“zellbiologieund physiologieder pflanzen” vorlesungsose2016 · 3 Ölkörperreifen am er...
TRANSCRIPT

1
“Zellbiologie und Physiologie der Pflanzen” Vorlesung SoSe 2016
Prof. Dr. Jörg Kudla (u. Mitarbeiter)
Prof. Dr. Antje von Schaewen
Prof. Iris Finkemeier
www.uni-muenster.de/Biologie.IBBP/agschaewen/lehre/index.html
VL-Folien unter: Nutzer: kurs
Kennwort: olympia
25.04.2016 Das pflanzliche Endomembran-System (und zelluläre Transportprozesse)
Für das Feld der Beobachtung gilt:Der Zufall
bevorzugt den vorbereiteten Geist
Louis Pasteur, Université de Lille, Decembre 7, 1854
Dans les champs de l‘ observationle hasard ne favorise que les esprits préparés

2
• Endoplasmatisches Retikulum (ER)
• Transportvesikel (mit verschiedenen Hüllproteinen), Coated Vesicles
• Golgi-Apparat (Golgi)
• trans-Golgi-Netzwerk (TGN) & Early
Endosome (EE)
• Plasmamembran (PM)
• Zellwand & Apoplast (extra-cellular
space)
• Late Endosome (PVC = Pre-Vacuolar
Compartment)
• Tonoplast (vakuoläre Membran)
• Vakuole (Zentralvakuole)
Das sekretorische System von Pflanzen
ER
Golgi-Apparat
Zellwand
Plasma-membran
Vakuole
Endosom (PVC)
Cytosol
Tonoplast
Kern
TGN
Vorlesung 2016
Abb.: aus Buchanan et al. (2000) „Biochemistry & Molecular Biology of Plants“ (Amer. Soc. of Plant Physiologists).
Transport-vesikel
Transport-vesikel
cis
medial
trans
Plasmodesmen verbinden Pflanzenzellen untereinander zu einem riesigen SYMPLASTEN
Vorlesung 2016
Abb. aus Buchanan et al. (2000) „Biochemistry & Molecular Biology ofPlants“ (Amer. Soc. ofPlant Physiologists)
• ER-Verbindung zur Kern-Hülle ⇒ „ER-to-nucleus
signaling“ (z.B. bei Stress)
• Rough ER (mit Ribosomen) Protein-Import und cotrans-lationale N-Glykosylierung
• ER-Lumen ⇒ Protein-faltung, Modifikation, Kontrolle
• Speichervesikel für Neutral-fette (Oil body), od. unlösliche Proteine (Protein body)
• Organell-Verankerung am Cytoskelet (Actin-binding
domain) ⇒ Gravitropismus!
• Kommunikation (inter-
zellulär) via Plasmodesmata
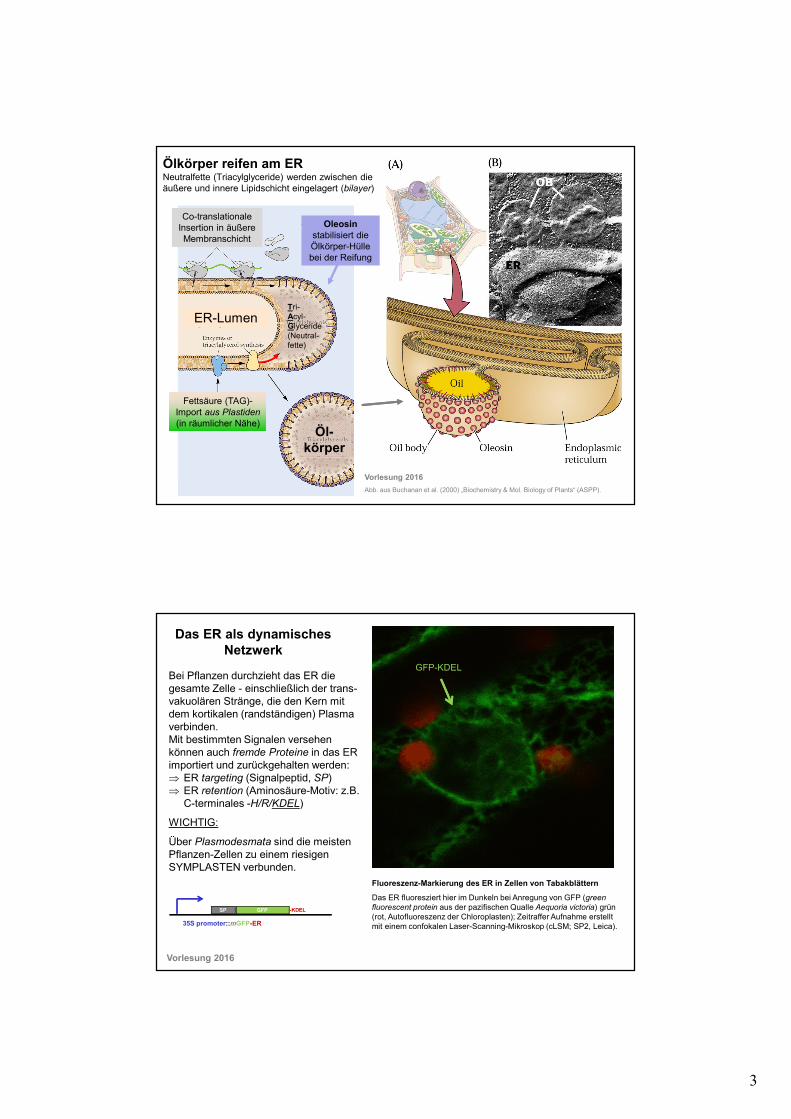
3
Ölkörper reifen am ER Neutralfette (Triacylglyceride) werden zwischen die äußere und innere Lipidschicht eingelagert (bilayer)
ER-LumenTri-Acyl-Glyceride(Neutral-fette)
Öl-körper
Fettsäure (TAG)-Import aus Plastiden
(in räumlicher Nähe)
Co-translationaleInsertion in äußere Membranschicht
Vorlesung 2016
Abb. aus Buchanan et al. (2000) „Biochemistry & Mol. Biology of Plants“ (ASPP).
Oleosinstabilisiert die Ölkörper-Hülle bei der Reifung
Bei Pflanzen durchzieht das ER die gesamte Zelle - einschließlich der trans-vakuolären Stränge, die den Kern mit dem kortikalen (randständigen) Plasma verbinden.Mit bestimmten Signalen versehen können auch fremde Proteine in das ER importiert und zurückgehalten werden: ⇒ ER targeting (Signalpeptid, SP)⇒ ER retention (Aminosäure-Motiv: z.B.
C-terminales -H/R/KDEL)
WICHTIG:
Über Plasmodesmata sind die meisten Pflanzen-Zellen zu einem riesigen SYMPLASTEN verbunden.
Fluoreszenz-Markierung des ER in Zellen von Tabakblättern
Das ER fluoresziert hier im Dunkeln bei Anregung von GFP (green
fluorescent protein aus der pazifischen Qualle Aequoria victoria) grün (rot, Autofluoreszenz der Chloroplasten); Zeitraffer Aufnahme erstellt mit einem confokalen Laser-Scanning-Mikroskop (cLSM; SP2, Leica).
Das ER als dynamisches Netzwerk
Vorlesung 2016
SP -KDELGFP
35S promoter::mGFP-ER
GFP-KDEL

4
Vorlesung 2016
Abb. aus Buchanan et al. (2000) „Biochemistry & MolecularBiology of Plants“ (ASPP)
Das sekretorische System von Pflanzen
rough ER
Cytosol
ER-Lumen
Protein-Import und -FaltungDie Interaktion N-terminaler Signalpeptide mit dem SRP (signal recogniton particle - als quasi zusätzliche ribosomaleUntereinheit) bewirkt kurzfristig ein Anhalten der Translation.
• SRP-Rezeptor: Rezeptor in der ER-Membran, an den das arretierte Ribosom bindet (⇒ docking protein)
• SRP release hebt den Translationsarrest auf. Das Signalpeptid wird im ER-Lumen von einer Signal-Peptidaseabgespalten. Danach erfolgen post-translationaleModifikationen und Chaperon-gestützte Proteinfaltung.
• Sec61 = Translokationskomplex (wassergefüllte „Pore“ = Membrankanal)
Signal-peptid
SRP = Signal Recognition Particle
gefaltetes Protein
„docking“
Membran-verankerte Proteine und ihre Topologie
Typ I Typ II Typ III
Typ IV Typ V Typ VI
Sanger-Modell, modifiziert nach: Howell & Crine (1996). Trends in Biochemical Sciences 21: 171.
C„innen“Cytosol
Lumen„außen“
N
Lumen„außen“
GPI-Anker
N
Spaltstelle
„innen“Cytosol
N C
N
N
Apoplast/
Zellwand
bzw.
Organell-
Lumen
Anheften im ER
Abspalten im
Apoplasten
C
CCytosol
Signal-
Anker

5
http://www.sigmaaldrich.com/etc/medialib/life-science/biochemicals/migrationbiochemicals1/GPI-Anchor.Par.0001.Image.580.gif
GPI-Anker (Detail)
Glycosyl-phosphatidyl-inositol-Anker, kurz: GPI-Anker
Protein
Im ER-Lumen können Zuckerbäumchen an bestimmte Asparagine (AsN, N) in der Polypeptidkette angeheftet werden. Dies erfolgt i.d.R. co-translational. Konsensus: AsN-X[Pro]-Ser/Thr (N-Glykane).
N-Glykane werden an der ER-Membran synthetisiert (Trägerlipid: Dolichol-Pyrophosphat) und im ER-Lumen mittels Oligosaccharyltransferase (OST) ‘en bloc‘ auf die wachsende Polypeptidkette transferiert.
Manche sekretierten Proteine werden im ER-Lumen glykosyliert
Vorlesung 2016
Abb. aus Buchanan et al. (2000) „Biochemistry & Molecular Biology of Plants“ (ASPP).
… AsN-X-Ser/Thr …
OSTOST =Oligo-saccharyl-Transferase

6
Synthese von N-Glykan-Vorstufen am „rauen“ ER
PPP
P
P
P
GDP
GDP
P P
PP
PP
PP
PP
P
UDP
mRNA
PP
UMP+UDP
GDP
GDP
GDPGDP
UDPUDP
PP
PP
N
Asn
Modifiziert nach: Abeijon & Hirschberg (1992). TIBS Vol. 17
Dolichol-Panchor
flip-flop
OST
UDP
GlcNAcManGlc
trapped
inside
activation
GPT =UDP-N-acetyl-glucosamine-dolichyl-phosphate: N-acetyl-glucosamine-1-phosphate transferase
Protein-Prozessierung, -Faltung und -Assemblierung
Das ER überwacht die Proteinreifung:
Prozessierung (Signalpeptidase), Faltung (Disulfidbrücken-Bildung etc.), und den GPI-Anker Transfer. Dabei sind verschiedene Helferproteine involviert (Chaperon-Komplexe).
Erst danach gelangen die Proteine zum Golgi-Apparat (per Vesikel-Transport), von wo aus sie auf weitere Zielkompartimente verteilt werden (Plasmamembran/Apoplast-Zellwand, Tonoplast/Vakuole).
Vorlesung 2016
Abb. Buchanan et al. (2000) „Biochemistry & Molecular Biology of Plants“ (Amer. Soc. of Plant Physiologists).
Proteine, die nicht korrekt gefaltet sind müssen abgebaut werden!
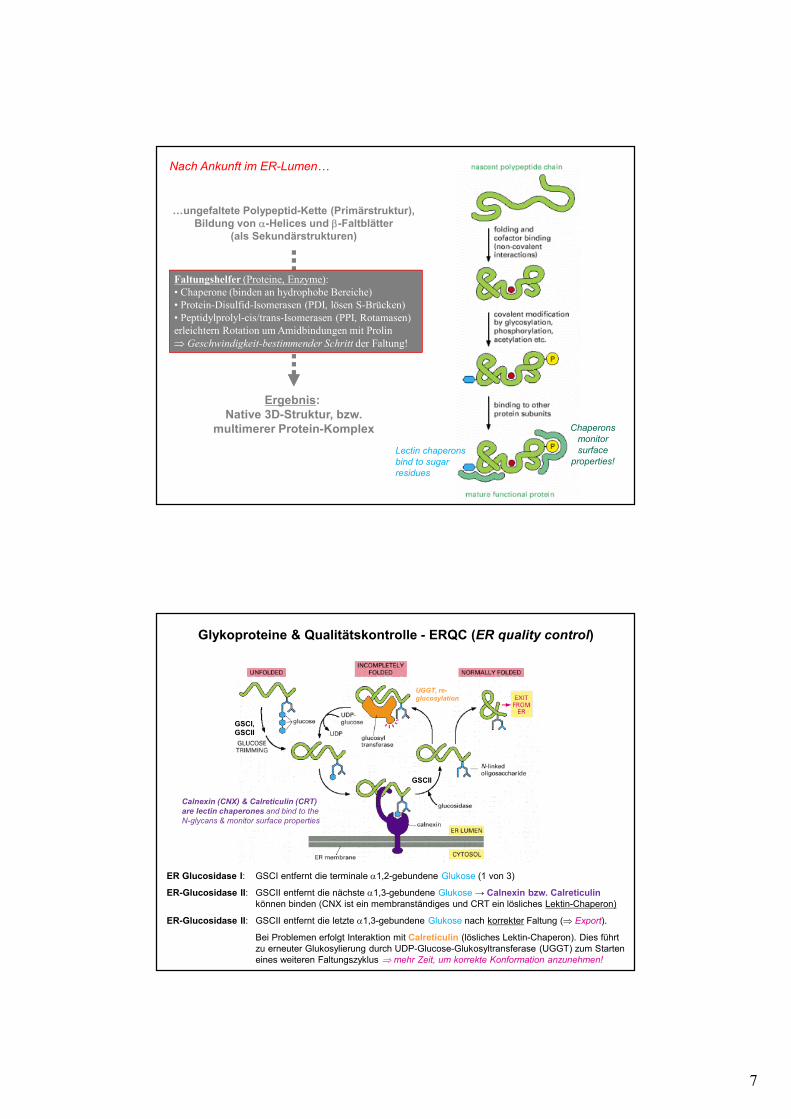
7
…ungefaltete Polypeptid-Kette (Primärstruktur),Bildung von α-Helices und β-Faltblätter
(als Sekundärstrukturen)
Ergebnis: Native 3D-Struktur, bzw.
multimerer Protein-Komplex
Nach Ankunft im ER-Lumen…
Faltungshelfer (Proteine, Enzyme):
• Chaperone (binden an hydrophobe Bereiche)
• Protein-Disulfid-Isomerasen (PDI, lösen S-Brücken)
• Peptidylprolyl-cis/trans-Isomerasen (PPI, Rotamasen)
erleichtern Rotation um Amidbindungen mit Prolin
⇒ Geschwindigkeit-bestimmender Schritt der Faltung!
Faltungshelfer (Proteine, Enzyme):
• Chaperone (binden an hydrophobe Bereiche)
• Protein-Disulfid-Isomerasen (PDI, lösen S-Brücken)
• Peptidylprolyl-cis/trans-Isomerasen (PPI, Rotamasen)
erleichtern Rotation um Amidbindungen mit Prolin
⇒ Geschwindigkeit-bestimmender Schritt der Faltung!
Chaperons
monitor
surface
properties!Lectin chaperons
bind to sugar
residues
Glykoproteine & Qualitätskontrolle - ERQC (ER quality control)
ER Glucosidase I: GSCI entfernt die terminale α1,2-gebundene Glukose (1 von 3)
ER-Glucosidase II: GSCII entfernt die nächste α1,3-gebundene Glukose → Calnexin bzw. Calreticulinkönnen binden (CNX ist ein membranständiges und CRT ein lösliches Lektin-Chaperon)
ER-Glucosidase II: GSCII entfernt die letzte α1,3-gebundene Glukose nach korrekter Faltung (⇒ Export).
Bei Problemen erfolgt Interaktion mit Calreticulin (lösliches Lektin-Chaperon). Dies führtzu erneuter Glukosylierung durch UDP-Glucose-Glukosyltransferase (UGGT) zum Starteneines weiteren Faltungszyklus ⇒ mehr Zeit, um korrekte Konformation anzunehmen!
UGGT, re-
glucosylation
Calnexin (CNX) & Calreticulin (CRT)
are lectin chaperones and bind to the
N-glycans & monitor surface properties
GSCI, GSCII
GSCII

8
Was geschieht wenn Faltungsprobleme peristieren?
Glucosidasen
Glucosen
Faltungsproblem• falsche Konformation (Mutation)
• einzelne Untereinheit(en) fehlen
⇒ hydrophobe Oberflächen!
ER-Chaperone wie BiP binden an misgefaltete Proteine und aktivieren die sogenannte
Unfolded Protein Response = UPR
Erhöhte Expression von Chaperonen (mehr Faltungshelfer).
Wird durch Stress (Hitze, Salz, Pathogene, etc.) induziert.
Tunicamycin (hemmt N-Glykanbildung) hat eine ähnliche Wirkung!
NH2
COOH
HN
AsN
Mannosen
Protein
GlcNAc
ER stress signaling discovered in mammals and yeast
Signal integration in the endoplasmic reticulum unfolded protein responseDavid Ron & Peter Walter (July 2007) Nature Reviews Molecular Cell Biology 8, 519-529 doi:10.1038/nrm2199
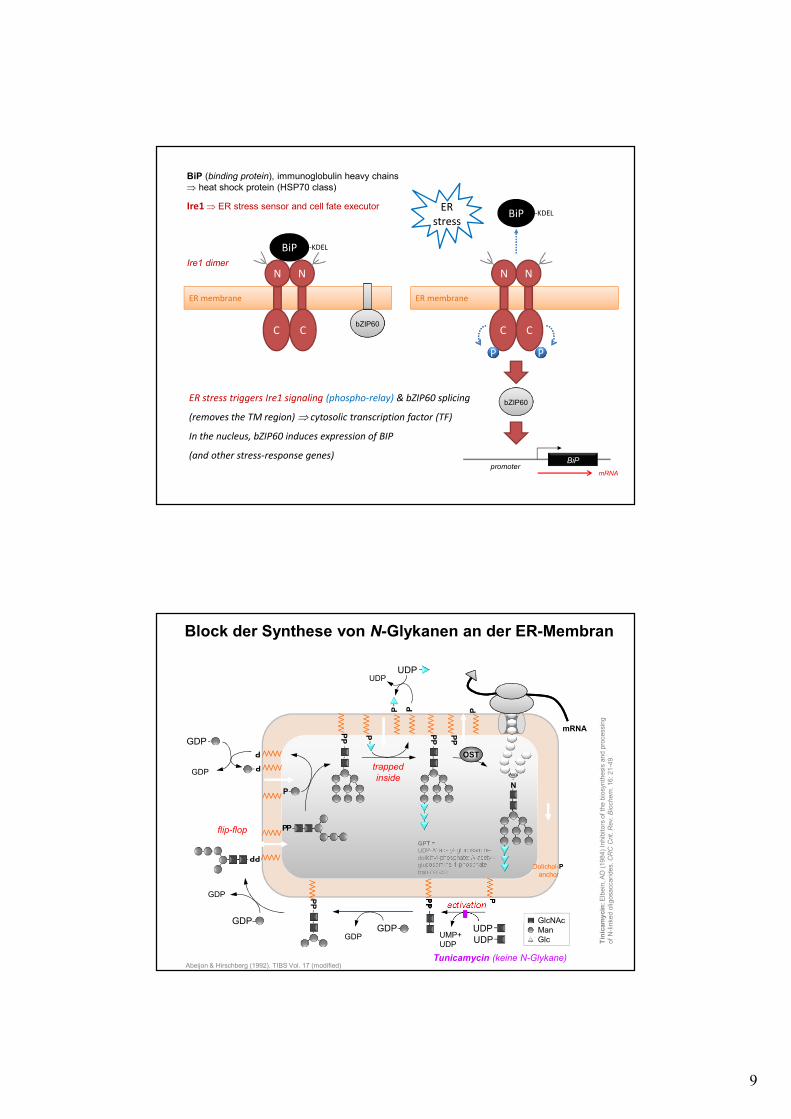
9
ER membrane
C C
N N
ER stress triggers Ire1 signaling (phospho-relay) & bZIP60 splicing
(removes the TM region) ⇒ cytosolic transcription factor (TF)
In the nucleus, bZIP60 induces expression of BIP
(and other stress-response genes)
ER
stress
PP
Ire1 ⇒ ER stress sensor and cell fate executor
Ire1 dimer
BiP -KDEL
ER membrane
C C
N N
BiP -KDEL
BiP (binding protein), immunoglobulin heavy chains⇒ heat shock protein (HSP70 class)
bZIP60
bZIP60
BiPpromoter
mRNA
Block der Synthese von N-Glykanen an der ER-Membran
Tin
icam
ycin
: Elb
ein
, AD
(1984)
Inhib
itors
ofth
ebio
synth
esi
sand
pro
cess
ing
ofN
-lin
ked
olig
osa
ccari
des.
CR
C C
rit.
Rev. B
iochem
. 16: 21-4
9.
Abeijon & Hirschberg (1992). TIBS Vol. 17 (modified)
PPP
P
P
P
GDP
GDP
P P
PP
PP
PP
PP
P
UDP
mRNA
PP
UMP+UDP
GDP
GDP
GDPGDP
UDPUDP
PP
PP
N
Asn
Dolichol-Panchor
flip-flop
OST
UDP
GlcNAcManGlc
trapped
inside
activation
GPT =UDP-N-acetyl-glucosamine-dolichyl-phosphate: N-acetyl-glucosamine-1-phosphate transferase
Tunicamycin (keine N-Glykane)

10
Was passiert mit endgültig falsch gefalteten Proteinen?
gefaltetes Glykoproteinmit Asparagin-gebundenerSeitenkette (N-Glykan)
Interaktion mit den ER-Lektinen Calnexin und Calreticulin via terminale Glucose-Reste (Faltungszyklen)
Entfernen von terminaler Mannose (Pos. 9 - im Faltungszyklus) durch ER Mannosidase I favorisiert Assoziation mit EDEM (ER-
degradation enhancing 1,2-mannosidase-like protein) und leitet so den Abbau ein: ER-Associated Degradation (ERAD).
⇒ Slow process (Mannose-timer model - Ari Helenius, 1994)
Fagioli & Sitia (2001) Glycoprotein Quality Control in the Endoplasmic Reticulum - MANNOSE TRIMMING BY
ENDOPLASMIC RETICULUM MANNOSIDASE I TIMES THE PROTEASOMAL DEGRADATION OF UNASSEMBLED
IMMUNOGLOBULIN SUBUNITS. The Journal of Biological Chemistry, 276: 12885-12892.
9
AUS: Fagioli & Sitia (2001)
Der Abbau falsch gefalteter Proteine erfolgt im Cytosol
nach Retro-Translokation durch den Sec61-Komplex
4 Schritte: - Substrat-Erkennung Chaperone bleiben gebunden (im ER-Lumen)- Rücktransport durch Sec61-Komplex (in der ER-Membran) - Deglykosylierung mittels Protein-N-Glykanase (PNGase, im Cytosol) - Markierung poly-Ubiquitinierung (im Cytosol)- Abbau in 26S-Proteasomen
Tsai et al. (2002) Retro-translocation of proteins from the Endoplasmic Reticulum into the cytosol. Nat. Rev. Mol. Cell Biol. 3:246-255.
Sec61
„Pore“
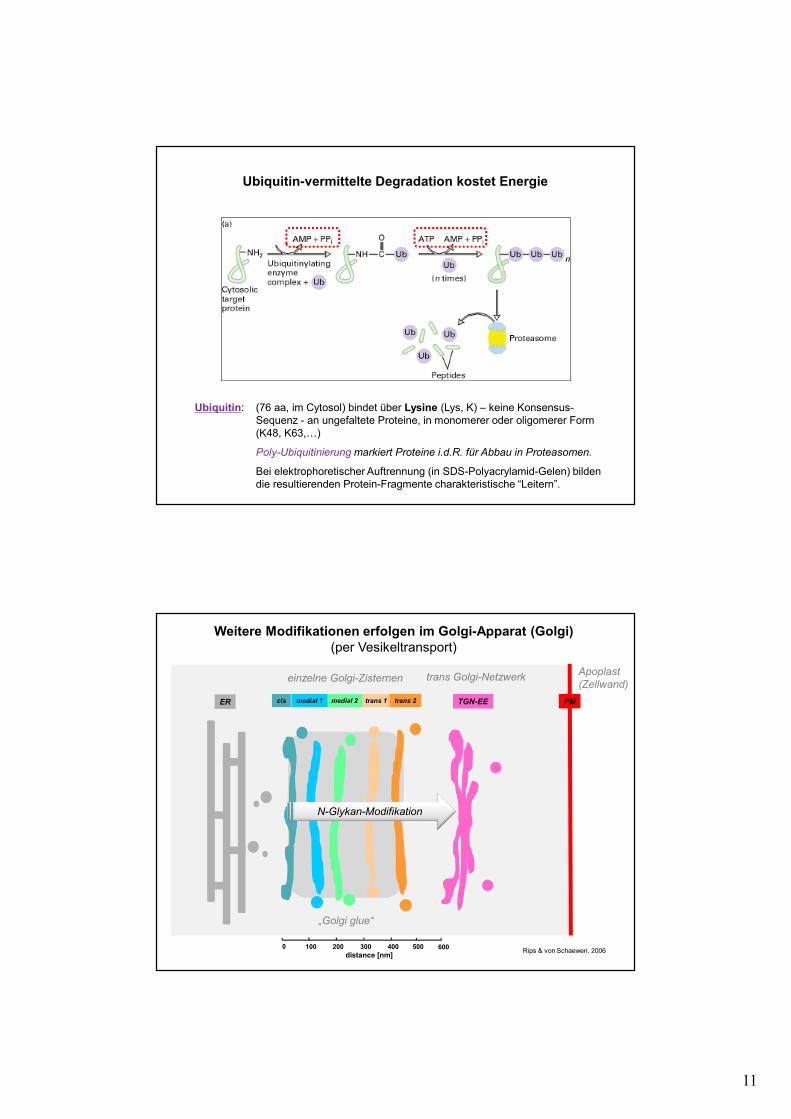
11
Ubiquitin-vermittelte Degradation kostet Energie
Ubiquitin: (76 aa, im Cytosol) bindet über Lysine (Lys, K) – keine Konsensus-Sequenz - an ungefaltete Proteine, in monomerer oder oligomerer Form (K48, K63,…)
Poly-Ubiquitinierung markiert Proteine i.d.R. für Abbau in Proteasomen.
Bei elektrophoretischer Auftrennung (in SDS-Polyacrylamid-Gelen) bildendie resultierenden Protein-Fragmente charakteristische “Leitern”.
„Golgi glue“
Weitere Modifikationen erfolgen im Golgi-Apparat (Golgi)(per Vesikeltransport)
cis trans 1medial 2 TGN-EEmedial 1
Rips & von Schaewen, 2006
trans 2
400distance [nm]
100 200 300 5000 600
ER
N-Glykan-Modifikation
einzelne Golgi-Zisternen trans Golgi-Netzwerk
PM
Apoplast
(Zellwand)

12
Die ER-Typ „high mannose“ N-Glykane werden im G-A zu „komplexen“ N-Glykanen modifiziert:
• Glykosidasen entfernen Zucker-Reste, Glykosyltransferasen heften NEUE an.
• Pflanzliche N-Glykane unterscheiden sich von Pilzen und Tieren (⇒ immunogenes Potential)
• Nicht alle N-Glykane sind nach Proteinfaltung zugänglich (⇒ „micro“-Heterogenität)
N-Glykan-Modifikation im Golgi-Apparat
Vorlesung 2016
Abb. Buchanan et al. (2000) „Biochemistry & Molecular Biology of Plants“ (Amer. Soc. of Plant Physiologists).
Man I
entfernt
Mannose-
Reste
GnT I
addiert
GlcNAc-
Rest
Man II
entfernt
Mannose-
Reste
XylT, FucT
& GnT II
addieren
Reste
GalT &
FucTc ⇒
Lewis a-
Epitope
Hexos-
aminidasen
entfernen
GlcNac
Sekretion
Vakuole
erhöht die Zugänglichkeit von Xyl und core Fuc
Verzweigung
am α1,6 –
Mannose-Arm !
Beispiel für Mikro-Heterogenität
ER-Typ „high mannose“ N-Glykane (in Taschen) ca. 2 kDa
Reife/prozessierte„complex type“N-Glykane (Oberfläche)ca. 1,3-1,7 kDa
AvS, 2009
SP
nach
Faltung
im ER

13
„high mannose“ Glykane in allen Eukaryonten
Pflanzliche N-Glykane des „komplexen“-Typs sind immunogen
Xyl- & Fuc-spezifische Antikörper
„komplexe“ Glykane in Pflanzen
„komplexe“ Glykane in Tieren
Arabidopsis cgl1-Mutante(complex glycosylation-less1)
von Schaewen et al. (1993),Kang et al. (2008), Frank et al. (2008)
cgl1-Mutanten
„complex glycan“-Spezifisches Kaninchen-Antiserum
bindet nicht anGkyoproteine
Gesamtprotein Immunoblot
RubisCOLSU (Blatt)
Sekretorische Proteine & Vesikeltransport
Vorlesung 2016
Abb. Buchanan et al. (2000) „Biochemistry & Molecular Biology of Plants“ (Amer. Soc. of Plant Physiologists).
ER-residente Chaperone, z.B.
zur Vakuole
zur Zellwand
Wie ist Verbleib im ER möglich?
Lösliche Proteine tragen ein C-terminales Signal für ER retention / retrieval (in Pflanzen -H/K/RDEL).
Ein spezifischer MEMBRAN-Rezeptor bindet diese Motive und bringt „entwischte“ Proteine von der cis-Seite des Golgi-Apparates zum ER zurück (retrograde protein
transport).
BiP -KDEL
Frage: Wie finden Vesikel zu
ihren ihre Zielorten?

14
…wider das Chaos
in Zellen…
Aus: „Frankfurter Allgemeine“ vom 17.10.2013
Säugerzellen Nervenzellen Hefezellen
Nobel-Preis 2013für „Medizin oder Physiologie“
SNAREs (SNAP-Rezeptoren) sind integrale Membranproteine auf der Oberfläche von Vesikeln und Zielorganellen
SNAP = Soluble N-ethylmaleimide-sensitive factor Attachment Protein
Das sogenannte SNARE-System
Vorlesung 2016
Abb. Buchanan et al. (2000) „Biochemistry & Molecular Biology of Plants“ (Amer. Soc. of Plant Physiologists).
Rab: GTP-bindende Proteinfamilie (GTPasen) fungieren als molekulare
Schalter im Vesikel-Transport.
GTP-Hydrolyse treibt die Fusionszyklen zwischen Vesikel- und Ziel-Membranen an (in Zellen muss es entsprechend viele verschiedene SNARE-Paare geben!)
• v-SNAREs: auf Vesikel-Membranen
• t-SNAREs: auf „target“-Membranen
N-ethylmaleimide, (NEM) bindet an Cystein-Reste

15
Der Golgi-Apparat & das trans-Golgi-Netzwerk (TGN)
Membranproteine und exponierte Hüllproteine (COPs, coat proteins) helfen beim Sortieren der transportierten Fracht:
z.B. bei Vesikel-Bildung, Abschnürung, Transport und Zielfindung:
• COP-Hüllen (ER und Golgi-Apparat)
• „glatte“ Hülle (zur Plasmamembran, Apoplast & Zellwand) Sekretion ⇒ default
• Clathrin-Hülle (zur Vakuole bzw. Tonoplast) ⇒ Rezeptor-vermittelt
Vorlesung 2016
Abb. Buchanan et al. (2000) „Biochemistry & Molecular Biology of Plants“ (Amer. Soc. of Plant Physiologists).
„cis“
„trans“
Zellwand(PM) Vakuole
(Tonoplast)
TGN-EE(pH-Abfall)
Golgi-„Stapel“
mit Vesikeln
Golgi-Matrix (in Pflanzen)
ER-Golgi-Transport-
Vesikel
Clathrin-Hülle
„glatte“Hülle
COP-Hülle
„protein sorting“
Golgi-Stapel bewegen sich in Pflanzen
In Pflanzenzellen ist der Golgi-Apparat über die gesamte Zelle verteilt (also „dispers“ organisiert).
Wie die Zeitraffer-Aufnahme zeigt, stehen Golgi-Stapel kaum still.
Man kann charakteristische „stop-
and-go movements“ beobachten, die wahrscheinlich der Aufnahme von Fracht dienen ⇒ Stoppen an ER-export sites (ERES).
GFP-Markierung von Golgi-Stapeln in Tabakzellen.
Durch Fusion einer Glykosyltransferase mit GFP leuchtet der GA grün (rot, Autofluoreszenz v. Chloroplasten; Aufn. Stephan Rips )
Vorlesung AvS 2016

16
shortfull
ER- und Golgi-Dynamik in Mesophyllzellen transgener Tabak-Pflanzen
ER
Golgi-Apparat
Serie auf der Stelle Serie durch die Zelle
The principle of receptor-mediated protein transport
Acceptor
compartment
releasebinding
Donor
compartment
recycling
sorting
VSR-ligand interaction is reversible, VSR transport is bi-directional
Courtesy of Dr. Peter Pimpl, Tübingen
17
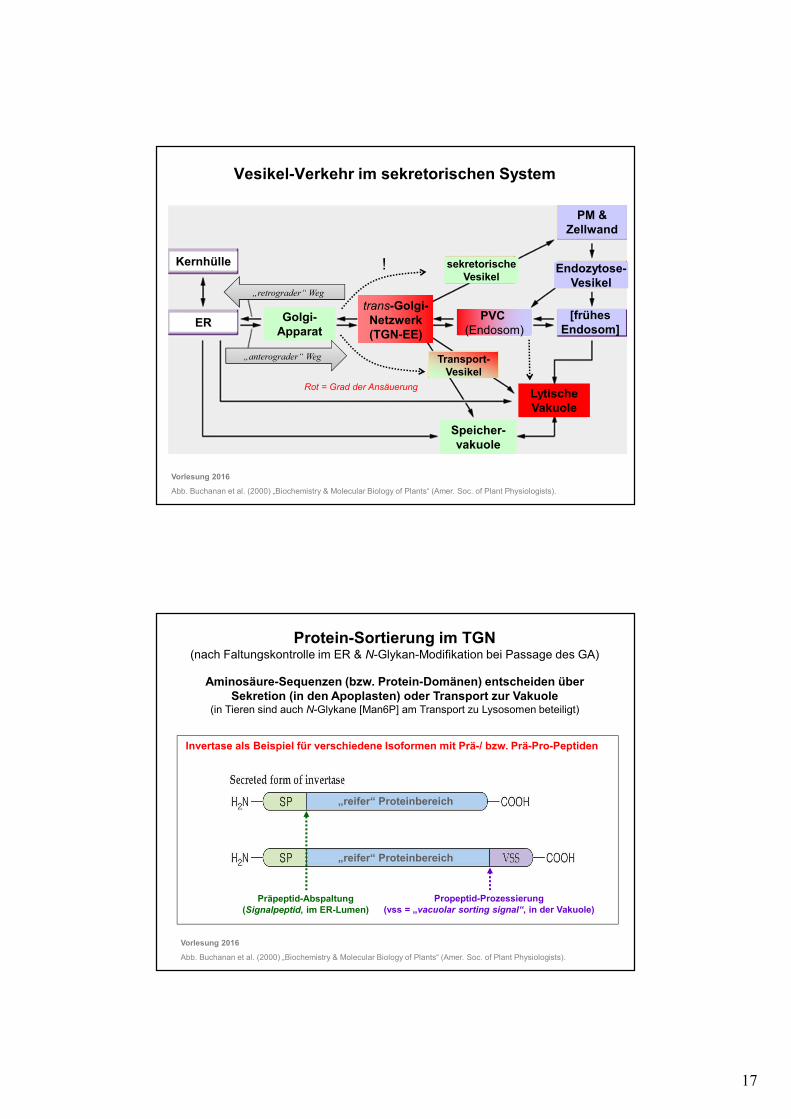
Vesikel-Verkehr im sekretorischen System
Vorlesung 2016
Abb. Buchanan et al. (2000) „Biochemistry & Molecular Biology of Plants“ (Amer. Soc. of Plant Physiologists).
ER Golgi-Apparat
trans-Golgi-Netzwerk(TGN-EE)
PM &Zellwand
LytischeVakuole
Speicher-vakuole
Kernhülle
PVC (Endosom)
[frühes Endosom]
Endozytose-Vesikel
sekretorischeVesikel
Transport-Vesikel
„retrograder“ Weg
„anterograder“ Weg
Rot = Grad der Ansäuerung
!
Protein-Sortierung im TGN (nach Faltungskontrolle im ER & N-Glykan-Modifikation bei Passage des GA)
Aminosäure-Sequenzen (bzw. Protein-Domänen) entscheiden über Sekretion (in den Apoplasten) oder Transport zur Vakuole
(in Tieren sind auch N-Glykane [Man6P] am Transport zu Lysosomen beteiligt)
Präpeptid-Abspaltung(Signalpeptid, im ER-Lumen)
Propeptid-Prozessierung(vss = „vacuolar sorting signal“, in der Vakuole)
Vorlesung 2016
Abb. Buchanan et al. (2000) „Biochemistry & Molecular Biology of Plants“ (Amer. Soc. of Plant Physiologists).
„reifer“ Proteinbereich
„reifer“ Proteinbereich
Invertase als Beispiel für verschiedene Isoformen mit Prä-/ bzw. Prä-Pro-Peptiden

18
Vakuolen
Lösliche Proteine gelangen i.d.R.
vom ER über den Golgi-Apparat,
Endosomen und das „prävakuoläre
Kompartiment“ (PVC) in lytische
Vakuolen (z.B. lytische Enzyme
vakuoläre Invertase,
Speicherproteine, Lektine, Protease-
inhibitoren,, etc.)
Vakuole
PVCManche Zellen enthalten verschiedene Vakuolentypennebeneinander:
- Saure Vakuolen (lytisch, für Stoff Um- und Abbau) - Neutrale Vakuolen (PSV, als Protein-Speicher)
sekretierte
Invertase
etc.
vakuoläre
Invertase,
etc.
sauer
neutral
MERKE:
Proteine können über unterschiedliche
Wege in PSV gelangen.
Protein-Speicher-Vakuolen (PSV) im Samen-Endoperm
Aleuron Zelle
V1, große Speichervakuole
V2, kleine lytische Vakuolen
A) Dichte Vesikel
B) Autophagie
Protein-Speicher-Vakuole
GArER
Vesikelfusion
PB-ER (protein body ER)

19
Beispiel: Speicherproteine des Reis-Endosperms
Prolamine 5% (Alkohol-löslich) kommen nur bei Monokotylen vor und fallen direkt nach Import und Faltung im ER-Lumen aus (akkumulieren zunächst dort).
Globuline 80% (Wasser-löslich) gelangen über den Golgi-Apparat in die PSV.
Indiz: Speicherproteine mit entweder rein ER- bzw. Golgi-typische N-Glykanen.
Vorlesung 2016Abb. Buchanan et al. (2000) „Biochemistry & Molecular Biology of Plants“ (Amer. Soc. of Plant Physiologists).
Nur ER-Typ
„high man“
N-Glykane
(GA bypass)
GA-Typ
„complex“
N-Glykane
Die pflanzliche Zellwand (Apoplast)
Ap
op
las
t
Cytosol
PM
20
Zusammenwirken
mehrerer Mechanismen:
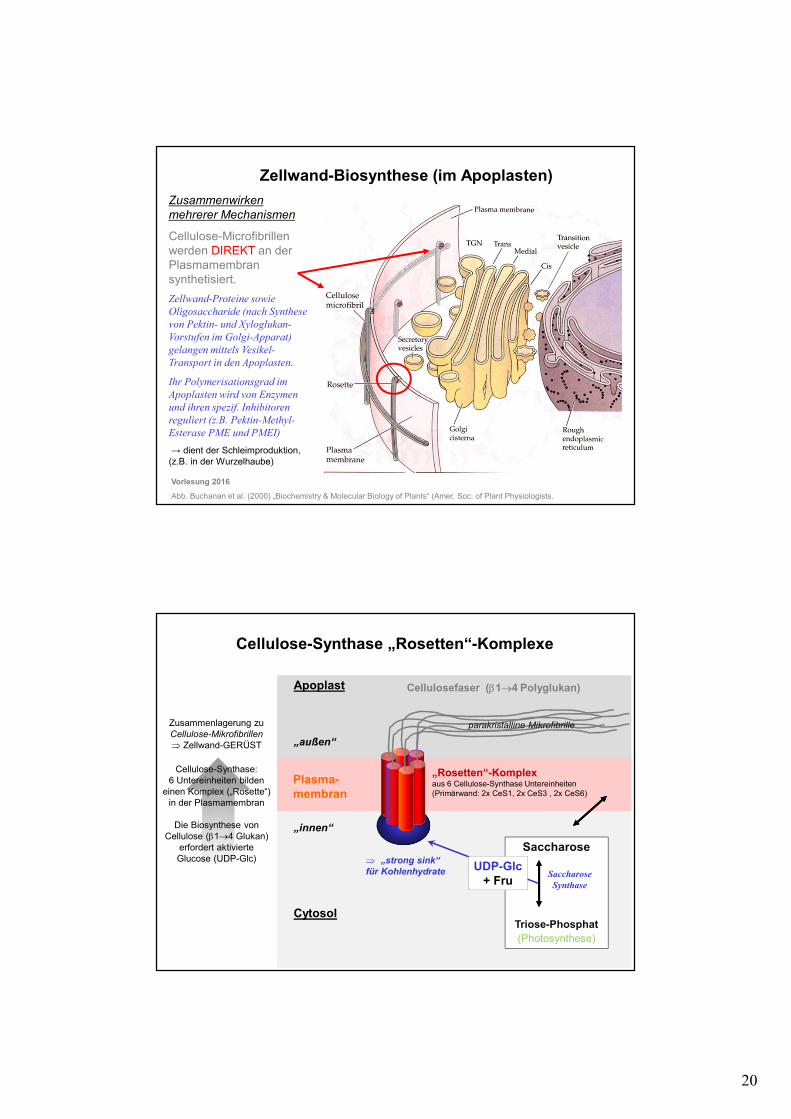
Cellulose-Microfibrillenwerden DIREKT an der Plasmamembran synthetisiert.
Zellwand-Biosynthese (im Apoplasten)
Vorlesung 2016
Abb. Buchanan et al. (2000) „Biochemistry & Molecular Biology of Plants“ (Amer. Soc. of Plant Physiologists.
Zellwand-Proteine sowie
Oligosaccharide (nach Synthese
von Pektin- und Xyloglukan-
Vorstufen im Golgi-Apparat)
gelangen mittels Vesikel-
Transport in den Apoplasten.
Ihr Polymerisationsgrad im
Apoplasten wird von Enzymen
und ihren spezif. Inhibitoren
reguliert (z.B. Pektin-Methyl-
Esterase PME und PMEI)
→ dient der Schleimproduktion, (z.B. in der Wurzelhaube)
Cellulose-Synthase „Rosetten“-Komplexe
Zusammenlagerung zu Cellulose-Mikrofibrillen
⇒ Zellwand-GERÜST
Cellulose-Synthase:6 Untereinheiten bilden
einen Komplex („Rosette“) in der Plasmamembran
Die Biosynthese von Cellulose (β1→4 Glukan)
erfordert aktivierte Glucose (UDP-Glc)
„Rosetten“-Komplex aus 6 Cellulose-Synthase Untereinheiten(Primärwand: 2x CeS1, 2x CeS3 , 2x CeS6)
Cellulosefaser (β1→4 Polyglukan)
Saccharose
Triose-Phosphat(Photosynthese)
UDP-Glc+ Fru
Plasma-membran
„innen“
Cytosol
Apoplast
„außen“
parakristalline Mikrofibrille
⇒ „strong sink“
für Kohlenhydrate Saccharose
Synthase

21
Im Golgi-Apparat werden verschiedene Oligo-
saccharid-Vorstufen für die Zellwand-MATRIX gebildet.
Hemicellulose-Vorstufen
(links, verzweigte
Xyloglukan-Ketten)
bzw.
Pektin-Vorstufen
(rechts, mit sauren Zuckern,
z.B. Galacturonsäure)
Später können die sauren Zuckerreste miteinander über divalente Kationen
(Ca2+, B3+) verbunden oder maskiert sein (Methyl-ester)
⇒ versch. Härtegrad!
Synthese von weiteren Oligosacchariden im Golgi-Apparat
Xyloglukan-Synthese Pektin-Synthese
Zellwand
Vorlesung 2016
Abb. Buchanan et al. (2000) „Biochemistry & Molecular Biology of Plants“ (Amer. Soc. of Plant Physiologists).
AGPs (Arabino-Galaktan-Proteine) - wichtig für die Entwicklung!
Extensine (Hydroxyprolin-reiche Proteine) - mechanische Wand-Eigenschaften, Signaling!
Vorlesung 2016
Abb. Buchanan et al. (2000) „Biochemistry & Molecular Biology of Plants“ (Amer. Soc. of Plant Physiologists.
Glykoproteine der Zellwand
Proteoglykane hoher Zuckeranteil, aber über OH-Gruppen verknüpft (d.h. via hydroxy-Pro und Ser/Thr)
AGP
Tomaten-Extensin (O-glykosyliert)
Linker-Region
Ceramid =
Abspaltung

22
Receptor-mediated MAPK signaling in plant growth & development
Figure 1
Xu & Zhang Trends in Plant Science (2015) 20, 56-64. DOI: (10.1016/j.tplants.2014.10.001) Copyright © 2014 Elsevier Ltd Terms and Conditions
Figure 2
Copyright © 2014 Elsevier Ltd Terms and ConditionsXu & Zhang Trends in Plant Science (2015) 20, 56-64. DOI: (10.1016/j.tplants.2014.10.001)
Receptor-like kinases (RLKs) perceive & transmit extracellular cues

23
Zusammenfassung:
Rolle des ER
• Protein-Import, N-Glykosylierung, Faltungskontrolle (ERQC), Export bzw. Abbau (ERAD), Retro-Translokation in das Cytosol und De-glykosylierung, Lysin-Ubiquitinierung und Abbau im Proteasom.
• Membranfluss (Lipide), anterograder und retrograder Vesikel-Transport ⇒ Zellwachstum (per Vesikelfusion mit Plasmamembran)
• Sekretion von hydrolytischen Enzymen in die Zellwand (Pathogen-Abwehr) und Sortierung in Valuolen (Stoff Um-/Abbau, Speicher)
• Bildung von Ölkörpern und Proteinspeichervakuolen (PSV) im Endosperm von Samen (Energie und Stickstoff für Nachkommen)
Zusammenfassung:
Rolle des Golgi-Apparates
• Modifikation von N-Glykanen (sekretierte Glykoproteine)
• Sekretion zur PM (default) Protein-Sortierung im TGN (zur Vakuole)
• Membran-Modifikation (z.B. Sterolbiosynthese am trans-Golgi/TGN)
• Synthese von Pektin- und Hemicellulose-Vorstufen (Zellwand-Matrix)
• „Schleim“-Produktion und Sekretion (z.B. Wurzelhaube)
Rolle des Apoplasten
• Zellwand-Synthese/Wachstum/Modifikation
• Transportweg für Saccharose (Hormone)
• Stress und Entwicklung „Signaling“ (nach Wahrnehmung)

24
Protein-Transport in Pflanzen
Sortieren der ER-importierten Proteinfracht (im TGN), gilt auch für integrale Membranproteine!
• Transport-Vesikel finden ihre Zielmembran durch Hüllproteine:
• COP II: anterograder Transport
• COP I: retrograder Transport
• „smooth“: zur Plasmamembran
• Clathrin-coated: Sorting, Vakuolen-Transport (TGN) und Endozytose (PM)
Merke: Das SNARE-System ermöglicht die (energieabhängige) Fusion von Vesikel- und Ziel-Membranen!
Vorlesung 2016
Abb. Buchanan et al. (2000) „Biochemistry & Molecular Biology of Plants“ (Amer. Soc. of Plant Physiologists).
Plastiden und Mitochondrien
Kern-kodierte Proteine werden im Cytosol an freien Ribosomentranslatiert und post-translational(ungefaltet) über die innere Hüllmembran transportiert.
Chaperone helfen beim Import:
Holdase-Funktion verhindert vorzeitiges Falten im Cytosol
Foldase-Funktion unterstützt Proteinfaltung im Stroma (nach Abspalten des Transitpeptids), sowie die Assemblierung von Komplexen.
Beispiel: Ribulose-1,5-bisphosphat Carboxylase/Oxygenase = RubisCO, aus 8 großen (LSU, Plastom-kodiert) und 8 kleinen (SSU) Untereinheiten.
Vorlesung 2016
Abb. Buchanan et al. (2000) „Biochemistry & Molecular Biology of Plants“ (Amer. Soc. of Plant Physiologists).
Abgrenzung:Protein-Import endosymbiontischer Organellen
RubisCO
LSU
RubisCO
SSU
Holdase
Foldase