apobecファミリーとその抗ウイルス活性jsv.umin.jp/camp/13th/13th_w1 slide.pdf ·...
TRANSCRIPT

APOBECファミリーとその抗ウイルス活性
第13回ウイルス学キャンプ in 湯河原
主催:ウイルス学キャンプ実行委員会
村松 正道
金沢大学 医 分子遺伝
平成28年8月30日
ニューウェルシティー湯河原

1. APOBECとは
2. HBVとAPOBEC
3. HPV16とAPOBEC
総論的
これまでにわかった事
未解決事象や問題点
4. 失敗や経験から学んだこと

deamination o
NH3
C
N
N
o DNA/RNA
NH2
C U C
N
N
o DNA/RNA
APOBEC family
Anti viral factor
(HIV-1)
APOBEC
HBV & HPV16
replication block
GCATATCCTCCGCCAACCGCACTCCA
APOBEC3
GUATATCUTUUGUUAACUGUACTCUA
Hypermutation
viral DNA

HBV
3.2kb
DNA virus Human hapatocytes
Hepatitis B virus (HBV)
cirrhosis
liver cancer

A3GのHBVへの抗ウイルス作用
IFNs A3G
A3G
UNG A3G UUUUU
A3G C−to-U
Plos Pathogens 2013

AIDの抗HBV作用(モデル)
TGF-b
TNF-a
IL-1b
AID
AID AID
C−to-U UUU
Liang et al PNAS 2013
Watashi et al JBC 2013
Liang et al Plos Pathogens
2016

pgRNA
RC-DNA
3.2kb
cccDNA
HBx,
HBs
p
p
☆患者病態では、どうなっている?
☆実際のウイルス排除にどの程度貢献しているか?
☆肝癌との関連性?
☆エスケープ変異体、薬剤耐性変異体との因果関係?
Cytokines APOBECs
hypermutate
hypermutate
HBV感染におけるAPOBEC未解決事象
degradation

deamination o
NH3
C
N
N
o DNA/RNA
NH2
C U C
N
N
o DNA/RNA
APOBEC family
APOBEC
HBV & HPV16
Anti viral factor
(HIV-1)
replication block
GCATATCCTCCGCCAACCGCACTCCA
APOBEC3
GUATATCUTUUGUUAACUGUACTCUA
Hypermutation
viral DNA

Human Papillomavirus type 16
(HPV16)
咽頭癌
年間3500人の
日本人女性が死亡。
タバコとともに
中咽頭癌の
主要原因

HPV16
排除 排除
自然排除
HPVによる発ガン機構
(10-30年の経過)
ウイルス持続感染
①ウイルスゲノム挿入
②ウイルス癌遺伝子
E6, E7の持続的発現
とp53, Rb抑制
③宿主ゲノム変異
episome ゲノム挿入
形質転換

仮説: APOBEC = 諸刃の剣
GCATATCCTCCGCCAACCGCACTCCA
AID・APOBEC
GCATATCCTCCGUCAACCGCACTCCA
感染防御能 発がん
病原体の多い環境では、
発がんリスクに目をつぶっても
あまりあるメリットを提供している。

HPV16
排除 排除
自然排除
APOBECが関わるとしたら?
(10-30年の経過)
ウイルス持続感染
抗ウイルス因子
としての
APOBECの役割
急性期 慢性期
変異源としてのAPOBECの役割
①ウイルスゲノム挿入
②ウイルス癌遺伝子
E6, E7の持続的発現
とp53, RB抑制
③宿主ゲノム変異
現時点では
この仮説は、
部分的知見があるのみ。

Pseudovirion系はあるものの
動物モデル
in vitro自然感染モデル not available
HPV16研究の現状
発ガン初期 event の研究にW12が用いられている。
感染初期
の研究は
難しい。
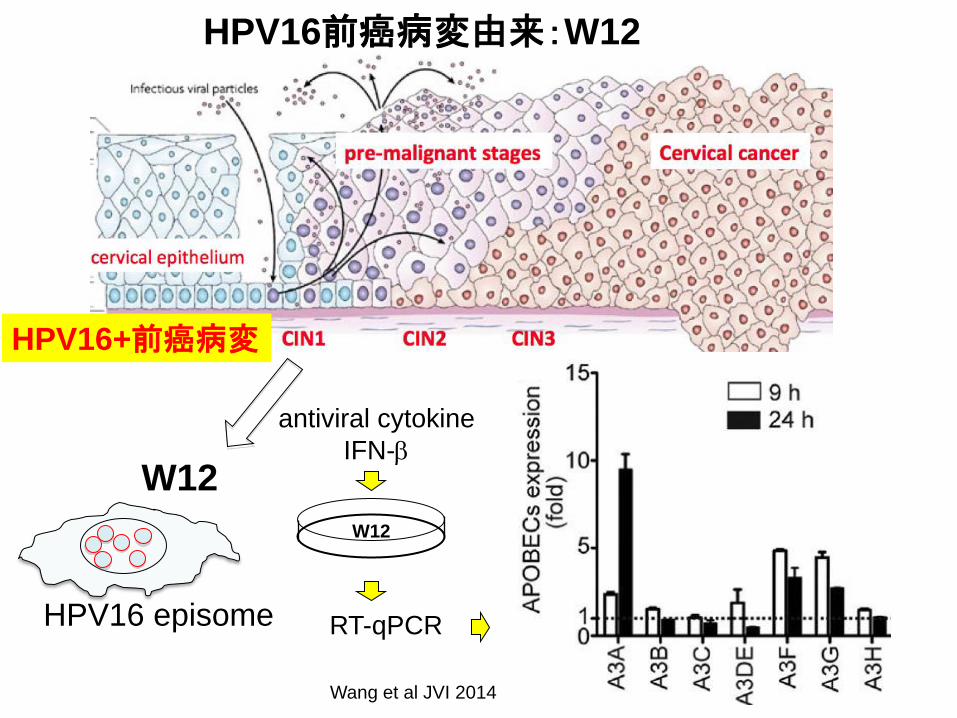
HPV16前癌病変由来:W12
W12
antiviral cytokine
IFN-b
W12
RT-qPCR HPV16 episome
HPV16+前癌病変
Wang et al JVI 2014

HPV16 episome
IFN-dependent hypermutation in W12 cells
W12 IFN-b
W12
3D-PCR
℃
non-treat
IFN-b
IFN処理でhypermutation観察可能。
G
C
G
C
C
G
A
T
T
A
C
G
G
C
A
T T
A
A
T
T
A
T
A
AT rich DNA
APOBEC hypermutation
(= Hypermutated DNA)
PCR変成温度

Overexpression of APOBECs induces hypermutation on HPV16 E2 gene
IF
Nb
E2 3D-PCR
GCATCCTAATTT!
CGTAGGATTAAA!
GCATUCTAATTT!
CGTAGGATTAAA!
GCAT--CTAATTT!
CGTAGGATTAAA!
; 0<2,=%
GCAT CTAATTT!
CGTAGGATTAAA!>&; !( /%?@?3%
A&B; 0CDE*F2
G0H!!,7=%3/?I ' 7
H,/?)!,7.%6I ' 7
*1=' @,7%2
; 0E
*<=' <#!!B<=' <; !
G1J %/+ K=?I ' 7
L
B%7' + ,6!>&; !MN,/?)!O!$' 2=P
QKJ J /%22,' 7!' .!!N,/?)!/%J ),6?I ' 7L
L
L
A&B!,7$,( ,=' /!
H /) (-03-8: ; <=>?3-F-
RS T=!,2!6' + J ' 2%5!' .!; F; U!; FDU!; F*U!; F>EU!; FVU!; FBU!; FGS!!
8S ; 0CDE*F2!J /' =%,72!?/%!?( )%!=' !%5,=!N,/K2!3%7' + %!,7!GTH!UGDHU!EDHU!GQH!?75!; ; HU!
%N%7!,7!G0H!W*X!/%3,' 7S!FS T7!( ?2%!/%J ?,/%5!J /' 6%22U!*!=' !A!6' 7N%/2,' 7!Y ' K)5!6/%?=%!>QD!M!5' K( )%<
2=/?75!( /%?@PS!!!
RS!; 0CDE*F; !?75!; 0CDE*FB!?/%!%ZJ /%22%5!,7!$K+ ?7!@%/?I 7' 61=%!6%))!),7%!M[ R8P!?75!,75K6%5!( 1!TV&!!!
! "#$%&#'(
9!
8!
\ !
] !
: !
R9!
R8!
; F; ! ; FD! ; F*! ; F>E! ; FV! ; FB! ; FG!
8:;<=>
3-$I#%$330+,-J.+1CK-
^$! 8\$!) *+, - ./$01'([ R8!6%))!6' 7=?,72!G0HR] !>&; !J /%2%7=!?=! ?J J /' Z,+ ?=%)1! R99! 6' J ,%2! ?75! =$%!2=?=%!' .!=$%!N,/?)!>&; !,2!+ ?,7=?,7%5!?=!%J ,2' + %!' N%/!?!7K+ ( %/!' .!J ?22?3%2S!
8?8L-8?M-) , C-8?N-) %$-$I #%$33$C-) , C-0, C' D$C-9"-OM7P-
IFN-b
+UGI overexpression
Wang et al JVI 2014

overexpression
Southern blot
HIVの時と比較すると
Mutationの頻度も、
2桁以上、低い事が判明。
ウイルス複製量は?
Wang et al JVI 2014

APOBEC3 抗ウイルス活性
Hypermutation
Hypermutation依存性
HIV-1
ウイルス成分に結合して
複製阻害する。
HIV-1での研究では、、
Hypermutation非依存性

293T
lipofection
pHPV16 (L1 + L2) ( capsid expressing vector)
p-Luciferase
Pseudovirion
+ APOBEC3
GFP
Pseudovirion
0
20
40
60
80
100
120
GFP A3A A3C A3F A3G
Infectivity
Infection Hela
100%
Lu
cife
rase
Ahasan et al BBRC 2015
ウイルス粒子集合
エントリー
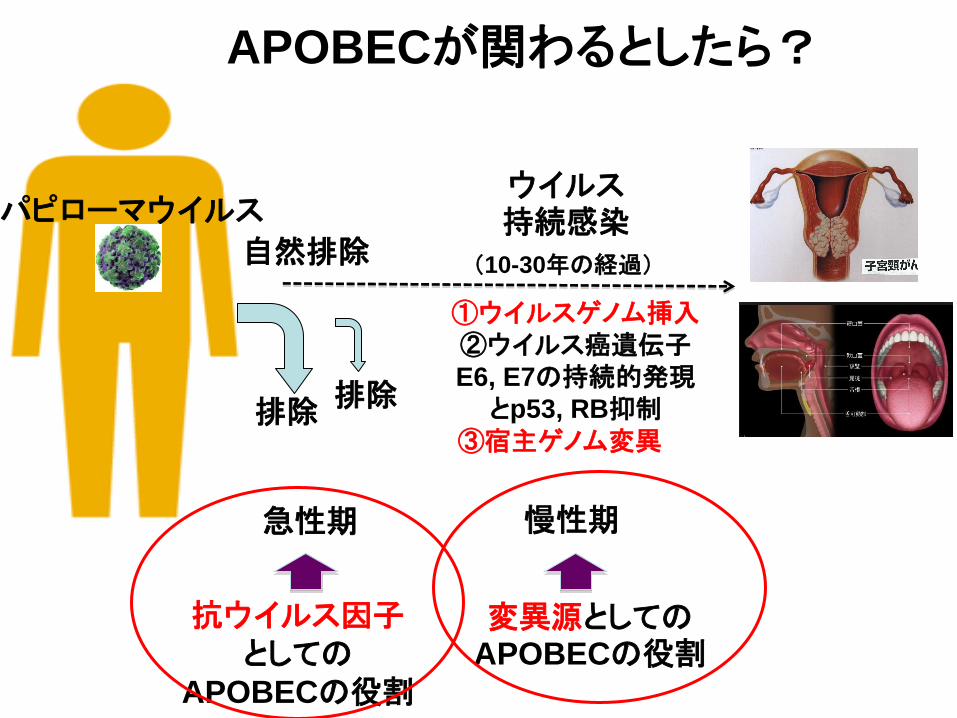
パピローマウイルス
排除 排除
自然排除
APOBECが関わるとしたら?
急性期
(10-30年の経過)
ウイルス持続感染
抗ウイルス因子
としての
APOBECの役割
慢性期
変異源としてのAPOBECの役割
①ウイルスゲノム挿入
②ウイルス癌遺伝子
E6, E7の持続的発現
とp53, RB抑制
③宿主ゲノム変異

HPV16+ CIN1 臨床検体
3D-PCR
HPV16 viral DNA Hypermutation
G
C
G
C
C
G
A
T
T
A
C
G
G
C
A
T T
A
A
T
T
A
T
A
AT rich DNA
APOBEC hypermutation
臨床検体でも
ウイルスDNAにAPOBEC活性産物を検出できる。
(= Hypermutated DNA)
control
DNA
hypermutated
Hypermutation
amplification at lower Tm
=
Wakae et al. Virology 2015

UGI A3A
- -
+ -
+ -
+ +
UGI A3A
- -
+ -
+ -
+ +
TP53 HPV16 E2
87
.0
86
.7
86
.1
85
.1
83
.9
83
.1
82
.4
82
.0
(℃)
87
.5
86
.9
86
.5
86
.1
85
.2
84
.5
84
.3
(℃)
↗ A G C T
A 0
G 56
C
T
178bp x 6 clones
↗ ︎ A G C T
A 0
G 74
C 24
T
165bp x 7 clones
↗ ︎ A G C T
A 0
G 0
C 49
T
165bp x 4 clones
APOBEC3A
HPV16+
UNG阻害
293T細胞
3D-PCR
APOBEC3Aは、宿主ゲノムにもhypermutationを入れうる。 Kondo et al. Oncogene in press
宿主ゲノムDNAにはhypermutationは起こりうるか?

HPV16
排除 排除
自然排除
現在、作業仮説の妥当性を検証中
急性期
(10-30年の経過)
ウイルス持続感染
抗ウイルス因子
としての
APOBECの役割
慢性期
変異源としてのAPOBECの役割
①ウイルスゲノム挿入
②ウイルス癌遺伝子
E6, E7の持続的発現
とp53, RB抑制
③宿主ゲノム変異

APOBECのプロジェクト紹介
は
ここまで。
以後は、失敗談へ

これまでに失敗や経験から学んだ事
1, 埋没するぐらい実験すべし!
しかし目指すものを忘れるな!
2, がむしゃらにサイエンスに打ち込んでください。
その姿勢はいずれは評価されます。
経験 1
経験 2
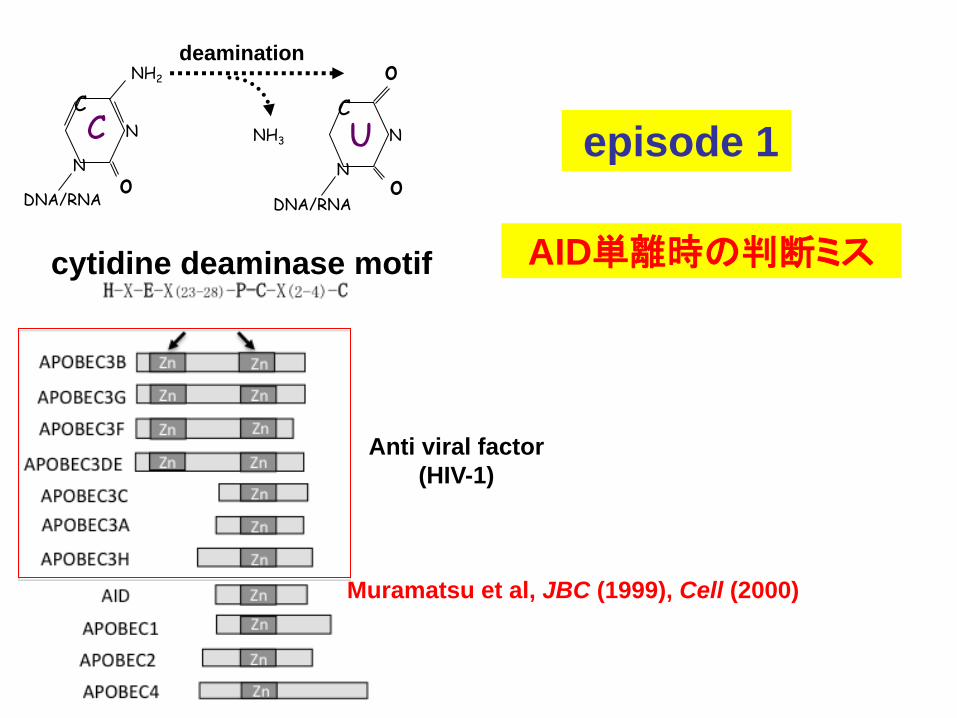
episode 1
o
NH3
C
N
N
o DNA/RNA
NH2
C U C
N
N
o DNA/RNA
deamination
cytidine deaminase motif
Muramatsu et al, JBC (1999), Cell (2000)
AID単離時の判断ミス
Anti viral factor
(HIV-1)

抗原刺激
Somatic hypermutation
V
クラススイッチ組換え
C C C C Cb Ca C
S S S Sb Sa S V
抗体重鎖 遺伝子
V
?
抗体の親和性成熟 IgG, IgA, IgE産生

抗原刺激
Somatic hypermutation
V
クラススイッチ組換え
C C C C Cb Ca C
S S S Sb Sa S V
抗体重鎖 遺伝子
V
AID
抗体の親和性成熟 IgG, IgA, IgE産生
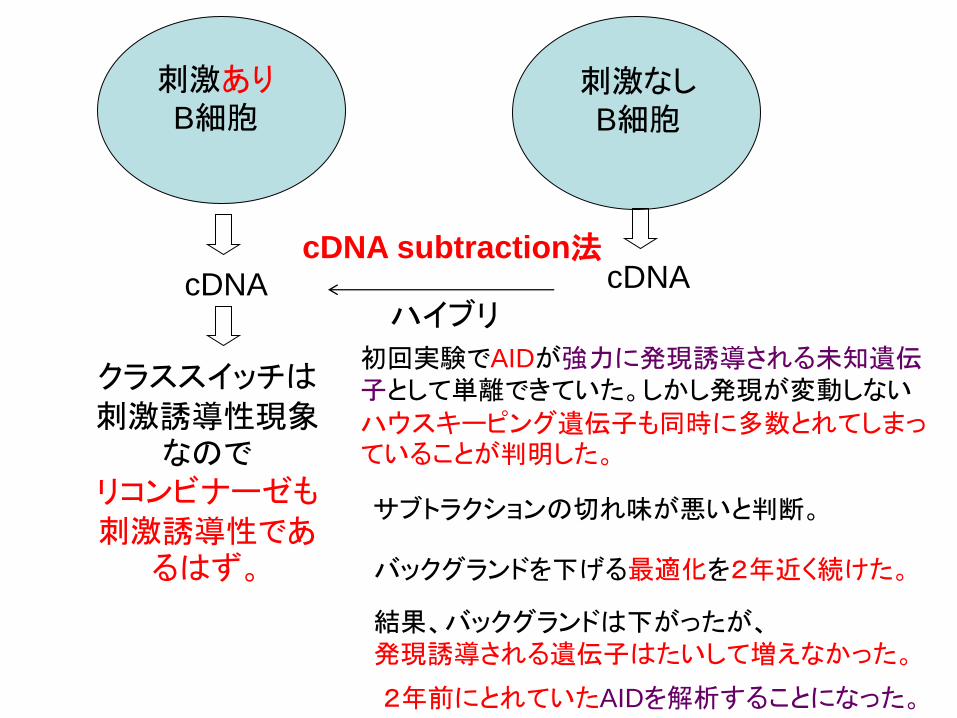
刺激あり
B細胞 刺激なし
B細胞
cDNA cDNA
ハイブリ
クラススイッチは
刺激誘導性現象なので
リコンビナーゼも
刺激誘導性であるはず。
初回実験でAIDが強力に発現誘導される未知遺伝子として単離できていた。しかし発現が変動しない
ハウスキーピング遺伝子も同時に多数とれてしまっていることが判明した。
cDNA subtraction法
結果、バックグランドは下がったが、
発現誘導される遺伝子はたいして増えなかった。
サブトラクションの切れ味が悪いと判断。
バックグランドを下げる最適化を2年近く続けた。
2年前にとれていたAIDを解析することになった。

これまでに失敗や経験から学んだ事
1, 方法論に凝りだすと目指すものを忘れがち!
何が最終目標か常に意識すべし!
2, がむしゃらにサイエンスに打ち込んでください。
その姿勢はいずれは評価されます。 経験 2

これまでに失敗や経験から学んだ事
1, 方法論に凝りだすと目指すものを忘れがち!
何が最終目標か常に意識すべし!
2, がむしゃらにサイエンスする姿勢は
必ず評価される。

Acknowledgements
分子遺伝研究室のみなさん
産婦人科の共同研究者 M. Nakamura, S. Kyo, and H. Fujiwara
耳鼻咽喉、頭頸部外科の共同研究者
S. Kondo, T. Yoshizaki
金沢大学