centro de investigación en alimentación y desarrollo, a.c. · al consejo nacional de ciencia y...
TRANSCRIPT

Centro de Investigación en Alimentación y Desarrollo, A.C.
EVALUACIÓN DE UN SISTEMA CERRADO PARA EL CULTIVO DE CAMARÓN BLANCO Litopenaeus
vannamei
POR:
CLAUDIA LIZETH LARA ESPINOZA
TESIS APROBADA POR LA:
COORDINACIÓN DE CIENCIA DE LOS ALIMENTOS
Como requisito parcial para obtener el grado de
MAESTRÍA EN CIENCIAS
Hermosillo, Sonora Septiembre de 2012

ii

iii

iv
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología, por haberme brindado el apoyo económico para realizar mis estudios de posgrado.
Al Centro de Investigación en Alimentación y Desarrollo, A.C. por
abrirme las puertas y darme la oportunidad de crecer profesionalmente y de ampliar mis conocimientos.
A la M.C. María del Carmen Bermúdez Almada por recibirme con los
brazos abiertos y darme la oportunidad de formar parte de este proyecto y darme siempre su apoyo y disponibilidad.
A la M.C. Angelica Espinosa Plascencia, por ser un gran modelo a
seguir que siempre estuvo conmigo en las buenas y en las malas, por darme siempre el apoyo y la seguridad de que todo iba a salir bien y a tiempo, por todos sus comentarios, observaciones y por enseñarme y capacitarme durante la realización del estudio.
A la Granja Acuícola La Borbolla, especialmente al Ing. César Eduardo
Patiño Patiño, al Biol. Adolfo Pérez Álvarez y a los Técnicos Juan Carlos Gastélum Domínguez, Luis Elguezabal y Ángel Palomares por todo el apoyo brindado y por todos los conocimientos aportados durante mi estancia en la granja. Gracias a ustedes fue más amena mi estancia. También agradezco a la Sra. Guadalupe Leal Noriega por consentirnos con las deliciosas comidas que nos preparaba y por ser una gran persona.
A mis asesoras las Doctoras Luz Vázquez Moreno y Evelia Acedo
Félix por sus atinadas observaciones y sugerencias que enriquecieron este trabajo.
A mi compañero y amigo Edgar Noris Rodríguez, quien me brindo
incondicionalmente todo su apoyo, tanto en el desarrollo de esta investigación en el Laboratorio de Análisis Biológicos como en nuestra estancia en la granja camaronícola. Amigo gracias!
Al Laboratorio de Biología Molecular de Plantas, en especial a la Dra.
Marisela Rivera Domínguez y a la Q.B. Karen Rosalinda Astorga Cienfuegos, por el apoyo técnico recibido y por compartir conmigo su experiencia en técnicas moleculares, que fue una parte muy importante de este trabajo, así como por la disposición que siempre me brindaron para utilizar el equipo,

v
reactivos y espacio en el laboratorio. Muchas gracias, por toda su ayuda y paciencia.
Al Laboratorio de Biotecnología Marina en especial al Dr. Francisco
Vargas Albores por proporcionarnos algunos de los reactivos utilizados en esta investigación y muy especialmente al cDr. Enrique Villalpando Canchola, por su amable disposición y todas las atenciones recibidas durante el desarrollo de una parte de la tesis, permitiéndome utilizar el equipo necesario para llevar a buen termino este trabajo. También quiero agradecer el apoyo incondicional de mi compañera y amiga M.C. Fabiola Valenzuela González, que fue de gran ayuda. Gracias por todos tus consejos y conocimientos compartidos conmigo.
A la Dra. Lorena Olivia Noriega Orozco, por apoyarnos con la donación
de las cepas de Vibrio utilizadas en esta investigación. Al Laboratorio de Genética y Biología Molecular de Plantas, en
especial a la Dra. María Auxiliadora Islas Osuna, la Dra. Carmen Contreras Vergara y al M.C. Angel Javier Ojeda Contreras por permitirme el uso del equipo de laboratorio y por su gran disposición.
A mi compañera Cindy Melina Colosio López por el apoyo técnico
brindado en el laboratorio y por la paciencia que me tuvo durante mi capacitación en técnicas microbiológicas.
Al Laboratorio de Inmunología y Microbiología en especial a la Dra.
Verónica Mata Haro por permitirme trabajar en su laboratorio y utilizar el equipo bajo su responsabilidad. Además, quiero agradecerle el compartir conmigo sus conocimientos, gracias por sus comentarios y sugerencias.
Al Laboratorio de Microbiología Molecular, en especial a la Q.B.
Rosalva Pérez Morales y Germán Jacquez Muñoz, por su disposición y apoyo técnico recibido durante mi capacitación en el uso del equipo del laboratorio.
Al Laboratorio de Bioquímica de Proteínas de Estrés en especial a la
M.C. Ciria Guadalupe Figueroa Soto por permitirme hacer uso del equipo de laboratorio.

vi
DDDDEDICATORIASEDICATORIASEDICATORIASEDICATORIAS
A DiosDiosDiosDios por ser esa luz que me ha guiado y protegido a lo largo de mi vida.
A mis padresmis padresmis padresmis padres José y Lupita por darme siempre su apoyo incondicional, por ser un gran
ejemplo a seguir y ayudarme a ser una mejor persona, por enseñarme a luchar para
seguir adelante, gracias!
A mis hermanosmis hermanosmis hermanosmis hermanos José Alberto, Emmanuel y Marco por estar siempre a mi lado, por
compartir tantos bonitos momentos conmigo que a lo largo de nuestras vidas hemos
pasado.
A mimimimi sosososobrinitobrinitobrinitobrinito que viene en camino que con tantas ansias espero, que ya hacía falta en
la familia para alegrarnos más la vida.
A mimimimi gran amigagran amigagran amigagran amiga Karla por darme siempre su apoyo, por abrirme las puertas de su
hogar y brindarme incondicionalmente su amistad.
A mi mi mi mi novionovionovionovio Juan Carlos por estar a mi lado y darme todo su amor y cariño en las
buenas y en las malas, por todo el apoyo y ánimo que me ha brindado, amor gracias!
A mis amigomis amigomis amigomis amigossss en especial a Anna, Marina, Max, Manolo “el plebe” por hacer mas
amena y divertida mi estancia en el CIAD y por brindarme su amistad incondicional a
lo largo de estos dos años, amigos gracias!

vii
CONTENIDO
Página
LISTA DE FIGURAS9999999999999999999999999...x
LISTA DE TABLAS 99999999999999999999999999xi
RESUMEN.99999999999999999999999999999.xii
ABSTRACT99999999999999999999999999999xiii
INTRODUCCIÓN999999999999999999999999999.1
ANTECEDENTES99999..999999999999999999999.3
La Problemática de la Camaronicultura en México9999999999993
Generalidades de las Bacterias del Género Vibrio99999.99999994
Características Fisiológicas y/o Morfológicas de la Familia Vibrionacea99999999999999999999999999..9..6
Aislamiento de Bacterias del Género Vibrio9999999999..999..7
Reacción en Cadena de la Polimerasa (PCR) para la Identificación de bacterias de Vibrio9999999999999..9999999...............8
Factores Abióticos que Intervienen en el Desarrollo del Camarón Blanco Litopenaues vannamei999999999999999999999........10
Temperatura9.999999999999999999999999..910
Salinidad999999999999999999999999999..911
Oxígeno Disuelto (OD)999999999999999999999.9.11
Potencial de Hidrógeno (pH)9999999999999999..999..12
Concentración de Nitrógeno Amoniacal Total (TAN)9999.99999..13
Concentración de Nitritos (NO2)999999999999999999..14
Concentración de Iones Minerales en el Agua del Estanque de Cultivo99.16
Calcio99999999999999999999999999999...18
Magnesio99999999999999999999999.9999919
Potasio9999999999999999999999999999.919
Uso de Antibióticos en la Acuicultura99999999999999999.21

viii
Características de Oxitetraciclina9999999..9999999999..22
Desarrollo de Resistencia Bacteriana por el Uso de Oxitetraciclina9..9923
Uso de Sistemas Cerrados para el Cultivo de Camarón999999999.24
HIPÓTESIS9999999999999999999999999999927
OBJETIVOS9999999999999999999999999999...28
Objetivo General..............................................................................................28
Objetivos Específicos9999...9999999999999999999.28
MATERIALES Y MÉTODOS999999999999999999999...29
Diseño Experimental999999...99999999999999999..29
Medición de Parámetros Fisicoquímicos9999...9999999999930
Determinación de Ganancia de Peso en Camarón99..99999999...30
Determinación de la Sobrevivencia en Camarón99...999999999..30
Concentración de los Iones Ca, Mg y K, Nitrógeno Amoniacal Total (TAN) y Nitritos (NO2) en el Agua del Estanque de cultivo9..999931
Determinación de Calcio99999999999999999..9999.31
Determinación de Potasio999999999999999..99999...32
Determinación de Magnesio...99999999999999..9999...32
Nitrógeno Amoniacal Total999.99999999999999999..33
Nitritos (NO2)9999999..9999999999999999999.33
Diagnóstico para la Detección de Lesiones Externas e Internas en Camarón.999999999999999999..9999999999..34
Análisis Bacteriológico en los Órganos del Camarón9999999999..36
Extracción de Hemolinfa999999.999999999999999..36
Extracción de Hepatopáncreas y Branquias de Camarón9999..999.37
Aislamiento de Bacterias del Género Vibrio9.99999.999999938
Detección de Especies de Vibrio Aisladas de Camarón Litopenaeus vannamei9999999999999999..999999...39
Extracción de ADN a partir de Aislamientos Bacterianos de Tejidos de Camarón99.999999999999999999999999...39
Determinación de la Concentración de ADN9.99999999999940
Reacción en Cadena de la Polimerasa (PCR)999999.999999.40
Electroforesis en Gel de Agarosa99999999..999999999..41

ix
Determinación de las Concentraciones Mínimas Inhibitorias (CMI) en Bacterias de Vibrio999999999..999999999999999..43
Análisis Estadístico9999999999999..99999999999.45
RESULTADOS Y DISCUSIÓN99999999999999999999...46
Parámetros Fisicoquímicos9999999999..9999999999...46
Determinación de Ganancia de Peso y Sobrevivencia en Camarón99...9.47
Concentración de Iones (Ca, Mg y K) en el Agua del Estanque99999...48
Concentración de Nitrógeno Amoniacal Total (TAN) y Nitritos (NO2) en el Agua del Estanque9.99999999999999999999999...51
Evaluación de Lesiones Internas y Externas en Camarón Litopenaeus vannamei99999999999999999999999......................54
Análisis Bacteriológico (Branquias, Hemolinfa y Hepatopáncreas) en Camarón Litopenaeus vannamei 99..999999999999999955
Cuenta Total Bacteriana en Tejidos de Camarón999999999..9..56
Recuento de Bacterias del Género Vibrio Aisladas de los Distintos Tejidos de Camarón 999.9.99999999999999...57
Detección de Especies de Vibrio Aisladas de Camarón de cultivo Litopenaeusvannamei...9999999999999999999999...59
Concentración Mínima Inhibitoria (CMI) de la Oxitetraciclina en Cepas de Vibrio Aisladas de Camarón999..99999999999999999..64
RESUMEN DE HALLAZGOS 99999999999999999999966
CONCLUSIONES99999999999999999999999999.67
BIBLIOGRAFÍA999999999999999999999999999.68

x
LISTA DE FIGURAS
Figura Página 1. Ciclo del Nitrógeno en un Estanque de Cultivo999999999999...16
2. Determinación de Iones en el Agua de Mar99999999999999.33
3. Procedimiento para la Extracción de Hemolinfa de Camarón Litopenaeus vannamei99999999999999999999999999999937
4. Extracción de los Órganos de Camarón Litopenaeus vannamei99999..38
5. Concentración de los Iones Ca, Mg y K en el Agua del Estanque Durante La Evaluación del Sistema Cerrado de Cultivo de Camarón.999949
6. Comportamiento de Nitrógeno Amoniacal Total (TAN) y Nitritos (NO2) Durante la Evaluación del Sistema para el Cultivo de Camarón99.................52
7. Amplificación mediante PCR del gen 16s RNA999999999999...60
8. Amplificación mediante PCR del gen de hemolisina en cepas de Vibrio.......61
9. Amplificación mediante PCR del gen toxR en cepas Vibrio9..9999.9...62

xi
LISTA DE TABLAS
Tabla Página
1. Criterios para Establecer el Grado de Severidad de las Lesiones en los Distintos Órganos de Camarón99999999999999999999..35
2. Condiciones de PCR Para Cada uno de los Oligonucleótidos Iniciadores Utilizados en la Identificación de Especies de Vibrio999999..42
3. Distribución de las Muestras en la Placa Para la Determinación de la Concentración Mínima Inhibitoria de la Oxitetraciclina en Bacterias de Vibrio...44 4. Parámetros Fisicoquímicos Evaluados en el Sistema Cerrado para el Cultivo de Camarón L. vannamei����..9999999999999999..9946 5. Evaluación del Daño en los Distintos Órganos de Camarón Litopenaeus vannamei Cultivado en un Sistema Cerrado99999999999999955 6. Cuenta Total Bacteriana en Tejidos de Camarón Litopenaeus vannamei9..56
7. Evaluación de Bacterias de Vibrio Sacarosa (+) Aisladas de Diferentes Tejidos de Camarón Litopenaeus vannamei999999999...9999.957 8. Concentración Mínima Inhibitoria de la Oxitetraciclina en Bacterias de Vibrio Aisladas de los Diferentes Tejidos de Camarón999999999.65

xii
RESUMEN
La camaronicultura es una actividad industrial importante para el estado
de Sonora. La producción de camarón en sistemas tradicionales disminuyó drásticamente en los tres últimos años, debido a la alta tasa de mortalidad de los organismos en cultivo, provocada por infecciones virales y bacterianas, lo que ocasionó pérdidas económicas cuantiosas. En esta investigación se estudió el desarrollo de camarón blanco L. vannamei en un Sistema de Cultivo Cerrado, evaluando aspectos fisicoquímicos, fisiológicos y microbiológicos. Se realizó un bioensayo durante 58 días en una granja comercial ubicada en Bahía de Kino, Sonora, empleando un estanque tipo invernadero de 0.04 Ha, sembrado con 126 organismos/m2. Se determinaron parámetros fisicoquímicos, como temperatura, oxígeno disuelto (OD), pH, salinidad y nitritos, así como la severidad de lesiones en los órganos del camarón, cuenta total bacteriana y de Vibrio en hemolinfa, hepatopáncreas y branquias y la sensibilidad a oxitetraciclina por dilución en microplacas. Además, se realizó la identificación de la especie por técnicas moleculares (PCR) en las cepas bacterianas de Vibrio aisladas de los tejidos de camarón. Los valores obtenidos en los parámetros fisicoquímicos del agua del estanque fueron: temperatura 30.44±1.0°C, OD 4.71±0.64 mg/L, pH 7.34±0.15 y salinidad 39.90±0.88‰. Los niveles de nitritos en el agua estuvieron por arriba del valor recomendado (0.01 mg/L), encontrándose en promedio una concentración de 4.84±4.40 mg/L, observando en los camarones una disminución en las mudas, aparición de cromatóforos en el intestino y necrosis en la cutícula. La concentración de iones en el agua fue de 168.80±14.10, 920.00±339.40 y 88.31±22.70 mg/L para Ca, Mg y K, respectivamente. Los camarones tuvieron un incremento en peso de 1.95±0.64 g/semana y una sobrevivencia del 74%. No se detectaron lesiones severas en los órganos de los camarones al término del bioensayo. Las cuentas bacterianas se mantuvieron por debajo de los niveles recomendados (5x103 UFC/g). La Concentración Mínima Inhibitoria para la oxitetraciclina fue de 0.39-100 µg/mL en bacterias aisladas de hepatopáncreas, 0.78-325 µg/mL en branquias y 0.78-75 µg/mL en hemolinfa. Se detectó mediante la técnica de PCR la presencia de bacterias del género Vibrio spp. y Vibrio harveyi, aisladas de los distintos tejidos de camarón. Los resultados obtenidos mostraron que el Sistema de Cultivo Cerrado permitió un mayor control de los parámetros evaluados, lo que favoreció el desarrollo de los organismos, por lo que se puede proponer este sistema como una alternativa viable para el cultivo de camarón Litopenaeus vannamei en la región. Palabras clave: camarón, Litopenaeus vannamei, sobrevivencia, Sistema de Cultivo Cerrado, oxitetraciclina.

xiii
ABSTRACT
Shrimp farming is an important industrial activity in the state of Sonora. Shrimp production in culture under traditional systems decreased in the last three years, due to the high mortality rate of the organisms, caused by viral and bacterial infections, causing substantial economic losses. In this research we studied an alternative of Shrimp production under a closed culture measuring a series of parameter physicochemical, physiological and microbiological. Bioassay was performed for 59 days in a commercial farm in Bahia de Kino, Sonora, using a type greenhouse pond of 0.04 ha, to a density of 126 organisms/m3. Measures included in this study were temperature, dissolved oxygen (DO), pH, salinity and nitrite, and the severity of organ damage shrimp, total bacterial count and Vibrio in hemolymph, hepatopancreas and gills and sensitivity to oxytetracycline by dilution on microplates. In addition, we also performed the species identification by molecular techniques (PCR) from bacterial strains Vibrio isolated from tissues of shrimp. Physicochemical parameters related to the pond water were temperature 30.44 ± 1.0 ° C, DO 4.71 ± 0.64 mg / L, pH 7.34 ± 0.15 and 39.90 ± 0.88 ‰ salinity. Nitrite levels in the water were 4.84±4.40 mg/L which was above the recommended value of 0.01 mg/L, this results were a decline in shrimp molts, chromatophores in the intestine and necrosis in the cuticle. The concentration of ions in water was 168.80±14.10, 920.00±339.40 and 88.31±22.70 mg/L for Ca, Mg and K, respectively. Furthermore, shrimp had an increase in weight of 1.95±0.64 g/week and a survival rate of 74%. No serious damage were detected in the organs of the shrimp at the end of the study. The bacterial counts were below of recommendations (5x103 CFU/g). The minimum inhibitory concentration for oxytetracycline was 0.39 to 100 µg/mL in bacteria isolated from hepatopancreas, from 0.78 to 325 µg/mL in gills and from 0.78 to 75 µg/mL in hemolymph. The species of bacteria detected from different tissues of shrimp were Vibrio spp. and Vibrio hemolisin gen and toxR. Given that closed culture system allowed greater control of the parameters evaluated and a better development of organisms, this system can be proposed as a viable alternative for Litopenaeus vannamei shrimp farming in the region.
Keywords: shrimp Litopenaeus vannamei, survival, closed culture system, oxytetracycline.

1
INTRODUCCIÓN
La acuacultura provee el 45 % de los productos acuícolas a nivel
mundial, siendo el camarón, el producto que mayormente se comercializa,
generando una producción mundial de aproximadamente seis millones de Ton
(FAO, 2009). En México, se ha registrado un crecimiento constante en la
industria camaronícola, representando una de las principales fuentes
alimenticias tanto para consumo nacional como de exportación. Actualmente, es
una de las actividades productivas más importantes en el estado de Sonora,
siendo el camarón de la especie Litopenaeus vannamei el mayormente
cultivado, debido al amplio conocimiento que se tiene en relación a su cultivo y
manejo. Sin embargo, una de las principales limitantes en el crecimiento y
sustentabilidad de esta actividad industrial, es la presencia de enfermedades,
tanto virales como bacterianas, en los sistemas de cultivo (COSAES, 2010).
En relación a las enfermedades bacterianas que afectan los cultivos de
camarón, las infecciones ocasionadas por bacterias del género Vibrio son las
más comunes, debido a que habitan de manera natural el ambiente marino y
son consideradas patógenos oportunistas que pueden infectar al camarón en
cualquiera de sus etapas de desarrollo (González, 2003). Ante esto, es
necesario aplicar medidas de bioseguridad y un monitoreo constante para la
detección de bacterias patógenas (Galaviz et al., 2007).
Debido a la difícil situación que representa mantener un control sobre las
enfermedades que afectan los cultivos de camarón, es que se plantea el uso de
sistemas de producción cerrados, como una alternativa viable para el cultivo de
camarón Litopenaeus vannamei. Estos sistemas proporcionan un ambiente
mejor controlado para promover el crecimiento de los organismos acuáticos,
mediante el monitoreo de parámetros que incluyen concentraciones de oxígeno
disuelto (OD), nitrógeno amoniacal total (TAN), nitritos, dióxido de carbono

2
(CO2), temperatura, potencial de Hidrógeno (pH) y los niveles de alcalinidad en
el sistema. Además, tienen un bajo impacto ambiental ya que se reutiliza el
agua, previo tratamiento físico, químico y biológico. También, se emplea menos
del 10 % del agua requerida en un sistema de producción convencional. Por
esta razón, desde hace más de tres décadas, se han realizado investigaciones
relacionadas con sistemas de cultivo cerrados o con recirculación de agua en la
industria acuícola y aproximadamente una década en la producción intensiva de
organismos (Timmons et al., 2002).

3
ANTECEDENTES
La Problemática de la Camaronicultura en México
La camaronicultura es un área que ha tenido gran desarrollo desde
finales del siglo pasado. Sin embargo, conforme se ha incrementado la
producción de camarón por unidad de superficie en los sistemas intensivos, la
susceptibilidad de los organismos a contraer enfermedades también se ha
aumentado, debido principalmente al estrés que sufren por las altas densidades
de siembra, o bien por la pobre calidad del agua y los alimentos deficientes, lo
cual ha generado un colapso de esta industria en distintos países como, China,
Tailandia, Indonesia, Taiwán, Ecuador y México (Flegel, 2012).
Normalmente, cuando se habla de agentes patógenos causantes de
cuantiosas pérdidas en la industria camaronícola, se hace referencia a las
infecciones virales, como el Virus de la Mancha Blanca (WSSV), el Virus de la
Cabeza Amarrilla (YHV), Virus del Síndrome de Taura (TSV) entre otros, debido
a la gran mortalidad que pueden ocasionar en un período corto (Galaviz-Silva et
al., 2004), Sin embargo, existen otros patógenos que también causan daños
considerables en la producción de camarón, como las infecciones ocasionadas
por las bacterias del género Vibrio.
Distintas especies de Vibrio han sido reportadas a nivel mundial como las
causantes de grandes pérdidas en la producción de camarón cultivado, ya que
han generado mortalidades que alcanzaron hasta el 100% de los organismos,
particularmente en la etapa postlarvaria y juvenil (Aguirre-Guzmán, 2004).
También, otras bacterias causan enfermedades en camarones marinos
(Álvarez et al., 2000; Rendón y Balcázar, 2003), como la bacteria intracelular
tipo rickettsia causante de la Hepatopancreatitis Necrotizante (NHP), que se

4
caracteriza por la inflamación y necrosis del hepatopáncreas de los camarones.
Estas bacterias atacan las células del epitelio del hepatopáncreas, provocando
en este órgano una coloración pálida, las células hepatopancreáticas aparecen
hipertrofiadas y con gran cantidad de bacterias en el citoplasma (Gómez-Gil et
al., 2001; Morales, 2004).
La infección denominada erosión bacteriana del caparazón se presenta
en todas las especies de peneidos, manifestándose por la aparición de
manchas cafés o negras ocasionadas por bacterias quitinolíticas. Esta
infección se ha asociado a Vibrio sp., Aeromonas sp., Spirillum sp. y
Flavobacterium sp., representando una amenaza para los camarones en cultivo,
cuando estos se encuentran en condiciones de estrés severo (Morales, 2004;
Santiago, 2009).
Esta problemática ha hecho necesario mantener monitoreos constantes
de la población bacteriana, para evitar devaluar la calidad del producto. Para
ello, la identificación bacteriana por medio de pruebas de diagnóstico,
determinan las características coloniales de las bacterias, generando
información que ayude a mantener un control sobre la proliferación de los
microrganismos y así lograr aplicar tratamientos preventivos efectivos
(Valenzuela-Salcedo et al., 2008).
Generalidades de las Bacterias del Género Vibrio
Las bacterias que conforman el género Vibrio se caracterizan por habitar
naturalmente el medio marino (Soto-Rodríguez et al., 2010). Estas bacterias
juegan un importante papel dentro del ciclo de nutrientes, mediante la
remineralización de nutrientes orgánicos como carbono (C), nitrógeno (N) y
fósforo (P) (Pruzzo et al., 2005; Leyton y Riquelme, 2008), sin representar un
riesgo para la salud de los organismos en cultivo, excepto cuando éste posee
alguna deficiencia nutricional, inmunológica o genética, y/o que esté siendo
cultivado bajo un estrés ambiental ocasionado por la pobre calidad del agua,

5
una elevada densidad de siembra, una alta temperatura del agua, baja
concentración del oxígeno disuelto y una baja tasa de recambio de agua
(Aguirre-Guzmán, 2004; Venkateswara-Rao, 2009).
Las bacterias del género Vibrio son patógenos oportunistas que pueden
tomar ventaja de los cambios ecológicos generados durante el cultivo,
provocando infecciones. También, forman parte importante de la población
bacteriana del camarón y del medio ambiente que lo rodea, convirtiéndose por
lo tanto, en una posible y constante fuente de infecciones para estos
organismos (Aguirre-Guzmán, 2004). La severidad de la infección en el
camarón dependerá de la especie y la cepa de Vibrio involucrada, el estado de
desarrollo y edad del camarón, así como de las condiciones ambientales
(Jayasree et al., 2006).
Las enfermedades generadas por estas bacterias han sido descritas
como: Vibriosis, Enfermedad Bacterial, Septicemia Bacteriana de los Peneidos,
Vibriosis de los Peneidos, Vibriosis Luminiscente y Enfermedad de las Patas
Rojas. Los camarones afectados por alguna especie patógena de Vibrio spp.,
muestran distintos signos de la enfermedad como letargia, intestino semi-vacío,
anorexia, flexión dorsal abdominal, fuerte colonización bacteriana en la cutícula,
opacidad en el músculo abdominal, cromatóforos visibles en la base de las
apéndices, nódulos melanizados en el hepatopáncreas, retardo en la
coagulación de la hemolinfa y reducción de hemocitos, esto principalmente en
la etapa larvaria (López, 1998).
Además, se puede observar un gran número de bacterias en la
hemolinfa durante una septicemia bacteriana, mediante la realización de
análisis microscópicos en fresco. Necrosis, granulomas, inflamación de órganos
(órgano linfoide, branquias, corazón, hepatopáncreas, etc.) son fáciles de
evaluar por medio de técnicas histológicas.
Las especies del género Vibrio que han sido involucradas en las
enfermedades que se presentan en el camarón son: Vibrio harveyi, V.
parahaemolyticus, V. cholerae, V. vulnificus, V. alginolyticus, V. damselae
(actualmente Photobacterium damselae), V. penaeicida, V. anguillarum, V.

6
nereis, V. campbelli, V. tubiashi y V. fluvialis, las cuales pueden afectar
cualquiera de las etapas de desarrollo del camarón (Aguirre-Guzman, 2004;
Kitiyodom et al., 2010).
Algunas especies de Vibrio expresan genes de toxicidad como el gen
toxR el cual fue descubierto primero como un regulador transcripcional positivo
que codifica la toxina de Vibrio cholera (Pang et al., 2006). Otro gen que puede
estar presente en las especies de Vibrio es el gen de hemolisina, en general
las bacterias que expresan este gen, son consideradas patógenas ya que
causan hemorragias, septicemia y diarrea en los hospederos. Algunos tipos de
hemolisinas se han caracterizado y los genes que las codifican han sido
clonados en Vibrio parahaemolyticus, V. cholera, V. hollisae, V. mimicus, V.
vulnificus y V. anguillarum (Zhang et al., 2001).
Los problemas que implican los brotes por bacterias del género Vibrio en
la industria del cultivo de camarón, ha encaminado a la búsqueda de nuevas
estrategias para prevenir brotes de enfermedad. Éstas incluyen el uso de la
llamada “agua-verde”, aplicación de probióticos, uso de reservorios y cloración
de estanques durante el periodo de cultivo (Caipang y Aguana, 2011).
Características Fisiológicas y/o Morfológicas de la Familia Vibrionacea
El género Vibrio ésta conformado aproximadamente por 135 especies
(http://www.vibriobiology.net), caracterizadas por ser bacilos Gram (-), con
movilidad por un flagelo polar, siendo la mayoría de ellas oxidasa (+) y no
producen esporas (Dikow, 2011). Son bacterias facultativas anaerobias,
teniendo ambos metabolismos respiratorio y fermentativo, reducen nitrato a
nitrito, fermentan D-glucosa la cual es utilizada como su única o principal fuente
de carbono y usan el amonio como su única fuente de nitrógeno. La mayoría de
las especies de este género bacteriano requieren sodio (Na) para crecer y
requieren de 0.5-3% de NaCl para tener un óptimo crecimiento. Todas las

7
especies de Vibrio excepto V. cholerae y V. mimicus requieren de Na para su
desarrollo (Pruzzo et al., 2005; Caipang y Aguana, 2011).
La mayoría de las bacterias de Vibrio pueden crecer en condiciones
levemente alcalinas, de preferencia en un intervalo de pH entre 7 y 8. Algunas
especies, incluyendo V. cholerae y V. metschnikovii pueden crecer a pH de 10.
En relación a sus requerimientos de temperatura, se puede decir que la
mayoría de las especies de Vibrio crecen adecuadamente a una temperatura de
18-22°C. Pero también existen algunas especies de este género que pueden
crecer a temperaturas de 0-4°C, mientras que otras pueden desarrollarse
perfectamente a temperaturas por arriba de los 45°C (Soto et al., 2009).
Aislamiento de Bacterias del Género Vibrio
Existen diversos medios de cultivo para el aislamiento de las bacterias
pertenecientes a este género, entre los cuales destacan el Agar Marino, el cual
es un medio no selectivo, donde casi todas las cepas de Vibrio pueden crecer y
es el más utilizado para la determinación de cuentas totales de Vibrio y
protobacterias. También es útil para el aislamiento y crecimiento de organismos
cuyo hábitat natural son ambientes marinos u otros ambientes con un alto
contenido de sodio (Na).
El agar Tiosulfato-Citrato-Bilis-Sacarosa (TCBS) es el medio de cultivo
más ampliamente utilizado para el aislamiento de cepas de Vibrio provenientes
del ambiente. Aunque no se debe descartar que en este medio de cultivo
también pueden crecer otras bacterias como Pseudomonas spp. y Aeromonas
spp. Se ha observado que cuando las bacterias de Vibrio utilizan la sacarosa
como fuente de carbono producen ácido, apareciendo las colonias en la placa
de cultivo de color amarillo (Vibrio alginolyticus, V. cholerae, V. angillarum, V.
fluvialis). Las bacterias de Vibrio que crecen sin utilizar sacarosa aparecen de
color verde (Vibrio parahaemolyticus, V. damsella, V. fisheri, V. mimicus, V.
bollisea, V. harveyi) (Ibarra et al., 2004).

8
Otro medio de cultivo que puede ser utilizado para el aislamiento y
crecimiento de este género es el agua peptonada alcalina. Ésta es utilizada
como medio de enriquecimiento para Vibrio cholerae y otras especies de Vibrio,
que a menudo crecen mejor que los microorganismos no deseados, empleando
un pH alto. La cantidad de NaCl utilizada para la preparación de agua
peptonada alcalina no está estandarizada pero usualmente está en el intervalo
de 0.5-1% y puede incrementar hasta 2% para mejorar el crecimiento de las
cepas de Vibrio aisladas del medio marino. Si el contenido de NaCl en el medio
de cultivo se omite, se hace más selectivo para el crecimiento de las especies
como V. cholerae y V. mimicus (Soto et al., 2009; Nishibuchi y DePaola, 2005).
Reacción en Cadena de la Polimerasa (PCR) para la Identificación de Bacterias de Vibrio
Los avances en la instrumentación y los descubrimientos en bioquímica y
biología molecular han permitido generar información genética de los
microrganismos, la cual, es utilizada como una herramienta para la
identificación y cuantificación de las bacterias que son de interés para el
hombre. Entre estos métodos destacan la Reacción en Cadena de la
Polimerasa (PCR), la cual ha sido aplicada en el diagnóstico de ciertas
infecciones en camarones peneidos (Olsen, 2000).
El descubrimiento de la PCR ha permitido la amplificación selectiva de
secuencias específicas de ADN que se encuentran en cantidades muy
pequeñas. Como su nombre lo indica, esta técnica analítica se basa en la
actividad de la enzima ADN polimerasa que es capaz de sintetizar una cadena
de ADN complementaria a otra ya existente. El método de PCR utiliza dos
secuencias cortas de oligonucleótidos llamadas iniciadores o cebadores que
son complementarios a los extremos de la región de ADN que se pretende
amplificar. La reacción incluye ADN que puede ser en varias formas, por
ejemplo, ADN purificado a partir de un lisado de tejido, iniciadores, la enzima
polimerasa y nucleótidos para formar nuevas copias. En cada ronda de la

9
reacción en un termociclador, la muestra de ADN es desnaturalizada, los
iniciadores se alinean a sus regiones complementarias y la enzima polimerasa
cataliza la adición de los nucleótidos al final de cada primer, creando así,
nuevas copias de la región blanco (McBeath et al., 2000).
Las principales ventajas del uso de PCR en la detección e identificación
de microrganismos radica en tres aspectos fundamentales: su sensibilidad,
especificidad y capacidad de procesamiento de un gran número de muestras,
además la obtención de los resultados es rápida (2 a 3 días) y no es necesario
el uso de material radioactivo (Rojas-Herrera y González-Flores, 2006).
Adicionalmente este método identifica microrganismos que no pueden
ser estudiados por técnicas convencionales o que no pueden cultivarse en
substratos artificiales.
Entre los factores que limitan la aplicación de la técnica de PCR para la
identificación de microrganismos patógenos en muestras de alimentos, destaca
la presencia de sustancias que pueden ejercer un efecto inhibitorio sobre la
reacción y pueden actuar a diferentes niveles durante el proceso de extracción
y amplificación de los ácidos nucleicos y eventualmente, conducir a la obtención
de falsos negativos (Rojas-Herrera y González-Flores, 2006).
Entre los compuestos que ejercen un efecto negativo, se encuentran la
hemoglobina, la lactoferrina, los polisacáridos, las grasas, proteínas y los iones
metálicos, presentes en las matrices alimenticias. La interferencia de estos
compuestos puede ser eliminada empleando un proceso de pre-enriquecimiento
o mediante la utilización de enzimas apropiadas (polimerasas) (Gentry et al.,
2002).
El uso de PCR para la detección de bacterias patógenas es sumamente
importante ya que estos organismos habitan los ambientes marinos. Una
detección precisa y temprana de estos patógenos es un factor crucial para el
éxito de las operaciones de cultivo y la acuacultura (Cunningham, 2002; Tarr et
al., 2007).

10
Factores Abióticos que Intervienen en el Desarrollo del Camarón Blanco Litopenaeus vannamei
El camarón de cultivo L. vannamei requiere ciertas condiciones
ambientales específicas como, temperatura, pH, oxígeno disuelto,
concentración de amonio y salinidad para un desarrollo óptimo. Por ello, es
importante medir y controlar dichos parámetros durante su ciclo de cultivo. Los
cambios drásticos en estos parámetros fisicoquímicos pueden causar estrés en
el camarón y favorecer la proliferación de patógenos, los cuales podrían
provocar infecciones, un lento crecimiento de los organismos o la muerte,
reflejándose esto en pérdidas económicas cuantiosas para el sector productivo
camaronícola (Venkateswara-Rao, 2009).
Temperatura
La temperatura es uno de los parámetros más importantes para el
desarrollo y sobrevivencia del camarón. Tiene un efecto significativo en el
metabolismo del organismo, principalmente en la actividad enzimática, consumo
de alimento y tasa de crecimiento. La temperatura óptima para el desarrollo del
camarón L. vannamei se modifica con la edad. De tal modo que los camarones
con un peso menor a 5 g requieren una temperatura de 30°C, mientras que los
de mayor peso requieren temperaturas cercanas a los 27°C (Cuzón et al., 2004;
Jiang et al., 2000).
Los camarones pueden soportar temperaturas menores a 18°C, pero
esto les ocasiona que dejen de alimentarse y entrar en un estado de letargo.
Temperaturas superiores a 30°C, podrían acelerar su metabolismo ocasionando
pérdida de peso, necesitando consumir más alimento balanceado. Además, los
camarones pueden mostrar un comportamiento poco común. La temperatura
máxima que puede asegurar la sobrevivencia de estos crustáceos es de 28°C,
siendo el intervalo ideal para su crecimiento de 27 a 31°C (Limsuwan, 2005).

11
Salinidad
La salinidad del agua es otro de los factores ambientales que debe
considerarse, ya que este parámetro determina la distribución de los
organismos en el sistema de cultivo. Cambios drásticos en la salinidad del agua
pueden alterar el crecimiento y la respuesta fisiológica de los organismos en
cultivo (Li et al., 2008; Sen et al., 2009).
Los camarones son organismos eurihalinos, esto es, que pueden
soportar variaciones amplias de salinidad, de tal modo que salinidades de 30 ‰
son apropiadas para la reproducción, eclosión y el desarrollo larvario, pero son
requeridas salinidades de 15 ‰ en la etapa postlarvaria y juvenil del camarón
(Auró y Ocampo, 2006).
La salinidad óptima para el crecimiento de las distintas especies de
crustáceos, dependerá de las diferencias en su capacidad osmótica (CO).
Especies con una alta hiper-CO crecen mejor en salinidades bajas, mientras
que especies con baja hiper-CO se desarrollan más en salinidades elevadas
(Brito et al., 2000).
El camarón de la especie L. vannamei tiene la capacidad de tolerar un
amplio rango de salinidad desde 0.5 hasta 50 ‰, convirtiéndola en una especie
particularmente adaptable, con un desarrollo óptimo a una salinidad de 20‰
(Roy et al., 2007; Li et al., 2007; Saoud et al., 2003)
Oxígeno Disuelto (OD)
El oxígeno disuelto en el agua es uno de los principales factores
causantes de estrés en la acuicultura, ya que tiene un efecto destacado en el
desarrollo de los organismos, afectando directamente en el consumo del
alimento y el metabolismo. Cambios bruscos en este parámetro afectan la
solubilidad y disponibilidad de algunos nutrientes (Venkateswara-Rao, 2009).

12
Gran parte del OD en el agua, proviene de la atmósfera y de la fotosíntesis. La
concentración de OD puede disminuirse por cambios de temperatura, la muerte
de fitoplancton dominante, la reproducción excesiva de zooplancton y la
descomposición o acumulación de materia orgánica en los estanques,
incluyendo el alimento no consumido y las heces (Jiang et al., 2005).
Los camarones del género L. vannamei requieren para su desarrollo
concentraciones de OD mayores a 3 mg/L, siendo la concentración óptima
cercana a los 5 mg/L. Un exceso de OD puede ser perjudicial y causar la
enfermedad conocida como “burbujas de gas”. (Venkateswara-Rao, 2009).
Potencial de Hidrógeno (pH)
En el agua de los estanques, los niveles de pH varían de 6.6 a 10.2
debido a la eliminación de dióxido de carbono (CO2), proveniente de la
fotosíntesis durante el día y de la liberación de CO2 por las plantas y animales
durante la noche. Una gran cantidad de organismos acuáticos se adaptan a un
valor específico de pH y pueden morir si éste cambia a un nivel mínimo de 4.
Los camarones en etapa juvenil son más sensibles a los cambios de pH y no
pueden sobrevivir dentro del agua con niveles extremadamente altos o bajos de
pH (9.6 o 4.5) (Li y Chen, 2008).
El pH es un parámetro químico de gran importancia ya que afecta el
metabolismo y otros procesos fisiológicos de los organismos cultivados. El
intervalo de pH 6.8-8.7 debe ser mantenido para un crecimiento y una
producción aceptable. Sin embargo, en cultivos semi-intensivos es mejor
mantener un intervalo entre 7.4-8.5. Los cambios de pH en los estanques está
principalmente influenciado por la concentración de CO2 y los iones en equilibrio
con este. El pH también puede ser alterado por: a) los ácidos orgánicos que son
producidos por bacterias anaeróbicas, b) los minerales ácidos como el ácido
sulfúrico, c) la aplicación de hidróxido de calcio (Venkateswara-Rao, 2009).

13
En los sistemas de cultivo de camarón L. vannamei es recomendable un
valor de pH ligeramente básico (alrededor de 7.5), similar al pH del protoplasma
de las células, lo que les permitirá a los organismos un mejor desarrollo en
todos los niveles tróficos (Venkateswara-Rao, 2009).
Concentración de Nitrógeno Amoniacal Total (TAN)
La mayor fuente de nitrógeno que ingresa a un sistema de cultivo
intensivo de camarón es a través del alimento, éste al ser ingerido por los
camarones pasa por procesos metabólicos, que le permiten la conversión en
biomasa y otra parte es excretada hacia el ambiente acuático (Sánchez y
Ching, 2003; Kuhn, 2010). El mecanismo de excreción del camarón es en forma
de compuestos nitrogenados totales, que están constituidos por el amoniaco
ionizado (NH4+) y el amoniaco no ionizado (NH3) (Chen, 2003; Isla, 2007). En
los crustáceos decápodos, el nitrógeno es excretado en mayor cantidad como
amonio (60-70%) y en pequeñas cantidades como aminoácidos, urea y ácido
úrico (Jiang et al., 2000).
La toxicidad del amonio en los organismos acuáticos se ha atribuido a la
forma química NH3, debido a su habilidad para difundirse rápidamente a través
de la membrana celular, mientras que la forma química NH4+ se considera no
tóxica o significativamente menos tóxica (Crab et al., 2007). La excreción de
amonio en los crustáceos es afectada por factores intrínsecos como la talla, el
estadio del ciclo de muda y por los factores extrínsecos como la temperatura, la
salinidad, pH y OD (Re et al., 2004; Pillay y Kutty, 2005).
Una de las principales reacciones de los organismos acuáticos, al
encontrarse en un medio con altos niveles de nitrógeno, puede ser el cese y/o
reducción de la alimentación, lo cual reduce la formación del amonio
metabólico, manifestándose efectos sub-letales en los organismos como la
reducción en la tasa de crecimiento y la sobrevivencia. Los resultados adversos
sobre el crecimiento pueden provenir de lo siguiente: (a) una absorción lenta o

14
reducida de oxígeno causada por daño en las branquias, (b) demanda de
energía adicional causada por el uso de vías alternativas de desintoxicación, (c)
perturbaciones en la regulación osmótica y (d) daños físicos en varios tejidos
(Arredondo et al., 2007).
Por ello, es importante monitorear continuamente los niveles de amonio
en el agua, de los estanques de cultivo, ya que es recomendable que los
valores no sean mayores a 1.2 y 6.5 mg/L, para postlarvas y juveniles
respectivamente, evitando de esta forma los efectos tóxicos que este
compuesto produce, obteniendo mejores resultados en el cultivo de camarón.
La subestimación de nitrógeno total derivado de la excreción, es un problema
potencialmente serio, principalmente en los sistemas cerrados con recirculación
de agua (Jiang et al., 2000). Entre los principales y más efectivos métodos
aplicados en los sistemas cerrados con recirculación de agua para la remoción
de nitrógeno amoniacal total (TAN) se encuentran: (1) el arrastre del aire,
proporcionando la denitrificación (2) el intercambio iónico y (3) la biofiltración,
siendo este último el mas frecuentemente utilizado en los sistemas cerrados por
su bajo costo, operación y mantenimiento (Franco et al., 2004; De la Mora et al.,
2003).
Concentración de Nitritos (NO2)
El amonio, es un compuesto derivado de la excreción de compuestos
nitrogenados. Mediante un proceso oxidativo llamado nitrificación son
convertidos a nitritos por la participación de las bacterias del género
Nitrosomonas y Nitrosolobus. Posteriormente, se transforman a nitratos, por la
acción de las bacterias del género Nitrobacter y Nitrosococcus (Figura 1). Las
bacterias nitrificantes viven de forma natural en los ambientes acuáticos
(Sánchez y Ching, 2003; Cervantes et al., 2000; Zhang et al., 2008).
Los nitritos pueden estar presentes en los sistemas de cultivo en niveles
tóxicos. Recientemente se ha puesto una mayor atención en estos compuestos,

15
ya que se han considerado contaminantes de los sistemas acuáticos. Durante el
proceso de nitrificación, los nitritos pueden aumentar su concentración en los
sistemas de cultivo camaronícolas, debido a un desbalance en las poblaciones
bacterianas encargadas de los procesos de nitrificación y denitrificación, aun
cuando se realicen recambios de agua frecuentes en los estanques. También,
la concentración de nitritos puede ser elevada en los cuerpos de agua que
reciben los efluentes nitrogenados de los distintos parques acuícolas (Frías-
Espericueta et al., 2000).
Generalmente, se detectan en los estanques de producción intensiva de
camarón, concentraciones altas de nitritos durante las estaciones de otoño y
primavera, que es cuando ocurren fluctuaciones en la temperatura, generando
una ruptura en el ciclo del nitrógeno, debido a la disminución del plancton y/o
actividad bacteriana, ya que éstos son los principales consumidores del amonio
presente en el agua del estanque (Sánchez y Ching, 2003).
El proceso de nitrificación ocurre más rápido a valores de pH de 7 a 8 y
temperaturas desde 25 hasta 35°C. Al disminuir la temperatura ambiental
ocurre una reducción en la actividad del plancton, ya sea en estanques con
fondo de tierra, recubierto con plástico y/o tipo invernaderos. Otros factores que
influyen son, la reducción de nutrientes, la presencia de un clima nublado, los
tratamientos químicos al agua o los recambios de agua con condiciones físico-
químicas diferentes a las del estanque de cultivo, pueden ocasionar que una
menor cantidad de amonio sea asimilado por las algas, recayendo esta función
en las bacterias nitrificantes existentes en el estanque de cultivo. Entre los
principales efectos tóxicos del NO2 destacan aquellos que tienen una relación
directa en el transporte de oxígeno, la oxidación de compuestos importantes y
aquellos que causan un daño a los tejidos como, branquias y otros órganos
internos (Arredondo et al., 2007; Tomasso, 2012).
El efecto de los nitritos en el crecimiento de los camarones
dulceacuícolas y marinos ha sido documentado por Armstrong et al. (1976) y
por Wickins (1976); quienes observaron que una concentración de nitritos de
1.8 y 6.4 mg/L puede provocar una reducción del 35 y 50%, en el crecimiento

16
de postlarvas de Penaeus indicus. Posteriormente, Chen et al. (1990),
determinaron que una concentración de 21.38 mg/L de NO2, redujo al 50% el
crecimiento de P. monodon.
Figura 1. Ciclo del nitrógeno en un estanque de cultivo
(Sánchez y Ching, 2003).
Concentración de Iones Minerales en el Agua del Estanque de Cultivo
La sobrevivencia y el crecimiento de los camarones en los estanques de
cultivo al aire libre es afectado por diversos factores como, el tipo de suelo, la
salinidad del agua, la especie del camarón o la salud de las postlarvas.
Además, uno de los factores de mayor importancia en el desarrollo de los
organismos es la adecuada concentración de los iones minerales en el agua,

17
debido a que éstos han sido relacionados con la sobrevivencia de los
camarones. Son esenciales para la transmisión de los impulsos nerviosos y
para las contracciones musculares, juegan un papel vital en el equilibrio ácido-
base corporal y consecuentemente regulan el pH de la sangre y otros fluidos
corporales, son constituyentes esenciales de las estructuras esqueléticas, tales
como huesos y cutículas. El agua marina utilizada para el cultivo de camarón
usualmente posee diferente composición iónica (Saoud et al., 2003).
La salinidad del agua depende de la concentración total de los iones
disueltos, que básicamente son siete, cuya concentración promedio en el agua
de mar es la siguiente: Na 10.5, Mg 1.45, Ca 400.0, K 370.0, Cloruro 19.0,
Sulfato 2.7 y Bicarbonato 142.0 mg/L. Entre los principales iones se encuentran
el Calcio (Ca), Magnesio (Mg) y Potasio (K) y cada uno de ellos juega un papel
fundamental en el desarrollo de los organismos en cultivo.
La salinidad promedio del agua de mar es de 34.5 ‰, disminuyendo
durante la temporada de lluvia y aumentando durante el verano, siendo en esta
época cuando el agua de mar tiene una mayor salinidad debido a la
concentración de los iones a causa del incremento en la temperatura del agua,
lo que provoca una mayor evaporación
(http://www.horcalsa.com/contents/content-files/revista1.pdf). En la etapa
postlarvaria es muy importante la capacidad osmoreguladora del camarón. Las
postlarvas PL15 y PL20 de camarón L. vannamei toleran mejor la baja salinidad
del agua que las postlarvas PL10. Este fenómeno ha sido descrito en otras
especies de peneidos, observándose que la tolerancia a la salinidad de las
postlarvas depende también de la especie de camarón. Un ejemplo de ello es la
especie de L. vannamei ecuatoriano el cual crece mejor a baja salinidad que la
misma especie cultivada en México (Saoud et al., 2003).
Por otro lado, la alcalinidad es la capacidad del agua para neutralizar
ácidos o aceptar protones. Ésta representa la suma de las bases que pueden
ser tituladas en una muestra de agua, dado que la alcalinidad de aguas
superficiales está determinada generalmente por el contenido de carbonatos,
bicarbonatos e hidróxidos, ésta se toma como un indicador de dichas especies

18
iónicas. El bicarbonato constituye la forma química de mayor contribución a la
alcalinidad. Históricamente, la alcalinidad ha sido utilizada como un indicador de
la productividad de lagos donde niveles de alcalinidad altos indicarían una
productividad alta y viceversa. Ésta debe ser superior a 75 mg/L en el agua de
los estanques. La alcalinidad es menor en estanques con suelos ácidos y baja
salinidad. Si la alcalinidad es menor a 40‰, se pueden provocar efectos en el
camarón, principalmente en el ciclo de muda. Si la alcalinidad es baja y se
combina con un pH menor a 7.5 se pueden presentar problemas de mortalidad
en el cultivo. Por otro lado, si la alcalinidad es elevada (200-300 ‰) y el pH es
mayor de 8.5 los organismos interrumpen su ciclo de muda (Chávez, 2009).
Calcio
El Ca es un elemento muy importante para el crecimiento y bienestar de
los camarones y necesario para la formación del nuevo exoesqueleto después
de la muda, endureciendo el caparazón del camarón. Este mineral es absorbido
a través del tracto gastrointestinal por transporte activo (Davis y Glatin, 1996) y
a través de las branquias de los crustáceos.
(http://www.fao.org/docrep/field/003/ab492s/AB492S04.htm).
El Ca estimula la contracción muscular, regula la transmisión del impulso
nervioso de una célula a otra, participa en la osmoregulación y además sirve
como cofactor en diversas reacciones enzimáticas (Hou et al., 2012); su
absorción puede ser facilitada por la acción de la lactosa (Davis y Glatin, 1996).
Una fuente de Ca para los crustáceos, se obtiene de la aplicación de
hidróxido de Ca(OH)2 a los estanques de cultivo, ésta es una práctica muy
frecuente y dentro de los beneficios que se obtienen con ello, se encuentra la
eliminación y control de algunos parásitos de peces y crustáceos. Además,
ayuda a mantener el agua con la turbidez adecuada, evitando aguas obscuras
que estresen al camarón, se incrementa el pH del suelo y se aumenta la

19
liberación de nutrientes de los fangos del estanque y la descomposición de la
materia orgánica (Chávez, 2009).
Magnesio
Otro de los minerales esenciales es el magnesio (Mg), el cual es un
componente esencial del exoesqueleto y del mantenimiento de la homeostasis
intra- y extracelular de los crustáceos (Davis y Glatin, 1996). Es esencial para la
respiración celular y sirve como cofactor o componente de distintos sistemas
enzimáticos importantes, en la regeneración celular y en el ciclo reproductivo
(Roy et al., 2007).
Este mineral es fácilmente absorbido a través del tracto gastrointestinal,
branquias, piel y aletas de peces y crustáceos. La disponibilidad y absorción de
Mg se ve reducida en presencia de fitatos y concentraciones elevadas de Ca.
Mantener niveles adecuados de Mg, es importante para que se lleve a cabo la
correcta actividad de la enzima Na+-K+-ATPasa, ya que el Mg sirve como
cofactor y está involucrado en el metabolismo de lípidos, proteínas y
carbohidratos actuando en numerosas reacciones enzimáticas y metabólicas
(Roy et al., 2007; http://www.fao.org/docrep/field/003/ab492s/AB492S04.htm).
Entre los signos de una deficiencia de este mineral se encuentran un
crecimiento pobre, anorexia, letargia, flacidez muscular, convulsiones y alta
mortalidad (Davis y Glatin, 1996).
Potasio
Otro de los iones de gran importancia en la célula es el potasio (K+), que
se encuentra en casi todos los fluidos y tejidos blandos del organismo, ya que
es el principal catión de los fluidos intracelulares y participa en la regulación de
la presión osmótica intracelular. El K+ desempeña una función vital manteniendo

20
el equilibrio ácido-base. Igualmente tiene un papel muy importante en el
metabolismo de los organismos (Chávez, 2009;
http://www.fao.org/docrep/field/003/ab492s/AB492S04.htm).
Niveles inadecuados de K+ en el agua, podrían afectar seriamente la
capacidad osmoreguladora de las células (Roy et al. ,2007). Tanto el K+ como el
Mg son iones esenciales para el crecimiento, sobrevivencia y función
osmoreguladora de los crustáceos. El K+ es importante en la activación de la
enzima Na-K-ATPasa, ya que su actividad enzimática está directamente
relacionada con la concentración de K+ y la regulación del volumen extracelular
(Roy et al., 2007; McGraw y Scarpa, 2003). Además, se ha demostrado que una
disminución en los niveles de K+ afectarían al camarón ocasionándole una
disminución del crecimiento, anorexia, letargo e incluso la muerte (Zhu et al.,
2006).
Una de las sustancias utilizadas para mantener niveles adecuados de K y
Mg en el agua, cuando se cultiva camarón a baja salinidad es la aplicación de
cloruro de potasio (KCl) y sulfato de potasio y magnesio ((K2SO4-MgSO4) (Roy
et al., 2007). McNevin et al. (2004), observaron en Alabama una mayor
producción de camarón en aguas con baja salinidad, al incrementar los niveles
de K+ (6,2 mg l-1) y Mg2+ (4,6 mg l-1) a 40 y 20 mg l-1, respectivamente,
utilizando cloruro de potasio (KCl) y sulfato de potasio y magnesio (K2SO4-
MgSO4). Diversos estudios han demostrado un incremento en la sobrevivencia
y ganancia de peso en los camarones, cuando se mantienen niveles adecuados
en las concentraciones de K+ y Mg2+ en el agua, así como de otros minerales
durante la aclimatación de postlarvas en agua con baja salinidad (Roy et al.,
2007). Los efectos fisiológicos que se pueden presentar durante el desarrollo de
los camarones debido a una deficiencia en la concentración de los iones en el
agua pueden ser serios, de ahí la importancia de monitorear constantemente
estos minerales, ya sea empleando métodos fotométricos o bien metodologías
más complejas como es la Espectrometría de Absorción Atómica. (Roy et al.,
2007).

21
Uso de Antibióticos en la Acuicultura
Existen en la actualidad más de 40,000 clases de antibióticos y
aproximadamente 80 de ellos son utilizados en la agricultura y en la industria
acuícola. Los antibióticos son los compuestos quimioterapéuticos más
importantes con capacidad para inhibir el crecimiento de microorganismos o
aniquilarlos (Lee et al., 2007; Allison et al., 2001). Se obtienen de manera
natural, semi-sintética o sintética y pueden ser aplicados vía parenteral, oral o
tópica (Kemper, 2008).
Los antimicrobianos se han utilizado en la producción animal con
diversos propósitos, como es el control y tratamiento de enfermedades
infecciosas, con fines profilácticos y como promotores de crecimiento, buscando
incrementar el peso de los animales o una mayor eficiencia alimentaria (Viola y
DeVincent, 2006). En la acuicultura, los antibióticos son utilizados
frecuentemente durante los ciclos de cultivo, tanto en los laboratorios de
reproducción como en las granjas de cultivo de camarón (Soto-Rodríguez et al.,
2006), con la finalidad de disminuir o controlar las enfermedades bacterianas
(Uno, 2004).
Cuando se presentan infecciones bacterianas en los camarones
peneidos, se aplican terapias empleando alimento adicionado con algún
antibiótico, sin embargo, el empleo de dietas medicadas en la acuicultura, ha
sido asociado con problemas ambientales y de salud humana. Entre éstos se
incluye, la generación de bacterias resistentes, la persistencia de los
antibióticos en el sistema acuático y los efectos sobre la composición
biogeoquímica del sedimento (Santiago et al., 2009).
También, es importante resaltar que la acumulación de residuos de
antibióticos en el músculo de camarón, puede alterar la flora intestinal y
provocar problemas de intoxicación o alergias en el consumidor (Santiago et al.,
2009). No es recomendable utilizar los antibióticos en la producción animal
como medida preventiva, ya que las bacterias desarrollan resistencia a ellos de
una manera muy rápida, provocando su ineficacia.

22
La aplicación de antibióticos sólo debe realizarse si existe un diagnóstico
adecuado de la situación y siempre bajo protocolos de control previamente
establecidos. Además, debe llevarse a cabo una evaluación del impacto por la
administración de dicho compuesto, conocer el patrón de uso, la hidrología del
área y las condiciones fisicoquímicas del agua. Se deben considerar las buenas
prácticas de manejo acuícola como prioridad para evitar la entrada de
patógenos a los sistemas de cultivo de camarón y utilizar antibióticos
únicamente como último recurso (Santiago et al., 2009).
En la selección de un antibiótico, el costo es un factor importante ya que
existen medicamentos muy efectivos contra algún patógeno, pero su precio los
hace prácticamente imposibles de utilizar a las dosis adecuadas. En el caso de
camarón, la vía de administración de antibióticos más frecuente es la oral,
utilizando el alimento como vehículo, el cual es incorporado a un medio marino
sumamente agresivo. Debido a esto, es importante que el antibiótico vaya
dentro de un “pellet” para mantener su estabilidad y protegerlo de factores como
lixiviación y unión a cationes trivalentes y divalentes. El personal responsable
de la granja debe verificar que los camarones consuman el alimento para poder
aplicar la terapia con el antibiótico ya que de no ser así, los resultados en el
control de la enfermedad serán nulos y sólo se contaminará el medio ambiente
(Santiago et al., 2009). En México, los antibióticos de uso común en la
camaronicultura son enrofloxacina, florfenicol y oxitetraciclina (Soto-Rodríguez
et al., 2006). Sin embargo, se conoce muy poco de los efectos terapéuticos de
estos antibióticos, en infecciones producidas por bacterias.
Características de la Oxitetraciclina
La oxitetraciclina fue el primer antibiótico aprobado por la Administración
de Medicamentos y Alimentos (FDA) para ser utilizado en la acuicultura (Reed
et al., 2004). Es un antibiótico bacteriostático perteneciente a la familia de las
tetraciclinas, conformado por una estructura tetracíclica. Posee un amplio

23
espectro contra bacterias Gram (+) y Gram (-), rickettsias y micoplasma. Es
utilizado en las granjas acuícolas en el tratamiento de vibriosis,
hepatopancreatitis necrotizante (NHP) y furunculosis (Mendoza-Patiño y
Campos-Sepúlveda, 2008), ejerciendo su efecto antibacterial a nivel del
ribosoma. Oxitetraciclina es producida por cepas de Streptomyces rimosus que
presenta determinantes de resistencia a este antibiótico (Santiago et al., 2009).
Desarrollo de Resistencia Bacteriana por el Uso de la Oxitetraciclina
La resistencia antimicrobiana se ha convertido en un gran problema de
salud y es definida como la capacidad de las bacterias de soportar
concentraciones de antibióticos mayores a las que les causa inhibición. Los
microorganismos desarrollan esta capacidad de adaptación, debido a sus cortos
tiempos de generación y sus constantes mutaciones (COSAES, 2010).
El desarrollo de resistencia bacteriana a antibióticos puede ser intrínseca
(perteneciente a la bacteria) o adquirida (Betancourt et al., 2003). La primera se
define como aquella que es característica de todos los miembros de una
especie o género dado. En muchos casos se refiere a la incapacidad de un
antibiótico para causar un efecto en el sitio diana de la célula La resistencia
natural o intrínseca no está relacionada con el uso de antibióticos, ya que se
manifiesta independientemente de la presencia de éstos (Woodford y Ellington,
2007).
La resistencia adquirida se debe a la modificación del material genético
de la bacteria. Puede aparecer como resultado de una mutación al azar de
genes localizados en los cromosomas o en sitios extracromosómicos o bien por
la transferencia de material genético desde otros microorganismos. Se relaciona
también con los procesos dinámicos, como los cambios en el transporte del
antibiótico dentro de la célula, cambios moleculares o producción de enzimas
que modifican o inactivan al antibiótico Este tipo de resistencia puede estar o no
influenciada por el uso de algún compuesto determinado, es decir, el uso de

24
antimicrobianos puede estimular la adquisición de resistencia como parte de un
proceso conocido como presión selectiva (Tenover, 2006).
El uso desmedido de antibióticos como profilácticos (Baquero et al.,
2009) y la práctica generalizada de administrar dosis bajas de antibióticos como
promotores de crecimiento en la producción animal, han favorecido el desarrollo
de bacterias resistentes (Tang y Moxon, 2001). La presencia residual de
antibióticos en alimentos de origen animal también puede contribuir a aumentar
la resistencia bacteriana (Álvarez et al., 2004). En el ambiente acuático, la
transferencia de material genético con factores de resistencia a antibióticos es
relativamente fácil mediante el proceso de conjugación de plásmidos y puede
ocurrir entre cepas del mismo o diferente género (González-Lama et al., 2001).
Las bacterias que desarrollan resistencia a oxitetraciclina, habitualmente
presentan resistencia a otros antibióticos y es adquirida por plásmidos. Los
mecanismos que dan lugar a la resistencia, pueden ser a través de la pérdida o
disminución de la permeabilidad de la pared celular bacteriana para el
antibiótico o la adquisición de una vía de salida dependiente de energía, un
menor acceso de oxitetraciclina al ribosoma bacteriano o a través de la síntesis
de enzimas bacterianas que metabolizan al antibiótico (Mendoza-Patiño y
Campos-Sepúlveda, 2008).
Uso de Sistemas Cerrados para el Cultivo de Camarón
La acuicultura tradicional requiere de una gran cantidad de agua y
grandes extensiones de terreno. En muchos países de América Latina el agua
es un recurso escaso debido al crecimiento poblacional y al cambio climático
entre otros factores. Por otro lado, la producción mundial generada por la
actividad acuícola ha ido en aumento constante duplicándose en la última
década (FAO, 2009).
Según la Organización Mundial para la Alimentación y la Agricultura
(FAO), las principales cuestiones que se deben abordar en la acuicultura están

25
relacionadas con la implementación de tecnología apropiada, recursos
financieros, el impacto que tendrá al medio ambiente por el desarrollo de esta
actividad y la manera de enfrentar las enfermedades que se presentan en este
sector (FAO, 2009).
En la acuicultura la implementación de sistemas de cultivo con
recirculación de agua engloban un conjunto de procesos y componentes que
permiten que el agua sea continuamente tratada y reutilizada. El uso racional
del agua es una de las principales ventajas que presentan estos sistemas ya
que el volumen de recambio es menor a un 10% diario del volumen total del
sistema (Jiménez, 2012).
Este tipo de sistema de producción acuícola, promueve el crecimiento de
los organismos ya que les proporciona un ambiente con mayor control de los
parámetros fisicoquímicos como la temperatura, el OD, TAN, CO2, pH, la
alimentación y principalmente el control de enfermedades, además de permitir
realizar cultivos de camarón en los meses de invierno. Los sistemas de cultivo
cerrados son considerados bioseguros, sustentables, con poco impacto
ecológico, en los cuales se puede producir camarón de alta calidad con una
eficiencia costo-beneficio (Timmons et al., 2002).
Los estanques empleados en estos sistemas de cultivo son de tierra,
pero también se pueden utilizar membranas de recubrimiento para reducir la
erosión y mejorar la calidad del agua. Usualmente, los estanques son pequeños
(0.1–1.0 ha) de forma cuadrada o redonda. La profundidad puede ser de hasta
1.5 m y la densidad de siembra varia desde 60 hasta 300 postlarvas/m2.
Entre las principales características que debe poseer un sistema cerrado de
producción acuícola se encuentran:
a) La remoción de sólidos conformados por los desechos producidos en el
sistema, como las heces y el alimento no consumido.
b) La biofiltración que es utilizada para controlar los compuestos
nitrogenados, producto del metabolismo de los organismos.
c) La aireación u oxigenación constante del agua.
d) La degasificación para la eliminación del CO2 acumulado en el sistema.

26
e) La implementación de tecnología para mantener la circulación del agua.
Algunas de las ventajas de los sistemas cerrados en acuacultura son:
• La reducción en la transmisión y propagación de enfermedades.
• La considerable disminución de los contaminantes vertidos al medio
ambiente.
• La optimización en el uso de recursos, como el agua, alimento, energía,
terreno, personal, etc.
• La obtención de niveles más altos de Factor de Conversión Alimenticia
(FCA).
• La programación más eficiente de la producción.
• Estos sistemas de producción pueden ser utilizados durante las
diferentes etapas de desarrollo de los organismos acuáticos (peces,
crustáceos y moluscos), tanto de agua dulce como salada (Jiménez,
2012).
Las desventajas de los sistemas cerrados en acuacultura son:
• Una inversión inicial de alto costo.
• Se requiere de personal calificado (Jiménez, 2012).

27
HIPÓTESIS
La utilización de un Sistema de Cultivo Cerrado de producción intensiva
de camarón Litopenaeus vannamei mejora el control de los parámetros
fisicoquímicos y el desarrollo de los organismos, disminuyendo las infecciones
por los microorganismos patógenos que habitan el ambiente marino.

28
OBJETIVOS
Objetivo General
Evaluar un sistema cerrado para el cultivo de camarón blanco
Litopenaeus vannamei mediante el estudio de aspectos fisicoquímicos,
fisiológicos y microbiológicos.
Objetivos Específicos
1. Determinar los parámetros fisicoquímicos en el agua de mar, que influyen en
el desarrollo de camarón Litopenaeus vannamei mantenido en un Sistema de
Cultivo Cerrado.
2. Realizar una evaluación fisiológica externa e interna del camarón cultivado en
el sistema cerrado.
3. Detectar mediante la técnica de PCR convencional las cepas de Vibrio
aisladas de los distintos tejidos de camarón cultivado.
4. Identificar empleando PCR las especies de Vibrio patógenas para el camarón
y para humanos
5. Determinar la sensibilidad a la oxitetraciclina de las bacterias de Vibrio
aisladas del Sistema de Cultivo Cerrado.

29
MATERIALES Y MÉTODOS
Diseño Experimental
Esta investigación se realizó en una granja camaronícola comercial
ubicada en Bahía de Kino, Sonora, empleando un estanque tipo invernadero
con dimensiones de 50 m de largo, 8 m de ancho y una profundidad de 1.2 m,
mantenido con aireación constante, mediante aireadores de paleta impulsados
por motores de 2 caballos de fuerza. Dicho estanque fue sembrado con una
densidad de 126 camarones por m2 correspondientes a la especie L. vannamei
en etapa juvenil con un peso inicial promedio de 13.64 g.
El estanque se llenó con agua de mar, realizando la medición de los
parámetros fisicoquímicos y tratando de mantener las condiciones siguientes: la
salinidad 39‰, pH (6.5-7.0) y la temperatura de 29-30°C.
Una vez colocados los organismos, se dió a todo el estanque un tiempo
de aclimatación de una semana, bajo las condiciones experimentales
establecidas. Durante ese tiempo, los organismos se alimentaron cinco veces al
día con una dieta comercial sin medicar con 35 % de proteína, a una biomasa
de 3%. Posteriormente, los camarones fueron mantenidos en experimentación
durante 58 días evaluando el incremento en peso y la mortalidad.

30
Medición de Parámetros Fisicoquímicos
La medición de los parámetros físicoquímicos del agua como, la
temperatura, el pH y el oxígeno disuelto, se realizó diariamente dos veces al día
(mañana y tarde), empleando un oxímetro YSI (YSI 55, Yellow Springs OH.
EUA) y un potenciómetro YSI (Ecosense pH10, Yellow Springs, OH, EUA). La
medición de la salinidad se realizó dos veces por semana empleando un
refractómetro Aquatic Eco-Systems (VitalSine SR-6, Apopka, FL, EUA).
Determinación de Ganancia de Peso en Camarón
Para la determinación de este parámetro, se llevó a cabo un registro
inicial del peso de los camarones antes de colocarlos en el estanque.
Posteriormente, se realizaron muestreos semanales en diferentes puntos del
estanque, con la finalidad de monitorear el desarrollo del camarón durante toda
la etapa experimental. Para ello se tomaron al azar 10 organismos cada
semana, los cuales fueron pesados empleando una balanza analítica digital
marca OHAUS® (Mod. Scout Pro SP202, Parsippany, NJ, EUA). Para obtener el
valor real de ganancia de peso en los organismos estudiados, se utilizó la
fórmula propuesta por Pan et al. (2007) definida como:
Ganancia de Peso= Peso Final-Peso Inicial/ Tiempo Experimental-No. de
organismos.
Determinación de la Sobrevivencia en Camarón
Para determinar la sobrevivencia de los camarones, se contabilizaron
diariamente los organismos muertos en el estanque. El cálculo se realizó
comparando el número de organismos colocados inicialmente menos los

31
camarones muertos, reportando este parámetro como porcentaje de
sobrevivencia (Li et al., 2007).
Concentración de los Iones Ca, Mg y K, Nitrógeno Amoniacal Total (TAN) y Nitritos (NO2) en el Agua del Estanque de Cultivo
Para la determinación de la concentración de los iones y nitritos en el
agua del estanque se utilizó un fotómetro YSI Ecosense 9500 y los “kits”
comerciales para los elementos calcio (Ca), potasio (K), magnesio (Mg), amonio
y nitritos. Se tomaron muestras de agua de 1000 mL del estanque, las cuales se
dejaron reposar de 15 a 20 min, para sedimentar la materia orgánica.
Posteriormente, el agua se filtró, colocando 10 mL en celdas de vidrio para cada
uno de los analitos. Se incluyó junto con las muestras un control negativo que
consistió en una celda de vidrio con 10 mL de agua de mar filtrada.
Determinación de Calcio
Se interpretó como la dureza de calcio (CaCO3), tomando 10 mL de agua
previamente filtrada, a la cual se le adicionó una tableta de “Calcicol 1” (Water
Test Tablets Palintest AP 188 AILAPHOT) disolviéndola completamente en la
muestra. Posteriormente, se agregó una tableta de “Calcicol 2” (Water Test
Tablets Palintest AP 188 AILAPHOT) y se agitó hasta disolverla totalmente,
luego se dejó reposar por 2 min para permitir el completo desarrollo del color y
se llevó a cabo la lectura en el fotómetro a una longitud de onda de 570 nm. El
resultado obtenido se multiplicó por el factor 0.4008 para expresar el resultado
en mg/L de Ca.

32
Determinación de Potasio
Se tomaron 10 mL de la muestra de agua previamente filtrada, a la cual
se le adicionó la tableta K (Water Test Tablets Palintest AP 189 AILAPHOT), se
disolvió completamente en el agua. Posteriormente se dejó reposar por 2 min,
tornándose el agua turbia, lo cual indicó la presencia de potasio, luego se leyó
la muestra en el fotómetro a una longitud de onda de 520 nm.
Determinación de Magnesio
Se colocó en una celda de vidrio 1 mL de agua previamente filtrada y 9
mL de agua desionizada, se adicionó una tableta de Magnecol (Water Test
Tablets Palintest AP 193 AILAPHOT). Ésta se disolvió completamente en el
agua y se dejó reposar por 2 min, para su posterior lectura en el fotómetro a
una longitud de onda de 520 nm.
Para llevar a cabo la lectura de los iones, primeramente se realizó la
calibración del fotómetro, iniciando con la lectura del control negativo.
Posteriormente, se leyó cada una de las celdas de vidrio con la muestra para
los distintos iones. La lectura fue obtenida digitalmente, expresándose la
concentración en mg/L. Cuando la concentración de los iones de Ca, K, y Mg en
el agua fue elevada, apareció en la pantalla del fotómetro la leyenda “mayor”
(>>), lo cual indicó que la muestra debería diluirse. Para K se tomó 1 mL de la
muestra de agua y 9 mL de agua desionizada, para Ca y Mg la dilución fue en
proporción 1:1 (v/v) empleando 5 mL de muestra y 5 mL de agua desionizada y
se procedió a hacer nuevamente la lectura en el fotómetro. El tiempo
transcurrido entre la primera y la segunda lectura no fue mayor a 2 min, ya que
puede generarse una lectura incorrecta. La Figura 2 muestra la coloración que
presentó cada uno de los elementos determinados.

33
(a) (b) (c) (d)
Fig. 2. Determinación de iones en el agua de mar a) Control negativo, b) Potasio, c) Calcio, d) Magnesio Nitrógeno Amoniacal Total (TAN)
Para llevar a cabo esta determinación se colocaron en una celda de
vidrio 10 mL de agua del estanque, posteriormente se agregó 10 g de sales de
litio, para prevenir la precipitación de sales y se mezcló hasta disolver el
reactivo. Luego se agregaron las tabletas amonio No. 1 y amonio No. 2 (Water
Test Tablets, Palintest AP 152, Reino Unido), se mezclaron hasta disolverse
completamente. La muestra se dejó reposar durante 10 min para permitir la
aparición de color y se realizó la lectura en el fotómetro a una longitud de onda
de 640 nm.
Nitritos (NO2)
Para esta determinación se colocaron 10 mL de agua del estanque en
una celda de vidrio, a la cual se le adicionó una tableta de NITRICOL (Water
Test Tablets Palintest AP 109, Reino Unido), se agitó la celda hasta obtener

34
una homogeneización total de la tableta en el agua. Posteriormente, se dejó
reposar la muestra durante 10 min a temperatura ambiente para llevar a cabo la
reacción, generándose una coloración, luego se realizó la lectura en el
fotómetro a una longitud de onda de 520 nm.
Diagnóstico para la Detección de Lesiones Externas e Internas en Camarón
Esta evaluación se llevó a cabo con la finalidad de detectar lesiones
externas e internas en los camarones mantenidos dentro del Sistema de Cultivo
Cerrado, tomándose diariamente tres camarones. La evaluación externa,
consistió en buscar lesiones o cambios de coloración en la cutícula y/o
pleópodos, así como alguna patología como necrosis en la cutícula.
Posteriormente, se extrajeron los órganos del camarón (hepatopáncreas,
intestino y branquias) y se realizaron las observaciones microscópicas en
fresco, para establecer el grado de severidad de las lesiones en los camarones,
ocasionadas por infecciones bacterianas o virales.
La forma de extraer cada uno de los órganos, se llevó a cabo haciendo
una incisión por la línea media del cuerpo del camarón, con unas tijeras para
disección, tomando los órganos con unas pinzas. Éstos se colocaron sobre un
portaobjetos y se adicionó una gota de solución salina. Se colocó un
cubreobjetos presionando cuidadosamente con una pinza y se observó la
muestra bajo el microscopio (Labomed, Mod. CX, California, EUA) utilizando los
lentes de 4, 10, 40 y 100 X.
La interpretación de las lesiones observadas al microscopio, se llevó a
cabo siguiendo los criterios establecidos por Lightner (1996), empleando
valores categóricos del cero al cuatro, siendo el valor de cero cuando no existió
una lesión en el órgano y el de cuatro, cuando se presentó un daño muy severo
(Tabla 1). Se promediaron los valores obtenidos de los tres camarones en cada
una de las observaciones, llegando a un valor único de dicha población. Este

35
procedimiento permitió determinar los órganos que son más afectados por las
infecciones en los camarones.
Tabla 1. Criterios para establecer el grado de severidad de las lesiones en los distintos órganos de camarón
Grado de severidad Branquias
1 Ausencia de protozoarios parásitos Menos del 10% de las branquias cubiertas por protozoarios parásitos 2
3 Menos del 50% de las branquias cubiertas por protozoarios parásitos El 100% de las branquias cubiertas por protozoarios parásitos 4
Intestino 1 Ausencia de Gregarinas
2 3
Menos de 10 gregarinas adultas
Entre 10 y 50 gregarinas adultas 50 o más gregarinas adultas 4
Lípidos
0 Ausencia total de esferas de lípidos
1 Leve presencia de esferas de lípidos
2 El 25% de cada túbulo con esferas de lípidos
3 El 50% de cada túbulo con esferas de lípidos
4 Todos los túbulos llenos de lípidos
Túbulos del
Hepatopáncreas 1 Túbulos lisos y con grado 4 de lípidos
2 Presencia leve de túbulos estrangulados, sin disminución de lípidos
3ª El 25% de los túbulos estrangulados en varias partes
3b El 50% de los túbulos estrangulados y secos
3c Túbulos secos, sin lípidos y con necrosis (Lightner, 1996).

36
Análisis Bacteriológico en los Órganos de Camarón Extracción de Hemolinfa
Se tomaron diariamente tres camarones del estanque de cultivo durante
toda la etapa experimental, a los cuales se les extrajo la hemolinfa. El
procedimiento consistió en colocar el camarón en posición ventro-dorsal (boca
arriba) dejando expuesta la zona de unión entre el cefalotórax y el abdomen
(seno hemolinfático ventral), luego se secó perfectamente el abdomen del
camarón con papel absorbente, posteriormente, se utilizó una jeringa de
insulina estéril con aguja hipodérmica de 1mL cargada con aproximadamente
100 µl del anticoagulante citrato de sodio al 10% (J.T. Baker, EUA). Se extrajo
asépticamente la muestra de hemolinfa introduciendo con cuidado el bisel de la
aguja hacia arriba y succionando al mismo tiempo hasta extraer un volumen de
100 µL de hemolinfa por cada camarón. La relación de la hemolinfa y el
anticoagulante fue 1:1 (v/v). La Figura 3 muestra la forma en la cual se extrajo
la hemolinfa del camarón y la posición en que éste y la jeringa deben colocarse.
Inmediatamente después de extraer la hemolinfa, 50 µL de ésta fueron
colocados en una placa con agar marino y 50 µL en una placa con agar
Tiosulfato Citrato Bilis Sucrosa (TCBS), ambas placas con los medios de cultivo
fueron homogeneizadas empleando una varilla de vidrio estéril, esto se realizó
rápidamente para evitar la coagulación de la hemolinfa.
Se incluyó una placa de agar marino y otra de agar TCBS con y sin el
anticoagulante como control. Se registró el volumen extraído de hemolinfa de
cada camarón, para ser incluido en los cálculos realizados para obtener las
Unidades Formadoras de Colonias (UFC) por mL.
UFC/mL= número de colonias/volumen de hemolinfa

37
Figura 3. Procedimiento para la extracción de hemolinfa de camarón Litopenaeus vannamei Extracción de Hepatopáncreas y Branquias de Camarón
Se extrajeron el hepatopáncreas (Hp) y las branquias de los camarones
de forma aséptica, haciendo un registro del peso de ambos órganos. En la
Figura 4 se ilustra la extracción de branquias y hepatopáncreas de los
camarones. El Hp y las branquias se diluyeron por separado en tubos con 9 mL
de agua peptonada alcalina (APW) (Difco, Diagnostic Systems, EUA),
conteniendo 3.5 % de NaCl, las diluciones utilizadas fueron de 101 y 102. Se
homogenizó la muestra en el tubo y se tomaron 100 µL, los cuales se
inocularon por extensión en superficie en una caja petri con Agar Marino y Agar
TCBS (Difco, Diagnostic Systems, USA) adicionado con 2.5 % de NaCl. El
análisis se realizó por duplicado, incubando las placas a 37° C durante 24 h en
una incubadora (VWR Scientific products, mod. 1545, EUA).
Se realizó el conteo de las UFC en las placas que presentaron un
crecimiento bacteriano entre 25 a 250 UFC/mL. Los resultados se expresaron
como Unidades Formadoras de Colonias por gramo (UFC/g). Posteriormente,

38
se continuó con el aislamiento de las cepas de interés de las placas de Agar
TCBS.
(a) (b)
Figura 4. Extracción de los órganos de camarón Litopenaeus vannamei, (a) branquias, (b) hepatopáncreas. Aislamiento de Bacterias del Género Vibrio
De las cajas petri con crecimiento bacteriano superior a 25 UFC/mL se
seleccionaron 10 colonias bacterianas, con características morfológicas de las
bacterias pertenecientes al género Vibrio. Éstas fueron sembradas en tubos con
Agar Soya Tripticasa (AST) (Bioxon) con 3.5 % de NaCl, los cuales se
incubaron a 37°C durante 24 h. Después de la incubación, los tubos se
mantuvieron en un lugar fresco y oscuro hasta su traslado al Laboratorio de
Análisis Biológicos, del Centro de Investigación en Alimentación y Desarrollo
AC., para su posterior identificación molecular empleando técnicas de Reacción
en Cadena de la Polimerasa. Los mismos cultivos bacterianos aislados fueron
empleados en la determinación de la Concentración Mínima Inhibitoria (CMI) de
la oxitetraciclina (OTC).

39
Detección de Especies de Vibrio Aisladas de Camarón Litopenaeus vannamei
Extracción de ADN a Partir de Aislamientos Bacterianos de Tejidos de Camarón
Para la extracción de ADN, se utilizó el kit comercial para purificación
Wizard® Genomic ADN (Promega Inc. Madison, WI. EUA). Se tomó un inóculo
de las bacterias aisladas del camarón, las cuales fueron mantenidas en los
tubos de AST, para ser resembradas en un medio de pre-enriquecimiento que
consistió en agua peptonada con 2% de NaCl. Se tomó 1mL del medio de pre-
enriquecimiento y se colocó en tubos Eppendorf de 1.7 mL (Sorenson®,
BioScience, Inc. Utah, EUA), los cuales se centrifugaron en una centrifuga
Allegra (64R, Beckman Coulter, EUA), a 15,000 x g durante 2 min para
precipitar las células. Se removió el sobrenadante y se agregaron 600 µL de
una solución de lisis de núcleo, pipeteando en repetidas ocasiones hasta que
las células se resuspendieron.
Posteriormente, los tubos Eppendorf fueron incubados en un baño de
agua (Precision Scientific, mod. 181, Chicago, ILL. EUA) a 80°C durante 5 min
provocando lisis en las células. Luego los tubos se enfriaron a temperatura
ambiente y se les adicionó 3 µL de una solución de RNasa, se invirtieron de 2 a
5 veces para mezclar perfectamente su contenido. Los tubos fueron incubados
en un baño de agua a 37°C por 15 min, dejándolos enfriar a temperatura
ambiente. Posteriormente, a los tubos con las células lisadas se les adicionaron
200 µL de una solución para precipitación de proteínas, éstos se agitaron
vigorosamente empleando un vortex mixer (Baynstead Thermolyne Tipo 37600)
a velocidad alta por 20 s o hasta lograr una mezcla homogénea de la solución
de precipitación de proteínas con las células lisadas.
Posteriormente, se dejaron reposar las muestras sumergidas en hielo
durante 5 min. Luego se centrifugaron los tubos en una centrifuga Allegra (64R,
Beckman Coulter, EUA) a 15,000 x g durante 3 min, transfiriendo el
sobrenadante conteniendo el ADN a tubos Eppendorf limpios de 1.7 mL que

40
contenían 600 µL de isopropanol (Fluka, EUA). Ambos componentes se
mezclaron cuidadosamente por inversión, hasta que las cadenas de ADN
formaron una masa visible. Los tubos fueron centrifugados nuevamente de
15,000 x g por 2 min.
Cuidadosamente se removió el sobrenadante y los tubos fueron secados
empleando papel absorbente limpio, luego se les adicionaron 600 µL de etanol
(Sigma-Aldrich, lote 05J02, EUA) al 70% y se invirtieron varias veces para lavar
el precipitado de ADN. Posteriormente, se centrifugaron de 15,000 x g por 2 min
y se aspiró cuidadosamente el etanol. Se eliminó el líquido de los tubos
utilizando papel absorbente limpio y se dejaron secar a temperatura ambiente
por un período de 10-15 min. Se adicionaron a los tubos 100 µL de una solución
para la resuspensión del ADN. Los tubos conteniendo las muestras fueron
incubados a 65°C en un baño de agua (Precision Scientific, mod. 181, Chicago,
ILL. EUA) por 1 h, mezclando periódicamente. Alternativamente, se puede
rehidratar el ADN incubando la solución durante toda la noche a temperatura
ambiente o a 4°C. Posteriormente, las muestras conteniendo el ADN fueron
almacenadas en refrigeración de 2-8°C.
Determinación de la Concentración de ADN
La medición de la concentración de ADN se llevó a cabo en un
espectrofotómetro NanoDrop ND-1000 (NanoDrop Tech. Wilmington, DE. EUA),
a una absorbancia de 260 nm, para ello se utilizó 1 µL de ADN de cada una de
las muestras de camarón.
Reacción en Cadena de la Polimerasa (PCR)
Para la identificación de las bacterias de Vibrio (Vibrio spp, V. cholerae,
V. vulnificus, V. harveyi, V. harveyi gen de hemolisina, V. harveyi toxR), se
utilizó el kit comercial GoTaq® PCR Core Systems (Promega Inc. 53711-5399,

41
Madison, WI. EUA) utilizando una concentración de 100 ng de ADN para todas
las muestras a un volumen final de 25 µL. La mezcla de reacción estuvo
conformada por: 1 µL de una mezcla de dNTP´s 10 mM (dATP, dGTP, dTTP,
dCTP), 2 µL de MgCl2 25 mM, 5 µL de 5x buffer GoTaq® con colorante verde,
0.25 µL de ADN polimerasa GoTaq® y de 0.9 a 1.5 µL de cada oligonucleótido
a una concentración de 50 pmoL.
La amplificación del ADN se llevó a cabo utilizando un Termociclador
Perkin Elmer (DNA Thermal Cycler 480, Norwalk, CT). Las condiciones de la
reacción de PCR para cada uno de los oligonucleótidos iniciadores se muestran
en la Tabla 2.
Electroforesis en Gel de Agarosa
Para comprobar la amplificación del material genético del producto
obtenido de la reacción de PCR, se realizó una electroforesis en gel de agarosa
(Sigma, Aldrich St Louis MO, EUA) al 2.0 %. El gel se preparó disolviendo 1 g
de agarosa en 50 mL de buffer TPE 1X (Tris borato 1M, ácido fosfórico 85% y
EDTA 0.5 M), se le adicionó 1 µL de bromuro de etidio (Sigma-Aldrich, Inc.,
EUA) a una concentración de 10 mg/mL. La electroforesis se realizó utilizando
una fuente de poder (Labnet International inc., EUA) ajustada a 100 volts y la
corrida se realizó durante 30 min. El marcador molecular utilizado fue ADN
Ladder (BIO-RAD, No 170-8200), conformado de 50-2000 bp. Una vez
transcurrido el tiempo de la electroforesis, los geles se observaron y
fotografiaron en un fotodocumentador (Gel doc® XR+ BIO-RAD, EUA) mediante
un transiluminador de luz ultravioleta (UV) conectado a una computadora
conteniendo el programa Image Lab software (BIO-RAD, EUA).

42
Tab
la 2
. C
ond
icio
ne
s de
PC
R p
ara
ca
da u
no
de
los
olig
on
ucl
eót
ido
s in
icia
do
res
util
iza
do
s e
n
la id
ent
ifica
ció
n d
e
esp
eci
es
de
Vib
rio
Bacterias
de Vibrio
Iniciador
Desnatura-
lización inicial
Desnatura-
lización
Alineación
Extensión
Extensión
final
Ciclos
Amplicón
(pb)
Vib
rio
sp
p
(Ta
rr e
t a
l.,
20
07
)
70
0F
- 5
´-C
GG
TG
AA
AT
GC
GT
AG
AG
AT
-3´
13
25
R-5
´-T
TA
CT
AG
CG
AT
TC
CG
AG
TT
C-3
´
9
3°C
/15
min
92
°C/4
0 s
57
°C/1
min
72
°C/1
.5 m
in
7
2°C
/7 m
in
3
5
66
3
V.
ch
ole
rae
(Ca
iffe
r e
t a
l.,
20
07
)
30
2F
- 5
´-C
TC
AG
AC
GG
GA
TT
TG
TT
AG
GC
AC
G-3
´ 3
02
R-
5 ´
- T
CT
AT
CT
CT
GT
AG
CC
CC
TA
TT
AC
G-3
´
9
4°C
/6 m
in
9
4°C
/1 m
in
6
0°C
/1 m
in
7
2°C
/1 m
in
7
2°C
/7 m
in
3
5
30
2
V.
vu
lnific
us
(Ka
ysn
er
y
De
Pa
ola
, 2
00
1)
51
9F
- 5
´-C
CG
CG
GT
AC
AG
GT
TG
GC
GC
A-3
´ 5
19
R-
5´-
CG
CC
AC
CC
AC
TT
TC
GG
GC
C-3
´
9
4°C
/10
min
94
°C/1
min
6
2°C
/1 m
in
7
2°C
/1 m
in
7
2°C
/10
min
25
5
19
V.
ha
rve
yi
(He
rná
nde
z y
Olm
os,
20
03)
20
48
F-
5´C
TG
TG
TA
CT
CA
CT
GT
TT
AT
C-3
´ 2
04
8R
- 5
´TC
TA
TC
TC
TG
TA
GC
CC
CT
AT
TA
CG
-3´
9
4°C
/3 m
in
9
5°C
/1 m
in
5
1°C
/1 m
in
7
2°C
/2 m
in
7
2°C
/10
min
25
2
04
8
V.
ha
rve
yi
ge
n
he
mo
lisin
a
(Co
ne
jero
y
He
dre
yda
, 2
00
4)
F-5
´-A
TC
AT
GA
AT
AA
AA
CT
AT
TA
CG
TT
AC
T-3
´ R
- 5
´-G
CT
TG
AT
AA
CA
CT
TT
GC
GG
T-3
´
9
5°C
/5 m
in
9
4°C
/1 m
in
5
3°C
/1 m
in
7
2°C
/1 m
in
7
2°C
/10
min
3
0
3
08
V.
ha
rve
yi
ge
n t
oxR
(C
on
eje
ro y
H
ed
reyda
, 2
00
3)
F-
5´-
TT
CT
GA
AG
CA
GC
AC
TC
AC
-3´
R-
5´-
TC
GA
CT
GG
TG
AA
GA
CT
CA
-3´
9
4°C
/3 m
in
9
4°C
/30
s
6
5°C
/30
s
7
2°C
/30
s
7
2°C
/10
min
3
0
3
90

43
Determinación de las Concentraciones Mínimas Inhibitorias (CMI) en Bacterias de Vibrio
Para llevar a cabo esta determinación se emplearon placas de
poliestireno de 96 pozos estériles con tapa de baja evaporación
(Corning®CellBIND®Surface) y un lector de microplacas (BIO-RAD, MOD. 550,
Philadelphia, EUA), micropipetas (FINPIPETTE®, Thermo Labsystems, EUA) de
diversos volúmenes y puntas de plástico estériles, una Incubadora (VWR
Scientific products, mod. 1545) a una temperatura de 30º C, tubos de ensayo de
13x100 mm con 3 mL del medio de cultivo Caldo Soya Tripticasa (CST) y tubos
de ensayo de 16x150 mm con 9 mL de solución amortiguadora de fosfatos
(PBS) estéril, así como tubos con 6 mL de Agar Soya Tripticasa (AST)
inclinados, en los cuales se inocularon las cepas bacterianas aisladas de los
distintos tejidos de camarón. En esta determinación se emplearon como control
(+) cepas tipificadas de V. harveyi, V. parahaemoliticus V. vulnificus y V.
mimicus y un estándar del antibiótico oxitetraciclina (OTC) de alta pureza,
adquirido de la casa comercial Sigma (Sigma-Aldrich Inc., St. Louis, MO. EUA).
Las cepas bacterianas mantenidas en AST fueron sembradas en cajas
petrí con agar TCBS adicionado con 2.5% de NaCl, e incubadas a 30°C durante
24 h, posteriormente, se seleccionaron tres colonias las cuales fueron
inoculadas en tubos conteniendo 3 mL de CST e incubadas a 30°C por 24 h.
Una vez obtenido el crecimiento bacteriano, se ajustó la concentración de las
bacterias utilizando solución amortiguadora de fosfatos estéril (PBS), hasta una
concentración aproximada de 1.5x108 UFC/mL empleando el nefelómetro de
MacFarland 0.5.
Mediante la técnica de dilución en tubo, se realizaron diluciones del
estándar de OTC en el diluyente PBS estéril, hasta obtener las concentraciones
de 100, 75, 50, 25, 10, 5, 2.5 y 1.25 µg/mL.
En cada uno de los pozos de la microplaca estéril fueron colocados 250
µL CST+20 µL de antibiótico y 2 µL del cultivo bacteriano, empleando una
micropipeta automática con puntas estériles y bajo condiciones estériles (cerca
del mechero). Las pruebas se realizaron por triplicado para cada una de las

44
concentraciones de OTC. Además, se incluyó en la placa una hilera de pozos
con 100 µL del estándar de OTC como control, así como un control (-) que
consistió en el medio de cultivo estéril, un control (+) conformado por el medio
de cultivo y el inóculo bacteriano con la finalidad de demostrar la viabilidad de la
bacteria. Posteriormente, se colocaron las tapas en las placas de poliestireno y
se sellaron con parafilm (American National Can®,Neenah, WI, EUA) para
evitar la evaporación. La Tabla 3 muestra el orden en que fueron colocadas las
muestras en la placa.
Las placas se incubaron a 30°C durante 24-48 h y una vez transcurrido
ese tiempo, se realizó la lectura de las mismas en el lector de ELISA a una
longitud de onda de 415 nm, midiendo la absorbancia a las 24 y 48 h de
incubación.
El resultado final de la Concentración Mínima Inhibitoria se obtuvo
restando a la lectura obtenida de los pozos que contienen el medio, la bacteria y
el antibiótico, menos el valor obtenido del medio de cultivo solo, y del
antibiótico. El resultado de esta resta dio un valor negativo, el cual fue
interpretado como la CMI.
Tabla 3. Distribución de las muestras en la placa para la determinación de la Concentración Mínima Inhibitoria de la oxitetraciclina en bacterias de Vibrio
1 2 3 4 5 6 7 8 9 10 11 12
A OTC OTC OTC OTC OTC OTC OTC OTC (-) (+)
B OTC OTC OTC OTC OTC OTC OTC OTC (-) (+)
C OTC OTC OTC OTC OTC OTC OTC OTC (-) (+)
D 100 75 50 25 10 5 2.5 1.25
E OTC OTC OTC OTC OTC OTC OTC OTC (-) (+)
F OTC OTC OTC OTC OTC OTC OTC OTC (-) (+)
G OTC OTC OTC OTC OTC OTC OTC OTC (-) (+)
H 100 75 50 25 10 5 2.5 1.25

45
Análisis Estadístico
Los datos obtenidos durante la evaluación de los diferentes parámetros
del sistema cerrado de cultivo de camarón, fueron analizados empleando
estadística descriptiva, con medidas de tendencia central y de dispersión.

46
RESULTADOS Y DISCUSIÓN
Parámetros Fisicoquímicos
Los parámetros fisicoquímicos evaluados en el sistema cerrado
empleado en el cultivo de camarón L. vannamei, fueron monitoreados
diariamente durante toda la fase experimental. En la Tabla 4 se muestran los
resultados de los parámetros fisicoquímicos evaluados.
Tabla 4. Parámetros fisicoquímicos evaluados en el sistema cerrado para el cultivo de camarón L. vannamei Sistema cerrado Sistema abierto*
Temperatura ( °C) 30.44± 2.09 27 a 31
Oxígeno disuelto (mg/L) 4.71 ± 0.64 3.5-7.5
pH 7.34 ± 0.15 6-8
Salinidad (‰) 39.90 ± 0.88 15-25
(*Condiciones óptimas de acuerdo a Auró y Ocampo, 2006; Ling-Xu et al. 2005; Venkateswara-Rao, 2009; Gutiérrez-Salazar et al., 2011)
Éstos se mantuvieron estables y dentro de los valores considerados
óptimos para el desarrollo de este crustáceo, excepto la salinidad la cual estuvo
en un nivel promedio de 39.9 ± 0.88 ‰. Este valor fue mayor al establecido
como adecuado para el desarrollo del camarón, que es de 15-25 ‰ (Gutiérrez-
Salazar et al., 2011). La elevada salinidad en el agua del estanque de cultivo,

47
pudo ser debido a que no se realizaron recambios constantes de agua durante
el tiempo que permaneció el camarón en estudio, solamente se completó el
volumen que se perdió por la evaporación. Li et al. (2008), reportaron que una
de las características del camarón L. vannamei es que tiene la capacidad de
tolerar amplios intervalos de salinidad que van desde 1-50 ‰, sin que esto sea
un factor limitante para su desarrollo.
Los valores de temperatura y pH obtenidos en este estudio fueron muy
similares a los reportados por Anaya (2005) quien llevó a cabo mediciones de
temperatura (28.8°C), pH (7.9), oxígeno disuelto (7.1 mg/L) y salinidad (28.3 ‰)
en un Sistema de Cultivo Cerrado para camarón. Los valores obtenidos de
salinidad y oxígeno disuelto fueron diferentes a los reportados por este autor.
Determinación de Ganancia de Peso y Sobrevivencia en Camarón
Los camarones evaluados, mantenidos en un Sistema Cerrado de Cultivo
tuvieron un incremento semanal promedio de 1.95 ± 0.64 g, registrando un peso
final de 27.56 g. El mayor incremento en peso se observó en la segunda
semana de iniciado el bioensayo. La ganancia en peso utilizando este sistema,
fue mayor a la obtenida por Anaya (2005), quien reportó un incremento en peso
semanal de 1.18 g en camarones cultivados en un Sistema de Cultivo Cerrado.
A su vez, estudios realizados en el Centro Nacional de Acuicultura e
Investigaciones Marinas (CENAIM), mostraron que en camarones cultivados en
estanques cerrados tipo invernadero, se obtuvieron en promedio aumentos
semanales de peso de 0.8 g.
En este estudio se considera que la ganancia en peso obtenida en los
camarones fue adecuada, ya que una de las principales características de los
Sistemas de Cultivo Cerrados es que el factor de conversión alimenticia sea
mejor, lo cual se ve reflejado en una mayor ganancia de peso en relación al
consumo de alimento.

48
Rojas et al. (2005) reportaron que incrementos en peso entre 0.85 a 1.20
g por semana son considerados adecuados. Cuando la ganancia de peso
promedio semanal es menor a 0.7 g, existe la posibilidad de que el estanque
esté siendo sub-alimentado, como resultado de una mayor sobrevivencia de la
estimada o un error en la densidad de siembra. Si el incremento de peso
promedio se encuentra entre 1.3 y 2.0 g, se debe cuidar que no exista una
sobrealimentación, como resultado de una densidad de siembra menor o a una
disminución de la población, debida a mortalidades.
Cultivos de camarón mantenidos con agua de mar, muestran que el
camarón blanco puede crecer hasta 1.19 g/semana, cuando las densidades de
siembra se encuentran entre 223-299 camarones/m2. De manera libre, esta
especie de camarón puede ser capaz de crecer hasta 1.4 g/semana, cuando las
densidades son de 2-3 camarones/m2 (Valenzuela et al., 2010).
La sobrevivencia obtenida en los organismos mantenidos en el Sistema
de Cultivo Cerrado fue de 74%, con una biomasa total generada de 1,032.5 kg,
en un área de cultivo de 0.04 ha, lo que significa un rendimiento de 25.8 ton/ha.
El incremento en la producción con el Sistema Cerrado de Cultivo, fue cinco
veces mayor al que generalmente se obtiene empleando los sistemas de cultivo
tradicionales (5-6 ton/ha).
La sobrevivencia obtenida con el sistema cerrado fue mayor a la obtenida
por Anaya (2005), la cual fue del 65%, en organismos mantenidos en un
Sistema de Cultivo Cerrado con recirculación de agua. También, se ha
reportado que con estos sistemas, se pueden obtener tasas de sobrevivencia
del 90 al 95%, de acuerdo a lo reportado por Belsher et al., 2011, en una granja
productora de camarón en Perú.
Concentración de los Iones (Ca, Mg y K) en el Agua del Estanque
La concentración de los iones Ca y K en el agua del estanque
permanecieron estables, con niveles promedio de 167.84±16.46 y 88.49±17.78

49
mg/L, respectivamente (Figura 5). El ion Mg mostró variaciones en su
concentración durante el transcurso del estudio, obteniéndose un nivel
promedio de 908.14±176.43 mg/L. La relación de los iones de Mg:Ca
recomendada como adecuada por Pan et al. (2007), para el desarrollo del
camarón es de 4.96:1, cuando los organismos son cultivados en agua con una
salinidad de 40 ‰. Este valor es muy similar a la relación obtenida en este
estudio, que fue de 5:1 para estos mismos iones. La Figura 5 muestra
gráficamente el comportamiento que presentó cada uno de los iones
monitoreados durante el desarrollo del camarón L. vannamei cultivado en el
sistema cerrado.
Figura 5. Concentración de los iones Ca, Mg y K en el agua del estanque durante la evaluación del Sistema Cerrado de Cultivo de camarón
El mantener una relación adecuada en la concentración de los iones en
el agua de los estanques es importante para el crecimiento del camarón, debido
a que éstos iones han sido asociados con la sobrevivencia y ganancia de peso
0
200
400
600
800
1000
1200
1400
1600
1800
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39 41 43 45 47 49
Co
nce
ntr
aci
ón
(m
g/L
)
Días de cultivo
Magnesio
Calcio
Potasio

50
de los organismos (Davis et al., 2005), además de su participación en la
activación de algunas enzimas que tienen un papel fundamental en el desarrollo
de los mismos (Saoud et al., 2003). El agua de mar posee diferente
composición iónica, pero se ha demostrado que una de las características del
camarón de la especie Litopenaeus vannamei es su capacidad
osmoreguladora. Por ello, es considerada como una especie eurihalina (Roy et
al., 2007; Jiang et al., 2005). Es importante considerar que la capacidad
osmoreguladora de los camarones se ve reducida naturalmente cuando éstos
se encuentran iniciando la etapa adulta o sub-adulta (Gong et al., 2004).
La relación encontrada entre los iones de Mg:K fue de 10.6:1, este valor
se estuvo por debajo del valor establecido por Roy et al., 2007, para agua de
mar con una salinidad de 40 ‰, que es de 36:1. Una deficiencia en la
concentración de K+ puede ser perjudicial para el camarón, ya que el K+ es uno
de los iones de mayor importancia para la célula, éste se encuentra en casi
todos los fluidos y tejidos blandos del organismo, y es considerado el principal
catión de los fluidos intracelulares, además de participar en la regulación de la
presión osmótica intracelular. El K+ participa en la activación de la enzima Na+-
K+-ATPasa, la cual actúa en la regulación del volumen extracelular, manteniendo
el equilibrio ácido-base, fundamental en el metabolismo de los organismos
(Chávez, 2009).
La deficiencia de K+ es un fenómeno muy común en el agua de los
estanques, principalmente en los cerrados, ya que este mineral se puede perder
por procesos de filtración o adsorción en el suelo del estanque o por
compuestos presentes en el agua como el carbono orgánico (Boyd et al., 2007).
Una de las principales formas de contrarrestar las pérdidas de éste y
otros iones minerales en los sistemas de cultivo es mediante la adición de
fertilizantes tales como el cloruro o muriato de potasio (KCl) que contiene
aproximadamente 50% de potasio y sulfato de potasio (K2SO4) (Boyd y
Wudtisin, 2011). Sin embargo, esta práctica es muy costosa debido a los
grandes volúmenes de agua existentes en las áreas de cultivo involucradas
(Zhu et al., 2006). Una alternativa más factible de mantener los niveles

51
adecuados de estos iones mejorando la capacidad osmoreguladora del
camarón es mediante la suplementación de estos iones en la dieta, esto puede
ser más eficiente y económicamente más viable, que la adición de los minerales
al agua de los estanques de cultivo (Gong et al., 2004).
Los efectos fisiológicos que puede presentar el camarón durante su
desarrollo por la deficiencia de iones en el agua pueden ser graves, tales como
anorexia, baja actividad, un crecimiento pobre e incluso la muerte, de ahí se
deriva la importancia de monitorear estos minerales en el agua del estanque
(Zhu et al., 2004).
Concentración de Nitrógeno Amoniacal Total (TAN) y Nitritos (NO2) en el
Agua del Estanque
La concentración del TAN en el agua del estanque en el Sistema de
Cultivo Cerrado permaneció constante durante toda la fase experimental,
obteniendo en promedio una concentración de 0.64 ± 0.6 mg/L. El nivel más
elevado registrado durante el estudio fue de 2.88 mg/L y el más bajo de 0.1
mg/mL (Figura 6).
Los niveles de nitritos en el agua del estanque del sistema se
incrementaron a partir del día 14 del período experimental, permaneciendo
elevados hasta casi el final del bioensayo con una concentración promedio de
4.84 ± 4.4 mg/L, siendo la concentración más elevada de 13.60 mg/L alcanzada
al día 17 del estudio (Figura 6).
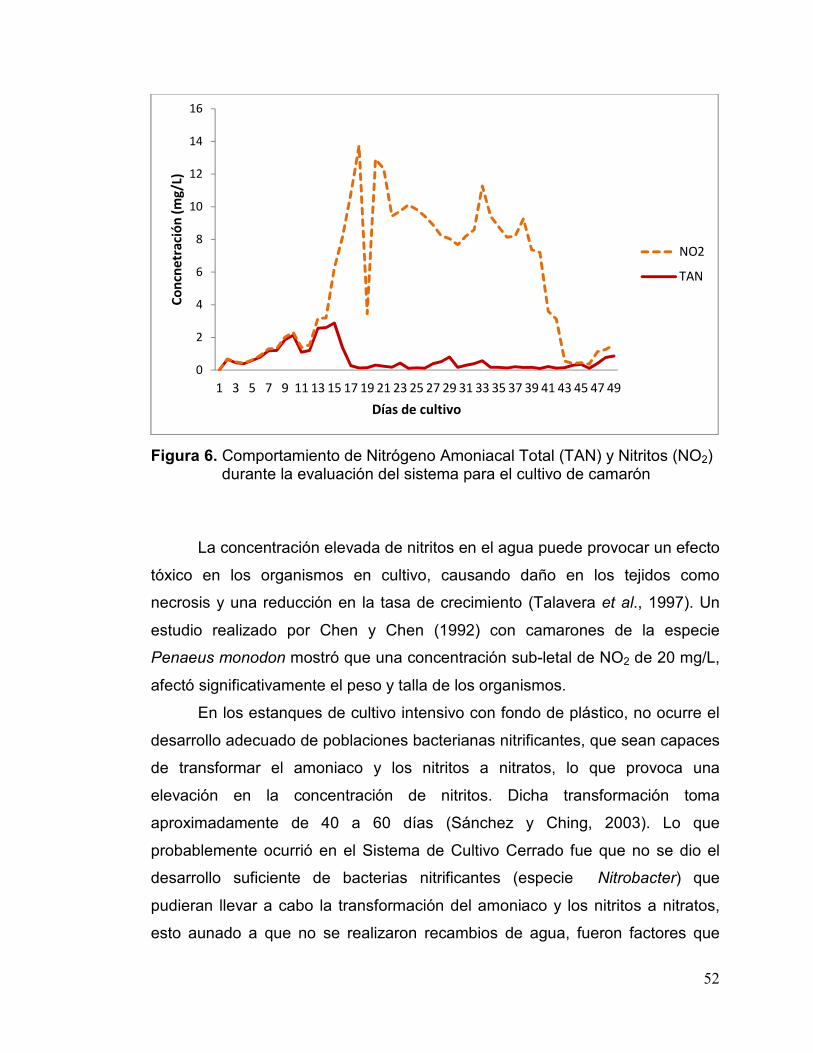
52
Figura 6. Comportamiento de Nitrógeno Amoniacal Total (TAN) y Nitritos (NO2) durante la evaluación del sistema para el cultivo de camarón
La concentración elevada de nitritos en el agua puede provocar un efecto
tóxico en los organismos en cultivo, causando daño en los tejidos como
necrosis y una reducción en la tasa de crecimiento (Talavera et al., 1997). Un
estudio realizado por Chen y Chen (1992) con camarones de la especie
Penaeus monodon mostró que una concentración sub-letal de NO2 de 20 mg/L,
afectó significativamente el peso y talla de los organismos.
En los estanques de cultivo intensivo con fondo de plástico, no ocurre el
desarrollo adecuado de poblaciones bacterianas nitrificantes, que sean capaces
de transformar el amoniaco y los nitritos a nitratos, lo que provoca una
elevación en la concentración de nitritos. Dicha transformación toma
aproximadamente de 40 a 60 días (Sánchez y Ching, 2003). Lo que
probablemente ocurrió en el Sistema de Cultivo Cerrado fue que no se dio el
desarrollo suficiente de bacterias nitrificantes (especie Nitrobacter) que
pudieran llevar a cabo la transformación del amoniaco y los nitritos a nitratos,
esto aunado a que no se realizaron recambios de agua, fueron factores que
0
2
4
6
8
10
12
14
16
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39 41 43 45 47 49
Co
ncn
etr
aci
ón
(m
g/L
)
Días de cultivo
NO2
TAN

53
aumentaron la acumulación de nitritos en el agua, provocando un efecto en los
organismos, manifestándose como necrosis en la cutícula. La Figura 6 muestra
el registro de TAN y NO2 durante la evaluación del sistema cerrado.
Otro de los elementos que favorecen el incremento de compuestos
nitrogenados como amonio y nitritos es el contenido proteico del alimento (Cai
et al., 1996). Un estudio realizado por Rosas et al. (1996) en el cual alimentaron
postlarvas de Penaeus setiferus, P. schmitti, P. duorarum y P. notialis con
dietas conteniendo diferente concentración de proteína, demostró que la
excreción de compuestos nitrogenados incrementó conforme aumentaba la
cantidad de proteína ingerida por el organismo.
El porcentaje de proteína contenida en la dieta del camarón utilizada en
este estudio fue de 35%, considerada adecuada para camarones de la especie
L. vannamei en etapa juvenil y adulta, por lo que se estima que el incremento
en la concentración de los nitritos en el agua fue debido principalmente a la
deficiencia de bacterias nitrificantes en el estanque más que al contenido
proteico de la dieta administrada.
El pH y la temperatura también son factores importantes en la reducción
de los niveles de nitritos en el agua (Re et al., 2004), pero en este estudio,
ambos parámetros permanecieron estables y dentro de los niveles óptimos para
el desarrollo del camarón, por lo que se considera que no influyeron en la
elevación de los niveles de nitritos.
Otra de las ventajas identificadas con este sistema, se relaciona con el
volumen de agua de mar utilizada durante el cultivo de los camarones, ya que
en el sistema tradicional, el consumo promedio mensual de agua es de 34,100
m3/ha (Reyes et al., 2009), mientras que en el Sistema Cerrado de Cultivo fue
de 12,000 m3/ha, lo que significa un consumo 3 veces menor que en el sistema
abierto. Este ahorro en la utilización del agua tiene un impacto favorable al
ecosistema, ya que se reducen las descargas de agua que son vertidas al
medio ambiente.

54
Evaluación de Lesiones Internas y Externas en Camarón Litopenaeus vannamei
Las observaciones microscópicas realizadas durante la evaluación de los
organismos mantenidos en el Sistema de Cultivo Cerrado, mostraron que en
general el estado de salud de los camarones fue adecuado, ya que no se
detectaron bacterias filamentosas o ectoparásitos en branquias.
En el intestino no se encontraron gregarinas ó gametocitos y en el
hepatopáncreas, los túbulos y la concentración de lípidos estuvieron dentro de
los valores establecidos como adecuados. El 17% del total de los camarones
analizados presentó túbulos totalmente estrangulados, 5% de los organismos
mostraron túbulos estrangulados en distintas regiones y solo un 4% exhibió
túbulos necrosados y secos. El 16% de los camarones presentó una
concentración lipídica inadecuada en el hepatopáncreas. La Tabla 5 muestra la
presencia de lesiones en los distintos órganos de camarón durante la
evaluación de su desarrollo en un Sistema de Cultivo Cerrado.
El hepatopáncreas juega un papel fundamental en las funciones
digestivas del camarón, pues es ahí donde se absorben y almacenan los
nutrientes, por tanto, cualquier disfunción en este órgano altera severamente el
desarrollo de los organismos (Gómez et al., 2001). El hepatopáncreas de
camarón puede ser afectado por bacterias como las del género Vibrio y por
virus. La enfermedad denominada Hepatopancreatitis Necrotizante (NHP), está
relacionada con un daño severo al hepatopáncreas, ocasionando una
disminución de la cantidad de lípidos presentes en este órgano y la presencia
de túbulos melanizados en cantidades variables, los cuales dan al
hepatopáncreas una coloración pálida con vetas obscuras indicativo de túbulos
melanizados, esto acompañado de una consistencia blanda y acuosa con fluido
en el centro (Lightner y Pantoja, 2001).
Cuando se realiza la determinación de lípidos es importante considerar si
el camarón se encuentra en etapa de muda o no, ya que los organismos toman
los lípidos que se encuentran en los túbulos del hepatopáncreas (el principal
sitio de almacenamiento de lípidos), como una fuente de energía para llevar a

55
cabo este proceso fisiológico (Gong et al., 2004). Por ello, es importante
monitorear este parámetro mediante observaciones microscópicas, ya que es
indicativo del estado de nutrición y salud del camarón.
Tabla 5. Evaluación del daño en los distintos órganos de camarón Litopenaeus vannamei cultivado en un sistema cerrado
Análisis Bacteriológico (Branquias, Hemolinfa y Hepatopáncreas) en Camarón Litopenaeus vannamei
Los recuentos bacterianos en camarón incluyeron la cuenta total
bacteriana y la determinación de bacterias presuntivas de Vibrio aisladas de los
tejidos.
Órgano Daño
Presencia/ Ausencia
de indicadores
(%) de daño
en los órganos
Necrosis Ausencia 0
Branquias Ectoparásitos Ausencia 0
(n=57) Bacterias Ausencia 0
Filamentosas
Túbulos
Túbulos lisos 74
Túbulos estrangulados 17
Hepatopáncreas Túbulos estrangulados en diferentes partes 5
(n=57) Túbulos estrangulados secos y con
necrosis 4
Lípidos
Concentración adecuada de lípidos 84
50% de túbulos con lípidos 14
25% de túbulos con lípidos 2
Intestino Gregarinas Ausencia 0
(n=57) Gametocitos Ausencia 0

56
Cuenta Total Bacteriana en Tejidos de Camarón
Las cuentas bacterianas en los diferentes tejidos de camarón
permanecieron relativamente bajas, siendo en hemolinfa donde se encontró la
menor cantidad de bacterias totales (4.3x102 UFC/mL), en branquias la mayor
con 1.6x104 UFC/g y en hepatopáncreas se contabilizaron en promedio 1.1x104
UFC/g (Tabla 6).
Gómez-Gil et al. (1998) establecieron que conteos bacterianos menores
de 103 UFC/g o mL de muestra, son considerados normales en larvas y
postlarvas de camarón, y recuentos superiores a 105 UFC/g o mL son
estimados como elevados en sistemas de cultivo abiertos. En este estudio, los
recuentos bacterianos totales obtenidos se encontraron en los niveles
considerados normales para camarón de esta especie. Esto es favorable ya que
el camarón fue cultivado en un sistema cerrado donde no hubo recambios de
agua, lo cual podría favorecer un incremento en el desarrollo bacteriano. Sin
embargo, esto no sucedió y se mantuvieron cuentas bacterianas bajas en los
camarones en cultivo.
Tabla 6. Cuenta Total Bacteriana en tejidos de camarón Litopenaeus Vannamei
Tejido Concentración (UFC/mL, g)
Hepatopáncreas 1.1 x 104
(n= 96)
Branquias 1.6 x 104
(n=32)
Hemolinfa 4.3 x 102
(n= 48)

57
Recuento de Bacterias del Género Vibrio Aisladas de los Distintos Tejidos de Camarón
Los recuentos promedio de bacterias presuntivas de Vibrio en los tejidos
del camarón permanecieron bajos, aunque se observó variabilidad durante el
estudio (Tabla 7). Se presentó una mayor incidencia de bacterias en branquias,
esto se explica por la función de filtración que cumple este órgano en el
camarón, ya que se encuentra en contacto directo con el agua del estanque,
que es donde las bacterias de Vibrio habitan de manera natural (Wu et al.,
2009). La mayor cantidad de UFC fueron amarillas, lo que las identifica como
cepas que utilizan la sacarosa como fuente de carbono con producción de
ácido. Algunas de las especies con estas características son V. alginolyticus, V.
cholerae, V. angillarum, V. fluvialis. La Tabla 7 muestra las cuentas bacterianas
obtenidas en los diferentes órganos del camarón, durante la evaluación del
Sistema de Cultivo Cerrado.
Las cuentas bacterianas obtenidas en hepatopáncreas de camarón
fueron mas bajas (4.4 x 103 UFC/g) que las obtenidas por Gómez-Gil et al.
(1998), donde encontraron 4.30x104 UFC/g de bacterias Vibrio en
Tabla 7. Evaluación de bacterias de Vibrio sacarosa (+) aisladas de diferentes tejidos de camarón Litopenaeus vannamei
Tejido Concentración (UFC/mL, g)
Hepatopáncreas 4.4 x 103
(n= 96)
Branquias 1.1 x 104
(n=32)
Hemolinfa 1.0 x 102
(n= 48)

58
hepatopáncreas de camarones en etapa juvenil mantenidos en tanques con un
sistema con recirculación de agua. Estos mismos autores reportaron en
hemolinfa conteos de 1.27x103 UFC/g de bacterias de Vibrio, los cuales fueron
más elevados a los de nuestro estudio (1.0x102 UFC/g). Se ha sugerido que la
presencia de bacterias en la hemolinfa es indicativo de septicemia y una
secuela común de estrés.
Gómez-Gil et al. (1998) estimó que los conteos de colonias bacterianas
amarillas presuntivas de Vibrio en hepatopáncreas menores a 105 UFC/g son
considerados aceptables en camarones que se encuentran en etapa juvenil y
adulta, mientras que recuentos bacterianos superiores a 105 UFC/g son
perjudiciales para los organismos. Los valores promedio obtenidos en nuestro
estudio estuvieron dentro del intervalo considerado como adecuado para este
género bacteriano.
También se ha reportado que cuentas bacterianas menores a 103
UFC/mL en hemolinfa de camarón son aceptables, por lo que los valores
obtenidos en este estudio fueron adecuados, ya que se encontraron muy por
debajo (Gómez-Gil et al., 1998).
Los conteos de bacterias totales y presuntivas del género Vibrio nos
permitió conocer si el estado de salud de los camarones era bueno, y que no
había alguna infección bacteriana que limitara su desarrollo. Es importante
reiterar que las bacterias del género Vibrio se encuentran de manera natural en
el agua marina de los estanques. Por ello, es relevante monitorear
constantemente los distintos parámetros de calidad del sistema de cultivo, como
es el agua del estanque, el estado de nutrición y salud del camarón, así como
los parámetros fisicoquímicos que pueden alterar el metabolismo de los
organismos, para detectar a tiempo enfermedades bacterianas que podrían
causar grandes pérdidas económicas a esta industria (Gómez-Gil et al., 1998).

59
Detección de Especies de Vibrio Aisladas de Camarón de cultivo Litopenaeus
vannamei
La microbiota presente en los organismos marinos incluido el camarón,
está relacionada con el ambiente en el que viven. Las bacterias del género
Vibrio se encuentran distribuidas ampliamente en ambientes costeros y
estuarinos y son causantes de numerosas enfermedades, dando como
resultado elevadas mortalidades de los organismos en los estanques.
El generar información relacionada con las especies bacterianas que
colonizan a los crustáceos es de suma importancia, para detectar
anormalidades en éstos y realizar un diagnóstico adecuado de las
enfermedades bacterianas que afectan al camarón (Gomez-Gil et al., 1998).
De los aislados bacterianos obtenidos en este estudio a partir de los
diferentes tejidos de camarón (branquias, hepatopáncreas y hemolinfa), se
obtuvo la amplificación de fragmentos de ADN que han sido reportados
previamente para Vibrio, como fueron Vibrio spp, Vibrio gen de hemolisina y
Vibrio toxR.
El fragmento de ADN amplificado del gen del 16s RNA presentó
aproximadamente 663 pares de bases y correspondió con la amplificación del
fragmento de ADN reportado por Tarr et al. (2007), para la identificación de
Vibrio spp. También, Al-Mouqati et al. (2012) han reportado la utilización del gen
del 16s RNA para la identificación de Vibrio spp.
Este fragmento de ADN se encontró en el 63% de las cepas que fueron
aisladas principalmente de branquias y hemolinfa de camarón. La Figura 7
muestra un gel de agarosa donde se aprecia una banda con un peso molecular
aproximado de 663 pb, que coincide con el peso molecular del marcador
utilizado.

60
Figura 7. Amplificación mediante PCR del gen 16s RNA Carril 1: Marcador de peso molecular Bio-Rad (50-2000 pb); Carriles 2,3,10,12,15,16,20 muestras de branquias; carriles 4,5,6,8,9,13,14, 17,18,19 muestras de hemolinfa; Carriles 7 y 11 muestras de hepatopáncreas. B: Branquias, Hp: Hepatopáncreas, He: Hemolinfa.
El fragmento de ADN del gen de hemolisina amplificado estuvo
conformado por aproximadamente de 308 pares de bases, lo cual fue reportado
en la identificación de V. harveyi gen de la hemolisina por Conejero y Hedreyda
(2004), así como por Zhang et al. (2001) y Parvathi et al. (2009), quienes
obtuvieron fragmentos de ADN con ese mismo peso molecular. En esta
investigación, la amplificación de este segmento de ADN se presentó en el 61%
de las cepas aisladas, siendo en hemolinfa donde se encontró con mayor
frecuencia.
En la Figura 8, se presenta un gel de agarosa donde se aprecia la banda
con un peso molecular de 308 pb correspondiente a la identificación de Vibrio
con gen de hemolisina.

61
Figura 8. Amplificación mediante PCR del gen de hemolisina en cepas de Vibrio Carril 1: Marcador de peso molecular Bio-Rad (50-2000 pb); Carriles 2,3,10,12,15,16,20 muestras de branquias; Carriles 4,5,6,8,9,13,14, 17,18,19 muestras de hemolinfa; Carriles 7 y 11 muestras de hepatopáncreas. B: Branquias, Hp: Hepatopáncreas, He: Hemolinfa.
Además, se obtuvo la amplificación de un fragmento de ADN del gen
toxR de Vibrio conformado por 390 pares de bases en 10 de las cepas aisladas
(Figura 9). Este fragmento de ADN correspondiente al gen toxR ha sido
reportado por Conejero y Hedreyda (2003). La amplificación de este gen se
presentó mayormente en hemolinfa. Las bandas obtenidas en el gel de agarosa
coinciden con las reportadas por estos autores al utilizar los mismos iniciadores,
lo que nos indica que se trata de esta especie. La Figura 9 muestra un gel de
agarosa, donde se aprecia la amplificación del fragmento de ADN con un peso
molecular de 390 pb que corresponde al gen toxR de Vibrio.

62
Figura 9. Amplificación mediante PCR del gen toxR en cepas de Vibrio Carril 1: Marcador de peso molecular Bio-Rad (50-2000 pb); Carriles 2,3,10,12,15,16,20 muestras de branquias; Carriles 4,5,6,8,9,13,14, 17,18,19 muestras de hemolinfa; Carriles 7 y 11 muestras de hepatopáncreas. B: Branquias, Hp: Hepatopáncreas, He: Hemolinfa.
El estudiar el género Vibrio es muy importante desde el punto de vista de
la salud humana, así como en relación a los organismos acuáticos, como es el
caso del camarón. El haber detectado en este estudio cepas de Vibrio con el
gen de hemolisina y toxR fue importante ya que éstos pueden ser empleados
como un marcador de la potencial patogenicidad de estas bacterias para el
camarón (Conejero y Hedreyda, 2004).
Gómez-Gil et al. (1998), aislaron bacterias a partir de hemolinfa de
camarones “aparentemente sanos”, observando que más del 50% de estas
bacterias aisladas correspondieron al género Vibrio spp. Por otro lado, Chen et
al. (1992), estudiaron dos eventos infecciosos en los cuales ocurrió una alta
mortalidad en camarones de la especie P. monodon; los aislamientos
bacterianos realizados de hepatopáncreas de los camarones mostraron que en
el primer evento el 73.4% de las bacterias correspondieron a Vibrio spp y en el
segundo evento infeccioso este género se presentó en un 84.6%. Las

63
principales especies de Vibrio identificadas fueron V. damsela con un 16.6% y
22.4% y V. harveyi en un 43.1% y 26.9%, respectivamente.
En esta investigación no se detectó Vibrio Cholerae, V. vulnificus y V.
harveyi en las muestras analizadas, lo cual es muy importante en relación a la
salud del consumidor, ya que estas especies son clínicamente significativas
como patógenos para humanos. Las infecciones causadas por estos
organismos están usualmente asociadas a la ingestión de productos marinos
contaminados y los síntomas que manifiesta el paciente van desde
gastroenteritis, infección a la piel, septicemia hasta la mortalidad en el caso de
pacientes inmunocomprometidos o con bajas defensas (Leyton y Riquelme,
2008).
Vibrio habita en aguas marinas, sedimentos y en la superficie de
animales marinos. Es también un miembro común de la flora intestinal natural
de vertebrados e invertebrados acuáticos (Pang et al., 2006). Esta bacteria es
considerada un patógeno dominante que puede infectar postlarvas,
ocasionando grandes mortalidades (Parvathi et al., 2009). Existen diversos
factores que contribuyen a la virulencia de esta especie bacteriana, entre los
que se incluye la producción de algunas enzimas como proteasas, lipasas,
fosfolipasas, hemolisinas y citotoxinas, entre otras (Austin et al., 2003; Zhang et
al., 2001).
Se han desarrollado algunos métodos de PCR para identificar aislados
de Vibrio como son el gen de la hemolisina y el gen toxR (Oakey et al., 2003),
los cuales han sido estudiados en esta especie (Franco y Hedreyda, 2006). Las
metodologías aplicadas en esta investigación para la detección e identificación
del gen de hemolisina y toxR, así como de Vibrio spp se basaron en los mismos
métodos utilizados por dichos autores.
El gen de la hemolisina es una exotoxina responsable de la lisis de
eritrocitos y daño en células hospederas. Las hemolisinas son producidas por
varias especies bacterianas que incluyen Escherichia coli, Pseudomonas
aeruginosa y vibrios. En la mayoría de los casos, existe evidencia experimental
y epidemiológica que sugiere que la expresión de genes como el de toxR y de

64
hemolisina está relacionado con la patogénesis de la enfermedad. Hemolisina
es la toxina más ampliamente distribuida entre las bacterias patógenas de
Vibrio y pueden ejercer distintos mecanismos durante procesos de infección
(Zhang y Austin, 2005).
Por otro lado, en bacterias del género Vibrio el gen toxR es un regulador
transmembranal de transcripción, el cual controla la expresión coordinada de
los genes de virulencia (Franco y Hedreyda, 2006). Este gen es considerado un
marcador taxonómico efectivo en la identificación de Vibrio spp. (Kim et al.,
1999); se encuentra presente en algunas especies de Vibrio como, V. cholerae,
V. parahaemolyticus, V. fischeri, V. vulnificus, V. alginolyticus, V. mimicus, entre
otros (Pang et al., 2006). Conejero y Hedreyda (2003), demostraron la utilidad
de analizar la secuencia del gen toxR para la diferenciación de Vibrio de otras
especies de Vibrio filogenéticamente relacionadas.
La persistencia y sobrevivencia de Vibrio en los estanques de cultivo de
camarón ha sido atribuida a su habilidad para formar biopelículas y a su
resistencia a desinfectantes y antibióticos (Austin y Zhang, 2006).
Concentración Mínima Inhibitoria (CMI) de la Oxitetraciclina en Cepas de Vibrio Aisladas de Camarón
La oxitetraciclina es uno de los antibióticos mas comúnmente utilizados
en la acuicultura, por su efectividad contra bacterias de Vibrio, que es el
microorganismo más comúnmente observado en los sistemas de cultivo de
camarón (Bray et al., 2006; Han et al., 2007). La Tabla 8 muestra los valores de
las Concentraciones Mínimas Inhibitorias (CMI) de OTC para Vibrio en los
diferentes tejidos del camarón cultivado en el sistema cerrado.

65
Tabla 8. Concentración Mínima Inhibitoria de la oxitetraciclina en bacterias de Vibrio aisladas de los diferentes tejidos de camarón
La sensibilidad a oxitetraciclina mostrada por las cepas bacterianas de
Vibrio, aisladas de camarón varió entre los distintos tejidos, siendo en las cepas
aisladas de branquias donde se requirió la mayor concentración del antibiótico
para provocar una inhibición, encontrándose en un intervalo de 0.78 a 325
µg/mL de OTC. En las bacterias aisladas de hemolinfa se obtuvo la menor
sensibilidad bacteriana, requiriéndose niveles de 0.78 a 75 µg/mL de OTC para
obtener un efecto inhibitorio.
Jayasree et al. (2006), aseveraron que bacterias del género Vibrio
aisladas de hemolinfa de camarón, fueron sensibles al antibiótico OTC. Por su
parte Santiago (2009), en un estudio realizado en la misma granja acuícola,
estableció valores de la CMI en un intervalo de 0.75-100 µg/mL para OTC en
bacterias de Vibrio aisladas de hepatopáncreas de camarón Litopenaeus
vannamei. En general estos valores fueron similares a los obtenidos en esta
investigación. La sensibilidad que muestran las bacterias a los antibióticos
puede variar entre las distintas especies, ya que dentro de un mismo sistema de
cultivo, existe gran diversidad de especies bacterianas que interactúan de
forma natural con los organismos. El uso continuo e irracional de antibióticos,
principalmente OTC en las granjas camaronícolas, ha ocasionado que las
bacterias desarrollen mecanismos de resistencia reduciendo la acción de este
antibiótico (Santiago et al., 2009; Han et al., 2007).
Tejido Concentración de OTC (µg/mL)
Hepatopáncreas 0.39 a 100
(n= 23)
Branquias 0.78 a 325
(n= 6)
Hemolinfa 0.78 a 75
(n= 7)

66
RESUMEN DE HALLAZGOS
El Sistema de Cultivo Cerrado permitió mantener controlados los
parámetros fisicoquímicos, excepto la salinidad y la concentración de nitritos,
los cuales estuvieron por arriba de los valores reportados como adecuados
para esta especie de camarón. La elevada salinidad, no influyó en la ganancia
de peso y la sobrevivencia de los organismos, mientras que las altas
concentraciones de nitrititos, causaron daño en la cutícula de camarón.
El 26 % de los camarones mantenidos en el Sistema de Cultivo Cerrado,
presentaron lesiones en hepatopáncreas en el intervalo de leve a moderado; el
16% de éstos mostró valores inadecuados de lípidos en dicho órgano. En
branquias no se detectaron ectoparásitos y bacterias filamentosas y en el
intestino no se observaron gametocitos y gregarinas.
Se detectó la presencia de bacterias del género Vibrio spp., Vibrio gen de
hemolisina y Vibrio gen toxR. La detección de estos genes es importante ya que
son indicativos de la potencial patogenicidad de estas bacterias que afectan al
camarón cultivado así como al humano.
Las cepas de Vibrio aisladas mostraron sensibilidad a oxitetraciclina con valores
de Concentración Mínima Inhibitoria en los intervalos de 0.78-75 µg/mL en
hemolinfa, para hepatopáncreas de 0.39-100 µg/mL y en branquias de 0.78-325
µg/mL, requiriéndose en las cepas aisladas de este ultimo órgano una mayor
concentración del antibiótico para causar un efecto inhibitorio.

67
CONCLUSIONES
El empleo del Sistema de Cultivo Cerrado permitió incrementar la
producción de camarón Litopenaeus vannamei en una menor área de cultivo,
manteniendo un adecuado desarrollo de los organismos y reduciendo el
impacto que esta actividad pueda causar al medio ambiente.
No se detectó Vibrio cholerae, V. vulnificus y V. harveyi en las muestras
de camarón analizadas, aspecto importante tanto para la salud de los
organismos cultivados como para el consumidor.
Se recomienda en los Sistemas de Cultivo Cerrados una vigilancia
estricta de la concentración de nitritos en el agua, ya que esto tiene un impacto
adverso en el desarrollo del camarón y puede afectar externamente la
apariencia del producto final en detrimento de su comercialización.
La implementación de este sistema, puede representar una alternativa
viable en el desarrollo y mejora de esta industria, de gran importancia
económica para la región y el país.

68
BIBLIOGRAFÍA
Aguirre-Guzmán, G. (2004). ¿Los Vibrios spp, son agentes patógenos importantes para el cultivo de camarón? Programa Nacional de Sanidad Acuícola y la Red de Diagnóstico. CONAPESCA. Universidad Autónoma de Tamaulipas. 1(25):1-3.
Allison D.G., Gilbert P., Lambert P.A. (2001). Antimicrobial action: basic principles. Molecular Medical Microbiology Sussman, M. Ed. pp. 585-590.
Al-Mouqati, S., Azad, I.S., Al-Baijan, D., Benhaji, A. (2012). Vibrio detection in market Seafood samples of Kuwait by biochemical (API20E) strips and its evaluation against 16s r DNA-based molecular methods.Reserch Journal of Biotechnology. 7(3):63-69.
Álvarez, J., Austin, B., Álvarez, A., Agurto, C. (2000). Especies de Vibrio y Aeromonas aisladas del intestino de camarones marinos sanos silvestres y cultivados en Venezuela. Veterinaria Tropical. 25(1): 5-27.
Álvarez, J.D., Agurto, C.P., Álvarez A.M., Obregón, J. (2004). Resistencia antimicrobiana en bacterias aisladas de tilapias, agua y sedimento en Venezuela. Revista Científica. 14(006): 1-17.
Anaya-Rosas, R.E. (2005). Cultivo de camarón blanco, Litopenaeus vannamei, Boone (1931), en sistema cerrado a alta densidad. Tesis de maestría. Centro de Investigación Científica y de Educación Superior de Ensenada.
Arredondo, J.L., Ingle, G., Guerrero, I., Ponce, J.T. Barriga, I. de los A. (2007). Tasas de remoción de amoniaco y nitritos en un sistema cerrado de recirculación de agua, bajo tres cargas de trucha arco iris Oncorhynchus mykiss. Revista Mexicana de Ingeniería Química. 6 (3): 301-308.
Armstrong, D.A., Stephenson, J.J. y Knight, A. (1976). Acute toxicity of nitrite of the giant Malasyan prawn Macrobrachium rosenbergii. Aquaculture, 9: 39-46.
Auró, A., Ocampo, L. (2006). El libro del camarón. México, D.F. pp. 83, 84, 87, 91, 203, 204.
Austin, B., Zhang, X. (2006). Vibrio harveyi: a significant pathogen of marine vertebrates and invertebrates. Letters in Applied Microbiology, 43: 119-124.
Austin, B., Pride, A.C., Rhodie, G.A. (2003) Association of a bacteriophage with virulence in Vibrio harveyi. Journal of Fish Diseases, 26:55-58.
Baquero, F., Martínez, J.L., Cantón, R. (2009). Antibiotics and antibiotic resistance in water environments. Current Opinion in Biotechnology, 19: 260-265.
Belsher, D., León, M., Quispe, M. (2011). Granja intensiva del Perú productora de camarón en invernadero. Industria Acuícola. 7(5):8-9.
Betancourt, O., Scarpa, C., Villagrán, K. (2003). Estudio de resistencia de cepas de Staphylococcus aureus aisladas de mastitis subclínica bovina frente a

69
cinco antibióticos en tres sectores de la IX región de chile (nota técnica). Revista Científica. 13(3): 413-417.
Boyd, C., Wudtisin, I. (2011). Possible Potassium and Magnesium Limitations for Shrimp Survival and Production in Low-Salinity, Pond Waters in Thailand. Journal of the World Aquaculture Society. 42 (6): 766-777.
Boyd, C.A., Boyd, C.E. Rouse, D.B. (2007). Potassium budget for inland, saline water shrimp ponds in Alabama. Aquacultural Engineering, 36:45–50.
Bray, W.A., Williams, R.R., Lightner, D.V., Lawrence, A.L. (2006). Growth, survival and histological responses of the marine shrimp, Litopenaeus vannamei, to three dosage levels of Oxytetracycline. Aquaculture, 258: 97-108.
Brito, R., Chimal, M.E., Rosas, C. (2000). Effect of salinity in survival, growth, and osmotic capacity of early juveniles of Farfantepenaeus brasiliensis (decapoda:penaeidae). Journal of Experimental Marine Biology and Ecology, 244: 253–263.
Caiffer, M. I., Terragno, R., González, F.S., Viñas, M.R., Pichel, M., Binsztein, N. (2007). Protocolos para la identificación y caracterización de Vibrio cholerae por PCR. En: Manual de procedimientos para aislamiento, identificación y caracterización de Vibrio cholerae.Organización Mundial de la Salud (WHO) Global Salm Surv. pp. 28-39.
Cai, Y., Wermerskirchen, J., Adelman, R. (1996). Ammonia excretion rate indicates dietary protein adequacy for fish. The progressive Fish-Culturist, 58: 124-127.
Caipang, C., Aguana, M. (2011). Conventional PCR assays for the detection of pathogenic Vibrio spp. in shrimp aquaculture in the Philippines. International Journal of the Bioflux Society. 4(3): 339-350.
Cervantes, F., Pérez, J., Gómez, Jorge. (2000). Avances en la Eliminación Biológica del Nitrógeno de las Aguas Residuales. Revista Latinoamericana de Microbiología, 42: 73-82.
Chávez, J. (2009). Manual de Análisis de Aguas: Criterios Biológicos en la Interpretación de los Análisis de Campo. pp. 2-3.
Chen, J.C. (2003). Manejo de desechos nitrogenados de ambientes acuícolas. Boletín Nicovita. 8 (2):1-4.
Chen, J.C., Lei, S.C. y Liu, P.C. (1990). Effect of ammonia and nitrite on Penaeus monodon juveniles. In: Hirano, Y. y Hanyu, C. (Eds.). The Second Asian Fisheries Forum. Tokyo, Japan. pp. 65-68.
Chen, J.C. y Chen, S.F. (1992). Effects of nitrite on growth and molting of Penaeus monodon exposed to ambient ammonia. Aquaculture, 109: 177-185.
Comité de Sanidad Acuícola (COSAES). (2010). El uso responsable de los antibióticos en las buenas prácticas de producción acuícola. Manual Técnico Servicio Nacional de Sanidad Inocuidad y Calidad Agroalimentaria y SAGARPA. p. 4.
Conejero, J.U., Hedreyda, C.T. (2003). Isolation and partial toxR gene of Vibrio harveyi and design of toxR-targeted PCR primer for species detection. Journal of Applied Microbiology, 95:602-611.

70
Conejero, J.U., Hedreyda, C.T. (2004). PCR detection of hemolysin (vhh) gene in Vibrio harveyi. Journal of General and Applied Microbiology, 50:137-142.
Crab, R., Avnimelech, Y., Defoirdt, T., Bossier, P., Verstraete, W. (2007). Nitrogen removal techniques in aquaculture for a sustainable production. Aquaculture, 270: 1-14.
Cunningham, C. (2002). Molecular diagnostics of fish and shellfish diseases: present status and potential use in disease control. Aquaculture, 206: 19-55.
Cuzón, G., Lawrence, A., Gaxiola, G., Rosas, Carlos., Guillaume, J. (2004). Nutrition of Litopenaeus vannamei reared in tanks or in ponds. Aquaculture, 235: 513-551.
Davis, D.A., Glatin, D.M. (1996). Dietary mineral requirements of fish and marine crustaceans. Reviews in Fisheries Science. 4(1):75-99.
Davis, D.E., Boyd, C.E., Rouse, D.B. (2005). Effects of Potassium, Magnesium and Age on Growth and Survival of Litopenaeus vannamei Post-Larvae Reared in Inland Low Salinity Well Waters in West Alabama. Journal of the World Aquaculture Society. 36 (3): 416-419.
De la Mora,G., Villarreal-Delgado, E., Arredondo-Figueroa, J., Ponce-Palafox, J., Barriga-Sosa, I. (2003). Evaluación de algunos parámetros de calidad del agua en un sistema cerrado de recirculación para la acuicultura, sometido a diferentes cargas de biomasa de peces. Hidrobiológica. 13(4): 247-253.
Dikow, R.B. (2011). Systematic relationships within the Vibrionaceae (Bacteria: Grammaproteobacteria): steps toward a phylogenetic taxonomy. Cladistics, 27: 9-28.
FAO. (2009). El estado mundial de la pesca y la acuicultura 2008. Roma, Italia. Flegel, T. (2012). Historic emergence, impact and current status of shrimp
pathogens in Asia. Journal of Invertebrate Pathology. 110(2): 166-173. Franco, P., Hedreyda, C. (2006). Amplification and sequence analysis of the full
length toxR gen in Vibrio harveyi. The Journal of General and Applied Micriobiology, 52: 281-287.
Franco, M.A., Blancheton, J.P., Deviller, G., Charrier, A., Le, J.Y. (2004). Effect of fish size and hydraulic regime on particulate organic matter dynamics in a recirculation aquaculture system: elemental carbon and nitrogen approach. Aquaculture, 239:179-198.
Frías-Espericueta, M.G., Harfush-Meléndez, M. y Páez-Osuna, F. (2000). Effects of ammonia on mortality and feeding of postlarvae shrimp Litopenaeus vannamei. Bulletin of Environmental Contamination and Toxicology, 65: 98-103.
Galaviz, S. L., Molina, G. Z. J., González, G. J. R., Ibarra, G. J. C. (2007). Patógenos que disminuyen la calidad del camarón de cultivo. Revista Salud Pública y Nutrición, 12:419–425.
Galaviz-Silva, L., Molina-Garza, Z.J., Alcocer-González, J.M., Rosales Encina, J.L., Ibarra-Gámez, J.C. (2004). White spot syndrome virus genetic variants detected in Mexico by a new multiplex PCR method. Aquaculture, 242:53–68.

71
Gentry, W. C., Hutcheson, H., Kim, L., Bolte, D., Traub, D. J. y Morley, P. (2002). Identification of two phylogenetically related organisms from feces by PCR for detection of Salmonella spp. Journal of Clinical Microbiology, 40: 1487-1492.
Gómez-Gil, B., Tron-Mayén, L., Roque, A., Turnbull, J.F., Inglis, V., Guerra-Flores, A.L. (1998). Species of Vibrio spp. isolated from hepatopancreas, hemolymph and digestive tract of a population of health juvenile Penaeus vannamei. Aquaculture, 163:1-9.
Gómez, G.B., Roque, A., Guerra, F.A. (2001). Enfermedades infecciosas más comunes en la camaronicultura en México y el impacto del uso de antimicrobianos. En: Páez-Osuna F. (ed.), Camaronicultura y Medio Ambiente. UNAM. pp. 315-346.
Gong, H., Jiang, D.H., Lightner, D.V., Collins, C., Brock, D. (2004). A dietary modification approach to improve the osmoregulatory capacity of Litopenaeus vannamei cultured in the Arizona desert. Aquaculture Nutrition, 10: 227–236.
González, B. J. (2003). Técnicas de bacteriología, análisis en fresco, calidad de agua y buenas prácticas de manejo y bioseguridad en granjas camaroneras. Programa de Capacitación. Mazatlán, Sinaloa.
González-Lama, Z., Soria, S., Díez, A., Tejedor M.T., Lupiola P. (2001). Susceptibilidad in vitro a antimicrobianos y producción de ß-lactamasas de Vibrio y Photobacterium marinos. Boletín del Instituto Español de Oceanografía. 17 (3 y 4): 233-237.
Gutiérrez-Salazar, G.J., Molina-Garza, Z.J., Hernández-Acosta, M., García-Salas, J.A., Mercado-Hernández, R., Galaviz-Silva, L. (2011). Pathogens in Pacific white shrimp (Litopenaeus vannamei Boone, 1931) and their relationship with physicochemical parameters in three different culture systems in Tamaulipas, México. Aquaculture, 321: 34-40.
Han, F., Walker, R., Janes, M., Priyawiwatkul, W. y Ge, B. (2007). Antimicrobial Susceptibilities of Vibrio parahaemolyticus and Vibrio vulnificus Isolates from Louisiana Gulf and Retail Raw Oysters. Applied and Environmental Microbiology. 73(21): 7096-7098.
Hernández, G., Olmos, J. (2003). Molecular identification of pathogenic and nonpathogenic strains of Vibrio harveyi using PCR and RAPD.
http://www.fao.org/docrep/field/003/ab492s/AB492S04.htm http://www.horcalsa.com/contents/content-files/revista1.pdf http://www.vibriobiology.net/ Hou, C., Wang, F., Shuanglin, D., Zhu, Y., Ting, Y. (2012). Effects of constant
Ca2+ concentration in salinity fluctuations on growth and energy budget of juvenile Litopenaeus vannamei. Aquaculture International, 20:177–188.
Ibarra, C., Reyes, J., Galavíz, L., Luna, C., Molina, Z.J., Tirado, M.S. (2004). Detección y prevalencia de Vibrio spp. en cultivo de camarón Litopenaeus vannamei en Sonora durante el ciclo 2003. Programa Nacional de Sanidad Acuícola y la Red de Diagnóstico. 2(26): 7-9.
Isla, M.M. (2007). Water quality in recirculating aquaculture systems for arctic charr (salvelinus alpinus L.) culture. Centro de Investigaciones Pesqueras. La Habana, Cuba.

72
Jayasree, L., Janakiram, P., Madhavi, R. (2006). Characterization of Vibrio spp. Associated with Diseased Shrimp from Culture Ponds of Andhra Pradesh (India). Journal of the World Aquaculture Society. 37 (4): 523-532.
Jiang, D.H., Lawrence, A.L., Neill, W.H., Gong, H. (2000). Effects of temperature and salinity on nitrogenous excretion by Litopenaeus vannamei juveniles. Journal of Experimental Marine Biology and Ecology, 253:193–209.
Jiang, L.X., Pan, L.G., Bo, F. (2005). Effect of dissolved oxygen on immuneparameters of the white shrimp Litopenaeus vannamei. Fish and Shellfish Immunology, 18:185-188.
Jiménez-Sáenz, A.J. (2012). Sistemas de recirculación en acuicultura: una visión y retos diversos para Latinoamérica. Industria Acuícola. 8 (5): 4-8.
Kaysner, C.A., DePaola, A. (2004). Bacteriological analytical manual. Capítulo 9. Food and Drug Administration (FDA). Ver: http://www.fda.gov/Food/ScienceResearch/LaboratoryMethods/BacteriologicalAnalyticalManualBAM/ucm070830.htm
Kemper, N. (2008). Veterinary antibiotics in the aquatic and terrestrial environment. Ecological indicators, 8: 1–13.
Kim, Y.B., Okuda, J., Matsumoto, C., Takahashi, N., Hashimoto, S., Nishibuchi, M. (1999). Identification of Vibrio parahaemolyticus at the species level by PCR targeted to the toxR gene. Journal of Clinical Microbiology, 37: 1173–1177.
Kitiyodom, S., Khemtong, S., WongtavatchaI, J., Chuanchuen, R. (2010). Characterization of antibiotic resistance in Vibrio spp. isolated from farmed marine shrimps (Penaeus monodon). Federation of European Microbiological Societies, 72: 219-227.
Kuhn, D.D., Smith, S.A., Boardman, G.D., Angier, M.W., Marsh, L., Flick, G.J. (2010). Chronic toxicity of nitrate to Pacific white shrimp, Litopenaeus vannamei: Impacts on survival, growth, antennae length, and pathology. Aquaculture, 309: 109-114.
Lee, J.B., Chung, H.H., Chung Y.H., Lee, K.G. (2007). Development of an analytical protocol for detecting antibiotic residues in various foods. Food Chemistry, 105: 1726–1731.
Leyton, Y., Riquelme, C. (2008). Vibrios en los sistemas marinos costeros. Revista de Biología Marina y Oceanografía. 43(3): 441-456.
Li, C.C., Chen, J.C. ( 2008). The immune response of white shrimp Litopenaeus vannamei and its susceptibility to Vibrio alginolyticus under low and high pH stress. Fish & Shellfish Immunology, 25: 701-709.
Li E., Chen L., Zeng C., Chen X., Yu N., Lai Q., Qin J. G. (2007). Growth, body composition, respiration and ambient ammonia nitrogen tolerance of the juvenile white shrimp Litopenaeus vannamei, at different salinities. Aquaculture, 265:385-390.
Li E., Chen L., Zeng C., Yu N., Xiong Z., Chen X., Qin J.G. (2008). Comparison of digestive and antioxidant enzymes activities, haemolymph oxyhemocyanin contents and hepatopancreas histology of white shrimp, Litopenaeus vannamei, at various salinities. Aquaculture, 274: 80-86.

73
Lightner, D.V. (1996). A Handbook of shrimp pathology and diagnostic procedures of diseases of cultured penaeid shrimp. World Aquaculture Society, Baton Riuge, Louisiana, USA. p. 304.
Lightner, D.V., Pantoja, C.R. (2001). Manual para el diagnóstico de enfermedades del camarón. Departamento de Agricultura de los Estados Unidos (USDA). pp.92.
Limsuwan, Ch. (2005). Cultivo intensivo del camarón blanco: Resumen de visitas y conferencias a camaroneras del Perú En: Boletín Nicovita Camarón de mar.
Ling-Xu, Lu- Qing, Fango-Bo. (2005). Effect of dissolved oxygen on inmune parameters of the white shrimp Litopenaeus vannamei. Fish and Shellfish Inmunology, 18: 185-188.
López-Torres, M.A. (1998). El género Vibrio y su importancia en la acuacultura. Boletín del Programa Nacional de Sanidad Acuícola y la Red de Diagnóstico. SEMARNAP. 1(3): 8-11.
McBeath, A.J.A., Burr, K.L.-A., Cunningham, C.O. (2000). Development and use of a DNA probe to confirmation of cDNA from infectious salmon anaemia virus (ISAV) in PCR products. The European Association of Fish Pathology, 20: 130-134.
McGraw, W.J., Scarpa, J. (2003). Minimum environmental potassium for survival of pacific white shrimp Litopenaeus vannamei (boone) in freshwater. Journal of Shellfish Research. 22(1): 263-267.
McNevil, A. A., Boyd, C.E., Silapajam, O., Silapajam, K. (2004). Ionic supplementation of pond waters for inland culture of marine shrimp. Journal of the World Aquaculture Society, 35:460-467.
Mendoza-Patiño N., Campos-Sepúlveda E. (2008). Actualidades farmacológicas Tetraciclinas. Revista de la Facultad Médica de la UNAM. 51(1): 29-32.
Morales, C.M.S. (2004). Enfermedades del camarón: Detección mediante análisis en fresco e histopatología. Ed. Trillas. México, D.F. pp 122.
Nishibuchi, M., DePaola, A. (2005). Vibrio species. En: Fratamico, P.M., Bhunia, A.K., Smith, J.L. (eds). Foodborne Pathogens Microbiology and Molecular Biology. Caister Academic Press. Wymondham. pp. 251-270.
Oakey, H.J., Levy, N., Bourne, D.G., Cullen, B., Thomas, A. (2003) The use of PCR to aid in the rapid identification of Vibrio harveyi isolates. Journal of Applied Microbiology, 95: 1293–1303.
Olsen, J .E. (2000). ADN-based methods for detection of food-borne bacterial pathogens. Food Research International, 33: 257-266.
Pan, L., Zhang, L., Liu, H. (2007). Effects of salinity and pH on ion-transport enzyme activities, survival and growth of Litopenaeus vannamei postlarvae. Aquaculture, 273:711-720.
Pang, L., Zhang, H., Zhong, Y., Chen, J., Li, Y., Austin, B. (2006). Identification of Vibrio harveyi using PCR amplification of the toxR gene. Letters in Applied Microbiology, 43:249–255.
Parvathi, A., George, J., Kumar, S. (2009). Prevalence and heterogeneity of hemolysin gene vhh among hatchery isolates of Vibrio harveyi in India. Current Microbiology, 59: 42-47.

74
Pillay, T.V.R. y Kutty, M.N. (2005). Aquaculture, Principles and Practices, 2nd Ed. Blackwell Publishing Ltd, Oxford, UK. p. 630.
Pruzzo, C., Huq, A., Colwell, R., Donelli, G. (2005). Pathogenic Vibrio Species in the Marine and Estuarine Environment. Oceans and Health: Pathogens in the Marine Environment. Belkin and Colwell, Springer, New York. pp. 228-238.
Re, A.D., Díaz, F., Sierra, E., Gómez-Jiménez, Silvia. (2004). Consumo de oxígeno, excreción de amonio y capacidad osmorreguladora de Litopenaeus stylirostris (stimpson) expuesto a diferentes combinaciones de temperatura y salinidad. Ciencias Marinas. 30(003): 443-453.
Reed, L.A., Siewicki, T.C., Shah, J.C. (2004).Pharmacokinetics of oxytetraciclyne in the white shrimp, Litopenaeus setiferus. Aquaculture, 232:11-28.
Rendón, L., Balcázar, J.L. (2003). Inmunología de camarones: Conceptos básicos y recientes avances. Revista AquaTIC, 19:27-33.
Reyes, J., Téllez, M., González, J. (2009). Situación actual y perspectivas del camarón en México. Boletín informativo de Fideicomiso Instituido en Relación con la Agricultura (FIRA). No. 3.pp 122.
Rojas, A.A., Haws, M.C. y Cabanillas, J.A. ed. (2005). Buenas Prácticas de Manejo Para el Cultivo de Camarón. The David and Lucile Packard Foundation. United States Agency for International Development (Cooperative Agreement No. PCE-A-00-95-0030-05).
Rojas-Herrera, R., González-Flores, T. (2006). Detección e identificación de bacterias causantes de enfermedades transmitidas por alimentos mediante la reacción en cadena de la polimerasa. Bioquimia. 31(2):69-76.
Rosas, C., Sánchez, A., Díaz, E., Soto, L.A., Gaxiola, G. y Brito, R. (1996). Effect of dietary protein on apparent heat increment and postprandial nitrogen excretion of P. setiferus, P. schmitti, P. duorarum and P. notialis postlarvae. Journal of the World Aquaculture Society, 27: 92-102.
Roy, L.A., Davis, D.A., Saoud, I.P., Henry, R.P. (2007). Effects of varying levels of aqueous potassium and magnesium on survival, growth, and respiration of the Pacific white shrimp, Litopenaeus vannamei, reared in low salinity waters. Aquaculture, 262: 461-469.
Sánchez, D., Ching, C. (2003). Nitrito en estanques de cultivo intensivo de camarón. Boletín Nicovita, 8:1-4.
Santiago, H.M.L. (2009). Acumulación de oxitetraciclina (OTC) en camarón de cultivo Litopenaeus vannamei y pruebas de sensibilidad en bacterias tipo Vibrio aisladas de un sistema de cultivo. Tesis de maestría. Centro de Investigación en Alimentación y Desarrollo, A.C.
Santiago, M.L., Espinosa, A., Bermúdez, M.C. (2009). Uso de antibióticos en la camaronicultura. Revista mexicana de ciencias farmacéuticas. 40(3): 22-32.
Saoud, P.I., Davis, A.D., Roise, B. D. (2003). Suitability studies of inland we water for Litopenaeus vannamei culture. Aquaculture, 217:373-383.
Sen, D., Fang, W., Hao, S., Guo, B., Dong S. (2009). Effects of Salinity Fluctuation Frequency on the Osmolarity, Na+-K+-ATPase Activity and

75
HSP70 Expression in Juvenile Chinese Shrimp, Fenneropenaeus chinensis. Oceanic and Coastal Sea Research. 8 (1): 71-76.
Soto,W., Gutierrez, J., Remmenga, M.D., Nishiguchi, M.K. (2009). Salinity and temperatura effects on physiological responses of Vibrio fischeri from diverse ecological niches. Microbial Ecology, 57:140-150.
Soto-Rodríguez, S., Gomez-Gil, B., Lozano, R., Roque, A. (2010). Density of vibrios in hemolymph and hepatopáncreas of diseased pacific white shrimp, Litopenaeus vannamei, from Northwestern México. Journal of the World Aquaculture Society,41: 76-83.
Soto-Rodríguez, S., Armenta, M., Gómez, B. (2006). Effects of enrofloxacin and florfenicol on survival and bacterial population in an experimental infection with luminescent Vibrio campbelli in shrimp larvae of Litopenaeus vannamei. Aquaculture, 255: 48-54.
Talavera,V., Zapata, L., Sánchez, D. (1997). Boletín Nicovita. Volumen 2, Edición 01. En: Boyd, C.E. 1990. Water quality in ponds for Aquaculture. Alabama Agricultural Experiment Station, Auburn University. Birmingham Publishing Co. p 482.
Tang, C.M., Moxon, E.R. (2001).The impact of microbial genomics on antimicrobial drug development. Annual Review of Genomics and Human Genetic. 2 (2): 59-69.
Tarr, C.L., Patel, J.S., Puhr, N.D., Sowers, E.G., Bopp, C.A., Strockbine, N.A. (2007). Identification of Vibrio isolates by a multiplex PCR assay and rpoB sequence determination. Journal of Clinical Microbiology. 45(1): 134-140.
Tenover, F.C. (2006). Mechanisms of antimicrobial resistance in bacteria. Association for Professionals in Infection Control and Epidemiology, Inc. and Elsevier, Inc. Atlanta, GA. pp. 1-8.
Timmons, M.B., Ebeling, J.M., Wheaton, F.W., Summerfelt, S.T. y Vinci, B. (2002). Sistemas de recirculación para la acuicultura. Fundación Chile. Vitacura, Santiago. p. 748.
Tomasso, J.R. (2012). Environmental nitrite and aquaculture: a perspective. Aquaculture International. Springer. Texas.
Uno, K. (2004). Pharmacokinetics of oxolonic acid and oxitetraciclyne in kuruma shrimp, penaeus japonicus. Aquaculture, 204: 1-11.
Valenzuela, W., Rodríguez, G., Esparza, H. (2010). Cultivo intensivo de camarón blanco Litonepenaeus vannamei en agua de pozo de baja salinidad como alternativa acuícola para zonas de alta marginación. Ra Ximhai. Red de Revistas Científicas de América Latina y el Caribe, España y Portugal. 6(1): 1-8.
Valenzuela-Salcedo, G.F., Huerta-Aldaz, N., López-Torres, M.A., Armenta-Ayón, L.A., Moreno-Bayón, J.P., Leyva-Miranda, G.A., García-Morales, C. (2008). Bacterias en camarón blanco Litopenaeus vannamei durante su cultivo. Boletín de PRONALSA. pp. 5-9.
Venkateswara-Rao, A. (2009). Vibriosis en la acuacultura del camarón. India. Ver: www.neospark.com
Viola, C., DeVincent, S.J. (2006). Overview of issues pertaining to the manufacture, distribution, and use of antimicrobials in animals and other

76
information relevant to animal antimicrobial use data collection in the United States. Preventive Veterinary Medicine, 73: 111-131.
Wickins, J.F. (1976). The tolerance of warm-water prawns to recirculated water. Aquaculture, 9:19-37.
Woodford, N., Ellington, M. J. (2007). The emergence of antibiotic resistance by mutation. Clinical Microbiology and Infection, 13: 5-18.
Wu, J., Chen, H., Huang, D. (2009). Histopathological Alterations in Gills of White Shrimp, Litopenaeus vannamei (Boone) After Acute Exposure to Cadmium and Zinc. Bulletin of Environmental Contamination and Toxicology, 82:90–95.
Zhang, X., Austin, B. (2005) A review: Hemolysins in Vibrio species. Journal of Applied Microbiology, 98: 1011–1019.
Zhang, X., Meaden, P., Austin, B. (2001). Duplication of hemolysin genes in a virulent isolate of Vibrio harveyi. Applied and Environmental Microbiology. 67(7): 3161-3167.
Zhang Y., J. Zhou, J. Guo, X. Zhang, L. Zhao and Sh. Yuan. (2008). Study on nitrite accumulation characteristics and nitrifying population dynamics at different growth environments. Proceedings of World Academy of Science, Engineering and Technology.
Zhu ,C., Dong, S., Wang, F., Huang, G. (2004). Effects of Na/K ratio in seawater on growth and energy budget of juvenile Litopenaeus vannamei. Aquaculture, 234: 485-496.
Zhu, C.B., Dong, S.L., Wang, F., Zhang, H.H. (2006). Effects of seawater potassium concentration on the dietary potassium requirement of Litopenaeus vannamei. Aquaculture, 258: 543–550.