venoms-based discovery of novel modulators of human ...381883/s4305838_mphil... · institute for...
TRANSCRIPT

Venoms-based discovery of novel modulators of human neuronal α7 and
α3* nicotinic acetylcholine receptors
Rosa Esther Prahl
MSc. Cell Biology
A thesis submitted for the degree of Master of Philosophy at
The University of Queensland in 2016
Institute for Molecular Bioscience

2
ABSTRACT Nicotinic acetylcholine receptors (nAChRs) are pentameric ligand-gated channels
that contain combinations of α and β subunits (heteromeric) or only five α subunits
(homomeric). To date, 12 subunits have been cloned. Although different types of
neuronal nAChRs are expected to occur in vivo, most structure-activity relationship
studies have been carried out for just a few neuronal subtypes. Clinical and basic
research has shown that cholinergic receptors play a role in several disorders of
the nervous system such as chronic pain, Alzheimer’s disease and addiction to
nicotine, alcohol and drugs. Unfortunately, the lack of selective modulators for
each subtype of nAChR makes their pharmacological characterization difficult,
which has slowed the development of therapeutic nAChR modulators with high
selectivity and absence of off-target side effects.
Animal venoms have proven to be an excellent natural source of bioactive
molecules with activity against ion channels. Venoms from different snakes and in
particular molluscs of the Conus genus have been an important source of
modulators of cholinergic receptors. However, spider and scorpion venoms have
been studied less extensively and there are only two examples of cholinergic
receptor modulators reported from arachnid venom. Thus, the primary goal of this
thesis was to identify novel modulators of the human ganglionic α3* nAChR
(where the asterisk indicates that β subunits form part of the heteropentameric
receptor) and the homomeric neuronal α7 nAChR using spider and scorpion
venoms.
“Hit” venoms from a selection of “modern” and “primitive” spiders were identified
via high-throughput screens against the α3* and α7 nAChRs endogenously
expressed in SH-SY5Y cells. A total of 71 spider venoms were screened, with
representatives from nine different taxonomic families. The resulting data showed
that 91% of the venoms presented some activity against the nAChR subtypes
tested and most of these were non-selective blockers of the α3* and α7 subtypes.
Toxins directed against vertebrate nAChRs likely evolved through frequent
interactions between spiders and small vertebrate predators. Our data revealed
that nAChR agonists are likely to be present in venom of the “primitive” spiders

3
and therefore were probably basally recruited in spider venom. In summary, the
screening of spider venoms against vertebrate cholinergic receptors provided
information about the origin and evolution of toxins against vertebrate receptors.
The search for nAChR modulators was extended to scorpion venoms that are
recognized as sources of toxins that affect primarily the properties of voltage-gated
sodium and potassium channels. In this work, venoms from Androctonus australis,
Androctonus crassicauda, Leirurus quinquestriatus and Grosphus grandidieri
(Buthidae) and Heterometus spinnifer (Scorpionidae) were screened against the
α3* and α7 nAChRs using SH-SY5Y cells. The venom of Heterometrus spinnifer
was found to contain an agonistic compound. Activity guided-fractionation allowed
isolation of the “hit” toxin, and it was identified by high-resolution mass
spectrometry as senecioylcholine. This non-peptide toxin has not previously been
identified in arachnids but it was previously isolated from some marine gastropods
(Muricidae). It shows high toxicity against vertebrates. Thus, this non-peptide toxin
is likely to function in defence against vertebrate predators.

4
DECLARATION BY AUTHOR
This thesis is composed of my original work, and contains no material previously
published or written by another person except where due reference has been
made in the text. I have clearly stated the contribution by others to jointly-authored
works that I have included in my thesis.
I have clearly stated the contribution of others to my thesis as a whole, including
statistical assistance, survey design, data analysis, significant technical
procedures, professional editorial advice, and any other original research work
used or reported in my thesis. The content of my thesis is the result of work I have
carried out since the commencement of my research higher degree candidature
and does not include a substantial part of work that has been submitted to qualify
for the award of any other degree or diploma in any university or other tertiary
institution. I have clearly stated which parts of my thesis, if any, have been
submitted to qualify for another award.
I acknowledge that an electronic copy of my thesis must be lodged with the
University Library and, subject to the policy and procedures of The University of
Queensland, the thesis be made available for research and study in accordance
with the Copyright Act 1968 unless a period of embargo has been approved by the
Dean of the Graduate School.
I acknowledge that copyright of all material contained in my thesis resides with the
copyright holder(s) of that material. Where appropriate I have obtained copyright
permission from the copyright holder to reproduce material in this thesis.

5
PUBLICATIONS DURING CANDIDATURE
No publications.
PUBLICATIONS INCLUDED IN THIS THESIS
No publications.

6
CONTRIBUTION BY OTHERS TO THE THESIS
Prof. Richard Lewis and Dr Irina Vetter (Institute for molecular Bioscience)
supported during the screening of “hit” toxins of the 71 venoms with their expertise
in FLIPR assays.
Chapter 1 Dr Volker Herzig (Institute for Molecular Bioscience) performed HPLC fractionation
of crude venom from Macrothele gigas.
Dr Irina Vetter (Institute for Molecular Bioscience) performed one FLIPR assay that
was mentioned in section 2.3.4.
Chapter 2 HPLC fractionation of crude venom from Heterometrus spinnifer was performed by
Dr Niraj Bende (Institute for Molecular Bioscience).
Dr Angela Salim identified the “hit” toxin from Heterometrus spinnifer as
senecioylcholine using high-resolution electrospray ionization mass spectrometry.
She also performed as well as the co-elution of the native compound with its
synthetic form, and two of its synthetic isomers (tigloylcholine and
angeloylcholine).
Pratik Neupane carried out the chemical synthesis of senecioylcholine,
tigloylcholine and angeloylcholine.
Angela and Pratik were involved in the writing of the protocol for identification and
chemical synthesis of senecioylcholine. Angela and Pratik are members of the
group of Prof. Robert Capon at The Institute for Molecular Bioscience.

7
STATEMENT OF PARTS OF THE THESIS SUBMITTED TO QUALIFY FOR THE AWARD OF
ANOTHER DEGREE
None.

8
ACKNOWLEDGEMENTS
I thank Prof. Glenn King, Prof. Richard Lewis, Prof. Robert Capon, Dr Amanda
Carozzi, Dr Irina Vetter, Dr Angela Salim, Pratik Neupane and Sassan Ranhama
for their time, efforts and suggestions during the course of my thesis.
This work was funding and supported by the following institutions: Postgraduate
studies of The University of Queensland (UQ), the UQ Institute for Molecular
Bioscience, and a program grant from the Australian National Health & Medical
Research Council.
I dedicate this thesis to the Holy Father, my husband Torben and my son Clouseau.

9
KEYWORDS
spider venom, scorpion venom, ligand gated calcium channel, α7 nicotinic
acetylcholine receptors, α3* nicotinic acetylcholine receptors, FLIPR, high
throughput screening, choline ester, senecioylcholine
AUSTRALIAN AND NEW ZEALAND STANDARD RESEARCH CLASSIFICATIONS
(ANZSRC)
ANZSRC code: 060101, Analytical Biochemistry, 80%
ANZSRC code: 060199, Biochemistry and Cell Biology not elsewhere classified,
20%
FIELDS OF RESEARCH (FOR) CLASSIFICATION
FoR code: 0601, Biochemistry and Cell Biology, 80%
FoR code: 0699, Other Biological Sciences, 20%

10
TABLE OF CONTENTS
Abstract ........................................................................................................................ 2
Declaration by author ................................................................................................... 4
Publications during candidature .................................................................................... 5
Publications included in this thesis ................................................................................ 5
Contribution by others to the thesis .............................................................................. 6
Statement of parts of the thesis submitted to qualify for the award of another degree . 7
Acknowledgements ....................................................................................................... 8
Keywords ...................................................................................................................... 9
Australian and New Zealand Standard Research Classifications (ANZSRC) ..................... 9
Fields of Research (FoR) Classification ........................................................................... 9
Table of Contents ......................................................................................................... 10
List of figures and Tables .............................................................................................. 12 List of abbreviations ................................................................................................................................................ 14
CHAPTER 1 — Introduction ........................................................................................... 16 1.1 Nicotinic acetylcholine receptors ........................................................................................................... 17 1.1.1 Homopentameric/heteropentameric architecture of neuronal nAChRs .................... 17 1.1.2 The use of toxins to elucidate the molecular architecture of nAChRs ....................... 19 1.1.3 Location and physiological function of nAChRs in vertebrates and invertebrates 21 1.1.4 Role of nAChRs in human disease ................................................................................................ 24
1.2 Modulators of nAChRs ............................................................................................................................... 26 1.2.1 Modulators of nAChRs as drugs and insecticides ................................................................. 26 1.2.2 Novel subtype-selective modulators of vertebrate nAChRs are required to develop accuracy pharmacological tools and potential therapeutic leads .............................. 28
1.3 Venom-derived modulators of nAChRs ............................................................................................. 32 1.3.1 Important role played by subtype-selective α-conotoxins in study of vertebrate nAChRs .......................................................................................................................................................................... 32 1.3.2 nAChR modulators from other venomous animals ............................................................... 34 1.3.3 Rationale for examining spider venoms as a source of novel nAChR modulators 34
1.4 Significance and aims of the current work ....................................................................................... 35 1.4.1 Significance .................................................................................................................................................. 35 1.4.2 Aims .................................................................................................................................................................. 36
CHAPTER 2 — nAChR modulators from spider venoms .................................................. 37 2.1 Introduction ....................................................................................................................................................... 37 2.2 Methods and Materials ............................................................................................................................... 39 2.2.1 Tissue culture reagents ......................................................................................................................... 39 2.2.2 Spider venoms ........................................................................................................................................... 40 2.2.3 Cell lines and culture .............................................................................................................................. 40 2.2.4 Preparation of the Calcium 4-dye ................................................................................................... 41 2.2.5 High-throughput FLIPR calcium assay ........................................................................................ 42 2.2.6 Activation of α3* nAChR by nicotine ............................................................................................. 43 2.2.7 Activation of α7 nAChR by choline in the presence of the positive allosteric modulator PNU-120596 ........................................................................................................................................ 45

11
2.2.8 Identification of blockers and agonists for α3* and α7 nAChR using the FLIPRTETRA system ................................................................................................................................................... 46 2.2.9 Data analysis ............................................................................................................................................... 49 2.2.10 Statistical Analyses ............................................................................................................................... 49
2.3 Results ................................................................................................................................................................ 50 2.3.1 Screening of 71 spider venoms using the high-throughput FLIPRTETRA system .. 50 2.3.2 Screening of venoms from “modern” spiders (Infraorder Araneomorphae) ........... 50 2.3.3 Screening of venoms from “primitive” spiders (Infraorder Mygalomorphae) ......... 55 2.3.4 Identification of α7/α3* nAChR agonists in the venom of theraphosid spiders .... 55 2.3.5 Venoms that selectively inhibit vertebrate α3* nAChRs .................................................... 58 2.3.6 Venoms that non-selectively block both α3* and α7 nAChRs ....................................... 61 2.3.7 Theraphosid venoms with no apparent effect on nAChRs ............................................... 63 2.3.8 Effect of non-theraphosid mygalomorph venom on vertebrate nAChRs ................. 64 2.3.9 Taxonomic distribution of nAChR modulators within Araneae ....................................... 64
2.4 Discussion ......................................................................................................................................................... 66 CHAPTER 3 — nAChR modulators from scorpion venom ................................................ 75 3.1 Introduction ....................................................................................................................................................... 75 3.1.1 Scorpion toxins and their pharmacological action ................................................................. 75
3.2 Methods .............................................................................................................................................................. 77 3.2.1 Figure 3.1 shows an example of the protocol as applied to scorpion venoms ..... 77 3.2.2 Scorpion venoms ...................................................................................................................................... 80 3.2.3 Venom fractionation ................................................................................................................................ 80 3.2.4 Discovery of a choline ester from the venom of H. spinnifer .......................................... 80 3.2.5 Determination of mass and purity ................................................................................................... 81 3.2.6 Identification of active compound using HRMS and LCMS ............................................. 81 3.2.7 Chemical synthesis of choline esters ............................................................................................ 82
3.3 Results ................................................................................................................................................................ 84 3.3.1 High-throughput FLIPR assay ........................................................................................................... 84 3.3.2 Isolation of the active compound from H. spinnifer venom .............................................. 87 3.3.3 Identification of the active compound by high-resolution mass spectrometry ...... 90
3.4 Discussion ......................................................................................................................................................... 92 CHAPTER 4 — Conclusions ............................................................................................ 94
Summary and future directions .................................................................................... 95
References ................................................................................................................... 97
Appendix I .................................................................................................................. 125

12
LIST OF FIGURES AND TABLES FIGURE 1.1. GENERAL ARCHITECTURE OF PENTAMERIC ACETYLCHOLINE RECEPTORS. ..................................... 18 FIGURE 1.2 α-CONOTOXINS AS RESEARCH TOOLS TO STUDY THE STRCUTURE AND FUNCTION OF NACHRS. ............................................................................................................................................................................................................... 21 FIGURE 1.3. STRUCTURES OF TOXINS AND TOXIN DERIVATIVES THAT TARGET NACHRS. .................................. 31 TABLE 1.1 AMINO ACID SEQUENCES OF SELECTED α-CONOTOXINS. SI CONOTOXIN FROM CONUS STRIATUS THAT TARGETS THE NEUROMUSCULAR SUBTYPE CHOLINERGIC RECEPTOR; MII CONOTOXIN FROM CONUS MAGUS THAT TARGETS WITH HETEROMERIC NACHRS WITH HIGHER AFFINITY THAN THE HOMOMERIC
SUBTYPE α7 NACHR; AUIA CONOTOXIN ISOLATED FROM CONUS AULICUS, WHICH TARGETS THE
NEURONAL HETEROMERIC SUBTYPE α3β4. THE ASTERISKS (*) REPRESENT C-TERMINAL AMIDATION. TABLE ADAPTED FROM (LEWIS, 2012; MOLGÓ, 2013). LOOP 1 AND LOOP2 ARE THE TWO INTERCYSTINE LOOPS. 33 α-CONOTOXIN ................................................................................................................................................................................... 33 FIGURE 2.1. BASIC TWO-ADDITION FLIPR PROTOCOL TO MEASURE CALCIUM RESPONSES EVOKED BY
ACTIVATION OF α7 OR α3* NACHRS. ..................................................................................................................................... 47 FIGURE 2.2. ‘TWO-ADDITION’ FLIPR PROTOCOL USED TO IDENTIFY NACHR AGONISTS. ................................... 48 FIGURE 2.3. SCREENING OF VENOMS FROM “MODERN” SPIDERS AGAINST HUMAN α3* AND α7 NACHRS USING A HIGH-THROUGHPUT FLIPR ASSAY. ......................................................................................................................... 52 TABLE 2.1. TAXONOMIC DISTRIBUTION OF NACHR MODULATORS FROM ARANEOMORPH SPIDER VENOMS. TWO SPECIES SPARASSIDAE SP. WERE ASSAYED; THE NUMBERS 1 AND 2 INDICATE THEY WERE COLLECTED IN TWO DIFFERENT AREAS IN PNG. VENOMS WITH HIGH BLOCKING EFFECT ARE INDICATED WITH TWO
SYMBOLS DENOTED AS √√, WHILE THOSE WITH PARTIAL EFFECT ARE INDICATED WITH ONE √. THE GREEN AREA INDICATES THAT OTHER EFFECTS ON THE VENOM IS NOT OBSERVED. ............................................................. 54 FIGURE 2.4. AGONISM OF HUMAN NACHRS BY THERAPHOSID VENOMS. .................................................................. 57 TABLE 2.2. APPARENT AGONISTIC EFFECT CAUSED BY VENOMS FROM FOUR THERAPHOSID SPIDERS. AFTER VENOM ADDITION, THE FLUORESCENT CALCIUM RESPONSES EVOKED BY NACHRS IN THE ABSENCE OR PRESENCE OF PNU WERE CONVERTER INTO AREA UNDER CURVE USING SCREENWORKS SOFTWARE, VERSION 3.1. THE VALUES ARE AVERAGES OF THREE INDEPENDENT EXPERIMENTS WITH N = 3. THE MEAN
OF THE CONTROL NICOTINE RESPONSE WAS 509 ± 50.40 WHILE THE CONTROL CALCIUM RESPONSE
ELICITED BY CHOLINE WAS 1088 ± 38.31 .............................................................................................................................. 58 TABLE 2.3. MYGALOMORPH SPIDERS WITH VENOMS THAT ARE LIKELY TO CONTAIN AGONISTS OF α7 NACHRS. N.D INDICATES THE EFFECT WAS NOT DETERMINED AND A FURTHER ANALYSIS IS REQUIRED TO
CONFIRM THAT THE VENOM EFFECT IS VIA ACTIVATION OF α7 NACHRS IN THE PRESENCE OF PNU. ............ 58 FIGURE 2.5. THERAPHOSID SPIDER VENOMS THAT PREFERENTIALLY BLOCK α3 NACHRS. .............................. 59 TABLE 2.4. THERAPHOSID SPIDERS WITH VENOMS THAT SELECTIVELY BLOCK α3* NACHRS. ......................... 59 SPECIES WITH CHARACTERISTIC COLORS ARE INDICATED. (M) AND (F) REFER TO GENDER. THE NUMBERS BESIDE THE NAME OF EACH THERAPHOSID SPIDER INDICATES THAT THEY ARE INDIVIDUALS FROM THE SAME SPECIES, ALTHOUGH WERE COLLECTED FROM DISTINCT LOCALITIES. ......................................................................... 59 FIGURE 2.6. MANY THERAPHOSID VENOMS NON-SELECTIVELY BLOCK BOTH α3* AND α7 NACHRS. ........... 61 TABLE 2.5. THERAPHOSID SPIDERS WITH VENOMS THAT NON-SELECTIVELY BLOCK BOTH α3* AND α7 NACHRS. (M) AND (F) REFER TO GENDER. THE NUMBERS BESIDE THE NAME OF EACH THERAPHOSID SPIDER INDICATES THAT THEY ARE INDIVIDUALS FROM THE SAME SPECIES, ALTHOUGH WERE COLLECTED FROM DISTINCT LOCALITIES. ........................................................................................................................................................ 62 FIGURE 2.7. FOUR TARANTULA VENOMS WITHOUT POTENT MODULATORS OF α3* OR α7 NACHRS. ............ 63 TABLE 2.6. VENOMS FROM AMERICAN THERAPHOSIDS THAT LACK OF MODULATORS OF HUMAN α3* AND α7 NACHRS. ............................................................................................................................................................................................ 64 FIGURE 2.8. SUMMARISE OF THE HIGH-THROUGHPUT SCREENING OF SPIDER VENOMS. .................................... 65 FIGURE 3.1. A. CRASSICAUDA VENOM CAN INDUCE CALCIUM RESPONSES ELICITED VIA ACTIVATION OF NAV CHANNELS. ......................................................................................................................................................................................... 79 FIGURE 3.2. FLIPR SCREENING OF SCORPION VENOMS AGAINST α7 AND α3* NACHRS. ................................ 85 FIGURE 3.3. NON-SELECTIVE ACTIVATION OF α7 AND α3* NACHRS EXPRESSED IN SH-SY5Y BY CRUDE VENOM FROM H. SPINNIFER. ....................................................................................................................................................... 86 FIGURE 3.4. HPLC FRACTIONATION OF CRUDE VENOM FROM H. SPINNIFER. CHROMATOGRAM RESULTING FROM FRACTIONATION OF CRUDE VENOM VIA C18 RP-HPLC. ..................................................................................... 87 FIGURE 3.5. ACTIVITY OF F10 FROM FRACTIONATION OF CRUDE VENOM FROM H. SPINNIFER. ....................... 88

13
FIGURE 3.6. PURIFICATION OF THE “HIT’ TOXIN BY RP-HPLC. ..................................................................................... 89 FIGURE 3.7. AGONISTIC ACTIVITY OF THE “HIT” TOXIN. ..................................................................................................... 89 FIGURE 3.8. CO-ELUTION OF BIOACTIVE COMPOUND FROM H. SPINNIFER VENOM AND SYNTHETIC CHOLINE ESTERS. .............................................................................................................................................................................................. 91 TABLE I. LIST OF SPIDERS WHOSE VENOMS WERE USED IN THIS WORK .................................................................. 125 TABLE II. LIST OF SCORPIONS WHOSE VENOMS WERE USED IN THIS WORK ........................................................... 128

14
List of abbreviations ACh Acetylcholine
AChBP acetylcholine binding protein
AuIB α-conotoxin AuIB
AFU arbitrary fluorescent units
BSA bovine serum albumin
Ca2+ calcium ion
Cav2.3 P/Q-type calcium channel
CHCA α-cyano-4-hydroxycinnamic acid
CNS central nervous system
Da Dalton
DRG dorsal root ganglion
EDTA ethylenediaminetetraacetic acid
ESIMS electrospray ionization-mass spectrometry
FBS foetal bovine serum
FITC fluorescein isothiocyanate
FLIPR fluorescent imaging plate reader
GABA
GABAARs
γ-aminobutyric acid
γ-aminobutyric acid type-A receptors
HEPES 4-(2-hydroexyethyl)-1-piperazineethane sulfonic acid
HILIC hydrophilic interaction liquid chromatography
HPLC high-performance liquid chromatographic
HRMS high- resolution mass spectrometry
kDa Kilodalton
LsIA α-conotoxin LsIA
MALDI-TOF matrix assisted laser desorption/ ionization time-of-
flight
MS mass spectrometry
NMR nuclear magnetic resonance
NSAID non-steroidal anti-inflammatory drugs
nAChRs nicotinic acetylcholine receptors
PNS peripheral nervous system

15
PNU-120596 1-(5-chloro-2,4-dimethoxyphenyl)-3-(5-methylisoxazol-
3-yl) urea
RLU relative light units
PSS physiological salt solution
RPMI Rosewell Park Memorial Institute
RP-HPLC reversed phase high-performance liquid
chromatography
SAR structure-activity relationship
TFA trifluoroacetic acid
TRITC Tetramethylrhodamine
TTX Tetrodotoxin

16
CHAPTER 1 — INTRODUCTION Recent studies indicate that imbalance of the cholinergic system in human brain
via nicotinic acetylcholine receptors (nAChRs) are linked to various neurological
disorders (AhnAllen, 2012; Chu, 2005; Nides, 2006; Tregellas, 2011; Yu, 1996)
while mutations in the nAChR can lead to frontal lobe epilepsy (for a review see
(Steilein, 2001)). These findings have prompted research into the development of
drugs that target neuronal cholinergic receptors. Nevertheless, most of the
chemical components under trial lack selectivity, or hardly improve the medical
condition (for reviews, see (Arneric, 2007; Jensen, 2005; Pammolli, 2011)). The
discovery of new therapeutic agents is currently a challenge because of the
complex arrangement of subunits of neuronal cholinergic receptors expressed in
human brain (Gotti, 2007) and the fact that their roles under physiological and
pathological conditions are not fully understood (for a review, see (Tuesta, 2011)).
To facilitate studies of the pharmacology of nAChRs, a search for new nAChR
modulators in spider and scorpion venoms was carried out using a high-
throughput FLIPR assay to rapidly identify “hit” venoms. The probability of finding
native cholinergic modulators was expected to be high because arachnid venoms
are estimated to contain about 1 million different components, and even though
less than 0.01% of these molecules have been studied (for reviews, see
(Escoubas, 2006b; King, 2014; King and Coaker, 2014; King and Hardy, 2013),
many of them are peptidic toxins that act on diverse types of ionic channels
(Meves, 1984; Quintero-Hernandez, 2013; Shahbazzadeh, 2007; Xie, 2012).
This chapter provides a review of what is known about the molecular structure of
nAChRs and their physiological functions in vertebrates and invertebrates.
Furthermore, I review the use of animal toxins as research tools to characterize
nAChRs, as well as their roles in the development of therapeutic drugs and
bioinsecticides.

17
1.1 NICOTINIC ACETYLCHOLINE RECEPTORS
1.1.1 Homopentameric/heteropentameric architecture of neuronal nAChRs nAChRs are members of the pentameric Cys-loop ligand-gated ion channel
superfamily that are activated by the neurotransmitter acetylcholine (ACh) (for a
review, see (Unwin, 2005)). Our current knowledge of the molecular structure of
neuronal nAChRs began with determination of the N-terminal sequence of the
nAChR isolated from the electric organ of the marbled ray Torpedo marmorata,
followed by the cloning of its subunits. These subunits were subsequently shown
to be closely related to cDNAs cloned from mammals that encoded 17 structurally
homologous subunits, denoted α1–α10, β1–β4, γ, δ and ε. Each subunit
comprises four α-helical transmembrane regions (M1–M4), of which M1-M3 are
conserved and M4 has a variable extracellular C-terminal sequence; a conserved
large N-terminal extracellular domain (ECD) of approximately 200 amino acid
residues; a short intracellular C-terminal tail (see Figure 1.1A); and a variable
cytoplasmic domain between the transmembrane regions M3 and M4 (Unwin,
2005). In addition, all subunits contain in the first extracellular domain a cysteine-
loop (Cys–loop) that consists of a pair of cysteines, and in the case of mammals,
the Cys-Cys pair is separated by 13 residues. This structural template describes
the molecular backbone of different types of nAChRs.
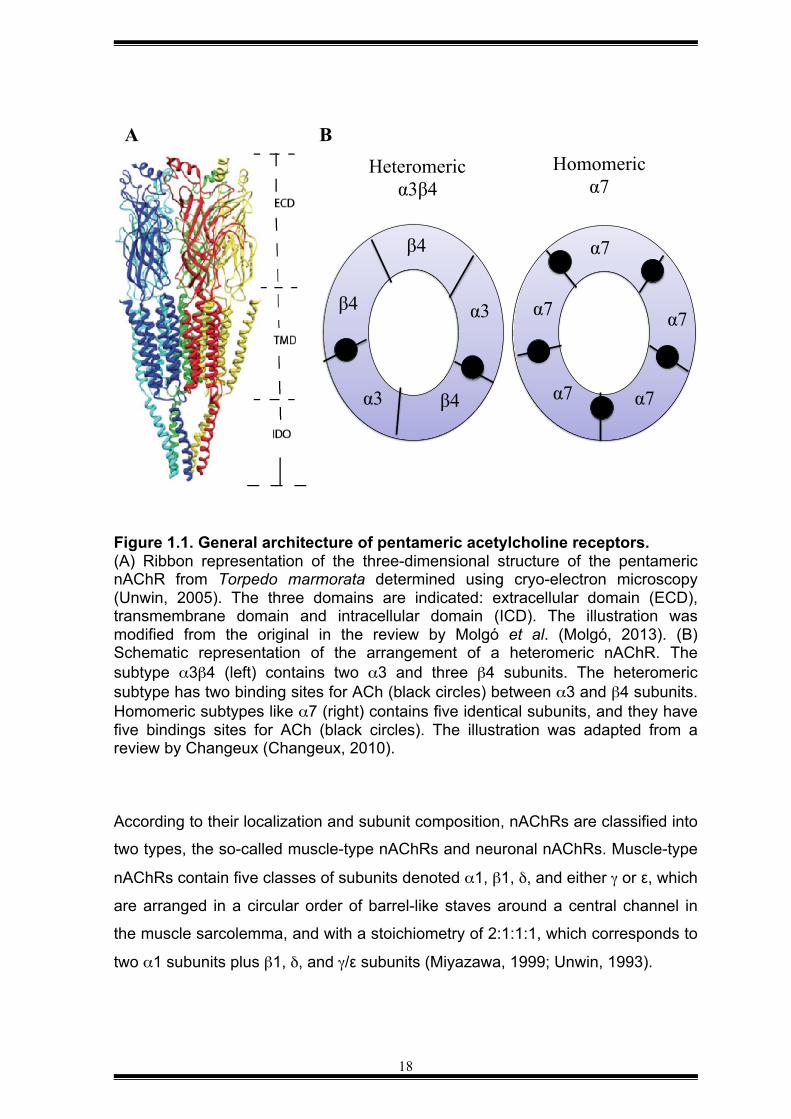
18
Figure 1.1. General architecture of pentameric acetylcholine receptors. (A) Ribbon representation of the three-dimensional structure of the pentameric nAChR from Torpedo marmorata determined using cryo-electron microscopy (Unwin, 2005). The three domains are indicated: extracellular domain (ECD), transmembrane domain and intracellular domain (ICD). The illustration was modified from the original in the review by Molgó et al. (Molgó, 2013). (B) Schematic representation of the arrangement of a heteromeric nAChR. The subtype α3β4 (left) contains two α3 and three β4 subunits. The heteromeric subtype has two binding sites for ACh (black circles) between α3 and β4 subunits. Homomeric subtypes like α7 (right) contains five identical subunits, and they have five bindings sites for ACh (black circles). The illustration was adapted from a review by Changeux (Changeux, 2010).
According to their localization and subunit composition, nAChRs are classified into
two types, the so-called muscle-type nAChRs and neuronal nAChRs. Muscle-type
nAChRs contain five classes of subunits denoted α1, β1, δ, and either γ or ε, which
are arranged in a circular order of barrel-like staves around a central channel in
the muscle sarcolemma, and with a stoichiometry of 2:1:1:1, which corresponds to
two α1 subunits plus β1, δ, and γ/ε subunits (Miyazawa, 1999; Unwin, 1993).
α7
Heteromeric α3β4
β4
β4
β4
α3
Homomeric α7
α7
α7
α7
α7
α3
A B

19
In contrast to the muscle-type nAChRs, the neuronal-type receptors are more
diverse. Neuronal nAChRs are encoded by nine α (α2–α7, α9, α8, α10) and three
β (β2–β4) subunit genes. In the case of sensory and neuronal mammalian tissues,
eleven subunits have been cloned, which are α2–α7, α9, α10, and β2–β4, but a
homologue of the avian α8 subunit has not been described (reviewed in
(Albuquerque, 2009; Nicke, 2004)). Neuronal nAChRs also contain five subunits
that form a functional receptor via combinations of different α and β subunits
(heteropentamers), such as α3β2 and α4β2. The ACh binding site is located at the
interface between an α subunit and an adjacent β subunit (see Figure 1.1B). Also,
a third subunit can be included in the receptor (α4)2(β2)2α5. In addition, the
pentamers can consist of only α subunits, such as homopentamers of α7, α9 or
α10 (Cooper, 1991); in these subtypes, the ACh binding site is located between α
subunits (see Figure 1B).
The molecular architecture of heteropentamers and homopentamers is similar
(see Figure 1.1B). Nevertheless, each subtype differs in function (Elgoyhen, 2001;
Zhou, 2003). If novel and selective modulators for heteropentameric and
homopentameric nAChRs were available some relevant questions in terms of the
functional characterization of neuronal nAChRs could be answered. For example,
the α6 and β3 subunits are co-expressed in brain areas such as the substantia
nigra and the ventral tegmental area (Quik and McIntosh, 2006) which leads to two
questions: firstly, why are these subunits co-expressed only in these areas of the
brain, and secondly, is co-expression of these subunits related to the
symptomatology of Parkinson’s disease or addiction to nicotine.
1.1.2 The use of toxins to elucidate the molecular architecture of nAChRs
Toxins derived from animal venoms have been a valuable source of
pharmacological tools for studying structure-activity relationships of nAChRs. For
example, the muscle-type nAChR was characterized for the first time by Changeux
and colleagues in 1970 thanks to the use of α-bungarotoxin, which was purified
from the venom of an elapid snake (Bungarus multicinctus), on membrane
preparations derived from the electric organ of the eel Electrophorus electricus

20
(Changeux, 1970). In addition, the proposed binding site for ACh in nicotinic
receptors, which lies in a cleft at the interface between the subunits, was
determined from x-ray crystallographic structures of the acetylcholine binding
protein (AChBP), a homolog of nAChRs, in complex with a variety of ligands (for a
review, see (Tsetlin and Hucho, 2009)). AChBP is a soluble protein that shares
sequence similarity to the ligand-binding site of α1 or α7 nAChRs. Following its
discovery, several crystal structures have been determined of the complex formed
AChBP and various α-conotoxins (see Figure 1.2A,B). For instance, co-
crystallization of the complexes formed between AChBP and the α-conotoxins IMI
and PnIA revealed that these peptide toxins bind at a similar spatial position in the
ACh binding domain despite having divergent primary structures (see Figure
1.2C). Moreover, it was shown that hydrophobic interactions between the ACh
ligand- binding domain, and a conserved proline residue in both conotoxins could
lead to a differential selectivity to nAChRs (for reviews, see (Lebbe, 2014b; Lewis,
2012)).
A large number of α-conotoxins have been used to characterise many of the
neuronal subtypes of nAChRs (for a review, see (Lebbe, 2014b)) and structural
studies of the α-conotoxins : nAChR interaction have been invaluable (Bourne,
2005; Lewis, 2012). Nowadays, we can predict what amino acids residues can be
modified to improve α-conotoxin affinity and selectivity. For example, it was shown
that the selectivity of α-conotoxin, Lo1a from Conus longuriunis (see Figure 1.1D)
could be switched from the neuronal α7 subtype to adult muscle-subtype nAChR
by deleting an Asp residue from its C-terminus (Lebbe, 2014a). An α-conotoxin
that can differentially target the muscle subtype over the neuronal subtype could
be used to engineered probes for imaging studies of nAChRs in the peripheral
nervous system. Because α marine cone snails represent the largest source of
tools for studying vertebrate nAChRs, most of the modulators discovered to date
are antagonists. In contrast, very few nAChRs agonists have been reported.
Therefore, further research is needed to obtain new, subtype-selective nAChRs
modulators that might be useful therapeutics and/or useful tools for addressing
fundamental questions about nAChRs.

21
Figure 1.2 α-Conotoxins as research tools to study the strcuture and function of nAChRs. (A) Top view of the X-ray structure of the complex between Aplysia californica AChBP (AcAChBP) and α-conotoxin PnIA (for a review see (Armishaw, 2010)). Each of the five subunits is shown in a different color (orange, green, blue, red and cyan). (B) A single subunit of AcAChBP interacting with an α-conotoxin , with the C-loop is an open position (cyan). The arrow indicates the difference in conformation between the C-loop in the open (cyan) and closed (red) conformations (for more details see review (Armishaw, 2010)). (C) Co-crystal structures between AcAChBP (transparent image, only two subunits shown for clarity) and α-conotoxin ImI. Residues Arg11 and Arg7 (labelled) that are the primary residues on ImI that interact with AcAChBP. Figure taken from Dutertre et al. (Dutertre, 2007). (D) NMR solution structure of α-conotoxin Lo1A. Disulfide bonds are shown in yellow. Image from Lebbe et al. (Lebbe, 2014a).
1.1.3 Location and physiological function of nAChRs in vertebrates and invertebrates In vertebrates, nAChRs are expressed in most cells of the central and peripheral
nervous system (Albuquerque, 2009; Cooper, 1991), and in non-neuronal cells
(Macklin, 1998; Plummer, 2005). It seems that the role of ACh and nAChRs in the
CNS is mainly modulatory because the cholinergic signal can be excitatory or
C D

22
inhibitory of synaptic pathways in different cerebral areas (for a review, see
(Picciotto, 2012)). The different roles of neuronal nAChRs are made possible by
different combinations of α and β (Gotti, 2007). To date, the function of each
nAChR subtype in each place of expression is still not fully understood.
Human imaging studies in vivo have allowed mapping of the distribution of
nAChRs in the human brain (Kimes, 2006). Cholinergic projections are widely
distributed in the brain. For example, neuronal nAChRs have been reported to be
expressed in the white matter (Ding, 2004) and are likely to be involved in the
regulation of neuronal excitability in the thalamocortical area (Bucher and
Goaillard, 2011). Another cerebral region is the ventral tegmental area where the
release of glutamate and dopamine are related to the reward responses generated
by nicotine administration (Picciotto and Kenny, 2013). It contains GABAergic,
glutamatergic, and dopaminergic neurons; and their activities are regulated by a
combination of α4β2* nAChRs expressed mainly in GABAergic neurons and α7
nAChRs expressed in terminals of the glutamatergic neurons (Mansvelder, 2002).
Neuronal nAChRs are not restricted to the postsynaptic membranes; they can also
be expressed along the neuronal body, including axons and presynaptic terminals,
and different subtypes of nAChRs can be co-expressed in a neuron (for a review,
see (Picciotto, 2012)). Other cholinergic projections come from the basal forebrain
complex to the cerebral cortex. Their terminals contain β2* nAChRs; and are
distributed across all the cerebral cortex. Activation of the receptor lead to release
of glutamate into the prefrontal cortex, and this mediates neuronal processes such
as memory, attention and cue detection (Parikh, 2010).
To date, it is known that in humans the distribution of nAChRs is not limited to the
nervous system. Cholinergic receptors are expressed in human aortic endothelial
cells (α3, α5, α7, α10, β2-4) (Wada, 2007), vascular smooth muscle cells (α2-5,
α7, α10) and immune cells, such as human platelets (α7) (Schedel, 2010); human
mononuclear leukocytes (α7, β2) (Albaugh, 2004), human macrophages (α1, α7,
α10), B-lymphocytes (α3-5, α7), T-lymphocytes (α3, α4, α7, β2, β4), (for a review,
see (Bauwens, 2015)). The function of these receptors is linked to homeostasis of
the inflammatory response by the cholinergic anti-inflammatory pathway. For

23
instance, when terminals of afferent fibres located in the vagus are stimulated by
pro-inflammatory stimuli, action potentials are generated, and this causes efferent
fibres to activate α7 expressed on macrophages by acetylcholine (for a review see
(Egea, 2015)). In the end, α7 control the production of pro-inflammatory cytokines
such as TNF-α and IL-6 (Zhang, 2015a) without affecting the levels of anti-
inflammatory cytokines by inhibiting NF-kB nuclear translocation and activating
several pathways including JAK2/STA3 (Li, 2015). However, the cholinergic anti-
inflammatory pathway is not fully understood (for a review, see (Egea, 2015)).
In invertebrates, nAChRs have been found in the central nervous system, where
they mediate the excitatory synapsis of ACh. Studies of insect brain nAChRs have
been limited because there has only been limited success with heterologous
expression of cloned insect nAChRs (Landsdell, 2012). The main problem is that
cholinergic currents are evoked at high concentrations of ACh and the inhibitory
currents are usually too small to be accurately detected with electrophysiology
experiments using Xenopus oocytes, although co-expression of insect nAChRs
with subunits of vertebrate nAChRs has partially facilitated the expression of insect
nAChRs (Landsdell, 2012). There are still important nAChRs from insect species
such as honeybee Apis mellifera that are important to study to design novel and
safe insecticides. Today, it is known insect nAChRs have similar molecular
structure to vertebrate nAChRs. For instance, in the genome of Drosophila
melanogaster, 10 nAChR subunit genes of nAChRs have been identified: seven
encoded the α subtype (Dα1-7), and three corresponded to β subtype (Dβ1-3)
(Jones, 2007). The sequences of Dα5, Dα6 and Dα7 subunits are similar to each
other, and to α7 subunit from vertebrate nAChRs (Grauso, 2002; Jones and
Satelle, 2009). The Dα6 subunit is the most similar to the vertebrate α7 nAChR
(Sattelle, 2005). This similarity may allow insect brains to serve as a model system
for studying brain insect as an alternative animal model to study the role of
nAChRs in different pathologies. For example, more than 40% of bee brain
consists of Kenyon cells that are the major cells of the cerebral structure, known
as mushroom bodies or corpora pedunculata. This structure processes multiple
sensory information derived from the antennal lobe (Akalal, 2006; McGuire, 2001).
It was shown that neonicotinoids cause up-regulation of nAChRs (Moffat, 2015);

24
and because nAChRs can cause mitochondrial depolarization it was suggested
the effect of these insecticides may be mediated in part by up-regulation of
nAChRs in Kenyon cells leading to mitochondrial dysfunction, memory deficits,
and poor navigation (Fischer, 2014; Williamson, 2013).
To summarize, nAChRs engaged in different physiological functions that are
guided by the variability of their subunits (McGehee and Role, 1995). Many efforts
have been made to determine the role of various nAChR subtypes in the nervous
system but most studies have focussed on the homomeric α7 and heteromeric α4
nAChRs (Koga, 2014; Schedel, 2010; Zhou, 2003). This is reflected in the
increasing number of drugs that target α7 and α4β2, which are under development
for treatment of several neurological conditions. However, the lack of specific
modulators for other nAChR subtypes makes it challenging to elucidate the role of
each subtype under both physiological and pathological conditions. Thus, there is
still a significant need to discover or develop highly selective modulators of
neuronal nAChRs.
1.1.4 Role of nAChRs in human disease
The role of ACh is as important as their molecular target, nAChRs. It is
synthesised by the enzyme choline acetyltransferase that transfers an acetyl
group from acetyl-CoA to choline (Korey, 1951). Its precursor, choline, is
endogenously produced, although it is also absorbed in the small intestine to
reach the levels required to accomplish several functions (Zeisel and da Costa,
2009). For example, choline is a precursor for the biosynthesis of constituents of
plasma membrane such as phosphatidylcholine and sphingomyelin (Zeisel, 1992;
Zeisel and da Costa, 2009). Furthermore, choline is a methyl group donor; and
methylation is needed to regulate of expression of genes and mediate biosynthetic
reactions (Zeisel and da Costa, 2009). A deficiency in choline has been related to
liver damage (Buchman, 1995) and impairment of cognitive functions in older
people (Fioravanti and Yanagi, 2005). Indeed, choline supplementation enhances
cognitive functions in patients with schizophrenia (Knott, 2015). De-regulation of
cholinergic function is linked to the function and expression of nAChRs. For

25
instance, chronic administration of nicotine can induce an increase in the number
of neurons containing α4β2 nAChRs in the glutamatergic subthalamic nucleus, a
cerebral area related to movement control (Xiao, 2015), or down-regulate the
function of other subtype nAChRs. Below is discussed different pre-clinical and
clinical evidence of the consequences of smoking, and links between de-regulation
of nAChRs and major symptoms in neurodegenerative diseases.
Post-mortem studies in the brain of people who suffer from Alzheimer’s and
Parkinson’s disease (Rinne, 1991), epilepsy and schizophrenia showed that the
density of α4β2 nAChRs is extremely low. The degeneration of cholinergic circuits
was confirmed in human brains after the development of nicotine radioligands for
imaging studies in vivo. For example, in vivo studies of brains of people suffering
from Alzheimer’s disease showed a decrease of α4β2 nAChRs (Kendziorra,
2010), and the density of nAChRs in arteries appeared to be affected as well
(Bauwens, 2015).
In patients suffering from Alzheimer’s disease and Parkinson’s disease, there is a
notable loss of cholinergic projections into the cerebral cortex, and this has been
considered as part of the reason for the impaired cognition observed in these
patients, although the loss of the cortical cholinergic activity in people with
Parkinson’s disease is notable (for a review see (Müller and Bohnen, 2013)). In
addition, depressive symptoms in patients with Parkinson’s disease is strongly
correlated to loss of α4β2 nAChRs in the neuronal frontal corticomesostriatal
circuitry, while those patients with mild cognitive impairment were associated with
low levels of α4β2 in the neuronal frontal corticobasal ganglia-limbic-cerebellar
circuitry that includes some areas of the former circuitry (Meyer, 2009).
Furthermore, α4β2* nAChRs are widely distributed in the brain and are not
restricted to cholinergic or dopaminergic neurons. Thus, down-regulation of this
heteromeric subtype may lead to dysfunction of other neuronal circuitries (Emre,
2003). Cholinergic deregulation in the hippocampus may be associated with
cognitive problems because this cerebral region is related to functions of learning
and memory (Kutlu and Gould, 2015) and this correlation between dysfunction of

26
neuronal circuitry and cognitive functions is observed in various neuronal disorders
(Drevets, 2008).
nAChRs have also been identified in immune cells, and up-regulation of its activity
in this cells has been linked to cardiovascular diseases such as atherosclerosis in
smokers (Aicher, 2003; Egleton, 2009; Wada, 2007). Recent evidence suggests
that the mechanism underlying this process could be activation of α7 nAChRs that
trigger transcription of cell-adhesion proteins on endothelial cells (Alamanda,
2012). These proteins, such as E-selectin, favour the attachment of monocytes,
and it might lead to abnormal formation of fibrous plaques in vascular smooth
muscle cells (Alamanda, 2012). Notably, the duration and amount of nicotine
administered via smoking are dependent variables, and when these reach critical
values it may facilitate the activation of numerous nAChR subtypes (Koga, 2014;
Wada, 2007) that are linked to signalling cascades (Wada, 2007) and cause
abnormal proliferation of neuronal and non-neuronal cells (He, 2014; Wada, 2007).
1.2 MODULATORS OF NACHRS
1.2.1 Modulators of nAChRs as drugs and insecticides
Several clinical trials are being carried out to evaluate potential drugs that target
nAChRs. For instance, some nicotine analogues are been used to facilitate
quitting smoking (Rollema, 2007). Varenicline (trade name Chantix®) is currently
being used for treatment of nicotine addiction and is a partial agonist of the α4β2
and α3* receptors. However, varenicline also activates the α7 nAChR (Mihalak,
2006), and it has been suggested that its mechanism of action involves is by
modulation of several nAChRs (Bordia, 2012; Lotfipour, 2012) that contributes to
decreasing the levels of dopamine that were previously increased by nicotine in
the limbic system (Corrigall, 1992, 1994), thus, preventing the reward response
induced by smoking (Mihalak, 2006; Rollema, 2007). Imaging studies in vivo of
smokers under treatment with varenicline showed that α4β2* nAChRs are
saturated after administration of the drug at low or high doses, and withdrawal
effects were not detected. Unfortunately, varenicline can also cause adverse
effects in smokers under treatment; nausea is very common and is one of the

27
primary reasons for withdrawal for treatment (Jorenby, 2006; Lam and Patel,
2007). The side-effect is likely to be mediated by activation of ganglionic nAChRs.
This is likely to be a common problem when using therapeutic agents that target
multiple subtypes of nAChRs distributed across the peripheral nervous system.
As mentioned previously, nAChRs are also important insecticide targets, and the
so-called neonicotinoid insecticides that target insect nAChRs are one of the most
successful classes of chemical insecticides. Nicotinoid insecticides are derived
from nicotine, the major alkaloid derived from leaves and stems of the tobacco
plant Nicotiana tabacum; nicotine is a secondary metabolite that is used by the
plant to defend against herbivorous insects (Isman, 2006). Nicotine was used to
control crop pests as far back as the 1700s (Siegwart, 2015) but its use was
limited because of toxic effects on mammals and only moderate potency against
insects. The neonicotinoids are chemically similar to nicotine but they have low
toxicity to mammals as they were engineered to bind selectively to insect nAChRs
(Brown, 2006). However, like the majority of chemical insecticides, most
neonicotinoids are broadly active against a wide range of insect pests, including
beneficial insects such bees (Déglise, 2002; Faucon, 2005). Honeybees are
efficient pollinators that mediate plant reproduction, and therefore it would be
highly desired to engineer insecticides that preferentially target nAChRs in insect
pests but not beneficial insects such as pollinators or predators of the targeted
pest.
Structure-activity relationship studies are helping to guide the development of
more selective neonicotinoid insecticides. Crystal strictures of the complex formed
between neonicotinoids and AChBP have been used to identify rational methods
of improving the potency and/or selectivity of particular neonicotinoids (Matsuda,
2009). For instance, Matsuda et al. (Matsuda, 2009) compared the complex
formed between imidacloprid, the most successful neonicotinoid, and α2β1
nAChRs from the honeybee Apis mellifera and the green peach aphid Myzus
persicae (Matsuda, 2009). They found a remarkable structural difference between
the binding sites for imidacloprid in the two nAChRs, with a hidden and broader
binding groove in the aphid nAChR compared with the bee receptor. The authors

28
suggested that linking imidacloprid to a molecular fragment that fits the groove of
the aphid nAChR could improve the selectivity for insect pests over bees.
To summarise, neonicotinoids are generally safe for humans but they can be toxic
for small vertebrates such as birds and beneficial insects such as honeybees.
Thus, further research is needed to develop more selective neonicotinoids with
reduced ecological impact.
1.2.2 Novel subtype-selective modulators of vertebrate nAChRs are required to develop accuracy pharmacological tools and potential therapeutic leads
The selectivity of some toxins for nAChRs has facilitated the development of
fluorescent binding probes and radioligands for use in biological sciences
(Hannan, 2015; Schmaljohann, 2006) and nuclear medicine (Schmaljohann, 2005,
2006), respectively. It has helped to determine the distribution of nAChRs in the
brain of rodents (Perry, 2002; Rueter, 2006) and humans (Ding, 2004; Kimes,
2006), including in people suffering from Alzheimer’s and Parkinson’s disease
(Schmaljohann, 2005, 2006). For example, α-bungarotoxin a 74-residue peptide
isolated from venom of Bungarus multicinctus, was used to affinity purify the
nAChR from the electric organ of T. marmorata (Changeux, 1970). α-Bungarotoxin
has also been useful for visualizing the distribution of nAChRs in the brain and the
in the electric organ from Torpedo californica, and for identifying clones expressing
epitopes from nAChR in Escherichia coli (Gershoni, 1987). Recently, it was shown
that Alexa Fluor 555-labelled α-bungarotoxin labels GABAA-Rs expressed in HEK-
293 cells (Hannan, 2015), and this binding is inhibited by bicuculline, a competitive
antagonist at GABAA-Rs, and d-tubocurarine, a non-selective nAChRs antagonist
and a competitive antagonist at GABAA-Rs (Caputi, 2003) (see Figure 1.2A-C).
This finding is not surprising because type-A GABAergic receptors are members of
the pentameric Cys-loop ligand-gated ion channel family, like nAChRs (Corringer,
2012). The selectivity of α-bungarotoxin has been used to discover new functional
subunits of nAChRs, such as the α7 subunit of nAChRs. The vertebrate neuronal
subunit α7 was cloned for the first time in 1990 by Couturier et al. (Couturier,

29
1990). They found the subunit formed an oligomeric channel in the membrane of
Xenopus laevis oocytes and it was activated by nicotine and choline, and
desensitised rapidly. This was a receptor similar to the heteropentameric nAChR
reported previously because α-bungarotoxin blocked its activation and was able to
bind the subunit α7. Furthermore, iodinated α-bungarotoxin has been used in
competition studies to evaluate the specificity of radioligands towards α7 nAChRs.
Nicotine radioligands are being used for imaging studies using positron emission
tomography (PET) (for a review, see (Bauwens, 2015)). Note that radioligands are
typically used for imaging studies at sub-therapeutic doses, and consequent safety
rarely poses an issue. The first nicotine-based radioligand developed to label
nAChR was [11C]-nicotine (Halldin, 1992) (see Figure 1.3E). It was synthesised at
high purity but had low specificity when tested on human brain in vivo (Nybäck,
1994). Other radioligands that are chemically related to nicotine are still under
development with some promising results, such as [18F]-ZW-104 and [18F]-AZAN-α
(for a review, see (Bauwens, 2015)) (see Figures 1.3F-G). The first probe is a
selective ligand for the β2 subunit of nAChR and has been tested pre-clinically.
[18F]-AZAN-α has been assayed in human studies with promising results (for a
review, see (Bauwens, 2015)). It reached the brain in less than 20 min after
injection, and selectively bound the α4β2 subtype (Wong, 2013). In this study the
specificity for α4β2 was evaluated through administration of varenicline, a
selective partial agonist of α4β2 that is used to aid smoking cessation (Rollema,
2007). The specificity of the radiotracer for the receptor was confirmed after the
drug blocked totally the uptake of [18F]-AZAN-α. Another radio-chemical tracer is
[18F]-flubatine, which is an analogue of epibatidine, an alkaloid isolated from the
skin of the frog Epipodobates tricolor (Fisher, 1994) (see Figures 1.3D,H).
A-85380 is another example of a radiotracer (Kimes, 2006; Sullivan, 1996) that
has high affinity for α4β2 nAChRs (for a review, see (Lotfipour, 2011)). It allowed
mapping of the distribution of nAChRs in the brain of smokers and non smokers
(Kimes, 2006). Furthermore, it has been used to confirm that expression of α4β2
in the brain of patients with Alzheimer’s disease and dementia is significantly
decreased, and is correlated with the degree of cognitive impairment (Colloby,

30
2010).
The development of agents to be used for human brain imaging in vivo has been
extended to design of tracers selective for the α7 subtype. Some promising results
has been obtained even where the homomeric α7 in the human brain is found in
low concentrations compared to α4β2. For example, [18F]-NS-10743 and [18F]-
ASEM (see Figure 1.3I-J) showed high affinity toward α7 and the distribution of
these radioligands were found in brain areas where α7 is densely expressed, such
as frontal cortex and hippocampus. To prove the specificity of the agents, the
specific ligand SSR180771, which is a selective and partial agonist at human and
rodent subtype α7, was used in competition studies. The tracers showed their
specificity for α7 when these were displaced by SSR180771.
As discussed above, part of the problem with nAChR modulators as drug leads,
insecticides, and pharmacological tools is their lack of selectivity. This has made it
difficult to dissect the role of nAChRs in different physiological processes (García,
2015) as well as their role in pathophysiological processes such as cognitive
dysfunction, depression, chronic anxiety, analgesia, inflammation and
neurodegenerative diseases. Thus, there is still a great need to develop novel
nAChR modulators that are both potent and selective. As discussed below, animal
venoms have provided some of the most potent and selective nAChR modulators
described to date, and represent a potential source of new nAChR modulators.

31
Figure 1.3. Structures of toxins and toxin derivatives that target nAChRs. (A) α-bungarotoxin. Image adapted from https://en.wikipedia.org/wiki/Alpha-Bungarotoxin. (B) Bicuculline. Image from http://halofanon.wikia.com/wiki/Bicuculline. (C) Tubocurarine chloride. Image from https://en.wikipedia.org/wiki/Tubocurarine_chloride. (D) Epibatidine. (E) 11C-nicotine. (F) [18F]-ZW-184. Image from http://www.iphc.cnrs.fr/dirac/productReadHit.php?id=923. (G) [18F]-AZAN-α. Image adapted from (Bauwens, 2015). (H) [18F]-Flubatine. Image adapted from (Bauwens, 2015). (I) [18F]-NS-10743. Image adapted from (Bauwens, 2015). (J) [18F]-ASEM. Image from Horti et al. (Horti, 2014). The pyridine and the pyrrolidine rings are shown in orange and green, respectively. The chemical structure of nicotine is similar to that from epibatidine such as pyridine rings and nitrogen atoms (in orange color). The tracers are shown in blue.
E) F)
A) B) C)
α-bungarotoxin Bicuculline Tubocurarine chloride
[18F]-NS-10743
D)
I) J)
Epibatidine
[18F]-AZEM
N
N Cl
N H3[11C]
N
11C-nicotine [18F]-ZW-104 [18F]-AZAN-α [18F]-Flubatine
N
O
18F G) H)
N
N
N
18F
N N
18F
N N
O 18F N
N N
18F O O
S

32
1.3 VENOM-DERIVED MODULATORS OF NACHRS 1.3.1 Important role played by subtype-selective α-conotoxins in study of
vertebrate nAChRs Venoms from marine cone snails (genus Conus) have been extensively studied,
although scarcely 0.1% of the predicted total number of venom compounds have
been characterized so far (Lebbe, 2014b). Conus venoms are replete with
disulfide-bridged peptides, named conopeptides or conotoxins that target a wide
range of voltage and ligand-gated ion channels, including nAChRs (Lebbe, 2014b;
Lewis, 2012; McIntosh, 1999). Indeed, the venoms of marine cone snails are the
largest natural source of potent and selective nAChR blockers (for reviews, see
(Lewis, 2012; McIntosh, 1999)). There are at least seven families α-conotoxins
that bind specific muscle and neuronal nAChR subtypes with high selectivity
(Wang, 2015). α-Conotoxins are the smallest conopeptides (generally 12–20
amino acid residues) that competitively binds to the ACh binding site of nAChRs.
Their general framework is denoted CC-Xm-C-Xn-C, where C represents a
cysteine residue and X corresponds to the two inter-cysteine loops that contain a
variable region of amino acid residues. The number of amino acid residues is
indicated as m/n and α-Conotoxins with a 3/5 loop size combination are usually
purified from the venom of Conus species that feed on fish and are typically active
on mammalian neuromuscular nAChRs, while other conopeptides with 4/3, 4/4,
4/5, 4/6 and 4/7 loop combinations preferentially act on mammalian neuronal
nAChRs (for reviews, see (Lebbe, 2014b; Lewis, 2012; Terlau and Olivera, 2004)).
See table 1.1 below.

33
Table 1.1 Amino acid sequences of selected α-conotoxins. SI conotoxin from Conus striatus that targets the neuromuscular subtype cholinergic receptor; MII conotoxin from Conus magus that targets with heteromeric nAChRs with higher affinity than the homomeric subtype α7 nAChR; AuIA conotoxin isolated from Conus aulicus, which targets the neuronal heteromeric subtype α3β4. The asterisks (*) represent C-terminal amidation. Table adapted from (Lewis, 2012; Molgó, 2013). Loop 1 and Loop2 are the two intercystine loops. α-conotoxin Amino acid sequence Selectivity
C Loop 1 C Loop 2 C*
SI (3/5 loop) I CC NPA C GPKYS C* Muscle type α12βγδ
MII (4/7 loop)
G CC SNP V C HLEHSNL C* Neuronal subtype α6α3β2β3>α3β2>α7
AuIA (4/7 loop)
G CC SYP P C FATNSDY C* Neuronal subtype α3β4
Several studies of the interaction between α-conotoxins and AChBP have
contributed to our understanding of how α-conotoxins can be selective for a
particular nAChR (Dutertre, 2007). Basically, the different electrostatic interactions
and hydrogen bonds between aromatic residues of α-conotoxins and amino acid
residues outside of the ACh binding pocket are found in the conotoxins that block
nAChRs while those conotoxins with fewer interaction are more likely to be
agonists (Dutertre, 2004). These studies have also helped develop a structural
model of the pharmacophore of α-conotoxins. It is delimitated by the two loops
between the cysteine residues. One is conserved and determines the binding to
the nAChR and the second loop consists of a variable sequence that determines
their selectivity for a particular subunit of the nAChR (for reviews, see (Lebbe,
2014b; Lewis, 2012; Terlau and Olivera, 2004)).

34
1.3.2 nAChR modulators from other venomous animals Snake venom is another large source of proteins and peptides that target nAChRs
(Brahma, 2015; Utkin, 2015). The most abundant proteins described so far in
elapid, colubrid and psammophid snakes are the three-finger toxins (3FTx)
(Sunagar, 2013) that contain between 60 to 74 amino acid residues with molecular
masses ranging between 6000 and 8000 Da (Manjunatha and Robin, 2010; Roly,
2014). Most 3FTxs target nAChRs. These toxins are divided into three types
differentiated by the number of cysteines and the inter-cysteine spacing. Type I
and type III α-neurotoxins with eight cysteine residues target neuromuscular
nAChRs. Type II α-neurotoxins contain an extra pair of cysteine residues which
form an additional disulfide bond that stabilises loop 2 (Fry, 2003; Sunagar, 2013).
This additional disulfide bond facilitates interaction with neuronal α7 and α9-10
nAChRs, while maintaining affinity for the neuromuscular α1 nAChR (Fry, 2003).
Another interesting group of 3FTxs are the so-called “non-conventional toxins” that
have an additional disulfide bond that is not in the central loop II but instead in the
N-terminal loop I (for a review see (Tsetlin, 2015)). Examples of non-conventional
toxins that block nAChRs are candoxin from Bungarus candidus and the weak
toxin (WTX) from Naja Kaouthia.
1.3.3 Rationale for examining spider venoms as a source of novel nAChR modulators Spiders are very known to produce venoms that contain diverse chemical
structures such as peptides up to 10 kDa, proteins, amino acids, and
acylpolyamines (for a review, see (Estrada, 2007)). This complex and
heterogeneous mixture of compounds allows spiders to prey on, and defend
themselves against a wide range of prey and predators. The different repertoire of
acylpolyamines appears to have evolved to allow predation of different species of
invertebrates and small vertebrates.
Acylpolyamines can selectively block ionotropic glutamate receptors from insects
and vertebrates including mammals (for a review see (Olsen, 2011)). Because
glutamate is the main neurotransmitter at the neuromuscular junctions in insects,

35
block of glutamatergic receptors by acylpolyamines induces paralysis (Piek, 1971;
Piek and Njio, 1975). However, acylpolyamines toxins are also reported to
antagonise of nAChRs (Willians, 1997). The basic backbone of acylpolyamines is
an aromatic acyl group and the polyamine chain that form the essential part of the
molecule (Estrada, 2007). These two features are generally present in
acylpolyamines derived from funnel-web, trap door and tarantula spiders
(Strømgaard, 2005). The length of the polyamine chain and the degree of
hydrophobicity determines the affinity for nAChRs.
Recently, a toxin denoted VdTX-1 was purified from the venom of the Brazilian
theraphosid spider Vitalius dubius (Rocha-E-Silva, 2013). This component has a
molecular weight of 728 Da and it non-competitively blocks nAChRs. Surprisingly,
this toxin was photosensitive, which might be why this toxin, and perhaps related
toxins in other spider venoms, was not identified in previous studies. In any case,
there is clearly some evidence for the presence of small-molecule nAChR
modulators in spider venoms, suggesting that these venoms should be explored
more systematically as a potential source of novel modulators.
1.4 SIGNIFICANCE AND AIMS OF THE CURRENT WORK 1.4.1 Significance nAChRs are considered a key molecular target for treatment of several
neurological and vascular disorders (Bauwens, 2015; van Enkhuizen, 2015). Key
knowledge about the pharmacology and distribution of nAChRs in the vertebrate
nervous system has been achieved thanks to the use of animal toxins as research
tools. However, we are still a long away from fully understanding of role of different
subtypes of nAChRs in neurons and non-neuronal cells due largely to the lack of
selective pharmacological modulators. Although animal venoms have proved to be
an excellent source of nAChR modulators, most work has been carried out using
venoms from snakes and cone snails, while spiders and scorpions remain largely
unstudied as a potential source of nAChR modulators. Since spiders and
scorpions use their venom to either hunt, or defend against, vertebrates, their
venoms represent a potential source of compounds that can modulate the activity

36
of vertebrate nAChRs. Thus, in this thesis, I aimed to explore arachnid venoms for
the presence of compounds that modulate the activity of vertebrate α3* and the
neuronal α7 nAChRs.
1.4.2 Aims Aim 1: Determine the taxonomic penetrance of nAChR modulators in venoms
from the two major infraorders of spiders: the “primitive” spiders (Mygalomorphae)
and “modern” spiders (Araneomorphae)
Aim 2: Isolate and pharmacologically characterize a non-peptide nAChR
modulator identified in venom of the scorpion “Heterometrus spinnifer”.

37
CHAPTER 2 — NACHR MODULATORS FROM SPIDER VENOMS 2.1 INTRODUCTION
Venomous animals use their venom to hunt prey and/or protect themselves
against predators (Billen, 2008; Olsen, 2011). Most animal venoms are a complex
mixture of salts, small organic compounds, peptides and proteins (Escoubas,
2006b; Zhang, 2015b), with activity against a wide range of receptors (Inserra,
2013; Wang, 2015) and ion channels (Billen, 2008; DeBin, 1993; Nirthanan, 2002).
Consequently, over the past 20 years, a number of venom molecules have been
purified which have proved useful clinically, to agronomy, or as research tools
(Harrison, 2014; King and Coaker, 2014; King and Hardy, 2013; Lewis, 2012;
Nicke, 2004; Windley, 2012b).
One potentially rich source of compounds active against ligand-gated channels,
which remains largely unexplored, is spider venoms (Nørager, 2014). It is
predicted that all venoms from the estimated 100,000 extant species of spiders
contains approximately 20 million bioactive molecules and scarcely 0.008% have
been characterized so far (for reviews, see (Estrada, 2007; King and Coaker,
2014; King and Hardy, 2013; Windley, 2012b)). This has been due in part to the
small amount of venom that can be obtained from arachnids, making traditional
biochemical characterization challenging. However, the development of high-
throughput functional assays has made it more feasible to functionally characterize
venom from small venomous animals (for a review, see (Vetter, 2011)).
There are two reasons why spider venoms were considered to be a likely possible
source of novel molecules that target human neuronal nAChRs. Firstly, each
venom is estimated to contain several hundred to more than a thousand molecules
in the desired molecular weight range of between 0.1 and 10 kDa (for reviews, see
(Escoubas, 2000, 2006a, b; Estrada, 2007; King and Hardy, 2013)), with most of
those already isolated having been shown to predominantly act on voltage- and
ligand-gated ion channels (for reviews, see (Escoubas, 2000, 2006a, b; King and
Hardy, 2013)). Secondly, even though spider toxins are designed to target mainly
insect receptors (because insects are the main prey of spiders) (Corzo, 2002;

38
Vassilevski, 2008), the similarity of invertebrate receptors to their counterparts in
vertebrates (Lansdell, 2012) suggests that modulators of human nAChR receptors
might be found in spider venoms.
Some authors have divided spider toxins into two major groups: acylpolyamines
and cysteine-rich peptide toxins (Liang, 2008; Vassilevski, 2009). The
acylpolyamines were first isolated from theraphosid spiders (for reviews, see
(Estrada, 2007; Strømgaard, 2001) and shown to cause fast paralysis in insects
(Escoubas, 2000) but they are found in many spider venoms (Moore, 2009;
Strømgaard and Mellor, 2004) They are known to block both ionotropic
glutamatergic receptors (Nørager, 2014; Strømgaard and Mellor, 2004) and
nAChR receptors in vertebrates and invertebrates (Mellor and Usherwood, 2004;
Strømgaard, 2001). Their potential therapeutic and insecticidal use has led
researchers to synthesise acylpolyamines (Strømgaard and Mellor, 2004) and
study their structure-activity relationships in detail in order to improve their affinity
for either glutamatergic (Kromann, 2002) or cholinergic receptors (Olsen, 2006).
Recently, a small molecule of 728 Da was isolated from the venom of the
theraphosid spider Vitalius dubius that blocked cholinergic receptors in vertebrate
nerve-muscle preparations (Rocha-E-Silva, 2013). The structure of the molecule
was not determined but its sensitivity to light and its small size suggests that it
likely to be a polyamine.
In addition to polyamines, other small molecules can be found in spider venoms,
such as amino acids (GABA, glutamate, acetylcholine), adenosine triphosphate
(ATP) and adenosine monophosphate (ADP) (for reviews, see (Estrada, 2007;
King and Hardy, 2013; Olsen, 2006, 2011). These molecules are endogenous
modulators of ion channels and receptors both in invertebrates and vertebrates but
their lack of selectivity makes these molecules unsuitable as drug leads (for a
review, see (King and Hardy, 2013)).
Most spider venoms are dominated by high concentrations of small cysteine-rich
peptides (King and Hardy, 2013). Most of these peptide toxins conform to a
structural scaffold known as an inhibitor cysteine knot (Nicke) or "knottin" fold (for
a review, see (King and Hardy, 2013)). Numerous spider-venom ICK peptides that

39
target ion channels and receptors have been identified in the venom of both
mygalomorph (Ayroza, 2012; Corzo, 2008) and araneomorph (Trachsel, 2012;
Vassilevski, 2013) spiders. In contrast, there is a small group of spiders whose
venoms contain cytolytic peptides that consist of linear amphipathic domains (for
reviews, see (King and Hardy, 2013; Windley, 2012a). These peptides, which are
devoid of disulphide bridges, have been identified in venoms of araneomorph
spiders from the families Ctenidae, Lycosidae and Oxyopidae (for reviews, see
(King and Hardy, 2013; Kuhn-Nentwig, 2003).
Apart from the discovery of huwentoxin-I (HWTX-I), which blocks vertebrate
cholinergic transmission, from venom of the theraphosid Haplopelma schmidti
(formerly Selenocosmia huwen Selenocosmia (now Ornithoctonus huwena) (Zhou,
1997), there is no evidence of spider-venom peptides with effects on cholinergic
receptors. In this work, a systematic approach was undertaken to screen 71 spider
venoms, extracted from representative species of both “primitive” and “modern”
spiders, for modulators of vertebrate neuronal nAChRs, using a high-throughput
FLIPRTETRA screening approach. It was hoped the data derived from the screening
of spider venoms would identify lead peptide toxins that could be further
characterised as potential therapeutic agents. However, no “hit” peptide toxin
could be identified. Nevertheless, cholinergic modulatory activity was identified in
venoms from a number of species and this data was used to map the taxonomic
distribution of modulators that target human neuronal nAChRs.
2.2 METHODS AND MATERIALS
This chapter describes the strategy followed to identify modulators of nAChRs in
crude spider venoms using a high-throughput FLIPR assay. The protocol used for
the high-throughput FLIPR assay was based on that described by Vetter and
Lewis (Vetter, 2012; Vetter and Lewis, 2010).
2.2.1 Tissue culture reagents
RPMI (Roswell Park Memorial Institute) medium, (+)-L glutamine; DPBS
(Dulbecco’s Phosphate Buffered Saline) without CaCl2 and MgCl2; 0.25 %

40
Trypsin/EDTA were from Gibco Life Technologies (Mulgrave, Vic, Australia).
Fetal Bovine Serum (FBS) were from Scientific Life (Clayton, Vic, Australia). PNU-
120596 (1-(5-chloro-2,4 -dimethoxyphenyl)-3-(5-methylisoxazol-3-yl) urea) was
from Sigma-Aldrich. FLIPRTETRA pipet tips, and Calcium-4 no-wash kit were from
Molecular Devices (Sunnyvale, CA, USA).
384-Well cell/imaging microplates (flat-well, clear-bottom black polystyrene) and
384-Well reagent microplates (clear-well, round-bottom polypropylene) were from
Corning® Incorporation (Corning, Lowell, MA, USA). Nicotine and choline were
from Sigma-Aldrich (Castle Hill, New South Wales, Australia), and were prepared
as a stock solution of 100 mM in MilliQ water. The composition of the Physiological
Salt Solution (PSS) used in all the assays was NaCl 140 mM, glucose 11.5 mM,
KCl 5.9 mM, MgCl2 1.4 mM, NaH2PO4 1.2 mM, NaHCO3 5 mM, CaCl2 1.8 mM,
HEPES 10 mM, and pH adjusted at 7.4 using NaOH (Vetter 2012).
2.2.2 Spider venoms
Crude venoms were extracted from spiders by Dr Volker Herzig (Institute for
Molecular Bioscience, The University of Queensland) via electrical stimulation of
the chelicerae. Venoms were then lyophilized and stored at –20°C. Before starting
fractionation of venoms using reverse-phase high-performance liquid
chromatography (RP-HPLC), crude venom (1 mg) was dissolved in 5% Buffer B
(90% CH3CN, 0.05% TFA in H2O).
Crude venom from 71 spiders was used in this research (see Appendix I for the list
of spiders whose venom was analysed for nAChR activity using the FLIPRTETRA
system). For the FLIPR screens, four different concentrations of spider venom
were prepared using MilliQ water as diluent. All venoms and toxins were dissolved
in MilliQ water before running the FLIPR calcium assay.
2.2.3 Cell lines and culture
In this project SH-SY5Y cells were used for all FLIPR screens because they are

41
reported to endogenously express the neuronal α7 nAChR and the “ganglionic”
nAChRs, named α3β2 and α3β4 (Dajas-Bailador, 2002). To facilitate the
description of these heteromeric receptors, an asterisk (*) is included to denote the
possibility of various β subunits in combinations with α3. The calcium responses
evoked by these subtypes after stimulation with choline and PNU-120596, and
nicotine, respectively, were characterized previously by Vetter and Lewis (Vetter
and Lewis, 2010).
SH-SY5Y human neuroblastoma cells (a kind gift from Victor Diaz, Max Planck
Institute for Experimental Medicine, Göttingen, Germany) were maintained at 37°C
with 5% (v/v) CO2 in RPMI medium which contained 2 mM L-glutamine and 15%
(v/v) FBS. Cells were passaged every 3–5 days using 0.25% trypsin/EDTA at a
dilution of 1:7. The SH-SY5Y cells were passaged a maximum of 10–20 times in
antibiotic-free medium, prior to use for assay. The SH-SY5Y cells used in this
project did not show irregular expression of α3* and α7 nAChRs as the control
calcium responses, which were obtained in each high-throughput screening, were
comparable to the control responses obtained by Vetter and Lewis (Vetter and
Lewis, 2010) during the characterization of α7 and α3* nAChRs expressed in SH-
SY5Y cells.
Two days prior to assay, a confluent culture of SH-SY5Y cells was collected by
trypsinization and resuspended in RPMI with 15% (v/v) FBS, then 20,000 SH-
SY5Y cells per well were seed in uncoated black–walled 384-well imaging plates
and maintained at 37°C and 5% CO2 for 48 h before running the experiment
(Vetter and Lewis, 2010). This time is the optimum to obtain a homogenous
monolayer of cells, thus avoiding problems associated with cellular differentiation
and changes in the levels of endogenous expression of receptors and ion
channels (Vetter, 2012).
2.2.4 Preparation of the Calcium 4-dye
A stock solution of calcium 4-dye was prepared according to the instructions of
Molecular Devices. Briefly, 10 ml of PSS is added to the component A to make a

42
10 x concentrated stock solution, then the solution is protected from light with
aluminium foil and stored in aliquots of 100 µL at –20°C. Aliquots can be reused,
at least once, if refrozen at –20°C immediately post-experiment.
2.2.5 High-throughput FLIPR calcium assay
The FLIPR-based calcium-influx assay has been shown to be a good high-
throughput approach to identify and characterize toxins derived from animal
venoms (Inserra, 2013; Jin, 2014; Wang, 2015). In this research project, a FLIPR-
based calcium assay was used to expedite the discovery of bioactive molecules
from complex mixtures of spider venoms.
71 spider venoms were screened using the fluorescent calcium FLIPR assay, and
either agonistic or antagonistic function at α7 and α3* nAChRs was measured
according to the methodology described by Vetter and Lewis (Vetter and Lewis,
2010). Briefly, 20,000 SH-SY5Y cells per well were seeded on uncoated black–
walled-384-well walled imaging plates and maintained at 37°C and 5% CO2 for 48
h before running the experiment. Once a 95% confluent monolayer was obtained,
the RPMI medium was removed from the cell plate by flicking it, the cells were
immediately loaded with Calcium 4-no-wash-dye (Molecular Devices), prepared as
a 1 x solution in PSS as indicated above, for 30 min at 37°C and 5% CO2. A
washing step after medium removal and after dye loading was not required as a
no-wash calcium dye was used and all the control calcium responses elicited by
cholinergic receptors stimulated by nicotine and choline were comparable to the
calcium responses obtained in previous work (Vetter and Lewis, 2010).
It is noteworthy that the 30 min of incubation time is optimal for dye-uptake, and
de-esterification of the acetoxymethyl ester (AM) by endogenous esterases
(Vetter, 2012; Vetter and Lewis, 2010). The cell preparation was always protected
from the light during the incubation to prevent photobleaching of the fluorophore.
After dye loading, the cell plate was transferred to the FLIPRTETRA fluorescence
plate reader and a protocol test signal was executed to obtain an appropriate level

43
of fluorescent baseline that is approximately 1,000 arbitrary fluorescence units with
a standard deviation less than 5%. In general, the excitation intensity was adjusted
in 80% and camera gain around 150. All fluorescent calcium responses were
recorded using a ‘two-addition’ protocol and a cooled CCD camera with excitation
wavelength of 470–495 nm and an emission wavelength of 515–575 nm for over a
period of 10 min.
After baseline fluorescence readings were recorded every second for 5 s, variable
amounts of crude venom (2 µg; 1 µg; 0.2 µg; 0.02 µg), venom fractions (1% of
venom fraction in relation to 1 mg of crude venom fractionated by RP-HPLC was
tested) or pure toxin (1% of pure toxin in relation to 1 mg of crude venom dissolved
in MilliQ water was tested) were added to the cell plate (addition 1) and the
calcium response was measured every second for 300 s. At the end of this time,
cells were subsequently stimulated with nicotine (30 µM) or choline (30 µM) +
PNU-120596 (10 µM) (addition 2), and readings continued every second for an
additional 300 s.
From the stock solution of nicotine (100 mM) and choline (100 mM), a working
solution for each agonist was prepared at 120 µM in MilliQ water. The final
concentration of nicotine or choline in each well of the plate that contains the SH-
SY5Y cells was 30 µM.
2.2.6 Activation of α3* nAChR by nicotine
The activation of the α3* nAChR by nicotine (30 µM) was measured using the
‘two-addition’ protocol mentioned above. This protocol is described in Section
2.2.8 and a schematic of the protocol is shown in Figure 2.1A.
The reagent plate with venoms or venom fractions takes a long time to prepare
when testing 71 venoms at different concentrations. To avoid loss of incubation
time once cells were loaded with calcium 4-dye, the reagent plate with venoms or
venom fractions was prepared before loading the cells.

44
After loading the SH-SY5Y cells, a fluorescent baseline was recorded for 5 s then
PSS was added to the cells (addition 1), followed by readings each second for
300s. During this recording time, calcium responses are not expected to be
different from the baseline. Finally, nicotine was added (addition 2) to yield a final
concentration in the assay of 30 µM, and the fluorescence was measured for 300
s. At the end of this time there is a transient increase in the fluorescent calcium
signal that corresponds to activation of α3* by the agonist nicotine.
To confirm that the observed calcium responses were elicited by activation of
nAChRs, d-tubocurarine, a non-selective blocker of nAChRs (Wenningmann and
Dilger, 2001), was added to the SH-SY5Y cells before stimulation of α3* by
nicotine. A stock solution of d-tubocurarine was prepared at 100 mM in MilliQ
water and its final concentration in the assay was 30 µM. All FLIPR experiments
included a positive control (calcium response elicited by activation of α3* by
nicotine) and a negative control that corresponds to inhibition of these responses
by d-tubocurarine.
Figure 2.1B shows the ‘two-addition’ protocol that was employed to detect
blockers of the calcium responses elicited by α3* nAChRs. After recording the
fluorescent baseline for 5 s, d-tubocurarine was added to the cells (addition 1),
followed readings each second for 300 s. No calcium responses are expected to
occur during this time. Then, a second fluorescent baseline was read for 5 s,
followed stimulation of α3* by nicotine (addition 2), and the fluorescent calcium
responses were measured for another 300 s. As expected, no increase in the
fluorescent calcium response was detected after nicotine addition. Figure 2.1D
shows a typical recording of the inhibition of nicotine-induced calcium responses
by d-tubocurarine.
The calcium responses caused by stimulation of α3* with nicotine were generally
smaller than those induced by choline. Furthermore, the kinetics of activation and
inactivation were slow and they can be detected on the FLIPR assay without
incubation of the SH-SY5Y cells with PNU-120596 (Figure 2.1C).

45
2.2.7 Activation of α7 nAChR by choline in the presence of the positive
allosteric modulator PNU-120596
The α7 nAChRs have fast kinetics of deactivation and long-lasting desensitization
that makes it difficult to measure its activity (Lopez, 1998). An experimental
strategy to detect its activity in SH-SY5Y cells was the addition of PNU-120596 (10
µM) in the Calcium 4-no-wash-dye solution, followed by incubation in SH-SY5Y
cells for 30 min at 37°C and 5% CO2 (Vetter and Lewis, 2010). The stock solution
of PNU-120596 was prepared at 100 mM in MilliQ water and 5% (v/v) DMSO, and
stored in aliquots of 20 µL at –20°C. Aliquots can be reused if refrozen at –20°C
immediately post-experiment.
The activation of the α7 nAChRs by choline (30 µM final concentration) was
measured basically using the same protocol as that for the study of the α3*
subtype. Briefly, after reading the fluorescent baseline for 5 s, PSS was added
(addition 1) to the cells and fluorescence measurements were made for a further
300s. Then, the agonist choline was added (addition 2) to stimulate the α7
nAChRs, followed by fluorescence measurements for another 300 s. As expected,
the activation of fluorescence nAChRs by choline elicited an increase in the
fluorescent calcium response that corresponds to an increase in the concentration
of intracellular calcium.
A typical trace of increase of intracellular calcium after stimulation of α7 nAChR by
choline is shown in Figure 2.1E. The positive allosteric modulator PNU-120596 (10
µM) was included in assays to delay the deactivation kinetics of α7 nAChR and
allow recording of the fluorescent Ca2+ responses (Vetter and Lewis, 2010).The
calcium responses are mostly caused by activation of α7 when PNU and choline
were used, although subtypes α3β2/α3β4/α5 are also activated and they were part
of the calcium response (Vetter and Lewis, 2010).
The same ‘two-addition’ protocol was used to confirm whether the calcium
responses were elicited by α7 nAChR. Briefly, d-tubocurarine (30 µM final
concentration) was added to the cells (addition 1) and the fluorescent calcium

46
response was read for 300 s, then choline was added (addition 2) to stimulate the
receptor, and a final reading of the fluorescence was carried out for another 300 s.
As expected, d-tubocurarine blocked the calcium response elicited by activation of
α7 nAChR (Figure 2.1F). The following Section 2.2.8 provides an explanation of
how the FLIPR assays were used to identify nAChR blockers and agonists in
arachnid venoms.
2.2.8 Identification of blockers and agonists for α3* and α7 nAChR using
the FLIPRTETRA system A ‘two-addition’ protocol was used to detect the presence of α3* and α7 blockers
in venoms or venom fractions. Crude venoms, venom fractions, or pure toxin were
added first to the cells, and then the fluorescence was measured for 300 s. Those
venoms or venom fractions that contain blockers inhibited the calcium responses
elicited by either α3* or α7 when they were stimulated by nicotine or choline
(addition 2), respectively.
On the other hand, an agonist compound contained in the venoms or venom
fractions caused an increase in the fluorescent calcium response before
stimulation of the receptors by agonists (addition 2). Inhibition by d-tubocurarine
(30 µM) was used to confirm that the observed calcium response was caused by
activation of the receptor rather than activation of voltage-gated calcium channels
(Sousa, 2013) or other endogenously expressed GPCRs or ion channels coupled
to an increase in intracellular Ca2+.

47
Figure 2.1. Basic two-addition FLIPR protocol to measure calcium responses evoked by activation of α7 or α3* nAChRs. (A) Schematic of the protocol to measure calcium responses caused by activation of the heteromeric α3* nAChR: SH-SY5Y cells were plated onto a 384-well plate and loaded with calcium 4-dye after discarding the RPMI medium. The method is based on two additions, and control experiments were always included in every assay. PSS or test compound was added first to the SH-SY5Y cells and the fluorescence measured for 300 s, then the α3* nAChR or α7 nAChR were stimulated by nicotine or choline (addition 2), respectively, and the resulting calcium responses were measured for another 300 s. (B) Schematic of protocol used to confirm that the calcium responses were elicited by activation of nAChRs. If the calcium responses observed in (A) were elicited by activation of α3* or α7 nAChRs they should be inhibited when d-tubocurarine (addition 1) was added before agonist addition, nicotine or choline, respectively. (C) Calcium responses recorded by the FLIPRTETRA system when α3* nAChRs were stimulated by nicotine (addition 2). (D) The calcium response was elicited by the activation of α3* because the addition of d-tubocurarine (addition 1) inhibited the calcium response elicited by the receptor when it was stimulated by nicotine (addition 2). (E) A typical calcium response elicited by activation of α7 nAChR using choline as agonist (addition 2). (F) The calcium response elicited by activation of subtype α7 nAChR using choline was inhibited when d-tubocurarine was added before the addition of choline (addition 2). The traces of control calcium responses are an average of one experiment with n = 4.
Ca2+dye+
SH(Sy5Y+cells+
addition+1+ addition+2+
PSS+ agonist+
A)#
Ca2++dye+
SH(Sy5Y+cells+
addition+1+ addition+2+
d(tubocurarine+ agonist+
B)#
0"
1"
2"
3"
4"
1" 151" 301" 451" 601"Maxim
un(calcium
(respon
se(
(RLU
)(
5me((seconds)(
addi5on(1((PSS)(
addi5on(2((Nico5ne)(
C)(
0"
1"
2"
3"
4"
1" 151" 301" 451" 601"Maxim
un(calcium
(respon
se(
(RLU
)(5me((seconds)(
addi5on(1((d=tubocurarine)(
addi5on(2((Nico5ne)(
D)(
0"
2"
4"
6"
8"
1" 151" 301" 451" 601"Maxim
un(calcium
(respon
se(
(RLU
)(
5me((seconds)(
F)(
addi5on(1((d=tubocurarine)(
addi5on(2((Choline)(
0"
2"
4"
6"
8"
1" 151" 301" 451" 601"Maxim
un(calcium
(respon
se(
(RLU
)(
5me((seconds)(
addi5on(2((Choline)(
E)(
addi5on(1((PSS)(

48
Figure 2.2. ‘Two-addition’ FLIPR protocol used to identify nAChR agonists. (A) Venom was added (addition 1), then fluorescent responses were measured for 300 s. After the cells were stimulated by nicotine or choline (addition 2), fluorescence measurements were continued for another 300 s. If any venom compound contains a blocker, the fluorescent calcium response induced by nicotine or choline (addition 2) will be inhibited. (B) If any venom compound contains an agonist there will be an increase in the fluorescence after addition 1. d-tubocurarine (30 µM) is added during the loading process, then venom is added (addition 1), if the following response is inhibited then the response was caused by activation of nAChRs. Finally, addition 2 occurs, followed a second reading for 300 s. (C) 1 µg of crude venom from H. hainanum was added (addition 1) to the SH-SY5Y cells, then the calcium fluorescent response was measured for 300 s. After choline was added (addition 2), the fluorescent signal was measured for other 300 s. (D) To confirm that the increase in fluorescence after venom addition was caused by cholinergic activity, d-tubocurarine (30 µM) was included in the Calcium 4-dye solution, and added to the cells. Then, the crude venom (H. hainanum) was added (addition 1) and the fluorescent response was measured. The response was inhibited, and after choline addition (addition 2) the response remained inhibited. The venom was assayed in the presence of PNU-120596 (10 µM). The traces shown in panels (C) and (D) are averages with n = 3.
0
1
2
3
1 151 301 451 601
Max
imun
calc
ium
resp
onse
(RLU
)
time (seconds)
crude venom H. hainanum 1 µg addition 1
0
1
2
3
1 151 301 451 601
Max
imun
calc
ium
resp
onse
(RLU
)
time (seconds)
crude venom H. hainanum 1 µg addition 1 choline 30 µM
addition 2 choline 30 µM addition 2
C) D)
Ca2+dye(
SH+Sy5Y(cells(
addition(1( addition(2(
Venom/(fraction(venom( agonist(
Ca2+(dye((+blocker(
addition(1( addition(2(
agonist(Venom/(fraction(venom(
A)( B)(

49
2.2.9 Data analysis
The 71 spider venoms were tested in triplicate in at least three independent
experiments. All data was analysed as followed: Raw fluorescence values
obtained after venom addition were subtracted from the baseline fluorescence
values measured during the reads between 0 and 5 s after starting the assay. In
addition, the raw data readings obtained after stimulation of the receptors by
nicotine or choline were subtracted from the baseline raw fluorescent values
registered between 300 and 305 s. Then, the resulting raw fluorescent reads were
converted to maximum calcium responses by calculating in relative light units the
maximum values reached over the baseline reads using the ScreenWorks,
software, version 3.1. Once the data was corrected using the baseline fluorescent
reads, it was exported and analysed using the software Prism, version 5.0.
Data are expressed as mean ± standard error of the mean (SEM) from n = 3
independent experiments or as the mean ± SEM of the values obtained in the
three independent experiments with a total n = 9.
FLIPR data were normalized to the maximum fluorescence calcium response
evoked by activation of α3* nAChR with nicotine (30 µM) or by activation of α7
nAChR with choline (30 µM) and indicated as FMax (%)
The fluorescent traces from the FLIPR data are reported as calcium response over
baseline that is corrected previously by subtracting the average of the
fluorescence elicited by the negative control (only components of buffer PPS)
using ScreenWorks software. This eliminates the artefacts caused by buffer
addition.
2.2.10 Statistical Analyses
Comparative analysis of the mean of calcium responses induced by crude venoms
of each species on α3* and α7 nAChRs versus control responses was carried out

50
through a standard one-way ANOVA and non-parametric statistical analysis, using
the analysis tool of GraphPad Prism®, version 5.0.
2.3 RESULTS
A high-throughput FLIPR screen was carried out to examine the effect of 12
araneomorph and 59 mygalomorph venoms (see Appendix I, Table 1-2 for a list of
species) on the calcium responses evoked by activation of α3* and α7 nAChRs
endogenously expressed in SH-SY5Y cells. Following the initial screen, at least
two additional screens were performed to enhance the statistical significance of
the data, although the data for each species was highly reproducible, indicating
that the venom composition of each species is constant across different pools of
venom. To facilitate analysis of the 71 spider venoms, the resulting data from
araneomorph spiders is presented first in accordance with an Araneae
phylogenetic tree. The second group corresponds to mygalomorph spiders, and
the data was compared in accordance with the geographical origin (state and
country) of each species. Then, common and different elements between both
suborders were described and later discussed (see discussion section) to
determine possible mechanisms of evolution of modulators for vertebrate neuronal
cholinergic receptors.
2.3.1 Screening of 71 spider venoms using the high-throughput FLIPRTETRA system
In all cases, unless specified, 2 µg of crude venom from each spider was tested
against α3* and α7 nAChRs endogenously expressed in human SH-SY5Y cells.
For the screening of these 71 spider venoms, a two-addition protocol was used
(for further details see the methods section).
2.3.2 Screening of venoms from “modern” spiders (Infraorder Araneomorphae)
A phylogenetic tree of the infraorder Araneomorphae is shown in Figure 2.3, each
superfamily is indicated on the main branches of the tree, and the name of each

51
representative spider considered in this study is indicated beside the superfamily
name.
To describe the results obtained with araneomorph spider venoms, the spiders
were group in three groups, and are described in order of evolutionary
appearance. The first group corresponds to the superfamily Austrochiloidea, and
the one species from this taxon is Hickmania troglodytes, found in Tasmania. The
spider makes and hangs beneath extensive sheet webs, and occasionally wraps
prey after biting. The crude venom of H. troglodytes at 2 µg did not modify the
calcium responses evoked in SH-SY5Y cells by the agonists nicotine and choline
(Figure 2.3A).
According to the tree, the superfamily Araneoidea appeared in the Cretaceous 145
million years ago after the family Austrochiloidea. Crude venoms from four
exemplars of the superfamily Araneoidea were used in this work: Cyrtophora
moluccensis, Eriophora transmarina, Nephila pilipes and Nephila plumipes. This is
a large taxon that consists of approximately 29% of the spiders described so far
(Griswold, 2005). Venom from E. transmarina and the two Nephila species
blocked both the α3* and α7 nAChRs with high potency whereas C. moluccensis
specifically blocked only the heteromeric α3* nAChRs (Figure 2.3A). This venom
had no effect on the choline response (Figure 2.3B), indicating that blockers are
likely to target mainly the α3*. In contrast, N. pilipes and N. plumipes venom had
equally potent inhibitory effects on α3* and α7 nAChRs, see Figures 2.3A and
2.3B, respectively.

52
Figure 2.3. Screening of venoms from “modern” spiders against human α3* and α7 nAChRs using a high-throughput FLIPR assay. The left panel shows a phylogenetic tree of the infraorder Araneomorphae. The name of the spider species whose venoms were used in this study is denoted next to the branch of each family. The tree was adapted from (Coddington and Levi, 1991); ma = millions of years. On the right panel is shown: A) 2 µg of crude venoms of the twelve araneomorph spiders were assayed in the absence of PNU-120596 (10 µM). B) 2 µg of venoms from the same araneomorph spiders were assayed in the presence of PNU-120596 (10 µM). Venom from H. troglodytes (Family Austrochiloidea) had no effect on α3* and α7 nAChRs in the absence and presence of PNU-120596, respectively. Venoms from E. transmarina, N. pilipes, and N. plumites (Superfamily araneoidea) had a potent blocking effect on α3* and α7 subtypes with the exception of venom from C. moluccensis that only had potent blocking effect on α3* nAChRs in the absence of PNU-120596. As seen on the phylogenetic tree (left panel), spiders from the RTA clade evolved after the araneoid spiders; and venoms from Sparassidae sp. (1) and (2), and Lycosidae sp. showed a partial blocking effect on α3* and α7 subtypes without and with PNU-120596, respectively, while venoms from H. jugulans, and Ancylometes sp. had no effect on nAChRs, although venom from V. sylvestriformis only had a partial blocking effect on α3* nAChRs in the absence of PNU-120596. Venom from M. australianus blocked completely α3* and α7 nAChRs in the absence and presence of PNU-120596, respectively. The data on panel A was normalized in relation to the maximum calcium response elicited with nicotine addition (30 µM) (positive control) in the absence of PNU-120596 (10 µM) and expressed in percentage while data from panel B was normalised in relation to the maximum
Austrochiloidea.
Araneoidea.
RTA.clade.
Dionychans.
Lycosidae.
ENTELEGYNAE(
ARAN
EOMORPHAE((
(“modern”(SPIDERS)(
Pisauridae.
Agelenidae.
Ctenidae.
Sparassidae.
Therididae.
Nephilidae.
H.#Jugulans#
Sparassidae#sp.1#Sparassidae#sp.2#
Lycosidae#sp.##
Ancylometes#sp.#V.#sylvestriformis#
M.#australianus#
H.#troglodytes#
C.#Moluccensis##E.#transmarina#N.#Pilipes#.N.#plumites#
ma.
200.
145.
65.
2.
Jurassic( Cretaceous(
d-tuboc
urarin
e (30
µM)
H. trog
lodyte
s
C. molu
ccensis
E. tran
smari
na
N. pilip
es
N. plumite
s
Sparas
sidae
sp. 1
Sparas
sidae
sp. 2
Lycosid
ae sp.
H. jugu
lans
Ancylom
etes s
p.
V. sylve
strifo
rmis
M. austr
alian
us
0
50
100
150
2 µg crude venom / araneomorph spider
Nor
mal
ized
calc
ium
resp
onse
(%)
PNU-120596 (10 µM)
********
****
*** ****
*
****
No effect on nAChRs
Partial effect on nAChRs
RTA CladeB)
d-tuboc
urarin
e (30
µM)
H. trog
lodyte
s
C. molu
ccensis
E. tran
smari
na
N. pilip
es
N. plumite
s
Sparas
sidae
sp. 1
Sparas
sidae
sp. 2
Lycosid
ae sp
.
H. jugu
lans
Ancylom
etes s
p.
V. sylve
strifo
rmis
M. austr
alian
us
0
50
100
150
2 µg crude venom / araneomorph spider
Nor
mal
ized
cal
cium
res
pons
e (%
)
***************
**
****
*
**
Partial effect on nAChRs
**
No effect on nAChRs
RTA CladeA)

53
calcium response elicited with choline addition (30 µM) (positive control) in the presence of PNU-120596. The negative control corresponds to the inhibition of the calcium response of nAChRs by d-tubocurarine addition at 30 µM in the absence or presence of PNU-120596 (10 µM). The values are averages of three independent experiments with n = 3. Dotted lines represent the control response in the absence of venoms. Using the unpaired t-test, the means between the effect of each venom and the control response without venom were compared. The number of (*) indicates the degree of significance at P < 0.05 probability level. Those values that lack of significance do not have the symbol (*). The third group of spiders used herein belong to the Retrolateral Tibial Apophysis
(RTA) clade, and venoms from six exemplars of this group were assayed. Table
2.1 list these species and their geographic distribution. Most species are from
Australasia, although the two ctenid species (Ancylometes sp. and Viridasius
sylvestriformis) are from Central and South America. Of the six venoms tested, the
two sparassid venoms (Sparassidae sp. 1 and Sparassidae sp. 2), and the one
lycosid venom (Lycosidae sp.) showed to have a partial blocking effect on α3* and
α7, while one sparassid venom (H. jugulans), and the two ctenid venoms had no
effect on vertebrate α3* and α7 nAChRs, although venom from V. sylvestriformis
had a partial blocking effect on α3*. In striking contrast, venom from the giant
water spider Megadolomedes australianus (family Pisauridae) blocked both
nAChR subtypes with high potency.

54
Table 2.1. Taxonomic distribution of nAChR modulators from araneomorph spider venoms. Two species Sparassidae sp. were assayed; the numbers 1 and 2 indicate they were collected in two different areas in PNG. Venoms with high blocking effect are indicated with two symbols denoted as √√, while those with partial effect are indicated with one √. The green area indicates that other effects on the venom is not observed.
Spider Effect on human neuronal α3* and α7 nAChRs Block of α3*
& α7 Block of α3*
No effect
Location
Infraorder Araneomorphae Family Austrochiloidea Hickmania troglodytes
√ Tasmania, Australia
Family Araneoidea Cyrtophora moluccensis
√√ Gardens across South East and coast from Queensland, Northern and western Australia
Eriophora transmarina
√√ Gardens across eastern and southern Australia
Nephila pilipes √√ Gardens Northern Australia, PNG, China, Japan
Nephila plumites √√ Sydney region RTA Clade Family Sparassidae Heteropoda jugulans
√ Queensland, Australia
Sparassidae sp. 1
√ Papua New Guinea
Sparassidae sp. 2
√ Papua New Guinea
Family Ctenidae Ancylometes sp. √ Central and South
America Viridasius sylvestriformis
√ Amazon, South America
Family Lycosidae Lycosidae sp. √ Western Australia Family Pisauridae Megadolomedes australianus
√√ South Eastern Australia

55
2.3.3 Screening of venoms from “primitive” spiders (Infraorder Mygalomorphae)
Mygalomorphs are the oldest group of spiders. They are known as “primitive”
spiders because they retain some ancestral morphological features, such as
downward pointing chelicera (Coddington and Levi, 1991). In this study, venoms
were derived from 59 mygalomorph spiders. The infraorder Mygalomorphae is
comprised of 15 taxonomic families (Bond, 2012), of which two were considered
here, namely Hexathelidae (1 species) and Theraphosidae (58 species).
Four mygalomorph venoms were found to contain nAChR agonists (Section 2.9.4),
24 venoms selectively blocked α3* nAChRs (Section 2.9.5), 26 venoms inhibited
both α3* and α7 nAChRs (Section 2.9.6), and four mygalomorph venoms were
found to have no effect on α3* or α7 nAChRs (Section 2.9.7). Each of these four
groups is discussed in the following sections.
2.3.4 Identification of α7/α3* nAChR agonists in the venom of theraphosid
spiders
The two-addition protocol was used to identify potential agonists of vertebrate
nAChRs. Addition of venom (1 or 2 µg per well) from Haplopelma hainanum,
Pamphobeteus sp., Sericopelma rubronites and Xenesthis sp. caused an
increased calcium response in the FLIPR assay both in the absence and presence
of PNU (Table 2.2). Based in a previous FLIPR assay performed by Dr Vetter
(data not showed), the fluorescent responses evoked by 1 µg of venom from H.
hainanum, Pamphobeteus sp., Sericopelma rubronites and in the absence of PNU
were not abolished with addition of d-tubocurarine, indicating that these venoms
lack of agonists of α3* nAChRs. Interestingly, the same venoms at 1 µg were
potentiated in the presence of PNU, although were no assayed in the presence of
d-tubocurarine (data not showed). Based on these previous results, I evaluated if
the calcium responses evoked for the same venoms at 1 µg and in the presence
of PNU were related to activation of the α7 nAChRs (Table 2.3). The venom
responses from H. hainanum, Pamphobeteus sp., S. rubronites, and Xenesthis sp.

56
were assayed with the previous addition of d-tubocurarine (Figure 2.4A-D). The
traces derived from the FLIPRTETRA system are presented to show the strategy
used to identify agonists in these venoms. After crude venom addition (addition 1)
at 1 µg, there was an increase of the calcium response that was extinguished at
the end of the 300 s (Figure 2.4B-D) although the venom from H. hainanum
caused an increase of the fluorescence that was sustained during this period of
time (0–300 s) (Figure 2.4A). Then, after choline addition (addition 2), the calcium
response was not different from that evoked by stimulation of the α7 nAChRs by
choline (positive control) (Figure 2.4B-D). Interestingly, venom from H. hainanum
caused complete inhibition of the calcium response evoked by choline stimulation
of the α7 nAChR (see Figure 2.4A). A further analysis was not carried out to
determine if the differences of calcium responses in the presence of PNU and
addition of d-tubocurarine were statistically significant due to the limitation of
amount of these venoms, although the venom from H. hainanum is likely to have
an agonist compound.

57
Figure 2.4. Agonism of human nAChRs by theraphosid venoms. All traces are averages of the maximum calcium responses over baseline (with n = 3) elicited by crude venom from (A) Haplopelma hainanum, (B) Pamphobeteus sp., (C) Sericopelma rubronites, and (D) Xenesthis sp.. All responses elicited between 5 s and 300 s are caused by venom addition. The responses after choline addition were measured between 300 s and 600 s. All crude venoms were tested at 1 µg/well in the presence of PNU-120596 (10 µM). Dotted lines of each graphic correspond to averages of calcium responses over baseline (with n=3) elicited by the venoms in the presence of d-tubocurarine (30 µM) and PNU-120596 (10 µM).
0
1
2
3
1 151 301 451 601
Max
imun
cal
cium
res
pons
e (R
LU)
time (seconds)
crude venom Xenesthis sp. 1 µg + PNU-120596 (10 µM)
crude venom Xenesthis sp. 1 µg + PNU-120596 (10 µM) + d-tubocurarine (30 µM)
time (seconds)
0
1
2
3
1 151 301 451 601
Max
imun
calc
ium
resp
onse
(RLU
)
time (seconds)
crude venom S. rubronitens 1 µg + PNU-120596 (10 µM) crude venom S. rubronitens 1 µg + PNU-120596 (10 µM) + d-tubocurarine (30 µM)
0
1
2
3
4
1 151 301 451 601Max
imun
calc
ium
resp
onse
(R
LU)
time (seconds)
crude venom Pamphobeteus sp . 1 µg + PNU-120596 (10 µM) crude venom Pamphobeteus sp. 1 µg + PNU-120596 (10 µM) + d-tubocurarine (30 µM)
0
1
2
3
1 151 301 451 601
Max
imun
cal
cium
resp
onse
(R
LU)
time (seconds)
crude venom H. hainanum 1 µg + PNU-120596 (10 µM) crude venom H. hainanum 1 µg + PNU-120596 (10 µM) + d-tubocurarine (30 µM) A)#
C)# D)#
B)#

58
Table 2.2. Apparent agonistic effect caused by venoms from four theraphosid spiders. After venom addition, the fluorescent calcium responses evoked by nAChRs in the absence or presence of PNU were converter into area under curve using ScreenWorks software, version 3.1. The values are averages of three independent experiments with n = 3. The mean of the control nicotine response was 509 ± 50.40 while the control calcium response elicited by choline was 1088 ± 38.31 Spider Calcium response
over baseline after venom addition at 2 µg
Calcium response over baseline after venom addition at 2 µg when PNU-120596 (10 µM) was included in the assay
Haplopelma hainanum
345.6 ± 136 822.2 ± 127
Pamphobeteus sp. 542.2 ± 164.2 517 ± 179 Sericopelma rubronites
241.1 ± 101.5 696 ± 137
Xenesthis sp. 325.5 ± 151.4 678.8 ± 80 Table 2.3. Mygalomorph spiders with venoms that are likely to contain agonists of α7 nAChRs. N.D indicates the effect was not determined and a further analysis is required to confirm that the venom effect is via activation of α7 nAChRs in the presence of PNU.
Location and affect on human α7 nAChRs Spider Agonistic effect Location Haplopelma hainanum √ Southern China and
Vietnam Pamphobeteus (platyomma) sp. (F)
N.D Ecuador
Sericopelma rubronites (F) N.D Panama Xenesthis sp. (F) “white colour”
N.D Colombia
2.3.5 Venoms that selectively inhibit vertebrate α3* nAChRs
24 theraphosid venoms were found to selectively block human α3* nAChRs
(Figure 2.5A). The 24 species are listed in Table 2.4 along with their geographic
distribution. These venoms did not have a substantial effect on calcium responses
in the FLIPR assay when PNU-120596 was included (Figure 2.5B).

59
Figure 2.5. Theraphosid spider venoms that preferentially block α3 nAChRs. First screening of venom from “primitive” spiders against human α3* and α7 nAChRs was carried out by using a high-throughout calcium assay. The calcium responses elicited by crude venom was calculated as the area under fluorescence curve normalized to the maximum α3* response evoked by nicotine (A) or stimulation of α7 by choline (B). Each value corresponds to averages of normalized calcium responses over baseline of at least three independent experiments and n = 3. All venoms were tested at 2 µg/well, in the absence (A) and presence (B) of PNU-120596. For a species identification see Table 2.4. Dotted lines represent the control response in the absence of venoms. On graphic A and B, the control responses corresponds to stimulation of α3* with nicotine (30 µM), and α7 choline (30 µM), respectively. Using one-way ANOVA, the means between each venom and the control responses (nicotine response were compared). The number of (*) indicates the degree of significance at P < 0.05 probability level. Those values that lack of significance do not have the symbol (*).
Table 2.4. Theraphosid spiders with venoms that selectively block α3* nAChRs. Species with characteristic colors are indicated. (M) and (F) refer to gender. The numbers beside the name of each theraphosid spider indicates that they are individuals from the same species, although were collected from distinct localities. Number Theraphosid Geographic distribution
1-A Avicularia metallica (F)
Guyana (coast), Suriname (coast)
2-B Avicularia ulrichea (F) All Brazil
3-C Avicularia sp. 1 (M) “purple” Peru-Amazonas
4-D Coremiocnemis tropix Australia
d-tu
bocu
rarin
e1-
A2-
B3-
C4-
D5-
E6-
F7-
G8-
H 9-I
10-J
11-K
12-L
13-M
14-N
15-O
16-P
17-Q
18-R
19-S
20-T
21-U
22-V
23-W
24-X
0
50
100
150
Nor
mal
ized
cal
cium
resp
onse
(%)A)
**** ****
****
**** *******
********
**
****
****
**********
*******
*****
******
****
****
***
****
****
d-tu
bocu
rarin
e1-
A2-
B3-
C4-
D5-
E6-
F7-
G8-
H 9-I
10-J
11-K
12-L
13-M
14-N
15-O
16-P
17-Q
18-R
19-S
20-T
21-U
22-V
23-W
24-X
0
50
100
150
Nor
mal
ized
cal
cium
resp
onse
(%) PNU-120596 (10 µM)
****
****
*******
B)

60
5-E Chilobrachys sp. 1 Malaysia-Penang island
6-F Grammostola grossa 8 (F)
Pampas from Argentina and Paraguay. Brazil south and north of Paraguay
7-G Grammostola iheringi (M) Brazilian south, Entre Rios
8-H Euathlus sp. 1 (F) “green” Chile
9-1 Euathlus sp. 2 (F) “red” Chile
10-J Euathlus vulpinus (F) “crème” Chile highland
11-K Orphnaecus philippinus
Philippines
12-L Orphnaecus sp. Philippines-Paqnai-island
13-M Paraphysa scrofa (F) Chile
14-N Pelinobius muticus West Papua New-Guinea (PNG)
15-O Poecilotheria striatus Sri Lanka, India
16-P Pterinuchilus murinus
Central, Eastern, Southern Africa
17-Q Selenocosmia crassipes
Australia-East Coast Queensland
18-R Selenocosmia effera East PNG
19-S Selenocosmia sp. 1 PNG
20-T Selenocosmia sp. 2 PNG
21-U Selenotholus foelschei Australia
22-V Thrixopelma cyaneolum (F) Peru
23-W Thrixopelma sp. (F) Peru – Cajamarca
24-X Thrixopelma lagunas sp. (F)
Peru – Lagunas

61
2.3.6 Venoms that non-selectively block both α3* and α7 nAChRs
26 theraphosid venoms were found to non-selectively block both α3* (Figure 2.6A)
and α7 nAChRs (Figure 2.6B). The 26 species from which these venoms were
obtained are listed in Table 2.5 along with their geographic distribution.
Figure 2.6. Many theraphosid venoms non-selectively block both α3* and α7 nAChRs. Normalised calcium responses obtained from FLIPR screen of venoms in the absence (A) and presence (B) of PNU-120596. Each value corresponds to averages of normalized calcium responses over baseline of at least three independent experiments and n = 3. For a list of species see Table 2.5. Dotted lines represent the control response in the absence of venoms. On graphic A and B, the control responses corresponds to stimulation of α3* with nicotine (30 µM), and α7 choline (30 µM), respectively. Using one-way ANOVA, the means between each venom and the control responses (nicotine response were compared). The number of (*) indicates the degree of significance at P < 0.05 probability level. Those values that lack of significance do not have the symbol (*).
d-tub
ocura
rine1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 2021 22 23 24 25 26
0
50
100
150
Nor
mal
ized
cal
cium
resp
onse
(%)
***
A)
********* ********* ************************
********* ************
***
************
d-tub
ocura
rine1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
0
50
100
150
Nor
mal
ized
cal
cium
resp
onse
(%)
PNU-120596 (10 µM)
* ***********
*
***
**** ********
B)
****
****
****
*
**
***********
********
******** ********
****
****
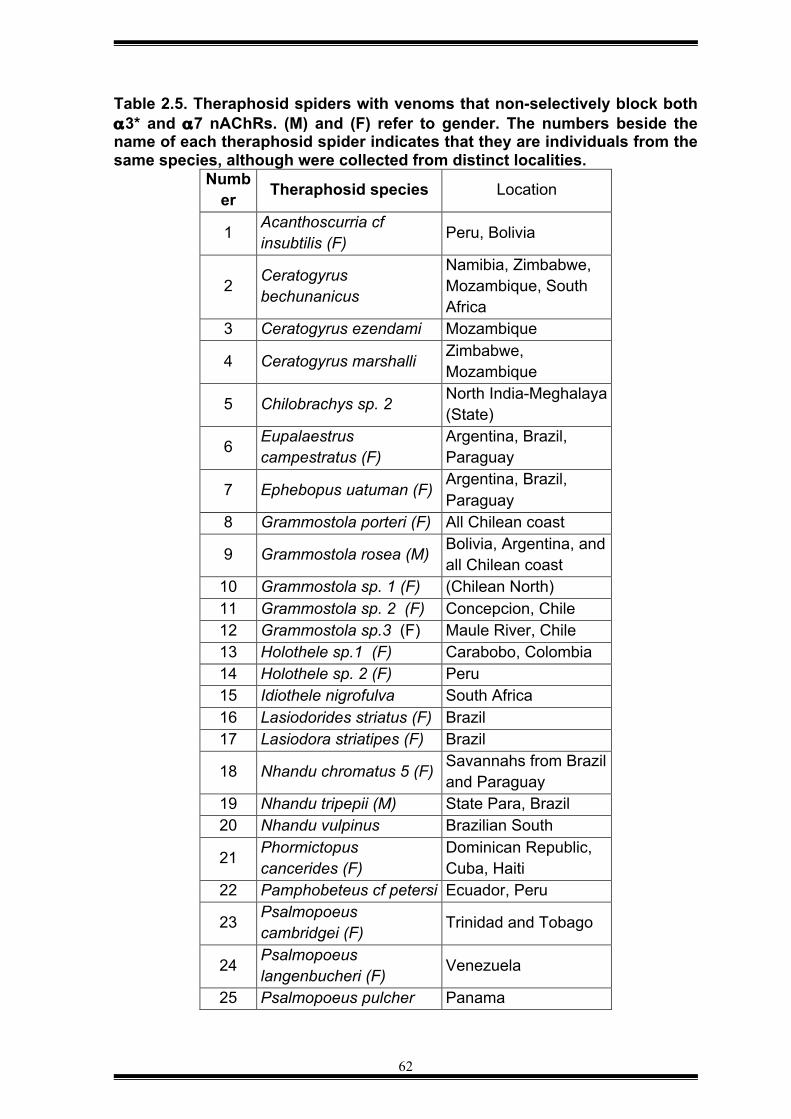
62
Table 2.5. Theraphosid spiders with venoms that non-selectively block both α3* and α7 nAChRs. (M) and (F) refer to gender. The numbers beside the name of each theraphosid spider indicates that they are individuals from the same species, although were collected from distinct localities.
Number Theraphosid species Location
1 Acanthoscurria cf insubtilis (F) Peru, Bolivia
2 Ceratogyrus bechunanicus
Namibia, Zimbabwe, Mozambique, South Africa
3 Ceratogyrus ezendami Mozambique
4 Ceratogyrus marshalli Zimbabwe, Mozambique
5 Chilobrachys sp. 2 North India-Meghalaya (State)
6 Eupalaestrus campestratus (F)
Argentina, Brazil, Paraguay
7 Ephebopus uatuman (F) Argentina, Brazil, Paraguay
8 Grammostola porteri (F) All Chilean coast
9 Grammostola rosea (M) Bolivia, Argentina, and all Chilean coast
10 Grammostola sp. 1 (F) (Chilean North) 11 Grammostola sp. 2 (F) Concepcion, Chile 12 Grammostola sp.3 (F) Maule River, Chile 13 Holothele sp.1 (F) Carabobo, Colombia 14 Holothele sp. 2 (F) Peru 15 Idiothele nigrofulva South Africa 16 Lasiodorides striatus (F) Brazil 17 Lasiodora striatipes (F) Brazil
18 Nhandu chromatus 5 (F) Savannahs from Brazil and Paraguay
19 Nhandu tripepii (M) State Para, Brazil 20 Nhandu vulpinus Brazilian South
21 Phormictopus cancerides (F)
Dominican Republic, Cuba, Haiti
22 Pamphobeteus cf petersi Ecuador, Peru
23 Psalmopoeus cambridgei (F) Trinidad and Tobago
24 Psalmopoeus langenbucheri (F) Venezuela
25 Psalmopoeus pulcher Panama

63
(F) 26 Theraphosinae sp. (F) Chile
2.3.7 Theraphosid venoms with no apparent effect on nAChRs Figure 2.7 summarizes FLIPR data obtained with venom from four American
theraphosids (Aphonopelma crinitum, Avicularia sp., Homoeomma sp. and
Pseudhapalopus sp.; Table 2.6). These venoms had only a minor effect on α3*
nAChRs (Figure 2.7A) and no effect on α7 nAChRs (Figure 2.7B).
Figure 2.7. Four tarantula venoms without potent modulators of α3* or α7 nAChRs. Normalized FLIPR responses resulting from a screen of crude venom against human α3* (A) and α7 nAChRs (B). All venoms were tested at 2 µg/well. Each value corresponds to averages of normalized calcium responses over baseline of at least three independent experiments and with n = 3. For legend see table 2.5. Dotted lines represent the control response in the absence of venoms. On graphic A and B, the control responses corresponds to stimulation of α3* with nicotine (30 µM), and α7 choline (30 µM), respectively. Using one-way ANOVA, the means between each venom and the control responses (nicotine response were compared. The number of (*) indicates the degree of significance at P < 0.05 probability level. Those values that lack of significance do not have the symbol (*).
d-tub
ocura
rine (
30 µM)
Aphon
opelm
a crin
itum
Avicula
ria sp
. 2
Homoe
omma s
p.
Pseuda
phalo
pus s
p.
0
50
100
150
Nor
mal
ized
resp
onse
(%)
*** **** ****
*
****
A)
d-tub
ocura
rine (
30 µM)
Aphon
opelm
a crin
itum
Avicula
ria sp
. 2
Homoe
omma s
p.
Pseudh
apalo
pus s
p.
0
50
100
150
Nor
mal
ized
resp
onse
(%)
****
B) PNU-120596 10 µM

64
Table 2.6. Venoms from American theraphosids that lack of modulators of human α3* and α7 nAChRs.
Theraphosid spider Geographic location Aphonopelma crinitum
(F) Northern Mexico
Avicularia sp. (F) 2 Bolivia-Rio Madre Homoeomma sp. (blue) Peru
Pseudhapalopus sp. (F)
South America
2.3.8 Effect of non-theraphosid mygalomorph venom on vertebrate nAChRs
Unfortunately, I only had access to one non-theraphosid mygalomorph venom for
this study, namely Macrothele gigas from the family Hexathelidae (see condensed
phylogenetic tree in Figure 2.8). M. gigas venom blocked both the α3* and α7
nAChR subtypes. At 2 µg, M. gigas venom reduced the nicotine-evoked calcium
response in the FLIPR assay by 28 ± 22% and the response evoked by choline in
the presence of PNU-120596 by 46 ± 27%.
2.3.9 Taxonomic distribution of nAChR modulators within Araneae
Figure 2.8 summarizes the data obtained from the high-throughput FLIPR-based
screen of spider venoms against human nAChRs. Figure 2.8A shows the
distribution of blockers, agonists and inactive venoms within the infra-orders
Mygalomorphae and Araneomorphae, while Figure 2.8B shows the distribution of
nAChR modulators mapped onto a condensed phylogenetic tree of spiders (order
Araneae).
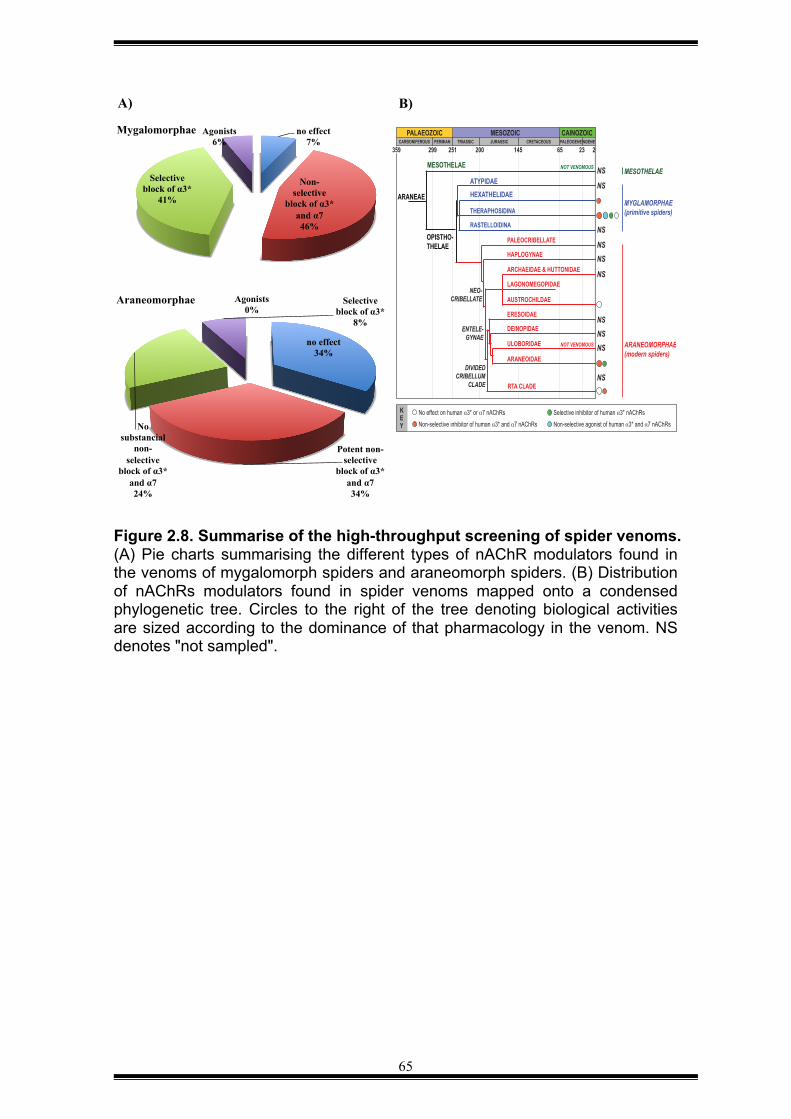
65
Figure 2.8. Summarise of the high-throughput screening of spider venoms. (A) Pie charts summarising the different types of nAChR modulators found in the venoms of mygalomorph spiders and araneomorph spiders. (B) Distribution of nAChRs modulators found in spider venoms mapped onto a condensed phylogenetic tree. Circles to the right of the tree denoting biological activities are sized according to the dominance of that pharmacology in the venom. NS denotes "not sampled".
PALAEOZOIC MESOZOIC CAINOZOICCARBONIFEROUS PERMIAN TRIASSIC JURASSIC CRETACEOUS PALEOGENE NGENE
299 251 200 145 65 23 2
ATYPIDAEHEXATHELIDAE
THERAPHOSIDINA
RASTELLOIDINA
LAGONOMEGOPIDAE
MESOTHELAE
ARANEAE
OPISTHO-THELAE
NOT VENOMOUS
ERESOIDAE
DEINOPIDAE
ULOBORIDAE
ARANEOIDAE
AUSTROCHILDAE
ARCHAEIDAE & HUTTONIDAE
HAPLOGYNAE
PALEOCRIBELLATE
RTA CLADE
ENTELE-GYNAE
NEO-CRIBELLATE
DIVIDEDCRIBELLUM
CLADE
MYGLAMORPHAE(primitive spiders)
ARANEOMORPHAE(modern spiders)
NOT VENOMOUS
MESOTHELAE
359
NS
NS
NS
NSNS
NS
NSNSNS
NS
No effect on human _3* or _7 nAChRs
Non-selective inhibitor of human _3* and _7 nAChRs
Selective inhibitor of human _3* nAChRs
Non-selective agonist of human _3* and _7 nAChRs
KEY
no effect 7%
Non-selective
block of α3* and α7 46%
Selective block of α3*
41%
Agonists 6%
Mygalomorphae
no effect 34%
Potent non-selective
block of α3* and α7 34%
No substancial
non-selective
block of α3* and α7 24%
Selective block of α3*
8%
Agonists 0%
Araneomorphae
A) B)

66
2.4 DISCUSSION
Today, it is recognized that an imbalance of the cholinergic pathways in humans
may lead to several pathologies (Picciotto, 2012). Therefore, nAChRs are now
molecular targets; and several studies have been carrying out to understand their
mechanisms of regulation, but few advances have been made (Bencherif, 2014;
Pammolli, 2011; Russo, 2014). In the field of agronomy, invertebrate nAChRs are
also targets of neonicotinoids that are used to control insect pests (Fischer, 2014;
Matsuda, 2009); and these are one of the most successful classes of chemical
insecticides ever developed, although it is no selective for insect pests, it has been
found to be lethal to other insects of importance in agriculture such as pollinators
(Moffat, 2015). Part of the problem is that many studies of cholinergic receptors
are limited by the number and type of selective modulators for nAChRs (Bauwens,
2015; Gotti, 2007).
Since spiders are the most successful insect predators on the planet (for a review
see (King and Hardy, 2013)), it would be surprising if they had not developed
nAChR modulators as part of their venom arsenal. To date, however, there are
very few reports of this pharmacological activity in spider venoms. We therefore
decided to screen spider venoms for nAChR modulators. Since cloned insect
nAChRs were not available, we performed screens against two subtypes of
vertebrate nAChRs (α3* and α7) based on the hypothesis that nAChR modulators
might also be useful for deterring vertebrate predators.
Several high-throughput FLIPR screenings were carried out to identify “hit”
venoms that were to contain promising lead compounds to further use as research
tools, or lead drugs. However, the data indicated that most of the spider venoms
contained predominantly blockers highly potent for subtype α3* nAChRs, and
blockers for α7 nAChRs; and a non-selective modulator was not considered to
isolate because adverse effects of some agents under current study are
associated with low selectivity for cholinergic receptors (Kalayci, 2013; Koga,
2014; Mihalak, 2006). Even when a toxin lead, which was expected not being
acylpolyamine type but selective for either α3* or α7 nAChRs, was not identified,

67
the resulting data was suitable to further analyse the distribution of modulators
across the phylogenetic tree of spiders.
Because spiders are venomous animals with more than 45,000 extant species
spread across 114 taxonomic families (see http://www.wsc.nmbe.ch/statistics/), it
was impossible in this study to sample broadly across this immense phylogenetic
tree, so we instead chose 71 species spread across the major suborder
Opisthothelae, which is comprised of the infraorders Araneomorphae ("modern"
spiders") and Mygalomorphae ("primitive spiders"). Note that Mesothelae, the
other taxonomic suborder with Araneae, comprises only ~100 extant species that
are thought to be non-venomous (for a review, see (Xu, 2015)).
With the caveat that it is difficult to make firm conclusions in the absence of deeper
taxonomic sampling across the phylogenetic tree, two trends emerge from the
data summary shown in Figure 2.8; and the discussion is based on the trends:
1) Although nAChR modulators can be found in distantly related species across
the phylogenetic tree, they were absent from 3 of the 6 venoms that we sampled
from the RTA clade as well as the single member of the family Austrochilidae that
we sampled. In addition, the 3 left venoms from the RTA clade had a partial
blocking effect on nAChRs. This indicates that modulation of vertebrate nAChRs
was either a basal pharmacology in spider venoms that was secondarily lost in
some species, or that it arose independently numerous times during Araneae
evolution. Deeper phylogenetic sampling in combination with biochemical
characterisation of the nAChR modulators found in spider venom will be required
to discriminate between these possibilities.
2) In this study, nAChR agonists, and specific blockers of the α3* subtype were
only found in the venom of theraphosid spiders, the largest family of mygalomorph
spiders with ~1000 species. Thus, mygalomorph venoms appear to be richer in
nAChR modulators than araneomorph venoms.
From the total of 71 spider venoms used in the present work, 12 belonged to
spiders of the suborder Araneomorphae. The resulting data indicated that

68
modulators for vertebrate nAChRs have evolved differently that were leading by
driven forces of natural selection such as geographical isolation and interactions
between preys and predators (Morgenstern, 2011; Zelanis, 2012). Probably, the
reason of absence of modulators in the venom of H. troglodytes is because it lives
in caves and underground drainages (Doran, 2001), and it is unlikely to encounter
vertebrate predators. Furthermore, they do not forage for insects instead they use
their long webs up to 1 metre of diameter to trap insects such as cave crickets.
The cheliceral structure of “modern” spiders evolved into an improved tool that can
be moved sideways, unlike fangs from “primitive” spiders that are moved up and
down 1. This feature in araneomorph spiders allows them grapping easily big preys
such as small vertebrates without using their venoms (Nyffeler and Pusey, 2014).
They attack their prey by crushing their fangs on the neck of preys that could be
small fish and lizards (Nyffeler and Pusey, 2014). In contrast, in mygalomorph
spiders, the position and movement of their cheliceraes do not allow
mygalomorphs to hunt preys bigger than big insects. Instead, they preferentially
use their venoms to immobilise them.
We could hypothesise that appearance of strong, and robust cheliceraes that are
moved sideways as forceps and the reduction of the length of “modern’ spiders
could lead to reduce the synthesis of modulators for vertebrate nAChRs in spider
venom. For instance, spider venoms with no potent blocking effect or no effect on
nAChRs belonged to spiders from the phylogenetic branch RTA clade, although in
the same phylogenetic clade, modulators for nAChRs could randomly appear on
other species. It can be evidenced when no substantial blocking effect on nAChRs
was observed in venoms from Ancylometes sp., V. sylvestriformis (Ctenidae), and
the lack of no modulators in venom from H. jugulans (Sparassidae), while venom
from Megadolomedes australianus contain potent blockers for nAChRs. The two
ctenid spiders studied in this research work have strong and long fangs that may
be used as defence. Moreover, they are found at night at the edge of water
sources in remote areas of the forest or the jungle (Nyffeler and Pusey, 2014)
1 Most information about the habitat, biology, and behaviour of spiders was obtained from web pages of The Australian Museum and http://www.arachne.org.au..

69
where they may have few contact to humans. Interestingly, M. australianus have
strong and large fangs and also potent blockers for vertebrate nAChRs. We could
speculate that toxins against nAChRs allowed that the spider could prey on fish,
which can be double of its size, tadpoles and insects (Nyffeler and Pusey, 2014).
An important physical adaptation in the spider is the presence of long legs that can
sense the vibrations of water when fish are nearby. Nevertheless, M. australianus
is relatively large because the length of its legs; and it may cause being seen
easily by natural enemies, and toxins against vertebrate nAChRs may be an
advantage.
Interestingly, our data showed that Araneomorph spiders from the araneoidea
family produced venoms with potent antagonists for vertebrate α3* and α7
nAChRs. For instance, two Australasian species of the genus Nephila, N. pilipes
and N. plumites showed to have in their venoms blockers for both α3* and α7
nAChRs. These compounds are likely to be acylpolyamines that have previously
been reported in other Nephila species such as Nephila madagascariencis (for a
review, see (Estrada, 2007)). Their preys are commonly insects, reviewed in
(Harvey, 2007) that are caught in their webs; because they produce a very strong
web between trees, on occasions small birds can get trap. Their natural enemies
are small vertebrates such as hummibirds (Vollrath, 1985). It seems modulators
against vertebrates could be evolved to protect themselves from them. Also,
humans could be considered as predators. Some native tribes from villages and
settlements from Thailand consider spiders of genus Nephila as food (for a review,
see (Harvey, 2007). Sometimes N. pilipes is found in house gardens and N.
plumipes can build their webs against buildings (for a review, see (Harvey, 2007))
and it makes these spiders more exposed to be grape from people; and it might
explain why venoms from N. pilipes and N. plumites contain potent blockers for
human nAChRs.
Mygalomorphae or “primitive” spiders
Because the suborder Mygalomorphae is considered an ancient monophyletic
group (Coddington and Levi, 1991), and its oldest fossil was dated to the Triassic

70
(Selden and Gall, 1992), this phylogenetic group can be particularly useful to
estimate the origin of modulators for vertebrate nAChRs.
From the 71 spider species used to initially searching for lead compounds, 59
venoms belonged to spiders of the suborder Mygalomorphae 2. The data showed
that most of the venoms blocked both subtype of mammalian nAChRs, and it is
likely the toxins were type acylpolyamines. Indeed the first acylpolyamine was
discovered in theraphosid spiders (for a review, see (Estrada, 2007)).
The only hexathelid species assayed herein was Macrothele gigas. Its venom
contained blockers for vertebrate α3* and α7 nAChRs. It also might contain an
agonist for α3* but an extra analysis of the active fraction may be carried out to
confirm the agonistic effect. It is likely blockers type acylpolyamines existed in the
venom of primitive spiders since the Triassic period because the ancestor of M.
gigas has been estimated to appear on that era. Interestingly, venoms from
Hexathelidae and Mygalomoprhae contain agonists and antagonists. This
similarity is not surprising because hexathelid and mygalomorph spiders are
closely related. It also suggests that a common ancestor could contain modulators
for vertebrate nAChRs in its venom. They are typically much larger than
araneomorphs, and they are consequently more prone to predation from
vertebrate predators such as birds, rodents and small reptiles.
On the other hand, a potential agonistic activity on nAChR was detected in venom
from Haplopelma hainanum. This spider lives in burrows that can reach up to 1
meter of depth. Its venom is categorized as highly venomous. The data herein
indicates this venom is likely to contain an agonistic component for nAChRs.
Probably, native acetylcholine may be present in the venom in large amounts
because there is a report of small molecules of a related species. For example, in
the venom of Haplopelma lividum was detected the polyamine spermine,
histamine, adenosine; and glutamic acid that was abundant in the venom fraction
of low molecular weight molecules (Moore, 2009). The plasticity of toxins to evolve
in accordance to environmental pressures, such as type of prey and type of
2 The data is summarised in Figure 2.8.

71
predators have been reported recently in the venom of Haplopelma hainanum
(Zhang, 2015b). It was shown that its intraspecific venom variations of blockers for
vertebrate nAChRs could be attributed to interaction gene-environment. In
addition, the presence of agonist for α7 nAChRs in venoms from Pamphobeteus
sp., Sericoplema rubronites and Xenesthis sp. remain to be confirmed. These
tarantulas are usually trade as pets; and if the presence of agonists for α7
nAChRs is due to the constant interaction with humans when these are in captivity
need to be evaluated. It would be interesting to further study venoms from these
tarantulas that naturally inhabit in the forest and the Amazon in South America.
According to the results of the present thesis, spider venoms from the family
theraphosidae, such as Selenocosmia crassipes, Selenocosmia effera,
Selenocosmia sp (Papua New Guinea), Coremiocnemis tropix, Selenotholus
foelschei contain potent toxins that block preferentially human neuronal α3*
nAChRs. In Australia, Selenocosmia sp. is known to cause severe pain in humans
and death in canines while venoms from Coremiocnemis tropix (spread out in
North Australia) and Selenotholus foelschei (inhabit only in Western Australia) are
shown to have a pre synaptic action at the neuromuscular junction in chicks
(Herzig, 2008). The analysis of our data indicates that these venoms may contain
compounds that preferentially act on the autonomic nervous system where the
“ganglionic” nAChR subtypes are expressed, and mediate the pain observed in
people bite by these spiders instead of targeting the nAChR muscle subtype.
On the other hand, theraphosid spiders from the new world studied herein showed
to contain blockers for vertebrate cholinergic receptors. Scarcely information is
available about venomic, diet and habitat of spiders from the new world, although,
some works have been carried out in some spiders assayed in this work (Bertani
and Fukushima, 2009).
In this study, the data showed that venom from Aphonopelma crinitum had some
blocking effect on α3* nAChRs but it was not significant. The presence of
acylpolyamines cannot be discarded because in a previous work two
acylpolyamines were discovered in the venom of Aphonopelma chalcodes

72
(Skinner, 1990). Aphonopelma crinitum, Pseudhapalopus sp. and Homoeomma
sp. may produce few blockers for α3* nAChRs with no effect on α7 nAChRs
because these are found in remote areas where there is little human activity . For
example, Aphonopelma crinitum lives in dry areas or desserts of North Mexico.
The plasticity of toxins to evolve in accordance to environmental pressures have
been reported recently in venoms of several species from Avicularia sp.,
Grammostola, sp., Holothele sp., Nhandu sp., and Psalmopoeus sp. (Zhang,
2015b). The diversity of antagonists for vertebrate nAChRs could be attributed to
interaction gene-environment. The Spiders Euathlus sp. and Thrixopelma sp.
inhabit along the western coast of Chile, and Thrixopelma cyaneolum inhabit the
coast along western Peru, all seemed to contain blockers for the subtype α3*
nAChRs. However, one venom was extracted from a Grammostola grossa species
that inhabits the Eastern Coast of Argentina. Its venom also had a blocking effect
on the subtype α3* nAChRs but the presence of modulators in its venom cannot
be attributed to the type of prey or predators that are found on the geographical
place where this species inhabits (the coast). G. grossa may be the same group of
spiders that originally inhabit the Amazons in Brazil that are spread around
adjacent regions of Brazil. At some point Grammostola species could evolved into
other specie that produce blockers for both subtypes of α3* and α7 nAChRs. This
is the case for Grammosotla rosea that is found in Bolivia, then, there could be a
spread of this species heading down to south America like the case of
Grammostola iheringi and Grammostola porteri that can be found in Chile and
Argentina. In general, few research are carried out about tarantula venoms, in
particular for those species found in Brazil, such as Acanthoscurria paulensis
(Mourão, 2013). Arboreal species Avicularia genus consist of 13 species and
these are found in Central and South America. Scarce studies about venoms from
Avicularia juruensis exist in the scientific literature (Ayroza, 2012). Avicularia
species are threatened for severe deforestation in the rainforest in Brazil and the
illegal trade to collectors (Bertani and Fukushima, 2009). It could explain why
several species of Avicularia has been spread across other regions around Brazil.
Interestingly, all species studied herein that are found in Brazil (in the forests of the
Amazonas) contain blockers for α3* nAChRs.

73
The two tarantulas Holothele sp. were collected originally from Colombia and
Peru. Their venoms contain blockers for both subtype nAChRs. Probably both
species are the same and these were spread around Colombia and northern Peru.
These tarantulas live in burrows, and blockers for cholinergic receptors could be
used for protection against small vertebrate predators. Like Holothele sp., the
theraphosid Nhandu vulpinus, Nhandu tripepii and Nhandu chromatus live in
burrows in the savannahs and tropical forests of South Brazil. Their venoms have
low toxicity and could be comparable to bee sting. Their diet is based on crickets,
mealworms and houseflies. It is likely the blocking effect on nAChRs is for
protection against small predators.
It is known from previous studies that nicotinic receptors are densely expressed in
insects and these share similar pharmacological profiles to counterparts
expressed in vertebrates (Sattelle, 2005). It suggests that the effect observed in
human nicotinic receptors caused by some spider venoms from the two major
lineages Mygalomorphae and Araneomorphae could share some similitude to
neuronal nicotinic cholinergic receptors from invertebrates, which known to be
expressed in neurons and cell bodies in insects. It is likely that selective toxins for
invertebrate nAChRs may be found because spiders based most of their diet on
insects.
The mechanisms of toxin evolution and diversity in spiders are currently not well
understood (Zhang, 2015b), although, this thesis provides with important insights
of the evolutionary role of toxins against mammalian cholinergic receptors. The
number of different spider venoms used on the present thesis is small to make
generalised conclusions. However, the results so far indicated that blockers for
vertebrate cholinergic receptors were likely to be present in old ancestor of
mygalomorph spiders. Inhibitors for human neuronal α3* and α7 nAChRs are the
major compounds in “primitive” spiders. These are likely to be acylpolyamines that
have previously been reported in “primitive” spiders (Skinner, 1990; Zhang,
2015b); and it implies that acylpolyamines have existed about 400 million years
ago, and are the simplest toxin to synthesise with high potency, unlike synthesis of
peptides or proteins. Furthermore, the data suggests that modulators for

74
vertebrate nAChRs were decreased notoriously when araneomorph spiders
appeared.
Our data also implies that primitive spiders produce a wide range of modulators for
vertebrate nAChRs that could include agonistic modulators on α7. It seems the
large size of mygalomorph spiders and their cheliceral position to subdue their
preys was no advantageous with the appearance of new vertebrate animals. It is
likely that in “primitive” spiders the role of toxins for both subtypes of mammalian
nAChRs is for defence. We could speculate also that in “modern” spiders
synthesis of agonists for α7 is energetically more expensive than no selective
antagonists for α3* and α7 nAChRs. After all, no selective blockers may lead to
the same outcome that warns vertebrate predators away.
To summarize, we can suggest that “modern” spiders (Araneopmorphae)
appeared with new physical characteristics to adapt them to new ecosystems; and
in parallel their venoms evolved with the appearance of different vertebrate
species, including small mammals. Araneomorph spiders retain the presence of
cheliceraes but it seems these were adapted to pinch the preys and avoid they
scape, instead of rely on their venoms to immobilise their preys, like in the case of
Mygalomorph spiders.
The resulting data of this thesis is a started point to further selection of venoms
with potential effect on cholinergic receptors of invertebrates. Extra screenings
using the FLIPR system, and including as positive control neonicotinoids may help
to choose “hit” venoms.
This is the first study of distribution of toxins from spider venoms against human
neuronal nAChRs, and it is hoped that it can serve to the discovery of novel toxins
against invertebrates as well as help to reveal the evolutional process of spider
toxins.

75
CHAPTER 3 — NACHR MODULATORS FROM SCORPION VENOM 3.1 INTRODUCTION
Like spiders, scorpion venoms contain a vast number of uncharacterized
compounds, and therefore they represent an untapped source of novel toxins with
potential applications in medicine, agronomy and as research tools (Harrison,
2014; King, 2014; King and Coaker, 2014; Wu, 2014). It has been estimated that
the molecules currently described from scorpion venoms represent less than 1%
of all components predicted to be in their venoms (Quintero-Hernandez, 2013).
Thus, we extended the search for modulators of vertebrate α7 and α3* nAChRs to
five scorpion venoms. As outlined below, this work led to the isolation of a non-
selective agonist of α7 and α3* nAChRs, from the venom of the giant forest
scorpion Heterometrus spinnifer.
3.1.1 Scorpion toxins and their pharmacological action Scorpions (Arthropoda: Chelicerata: Arachnida: Scorpiones) use their venoms to
immobilise prey and/or to defend against predators or competitors. The common
biological function of these venoms is their action on a variety of voltage-gated ion
channels in invertebrates and small vertebrates. In particular, scorpion venoms are
rich in modulators of both insect and vertebrate voltage-gated sodium (for a
review, see (Egea, 2015)) and Potassium (Kv) (Chugunov, 2013; Morales-Lázaro,
2015; Quintero-Hernandez, 2013; Swartz, 2013). The most toxic scorpion venoms
for mammals are from Androctonus sp. and Leirurus quinquestriatus (paleotropical
especies) and Centruroides sp. and Tityus sp. (neotropical especies), and these
venoms have been extensively studied due to their clinical relevance (DeBin,
1993; Diaz, 2009; Kularatne and Senanayake, 2014; Quintero-Hernandez, 2013;
Rjeibi, 2011). Most of the components purified and identified from scorpions to
date are toxins that act on Nav (Forsyth, 2012; Meves, 1984; Zhu, 2012), Kv
(Camargos, 2011; Nirthanan, 2005; Xie, 2012), Cav (Norton and McDonough,
2008; Shahbazzadeh, 2007) and chloride channels (DeBin, 1993; Nie, 2012;
Rjeibi, 2011; Zheng, 2000). In particular, toxins that target Nav channels that are
largely responsible for the symptomatology of mammalian envenomation

76
(Caliskan, 2012; Clot-Faybesse, 2000; Quintero-Hernandez, 2013) and paralysis
in insects (Billen, 2008; DeBin, 1993; Forsyth, 2012). However, the effect of
scorpion venom on nAChRs has been scarcely documented.
Androctonin, a disulphide-bridged peptide isolated from the hemolymph, but not
the venom of Androctonus australis, shows some similarity to a nAChR named α-
conotoxin SII, but its effect on nAChRs has not been determined (Ehret-Sabatier,
1996).
A non-toxic venom extract from the scorpion Buthus occitanus tunetanus contains
a component that depolarises muscle cells via an effect on nAChRs (Cheikh,
2007), but the authors did not isolate the active component.
Venoms from scorpions reported as non-threatening to humans have not been
extensively studied. Interestingly, venom from the giant forest scorpion
Heterometrus spinnifer was reported to contain high concentration of acetylcholine
and norepinephrine (Nirthanan, 2002). The authors suggested that acetylcholine
induces muscle contraction in rat nerve-smooth muscle preparations through
activation of muscarinic receptors. However, all of the bioassays were carried out
with a venom fraction that contained all molecules from the venom with molecular
masses less than 3 kDa, and the activity was not compared with pure
acetylcholine.
From our previous screening of 71 arachnid venoms for searching modulators for
nAChR using a high-throughput FLIPR calcium assay, we detected an agonistic
activity elicited from crude venom of H. spinnifer. We continued further to isolate
the “hit” toxin responsible for the activity because venom compounds seem to
have promising applications in therapy (Nie, 2012; Yang, 2011) and likely in
agronomy (Morales-Lázaro, 2015 #7), as mentioned before the venom is devoid of
harm in humans (Kularatne and Senanayake, 2014) and has not been extensively
studied due to the lack of clinical interest (Cheikh, 2007; Nirthanan, 2002).
The agonistic compound was isolated, and identified as a choline ester, named
senecioylcholine. It activates human neuronal nAChRs endogenously expressed

77
in SH-SY5Y cells and the synthetic form did not show insecticidal effect. It is
known ester of choline occurs naturally (Erspamer and Glässer, 1957; Erspamer,
1953; Florey, 1963; Keyl, 1957), and mainly purified from hypobranchial gland of
gastropods (Keyl and Whittaker, 1958; Roseghini, 1996). Even when acetylcholine
is a choline ester present in invertebrate nervous system (Florey, 1963),
senecioylcholine has not been before identified in scorpion venoms. Few
pharmacological studies have been done for senecioylcholine and its isomer
named tygloylcholine (Roseghini, 1996). Furthermore, there are discrepancies on
the function of choline ester in the venom of gastropods as mechanism of defence
(Erspamer and Glässer, 1957; Roseghini, 1996) and in the correct identification of
senecioylcholine or tygloylcholine (Baker and Duke, 1976, 1978).
The toxicity of senecioylcholine reported in vertebrates (Roseghini, 1996) and its
non-selective agonistic effect on human neuronal nAChRs suggest its role in the
venom is highly for defence against small vertebrates. The discovery of
senecioylcholine in the venom of a scorpion, which uses its chelae to subdue its
preys, makes a significant contribution in the evolution of venom compounds as
defensive weapons, and it is the first evidence of a convergent evolution between
scorpions and molluscs. To provide with a complete base for future research
works in interactions prey-predator and evolution, this study was extended up to
the pharmacological characterization of senecioylcholine and its isomers
tygloylcholine and angeloylcholine, which could be present in other arachnid
venoms.
3.2 METHODS
We used the high-throughput FLIPR assay outlined in Chapter 2 to screen five
scorpion venoms for modulators of vertebrate α7 and α3* nAChRs. For details of
the procedures used to identify “hit” venoms and purify bioactive venom
components see the Section 2.5 and 3.3.2, respectively.
3.2.1 Figure 3.1 shows an example of the protocol as applied to scorpion venoms

78
Figure 3.1A, shows the fluorescent response obtained by stimulation of α3*
nAChRs with nicotine, and inhibition of this response by d-tubocurarine. Figure
3.1B shows that the calcium fluorescence response is evoked mainly by activation
of α3* nAChRs, which was inhibited by d-tubocurarine, and this nicotine response
is independent of activation of Nav channels. On Figure 3.1C is shown the
fluorescent response evoked by crude venom from A. crassicauda, which induces
a robust increase in fluorescence prior to the addition of nicotine. The venom-
induced increase in fluorescence was not substantially inhibited by d-tubocurarine
(Figure 3.1D) but it was notably by TTX (Figure 3.1 E), a non-selective Nav
channel inhibitor (Figure 3.1E), Thus, the robust increase in fluorescence induced
by A. crassicauda venom is likely due to the presence of an agonist of Nav
channels endogenously expressed in SH-SY5Y cells.

79
Figure 3.1. A. crassicauda venom can induce calcium responses elicited via activation of Nav channels. (A) Fluorescent responses elicited by stimulation of α3* nAChRs with nicotine, and inhibition of this response by d- tubocurarine. (B) The calcium fluorescent response observed is mediated mostly by activation of α3* nAChRs with nicotine (30 µM). The inhibitor for Nav channels, TTX, does not inhibit the calcium response. (C) Addition of crude venom (2 µg) from A. crassicauda elicited fluorescent response (black trace), prior to the activation of α3* nAChRs with nicotine (blue trace). The venom-induced increase in fluorescence was not substantially inhibited by d-tubocurarine (panel D) but it was significantly inhibited by 2 µM TTX (panel E).
0
1
2
3
4
1 151 301 451 601time (seconds)
A. crassicauda venom 2 µg
TTX 2 µM + A. crassicauda venom 2 µg
E)
Max
imun
cal
cium
resp
onse
(RLU
)
0
1
2
3
1 151 301 451 601
Max
imun
cal
cium
resp
onse
(R
LU)
time (seconds)
nicotine 30 µM d-tubocurarine 30 µM
A)
0
1
2
3
1 151 301 451 601
Max
imun
cal
cium
resp
onse
(R
LU)
time (seconds)
nicotine 30 µM TTX (2 µM)
B)
0
1
2
3
4
1 151 301 451 601
Max
imun
cal
cium
resp
onse
(R
LU)
time (seconds)
A. crassicauda venom 2 µg nicotine 30 µM
C)
0
1
2
3
4
1 151 301 451 601M
axim
un c
alci
um re
spon
se
(RLU
)
time (seconds)
A. crassicauda venom 2 µg d-tubocurarine 30 µM + A. crassicauda venom 2 µg
D)

80
3.2.2 Scorpion venoms
Dr Volker Herzig kindly provided five scorpion venoms for screening against
vertebrate α7 and α3* nAChRs. The list of scorpion species used in this study is
given in Appendix I.
3.2.3 Venom fractionation
Crude scorpion venom (1 mg) was dissolved in 1 ml of 5% of buffer B (90%
acetonitrile containing 0.05% TFA) and centrifuged at 5,000 g for 15 min at 4°C.
After centrifugation, the supernatant was loaded onto a Phenomenex C18 reverse-
phase HPLC (RP-HPLC) column Jupiter 250 x 4.60 mm; 5 µm particle size; 300 Å
pore size and fractionation performed using Shimadzu HPLC system. Venom
fractionation was performed using a flow rate of 1 ml/min and the following
gradient of Buffer B in Buffer A (Milli-Q water + 0.05% TFA): 0–5 min: 5% Buffer B;
5–10 min: 5–20% Buffer B; 10–50 min: 20–40% Buffer B; 50–55 min: 40–80%
Buffer B; 55–57 min: 80% Buffer B.
The elution profile was recorded at 214 nm and 280 nm, and fractions were
collected manually based on 214 nm absorbance. At the end of the fractionation
the samples were frozen, lyophilized, dissolved in Milli-Q water, and stored at –
20°C until required.
3.2.4 Discovery of a choline ester from the venom of H. spinnifer
We isolated the active fraction from H. spinnifer venom using two chromatographic
steps of purification using the FLIPR high-throughput calcium assay described in
the previous section. The active fraction from the initial venom fractionation using
a Jupiter C18 RP-HPLC column was further fractionated on a Grace Discovery
Science Vydac® 218TP C18 RP-HPLC column 250 x 4.60 mm; 5 µm particle size;
and 300 Å pore size using a flow rate of 1 ml/min and the following gradient: 0–5
min: 5% Buffer B; 5–10 min: 5–10% Buffer B; 10–50 min: 10–30% Buffer B; 50–55
min: 30–80% Buffer B; 55–57 min: 80% Buffer B.

81
3.2.5 Determination of mass and purity
The active component isolated from H. spinnifer venom was dissolved in 5% of
acetonitrile (grade HPLC) and analysed via matrix-assisted laser
desorption/ionization (MALDI) mass spectrometry (MS) performed on an Applied
Biosystems mass spectrometer operating in the positive ion linear mode. The
sample (0.4 µL) was mixed on a MALDI plate with 0.4 µl of a-cyano-4-
hydroxycinnamic acid (CHCA) matrix (6 mg/mL) dissolved in acetonitrile/ MilliQ
water /formic Acid (50:45:5, v/v).
The MALDI-TOF spectra did not reveal any masses higher than 1 kDa. Thus, the
active fraction was further analysed via LCMS using an Agilent 1100 Series
separations module equipped with diode array detector and Agilent 1100 Series
LC/MSD mass detector in both positive and negative ion modes. The sample was
run on an Agilent Zorbax-C8 column 4.6 mm x 150 mm, 5 µm particle size using a
gradient of 10–100% methanol/H2O (with constant 0.05% formic acid modifier)
over 15 min, using a flow rate of 1 mL/min.
3.2.6 Identification of active compound using HRMS and LCMS
High-resolution (HR) electrospray ionisation (ESI) MS measurements were
obtained on a Bruker micrOTOF with an ESI probe by direct infusion in acetonitrile
at 3 µL/min using sodium formate clusters as an internal calibrant. The active
venom yielded m/z 186.1504 [M]+ (calculated for C10H20NO2, 186.1489).
LC/MS was performed on native compound co-injected with synthetic choline
esters using an Agilent 1100 series LC/MS with an Agilent Zorbax-C18 aqua
column 4.6 x 150 mm, 5 µm particle size. Isocratic elution was performed using
4% methanol/H2O (with constant 0.02% formic acid modifier) at a flow rate of 1
mL/min, with single ion extraction at 186 [M+].

82
3.2.7 Chemical synthesis of choline esters 3.2.7.1 Synthesis of chlorocholine chloride
Choline chloride (279 mg, 2 mmol) was dried under high vacuum at 200°C for 2 h.
SOCl2 (2 mL) was added and the solution was heated under reflux for 1 h. After
cooling, solvent was evaporated to dryness. Solid residue was redissolved in
methanol (3 x 2 mL), and re-evaporated to dryness. The resultant white solid was
dried under high vacuum at 80°C for 2 h.
3.2.7.2 Synthesis of tigloylcholine
To a solution of tiglic acid (200 mg, 2 mmol) in a dioxane–water (3:2) mixture (10
mL), a solution of K2CO3 (138 mg, 1 mmol in water 10 mL) was added. After
stirring for 10 min, the mixture was evaporated to dryness and again dried under
high vacuum at 70 °C for 1 h. Chlorocholine chloride and anhydrous DMSO were
added and the mixture heated at 60 °C for 48 h. DMSO was removed by vacuum
distillation and the residue was purified by reverse phase column chromatography
to afford tigloylcholine as a light brown hygroscopic solid. 1H NMR (600 MHz,
CD3OD): δ 6.92 (qq, 7.1, 1.3 Hz, H-3), 4.59 (m, H2-2ʹ′), 3.75 (m, H2-1ʹ′), 3.22 (s,
NMe3), 1.85 (dd, 1.3, 1.1 Hz, H3-5), 1.82 (dq, J = 7.1, 1.1 Hz, H3-4). 13C NMR (150
MHz, CD3OD): δ 168.4 (C-1), 140.3 (C-3), 129.1 (C-2), 66.4 (C-2ʹ′), 59.1 (C-1ʹ′),
54.60 (NMe), 54.58 (NMe), 54.55 (NMe), 14.6 (C-4), 12.3 (C-5). HRMS calc. for
C10H20NO2+ 186.1489, found m/z = 186.1491.
ClNMe3
ClHONMe3
Cl
SOCl2
70 o C, 1h
ONMe3OH
O Oi) K2CO3, dioxane/H2O, rt
DMSO, 60 oC, 48 hCl
NMe3 Clii)
1'
2'
Cl
12
3
4
5

83
3.2.7.3 Synthesis of senecioylcholine
As described for the synthesis of tigloylcholine, starting with choline chloride (2
mmol, 279 mg) and 2-methyl butenoic acid (2 mmol, 200 mg), senecioylcholine
was synthesized and purified by reverse phase column chromatography, to afford
senecioylcholine as a white hygroscopic solid. 1H NMR (600 MHz, CD3OD): δ 5.76
(qq, 1.2, 1.2 Hz, H-2), 4.54 (m, H2-2ʹ′), 3.73 (m, H2-1ʹ′), 3.22 (s, NMe3), 2.18 (d, 1.2
Hz, H3-4), 1.93 (d, 1.2 Hz, H3-5), 13C NMR (150 MHz, CD3OD): δ 166.8 (C-1),
161.2 (C-3), 115.8 (C-2), 66.4 (C-2ʹ′), 58.1 (C-1ʹ′), 54.66 (NMe), 54.63 (NMe), 54.61
(NMe), 27.6 (C-4), 20.6 (C-5). HRMS calc. for C10H20NO2+ 186.1489, found m/z =
186.1485.
3.2.7.4 Synthesis of angeloylcholine
As described for the synthesis of tigloylcholine, starting with choline chloride (2
mmol, 279 mg) and angelic acid (2 mmol, 200 mg), angeloylcholine was
synthesized and purified by reverse phase column chromatography, to afford
angeloylcholine as a white hygroscopic solid. 1H NMR (600 MHz, δ 6.23 (qq, 7.2,
1.3 Hz, H-3), 4.60 (m, H2-2ʹ′), 3.75 (m, H2-1ʹ′), 3.22 (s, NMe3), 2.00 (dq, 7.2, 1.3 Hz,
H3-4), 1.91 (dq, J = 1.3, 1.3 Hz, H3-5). 13C NMR (150 MHz, CD3OD): δ 167.9 (C-1),
141.7 (C-3), 128.0 (C-2), 66.4 (C-2ʹ′), 58.6 (C-1ʹ′), 54.59 (NMe), 54.56 (NMe), 54.54
(NMe), 20.8 (C-4), 16.2 (C-5). HRMS calc. for C10H20NO2+ 186.1489, found m/z =
186.1483.
ONMe3OH
O Oi) K2CO3, dioxane/H2O, rt
DMSO, 60 oC, 48 hCl
NMe3 Clii)
1'
2'1
23
4
5
Cl
ONMe3OH
O Oi) K2CO3, dioxane/H2O, rt
DMSO, 60 oC, 48 hCl
NMe3 Clii)
Cl
1'
2'1
23
4
5

84
3.3 RESULTS
3.3.1 High-throughput FLIPR assay
To speed the venom-based discovery of “hit” biomolecules from five scorpion
venoms a high-throughput FLIPR assay was performed (King, 2014; Vetter, 2011).
Five venoms derived from scorpions of the family Buthidae were screened using
the high-throughput FLIPR assay described in Chapter 2. At 2 µg, none of the
venoms had a significant effect on vertebrate α7 or α3* nAChRs (Figure 3.2).
However, crude venom (1 or 2 µg) from H. spinnifer elicited a transient calcium
response that was potentiated by 50% in the presence of the positive allosteric
modulator PNU-120596 (Figure 3.3A). This response was completely blocked by
d-tubocurarine (Figure 3.3B) indicating that the venom activates neuronal
nAChRs. The total elimination of the calcium response in the presence of PNU-
120596 by d-tubocurarine suggests that the agonistic effect observed is mediated
totally by α3* and α7 nAChRs. Two other concentrations of venoms were tested at
0.02 µg or at 0.2 µg (data not shown), and no effect was observed. The agonistic
effect of this venom is therefore dose-dependent.

85
Figure 3.2. FLIPR screening of scorpion venoms against α7 and α3* nAChRs. 1 µg of crude venoms from (A) L. quinquestriatus, (B) A. australis, (C) A. crassicauda, (D) G. grandidieri and (E) H. spinnifer was added to loaded- SH-SY5Y cells in the presence of PNU-120596. Calcium responses were then measured for 300 s, followed by choline addition, and recording of calcium response for a further another 300 s. To show the participation of cholinergic receptors in the calcium responses following venom addition, the cells were treated by d-tubocurarine before starting the assay. There was no effect of d-tubocurarine in the responses caused by venom from (A) L. quinquestriatus (trace in dashed lines and orange colour), (B) A. australis (trace in dashed lines and dark blue colour), and (C) A. crassicauda (trace in red colour). However, d-tubocurarine caused significant inhibition of the calcium response elicited by venoms from (D) G. grandidieri (trace in dashed lines and black colour) and (E) H. spinnifer (trace in blue colour). Traces are the average of at least three wells.
0
1
2
3
4
1 151 301 451 601
Max
imun
cal
cium
resp
onse
(RLU
)
time (seconds)
crude venom G.grandidieri 1 µg + PNU-120596 (10 µM)
crude venom G. grandidieri 1 µg + PNU-120596 (10 µM) + d-tubocurarine (30 µM)
0
1
2
3
1 151 301 451 601
Calc
ium
resp
onse
(RLU
)
time (seconds)
venom H. spinnifer 1 µg + PNU d-tubocurarine 30 µM
0
1
2
3
4
1 151 301 451 601Max
imun
calci
um re
spon
se (R
LU)
time (seconds)
A. crassicauda crude venom 2 µg d-tubocurarine 30 µM+ venom A. crassicauda 2 µg
C)
0
1
2
3
1 151 301 451 601M
axim
un
cal
ciu
m r
esp
on
se (
RLU
)
time (seconds)
crude venom A. australis 1 µg + PNU-120596 (10 µM) crude venom A. australis 1 µg + PNU-120596 (10 µM) + d-tubocurarine (30 µM)
0
1
2
3
1 151 301 451 601Ma
xim
un
ca
lciu
m r
esp
on
se
(RLU
)
time (seconds)
crude venom L. quinquestriatus 1 µg + PNU-120596 (10 µM) crude venom L. quinquestriatus 1 µg + PNU-120596 (10 µM) + d-tubocurarine (30 µM)
A)#
D)#
B)# C)#
E)#

86
Figure 3.3. Non-selective activation of α7 and α3* nAChRs expressed in SH-SY5Y by crude venom from H. spinnifer. (A) 2 µg of crude venom from H. spinnifer induced an increase of intracellular calcium in SH-SY5Y cells (blue trace) that was potentiated by PNU-120596 (red trace). (B) The calcium response elicited by 1 µg of crude venom from H. spinnifer in the presence of PNU-120596 (red trace) was totally blocked by d-tubocurarine (black trace in black colour). Traces are the average of at least three wells.
0
1
2
3
4
5
6
1 51 101 151 201 251 Calcium re
spon
se (R
LU)
Tme (seconds)
2 µg venom
2 µg venom + PNU
A)
0
1
2
3
1 51 101 151 201 251 301
Calcium re
spon
se (R
LU)
Tme (seconds)
1 µg venom + PNU120596
1 µg venom + PNU120596 + d-‐tubocurarine
B)

87
3.3.2 Isolation of the active compound from H. spinnifer venom
1 mg of crude venom from H. spinnifer was separated by RP-HPLC (Figure 3.4)
and fractions were manually collected. Each resulting fraction at 0.42 µg of total
crude venom was evaluated using the FLIPR assay to evaluate its impact on
activation of nAChRs using nicotine or choline plus the positive allosteric
modulator PNU-120596 to stimulate the heteromeric subtypes α3β2/α3β4 and the
homomeric subtype α7 nAChR, respectively. Only fraction 10, which eluted at
9.49% acetonitrile in the C18 RP-HPLC separation (inset, Figure 3.4), activated
the α3* subtype as its activity was substantially inhibited (66%) by d-tubocurarine
(30 µM) (Figure 3.5). Furthermore, the increased response in the presence of
PNU-120596 (Figure 3.5) indicates that α7 nAChRs are also activated by fraction
10. Thus, fraction 10 contains the agonistic molecule that is responsible for the
calcium response elicited by crude H. spinnifer venom.
Figure 3.4. HPLC fractionation of crude venom from H. spinnifer. Chromatogram resulting from fractionation of crude venom via C18 RP-HPLC. The fraction highlighted in black (F10) activated α7 and α3* nAChRs in the FLIPR assay. The inset shows an expansion of the region containing F10, which eluted at ∼ 13.6 min.
20 40 60
0
1
2
3
Time (min)
Abs
orba
nce
214
nm
12 14 16
0
1
2
3
Time (min)
Abs
orba
nce
214
nm
Fraction 10

88
Figure 3.5. Activity of F10 from fractionation of crude venom from H. spinnifer. Active fraction 10 (F10) (1% from initial fractionation of 1 mg of crude venom) was assayed against vertebrate nAChRs using the FLIPR. F10 induced a maximum fluorescent response similar to that evoked by nicotine. The response was inhibited by d-tubocurarine and potentiated by PNU-120596. The difference are significant with a P < 0.05. One-way ANOVA was used to compare the means of each experimental treatment in relation to the control response (nicotine response). The values are averages of three independent experiments with n = 3. Dotted lines represent the control response in the absence of venoms. Fraction 10 (72 µg) was further fractionated using an analytical RP-HPLC
(GRACE-Vydac C18 column RP-HPLC) (Figure 3.6) and the resulting fractions
collected manually. Each of the active fractions were then analysed using the
FLIPR assay, for activity against vertebrate nAChRs. Only two fractions were
active in the FLIPR assay, which eluted at 11.1 min (fraction 4) and 13.6 min
(fraction 7) (Figure 3.6). Unfortunately, further study of fraction 7 was not possible
due to its low abundance as evidenced by its very low absorbance at 214 nm.
Thus, further studies focused on fraction 4. Assay of fraction 4 (77.35 µM) in the
FLIPR assay revealed that it induced a fluorescent response that was completely
inhibited by 30 µM d-tubocurarine and potentiated by 10 µM PNU-120596 (Figure
3.7). We therefore focussed attention on identification of the active component in
this fraction.
F10 (4
.2 µg)
F10 (4
.2 µg)
d-tuboc
urarin
e (30
µM)
F10 (4
.2 µg)
PNU1205
96 (1
0 µM
)0
100
200
300%
FM
ax
****
****
****

89
Figure 3.6. Purification of the “hit’ toxin by RP-HPLC. Chromatogram resulting from separation of fraction 10 (72 µg) on an analytical C18 RP-HPLC. The “hit” toxin (fraction 4) eluted at 11.1 min (10.35% Buffer B), and fraction 7 eluted at 13.6 min (12.9% Buffer B).
Figure 3.7. Agonistic activity of the “hit” toxin. Fraction 4 (77 µM) from analytical HPLC induced a fluorescent response that was totally blocked by d-tubocurarine and by PNU-120596. This suggests that fraction 4 contains the “hit” toxin that is primarily responsible for the non-selective agonism of α3* and α7 nAChRs by H. spinnifer venom. The difference are significant with a P < 0.05. One-way ANOVA was used to compare the means of each experimental treatment in relation to the control response (nicotine response). The values are averages of three independent experiments with n = 3. Dotted lines represent the control response in the absence of venoms.
d-tuboc
urarin
e (30
µM)
Native
senec
ioylch
oline (
77 µM
)
Native
senec
ioylch
oline (
77 µM
)
d-tuboc
urarin
e (30
µM)
Native
senec
ioylch
oline (
77 µM
)
PNU1205
96 (1
0 µM
)
0
50
100
150
200
250
% F
Max
****
****
****
****
0 20 40
0
100
200
300
Time (min)
Abs
orba
nce
(214
nm
)
Frac
tion
4
11.1
Frac
tion
7

90
3.3.3 Identification of the active compound by high-resolution mass spectrometry
Analysis of fraction 4 using MALDI-TOF-MS revealed no masses larger than 1
kDa; it is difficult to detect masses smaller than this via MALDI-TOF-MS due to the
noise caused by the matrix (CHCA). Thus, the sample was further analysed using
HR-MS; this revealed a single compound with m/z 186.1489. Database searching
revealed that the mass formula of C10H20N1O2 corresponds to a previously
described choline ester named senecioylcholine (Keyl, 1957; Roseghini, 1996;
Whittaker, 1960). However, there are two isomers of senecioylcholine,
tigloylcholine and angeloylcholine, with identical mass formula and it is impossible
to determine, which of these isomers corresponds to the native venom-derived
compound based on HR-MS data alone. Thus, all three isomers were chemical
synthesized and LC-MS co-elution studies were performed to identify the native
compound. As shown in Figure 3.8, senecioylcholine but neither of the two
isomers, co-elutes with the active nAChR agonist purified from H. spinnifer venom.
Thus, we conclude that the α3*/α7 nAChR agonist in H. spinnifer venom is
senecioylcholine.

91
Figure 3.8. Co-elution of bioactive compound from H. spinnifer venom and synthetic choline esters. LC-MS chromatograms of synthetic choline esters (top three panels), the α3*/α7 nAChR agonist from H. spinnifer venom (bottom panel). LC was performed using a Zorbax C18-aqua column (4.6 × 150 mm), 5 µm particle size with a gradient of methanol + 0.02% formic acid at a flow rate of 1 ml/min.
0 5000
10000 15000
0 2500 5000 7500 10000 12500
0
2000 4000 6000 8000
10000
1000 2000 3000 4000 5000
1 2 3 4 5 6 7 8 9 0
5000 10000 15000
0
synthetic angeloylcholine
synthetic tigloylcholine
synthetic senecioylcholine
native senecioylcholine
co-injection of native + synthetic senecioylcholine
Retention time (min)
tota
l ion
cou
nt

92
3.4 DISCUSSION
H. spinnifer is a scorpion from the Scorpionidae family. It is widely distributed in
South-eastern Asia but its venom has been poorly studied due to its innocuous
effect on humans. However, it is still a rich source of mostly uncharacterised
compounds that could be useful as novel modulators of nAChRs with clinical or
agronomic relevance. In this study, we used activity-guided fractionation to isolate
a novel agonist of vertebrate nAChRs from the venom of H. spinnifer. By using
HR-MS in combination with chemical synthesis, we identified this agonist as
senecioylcholine, a choline ester. We found that senecioylcholine is a non-
selective agonist of both α3β2/α3β4 and α7 nAChR subtypes endogenously
expressed in SH-SY5Y cells.
Choline esters are secreted by the hypobranchial gland of predatory marine
gastropods from the family Muricidae (phylum Mollusca), for a review, see
(Benkendorff, 2013) where they are postulated to play a role in predation and/or
defence against predators. They have also been detected in the ventral nerve cord
of the American lobster Homarus americanus (phylum Arthropoda, class
Crustacea) (Keyl, 1957). The main choline ester identified in Mollusks is murexine
(Erspamer and Glässer, 1957) although senecioylcholine has also been detected
in significant quantities in the hypobranchial gland of Dicathais orbita. A more
recent study identified tigloylcholine, an isomer of senecioylcholine, muricids of the
genus Thais (Shiomi, 1998). Murexine and tigloylcholine are both lethal to mice,
with intravenous administration leading to LD50 values of 8.5 mg/kg and 0.92
mg/kg (Shiomi, 1998), respectively, for a review, see (Benkendorff, 2013). Like
murexine, senecioylcholine is also a neuromuscular blocking compound with
nicotine action on vertebrates (Roseghini, 1996). Interestingly, however, even high
doses of senecioylcholine have no effect on the fruit fly Drosophila melanogaster
(data not shown). Thus, it is likely that senecioylcholine is used by H. spinnifer for
defence against vertebrate predators.
A few bioactive compounds have previously been isolated from H. spinnifer venom
k-KTx1.3, a non-toxic peptidic modulator of Kv channels (Nirthanan, 2005), and six
novel antimicrobial peptides (Nie, 2012; Wu, 2014). H. spinnifer venom was also

93
shown to contain high concentrations of acetylcholine (∼80 µM) and this was
proposed to be responsible for the reversible venom-evoked contracture of chick
biventer cervicis muscle that was blocked by d-tubocurarine (Nirthanan, 2002).
However, all the bioassays were carried out using a venom fraction that contained
all molecules with molecular masses less than 3 kDa. Thus, it is likely that
senecioylcholine, which was identified in H. spinnifer venom in the current study,
mighty be at least partially responsible for the observed contracture of chick
biventer cervicis muscle. The discovery of senecioylcholine in both scorpion
venom (phylum Arthropoda) and predatory marine gastropods (phylum Mollusca)
suggests that these phylogenetically unrelated organisms have convergently
evolved the use of choline esters to defend themselves against vertebrate
predators. It remains to be seen whether choline esters are ubiquitous in scorpion
venoms or restricted to specific families or genera. However, the fact that only 2 of
the scorpion venoms tested in this study appear to contain agonists of human
α3β2/α3β4 and α7 nAChRs suggests that senecioylcholine and related choline
esters are not widely distributed in scorpion venoms.

94
CHAPTER 4 — CONCLUSIONS Venomous animals use their venom to hunt preys and/or protect themselves
against predators or competitors (Olsen, 2011). Arachnid venoms are complex
mixtures of salts, small organic molecules, peptides and proteins that target a wide
range of neuronal receptors and ion channels (Billen, 2008; Estrada, 2007;
Morales-Lázaro, 2015; Quintero-Hernandez, 2013). A large number of modulators
of voltage-gated ion channels have been isolated from the venom of spiders and
scorpions, but there are very few reports of arachnid venom components
modulating the activity of nAChRs. This is surprising given that neonicotinoid
insecticides are one of the most successful classes of chemical insecticides, which
suggests that targeting of insect nAChRs would be a useful pharmacology for
arachnid venoms. Thus, the present work was carried out to survey the prevalence
of nAChR modulators in arachnid venoms.
Until recently, most characterizations of arachnid venoms used traditional
biological assays that require large amounts of venom or venom fraction, which
hampered studies of small venomous animals such as arachnids. In the current
study we used a high-throughput FLIPR assay that enabled screens to be rapidly
performed against human α3* and α7 nAChRs using only a few micrograms of
venom. This approach allowed simultaneous screening of 76 arachnid venoms at
four different concentrations.
The data were reproducible even when different pools of venoms were used. This
suggests that nAChR modulators in arachnid venoms are maintained even when
they are kept in captivity. Our data revealed that spider venoms are a rich source
of nAChR modulators, and that at least some scorpion venoms contain choline
esters that agonize vertebrate nAChRs.
We propose that these nAChR modulators are primarily used for defence against
vertebrate predators. It would be of great interest in future to perform high-
throughput screens of arachnid venoms against insect nAChRs. Predation of
insects is the primary role of arachnid venoms and therefore we might expect to
find an even greater abundance and variety of compounds in these venoms that

95
modulate the activity of invertebrate nAChRs. Nevertheless, the current results
strongly support the view that arachnid venoms are a novel source of modulators
of ligand-gated ion channels with potential biotechnological applications.
SUMMARY AND FUTURE DIRECTIONS
The current work revealed, for the first time, that many spider venoms (65 of 71
tested in this study) contain modulators of vertebrate nAChRs. Moreover, 31 of the
65 active venoms contained non-selective modulators that affected both α3* and
α7 nAChRs, whereas a smaller number (25) were selective modulators of
vertebrate α3* nAChRs. This suggests that during the course of evolution, non-
selective modulators of vertebrate nAChRs were selected for the purposes of
defence against vertebrate predators. However, future studies aimed at screening
a taxonomically broader range of spiders against a larger panel of vertebrate
nAChR subtypes will be required to address this hypothesis.
The current study also identified a non-selective agonist of vertebrate nAChRs in
the venom of the giant forest scorpion Heterometrus spinnifer (Scorpionidae). In
contrast with most channel modulators found in arachnid venoms, this nAChR
modulator was identified as senecioylcholine, a choline ester. Senecioylcholine
was previously identified in the hypobranchial gland of some predatory marine
gastropods (family Muricidae) but this is the first report of it being present in
scorpion venom. H. spinnifer is one of the largest scorpion species but its venom
is no lethal to humans. However, envenomation of humans by H. spinnifer leads to
intense pain and inflammation, and senecioycholine may contribute to these
symptoms. Thus, this study provides the first evidence for convergent evolution of
a non-peptide toxin directed against vertebrate nAChRs. It would be interesting in
future studies to examine whether senecioylcholine and related choline ester are
found more broadly in scorpion venoms, particularly in species that are not
relevant in clinic.
While the current study focussed on vertebrate nAChRs, it would be of great
interest in future to examine whether arachnid venoms also target nAChR

96
subtypes found in their major prey, namely insects. Compounds that specifically
targeted insect, but not vertebrate, nAChRs would be of significant interest as
bioinsecticide leads. In this respect, it might therefore be interesting to examine
venoms that were shown in this study to be inactive against vertebrate nAChRs,
such as those from the araneomorph spider Heteropoda jugulans and the
mygalomorph spider Aphonopelma crinitum.
Most venom research to date has focussed on animals of medical relevance, but
this study has highlighted the value of studying venoms with non-lethal effects on
humans when searching for novel compounds. The continued development of
high-throughput screening methods and analytical tools that require only small
amounts of venom will provide more opportunities in future for exploring the vast
pharmacological repertoire encoded within the venoms of small venomous animals
such as spiders and scorpions.

97
REFERENCES
1. AhnAllen, C.G., 2012. The role of the α7 nicotinic receptor in cognitive
processing of persons with schizophrenia. Curr. Opin. Psychiatry 25,
103 –108.
2. Aicher, A., 2003. Nicotine strongly activates dendritic cell-mediated
adaptive immunity: potential role for progression of atherosclerosis.
Circulation 107, 604–611.
3. Akalal, D.B., Wilson, C., Zong, L., Tanaka, N., Ito, K. and Davis, R.,
2006. Roles for Drosophila mushroom body neurons in olfactory
learning and memory. Learning and Memory 13, 659–668.
4. Alamanda, V., Singh, S., Lawrence, N.J. and Chellapan, S.P., 2012.
Nicotine-mediated induction of E-selectin in aortic endothelial cells
requires Src kinase and E2F1 transcriptional activity. Biochem.
Biophys. Res. Commun. 418, 56–61.
5. Albaugh, G., Bellavance, E., Strande, L., Heinburger, S., Hewitt, C.W.
and Alenxander, J.B., 2004. Nicotine induces mononuclear leukocyte
adhesion and expression of adhesion molecules, VCAM and ICAM, in
endothelial cells in vitro. Ann. Vasc. Surg. 18, 302–307.
6. Albuquerque, E.X., Pereira, E.F.R., Alkondon, M. and Rogers, S.W.,
2009. Mammalian nicotinic acetylcholine receptors: from structure to
function. Physiol. Rev. 89, 73–120.
7. Armishaw, C.J., 2010. Synthetic α-conotoxin mutants as for studying
nicotinic acetylcholine receptors and in the development of novel drug
leads. Toxins 2, 1471–1499.
8. Arneric, S.P., Holladay, M. and Williams, M., 2007. Neuronal nicotinic
receptors: a perspective on two decades of drug discovery research.
Biochem. Pharmacol. 74, 1092 –1101.

98
9. Ayroza, G., Ferreira, I.L., Sayegh, R.S., Tashima, A.K. and da Silva
Junior P.I., 2012. Juruin: an antifungal peptide from the venom of the
amazonian pink toe spider, Avicularia juruensis, which contains the
inhibitory cystine knot motif. Front. Microbiol. 3, 324.
10. Baker, J.T., Duke, C.C., 1976. Isolation of choline and choline ester
salts of tyrindoxyl sulphate from the marine molluscs Dicathais orbita
and Mancinella keineri. Tetrahedon Lett. 17, 1233–1234.
11. Baker, J.T., Duke, C.C., 1978. N-methylmurexine a naturally occuring
marine compound? Tetrahedon Lett. 19, 5047–5048.
12. Bauwens, M., Mottaghy, F.M. and Bucerius, J., 2015. PET imaging of
the human nicotinic cholinergic pathway in atherosclerosis. Curr.
Cardiol. Rep. 17, 67.
13. Bencherif, M., Narla, S.T. and Stachowiak, M.S., 2014. Alpha7
neuronal nicotinic receptor: a pluripotent target for diseases of the
central nervous system. CNS Neurol. Disord. Drug Targets 13, 836–
845.
14. Benkendorff, K., 2013. Natural product research in the Australian
marine invertebrate Dicathais orbita. Mar. Drugs 11, 1370–1398.
15. Bertani, R., Fukushima, C.S., 2009. Description of two new species of
Avicularia Lamarck 1818 and redescription of Avicularia diversipes
(C.L. Koch 1842) (Araneae, Theraphosidae, Aviculariinae)-three
possibly threatened Brazilian species. Zootaxa 2223, 25–47.
16. Billen, B., Bosmans, F. and Tytgat, J., 2008. Animal peptides targeting
voltage-activated sodium channels. Curr. Pharm. Des. 14, 2492–2502.
17. Bond, J.E., Hendrixson, B.E., Hamilton, C.A. and Hedin, M., 2012. A
reconsideration of the classification of the spider infraorder
Mygalomorphae (Arachnida: Araneae) based on three nuclear genes
and morphology. PLos One 7, e38753.

99
18. Bordia, T., Hrachova, M., Chin, M., McIntosh, J.M. and Quik, M., 2012.
Varenicline is a potent partial agonist at α6β2* nicotinic acetylcholine
receptors in rat and monkey striatum. Neuropharmacology 342, 327–
334.
19. Bourne, Y., Talley, T. T., Hansen, S. B., Taylor, P. and Marchot, P. ,
2005. Crystal structure of a Cbtx-AChBP complex reveals essential
interactions between snake α-neurotoxins and nicotinic receptors.
EMBO J. 24, 1512–1522.
20. Brahma, R.K., McCleary, R.J., Kini, R.M. and Doley, R., 2015. Venom
gland transcriptomics for identifying, cataloging, and characterizing
venom proteins in snakes. Toxicon 93, 1–10.
21. Brown, L.A., Ihara, M., Buc kingham, S.D., Matsuda, K. and Sattelle,
D.B., 2006. Neonicotinoid insecticides display partial and super agonist
actions on native insect nicotinic acetylcholine receptors. J.
Neurochem. 99, 608–615.
22. Bucher, D., Goaillard, J.M., 2011. Beyond faithful conduction: short-
term dynamics, neuromodulation, and long-term regulation of spike
propagation in the axon. Prog. Neurobiol. 94, 307–346.
23. Buchman, A.L., Dubin, M., Moukarzel, A., Jenden, D., Roch, M.H.,
Rice, K., Gorbein, J. and Ament, M.E., 1995. Choline defficiency: a
cause of hepatic steatosis during parenteral nutrition that can be
reversed with intravenous choline supplementation. Hepatology 22,
1399–1403.
24. Caliskan, F., Quintero-Hernandez, V., Restano-Cassulini, R., Baptista,
C.V., Zamudio, F.Z., Coronas, F.I. and Possani, L., 2012. Turkish
scorpion Buthacus macrocentrus: general characterization of the
venom and description of Bu1, a potent mammalian Na+-channel α-
toxin. Toxicon 59, 408–415.

100
25. Camargos, T.S., Restano-Cassulini, R., Possani, L.D., Peigneur, S.,
Tytgat, J., Schwartz, C.A. Alves, E.M., de Freitas, S.M., and Schwartz,
E.F., 2011. The new kappa-KTx2.5 from the scorpion Opisthacanthus
cayaporum. Peptides 32, 1509–1517.
26. Caputi, L., Bengtson, C.P., Guatteo, E., Bernardi, G. and Mercuri, N.B.,
2003. D-tubocurarine reduces GABA responses in rat substantia nigra
dopamine neurons. Synapse 47, 236–239.
27. Changeux, J., Kasai, M. and Lee, C. , 1970. Use of a snake venom
toxin to characterize the cholinergic receptor protein. Proc. Natl. Acad.
Sci. U. S. A. 67, 1241–1247.
28. Changeux, J.P., 2010. Nicotine addiction and nicotinic receptors:
lessons from genetically modified mice. Nat. Rev. Neurosci. 11, 389–
401.
29. Cheikh, A., Benkhalifa, R., Potreau, D., Raymond, G., El Ayeb, M. and
Cognard, C., 2007. A novel depolarizing activity of scorpion venom
fraction M1 due to activation of skeletal muscle nicotinic receptors.
Toxicon 49, 117–122.
30. Chu, L.W., Ma, E.S., Lam, K.K., Chan, M.F. and Lee, D.H., 2005.
Increased α7 nicotinic acetylcholine receptor protein in Alzheimer' s
disease patients. Dement. Geriatr. Cogn. Disord. 19, 106–112.
31. Chugunov, A.O., Koromyslova, A.D., Berkut, A.A., Peigneur, S., Tytgat,
J., Polyansky, A.A., Pentkovsky, V.M., Vassilevski, A.A., Grishin, E.V.
and Efremov, R.G., 2013. Modular organization of α- toxins from
scorpion venom mirrors domain structure of their targets, sodium
channels. J. Biol. Chem. 288, 19014–19027.
32. Clot-Faybesse, O., Guieu, R., Rochat, H. and Devaux, C., 2000.
Toxicity during early development of the mouse nervous system of a
scorpion neurotoxin active on sodium channels. Life Sci. 66, 185–192.

101
33. Coddington, J.A., Levi, H.W., 1991. Systematics and evolution of
spiders (Araneae). Annu. Rev. Ecol. Evol. Syst. 22, 565–592.
34. Colloby, S.J., Perry, E.K., Pakrasi, S., Pimlott, S.L., Wyper, D.J.,
McKeith, I.G., Williams, E.D. and O'Brien, J.T., 2010. Nicotinic 123I-
5IA-85380 single photon emission computed tomography as a
predictor of cognitive progression in Alzheimer's disease and dementia
with Lewy bodies. Am. J. Geriatr. Psychiatry 18, 86–90.
35. Cooper, E., Couturier, S. and Ballivet, M. , 1991. Pentameric structure
and subunit stoichiometry of a neuronal nicotinic acetylcholine
receptor. Nature 350, 235–238.
36. Corrigall, W.A., Coen, K.M. and Adamson, K.L., 1994. Self-
administered nicotine activates the mesolimbic dopamine system
through the ventral tegmental area. Brain Res. 653, 278–284.
37. Corrigall, W.A., Franklin, K.B., Coen, K.M. and Clarke, P.B., 1992. The
mesolimbic dopaminergic system is implicated in the reinforcing effects
of nicotine. Psychopharmacology (Berl) 107, 285–289.
38. Corringer, P.J., Poitevin, F., Prevost, M.S., Sauguet, L., Delarue, M.
and Changeux, J.P., 2012. Structure and pharmacology of pentameric
receptor channels: from bacteria to brain. Structure 20, 941–956.
39. Corzo, G., Diego-Garcia, E., Clement, H., Peigneur, S., Odell, G.,
Tytgat, J., Possani, L.D. and Alagon, A., 2008. An insecticidal peptide
from the theraphosid Breachypelma smithi spider venom reveals
common molecular features among spider species from different
genera. Peptides 29, 1901–1908.
40. Corzo, G., Villegas, E., Gómez-Lagunas, F., Possani, L.D.,
Belokoneva, O.S., and Nakajima, T., 2002. Oxyopinins, large
amphipathic peptides isolated from the venom of the wolf spider
Oxyopes kitabensis with cytolytic properties and positive insecticidal

102
cooperativity with spider neurotoxins. J. Biol. Chem. 277, 23627–
23637.
41. Couturier, S., Bertrand, D., Matter, J.M., Hernandez, M. C., Bertrand,
S., Millar, N., Valera, S., Barkas, T. and Ballivet, M., 1990. A neuronal
nciotinic acetylcholine receptor subunit (α 7) is developmentally
regulated and forms a homo-oligomeric channel blocked by α-BTX.
Neuron 5, 847–856.
42. Dajas-Bailador, F.A., Mogg, A.J. and Wonnacott, S., 2002. Intracellular
Ca2+ signals evoked by stimulation of nicotinic acetylcholine receptors
in SH-SY5Y cells: contribution of voltage-operated Ca2+ channels and
Ca2+ stores. J. Neurochem. 81, 606–614.
43. DeBin, J.A., Maggio, J.E. and Strichartz, G.R., 1993. Purification and
characterization of chloro-toxin, a chloride channel ligand from the
venom of the scorpion. Am. J. Physiol. 264, C361–C369.
44. Déglise, P., Grünewald, B. and Gauthier, M., 2002. The insecticide
imidacloprid is a partial agonist of the nicotinic receptor of homneybee
kenyon cells. Neurosci. Lett. 321, 13–16.
45. Diaz, P., D'Suze, G., Salazar, V., Sevcik, C., Shannon, J.D., Sherman,
N.E. and Fox, J.W., 2009. Antibacterial activity of six novel peptides
from Tityus discrepans scorpion venom. A fluorescent probe study of
microbial membrane Na+ permeability changes. Toxicon 54, 802–817.
46. Ding, Y.S., Fowler, J.S., Logan, J., Wang, G.J., Telang, F., Garza, V.,
Biegon, A., Pareto, D., Rooney, W., Shea, C., Alexoff, D., Volkow, N.D.
and Vocci, F. , 2004. 6-[18F]Fluoro-A-85380, a new PET tracer for the
nicotinic acetylcholine receptor: studies in the human brain and in vivo
demonstration of specific binding in white matter. Synapse 53, 184–
189.

103
47. Doran, N.E., Richardson, A.M.M. and Swain, R., 2001. The
reproductive behaviour of Hickmania troglodytes, the tasmanian cave
spider (Araneae, Austrochilidae). J. Zool. 253, 405–418.
48. Drevets, W.C., Price, J.L. and Furey, M.L., 2008. Brain structural and
functional abnormalities in mood disorders: implications for
neurocircuitry models of depression. Brain Struct. Funct. 213, 93–118.
49. Dutertre, S., Nicke, A., Tyndall, J.D. and Lewis, J.R., 2004.
Determination of α-conotoxin binding modes on neuronal nicotinic
acetylcholine receptors. J. Mol. Recognit. 17, 339–347.
50. Dutertre, S., Ulens, C., Buttner, R., Fish, A., van Elk, R., Kendel, Y.,
Hopping, G., Alewood, P.F., Schroeder, C., Nicke, A., Smit, A.B.,
Sixma, T.K. and Lewis, J.R., 2007. AChBP-targeted α-conotoxin
correlates distinct binding orientations with nAChR subtype selectivity.
EMBO J. 26, 3858–3867.
51. Egea, J., Buendia, I., Parada, E., Navarro, E., León , R. and Lopez,
M.G., 2015. Anti-inflammatory role of microglial alpha7 nAChRs and its
role in neuroprotection. Biochem. Pharmacol. 97, 463–472.
52. Egleton, R.D., Brown, K.C. and Dasgupta, P., 2009. Angiogenic activity
of nicotinic acetylcholine receptors: implications in tobacco-related
vascular diseases. Pharmacol. Ther. 121, 205–223.
53. Ehret-Sabatier, L., Loew, D., Goyffon, M., Fehlbaum, P., Hoffmann,
J.A., van Dorsselaer, A. and Bulet, P., 1996. Characterization of novel
cysteine-rich antimicrobial peptides from scorpion blood. J. Biol. Chem.
271, 29537–29544.
54. Elgoyhen, A.B., Vetter, D.E., Katz, E., Rothlin, C.V., Heineman, S.F.
and Boulter, J., 2001. α10: a determinant of nicotinic cholinergic
receptor function in mammalian vestibular and cochlear
mechanosensory hair cells. Proc. Natl. Acad. Sci. U. S. A. 98, 3501–
3506.

104
55. Emre, M., 2003. Dementia associated with Parkinsons's disease.
Lancet Neurol. 2, 229–237.
56. Erspamer, V., Glässer, A., 1957. The pharmacological actions of
murexine (urocanylcholine). Brit. J. Pharmacol. 12, 176–184.
57. Erspamer, V.a.B., O., 1953. Isolation of murexine from extracts of
hypobranchial glands of Murex trunculus and its identification as beta-
[imidazolyl-4(5)]-acryl-choline]. Biochem. Z. 324, 66–73.
58. Escoubas, P., 2006a. Mass spectrometry in toxinology: a 21st-century
technology for the study of biopolymers from venoms. Toxicon 47,
609–613.
59. Escoubas, P., Diochot, S. and Corzo, G., 2000. Structure and
pharmacology of spider venom neurotoxins. Biochimie 82, 893–907.
60. Escoubas, P., Sollod, B. and King, G.F., 2006b. Venom landscapes:
minig the complexity of spider venoms via a combined cDNA and mass
spectrometric approach. Toxicon 47, 650–663.
61. Estrada, G., Villegas, E. and Corzo, G., 2007. Spider venoms: a rich
source of acylpolyamines and peptides as new leads for CNS drugs.
Nat. Prod. Rep. 24, 145–161.
62. Faucon, J.P., Aurières, C., Drajnudel, P., Mathieu, L., Ribière, M.,
Martel, A.C., Zeggane, S., Chauzat, M.P. and Aubert, M.F., 2005.
Experimental study on the toxicity of imidacloprid given in syrup to
honey bee (Apis mellifera) colonies. Pest. Manag. Sci. 61, 111–125.
63. Fioravanti, M., Yanagi, M., 2005. Cytidinediphosphocholine (CDP-
choline) for cognitive and behavioural disturbances associated with
chronic cerebral disorders in the elderly. Cochrane Database Syst.
Rev. 2, CD000269.

105
64. Fischer, J., Müller, T., Spatz, A.K., Greggers, U., Grünewald, B. and
Menzel, R., 2014. Neonicotinoids interfere with specific components of
navigation in honeybees. PLos One 9, e91364.
65. Fisher, M., Huangfu,D., Shen, T.Y. and Guyenet, P.G., 1994.
Epibatidine: an alkaloid from the poison frog Epipedobates tricolor is a
powerful ganglionic depolarizing agent. J. Pharmacol. Exp. Ther. 270,
702–707.
66. Florey, E., 1963. Acetylcholine in invertebrate nervous system. Can. J.
Biochem. Physiol. 41, 2619–2626.
67. Forsyth, P., Sevcik, C., Martinez, R., Castillo, C. and D'Suze, G., 2012.
Bactridine's effects on DUM cricket neurons under voltage clamp
conditions. J. Insect. Physiol. 58, 1676–1685.
68. Fry, B.G., Wüster, W., Kini, R.M., Brusic, V., Khan, A., Venkataraman,
D. and Rooney, A.P., 2003. Molecular evolution and phylogeny of
elapid snake venom three-finger toxins. J. Mol. Evol. 57, 110–129.
69. García, A.P., Aitta,-Aho, T., Schaaf, L., Heeley, N., Heuschmid, L., Bai,
Y., Barrantes, F.J. and Apergis-Schoute, J., 2015. Nicotinic α4
receptor-mediated cholinergic influences on food intake and activity
patterns in hypothalamic circuits. PLos One 10, e0133327.
70. Gershoni, J.M., 1987. Expression of the α-bungarotoxin binding site of
the nicotinic acetylcholine receptor by Escherichia coli transformants.
Proc. Natl. Acad. Sci. U. S. A. 84, 4318–4321.
71. Gotti, C., Moretti, M., Gaimarri, A., Zanardi, A., Clementi, F. and Zoli,
M., 2007. Heterogeneity and complexity of native brain nicotinic
receptors. Biochem. Pharmacol. 74, 1102 –1111.
72. Grauso, M., Reenan, R.A., Culetto, E. and Sattelle, D.B., 2002. Novel
putative nicotinic acetylcholine receptor subunit genes, Dα5, Dα6 and
Dα7, in Drosophila melanogaster identify a new and highly conserved

106
target of adenosine deaminase acting on RNA-mediated A-to-I pre-
mRNA editing. Genetics 160, 1519–1533.
73. Griswold, C.E., Ramirez, M.J., Coddington, J.A. and Norman, I.P.,
2005. Atlas of phylogenetic data for entelegyne spiders (araneae:
araneomorphae: entelegynae) with comments on their phylogeny.
Proc. Calif. Acad. Sci. 56, 1–324.
74. Halldin, C., Nagren, K., Swahn, C-G., Langström, B. and Nybäck, H.,
1992. (S)- and (R)- (11C) nicotine and the metabolite (R/S)-[11C]
nicotine. Preparation, metabolite studies and in vivo distribution in the
human brain using PET. Nucl. Med. Biol. 19, 871–880.
75. Hannan, S., Mortensen, M. and Smart, T.G., 2015. Snake neurotoxin
α-bungarotoxin is an antagonist at native GABAA receptors.
Neuropharmacology 93, 28–40.
76. Harrison, P., Abdel-Rahman, M.A. and Miller, K., 2014. Antimicrobial
peptides from scorpion venoms. Toxicon 88, 115–137.
77. Harvey, M.S., Austin, A.D. and Adams, M., 2007. The systematics and
biology of the spider genus Nephila (Araneae: Nephilidae) in the
australasian region. Invertebr. Syst. 21, 407–451.
78. He, F., Li, B., Zhao, Z., Zhou, Y., Hu, G., Zou, W., Hong, W., Zou, Y.,
Jiang, C., Zhao, D. and Ran, P., 2014. The pro-proliferative effects of
nicotine and its underlying mechanism on rat airway smooth muscle
cells. PLos One 9, e93508.
79. Herzig, V.a.H., W.C., 2008. Neurotoxic and insecticidal properties of
venom from the australian theraphosid spider Selenotholus foelschei.
NeuroToxicology 29, 471–475.
80. Horti, A.G., Gao, Y., Kuwabara, H., Wang, Y., Abazyan, S., Yasuda,
R.P., Tran, T., Xiao, Y., Sahibzada, N., Holt, D.P., Kellar, K.J.,
Pletnikov, M.V., Pomper, M.G., Wong, D.F. and Dannals, R.F., 2014.

107
18F-ASEM, a radiolabeled antagonist for imaging the α7-nicotinic
acetylcholine receptor with PET. J. Nucl. Med. 55, 672–677.
81. Inserra, M., Kompella, S.N., Vetter, I., Brust, A., Daly, N.L., Cuny, H.,
Craik, D.J., Alewood, P.F., Adams, D.J. and Lewis, R.J., 2013.
Isolation and characterization of α -conotoxin LsIA with potent activity
at nicotinic acetylcholine receptors. Biochem. Pharmacol. 86, 791–799.
82. Isman, M., 2006. Botanical insecticides, deterrents, and repellents in
modern agriculture and an increasingly regulated world. Annu. Rev.
Entomol. 51, 45–66.
83. Jensen, A.A., Frølund, B., Liljefors, T. and Krogsgaard-Larsen, P.,
2005. Neuronal nicotinic acetylcholine receptors: structural revelations,
target identifications, and therapeutic inspirations. J. Med. Chem. 48,
4705–4745.
84. Jin, A.-H., Vetter, I., Dutertre, S., Abraham, N., Emidio, N.B., Inserra,
M., Murali, S.S., Christie, M. J., Alewood, P. and Lewis, R., 2014. MrIC,
a novel α-conotoxin agonist in the presence of PNU at endogenous α7
nicotinic acetylcholine receptors. Biochemistry 53, 1–3.
85. Jones, A.K., Brown, A.M. and Satelle, D.B., 2007. Insect nicotinic
acetylcholine receptor gene families: from egentic model organisms to
vector, pest and beneficial species. Invert. Neurosci. 7, 67–73.
86. Jones, A.K., Satelle, D.B., 2009. Diversity of insect nicotinic
acetylcholine receptor subunits. Adv. Exp. Med. Biol. 683, 25–43.
87. Jorenby, D.E., Hays, J.T., Rigotti, N.A., Azoulay, S., Watsky, E.J.,
Williamns, K.E., Billing, C.B., Gong, J., Reeves, K.R. and Vareniclne
phase 3 study group, 2006. Efficacy of varenicline, and alpha4beta2
nicotinic acetylcholine receptor partial agonist, vs placebo or sustained-
release bupropion for smoking cessation: a randomized controlled trial.
JAMA 296, 56–63.

108
88. Kalayci, A., Eren, A., Kocabay, G., Karabay, C.Y., Gecmen, C. and
Kirma, C., 2013. Varenicline-induced coronary thrombosis. Ann.
Pharmacother. 47, 1727–1729.
89. Kendziorra, K., Wolf, H., Meyer, P.M., Barthel, H., Hesse, S., Becker,
G.A., luthardt, J., Schildan, A., Patt, M., Sorger, D., Seese, A., Gertz,
H.J. and Sabri, O., 2010. Decreased cerebral alpha4beta2* nicotinic
acetylcholine receptor availability in patients with mild cognitive
impairment and Alzheimer's disease assessed with positron emission
tomography. Eur. J. Nucl. Med. Mol. Imaging 38, 515–525.
90. Keyl, M.J., Michaelson, I.A. and Whittaker, V.P., 1957. Physiologically
active choline esters in certain marine gastropods and other
invertebrates. J. Physiol. 139, 434–454.
91. Keyl, M.J., Whittaker, V.P., 1958. Some pharmacological properties of
murexine (urocanoylcholine). Br. J. Pharmacol. Chemother. 13, 103–
106.
92. Kimes, A.S., Horti, A.G., London, E.D., Chefer, S.I., Contoreggi, C.,
Ernst, M., Friello, P., Koren, A.O., Kurian, V., Matochik, J.A., Pavlova,
O., Vaupel, D.B. and Mukhin, A.G., 2006. 2-[18F]F-A-85380: PET
imaging of brain nicotinic actylcholine receptors and whole body
distribution in humans. FASEB J. 17, 1331–1333.
93. King, G.F., 2014. The future of venoms-based drug discovery: an
interview with Glenn King. Future Medicinal Chemistry 16, 1613–1615.
94. King, G.F., Coaker, H., 2014. The future of venoms-based drug
discovery: an interview with Glenn King. Future Med. Chem. 6, 1613–
1615.
95. King, G.F., Hardy, M.C., 2013. Spider-venom peptides: structure,
pharmacology, and potential for control of insects pests. Annu. Rev.
Entomol. 58, 475–496.

109
96. Knott, V., de la Salle, S., Choueiry, J., Impey, D., Smith, D., Smith, M.,
Beaudry, E., Saghir, S., LLivitsky, V. and Labelle, A. , 2015.
Neurocognitive effects of acute choline supplementation in low,
medium and high performer. Pharmacol. Biochem. Behav. 131, 119–
129.
97. Koga, M., Kanaoka, Y., Ohkido, Y., Kubo, N., Ohishi, K., Sugiyama, K.,
Yamauchi, A. and Kataoka, Y., 2014. Varenicline aggravates plaque
formation through α7 nicotinic acetylcholine receptors in ApoE KO
mice. Biochem. Biophys. Res. Commun. 455, 194–197.
98. Korey, S.R., de Braganza, B. and Nachmansohn, D., 1951. Choline
acetylase. V. Esterifications and transacetylations. J. Biol. Chem. 189,
705–715.
99. Kromann, H., Krikstolaityte, S., Andersen, A.J., Andersen, K.,
Krogsgaard-Larsen, P., Jaroszewski, J.W., Egebjeerg, J. and
Strømgaard, K., 2002. Solid-phase synthesis of polyamine toxin
analogues: potent and selective antagonists of Ca2+-permeable AMPA
receptors. J. Med. Chem. 45, 5745–5754.
100. Kuhn-Nentwig, L., 2003. Antimicrobial and cytolytic peptides of
venomous arthropods. Cell Mol. Life Sci. 60, 2651–2668.
101. Kutlu, M.G., Gould, T.J., 2015. Nicotinic receptors, memory, and
hippocampus. Curr. Top. Behav. Neurosci. 23, 137–163.
102. Lam, S., Patel, P.N., 2007. Verenicline: a selective alpha4beta2
nicotinic acetylcholine receptor partial agonist approved for smoking
cessation. Cardiol. Rev. 15, 154–161.
103. Landsdell, S.J., Collins, T., Goodchild, J. and Millar, N.S., 2012.
The Drosophila nicotinic acetylcholine receptor subunits Dα5 and Dα7
form functional homomeric and heteromeric ion channels BMC
Neurosci. 13, 73.

110
104. Lansdell, S.J., Collins, T., Goodchild, J. and Millar, N.S., 2012.
The drosophila nicotinic acetylcholine receptor subunits Dα5 and Dα7
form functional homomeric and heteromeric ion channels BMC
Neurosci. 13, 73.
105. Lebbe, E.K., Peigneur, S., Maiti, M., Devi, P., Ravichandran, S.,
Lescrinier, E., Ulens, C., Waelkens, E., D'Souza, L., Herdewijn, P. and
Tytgat, J., 2014a. Structure-function elucidation of a new α-conotoxin,
Lo1a, from Conus longurionis. J. Biol. Chem. 289, 9573–9583.
106. Lebbe, E.K.M., Peigneur, S., Wijesekara, I. and Tytgat, J.,
2014b. Conotoxins targeting nicotinic acetylcholine receptors: an
overview. Mar. Drugs 12, 2970–3004.
107. Lewis, R.J., Dutertre, S., Vetter, I. and Christie, M.J. , 2012.
Conus venom peptide pharmacology. Pharmacol. Rev. 64, 259-–298.
108. Li, T., Wu, S., Zhang, H., Wang, Y., Luo, H., Zuo, X. and Xiao,
X., 2015. Activation of nicotinic receptors inhibits TNF-α-induced
production of pro-inflammatory mediators through the JAK2/STAT3
signaling pathway in fibroblast-like synoviocytes. Inflammation 38,
1424–1433.
109. Liang, S., 2008. Proteome and peptidome profiling of spider
venoms. Expert review of proteomics 5, 731–746.
110. Lopez, M.G., Montiel, C., Herrero, C.J., Garcia-Palomero, E.,
Mayorgas, I., Hernandez-Guijo, J.M., Villaroya, M., Olivares, R.,
Gandia, L., McInstosh, J.M., Olivera, B.M. and Garcia, A.G., 1998.
Unmasking the functions of the chromaffin cell α7 nicotinic receptor by
using short pulses of acetylcholine and selective blockers. Proc. Natl.
Acad. Sci. U. S. A. 95, 14184–14189.
111. Lotfipour, S., Mandelkem, M. and Brody, A.L., 2011.
Quantitative molecular imaging of neuronal nicotinic acetylcholine

111
receptors in the human brain with A-85380 radiotracers. Curr. Med.
Imaging Rev. 7, 107–112.
112. Lotfipour, S., Mandelkem, M., Alvarez-Estrada, M. and Brody,
A.L., 2012. A single administration of low-dose varenicline saturates
α4β2* nicotinic acetylcholine receptors in the human brain.
Neuropsychopharmacology 37, 1738–1748.
113. Macklin, K.D., Maus, A.D., Pereira, E.F., Albuquerque, E.X. and
Conti-Fine, B.M., 1998. Human vascular endothelial cells express
functional nicotinic acetylcholine receptors. J. Pharmacol. Exp. Ther.
287, 435–439.
114. Manjunatha, K.R., Robin, D., 2010. Structure, function and
evolution of three-finger toxins: mini proteins with multiple targets.
Toxicon 56, 855–867.
115. Mansvelder, H.D., Keath, J.R. and McGehee, D.S., 2002.
Synaptic mechanisms underlie nicotine-induced excitability of brain
reward areas. Neuron 33, 905–919.
116. Matsuda, K., Kanaoka, S., Akamatsu, M. and Sattelle, D.B.,
2009. Diverse actions and target-site selectivity of neonicotinoids:
structural insights. Mol. Pharmacol. 76, 1–10.
117. McGehee, D.S., Role, L.W., 1995. Physiological diversity of
nicotinic acetylcholine receptors expressed by vertebrate neurons.
Annu. Rev. Physiol. 57, 521–546.
118. McGuire, S., Le, P. and Davis, R., 2001. The role of Drosophila
mushroom body signalling in olfactory memory. Science 17, 1330–
1333.
119. McIntosh, J.M., Santos, A.D. and Olivera, B.M., 1999. Conus
peptides targeted to specific nicotinic acetylcholine receptor subtypes.
Annu. Rev. Biochem. 68, 59–88.

112
120. Mellor, I.R., Usherwood, P.N., 2004. Targeting ionotropic
receptors with polyamine-containing toxins. Toxicon 43, 493–508.
121. Meves, H., Simard, J.M. and Watt, D.D., 1984. Voltage-
dependent effect of a scorpion toxin on sodium current inactivation.
Eur. J. Physiol. Pflugers Arch. 402, 24–33.
122. Meyer, P.M., Strecker, K., Kendziorra, K., Becker, G., Hesse, S.,
Woelpl, D., Hensel, A., Patt, M., Sorger, D., Wegner, F., Lobsien, D.,
Barthel, H., Brust, P., Gertz, H.J. Sabri, O. and Schwartz, J., 2009.
Reduced alhpa4beta2*-nicotinic acetylcholine receptor binding and its
relationship to mild cognitive and depressive symptoms in Parkinson's
disease. Arch. Gen. Psychiatry. 66, 866–877.
123. Mihalak, K.B., Carroll, F.I. and Luetje, C.W., 2006. Varenicline is
a partial agonist at alpha4beta2 and a full agonist at alpha7 neuronal
nicotinic receptors. Mol. Pharmacol. 70, 801–805.
124. Miyazawa, A., Fujiyoshi, Y., Stowell, M. and Unwin, N. , 1999.
Nicotinic acetylcholine receptor at 4.6Å resolution: transverse tunnels
in the channel wall. J. Mol. Biol. 288, 765–786.
125. Moffat, C., Pacheco, J.G., Sharp, S., Samson, A.J., Bollan, K.A.,
Huang, J., Buckland, S.T. and Connolly, C.N., 2015. Chronic exposure
to neonicotinoids increases neuronal vulnerability to mitochondrial
dysfunction in the blumblebee (Bombus terrestris). FASEB J. 29,
2112–2119.
126. Molgó, J., Rómulo, A., Benoit, E. and Iorga, B.I., 2013. Physical
and virtual screening methods for marine toxins and drug discovery
targeting nicotinic acetylcholine receptors. Expert. Opin. Drug Discov.
8, 1203–1223.
127. Moore, S., Smyth, W.F., Gault, V.A., O'Kane, E. and McClean,
S., 2009. Mass spectrometric characterization and quantitation of
selected low molecular mass compounds from the venom of

113
Haplopelma lividum (Theraphosidae). Rapid Commun. Mass
Spectrom. 23, 1747–1755.
128. Morales-Lázaro, S.L., Hernández-García, E., Serrano-Flores, B.
and Rosenbaum, T. , 2015. Organic toxins as tools to understand ion
channels mechanisms and structure. Curr. Top. Med. Chem. 15, 581–
603.
129. Morgenstern, D., Rohde, B.H., King, G.F., Tal, T., Sher,D. and
Zlotkin, E., 2011. The tale of a resting gland: transcriptome of a replete
venom gland from the scorpion Hottentotta judaicus. Toxicon 57, 695–
703.
130. Mourão, C.B., Heghinian, M.D., Barbosa, E.A., Marí, F., Bloch,
C. Jr., Restano-Cassulini, R., Possani, L.D. and Schwartz, E.F., 2013.
Characterization of a novel peptide toxin from Acanthoscurria paulensis
spider venom: a distinct cysteine assignment to the HWTX-II family.
Biochemistry 52, 2440–2452.
131. Müller, M.L.T.M., Bohnen, N., 2013. Cholinergic dysfunction in
Parkison's disease. Curr. Neurol. Neurosci. Rep. 13, 377.
132. Nicke, A., Wonnacott, S. and Lewis, R., 2004. α-conotoxins as
tools for the elucidation of structure and function of neuronal nicotinic
acetylcholine receptor subtypes. Eur. J. Biochem. 271, 2305–2319.
133. Nides, M., Oncken, C., Gonzales, D., Rennard, S., Watsky, E.J.,
Anziano, R. and Reeves, K.R., 2006. Smoking cessation with
varenicline, a selective α4β2 nicotinic receptor partial agonist: results
from a 7-week, randomized, placebo- and bupropion-controlled trial
with 1-year follow-up. Arch. Intern. Med. 166, 1561–1568.
134. Nie, Y., Zeng, X.C., Yang, Y., Luo, F., Luo, X., Wu, S., Zhang, L.
and Zhou, J., 2012. A novel class of antimicrobial peptides from the
scorpion Heterometrus spinnifer. Peptides 38, 389–394.

114
135. Nirthanan, S., Pil, J., Abdel,-Mottaleb, Y., Gopalakrishnakone,
P., Joseph, J.S., Sato, K. and Tytgat, J., 2005. Assignment of voltage-
gated potassium channel blocking activity to kappa-KTx1.3, a non-toxic
homologue of kappa-hefutoxin-1, from Heterometrus spinifer venom.
Biochem. Pharmacol. 69, 669–678.
136. Nirthanan, S., Pil, J., Joseph, J.S., Gopalakrishnakone, P.,
Joseph, J.S., Khoo, H.E., Cheah, L.S. and Gwee, M.C.E., 2002.
Biochemical and pharmacological characterization of the venom of the
black scorpion Heterometrus spinnifer. Biochem. Pharmacol. 63, 49–
55.
137. Nørager, N.G., Poulsen, M.H., Jensen, A.G., Jeppesen, N.S.,
Kristensen, A.S. and Strørmgaard, K., 2014. Structure-activity
relationships studies of N-methylated and N-hydroxylated spider
polyamine toxins as inhibitors of ionotropic glutamate receptors. J.
Med. Chem. 57, 4940–4949.
138. Norton, R.S., McDonough, S.I., 2008. Peptides targeting
voltage-gated calcium channels. Curr. Pharm. Des. 14, 2480–2491.
139. Nybäck, H., Halldin, C., Ahlin, A., Curvall, M. and Eriksson, L. ,
1994. PET studies of the uptake of (S)- and (R)-11C-nicotine in the
human brain: difficulties in visualizing specific receptor binding in vivo.
Psychopharmacology 115, 31–36.
140. Nyffeler, M., Pusey, B.J., 2014. Fish predation by semi-auqatic
spiders: a global pattern. PLos One 9, e99459.
141. Olsen, C.A., Kristensen, A.S. and Strømgaard, K., 2011. Small
molecules from spiders used as chemical probes. Angew. Chem. Int.
Ed. 50, 11296–11311.
142. Olsen, C.A., Mellor, I.R., Wellendorph, P., Usherwood, P.N.,
Witt, M., Franzyk, H. and Jaroszewski, J.W., 2006. Turning wasp toxin
structure for nicotinic receptor antagonism: cyclohexylalanine-

115
containing analogues as potent and voltage-dependent blockers.
Chem. Med. Chem. 1, 303–305.
143. Pammolli, F., Magazzini, L. and Riccaboni, M. , 2011. The
productivity crisis in pharmaceutical R & D. . Nat. Rev. Drug Discov.
10, 428–438.
144. Parikh, V., Ji, J., Decker, M.W. and Sarter, M., 2010. Prefrontal
beta2 subunit-containing and alpha7 nicotinic acetylcholine receptors
differentially control glutamatergic and cholinergic signaling. J.
Neurosci. 30, 3518–3530.
145. Perry, D.C., Xiao, Y., Nguyen, H.N., Musachio, J.L., Dávila-
García, M.I. and Kellar, K.J., 2002. Measuring nicotinic receptors with
characteristics of alpha4beta2, alpha3beta2 and alpha3beta4 subtypes
in rat tissues by autoradiography. J. Neurochem. 82, 468–481.
146. Picciotto, M.R., Higley, M.J. and Mineur, Y.S., 2012.
Acetylcholine as a neuromodulator: cholinergic signaling shapes
nervous system function and behavior. Neuron 76, 116–129.
147. Picciotto, M.R., Kenny, P.J., 2013. Molecular mechanisms
underlying behaviors related to nicotine addiction. Cold Spring Harb.
Perspect. Med. 3, a012112.
148. Piek, T., Mantel, P. and Engels, E., 1971. Neuromuscular block
in insects caused by the venom of the digger wasp Philanthus
triangulum F. Comp. Gen. Pharmacol. 2, 317–331.
149. Piek, T., Njio, K.D., 1975. Neuromuscular block in honeybees by
the venom of the bee wolf wasp (Philanthus triangulum F.). Toxicon 13,
199–201.
150. Plummer, H.K.r., Dhar, M. and Schuller, H.M., 2005. Expression
of the alpha7 nicotinic acetylcholine receptor in human lung cells.
Respir. Res. 6, 29.

116
151. Quik, M., McIntosh, J.M., 2006. Striatal α-6* nicotinic
acetylcholine receptors: potential targets for Parkinson's disease
therapy. J. Pharmacol. Exp. Ther. 316, 481–489.
152. Quintero-Hernandez, V., Jimenez-Vargas, J.M., Gurrola, G.B.,
Valdivia, H.H.F. and Possani, L., 2013. Scorpion venom components
that affect ion-channels function. Toxicon 76, 328–342.
153. Rinne, J.O., Myllykylä., Lönnberg, P. and Marjamäki, P., 1991. A
postmortem study of brain nicotinic receptors in Parkinson's and
Alzheimer's disease. Brain. Res. 547, 167–170.
154. Rjeibi, I., Mabrouk, K., Mosrati, H., Berenguer, C., Mejdoub, H.,
Villard, C., Laffitte, D., Ouafik, L., Luis, J., Elayeb, M. and Srairi-Abid,
N., 2011. Purification, synthesis and characterization of AaCtx, the first
chlorotoxin-like peptide from Androctonus australis scorpion venom.
Peptides 32, 656–663.
155. Rocha-E-Silva, T.A., Rostelato-Ferreira, S., Leite, G.B., DaSilva
P.I J.R., Hyslop, S. and Rodrigues-Simioni, L. , 2013. VdTX1, a
reversible nicotinic receptor antagonist isolated from venom of the
spider Vitalius dubius (Theraphosidae). Toxicon 70, 135–141.
156. Rollema, H., Coe, J.W., Chambers, L.K., Hurst, R.S., Stahl, S.M.
and Williams, K.E., 2007. Rationale, pharmacology and clinical efficacy
of partial agonists of alpha4beta2 nACh receptors for smoking
cessation. Trends Pharmacol. Sci. 28, 316–325.
157. Roly, Z.Y., Islam, M.M. and Reza, M.A., 2014. A comparative in
silico characterization of functional and physicochemical properties of
3FTx (three finger toxin) proteins from four venomous snakes.
Bioinformation 10, 281–287.
158. Roseghini, M., Severini, C., Erspamer, G.F. and Erspamer, V.,
1996. Choline esters and biogenic amines in the hypobranchial gland

117
of 55 molluscan species of the neogastropod muricoidea superfamily.
Toxicon 34, 33–35.
159. Rueter, L.E., Donnelly-Roberts, D.L., Curzon, P., Briggs, C.A.,
Anderson, D.J. and Bitner, R.S., 2006. A-85380: a pharmacological
probe for the preclinical and clinical investigation of the alphabeta
neuronal nicotinic acetylcholine receptor. CNS Drug Rev. 12, 100–112.
160. Russo, P., Del Bufalo, A., Milic, M., Salinaro, G., Fini, M. and
Cesario, M., 2014. Cholinergic receptors as target for cancer therapy in
a systems medicine perspective. Curr. Mol. Med. 14, 1126–1138.
161. Sattelle, D.B., Jones, A.K., Sattelle, B.M., Matsuda, K., Reenan,
R. and Biggin, P.C., 2005. Edit, cut and paste in the nicotinic
acetylcholine receptor gene family of Drosophila melanogaster.
Bioessays 27, 366–376.
162. Schedel, A., Thornton, S., Schloss, P., Kluter, H. and Bugert, P.,
2010. Human platelets express functional alpha7-nicotinic
acetylcholine receptors. Arterioscler. Thromb. Vasc. Biol. 31, 928–934.
163. Schmaljohann, J., Gündisch, D., Minnerop. M., Bucerius, J.,
Joe, A., Guhlke, S., Biersack, H.J. and Wüllner, U., 2006. In vitro
evaluation of nicotinic acetylcholine receptors with 2-[18F]F-A85380 in
Parkinson's disease. Nucl. Med. Biol. 33, 305–309.
164. Schmaljohann, J., Gündisch, D., Minnerop. M., Joe, A.,
Bucerius, J., Dittmar, C., Jessen, F., Guhlke, S. and Wüllner, U., 2005.
A simple and fast method for the preparation of n.c.a. 2-[18F]F-A85380
for human use. Appli. Radiat. Isot. 63, 433–435.
165. Selden, P.A., Gall, J.C., 1992. A triassic Mygalomorph spider
from the northern Vosges, France. Paleontology 35, 211–235.
166. Shahbazzadeh, D., Srairi-Abid, N., Feng, W., Ram, N.,
Borchani, L., Ronjat, M., Akbari, A., Pessah, I.N., De Waard, M. and El
Ayeb, M., 2007. Hemicalcin, a new toxin from the Iranian scorpion

118
Hemiscorpius lepturus which is active on ryanodine-sensitive Ca2+
channels. Biochem. J. 404, 89–96.
167. Shiomi, K., Ishii, M., Shimakura, K., Nagashima, Y. and Chino,
M., 1998. Tygloylcholine: a new choline ester toxin from the
hypobranchial gland of two spsecies of muricid gastropods (Thais
clavigera and Thais bronni). Toxicon 36, 795–798.
168. Siegwart, M., Graillot, B., Blachere Lopez, C., Besse, S., Bardin,
M., Nicot, P.C. and Lopez-Ferber, M., 2015. Resistance to bio-
insecticides or how to enhance their sustainability: a review. Front.
Plant. Sci. 6, 381.
169. Skinner, W.S., Dennis, P.A., Lui, A., Carney, R.L. and Quistad,
G.B., 1990. Chemical characterization of acylpolyamine toxins from
venom of a trap-door and two tarantulas. Toxicon 28, 541–546.
170. Sousa, S.R., Vetter, I., Ragnarsson, L. and Lewis, R., 2013.
Expression and pharmacology of endogenous Cav channels in SH-
SY5Y human neuroblastoma cells. PLos One 8, e59293.
171. Steilein, O.K., 2001. Genes and mutations in idiopathic epilepsy.
Am. J. Med. Genet. 106, 139–145.
172. Strømgaard, K., Andersen, K., Krogsgaard-Larsen, P. and
Jaroszewski, J.W., 2001. Recent advances in the medicinal chemistry
of polyamine toxins. Mini Rev. Med. Chem. 1, 317–338.
173. Strømgaard, K., Jensen, L.S. and Vogensen, S.B., 2005.
Polyamine toxins: development of selective ligands for ionotropic
receptors. Toxicon 45, 249–254.
174. Strømgaard, K., Mellor, I., 2004. AMPA receptor ligands:
synthetic and pharmacological studies of polyamines and polyamine
toxins. Med. Res. Rev. 24, 589–620.

119
175. Sullivan, J.P., Donnelly-Roberts, D., Briggs, C.A., Anderson,
D.J., Gopalakrishnan, M., Piattoni-Kaplan, M., Campbell, J.E.,
McKenna, D.G., Molinari, E., Hettinger, A.M., Garvey, D.S., Wasicak,
J.T., Holladay, M.W., Williams, M. and Americ, S.P., 1996. A-85380 [3-
2(S)-azetidinylmethoxy) pyridine]: in vitro pharmacological properties of
a novel, high affinity alpha4 beta 2 nicotinic acetylcholine receptor
ligand Neuoropharmacology 35, 725–734.
176. Sunagar, K., Jackson, T.N.W, Undheim, E.A.B., Ali, S.A.,
Antunes, A. and Fry, B.G., 2013. Three-fingered RAVERs: rapid
accumulation of variations in exposed residues of snake venom toxins.
Toxins (Basel) 5, 2172–2208.
177. Swartz, K.J., 2013. The scorpion toxin and the potassium
channel. elife 2.
178. Terlau, H., Olivera, B.M., 2004. Conus venoms: a rich source of
novel ion channel-targeted peptides. Physiol. Rev. 84, 41–68.
179. Trachsel, C., Siegemund, D., Kämpfer, U., Kopp, L.S., Bühr, C.,
Grossmann, J., Lüthi, C., Cunningham, M., Nentwig, W., Kuhn-
Nentwig, L., Schurch, S. and Schaller, J., 2012. Multicomponent
venoms of the spider Cupiennius salei: a bioanalytical investigation
applying different strategies. FEBS J. 279, 2683–2694.
180. Tregellas, J.R., Tanabe, J., Rojas, D.C., Shatti, S., Olincy, A.,
Johnson, L., Martin, L.F., Soti, F., Kem, W.R., Leonard, S. and
Freedman, R., 2011. Effects of an α7 -nicotinic agonist on default
network activity in schizophrenia. Biol. Psychiatry. 69, 7–11.
181. Tsetlin, V., Hucho, F., 2009. Nicotinic acetylcholine receptors at
atomic resolution. Curr. Opin. Pharmacol. 9, 306–310.
182. Tsetlin, V.I., 2015. Three-finger snake neurotoxins and Ly6
proteins targeting nicotinic acetylcholine receptors: pharmacological

120
tools and endogenous modulators. Trends Pharmacol. Sci. 36, 109–
123.
183. Tuesta, L., Fowler, C.D. and Kenny, P.J., 2011. Recent
advances in understanding nicotinic receptor signalling mechanisms
that regulate drug self-administration behavior. Biochem. Pharmacol.
82, 984–995.
184. Unwin, N., 1993. Nicotinic acetylcholine receptor at 9Å
resolution J. Mol. Biol. 229, 1101–1124.
185. Unwin, N., 2005. Refined structure of the nicotinic acetylcholine
receptor at 4Å resolution. J. Mol. Biol. 346, 967–989.
186. Utkin, Y.N., 2015. Animal venom studies: current benefits and
future developments. World J. Biol. Chem. 6, 28-–33.
187. van Enkhuizen, J., Janowsky, D.S., Olivier, B., Minassian, A.,
Perry, W., Young, J.W. and Geyer, M.A., 2015. The catecholaminergic-
cholinergic balance hypothesis of bipolar disorder revisited. Eur. J.
Biochem. 753, 114–126.
188. Vassilevski, A.A., Kozlov, S.A. and Grishin, E.V., 2009.
Molecular diversity of spider venom. Biochemistry (Moscow) 74, 1505–
1534.
189. Vassilevski, A.A., Kozlov, S.A., Samsonova, O.V., Egorova,
N.S., Karpunin, D.V., Pluzhnikov, K.A., Feofanov, A.V. and Grishin,
E.V., 2008. Cyto-insectotoxins, a novel class of cytolytic and
insecticidal peptides from spider venom. Biochem. J. 411, 687–696.
190. Vassilevski, A.A., Sachkova, M.Y., Ignatova, A.A., Kozlov, S.A.,
Feofanov, A.V. and Grishin, E.V., 2013. Spider toxins comprising
disulfide-rich and linear amphipathic domains: a new class of
molecules identified in the lynx spider Oxyopes takobius. FEBS J. 280,
6247–6261.

121
191. Vetter, I., 2012. Development and optimization of FLIPR high
throughput calcium assays for ion channels and GPCRs. Adv. Exp.
Med. Biol. 740, 45–82.
192. Vetter, I., Davis, J.L., Rash, L.D., Anagi, R., Mobli, M., Alewood,
P.F., Lewis, R. and King, G.F., 2011. Venomics: a new paradigm for
natural products-based drug discovery. Amino acids 40, 15–28.
193. Vetter, I., Lewis, R.J., 2010. Characterization of endogenous
calcium responses in neuronal cell lines. Biochem. Pharmacol. 79,
908–920.
194. Vollrath, F., 1985. Web spider's dilemma: a risky move or site
dependent growth. Oecologia 68, 69–72.
195. Wada, T., Naito, M., Kenmochi, H. and Sasaoka, T., 2007.
Chronic nicotine exposure enhances insulin-induced mitogenic
signaling via up-regulation of alpha7 nicotinic receptors in isolated rat
aortic smooth muscle cells. Endocrinology 148, 790–799.
196. Wang, S., Zhao, C., Liu, Z., Wang, X., Liu, N., Du, W. and Dai,
Q., 2015. Structural and functional characterization of a novel α-
conotoxin Mr1.7 from Conus marmoreus targeting neuronal nAChR
α3β2, α9 α10 and α6/ α3β2β3 subtypes. Mar. Drugs 13, 3259–3275.
197. Wenningmann, I., Dilger, J.P., 2001. The kinetics of inhibition of
nicotinic acetylcholine receptors by (+)-tubocurarine and pancuronium.
Mol. Pharmacol. 60, 790–796.
198. Whittaker, V.P., 1960. Pharmacologically active choline esters in
marine gastropods. Ann. N. Y. Acad. Sci. 90, 695–705.
199. Williamson, S.M.a.W., G.A., 2013. Exposure to multiple
cholinergic pesticides imparirs olfactory learning and memory in
honeybees. J. Exp. Biol. 216, 1799–1807.

122
200. Willians, K., 1997. Interactions of polyamines with ion-channels.
Biochem. J. 325, 289–297.
201. Windley, M., Herzig, V., Dziemborowicz, S.A., Hardy, M., King,
G.F. and Nicholson, G.M., 2012a. Spider-venom as bioinsecticides.
Toxins (Basel) 4, 191–227.
202. Windley, M.J., Herzig, V.,Dziemborow, S.A., Hardy, M., King,
G.F. and Nicholson, G.M., 2012b. Spider-venom peptides as
bioinsecticides. Toxins 4, 191–227.
203. Wong, D.F., Kuwabara, H., Kim, J., Brasic, J.R., Chamroonrat,
W., Gao, Y., Valentine, H., Willis, W., Mathur, A., McCaul, M.E., Wand,
G., Gean, E.G., Dannals, R.F. and Horti, A.G., 2013. PET imaging of
high-affinity alpha4beta2 nicotinic acetylcholine receptors in humans
with 18F-AZAN, a radioligand with optimal brain kinetics. J. Nucl. Med.
54, 1308–1314.
204. Wu, S., Nie, Y., Zeng, X.C., Cao, H., Zhang, L., Zhou, L., Yang,
Y., Luo, X. and Liu, Y., 2014. Genomic and functional characterization
of three new venom peptides from the scorpion Heterometrus spinifer.
Peptides 53, 30–41.
205. Xiao, C., Miwa, J.M., Henderson, B.J., Wang, Y., Deshpande,
P., McKinney, S.L. and Lester, H.A., 2015. Nicotinic receptor subtype-
selective circuit patterns in the subthalamic nucleus. J. Neurosci. 35,
3734–3746.
206. Xie, S., Feng, J., Yu, C., Li, Z., Wu, Y., Cao, Z., Li, W., He, X.,
Xiang, M and Han, S., 2012. Identification of a new specific Kv1.3
channel blocker Ctri9577, from the scorpion Chaerilus tricostatus.
Peptides 36, 94–99.
207. Xu, X., Liu, F., Chen, J., Ono, H., Li, D. and Kuntner, M., 2015.
A genus-level taxonomic review of primitively segmented spiders
(Mesothelae, Liphistiidae). Zookeys 488, 121–151.

123
208. Yang, R., Zhao, Z., He, Y., Wu, L., Cai, D., Hong, W., Wu, Y.,
Cao, Z., Zheng, C. and Li, W., 2011. A new natural α-helical peptide
from the venom of the scorpion Heterometrus petersii kills HCV.
Peptides 32, 11–19.
209. Yu, D., Zhang, L., Eisele, J.L., Bertrand, D., Changeux, J.P. and
Weight, F.F., 1996. Ethanol inhibition of nicotinic acetylcholine type α7
receptors involves the amino-terminal domain of the receptor. Mol.
Pharmacol. 50, 1010–1016.
210. Zeisel, S.H., 1992. Choline: an important nutrient in brain
development, liver, function and carcinogenesis. J. Am. Coll. Nutr. 11,
473–481.
211. Zeisel, S.H., da Costa, K.A., 2009. Choline: an essential nutrient
for public health. Nutr. Rev. 67, 615–623.
212. Zelanis, A., Andrade-Silva, D., Rocha, M.M., Furtado, M.F.,
Serrano, S.M., Junqueira-de-Azevedo, I.L. and Ho, P.L., 2012. A
transcriptomic view of the proetome variability of newborn and adult
Bothrops jararaca snake venoms. PLoS Negl. Trop. Dis. 6.
213. Zhang, W., Liu, Y., Hou, B., Gu, X. and Ma, Z., 2015a. Activation
of spinal alpha-7 nicotinic acetylcholine receptor attenuates
remifentanil-induced postoperative hyperalgesia. Int. J. Clin. Exp. Med.
8, 1871–1879.
214. Zhang, Y.Y., Huang, Y., He, Q.Z., Luo, J., Zhu, L., Lu, S.S., Liu,
J.Y., Huang, P.F., Zeng, X.Z. and Liang, S.P., 2015b. Structural and
functional diversity of peptide toxins from tarantula Haplopelma
hainanum (Ornithoctonus hainana) venom revealed by transcriptomic,
peptidomic and patch-clamp approaches. J. Biol. Chem. 290, 14192–
14207.
215. Zheng, X.C., Li, W.X., Zhu, S.Y., Peng, F., Zhu, Z.H., Wu, K.L.
and Yiang, F.H., 2000. Cloning and characterization of a cDNA

124
sequence encoding the precursor of a chlorotoxin-like peptide from the
Chinese scorpion Buthus martensii Karsch. Toxicon 38, 1009–1014.
216. Zhou, P.A., Xie, X.J., Li, M., Yang, D.M., Xie, Z.P., Zong, X. and
Liang, S.P., 1997. Blockade of neuromuscular transmission by
huwentoxin-I, purified from the venom of the chinese bird spider
Selenocosmia huwena. Toxicon 35, 39–45.
217. Zhou, Y., Nelson, M.E., Kuryatov, A., Choi, C., Cooper, J. and
Lindstrom, J., 2003. Human α4β2 acetylcholine receptors formed from
linked subunits. J. Neurosci. 23, 9004–9015.
218. Zhu, S., Peigneur, S., Gao, B., Lu, X., Cao, C. and Tytgat, J.,
2012. Evolutionary diversification of Mesobuthus α- scorpion toxins
affecting sodium channels. Mol. Cell Proteomics 11, 1–18.
219. Kularatne, S.A.M., Senanayake, N., 2014. Venomous snake
bites, scorpions, and spiders, in: Ferro, J.B.a.J.M. (Ed.), Handbook of
Clinical Neurology. Elsevier B.V., Sri Lanka.

125
APPENDIX I Table I. List of spiders whose venoms were used in this work
Araneomorphae (“modern” spiders)
Abbreviations
No effect on α3* and α7 nAChR
Austrochiloidea family
1. Hickmania troglodytes H. troglodytes
Blocking effect on α3* nAChR
Araneoidea family
2. Cyrtophora moluccensis C. moluccensis
Blocking effect on α3* and α7
nAChR
Araneoidea family
3. Eriophora transmarina E. transmarina
Genus Nephila
4. Nephila pilipes N. pilipes
5. Nephila plumipes N. plumites
No effect on α3* and α7 nAChR
RTA Clade
Superfamily Ctenoidea
6. Ancylometes sp. Ancylometes sp.
7. Viridasius sp. (sylvestriformis) Viridasius sp.
Sparassidae family
8. Heteropoda jugulans H. jugulans
Partial blocking effect on α3* and α7 nAChR
RTA Clade
Superfamily Lycosoidea
9. Lycosidae sp. Lycosidae sp.
Sparassidae family
10. Sparassidae sp. 1 Sparassidae sp. 1
11. Sparassidae sp. 2 Sparassidae sp. 2

126
Blocking effect on α3* and α7
nAChR
RTA Clade
Pisauridae family
12. Megadolomedes M. australianus
Mygalomorphae (“primitive” spiders) Abbreviations
Blocking effect on neuronal α3*
and α7 nAChR
1. Macrothele gigas M. gigas
2. Acanthoscurria cf insubtilis A. insubtilis
3. Ceratogyrus bechunanicus C. bechunanicus
4. Ceratogyrus marshalli C. marshalli
5. Ceratogyrus ezendami C. ezendami
6. Eupalaestrus campestratus E. campestratus
7. Ephebopus uatuman E. uatuman
8. Chilobrachys sp. 2 Chilobrachys sp. 2
9. Grammostola porteri G. porteri
10. Grammostola rosea G. rosea
11. Grammostola sp. (Chilean
North) Grammostola sp. 1
12. Grammostola sp. (Concepcion,
Chile) Grammostola sp. 2
13. Grammostola sp. (Maule River,
Chile) Grammostola sp. 3
14. Holothele sp. (Carabobo,
Colombia) Holothele sp. 1
15. Holothele sp. (Peru) Holothele sp. 2
16. Idiothele nigrofulva I. nigrofulva
17. Lasiodorides striatus L. striatus
18. Lasiodora striatipes L. striatipes
19. Nhandu chromatus N. chromatus
20. Nhandu tripepii N. tripepii

127
21. Nhandu vulpinus N. vulpinus
22. Phormictopus cancerides P. cancerides
23. Pamphobeteus cf petersi Pamphobeteus cf petersi
24. Psalmopoeus cambridgei P. cambridgei
25. Psalmopoeus pulcher P. pulcher
26. Psalmopoeus langenbucheri P. langenbucheri
27. Theraphosinae sp. (Chile) Theraphosinae sp.
Potent blocking effect on α3*
nAChR
28. Avicularia metallica A. metallica
29. Avicularia ulrichea A. ulrichea
30. Avicularia sp. (Amazonas
Purple, Peru) Avicularia sp. 1
31. Coremiocnemis tropix C. tropix
32. Chilobrachys sp. (Penang,
Malaysia) Chilobrachys sp. 1
33. Euathlus sp. (green, chile) Euathlus sp. 1
34. Euathlus sp. (“red”=”fire”,
Chile) Euathlus sp. 2
35. Euathlus vulpinus (crème,
Chile highland) E. vulpinus
36. Grammostola grossa G. grossa
37. Grammostola iheringi G. iheringi
38. Orphnaecus philippinus O. philippinus
39. Orphnaecus sp. (‘treedweller’,
Paqnai-Island, Phillippines)
Orphnaecus sp.
40. Paraphysa scrofa P. scrofa
41. Pelinobius muticus P. muticus
42. Poecilotheria striatus P. striatus
43. Pterinochilus murinus P. murinus
44. Selenocosmia crassipes S. crassipes
45. Selenocosmia effera S. effera
46. Selenocosmia sp. (Papua Selenocosmia sp. 1

128
New-Guinea) 47. Selenocosmia sp. Selenocosmia sp. 2
48. Selenotholus foelschei S. foelschei
49. Thrixopelma cyaneolum
(PERU) T. cyaneolum
50. Thrixopelma sp. (Chile) Thrixopelma sp.
51. Thrixopelma lagunas sp.
(Peru) T. lagunas
Potential agonistic effect on α7 nAChR. Further analysis is required to
confirm the venom effect on subtype α7.
52. Haplopelma hainanum H. hainanum
53. Pamphobeteus sp.
(platyomma) Pamphobeteus sp.
54. Sericopelma rubronites S. rubronites
55. Xenesthis sp. (‘white’,
Colombia) Xenesthis sp.
No effect on α3* and α7 nAChR
56. Aphonopelma crinitum A. crinitum
57. Avicularia sp. (Rio Madre,
Bolivia) Avicularia sp. 2
58. Homoeomma sp. (blue) Homoeomma sp.
59. Pseudhapalopus sp.
(langhaarig) Pseudhapalopus sp.
Table II. List of scorpions whose venoms were used in this work
Abbreviations
Buthidae family
1. Androctonus australis A. australis
2. Androctonus crassicauda A. crassicauda
3. Leirurus quinquestriatus L. quinquestriatus
4. Grosphus grandidieri G. grandidieri
Scorpionidae family

129
Agonistic effect on α3* and α7
nAChR
1. Heterometrus spinnifer H. spinnifer