uptake of α-aminoisobutyric acid in pig spleen lymphocytes stimulated by sodium periodate
TRANSCRIPT

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS Vol. 187, No. 2, April 30, pp. 346-354, 1978
Uptake of cr-Aminoisobutyric Acid in Pig Spleen Lymphocytes Stimulated by Sodium Periodate
SOON-HO LEE, HIDEHIKO NAKAGAWA, VIJAY K. KALRA, AND
ARNOLD F. BRODIE
Department of Biochemistry, University of Southern California, School of Medicine, Los Angeles, California 90033
Received August 5, 1977; revised October 28, 1977
The effect of sodium periodate on the ability of pig spleen lymphocytes to transport the nonmetabolixable amino acid, o-aminoisobutyric acid, was studied. NaIOd-treated cells exhibited a lowered rate of uptake of a-aminoisobutyric acid in contrast to phytohemaggh- tinin- and concanavalin A-treated cells. However, when periodate-treated cells were prein- cubated with untreated cells for 2 h, the mixed cells exhibited twofold stimulation in the uptake of a-aminoisobutyric acid as compared to untreated cells. The increased uptake of cy-aminoisobutyric acid in mixed cells was due to a change in the V but not in the K,. The observed increased uptake of cu-aminoisobutyric acid in mixed cells was inhibited (24%) by ouabain, although the level of uptake in untreated and NaIOd-treated cells was not affected. Na+,K+-ATPase activity in mixed cells, which was ouabain sensitive, was stimulated 56%. Studies also showed that there was a decrease in the fluorescence polarization (P value) of diphenyl hexatriene in mixed cells (P = 0.21) as compared to untreated cells (P = 0.24). These results demonstrate that NaI04 treatment induces a change in the lymphocyte cell membrane and transport of a-aminoisobutyric acid. Incubation of NaIOd-treated cells with untreated cells is required for the stimulatory effect in the uptake of cr-aminoisobutyric acid, and the stimulation appears to be due to changes in Na+,K+-ATPase activity and membrane fluidity.
Sodium periodate has been shown to in- duce blastogenesis in rat lymph node cells (l), mouse spleen, lymph node, and mouse thymus cells (2,3), rabbit lymph node cells (4), calf lymph node cells (5), and human peripheral blood lymphocytes (6-10). The mechanism of the blastogenesis by NaI04 in the induction of lymphocyte transfor- mation is not known. Novogrodsky and Katchlaski (2) suggested that the NaI04 target site is a glycoprotein complex con- taining sialic acid that yields an aldehyde moiety by oxidation. NaI04 was also re- ported to inactivate the blood group recep- tors on the human erythrocyte surface (ll), thereby altering their antigenicity and the subsequent immune response to them (12). Recently, O’Brien et ul. (8, 9) showed that NaIOd-treated human peripheral blood lymphocytes, when cultured with untreated autologous lymphocytes, resulted in the blastogenic transformation of the untreated
cells as measured by [‘Hlthymidine incor- poration into DNA.
The chemical modification of the lym- phocyte membrane may also affect some biological functions, such as the activity of membrane-bound enzymes and the uptake of solutes. Van den Berg and Betel (13, 14) have shown that the transport of cr-amino- isobutyric acid (AIB)’ in rat lymphocytes was stimulated (two- to sixfold) when lym- phocytes were treated with plant lectins such as phytohemagglutinin (PHA) or con- canavalin A (Con A). These authors sug- gested that the early increase in cr-amino-
’ Abbreviations used: AIB, a-aminoisobutyric acid; TBS, Tris-buffered saline (10 mM Tris buffer, pH 7.4, 0.15 M NaCl); DPH, 1,6-diphenyl-1,3,5-hexatriene; PBS, phosphate-buffered saline (NaCl, 8.0 g; KCl, 0.2 g; Na2HP0,, 1.15 g; KHzPO+ 0.2 g; CaCL, 0.1 g; MgCb, 0.1 g; and water to a final volume of 1 liter); ConA, concanavalin A; PHA, phytohemagglutinin; pCMBS, p-chloromercuribenzenesulfonic acid.
346
0003-9861/78/1872-0346$02.00/O Copyright 0 1978 by Academic Press, Inc. All rights uf reproduction in any form reserved.

UPTAKE OF a-AMINOISOBUTYRIC ACID IN PIG SPLEEN 347
isobutyric acid transport in rat lympho- cytes stimulated with Con A was not caused by a simple interaction of Con A with the lymphocyte membrane; it required the par- ticipation of energy metabolism and protein synthesis but not of RNA or DNA synthesis (15). Mendelsohn et al. (16) also showed an increased uptake of a-aminoisobutyric acid in human lymphocytes treated with phy- tohemagglutinin. It has been shown that insulin (17) stimulates while cortisol (18) inhibits the uptake of cu-aminoisobutyric acid in rat lymphocytes. However, little is known regarding the effect of chemical modification of the lymphocyte membrane upon the transport processes.
The results presented in this communi- cation show that sodium periodate treat- ment of pig spleen lymphocytes results in a decreased level of uptake of a-aminoiso- butyric acid. However, incubation of so- dium periodate-treated lymphocytes with untreated autologous lymphocytes as com- pared to NaIOA-treated or untreated cells alone resulted in an increased level of up- take of AIB and also in an increased fluidity of the membrane as measured by the fluo- rescence polarization of diphenyl hexa- triene.
MATERIALS AND METHODS
Preparation of lymphocytes. The pig spleens were minced in Tris-buffered saline (10 mM Tris-buffer, pH 7.4, containing 0.15 M NaCl; TBS) and passed through a 200-mesh sieve. The isolated lymphocytes were washed three times with the same buffer by centrifu- gation at 300g for 10 min. The cell suspensions were layered over a 2:l mixture of 9% (w/v) Ficoll (Phar- macia Fine Chemicals) and 29.9% (v/v) Hypaque (Winthrop Laboratories, N.Y.) and centrifuged at 300g for 30 min. The cells at the interface were collected, resuspended, and washed twice with TBS. The aver- age viability was determined by trypan blue dye exclu- sion, and the lymphocytes were counted in a standard hemocytometer. Aliquots of the suspension were cen- trifuged to obtain the desired number of lymphocytes for experiments.
Treatment of lymphocytes with NaIO4. Lympho- cytes were suspended in TBS containing 1 mM NaI04 (or other concentrations as specified in the tables and figures) and incubated at 4°C for 30 min. The NaIOh- treated cells were removed by centrifugation and washed with TBS. For preparation of mixed cells, NaIOd-treated lymphocytes and untreated autologous lymphocytes were mixed in a 1:2 proportion in
Hanks-Eagle medium containing 15% calf serum and incubated at 37°C for 2 h, or for other specified periods where indicated. The final concentration of cells in the incubation mixture was 4 to 6 x 10s cells/ml. After the incubations, mixed cells were washed with 5 vol of TBS.
Determination of AIB uptake. The TBS-washed cells were resuspended in 0.5 ml of Kreb’s buffer (containing 3 mM phosphate buffer, pH 7.2, 0.13 M
NaCl, 5 mM KCl, 1.25 mM CaCh, 1 mM MgClz, and 5.5 mM glucose). Amino acid uptake was measured by the addition of 0.5 pCi/rnl of [‘4C]AIB (final concentration, 0.5 mM) and incubation was carried out at 37°C. At indicated time intervals, aliquots of the cell suspension were removed and immediately diluted in 3.0 ml of ice-cold TBS. The suspension was rapidly filtered on a glass-fiber filter (Whatman, GF/A). The dilution and filtration took less than 10 s for completion. After washing the filters twice with the same buffer, the filter was dried and the radioactivity retained by the filters was counted in 10 ml of Bray’s solution in a Nuclear Chicago liquid scintillation counter.
Estimation of intracellular fluid space. The intra- cellular fluid space of the lymphocytes was calculated from the differences between the total 3HzO-permea- ble space and the dextran [“Clcarboxyl impermeable space (19).
Measurement of membrane fluidity. The fluores- cent probe used in this study, l,&diphenyl-1,3,5-hex- atriene (DPH), has been established as a probe for the hydrocarbon region of the lipid layer of biological membranes and liposome system (20, 21). Lympho- cytes (2 X lo6 cells) were incubated with 2 ml of labeling solution (2 PM DPH dispersion) in phosphate- buffered saline, pH 7.4 (PBS). After 60 min of incu- bation at 25°C cells were washed twice with TBS to remove excess probe and resuspended in 2.5 ml of TBS. Unlabeled control cells were prepared by incu- bating cells with a solution of tetrahydrofuran in PBS (1:lOOO) under the same conditions.
Fluorescence polarization was measured with an MPF-4 Perkin-Elmer spectrofluorometer equipped with polarizers and a Thermostatic cell holder. Exci- tation was carried out at a 360-nm wavelength while the emission maximum was set at 426 nm. The exci- tation was carried out for a 10-s period in order to reduce the reversible photoisomerization process. The samples were stirred with a plastic stirring rod before taking the polarization values in order to obtain a uniform distribution of cells. The Thermostatic bath (Haake) was used to control the temperature of the cuvette. The temperature was recorded in the cuvette using a Thermistor probe. The fluorescence polariza- tion value (P) was calculated from the following equa- tion (22, 23):
p = L - (Ih” x LhlZhh) 1”” + (zhv x kh/lhh)
348 LEE ET AL.
where I is the corrected fluorescence and the sub- scripts v and h indicate vertical and horizontal ori- entation of the excitation and analyzer polarizers, re- spectively. The corrected fluorescence (4 was ob- tained by subtracting the intensity of light obtained with unlabeled control cells from the observed inten- sity of the labeled cells. In this manner the error due to the scattering of the polarized excited light was minimized. The fluorescence polarization measure- ment has an error of *6%.
Assay of Na+,K+-ATPase actiuity. ATPase activity was carried out in a reaction mixture containing sam- ple solution (0.5 to 1.0 mg of protein), 10 mM Tris-HCl, pH 7.4,0.15 M NaCl, 0.15 M KCl, 5 mM MgC12, 10 mM ATP, and water to a final volume of 1.0 ml. To estimate ouabain inhibition, the samples were prein- cubated with 1 mM ouabain for 10 min at 37°C before the addition of ATP. The reaction was terminated by the addition of 1.0 ml of 10% trichloroacetic acid. Released inorganic phosphate was estimated by the method of Fiske and SubbaRow (24).
Protein estimation. Protein concentration was de- termined by the method of Lowry et al. (25), with bovine serum albumin as standard.
Other materials. [‘%]AIB, 3Hz0, and dextran [‘%]carboxyl were purchased from New England Nu- clear Corp.; NaI04, AIB, and Con A from Sigma Chem- ical Co.; Hanks-Eagle medium was from Microbiolog- ical Associates; phytohemagglutinin and calf serum were obtained from Difco Labs. All other chemicals were of reagent grade.
RESULTS
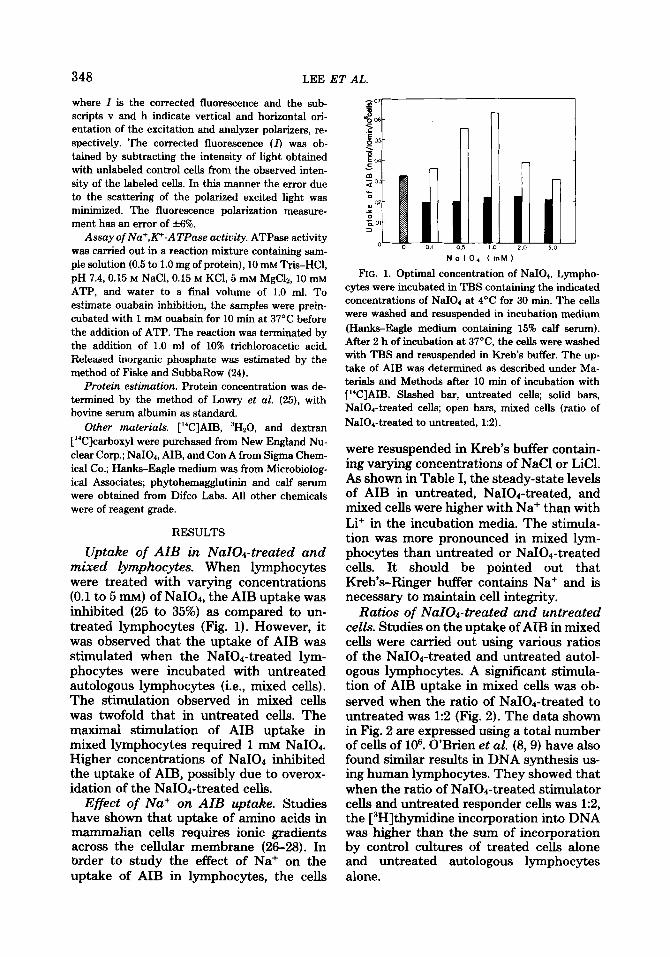
Uptake of AIB in NaIOd-treated and mixed lymphocytes. When lymphocytes were treated with varying concentrations (0.1 to 5 mu) of NaI04, the AIB uptake was inhibited (25 to 35%) as compared to un- treated lymphocytes (Fig. 1). However, it was observed that the uptake of AIB was stimulated when the NaIOd-treated lym- phocytes were incubated with untreated autologous lymphocytes (i.e., mixed cells). The stimulation observed in mixed cells was twofold that in untreated cells. The maximal stimulation of AIB uptake in mixed lymphocytes required 1 mM NaI04. Higher concentrations of NaI04 inhibited the uptake of AIB, possibly due to overox- idation of the NaIOr-treated cells.
Effect of Na+ on AIB uptake. Studies have shown that uptake of amino acids in mammalian cells requires ionic gradients across the cellular membrane (26-28). In brder to study the effect of Na+ on the uptake of AIB in lymphocytes, the cells
NulO, (mh4)
FIG. 1. Optimal concentration of NaI04. Lympho- cytes were incubated in TBS containing the indicated concentrations of NaIOd at 4’C for 30 min. The cells were washed and resuspended in incubation medium (Hanks-Eagle medium containing 15% calf serum). After 2 h of incubation at 37”C, the cells were washed with TBS and resuspended in Kreb’s buffer. The up- take of AIB was determined as described under Ma- terials and Methods after 10 min of incubation with [“CIAIB. Slashed bar, untreated cells; solid bars, NaIOd-treated cells; open bars, mixed cells (ratio of
NaIOd-treated to untreated, 1:2).
were resuspended in Kreb’s buffer contain- ing varying concentrations of NaCl or LiCl. As shown in Table I, the steady-state levels of AIB in untreated, NaIO&reated, and mixed cells were higher with Na+ than with Li+ in the incubation media. The stimula- tion was more pronounced in mixed lym- phocytes than untreated or NaIOr-treated cells. It should be pointed out that Kreb’s-Ringer buffer contains Na+ and is necessary to maintain cell integrity.
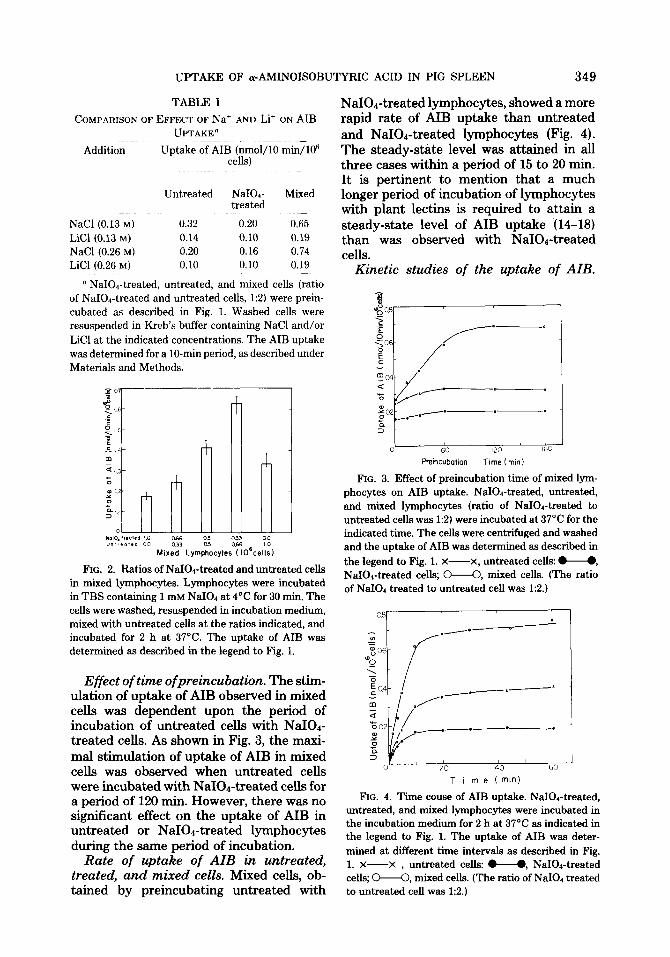
Ratios of NaIO.&reated and untreated cells. Studies on the uptake of AIB in mixed cells were carried out using various ratios of the NaIOr-treated and untreated autol- ogous lymphocytes. A significant stimula- tion of AIB uptake in mixed cells was ob- served when the ratio of NaIOd-treated to untreated was 1:2 (Fig. 2). The data shown in Fig. 2 are expressed using a total number of cells of 106. O’Brien et al. (8,9) have also found similar results in DNA synthesis us- ing human lymphocytes. They showed that when the ratio of NaIOd-treated stimulator cells and untreated responder cells was 1:2, the [3H]thymidine incorporation into DNA was higher than the sum of incorporation by control cultures of treated cells alone and untreated autologous lymphocytes alone.
UPTAKE OF (u-AMINOISOBUTYRIC ACID IN PIG SPLEEN 349
TABLE I
COMPARISONOFEFFIXTOF Na+ AND Li+ ONAIB UPTAKE"
Addition Uptake of AI~e(;iol/lO min/lb
Untreated NaI04- Mixed treated
-~ NaCl (0.13 M) 0.32 0.20 0.65 Licl (0.13 M) 0.14 0.10 0.19 NaCl (0.26 M) 0.20 0.16 0.74 LiCl (0.26 M) 0.10 0.10 0.19
a NaI04-treated, untreated, and mixed cells (ratio of NaIOd-treated and untreated cells, 1:2) were prein- cubated as described in Fig. 1. Washed cells were resuspended in Kreb’s buffer containing NaCl and/or
LiCl at the indicated concentrations. The AIB uptake was determined for a lo-min period, as described under Materials and Methods.
Mixed Lymphocytes ( IO’cells)
FIG. 2. Ratios of NaIO&reated and untreated cells in mixed lymphocytes. Lymphocytes were incubated in TBS containing 1 mM NaI04 at 4°C for 30 min. The cells were washed, resuspended in incubation medium, mixed with untreated cells at the ratios indicated, and incubated for 2 h at 37°C. The uptake of AIB was determined as described in the legend to Fig. 1.
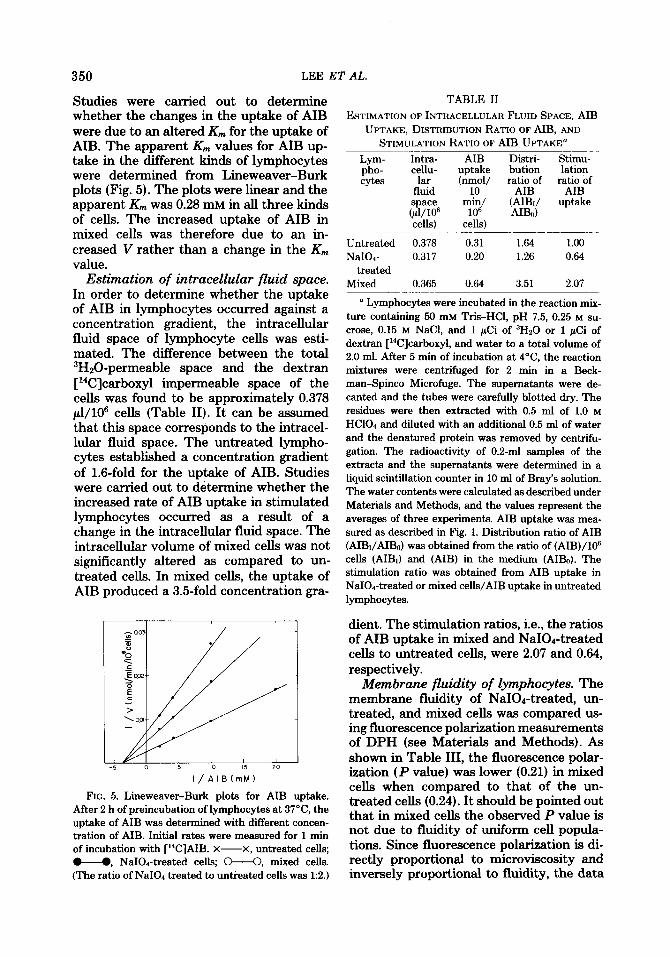
Effect of time ofpreincubation. The stim- ulation of uptake of AIB observed in mixed celIs was dependent upon the period of incubation of untreated cells with NaIOd- treated cells. As shown in Fig. 3, the maxi- mal stimulation of uptake of AIB in mixed cells was observed when untreated cells were incubated with NaIO1-treated cells for a period of 120 min. However, there was no significant effect on the uptake of AIB in untreated or NaIOd-treated lymphocytes during the same period of incubation.
Rate of uptake of AIB in untreated, treated, and mixed cells. Mixed cells, ob- tained by preincubating untreated with
NaIOr-treated lymphocytes, showed a more rapid rate of AIB uptake than untreated and NaIOd-treated lymphocytes (Fig. 4). The steady-state level was attained in all three cases within a period of 15 to 20 min. It is pertinent to mention that a much longer period of incubation of lymphocytes with plant lectins is required to attain a steady-state level of AIB uptake (14-18) than was observed with NaIOb-treated cells.
Kinetic studies of the uptake of AIB.
1 I c 6C IX ItiC
Freincubotion Time (min)
FIG. 3. Effect of preincubation time of mixed lym- phocytes on AIB uptake. NaIOa-treated, untreated, and mixed lymphocytes (ratio of NaIOd-treated to untreated cells was 1:2) were incubated at 37°C for the indicated time. The cells were centrifuged and washed and the uptake of AIB was determined as described in
the legend to Fig. 1. X-X, untreated cells: u, NaIOd-treated cells; M, mixed cells. (The ratio of NaI04 treated to untreated cell was 1:2.)
T i me (mln)
FIG. 4. Time cause of AIB uptake. NaIOI-treated, untreated, and mixed lymphocytes were incubated in the incubation medium for 2 h at 37°C as indicated in the legend to Fig. 1. The uptake of AIB was deter-
mined at different time intervals as described in Fig. 1. X-X , untreated cella w, NaIOd-treated cells; M, mixed cells. (The ratio of NaI04 treated to untreated cell was 1:2.)
350 LEE ET AL.
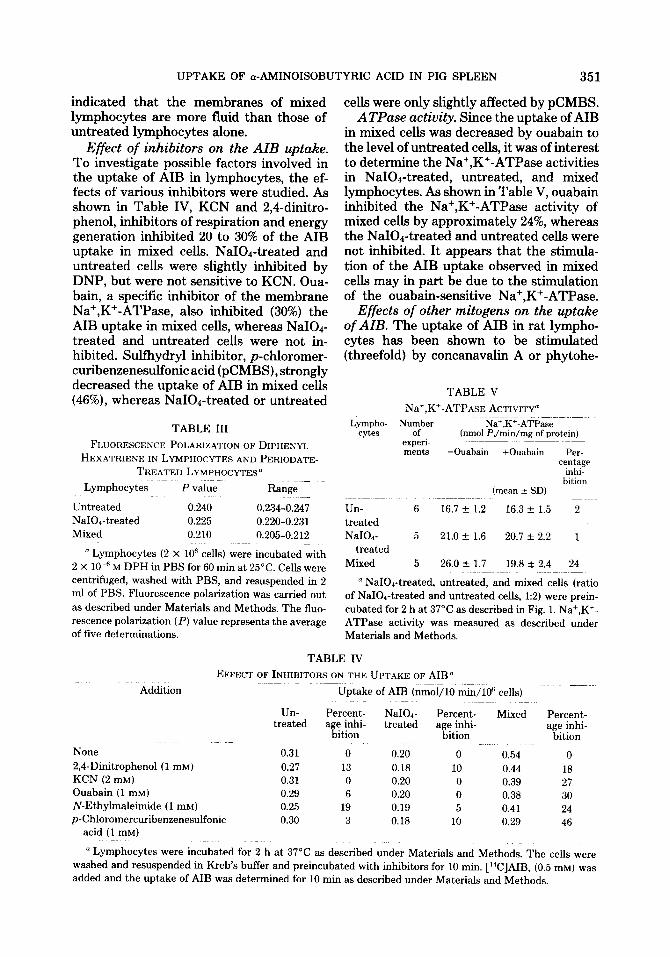
Studies were carried out to determine whether the changes in the uptake of AIB were due to an altered K, for the uptake of AIB. The apparent K, values for AIB up- take in the different kinds of lymphocytes were determined from Lineweaver-Burk plots (Fig. 5). The plots were linear and the apparent K, was 0.28 mM in all three kinds of cells. The increased uptake of AIB in mixed cells was therefore due to an in- creased V rather than a change in the K,,, value.
Estimation of intracellular fluid space. In order to determine whether the uptake of AIB in lymphocytes occurred against a concentration gradient, the intracellular fluid space of lymphocyte cells was esti- mated. The difference between the total 3HzO-permeable space and the dextran [‘4C]carboxyl impermeable space of the cells was found to be approximately 0.378 $/lo6 cells (Table II). It can be assumed that this space corresponds to the intracel- lular fluid space. The untreated lympho- cytes established a concentration gradient of 1.6-fold for the uptake of AIB. Studies were carried out to determine whether the increased rate of AIB uptake in stimulated lymphocytes occurred as a result of a change in the intracellular fluid space. The intracellular volume of mixed cells was not significantly altered as compared to un- treated cells. In mixed cells, the uptake of AIB produced a 3.5-fold concentration gra-
I/AIB(mM)
FIG. 5. Lineweaver-Burk plots for AIB uptake. After 2 h of preincubation of lymphocytes at 37°C the uptake of AIB was determined with different concen- tration of AIB. Initial rates were measured for 1 min of incubation with [W]AIB. X-X, untreated cells; M, NaIOd-treated cells; W, mixed cells. (The ratio of NaI04 treated to untreated cells was 1:2.)
TABLE II
ESTIMATION OF INTRACELLULAR FLUID SPACE, AIB UPTAKE, DISTRIBUTION RATIO OF AIB, AND
STIMULATION RATIO OF AIB UPTAKE=
yh:: Intra- AIB Distri- Stimu- cellu- untake bution lation
cytes lar (nmol/ ratio of ratio of fluid 10 AIB AIB space min/ (ul/lO~ lofi a”:‘:
uptake 0
cells) cells)
Untreated 0.378 0.31 1.64 1.00 NaIOd- 0.317 0.20 1.26 0.64
treated Mixed 0.365 0.64 3.51 2.07
D Lymphocytes were incubated in the reaction mix- ture containing 50 mM Tris-HCl, pH 7.5, 0.25 M SU-
crose, 0.15 M NaCl, and 1 pCi of 3Hs0 or 1 PCi of dextran [i4C]carboxyl, and water to a total volume of 2.0 ml. After 5 min of incubation at 4”C, the reaction mixtures were centrifuged for 2 min in a Beck- man-Spinco Microfuge. The supernatants were de- canted and the tubes were carefully blotted dry. The residues were then extracted with 0.5 ml of 1.0 M HClOd and diluted with an additional 0.5 ml of water and the denatured protein was removed by centrifu- gation. The radioactivity of 0.2-ml samples of the extracts and the supernatants were determined in a liquid scintillation counter in 10 ml of Bray’s solution. The water contents were calculated as described under Materials and Methods, and the values represent the averages of three experiments. AIB uptake was mea- sured as described in Fig. 1. Distribution ratio of AIB (AIBi/AIBs) was obtained from the ratio of (AIB)/106 cells (AIBI) and (AIB) in the medium (AI&). The stimulation ratio was obtained from AIB uptake in NaIOd-treated or mixed cells/AIB uptake in untreated lymphocytes.
dient. The stimulation ratios, i.e., the ratios of AIB uptake in mixed and NaIOd-treated cells to untreated cells, were 2.07 and 0.64, respectively.
Membrane fluidity of lymphocytes. The membrane fluidity of NaI04-treated, un- treated, and mixed cells was compared us- ing fluorescence polarization measurements of DPH (see Materials and Methods). As shown in Table III, the fluorescence polar- ization (P value) was lower (0.21) in mixed cells when compared to that of the un- treated cells (0.24). It should be pointed out that in mixed cells the observed P value is not due to fluidity of uniform cell popula- tions. Since fluorescence polarization is di- rectly proportional to microviscosity and inversely proportional to fluidity, the data
UPTAKE OF a-AMINOISOBUTYRIC ACID IN PIG SPLEEN 351
indicated that the membranes of mixed lymphocytes are more fluid than those of untreated lymphocytes alone.
Effect of inhibitors on the AIB uptake. To investigate possible factors involved in the uptake of AIB in lymphocytes, the ef- fects of various inhibitors were studied. As shown in Table IV, KCN and 2,4-dinitro- phenol, inhibitors of respiration and energy generation inhibited 20 to 30% of the AIB uptake in mixed cells. NaIOd-treated and untreated cells were slightly inhibited by DNP, but were not sensitive to KCN. Oua- bain, a specific inhibitor of the membrane Na+,K+-ATPase, also inhibited (30%) the AIB uptake in mixed cells, whereas NaIOd- treated and untreated cells were not in- hibited. Sulfhydryl inhibitor, p-chloromer- curibenzenesulfonic acid (pCMBS), strongly decreased the uptake of AIB in mixed cells (46%), whereas NaI04-treated or untreated
TABLE III
FI.UOHESCENCF: POLARIZATION OF DIPHENYI,
HEXATRIENE IN LYMPHOCYTES AND PERIODATE- TREATED LYMPHOCYTES~
Lymphocytes P value Range
Untreated 0.240 0.234-0.247 NaIOd-treated 0.225 0.220-0.231 Mixed 0.210 0.205-0.212
” Lymphocytes (2 X 10fi cells) were incubated with 2 X 10efi M DPH in PBS for 60 min at 25°C. Cells were centrifuged, washed with PBS, and resuspended in 2 ml of PBS. Fluorescence polarization was carried out as described under Materials and Methods. The flue- rescence polarization (P) value represents the average of five determinations.
cells were only slightly affected by pCMBS. ATPase activity. Since the uptake of AIB
in mixed cells was decreased by ouabain to the level of untreated cells, it was of interest to determine the Na+,K+-ATPase activities in NaIO.&reated, untreated, and mixed lymphocytes. As shown in Table V, ouabain inhibited the Na+,K+-ATPase activity of mixed cells by approximately 24%, whereas the NaIO.+-treated and untreated cells were not inhibited. It appears that the stimula- tion of the AIB uptake observed in mixed cells may in part be due to the stimulation of the ouabain-sensitive Na+,K+-ATPase.
Effects of other mitogens on the uptake of AIB. The uptake of AIB in rat lympho- cytes has been shown to be stimulated (threefold) by concanavahn A or phytohe-
TABLE V
Na’,K’-ATPAsE ACTIVITY”
Lympho- Number Na+,K+-ATPase cytes of (nmol P,/min/mg of protein)
experi- ~ ~~~ merits -0uabain +Ouabain Per-
centage inhi-
bition (mean + SD)
Un- 6 16.7 + 1.2 16.3 f 1.5 2 treated NaIOd- 5 21.0 + 1.6 20.7 -c 2.2 1
treated
Mixed 5 26.0 + 1.7 19.8 + 2.4 24
” NaIOd-treated, untreated, and mixed cells (ratio of NaI04-treated and untreated cells, 1:2) were prein- cubated for 2 h at 37°C as described in Fig. 1. Na+,K+- ATPase activity was measured as described under Materials and Methods.
Addition
TABLE IV
EFFECT OF INHIBITORS ON THE UPTAKE OF AIB”
Uptake of AIB (nmol/lO min/l@ cells)
Un- Percent- NaIOd- Percent- Mixed treated age inhi- treated
bition age inhi-
bition
Percent- age inhi-
bition
None
2,4-Dinitrophenol (1 mM) KCN (2 mM) Ouabain (1 mM)
N-Ethylmaleimide (1 mM) p-Chloromercuribenzenesulfonic
acid (1 mM)
0.31 0 0.20 0 0.54 0
0.27 13 0.18 10 0.44 18 0.31 0 0.20 0 0.39 27 0.29 6 0.20 0 0.38 30 0.25 19 0.19 5 0.41 24 0.30 3 0.18 10 0.29 46
” Lymphocytes were incubated for 2 h at 37°C as described under Materials and Methods. The cells were washed and resuspended in Kreb’s buffer and preincubated with inhibitors for 10 min. [‘%]AIB, (0.5 mM) was added and the uptake of AIB was determined for 10 min as described under Materials and Methods.
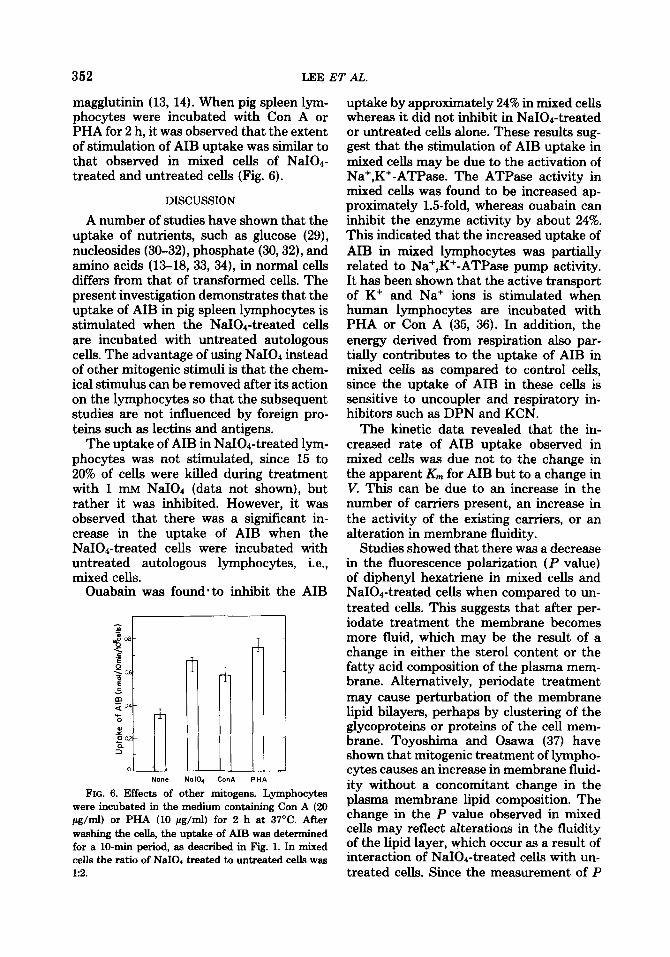
352 LEE ET AL.
magglutinin (13, 14). When pig spleen lym- phocytes were incubated with Con A or PHA for 2 h, it was observed that the extent of stimulation of AIB uptake was similar to that observed in mixed cells of NaI04- treated and untreated cells (Fig. 6).
DISCUSSION
A number of studies have shown that the uptake of nutrients, such as glucose (29), nucleosides (30-32)) phosphate (30,32), and amino acids (13-18, 33, 34), in normal cells differs from that of transformed cells. The present investigation demonstrates that the uptake of AIB in pig spleen lymphocytes is stimulated when the NaIOd-treated cells are incubated with untreated autologous cells. The advantage of using NaI04 instead of other mitogenic stimuli is that the chem- ical stimulus can be removed after its action on the lymphocytes so that the subsequent studies are not influenced by foreign pro- teins such as lectins and antigens.
The uptake of AIB in NaIOr-treated lym- phocytes was not stimulated, since 15 to 20% of cells were killed during treatment with 1 mu NaI04 (data not shown), but rather it was inhibited. However, it was observed that there was a significant in- crease in the uptake of AIB when the NaIO&reated cells were incubated with untreated autologous lymphocytes, i.e., mixed cells.
Ouabain was found* to inhibit the AIB
Ncmc NaIO+ ConA PHA
FIG. 6. Effects of other mitogens. Lymphocytes were incubated in the medium containing Con A (20 pg/mI) or PHA (10 pg/mI) for 2 h at 37°C. After washing the cells, the uptake of AIB was determined for a lO-min period, as described in Fig. 1. In mixed ceils the ratio of NaIOd treated to untreated ceIIs was 1:2.
uptake by approximately 24% in mixed cells whereas it did not inhibit in NaI04-treated or untreated cehs alone. These results sug- gest that the stimulation of AIB uptake in mixed cells may be due to the activation of Na+,K+-ATPase. The ATPase activity in mixed cells was found to be increased ap- proximately L&fold, whereas ouabain can inhibit the enzyme activity by about 24%. This indicated that the increased uptake of AIB in mixed lymphocytes was partially related to Na+,K+-ATPase pump activity. It has been shown that the active transport of K+ and Na+ ions is stimulated when human lymphocytes are incubated with PHA or Con A (35, 36). In addition, the energy derived from respiration also par- tially contributes to the uptake of AIB in mixed cells as compared to control cells, since the uptake of AIB in these cells is sensitive to uncoupler and respiratory in- hibitors such as DPN and KCN.
The kinetic data revealed that the in- creased rate of AIB uptake observed in mixed cells was due not to the change in the apparent K,,, for AIB but to a change in V. This can be due to an increase in the number of carriers present, an increase in the activity of the existing carriers, or an alteration in membrane fluidity.
Studies showed that there was a decrease in the fluorescence polarization (P value) of diphenyl hexatriene in mixed cells and NaIOr-treated cells when compared to un- treated cells. This suggests that after per- iodate treatment the membrane becomes more fluid, which may be the result of a change in either the sterol content or the fatty acid composition of the plasma mem- brane. Alternatively, periodate treatment may cause perturbation of the membrane lipid bilayers, perhaps by clustering of the glycoproteins or proteins of the cell mem- brane. Toyoshima and Osawa (37) have shown that mitogenic treatment of lympho- cytes causes an increase in membrane fluid- ity without a concomitant change in the plasma membrane lipid composition. The change in the P value observed in mixed cells may reflect alterations in the fluidity of the lipid layer, which occur as a result of interaction of NaIOd-treated cells with un- treated cells. Since the measurement of P

UPTAKE OF cr-AMINOISOBUTYRIC ACID IN PIG SPLEEN 353
value was not carried out on uniform cell populations but on a mixture of treated and untreated cells, it is difficult to ascertain the contribution of each population of cells to the fluidity of the membrane. Although changes in the fluidity and the stimulation of AIB uptake occur in mixed cells, the exact relationship between the two is not clear.
Some possible mechanisms have been proposed for the NaIOd-induced transfor- mation of lymphocytes. Novogrodsky and Katchlaski (2) suggested that NaI04 in- duces a specific surface conformational change in membrane glycoproteins, as dis- cussed earlier. O’Brien et al. (8, 9) have speculated that the NaIOd-treated cells might be recognized by autologous lympho- cytes as foreign and that the production of aldehyde groups on the surface of treated cells is responsible for the transformation of untreated cells by cellular interaction or intracellular contacts. Recently, Monlian et al. (10) demonstrated that a soluble growth factor was released into the medium from NaI04-treated human peripheral blood lymphocytes. This lymphocyte growth fac- tor can stimulate DNA synthesis when added to native autologous cells. It is per- tinent to mention that O’Brien et al. (8, 9) failed to find a mitogenic factor released into the medium by NaIOd-treated lympho- cytes. The results presented in this paper, namely, the early stimulation of AIB up- take in pig lymphocytes in mixed NaIOd- treated and untreated cells appears to be mediated by cell-cell contact since the sol- uble mitogenic factor is released into the media after long periods (96 to 120 h) of incubation of lymphocytes with periodate (10).
ACKNOWLEDGMENTS
We are pleased to acknowledge the supply of pig spleens from Clougherty Packing Company. We also thank Drs. J. W. Parker, and R. L. O’Brien, for reading and criticizing the manuscript prior to submission for publication.
This work was supported by Grants from the Na- tional Institutes of Health, United States Public Health Service (AI 05637), the National Science Foun- dation (GB 32351 X), and the Hastings Foundation at the University of Southern California, School of Med- icine. Work by S. H. Lee, was done during the tenure
of a Cancer Research Training Fellowship under NIH Grant CA 05297 at the University of Southern Cali- fornia, School of Medicine.
REFERENCES
1. NOVOGRODSKY, A., AND KATCHLASKI, E. (1971) FEBS Lett. 12.297-300.
2. NOVOGRODSKY, A., AND KATCHLASKI, E. (1972) Proc. Nat. Acad. Sci. USA 69.3207-3210.
3. NOVODRODSKY, A., AND GERY, I. (1972) J. Zm- munol. 109, 1278-1281.
4. ONO, M., AND HOGUMI, M. (1973) Biochem. Bio- phys. Res. Commun. 53.342-349.
5. PRESANT, C. A., AND PARKER, S. (1976) J. Biol. Chem. 251, 1864-1870.
6. PARKER, J. W., O’BRIEN, R. L., LUKES, R. J., AND STEINER, J. (1972) Lancet 1, 103-104.
7. PARKER, J. W., O’BRIEN, R. L., STEINER, J., AND PAOLILLI, P. (1973) Exp. Cell Res. 78, 279-286.
8. O’BRIEN, R. L., PARKER, J. W., PAOI,ILLI, P., AND STEINER, J. (1974) in Proceedings of the 8th Leuc. Culture Conference, pp. 19-24.
9. O’BRIEN, R. L., PARKER, J. W., PAOLILLI, P., AND STEINER, J. (1974) J. Immunol. 112,1884-1889.
10. MONALIAN, T. M., FROST, A. F., AND ABELL, C. W. (1976) Biochem. Biophys. Res. Commun. 73, 1115-1121.
11. MORGAN, W. T. J., AND WATKINS, W. M. (1951)
Brit. J. Exp. Pathol. 32, 34-48. 12. STEWART, F. S. (1949) J. Pathol. Bacterial. 61,
456-458. 13. VAN DEN BERG, K. J., AND BETEL, I. (1971) Exp.
Cell Res. 66, 257-259. 14. VAN DEN BERG, K. J., AND BETEL, I. (1973) Exp.
Cell Res. 76,63-72. 15. VAN DEN BERG, K. J., AND BETEL, I. (1974) Exp.
Cell Res. 84,412-418. 16. MENDELSOHN, J., SKINNER, S. A., AND KORN-
FELD, S. (1971) J. Clin. Znuest. 50.818-826. 17. GOLDFINE, I. D., GARDNER, J. D., AND NEVILLE,
D. M. JR. (1972) J. Biol. Chem. 241,6919-6926. 18. BARAN, D. T., PECK, W. A., FRENGLEY, P. A.,
AND LICHTMAN, M. A. (1973) Biochim. Biophys. Acta 307,627-639.
19. HUNTER, G. R., AND BRIERLEY, G. P. (1969) B&him. Biophys. Acta 180.68-80.
20. SHINITZKI, M., AND INBAR, M. (1974) J. MOE. Biol. 85, 603-615.
21. SHINITZKI, M., AND BARENHOLZ, T. (1974) J. Biol. Chem. 242,2652-2657.
22. AZUMI, T., AND MCGLEEN, S. P. (1962) J. Chem. Phys. 37, 2413-2420.
23. CHEN, R. F., AND BOWMAN, R. L. (1965) Science 147, 729-732.
24. FISKE, C. H., AND SUBBAROW, Y. (1925) J. Biol. Chem. 66,375-W.
25. LOWRY, 0. H., ROSEBROUGH, N. J., FARR, A. L., AND RANDALL, R. J. (1951) J. Biol. Chem. 193, 265-275.

354 LEE ET AL.
26. CHRISTENSEN, H. N. (1970) in Membranes and 32. ROSENGURT, E., AND JIMENEZ DE ASUA, L. (1973) Ion Transport (Bittar, E. E., ed.), Vol. 1, pp. Proc. Nat. Acad. Sci. USA 70.3609-3612. 365-394, Wiley-Interscience, London.
27. OXENDER, D. L. (1972) Annu. Rev. B&hem. 41,
777-814. 28. SCHULTZ, S. G., AND CURRAN, P. F. (1970) Phys-
iol. Rev. 60, 637-718. 29. HATANAKA, M. (1974) Biochim. Biophys. Acta
355.77-104.
33.
34.
35.
FOSTER, D. O., AND PARDEE, A. B. (1969) J. Biol. Chem. 244,2675-2681.
ISSELBACHER, K. J. (1972) Proc. Nat. Acad. Sci. USA 69,565~589.
QUASTEL, M. R., AND KAPLAN, J. G. (1970) Exp. Cell Res. 63,230-233.
30. CUNNINGHAM, D. D., AND PARDEE, A. B. (1969) 36. AVERDUNK, R., AND LAUF, P. K. (1975) Exp. Cell Proc. Nat. Acad. Sci. USA 64, 1049-1056. Res. 93.331-342.
31. QUINLAN, D. C., AND HOCHSTADT, S. (1974) Proc. 37. TOYOSHIMA, S., AND OSAWA, T. (1976) Erp. Cell Nat. Acad. Sci. USA 71,5000-5003. Res. 102.438-441.