective and unable to producevirions, whereas one out of two wt-501 particlesis similarly defective). The mutant DNA is as-sumed to be genetically and physically homo-geneous in the first hypothesis and heteroge-neous, as found with SV40 defective DNA (31),in the second. At present we favor the firsthypothesis and consider pm-522 to be a homo-geneous population having a lower frequency(about one-third of the wild type) of completingthe single cycle of productive infection in a low-multiplicity infection for the following reasons:(i) that pm-522 DNA was as homogeneous aswild type BKV and nondefective SV40 DNAswith respect to size and base sequence; (ii) thatsome HEK cells were abortively infected withina plaque presumably initiated by a single viable
TJ Vi{(lo'
on April 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

BKV MUT'ANT' NI)UCING HAMST'ER INSUIANOMAS 941
particle; and (iii) that the proportion of sparselyfluorescent T-antigen positive cells did notchange much among various samples (data notshown). It is possible that the appearance ofweakly fluorescent T-antigen positives in thewild-type infection results from a delayed reac-
tion of infected cells and virions are made inthem afterwards. It seems unlikely, however,that pm-522 just induces a slower cycle of infec-tion with a later appearance of strongly fluores-cent nuclei for T-antigen and V-antigen. If suchwere the case, there would be little difference inspecific plaque-forming activities between pm-522 and the wild type.
In the previous studies (21, 22) a series ofuncloned BKV samples was found to inducevarious hamster tumors, each at different fre-quencies among the samples. Those samplescould be classified as two groups, the one withhigh insulinoma-inducing capacity and the otherwith little such capacity. In the present study weincluded one uncloned sample (P23), whose pa-
rental stock belonged to the insulinoma-inducinggroup, in the test of tumorigenicity. The resultsobtained with P23 were those expected of thisgroup, indicating that the present results withthe two cloned viruses can be directly comparedwith the previous ones. Wild-type 501, whichhad been isolated from an uncloned insulinoma-inducing stock, induced brain tumors and osteo-sarcomas but no insulinomas (Table 2). Thispattern resembles those obtained with unclonedsamples belonging to the group that induced fewinsulinomas. These results strongly suggest thatthe wild-type BKV lacks that capacity and thatthe uncloned samples used in the previous stud-ies were indeed the mixtures of mutants capableof inducing insulinomas and the wild type atdifferent ratios. A few more wild-type clones likewt-501 are now being tested for tumorigenicity.On the other hand, pm-522 showed an inter-
esting pattern of tumor induction. It inducedbrain tumors and insulinomas mostly between 3and 4 months after inoculation, significantly ear-lier than wt-501 (3 to 8 months) and the un-
cloned virus (3 to 7 months). Although pm-522grew somewhat more poorly than the wild typein HEK cells, its tumorigenic capacity appears
to be higher than that of the wild type. We needfurther detailed studies on the interaction be-tween pm-522 and HEK cells or hamster cells toanswer the question of how the mutation in theputative leader segment for late mRNA is re-
lated to the induction of insulinoma in hamstersand to fully understand the mechanism of thatinduction. It is also possible that, besides thedeletion-insertion, there are other differencesbetween pm-522 and wild-type genomes not de-
tected by the methods used in this study andthat one such change in pm-522 is responsiblefor its characteristic pattern of tumorigenesis.The present results, although they demon-
strate the presence of an insulinoma-inducingBKV mutant, do not rule out the possibility thatthe uncloned stocks contain other mutants ca-pable of inducing insulinomas or other specifictumors. In fact, we expect from the previousresults (21) that there are some defective mu-tants having different tumor-inducing spectra,for instance, a mutant capable of inducing osteo-sarcoma but no brain tumor. Attempts to isolatesuch mutants are now in progress.
ACKNOWLEDGMENTSWe thank Takeshi Muto, National Institute of Health,
Tokyo, for the diagnosis of tumors and Nobuo Yamaguchi,Institute of Medical Science, University of Tokyo, for criticalcomments on the manuscript. We are also indebted to IsamuTagaya, Department of Enteroviruses, National Institute ofHealth, Tokyo, for his support.
This study was supported in part by Grant-in-Aid forCancer Research 301019, 301028, 301038, and 301528 from theMinistry of Education, Science, and Culture, Japan (to S.W.,K.Y., and S.U.) aid by a grant from the Japanese Society forPromotion of Cancer Research (to S.U.).
LITERATURE CITED
1. Corallini, A., G. Altavilla, M. G. Cecchetti, G. Fabris,M. P. Grossi, P. G. Balboni, G. Lanza, and G.Barbanti-Brodano. 1978. Ependymomas, malignanttumors of pancreatic islets, and osteosarcomas inducedin hamsters by BK virus, a human papovavirus. J. Natl.Cancer Inst. 61:875-883.
2. Corallini, A., G. Barbanti-Brodano, W. Bortoloni, I.Nenci, E. Cassai, M. Tampieri, M. Portolani, andM. Borgatti. 1977. High incidence of ependymomasinduced by BK virus, a human papovavirus. J. Natl.Cancer Inst. 59:1561-1564.
3. Costa, J., C. Yee, T. S. Tralka, and A. S. Rabson.1976. Hamster ependymomas produced by intracerebralinoculation of a human papovavirus (MMV). J. Natl.Cancer Inst. 56:863-864.
4. Davis, R. W., M. Simon, and N. Davidson. 1971. Elec-tron microscope heteroduplex methods for mappingregions of base sequence homology in nucleic acids.Methods Enzymol. 21:413-428.
5. Dhar, R., C.-J. Lai, and G. Khoury. 1978. Nucleotidesequence of the DNA replication origin for humanpapovavirus BKV: sequence and structural homologywith SV40. Cell 13:345-358.
6. Dhar, R., I. Seif, and G. Khoury. 1979. Nucleotidesequence of the BK virus DNA segment encoding smallt antigen. Proc. Natl. Acad. Sci. U.S.A. 76:565-569.
7. Dougherty, R. M. 1976. Induction of tumors in Syrianhamsters by a human renal papovavirus, RF strain. J.Natl. Cancer Inst. 57:395-400.
8. Freund, J., G. di Mayorca, and K. N. Subramanian.1979. Mapping and ordering of fragments of BK virusDNA produced by restriction endonucleases. J. Virol.29:915-925.
9. Gardner, S. D., A. M. Field, D. V. Coleman, and B.Hulme. 1971. New human papovavirus (B.K.) isolatedfrom urine after renal transplantation. Lancet i:1253-1257.
10. Howley, P. M., G. Khoury, J. C. Byrne, K. K. Take-moto, and M. A. Martin. 1975. Physical map of the
X'oi. ;32, 1979
on April 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

942 N A I'ANABE E'l' Al.
BK virus genome. J. Virol. 16:959-973.11. Howley, P. M., M. F. Mullarkey, K. K. Takemoto, and
M. A. Martin. 1975. Characterization of human papo.vavirus BK DNA. J. Virol. 15:173-181.
12. Khoury, G., P. M. Howley, C. Garon, M. F. Mullarkey,K. K. Takemoto, and M. A. Martin. 1975. Hotnolgoyand relationship between the genomes of papovaviruses,BK virus and simian virus 40. Proc. Natl. Acad. Sci.U.S.A. 72:2563-2567.
13. Mertz, J. E., and P. Berg. 1974. Viable deletion mutantsof simian virus 40: selective isolation by means of arestriction endonuclease from Hemophilus parainflu-enzae. Proc. Natl. Acad. Sci. U.S.A. 71:4879-4883.
14. Nase, L. M., M. Karkkainen, and R. A. Mantyjarrvi.1975. Transplantable hamster tumors induced with theBK virus. Acta Pathol. Microbiol. Scand. Sect. B 83:347-352.
15. Newell, N., C.-J. Lai, G. Khoury, and T. J. Kelly, Jr.1978. Electron microscope study of the base sequencehomology between simian virus 40 and human papo-vavirus BK. J. Virol. 25:193-201.
16. Osborn, J. E., S. M. Robertson, B. L. Padgett, D. L.Walker, and B. Weisblum. 1976. Comparison of JCand BK human papovaviruses with simian virus 40:DNA homology studies. J. Virol. 19:675-684.
17. Reddy, V. B., B. Thimnlappaya, R. Dhar, K. N. Sub-ramanian, B. S. Zain, J. Pan, P. K. Ghosh, M. L.Celma, and S. M. Weissman. 1978. The genome ofsimian virus 40. Science 200:494-502.
18. Shah, K. V., R. W. Daniel, and J. D. Standberg. 1975.Sarcoma in a hamster inoculated with BK virus, ahuman papovavirus. J. Natl. Cancer Inst. 54:945-950.
19. Shenk, T. E., J. Carbon, and P. Berg. 1976. Construc-tion and analysis of viable deletion mutants of simianvirus 40. J. Virol. 18:664-671.
20. Sugden, B., B. DeTroy, R. J. Roberts, and J. Sam-brook. 1975. Agarose slab-gel electrophoresis equip-ment. Anal. Biochem. 68:36-46.
21 ITehida. S.. S. Watanabe, T. Aizawa, A. Furuno, and
T. Muto. 1979. Polyoncogenicity and insulinoma-induc-ing ability of BK virus, a human papovavirus, in Syriangolden hamsters. J. Natl. Cancer Inst. 63:119-126.
22. Uchida, S., S. Watanabe, T. Aizawa, K. Kato, A.Furuno, and T. Muto. 1976. Induction of papillaryependymomas and insulinomas in the Syrian goldenhamster by BK virus, a human papovavirus. Gann 67:857-865.
23. Uchida, S., S. Watanabe, and M. Kato. 1966. Incom-plete growth of simian virus 40 in African green monkeykidney culture induced by serial undiluted passages.Virology 28:135-141.
24. van der Noordaa, J. 1976. Infectivity, oncogenicity andtransforming ability of BK virus and BK virus DNA. J.Gen. Virol. 30:371-373.
25. Villarreal, L. P., R. T. White, and P. Berg. 1979.Mutational alterations within the simian virus 40 leadersegment generate altered 16S and 19S mRNA's. J. Virol.29:209-219.
26. Watanabe, S. 1975. Virus DNA synthesizing ability of Tantigen-forming defective SV40 produced by successiveundiluted passages. J. Gen. Virol. 26:49-57.
27. Yang, R. C. A., and R. Wu. 1978. BK virus DNA:cleavage map and sequence analysis. Proc. Natl. Acad.Sci. U.S.A. 75:2150-2154.
28. Yang, R. C. A., and R. Wu. 1979. BK virus DNAsequence: extent of homology with simian virus 40DNA. Proc. Natl. Acad. Sci. U.S.A. 76:1179-1183.
29. Yoshiike, K., and V. Defendi. 1977. Presence of deletionmolecules in human wart virus DNA. J. Virol. 21:415-418.
30. Yoshiike, K., and V. Defendi. 1977. Addition of extraDNA sequences to simian virus 40 DNA in vivo. J.Virol. 23:323-337.
31. Yoshiike, K., A. Furuno, S. Watanabe, S. Uchida, K.Matsubara, and Y. Takagi. 1974. Characterization ofdefective simian virus 40 DNA: comparison betweenlarge-plaque and small-plaque types. Cold Spring Har-bor Symp. Quant. Biol. 39:85-93.
J. VlRoi,.
on April 13, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from