lucas rodrigues santos estrutura da fauna de bivalves (mollusca ... · a minha mãe “edjane...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE SERGIPE
CENTRO DE CIÊNCIAS BIOLÓGICAS E DA SAÚDE
DEPARTAMENTO DE ECOLOGIA – DECO
Lucas Rodrigues Santos
Estrutura da fauna de Bivalves (Mollusca) associados às macroalgas provenientes da plataforma continental ao longo das bacias
sedimentares de Sergipe-Alagoas e Jacuípe
São Cristóvão
2017.1

2
UNIVERSIDADE FEDERAL DE SERGIPE
CENTRO DE CIÊNCIAS BIOLÓGICAS E DA SAÚDE
DEPARTAMENTO DE ECOLOGIA – DECO
Lucas Rodrigues Santos
Estrutura da fauna de Bivalves (Mollusca) associados às macroalgas provenientes da plataforma continental ao longo das bacias
sedimentares de Sergipe-Alagoas e Jacuípe
Trabalho de conclusão de curso apresentado ao Departamento de Ecologia da Universidade Federal de Sergipe como parte dos requisitos para obtenção do título de Bacharel em Ecologia, desenvolvido sob a orientação da Profa. Dra. Carmen Regina Parisotto Guimarães e Coorientação de José Weverton Santos de Souza.
São Cristóvão
2017.1

3

4
AGRADECIMENTOS
Foram 5 longos anos de sufoco, de eita atrás de eita e de muitos momentos dos quais quis abandonar tudo. Porém, olhando de lá pra cá, o único sentimento que consigo ter é orgulho. Orgulho da pessoa que me tornei hoje e do profissional que pretendo ser daqui em diante.
Então, primeiramente, agradeço a Deus pelas oportunidades e por todo aprendizado adquirido durante todo esse tempo na graduação, não só dentro, como fora da faculdade também. Pelos momentos dos quais retirei forças e acreditei em mim mesmo, ultrapassando todas as minhas limitações. Então, “MUITO OBRIGADO!”.
Agradeço a toda minha família;
A minha mãe “Edjane Rodrigues Santos” e ao meu pai “Valdikson da Vitória Santos”, muito obrigado por todo o apoio e por me aturarem nos meus momentos de surtos (risos). Ao meu irmão “Valdikson-Jr” que nem faz ideia do quanto me ajudou direta e indiretamente (muito obrigado pelos 5 anos de internet pagos ;] ).
Aos meus avós, os quais sempre foram os pilares da minha família; a todos os meus tios e tias que sempre os vi como inspiração e a todos os meus primos, especialmente a Débora, Hellen e Isaac, que sempre estiveram comigo em todos os momentos especiais da minha vida.
A todos os meus amigos fora e dentro da UFS;
Aos meus amigos Américo, Gersieny, Lara e Suellen, os quais estão comigo dês do ensino médio, sempre me aturando, dando conselhos, papos furados e sempre rendendo muitas risadas.
As minhas irmãs de Laboratório/Iniciação Científica/TCC/Sofrimento Galdêndia e Ana Paula. Muito obrigado pelo apoio nos momentos mais difíceis, pelas risadas diárias e pela companhia sempre muito bem vinda. “Sim, Conseguimos!” kkk.
Aos meus amigos de graduação: Bruna, Karine, Léo, Monize e Susi, que sempre estiveram comigo nas atividades do curso, assim como nos melhores momentos da minha estadia na UFS. As minhas amigas Luiza e Tâmara pela parceria e gestos de carinho sempre que possível. Também agradeço a toda galera da “Ecologia de campo”.
A toda equipe do Laboratório de Bentos Costeiro (LABEC);
A Ilma, muito obrigado por todas as boas conversas acompanhadas do cafezinho da tarde. Aos Irmãos Cosme e Damião, e a todos os estagiários dos quais tive convívio durante todos os meus 3 anos de estadia no LABEC.

5
A minha orientadora Profa. Dra. Carmen Regina Parisotto Guimarães, por ter aceitado, literalmente, o desafio de me orientar durante esses 2 últimos anos e ter acreditado no meu potencial. Muito obrigado pelos inúmeros puxões de orelha (sempre necessários), pelos momentos de motivação e também pelos melhores conselhos tanto acadêmicos quanto pessoais (sempre lembrarei deles).
Ao meu coorientador José Weverton Santos de Souza, meu melhor professor de estatística e de ecologia “EVER”, muito obrigado por me ajudar nos momentos mais importantes da minha graduação. Sempre lembrarei de você como um “Influencer” para minha futura carreira de ecólogo (risos).
A minha banca avaliadora, Luana Marina de Castro Mendonça e Mariana Andrade Oliveira de Carvalho, por terem aceitado o convite e pelas contribuições na minha monografia.
A todos os meus professores, pelo aprendizado passado, em especial aos professores do Departamento de Ecologia.
A Setuko Masunari, pela disponibilização do material bibliográfico, em um dos meus momentos de desespero.
E, por fim, agradeço a PETROBRAS/CENPES pela coleta dos dados, ao Laboratório de Bentos Costeiro pela disponibilização de todo o material analisado e a Universidade Federal de Sergipe pela oportunidade.

6
ÍNDICE DE FIGURAS
Figura 1: Mapa contemplando a área de estudo com as divisões das porções marinhas das Bacias de Sergipe-Alagoas e do Jacuípe. Retirado de ANP (2017). ........................................ 16 Figura 2: Mapa identificando as estações amostrais nas Bacias sedimentares de Sergipe-Alagoas e Jacuípe, identificadas pelos transectos de A até o H e suas respectivas isóbatas de 10, 25 e 50 m, coletadas no ano de 2011. ................................................................................. 17 Figura 3: Representação dos 10 táxons mais abundantes da fauna de bivalves associados às macroalgas provenientes das bacias sedimentares de Sergipe/Alagoas e Jacuípe, referentes aos períodos seco e chuvoso do ano de 2011. Legenda: Bivalvia = Bivalvia não identificado. .... 20 Figura 4: Variação sazonal e desvio padrão dos índices ecológicos riqueza, abundância, diversidade e equitatividade, e o nível de significância (p) obtido a partir do teste Mann-Whitney, da fauna de bivalves associados às macroalgas provenientes das bacias sedimentares de Sergipe/Alagoas e Jacuípe, referentes aos períodos seco e chuvoso do ano de 2011. Legenda: * = variável significativa. ......................................................................................... 23 Figura 5: Escalonamento multidimensional não métrico (nMDS) agrupando a abundância por estações e períodos sazonais da fauna de bivalves associados às macroalgas provenientes das bacias sedimentares de Sergipe/Alagoas e Jacuípe, referentes aos períodos seco e chuvoso do ano de 2011. .............................................................................................................................. 24 Figura 6: Variação e desvio padrão dos índices ecológicos de riqueza, abundância, diversidade e equitatividade, e o nível de significância (p) obtido a partir do teste Kruskal-Wallis, da fauna de bivalves associados às macroalgas provenientes das bacias sedimentares de Sergipe/Alagoas e Jacuípe, referente as isóbatas de 10, 25 e 50 m amostradas nos período seco e chuvoso do ano de 2011. Legenda: * = variável significativa. ...................................... 25 Figura 7: Escalonamento multidimensional não métrico (nMDS) agrupando a abundância por estações e isóbatas de 10, 25 e 50 m da fauna de bivalves associados às macroalgas provenientes das bacias sedimentares de Sergipe/Alagoas e Jacuípe, referentes aos períodos seco e chuvoso do ano de 2011. ............................................................................................... 26 Figura 8: Variação e desvio padrão dos índices ecológicos de riqueza, abundância, diversidade e equitatividade, e o nível de significância (p) obtido a partir do teste Kruskal-Wallis, da fauna de bivalves associados às macroalgas provenientes das bacias sedimentares de Sergipe/Alagoas e Jacuípe, referentes aos transectos amostrados nos períodos seco e chuvoso do ano de 2011. .......................................................................................................... 27 Figura 9: Escalonamento multidimensional não métrico (nMDS) agrupando a abundância por estações e transectos da fauna de bivalves associados às macroalgas provenientes das bacias sedimentares de Sergipe/Alagoas e Jacuípe, referentes aos períodos seco e chuvoso do ano de 2011. ......................................................................................................................................... 28 Figura 10: Diagrama de ordenação CCA (análise de correspondência canônica) dos pontos amostrais e das variáveis ambientais coletadas na bacia sedimentar de Sergipe/Alagoas e Jacuípe, referente ao período seco e chuvoso do ano de 2011. ................................................ 30 Figura 11: Diagrama de ordenação CCA (análise de correspondência canônica) da fauna de bivalves e das variáveis ambientais coletadas na bacia sedimentar de Sergipe/Alagoas e Jacuípe, referente ao período seco e chuvoso do ano de 2011. Legenda das espécies: Cop -

7
Corbula operculata, Csw - Caryocorbula swiftiana, Cch - Caryocorbula chittyana, Aze - Arca zebra, Aan - Anadara aff. notabilis, An1 - Anadara sp. 1, An2 - Anadara sp. 2, Bdo - Barbatia domingensis, Asa - Amygdalum sagittatum, Mla - Musculus lateralis, Dvi - Dacrydium vitreum, Cde - Crenella decussata, My1 - Mytilidae sp. 1, My2 - Mytilidae sp. 2, Sbe - Spathochlamys benedicti, Lba - Leptopecten bavayi, Lsp - Lindapecten sp., Pim - Pinctada imbricata, Pco - Pteria colymbus, Can - Cratis antillensis, Csp - Cratis sp., Gce - Gouldia cerina, Nco - Nuculana concêntrica, Clu - Crassinella lunulata, Sfr - Sphenia
fragilis, Lhi - Lamychaena hians, Aad - Arcopsis adamsi, Osp - Ostreidae sp., Ung - Ungulinidae sp., Li1 - Limidae sp. 1, Li2 - Limidae sp. 2, Biv - Bivalvia não identificado. ... 31
ÍNDICE DE TABELAS
Tabela 1: Abundância total de indivíduos indicados por família, gênero e espécie da fauna de bivalves associados às macroalgas provenientes das bacias sedimentares de Sergipe/Alagoas e Jacuípe, referentes aos períodos seco e chuvoso do ano de 2011............................................. 21 Tabela 2: Fauna geral de bivalves associados às macroalgas das bacias sedimentares de Sergipe/Alagoas e Jacuípe amostrados no ano de 2011, indicando o período sazonal, transectos e suas respectivas isóbatas onde foram encontrados. .............................................. 28 Tabela 3: Variáveis ambientais utilizadas na análise de correspondência canônica (CCA) com os valores de explicação (%) e nível de significância (p) de seus respectivos eixos. .............. 31 Tabela 4: Modelos lineares generalizados (GLM) realizados entre a abundância dos bivalves e os parâmetros ambientais amostrados, incluindo os seus respectivos os valores de significância (p) dos parâmetros ambientais e suas interações, da bacia sedimentar de Sergipe/Alagoas e Jacuípe, referente ao período seco e chuvoso do ano de 2011. Legenda: * = variável significativa................................................................................................................. 32

8
SUMÁRIO
1. INTRODUÇÃO ................................................................................................................ 10
2. REFERENCIAL TEÓRICO ............................................................................................. 11
2.1. O ambiente fital ......................................................................................................... 11
2.2. Classe Bivalvia e sua relação com o ambiente fital ................................................... 13
3. OBJETIVOS...................................................................................................................... 15
3.1. Geral ........................................................................................................................... 15
3.2. Específicos ................................................................................................................. 15
4. METODOLOGIA ............................................................................................................. 15
4.1. Área de estudo ........................................................................................................... 15
4.2. Procedimentos de campo ........................................................................................... 16
4.3. Procedimentos de laboratório .................................................................................... 17
4.4. Análise dos dados ...................................................................................................... 17
4.5. Programas estatísticos ................................................................................................ 20
5. RESULTADOS ................................................................................................................. 20
5.1. Composição da fauna de bivalves .............................................................................. 20
5.2. Distribuição temporal da fauna de bivalves ............................................................... 22
5.3. Distribuição espacial da fauna de bivalves pelas isóbatas ......................................... 24
5.4. Distribuição espacial da fauna de bivalves pelos transectos ...................................... 26
5.5. Correlação da fauna com os parâmetros ambientais .................................................. 29
6. DISCUSSÃO ..................................................................................................................... 32
7. CONSIDERAÇÕES FINAIS ............................................................................................ 35
8. REFERÊNCIAS ................................................................................................................ 36

9
RESUMO
As macroalgas formam um rico ambiente marinho e bentônico, com os seus diversos
organismos associados. Dentre os grupos animais componentes deste ambiente, os moluscos
são citados como um dos mais representativos, sendo a classe Bivalvia uma das mais
abundantes. Nesse sentido, o objetivo deste trabalho foi conhecer a composição taxonômica e
a distribuição espacial e temporal dos bivalvia associada às macroalgas encontradas na região
da plataforma continental das bacias sedimentares de Sergipe-Alagoas e Jacuípe. O material
analisado é proveniente de arrastos pesqueiros realizados com rede camaroneira, coletados em
duas campanhas oceanográficas, envolvendo os períodos seco (fevereiro) e chuvoso (julho) de
2011, na qual foram amostradas 24 estações distribuídas em oito transectos. A fauna de
bivalves esteve representada por uma abundância total de 1.384 indivíduos, distribuídos em
20 famílias, 26 gêneros e 28 espécies. Dentre as espécies mais abundantes, a Arca zebra
(Arcidae) foi a mais representativa em ambos os períodos sazonais, compondo 40% do total
dos organismos coletados. A fauna também variou em função do tempo e do espaço, e os
principais parâmetros ambientais explicativos dessa variação foram a profundidade,
salinidade e o fundo sedimentar do tipo arenoso.
PALAVRAS-CHAVE: macroalgas, bentos, malacofauna, bivalves, ecologia de comunidades

10
1. INTRODUÇÃO
Os Mollusca representam o segundo maior filo animal, com cerca de 150.000 espécies
descritas e, são representados atualmente, por oito classes: Caudofoveata, Solenogastres,
Scaphopoda, Polyplacophora, Monoplacophora, Cephalopoda, Gastropoda e Bivalvia. Todos
estes são presente e bem-sucedidos na vida marinha (Rupert & Barnes 2005, Amaral et al.
2003). São participantes de vários níveis da cadeia trófica, tanto como presas quanto como
predadores, possuindo importante papel na organização de várias comunidades marinhas
(Rocha 2011).
Dentre os variados habitats no qual o filo Mollusca ocorre está o ambiente de
plataforma continental, que pode ser caracterizado como um prolongamento submerso da
maior parte dos continentes e tem o seu fim com o aumento do declive, denominado quebra
de plataforma, sendo este também o seu limite externo (Coutinho 2000). Plataformas
continentais são caracteristicamente ambientes altamente complexos e dinâmicos, que
comportam uma grande diversidade de organismos marinhos e são de suma importância para
a manutenção e equilíbrio ecológico dos oceanos (Manso et al. 2004).
A plataforma continental brasileira possui suas águas superficiais formadas
basicamente por águas tropicais e subtropicais em suas afinidades oceanográficas, faunísticas
e florísticas (Lana et al. 1996). A complexidade desta pode ser aumentada por fenômenos
oceanográficos ali ocorrentes, sejam eles periódicos ou regulares, como por exemplo a
ocorrência de ressurgências, pela influência terrígena dada pelo carreamento de material para
os estuários e a sua descarga no ambiente marinho, e também pela sua grande
heterogeneidade de fundos ao longo de todo litoral brasileiro, no qual podem ocorrer
ambientes recifais, lamosos ou arenosos, dentre outras várias fácies sedimentares possíveis.
Estes fenômenos criam uma ampla variabilidade na diversidade e biomassa dos organismos,
também refletida na distribuição espaço-temporal das comunidades, podendo inclusive, afetar
o estabelecimento e a distribuição de ambientes como fital e/ou da fauna associada ao mesmo
(Lana et al. 1996).
O fital, na visão de Nascimento & Rosso (2007), pode ser definido como o conjunto
de organismos que habitam o macrofitobentos, usando-o como substrato. As macroalgas
presentes neste ambiente fornecem microhabitats para uma grande diversidade da fauna e
flora epibêntica e, pode ser utilizada como abrigo, refúgio, moradia, berçário, alimentação,
proteção contra a luminosidade, ressecamento e também como pastagem para os diversos
organismos associados a esse ambiente, inclusive, alguns animais de interesse comercial para

11
o homem, como por exemplo os moluscos, peixes e crustáceos (Alves & Araújo 1999,
Masunari & Forneris 1981, Nascimento & Rosso 2007, Nascimento 2007).
Variações espaço-temporais das comunidades epifaunais estão atribuídas
conjuntamente aos componentes bióticos e abióticos (Johnson & Scheibling 1987), sendo a
interação destes componentes bastante clara no fital, pois fatores como a predação,
competição, disponibilidade de nutrientes, luz, temperatura, nível batimétrico, características
hidrodinâmicas, sazonalidade, quantidade de sedimento retido nas frondes, textura, cor e
compostos químicos das macroalgas, podem influenciar a estadia das comunidades presentes
(Masunari 1987, Chemello & Millazo 2002, Nascimento & Rosso 2007).
Diversos trabalhos têm demonstrado a variada fauna de invertebrados associada às
macroalgas marinhas (Tararam & Wakabara 1981, Johnson & Scheibling 1987, Viejo 1999,
Nascimento & Rosso 2007), e os bivalves apresentam-se como um destes componentes
(Jacobucci et al. 2006, Bezerra 2011, Fernandes 2014), sendo o segundo maior grupo
representativo do filo Molusca no fital, tanto em abundância quanto em diversidade, atrás
apenas dos gastrópodes (Almeida 2007, Bezerra 2011).
Apesar de alguns trabalhos apresentarem a importância das macroalgas para as
diversas comunidades de organismos associadas a elas, ainda há pouco conhecimento
relacionado às características estruturadoras destas comunidades (Queiroz 2011). Sendo
assim, o conhecimento de alguns padrões de estruturação, tais como os índices biológicos de
abundância, riqueza, diversidade e uniformidade, podem funcionar como uma importante
ferramenta para o monitoramento e aplicação de manejo de determinadas comunidades
marinhas. Entretanto, apesar da sua significativa importância para o bentos, as comunidades
de bivalves ainda têm sido pouco estudada no que diz respeito a estes parâmetros
estruturadores e, estes podem ser aplicados com o intuito de obter informações e promover a
comparação intra e entre diferentes habitats (Soares-Gomes & Pires-Vanin 2003).
Diante disso, e com base na importância ecológica das macroalgas, o presente trabalho
visou ampliar o conhecimento sobre a biodiversidade de bivalves associada a este ambiente
nas bacias sedimentares de Sergipe-Alagoas e Jacuípe. O levantamento destas informações
poderá, posteriormente, servir como subsídio à tomada de decisões no gerenciamento costeiro
da área de estudo.
2. REFERENCIAL TEÓRICO
2.1. O ambiente fital

12
O fital pode ser definido como um terceiro ambiente existente dentro do ecossistema
marinho litoral, diferindo-se dos ambientes pelagial e bental, pois, a composição e
distribuição dos organismos associados a uma determinada população de alga difere-se da do
sedimento e da coluna d’água (Remane, 1933 apud Masunari & Forneris 1981). Este
ambiente é dominado por algas talosas e gramas marinhas (Edgar & Moore 1986) as quais
comportam uma diversificada fauna e flora (Tararam & Wakabara 1981, Johnson &
Scheibling 1987).
As macroalgas marinhas são organismos fotossintetizantes comumente distribuídas ao
longo de todo litoral brasileiro, sendo mais abundantes e diversificadas em regiões de águas
transparentes e substratos rochosos (Oliveira et al. 2002). Na costa nordeste do país as
macroalgas são predominantes, pois a costa apresenta um menor aporte de sedimentos e de
água doce, ocasionado pela ausência de grandes rios (Figueiredo et al. 2008). No entanto,
diferente dos demais estados, na costa de Sergipe não é encontrado o mesmo padrão, já que
ela apresenta um intenso aporte fluvial devido a presença da desembocadura de cinco rios ao
longo da costa, destacando-se o do rio São Francisco, cuja pluma sedimentar é transportada
do norte para o sul do estado (Guimarães 2010).
Os bancos de macroalgas marinhas são representados basicamente por dois principais
grupos taxonômicos, sendo eles: Rhodophyta (algas vermelhas) e Chlorophyta (algas verdes)
(Orfanidis et al. 2001). Apesar das peculiaridades de cada grupo, possuem como estrutura
comum o corpo formado por um talo e, as variações do seu formato, dão origem as diversas
formações vegetais marinhas conhecidas, a exemplo das algas incrustantes, filamentosas e as
foliáceas (Muniz et al. 2013).
As macroalgas desempenham importante função ecológica no ambiente marinho,
estando entre os principais produtores primários da zona costeira e participando da base da
cadeia trófica desse ecossistema. Além disso, possuem forte relação com uma variedade de
invertebrados e peixes, por oferecerem um ambiente propicio para os mesmos, como recurso
espacial, proteção e locais de alimentação (Muniz et al. 2013, Nascimento & Rosso 2007,
Nascimento 2007, Costa et al. 2012).
Fatores biológicos, físicos e químicos podem intervir na organização e distribuição das
comunidades biológicas marinhas (Johnson & Scheibling 1987), diante disso, diversos
trabalhos têm evidenciado a influência desses fatores em diferentes fitais (Masunari 1987,
Chemello & Millazo 2002, Nascimento & Rosso 2007, Lacerda et al. 2009).
Dentre as variadas relações existentes entre o fital e as macroalgas, a variação da
complexidade estrutural do habitat está entre os assuntos mais abordados, sendo a textura,

13
forma, arquitetura ou estrutura da superfície da alga, podem influenciar a complexidade desse
ambiente (Nascimento 2007). Chemello & Milazzo (2002) relatam a relação da complexidade
da alga com a diversidade de organismos associados, de modo que, algas mais complexas
estruturalmente apresentaram uma maior diversidade, ao contrário das algas menos
complexas. Já Masunari & Forneris (1981) comentam que as adaptações morfológicas
sofridas pelos organismos componentes do fital estão relacionadas à grande heterogeneidade
dos indivíduos associados a alga substrato.
A fauna encontrada associada ao fital pode ser classificada a partir dos seus diversos
tamanhos em micro, meio, macro e megafauna. Esta fauna é representada por organismos
vágeis e sésseis muito diversos e com diferentes hábitos alimentares (Nascimento & Rosso
2007, Almeida 2007). Estudos sobre a fauna associada aos fitais do Brasil concentram-se na
região Sudeste do país, alguns destes relacionados a grupos específicos de organismos
marinhos, a exemplo de Boffi (1972), Tararan & Wakabara (1981), Masunari (1988),
Jacobucci & Leite (2002), Guth (2004) e Jacobucci et al. (2006). No litoral nordestino poucos
trabalhos foram realizados sobre as comunidades fitais, dentre eles, alguns exemplos como o
trabalho de Queiroz et al. (2007), Pereira et al. (2010) e Barros (2015). Entretanto, nenhum
dos trabalhos aqui citados, apresentou como metodologia adotada o arrasto pesqueiro para a
obtenção da fauna associada as macroalgas.
2.2.Classe Bivalvia e sua relação com o ambiente fital
A classe Bivalvia tem o seu nome devido a presença de duas valvas calcárias externas
que protegem a parte mole do corpo do animal (Tunnell-Jr et al. 2010). Também conhecida
como Pelecypoda, esta classe inclui cerca de 20.000 espécies viventes de organismos
marinhos e também de água doce. No ambiente marinho, possui representantes em todas as
profundidades (Brusca & Brusca 2007).
Os bivalves se dividem em três grupos morfológicos que podem ser diferenciados por
peculiaridades de suas brânquias e pelo hábito alimentar, sendo eles: os protobrânquios; os
lamelibrânquios e os septibrânquios. Acredita-se que os protobrânquios (comedores seletivos
de depósitos) sejam os bivalves existentes mais primitivos; por outro lado, os lamelibrânquios
(comedores de suspensão) estão a grande maioria das espécies atuais. Por fim, os
septibrânquios são citados como um grupo reduzido bem especializado, possuindo hábito
alimentar carnívoro, detritívoro ou necrófago (Rupert & Barnes 2005).
Em relação ao seu habitat, podem ser encontrados associados a diferentes tipos de
substratos (areia, lama, cascalho, rocha, madeira, ervas marinhas, algas ou marismas),

14
podendo possuir modo de vida séssil ou infaunal (presos pelo bisso ou cimentados), epifaunal
ou de vida livre (Amaral et al. 2003, Mikkelsen & Bieler 2008).
No ambiente fital, a fauna presente encontra nas macroalgas substrato para fixação,
recurso alimentar e locais para sua morada e abrigo (Masunari 1987), dentre os invertebrados
associados ao ambiente, os moluscos são um dos grupos mais representativos, destacando-se a
classe bivalvia como uma das mais abundantes e diversificadas do ambiente (Almeida 2007,
Bezerra 2011). Fernandes (2014) comenta sobre a importância ecológica da malacofauna,
com destaque para os bivalves, já que os mesmos estão intimamente relacionados com o
fornecimento do fluxo de energia nos diferentes níveis tróficos das teias alimentares marinhas.
Castro & Huber (2012) relatam a relação da alta produtividade primária dos ambientes
vegetados e das comunidades animais presentes, sendo que, em ambientes de bancos de
fanerógamas e de campos de algas pardas gigantes, os animais associados podem aproveitar-
se destas de várias formas, mesmo não as consumindo diretamente. Estes autores destacam os
poliquetos depositívoros, pepinos-do-mar e os moluscos bivalves como alguns dos
consumidores diretos dos detritos formados pelas algas mortas, sendo estes detritos também
uma importante fonte de alimento para as diversas comunidades do ambiente marinho.
Peterson & Heck-Jr (2001) comparando dois ambientes, um não vegetado e outro
vegetado com bancos da alga Thalassia testudinum, observaram o aumento da taxa da
sobrevivência de uma comunidade de bivalve quando associados à alga, já que a mesma
permitiu recurso espacial e habitat propício para o estabelecimento desses organismos.
Soares-Gomes & Pires-Vanin (2003) citam a composição e os padrões de estruturação
como uma das etapas cruciais para o entendimento das diversas comunidades. Porém, no
Brasil, ainda existem poucos trabalhos publicados no que diz respeito à malacofauna
associada ao fital evidenciando tais padrões, principalmente no litoral nordestino. Dentre
esses trabalhos, alguns merecem destaque, a exemplo de Alves & Araújo (1999), que
descreveram a estrutura de comunidades de moluscos associados ao fital Halodule wrightii da
costa leste da Ilha de Itamaracá (PE); Almeida (2007) que realizou o levantamento da
malacofauna associada a macroalga Sargassum spp. no Pontal do Cupe, Ipojuca (PE); Veras
(2011) por caracterizar quali e quantitativamente a malacofauna associada a macroalga
Pterocladiella caerulescens (Rhodophyta, Pterocladiaceae) na zona entremarés da Praia da
Pedra Rachada, em Paracuru (CE) e Bezerra (2011), que realizou o levantamento da
malacofauna associada a macroalga Halimeda Opuntia (Linnaeus) no Pontal do Cupe, Ipojuca
(PE).

15
3. OBJETIVOS
3.1. Geral
Conhecer a composição taxonômica e a distribuição espacial e temporal dos bivalvia
associados às macroalgas da plataforma continental na região das bacias sedimentares de
Sergipe-Alagoas e Jacuípe.
3.2. Específicos
� Inventariar a fauna de bivalves associada às macroalgas;
� Caracterizar a distribuição espaço-temporal da fauna dos bivalves associada às
macroalgas a partir dos descritores ecológicos de riqueza, abundância, diversidade e
equitatividade;
� Investigar a influência dos parâmetros ambientais amostrados sobre a fauna de
bivalves associados às macroalgas;
4. METODOLOGIA
4.1. Área de estudo
A área de estudo compreende a porção marinha das Bacias de Sergipe-Alagoas e do
Jacuípe (Figura 1).
A Bacia de Sergipe-Alagoas está situada na margem continental da região Nordeste do
Brasil, e possui uma área total de 44.370Km², sendo a porção marítima equivalente a
31.750Km² e a porção terrestre a 12.620Km². É limitada ao norte, com a Bacia de
Pernambuco-Paraíba pelo Alto de Maragogi e ao sul, com a bacia de Jacuípe pela Plataforma
de Estância na porção terrestre e pelo sistema de falhas do Vaza-Barris na porção marítima
(Haeser 2015).
Por sua vez, a Bacia de Jacuípe está situada na margem continental Leste do Brasil, e é
predominantemente marítima, cuja área total aproxima-se de 28.000km², limitando-se ao
norte, pela Bacia Sergipe-Alagoas e ao sul, pela Bacia de Camamu-Almada pelo sistema de
falhas de Itapuã (Freitas & Ferreira 2015).
A plataforma continental de Sergipe apresenta uma configuração diferente dos demais
estados do Nordeste/Leste do Brasil, sendo considerada uma plataforma rasa e estreita, com
recortes em frente aos cânions do São Francisco e do Japaratuba os quais ficam distantes
aproximadamente 12Km da costa, possuindo um intenso aporte fluvial provocado pela
desembocadura de cinco complexos estuarinos com destaque para o do rio São Francisco,
cuja pluma sedimentar é transportada do norte ao sul do estado (Guimarães 2010).

16
O regime pluviométrico da área de estudo é caracterizado por chuvas mais abundantes
no período outono-inverno (UFS/SEPLAN 1979).
Figura 1: Mapa contemplando a área de estudo com as divisões das porções marinhas das Bacias de Sergipe-Alagoas e do Jacuípe. Retirado de ANP (2017).
4.2. Procedimentos de campo
Todo o material utilizado neste estudo é proveniente do Projeto – “Caracterização
Ambiental da Bacia de Sergipe e sul de Alagoas (MARSEAL)”, coordenado pela
PETROBRAS/CENPES e desenvolvido por uma equipe de pesquisadores da Universidade
Federal de Sergipe (UFS).
As macroalgas foram analisadas a partir das amostras da megafauna bêntica. Este
material foi obtido a partir de duas campanhas amostrais realizadas durante o período seco
(ARR1), no mês de fevereiro de 2011 e durante o período chuvoso (ARR2), em julho do
mesmo ano. Foram amostradas 24 estações, distribuídas em oito transectos, denominados de
norte para sul de A à H contemplando as isóbatas de 10, 25 e 50 m (Figura 2), as coordenadas
dos polígonos estão representadas na pela tabela 1.
As amostras foram obtidas em arrastos pesqueiros realizados com uma rede de portas
do tipo camaroneira. Ainda em campo, as macroalgas foram separadas do restante do
material, acondicionadas em isopor com gelo e, posteriormente, levadas para o Laboratório de
Ecossistemas Costeiro (LABEC) do Departamento de Biologia da UFS.
Todas as variáveis ambientais (tipo do sedimento de fundo, temperatura, salinidade e a
profundidade) de cada estação foram cedidas pelo LABEC.

17
Figura 2: Mapa identificando as estações amostrais nas Bacias sedimentares de Sergipe-Alagoas e Jacuípe, identificadas pelos transectos de A até o H e suas respectivas isóbatas de 10, 25 e 50 m, coletadas no ano de 2011.
4.3. Procedimentos de laboratório
No Laboratório, as macroalgas foram lavadas sobre uma peneira com malha de 500µm
e, posteriormente, o material resultante foi fixado em formol 10%. As amostras foram triadas
a olho nu para retirada dos organismos associados. Em seguida os organismos foram
separados em grandes grupos taxonômicos sob microscópio estereoscópico e acondicionados
em frascos contendo álcool 70%, para sua conservação.
Os moluscos foram separados da fauna geral e triados com o auxílio do microscópio
estereoscópico Leica EZ4. Posteriormente os Bivalvia foram separados dos demais Mollusca
e identificados até o menor nível taxonômico possível de acordo com bibliografia pertinente
baseada em Rios (1994, 2009), Mikkelsen & Bieler (2008), entre outros. Para este estudo,
foram apenas considerados os indivíduos contendo concha e parte mole.
A nomenclatura das espécies encontradas foi atualizada pelo website de registro de
espécies marinhas WoRMS – World Register of Marine Species.
4.4. Análise dos dados
A análise de dados envolveu o cálculo da frequência relativa das espécies encontradas
e dos descritores ecológicos riqueza (Nibbaken 1982), abundância, diversidade (Shannon-
Wiener) e equitatividade (Pielou, 1975, 1969). Posteriormente, esses parâmetros foram

18
utilizados nas análises da distribuição espacial e temporal da fauna dos bivalves e a correlação
com os dados ambientais.
� Frequência relativa (Fr%)
A frequência relativa (Fr) é a relação entre o número de indivíduos de uma espécie e o
número total de indivíduos da amostra. Dada em porcentagem (%), esta é expressa pela
seguinte equação:
�� = �
��100
onde: n = número de indivíduos de determinada espécie;
T = número total de espécies da amostra.
� Índices de riqueza, abundância, diversidade e equitatividade
A riqueza (S) constituiu a contagem do número de espécies presentes em cada estação.
Já a abundância (N) foi obtida a partir do somatório do número de indivíduos coletados.
Para o cálculo da diversidade foi utilizado o índice de Shannon-Wiener (H’), o mesmo
leva em consideração a riqueza e a uniformidade na abundância, dando maior peso para as
espécies raras devido as variações da abundância, e é expressa pela seguinte equação:
� =∑(��� ln ��)
onde: pi = é a proporção da espécie (i), estimada como ni/N (no qual ni é o número de
indivíduos encontrados na estação e N é o número total de indivíduos total).
A diversidade é dada em bits/ind., sendo considerada baixa quando < 1, intermediária
entre 1 e 3 e elevada quando > 3.
A uniformidade ou equitatividade (J) é calculada a partir do índice de Pielou (1975,
1969), o mesmo leva em consideração a razão da diversidade obtida (H’) e o número de
espécies (S), esta é expressa pela seguinte equação:
� =′
���
onde: H’ = diversidade obtida;

19
S = número total de espécies.
O valor de J varia de 0 a 1, sendo que os valores próximos a 1 representam
distribuição equitativa do número de indivíduos das espécies presentes.
� Análise espaço-temporal e correlação da fauna com os parâmetros ambientais
A normalidade dos dados dos descritores ecológicos foi verificada a partir do teste de
Shapiro-Wilk, adotando-se o valor de significância de p < 0,05. Dos 4 descritores testados
(riqueza, abundância, diversidade e equitatividade), todos não corresponderam ao pressuposto
de normalidade e, estes apresentaram distribuição não-paramétrica dos dados.
A posteriori, com o objetivo de normalizar a distribuição dos dados, todos os
descritores foram transformados com a fórmula matemática (log+1). Mesmo após a
transformação, a distribuição dos dados continuou não-paramétrica, então foram aplicados
testes não-paramétricos para análise dos dados.
O teste não paramétrico de Mann-Whitney foi utilizado para verificar a variabilidade
temporal entre os períodos seco e chuvoso, enquanto para verificar a variabilidade espacial
entre as isóbatas e os transectos foi utilizado o teste não paramétrico de Kruskal-Wallis
seguido do teste U para comparação par-a-par. Foi adotado para ambos os testes o mesmo
valor de significância (p < 0,05).
Foi realizada um escalonamento multidimensional não-métrico (nMDS) utilizando o
índice de Bray-Curtis, com o intuito de comparar a composição da fauna através da
dissimilaridade das abundâncias dos táxons temporalmente (períodos seco e chuvoso) e
espacialmente (entre as isóbatas e transectos). Posteriormente foi feita uma análise de
similaridade (ANOSIM) buscando identificar diferenças significativas na ordenação nMDS.
Foi realizada uma análise de correspondência canônica (CCA) utilizando os
parâmetros ambientais e os táxons com abundância superior a 1, com o intuito de encontrar
possíveis correlações entre os parâmetros ambientais com a abundância dos organismos.
Posteriormente foi feito um teste de Permutação randomizado 9999 vezes para verificar a
significância dos eixos, enquanto a significância dos parâmetros ambientais foi testada por
modelos lineares generalizados (GLM) com a distribuição de Poisson.
Para análises multivariadas da ordenação nMDS e CCA, assim como o teste GLM, foi
aplicada logaritmização (log+1) para corrigir possíveis assimetria na distribuição dos dados,
enquanto para a abundância foi tirada a raiz da proporção de cada táxon aplicando o método
de transformação de Hellinger.

20
4.5. Programas estatísticos
Para o cálculo dos descritores ecológicos (riqueza, abundância, diversidade e
equitatividade) e da análise de correspondência canônica (CCA) foi utilizado o software
PAST 2.16. O teste de Shapiro-Wilk, Mann-Whitney, Kruskal-Wallis e os modelos lineares
generalizados (GLM) foram realizados no software R 3.3.1. O escalonamento
multidimensional não-métrico (nMDS) e o teste de similaridade (ANOSIM) foram feitos a
partir do software PRIMER 5.0. Já para a elaboração dos gráficos foi utilizado o software
Graph PadPrism 5.0.1.
5. RESULTADOS
5.1. Composição da fauna de bivalves A fauna de bivalves obtida esteve representada por uma abundância total de 1.384
indivíduos distribuídos em 46 táxons. Os organismos coletados pertencem a 20 famílias, 26
gêneros e 28 espécies (Tabela 1).
As famílias Arcidae e Mytilidae foram as que apresentaram maior riqueza (S = 7,
cada), seguida de Corbulidae (S = 6), em contrapartida, 10 famílias foram representadas
apenas por 1 táxon (Semelidae, Nuculanidae, Propeamussiidae, Crassatellidae, Myidae,
Noetiidae, Ostreidae, Ungulinidae, Chamidae e Spondylidae).
Em termos de abundância, a família Arcidae foi a mais numerosa (N = 906 ind. e Fr =
65,4), seguida de Pteriidae (N = 123 ind. e Fr = 8,8) e por Mytilidae (N = 97 ind. e Fr = 7%),
que juntas representaram 81,2% das famílias distribuídas (Figura 3).
Figura 3: Representação dos 10 táxons mais abundantes da fauna de bivalves associados às macroalgas provenientes das bacias sedimentares de Sergipe/Alagoas e Jacuípe, referentes aos períodos seco e chuvoso do ano de 2011. Legenda: Bivalvia = Bivalvia não identificado.

21
Já a espécie mais abundante foi Arca zebra (N = 549 ind. e Fr. 40%), seguida da
Anadara sp.1 (N = 311 ind. e Fr = 22,5%) e por Pinctada imbricata (N = 117 ind. e Fr =
8,5%) que juntas representaram 71% da abundância total dos indivíduos encontrados,
enquanto as outras 25 espécies, com frequência menor que 8%, representaram 29% dos
indivíduos.
A diversidade geral dos bivalves coletados foi de 2,083 bits/ind, e a equitatividade de
0,544, ambos os valores dos índices foram considerados intermediários.
Tabela 1: Abundância total de indivíduos indicados por família, gênero e espécie da fauna de bivalves associados às macroalgas provenientes das bacias sedimentares de Sergipe/Alagoas e Jacuípe, referentes aos períodos seco e chuvoso do ano de 2011.
Família Espécie Sigla Abundância
Corbulidae Corbula operculata (Philippi, 1848) Cop 50
Corbula sp. Cosp 1
Caryocorbula contracta (Say, 1822) Cco 1
Caryocorbula aff. contracta Cac 1
Caryocorbula swiftiana (Adams, 1852) Csw 2
Caryocorbula chittyana (Adams, 1852) Cch 6
Arcidae Arca zebra (Swainson, 1833) Aze 549
Anadara aff. notabilis Aan 8
Anadara sp. 1 An1 311
Anadara sp. 2 An2 6
Barbatia domingensis (Lamarck, 1819) Bdo 29
Barbatia aff. cândida Bac 1
Fugleria tenera (C. B. Adams, 1845) Fte 1
Mytilidae Amygdalum sagittatum (Rehder, 1935) Asa 2
Musculus lateralis (Say, 1822) Mla 24
Botula fusca (Gmelin, 1791) Bfu 1
Dacrydium vitreum (Møller, 1842) Dvi 43
Crenella decussata (Montagu, 1808) Cde 12
Mytilidae sp. 1 My1 2
Mytilidae sp. 2 My2 13
Pectinidae Spathochlamys benedicti (Verrill & Bush in Verrill, 1897)
Sbe 11
Leptopecten bavayi (Dautzenberg, 1900) Lba 5
Lindapecten sp. Lsp 2
Pteriidae Pinctada imbricata (Röding, 1798) Pim 117
Pteria colymbus (Röding, 1798) Pco 6
Philobryidae Cratis antillensis (Dall, 1881) Can 55
Cratis sp. Csp 6
Veneridae Gouldia cerina (C. B. Adams, 1845) Gce 5
Cyclinella sp. Cysp 1
Veneridae sp. Vsp 1

22
Lyonsiidae Lyonsia sp. Lsp 1
Semelidae Cumingia lamellosa (Sowerby I, 1833) Cla 1
Nuculanidae Nuculana concentrica (Say, 1824) Nco 2
Propeamussiidae Parvamussium pourtalesianum (Dall, 1886) Ppo 1
Crassatellidae Crassinella lunulata (Conrad, 1834) Clu 6
Myidae Sphenia fragilis (H. Adams & A. Adams, 1854) Sfr 19
Gastrochaenidae Lamychaena hians (Gmelin, 1791) Lhi 2
Gastrochaenidae sp. Gsp 1
Noetiidae Arcopsis adamsi (Dall, 1886) Aad 6
Ostreidae Crassostrea sp. Crsp 1
Ostreidae sp. Osp 15
Ungulinidae Ungulinidae sp. Ung 8
Limidae Limidae sp. 1 Li1 3
Limidae Limidae sp. 2 Li2 2
Chamidae Chamidae sp. Cha 1
Spondylidae Spondylidae sp. Spo 1
Bivalvia não identificado
- Biv 42
5.2. Distribuição temporal da fauna de bivalves
Dentre todos os descritores ecológicos utilizados, apenas o descritor de abundância
diferiu significativamente (p = 0,01) entre os dois períodos sazonais amostrados, com a maior
abundância obtida no período chuvoso (N = 1.108 ind.), e a menor no período seco (N = 276
ind.) (Figura 4).
Dos táxons encontrados, 15 ocorreram exclusivamente no período seco, 15
exclusivamente no chuvoso e 16 em ambos (Tabela 2).

23
Figura 4: Variação sazonal e desvio padrão dos índices ecológicos riqueza, abundância, diversidade e equitatividade, e o nível de significância (p) obtido a partir do teste Mann-Whitney, da fauna de bivalves associados às macroalgas provenientes das bacias sedimentares de Sergipe/Alagoas e Jacuípe, referentes aos períodos seco e chuvoso do ano de 2011. Legenda: * = variável significativa.
A partir do teste ANOSIM, não foi evidenciada variação significativa na composição
dos táxons entre os períodos seco e chuvoso (ANOSIM, p = 0,15 e R Global = 0,06). A
similaridade sazonal encontrada no teste anterior também ficou expressa no gráfico gerado
pela ordenação nMDS (Figura 5).

24
Figura 5: Escalonamento multidimensional não métrico (nMDS) agrupando a abundância por estações e períodos sazonais da fauna de bivalves associados às macroalgas provenientes das bacias sedimentares de Sergipe/Alagoas e Jacuípe, referentes aos períodos seco e chuvoso do ano de 2011.
5.3. Distribuição espacial da fauna de bivalves pelas isóbatas
Na distribuição espacial ao longo do gradiente batimétrico (isóbatas), foi evidenciado
que apenas o descritor ecológico de riqueza apresentou diferença significativa (p = 0,03)
(Figura 6). A isóbata de 50 m, região de maior profundidade, foi encontrada o maior número
de táxons (S = 31 táxons), seguida pela de 25 m (S = 18 táxons) e pela de 10 m (S= 15
táxons).
Apesar de a abundância não ter apresentado diferença significativa entre as isóbatas, o
maior número de indivíduos foi encontrados na isóbata de 50 m (N = 833 ind.), seguida pela
de 10 m (N = 411 ind.) e pela de 25 m (N = 140 ind.).
Também foi observada uma tendência de aumento dos descritores ecológicos de
diversidade e equitatividade nas isóbatas mais profundas, de modo que, a diversidade foi
considerada intermediária na isóbata de 50 m (H’ = 1,02) e baixa nas demais 25 m (H’ = 0,71)
e 10 m (H’ = 0,41), enquanto as isóbatas de 50 e 25 m foram consideradas mais equitativas (J
= 0,55 e 0,54 respectivamente) em comparação com a de 10 m (J = 0,33) (Figura 5). Dos
táxons encontrados 6 ocorreram exclusivamente na isóbata de 10 m, 5 na 25 m e 20 na de 50
m. O restante dos táxons foram amostrados em duas ou em todas as isóbatas (Tabela 2).

25
Figura 6: Variação e desvio padrão dos índices ecológicos de riqueza, abundância, diversidade e equitatividade, e o nível de significância (p) obtido a partir do teste Kruskal-Wallis, da fauna de bivalves associados às macroalgas provenientes das bacias sedimentares de Sergipe/Alagoas e Jacuípe, referente as isóbatas de 10, 25 e 50 m amostradas nos período seco e chuvoso do ano de 2011. Legenda: * = variável significativa.
O teste ANOSIM não evidenciou variação significativa da composição dos táxons
entre as isóbatas (ANOSIM, p = 0,40 e R = 0,01). A similaridade dos táxons encontrada no
teste anterior também ficou expressa no gráfico gerado pela ordenação nMDS (Figura 7).

26
Figura 7: Escalonamento multidimensional não métrico (nMDS) agrupando a abundância por estações e isóbatas de 10, 25 e 50 m da fauna de bivalves associados às macroalgas provenientes das bacias sedimentares de Sergipe/Alagoas e Jacuípe, referentes aos períodos seco e chuvoso do ano de 2011.
5.4. Distribuição espacial da fauna de bivalves pelos transectos
As espécies Arca zebra e Pinctada imbricata foram as mais bem distribuídas ao longo
das bacias sedimentares de Sergipe/Alagoas e Jacuípe, ocorrendo em 6 dos 8 transectos
amostrados (A, B, D, F, G e H) (Tabela 2).
Com relação a variabilidade espacial entre os transectos, nenhum dos descritores
ecológicos apresentou diferença significativa (p > 0,05) (Figura 8).
Embora não tenha sido evidenciada variação significativa, a riqueza mostrou-se
numericamente maior nos transectos D (S = 36 táxons), seguido do F (S = 29 táxons), A (S =
19 táxons), H (S = 17 táxons), B (S = 16 táxons), G (S = 12 táxons), E (S = 3 táxons) e o C (S
= 1 táxons).
A abundância mostrou-se maior no transecto D (N = 533 ind.), seguido do H (N = 348
ind.), F (N = 308 ind.), A (N = 87 ind.), G (N = 57 ind.), B (N = 45 ind.), E (N = 5 ind.) e pelo
C (N = 1 ind.).
A diversidade foi considerada intermediária apenas no transecto D (H’ = 1, 23) e baixa
nos demais transectos. Já em relação a equitatividade, apenas os transectos A, D e F foram
considerados equitativos (J = 0,68, 0,61 e 0,50 respectivamente) (Figura 7).

27
Figura 8: Variação e desvio padrão dos índices ecológicos de riqueza, abundância, diversidade e equitatividade, e o nível de significância (p) obtido a partir do teste Kruskal-Wallis, da fauna de bivalves associados às macroalgas provenientes das bacias sedimentares de Sergipe/Alagoas e Jacuípe, referentes aos transectos amostrados nos períodos seco e chuvoso do ano de 2011.
O teste ANOSIM não evidenciou nenhuma variação significativa na composição dos
táxons entre os transectos amostrados ao longo das bacias (ANOSIM, p = 0,40 e R = - 0,01).
A similaridade dos táxons encontrada no teste anterior também ficou expressa no gráfico
gerado pela ordenação nMDS (Figura 9).

28
Figura 9: Escalonamento multidimensional não métrico (nMDS) agrupando a abundância por estações e transectos da fauna de bivalves associados às macroalgas provenientes das bacias sedimentares de Sergipe/Alagoas e Jacuípe, referentes aos períodos seco e chuvoso do ano de 2011.
Tabela 2: Fauna geral de bivalves associados às macroalgas das bacias sedimentares de Sergipe/Alagoas e Jacuípe amostrados no ano de 2011, indicando o período sazonal, transectos e suas respectivas isóbatas onde foram encontrados.
Táxons Período seco Período chuvoso
Transectos Isóbatas (m) Transectos Isóbatas (m) Corbula operculata D e F 25 e 50 A e G 50 Corbula sp. D 50 _ _ Caryocorbula contracta F 50 _ _ Caryocorbula aff. Contracta D 50 _ _ Caryocorbula swiftiana F 25 D 25 Caryocorbula chittyana C e F 25 _ _ Arca zebra B, D, F, G e H 25 e 50 A, B, D, F e H 25 e 50 Anadara aff. notabilis D 50 D, F e G 25 e 50 Anadara sp 1 _ _ E, G e H 10 e 25 Anadara sp 2 _ _ A, B, E e H 10, 25 e 50 Barbatia domingensis F 50 D 50 Barbatia aff. cândida A 10 _ _ Fugleria tenera _ _ D 25 Amygdalum sagittatum _ _ H 10 Musculus lateralis B e F 25 e 50 A, D e F 10, 25 e 50 Botula fusca F 50 _ _ Dacrydium vitreum F 50 B, D e F 50 Crenella decussata A, D e F 50 A e G 50 Mytilidae 1 _ _ H 10 Mytilidae 2 _ _ H 10 Spathochlamys benedicti B, D e F 25 e 50 A, B, D e F 50 Leptopecten bavayi A 10 e 25 H 10

29
Lindapecten sp. F 25 _ _ Pinctada imbricata A, B, D e F 10, 25 3 50 A, B, D, F, G e
H 10, 25 e 50
Pteria colymbus _ _ A e D 10 e 25 Cratis antillensis B e D 50 B e D 50 Cratis sp. D e F 50 _ _ Gouldia cerina B 50 D e G 25 e 50 Cyclinella sp. D 50 _ _ Veneridae sp. A 50 _ _ Lyonsia sp. D 50 _ _ Cumingia lamellosa D 25 _ _ Nuculana concentrica D 50 E 50 Parvamussium pourtalesianum
D 50 _ _
Crassinella lunulata F 50 B, D, F e G 25 e 50 Sphenia fragilis A 10 G e H 10 e 25 Lamychaena hians _ _ A e H 10 e 50 Gastrochaenidae sp. _ _ B 50 Arcopsis adamsi _ _ A, D e F 10 e 50 Crassostrea sp. A 10 _ _ Ostreidae sp. _ _ G e H 10 Ungulinidae sp. F 50 _ _ Limidae 1 _ _ B e D 50 Limidae 2 _ _ D 50 Chamidae sp. _ _ F 50 Spondylidae sp. _ _ D 50 Bivalvia - não identificado A, B, D e F 10, 25 e 50 A, B, D, E, F, G
e H 10, 25 e 50
5.5. Correlação da fauna com os parâmetros ambientais
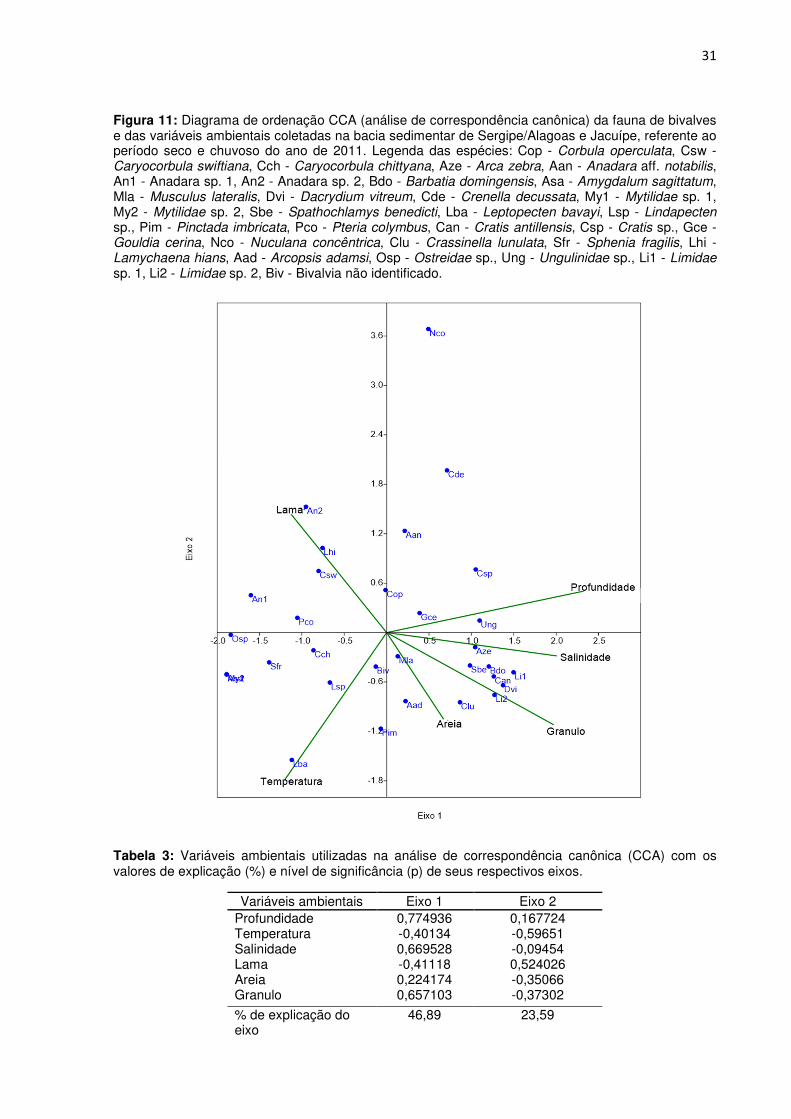
Ao relacionar a abundância dos bivalves pelas estações com as seis variáveis
ambientais (profundidade, temperatura, salinidade, lama, areia e granulo) a partir da análise
de correspondência canônica (CCA), foi possível evidenciar duas associações faunísticas
distintas representadas pelos dois primeiros eixos.
Apenas o eixo 1 foi significativo na estruturação da fauna (p = 0,0008) explicando
46% da distribuição dos bivalves e, os parâmetros ambientais mais relacionados a este eixo
foram a profundidade, salinidade e os sedimentos de fundo do tipo granulo e de areia (Tabela
3). Por sua vez, este eixo envolveu as estações A3, B2, B3, D2, D3, F3 e G3 (período seco), e
as estações B2, B3, D3, G3, F2 e F3 (período chuvoso) (Figura 9). Nesta associação esteve
presente a espécie mais representativa da área de estudo, sendo esta a Arca zebra. Outros
táxons também estiveram presentes de forma representativa a exemplo da Anadara sp 1,
Cratis antillensis e Dacrydium vitreum (Figura 10).
O eixo 2, por sua vez não significativo estatisticamente (p = 0,3973), explicou 23% da
distribuição dos bivalves e, os parâmetros ambientais mais relacionados a este foram a
temperatura e o sedimento de fundo do tipo lama. O eixo envolveu A1, C2 e F2 (período
seco), e as estações A1, A3, B1, D2, E1, E2, G1, G2, H1 e H2 (período chuvoso) (Figura 9).

30
A maior abundância desta associação esteve representada por Pinctada imbricata (Figuras
10).
Quando comparada a abundância com os parâmetros ambientais a partir dos modelos
lineares generalizados (GLM) (Tabela 4), foi observado que apenas o sedimento de fundo
arenoso foi significativo isoladamente (p = 0,00101), porém, quando feita a interação entre as
seis variáveis ambientais amostradas, três interações mostraram-se significativas, sendo estas
a profundidade com a salinidade (p = 0,01232), areia e granulo (p = 0,03229) e a
profundidade, temperatura e salinidade (p = 0,01145), todas estas positivas com a distribuição
da fauna de bivalves nas bacias sedimentares amostradas.
Figura 10: Diagrama de ordenação CCA (análise de correspondência canônica) dos pontos amostrais e das variáveis ambientais coletadas na bacia sedimentar de Sergipe/Alagoas e Jacuípe, referente ao período seco e chuvoso do ano de 2011.
31
Figura 11: Diagrama de ordenação CCA (análise de correspondência canônica) da fauna de bivalves e das variáveis ambientais coletadas na bacia sedimentar de Sergipe/Alagoas e Jacuípe, referente ao período seco e chuvoso do ano de 2011. Legenda das espécies: Cop - Corbula operculata, Csw - Caryocorbula swiftiana, Cch - Caryocorbula chittyana, Aze - Arca zebra, Aan - Anadara aff. notabilis, An1 - Anadara sp. 1, An2 - Anadara sp. 2, Bdo - Barbatia domingensis, Asa - Amygdalum sagittatum, Mla - Musculus lateralis, Dvi - Dacrydium vitreum, Cde - Crenella decussata, My1 - Mytilidae sp. 1, My2 - Mytilidae sp. 2, Sbe - Spathochlamys benedicti, Lba - Leptopecten bavayi, Lsp - Lindapecten sp., Pim - Pinctada imbricata, Pco - Pteria colymbus, Can - Cratis antillensis, Csp - Cratis sp., Gce - Gouldia cerina, Nco - Nuculana concêntrica, Clu - Crassinella lunulata, Sfr - Sphenia fragilis, Lhi - Lamychaena hians, Aad - Arcopsis adamsi, Osp - Ostreidae sp., Ung - Ungulinidae sp., Li1 - Limidae sp. 1, Li2 - Limidae sp. 2, Biv - Bivalvia não identificado.
Tabela 3: Variáveis ambientais utilizadas na análise de correspondência canônica (CCA) com os valores de explicação (%) e nível de significância (p) de seus respectivos eixos.
Variáveis ambientais Eixo 1 Eixo 2
Profundidade 0,774936 0,167724 Temperatura -0,40134 -0,59651 Salinidade 0,669528 -0,09454 Lama -0,41118 0,524026 Areia 0,224174 -0,35066 Granulo 0,657103 -0,37302
% de explicação do eixo
46,89 23,59

32
Significância do eixo (p)
0,0008 0,3973
Tabela 4: Modelos lineares generalizados (GLM) realizados entre a abundância dos bivalves e os parâmetros ambientais amostrados, incluindo os seus respectivos os valores de significância (p) dos parâmetros ambientais e suas interações, da bacia sedimentar de Sergipe/Alagoas e Jacuípe, referente ao período seco e chuvoso do ano de 2011. Legenda: * = variável significativa.
Parâmetros Abundância (p)
Profundidade 0,38671
Temperatura 0,34357
Salinidade 0,24432
Lama 0,05490
Areia 0,00101*
Granulo 0,62971
Profundidade:Temperatura 0,85081
Profundidade:Salinidade 0,01232*
Temperatura:Salinidade 0,09929
Profundidade:Lama 0,05120
Temperatura:Lama 0,50842
Salinidade:Lama 0,39831
Profundidade:Areia 0,7269
Temperatura:Areia 0,78722
Salinidade:Areia 0,76690
Lama:Areia 0,84388
Profundidade:Granulo 0,14857
Temperatura:Granulo 0,50967
Salinidade:Granulo 0,55612
Lama:Granulo 0,13964
Areia:Granulo 0,03229*
Profundidade:Temperatura:Salinidade 0,01145*
Profundidade:Temperatura:Lama 0,20765
Profundidade:Salinidade:Lama 0,99757
Temperatura:Salinidade:Lama 0,12626
Profundidade:Temperatura:Areia 0,73695
Profundidade:Salinidade:Areia 0,30485
Temperatura:Salinidade:Areia 0,01511*
Profundidade:Lama:Areia 0,78246
6. DISCUSSÃO
A fauna encontrada associada às macroalgas do fital das bacias sedimentares de
Sergipe/Alagoas e Jacuípe apresentou uma riqueza de 46 táxons, ampliando o número de
famílias e espécies não só para a costa de Sergipe, já que a metodologia utilizada para
obtenção dos bivalves foram a partir de arrastos pesqueiros, e estes não fazem parte da

33
metodologia usual para estudo dos organismos associados às macroalgas componentes dos
fitais.
Embora poucos estudos recentes abordando a distribuição da malacofauna associada
às macroalgas componentes do fital tenham sido feitos na região Nordeste do Brasil, Bezerra
(2011) evidenciou 14 espécies de bivalves associados à macroalga Halimeda opuntia na praia
do Cupe em Ipojuca (PE), Veras (2011) evidenciou 11 espécies de Bivalvia associados a
macroalga Pterocladiella caerulescens na zona entremarés da praia da pedra rachada (CE) e
Almeida (2007) evidenciou 6 espécies de bivalves associados a macroalga Sargassum sp. em
Pontal do Cupe (PE).
Rocha & Martins (1998) comparando a distribuição da malacofauna em seis diferentes
tipos de substratos na plataforma continental do Ceará observaram um maior número de
espécies associadas ao substrato formado por algas calcárias, incluindo 18 espécies de
bivalves. Estes mesmos autores ainda comentam que a maior riqueza de indivíduos associados
a este tipo de alga deveu-se a mesma propiciar maior disponibilidade de superfícies para
fixação, refúgio contra predadores e alimento disponível.
Dois trabalhos realizados na costa do Nordeste Brasileiro também encontraram a
ocorrência da espécie Arca zebra, a mais abundante numericamente deste estudo, sendo o
primeiro registrado por Silva (2014) entre as profundidades de 6 e 60 m na plataforma
continental da região semiárida do Brasil, incluindo os estados do Ceará, Piauí e o leste do
Maranhão, porém as abundâncias e o tipo de substrato onde foram encontradas não foram
descritos no estudo. O segundo trabalho foi registrado por Rocha & Martins (1998) na
plataforma continental do Ceará, onde a espécie ocorreu entre as profundidades de 10 a 27 m
em substratos de areias quartozas e de algas calcárias, compondo cerca de 22% da
composição dos organismos encontrados.
É válido ressaltar que todos os trabalhos citados apresentaram áreas de amostragem de
variadas extensões e diferentes esforços amostrais, além do ambiente de coleta ter sido
diverso, grande parte dos trabalhos encontrados se referem a fitais de zonas de entre-marés e
não foram encontrados trabalhos discutindo os fitais associados a arrastos pesqueiros, objeto
do presente trabalho.
Parte dos indivíduos contabilizados neste estudo representavam juvenis, com conchas
sem as ornamentações características da espécie, o que contribuiu para a não identificação ao
menor nível taxonômico possível de algumas espécies.

34
A ocorrência de indivíduos em estado juvenil evidenciada poderia indicar que o fital
formado pelas macroalgas da região poderia ser um local de reprodução e berçário para tais
espécies, assim como relatado nos estudos de Almeida (2007), Veras (2011) e Bezerra (2011).
A fauna de bivalves mostrou-se se mais abundante no período chuvoso, quando
comparado com o seco. Guimarães (2010) estudando a estrutura e dinâmica da comunidade
megabêntica da plataforma continental de Sergipe, também encontrou variabilidade temporal
na fauna de moluscos, marcada pela alta abundância destes no período chuvoso.
A área de estudo do presente trabalho é caracterizada por não apresentar as quatro
estações do ano bem definidas, mas a sazonalidade é refletida pela variação pluviométrica
caracterizada por chuvas mais abundantes no período outono-inverno (UFS/SEPLAN 1979).
Guimarães (2010) ainda comenta sobre a ocorrência de dois períodos distintos no estado de
Sergipe, seco e chuvoso, estes influenciados pela variação da pluviosidade ao longo do ano,
com maior pluviosidade no inverno e pelas entradas de massas de ar mais frias no verão com
pequenas resurgências a partir dos cânions do São Francisco e Japaratuba, sendo esses
fenômenos ao longo do ano, relevantes na distribuição dos organismos na plataforma
continental.
O sedimento de fundo pareceu ser o principal estruturador da fauna coletada, de modo
que, assim como observado por Guimarães (2010), onde nas regiões próximas à costa da
plataforma continental de Sergipe há o predomínio de lamas terrígenas e águas mais turvas,
ambiente não muito propício para o estabelecimento e desenvolvimento das macroalgas, por
outro lado, nas regiões mais afastadas dos aportes fluviais com águas claras e com a
ocorrência de manchas de sedimentos mais grossos, a presença das macroalgas pode ser mais
evidente e estas podem oferecer microhabitats propícios para a colonização de diversas
espécies na plataforma, inclusive de bivalves. Em estudo realizado da plataforma continental
semiárida no Nordeste do Brasil, Silva (2014) notou a predominância de banco de algas em
regiões constituídas em sedimentos do tipo cascalho, de areias siliciclásticas e carbonáticas,
de tal forma que, os moluscos também estiveram associados a estes.
As maiores abundâncias e riqueza foram encontradas concentradas nas isóbatas mais
profundas e fora da influencia do sedimento de lama fluvial (transectos D e F), relacionadas
aos sedimentos mais grossos. Diversos trabalhos também evidenciaram a influencia do
substrato de fundo como responsável pela distribuição de comunidades bentônicas, a exemplo
de Pires-Vanin (1989) na plataforma continental da região Norte do estado de São Paulo;
Rocha & Martins (1998) na plataforma continental do Ceará; Gomes (2006) na plataforma
continental e talude superior de Cabo Frio, Rio de Janeiro; Guimarães (2010) na plataforma

35
continental de Sergipe e Silva (2014) na plataforma continental da região semiárida do
Nordeste do Brasil que estudou a distribuição de moluscos bentônicos e sua relação com o
sedimento de fundo, inclusive de algas.
A variação da profundidade apresentou relevância ao evidenciar uma separação dos
táxons entre os ambientes de 10 e 25 m do ambiente de 50 m, de modo que a riqueza diferiu
significativamente, esta maior na isóbata de 50 m, enquanto a abundância e diversidade da
fauna tiveram uma tendência de aumento nas isóbatas mais profundas. É possível inferir que
esta variação tenha ocorrido devido a heterogeneidade de tipos de fundos onde os sedimentos
mais grossos encontram-se nas isóbatas mais profundas.
Soares-Gomes e Pires-Vanin (2003) também evidenciaram maiores abundância e
diversidade da comunidade de bivalves na isóbata de 50 m na porção interna da plataforma
continental do norte de São Paulo. Soares-Gomes & Fernandes (2005) na plataforma
continental de Cabo frio, Rio de Janeiro, observaram o mesmo padrão para a comunidade de
bivalves amostrada entre as isóbatas de 45 e 60 m, porém nos trabalhos citados não foram
constatados o substrato algal nos seus resultados. Soares-Gomes & Fernandes (2005) ainda
associaram o padrão encontrado desses índices a maior heterogeneidade espacial nessas
regiões, seja esta relacionada a disponibilidade de alimento ou aos microhabitats existentes
nestas.
Assim como também existem diferentes fundos sedimentares nas diferentes isóbatas, a
plataforma continental de Sergipe também é fortemente influenciada pela pluma sedimentar
carreada pelo rio São Francisco e, esta pareceu atuar negativamente sobre o transecto mais
próximo à sua foz (C), assim como as isóbatas mais rasas ao longo de toda a plataforma do
estado de Sergipe. Acredita-se que, a influência terrígena exercida pela pluma sedimentar
tenha sido o motivo da baixa riqueza e abundância das espécies de bivalves por não
oferecerem às macroalgas substrato propicio para seu crescimento.
Foi observada também três interações significativas distintas encontradas a partir da
correlação dos parâmetros ambientais com a abundância dos bivalves, as duas primeiras
envolvendo o aumento da temperatura e do teor de salinidade com o decorrer do aumento da
profundidade, e a terceira envolvendo os tipos de fundo formados pela areia e pelo granulo.
Todas estas interações apresentaram-se positivas, com boa parte da fauna de bivalves
encontrada nas estações envolvidas com estas características.
7. CONSIDERAÇÕES FINAIS

36
- A fauna dos moluscos bivalves encontrada associada às macroalgas do fital das bacias
sedimentares de Sergipe/Alagoas e Jacuípe esteve representada por 46 táxons que, dentre
estes, a espécie Arca zebra foi a mais representativa, dominando numericamente ambos os
períodos sazonais amostrados.
- A maior abundância de organismos foi encontrada no período sazonal chuvoso e, este
ocorrido pode estar relacionado as pequenas variações dos fatores abióticos ao longo do ano,
especificamente nos dois únicos períodos existentes (seco e chuvoso) da área de estudo.
- O sedimento de fundo, a profundidade e a salinidade pareceram ser os principais fatores
estruturadores da fauna encontrada, de modo que, a abundância e diversidade dos bivalves
tiveram uma tendência de aumento nas isóbatas mais profundas. É possível inferir que esta
variação tenha ocorrido devido a maior heterogeneidade de fundos sedimentares encontrados
na plataforma nas isóbatas mais profundas e dos microhabitats disponíveis nestes ambientes.
- A ocorrência de indivíduos em estado juvenil evidenciada poderia indicar que o fital
formado pelas macroalgas da região poderia ser um local de reprodução e berçário para tais
espécies, assim como relatado em vários estudos anteriores descrevendo as comunidades de
bivalves associadas às macroalgas.
8. REFERÊNCIAS
ALMEIDA, S.M. 2007. Malacofauna associada ao fital de Sargassum spp no Pontal do Cupe,
Ipojuca, PE. Dissertação de mestrado, Universidade Federal de Pernambuco, Recife.
ALVES, M.S. & ARAUJO, M.J.G. 1999. Moluscos associados ao fital Halodule wrightii
Aschers na Ilha de Itamaracá – PE. Trab. Oceanog. Univ. Fed. PE. 27(1):91-97.
AMARAL, A.C.Z, LANA, P.C., FERNANDES, F.C. & COIMBRA, J.C. 2003. Programa
REVIZEE Score Sul – Bentos. Biodiversidade bêntica da região sul-sudeste da costa
brasileira. MMA. São Paulo.
ANP – Agência Nacional do Petróleo, Gás Natural e Biocombustíveis.2017. Disponível em: http://www.anp.gov.br/wwwanp/publicacoes acesso: 12 de Outubro de 2017.
BARROS, G. 2015. Estrutura e composição da malacofauna associada à macroalgas em um ambiente recifal neotropical. Trabalho de conclusão de curso, Universidade Estadual da Paraíba, Campina Grande.
BEZERRA, M.G. 2011. Malacofauna associada ao fital de Halimeda opuntia (Linnaeus)
J.V.L amouroux no Pontal do Cupe, Ipojuca – PE, Brasil. Dissertação de mestrado,
Universidade Federal de Pernambuco, Recife.

37
BOFFI, K.H. 1972. Ecological aspects of ophiuroids from phytal of S. W. Atlantic Ocean warm waters. Mar. Biol. 15(4):316-328.
BRUSCA, R.C. & BRUSCA, G.J. 2007. Invertebrados. 2° ed. Editora Guanabara-Koogan,
Rio de Janeiro.
CAPÍTOLI, R.R. & BEMVENUTI, C. 2004. Distribuição batimétrica e variações de diversidade dos macroinvertebrados bentônicos da plataforma continental e talude superior no extremo Sul do Brasil. Atlântica, Rio Grande. 26 (1):27-43.
CASTRO, P. & HUBER, M.E. 2012. Biologia marinha. 8. ed. AMGH, Porto Alegre.
CHEMELLO, R. & MILAZZO, M. 2002. Effect of algal architecture on associated fauna:
some evidence from phytal molluscs. Marine Biology. 140:981-990.
COSTA, I.O., CAIRES, T.A., FILHO, G.H.P. & NUNES, J.M.C. 2012. Macroalgas
bentônicas associadas a bancos de Hypnea musciformis (Wulfen) J.V. Lamour. (Rhodophyta
– Gigartinales) em duas praias do litoral baiano. Acta bot. bras. 26(2): 493-507.
COUTINHO, P.N. 1995-2000. Programa REVIZEE. Oceanografia geológica da região
nordeste. MMA/SMA.
EDGAR, G.J. & MOORE, P.G. 1986. Macro-algae as habitats for motile macrofauna. In
Monografias Biologicas. Simposio Internacional. Usos Funciones Ecologicas de las Algas
Marinas Bentonicas. 255-277.
FERNANDES, M.C. 2014. Estrutura da comunidade de Gastropoda e Bivalvia (Mollusca)
associados às algas pardas do gênero Sargassum C. agardh, 1820 (Sargassaceae) do canal de
São Sebastião, litoral norte do estado de São Paulo. Dissertação de mestrado, Universidade
Estadual de Campinas, Campinas.
FIGUEIREDO, M.A.O., HORTA, P.A., PEDRINI, A.G. & NUNES, J.M.C. 2008. Benthic
algae of the coral reefs of Brazil: a literature review. Oecol. Brasil. 12:258-269.
FREITAS, V. A. & FERREIRA, A. L. 2015. Bacia de Jacuípe: Sumário Geológico e Setores em Oferta. Agência Nacional do Petróleo.13° rodada de licitações.
GOMES, M. F. 2006. Variações espaciais e sazonais na composição e estrutura da comunidade macrobêntica na plataforma continental e talude superior de Cabo Frio, Rio de Janeiro, Brasil. Dissertação de mestrado, Universidade de São Paulo, São Paulo.

38
GUIMARÃES, C.R.P. 2010. Composição e distribuição dos sedimentos superficiais e da
fauna bêntica na plataforma continental de Sergipe. Tese de doutorado, Universidade Federal
da Bahia, Salvador.
GÜTH, A. Z., 2004. A comunidade fital: variação espacial e nictemeral da epifauna, especialmente anfípodos, associada à alga parda Sargassum spp. em quatro praias de Ubatuba, litoral Norte do estado de São Paulo. Dissertação de mestrado, Universidade Estadual de Campinas, Campinas.
HAESER, B. 2015. Bacia de Sergipe-Alagoas: Sumário Geológico e Setores em Oferta. Agência Nacional do Petróleo.13° rodada de licitações.
JACOBUCCI, G.B. & LEITE, F.P.P. 2002. Distribuição vertical e flutuação sazonal da
macrofauna vágil associada a Sargassum cymosum C. Agardh, na praia do Lázaro, Ubatuba,
São Paulo, Brasil. Revista Brasileira de Zoologia. 19:87-100.
JACOBUCCI, G.B.; GÜTH, A.Z.; TURRA, A.; MAGALHÃES, C.A.; DENADAI, M. R.;
Chaves, A.M.R. & SOUZA, E.C.F. 2006. Levantamento de Mollusca, Crustacea e
Echinodermata associados a Sargassum spp. na ilha da queimada pequena, estação ecológica
dos Tupiniquins, litoral Sul do estado de São Paulo, Brasil. Biota Neotropica. 6(2):1-8.
JOHNSON, S.C. & SCHEIBLING, R.E. 1987. Structure and dynamics of epifaunal
assemblages on intertidal macroalgae Ascophyllum nodosum and Fucus vesiculosus in Nova
Scotia, Canada. Mar. Eco. Prog. Ser. 37:209-227.
LACERDA, M.B., DUBIASKI-SILVA, J. & MASUNARI, S. 2009. Malacofauna de três fitais da Praia de Caiobá, Matinhos, Paraná. Acta Biológica Paranaense. 38(1-2):59-74.
LANA, P.C., CAMARGO, M.G., BROGIM, R.A. & ISAAC, V.J. 1996. Programa
REVIZEE. O bentos da costa brasileira. Avaliação crítica e levantamento bibliográfico (1858-
1996). MMA/CIRM/FEMAR. Rio de Janeiro.
MANSO, V.A.V.,VALENÇA, L.M.M., COUTINHO, P. N. & GUERRA, N. C. 2004.
Sedimentologia da Plataforma Continental. In Oceanografia um cenário tropical (E.
ESKINAZI-LEÇA, S. NEUMANN-LEITÃO & F.M. COSTA (org.). Bagaço, Recife, v. 1, p.
59-86.
MASUNARI, S. & FORNERIS, L. 1981. O ecossistema fital: uma revisão. In: Academia
Brasileira de Ciências (Ed.). Seminários de Biologia Marinha. Rio de Janeiro, p. 149-172.

39
MASUNARI, S. 1988. A associação entre Crepiscula aculeta (Gastropoda, Calyptraeidae) a alga calcárea Amphiroa beauvouisii, na Baía de Santos, São Paulo, Brasil. Revta bras. Zool. 5(2):293-310.
MASUNARI, S. Ecologia das comunidades fitais. 1987. In 1° simpósio sobre ecossistemas da
costa sul e sudeste brasileira-síntese dos conhecimentos. Academia de Ciências do Estado de
São Paulo. Cananéia, p. 195-310.
MIKKELSEN, P. M.; BIELER R. 2008. Seashells of southern Florida. Living marine mollusks of the Florida Keys and adjacent regions. Bivalves. Princeton University Press, Oxford.
MUNIZ, R.A., REIS, R.P., MARROIG, R. & AMADO-FILHO, G. 2013. Algas marinhas do monumento natural das ilhas Cagarras. In História, Pesquisa e Biodiversidade do Monumento Natural das Ilhas Cagarras – Série Livros 48. (F. MORAES, A. BERTONCINI & A. AGUIAR). 1 ed. Museu Nacional, Rio de Janeiro. p 49-61.
NASCIMENTO, E. 2007. O ecossistema fital: uma abordagem por fractais. In Anais do VIII
Congresso de Ecologia do Brasil, Caxambu/MG. p. 1.
NASCIMENTO, E.F.I. & ROSSO, S. 2007. Fauna associada à macroalgas marinhas
bentônicas (Rhodophyta e Phaeophyta) da região de São Sebastião, São Paulo. Universidade
de São Paulo, Instituto de Biociências. 38-52.
NIBBAKEN, J.W. 1982. Marine Biology: an ecological approach. Harper & Row, New York.
OLIVEIRA, E.C. 2002. Macroalgas marinhas da costa brasileira: estado do conhecimento,
usos e conservação biológica. In Biodiversidade, conservação e uso sustentável da flora do
Brasil (E.L. ARAÚJO, A.N. MOURA, E.V.S.B. SAMPAIO, L.M.S. GESTINARI & J.M.T.
CARNEIRO. UFRPE: Imprensa Universitária, Recife, p. 122-126.
ORFANIDIS, S., PANAYOTIDIS, P. & STAMATIS, N. 2001. Ecological evalution of
transitional and coastal Waters: A marine benthic macrophytes-based model. Mediterranean
Marine Science. 2:45-65.
PEREREIRA, P.H.C., FERREIRA, P.B. & REZENDE, S.M. 2010. Community structure of the ichthyofauna associated with seagrass beds (Halodule wrightii) in Formoso river estuary – Pernambuco, Brazil. In Anais da Academia Brasileira de Ciências. 82(3):617-628.
PETERSON, B.J. & HECK JR, K.L. 2001. Positive interactions between suspension-feeding
bivalves and seagrass - a facultative mutualism. Mar Ecol Prog Ser. 213:143-155.

40
PIELOU, E.C. 1969. An introdution to Mathematical Ecology. Wilwy-Intercience, New York.
PIELOU, E.C. 1975. Ecological Diversity. John Willey & Sons, Inc., New York.
PIRES-VANIN A.M.S. 1989. Estrutura e dinâmica da megafauna bêntica na plataforma continental da região Norte do Estado de São Paulo, Brasil. Tese de doutorado. Universidade de São Paulo, São Paulo.
QUEIROZ, A.C.M., LIMA, M.S. & MARTINS, I.X. 2007. Levantamento preliminar da fauna associada ao fital na praia do Pacheco, Caucaia, Ceará. In Anais do XII Congresso Latin-Americano de Ciências do Mar, Florianópolis. p. 1-3.
QUEIROZ, R.N.M. 2011. Inventário dos moluscos associados à macroalgas do gênero
Gracilaria (Rhodophyta), do manguezal hipersalino do Rio Tubarão (Macau, Rio Grande do
Norte). Trabalho de conclusão de curso, Universidade Estadual da Paraíba, Campina Grande.
REMANE, A. 1933. Verteilung und Organisation der benthonishen Microfauna der Kieler Bucht. Helgol. Wiss. Meeresunter. 21:161-221.
RIOS, E.C. 1994. Seashells of Brazil. 2 ed. Editora FURG, Rio Grande-RS.
RIOS, E.C. 2009. Compendium of Brazilian Sea Shells. Evangraf, . Rio Grande-RS.
ROCHA, C. A. & MARTINS, I. X. 1998. Estudo da macrofauna bentônica na plataforma continental do litoral oeste do estado do Ceará, Brasil. Arquivos de Ciências do Mar. 31:65-72.
ROCHA, V.P. 2011. Morfometria e anatomia de arcídeos (mollusca, bivalvia) da costa norte-
nordeste do Brasil. Dissertação de mestrado, Universidade Federal do Ceará, Fortaleza.
RUPPERT, E.E., BARNES, R.D & FOX, R.S. 2005. Zoologia dos invertebrados: uma
abordagem funcional-evolutiva. 7. ed. Roca, São Paulo, SP.
SILVA, A.F. 2014. Distribuição dos moluscos bentônicos e sua relação com o sedimento na plataforma continental da região semiárida do Nordeste do Brasil. Tese de doutorado, Universidade Federal do Ceará, Fortaleza.
SOARES-GOMES, A. & FERNANDES, F.C. 2005. Spatial distribution of bivalve mollusc assemblages in the upwelling ecosystem of the continental shelf of Cabo Frio, Rio de Janeiro, Brazil. Revista Brasileira de Zoologia. 22(1):73–80.
SOARES–GOMES, A. & PIRES–VANIN, A.M. 2003. Padrões de abundância, riqueza e
diversidade de moluscos bivalves na plataforma continental ao largo de Ubatuba, São Paulo,
Brasil: uma comparação metodológica. Rev. Bras. Zool. 20:717–725.

41
TARARAN, A.S. & WAKABARA, Y. 1981. The mobile fauna especially Gammaridea of
Sargassum cymosum. Mar. Ecol. Progress Series. 5:157-163.
TUNNELL-JR, J.W., ANDREWS J., BARRERA, N.C. & MORETZSOHN, F. 2010.
Encyclopedia of Texas Seashells - Identification, Ecology, Distribution & History. Harte
Research Institute for Gulf of Mexico Studies Series, College station.
UFS/SEPLAN. 1979. Atlas de Sergipe. Aracaju. UFS/SEPLAN. 95p.
VERAS, D.R.A. 2011. Moluscos associados à macroalga Pterocladiella caerulescens
(Rhodophyta, Pterocladiaceae) na zona entremarés da Praia da Pedra Rachada, Paracuru,
Ceará, Nordeste do Brasil. Dissertação de mestrado, Universidade Federal do Ceará,
Fortaleza.
VIEJO, R.M. 1999. Mobile epifauna inhabiting the invasive Sargassum muticum and two local seaweeds in Northern Spain. Aquat Bot. 64:131–149.