santiago bustamante sanint - javeriana.edu.co · 4 agradecimientos quiero agradecer a la pontificia...
TRANSCRIPT

1
Modelado de especies invasoras, caso de estudio: pérdida del espejo de agua en la laguna de
Fúquene por invasión del buchón (Eichhornia crassipes).
Santiago Bustamante Sanint
PONTIFICIA UNIVERSIDAD JAVERIANA
FACULTAD DE INGENIERÍA
BOGOTÁ D.C
2010

2
Modelado de especies invasoras, caso de estudio: pérdida del espejo de agua en la laguna de
Fúquene por invasión del buchón (Eichhornia crassipes).
Santiago Bustamante Sanint
Ecólogo
Trabajo de Grado presentado como requisito para optar
al título de Magister en Hidrosistemas
DIRECTOR:
EFRAÍN ANTONIO DOMÍNGUEZ CALLE
Ing. Hidrólogo, MSc. Ecología Hidrometeorológica
PhD. en Ciencias Técnicas
CODIRECTOR:
ANDRES ETTER
Biólogo, MSc. Land Ecology
PhD. En Ecología
PONTIFICIA UNIVERSIDAD JAVERIANA
FACULTAD DE INGENIERÍA
BOGOTÁ D.C
2010

3
A la memoria de mi abuelo Guillermo Sanint Botero

4
Agradecimientos
Quiero agradecer a la Pontificia Universidad Javeriana, al cuerpo docente de la Maestría en
Hidrosistemas de la Facultad de Ingeniería Civil, al departamento de Ecología y Territorio de la
Facultad de estudios ambientales y Rurales, especialmente al director de este trabajo, Efraín
Antonio Domínguez Calle y al codirector Andrés Etter. Agradezco a los miembros del proyecto
―Modelamiento de procesos de invasión biológica de especies vegetales en la Región Andina
(Colombia): una aproximación a múltiples escalas‖, a Armando Sarmiento, Jaime Burbano y
Juan Sebastián Lozano por sus múltiples aportes para la elaboración de esta investigación. Así
mismo, agradezco a Natalia García Mora por la corrección de estilo de este documento y su
apoyo moral y sentimental durante todo el transcurso de la maestría; a mi abuelo Guillermo
Sanint Botero por financiar mis estudios y a mi mamá Hilda María Sanint Salazar por sus aportes
y comentarios durante el desarrollo de este trabajo.
Este trabajo se realizó como requisito para obtener el grado de Maestría en Hidrosistemas. La
investigación fue enmarcada en el proyecto dirigido por Andrés Etter ―Modelamiento de
procesos de invasión biológica de las especies vegetales en la Región Andina (Colombia): una
aproximación a múltiples escalas‖ que fue financiado por Colciencias (CT 239-08) y
desarrollado en el departamento de Ecología y Territorio de la Facultad Estudios Ambientales y
Rurales de la Pontificia Universidad Javeriana.

5
Índice
Agradecimientos ............................................................................................................................. 4
Índice............................................................................................................................................... 5
Índice de tablas ............................................................................................................................... 6
Índice de figuras .............................................................................................................................. 7
1 Introducción ............................................................................................................................ 8
2 Objetivos ............................................................................................................................... 13
2.1 Objetivo general ............................................................................................................. 13
2.2 Objetivos específicos...................................................................................................... 13
3 Marco referencial .................................................................................................................. 14
3.1 El Buchón ....................................................................................................................... 14
3.2 Las invasiones biológicas ............................................................................................... 15
3.3 Modelado ........................................................................................................................ 18
3.4 Sistemas de información geográfica .............................................................................. 22
3.5 Autómatas celulares ....................................................................................................... 23
4 Región geográfica del estudio ............................................................................................... 29
4.1 La cuenca Ubaté – Suárez .............................................................................................. 31
4.1.1 Usos e importancia económica de la laguna ........................................................... 32
5 Metodología .......................................................................................................................... 33
5.1 Caracterización de la base empírica. .............................................................................. 35
5.2 Caracterización y aplicación del modelo de autómatas celulares .................................. 38
6 Resultados ............................................................................................................................. 42
6.1 Crecimiento histórico del buchón 1955-2007 ................................................................ 42
6.2 Descripción del modelo .................................................................................................. 46
7 Discusión............................................................................................................................... 51
8 Conclusiones y recomendaciones ......................................................................................... 56
9 Anexos .................................................................................................................................. 58
10 Bibliografía ........................................................................................................................... 59

6
Índice de tablas
Tabla 1. Ejemplo de función de transición de un autómata celular. ............................................. 28
Tabla 2. Demanda hídrica anual de la cuenca de los ríos Ubaté y Suárez ................................... 32
Tabla 3. Reglas del autómata celular ............................................................................................ 40
Tabla 4. Tabla de contingencia ..................................................................................................... 41
Tabla 5. Superficie observada del agua y del buchón entre 1955 y 2007. ................................... 43

7
Índice de figuras
Figura 1. Fotografía de un parche de buchón en la laguna de Fúquene. ....................................... 14
Figura 2. Diagrama de flujo de las variables que intervienen en el proceso de invasión. ........... 18
Figura 3. Trilogía de Samarsky (1997) ........................................................................................ 19
Figura 4. Esquema general de un modelo matemático ................................................................. 20
Figura 5. Esquema general de los tipos de problemas en modelación.......................................... 21
Figura 6.Tipo información que se maneja en un SIG .................................................................. 23
Figura 7. Posibles estados de un autómata celular de dos dimensiones ....................................... 26
Figura 8.Vecindades de un autómata celular de 2 dimensiones ................................................... 27
Figura 9. Representación gráfica de las funciones de transición de la Tabla 1 ............................ 28
Figura 10. Ejemplo de un autómata celular de 2 dimensiones ..................................................... 28
Figura 11. Mapa hidrográfico de la laguna de Fúquene. .............................................................. 30
Figura 12. Diagrama de flujo indicando la metodología que se utilizó en la investigación. ........ 34
Figura 13. Fotografías aéreas con sus respectivos años ................................................................ 36
Figura 14. Proceso de interpretación de imágenes y generación de mapas .................................. 37
Figura 15. Batimetría de la laguna de Fúquene ............................................................................ 38
Figura 16. Vecindad de Von Neumann......................................................................................... 39
Figura 17. Porcentaje de cobertura de las macrófitas y el espejo de agua en la laguna de Fúquene
observada entre 1955 y 2007 ........................................................................................................ 43
Figura 18. Mapa de cobertura de la laguna de Fúquene ............................................................... 45
Figura 19. Diagrama del flujo que representa el modelo. ............................................................. 47
Figura 20. Resultados gráficos de modelo desde 1955 hasta 2007............................................... 50
Figura 21. Validación de los resultados simulados contra los observados en cada año. .............. 52
Figura 22. Pronostico del crecimiento del buchón entre los años 2010 y 2035............................ 54

8
1 Introducción
El rápido crecimiento de la población humana ha tenido como consecuencia la transformación de
los ecosistemas naturales. En muchos casos estas transformaciones han llevado a procesos de
invasión de plantas exóticas en los ecosistemas naturales (Drake and Lodge 2006). El proceso de
invasión altera la estructura y la función de los ecosistemas causando pérdidas económicas y
ecológicas (Higgins and Richardson 1996, Catford et al. 2009). Los ecosistemas acuáticos no son
la excepción, siendo el buchón Eichhornia crassipes (Mart.) Solms. una de las plantas invasoras
que genera problemas de diferentes índoles (Bock 1969, Barrett and Forno 1982, Charudattan
1986, Mailu 2000, Masifwa et al. 2001, Njiru et al. 2002, Williams et al. 2005, Spencer et al.
2006, Colasanti et al. 2007, Greenfield et al. 2007, Kateregga and Sterner 2007).
El buchón es originario de la cuenca del río Amazonas y ha invadido diversos cuerpos de agua
en los trópicos, las zonas subtropicales y en los climas templados (Bock 1969); su amplia
distribución se caracteriza por poseer una alta tasa de crecimiento, el número de retoños que
posee una planta puede doblarse en tan sólo una semana (Ren and Zhang 2007). Esta especie ha
causado considerables daños al medio ambiente afectando la entrada de la luz, desplazando
especies nativas, dificultando o imposibilitando la navegación y alterando la dinámica natural de
la vegetación lacustre y palustre (Bock 1969, Charudattan 1986, Williams et al. 2005, Franco
Vidal et al. 2007, Kateregga and Sterner 2007);
En las sucesiones vegetales de los cuerpos de agua, el buchón es una planta pionera cuyo hábito
estolonífero crea ―islas‖ densas flotantes que se desarrollan desde los márgenes hacia el centro
del lago. Esta planta brinda un medio favorable para el crecimiento de las hidrófitas emergentes
como las gramíneas y las ciperáceas que estabilizan y ligan al margen estas ―islas‖; estos
cambios culminan con el establecimiento de las especies terrestres (CAR 2000).
Uno de los casos más dramáticos alrededor del mundo y mejor documentado de la invasión del
buchón, es el del lago Victoria (África), en donde el costo del control buchón para Kenia,
Tanzania y Uganda ha sido del orden de miles de millones de dólares (Mailu 2000) debido a
problemas como: (i) reducción de los peces por desoxigenación del agua, a pesar de que puede

9
generar refugio para pequeños peces y aumentar la riqueza de las especies generalistas comunes
en muchos cuerpos de agua (Vitousek 1990, Kateregga and Sterner 2007). (ii) interferencia
física con la pesca y el transporte, (iii) impedimento para suplir el agua en los sistemas urbanos,
rurales e hidroeléctricos, (iv) provisión de hábitat para las especies peligrosas como los caracoles
que causan esquistosomiasis, proliferación de zancudos que transmiten enfermedades y de
serpientes venenosas (Wortmann et al. 1998, Chen et al. 2002, Kateregga and Sterner 2007).
En Colombia el buchón ha invadido diversos cuerpos de agua generando pérdidas millonarias. Se
ha reportado en el embalse de Chivor y el Guavio donde es una problemática por el incremento
de los costos en la producción de energía. En el embalse del Muña el buchón se utiliza para la
captura de nutrientes y descontaminación del agua. Sin embargo no se han realizado estudios
acerca del impacto y distribución a nivel nacional.
En Fúquene los principales trabajos han sido:
Un plan de mejoramiento ambiental regional para la cuenca de la laguna (Corporación
Autónoma Regional (CAR) – Agencia de Cooperación Ambiental del Japon (JICA)
2000)
La clasificación y estado actual de los hábitats de la laguna y las implicaciones para su
manejo (Franco Vidal et al. 2007)
La caracterización biofísica ecológica y sociocultural (Instituto De Investigación De
Recursos Biológicos Alexander Von Humboldt 2004)
Una estrategia para el manejo ambiental realizado por el Consejo Nacional de Política
Económica y Social (CONPES 2006).
Los problemas de la laguna de Fúquene están asociados con la transformación de los bosques
nativos y los humedales a pastizales, y la desecación de la laguna y la adecuación de las tierras

10
con fines agrícolas y ganaderos lo que ha generado un alto impacto negativo sobre el ecosistema
(CAR 2000).
Las transformaciones en la laguna de Fúquene han ocurrido a diferentes escalas espaciales y
temporales, tal es el caso del cambio en el borde lagunar siendo pocos los fragmentos que
conservan una transición natural de la tierra firme al agua, esto ha favorecido el proceso de
eutrofización. La más notoria de estas intervenciones fue la construcción del canal perimetral que
trajo consigo grandes repercusiones sobre la estructura y el funcionamiento del ecosistema. El
canal se abrió con el propósito de evitar la desecación causada por los campesinos y los
terratenientes quienes rellenaban las zonas someras para apropiarse de terrenos de la laguna, con
el fin de aumentar el tamaño de sus hatos ganaderos. La construcción del canal se hizo
excavando y depositando el sedimento extraído en el borde que creó una barrera adicional entre
la ronda y el agua. Debido a esto y a otras acciones humanas, el buchón y otras macrófitas han
crecido notoriamente (CAR 2000).
La expansión del buchón ha estado favorecida por el aporte de gran cantidad de nutrientes
provenientes de la cuenca del río Ubaté (Instituto De Investigación De Recursos Biológicos
Alexander Von Humboldt 2004). Esta planta es la principal causa de la pérdida del espejo de
agua en la laguna de Fúquene, allí llegó como resultado de una acción de manejo que buscaba
disminuir el exceso de los nutrientes del agua para revertir los síntomas de la eutrofización
(Franco Vidal et al. 2007).
La Corporación Autónoma Regional de Cundinamarca - CAR, ha dedicado varios proyectos a la
recuperación ambiental de la laguna y su cuenca desde 1961. Las principales acciones han sido:
la realización del canal perimetral, mencionado anteriormente; el proyecto Checua, el proyecto
de Japan International Cooperation Agency - JICA, los programas de reforestación de la cuenca
para disminuir la escorrentía de los sedimentos a la laguna, la protección de páramos y los
programas de educación ambiental (CAR 2000). Según el CONPES (2006), la CAR planeó
destinar 2652 millones de pesos, entre los años 2007-2010, a actividades de extracción mecánica,
la implementación de control biológico y químico para las malezas acuáticas en los canales
principales, secundarios y en el espejo de agua de la laguna.

11
En el presente estudio se desarrolló una herramienta para simular la pérdida del espejo de agua
de la laguna de Fúquene. Históricamente, el modelado matemático y computacional en ecología
ha sido utilizado para simular el crecimiento poblacional mediante modelos agregados como el
de Lotka-Volterra. Sin embargo, los modelos agregados no tienen en cuenta la distribución
espacial de los individuos y asumen que la distribución es uniforme en todo el terreno.
En la realidad las poblaciones presentan diferentes patrones de distribución: cuando los
individuos tienen una distribución agregada, la competencia intraespecífica es mucho mayor que
cuando la distribución es uniforme o aleatoria y como consecuencia, las tasas de crecimiento de
la población son mucho menores (Begon et al. 1996).
En el caso del buchón su población crece de manera agregada desde los bordes de la laguna
hacia el centro. Cuando una planta está rodeada por otras de la misma especie, la competencia es
más alta y los recursos como la luz se verán reducidos, lo que a su vez disminuirá la actividad
fotosintética y como resultado tendrá un menor crecimiento que una planta aislada (Begon et al.
1996).
Es importante por esta razón, trabajar con modelos que tengan en cuenta la distribución espacial,
lo que se facilita gracias a la capacidad del procesamiento y alto rendimiento de las
computadoras actuales que hacen posible manejar los datos a grandes escalas y con alta
resolución espacial (Qu et al. 2008). Estas técnicas involucran autómatas celulares, modelos
basados en individuos y esquemas de caja ―box-based‖. El uso de autómatas celulares como un
tipo de modelado espacial, permite simular la pérdida del espejo de agua y arroja resultados para
direccionar decisiones acertadas en el establecimiento de los planes de acción. Gracias a esto, se
han desarrollado modelos que involucran la distribución espacial en muchas disciplinas (Qu et
al. 2008).
Los autómatas celulares han sido aplicados en muchos campos incluyendo la evolución del uso
del paisaje en patrones urbanos (White and Engelen 2000, Soares-Filho et al. 2002), la dispersión
del fuego en los bosques (Karafyllidis and Thanailakis 1997), la dinámica de las poblaciones de

12
animales (Gronewold and Sonnenschein 1998) y la dinámica de los cambios de cobertura de la
vegetación (Balzter et al. 1998, Facon et al. 2006), entre otros.
El propósito de este trabajo es desarrollar un modelo de autómatas celulares para simular la
invasión del buchón en la laguna de Fúquene (1950-2030). Se construyó una base empírica
multitemporal de 8 mapas a partir de fotografías aéreas (1950-2007), dichos mapas se utilizaron
como base para la calibración de los parámetros del modelo. Finalmente, se estimó la fecha
aproximada de la pérdida del espejo de agua de la laguna teniendo en cuenta el equivalente de
los pasos de tiempo ( modelados a tiempo real.

13
2 Objetivos
2.1 Objetivo general
Modelar el proceso de invasión espacial del buchón (Eichhornia crassipes) en la laguna de
Fúquene.
2.2 Objetivos específicos
Elaborar los mapas de cobertura de la laguna de Fúquene entre los años 1950 y 2007.
Modelar la población del buchón y simular su crecimiento por medio de autómatas
celulares y modelos de ecuaciones diferenciales ordinarias EDO acopladas.
Validar el modelo contra los mapas empíricos elaborados a partir de las fotografías aéreas
de los diferentes años identificando el margen de error.
Estimar la fecha probable de la pérdida del espejo de agua a partir de la predicción de la
modelación.

14
3 Marco referencial
3.1 El Buchón
El Buchón Eichhornia crassipes (Mart.) Solms (Pontenderiaceae) es una planta acuática con
distribución cosmopolita, nativa de la cuenca del río Amazonas en Sur América, tiene flores de
color entre azul a purpura con puntos amarillos en los pétalos y sus hojas son redondeadas
(Figura 1). El buchón se caracteriza por tener crecimiento modular y formar densos parches
flotantes de los que no se puede obtener información demográfica detallada sin hacer análisis
genéticos (Ge et al. 1999). Por ejemplo, en un parche con muchos módulos que consiste de uno o
más clones, no hay forma de diferenciar a los individuos de los clones, ni su distribución
espacial. La dispersión de los clones tiene un importante impacto en los estudios de ecología
vegetal (Fischer et al. 2000) ya que se cree que da diversidad y estructura de los clones juega un
papel crucial en el éxito reproductivo ―fitness‖ de las especies vegetales (Bock 1969, Eckert et
al. 2003).
Figura 1. Fotografía de un parche de buchón en la laguna de Fúquene.

15
El buchón tiene una tasa de crecimiento muy rápida al punto que puede llegar a duplicar el
número de retoños en tan solo una semana. Debido a sus flores vistosas, a la capacidad de
absorber y capturar nutrientes del agua, ha sido dispersado a cuerpos de agua en todo el mundo
en donde se ha vuelto una plaga con impactos económicos negativos (Navarro and Phiri 2000).
Dada la amplia distribución del buchón y su problemática a nivel mundial, se han realizado
investigaciones para encontrar usos y beneficios económicos a partir de su aprovechamiento
(Johnston 1991). El buchón puede ser utilizado como fertilizante en los sistemas agrarios, como
comida animal, para producir biogás, para producir papel e incluso como material de
construcción. El buchón puede ser usado para controlar la contaminación en cuerpos de agua, ya
que tiene una tasa de absorción de metales pesados muy alta. A pesar de todos estos usos, las
tecnologías para aprovechar el buchón no son económicamente competitivas y por esta razón no
son un mecanismo eficiente de control.
3.2 Las invasiones biológicas
Para que una especie pueda ser considerada invasora debe pasar a través de una variedad de
filtros ambientales, y la capacidad de superar cada uno de estas fases depende de distintas
características.
Primero una especie debe cruzar barreras geográficas y ocupar nuevas áreas. La habilidad de que
una especie pase a través la fase de transporte depende de la tasa a la cual se propaga. Segundo,
la especie deber sobrevivir y tolerar las condiciones ambientales de la zona de llegada. Tercero,
la especie debe adquirir los recursos críticos, sobrevivir a las interacciones con los enemigos
naturales y establecer relaciones mutualistas en la nueva área. Finalmente, la especie debe poder
dispersarse y establecer nuevas poblaciones en nuevos sitios a través del paisaje (Figura 1)
(Hellmann et al. 2008).

16
Figure 1. Modelo conceptual del proceso de invasión de especies.
El rango y la frecuencia de muchas especies se ha incrementado, más allá de los rangos naturales
y poblacionales (Catford et al. 2009), estas especies se definen como especies invasoras. (Catford
et al. 2009). Las especies invasoras afectan negativamente a las especies nativas, la dinámica de
las comunidades, la estructura y la función de los ecosistemas (Blossey and Notzold 1995,
Lonsdale 1999, Alpert et al. 2000, Keane and Crawley 2002, Marco et al. 2002, Cannas et al.
2004, Richardson and Pysek 2006, Catford et al. 2009).
El incremento del comercio internacional ha aumentado considerablemente las distancias de las
migraciones y ha ocasionado que las especies alcancen lugares en los que nunca han estado
(Catford et al. 2009). La probabilidad de que una especie introducida logre establecerse y
competir con las especies residentes es extremadamente bajo, a menos que: (i) las condiciones
ambientales hayan cambiado recientemente, (ii) la especie invasora ocupe un nicho que no ha
sido ocupado por ninguna especie residente y (iii) la especie escape de los predadores y los

17
patógenos que permiten que la especie invasora tenga un éxito reproductivo ―fitness‖ igual o
mayor al de las especies nativas (Blossey and Notzold 1995, Keane and Crawley 2002).
A grandes rasgos se ha observado que las características que determinan que una especie sea
invasora son: un rápido paso de la fase vegetativa a la fase de flor y la capacidad de dispersarse
efectivamente sobre distancias cortas y largas (Moles et al. 2008). Las invasiones son la suma de
tres factores: presión de propagación, factores bióticos y factores abióticos (Figura 2).
Los factores bióticos se refieren los resultados de las interacciones como simbiosis, mutualismo
y competencia entre la especie invasora y las especies nativas. Las especies nativas pueden ganar
o perder interacciones cuando una nueva especie entra a una nueva área. Los factores abióticos
se refieren a que las condiciones y recursos ambientales sean propicios para la supervivencia,
crecimiento y reproducción de las especies invasoras (Moles et al. 2008).
Generalmente el alto nivel de disturbio y la gran productividad del ecosistema (tasas de
crecimiento) facilitan la invasión de las especies. Las especies invasoras son especialmente
exitosas en estados tempranos de sucesión y pueden superar a las especies nativas en ambientes
ricos en recursos (Moles et al. 2008).
A pesar de las generalizaciones mencionadas anteriormente hay que ser precavido, ya que no
siempre se cumplen. Se debe tener en cuenta el contexto en el que ocurre una invasión biológica
(Moles et al. 2008). Muchos estudios han buscado patrones para generalizar el éxito de las
especies invasoras, desafortunadamente muy pocos patrones han emergido de estos trabajos
(Moles et al. 2008).

18
Figura 2. Diagrama de flujo de las variables que intervienen en el proceso de invasión.
3.3 Modelado
Un modelo matemático es entendido como una simplificación de un proceso o fenómeno del
mundo real, expresada con ayuda de simbolismos matemáticos. La aplicación de la modelación
matemática consiste en el reemplazo del objeto cognitivo por su imagen matemática (modelo
matemático) la cual, implementada en algoritmos lógico – numéricos en un ordenador, permite
estudiar las cualidades del proceso original. Este método de cognición conjuga las ventajas de la
teoría y del experimento (Domínguez 2010). El proceso de modelación matemática de cualquier
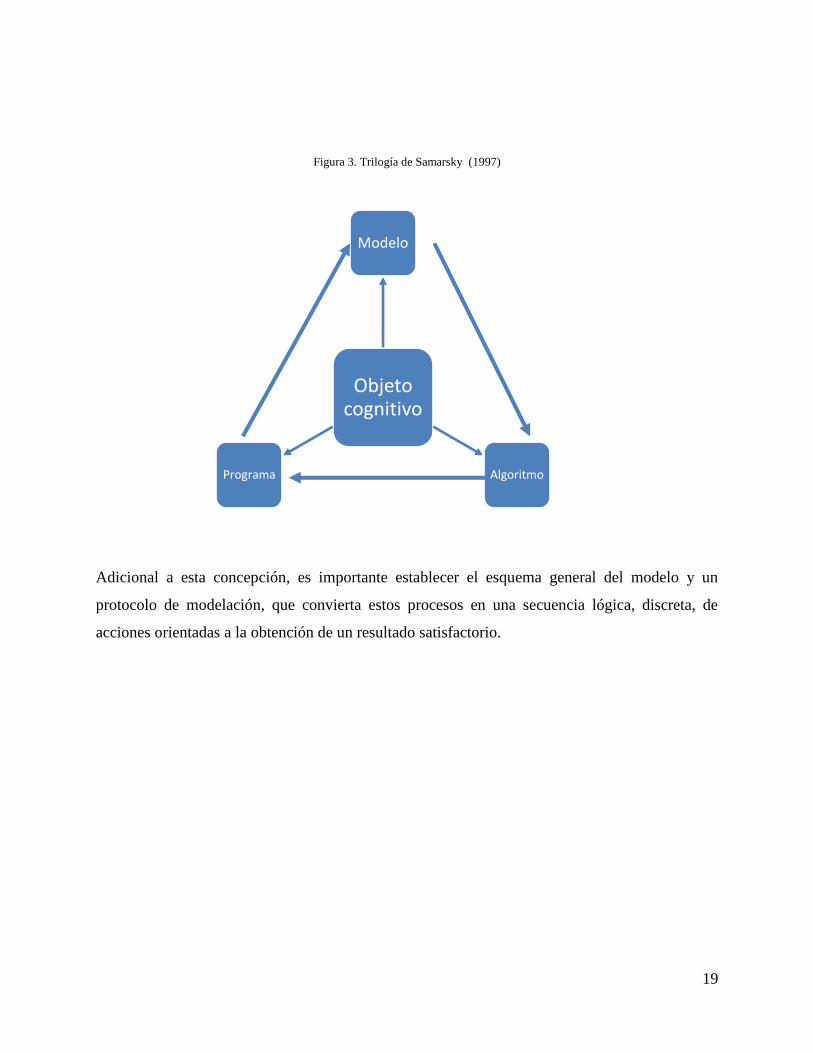
objeto cognitivo consiste en un plan de trabajo preciso que se enmarca en tres (3) etapas que
conforman la trilogía modelo – algoritmo – programa. La descripción secuencial de la trilogía
comenzaría al escoger o construir el equivalente al objeto cognitivo, expresado con
formulaciones matemáticas inferidas del conocimiento previo de los procesos en estudio. La
segunda etapa es escoger o desarrollar el algoritmo de cálculo, que permitirá plasmar el modelo
en un ordenador. El modelo debe permitir la aplicación de métodos numéricos para obtener la
precisión requerida, sin distorsionar las cualidades básicas originales. La tercera etapa es crear
los programas computacionales, que traducen el modelo y el algoritmo a un lenguaje entendible
por el ordenador (Figura 3) (Domínguez 2010).
Presión de
propagación
Factores
bióticos Factores
abióticos
Invasión
Intervención
antrópica
Intervención
antrópica
Intervención
antrópica
19
Figura 3. Trilogía de Samarsky (1997)
Adicional a esta concepción, es importante establecer el esquema general del modelo y un
protocolo de modelación, que convierta estos procesos en una secuencia lógica, discreta, de
acciones orientadas a la obtención de un resultado satisfactorio.
Objeto cognitivo
Modelo
Algoritmo Programa

20
Figura 4. Esquema general de un modelo matemático
Fuente: (Domínguez 2010)
En la Figura 4 se presenta la estructura del esquema general de los modelos matemáticos, en
contexto las entradas -analogía de las causas- representan las variables que activan el flujo de
materia, energía o información en el sistema. Las salidas están conformadas por el vector de
variables que caracterizan el estado del sistema. La estructura del modelo -un mecanismo de
transformación de las entradas en salidas- está conformado por:
El operador: es la estructura matemática que establece las reglas de correspondencia entre
las entradas y las salidas.
Los parámetros: son las magnitudes encargadas de describir las características físicas y
funcionales del sistema.
Las variables de estado: son las magnitudes que representan el estado del sistema y cuya
evolución espacio temporal constituye las salidas del modelo.
Entrada Salida
Función de retroalimentación
Analogía
de causa
Analogía
de efecto
Analogía del sistema
Parámetros Variables
de estado
OPERADOR
ESTRUCTURA DEL MODELO
Analogía de la influencia inversa

21
La función de retroalimentación: mecanismo que refleja el grado en el que las salidas del
modelo influyen en el transcurrir de los procesos del sistema. Cualidad intrínseca del
operador del modelo. Generalmente representa el orden de no linealidad del proceso en
estudio.
En la modelación matemática, se resuelven dos tipos de problemas: el problemas directo y el
problema inverso (Figura 5). El problema directo es la situación ideal en la modelación
matemática, ya que se conocen las entradas, las variables de estado, el operador y los parámetros;
lo único desconocido son las salidas. Se establece como matemáticamente correcto, si en el
problema directo, la solución existe, es única y estable. El problema inverso está dividido en 3
clases: 1) No se conocen los parámetros, el resolverlo establece encontrar los parámetros
idóneos, que aplicados al operador matemático y utilizando las entradas conocidas produce las
salidas observadas, 2) Se desconocen las entradas, a través de este problema se intenta
reconstruir las causas que produjeron determinados efectos, 3) El elemento desconocido es el
operador matemático. Resolver este problema significa encontrar las ecuaciones que mejor
transforman las entradas en salidas.
Figura 5. Esquema general de los tipos de problemas en modelación.
Problema
directo
Operador
Parámetros
Variables
Problema
inverso
Calibración
Operador
????????
Variables
Reconstrucción
Operador
Entradas
Entradas
???????
salidas
salidas
???????

22
Parámetros
Variables
Identificación de
operadores
????????
Parámetros
Variables
Fuente: (Caicedo Carrascal 2008, Domínguez 2010)
3.4 Sistemas de información geográfica
Los sistemas de información geográfica son una herramienta computarizada que permite la
captura, manejo, análisis, y representación en mapas de información geográficamente
referenciada. Las Funcionalidades de un SIG son colectar, guardar y recuperar información
geográficamente localizada, identificar localizaciones especificas en un espacio determinado,
analizar información espacialmente relacionada como base para tomar decisiones respecto a la
gestión del escenario en cuestión, facilitar la generación información que puede ser utilizada
también en modelos para la evaluación de impactos de los instrumentos de política sobre la
unidad territorial analizada y mostrar exhibir y diseñar gráfica y numéricamente el escenario
analizado (Zuñiga 2004).
Los SIG permiten representar entidades del mundo real. Para esto se utilizan objetos. Los objetos
de los SIG pueden ser agrupados en tres grandes categorías: 1. Objetos físicos como carreteras y
causes de agua 2. Objetos que representan gradientes continuos como cambios de temperatura,
altitud, concentración de nutrientes humedad, etc. y 3. Objetos categóricos como los tipos de
vegetación, zonas climáticas o grupos de edades (Bernhardsen 2002).
Los datos geográficos pueden ser divididos en dos grandes características: la geometría y los
atributos. La geometría a su vez puede ser de tres tipos puntos, líneas y área. Los puntos se
utilizan para identificar una localidad específica, las líneas para representar ríos, carreteras o
delimitar zonas y las áreas se utilizan para representar coberturas o características de una
superficie (Bernhardsen 2002). Los atributos se refieren a información acerca del objeto
representado por la geometría. Pueden ser datos cuantitativos o cualitativos (Figura 6)
(Bernhardsen 2002).
Entradas salidas

23
Figura 6.Tipo información que se maneja en un SIG
(Bernhardsen 2002)
La integración de Sistemas de información geográfica (SIG) con simulación por computadores
involucrando las características del medio ambiente para la simulación de invasiones biológicas
comprende tres pasos (Ming and Albrecht 2004):
1. Seleccionar los modelos que capturen los estados, los procesos y las relaciones de las
características que afectan las invasiones biológicas.
2. Evaluar patrones espacio-temporales y explorar las implicaciones de las interacciones
entre la respuesta de los organismos y la heterogeneidad del paisaje.
3. Predecir y simular la evolución espacio- temporal de las invasiones biológicas.
3.5 Autómatas celulares
Una AC es una red discreta de celdas autónomas.
Datos que se manejan en un SIG
geometria
Puntos
Lineas
Poligonos
Atributos
Cuantitativos
Ordinal
intervalos
tasas
Cualitativos
Nominal

24
Celda Autónoma: una celda autónoma es un objeto localizado en una red discreta y provisto de
un conjunto de reglas de autonomía.
Regla de Autonomía, f: Una regla de autonomía es una regla que establece unívocamente una
correspondencia entre el estado interno de un objeto (una celda del autómata en el caso de un
AC) y el estado de la vecindad del objeto (vecindad de la celda en el caso de un AC).
Matemáticamente: : v Cf es decir, f es la regla que produce el estado C de la celda
"C" a partir del estado V de la vecindad V de "C".
Vecindad de una celda en un AC: La vecindad de una celda "C" en un AC es el conjunto de
celdas cuyo estado influye en el estado de "C". Generalmente, la influencia de un vecino sobre
una celda depende de su distancia geométrica a "C", por lo que se habla de "primeros vecinos",
"segundos vecinos", etc.
Un autómata celular tienen al menos 5 propiedades: 1) su dimensión: el número de coordenadas
espaciales de cada punto de la red (su número de grados de libertad), 2) la medida de cada una de
sus dimensiones, en este caso su ancho "A" y su largo "L": los rangos de variación de los índices
"i", i=1,...,A y "j", j=1,...L que identifican la localización de cada celda Cij 3) la regla que define
la vecindad para cada celda y su tamaño: el número de celdas vecinas a Cij 4) el espacio de
estados de las celdas: el conjunto de posibles estados que la celda puede tomar, 5) las reglas de
evolución localizadas: las funciones f que definen la actualización del estado de la celda en cada
paso de tiempo (Qu et al. 2008). Además un AC puede ser caracterizado por 1) el paralelismo:
todos los estados son actualizados al mismo tiempo, 2) la homogeneidad: todas las celdas siguen
las mismas reglas de evolución, 3) la localidad: la celda sólo toma información de las celdas
vecinas y puede ser afectada sólo por ellas (Qu et al. 2008).
Las condiciones de frontera, pueden ser de tres tipos (Schiff 2007):

25
Periódicas (también conocidas como cíclicas): en este tipo de condición los bordes se
tocan entre ellos, es decir la celda del extremo derecho sería vecina a la celda del extremo
izquierdo, y la celda de arriba sería vecina a la celda de abajo, en este caso la grilla
tendría forma de una dona.
Reflectivas: en este caso los bordes de la red tendrían los mismos valores que la celda que
limita con el borde.
Fijas: los bordes presentan valores asignados y se mantienen constantes a lo largo de la
simulación.
Un autómata celular sirve como modelo de un sistema natural. Los estados de las celdas
evolucionan en el tiempo de acuerdo a sus reglas de autonomía. Para un proceso de tiempo
discreto, cada configuración de un autómata celular (su estado) en un tiempo determinado
implica una única configuración en el tiempo siguiente, sin embargo cada configuración del
autómata en un tiempo dado puede tener múltiples predecesores, lo que indica que el proceso es
irreversible. Esto nos lleva a una propiedad importante que tienen los autómatas celulares que es
la irreversibilidad; es decir que entre una configuración y la siguiente hay pérdida de información
(Martin et al. 1984, Kier et al. 2005).
Los modelos de autómatas celulares definen las interacciones entre las especies a nivel local; los
patrones complejos a escala global surgen través de la evolución de los modelos de los
autómatas celulares (Wootton 1988).
Un autómata celular consiste en una grilla de celdas, la mayoría de las veces son cuadradas pero
pueden ser hexagonales o de otras formas. Cada celda puede tener k diferentes estados. Donde k
es un número finito mayor o igual a 2. En el caso más simple la celda puede tener valor de cero y
uno, representados gráficamente por negro y blanco, respectivamente. La grilla puede tener n
(≥1) dimensiones, pero lo más común en el trabajo de autómatas celulares es trabajar con una o
dos dimensiones. Si asumimos que las celdas son cuadradas y que trabajamos con una dimensión,

26
se forma una fila de celdas adyacentes (Figura 7). En principio el número de celdas de un arreglo
es infinito, pero en términos prácticos, se utiliza un arreglo suficientemente largo para ilustrar el
comportamiento del fenómeno en cuestión. Y se imponen ciertas condiciones para las celdas que
limitan con los bordes. En algunos casos se pueden generar restricciones intencionales para
limitar el tamaño de la grilla (Schiff 2007).
Figura 7. Posibles estados de un autómata celular de dos dimensiones
Los ocho posibles estados con r = 1 y k = 2, se está usando la convención más común 0 es negro y 1 es blanco donde r es rango y
k el número de estados.
Siguiendo estas ideas se busca que la grilla evolucione, para esto se necesitan cambios en el
tiempo. Esto es hecho de una manera inusual, se considera que los cambios de estado de las
celdas ocurren en momentos discretos de tiempo. Así, para periodos de tiempo
que pueden ser entendidos como el clic de un reloj. El tiempo cero generalmente es reservado
para las condiciones iniciales antes de que haya un cambio en el estado de las celdas de la grilla.
Otro elemento que se necesita para que el autómata evolucione es una función de transferencia
local la cual indica cómo cada celda cambia en un instante de tiempo basada en ella misma y en
sus vecinas en el tiempo anterior. Este cambio ocurre para todas las celdas de la grilla
sincrónicamente. Las funciones de transición pueden ser determinísticas o probabilísticas. La
grilla de celdas, el conjunto de estados disponibles, junto con las funciones de transferencia, son
llamados autómata celular, que es una configuración determinada de una celda en un estado de
tiempo particular (Schiff 2007). Un autómata tiene tres características fundamentales:
Uniformidad: todas las celdas se actualizan con el mismo conjunto de reglas.
Sincronismo: todas las celdas se actualizan simultáneamente.
Localidad: las reglas se aplican en cada localidad.

27
La mayoría de las características de la dinámica de los autómatas celulares pueden ser
encontradas en los autómatas de una dimensión. En estos se define la vecindad de las celdas c
con un radio (rango) r, y se toman r. celdas hacia la derecha y r celdas a la izquierda de la celda c.
Incluyendo la celda c, la vecindad contiene 2r+1 celdas. En el caso más simple r = 1 y k = 2, se
tendrían tres celdas vecinas con los diferentes estados 0 y 1 para cada celda, que pueden ser
expresados en 23 = 8 diferentes posibilidades (Figura 7 y Figura 8). Como regla general se
tendrían estados de vecindad con una dimensión.
Figura 8.Vecindades de un autómata celular de 2 dimensiones
Los estados de la celda central c y sus vecinas y
en el estado de tiempo t.
El estado de la celda c en el estado de tiempo siguiente es función de la celda y las vecinas en el
tiempo t (Figura 8)
Ecuación 1. Estado de la celda c en el tiempo t+1, f es la función de transición.
[
]
El ejemplo de la función de transición se puede ver en la Tabla 1 y de e manera gráfica se puede representar como se muestra en
la Figura 9. La
Figura 10 es un ejemplo de un autómata celular de dos dimensiones que sigue las reglas de la
Tabla 1 representados en la Figura 9. En él se ve como con unas reglas sencillas y unas
condiciones iniciales de solo una celda negra, cada nueva línea representa la evolución del
modelo en un paso de tiempo da como resultado una estructura compleja de la cual emergen
patrones y figuras con dimensión fractal.

28
Tabla 1. Ejemplo de función de transición de un autómata celular.
1 1 1 1
1 1 0 0
1 0 1 0
1 0 0 1
0 1 1 0
0 1 0 1
0 0 1 1
0 0 0 0
Figura 9. Representación gráfica de las funciones de transición de la Tabla 1
Figura 10. Ejemplo de un autómata celular de 2 dimensiones
Los AC presentan los fenómenos bajo estudio en una grilla de celdas distribuidas en el espacio,
el estado de las celdas afectadas por las vecinas y las reglas de transición. Las celdas son la

29
unidad más pequeña en los AC. Su estado cambia bajo las leyes impuestas por las funciones de
transición de acuerdo a las características de las celdas vecinas. Mientras que los vecinos afectan
los AC, en necesario tener unas reglas de transición claras que sean en pivote de la realidad del
modelo del fenómeno estudiado. Las reglas deben aplicar a cada celda, estado y vecindad. El
cambio de estado debe ser local (Silva et al. 2008).
Los autómatas celulares esquematizan el espacio en una grilla de acuerdo a la escala del sistema
estudiado, cada celda tiene algunas propiedades y toma valores en estados finitos. El valor es
actualizado en tiempo discreto según unas reglas predefinidas que son funciones del estado
actual de la celda misma y las vecinas (Qu et al. 2008).
Los modelos basados en individuos toman cada especie como el objeto de estudio para describir
sus propiedades (edad, genero, tamaño etc.) y acciones (Qu et al. 2008).
Los modelos de caja estudian un espacio y el comportamiento o el mecanismo del objeto de
estudio, considerado homogéneo dentro de la caja. Cada caja implementa su propio conjunto de
ecuaciones dinámicas (masa, flujos de energía) (Qu et al. 2008).
4 Región geográfica del estudio
La laguna de Fúquene (Latitud 5º 29’50‖; 5º 25’26‖ N & Longitud 73º 46’49‖; 73º 42’44‖ O) se
encuentra ubicada a 80 km al norte de Bogotá, en el municipio de Fúquene en el departamento de
Cundinamarca en la cordillera oriental de Colombia. La precipitación promedio de la cuenca es
de 962.5 mm año-1
, con una temperatura media de 13°C, y un clima frío semi-húmedo (Franco
Vidal et al. 2007).
La laguna de Fúquene es surtida por los ríos Ubaté, Susa y Fúquene principalmente. El río Ubaté
desemboca en la laguna por la zona sur, recoge las aguas de los drenajes provenientes de las
lagunas de Cucunubá y Palacio (Figura 11). La laguna de Fúquene tiene un volumen máximo
operativo de 36.8 millones de metros cúbicos (CONPES 2006).

30
La laguna de Fúquene hace parte de los humedales interiores de Colombia, estos son ecosistemas
estratégicos y vitales para el desarrollo presente y futuro de la Nación. Los humedales son
ecosistemas que cumplen múltiples funciones interrelacionadas, prestando diversos servicios
ambientales con un carácter dinámico, por lo tanto, sus componentes y procesos se deben
conservar y sostener (CONPES 2006).
Figura 11. Mapa hidrográfico de la laguna de Fúquene.
Fuente IGAC
N
Laguna de
Fúquene

31
4.1 La cuenca Ubaté – Suárez
La laguna de Fúquene, tiene una superficie promedio de 3100 has. y una profundidad media
estimada de 2.3m, tiene un volumen máximo operativo de 36.8 millones de metros cúbicos,
correspondiente a una cota de 2539 m.s.n.m. El sistema lagunar es el principal componente del
sistema hidrográfico de esta región y se considera una reserva ecológica, económica, social y
cultural de importancia nacional (CAR 2000, CONPES 2006).
En el sistema lagunar de Fúquene, ubicado en el valle de Úbate, se han realizado
transformaciones a diferentes escalas espaciales y temporales, todas ellas encaminadas a la
desecación de la laguna y adecuación de tierras para usos económicos, con un alto impacto
negativo sobre el ecosistema. En la actualidad la cuenca del río Ubaté se caracteriza por
evidenciar índices de escasez y vulnerabilidad del recurso hídrico. En el valle del río Ubaté, en
épocas de verano los caudales se reducen y en épocas de invierno los mismos aumentan
produciendo inundaciones, dada la baja capacidad de drenaje, producto de las condiciones
naturales del terreno. El marcado déficit hídrico que en épocas de verano llega en algunas áreas a
15.85 millones de m3/año, está asociado principalmente a la baja capacidad de regulación del
recurso hídrico en la cuenca (CONPES 2006).
Ningún diagnóstico puede pasar por alto la decisión del Estado y de los particulares de
desarrollar obras de desecación iniciadas desde el año 1822. A partir de este año todos los
esfuerzos se encaminaron a optimizar sistemas de drenaje que garantizaran la ocupación de
nuevas áreas de terreno circundante a la Laguna de Fúquene para su aprovechamiento
económico. El primer registro cartográfico que da cuenta de este proceso de desecación,
corresponde al año 1933, cuyo plano se presenta a continuación, elaborado para el Ministerio de
Industria y Trabajo, por el Jefe de la sección de minas y petróleo de la época, Ingeniero E. Santos
Potes (CONPES 2006).
La mayoría de las quebradas y drenajes son interceptados por el canal perimetral sin permitir el
aporte natural de aguas de calidad aceptable a la Laguna de Fúquene, lo que permite pensar en
adecuaciones hidráulicas y propuestas de operación, para lograr nuevamente que dichos caudales
transiten por la Laguna de Fúquene. Esta labor implica modificar la estructura del canal

32
perimetral para dar paso a un jarillón perimetral y la conformación de orillas, sin poner en riesgo
suelos productivos en sitios donde hoy se permite el desarrollo agropecuario (CONPES 2006).
4.1.1 Usos e importancia económica de la laguna
Los recursos hídricos superficiales de la cuenca del río Ubaté – Suárez, son utilizados para el
abastecimiento de áreas de irrigación y en una menor proporción, para el consumo municipal y la
ganadería (CONPES 2006).
El consumo de agua para ganadería, se estima en 0.06 m3/s. El agua proviene del sistema de
canales y drenajes del distrito de riego, la demanda total de agua para los acueductos municipales
se calcula en 10.8 millones de m3/año e involucra el uso doméstico, institucional e industrial,
además incluye las áreas urbanas y algunas rurales, abastecidas con el sistema de acueducto
municipal. El resto de áreas se abastecen con acueductos veredales. En total la demanda de agua
se estima en 127 millones de m3/año (Tabla 2) (CONPES 2006).
Los principales cultivos en el área irrigada corresponden a pastos, trigo, cebada, maíz y tomate.
La demanda total de agua superficial en los campos de cultivo, en la totalidad de los bloques
irrigados, se estima en 4.03 m3/s, correspondientes a 127 millones de m
3/año (Tabla 2).
Tabla 2. Demanda hídrica anual de la cuenca de los ríos Ubaté y Suárez
USO m3/s VOLUMEN ANUAL (m
3)
Agrícola 3.39 106907040
Pecuario 0.06 1892160
Doméstico 0.34 10816848
Industrial 0.24 7568640
Total 4.03 127184688 (CONPES 2006).
El sistema hidrográfico del Valle de Ubaté y Chiquinquirá, contiene tres secciones: alta, media y
baja. La parte alta está conformada al sur desde el nacimiento del río Ubaté, uniéndose aguas
abajo con otros tributarios mayores como los ríos Suta y Lenguazaque, y drenajes provenientes

33
de las lagunas de Cucunubá y Palacio, hasta descargar al norte, en la laguna de Fúquene
(CONPES 2006).
La construcción del canal perimetral en la laguna de Fúquene ha generado una disminución del
agua que entra a la laguna ya que los canales son interceptados por dicho canal, convirtiéndose
en un ecosistema que acumula altas concentraciones de nutrientes que generan una de las
principales problemáticas, la proliferación de malezas acuáticas como el buchón y la elodea
(CONPES 2006).
5 Metodología
La metodología se divide en 4 partes, la primera es una revisión de bibliografía sobre el buchón,
sistemas de información geográfica, especies invasoras y autómatas celulares. Luego se generó la
cartografía de las coberturas a partir de las fotografías aéreas obtenidas en el IGAC.
Posteriormente se realizó un programa de computador que simula el crecimiento del buchón y la
pérdida del espejo de agua de la laguna de Fúquene. Finalmente el modelo se validó contra las
fotografías y se obtuvieron los resultados que constan de las imágenes simuladas, el área de
buchón para cada año y las reglas de modelo, el cambio en el tiempo ∆t (delta de t); con su
respectivo análisis (Figura 12).

34
Figura 12. Diagrama de flujo indicando la metodología que se utilizó en la investigación.
Invasión del Buchón
Revisión de bibliografía
Buchón SIGAutómatas Celulares
Laguna de fuquene
Análisis del Paisaje
Identificación de cobertura
Modelo de Automatas
Ajuste de la dimensión de la
grilla
Ajuste de la dimensión de la
celda
Ajuste de las condiciones
iniciales
Ajuste de reglas
Identificación de la duracion de los pasos de tiempo
Interpretación de fotografías (1995 1956, 1978,1982, 1983, 1989, 1992
y 2007)
ValidacionNo aceptable ResultadosAceptable
Analisis
Mapas de cobertura
Prediccion
A continuación se describirá detalladamente la metodología utilizada en el proyecto y que se
muestra en la Figura 12:

35
La revisión bibliográfica se realizó en las bases de datos Science Direct (Elsevier), ISI Web of
Science, Proquest, JSTOR (Journal Storage), ProQuest (Todas las bases), principalmente.
También se consultó ―Google books‖ y ―Google scholar‖. La búsqueda incluyó artículos sobre el
buchón, autómatas celulares, sistemas de información geográfica y especies invasoras. Cuando
se encontraba un artículo pertinente se leía, resumía y se seleccionaban nuevas referencias de su
bibliografía. Se seleccionaron en total 114 artículos los cuales se indexaron en Endnote x4.
5.1 Caracterización de la base empírica.
Como fuente de información para caracterizar el crecimiento del buchón y la pérdida del espejo
de agua se indagaron imágenes producidas por sensores remotos: imágenes satelitales de radar y
de falso color y fotografías aéreas, seleccionándose estas últimas por tener la mejor resolución y
periodicidad de la zona de estudio; además de ser más económicas. Las fotografías se obtuvieron
de la base de datos del Instituto Geográfico Agustín Codazzi, se revisaron los vuelos fotográficos
ubicados sobre la laguna de Fúquene y se seleccionaron las fotografías en las que aparecía la
laguna total o parcialmente. En el Anexo 1 hay un listado de las fotografías seleccionadas, la
fecha a la que corresponden y la escala en las que fueron tomadas.
Cada una de las fotografías se georreferenció utilizando como base una imagen satelital de 2007
que tenía las coordenadas del área. En algunos casos la laguna se observaba en una sola
fotografía, en otros casos se necesitaban varias fotografías para conformar la imagen completa de
la laguna, así que se unieron las fotografías que correspondían al mismo año para este fin, esto se
realizó con el programa ENVI 4.3. De esta manera se obtuvo una imagen completa de la laguna
para los años 1955, 1956, 1978, 1982, 1983, 1992 y 2007 (Figura 13).

36
Figura 13. Fotografías aéreas con sus respectivos años
1955
1956
1978
1982
1983
1989
1992
2003
2007
Fuente: IGAC, 2009.

37
Posteriormente, se procedió a modelar espacialmente la distribución del buchón teniendo en
cuenta los elementos propuestos por Pearson (2007):
Un mapa raster formado por una grilla de celdas a una resolución determinada, que
representa el área de estudio modelada.
La variable independiente que corresponde a la distribución conocida de la especie.
Un conjunto de variables ambientales colectadas para caracterizar cada celda. En este
caso se utilizó la profundidad y el porcentaje de buchón de las celdas vecinas.
La probabilidad de encontrar un buchón en una celda es una función de las variables que
se encuentran en la celda, y el porcentaje de buchón de las celdas vecinas.
Para delimitar y cuantificar la distribución espacial del buchón se interpretaron las imágenes y
se identificaron las zonas de cada imagen en las que se encontraba cada cobertura: agua, buchón
y tierra firme. En este punto se obtuvieron los mapas de coberturas de la laguna para cada año.
Posteriormente se generaron mapas con polilíneas utilizando el programa ArcMap 9.3 y se
convirtieron a raster con tamaño de pixel de 25m x 25m, para luego ser importados por el
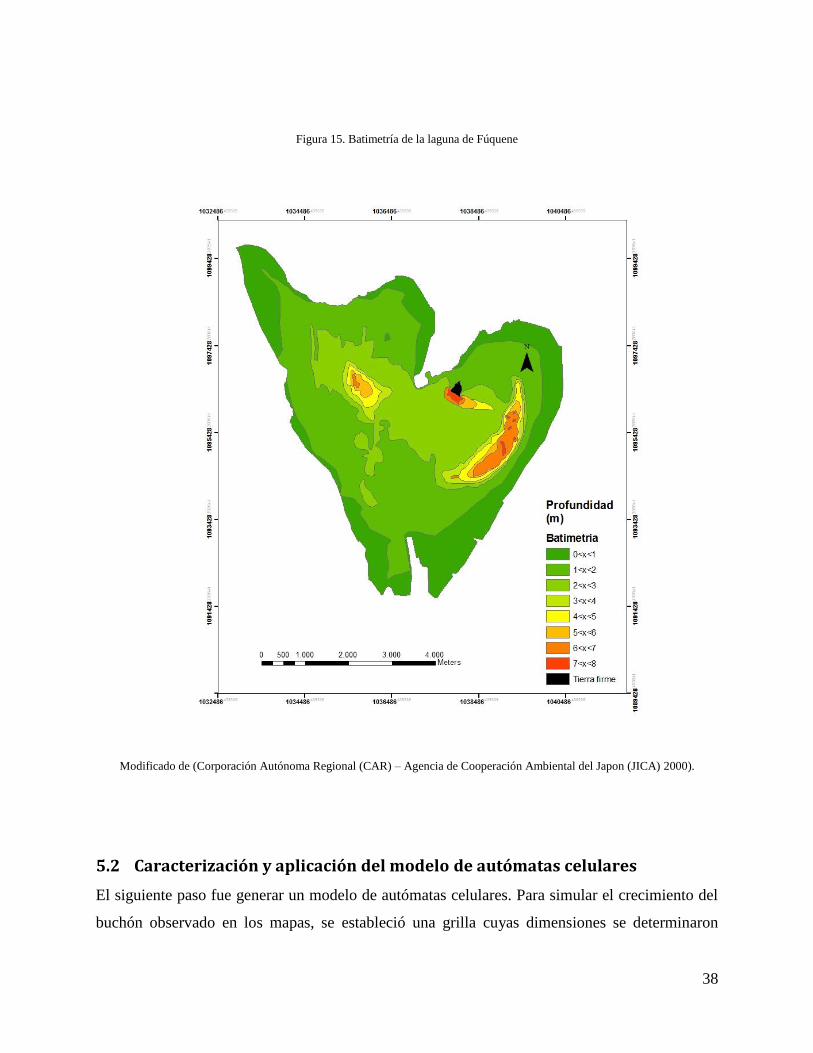
modelo del que se va a hablar más adelante. Se digitalizó una imagen raster con la batimetría
para incorporarla como insumo del modelo (Figura 15).
Figura 14. Proceso de interpretación de imágenes y generación de mapas
Fotografía aérea
Polilíneas sobre la fotografía aérea
Mapa raster
38
Figura 15. Batimetría de la laguna de Fúquene
Modificado de (Corporación Autónoma Regional (CAR) – Agencia de Cooperación Ambiental del Japon (JICA) 2000).
5.2 Caracterización y aplicación del modelo de autómatas celulares
El siguiente paso fue generar un modelo de autómatas celulares. Para simular el crecimiento del
buchón observado en los mapas, se estableció una grilla cuyas dimensiones se determinaron

39
teniendo en cuenta la escala y la resolución de las fotografías y el margen de error de la
georreferenciación. La laguna tiene unas dimensiones de 8150m de norte a sur y 7550m de
oriente a occidente, lo que corresponde a una matriz de celdas cuadradas de 25m de lado de 326
filas x 302 columnas.
Después de tener bien definidas las dimensiones de la grilla y el tamaño de las celdas, se definió
la vecindad, tomando como criterio la vecindad de Von Neumann: para cualquier celda
denotada ; en una grilla de dos dimensiones, los vecinos son definidos según la
Ecuación 2. (Figura 16).
Figura 16. Vecindad de Von Neumann.
Los vecinos se encuentran en cuatro direcciones, arriba, abajo, derecha e izquierda.
Ecuación 2 . Vecindad de Von Neumann
( ) { | | | | } { }
Luego se asignaron las condiciones iniciales, para esto se utilizó el mapa raster del primer año:
1955. Dado que los autómatas celulares utilizan periodos de tiempo discretos, es necesario
definir cuánto dura cada paso de tiempo ( ) en el que van a cambiar los valores de cada celda;
en este caso particular el corresponde a un año. Esto se asignó de manera arbitraria, pero los
parámetros se ajustaron teniendo en cuenta este valor y así se obtuvieron las tasas intrínsecas de
crecimiento anual r y de contagio.

40
En éste estudio se determinaron las reglas para las celdas que limitan con el borde de la laguna,
se estableció que las celdas del borde eran fijas y siempre presentaban buchón, ya que por las
corrientes y los vientos cualquier segmento de planta que se desprende termina en el borde y
sirve de semilla para el crecimiento del buchón.
Luego, se establecieron las reglas que definen el estado de una celda después de un paso de
tiempo (Tabla 3).
Tabla 3. Reglas del autómata celular
Condición si: Condición en t+1
La celda está vacía y los vecinos están vacíos. La celda permanece vacía.
La celda está vacía y algún vecino está lleno de
buchón.
La celda se contagia de una cantidad de
buchón en función de la profundidad.
La celda está en contacto con el borde. La celda se contagia de una cantidad de
buchón en función de la profundidad.
La celda está ocupada. La población crece siguiendo el modelo
logístico, en el cual el r es función de la
profundidad.
Después de tener definidos el tamaño de la celda, el tamaño de la grilla, las condiciones iniciales
y las reglas; se utilizó el algoritmo de optimización de Solver V10.0 para ajustar la tasa de
intrínseca de crecimiento r y la tasa de contagio a el , de esta manera se encontraron las tasas
que mejor se ajustaban a las características de crecimiento del buchón en la laguna de Fúquene.
Sin embargo no se logró acoplar el Solver con el modelo para parametrizarlo, así que se utilizó
un algoritmo que a partir de prueba y error, generando números aleatorios, seleccionaba los
mejores parámetros después de 100.000 iteraciones.
Para validar el modelo se recurrió a un coeficiente de similitud simple (Ecuación 3) y una
función objetivo (Ecuación 4). La tabla de contingencia da el porcentaje de similitudes entre dos
mapas, una simulada y una observada; se usa para ver qué tipo de errores son cometidos (Tabla
4) (Krebs 1989, Pearson 2007, WWRP/WGNE Joint Working Group on Verification 2009). Una

41
predicción perfecta produce sólo verdaderos positivos y verdaderos negativos, y no producen ni
falsos positivos ni falsos negativos.
Ecuación 3. Coeficiente de similitud simple.
Ecuación 4. Función objetivo:
∑
Tabla 4. Tabla de contingencia
Observado
Presencia de buchón Ausencia de buchón
Simulado
Presencia de buchón verdadero positivo falsos positivos
Ausencia de buchón falsos negativos d verdadero negativo
Se compararon cada una de las ocho fotografías observadas con las simuladas, correspondientes
al mismo año, tomando cada una de las celdas de la imagen simulada con la celda análoga en la
imagen observada. Se contabilizó el número de verdaderos positivos denotados por , número de
falsos positivos , número de falsos negativos y número de verdaderos negativos . Un
verdadero positivo es cuando una celda específica del mapa raster, de un año determinado (
observada), registra la presencia del buchón y la celda correspondiente a la misma posición del
mapa simulado para el mismo año ( simulada) también presenta buchón. Un falso positivo
ocurre cuando la celda simulada presenta buchón y la observada no. Un falso negativo ocurre
cuando la celda simulada no presenta buchón y la celda observada si, y un verdadero negativo
ocurre cuando ninguna de las dos presenta buchón. Así, al comparar todas las celdas de una foto

42
observada y una simulada se obtiene una cantidad de verdaderos positivos , falsos positivos b,
falsos negativos c y verdaderos negativos .
A partir de la tabla de contingencia (Tabla 4) se aplicó el coeficiente de similitud simple para
comparar las imágenes simuladas con las observadas, este coeficiente tiene valores de similitud
entre cero y uno, cero significa que las imágenes son completamente diferentes y uno que son
iguales (Ecuación 4) (Krebs 1989, Pearson 2007, WWRP/WGNE Joint Working Group on
Verification 2009). Se obtuvo un valor del coeficiente de similitud por cada par de fotografías
(simulada y observada) es decir se obtuvieron ocho valores, los que se sumaron y se dividieron
entre el número de fotografías (8) para estandarizar el error. Este procedimiento se realizó
100.000 veces, cambiando los parámetros, r y tasa de contagio, de forma aleatoria cada vez. Se
tomó el menor de los resultados y luego se graficaron los resultados de la simulación y
analizaron los resultados.
6 Resultados
6.1 Crecimiento histórico del buchón 1955-2007
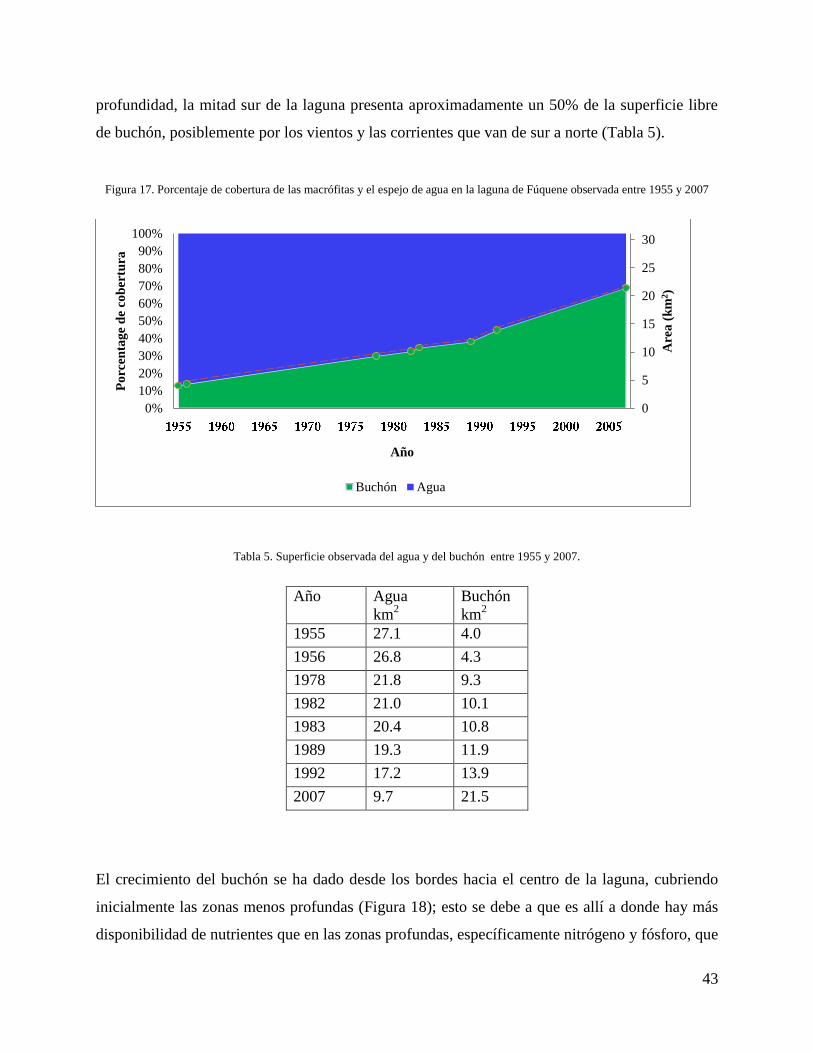
Desde el año 1955 el buchón ha tenido un crecimiento exponencial que ha cubierto
progresivamente el espejo de agua de la laguna de Fúquene, en el 2007 (año de la última foto
conseguida para la zona) dicho cubrimiento se acercó al 70% de la superficie de la laguna
(Figura 17). La suma de la superficie de agua y buchón para cada año siempre suman 31.1 km2,
correspondientes al área de la laguna. El área cubierta por el buchón ha pasado de 4 km2 en 1955
a 21.4 km2 en el 2007. El crecimiento del buchón de 1955 a 1956 fue de 304.616 m
2; de 1956 a
1978 fue de 4.978.345 m2, para este momento el área con una profundidad menor a un metro
estaba invadida por el buchón. De 1978 a 1982 el crecimiento fue de 815.710 m2; de 1982 a 1983
fue de 653.295 m2; de 1983 a 1989 fue de 1.096.759m
2; de 1989 a 1992 de 2.053.897 m
2, en este
periodo el cuarto noroccidental de la laguna con profundidades menores a dos metros se
encontraba totalmente cubierta de buchón, esto, posiblemente, obedece a la dirección de la
corriente, ya que el desfogue de la laguna está localizado en este sector. El crecimiento del
buchón entre los años 1992 y 2007 fue de 7.521.277 m2, en este punto la mitad norte de la laguna
se encuentra completamente cubierta, excepto los sectores con más de tres metros de
43
profundidad, la mitad sur de la laguna presenta aproximadamente un 50% de la superficie libre
de buchón, posiblemente por los vientos y las corrientes que van de sur a norte (Tabla 5).
Figura 17. Porcentaje de cobertura de las macrófitas y el espejo de agua en la laguna de Fúquene observada entre 1955 y 2007
Tabla 5. Superficie observada del agua y del buchón entre 1955 y 2007.
Año Agua
km2
Buchón
km2
1955 27.1 4.0
1956 26.8 4.3
1978 21.8 9.3
1982 21.0 10.1
1983 20.4 10.8
1989 19.3 11.9
1992 17.2 13.9
2007 9.7 21.5
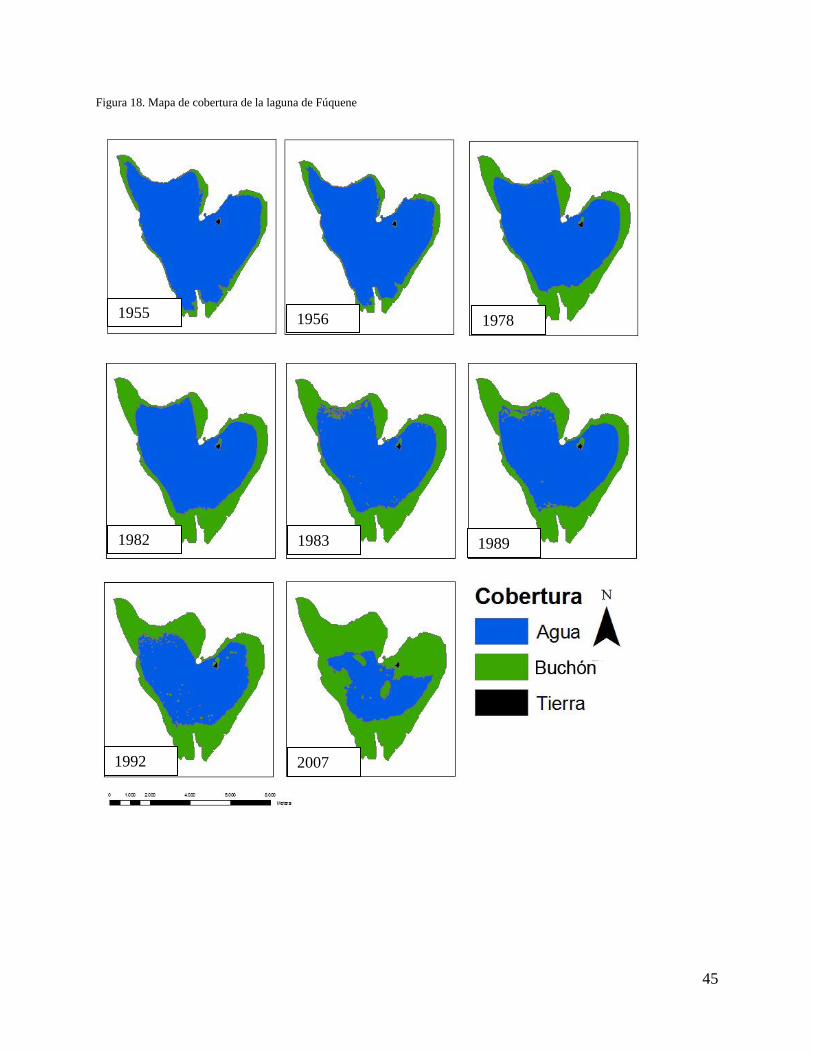
El crecimiento del buchón se ha dado desde los bordes hacia el centro de la laguna, cubriendo
inicialmente las zonas menos profundas (Figura 18); esto se debe a que es allí a donde hay más
disponibilidad de nutrientes que en las zonas profundas, específicamente nitrógeno y fósforo, que
0
5
10
15
20
25
30
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Are
a (
km
2)
Po
rcen
tag
e d
e co
ber
tura
Año
Buchón Agua

44
son los principales recursos de los que depende el crecimiento de la macrófitas. El mismo patrón
es notorio en el costado norte de la isla, que es más somero que el costado sur. Se hace posible
observar dicho comportamiento al sobreponer los mapas de la cobertura y la batimetría de la
laguna (Figura 15). De esta manera, en los sectores con profundidades entre cuatro y ocho
metros no se ha establecido el buchón; a diferencia de los sectores con profundidades entre los
cero y cuatro metros (Figura 15).
45
Figura 18. Mapa de cobertura de la laguna de Fúquene
1955 1956
1983
2007 1992
1989 1982
1978

46
6.2 Descripción del modelo
Para simular el crecimiento y distribución del buchón en la laguna de Fúquene de manera
espacialmente explícita, se realizó el programa basado en el algoritmo representado en el
diagrama de flujo de la Figura 19.
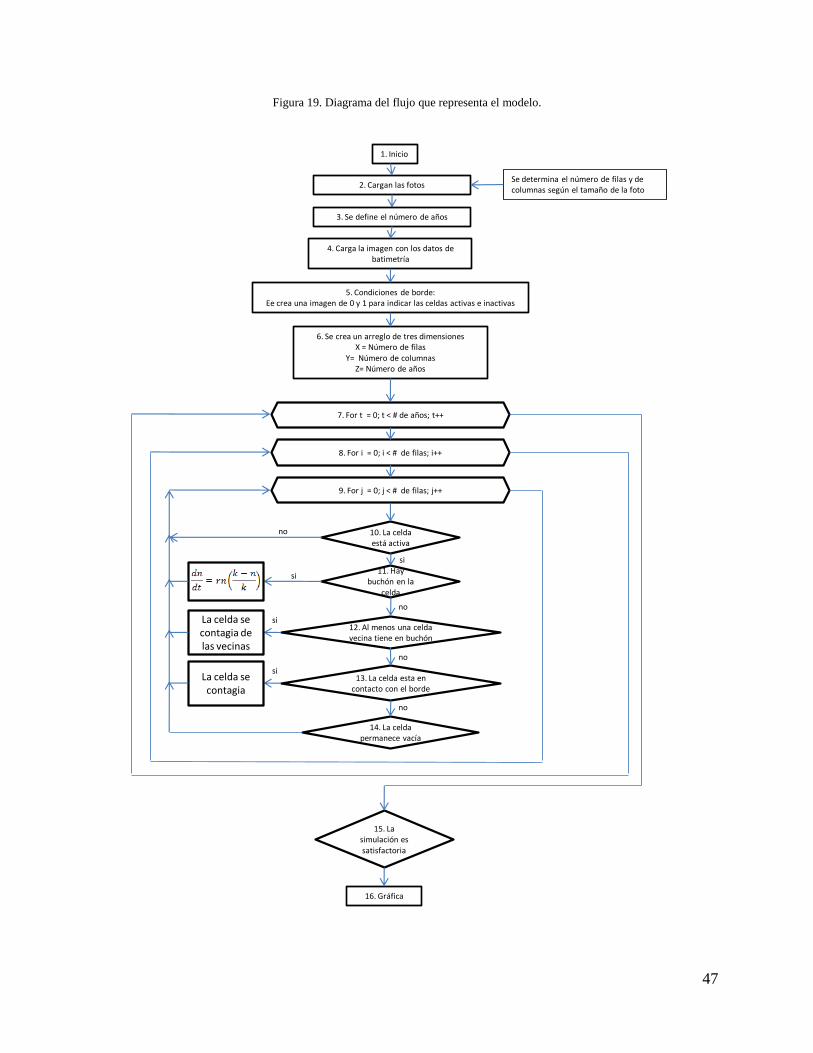
47
Figura 19. Diagrama del flujo que representa el modelo.
1. Inicio
2. Cargan las fotosSe determina el número de filas y de columnas según el tamaño de la foto
4. Carga la imagen con los datos de batimetría
5. Condiciones de borde: Ee crea una imagen de 0 y 1 para indicar las celdas activas e inactivas
6. Se crea un arreglo de tres dimensionesX = Número de filas
Y= Número de columnasZ= Número de años
7. For t = 0; t < # de años; t++
8. For i = 0; i < # de filas; i++
9. For j = 0; j < # de filas; j++
10. La celda está activa
11. Hay buchón en la
celda
12. Al menos una celda vecina tiene en buchón
14. La celda permanece vacía
15. La simulación es satisfactoria
16. Gráfica
3. Se define el número de años
13. La celda esta en contacto con el borde
si
si
si
si
no
no
no
no
La celda se contagia de las vecinas
La celda se contagia

48
Inicialmente, se cargaron las imágenes raster generadas en ArcMap 9.3 que representan la laguna
entre 1955 y 2007 (Figura 18), la imagen de la batimetría (Figura 15) y una imagen raster
compuesta de unos y ceros que determina el área de la laguna y las islas; luego se definieron el
número de años a simular (paso 2-4 Figura 19).
Posteriormente se identificaron las dimensiones de la grilla y de las celdas, éstas estaban
estrechamente relacionadas con la escala y la resolución a la que se estudió el proceso. Se utilizó
una grilla de 302 columnas y 326 filas, con celdas cuadradas de 25m de lado cada una; esto
correspondió a un área de 7550 x 8150m representado por 98.452 celdas. De las cuales 49.818
corresponden a celdas activas, es decir que son evaluadas en cada paso de tiempo; y 48.634 son
celdas inactivas, que representan tierra firme y en ellas no crece el buchón, por lo que su valor es
constante. Las celdas activas, que son las que corresponden al área de la laguna, pueden tener
valores entre 0% y 100%; donde 0 (cero) indica que la celda está libre de buchón y el resto de los
valores indican el porcentaje de ocupación de la celda por el buchón. Posteriormente, se creó un
arreglo de tres dimensiones (i, j, t) donde i y j representan las filas y las columnas,
respectivamente, y t el tiempo (paso 6 Figura 19).
En este punto se asignaron los valores de r para la ecuación logística y el de la tasa de contagio
para el autómata celular, esto para cada una de las profundidades de cada celda (paso 7 Figura
19).
Para cada paso de tiempo t se recorre cada una de las celdas en la posición (i, j) (pasos 8-10 de la
Figura 19). En cada celda interactúan dos modelos acoplados; el primero es un modelo logístico
que habita en cada una de las celdas, este modelo representa el crecimiento del buchón; el otro es
un modelo de autómatas celulares que determina la tasa de contagio entre las celdas, que
representa la dispersión del buchón en la laguna. El modelo logístico está definido por la
Ecuación 5, en donde es la tasa intrínseca de crecimiento anual, es el porcentaje de
ocupación de la celda por el buchón y % correspondiente a la capacidad de carga de la
celda, dada en porcentaje de superficie ocupada. La tasa intrínseca de crecimiento es función
de la profundidad de la laguna en la zona donde se encuentra la celda (pasos 11-12 Figura 19).

49
Ecuación 5. Modelo de crecimiento logístico de Lotka - Voltera
(
)
En el modelo de autómatas celulares la llegada del buchón a una celda en un periodo es función
de la profundidad, el valor de dicha celda, y de las celdas vecinas en el tiempo anterior (paso 13-
15 Figura 19, Ecuación 6). Si la celda está en contacto con el borde se contagiará ya que se
asume que el borde es una fuente de contagio (Ecuación 6).
Ecuación 6. Variables que determinan el valor de la celda
{
}
A continuación, el algoritmo calcula el coeficiente de similitud simple (Ecuación 4) para
comparar las imágenes simuladas con las observadas. Luego aplica la función objetivo (Ecuación
4), este proceso se realiza 100.000 veces y se toman los valores de los parámetros que
corresponden a la simulación que mejor se ajusta a los datos observados (pasos 7-15 Figura 19).
Si sí se parecen, se gráfica (Figura 20) y se da por terminado el algoritmo.

50
Figura 20. Resultados gráficos de modelo desde 1955 hasta 2007
1955
1962
1963
1970
1971
1978
1979
1986
1987
1994
1995
2001
2002
2007

51
7 Discusión
El principal objetivo de este trabajo fue generar un modelo de autómatas celulares que permitió
explorar el crecimiento del buchón y las dinámicas del proceso de invasión en la laguna de
Fúquene, incorporando las siguientes variables: la profundidad del cuerpo de agua, la presencia o
ausencia de buchón, la tasa de crecimiento y de dispersión del buchón y la forma la laguna.
El modelo reprodujo el crecimiento del buchón acorde con la teoría que plantea que el buchón
crece de los bordes hacia el centro (Masifwa et al. 2001). En la validación del modelo al
comparar las imágenes simuladas con las observadas (Figura 21), el índice de similitud usando la
función objetivo (Ecuación 4), correspondió a un porcentaje promedio histórico del 76%. Se
observa que el buchón modelado tiene un comportamiento similar al buchón observado. El
centro de la laguna y los bordes fueron correctamente predichos, en los bordes siempre se
encuentran verdaderos positivos y en el centro se observan verdaderos negativos. No obstante, en
las zonas del límite entre el buchón y el agua se presentaron en su mayoría falsos positivos. En la
simulación del año 2007 fue notoria la aparición de falsos negativos, causados por una isla de
buchón ubicada en la zona central de la laguna (Figura 21).
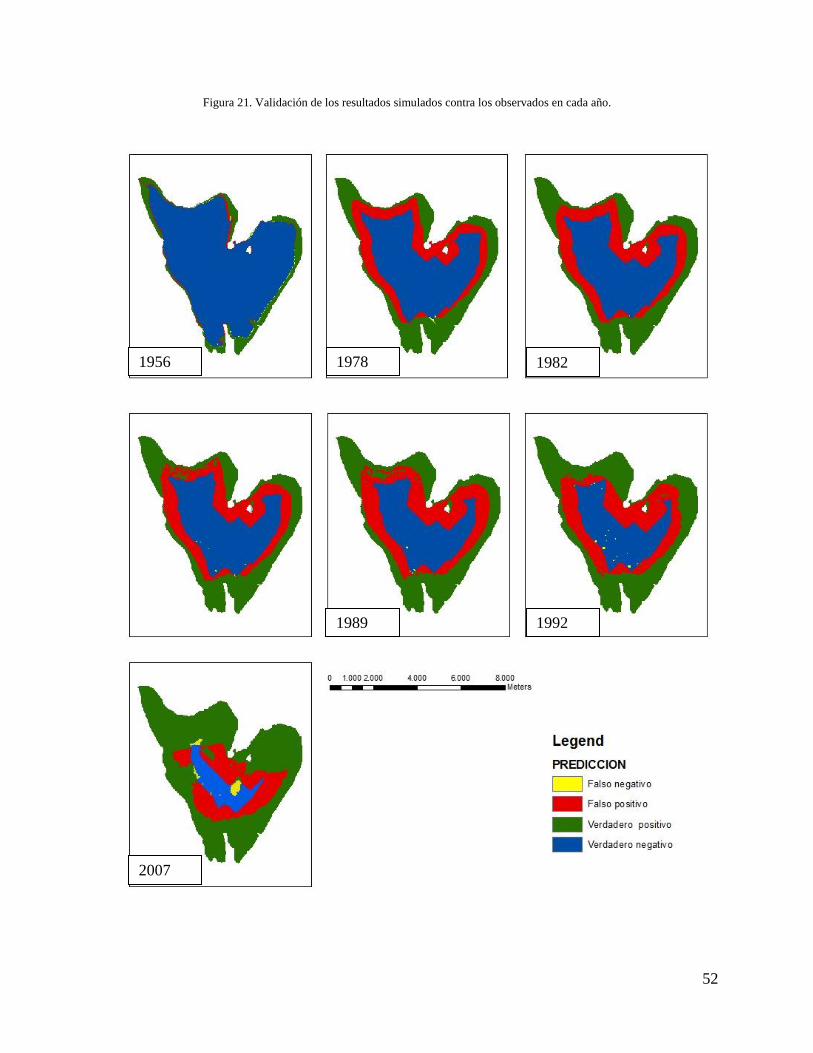
52
Figura 21. Validación de los resultados simulados contra los observados en cada año.
1992 1989
2007
1982 1978 1956

53
Si se toma la última fotografía, correspondiente al año 2007, como condición para la simulación,
se obtiene que para aproximadamente el año 2035 el buchón cubrirá totalmente el espejo de agua
siguiendo un patrón de crecimiento logístico hasta alcanzar la capacidad de carga, es decir el
cubrimiento del espejo de agua de la laguna (Figura 22). Por esta razón es indispensable tomar
acciones correctivas, para controlar el crecimiento del buchón y evitar que la laguna de Fúquene
sufra el mismo destino que la laguna de Palacio la cual perdió su espejo de agua, ya no es
navegable y murieron todos sus peces por desoxigenación (CAR 2000).
El uso de AC trae gran número de beneficios a los estudios de la dinámica de la vegetación, pero
la correcta implementación de los resultados requiere tener en cuenta las limitaciones, ya que los
AC deben ser entendidos como una herramienta más y no como el remplazo de las herramientas
tradicionales como ecuaciones de derivadas parciales, ecuaciones diferenciales ordinarias y
modelos conceptuales (Colasanti et al. 2007).
El modelo tiene limitaciones espaciales y temporales. En la presente investigación al trabajar a
una escala intermedia, no se tuvieron en cuenta las alteraciones producidas a mayor escala como
la incorporación de la productividad bajo los efectos de la competencia inter e intraespecífica, el
incremento modular, el incremento de biomasa de cada individuo etc. Por otro lado, al trabajar a
una menor escala se podrían estudiar el proceso de dispersión del buchón entre diferentes
cuerpos de agua, a una escala departamental, nacional, continental o global.
En cuanto a la escala temporal este modelo maneja un cambio en el tiempo mínimo de un año,
por esta razón no se pueden observar alteraciones diarias ni mensuales, así como los cambios
estacionales que ocurren durante un año.
A pesar de estas limitaciones al utilizar una escala que abarcara la laguna con un tamaño de celda
de 25m de lado, el modelo se ajustó satisfactoriamente a los resultados observados y esperados;
logró reproducir el crecimiento del buchón como se observa en la naturaleza: desde los bordes de
la laguna hacia el centro, y reprodujo el crecimiento del buchón de una manera logística de
acuerdo con las teorías existente (Begon et al. 1996).

54
Figura 22. Pronostico del crecimiento del buchón entre los años 2010 y 2035
Otra de las limitantes del modelo es que no tiene en cuenta fenómenos estocásticos como
tormentas, inundaciones, heladas, que pueden provocar altos índices de mortalidad del buchón, y
pueden incrementar los errores de la simulación.
Para lograr una buena modelación, se deben identificar las variables a las que el fenómeno
estudiado es más sensible y que respondan a la física y la biología del proceso estudiado. Sin
embargo, uno de los inconvenientes limitantes más importantes para realizar modelos, es la falta
de disponibilidad y la mala calidad de los datos. Lamentablemente en Colombia la disponibilidad
de la mayoría de los datos biológicos y físico-químicos es muy limitada debido a los pocos
2010 2015 2020
2035 2030 2025

55
recursos de investigación y monitoreo y a la falta de constancia al realizar las mediciones.
Sumado a esto, los datos existentes están dispersos en estudios de difícil acceso y en muchos
casos a pesar de ser públicos son muy costosos. Por estas razones no se pudo acceder a datos
como la concentración del nitrógeno y el fósforo en diferentes sectores de la laguna, relevantes
para estimar las tasas de crecimiento del buchón (Bock 1969). Tampoco se tuvo en cuenta la
intensidad y dirección del viento que transporta las islas de buchón de un lado a otro de la laguna
dependiendo la época del año.
Por las razones mencionadas, es indispensable aumentar los esfuerzos para generar bases de
datos centralizadas de fácil acceso y bajo costo, que aumentarían considerablemente la calidad y
la cantidad de las investigaciones realizadas en el país.
La característica más destacable de este trabajo fue la validación espacialmente explícita de los
resultados del modelo contra los mapas obtenidos de las fotografías históricas. De esta manera se
tiene certeza del margen de error del modelo y de las zonas de la laguna en las que este ocurrió.
Lo anterior diferencia a este trabajo de muchos otros estudios de modelación ecológica que no
incluyen una validación debido a que son trabajos teóricos y que no poseen datos empíricos de
alguna especie en particular como es el caso de Aassine y Jaï (2002) que realizaron un modelo
teórico de la evolución espacio-temporal de la propagación de especies vegetales. Arii y Parrott
(2006) que estudiaron el proceso de colonización y las habilidades de competencia de especies
exóticas invasivas. Otros trabajos de modelación de autómatas celulares si utilizaron datos
empíricos, pero no lograron hacer validaciones debido a que no disponían de información
histórica de las especies con que trabajaron Balzter et al. (1998) crearon un modelo aglomerado
del crecimiento poblacional de tres especies vegetales Glechoma hederácea, Lolium perenne y
Trifolium repens, con autómatas celulares y cadenas de Markov. Ellos validaron el modelo y los
resultados se ajustaron correctamente a Glechoma hederácea mas no para las otras dos especies.
Sebert-Cuvillier (2009) realizaron un modelo estocástico, espacialmente explícito de la
demografía de la planta Prunus serotina.

56
8 Conclusiones y recomendaciones
Los autómatas celulares son una herramienta útil para predecir y simular la dinámica de los
procesos, en los que la distribución espacial juega un papel relevante, como es el caso de la
distribución de las especies; mas no quiere decir que los autómatas celulares vayan a remplazar a
todas las otras herramientas de la modelación, el mundo es muy complejo y se necesita de todas
las herramientas posibles para aproximarse cada vez más al entendimiento y la predicción de los
procesos y las dinámicas de la naturaleza.
La definición de las vecindades usadas en un AC debe ser tomada con extrema cautela, ya que
puede alterar drásticamente los resultados de la simulación. En esta investigación, después de
probar con las vecindades de Moore, Moore extendido y Von Neumann, la regla de las
vecindades que mejor se ajustó al crecimiento del buchón fue la de Von Neumann (Balzter et al.
1998, Adamatzky et al. 2008), esta última utilizada en la simulación final por ser esta la que más
se acercó a las fotografías observadas.
La integración de autómatas celulares con modelos de EDO como el logístico Lotka-Voltera, ha
mostrado ser una buena forma de modelar procesos de expansión de las especies invasoras.
El uso de AC trae gran número de beneficios a los estudios de la dinámica de vegetación, pero la
correcta implementación de sus resultados requiere un entendimiento de sus limitaciones. Los
AC deben ser entendidos como una herramienta más y no como el remplazo de las herramientas
tradicionales.
Los AC permiten investigar procesos emergentes que son atribuidos al comportamiento colectivo
del sistema pero que se originan desde una descripción a nivel individual, por ejemplo la
resiliencia, la capacidad de invadir, biodiversidad, etc.

57
Se recomienda implementar el modelo en diversos cuerpos de agua con diferentes condiciones
para mejorar el entendimiento del proceso de pérdida del espejo de agua, probar con diferentes
tamaños de celda e involucrar variables ambientales como gradientes de nutrientes,
principalmente nitrógeno y fosforo.
La profundidad es una variable relevante para determinar la tasa de crecimiento del buchón y las
sucesiones naturales de plantas en los medios acuáticos, ya que la concentración de los nutrientes
de los que dependen estas plantas son función de la profundidad, debido a las tasas de
sedimentación e intercambio de los nutrientes con el fondo del lago (Johnston 1991).
Como estudios complementarios se recomienda estudiar el cambio histórico de la
evapotranspiración en un cuerpo de agua mientras que está siendo cubierto por macrófitas como
el buchón, y se puede tomar como insumo los resultados del modelo del crecimiento del buchón.
Si se desea hacer extracción de buchón de la laguna de Fúquene, teniendo en cuenta los
resultados del modelo y la simulación, entonces la mejor forma sería desde el centro de la laguna
hacia los bordes y así no se aumenta la frontera de crecimiento del buchón. Si la extracción se
realiza desde los bordes de la laguna se generan nuevas zonas de crecimiento y el buchón se
reproducirá más rápido y cubrirá más superficie de agua, por lo tanto el tiempo de poblamiento
de la laguna sería el mismo que si no se hiciera extracción.
Dada la complejidad de los procesos naturales, y con el ánimo de comprender sus dinámicas, es
necesaria la implementación de protocolos para tomar mediciones estandarizadas; de manera que
estén agrupadas en bases de datos de fácil acceso a la comunidad científica y faciliten el
desarrollo de modelos que se ajusten a la realidad.
En la laguna de Fúquene se encuentran cerca de 15 especies de micrófitos las cuales están
compitiendo entre sí y contribuyen al proceso de perdida de del espejo de agua. Sin embargo este
modelo no tiene en cuenta todas estas interacciones. Por esta razón se recomienda que en futuros
estudios se involucren estas especies y la dinámica de la competencia interespecifica.

58
9 Anexos
Los otros anexos se entregan encuentran en el CD:
1. El software realizado para modelar el crecimiento del buchón en la laguna de Fúquene.
2. El código de java en el que se desarrolló el software.
Anexo 1. Fotografías aéreas de la laguna de Fúquene IGAC
VUELO FECHA SOBRE FOTO Escala
M45 27 de enero de 1955 143A 4183 1:60000
M52 20 de enero de 1956 167 5040 1:60000
M52 20 de enero de 1956 167 5041 1:60000
c1054 21 de febrero de 1962 22189 234 1:20000
c1054 21 de febrero de 1962 22189 236 1:20000
c1054 21 de febrero de 1962 22185 120 1:20000
c1054 21 de febrero de 1962 22185 122 1:20000
c1058 13 de marzo de 1962 22217 1266 1:20000
c1058 13 de marzo de 1962 22217 1267 1:20000
C1822 2 de febrero de 1978 29294 350 1:26800
C1822 2 de febrero de 1978 29294 352 1:26800
C2050 20 de enero de 1982 31412B 50 1:19500
C2050 20 de enero de 1982 31412B 52 1:19500
C2050 20 de enero de 1982 31412B 54 1:19500
C2051 20 de enero de 1982 31416B 29 1:18800
C2051 20 de enero de 1982 31416B 31 1:18800
C2051 20 de enero de 1982 31416B 32 1:18800
C2064 15 de octubre de 1982 31545 168 1:25700
C2064 15 de octubre de 1982 31545 170 1:25700
C2072 9 de enero de 1983 31654 99 1:20320
C2072 9 de enero de 1983 31654 101 1:20320
C2072 9 de enero de 1983 31655 118 1:19700
C2072 9 de enero de 1983 31655 120 1:19700
C2072 9 de enero de 1983 31653 84 1:20320
C2072 9 de enero de 1983 31653 85 1:20320
C2378 16 de noviembre de 1989 35131 35 1:50000
C2378 16 de noviembre de 1989 35131 36 1:50000
c2471 1 de febrero de 1992 36260 60 1:34200
59
VUELO FECHA SOBRE FOTO Escala
c2471 1 de febrero de 1992 36260 61 1:34200
c2471 1 de febrero de 1992 36260 62 1:34200
C2611 31 de enero de 1997 37690 31 1:45000
C2697 26 de octubre de 2003 39029 234 1:19250
C2697 26 de octubre de 2003 39029 235 1:19250
C2697 26 de octubre de 2003 39029 236 1:19250
C2799 1 de febrero de 2007 40774 99 1:39070
C2799 1 de febrero de 2007 40774 101 1:39071
10 Bibliografía
Aassine, S. and M. C. El Jaï. 2002. Vegetation dynamics modelling: a method for coupling local
and space dynamics. Ecological Modelling 154:237-249.
Adamatzky, A., R. Alonso-Sanz, and A. Lawniczak. 2008. Automata-2008: Theory and
Applications of Cellular Automata.
Alpert, P., E. Bone, and C. Holzapfel. 2000. Invasiveness, invasibility and the role of
environmental stress in the spread of non-native plants. Perspectives in Plant Ecology,
Evolution and Systematics 3:52-66.
Arii, K. and L. Parrott. 2006. Examining the colonization process of exotic species varying in
competitive abilities using a cellular automaton model. Ecological Modelling 199:219-
228.
Balzter, H., P. Braun, and W. Köhler. 1998. Cellular automata models for vegetation dynamics.
Ecological Modelling 107:113-125.
Barrett, S. and I. Forno. 1982. Style morph distribution in New World populations of Eichhornia
crassipes (Mart.) Solms-Laubach (water hyacinth). Aquatic Botany 13:299-306.
Begon, M., J. Harper, and C. Townsend. 1996. Ecology: individuals, populations, and
communities. Wiley-Blackwell.
Bernhardsen, T. 2002. Geographic information systems: an introduction. John Wiley & Sons Inc.
Blossey, B. and R. Notzold. 1995. Evolution of Increased Competitive Ability in Invasive
Nonindigenous Plants: A Hypothesis. The Journal of Ecology 83:887-889.
Bock, J. 1969. Productivity of the water hyacinth Eichhornia crassipes (Mart.) Solms.
Ecology:460-464.
Caicedo Carrascal, F. M. 2008. Asimilación de precipitación estimada por imágenes de satélite
en modelos hidrológicos aglutinados y distribuidos, caso de estudio afluencias al embalse
de Betania (Huila, Colombia). Pontificia Universidad Javeriana, Bogota D.C.
Cannas, S., D. Marco, S. Páez, and M. Montemurro. 2004. Modelado de invasiones biologicas:
dinámica poblacional y formación de patrones espaciales. Julio A. Hernández and Andrés
Pomi (Ed. Dirac, 2004).
CAR. 2000. Fúquene: El Lecho de la Zorra. Corporación Autónoma Regional de Cundinamarca,
Bogotá, Colombia.
Catford, J., R. Jansson, and C. Nilsson. 2009. Reducing redundancy in invasion ecology by
integrating hypotheses into a single theoretical framework. Diversity and Distributions
15:22-40.

60
Charudattan, R. 1986. Integrated control of waterhyacinth (Eichhornia crassipes) with a pathogen,
insects, and herbicides. Weed Science 34:26-30.
Chen, Q., A. E. Mynett, and A. W. Minns. 2002. Application of cellular automata to modelling
competitive growths of two underwater species Chara aspera and Potamogeton pectinatus
in Lake Veluwe. Ecological Modelling 147:253-265.
Colasanti, R., R. Hunt, and L. Watrud. 2007. A simple cellular automaton model for high-level
vegetation dynamics. Ecological Modelling 203:363-374.
CONPES. 2006. ESTRATEGIA PARA EL MANEJO AMBIENTAL DE LA CUENCA UBATÉ -
SUÁREZ.in D. N. d. Planeación, editor., Bogotá, D.C
Corporación Autónoma Regional (CAR) – Agencia de Cooperación Ambiental del Japon (JICA).
2000. Plan de mejoramiento ambiental regional para la cuenca de la laguna de Fúquene
Santa Fé de Bogotá.
Domínguez, E. 2010. Modelación matemática, Una introducción al método. Bogota.
Drake, J. and D. Lodge. 2006. Allee effects, propagule pressure and the probability of
establishment: risk analysis for biological invasions. Biological Invasions 8:365-375.
Eckert, C., K. Lui, K. Bronson, P. Corradini, and A. Bruneau. 2003. Population genetic
consequences of extreme variation in sexual and clonal reproduction in an aquatic plant.
Molecular Ecology 12:331-344.
Facon, B., B. Genton, J. Shykoff, P. Jarne, A. Estoup, and P. David. 2006. A general eco-
evolutionary framework for understanding bioinvasions. Trends in Ecology & Evolution
21:130-135.
Fischer, M., R. Husi, D. Prati, M. Peintinger, M. Kleunen, and B. Schmid. 2000. RAPD variation
among and within small and large populations of the rare clonal plant Ranunculus reptans
(Ranunculaceae). American Journal of Botany 87:1128.
Franco Vidal, L., A. Villa, and A. Sarmiento. 2007. Clasificación y estado actual de los hábitats
de humedal de las lagunas de Fúquene, Cucunubá y Palacio: Implicaciones para su
manejo in L. Franco Vidal and G. Andrade Pérez, editors. Fúquene, Cucunubá y Palacio.
Conservación de la biodiversidad y manejo sostenible de un ecosistema lagunar andino.
Fundacion Humedales e Instituto de Investigacion de Recursos Biológicos Alexander von
Humboldt Bogota D.C.
Ge, S., K. Wang, and M. Dong. 1999. Genetic diversity and clonal structure of Hedysarum laeve
in MoUs Sandland. Acta Bot. Sin 41:301–306.
Greenfield, B., G. Siemering, J. Andrews, M. Rajan, S. Andrews, and D. Spencer. 2007.
Mechanical shredding of water hyacinth (Eichhornia crassipes): effects on water quality
in the Sacramento-San Joaquin River Delta, California. Estuaries and Coasts 30:627-640.
Gronewold, A. and M. Sonnenschein. 1998. Event-based modelling of ecological systems with
asynchronous cellular automata. Ecological Modelling 108:37-52.
Hellmann, J., J. Byers, B. Bierwagen, and J. Dukes. 2008. Five potential consequences of climate
change for invasive species. Conservation Biology 22:534-543.
Higgins, S. and D. Richardson. 1996. A review of models of alien plant spread. Ecological
Modelling 87:249-265.
Instituto De Investigación De Recursos Biológicos Alexander Von Humboldt. 2004.
Caracterización biofísica, ecológica y sociocultural del complejo de humedales del Valle
de Ubaté: Fúquene, Cucunubá y Palacio. Fundacion humedales e Instituto De
Investigación De Recursos Biológicos Alexander Von Humboldt, Bogota.

61
Johnston, C. 1991. Sediment and nutrient retention by freshwater wetlands: effects on surface
water quality. Critical Reviews in Environmental Science and Technology 21:491-565.
Karafyllidis, I. and A. Thanailakis. 1997. A model for predicting forest fire spreading using
cellular automata. Ecological Modelling 99:87-97.
Kateregga, E. and T. Sterner. 2007. Indicators for an invasive species: Water hyacinths in Lake
Victoria. Ecological Indicators 7:362-370.
Keane, R. M. and M. J. Crawley. 2002. Exotic plant invasions and the enemy release hypothesis.
Trends in Ecology & Evolution 17:164-170.
Kier, L. B., P. G. Seybold, and C.-K. Cheng. 2005. Cellular automata. Pages 9-38 Cellular
automata modeling of chemical systems : a textbook and laboratory manual. Springer,
Dordrecht.
Krebs, C. 1989. Ecological methodology. Harper & Row New York.
Lonsdale, W. 1999. Global patterns of plant invasions and the concept of invasibility. Ecology
80:1522-1536.
Mailu, A. 2000. Preliminary Assessment of the Social, Economic and Environmental Impacts of
Water Hyacinth in Lake Victoria Basin and Status of Control. Pages 130-139. ACIAR;
1998.
Marco, D., S. Páez, and S. Cannas. 2002. Species invasiveness in biological invasions: a
modelling approach. Biological Invasions 4:193-205.
Martin, O., A. Odlyzko, and S. Wolfram. 1984. Algebraic properties of cellular automata.
Communications in mathematical physics 93:219-258.
Masifwa, W., T. Twongo, and P. Denny. 2001. The impact of water hyacinth, Eichhornia
crassipes (Mart) Solms on the abundance and diversity of aquatic macroinvertebrates
along the shores of northern Lake Victoria, Uganda. Hydrobiologia 452:79-88.
Ming, P. and J. Albrecht. 2004. Integrated Framework for the Simulation of Biological Invasions
in a Heterogeneous Landscape. Transactions in GIS 8:309-334.
Moles, A. T., M. A. M. Gruber, and Stephen P. Bonser. 2008. A new framework for predicting
invasive plant species. Journal of Ecology 96:13-17.
Navarro, L. and G. Phiri. 2000. Water hyacinth in Africa and the Middle East: a survey of
problems and solutions. Idrc.
Njiru, M., A. Othina, A. Getabu, D. Tweddle, and I. Cowx. 2002. Is the invasion of water
hyacinth, Eichhornia crassipes Solms(Mart.), a blessing to Lake Victoria fisheries?
Management and Ecology of Lake and Reservoir Fisheries.:255-266.
Pearson, R. G. 2007. Species' Distribution Modeling for Conservation Educators and
Practitioners. Synthesis. American Museum of Natural History. Available at
http://ncep.amnh.org.
Qu, S., Q. Chen, and F. Recknagel. 2008. Cellular automata based simulation of random versus
selective harvesting strategies in predator-prey systems. Ecological Informatics 3:252-
258.
Ren, M. X. and Q. G. Zhang. 2007. Clonal diversity and structure of the invasive aquatic plant
Eichhornia crassipes in China. Aquatic Botany 87:242-246.
Richardson, D. and P. Pysek. 2006. Plant invasions: merging the concepts of species
invasiveness and community invasibility. Progress in Physical Geography 30:409.
Schiff, J. 2007. Cellular automata: a discrete view of the world. John Wiley & Sons, Inc,
Auckland, Canada.

62
Sebert-Cuvillier, E., M. Simonet, V. Simon-Goyheneche, F. Paccaut, O. Goubet, and G. Decocq.
2009. PRUNUS: a spatially explicit demographic model to study plant invasions in
stochastic, heterogeneous environments. Biological Invasions:1-24.
Silva, E. A., J. Ahern, and J. Wileden. 2008. Strategies for landscape ecology: An application
using cellular automata models. Progress in Planning 70:133-177.
Soares-Filho, B., G. Coutinho Cerqueira, and C. Lopes Pennachin. 2002. DINAMICA—a
stochastic cellular automata model designed to simulate the landscape dynamics in an
Amazonian colonization frontier. Ecological Modelling 154:217-235.
Spencer, D., G. Ksander, M. Donovan, P. Liow, W. Chan, B. Greenfield, S. Shonkoff, and S.
Andrews. 2006. Evaluation of Waterhyacinth Survival and Growth in the Sacramento
Delta, California, Following Cutting. Journal of Aquatic Plant Management 44:50-60.
Vitousek, P. 1990. Biological invasions and ecosystem processes: towards an integration of
population biology and ecosystem studies. Oikos 57:7-13.
White, R. and G. Engelen. 2000. High-resolution integrated modelling of the spatial dynamics of
urban and regional systems. Computers, Environment and Urban Systems 24:383-400.
Williams, A., H. Duthie, and R. Hecky. 2005. Water hyacinth in Lake Victoria: Why did it
vanish so quickly and will it return? Aquatic Botany 81:300-314.
Wootton, J. 1988. Local interactions predict large-scale pattern in empirically derived cellular
automata. Naturwissenschaften 75:618-621.
Wortmann, J., J. Hearne, and J. Adams. 1998. Evaluating the effects of freshwater inflow on the
distribution of estuarine macrophytes. Ecological Modelling 106:213-232.
WWRP/WGNE Joint Working Group on Verification. 2009. NCAR Advanced Study Program
Summer Colloquium Forecast Verification in the Atmospheric Sciences and Beyond June
6-18, 2010.
Zuñiga, F. 2004. Técnicas de muestreo para manejadores de recursos naturales. Unam.