rapport stage pm
TRANSCRIPT

Biogenèse des Signaux PeptidiquesUniversité Pierre et Marie Curie - ER3 BIOSIPE
Responsables de stage : Sandrine Cadel, Thierry Foulon
Parcours « Molécules et Cibles Thérapeutiques »Année 2009-2010
Patricia Masson
1
Recherche d’acides aminés impliqués dans la spécificité de reconnaissance du substrat et dans le mécanisme
catalytique de l’Aminopeptidase B

L’aminopeptidase B (EC 3.4.11.6)
• Enzyme monomérique : 72 kDa
• Ubiquitaire chez les vertébrés
• Zn2+-métallopeptidase
• HEXXHX18 E - Famille M1
• Enlève les résidus basiques (R, K) en NH2 terminal des peptides (Arg-enképhalines…)
• Activation par les ions chlore
• Phylogénétiquement très proche de la LTA4 hydrolase (33% identité, 48% similarité de séquence)
2
• Présente une activité résiduelle leucotriène A4 hydrolase in vitro
• Enzyme bifonctionnelle
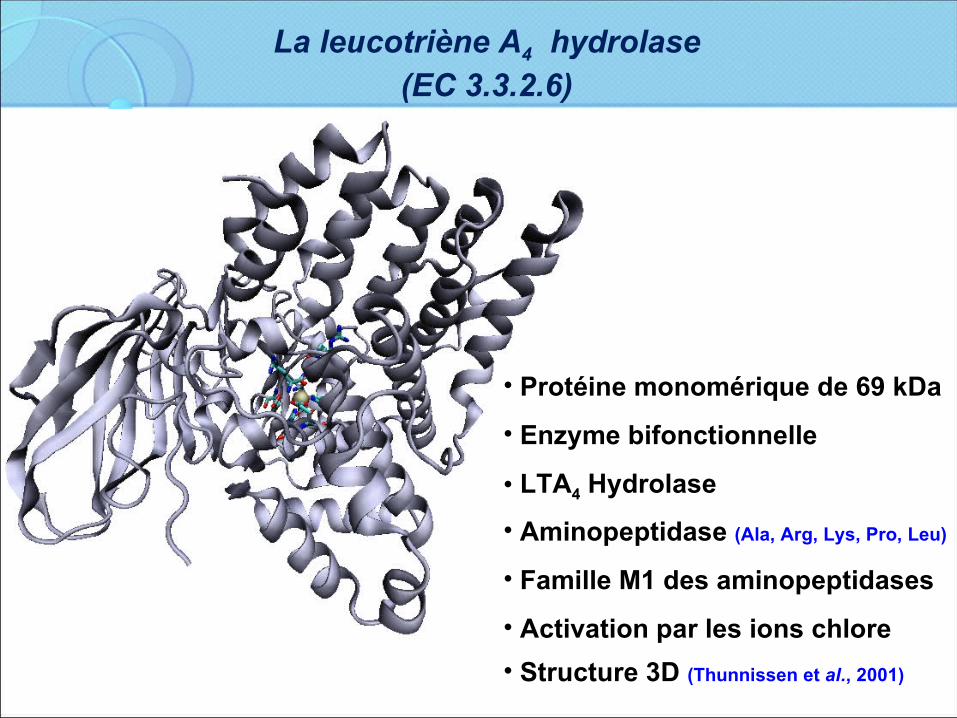
• Protéine monomérique de 69 kDa
• Enzyme bifonctionnelle
• LTA4 Hydrolase
• Aminopeptidase (Ala, Arg, Lys, Pro, Leu)
• Famille M1 des aminopeptidases
• Activation par les ions chlore
• Structure 3D (Thunnissen et al., 2001)
La leucotriène A4 hydrolase (EC 3.3.2.6)
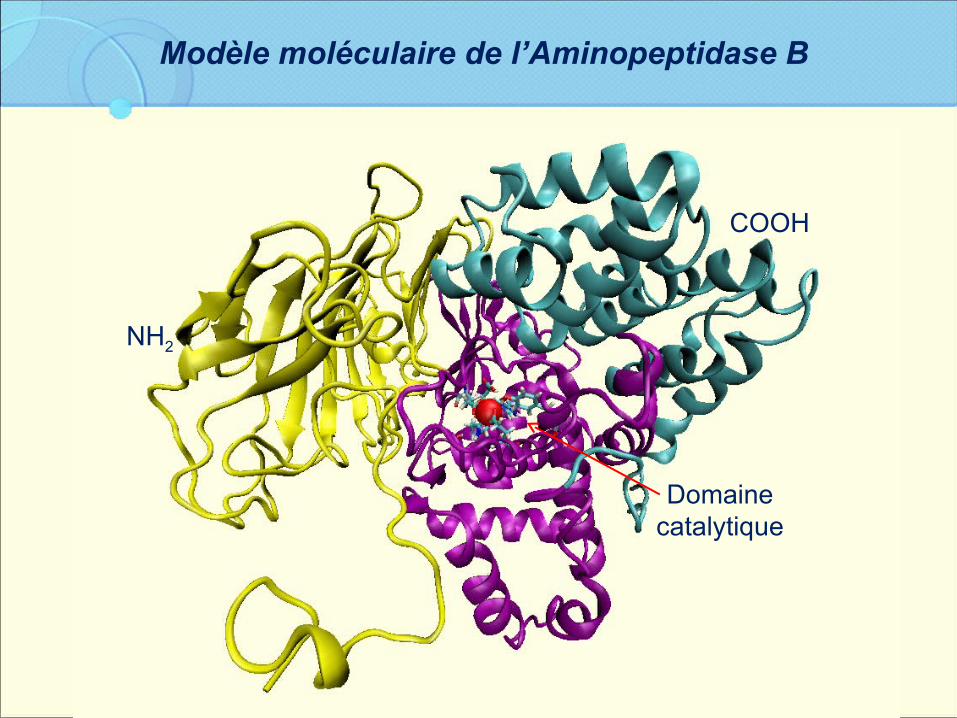
Modèle moléculaire de l’Aminopeptidase B
NH2
Domaine catalytique
COOH
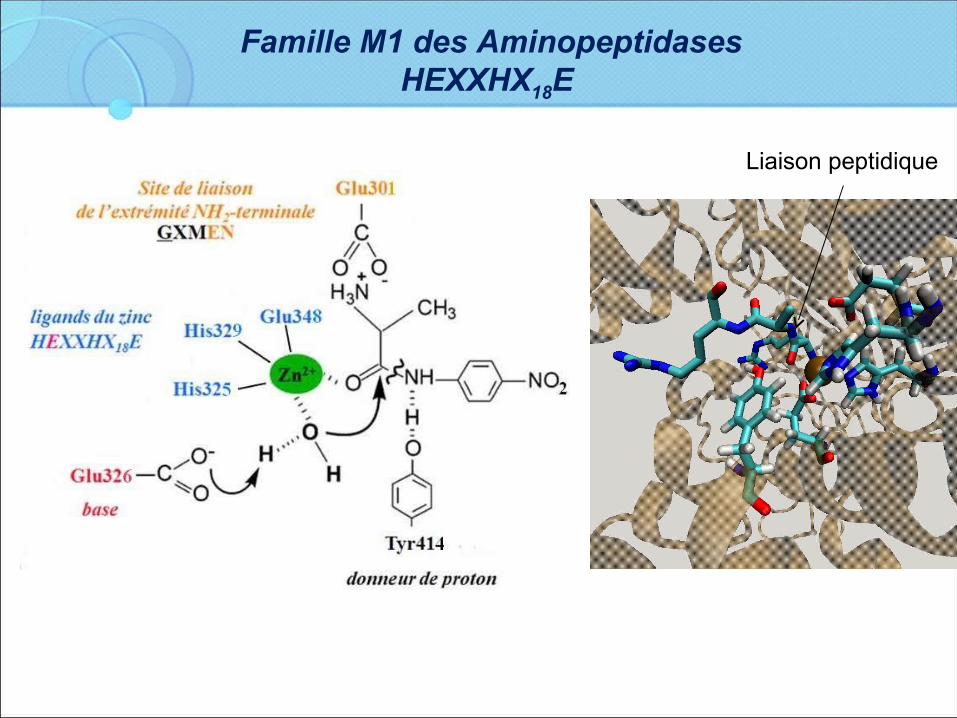
Famille M1 des Aminopeptidases HEXXHX18E
Liaison peptidique
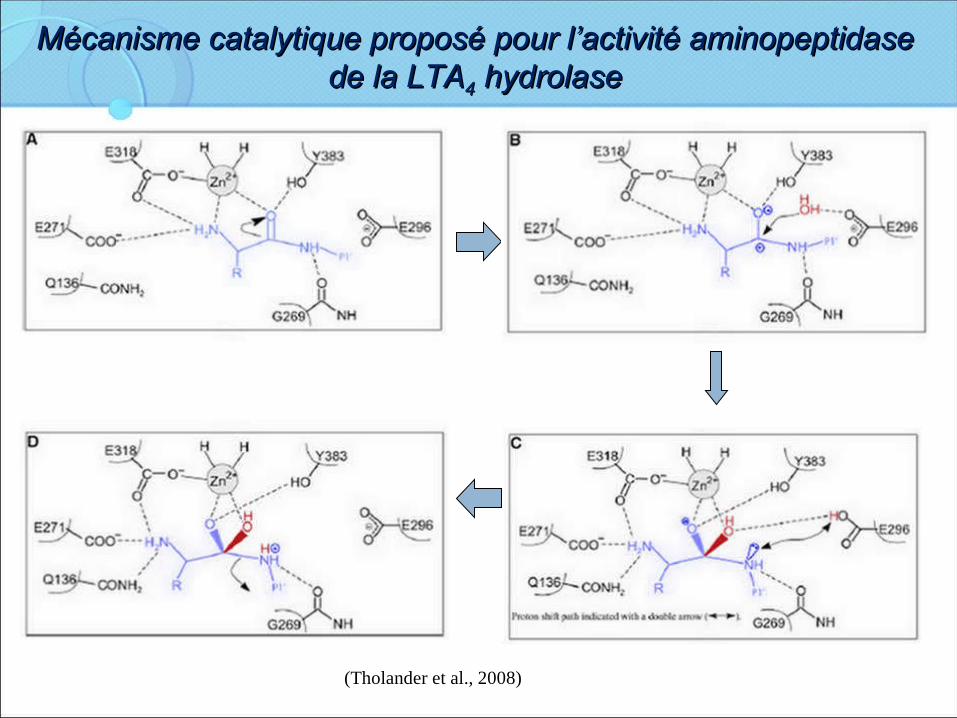
Mécanisme catalytique proposé pour l’activité aminopeptidase Mécanisme catalytique proposé pour l’activité aminopeptidase de la LTAde la LTA44 hydrolase hydrolase
(Tholander et al., 2008)
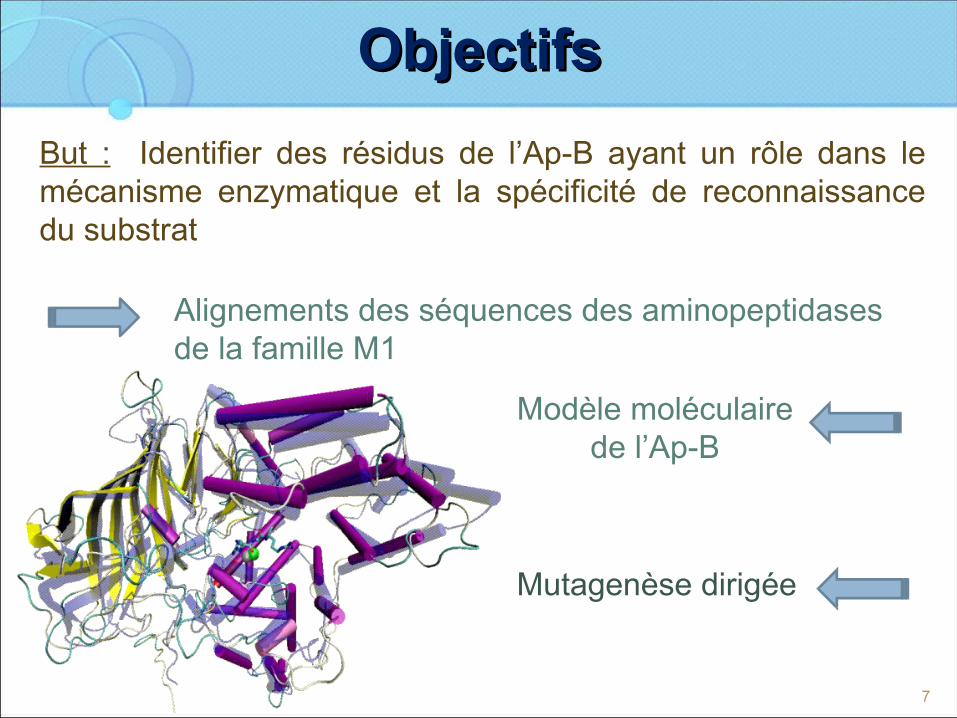
ObjectifsObjectifs
But : Identifier des résidus de l’Ap-B ayant un rôle dans le mécanisme enzymatique et la spécificité de reconnaissance du substrat
Mutagenèse dirigée
Alignements des séquences des aminopeptidases de la famille M1
Modèle moléculaire de l’Ap-B
7
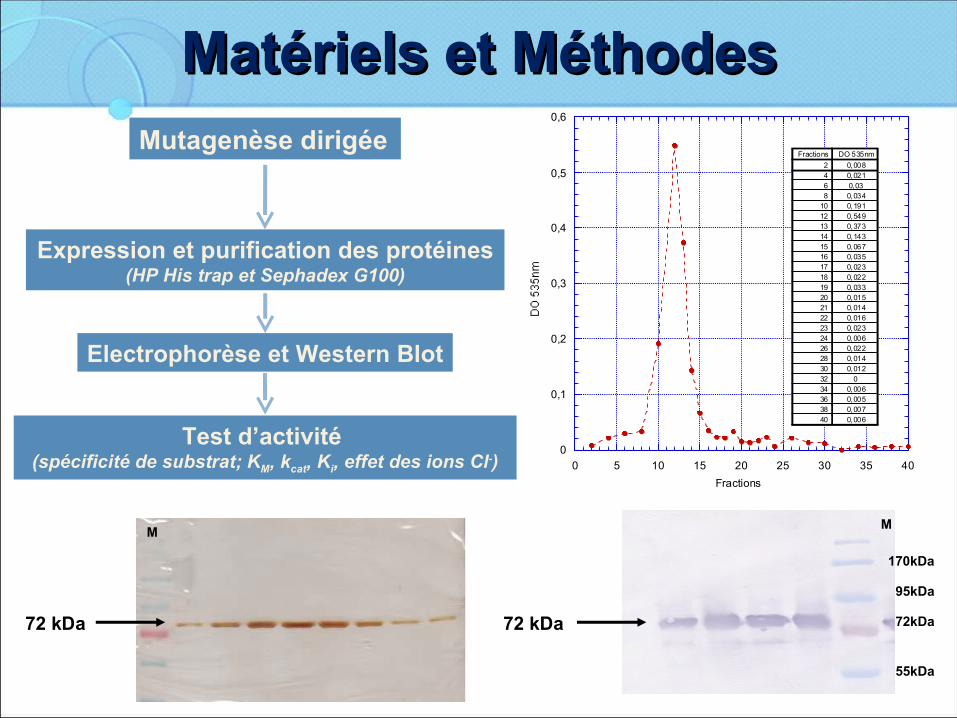
Matériels et MéthodesMatériels et MéthodesMutagenèse dirigée
Expression et purification des protéines (HP His trap et Sephadex G100)
Test d’activité (spécificité de substrat; KM, kcat, Ki, effet des ions Cl-)
Electrophorèse et Western Blot
M
72 kDa
M
72 kDa
170kDa
95kDa
72kDa
55kDa
0
0,1
0,2
0,3
0,4
0,5
0,6
0 5 10 15 20 25 30 35 40
D404V Colonne NiDO 535nm
Fractions
DO 535nmFractions
0,00820,02140,0360,03480,191100,549120,373130,143140,067150,035160,023170,022180,033190,015200,014210,016220,023230,006240,022260,014280,01230
0320,006340,005360,007380,00640
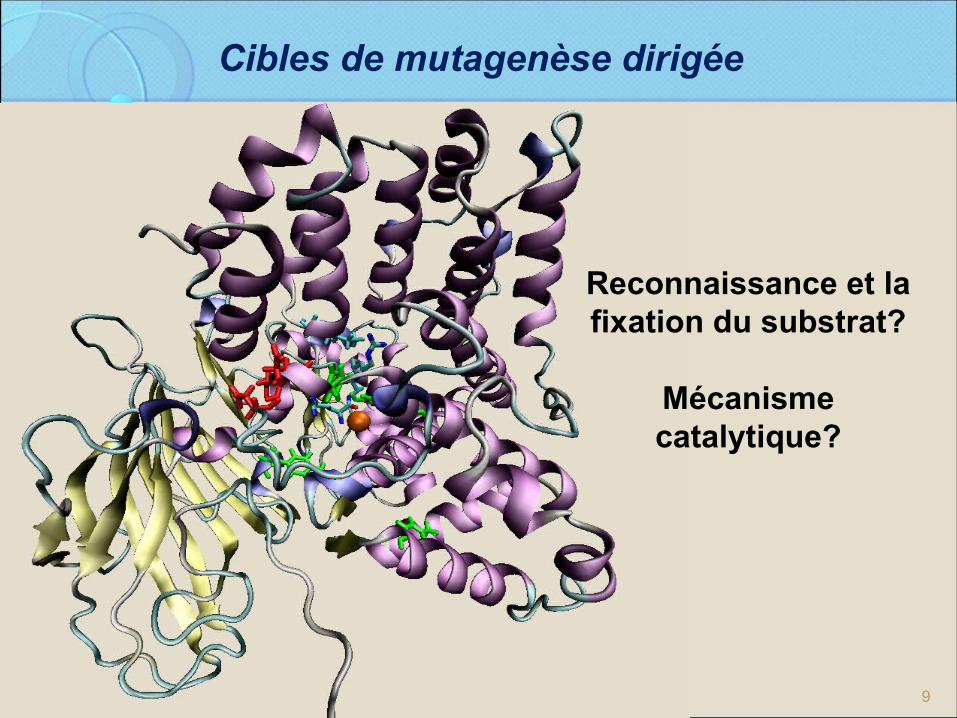
9
Cibles de mutagenèse dirigée
Reconnaissance et la fixation du substrat?
Mécanisme catalytique?
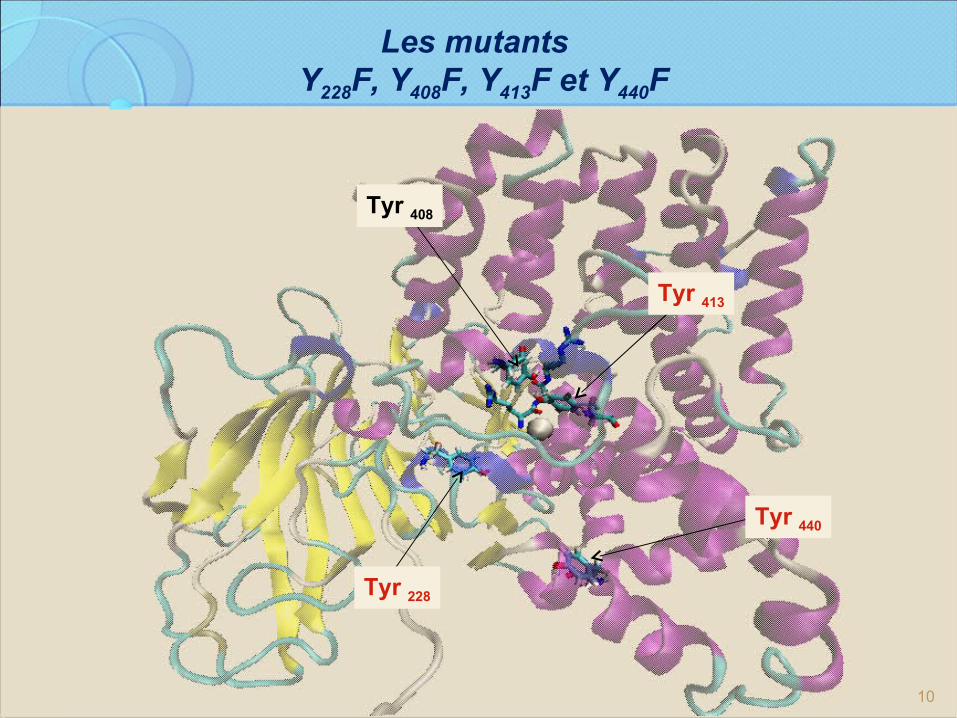
10
Les mutants Y228F, Y408F, Y413F et Y440F
Tyr 408
Tyr 413
Tyr 228
Tyr 440

Les mutants Y408F et Y440F
0
0,4
0,8
1,2
1,6
0 2 4 6 8 10
Y409F Effet du NaCl
DO 535nm + NaClDO 535nm - NaCl
Quantité d'enzyme (µl)0
0,2
0,4
0,6
0,8
1
1,2
0 2 4 6 8 10
Y441F Effet du NaCl
DO 535nm + NaClDO 535nm - NaCl
Quantité d'enzyme (µl)
• Effet des ions chlore
Augmentation de l’activité d’un facteur 3 en présence de 150 mM NaCl
0
0,2
0,4
0,6
0,8
1
1,2
0 2 4 6 8 10
Y441F Effet du NaCl
DO 535nm + NaClDO 535nm - NaCl
Quantité d'enzyme (µl)
Y408F Y440F
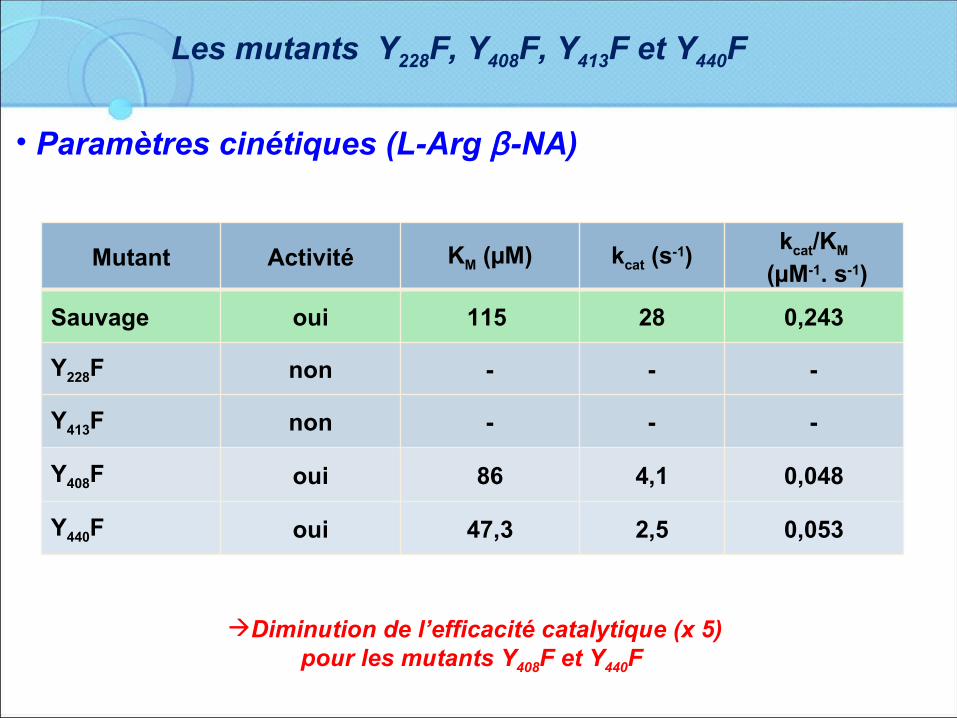
Mutant Activité KM (µM) kcat (s-1)kcat/KM
(µM-1. s-1)
Sauvage oui 115 28 0,243
Y228F non - - -
Y413F non - - -
Y408F oui 86 4,1 0,048
Y440F oui 47,3 2,5 0,053
• Paramètres cinétiques (L-Arg β-NA)
Diminution de l’efficacité catalytique (x 5) pour les mutants Y408F et Y440F
Les mutants Y228F, Y408F, Y413F et Y440F
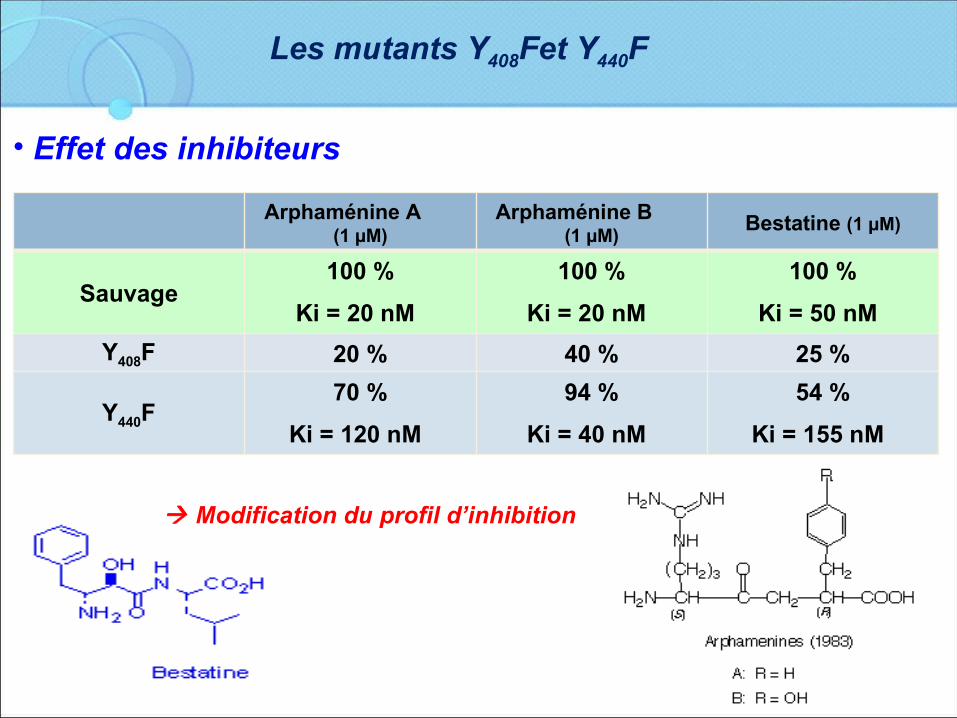
Arphaménine A (1 µM)
Arphaménine B (1 µM)
Bestatine (1 µM)
Sauvage100 %
Ki = 20 nM 100 %
Ki = 20 nM 100 %
Ki = 50 nM Y408F 20 % 40 % 25 %
Y440F70 %
Ki = 120 nM 94 %
Ki = 40 nM 54 %
Ki = 155 nM
• Effet des inhibiteurs
Modification du profil d’inhibition
Les mutants Y408Fet Y440F
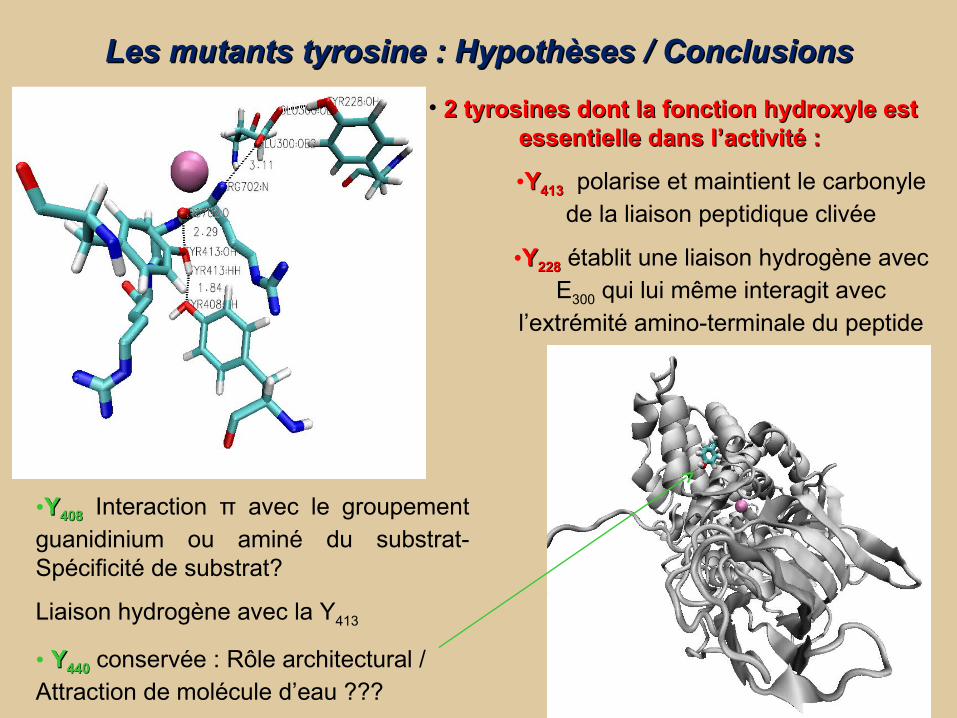
Les mutants tyrosine : Hypothèses / ConclusionsLes mutants tyrosine : Hypothèses / Conclusions
• 2 tyrosines dont la fonction hydroxyle est 2 tyrosines dont la fonction hydroxyle est essentielle dans l’activité : essentielle dans l’activité :
•YY413413 polarise et maintient le carbonyle de la liaison peptidique clivée
•YY228228 établit une liaison hydrogène avec E300 qui lui même interagit avec
l’extrémité amino-terminale du peptide
•YY408408 Interaction π avec le groupement guanidinium ou aminé du substrat- Spécificité de substrat?
Liaison hydrogène avec la Y413
• YY440440 conservée : Rôle architectural / Attraction de molécule d’eau ???
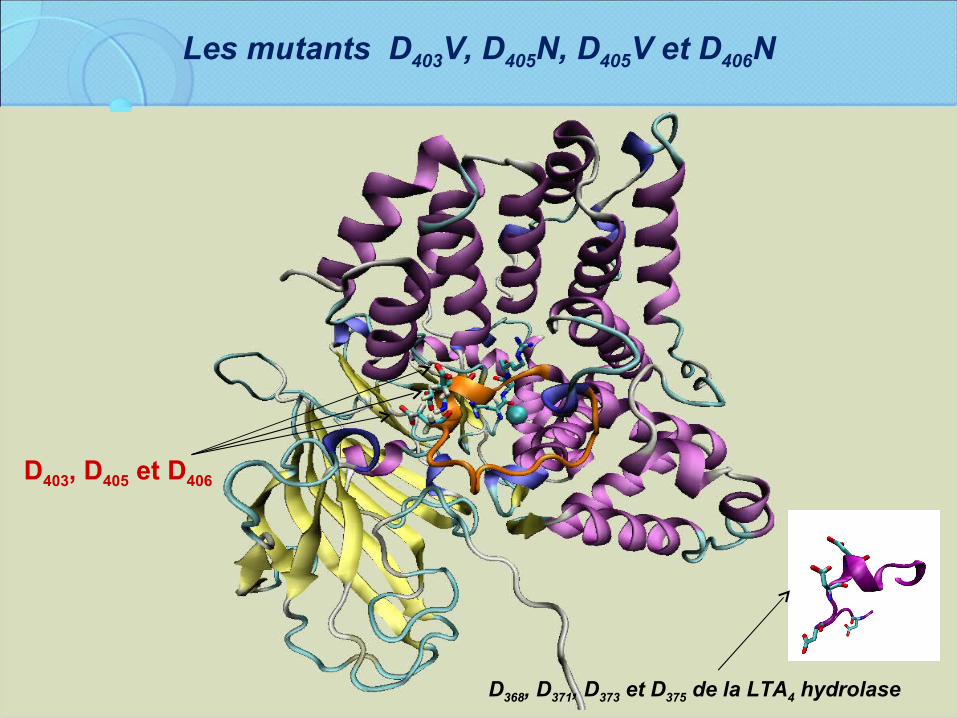
Les mutants D403V, D405N, D405V et D406N
Gel
D403, D405 et D406
D368, D371, D373 et D375 de la LTA4 hydrolase
16
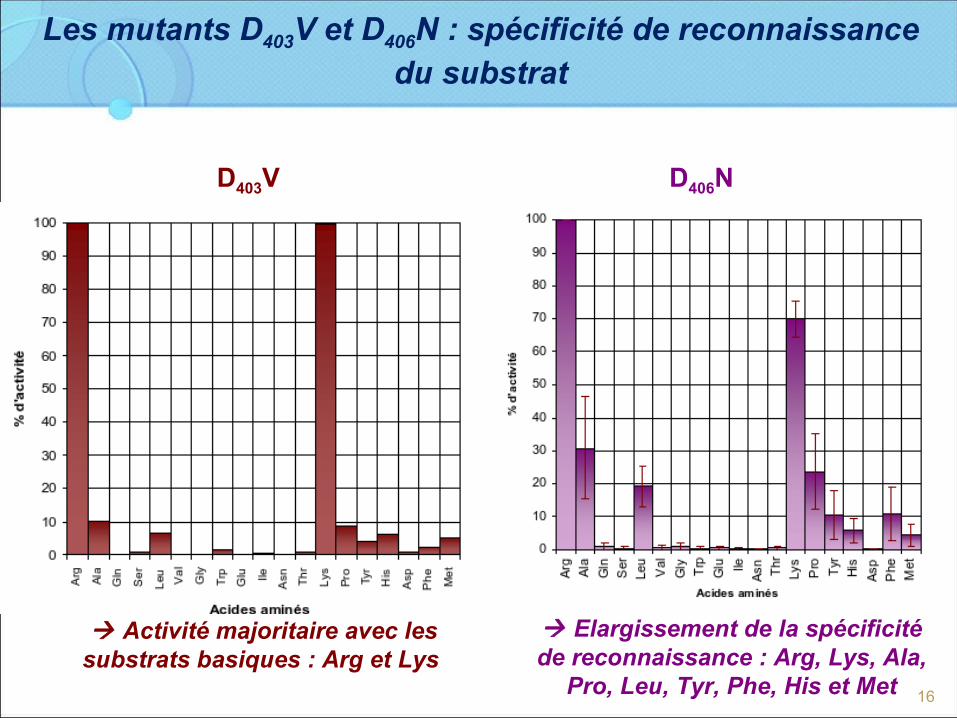
Les mutants D403V et D406N : spécificité de reconnaissance du substrat
D406ND403V
Activité majoritaire avec les substrats basiques : Arg et Lys
Elargissement de la spécificité de reconnaissance : Arg, Lys, Ala,
Pro, Leu, Tyr, Phe, His et Met
17
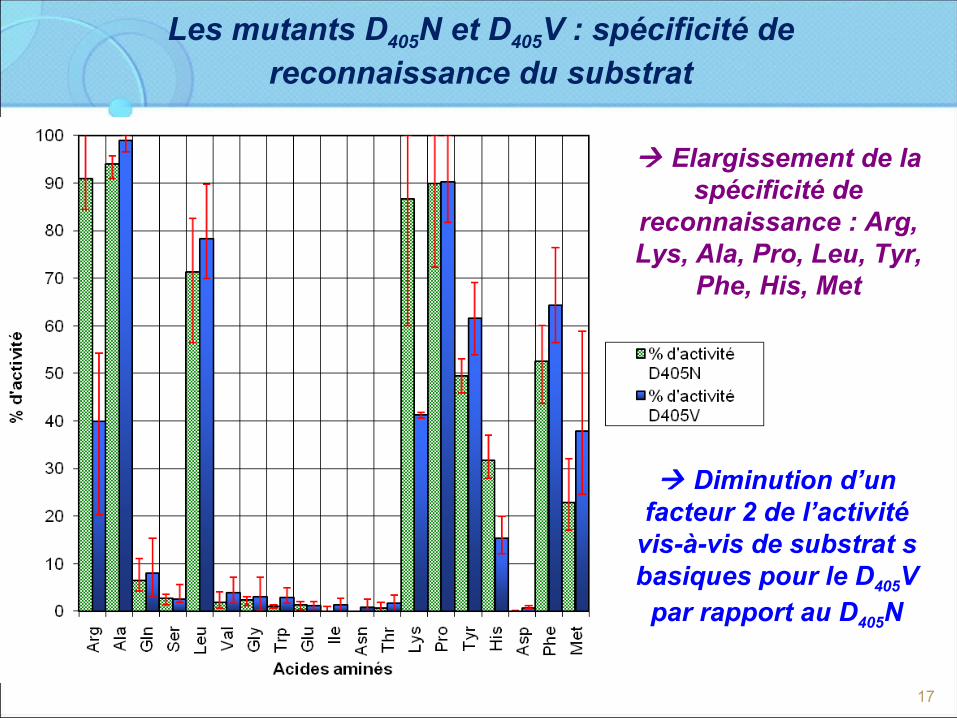
Les mutants D405N et D405V : spécificité de reconnaissance du substrat
Diminution d’un facteur 2 de l’activité
vis-à-vis de substrat s basiques pour le D405V
par rapport au D405N
Elargissement de la spécificité de
reconnaissance : Arg, Lys, Ala, Pro, Leu, Tyr,
Phe, His, Met
18
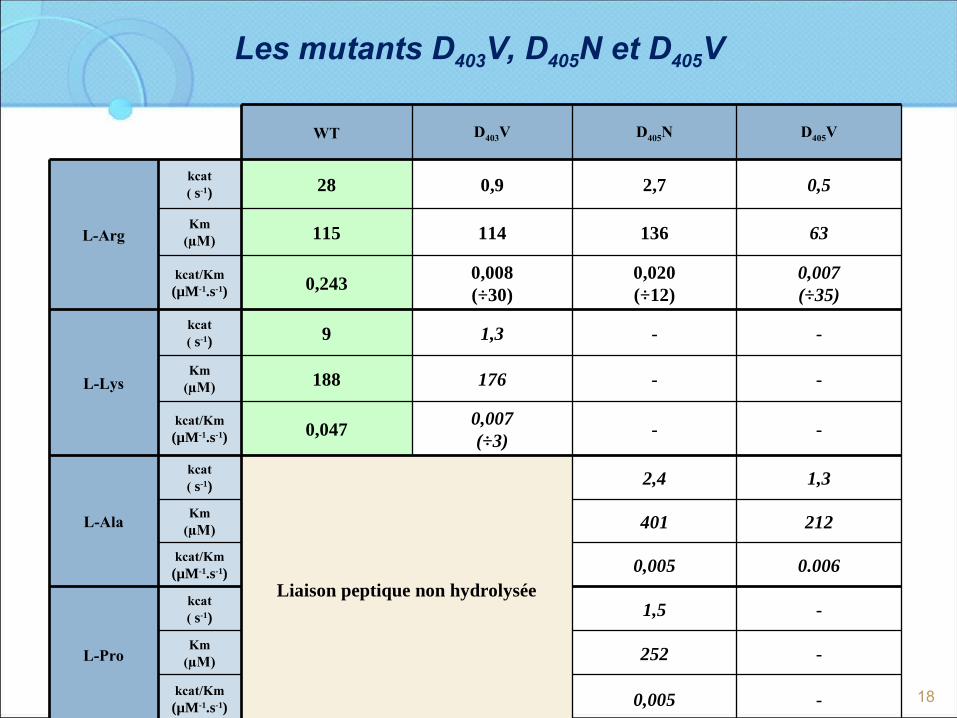
Les mutants D403V, D405N et D405V
WT D403V D405N D405V
L-Arg
kcat( s-1) 28 0,9 2,7 0,5
Km(µM) 115 114 136 63
kcat/Km(µM-1.s-1) 0,243
0,008(÷30)
0,020(÷12)
0,007(÷35)
L-Lys
kcat( s-1) 9 1,3 - -
Km(µM) 188 176 - -
kcat/Km(µM-1.s-1) 0,047
0,007(÷3)
- -
L-Ala
kcat( s-1)
Liaison peptique non hydrolysée
2,4 1,3
Km(µM) 401 212
kcat/Km(µM-1.s-1) 0,005 0.006
L-Pro
kcat( s-1) 1,5 -
Km(µM) 252 -
kcat/Km(µM-1.s-1) 0,005 -
19
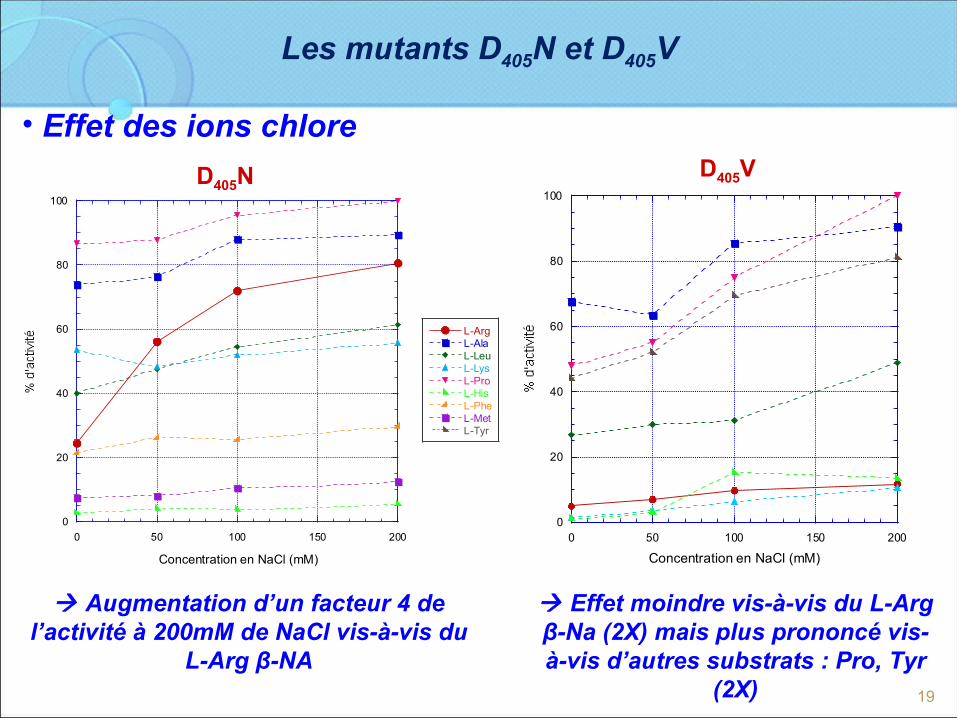
Les mutants D405N et D405V
• Effet des ions chlore
0
20
40
60
80
100
0 50 100 150 200
D406N : % d'activité = f (Concentration en NaCl)
L-ArgL-AlaL-LeuL-LysL-ProL-HisL-PheL-MetL-Tyr
Concentration en NaCl (mM)
0
20
40
60
80
100
0 50 100 150 200
D406V : % d'activité = f ( Concentration en NaCl)
Concentration en NaCl (mM)
Augmentation d’un facteur 4 de l’activité à 200mM de NaCl vis-à-vis du
L-Arg β-NA
Effet moindre vis-à-vis du L-Arg β-Na (2X) mais plus prononcé vis-à-vis d’autres substrats : Pro, Tyr
(2X)
D405ND405V
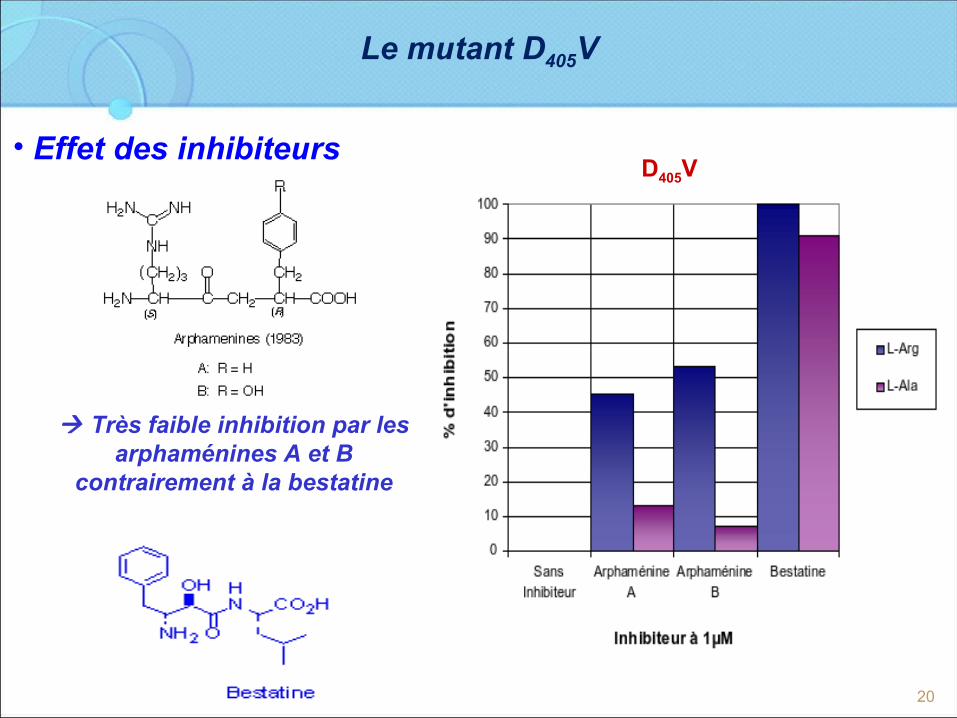
20
Le mutant D405V
• Effet des inhibiteurs
Très faible inhibition par les arphaménines A et B
contrairement à la bestatine
D405V
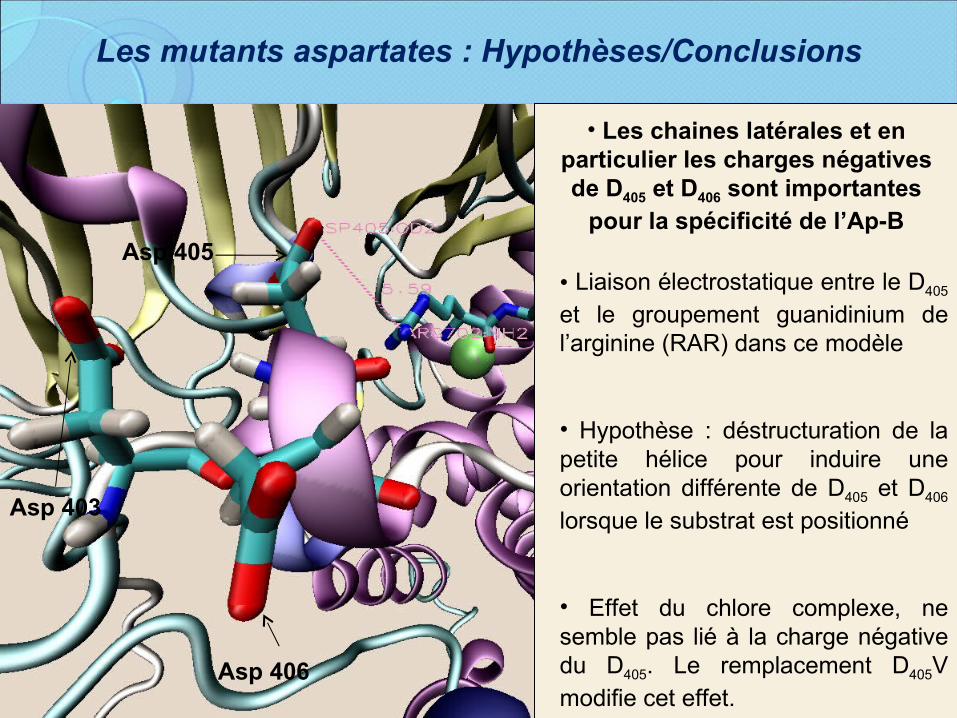
Les mutants aspartates : Hypothèses/Conclusions
• Liaison électrostatique entre le D405 et le groupement guanidinium de l’arginine (RAR) dans ce modèle
• Hypothèse : déstructuration de la petite hélice pour induire une orientation différente de D405 et D406 lorsque le substrat est positionné
• Effet du chlore complexe, ne semble pas lié à la charge négative du D405. Le remplacement D405V modifie cet effet.
Asp 405
Asp 403
Asp 406
• Les chaines latérales et en particulier les charges négatives de D405 et D406 sont importantes
pour la spécificité de l’Ap-B
PerspectivesPerspectivesConfirmer les résultats obtenus
Effet du chlore : paramètres cinétiques (KM, kcat) en fonction du substrat et de la mutation
Concevoir d’autres mutants:Simples: F296Y, K599G, Y408G…Doubles: F296Y/K599R; D405N/D406N, D405V/D406V …Triples: D403A/D405A/D406A….
Activité vis-à-vis de différents peptides
Informations structurales (dichroïsme circulaire, spectroscopie de fluorescence)
Structure 3D avec un inhibiteur confirmation du modèle proposé

À plus long terme : À plus long terme :
Décrypter les mécanismes catalytiques
Différences entre ApB et la LTA4 hydrolase(bifonctionnalité)
Conception d’ inhibiteurs spécifiques de chacune des activités de l’ApB Cibler pathologies
Identifier les substrats peptidiques physiologiques de l’ApB
24
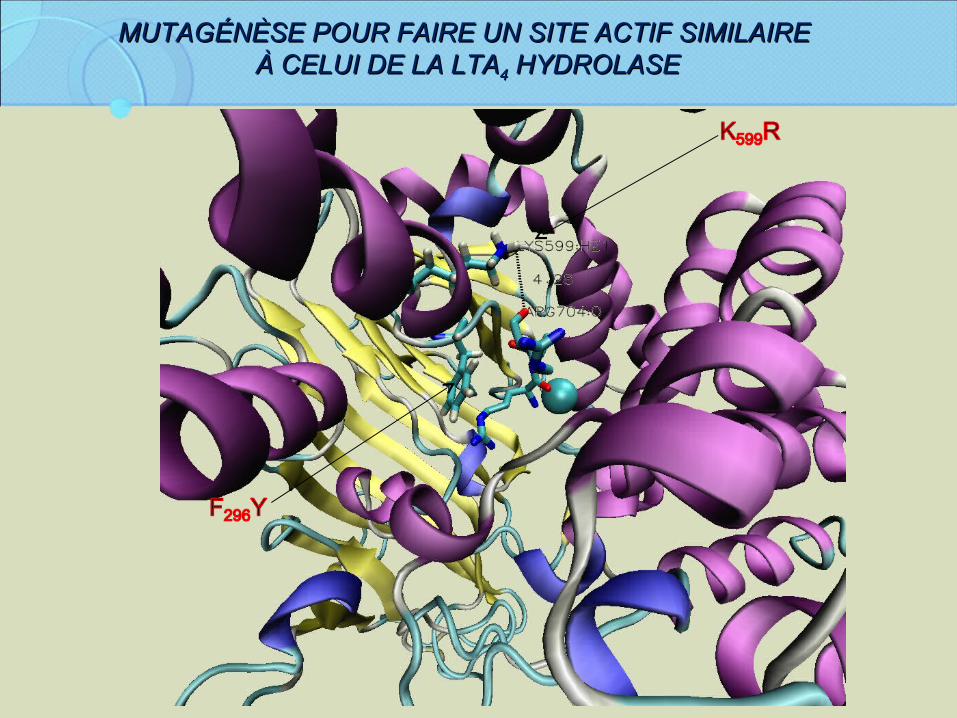
MUTAGÉNÈSE POUR FAIRE UN SITE ACTIF SIMILAIREMUTAGÉNÈSE POUR FAIRE UN SITE ACTIF SIMILAIRE À CELUI DE LA LTAÀ CELUI DE LA LTA44 HYDROLASE HYDROLASE
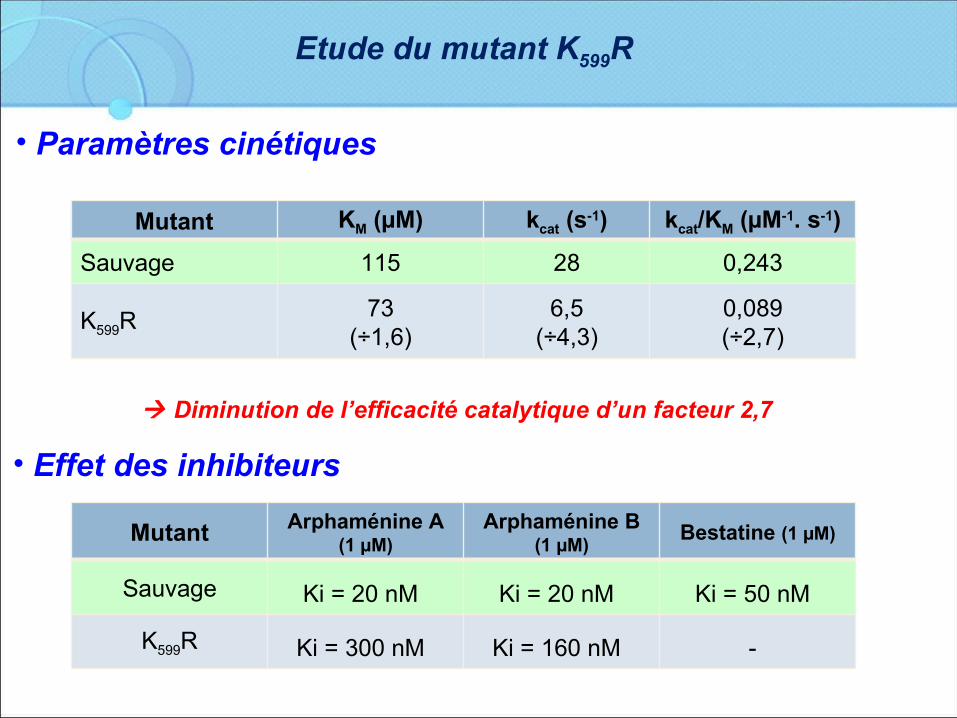
Etude du mutant K599R
Mutant KM (µM) kcat (s-1) kcat/KM (µM-1. s-1)
Sauvage 115 28 0,243
K599R73
(÷1,6)6,5
(÷4,3)0,089(÷2,7)
• Paramètres cinétiques
Diminution de l’efficacité catalytique d’un facteur 2,7
• Effet des inhibiteurs
Mutant Arphaménine A (1 µM)
Arphaménine B (1 µM)
Bestatine (1 µM)
Sauvage Ki = 20 nM Ki = 20 nM Ki = 50 nM K599R Ki = 300 nM Ki = 160 nM -
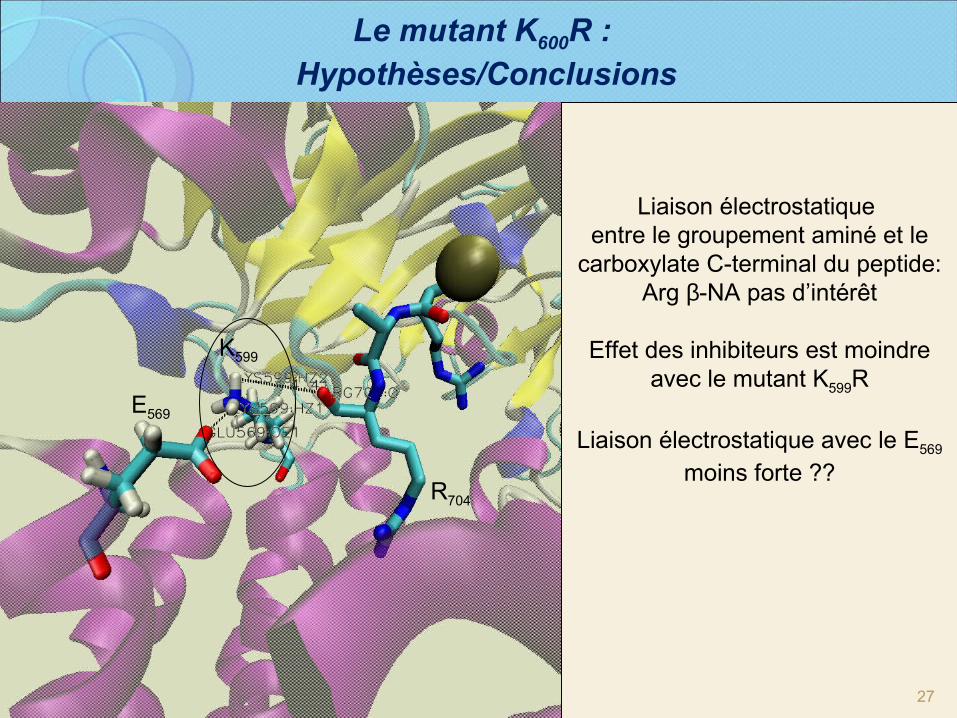
27
Liaison électrostatique entre le groupement aminé et le
carboxylate C-terminal du peptide:Arg β-NA pas d’intérêt
Effet des inhibiteurs est moindre avec le mutant K599R
Liaison électrostatique avec le E569 moins forte ??
K599
E569
R704
Le mutant K600R : Hypothèses/Conclusions