secretion of ferritin protein of periserrula leucophyryna in bacillus · · 2016-07-19secretion...
TRANSCRIPT

Korean Society for Biotechnology and Bioengineering Journal 31(2): 105-112 (2016)http://dx.doi.org/10.7841/ksbbj.2016.31.2.105 ISSN 1225-7117 / eISSN 2288-8268
고초균에서 흰이빨참갯지렁이 페리틴 단백질의 분비 및 사료 효율성
최장원*
Secretion of Ferritin Protein of Periserrula leucophyryna in Bacillus
subtilis and Its Feed Efficiency
Jang Won Choi*
Received: 11 April 2016 / Revised: 28 April 2016 / Accepted: 28 April 2016
© 2016 The Korean Society for Biotechnology and Bioengineering
Abstract: Ferritin is known to regulate iron metabolism and
maintain iron in a variety of the eukaryotic organisms. The
region encoding the mature ferritin (0.47 kb, H-type) of Peris-
errula leucophryna was amplified using the designed primers
including restriction enzyme site and termination codon and
subcloned in frame to the pRBAS secretion vector containing
the signal sequence, RBS, and promoter of amylase gene (E.
coli-Bacillus shuttle vector), resulting in recombinant pRBAS-
PLF vector. Recombinant ferritin (18 kDa) was correctly pro-
cessed and secreted from Bacillus subtilis LKS strain harbor-
ing the pRBAS-PLF vector and quantitatively analyzed by
SDS-PAGE and western blot, respectively. Secretion of the
ferritin was optimized by culture conditions (host, medium,
temperature, nitrogen source) in 3 L batch culture and 5 L jar
fermenter. Finally. the ferritin was largely produced using 50
L fermenter as the following conditions; at 30oC, 150 rpm, 1
vvm in Bacillus subtilis LKS using PY medium. The secreted
ferritin was maximally measured (approximately 177.6 ug/
ml) when the cell density reached to 14.4 at OD600 (20 h incu-
bation). The iron binding activity was confirmed by Perls'
staining in 7.5% non-denaturing gel, indicating that the multi-
meric ferritin (composed of 24 subunits) was formed in the
culture broth after secretion. Biologically, the culture broth
and powder type containing ferritin were tested for possibil-
ity as feed additive in chicken broiler. As a result, the ferritin
stimulated the growth of chick broil and improved feed effi-
ciency and production index.
Keywords: Ferritin, Periserrula leucophryna, Perls’ stain, feed
additive, production index
1. INTRODUCTION
페리틴 (Ferritin)은 세포 내의 대표적인 철 저장 단백질로 거
의 모든 생물 종에 존재하며 지금까지 사람을 포함해서 동물,
식물 및 미생물 등 여러 생물체로부터 분리되었다 [1]. 대부
분의 척추동물의 페리틴 단백질은 24개의 subunit으로 이루
어져 있으며 (MW 450 kDa), 분자 중심에 형성된 내공 (core)
에는 옥시하이드로사이드 폴리머 (oxyhydroxied polymer)가
있어 분자 당 헤모글로빈 1,200 분자를 생합성 할 수 있고
4,500개의 철 원자를 결합할 수 있다 [2,3]. 페리틴의 주된 기
능은 내공에 철을 Fe+3 상태로 보유함으로써 철을 함유하는
단백질이 합성될 때 철을 공급하고, 유리된 철에 의한 세포
내 독성으로부터 세포를 보호할 수 있고, 또한 철을 필요로
할 때는 Fe+2 상태로 환원시켜 공급한다 [4,5].
척추동물의 페리틴은 기능적 및 유전적으로 다른 두 가지
형태의 subunit, H (heavy/heart) type과 L (light/liver) type으로
구성되어 있으며 조직에 따라 그 조성 비율이 다른 여러 이
소페리틴 (isoferritin)으로 존재한다. L type subunit은 페리틴
분자의 내공을 형성하는데 관여하며, H type subunit은 Fe+2
의 흡수에 필요한 페록시다제 (ferroxidase) 활성을 가지고 있
어 페리틴 흡수 촉진 및 Fe+2의 산화 촉진 활성을 가지고 있
1대구대학교 생명환경대학 바이오산업학과1Department of Bioindustry, Daegu University, Gyeongbuk 712-714,KoreaTel: +82-53-850-6756, Fax: +82-53-210-8850e-mail: [email protected]
연구논문

106 Korean Society for Biotechnology and Bioengineering Journal 31(2): 105-112 (2016)
고, 또한 H 형이 풍부한 페리틴은 철분 운반의 기능이 높은
것으로 추측된다 [6,7]. 무기물 철분 제제는 철분 흡수 효율이
낮고 위염, 소화장애 등 부작용이 심해 철분 결핍증의 주요
대상이 유아 및 임산부임을 고려할 때 사용에 많은 제약을
갖는다. 이를 대체할 제제는 페리틴으로 철분 결핍성 빈혈의
치료에 쓰이고 있어 무기물 제제보다 부작용이 작고 훨씬 효
과가 있는 것으로 알려져 있다. 현재까지 효모 및 대장균에
서 유전자 재조합 페리틴의 생산을 위한 다양한 연구가 진행
되어 왔지만 [8-12], 대부분 페리틴의 이형집합체의 발현 및
정제에 관한 것이었고, 또한 사료첨가제로 사용하기 위한 시
도는 있었지만 발현 수율이 낮고 생물학적 활성 면에서도 그
다지 좋은 효과를 주지 못한 것으로 보고되었다.
흰이빨참갯지렁이 (Periserrula leucophryna)는 강화도 갯
벌 지역에 사는 다모류의 일종으로 분류학적으로는 전 세계
1속 1종으로 알려진 우리나라 고유의 생명체이며, 최장 길이
가 2~2.5 m까지 자라며 갯벌 환경 생태 및 순환에 지대한 역
할을 하는 것으로 알려져 있다 [13]. 흰이빨 참갯지렁이로부
터 페리틴 유전자가 분리되었고 (GenBank accession number
DQ207752), 페리틴 단백질은 신호서열 17개 아미노산을 포
함하여 174개 아미노산으로 구성된 18.2 kDa의 H-type sub-
unit 24개가 결합하여 multimer를 형성하는 것으로 보고되었
으며 [14], 그 shell내에 4,500 개의 철원자(3가 철)를 저장할
수 있는 철 저장 단백질로 무기물 제제보다 부작용이 작고
훨씬 효과가 있는 것으로 알려져 있다. 따라서 갯벌생명체로
부터 분리한 한 종류의 페리틴 (H-type) 유전자를 이용하여
효율적으로 고발현, 고분비 시스템을 개발하여 페리틴을 대
량생산하고, 생산된 발효액을 사용하여 용도에 따른 제형화
를 거쳐 기존의 사료에 첨가하여 생물학적 활성을 유도하는
기술의 개발이 필요한 실정이다.
따라서 본 연구에서는 우리나라 갯벌에만 서식하는 토착
갯지렁이 흰이빨참갯지렁이 유래 페리틴 성분을 고초균-벡
터 조합의 발현 및 분비시스템에 적용하여 대량생산체계를
구축하고 발효액에서 철분결합단백질을 손쉽게 회수하는
간편한 공정으로 경제성을 갖는 동물용 철분 사료첨가제를
개발하고자 한다.
2. MATERIALS AND METHOD
2.1. 재료 및 시약
제한효소 및 T4 DNA ligase는 KOSCHEM (Sungnam, Korea)
으로부터, Taq polymerases는 Applied Biosystem (Foster, CA,
USA), NEB (Beverly, MA, USA), 및 Biotools (Madrid, Spain)
로부터 구입하여 사용하였고, 올리고머는 Bioneer (Daejeon,
Korea)로부터 합성하여 사용하였다. Bacto-agar, tryptone, and
yeast extract는 Merck사 (Lutterworth, Germany), Immobilon-P
PVDF membrane은 Millipore사 (Bedford, MA, USA), QIAEX
agarose gel extraction kit는 Qiagen사 (Hilden, Germany), ECL
western blotting system은 Amersham Pharmacia Biotech사
(Piscataway, NJ, USA)로부터 구입하여 사용하였다. 일차 및
이차 항체는 Santa Cruz Biotechnology사 (Santa Cruz, CA
USA)사로부터, 나머지는 시약들은 분석용 수준급을 사용하
였다.
2.2. Bacterial strains, 배지 및 plasmid
E. coli XL1-Blue MRF (F', proAB, lacIqZM15, thi, recA, gyrA,
relA, supE, Tn10) (Stratagene, La Jolla, CA)는 형질전환 및 플
라스미드 증폭을 위한 기본 숙주로 사용하였고, pGEM-T Easy
vector (Promega, Madison, USA)는 PCR 산물을 클로닝하기
위하여, Bacillus subtilis LKS (nprR2, nprE18, aprA3, amyE)
균주 [15] 및 Bacillus subtilis 168 (wild type, GRAS and gen-
ome sequenced strain)은 외래단백질 발현 및 분비를 위한 숙
주세포로 사용하였다. 페리틴 유전자의 발현 및 분비를 위하
여 Bacillus 유래의 알칼리성 단백질분해효소 유전자의 프로
모터와 신호서열 [16]을 함유하는 pRBAS벡터는 pRB374 벡
터 [17]로부터 자체적으로 구성되었고, 흰이빨참갯지렁이 페
리틴 유전자 [14]를 도입하여 재조합 pRBAS-PLF 벡터를 본
연구에서 구성하였다. 플라스미드가 도입된 대장균은 37oC
에서 항생제 ampicillin (50 µg/mL)이 첨가된 Luria-Bertani
(LB) 배지에서 [18] 키웠고, 재조합벡터가 도입된 Bacillus 균
은 30oC 또는 37oC에서 항생제 kanamycin (50 µg/mL)이 첨가
된 PY 배지 [19]에서 배양하였다.
2.3. 분비벡터 구성 및 고초균 형질전환
분비벡터 구성을 위하여 PCR을 위한 primer들은 염기 서열
이 완전히 밝혀진 B. subtilis로부터 알카리성 단백질 분해효
소의 promoter, SD 서열, signal sequence 서열을 포함하도록
forward primer F1(5’-GGGGAATTCTGCTTGGCGAATGTT
CATCT-T3')와 reverse primer R1(5’-GGGAAGCTTAGCCT
GCGCAGACATGTTGCT-3')를 합성하여 염색체 DNA를 주
형으로 Taq DNA 중합효소를 이용하여 95oC에서 5분간 pre-
denaturation 시키고, 95oC에서 1분, 45oC에서 1분, 72oC에서
2분의 조건으로, 30 cycle의 PCR 반응을 실시한 후 얻어진
PCR 산물은 pGEM-Teasy 벡터 (Promega, Madison, USA)에
클로닝하여 E. coli XL1-Blue MRF'로 형질 전환하였다 [18].
얻어진 형질전환체들로부터 플라스미드를 추출하여 DNA
크기 및 제한효소 절단에 의해 재조합된 벡터를 확인한 다음,
최종적으로 DNA 염기서열을 분석하였으며 pGEMAS로 명
명하였다. 대장균-고초균 shuttle vector인 pRB374 [17]를
EcoRI 및 HindIII 제한효소로 잘라 5.9 kb 크기의 DNA 조각
을 전기영동에 의해 분리하고, 위에서 구성된 pGEMAS도 같
은 효소로 처리하여 537 bp 크기의 단편을 확보한 다음 liga-
tion하여 pRBAS 분비벡터를 구성하여 페리틴 분비 생산을 위
한 vector로 사용하였다. 고초균의 형질전환은 SPMM-I (Spi-
zizen's minimal medium)과 SPMM-II 배지를 사용하는 Sadaie
and Kada [20] 방법에 의해 행하였고 kanamycin (50 µg/mL)
이 들어간 고체 LB 배지에서 형질전환체를 선별하였다.

Secretion of Ferritin Protein in Bacillus subtilis 107
2.4. 단백질 발현 분비 및 대량배양
얻어진 형질전환체는 고초균 배양 및 분비시스템을 통하여
페리틴 생산 및 조건 확립을 위하여 고초균 배양에 추천되는
LB [18], PY [19] 및 MSR (modified super rich) 배지 [21]를 선
택하여 여러 배지 조성, 배양 온도, 배양시간, aeration 등의 변
화에 따라 전 배양 (10 mL, 300 mL)을 통하여 균을 키워 3 L
baffled flask (working vol. 1 L), 5 L jar fermenter (working vol.
3 L, KoBiotech. Korea), 50 L fermenter (working vol. 30 L, 진
주 바이오센터, Korea)를 이용하여 1%정도 접종한 다음 페리
틴 생산균주의 최적 조건들을 조사하였다.
2.5. SDS-PAGE 및 western blot 분석
발효 후 얻어진 배양액은 알콜로 농축하여 단백질 pellet을 얻
어 건조한 다음, Lammlie [22]의 loading buffer (0.05 M Tris-
HCl, pH 6.8, 0.1 M DTT, 2% SDS, 10% glycerol, and 0.1%
bromphenol blue)에 현탁하여 100oC에서 1분 가열 후 12.5%
SDS (sodium-dodecyl sulfate) PAGE (polyacrylamide gel elec-
trophoresis)에 의해 분석하였고, 전체 분비단백질의 양은 bo-
vine serum albumin을 표준단백질로 사용하여 Bradford pro-
tein assay kit (Bio-Rad, USA)에 의해 정량화하였다 [22]. 배양
액 내의 단백질들을 SDS-PAGE에서 fractionation후, Trans-
Blot SD apparatus (Bio-Rad, USA)를 이용하여 gel로부터
PVDF membrane으로 단백질들을 이동시킨 다음, TBST (10
mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.1% Tween-20)에 5%
로 농도로 첨가된 skim milk 용액을 이용하여 4°C에서 over-
night blocking 시켰다. 그 후, 1차 항체인 rabbit polyclonal
anti-ferritin antibody (1,000:1 dilution, Santa Cruz Biotechno-
logy)로 2시간동안 반응시킨 다음, 결합되지 않은 1차 항체는
TBST 용액으로 세척하여 제거하였고, 다시 2차 항체인 horse-
radish peroxidase-conjugated anti-rabbit IgG secondary anti-
body (1:1000, Santa Cruz Biotechnology)로 2시간 동안 반응
한 다음, ECLTM Western blotting detection system (Amersham
Pharmacia Biotech, USA)을 이용하여 develop하여 X-ray필름
에 감광하였다. 그 blot은 scanning 하여 ChemiImager™ (Al-
pha Innotech Corporation, San Leandro, USA)로 단백질 밴드
의 intensity를 측정하였다.
2.6. Perls’ staining에 의한 철분 단백질 확인
분비된 페리틴의 생물학적 활성을 조사하기 위하여, 페리틴
분자의 철 결합능은 Perls' staining 방법 [24-26]에 의해 확인
되었다. 즉, 배양액 일정량을 western blot용 시료와 Perls’
staining용 시료, duplicate를 알콜로 농축하고 10,000 × g에서
10분간 원심분리하여 단백질 pellet을 얻어 건조한 다음, Lam-
mlie [22]의 loading buffer (0.05 M Tris-HCl, pH 6.8, 0.1 M
DTT, 2% SDS, 10% glycerol, and 0.1% bromphenol blue)와
SDS와 DTT를 포함하지 않는 loading buffer에 현탁하여 12.5%
SDS-PAGE와 7.5% non-denaturing polyacrylamide gel (native
gel)에서 각각 전기영동을 수행하였다. 그 후 native gel은 sol-
ution A (1 ml HCl in 24 ml DW)와 solution B (1 g potassium
ferricyanide in 25 mL DW)의 혼합액에서 15분간 염색하고 물
로 여러 번 세척한 다음, DAB (Diaminobenzide) 기질 용액 (25
mg DAB, 2.5 mL 1 M Tris-HCl, pH 7.5, and 40 µL of H2O2 in
total volume 50 mL)과 30분간 반응하여 철분을 함유하는 페
리틴 중합체의 불용성 진한갈색 band를 확인하였다.
2.7. 육계를 이용한 field test
Ferritin이 도입된 고초균을 PY배지 (1% peptone, 0.5% yeastex
LSP, 2% monodex, all trace elements)에서 50 L fermenter를 이
용하여 대량 배양한 다음 배양액을 원심분리에 의해 회수하
여 분비된 페리틴 농도를 SDS-PAGE, western blot, Bradford
단백질 정량 등에 의해 측정하였다. 이렇게 얻어진 페리틴 단
백질을 같은 조성의 액체배지로 적절히 희석하여 (~1-10 ug/
mL) 자동급수 시설을 갖춘 육계농장 (경북 김천시)의 협조를
받아 공급 음용수에 첨가하여 액상의 사료첨가제가 육계 성
장에 미치는 영향을 조사하였다. 또한 상기 배양에 의해 얻어
진 배양액에 말분 (wheat sorts, 대한제분, Korea)을 섞어 동결
건조에 의해 파우더 형태의 사료첨가제를 제조하여 같은 시
설의 협조를 받아 본사료에 첨가하는 방식으로 투여하여 고
형분 상태 페리틴의 사료요구율 (FE, feed efficiency) 및 생산
지수 (PI, production index)를 산출하여 대조군과 비교하였다
[27].
3. RESUTLS AND DISCUSSION
3.1. 분비벡터 구축
고초균 클로닝 벡터인 5.9 kb 크기의 E. coli-Bacillus shuttle
vector인 pRB374 [17]를 이용하여 기존에 확보하고 있는 alka-
line protease promoter (0.45 kb), 리보솜결합서열 (GGAGAG
GG)과 신호서열 (87 bp)을 [16] 설계된 primer를 이용하여
PCR 증폭 후 pRB374벡터의 EcoRI 및 HindIII 제한효소 부위
에 도입하여 6.43 kb의 분비벡터를 조성하였고 (pRBAS vec-
tor), 구성된 분비벡터 pRBAS를 HindIII로 잘라 전기영동에
의해 6.43 kb 크기의 벡터 단편을 분리하였다. 흰이빨참갯지
렁이의 페리틴 유전자가 클로닝되어 있는 4.07 kb 크기의 pLF
클론 [14]을 주형으로 하여, pRBAS 벡터의 분비서열과 frame
이 맞도록 디자인하여 합성한 Fer-F1 (5’-GGGAAGCTTGA
AGCTGGAATCAACAAACAG-3’) 및 Fer-R1 primer (5’-CC
CAAGCTTCTAGCTCTTGAGTTCCTTGTC-3’) set로 PCR 증
폭하여 0.49 kb 크기의 DNA 단편을 확보하고 HindIII로 잘라
전기영동에 의해 DNA를 elution 하였다. 확보한 분비벡터와
페리틴 mature form DNA를 농도 측정 후 ligation하여 대장균
에 형질전환한 다음, 항생제 내성을 보이는 형질전환체를 분
석하여 insert DNA가 들어간 형질전환체를 선별하고 plasmid
DNA를 추출하여 최종적으로 염기서열 분석에 의해 in frame
하에서 정확하게 페리틴 유전자가 도입된 것을 확인하고
pRBAS-PLF (6.9 kb)로 명명하였다 (Fig. 1).

108 Korean Society for Biotechnology and Bioengineering Journal 31(2): 105-112 (2016)
3.2. 형질전환체를 이용한 발현 유도 및 분비
구성된 pRBAS-PLF 분비벡터는 SPMM-I (Spizizen's minimal
medium)과 SPMM-II 배지를 사용하는 Sadaie and Kada [20]
방법에 의해 고초균 (Bacillus subtilis LKS 및 Bacillus subtilis
168)으로 형질전환하였고, kanamycin (50 µg/mL)이 들어간
고체 LB 배지에서 1차 형질전환체를 선별한 다음, plasmid를
분리하여 DNA 염기서열 분석을 통하여 페리틴 유전자의 도
입 및 클로닝 부위의 신호서열과 frame이 정확하게 도입되었
는지 여부를 확인하였다. 페리틴 유전자는 단백질의 mature
form에 대하여 encoding하며 157개의 아미노산에 대한 정보
를 가지고 있고 [14], amylase 신호서열의 연결부위에 HindIII
제한효소부위에서 유래된 AAG (Lys), CTT (Leu) 2개의 아미
노산이 첨가되어 159개의 아미노산으로 구성된 페리틴 (약
18 kDa)이 세포 외로 분비될 것으로 예상하였다.
기초적인 배양조건을 확립하기 위하여 적절한 균주, 온도,
배지, 배양시간 등의 1차적 조건들을 3 L baffled flask 내에 1
liter의 액상배지에서 batch culture를 통하여 조사하였다. 균
주는 Bacillus subtilis LKS 및 Bacillus subtilis 168을 사용하였
고, 배지는 LB [18], PY [19] 및 MSR [21] 배지를, 접종량은 1-
4%, 배양온도는 30oC 및 37oC, 배양시간은 24-48시간 등, 배
양시간에 따라 배양액을 수집하여 단백질을 분석한 결과, 균
주는 pRBAS-PLF 분비벡터를 함유하는 Bacillus subtilis 168
에서보다는 Bacillus subtilis LKS에서, 접종량은 1%, 배양온
도는 30oC, 배지는 LB에서 보다는 PY 및 MSR 배지에서 더
많은 양의 페리틴이 분비되는 것으로 나타났으며, 특히 24시
간 배양에서는 PY 배지가 MSR 배지보다 조금 더 많은 양의
페리틴을 생산하는 것으로 나타났다 (data not shown). 또한
배양액의 단백질 분석 결과 페리틴 단백질의 amylase 신호서
열이 processing되어 세포 바깥으로 단백질 분비가 일어나는
것으로 확인하기 위하여, 즉 페리틴이 생성되어 세포 내에 축
적되는지 또는 세포 외로 분비되는지를 알아보기 위하여 상
기 실험 시료 24시간 및 36시간 배양 샘플을 수집하여 1 mL
을 취한 다음, cell pellet과 상등액을 분리하여 cell pellet은
sonication에 의해 분쇄하고, 상등액은 에탄올로 단백질을 침
전시켜 SDS-PAGE와 western blot에 의해 분석한 결과, Fig.
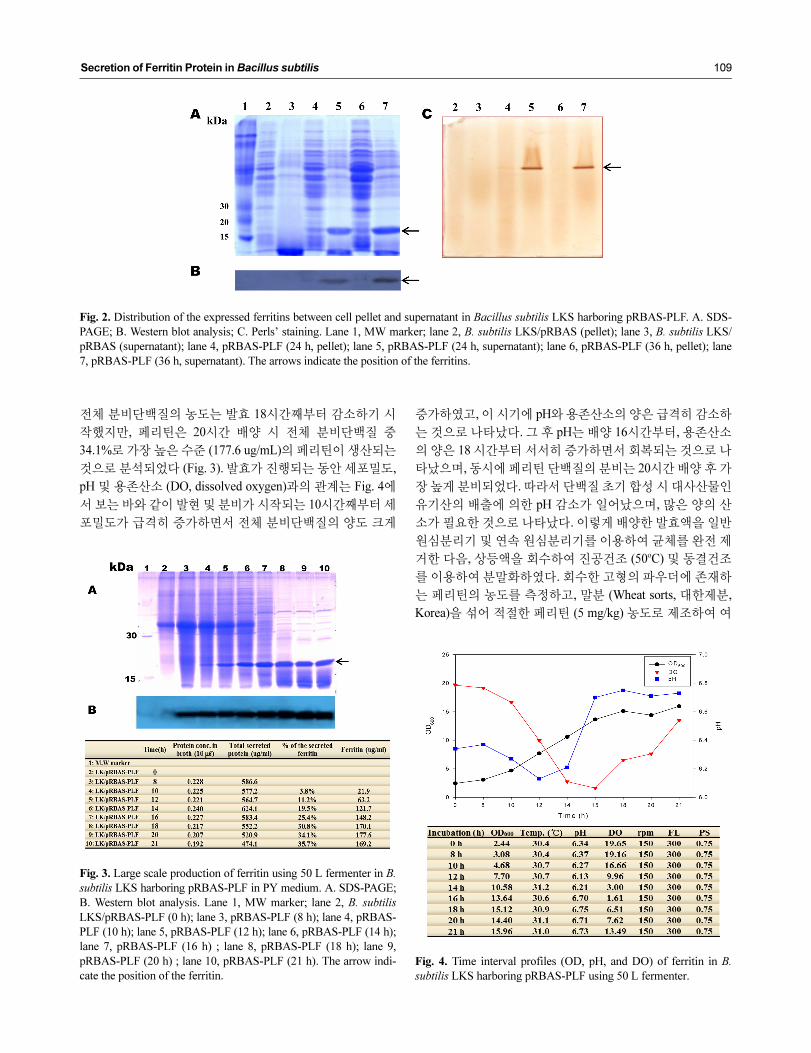
2A, 2B에서 보는 바와 같이 수용성 페리틴은 cell pellet frac-
tion에서는 거의 검출되지 않았고, 완전하게 세포 외로 분비
되는 것을 확인하였다. 따라서 구축한 분비벡터의 amylase
promoter의 전사가 효율적으로 작동하여 생산된 페리틴의 N-
말단 부위에 연결되어 있는 amylase 신호서열이 정확하게 제
거되어 활성적 형태의 페리틴이 분비되었음을 보여준다
(Fig. 2A, 2B).
Batch culture에서 확립된 배양 조건이 대량생산을 위한 fer-
menter 조건과 정확하게 같지는 않지만 적정온도 (30oC), 배
지조성 (PY + peptone, 1-4%), 배양시간 (24-36 h) 등을 바탕
으로 5 L jar fermenter (working vol. 3 L)에서 대량생산을 위
한 조건을 확립하였다. 페리틴 유전자를 함유하는 분비벡터,
pRBAS-PLF가 도입된 고초균 (B. subtilis LKS)을 LB 배지에
서 seed culture하여 PY 배지 3리터에 최종적으로 1%가 되도
록 접종하고, 30oC, 300 rpm에서 1 vvm의 공기 주입속도로 배
양하여 시간에 따라 배양액을 수집하여 단백질을 분석한 결
과, 24-36시간 배양했을 때 가장 좋은 수율의 최대 생산량
(85-90 ug of ferritin/mL)을 얻었다 (data not shown). 그 결과
얻어진 결과들을 참고로 하여 50 L fermenter에서 대량 배양
(working vol. 30 L)을 시도하였다. 질소원으로 사용한 peptone
의 경우 1-4% 농도로 사용하였을 때 농도가 증가하면서 페리
틴 단백질이 약간씩 증가하였지만 대량생산의 경우 경제성
및 페리틴 생산성을 고려하여 1% peptone을 사용하였고, 배
양시간도 기기 사용 비용의 절감을 위하여 충분한 수율로 페
리틴이 생산되는 24시간까지 배양을 시도하였다.
3.3. 대량배양 및 단백질 분석
50 L (working vol. 30 L, 1% soypeptone) 발효조의 경우 균 접
종량은 1% (300 mL)로 접종한 다음, 임펠라 회전속도는 150
rpm에서, 1 vvm의 공기 주입속도로 30oC에서 발효를 진행하
면서 8시간 배양 후부터 매 2시간 별로 시료를 일정량 (~100
mL) 채취하였다. 각각의 시료 0.5 mL에 에탄올을 첨가하여
단백질을 회수한 다음 12.5% SDS-PAGE와 western blot에 의
해 분비된 페리틴 단백질의 생산효율을 imageanalyzer에 scan-
ning하여 분석하였다. Fig. 3에서 보듯이 발현 및 분비는 배양
후 10 시간째 (전체 분비단백질의 3.8%)부터 본격적으로 시
작되었고 배양시간에 비례하여 증가하는 것으로 나타났다.
Fig. 1. Construction of the secretion vector containing mature form
DNA of ferritin from Periserrula leucophryna.
Secretion of Ferritin Protein in Bacillus subtilis 109
전체 분비단백질의 농도는 발효 18시간째부터 감소하기 시
작했지만, 페리틴은 20시간 배양 시 전체 분비단백질 중
34.1%로 가장 높은 수준 (177.6 ug/mL)의 페리틴이 생산되는
것으로 분석되었다 (Fig. 3). 발효가 진행되는 동안 세포밀도,
pH 및 용존산소 (DO, dissolved oxygen)과의 관계는 Fig. 4에
서 보는 바와 같이 발현 및 분비가 시작되는 10시간째부터 세
포밀도가 급격히 증가하면서 전체 분비단백질의 양도 크게
증가하였고, 이 시기에 pH와 용존산소의 양은 급격히 감소하
는 것으로 나타났다. 그 후 pH는 배양 16시간부터, 용존산소
의 양은 18 시간부터 서서히 증가하면서 회복되는 것으로 나
타났으며, 동시에 페리틴 단백질의 분비는 20시간 배양 후 가
장 높게 분비되었다. 따라서 단백질 초기 합성 시 대사산물인
유기산의 배출에 의한 pH 감소가 일어났으며, 많은 양의 산
소가 필요한 것으로 나타났다. 이렇게 배양한 발효액을 일반
원심분리기 및 연속 원심분리기를 이용하여 균체를 완전 제
거한 다음, 상등액을 회수하여 진공건조 (50oC) 및 동결건조
를 이용하여 분말화하였다. 회수한 고형의 파우더에 존재하
는 페리틴의 농도를 측정하고, 말분 (Wheat sorts, 대한제분,
Korea)을 섞어 적절한 페리틴 (5 mg/kg) 농도로 제조하여 여
Fig. 2. Distribution of the expressed ferritins between cell pellet and supernatant in Bacillus subtilis LKS harboring pRBAS-PLF. A. SDS-
PAGE; B. Western blot analysis; C. Perls’ staining. Lane 1, MW marker; lane 2, B. subtilis LKS/pRBAS (pellet); lane 3, B. subtilis LKS/
pRBAS (supernatant); lane 4, pRBAS-PLF (24 h, pellet); lane 5, pRBAS-PLF (24 h, supernatant); lane 6, pRBAS-PLF (36 h, pellet); lane
7, pRBAS-PLF (36 h, supernatant). The arrows indicate the position of the ferritins.
Fig. 3. Large scale production of ferritin using 50 L fermenter in B.
subtilis LKS harboring pRBAS-PLF in PY medium. A. SDS-PAGE;
B. Western blot analysis. Lane 1, MW marker; lane 2, B. subtilis
LKS/pRBAS-PLF (0 h); lane 3, pRBAS-PLF (8 h); lane 4, pRBAS-
PLF (10 h); lane 5, pRBAS-PLF (12 h); lane 6, pRBAS-PLF (14 h);
lane 7, pRBAS-PLF (16 h) ; lane 8, pRBAS-PLF (18 h); lane 9,
pRBAS-PLF (20 h) ; lane 10, pRBAS-PLF (21 h). The arrow indi-
cate the position of the ferritin.
Fig. 4. Time interval profiles (OD, pH, and DO) of ferritin in B.
subtilis LKS harboring pRBAS-PLF using 50 L fermenter.

110 Korean Society for Biotechnology and Bioengineering Journal 31(2): 105-112 (2016)
러 축종의 사육에 필요한 철분강화제 및 신진대사 촉진제로
사용하였다.
3.4. 생물학적 활성
3.4.1. 철 결합능 분석 (Perls’ staining)
대부분의 페리틴은 고분자량의 multimer 단백질 (24 subunits)
이며 분자 중심에 형성된 내공(core)에는 옥시하이드로사이
드 폴리머(oxyhydroxied polymer)가 있어 분자 당 헤모글로빈
1,200분자를 생합성할 수 있고, 내부 표면의 여러 지점에서 3
가 철과 결합하여 철-단백질 상호작용하는 경계면을 형성하
며, 4,500개의 철 원자를 함유할 수 있다 [2-3, 28]. 그 경계면
은 내공의 핵형성 부위이며, subunit의 dimer가 형성되는 지
점에 위치하여 그 연결부위를 통하여 철이 이동하게 된다
[29,30]. 페리틴이 생성되어 고초균 바깥으로 분비되는 성상
이 monomer로 분비됨과 동시에 24개의 monomer들이 결합
하여 multimer를 형성하고, 또한 철 결합 능을 가지고 있는지
조사하기 위하여, Fig. 2A & B에서 보는 시료에 대하여 SDS
(sodium dodecyl sulfate)와 DTT (dithiothreitol)를 포함하지 않
는 native non-denaturating gel (7.5%)을 사용하여 분석한 결
과, Fig. 2C에서 보는 바와 같이 약 18 kDa의 monomer 페리틴
이 SDS-PAGE에서 확인되었으며, 철분을 함유하는 페리틴
중합체 형태는 non-denaturing gel (7.5%) 위쪽 부분의 고분자
량 위치에서 DAB (diaminobenzide) 기질에 의한 염색 후 진
한 갈색을 나타내는 band로 확인되었고 또한 철을 결합하는
활성도 유지하고 있는 것으로 분석되었다 (Fig. 2C). 결과적
으로 분비된 페리틴은 24개의 subunit으로 구성된 큰 구형의
거대분자를 형성하고 이때 생긴 내부 공간에 4,500개의 철 원
자를 축적할 수 있고, 또한 헥사시아노철산염 이온은 (K4Fe
(CN)6) 페리틴 중합체의 내부공간에 존재하는 3가 철 Fe3+과
결합하여, 군청색의 불용성 색소인 Prussian Blue로 알려진
potassium ferric ferrocyanide를 형성한다. 이는 생명체에 유해
한 2가 철 이온(Fe2+)은 3가 철이온처럼 Prussian Blue 반응을
하지 않으므로 페리틴 중합체는 3가 철이온을 포집하는 것을
의미한다.
3.4.2. 육계 필드시험
Ferritin이 도입된 고초균을 대량배양하여 원심분리에 의해
상등액을 회수하여 분비된 페리틴의 양을 측정한 다음, 재료
및 방법에 따라 본 사료에 0.1% 정도로 투여할 수 있도록 액
상 및 파우더 형태의 사료첨가제를 제조하였다. 경상북도 김
천시에 위치한 자동 급수 시설을 갖춘 육계농장의 협조를 받
아 페리틴 함유 사료첨가제의 생물학적 활성을 측정하였다.
Table 1에서 보는 바와 같이 대조구 및 시험구의 경우 9,000
수의 병아리를 사용하였고, 병아리가 입고된 후 2-3일 정도의
적응기간 후부터 액상의 페리틴 (5 ug/mL) 함유 사료첨가제
를 음용수의 0.1% 정도 되게 출고 직전까지 투여하였다. 출
고시 전체중량 (TBW)은 시험구의 경우 14,110 kg (대조구
12,912 kg)이었고, 평균 한 수당 체중은 시험구는 1.59 kg으로
대조구 1.47 kg보다 8.2% 정도 증체 효과를 나타냈다. 일당 체
중 증가율은 시험구에서 46.76 g이며 대조구의 경우 43.23 g
으로 시험구에서 높게 나타났으며, 시험구에서는 전체 사료
량이 21,871 kg (2.46 kg/수) 소모되었고, 대조구에서는 22,467
kg (2.56 kg/수) 소진되었다. 전체적인 사료요구율 (FE= total
feed intake/total body weight)은 시험구에서 1.55, 대조구의 경
우 1.74로서 시험구에서 약 11% 정도 개선된 효과를 보이는
것으로 나타났다. 또한 생산지수 [PI (production index) : (ave-
rage body weight × rate of survival)/(feed efficiency × culturing
period) × 100]의 경우 시험구에서는 297.5이며, 대조구에서는
242.5로 페리틴 함유 사료첨가제 투여 시 생산성이 약 22.7%
정도 크게 증가한 것으로 나타났다. 사육기간 동안 필요한 음
용수의 양과 액상의 페리틴 양을 측정한 결과, 1 마리가 필요
한 음용수는 34일동안 약 5,432 mL이였고 9,000마리의 경우
48,888리터의 음용수가 필요하였으며, 따라서 액상의 페리틴
(5 ug/mL)은 약 49리터 정도가 필요한 것으로 나타났다.
Table 1. Field test for biological activity of ferritin (liquid type) in farm animal (chicken)
SampleNo. Input
animal
Culturing
Period (days)
No. Output
animal
Rate of
Survival (%)
TBW
(kg)
ABW
(kg)
ADG
(g)
TFI
(kg)FE PI
Control group 9,000 34 8,784 97.6 12,912 1.47 43,23 22,467 1.74 242.5
Treated group 9,000 34 8,874 98.6 14,110 1.59 46.76 21,871 1.55 297.5
Liquid feed containing ferritin (5 mg ferritin/L culture broth) was supplied to final concentration of 0.1% in drinking water after input.
ADG (Average daily gain): ABW (Average body weight)/Culturing Period
FE (Feed efficiency): TFI (Total feed intake)/ TBW (Total body weight)
PI (Production index): (ABW × Rate of Survival)/(FE × Culturing Period) × 100
Table 2. Field test for biological activity of ferritin (powder type) in farm animal (chicken)
SampleNo. Input
animal
Culturing
Period (days)
No. Output
animal
Rate of
Survival (%)
TBW
(kg)
ABW
(kg)
ADG
(g)
TFI
(kg)FE PI
Control group 10,200 30 9,690 95.0 14,922 1.54 51.3 25,008 1.68 290.3
Treated group 9,900 30 9,700 98.0 15,617 1.61 53.7 23,998 1.54 341.5
Powder containing ferritin (5 mg ferritin/kg wheat sorts ) was supplied to final concentration of 0.1% in main feed after input.
ADG (Average daily gain): ABW (Average body weight)/Culturing Period
FE (Feed efficiency): TFI (Total feed intake)/ TBW (Total body weight)
PI (Production index): (ABW × Rate of Survival)/(FE × Culturing Period) × 100

Secretion of Ferritin Protein in Bacillus subtilis 111
파우더 형태의 페리틴을 투여한 육계시험의 경우, 대조구
및 시험구의 경우 각각 10,200 및 9,900 수의 병아리를 사용
하였고, 병아리가 입고된 직후부터 출고 전까지 본사료에
0.1% 정도 혼합하여 투여하였다. 출고시 전체중량 (TBW)은
시험구의 경우 15,617 kg (대조구 14,922 kg)이고, 한 수당 평
균체중은 시험구는 1.61 kg으로 대조구 1.54 kg 보다 4.5% 정
도 증체 효과를 나타냈다. 일당 체중 증가율은 시험구에서
53.7 g이었고, 대조구의 경우 51.3 g으로 시험구에서 조금 높
게 나타났으며, 시험구에서는 전체 사료량이 23,998 kg (2.42
kg/수) 소모되었고, 대조구에서는 25,008 kg (2.45 kg/수) 소진
되었다. 전체적인 사료요구율은 (FE)은 시험구에서 1.54, 대
조구의 경우 1.68로서 시험구에서 약 9.1% 정도 개선된 효과
를 보이는 것으로 나타났다. 또한 생산지수 (PI)의 경우 시험
구에서는 341.5이며, 대조구에서는 290.3 으로 생산성이 약
17.6% 정도 증가한 것으로 나타났다. 사육기간 동안 페리틴
투여군에 있어서 무게변화는 4주째 들어가면서 크게 증가하
였고 또한 사료섭취량도 비례하여 증가하였다. 사육기간 30
일동안 42 g에서 시작하여 출하 시 평균체중이 1.61 kg으로
1,566 g 증가하였으며, 한 마리가 평균적으로 먹는 사료 양은
2,424 g으로 측정되었다. 따라서 시험구 9,900수의 육계 사육
에 필요한 사료 총 양은 23,998 kg이 필요하였으며, 페리틴 파
우더 (5 mg/kg 말분)는 약 24 kg이 필요한 것으로 나타났다.
전체적으로 볼 때 육계 사육농장의 환경 조건 및 입수되는 병
아리의 초기 건강상태에 따라, 또는 페리틴의 액상 및 파우더
형태의 성상에 따라 약간의 성장효과에 있어서 차이는 있지
만 전반적으로 페리틴의 액상 및 파우더 형태 둘 다 육계 사
육에 좋은 성장 효과를 보이는 것으로 나타났으며, 이는 철분
단백질에 의한 원활한 산소 공급에 의해 신진대사 촉진 효과
라고 추정되어진다 [2].
4. CONCLUSION
흰이빨참갯지렁이로부터 페리틴 유전자를 분리하여 amyl-
ase promoter, 리보솜결합부위, 신호서열을 포함하는 E. coli-
Bacillus shuttle vector에 도입하여 단백질분해효소가 결여된
GRAS (generally regarded as safe) 균주인 고초균에서 발현 및
분비를 시도하였다. 그 결과 배양조건 최적화에 의해 높은 수
율의 활성 페리틴이 배양액으로 분비되었고 [31], 24 subunit
로 구성된 페리틴 중합체 형성을 확인하였다. 철분 단백질을
산업적으로 생산하기 위하여 고초균을 이용하였고 [32], 또
한 배양액으로 단백질을 분비하여 배양액만 사료제제로서
활용하기 위한 시스템을 구축하였다. 생물학적 활성을 검증
하기 위하여 액상 및 파우더 형태의 사료첨가제를 제조하여
육계에 투여하였을 때 체중증가, 사료요구율 및 생산지수 등
이 크게 증가하였고, 사료첨가제로서 충분한 경쟁력을 가질
수 있을 것이라 전망되며, 여러 축종의 동물에 투여하여 기초
data를 확보할 예정이며 사료 산업 뿐만 아니라 철 결핍 예방
및 치료용을 위한 의약소재산업, 식품산업 등의 분야에서 유
용하게 사용할 수 있을 것이라 전망한다. 따라서 차후에는 생
산성을 증진시키기 위하여 여러 가지 promoter 및 신호서열
조합에 의한 발현 분비 최적화 실험을 시도한 후에, 산업적
용도로 사용하기 위하여 pilot scale에서 대량 생산 조건을 확
립하고 안정성을 확보할 예정이다.
Acknowledgements
이 논문은 2011학년도 대구대학교 학술연구비 지원에 의하
여 연구되었음.
REFERENCES
1. Andrew, S. C., P. Arosio, W. Bottke, J. F. Briat, M. von Darl, P. M.
Harrison, J. P. Laulhe`re, S. Levi, S. Lobreaux, and S. J. Yewdall
(1992) Structure, function, and evolution of ferritins. J. Inorg. Bio-
chem. 47: 161-174.
2. Theil, E. C. (1987) Ferritin: structure, gene, regulation, and cellu-
lar function in animals, plants, and microorganisms. Ann. Rev. Bio-
chem. 56: 289-315.
3. Harrison, P. M. and P. Arosio (1996) The ferritins: molecular prop-
erties, iron storage function and cellular regulation. Biochim. Bio-
phys. Acta 1275: 161-203.
4. Baynes, R. D. and T. H. Bothwell (1990) Iron deficiency. Ann. Rev.
Nutr. 10: 133-148.
5. Theil, E. C. (1990) Regulation of ferritin and transferrin receptor
mRNAs. J. Biol. Chem. 265: 4771-4774.
6. Lawson, D. M., A. Treffry, P. J. Artymiuk, P. M. Harrison, S. J.
Yewdall, A. Luzzago, G. Cesareni, S. Levi, and P. Arosio (1989)
Identification of the ferroxidase centre in ferritin. FEBS Lett. 254:
207-210.
7. Stefanini, S., E. Chiancone, P. Arosio, A. Finazzi-Agrò, and E. An-
tonini (1982) Structural heterogeneity and subunit composition of
horse ferritins. Biochem. 21: 2293-2299.
8. Park, H. K., Y. S. Huh, J. W. Lee, and I. H. Kim (2002) Purifica-
tion of Heavy Chain Ferritin (FH) from Recombinant E. coli. Kor.
J. Biotechnol. Bioeng. 17: 44-451.
9. Lee, J., S. W. Kim, Y. H. Kim, and J. Y. Ahn (2002) Active human
ferritin H/L-hybrid and sequence effect on folding efficiency in Es-
cherichia coli. Biochem. Biophys. Res. Commun. 298: 225-229.
10. Chang, Y. J., C. U. Park, and K. S. Kim (2004) Iron uptake by the
recombinant yeasts producing ferritin heteropolymers. Kor. J. Bio-
technol. Bioeng. 19: 352-357.
11. Lee, J. L., R. E. Levin, and H. Y. Kim (2008) Improved coexpres-
sion and multiassembly properties of recombinant human ferritins
subunit in Escherichia coli. J. Microbiol. Biotechnol. 18: 926-932.
12. Seo, H. Y., Y. J. Chung, S. J. Kim, C. U. Park, and K. S. Kim
(2003) Enhanced expression and functional characterization of the
human ferritin H- and L-chain genes in Saccharomyces cerevisiae.
Appl. Microbiol. Biotechnol. 63: 57-63.
13. Brands, S. J. (comp.) (1989-2002) Systema Naturae 2000. Amster-

112 Korean Society for Biotechnology and Bioengineering Journal 31(2): 105-112 (2016)
dam, The Netherlands. [http://sn2000.taxonomy.nl/]
14. Jeong, B. R., S. M. Chung, N. J. Baek, K. B. Koo, H. S. Baik, H. S.
Joo, C. S. Chang, and J. W. Choi (2006) Characterization, cloning,
and expression of the ferritin gene from the Korean polychaete,
Periserrula leucophryna. J. Microbiol. 44: 54-63.
15. Kim, S. I., J. W. Choi, and S. Y. Lee (1997) Effects of pleiotrophic
mutations, degUh and spoOA, on the production of foreign pro-
teins using the heterologous secretion system of Bacillus subtilis.
Mol. Cells. 7: 158-164.
16. Joo, H. S. and J. W. Choi (2011) Cloning and expression of a alka-
line protease from Bacillus clausii I-52. J. Agri. Life Sci. 45: 201-
212.
17. Shi, X. L., M. Q. Feng, Y. J. Zhao, X. Guo, and P. Zhou (2008)
Overexpression, purification and characterization of a recombinant
secretary catalase from Bacillus subtilis. Biotechnol. Lett. 30: 181-
186.
18. Sambrook, J., E. F. Fritsch, and T. Maniatis (1989) In Molecular
cloning, A Laboratory Manual, Cold Spring Harbor Laboratory
Press.
19. Lee, M. H., J. J. Song, Y. H. Choi, S. P. Hong, E. H. Rha, K. Kim,
S. G. Lee, S. C. Lee, H. Poo, Y. B. Seu, and M. H. Sung (2003)
High-level expression and secretion of Bacillus pumilus lipase B26
in Bacillus subtilis chungkookjang. J. Microbiol. Biotechnol. 13:
892-896.
20. Sadaie, Y. and T. Kada (1983) Formation of competent Bacillus
subtilis cells. J. Bacteriol. 153: 813-821.
21. Park, I. S. and B. G. Kim (2001) Media optimization and compari-
son of fermentation type for overproduction of staphylokinase in
Bacillus subtilis WB700. Kor. J. Biotechnol. Bioeng. 16: 415-419.
22. Laemmli, U. K. (1970) Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature 227: 680-685.
23. Bradford, M. M (1976) A rapid and sensitive method for the quan-
tification of microgram quantities of protein utilizing the principle
of protein-dye binding. Anal. Biochem. 72: 248-254.
24. Perls, M. (1867) Nachweis von eisenoxyd in gweissen pigmenten.
Virchows Arch. 39: 42-48.
25. Roschzttardtz, H., G. Conejero, C. Curie, and S. Mari (2009) Iden-
tification of the endodermal vacuole as the iron storage compart-
ment in the Arabidopsis embryo. Plant Physiol. 151:1329-1338.
26. Seo, H. Y., Y. J. Chung, S. J. Kim, C. U. Park, and K. S. Kim
(2003) Enhanced expression and functional characterization of the
human ferritin H- and L-chain genes in Saccharomyces cerevisiae.
Appl. Microbiol. Biotechnol. 63: 57-63.
27. Owens, F. N., D. S. Secrist, W. J. Hill, and D. R. Gill (1997) The
effect of grain source and grain processing on performance of
feedlot cattle: a review. J. Anim. Sci. 75: 868-879.
28. Massover, W. H. and J. M. Cowley (1973) The ultrastructure of fer-
ritin macromolecules, The lattice structure of the core crystallites.
Proc. Natl. Acad. Sci. USA 70: 3847-3851.
29. Banyard, S. H., D. K. Stammers, and P. M. Harrison (1980) Elect-
ron density map of apoferritin at 2.8-A resolution. Nature 288: 298-
300.
30. Yang, C. Y., A. Meagher, B. H. Huynh, D. E. Sayers, and E. C.
Theil (1986) Iron(III) clusters bound to horse spleen apoferritin: An
X-ray absorption and Mossbauer spectroscopy study that shows
that iron nuclei can form on the protein. Biochem. 26: 497-503.
31. Sarvas, M., C. R. Harwood, S. Bron, and J. M. van Dijl (2004) Post-
translocational folding of secretory proteins in gram positive bacte-
ria. Biochim. Biophys. Acta. 1694: 311-327.
32. Schallmey, M., A. Singh, and O. P. Ward (2004) Developments in
the use of Bacillus species for industrial production. Can. J. Micro-
biol. 50: 1-17.