veränderungen der membranpermeabilität, des kohlenhydratgehaltes, des lipidgehaltes and der...
TRANSCRIPT

Institut fur Pflanzenpathologie und Pflanzenschutz der Universitat Gottingen, Bundesrepublik Deutschland
Veranderungen der Membranpermeabilitat, des Kohlenhydratgehaltes, des Lipidgehaltes und der Lipidzusammensetzung in Keimwurzelspitzen von Zea mays L. nach Behandlung mit Diclofop-methyl
Changes in Membrane Permeability, Carbohydrate Content, Lipid Content, and Lipid Composition in Root Tips from Zea mays after Treatment with Diclofop-methyl
H. H. HOPPE
Mit 4 Abbildungen
Eingegangen am 11. Juni 1980 . Angenommen am 19. August 1980
Summary
To get some information on the mode of action of diclofop-methyl (methyl 2-[4-(2',4'-dichlorophenoxy)phenoxyJpropanate) the effect of the herbicide on physiological activities and cell constituents of root tips isolated from untreated and herbicide treated seedlings of Zea mays was studied. Experiments were carried Out with 3 concentrations of diclofop-methyl: 3 X 10-7 M, which was subletal allowing further root growth, 3 X 10-6 ]\'1 the minimum inhibition concentration, and 3 X 10-5 M. The herbicide had no effect on respiratory 02-uptake, protein and nucleic acid content. It caused an accumulation of alcohol soluble carbohydrates and an increase in leakage of amino compounds from treated root tips. The herbicide induced a reduction of phospholipid and trigyceride content and an
Diese Arbeit ist ein Teil der bei der Landwirtschaftlichen Fakultat der Georg-AugustUniversitat zu Gottingen vorgelegten Habilitationsschrift «Untersuchungen zum Wirkungsmechanismus herbizider Phenoxy-Phenoxy-Propionsaurederivate».
Abkiirzungen: P A = Phosphatidsaure (phosphatidic acid); PS = Phosphatidylserin (phosphatidylserine); PI Phospatidylinosit (phosphatidylinositol); PCH Phosphatidylcholin (phosphatidylcholine); PG = Phosphatidylglycerin (phosphatidylglycerol); PE Phosphatidylathanolamin (phosphatidylethanolamine); N-Ac-PE N-Acyl-Phosphatidylathanolamin (N-acyl-phosphatidylethanolamine); MGG Monogalactosyldiglycerid (monogalactosyl diglyceride); DPG = Diphosphatidylglycerin (diphosphatidylglycerol); 16: 0 = Palmitinsaure (palmitic acid); 18: 0 = Stearinsaure (stearic acid); 18: 1 = tJlsaure (oleic acid); 18: 2 = Linolsaure (linoleic acid); 18: 3 Linolensaure (linolenic acid); DC = Dunnschichtchromatographie; FGW = Frischgewicht; Pi = anorganisches Phosphat; TCA = Trichloressigsaure.
Z. P/lanzenphysiol. Bd. 100. S. 415-426. 1980.

416 H . H. HOPPE
increase in glycolipid content. The decrease of phospholipids was due mainly to phosphatidylethanolamine and phosphatidylcholine and could probably be the cause for the increase in membrane permeability. The accumulation of glycolipids could be explained by an increase in the content of sterol glycosides and twO other unknown glycolipids which contained no fatty acids. Diclofop-methyl caused a shift in percentual fatty acid distribution. Herbicide treated root tips showed compared with the controls a lower content of oleic and palmitic acid and a higher content of linolenic acid. This shift in fatty acids was observed in the total lipid extract as well as in several individual polar lipids. Effects on membrane permeability and on lipid content and lipid composition increased with increasing herbicide concentrations and increasing incubation times. They appeared early enough to be responsible for the inhibition of root growth observed after treatment of maize seedlings with diclofop-methyl.
Key words: diclofop-methyl, membrane permeability, carbohydrate content, lipid content, root tips, Zea mays.
Einleitung
Diclofop-methyl (2-[ 4-(2',4'-Dichlorphenoxy)-phenoxy] -propionsauremethyi-ester, Handelspraparat: Illoxan®) gehort zur Gruppe der selektiven, ausschlieBlich gegen Graser wirksamen herbiziden Di(phenoxy)propionsaurederivate, die von der Hoechst AG entwickelt und erstmals 1975 der Offentlichkeit vorgestellt wurden (LANGELUDDEKE et al., 1975; SCHWERDTLE et al., 1975). Das Herbizid fuhrt nach der Behandlung von Maiskeimlingen durch eine Hemmung der Zellteilung und Zellstreckung zu einer Reduzierung des Keimwurzelwachstums. Hohere Herbizidkonzentrationen (10-6 M) leiten an den Keimwurzeln eine Nekrose ein, die im Bereich der Teilungs- und Streckungszonen beginnt und sich dann bis zum Korn hin ausbreitet (HOPPE, 1980). Urn die Ursa chen dieser Effekte auf die Keimwurzeln von Zea mays zu erfassen und damit Anhaltspunkte fur den Wirkungsmechanismus des Herbizids zu bekommen, wurde in der vorliegenden Arbeit der EinfluB von Diclofop-methyl auf den respiratorischen O 2- Verbrauch, den Efflux von Aminoverbindungen, den Kohlenhydrat-, Protein-, Nukleinsaure- und Lipidgehalt der Keimwurzelspitzen von Zea mays iiberpriift. Die Versuche wurden mit 3 Konzencrationen von Diclofop-methyl durchgefiihrt, einer subletalen Konzentration (5 X 10-7 M), die noch ein weiteres Wurzelwachstum zulieg, und 2 hoheren Konzentrationen (5 X 10- 6 M und 5 X 10-5 M), die nach einer 12stiindigen Einwirkungsdauer das Wurzelwachstum vollig hemmten (HOPPE 1980).
Material und Methoden
Die Anzucht und Herbizidbehandlung der Maiskeimlinge wurden bereits beschrieben (HOPPE, 1980). Zu verschiedenen Terminen nach der Herbizidbehandlung der Maiskeimlinge wurden 1,5 cm lange Wurzelspitzen von den Keimwurzeln abgetrennt und fiir Respirationsmessungen verwendet oder auf verschiedene Inhaltsstoffe analysiert. Der O2- Verbrauch der Wurzelspitzen wurde polarographisch mit Hilfe einer «Clarb-Elektrode (Yellow Springs Instument Co., USA) verfolgt (LESSLER, 1972). Zur ErmittIung des Gehaltes an pufferloslichen Proteinen wurden die Wurzelspitzen mit Tris-HCl-Puffer (0,1 M, pH 7,5) ex-
Z. Pflanzenphysiol. Ed. 100. S. 415-426. 1980.

Veranderungen in Zea nach Diclofop-methyl-Behandlung 417
trahiert und die Proteine nach einer TCA-Fallung quantitativ bestimmt (LOWRY, 1951). Die Extraktion der N ukleinsauren erfolgte nach WILCOCKSON und HULL (1974), ihre quantitative Bestimmung uber die Ermittlung des Extinktionsverhaltnisses E260/E280 (PEACOCK und DINGMAN, 1967).
Zur Bestimmung des alkoholloslichen Gehaltes an reduzierenden Verbindungen und Gesamtkohlenhydraten wurden 50 Wurzelspitzen mit 60 ml 800f0igem Methanol extrahiert. Die quantitative Bestimmung der reduzierenden Verbindungen erfolgte nach MILLER (1959), die der Gesamtkohlenhydrate nach DUBOIS et aI. (1956).
Zur Bestimmung des Nettoeffluxes von Aminoverbindungen wurden 40 Wurzelspitzen unter sterilen Bedingungen 3, 6 oder 12 Std. lang in 10-4 M CaCl2-Losung oder den entsprechenden Herbizidlosungen bei 26 DC im Wasserbad geschuttelt. Nach dem Dekantieren der Losungen wurden die Wurzelspitzen erneut in 30 ml bidestilliertem H 20 geschuttelt, dem zur Bestimmung der gesamten diffundierbaren Aminoverbindungen 0,2 ml CHCl3 zugesetzt war. Diese Behandlung fi.ihrte zu einem raschen Austritt der noch im Gewebe vorhandenen Aminoverbindungen, der nach 3 Std. beendet war. Die Bestimmung des AminoN-Gehaltes erfolgte nach YEMM und COCKING (1955).
Zur Bestimmung des Lipidgehaltes und des Lipidspektrums wurden 2-4 g (FGW) Wurzelspitzen in Anlehnung an BLIGH und DYER (KATES, 1972) extrahiert. In Anteilen der Lipidextrakte wurden die Fettsaurezusammensetzung (HUSEK, 1969), sowie die Gehalte an lipidgebundenen Phosphaten (BARTLETT, 1969) und Kohlenhydraten (DUBOIS et aI., 1956) bestimmt. Die nach den Gesamtanalysen verbliebenen Lipidextrakte wurden eingeengt und fur die DC an Kieselgel-Fertigplatten (Kieselgel 60 F254 , Merck, Darmstadt) verwendet. Die Auftrennung der neutralen Lipide erfolgte eindimensional mit Hexan : Diathylather : Essigsaure (960f0ig) = 75 : 25 : 2 (KATES, 1972). Die polaren Lipide wurden zweidimensional unter Verwendung folgender Laufmittelsysteme aufgetrennt (HOPPE und HEITEFUSS, 1974 a):
I: CHCla : CH30H : NH40H (250f0ig) : H 20 = 82 : 38 : 3: 2 II: CHCl3 : CH30H : CH3-COOH (960f0ig) : H 20 = 85 : 12.5 : 12.5 : 3
Die Lipide wurden mit Joddampfen angefarbt oder nach dem Bespruhen mit 0,6 Ofo K2Cr04 in 30 % iger H 2S04 durch Erhitzen bei 120 DC sichtbar gemacht. Die Identifizierung der einzelnen Lipide erfolgte durch Vergleich der relativen Rf-Werte mit den Ergebnissen anderer Autoren, mit Spruhreagenzien fur Phosphatester, NH2-haltige Lipide, Cholinphosphatide und Sterine (KATES, 1972; HOPPE und HEITEFUSS, 1974 a) sowie mit Hilfe verschiedener Referenzsubstanzen (HOPPE und HEITEFUSS, 1974 a). Urn die auf diese Weise analytisch erfaEbaren Phospholipide (PCH, PE, PI, PG) quantitativ zu bestimmen, wurden pro Extrakt 3 Chromatogramme angefertigt, die Phospholipidflecke in Zentrifugenrohrchen gesammelt und in Gegenwart des Kieselgels Phosphatbestimmungen durchgefuhrt (HOPPE und HEITEFUSS, 1974 a). Bei den Versuchen, bei denen nach der DC das Fettsaurespektrum der Lipide aufgenommen wurde, wurden den Laufmitteln Butylhydroxytoluol als Antioxidans zugesetzt und die Lipide durch Bespruhen mit Dichlorfluorescein sichtbar gemacht. Die Lipide wurden in Gegenwart des Kieselgels nach der Methode von HUSEK (1969) umgeestert und gaschromatographisch nachgewiesen (HOPPE und HEITEFUSS, 1974 b).
Ergebnisse
1. O2- Verbrauch, Protein- und N uklcinsauregehalt unbehandelter und herbizidbehandelter Keimwurzelspitzen von Zea mays
Diclofop-methyl fiihrte beim Bezug auf das FGW der Wurzelspitzen innerhalb der ersten 40 Std. nach der Herbizidbehandlung auch bei Verwendung hoher Her-
Z. Pflanzenphysiol. Bd. 100. S. 415-426. 1980.
418 H. H. HOPPE
bizidkonzentrationen nicht zu eindeutigen Veri:inderungen des O2- Verbrauches, des Protein- oder des Nukleinsauregehahes. Es konnten daher keine Hinweise fur einen direkten Eingriff des Herbizids in diese Stoffwechselbereiche gefunden werden.
2. Kohlenhydratgehalt unbehandelter und herbizidbehandelter Keimwurzelspitzen von Zea mays
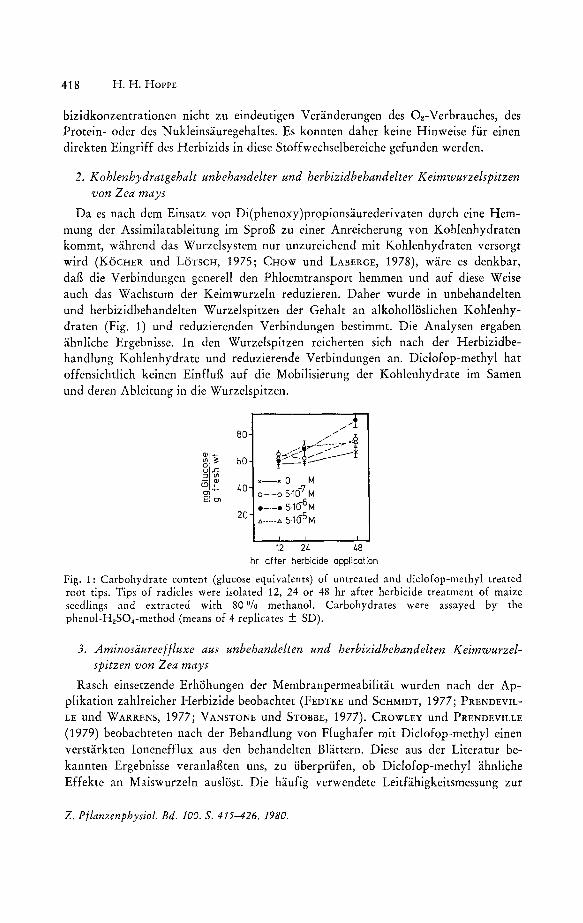
Da es nach dem Einsatz von Di(phenoxy)propionsaurederivaten durch eine Hemmung der Assimilatableitung im Sprog zu einer Anreicherung von Kohlenhydraten kommt, wahrend das Wurzel system nur unzureichend mit Kohlenhydraten versorgt wird (KOCHER und LaTSCH, 1975; CHOW und LABERGE, 1978), ware es denkbar, dag die Verbindungen generell den Phloemtransport hemmen und auf diese Weise auch das Wachstum der Keimwurzeln reduzieren. Daher wurde in unbehandelten und herbizidbehandelten Wurzelspitzen der Gehalt an alkoholloslichen Kohlenhydraten (Fig. 1) und reduzierenden Verbindungen bestimmt. Die Analysen ergaben ahnliche Ergebnisse. In den Wurzelspitzen reicherten sich nach der Herbizidbehandlung Kohlenhydrate und reduzierende Verbindungen an. Diclofop-methyl hat offensichtlich keinen Einflug auf die Mobilisierung der Kohlenhydrate im Samen und deren Ableitung in die Wurzelspitzen.
80
50
x-x 0 M 40 0--0 5107 M
• __ .5105M 20 6 ..... 65.105M
12 24 48
hr after herbicide application
Fig. 1: Carbohydrate content (glucose equivalents) of untreated and diclofop-methyl treated root tips. Tips of radicles were isolated 12, 24 or 48 hr after herbicide treatment of maize seedlings and extracted with 80 % methanol. Carbohydrates were assayed by the phenol-H2S04-method (means of 4 replicates ± SD).
3. Aminosaureeffluxe aus unbehandelten und herbizidbehandelten Keimwurzelspitzen von Zea mays
Rasch einsetzende Erhohungen der Membranpermeabilitat wurden nach der Applikation zahlreicher Herbizide beobachtet (FEDTKE und SCHMIDT, 1977; PRENDEVILLE und WARRENS, 1977 ; VANSTONE und STOBBE, 1977). CROWLEY und PRENDEVILLE (1979) beobachteten nach der Behandlung von Flughafer mit Diclofop-methyl einen verstarkten Ionenefflux aus den behandelten Blattero. Diese aus der Literatur bekannten Ergebnisse veranlagten uns, zu uberprufen, ob Diclofop-methyl ahnliche Effekte an Maiswurzeln auslOst. Die haufig verwendete Leitfahigkeitsmessung zur
z. P/lanzenphysiol. Ed. 100. S. 415-426. 1980.
Veranderungen in Zea nach Diclofop-methyl-Behandlung 419
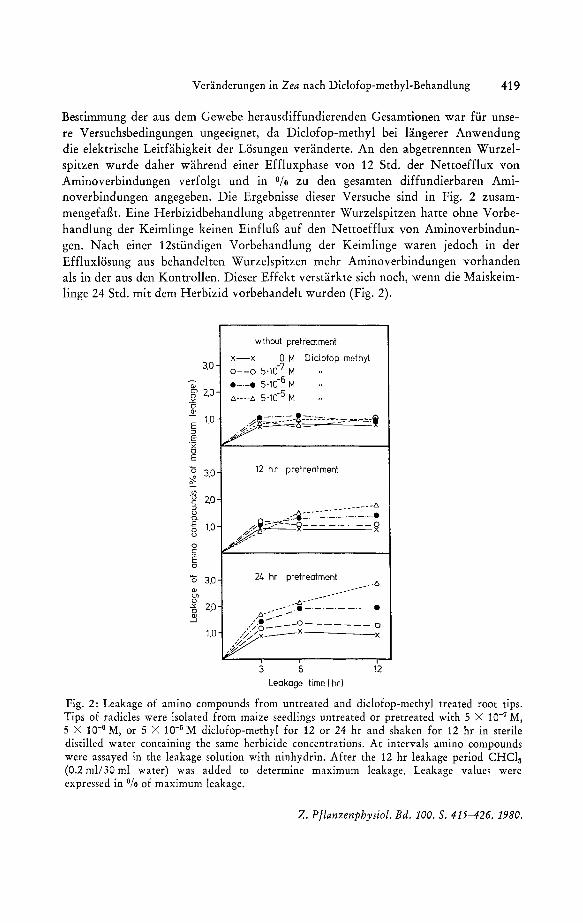
Bestimmung der aus dem Gewebe herausdiffundierenden Gesamtionen war fur unsere Versuchsbedingungen ungeeignet, da Diclofop-methyl bei langerer Anwendung die elektrische Leitfahigkeit der Losungen veranderte. An den abgetrennten Wurzelspitzen wurde daher wahrend einer Effluxphase von 12 Std. der Nettoefflux von Aminoverbindungen verfolgt und in Ofo zu den gesamten diffundierbaren Aminoverbindungen angegeben. Die Ergebnisse dieser Versuche sind in Fig. 2 zusammengefafh. Eine Herbizidbehandlung abgetrennter Wurzelspitzen hatte ohne Vorbehandlung der Keimlinge keinen Einflug auf den Nettoefflux von Aminoverbindungen. Nach einer 12stundigen Vorbehandlung der Keimlinge waren jedoch in der Effluxlosung aus behandelten Wurzelspitzen mehr Aminoverbindungen vorhanden als in der aus den Kontrollen. Dieser Effekt verstarkte sich noch, wenn die Maiskeimlinge 24 Std. mit dem Herbizid vorbehandelt wurden (Fig. 2).
3,0
OJ g> 2,0
"" o -"! E 1,0 ::> .~ x o E '0 'if. 3,0
(f)
-g 2.0 ::> o 0-
~ 1,0
o c E o
'0 3,0 QJ
Ol o -a 2,0 ~
1,0
without pretreatment
x-x 0 M Diclofop-methyl 0--0 5'10-7 M .--. 5,10-6 M 6----6 5,10-5 M
12 hr pretreatment
24 hr pretreatment
3 6
Leakage time ( hr)
12
Fig. 2: Leakage of amino compounds from untreated and diclofop-methyl treated root tips. Tips of radicles were isolated from maize seedlings untreated or pretreated with 5 X 10-7 M, 5 X 10-6 M, or 5 X 10-5 M diclofop-methyl for 12 or 24 hr and shaken for 12 hr in sterile distilled water containing the same herbicide concentrations. At intervals amino compounds were assayed in the leakage solution with ninhydrin. After the 12 hr leakage period CHCl3
(0.2 mli30 ml water) was added to determine maximum leakage. Leakage values were expressed in % of maximum leakage.
Z. PJlanzenphysiol. Bd. 100. S. 415-426. 1980.

420 H . H. HOPPE
Aufgrund der hier dargestellten Ergebnisse kann ausgeschlossen werden, daB Diclofop-methyl die biologischen Membranen zerstort. Diclofop-methyl verursachte zwar eine relativ fruh einsetzende Erhohung des Nettoeffluxes von Aminoverbindungen. Dieser Effekt war aber quantitativ zu gering, urn als alleinige Ursache der herbiziden Wirkung angesehen werden zu konnen: Unter unseren Bedingungen traten nach der Herbizidbehandlung hochstens 3 Ofo der diffundierbaren Aminoverbindungen aus dem Gewebe aus.
4. Phospholipide und Glykolipide unbehandelter und herbizidbehandelter Keimwurzelspitzen von Zea Mays
Urn den Gehalt an lipidgebundenem Phosphat und lipidgebundenen Kohlennydraten in den Wurzelspitzen von Zea mays zu ermitteln, wurden in Teilen der Lipidextrakte Phosphat- oder Kohlenhydratbestimmungen durchgefiihrt. Bei Bezug auf 1 g FGW war der Gesamtphospholipidgehalt der herbizidbehandelten Wurzelspitzen geringer als der der unbehandelten, wahrend der Gehalt an lipidgebundenen Kohlenhydraten durch die Herbizidbehandlung erhoht wurde. (Fig. 3). Beide Effekte deuteten sich bereits 12 Std. nach der Herbizidbehandlung an und verstarkten sich mit zunehmender Einwirkungsdauer von Diclofop-methyl.
0.50
°1_ 0.1.5 <.9;:
~~ 0.30 ""w :.::il.::
0l0l EI 0.15
100
or 75 I ;:
~L ""III :.::il~ 50 Ol :::1\.0>
25
.. ··A ~>b;,;.:J~9 .~~--l 1
.-xO
0--0 5'10-7 M .--.5·105 M 6 ...... 65·10-5 M
I
.€~::-_+ ___ --i .~",:::.~~:::----- §
~-'~':::;:::t x-x 0 M
0--05-10-7 M .--.510-5 M 6 ...... 65-10-5 M
5 12 21.. 48 hr after herbicide application
Fig. 3: Content of lipidbound carbohydrates and phosphate in untreated and diclofop-methyl treated root tips. Tips of radicles were isolated 6, t 2, 24, or 48 hr after herbicide treatment of maize seedlings and extracted for lipids. Lipidbound phosphate was assayed in the total lipid extract by determination of Pi after acid digestion. Lipidbound carbohydrates were assayed by the phenol-H2S04-method and expressed in galactose equivalents (means of 3 replicates ± SD).
Z. Pjlanzenphysiol. Bd. 100. S. 415-426. 1980.

Veranderungen in Zea nach Diclofop-methyl-Behandlung 421
A
14 17
10 13 12 16
11 19
6 9
<..../ 20
3 ,..-4
<::::) 2 5
1
B
Fig. 4: Thin-layer chromatogramm of polar lipids from untreated (A) and diclofop-methyl treated (B, 5 X 10-6 M) root tips. Lipids were extracted 24 hr after herbicide treatment on the basis of equal fresh wt and visualized by charring at 120 °C after spraying with 0.6010 K2Cr04 in 30010 H 2S04 • 1: start; 3: PI; 4: Ps; 5: PA; 6: PCH; 7: PE; 11: PG; 15: sterol glycosides; 16: DPG; 17: MGG+N-Ac-PE; 18: neutral lipids; 19: freee fatty acids; 2, 8,9, 10, 12, 13, 14, 20: not identified .

422 H. H. HOPPE
Die Abnahme des Phospholipidgehaltes wurde auch bei visueller Beurteilung der Diinnschichtchromatogramme deutlich (Fig. 4). Urn den Effekt quantitativ genauer erfassen zu konnen, wurden nach der diinnschichtchromatographischen Trennung die analytisch erfaBbaren Phospholipidflecke (PCH, PE, PI, PG) von den Chromatogrammen abgekratzt und in Gegenwart des Kieselgels Phosphatbestimmungen durchgefiihrt. Vierundzwanzig Std. nach der Herbizidbehandlung war in den behandelten Wurzel spitz en vor allem der PCH- und PE-Gehalt vermindert, wahrend der Gehalt des PG und PI nicht abgenommen hatte (Tab. 1). Den Analysen des Gesamtgehaltes entsprechend war die Abnahme des PCH- und PE-Gehaltes bereits 12 Std. nach der Herbizidbehandlung zu erkennen. Die Reduzierung des Gesamtphospholipidgehaltes konnte damit in erster Linie auf das PCH und PE zuriickgefiihrt werden.
Table 1: Phospholipid content ( !.I g lipid-P /g fresh wt) of untreated and herbicide treated root tips. Tips of radicles were isolated 12 or 24 hr after herbicide treatment of the seedlings and extracted for lipids. Phospholipids were isolated by TLC and assayed by determination of PI after acid digestion.
12 hr after treatment 24 hr after treatment
untreated 5XI0-6M untreated 5x 10-6M diclofop-methyl diclofop-methyl
PCH 24 15 21 12 PE 22 16 19 9 PI 10 10 7 7 PG 6 6 4 3
Die Zunahme des Gehaltes an lipidgebundenen Kohlenhydraten im herbizidbehandelten Gewebe konnte auf Grund visueller Beurteilung der diinnschichtchromatographischen Trennungen auf die Steringlycoside sowie auf die Verbindungen 9 und 10 zuriickgefiihrt werden (Fig. 4). Die von uns nicht identifizierten Verbindungen 9 und 10 wurden im Gegensatz zu den iibrigen polaren Lipiden unter der UV -Lampe sehr deutlich als dunkle Flecke vor griinlich fluoreszierendem Hintergrund sichtbar. Die Verbindungen enthielten keine Fettsauren und kein Phosphat und waren auch nicht mit 32Pi, 14C-Acetat, l4C-tllsaure oder l4C-Malonsaure zu markieren. Nur bei Verwendung von l4C-Glucose als Vorstufe wurde ein hoher Anteil der 14C-Aktivitat in die Verbindungen 9 und 10 eingebaut (HOPPE, unveroffentlicht). MOROHASHI und BANDURSKI (1976) isolierten aus Maisembryonen 2 polare Glykolipide, die nach den Steringlykosiden am starksten mit l4C-Glucose markiert wurden und kein Phosphat enthielten. Die Autoren vermuten, daB die Verbindungen in ahnlicher Weise wie glykosylierte Phosphoprenole als Obertrager von Zukkerresten an der Biosynthese von Polysacchariden und Zellwandmaterial beteiligt sein konnten. Die Glykolipide konnten auf Grund ihres Verhaltens bei der Chromatographie mit den von uns nachgewiesenen Verbindungen 9 und 10 identisch sem.
Z. P/lanzenphysiol. Bd. 100. S. 415-426. 1980.

Veranderungen in Zea nach Diclofop-methyl-Behandlung 423
5. Neutrallipide unbehandelter und herbizidbehandelter Keimwurzelspitzen von Zea mays
Das dunnschichtchromatographisch ermittelte Spektrum der neutralen Lipide der Wurzelspitzen von Zea mays entsprach groihenteils den Angaben in der Literatur (HITCHCOCK und NICHOLS, 1971). Sterine und Sterinester waren mengenma~ig am starksten in der Neutrallipidfraktion vertreten. Daneben wurden Triglyceride, Diglyceride, C-Methyl- und C4-Dimethylsterine nachgewiesen. Die Herbizidbehandlung fuhrte zu einer Abnahme des Trigyceridgehaltes und in einigen Fallen auch zu einer Abnahme des Diglyceridgehaltes. Die Abnahme des Diglyceridgehaltes konnte nicht in allen Fallen nachgewiesen werden, weil bereits das unbehandelte Gewebe einen sehr geringen Diglyceridgehalt aufwies.
6. Fettsaurespektrum unbehandelter und herbizidbehandelter Keimwurzelspitzen von Zea mays
In Tabelle 2 ist der prozentuale Anteil der einzelnen Fettsaurekomponenten der Lipidextrakte aus unbehandelten und behandelten Wurzelspitzen dargestellt. In den Lipidextrakten wurden Palmitinsaure (16 : 0), Stearinsaure (18 : 0), Olsaure (18 : 1), Linolsaure (18 : 2) und Linolensaure (18 : 3) nachgewiesen. Die Linolsaure war mit ca. 80 % aller Fettsaurekomponenten am starksten vertreten. Ahnliche Fettsaurezusammensetzungen wurden auch von anderen Autoren in den Lipiden aus Wurzelmaterial von Mais (GIBRAT & ROSSIGNOL, 1977) und Spinat (SUKHIJA et al., 1976) sowie aus Kartoffelknollen (GALLIARD, 1968) festgestellt, wahrend in den Lipiden der Wurzeln wei~er Ruben (Brassica campestris var. rapa) vor allem Linolensaure
Table 2: Percentual distribution of fatty acids in lipid extracts from untreated and herbicide treated root tips. Tips ofradicles were isolated 12, 24 or 48 hr after herbicide treatment of the seedlings and extracted for lipids. Fatty acids were analysed as methyl esters by gas liquid chromatography.
hr after treatment Diclofop-methyl 16:0 18:0 18:1 18:2 18:3
12 untreated 8.55 1.49 2.96 80.53 6.50 5 x 10-' M 9.56 1.54 1.46 80.44 7.06 5x 10-6M 8.63 1.44 0.83 81.19 7.94 5x 10-5M 8.58 1.73 0.92 80.67 8.01
24 untreated 9.04 1.67 3.72 79.91 5.71 5 x IO-'M 8.68 1.66 2.09 81.31 6.26 5 x 10-6 M 8.17 1.69 1.15 79.36 9.69 5 x 10-5 M 7.41 1.99 1.09 78.68 10.83
48 untreated 11.52 1.43 3.66 78.41 4.99 5 x 10-7 M 8.70 1.72 1.82 76.90 10.86 5XIO-6M 8.58 2.06 1.02 76.78 11.56 5x 10-5M 7.61 1.98 0.94 77.82 11.70
Z. PJlanzenphysiol. Bd. 100. S. 415-426. 1980.

424 H. H. HOPPE
vorkam (LEPAGE, 1967). Die Herbizidbehandlung fuhrte zu deutlichen Verschiebungen des Fettsaurespektrums der Lipidextrakte. Besonders auffallig war die Abnahme des Olsaureanteiles in den herbizidbehandelten Wurzelspitzen (Tab. 2). Dieser Effekt war auch bei der subletalen Herbizidkonzentration (5 X 10-7 M) bereits 12 Std. nach der Behandlung deutlich sichtbar, zu einem Zeitpunkt, bei dem die Hemmung des Wurzelwachstums gerade einsetzte (HOPPE, 1980). Neben der Abnahme des Anteiles der Olsaure wurde eine Zunahme des Linolensaureanteiles und etwas spater (24 und 48 Std. nach der Behandlung) auch eine Abnahme des Palmitinsaureanteiles beobachtet.
Urn zu klaren, ob die Verschiebung des Fettsaurespektrums generell bei allen Lipiden auftrat oder ob davon vor allem die Lipide betroffen waren, deren Gehalt reduziert wurde, wurde das Fettsaurespektrum der starker konzentrierten Lipide PCH, PE, PI und MGG+ N-Ac-PE aufgenommen. Das Fettsaurespektrum der einzelnen Lipide war sehr ahnlich (Tab. 3) und entsprach dem des Gesamtlipidextraktes (Tab. 2). Von der herbizidinduzierten Verschiebung des Fettsaurespektrums waren auch die Lipide betroffen, deren Gehalt durch die Herbizidbehandlung nicht verandert wurde.
Table 3: Percentual distribution of fatty acids in individual lipids from untreated and herbicide treated root tips. Tips of radicles were isolated 24 hr after herbicide treatment of maize seedlings and extracted for lipids. Lipids were isolated by TLC and fatty acids were analysed by gas liquid chromatography.
Lipid Diclofop-methyl 16:0 18:0 18: 1 18:2 18: 3
PCH untreated 7.58 0.58 4.97 80.60 6.29 5x 10-6M 6.85 0.69 1.06 79.79 11.57
PE untreated 10.31 0.65 2.17 84.23 2.69 5x10-6M 10.38 0.63 0.77 83.68 4.63
PI untreated 17.78 1.02 5.30 70.79 5.10 5 X 10-6 M 16.87 1.76 1.17 71.75 8.50
MGG+ untreated 7.14 0.46 3.92 81.91 5.56 N-Ac-PE 5 x 10-6 M 7.15 0.68 0.70 82.05 9.46
Diskussion
Bei den in dieser Arbeit beschriebenen Versuchen war eine unterschiedliche Beeinflussung des Protein- und Nukleinsauregehaltes sowie des Lipidgehaltes und des Fettsaurespektrums durch das Herbizid zu erkennen. Im Gegensatz zum Proteinund Nukleinsauregehalt war der Phospholipidgehalt der herbizidbehandelten Wurzelspitzen bereits 12 Std. nach der Herbizidbehandlung reduziert, wenn die Analysendaten auf das FGW bezogen wurden (Fig. 3, 4; Tab. 1). Phospholipide sind fur die diffusionshemmende Wirkung biologischer Membranen unerlaGlich, und eine Abnahme des Phospholipidgehaltes hat in der Regel eine Abschwachung dieser Wir-
z. P/lanzenphysiol. Bd. 100. S. 415-426. 1980.

Veranderungen in Zea nach Diclofop-methyl-Behandlung 425
kung der Membranen zur Folge (SIMON, 1974). Die Abnahme des Phospholipidgehaltes kann daher als Ursache fUr den erhohten Efflux von Aminoverbindungen aus den herbizidbehandelten Wurzelspitzen angesehen werden, denn dieser Effekt trat ebenfalls 12 Std. nach der Herbizidbehandlung in Erscheinung (Fig. 2).
Wahrend die Behandlung der Maiskeimlinge mit Diclofop-methyl zu einer Abnahme des Phospholipidgehaltes fiihrte, reicherten sich Glykolipide in den herbizidbehandelten Wurzelspitzen an (Fig. 3, 4). Diese Veranderung konnte in Beziehung stehen zur Anreicherung von alkoholloslichen Kohlenhydraten in herbizidbehandelten Wurzelspitzen (Fig. 1) und Sproiheilen (KOCHER und LOTZSCH, 1975; CHOW und LABERGE, 1978) sowie zur erhohten Absonderung kohlenhydrathaltiger Schleime durch herbizidbehandelte Keimwurze!n von Zea mays (HOPPE, 1980).
Zeitlich parallel mit den Veranderungen im Phospholipid- und Glykolipidgehalt verlief die Verschiebung des Fettsaurespektrums. Die Abnahme des Anteiles der 01-saure- und Palmitinsaure sowie die Anreicherung von Linolensaure in den herbizidbehandelten Wurzelspitzen konnte darauf hindeuten, dag das Herbizid die Neusynthese von Fettsauren reduziert, wahrend die Verlangerung des Palmityl-ACP und die Desaturierung des Oleyl-ACP unbeeinflugt bleibel1 (STUMPF, 1977).
Die Veranderul1gen im Phospholipid- und Glykolipidgehalt und die Verschiebungen des Fettsaurespektrums verstarkten sich mit zunehmender Herbizidkonzentration und zunehmender Einwirkungsdauer von Diclofop-methyl. Sie traten augerdem friih genug auf, urn als Ursache fiir die Wachstumshemmung der Keimwurzeln in Frage zu kommen, denn auch der Einflug des Herbizids auf das Wurzelwachstum setzte 12 Std. nach der Herbizidbehandlung ein (HOPPE, 1980). Diclofop-methyl ware damit den Verbindungen zuzurechnen, die iiber einen Eingriff in den Lipidstoffwechsel herbizid wirksam werden (RIVERA und PENNER, 1979). In weiterfiihrenden Versuchen soli versucht werden, den Eingriff von Diclofop-methyl in den Lipidstoffwechsel genauer zu charakterisieren und zu iiberpriifen, ob es sich beim Einflug von Diclofop-methyl auf den Lipidstoffwechsel urn eine direkte oder indirekte Wirkung des Herbizids handelt.
Herrn Prof. Dr. R. HEITEFUSS danke ich flir die kritische Durchsicht des Manuskriptes und Frau S. GORLT flir die sorgfaltige Mitarbeit bei der Durchflihrung der Versuche. Der Hoechst AG danke ich flir die Dberlassung des reinen Wirkstoffes Diclofop-methyl.
Literatur
BARTLETT, G. R.: J. BioI. Chern. 234, 466 (1959). CHOW, P. N. P. and D. E. LABERGE: J. Agric. Food Chern. 26,1134 (1978). CROWLEY, J. and G. N. PRENDEVILLE: Can. J. Plant. Sci. 59, 275 (1979). DUBOIS, M., K. A. GILLES, J. K. HAMILTON, P. A. REBERS, and F. SMITH: Anal. Chern. 28,
350 (1956). FEDTKE, C. and R. R. SCHMIDT: Weed Res. 17, 233 (1977). GALLIARD, T.: Phytochem. 7, 1907 (1968). GIBRAT, R. and M. ROSSIGNOL: C. R. Acad. Sc. Paris, 285,1171 (1977). HITCHCOCK, C. and B. W. NICHOLS: Plant lipid biochemistry, Academic Press, London,
1971.
Z. PJlanzenphysiol. Bd. 100. S. 415-426. 1980.

426 H. H. HOPPE
HOPPE, H. H.: Weed Res. im Druck (1981). HOPPE, H. H. and R. HElTEFUSS: Physio!. Plant Patho!' 4,11 (1974 a). - - Physio!. Plant Patho!' 4, 25 (1974 b). HUSEK, P.: Z. klin. Chern. u. klin. Biochem. 6, 627 (1968). KATES, M.: Techniques of lipidology, North-Holland Publishing Company, Amsterdam,
London, 1972. KOCHER, H. und K. LOTZSCH: Proc. Europ. Weed Res. Soc. Symp. 430 (1975). LANGELUDDEKE, P., W. BECKER, H. H. NESTLER und F. SCHWERDTLE: Mitt. der Bio!. Bun-
desanst. 165, 169 (1975). LEPAGE, M.: Lipids 2, 244 (1967). LESSLER, M. A.: Methods in Cell Physio!. 5,199 (1972). LOWRY, O. H., N. J. ROSENBROUGH, A. L. FARR, and R. J. RANDALL: J. Bio!. Chern. 193,
265 (1951). MILLER, G. L.: Ana!. Chern. 31, 426 (1959). MOROHASHl, Y. and R. S. BANDURSKI: Plant Physio!. 57, 846 (1976). PEACOCK, A. G. and C. W. DINGMAN: Biochemistry 6,1818 (1967). PRENDEVILLE, G. N. and G. F. WARRENS: Weed Res. 30, 251 (1977). RIVERA, C. M. and D. PENNER: Res. Rev. 70,45 (1979). SCHWERDTLE, F., H. LEDITSCHKE, H. SCHONOWSKY und P. LANGE-LuDDEKE: Mitt. Bio!.
Bundesanst. 165, 171 (1975). SIMON, E. W.: New Phyto!' 73, 377 (1974). STUMPF, P. K.: Lipid biosynthesis in developing seeds. In: M. TEVINI and H. K. LICHTEN
THALER (Eds.): Lipids and lipid polymers in higher plants, 75-84. Springer-Verlag, Berlin, Heidelberg, New York, 1977.
SUKHlJA, P. S., J. S. SITAL, R. K. RAHEJA and 1. S. BHATIA: Physio!. Plant. 38, 221 (1976). VANSTONE, D. E. and E. H. STOBBE: Weed Sci. 25, 352 (1977). WILCOCKSON, J. and R. HULL: J. Gen. Viro!' 23, 107 (1974). YEMM, E. W. and E. C. COCKING: The Analyst 80, 209 (1955).
H. H. HOPPE, Institut fur Pflanzenpathologie und Pflanzenschutz, Universirat Gottingen, Grisebachstr. 6, D-3400 Gottingen.
z. PJlanzenphysiol. Bd. 100. S. 415-426. 1980.