prezentace aplikace powerpoint - univerzita...
TRANSCRIPT

Kap. 6
Vývojové procesy určované
polohou ve vztahu k určité
buňce, pletivu či orgánu
(Origins of periodic patterns)


Ekvivalentní a neekvivalentní buňkyu živočichů a rostlin
• laterální (bočná) specifikace(vymezení)
• indukce
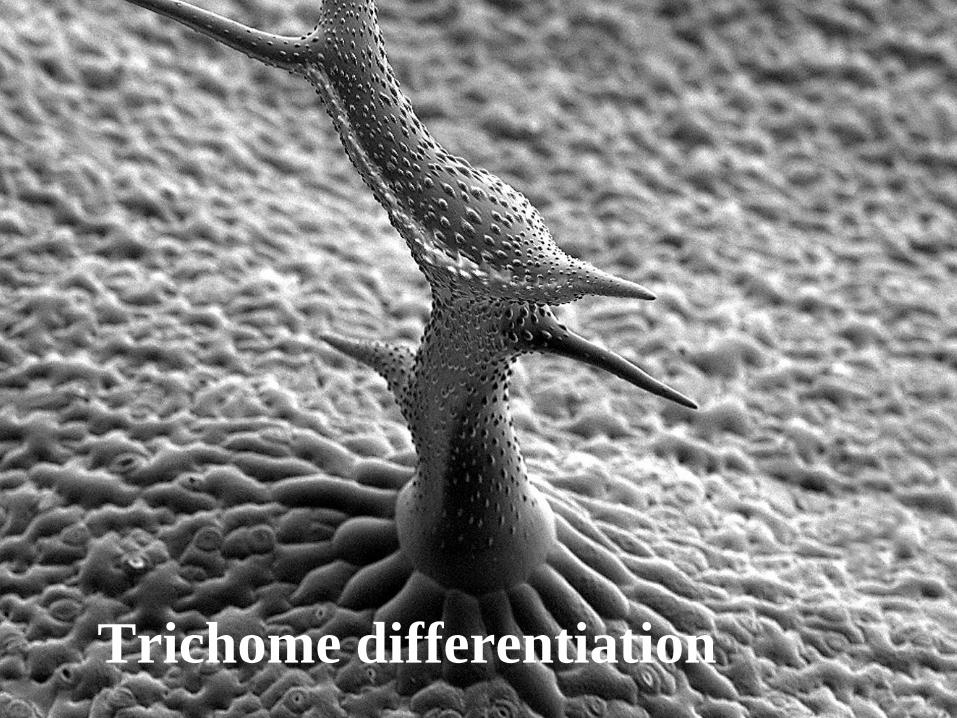
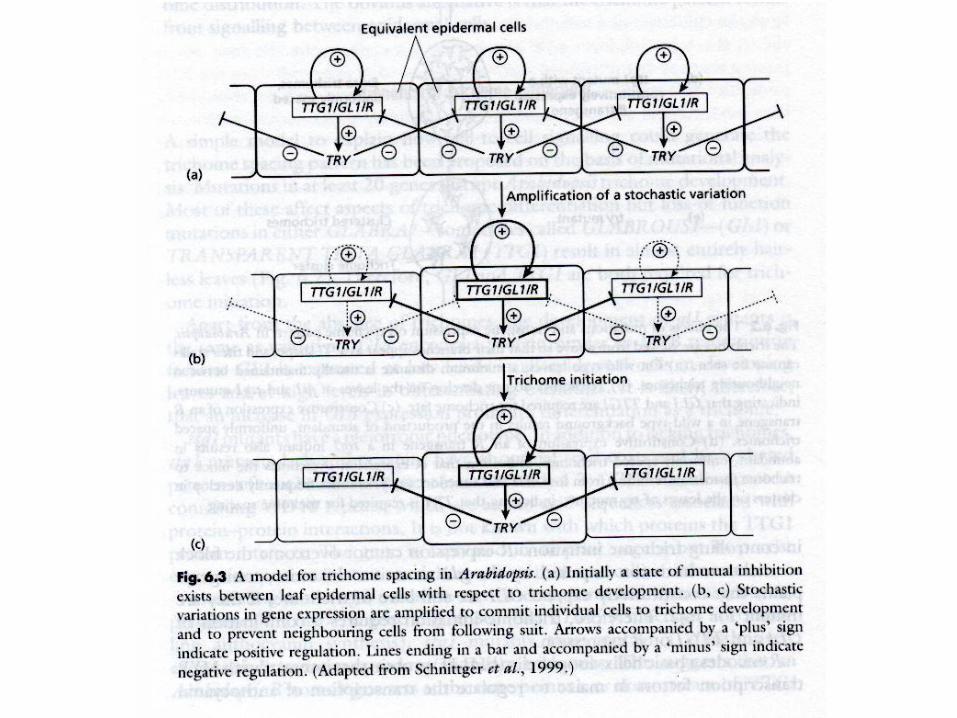
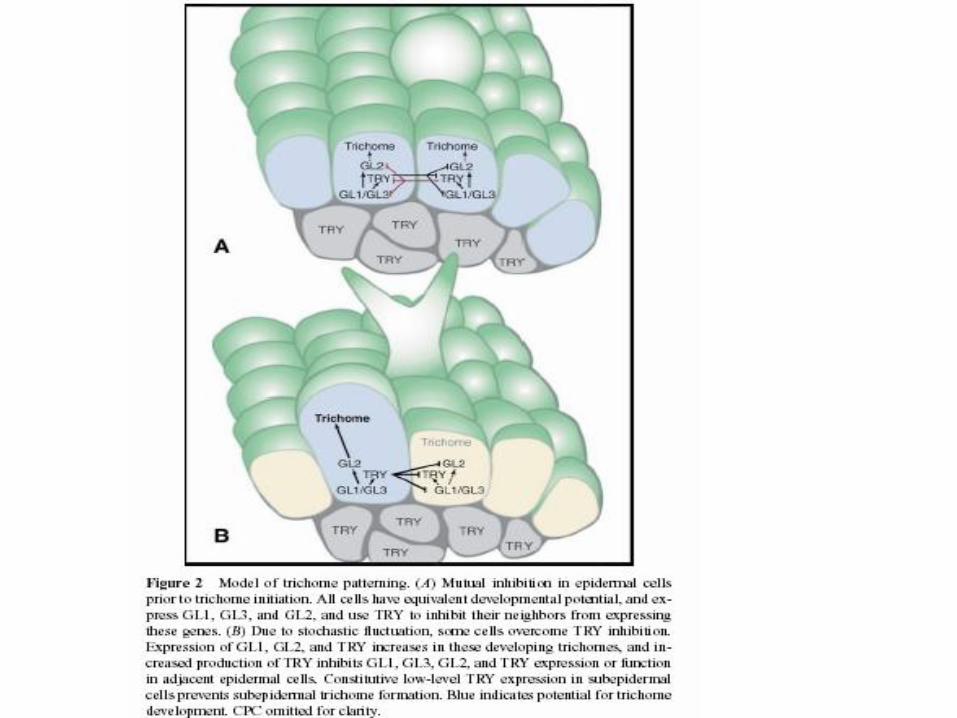
Trichome differentiation
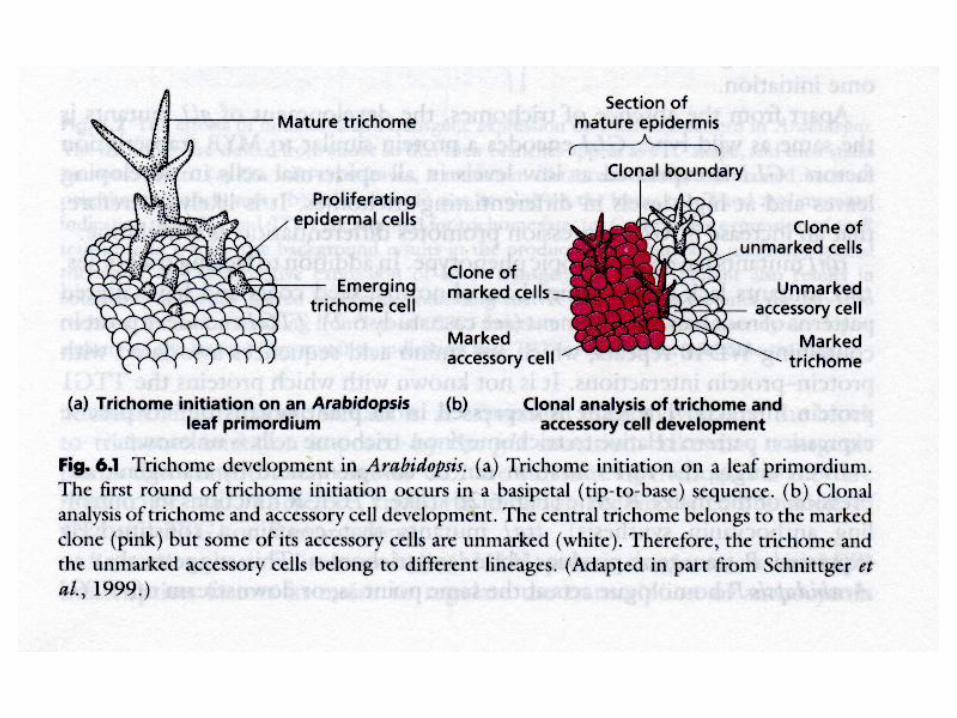
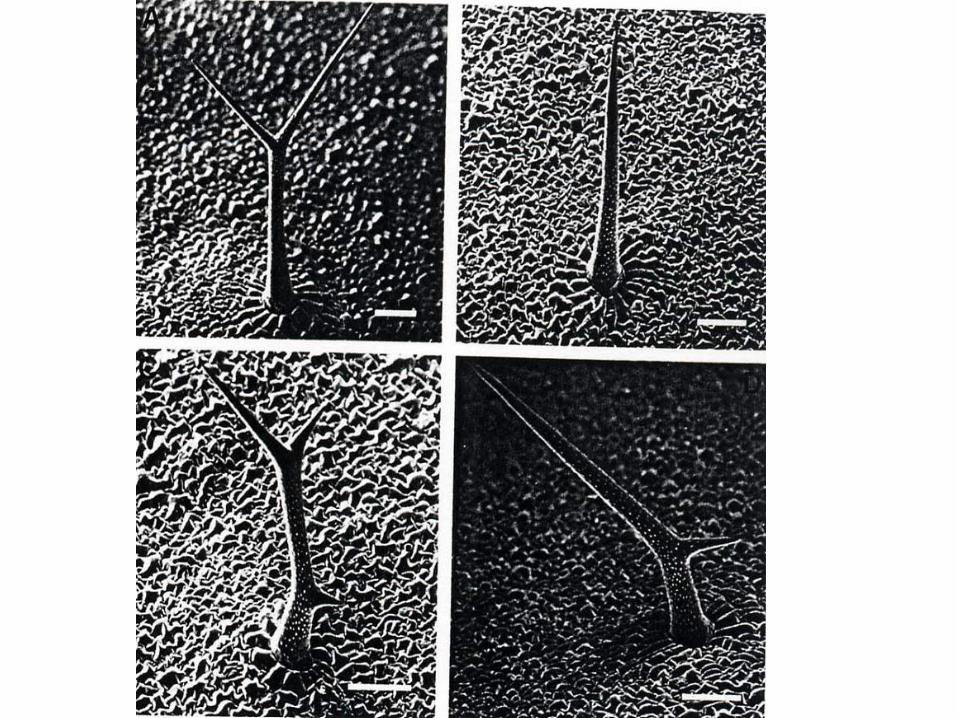
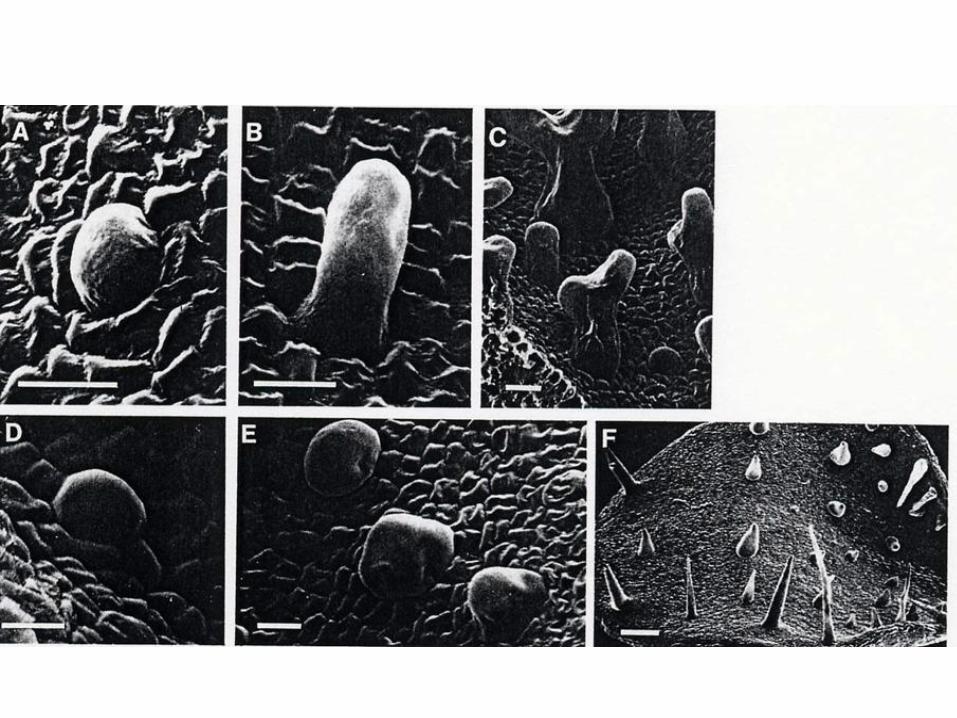
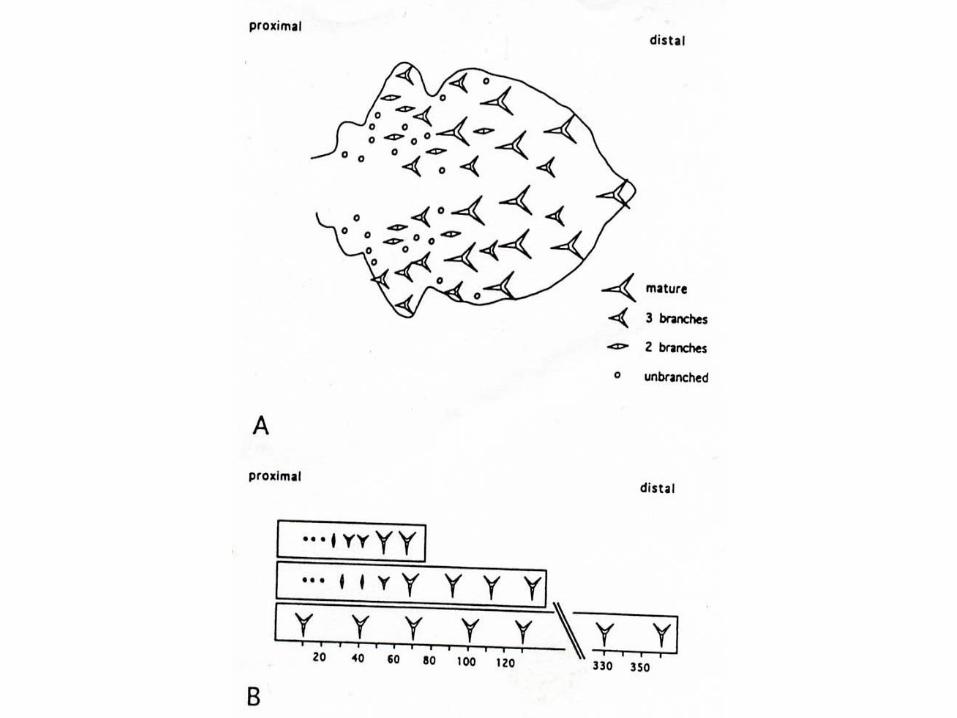
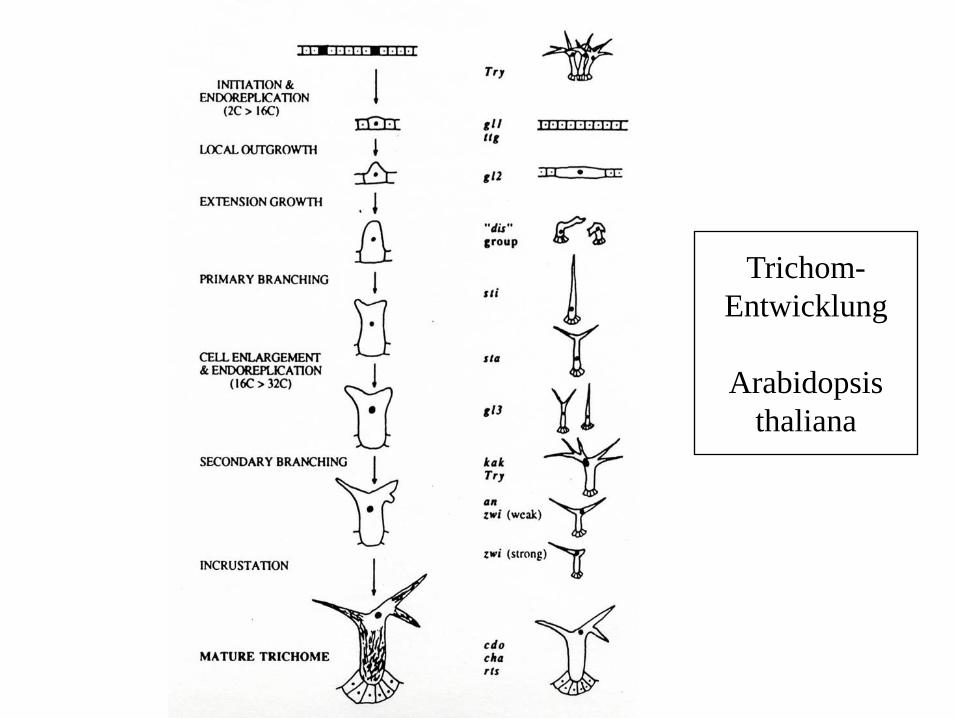
Trichom-
Entwicklung
Arabidopsis
thaliana
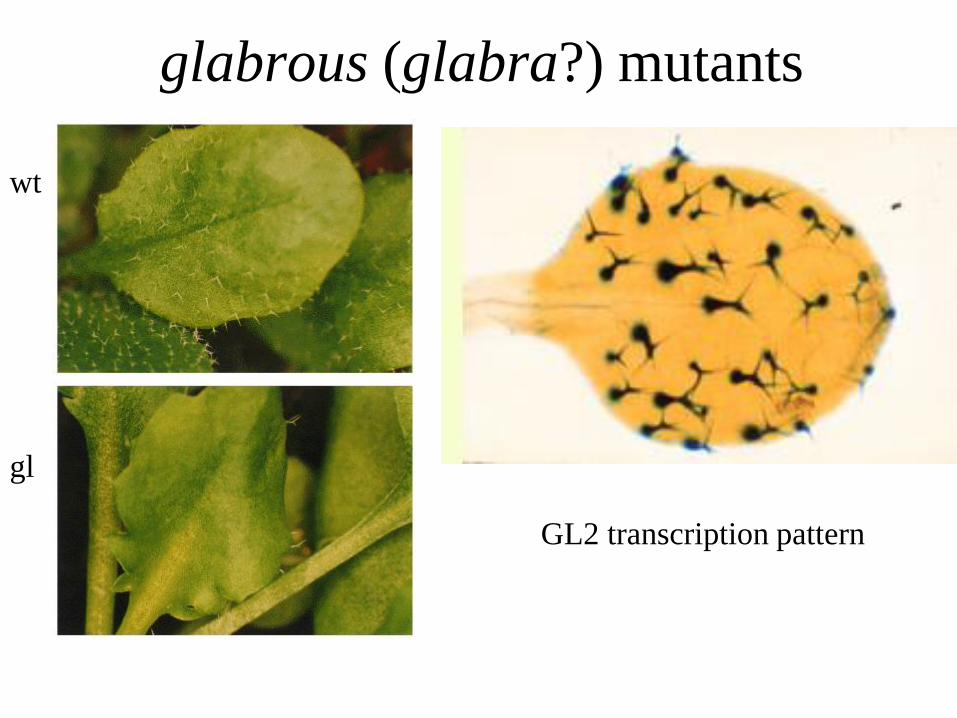
glabrous (glabra?) mutants
GL2 transcription pattern
wt
gl
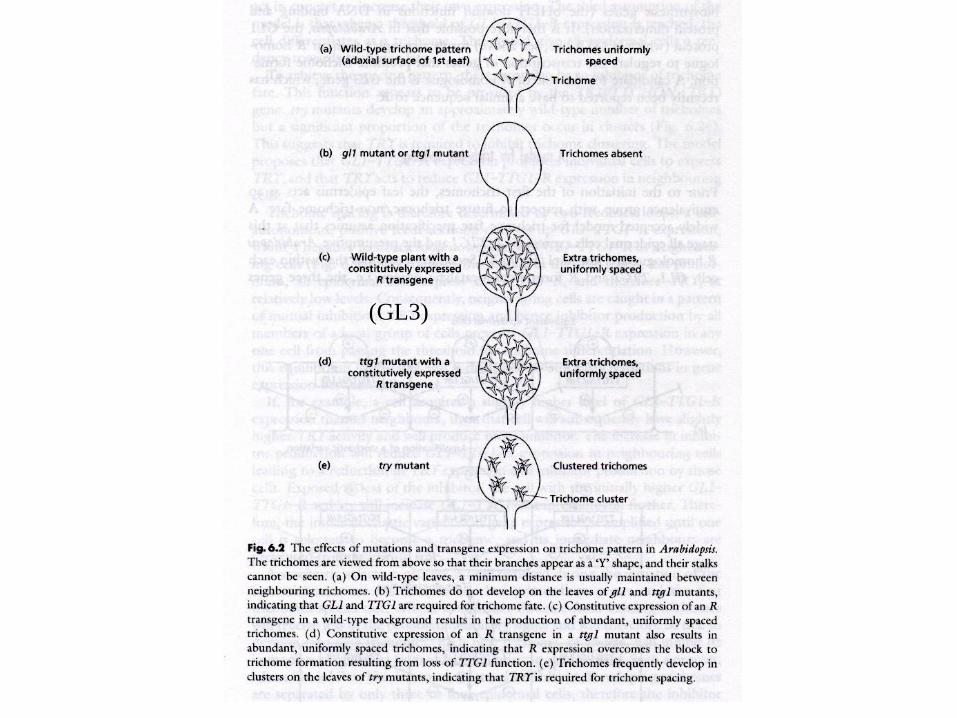
(GL3)

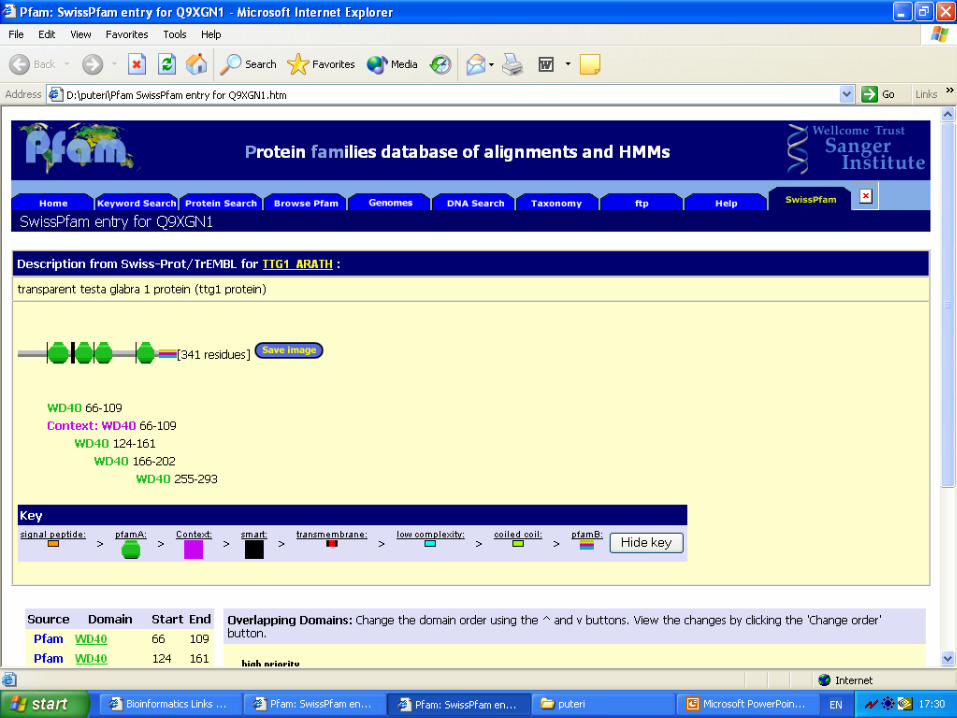
TTG1•The protein is mapped to the
transparent testa glabra1 locus.
•This locus regulates several
developmental and biochemical
pathways in Arabidopsis, including
the formation of hairs on leaves,
stems, and roots, and the production
of seed mucilage and anthocyanin
pigments (purple coloured seedlings).
•This protein is similar to AN11, a
regulator of anthocyanin biosynthesis
in petunia, and is more distantly
related to those of the β-subunits of
heterotrimeric G proteins, which
suggests a role for TTG1 in signal
transduction to downstream
transcription factors.

TTG1
• FUNCTION – it may regulate MYC transcription factors involved in trichome and root hair development, seed mucilage production, and anthocyanin biosynthesis by acting at the dihydorflavonol-4- reductase (DFR) step or it may regulate pathways that involve MYC factors
• SUBCELLULAR LOCATION - cytoplasmic.
• TISSUE SPECIFICITY - Roots, leaves, stems, meristems, flowers and flower buds.
• SIMILARITY - Contains 4 WD40 repeats (Proteins containing WD40 repeats are involved in a number of different types of regulatory roles, such as signalling, cell cycle regulation, transcriptional repression, vesicular trafficking and RNA processing)

Post Translational Modifications
Predicted by PROSITEMDNSAPDSLSRSETAVTYDSPYPLYAMAFSSLRSSSGHRIAVGSFLEDYNNRIDILSFDSDSMTVKPLPNLSFEHPYPPTKLMFSPPSLRRPSSGDLLASSGDFLRLWEINEDSSTVEPISVLNNSKTSEFCAPLTSFDWNDVEPKRLGTCSIDTTCTIWDIEKSVVETQLIAHDKEVHDIAWGEARVFASVSADGSVRIFDLRDKEHSTIIYESPQPDTPLLRLAWNKQDLRYMATILMDSNKVVILDIRSPTMPVAELERHQASVNAIAWAPQSCKHICSGGDDTQALIWELPTVAGPNGIDPMSVYSAGSEINQLQWSSSQPDWIGIAFANKMQLLRV
• N-glycosylation sites
• tyrosine sulphate site
• cAMP- and cGMP- dependent protein kinase phosphorylation sites
• protein kinase C phosphorylation sites
• casein kinase II phosphorylation sites
• N-myristoylation sites in bold
• WD repeats region is underlined
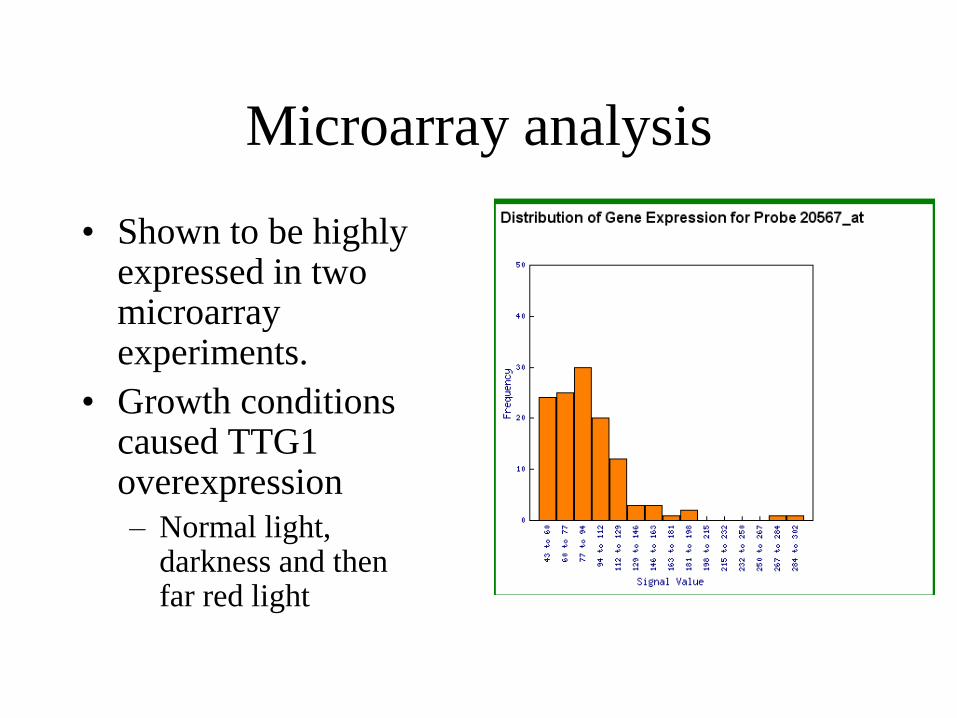
Microarray analysis
• Shown to be highly expressed in two microarray experiments.
• Growth conditions caused TTG1 overexpression
– Normal light, darkness and then far red light

Summary
• Arabidopsis thaliana transparent testa glabra 1 protein (TTG1)
• Encodes a protein of 341 amino acids localised to chromosome 5p and consists of 3 exons
• TTG1 protein does not act directly as a transcription factor but binds to other proteins to promote the initiation of trichomes in leaves and stems, it is also involved in regulating the transcription factors involved in seed mucilage production and anthocyanin biosynthesis.

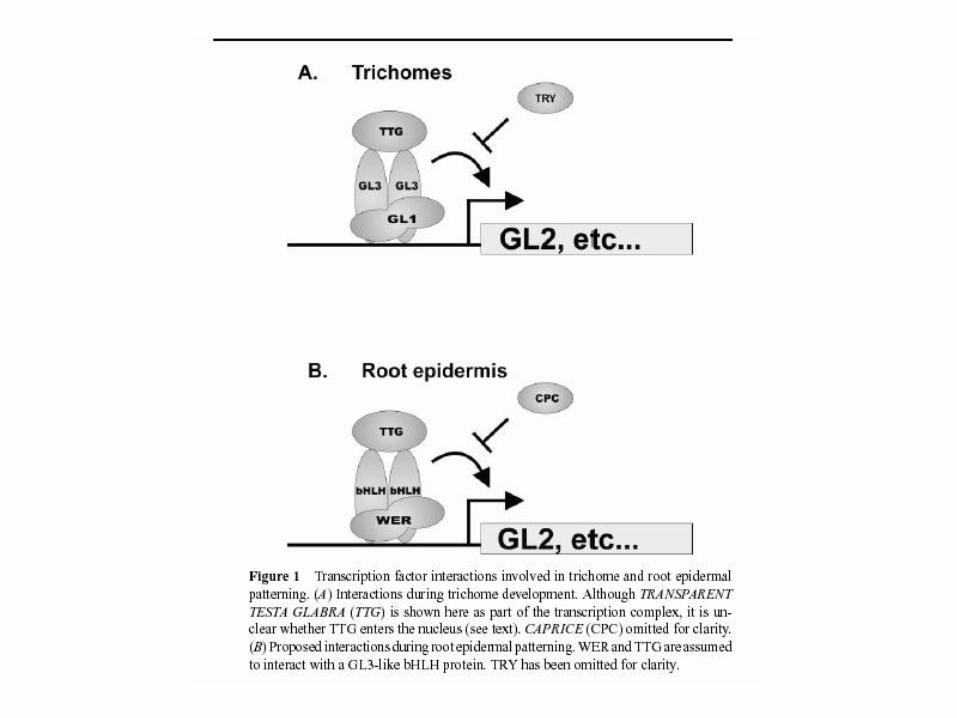
GLABRA1
• transcription factor – MYB related
• bHLH, MYB interactor
GL3 … and maize R

Triptychon (TRY)
• MYB-related
• inhibits trichome
differentiation
• mutant - clusters

Alan TURING
1912-1954
1951-2 - Reakčně-difusní model



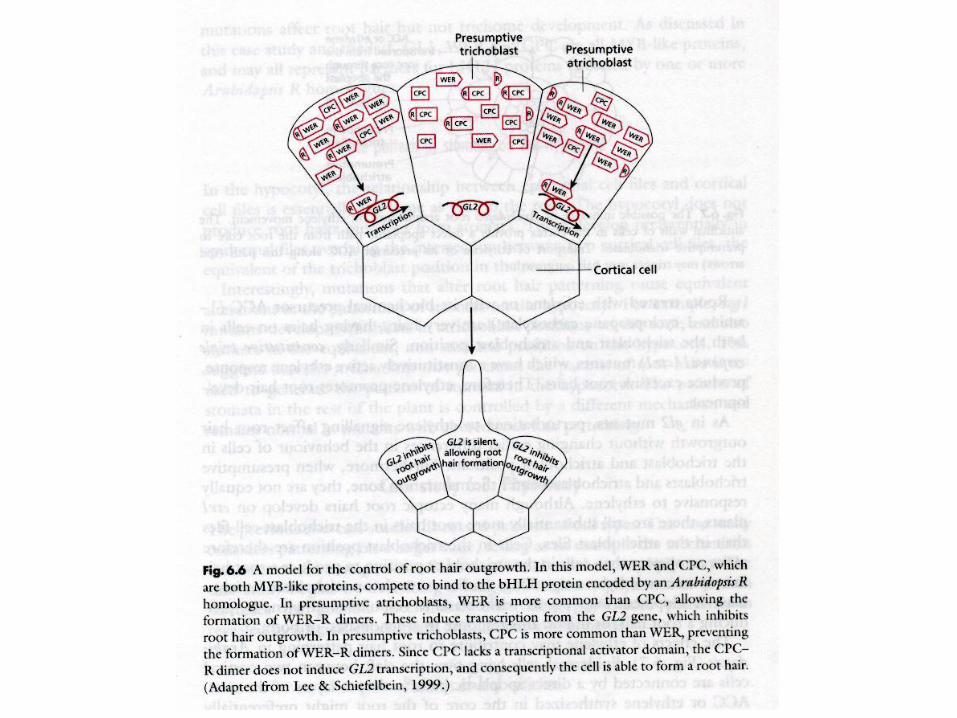
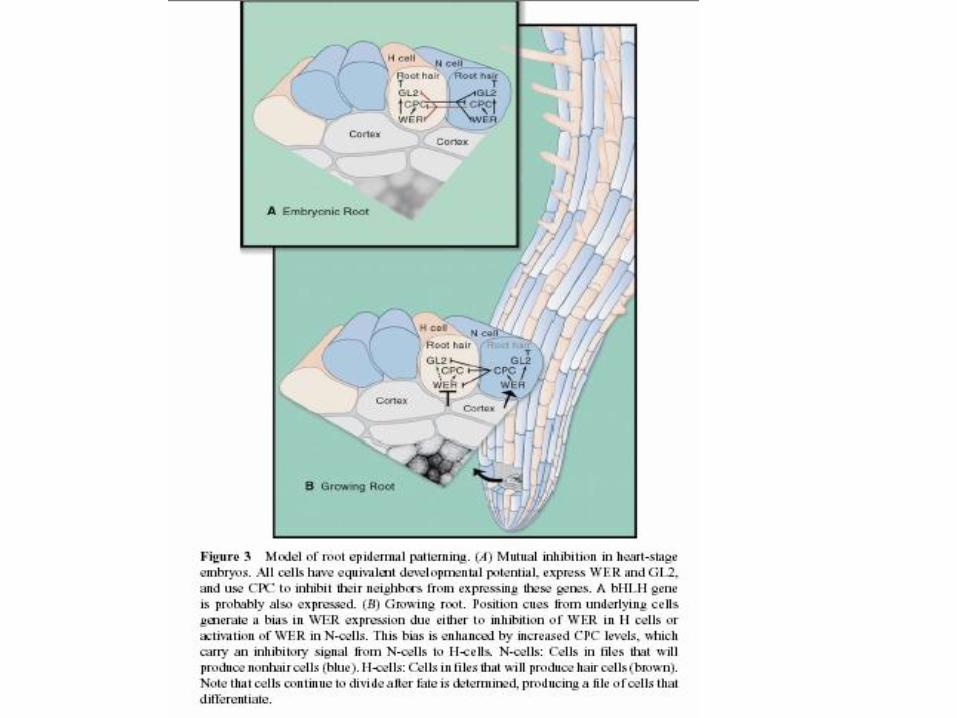
Root hair differentiation
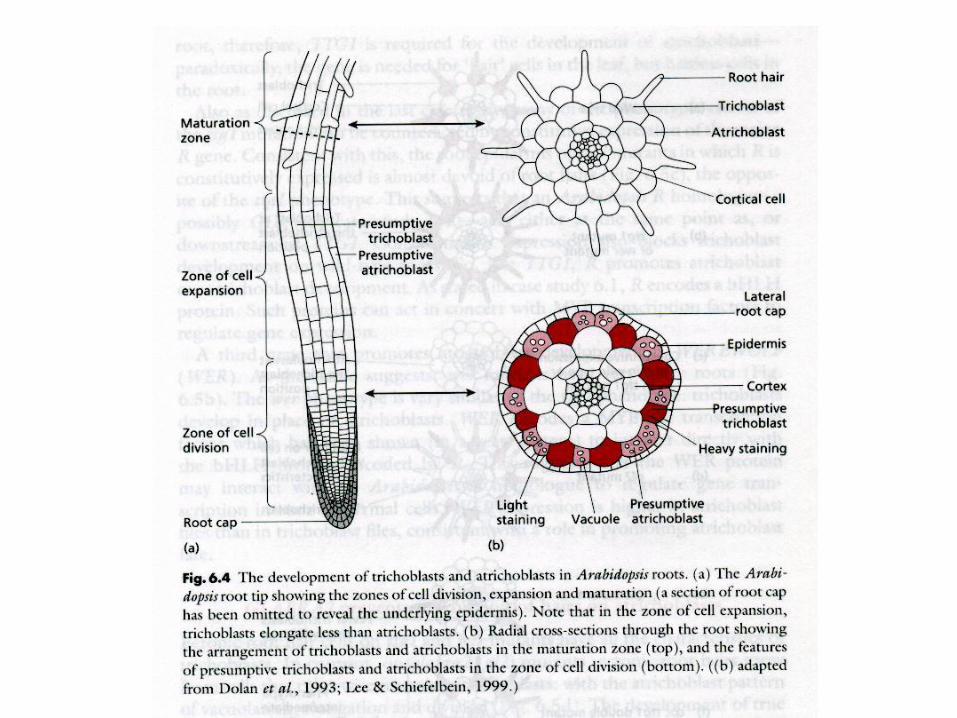



GL2 brání růstu vlášení na pozici
atrichoblastů
GL2 = homeodomain txn factor

GL2 exprese
• Photograph of wild-
type (left), ttg mutant
(center), and wer
mutant (right) roots
bearing the
GL2::GUS
transgene. Note that
ttg and wer
dramatically reduce
GL2 gene
expression.
(Schiefelbein lab)
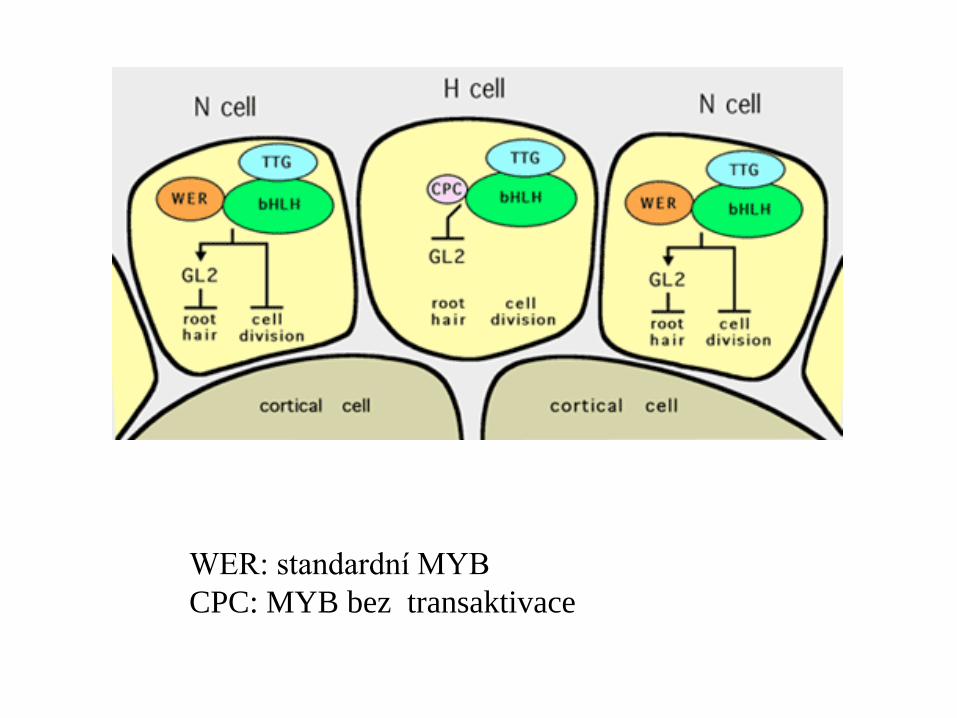
WER: standardní MYB
CPC: MYB bez transaktivace

CPC se ovšem exprimuje
přednostně v atrichoblastech
působí buněčně neautonomně –
transportuje se jako „aktivátor“ trichoblastů
do sousedních buněk trichoblastů
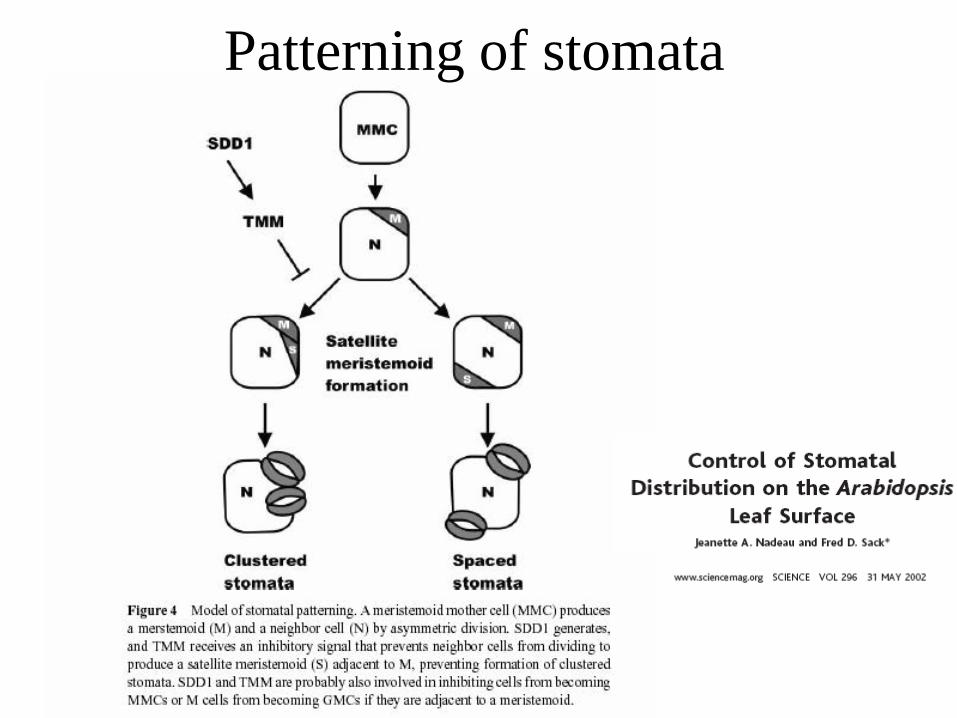
Patterning of stomata

Vasculature patterning
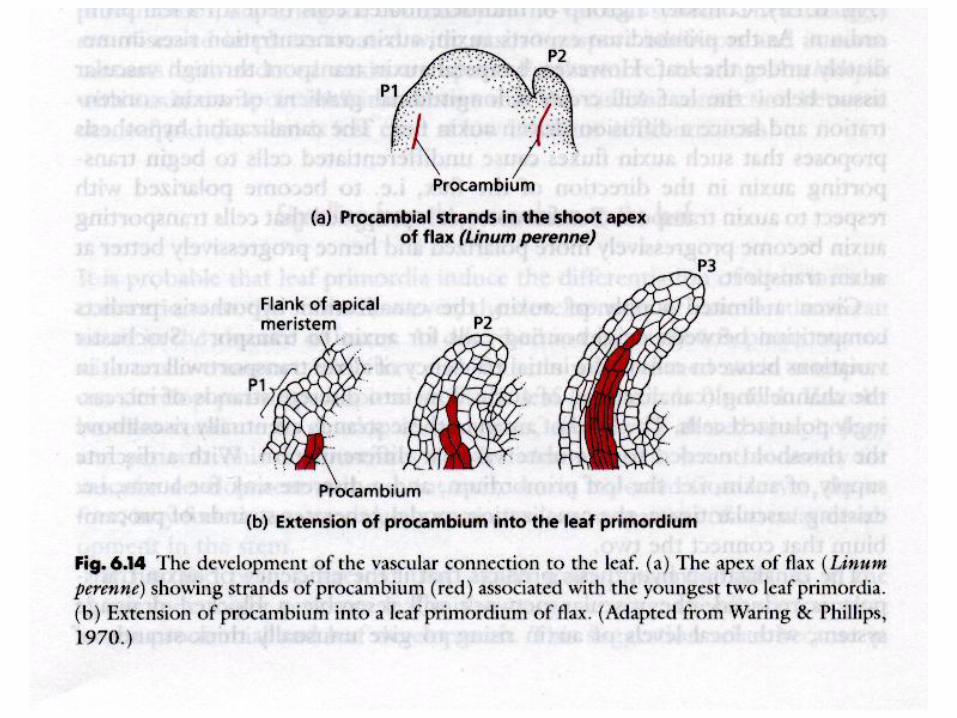
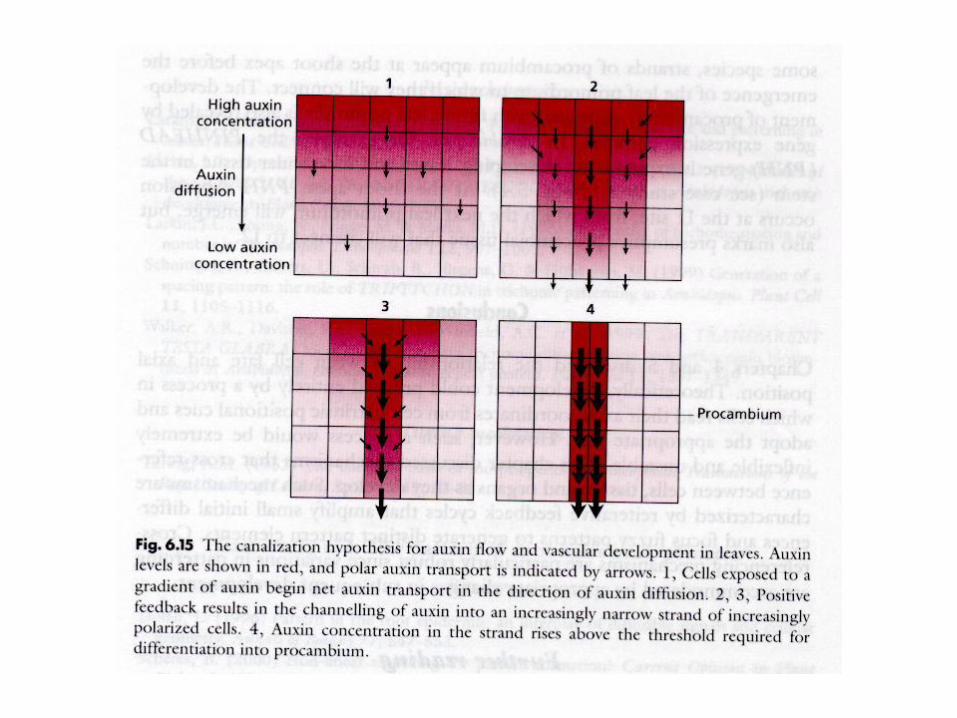

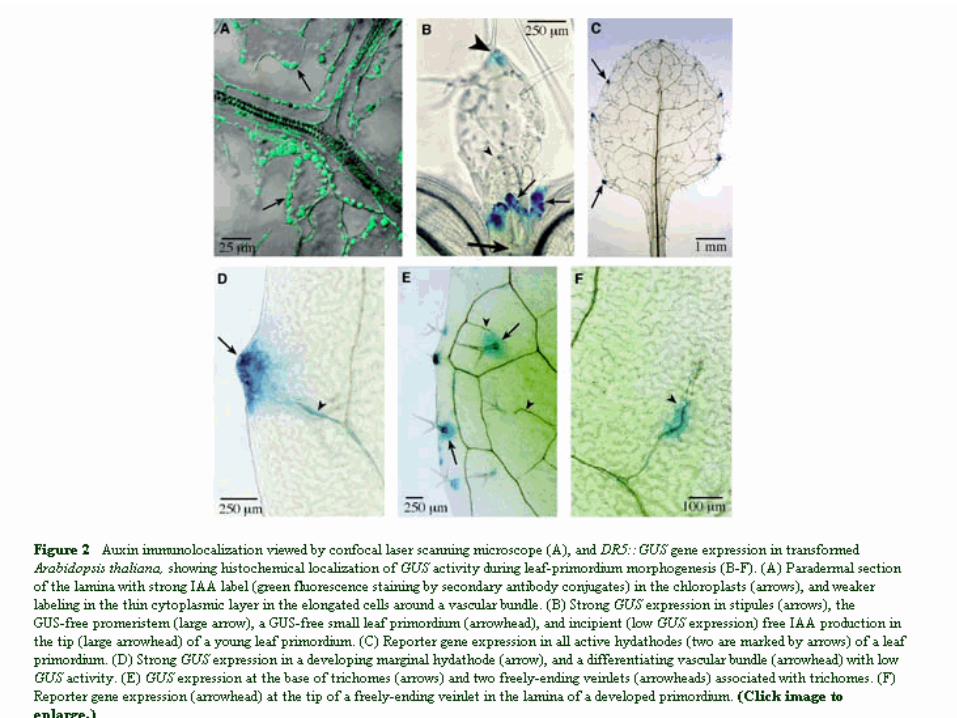


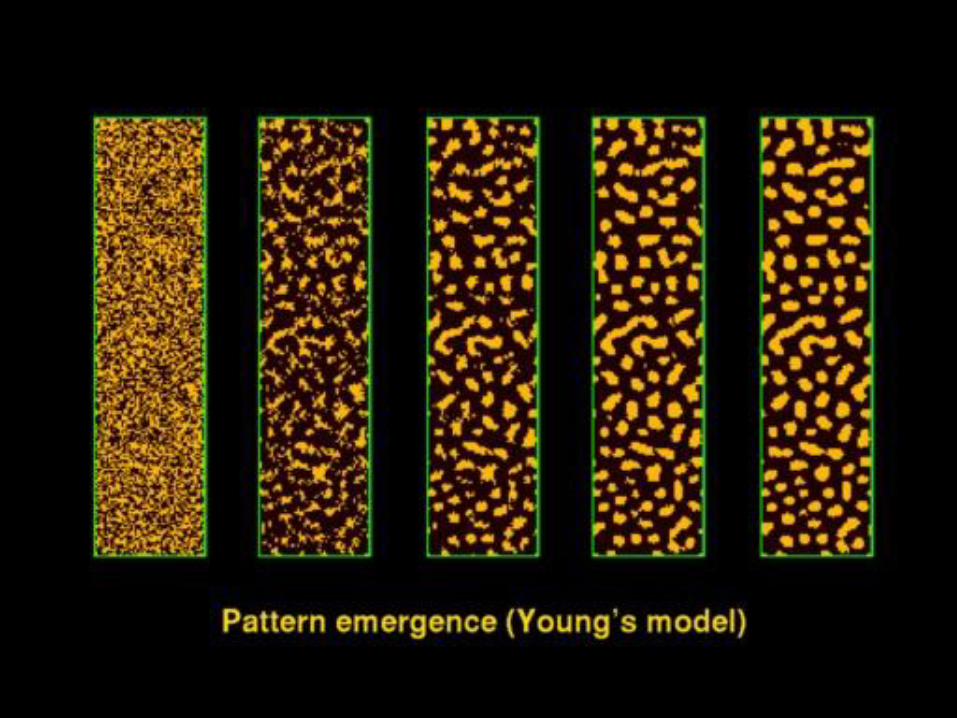
reakčně-difusní model
(http://texturegarden.com/)

Phyllotaxis
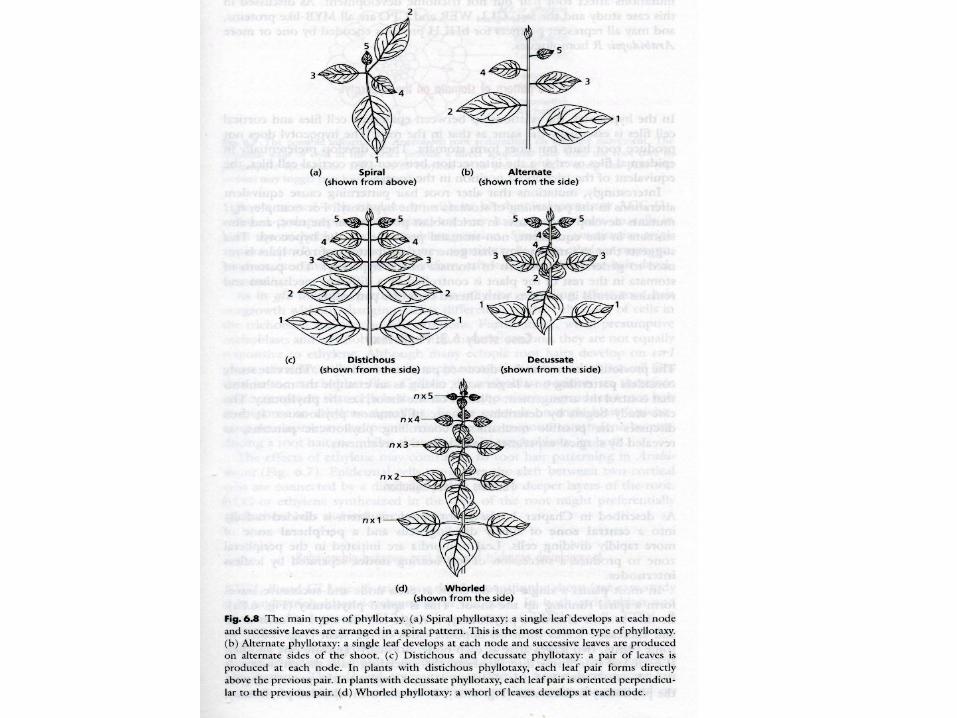
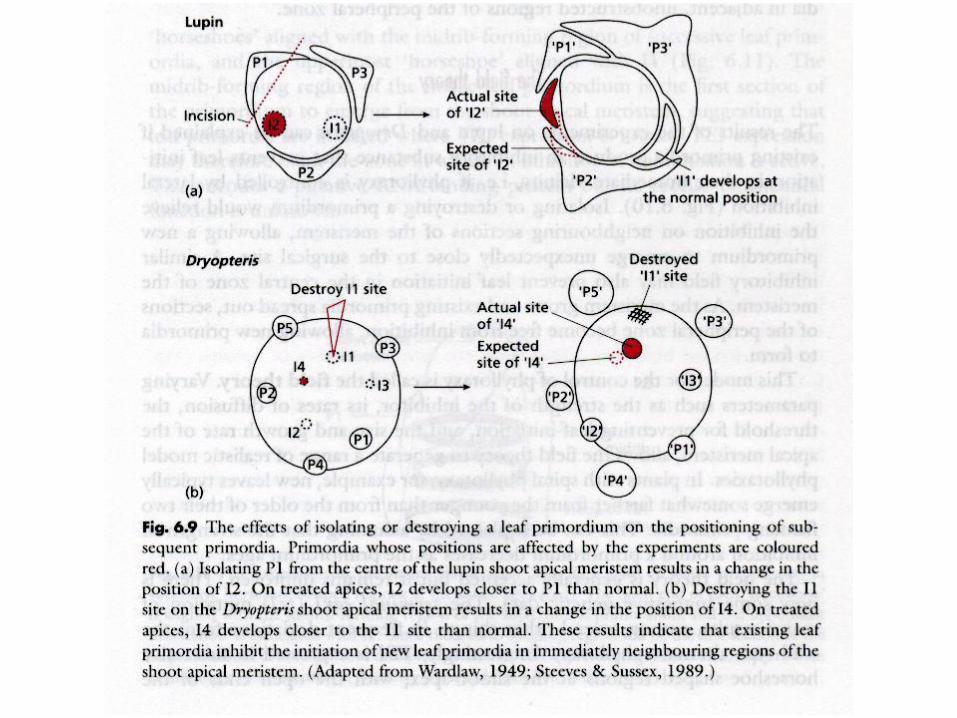
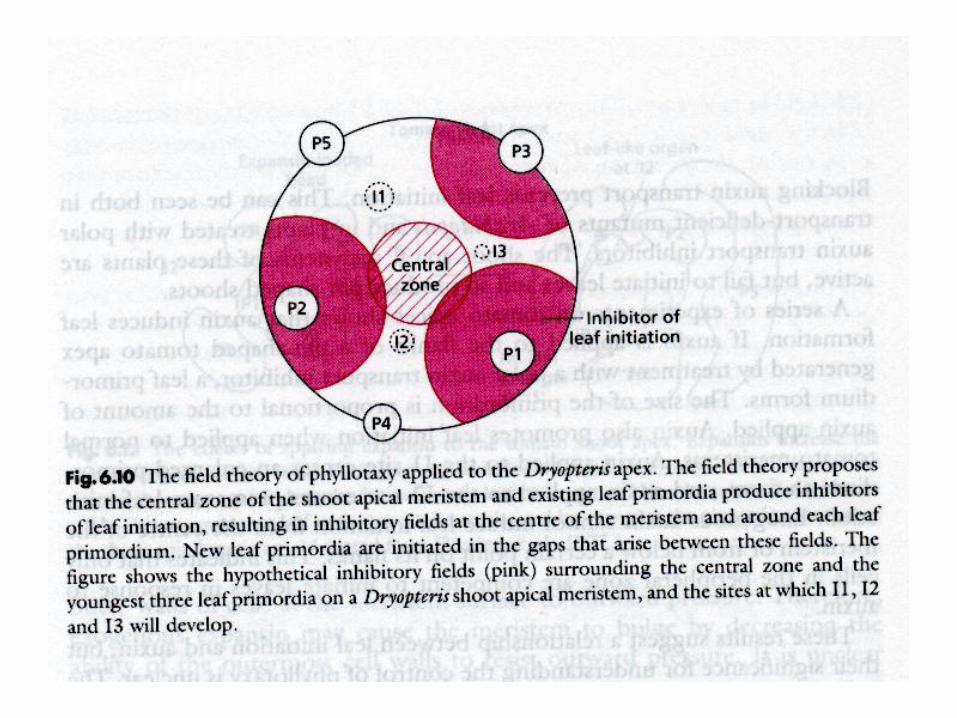


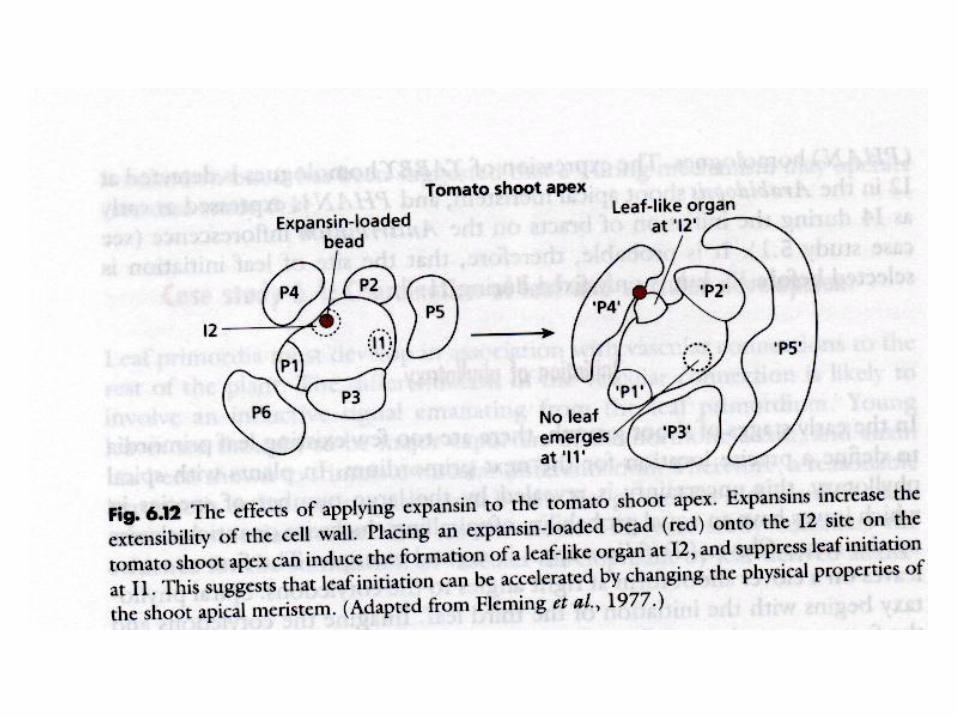

Alan TURING
1912-1954
1951? – nepublikovaná teorie
fylotaxe

1941 discusses daisy patterns & fir cones with
Joan Clarke
1947-1948 attends Cambridge undergraduate
physiology lectures
Feb 1951 Manchester Mark I machine installed
Nov 1951 paper on the ‘simple case’ - ‘Chemical basis
of morphogenesis’
1952 ‘Had quite a jolly time lecturing on fir
cones’
1952-1954 Drafts part of the Morphogen Theory of
Phyllotaxis, finally published in 1992.
Jonathan Swinton 2004. See http://www.swintons.net/jonathan/Turing/

Morphogenesis: Collected Works of AM Turing, Volume 3,
ed PT Saunders, North-Holland, 1992. This contains:
•An introduction by Saunders
•A reprint of The chemical basis of morphogenesis
•A diffusion-reaction theory of morphogenesis in plants
•The morphogen theory of phyllotaxis I
•The morphogen theory of phyllotaxis II
•Outline of the development of the daisy

‘my mathematical theory of embryology...is yielding to treatment, and it will so far as I can see, give satisfactory explanations of
(i) gastrulation
(ii) polygonally symmetrical structures, e.g. starfish, flowers
(iii) leaf arrangements, in particular the way the Fibonacci series (0,1,1,2,3,5,8,13,...) comes to be involved
(iv) colour patterns on some animals, e.g. stripes, spots and dappling
(v) pattern on nearly spherical structures such as some Radiolara...’
A. Turing, 8th Feb 1951

Fylotaxe a Fibonacciho posloupnosti
Turing archive Church, 1904

Dějiny fylotaxe - počátky
• 370-285 BC Theophrastus – pravidelné uspořádání listů
• 1202 Leonardo Fibonacci – Fibonacciho posloupnost <1, 1, 2, 3, 5, ... F(k), F(k+1)> (mimo souvislost s fylotaxí)
• 1754 Charles Bonnet – popis fylotaktic. spirál v moderním smyslu
• 1830-35 Schimper a Braun – formalizace popisu; od té doby pokusy o matematické uchopení
• 1882 Julius Sachs odmítá matematic. modely
• ... ale modelování pokračuje ... i pokusy

D´Arcy Wenworth
Thompson
1860-1948
1917 – On Growth and Form

Stems, cones and flowers
8 green
parastichies
13 red
parastichies
Church, 1904
This presentation is copyright Jonathan Swinton 2004. See http://www.swintons.net/jonathan/Turing/TuringCopyright.html for details and copyright owners of the images contained.

Experiment Douadyho a Coudera
(1992)
Drops of ferrofluids are deposited periodically at
the center of a circular dish filled with silicon oil.
Under the effect of magnets surrounding the dish,
the drops repel one another and are attracted to the
edge of the dish. To Douady and Couder’s own
amazement, when the magnetic field was slowly
reduced during the process, the drops self-organized
into Fibonacci phyllotactic patterns.

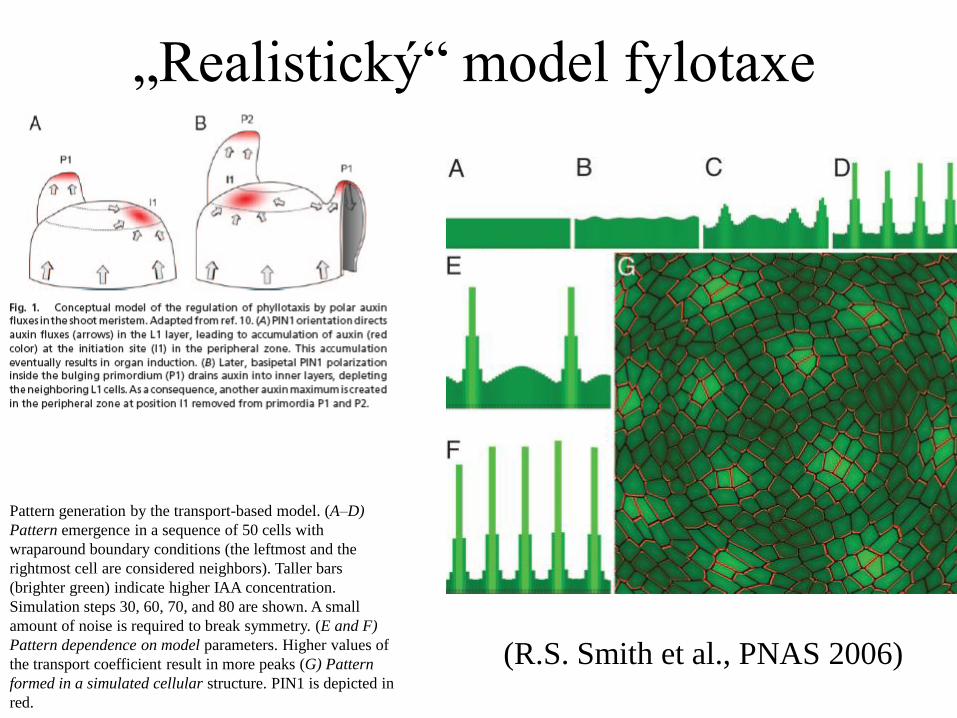
„Realistický“ model fylotaxe
Pattern generation by the transport-based model. (A–D)
Pattern emergence in a sequence of 50 cells with
wraparound boundary conditions (the leftmost and the
rightmost cell are considered neighbors). Taller bars
(brighter green) indicate higher IAA concentration.
Simulation steps 30, 60, 70, and 80 are shown. A small
amount of noise is required to break symmetry. (E and F)
Pattern dependence on model parameters. Higher values of
the transport coefficient result in more peaks (G) Pattern
formed in a simulated cellular structure. PIN1 is depicted in
red.
(R.S. Smith et al., PNAS 2006)

Shrnutí
• Specifikace typů buněk (pletiv, orgánů) v rámci rostlinného organismu je dána neustále probíhající vzájemnou signalizací-ovlivňováním vyvíjejících/diferencujících se buněk, pletiv a orgánů, které si tak vzájemně poskytují poziční informaci.
• Takové procesy jsou charakterizovány opakovanými zpětnovazebnými cykly, které amplifikují malé počáteční rozdíly. Vzájemná komunikace umožňuje pozdější úpravu počátečních odchylek od pravidelného uspořádání.
• Typickým jevem je repetitivní pravidelnost, kterou lze modelovat

Soběpodobnost a algoritmická
povaha rostlinného těla
• aneb k čemu jsou vizuální modely

J.W. Goethe o algoritmické povaze
rostliny
Vorwärts und rückwärts ist die Pflanze immer
nur Blatt.17.5.1787
Es mag nun die Pflanze sprossen, blühen oder
Früchte bringen, so sind es doch nur immer
dieselbiger Organe, welche in vielfältigen
Bestimmungen und unter oft veränderten
Gestalten die Vorschrift der Natur erfüllen.1790

Aristid
Lindenmayer
1925-1989
1968 - Mathematical models for
cellular interaction in development.

http://algorithmicbotany.org/
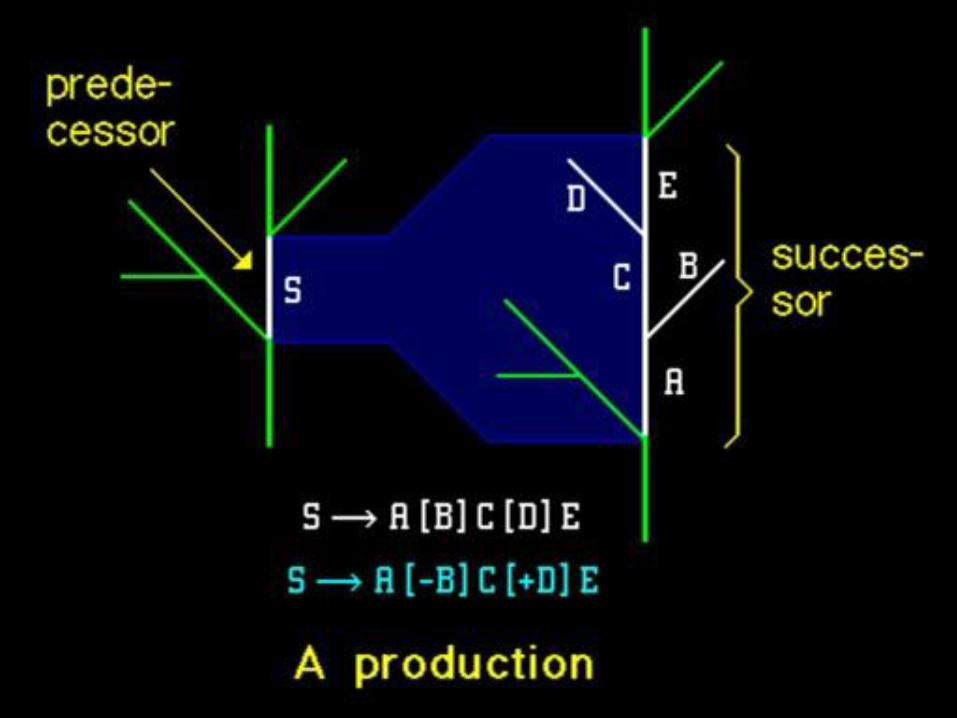
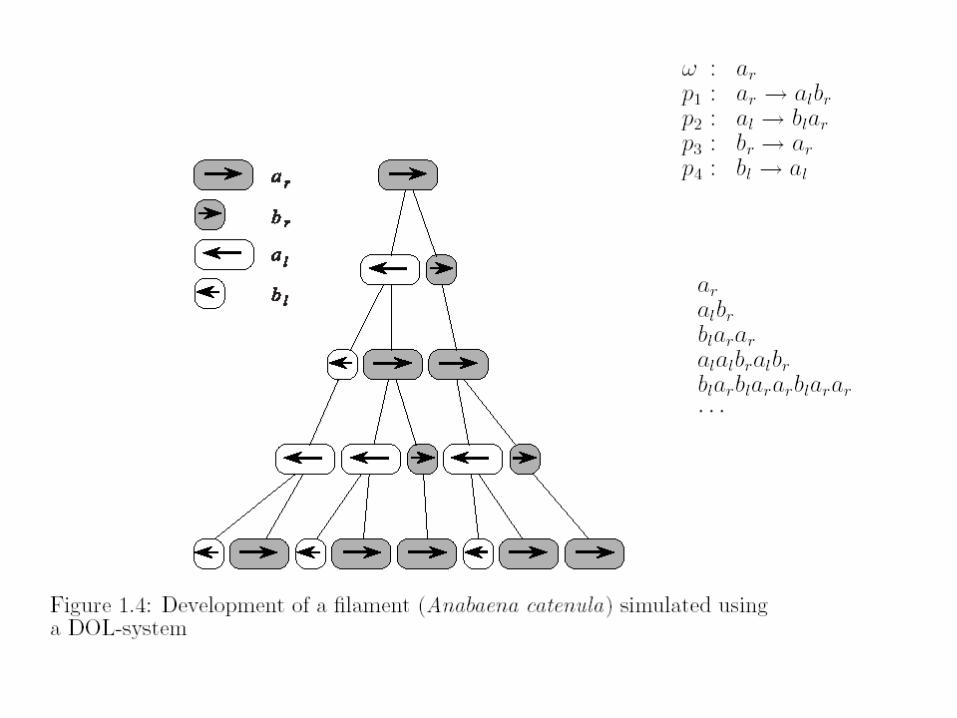
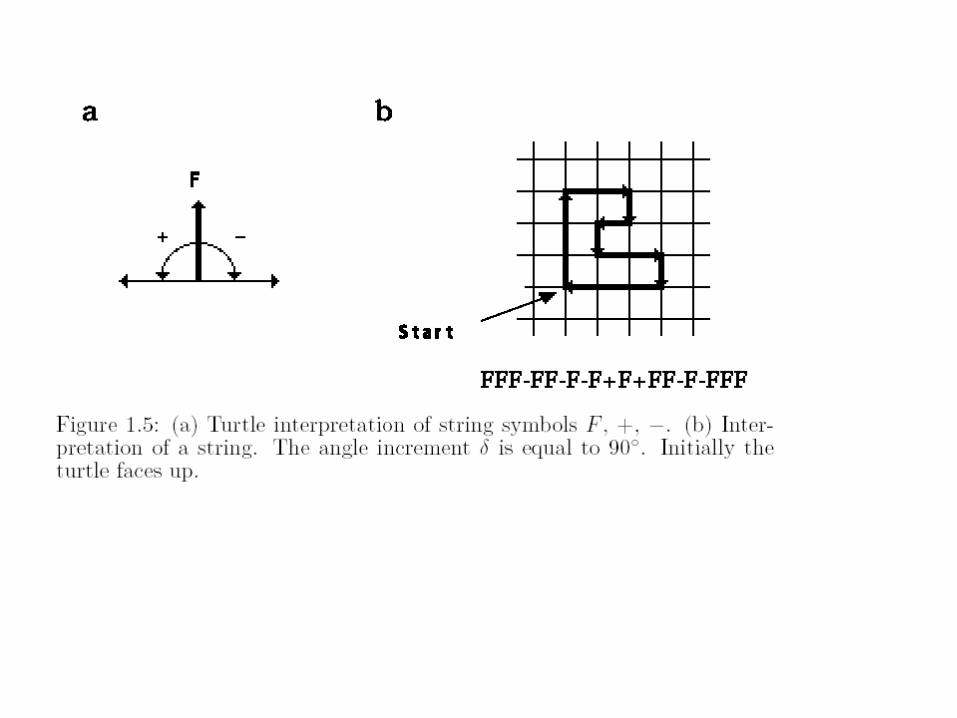
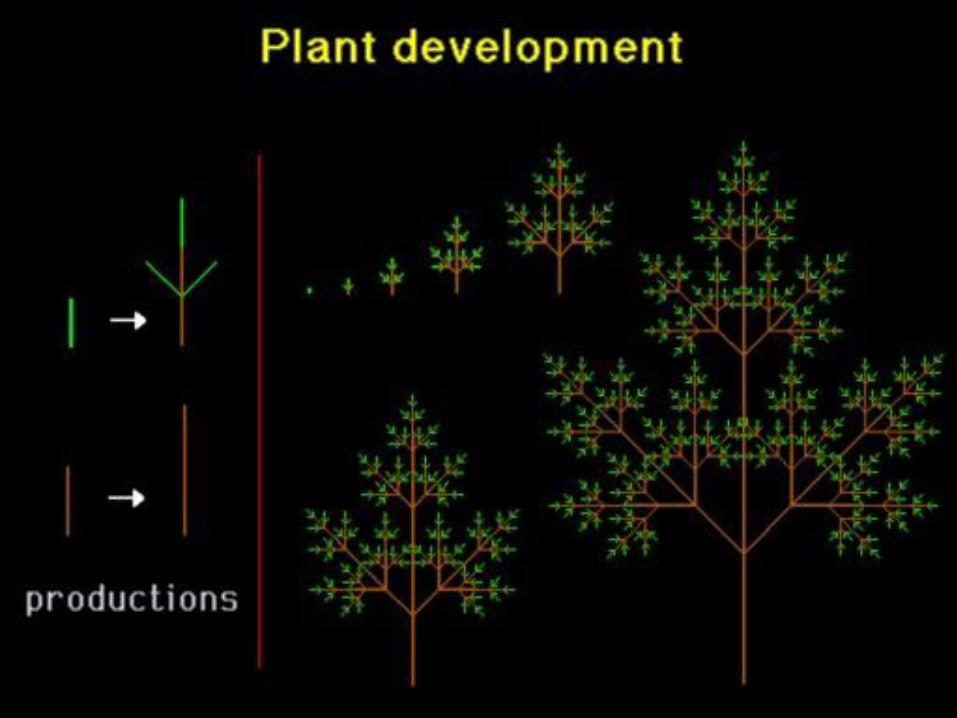


„Náhodný“ pseudorealismus ...

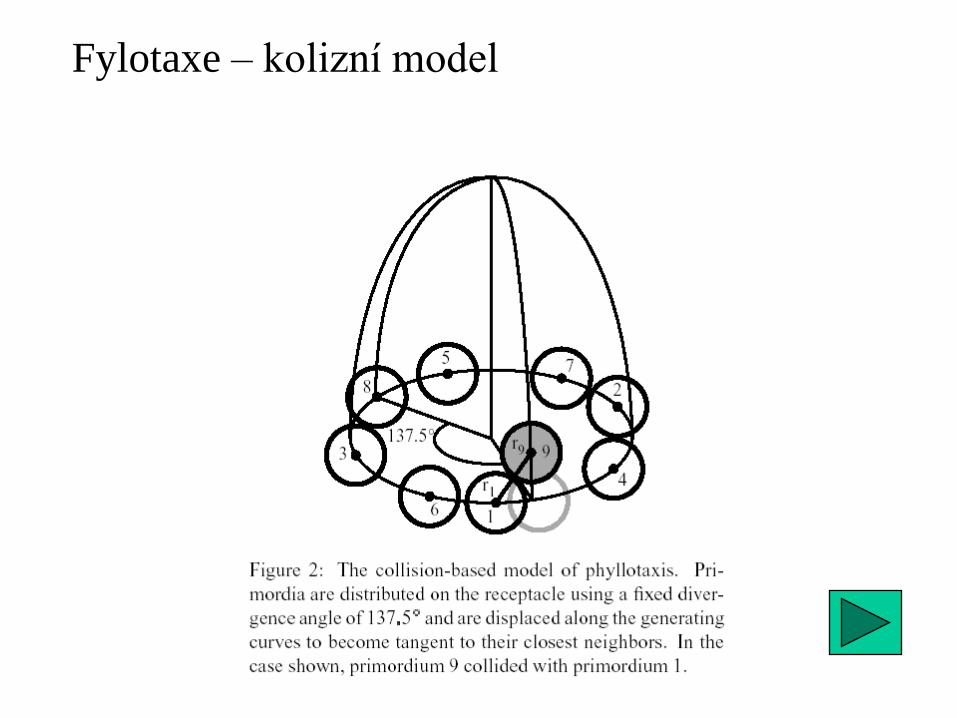
Ještě jednou fylotaxe ...
(pravidelnost není na škodu)




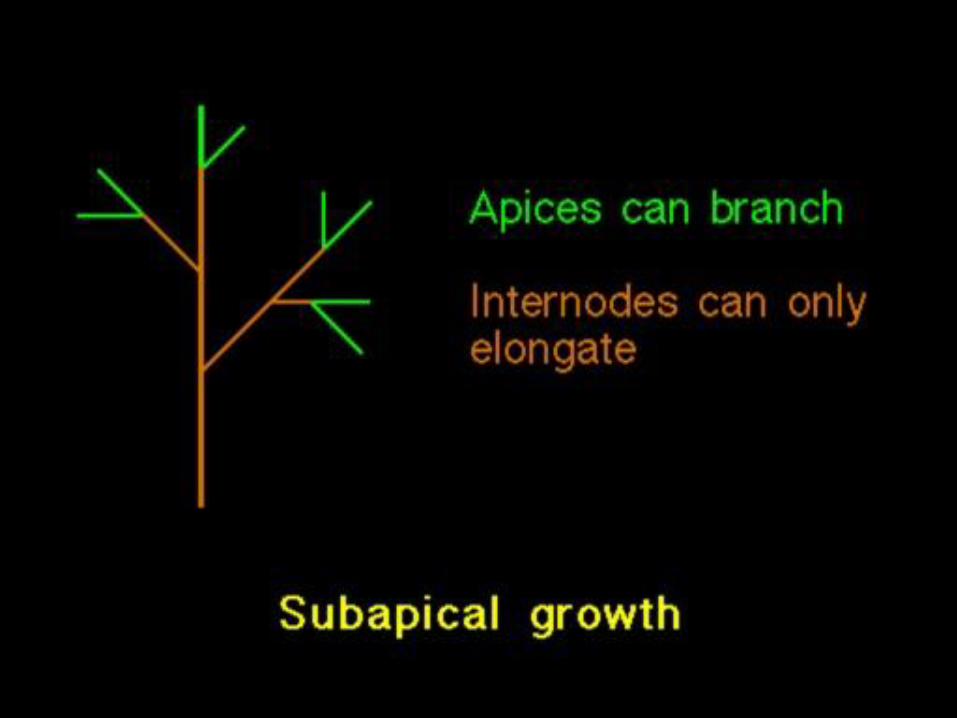
Architektura bylin
(jak zapracovat signály)
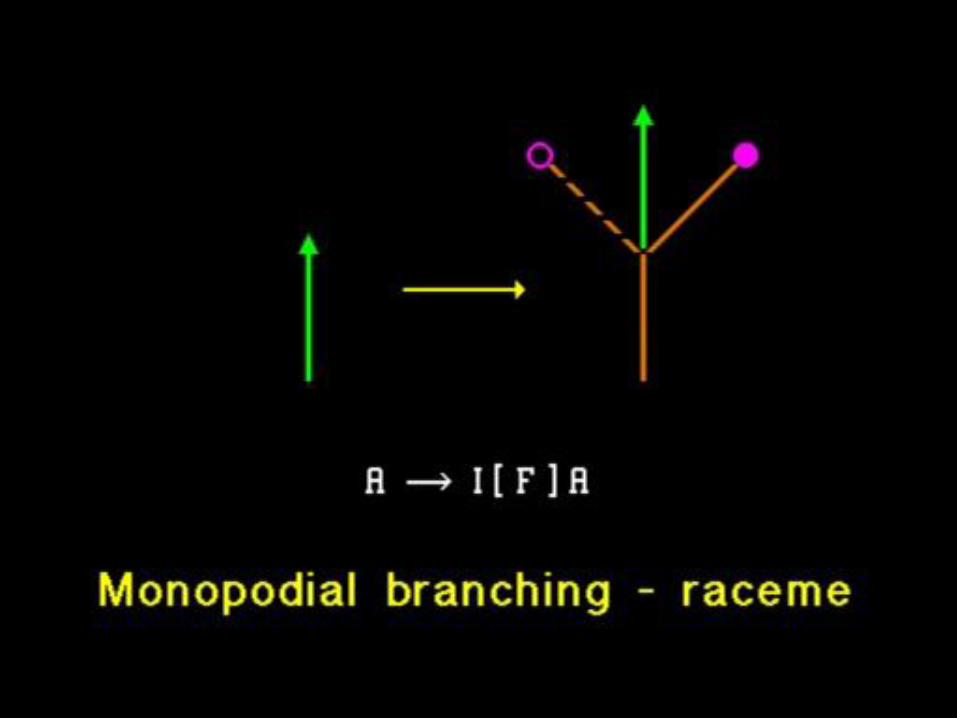

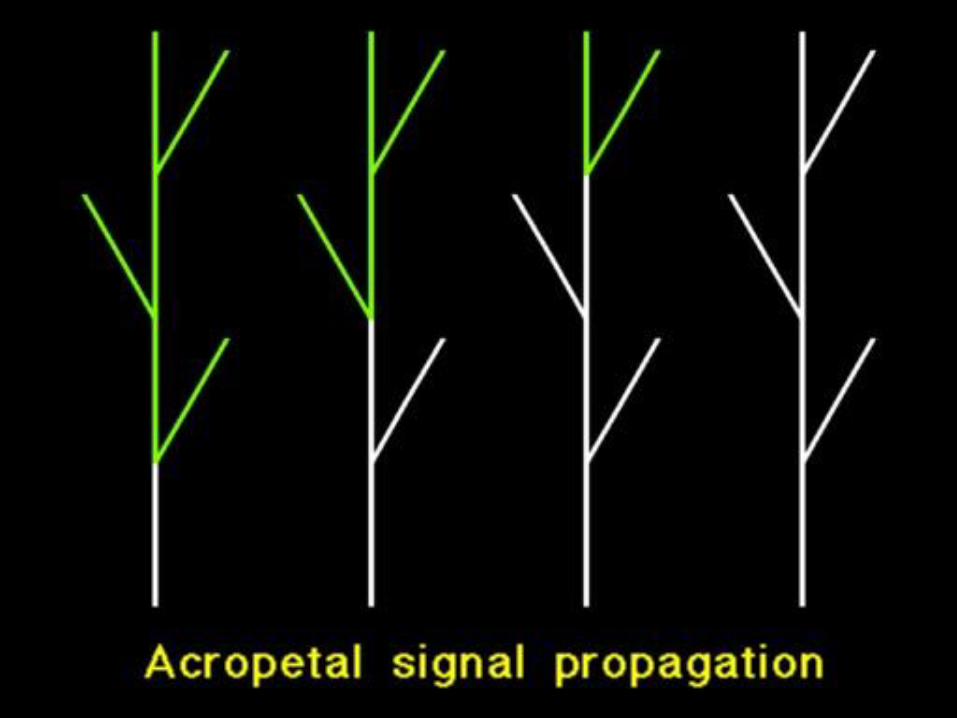
Šíření signálů rostlinou
• Akropetální kvetení
• Basipetální kvetení
• Architektura Mycelis
muralis – schématická
a „realistická“

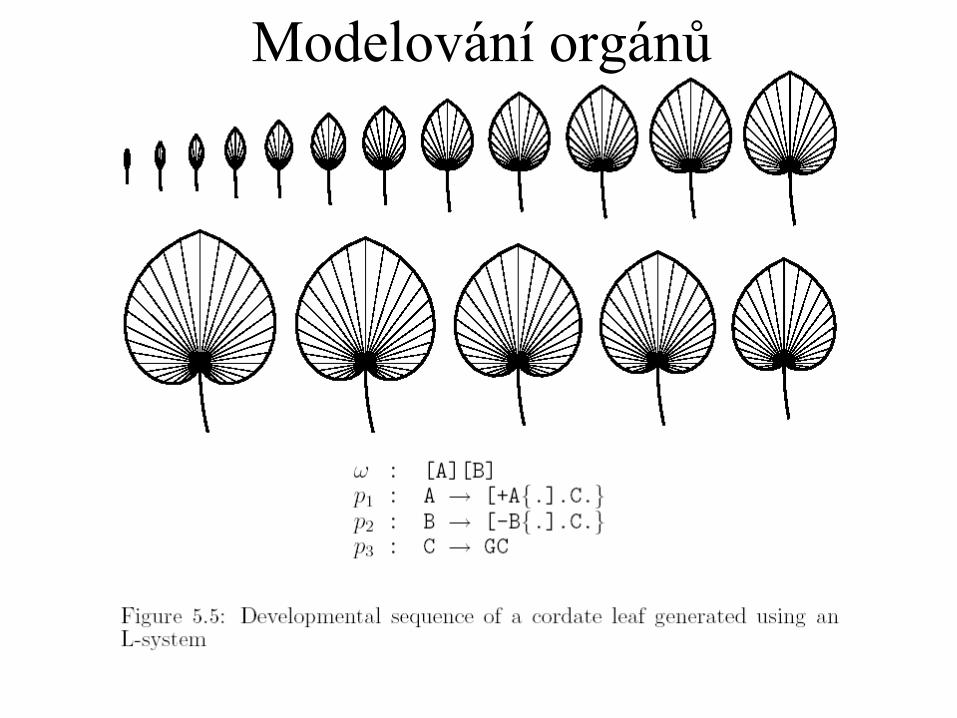
Modelování orgánů
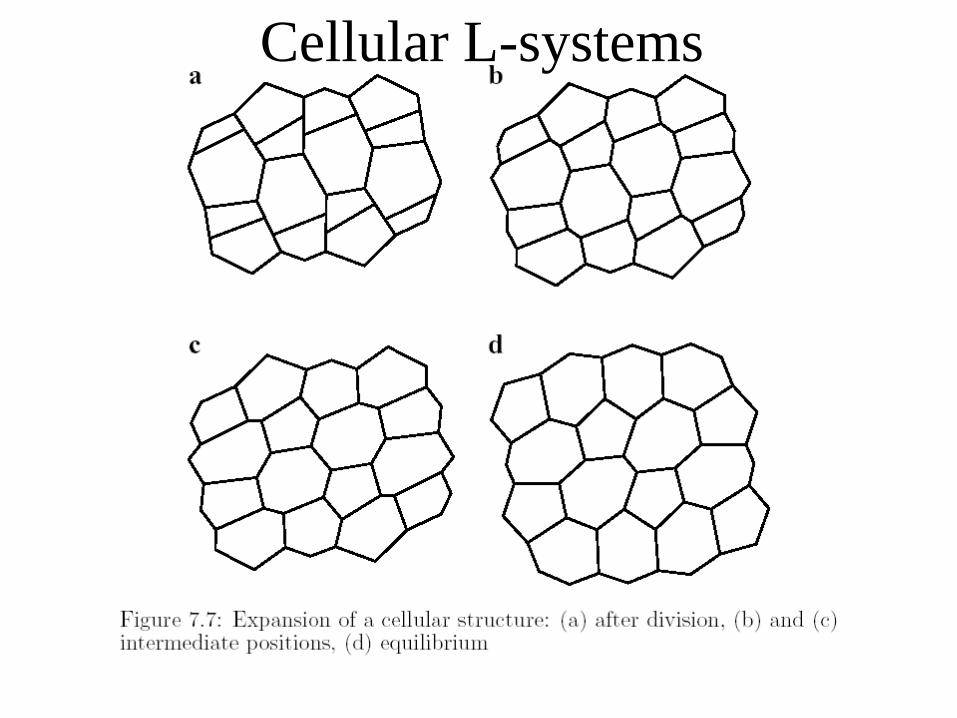
Cellular L-systems


Přidejme spojitý čas ...
• Složený list
• Apex – Ailanthus
• Kvetení – zvonek
Rostliny v prostředí
(cestou k ekosystému)
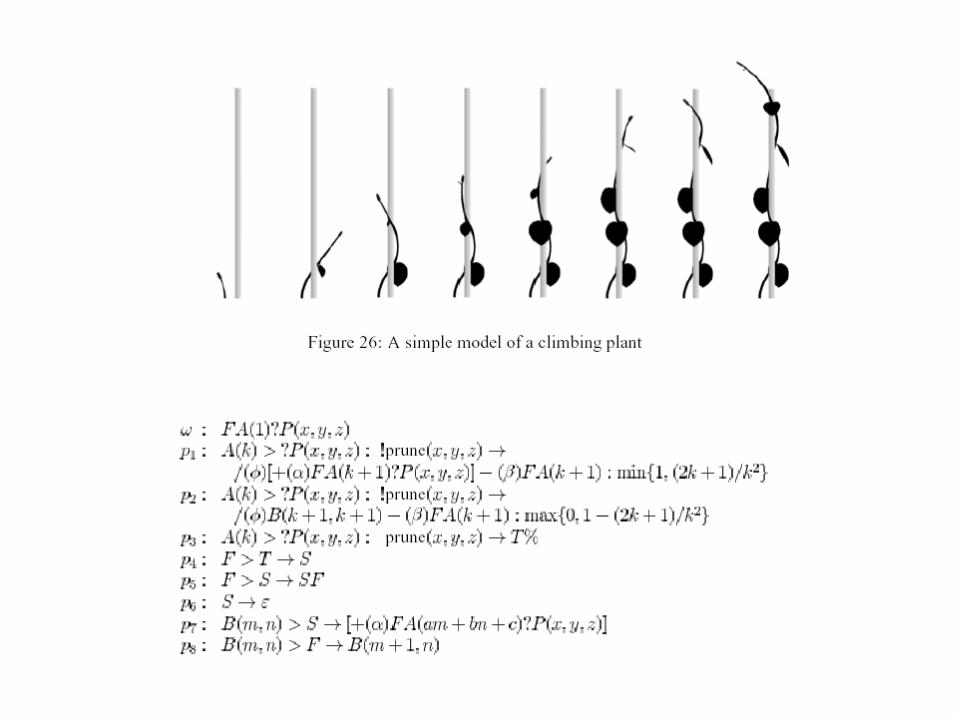
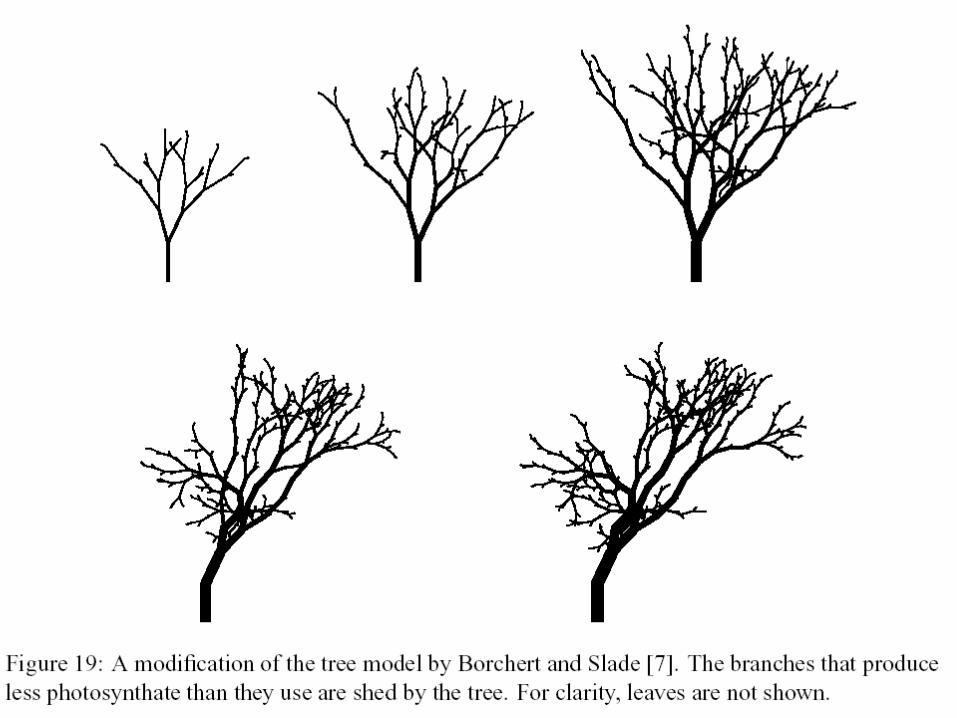

Interakce s prostředím
• Klonální jetel a světlo
• Kompetice větví o
světlo
• Kořen a voda v půdě
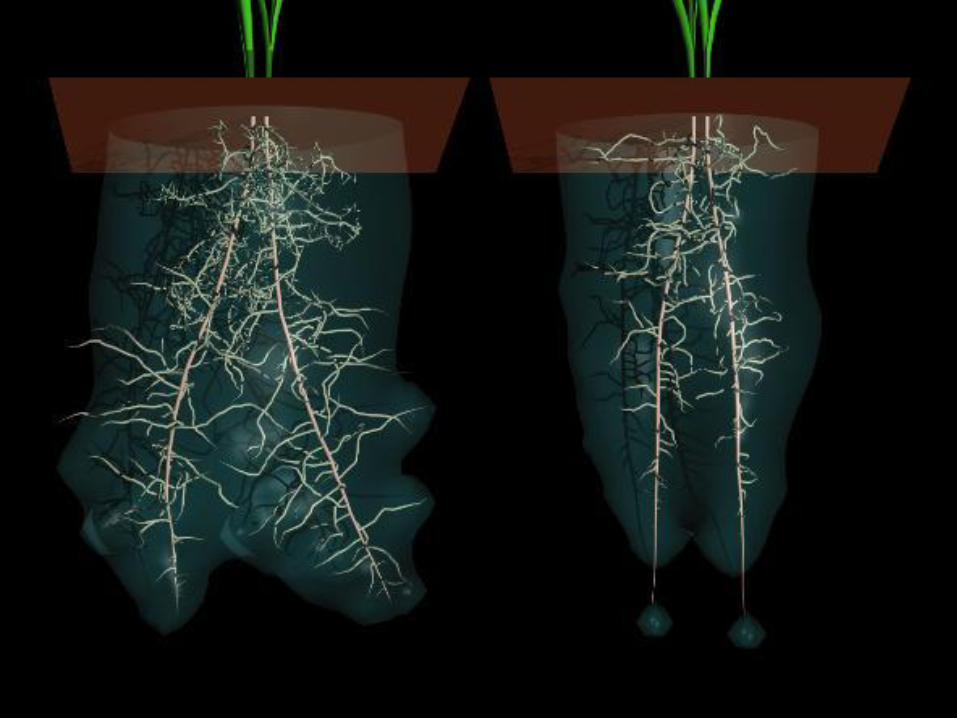


…anebo?
• P.Prusinkiewicz: modelování vývojových
mutantů, model fylotaxe uvažující
relokalizaci PIN (PNAS 2006)
• O.Leyser + P.Prusinkiewicz: identifikace
nového regulátoru dormance pupenů
(MAX)
• see e.g. Berleth et al. TiPS 2007