6. metapopulaciona genetika i evolucijanasport.pmf.ni.ac.rs/materijali/2629/masterkonzppt6.pdf ·...
TRANSCRIPT

6. Metapopulaciona genetika i evolucija
Prof dr Jelka Crnobrnja Isailović
Konzervaciona biologija 6.

Konzervaciona biologija 6.
Metapopulacija se pojavljuje kao termin još u
tekstu Richard-a Levins-a (Ričard Luins) 1970,
kao skup više lokalnih populacija koje sve
poseduju osobinu izvesnog izumiranja u nekom
bliskom budućem vremenu, ali i osobinu
ponovnog ustanovljavanja i to upravo od
strane jedinki koje migriraju iz okolnih
lokalnih populacija.

Konzervaciona biologija 6.
Opstanak lokalnih populacija u definisanom
geografskom prostoru uslovljen je postojanjem
komunikacije između njih, tj mogućnošću
uspešne migracije, što definiše metapopulaciju
kao osnovnu jedinicu u okviru koje je opstanak
vrste na tom prostoru moguć na duže staze.

Konzervaciona biologija 6.
METAPOPULACIJA U EVOLUCIONOJ BIOLOGIJI
Skup dema ili lokalnih populacija (sinonim) ograničene veličine
unutar kojih je prisutno ukrštanje po principu slučajnosti,
a između kojih postoji ograničen protok gena (Tucić, 2003)

Konzervaciona biologija 6.
METAPOPULACIJA U EVOLUCIONOJ BIOLOGIJI
Usled dinamike izumiranja i obnavljanja lokalnih populacija,
stepen međupopulacione genetičke diferencijacije, tj nivo
genetičke strukturiranosti metapopulacije, nije veliki, bez obzira na
ograničen protok gena između lokalnih populacija

Genetička strukturiranost populacija
HT
HI2
HS2 HI1 HS1
HI3 HS3 HI4
HS4
HI = prosečna dobijena heterozigotnost unutar fragmenata
Hs = prosečna očekivana HW heterozigotnost po fragmentu
Ht = očekivana HW heterozigotnost za ukupnu populaciju (pulirano)
Konzervaciona biologija 6.

Genetička strukturiranost populacija
HT
HI2
HS2 HI1 HS1
HI3 HS3 HI4
HS4
FIS = 1 – (HI/HS)
FST = 1 – (HS/HT)
FIT = 1 – (HI/HT)
Konzervaciona biologija 6.

Konzervaciona biologija 6.
METAPOPULACIJA U EVOLUCIONOJ BIOLOGIJI

Konzervaciona biologija 6.
METAPOPULACIJA U EVOLUCIONOJ BIOLOGIJI
“korak-po-korak”“Source- sink” Metapopulacija
Nema aktuelnogProtoka gena
Genetičkiuniformna

Konzervaciona biologija 6.
GENETIČKA OSNOVA I MIGRACIJE
INTENZITET MIGRACIJE:
-Abiotički faktori
-Biotički faktori
-Stepen genetičke varijabilnosti za migratorno ponašanje

Konzervaciona biologija 6.
GENETIČKA OSNOVA I MIGRACIJE
MIGRANTI:
Neslučajan genetički uzorak jedinki lokalne populacije

Konzervaciona biologija 6.
GENETIČKA OSNOVA I MIGRACIJE

Konzervaciona biologija 6.
GENETIČKA OSNOVA I MIGRACIJE

Konzervaciona biologija 6.
GENETIČKA OSNOVA I MIGRACIJE
Proces migriranja, kada traje duži vremenski period,
izaziva promenu stope migracije u populaciji.

Konzervaciona biologija 6.
GENETIČKA OSNOVA I MIGRACIJE
EMIGRACIJA >> imigracija
Izolovane populacije
Smanjenje stope migracije

Konzervaciona biologija 6.
GENETIČKA OSNOVA I MIGRACIJE
emigracija << IMIGRACIJA
male deme u blizini velikih
povećanje stope migracije

Konzervaciona biologija 6.
GENETIČKA OSNOVA I MIGRACIJE
Evolutivna promena unutar metapopulacije:
Model
Konstantan broj lokalnih populacijaKonstantne razlike u veličini lokalnih populacijaKonstantne razlike u kvalitetu fragmenataCena migracije postoji
Mehanizam povećanja adaptivne vrednosti jedinke(variranje adaptivne vrednosti jedinki u vremenu,variranje veličine lokalne populacije u vremenu)

Konzervaciona biologija 6.
GENETIČKA OSNOVA I MIGRACIJE
Evolutivna promena unutar metapopulacije:
Model
Konstantan broj lokalnih populacijaKonstantne razlike u veličini lokalnih populacijaKonstantne razlike u kvalitetu fragmenataCena migracije postoji
Mehanizam povećanja adaptivne vrednosti jedinke(variranje adaptivne vrednosti jedinki u vremenu,variranje veličine lokalne populacije u vremenu)
Migracija u demusa budućnošću;
migracija u prazanfragment

Konzervaciona biologija 6.
GENETIČKA OSNOVA I MIGRACIJE
SELEKCIJA UNUTAR METAPOPULACIJE
Lokalni nivo
OSOBINE ŽIVOTNE ISTORIJE
Populacioni nivo

Konzervaciona biologija 6.
GENETIČKA OSNOVA I MIGRACIJE
SELEKCIJA UNUTAR METAPOPULACIJE
Lokalni nivo
STOPA MIGRACIJE
Populacioni nivo

Konzervaciona biologija 6.
GENETIČKA OSNOVA I MIGRACIJE
SELEKCIJA UNUTAR METAPOPULACIJE
Fragmentacija staništa povećava cenu (rizik) migracije
Raste unutarpopulaciona selekcija PROTIV “migratornih” alela
Smanjuje se veličina lokalnih populacija
Povećava se intenzitet izumiranja lokalnih populacija
Povećava se variranje adaptivne vrednosti unutar metapopulacije
Intenzivira se selekcija za održavanje i povećanje učestalosti “migratornih” alela na metapopulacionom nivou


Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
Uticaj evolucionih faktora na metapopulacionu dinamiku još uvek je
nedovoljno istražen.

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
Uticaj evolucionih faktora na metapopulacionu dinamiku još uvek je
nedovoljno istražen.

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
EFEKAT INBRIDINGA
30% manja stopa izleganja jaja
Melitaea cinxia

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
EFEKAT “PROČIŠĆAVANJA” OD ŠTETNIH RECESIVNIH ALELA I METAPOPOPULACIJA
Veoma male populacije se ne mogu “pročistiti” jer se dešava fiksiranje alela sa ne tako jakim štetnim efektima,
koji se prenose i putem protoka gena između lokalnih populacija
a i povećani inbriding usled male veličine populacije,odnosno efekta slučajnog oscilovanja učestalosti alela
može brzo dovesti do inbridne depresije!

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
VARIJANSNA EFEKTIVNA VELIČINA POPULACIJE(Haldane, 1939)
Ne (v) = 4N/(Vk+2)

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
VARIJANSNA EFEKTIVNA VELIČINA POPULACIJE(Haldane, 1939)
Ne (v) = 4N/(Vk+2)
N = veličina populacije dobijena cenzusomVk = varijansa broja gameta koje proizvede
generacija roditelja

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
VARIJANSNA EFEKTIVNA VELIČINA POPULACIJE(Haldane, 1939)
Ne (v) = 4N/(Vk+2)

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
VARIJANSNA EFEKTIVNA VELIČINA POPULACIJE(Crow i Kimura, 1970)
Ne (v) = (Nk - k) / (1 + Vk/k)

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
VARIJANSNA EFEKTIVNA VELIČINA POPULACIJE(Crow i Kimura, 1970)
Ne (v) = (Nk - k) / (1 + Vk/k)
N = veličina populacije dobijena cenzusomk = prosečan broj potomaka po jedinkiVk = populaciona varijansa k

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
VARIJANSNA EFEKTIVNA VELIČINA POPULACIJE(Crow i Kimura, 1970)
Ne (v) = (Nk - k) / (1 + Vk/k)
Kada je reproduktivni uspeh jedinki slučajan, Vk=k=2 iNe (v) = N-1 približno Ne (v) = N

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
VARIJANSNA EFEKTIVNA VELIČINA POPULACIJE(Crow i Kimura, 1970)
Ne (v) = (Nk - k) / (1 + Vk/k) Vk=k=2
Ne (v) = (Nk - k) / (1 + 1)Ne (v) = (Nk - k) /2Ne (v) = (N - 1)k / 2Ne (v) = N - 1

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
Uticaj populacionog obrta:
Brzina i intenzitet izumiranja pojedinačnih elemenata metapopulacije
Veći uticaj populacionog obrta je ukoliko su deme veće
Izumiranje velikih dema intenzivnije smanjuje ukupnu genetičku varijabilnostmetapopulacije

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
Izumiranje dema
Ustanovljavanje malih propagula
Efekat osnivača

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
Izumiranje dema
Ustanovljavanje malih propagula
Efekat osnivača

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
Metapopulacije:
Kombinovani efekat uticaja Nm i e/c na FST

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
Metapopulacije:
Nm< < 1,
e/c → FST , ako
k<((2Nm/(1-φ) + 0.5)

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
Metapopulacije:
k<((2Nm/(1-φ) + 0.5)
k - broj jedinki koji obnavlja datu lokalnu populaciju,
N - ravnotežna veličina lokalnih populacija
φ - verovatnoća da su dva genska alela u grupi k poreklom iz iste populacije.

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
Metapopulacije:
k<((2Nm/(1-φ) + 0.5)
k ≈ Nm
e/c → FST

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
Metapopulacije:
k<((2Nm/(1-φ) + 0.5)
Intenzivan populacioni obrt →visoke vrednosti Nm
Nema genetičke strukturiranosti metapopulacije

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
Metapopulacije:
FST(sadašnji) = 1/2k +φ(1-1/2k) FST(prethodni)
pod uslovom
N>>k

Konzervaciona biologija 6.
GENETIČKA OSNOVA I IZUMIRANJE LOKALNIH
POPULACIJA
Metapopulacije:
FST(sadašnji) = 1/2k +φ(1-1/2k) FST(prethodni)
φ~ 0, k određuje FST
φ~ 1, nema mešanja alela iz različitih dema prilikom rekolonizacije, FST


Konzervaciona biologija 6.
RAJTOVA TEORIJA POMIČNE RAVNOTEŽE KAO PRIMER DINAMIKE
GENETIČKE STRUKTURE NA METAPOPULACIONOM NIVOU
Kompleksi malih dema povezani protokom gena
Interakcija delovanja genetičkog drifta i selekcije
Izumiranja i rekolonizacije pojedinačnih dema su realne

Konzervaciona biologija 6.
RAJTOVA TEORIJA POMIČNE RAVNOTEŽE KAO PRIMER DINAMIKE
GENETIČKE STRUKTURE NA METAPOPULACIONOM NIVOU
FAZA 1. Genetički drift oblikuje lokalne genetičke strukture
FAZA 2.Fiksiranje adaptivnih genskih kombinacija putem selekcije
FAZA 3. Interdemno širenje adaptivnih genskih kombinacija protokom gena;metapopulacija napreduje ka “adaptivnom vrhu”

Konzervaciona biologija 6.
RAJTOVA TEORIJA POMIČNE RAVNOTEŽE KAO PRIMER DINAMIKE
GENETIČKE STRUKTURE NA METAPOPULACIONOM NIVOU
FAZA 3.
Zbog epistatičkih genskih interakcija, različite adaptivne genske kombinacije
mogu imati istu (maksimalnu) adaptivnu vrednost .
“Okupaciju” tj “pomeranje adaptivnog vrha” sprečava snažan protok gena (Nm ≥ 1)

Konzervaciona biologija 6.
RAJTOVA TEORIJA POMIČNE RAVNOTEŽE KAO PRIMER DINAMIKE
GENETIČKE STRUKTURE NA METAPOPULACIONOM NIVOU
FAZA 3.
Zbog epistatičkih genskih interakcija, različite adaptivne genske kombinacije
mogu imati istu (maksimalnu) adaptivnu vrednost .
“Okupaciju” tj “pomeranje adaptivnog vrha” sprečava snažan protok gena (Nm ≥ 1),“okupacija” migranata iz brojnijih populacija sa nižom prosečnom adaptivnom vrednošću

Konzervaciona biologija 6.
RAJTOVA TEORIJA POMIČNE RAVNOTEŽE KAO PRIMER DINAMIKE
GENETIČKE STRUKTURE NA METAPOPULACIONOM NIVOU
FAZA 3.
“Range shift” – pomeranje areala
Pleistocenske glacijacije

Konzervaciona biologija 6.
RAJTOV “OSTRVSKI” MODEL POPULACIONE STRUKTURE KAO
PRIMER DINAMIKE EFEKTIVNE VELIČINE POPULACIJE NA
METAPOPULACIONOM NIVOU
Modeli populacione strukture
Ostrvski model - Nm < 1; FST > 0.20

Konzervaciona biologija 6.
RAJTOV “OSTRVSKI” MODEL POPULACIONE STRUKTURE KAO
PRIMER DINAMIKE EFEKTIVNE VELIČINE POPULACIJE NA
METAPOPULACIONOM NIVOU
Rajt (Wright, 1943) je definisao efektivnu veličinu metapopulacije
koja se sastoji od idealnih n subpopulacija u kojima je Ne = N, kao
NeT ≈ NT/(1- FST),
NT = nN tj ukupan broj jedinki u metapopulaciji.

Konzervaciona biologija 6.
RAJTOV “OSTRVSKI” MODEL POPULACIONE STRUKTURE KAO
PRIMER DINAMIKE EFEKTIVNE VELIČINE POPULACIJE NA
METAPOPULACIONOM NIVOU
NeT ≈ NT/(1- FST),
FST > 0 NeT > NT
u ravnotežnom stanju FST ≈ 1/(1 + 4Nem),
NeT/NT ≈ 1 + 1/4Nem

Konzervaciona biologija 6.
RAJTOV “OSTRVSKI” MODEL POPULACIONE STRUKTURE KAO
PRIMER DINAMIKE EFEKTIVNE VELIČINE POPULACIJE NA
METAPOPULACIONOM NIVOU
U uslovima gde su pogodni fragmenti staništa izrazito izolovani, njihov kapacitet sredine podjednak, i ako lokalna regulacija populacije održava veličinu populacije stabilnom,
fragmentacija može ograničiti varijansu reproduktivnog uspeha i tako voditi ka većoj NeT .

Konzervaciona biologija 6.
RAJTOV “OSTRVSKI” MODEL POPULACIONE STRUKTURE KAO
PRIMER DINAMIKE EFEKTIVNE VELIČINE POPULACIJE NA
METAPOPULACIONOM NIVOU
Isti efekat mogu imati specifični dizajn rezervata prirode i programi uzgajanja u zatočeništvu,
putem izjednačavanja reproduktivnog uspeha u subpopulacijama.
Konzervaciona biologija 6.
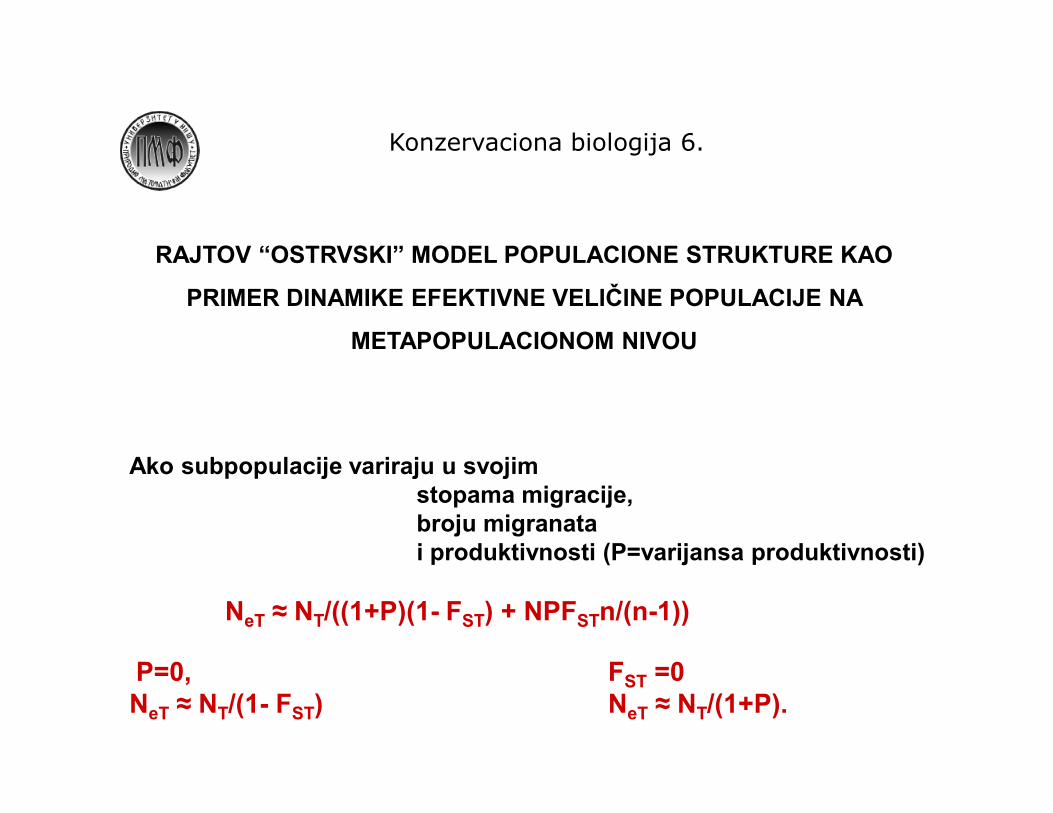
RAJTOV “OSTRVSKI” MODEL POPULACIONE STRUKTURE KAO
PRIMER DINAMIKE EFEKTIVNE VELIČINE POPULACIJE NA
METAPOPULACIONOM NIVOU
Ako subpopulacije variraju u svojim stopama migracije, broju migranata i produktivnosti (P=varijansa produktivnosti)
NeT ≈ NT/((1+P)(1- FST) + NPFSTn/(n-1))
P=0, FST =0NeT ≈ NT/(1- FST) NeT ≈ NT/(1+P).

Konzervaciona biologija 6.
RAJTOV “OSTRVSKI” MODEL POPULACIONE STRUKTURE KAO
PRIMER DINAMIKE EFEKTIVNE VELIČINE POPULACIJE NA
METAPOPULACIONOM NIVOU
NeT ≈ NT/((1+P)(1- FST) + NPFSTn/(n-1))
P=0, FST =0NeT ≈ NT/(1- FST) NeT ≈ NT/(1+P).
Fragmentacija metapopulacije, čak iako nema genetičke diferencijacije, utiče na smanjenje efektivne veličine populacije zbog postojanja varijanse produktivnosti subpopulacija (P>0).

Konzervaciona biologija 6.
RAJTOV “OSTRVSKI” MODEL POPULACIONE STRUKTURE KAO
PRIMER DINAMIKE EFEKTIVNE VELIČINE POPULACIJE NA
METAPOPULACIONOM NIVOU
Model koji obuhvata interdemni genetički drift (variranje produktivnosti subpopulacija i tako i doprinosa skupu migranata) generisan nakupljanjem slučajnih razlika u reproduktivnom uspehu između jedinki i pretpostavljenim inbridingom unutar subpopulacija, izraženim preko FIS koeficijenta, opisan je
jednačinom:
NeT ≈ NT/((1+ FIS)(1+ FST) - 2 FISFST)

Konzervaciona biologija 6.
RAJTOV “OSTRVSKI” MODEL POPULACIONE STRUKTURE KAO
PRIMER DINAMIKE EFEKTIVNE VELIČINE POPULACIJE NA
METAPOPULACIONOM NIVOU
FIS =0
NeT ≈ NT/(1+ FST)

Konzervaciona biologija 6.
RAJTOV “OSTRVSKI” MODEL POPULACIONE STRUKTURE KAO
PRIMER DINAMIKE EFEKTIVNE VELIČINE POPULACIJE NA
METAPOPULACIONOM NIVOU
Proces izumiranja/rekolonizacije može drastično da umanji efektivnu veličinu metapopulacije, naročito ako je broj naseljenih fragmenata staništa tako mali da je metapopulacija u opasnosti od izumiranja.

Konzervaciona biologija 6.
RAJTOV “OSTRVSKI” MODEL POPULACIONE STRUKTURE KAO
PRIMER DINAMIKE EFEKTIVNE VELIČINE POPULACIJE NA
METAPOPULACIONOM NIVOU
Ako je varijansa produktivnosti metapopulacije rezultat izumiranja irekolonizacije subpopulacija, odnos između efektivne veličinemetapopulacije i koeficijenta međupopulacione genetičkediferencijacije biće:
NeT ≈ n/4(m+e)FST

Konzervaciona biologija 6.
RAJTOV “OSTRVSKI” MODEL POPULACIONE STRUKTURE KAO
PRIMER DINAMIKE EFEKTIVNE VELIČINE POPULACIJE NA
METAPOPULACIONOM NIVOU
NeT ≈ n/4(m+e)FST
U ovakvom modelu, efektivna veličina metapopulacije bićeproporcionalna broju subpopulacija (n), a ne veličini subpopulacije(N) ili ukupnoj veličini metapopulacije (NT).