identification of bambi, an inhibitor of tgf- β signaling ...bambi and that bambi expression is...
TRANSCRIPT

1
Identification of BAMBI, an inhibitor of TGF-β signaling, as a target of the
β-catenin pathway in colorectal tumor cells
Takashi Sekiya*, Shungo Adachi*, Kazuyoshi Kohu*††††, Tatsuya Yamada‡‡‡‡, Osamu
Higuchi*¶¶¶¶, Yoichi Furukawa§§§§, Yusuke Nakamura§§§§, Tsutomu Nakamura*,
Kousuke Tashiro$$$$, Satoru Kuhara$$$$, Susumu Ohwada‡‡‡‡ and Tetsu Akiyama*####
*Laboratory of Molecular and Genetic Information, Institute of Molecular and
Cellular Biosciences, The University of Tokyo, 1-1-1 Yayoi, Bunkyo-ku, Tokyo
113-0032, Japan.§§§§Laboratory of Molecular Medicine, Human Genome Center, Institute of Medical
Science, The University of Tokyo, 4-6-1 Shirokanedai, Minato-ku, Tokyo 108-8639,
Japan.‡‡‡‡Second Department of Surgery, Gunma University School of Medicine, 3-39-22
Showa-machi, Maebashi, Gunma 371, Japan.$$$$Laboratory of Molecular Gene Technics, Department of Genetic Resources
Technology, Faculty of Agriculture, Kyushu University, Hakozaki 6-10-1, Higashi-
ku, Fukuoka, 812-8581, Japan.
Present Address: ††††Department of Molecular Immunology, Institute of
Development, Aging, and Cancer, Tohoku University, Sendai, Japan.¶¶¶¶Department of Cell Regulation, Medical Research Institute, Tokyo Medical and
Dental University, Bunkyo-ku, Tokyo 113-8510, Japan.
Running Title: Induction of BAMBI by β-catenin signaling
####To whom correspondence should be addressed.
E-mail:[email protected]
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

2
Summary
The Wnt signaling pathway is activated in most human colorectal tumors.
Mutational inactivation in the tumor suppressor adenomatous polyposis coli (APC),
as well as activation of β-catenin causes the accumulation of β-catenin, which in
turn associates with the TCF/LEF family of transcription factors and activates
transcription of their target genes. Here we show that β-catenin activates
transcription of the BMP and activin membrane-bound inhibitor (BAMBI)/NMA
gene. The expression level of BAMBI was found to be aberrantly elevated in most
colorectal and hepatocellular carcinomas relative to the corresponding non-
cancerous tissues. Expression of BAMBI in colorectal tumor cell lines was
repressed by a dominant-negative mutant of TCF-4 or by ICAT, an inhibitor of β-
catenin-TCF interaction, suggesting that β-catenin is responsible for the aberrant
expression of BAMBI in colorectal tumor cells. Furthermore, overexpression of
BAMBI inhibited the response of tumor cells to transforming growth factor-
β (TGF-β) signaling. These results suggest that β-catenin interferes with TGF-β-
mediated growth arrest by inducing the expression of BAMBI, and this may
contribute to colorectal and hepatocellular tumorigenesis.
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

3
Introduction
The conversion of an intestinal epithelial cell into a fully transformed, metastatic cancer
cell requires mutations in multiple proto-oncogenes and tumor suppressor genes (1,2).
Mutations in the tumor suppressor APC and β-catenin occur during the early stages of
this process. The product of the APC gene interacts with various proteins including β-
catenin, Axin, Rac-specific guanine nucleotide exchanger Asef, kinesin superfamily-
associated protein 3 (KAP3), EB1, microtubules and the human homolog of the
Drosophila Discs large (hDLG) (3-11). Through interacting with these molecules,
APC functions in multiple signaling pathways, thereby regulating Wnt signaling, actin
and microtubule cytoskeletal networks, cell morphology and migration (8,12-18).
β-catenin is an essential component of the Wnt signaling pathway and plays
important roles in development and tumorigenesis (19-22). Wnt signaling promotes
the stabilization and accumulation of β-catenin, which in turn interacts with the
TCF/LEF family of transcription factors and activates transcription of downstream
genes such as c-Myc, cyclin D1 and Axin2 (23-27). APC interacts with β-catenin,
glycogen synthase kinase-3β (GSK-3β), casein kinase 1α and Axin or the closely
related factor conductin/Axil (3-7,28-30). By recruiting β-catenin into this multi-
protein complex, APC promotes its proteasome-mediated degradation, hence mutations
in APC or β-catenin result in the accumulation of β-catenin in colorectal tumor cells
(31,32). Mutations in β-catenin and Axin have also been identified in many other
types of tumors, including hepatocellular carcinoma, ovarian cancer and endometrial
cancer (33-35). Thus, constitutive activation of β-catenin-TCF-mediated transcription
is believed to be a critical step in the tumorigenesis of various types of tumors.
The transforming growth factor-β (TGF-β) pathway inhibits the growth of
multiple epithelial cell types, and loss of this negative regulation is believed to
contribute to tumor development, including colorectal tumorigenesis. Indeed,
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

4
inactivating mutations in the type II receptor (TβRII), Smad2 and Smad4 have been
reported to occur during colorectal tumorigenesis. In addition, it has been reported
that the TGF-β receptor is often downregulated in tumor cells, or otherwise unavailable
at the cell surface, thus allowing tumor cells to escape the growth inhibitory activities of
TGF- β (36,37).
TGF-β signals through a heteromeric cell-surface complex of two types of
transmembrane serine/threonine kinases, the type I and type II receptors (TβRI and
TβRII). TβRII activates TβRI, which in turn propagates the TGF-β signal by
phosphorylating the Smads, Smad2 and Smad3 (38,39). Receptor-mediated
phosphorylation of Smads induces their association with Smad4, followed by
translocation into the nucleus, where these complexes activate transcription of target
genes, including CDK inhibitors p15INK4B and p21CIP1 (40,41).
ICAT is a β-catenin-interacting protein that interferes with the interaction between
β-catenin and TCF, and inhibits the proliferation of colorectal tumor cells containing
mutations in APC or β-catenin and hepatocellular carcinoma cells containing mutations
in Axin (42-45). In the present study, we examined the effects of ICAT on gene
expression in the human colorectal tumor cell line SW48, in which β-catenin-TCF4-
mediated transcription is aberrantly activated due to a mutation in β-catenin. We
found that ICAT suppresses expression of the BAMBI (BMP and activin membrane-
bound inhibitor). Furthermore, we found that β-catenin activates transcription of
BAMBI and that BAMBI expression is aberrantly elevated in most colorectal and
hepatocellular carcinomas, compared to the corresponding non-cancerous tissues. The
product of the BAMBI gene is a transmembrane protein that lacks an intracellular kinase
domain but has sequence similarity to the extracellular domain of TβRI and thereby
inhibits TGF-β signaling by forming heterodimer with TβRII (46). Our data suggest
that β-catenin-mediated overexpression of BAMBI may be one of the strategies by
which tumor cells escape the growth inhibitory activities of TGF-β.
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

5
Experimental Procedures
Construction of plasmids and recombinant viruses. For construction of the
BAMBI-luciferase reporter plasmids, a human BAC clone RP13-43N24, which contains
the entire promoter region and several exons and introns, was purchased from BACPAC
Resources. The longest construct, 7.1 kb, and deletion constructs were generated by
cloning each sequence into the appropriate enzyme sites in the pGL3-basic vector
(Promega). Site-directed mutagenesis of each TCF/LEF binding elements was created
by standard PCR techniques using Pyrobest DNA polymerase (TAKARA), with the
following primers (substituted nucleotides are written in bold-face); for TBE1, 5'-
GCTGCAGAGGATTGATTAGCGGTAG-3' and 5'- CTACCGCTAATCAATCCTCTG
CAGC-3', for TBE2, 5'- CTCTGTGTCTAGTTAAATGTATCTCTG-3' and 5'- CAGAG
ATACATTTAACTAGACACAGAG-3', for TBE3, 5'- CTCTAAGTGTAGTTATATCT
CTGAATG-3' and 5'- CATTCAGAGATATAACTACACTTAGAG-3', for TBE4, 5'-
CTGGAAATATAGAAAGCGGGCAGAAC-3' and 5'- GTTCTGCCCGCTTTCTATAT
TTCCAG-3', for TBE5, 5'- CTAAAAGTTCATGCAGTTAAATTTGGG-3' and 5'-
CCCAAATTTAACTGCATGAACTTTTAG-3', for TBE1', 5'-CCGTGCTGCTCGCCA
AATGTGGGTGCC-3' and 5'-GGCACCCACATTTGGCGAGCAGCACGG-3', for
TBE2', 5'-CGCCGCGGGGATTTGGCGCCGGCTGC-3' and 5'-GCAGCCGGCGCCA
AATCCCCGCGGCG-3', for TBE3', 5'-GTTGCTGTGTTATTAGCAGTCGCGGCG-3'
and 5'-CGCCGCGACTGCTAATAACTCTGCAAC-3', for TBE4', 5'- CGGACTCCGA
GCTAATGGCAGGCGCTG-3' and 5'-CAGCGCCTGCCATTAGCTCGGAGTCCG-3'.
pTOP-tk-luciferase and pFOP-tk-luciferase were kindly provided by V. Korinek and H.
Clevers (University Medical Center, Utrecht, The Netherlands). The adenoviruses
encoding myc-ICAT and LacZ were generated as previously described (43). Ad-Flag-β-
catenin S33Y was constructed using the pAdenoXTM Expression system (Clontech)
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

6
according to the manufacturer's directions. BAMBI expression plasmids: the full-
length BAMBI-GFP fusion expression vector pEGFP-N3-hBAMBI was constructed by
cloning RT-PCR products into the pEGFP-N3 vector (Clontech). A variant of this
vector, containing BAMBI lacking amino acids 20-131 (BAMBI∆N) was constructed
similarly.
Cells and clinical materials. All cells were cultured as monolayers in appropriate
media [DMEM (NISSUI) for COS-1, DU145, HepG2 and Alexander cells; Leibovit's L-
15 (SIGMA) for SW48 and SW480 cells, and McCoy's 5A (SIGMA) for HCT116 cells]
supplemented with 10% fetal bovine serum (JCS). All cells were maintained at 37°C
in an atmosphere of humidified air with 5% CO2. Cancerous tissues and corresponding
noncancerous tissue samples were excised during surgery at the Gunma University
Hospital, after informed consent had been obtained.
Northern blotting. Total RNA was extracted using NucleoSpin (MACHEREY-
NAGEL). Total RNA (20 µg per lane) was electrophoresed on a formaldehyde-
containing 1% agarose gel, transferred onto a Hybond-N+ (Amersham Pharmacia
Biotech), and hybridized with 32P-labeled cDNA probe specific for indicated genes, and
examined via BAS1500 (FUJI FILM).
Antibodies. Antibodies to the amino- and carboxy-terminal regions of BAMBI were
prepared by immunizing rabbits with bacterially-expressed, GST fusion proteins
containing human BAMBI amino acids 45-147 and 177-241, respectively. Rabbit
polyclonal antibody to ICAT was prepared as described (42). Mouse monoclonal
antibody to α-tubulin was purchased from OncogeneTM. Mouse monoclonal antibody
to FLAG (M2) and β-galactosidase (Ab-1) were purchased from SIGMA and
CALBIOCHEM, respectively. Immunoblotting was performed as described
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

7
previously (11).
Luciferase reporter assays. Cells were plated in 12-well dishes 18 h prior to
transfection. Transfections were performed with Lipofectamine PLUS (Invitrogen)
according to the manufacturer's protocol. Luciferase assays were performed using the
Dual-Luciferase Reporter Assay System (Promega) following the manufacturer's
protocol.
Semi-quantitative RT-PCR assay. Total RNA was extracted from cancerous and
corresponding noncancerous tissues, using NucleoSpin (MACHEREY-NAGEL). First
strand cDNA was synthesized using random hexamers and Superscript II reverse
transcriptase (Invitrogen). One µl of cDNA from each sample was used for PCR with
the PTC-2000 Peltier Thermal Cycler (MJ Research). All of the reactions were
performed in 25 µl volumes and amplified for 3 min at 94°C for initial denaturation,
followed by appropriate cycles at 94°C for 30 s, 62°C for 30 s, and 72°C for 1 min. The
PCR products were electrophoresed on a 1% agarose gel, and detected by EtBr staining.
Immunohistochemical analysis. Immunostaining of paraffin-embedded tissue
sections was performed according to standard procedures. In brief, sections were
deparaffinized in xylene, and then rehydrated into distilled water using graded ethanols.
Antigens were retrieved by microwaving the slides in citrate buffer (pH 6.0) for 15 min.
Sections were incubated with the indicated antibodies overnight at 4°C. Staining
patterns obtained with the antibodies to β-catenin and BAMBI were visualized with
RITC-labeled anti-mouse antibody, and FITC-labeled anti-rabbit antibodies,
respectively. The sections were visualized and photographed with a confocal
microscope. In Fig. 2C (insets, right), COS-1 cells were transfected with FLAG-
tagged TCF4-∆C and stained with ant-FLAG antibody followed by FITC-labelled anti-
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

8
rabbit secondary antibody. The nucleus was stained with TOTO-3 (Molecular Probes).
Colony formation assay. One day before seeding, DU145 cells were transfected with
the indicated plasmids, using Lipofectamine2000 (Invitrogen) according to the
manufacturer's protocol. Cells (5 x 105 cells) were seeded in each 10 cm dishes and
cultured in the presence or absence of 1 ng/ml TGF-β1 (R&D), under the selection with
400 µg/ml G418 (GIBCO). Medium change and TGF-β1 stimulation were performed
every 5 days. Three weeks after seeding, colonies were either stained with methylene
blue, or isolated for further experiments.
Results and Discussion
Using a cDNA microarray approach, we examined the effects of ICAT on gene
expression in SW48 cells. Infection of SW48 cells with a recombinant adenovirus
encoding ICAT (Ad-ICAT) induced changes in the expression levels of a number of
genes, including c-MYC, AXIN2 and BCL-XL. We found that one of the genes
repressed by Ad-ICAT is BAMBI.
To confirm that BAMBI expression is indeed repressed by ICAT, we examined
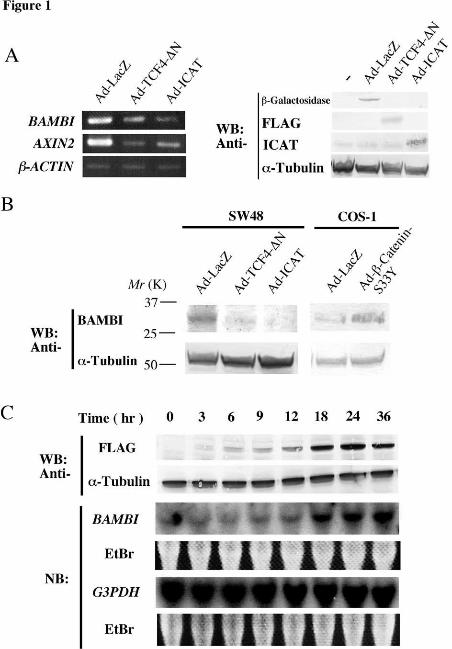
BAMBI expression in SW48 cells infected with Ad-ICAT by RT-PCR analysis (Fig. 1A,
left). We found that BAMBI was expressed in SW48 cells and repressed significantly
after Ad-ICAT infection similar to the known β-catenin target gene, AXIN2. In these
experiments, expression of adenovirus-transduced genes, ICAT, TCF-4-∆N (see below)
and β-galactosidase, was confirmed by immunoblotting analysis (Fig. 1A, right). We
also examined expression of BAMBI protein using antibodies directed against the
amino-terminal and carboxy-terminal regions of human BAMBI: viz. anti-BAMBI-N-
ter and anti-BAMBI-C-ter, respectively. Immunoblotting analysis of the lysates from
SW48 cells infected with Ad-ICAT revealed that BAMBI protein expression was also
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

9
repressed by ICAT (Fig. 1B).
These results suggest that the expression of BAMBI is regulated by β-catenin.
We therefore examined the effect of an active mutant of β-catenin, β-catenin-S33Y, on
the expression of BAMBI. β-catenin-S33Y is a mutant that was initially identified in
human colorectal tumors, and contains a tyrosine (Y) in pace of the normal serine at
residue 33 (S33), a change that renders the protein resistant to APC-mediated
degradation (34). Northern blot analysis of RNA isolated from COS-1 cells showed
that BAMBI expression is increased following infection with an adenovirus encoding β-
catenin-S33Y (Fig. 1C). Immunoblot analysis demonstrated a similar increase in
BAMBI protein expression by expression of β-catenin-S33Y (Fig. 1B). Induction of
BAMBI was evident within 6 h after Ad-β-catenin-S33Y infection and an
approximately five-fold increase was observed at 24 h after infection. To further
confirm that expression of BAMBI in SW48 cells is induced by β-catenin-TCF4-
mediated transactivation, we examined the effect of a dominant-negative mutant of
TCF-4, TCF-4-∆N, which lacks the amino-terminal β-catenin-binding domain but
contains the carboxy-terminal DNA-binding domain. Expression of BAMBI at both
the mRNA and protein levels in SW48 cells was found to be repressed by expression of
TCF-4-∆N (Fig, 1A, B). These results suggest that expression of BAMBI is regulated
by β-catenin-TCF4-mediated transactivation and that the expression of BAMBI in
SW48 cells is induced by mutated β-catenin.
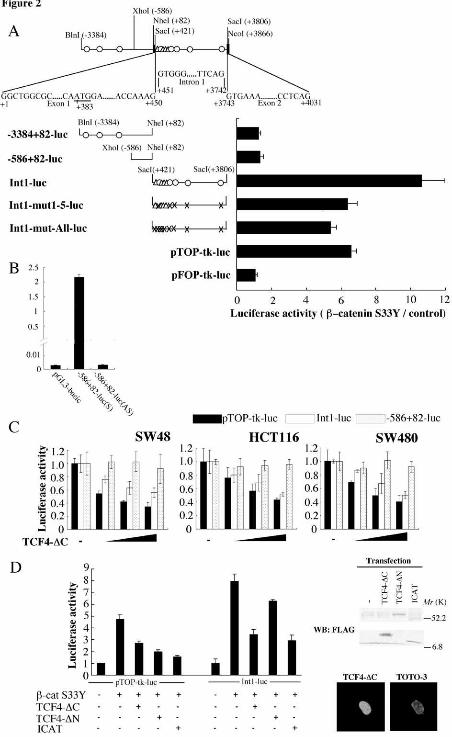
We next investigated whether transcription of BAMBI is directly regulated by β-
catenin. We isolated a BlnI (-3384)-NcoI (+3866) genomic fragment encompassing
the upstream and downstream regions of the transcription start site (Fig. 2A) and
examined whether the BAMBI promoter is contained within this fragment. When a
XhoI(-586)-NheI (+82) fragment was inserted upstream of a luciferase reporter gene in
the sense and anti-sense orientations, and transfected into COS-1 cells, the sense
construct exhibited about 170-fold higher luciferase activity than the anti-sense
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

10
construct or empty vector (Fig. 2B). Thus, the BAMBI promoter is presumably
contained within this fragment. However, the activity of the reporter containing this
BAMBI promoter region (-586+82-luc in Fig. 2A) was not enhanced by cotransfection
with β-catenin-S33Y (Fig. 2A). Similarly, β-catenin-S33Y did not stimulate the
activity of a reporter construct that contains the further upstream regions containing at
least 3 consensus sites for TCF-binding (-3384+82-luc in Fig. 2A). We therefore
examined the β-catenin responsiveness of the region downstream of the BAMBI
promoter [SacI(+421)-SacI(+3806) fragment]. We inserted this fragment upstream of
the SV40 minimal promoter in a luciferase reporter vector (Int1-luc in Fig. 2A) and
examined the construct for responsiveness to β-catenin. When transfected into COS-1
cells, activity of this reporter was found to be enhanced significantly by β-catenin-S33Y
(Fig. 2A). These results suggest that this region, which encompasses intron 1, is
responsible for β-catenin-mediated transactivation. Intron 1 contains at least 5
consensus sites for TCF-binding, and synthetic oligonucleotides containing each of
these sites demonstrated specific binding to the DNA-binding region of TCF-4, fused to
glutathione-S-transferase (GST), in an electrophoretic mobility-shift assay (data not
shown).
We next examined whether transcription of BAMBI is activated by the
endogenous β-catenin-TCF complexes using the colorectal tumor cell lines SW480,
HCT116 and SW48. SW480 contains mutated APC and wild-type β-catenin, whereas
HCT116 and SW48 possesses wild-type APC and mutated β-catenin. The activity of
Int1-luc but not –586+82-luc was repressed in a dose-dependent manner in SW480,
HCT116 and SW48 cells by a dominant-negative mutant of TCF-4, TCF-4-∆C, which
contains the amino-terminal β-catenin-binding domain but lacks the carboxy-terminal
DNA-binding domain (Fig. 2C). This mutant was constructed as a fusion to the
nuclear localization signal of SV40, and was found to localize efficiently to the nucleus
(Fig. 2D). Under these experimental conditions, TCF-∆C also repressed the activity of
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

11
pTOP-tk-luciferase, which contains optimal TCF-binding sites upstream of a luciferase
reporter gene, in these cell lines.
These results imply that BAMBI is a direct target for β-catenin-TCF-mediated
transactivation. However, we noticed that even when all five consensus TCF-binding
sites in Int1-luc were mutated, some β-catenin responsiveness remained (Int1-mut1-5-
luc in Fig. 2A). In addition to these five sites, we also mutated four sites in Int1-luc,
which diverge one nucleotide from the TCF-binding consensus sequence. This
construct (Int1-mut-All-luc in Fig. 2A) exhibited slightly lower activity than Int1-mut-5-
luc, suggesting that these sites contribute only a little to β-catenin-mediated
transactivation. Furthermore, the β-catenin-mediated transactivation of Int1-luc was
only weakly inhibited by TCF-4-∆N, whereas it was significantly inhibited by ICAT as
well as TCF-4-∆C (Fig. 2D). The weak effect of TCF-4-∆N is not due to low levels of
expression, since immunoblotting analysis revealed that TCF-4-∆N and TCF-4-∆C were
expressed at similar levels in these experiments. On the other hand, TCF-4-∆N as well
as TCF-4-∆C and ICAT exhibited significant inhibitory effects on the β-catenin-
mediated transactivation of pTOP-tk-luciferase. TCF-4-∆N differs from TCF-4-∆C in
that the former blocks TCF-binding to DNA, whereas TCF-4-∆C is able to inhibit the
interaction of a number of proteins, including β-catenin. These results suggest that
BAMBI may be transcriptionally regulated not only by β-catenin-TCF complexes, but
by β-catenin complexed to other proteins. This finding is consistent with recent
reports showing that complexes of β-catenin with partners other than TCF can also
transactivate target genes (47-50).
Northern blotting analysis revealed that BAMBI is expressed at very low levels in
human colon, small intestine, thymus and peripheral bloods, at moderate levels in brain,
skeletal muscle, liver and lung, and at high levels in heart, spleen, kidney and placenta
(data not shown). Since β-catenin accumulates in most human colorectal tumors due
to mutations in APC or β-catenin, we expected BAMBI expression to be up-regulated in
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

12
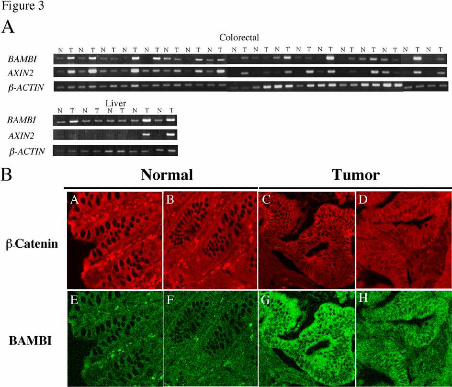
colorectal tumors. We examined BAMBI expression in 18-paired colorectal tumors
and adjacent non-cancerous tissues by semi-quantitative RT-PCR analysis. In 13 of 18
(72%) cases, the expression level of BAMBI was very high in colorectal tumors but was
very low in the corresponding non-cancerous tissues (Fig. 3A). In the other 5 cases,
BAMBI expression was almost equal to or lower than that of the non-cancerous tissues.
Immunostaining experiments also demonstrated that BAMBI is expressed at high levels
in colorectal tumors but at very low levels in non-cancerous tissues (Fig. 3B).
Furthermore, we found that BAMBI expression is higher in hepatocellular carcinomas
than in adjacent normal tissues in 3 of 5 cases, consistent with the fact that AXIN1 as
well as β-catenin is mutated in a certain subset of hepatocellular carcinomas (35,51).
Consistent with the fact that expression of AXIN2 is activated by β-catenin and up-
regulated in colorectal tumors (25,27), we observed that the expression patterns of
BAMBI and AXIN2 were similar in most tumors examined.
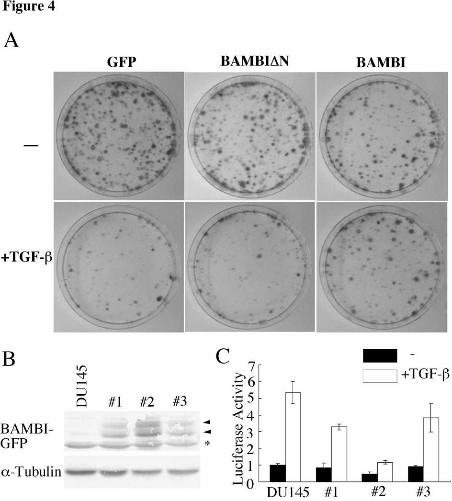
Since BAMBI has been shown to act as an antagonist of TGF-β signaling (46,52),
we examined the effect of overexpression of BAMBI on the growth of tumor cells in the
presence of TGF-β. In these experiments, we used the prostate tumor cell line DU145,
because it responds to TGF-β and does not exhibit aberrant β-catenin-mediated
transactivation (data not shown), suggesting that it does not possess mutations in APC,
β-catenin or AXIN. We transfected BAMBI into DU145 cells and performed a colony
formation assay in the presence or absence of TGF-β. TGF-β strongly inhibited
colony formation among DU145 cells that were transfected with GFP or mutant
BAMBI-GFP lacking the amino-terminal extracellular domain (BAMBI∆N-GFP). In
contrast, BAMBI-GFP-transfected cells were resistant to the growth inhibitory effects of
TGF-β (Fig. 4A). Immunoblotting analysis of BAMBI-GFP-transfected cells
propagated from TGF-β-resistant colonies revealed that BAMBI-GFP is indeed
expressed in these cells (Fig. 4B). We also examined the effect of BAMBI-GFP on
TGF-β-mediated transactivation in DU145 cells using the reporter p3TP-lux, which
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

13
contains a TGF-β-responsive element from the PAI-1 promoter placed upstream of a
luciferase reporter gene. Luciferase assay revealed that TGF-β-mediated
transactivation is repressed by exogenously expressed BAMBI-GFP (Fig. 4C). These
results imply that BAMBI has the potential to inhibit TGF-β-mediated transactivation
and cell growth inhibition.
Loss of TGF-β responsiveness provides an advantage for developing tumors
(36,37). Most colorectal tumors possess mutations in a component of the TGF-β
signaling pathway such as TβRII, SMAD2 or SMAD4. Our results show that, in
addition, an inhibitor of TGF-β signaling, BAMBI, is induced by β-catenin and
overexpressed in colorectal and hepatocellular carcinoma cells. In the multistep
progression into cancer, mutations in the Wnt signaling pathway generally precede
those in other tumor suppressor genes and oncogenes (1,20). It is therefore interesting
to speculate that overexpression of BAMBI may allow adenomas and tumor cells to
escape the growth inhibitory activities of TGF-β until a component of TGF-β signaling
is mutated. BAMBI also interferes with signaling pathways other than that mediated
by the SMADs, thus its overexpression may be more advantageous to oncogeneic
transformation than mutation of the SMADs alone. It is also possible that β-catenin-
induced BAMBI expression plays a role in embryogenesis. These possibilities could
be examined using BAMBI knockout mice, as well as APC and BAMBI double knockout
mice. Furthermore, our findings suggest that BAMBI may have the potential as a
target for therapy using monoclonal antibodies (53). Development of monoclonal
antibodies to BAMBI as anti-tumor reagents is currently under-way in our laboratory.
Acknowledgment
We thank I. Saito and Y. Kanegae for helping us to prepare the recombinant
adenoviruses; K. Miyazono for p3TP-lux; M. Lamphier and T. Okabe for reading the
manuscript. Supported by Grants-in-Aid for Scientific Research on Priority Areas and
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

14
the Organization for Pharmaceutical Safety and Research.
References
1. Kinzler, K. W., and Vogelstein, B. (1996) Cell 88887777, 159-170
2. Nakamura, Y. (1995) J Cancer Res Clin Oncol 111122221111, 529-534
3. Rubinfeld, B., Souza, B., Albert, I., Muller, O., Chamberlain, S. H., Masiarz, F. R.,
Munemitsu, S., and Polakis, P. (1993) Science 222266662222, 1731-1734
4. Su, L. K., Vogelstein, B., and Kinzler, K. W. (1993) Science 222266662222, 1734-1737
5. Kishida, S., Yamamoto, H., Ikeda, S., Kishida, M., Sakamoto, I., Koyama, S., and
Kikuchi, A. (1998) J Biol Chem 222277773333, 10823-10826
6. Nakamura, T., Hamada, F., Ishidate, T., Anai, K., Kawahara, K., Toyoshima, K., and
Akiyama, T. (1998) Genes Cells 3333, 395-403
7. Hamada, F., Tomoyasu, Y., Takatsu, Y., Nakamura, M., Nagai, S., Suzuki, A., Fujita,
F., Shibuya, H., Toyoshima, K., Ueno, N., and Akiyama, T. (1999) Science 222288883333,
1739-1742
8. Kawasaki, Y., Senda, T., Ishidate, T., Koyama, R., Morishita, T., Iwayama, Y.,
Higuchi, O., and Akiyama, T. (2000) Science 222288889999, 1194-1197
9. Jimbo, T., Kawasaki, Y., Koyama, R., Sato, R., Takada, S., Haraguchi, K., and
Akiyama, T. (2002) Nat Cell Biol 4444, 323-327
10. Su, L. K., Burrell, M., Hill, D. E., Gyuris, J., Brent, R., Wiltshire, R., Trent, J.,
Vogelstein, B., and Kinzler, K. W. (1995) Cancer Res 55555555, 2972-2977
11. Matsumine, A., Ogai, A., Senda, T., Okumura, N., Satoh, K., Baeg, G. H., Kawahara,
T., Kobayashi, S., Okada, M., Toyoshima, K., and Akiyama, T. (1996) Science 222277772222,
1020-1023
12. Fearnhead, N. S., Britton, M. P., and Bodmer, W. F. (2001) Hum Mol Genet 11110000,
721-733
13. Rosin-Arbesfeld, R., Ihrke, G., and Bienz, M. (2001) Embo J 22220000, 5929-5939
14. Nathke, I. S., Adams, C. L., Polakis, P., Sellin, J. H., and Nelson, W. J. (1996) J Cell
Biol 111133334444, 165-179
15. Mimori-Kiyosue, Y., Shiina, N., and Tsukita, S. (2000) J Cell Biol 111144448888, 505-518
16. Munemitsu, S., Souza, B., Muller, O., Albert, I., Rubinfeld, B., and Polakis, P. (1994)
Cancer Res 55554444, 3676-3681
17. Smith, K. J., Levy, D. B., Maupin, P., Pollard, T. D., Vogelstein, B., and Kinzler, K. W.
(1994) Cancer Res 55554444, 3672-3675
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

15
18. Kawasaki, Y., Sato, R., and Akiyama, T. (2003) Nat Cell Biol 5555, 211-215
19. Cadigan, K. M., and Nusse, R. (1997) Genes Dev 11111111, 3286-3305
20. Polakis, P. (2000) Genes Dev 11114444, 1837-1851
21. Bienz, M., and Clevers, H. (2000) Cell 111100003333, 311-320
22. Peifer, M., and Polakis, P. (2000) Science 222288887777, 1606-1609
23. He, T. C., Sparks, A. B., Rago, C., Hermeking, H., Zawel, L., da Costa, L. T., Morin, P.
J., Vogelstein, B., and Kinzler, K. W. (1998) Science 222288881111, 1509-1512
24. Tetsu, O., and McCormick, F. (1999) Nature 333399998888, 422-426
25. Yan, D., Wiesmann, M., Rohan, M., Chan, V., Jefferson, A. B., Guo, L., Sakamoto, D.,
Caothien, R. H., Fuller, J. H., Reinhard, C., Garcia, P. D., Randazzo, F. M., Escobedo,
J., Fantl, W. J., and Williams, L. T. (2001) Proc Natl Acad Sci U S A 99998888, 14973-
14978
26. Leung, J. Y., Kolligs, F. T., Wu, R., Zhai, Y., Kuick, R., Hanash, S., Cho, K. R., and
Fearon, E. R. (2002) J Biol Chem 222277777777, 21657-21665
27. Lustig, B., Jerchow, B., Sachs, M., Weiler, S., Pietsch, T., Karsten, U., van de
Wetering, M., Clevers, H., Schlag, P. M., Birchmeier, W., and Behrens, J. (2002) Mol
Cell Biol 22222222, 1184-1193
28. Rubinfeld, B., Albert, I., Porfiri, E., Fiol, C., Munemitsu, S., and Polakis, P. (1996)
Science 222277772222, 1023-1026
29. Liu, C., Li, Y., Semenov, M., Han, C., Baeg, G. H., Tan, Y., Zhang, Z., Lin, X., and He,
X. (2002) Cell 111100008888, 837-847
30. Behrens, J., Jerchow, B. A., Wurtele, M., Grimm, J., Asbrand, C., Wirtz, R., Kuhl, M.,
Wedlich, D., and Birchmeier, W. (1998) Science 222288880000, 596-599
31. Aberle, H., Bauer, A., Stappert, J., Kispert, A., and Kemler, R. (1997) Embo J 11116666,
3797-3804
32. Sakanaka, C., Weiss, J. B., and Williams, L. T. (1998) Proc Natl Acad Sci U S A 99995555,
3020-3023
33. Munemitsu, S., Albert, I., Rubinfeld, B., and Polakis, P. (1996) Mol Cell Biol 11116666,
4088-4094
34. Morin, P. J., Sparks, A. B., Korinek, V., Barker, N., Clevers, H., Vogelstein, B., and
Kinzler, K. W. (1997) Science 222277775555, 1787-1790
35. Satoh, S., Daigo, Y., Furukawa, Y., Kato, T., Miwa, N., Nishiwaki, T., Kawasoe, T.,
Ishiguro, H., Fujita, M., Tokino, T., Sasaki, Y., Imaoka, S., Murata, M., Shimano, T.,
Yamaoka, Y., and Nakamura, Y. (2000) Nat Genet 22224444, 245-250
36. Derynck, R., Akhurst, R. J., and Balmain, A. (2001) Nat Genet 22229999, 117-129
37. Massague, J., Blain, S. W., and Lo, R. S. (2000) Cell 111100003333, 295-309
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

16
38. Massague, J., and Chen, Y. G. (2000) Genes Dev 11114444, 627-644
39. Miyazono, K., ten Dijke, P., and Heldin, C. H. (2000) Adv Immunol 77775555, 115-157
40. Hannon, G. J., and Beach, D. (1994) Nature 333377771111, 257-261
41. Datto, M. B., Li, Y., Panus, J. F., Howe, D. J., Xiong, Y., and Wang, X. F. (1995) Proc
Natl Acad Sci U S A 99992222, 5545-5549
42. Tago, K., Nakamura, T., Nishita, M., Hyodo, J., Nagai, S., Murata, Y., Adachi, S.,
Ohwada, S., Morishita, Y., Shibuya, H., and Akiyama, T. (2000) Genes Dev 11114444,
1741-1749
43. Sekiya, T., Nakamura, T., Kazuki, Y., Oshimura, M., Kohu, K., Tago, K., Ohwada, S.,
and Akiyama, T. (2002) Cancer Res 66662222, 3322-3326
44. Daniels, D. L., and Weis, W. I. (2002) Mol Cell 11110000, 573-584
45. Graham, T. A., Clements, W. K., Kimelman, D., and Xu, W. (2002) Mol Cell 11110000,
563-571
46. Onichtchouk, D., Chen, Y. G., Dosch, R., Gawantka, V., Delius, H., Massague, J., and
Niehrs, C. (1999) Nature 444400001111, 480-485
47. Tice, D. A., Szeto, W., Soloviev, I., Rubinfeld, B., Fong, S. E., Dugger, D. L., Winer,
J., Williams, P. M., Wieand, D., Smith, V., Schwall, R. H., Pennica, D., and Polakis, P.
(2002) J Biol Chem 222277777777, 14329-14335
48. Xu, L., Corcoran, R. B., Welsh, J. W., Pennica, D., and Levine, A. J. (2000) Genes
Dev 11114444, 585-595
49. Shtutman, M., Zhurinsky, J., Oren, M., Levina, E., and Ben-Ze'ev, A. (2002) Cancer
Res 66662222, 5947-5954
50. Kioussi, C., Briata, P., Baek, S. H., Rose, D. W., Hamblet, N. S., Herman, T., Ohgi, K.
A., Lin, C., Gleiberman, A., Wang, J., Brault, V., Ruiz-Lozano, P., Nguyen, H. D.,
Kemler, R., Glass, C. K., Wynshaw-Boris, A., and Rosenfeld, M. G. (2002) Cell 111111111111,
673-685
51. Taniguchi, K., Roberts, L. R., Aderca, I. N., Dong, X., Qian, C., Murphy, L. M.,
Nagorney, D. M., Burgart, L. J., Roche, P. C., Smith, D. I., Ross, J. A., and Liu, W.
(2002) Oncogene 22221111, 4863-4871
52. Tsang, M., Kim, R., de Caestecker, M. P., Kudoh, T., Roberts, A. B., and Dawid, I. B.
(2000) Genesis 22228888, 47-57
53. Carter, P. (2001) Nat Rev Cancer 1111, 118-129
54. Suzuki, Y., Yamashita, R., Nakai, K., and Sugano, S. (2002) Nucleic Acids Res 33330000,
328-331
Figure Legends
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

17
Fig. 1. Expression of BAMBI is regulated by β-catenin-TCF-mediated
transactivation. (A) Left, Semi-quantitative RT-PCR analysis of RNA from SW48
cells infected with Ad-LacZ, Ad-ICAT or Ad-TCF-4-∆N using primers specific for
BAMBI, AXIN2 or β-ACTIN. Right, Expression of adenovirus-transduced genes were
detected by immunoblotting analysis. TCF-4-∆N was tagged with FLAG-tag and
detected with anti-FLAG antibody. Cells were infected with the indicated
adenoviruses at a MOI 100. (B) Immunoblotting analysis of lysates from SW48 cells
infected with Ad-LacZ, Ad-ICAT or Ad-TCF-4-∆N, or from COS-1 cells infected with
Ad-LacZ or Ad-Flag-β-catenin-S33Y, with anti-BAMBI-N-ter antibody or anti-α-
tubulin antibody. Cells were infected with adenoviruses at a MOI 100. (C) Time
course of BAMBI upregulation following Ad-Flag-β-catenin-S33Y infection of COS-1
cells. Total RNA was isolated at the indicated times and analyzed by Northern blot.
Two different blots were hybridized with the BAMBI and G3PDH probes, with EtBr
staining serving as a loading control. Lysates prepared at the indicated times were
subjected to immunoblotting analysis with FLAG antibody or anti-α-tubulin antibody.
Cells were infected with adenoviruses at a MOI 20.
Fig. 2. β-catenin-TCF-mediated transactivation of the BAMBI promoter. (A)
Schematic representation of the BAMBI promoter region. O, consensus TCF-binding
sites (CTTTGA/TA/T); ∆, potential TCF-binding sites, which differ from the consensus
TCF-binding sites by one base; X, mutated TCF-binding sites (three nucleotides in O
and two nucleotides in ∆ were mutated). The nucleotide sequence of this region is
available from the NCBI database (accession no. AL390996). The transcription
initiation site was adopted from DBTSS [Database of Transcriptional Start Sites (54)].
The fragments indicated in the figure were inserted directly upstream of the luciferase
gene in pGL3 (-3384+82-luc and –586+82-luc) or upstream of the SV40 minimal
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

18
promoter in a luciferase reporter vector (Int1-luc, Int1-mut1-5-luc and Int1-mut-All-luc),
respectively. COS-1 cells were transfected with β-catenin-S33Y (0.8 µg) along with
the indicated promoter construct (0.4 µg), and luciferase activity was measured.
pTOP-tk- and pFOP-tk-luciferase (0.4 µg) were used as positive and negative controls,
respectively. Error bars represent the standard deviation of triplicate assays. (B) A
fragment containing nucleotides –586 to +82 was inserted in the sense or anti-sense
orientation into pGL3 [-586+82-luc(S) or –586+82-luc(AS), respectively]. COS-1
cells were transfected with these reporter constructs and luciferase activity was
measured. (C) Effects of a dominant-negative mutant of TCF-4 on the activity of the
BAMBI promoter in colorectal tumor cells. SW480, HCT116 and SW48 cells were
transfected with a luciferase reporter plasmid (0.4 µg of pTOP-tk-luc, Int1-luc or
–586+82-luc) and increasing amounts of TCF-4-∆C (0, 0.1, 0.4, and 1.2 µg), and
luciferase activity were measured. (D) (Left) Effects of dominant-negative mutants of
TCF-4 and ICAT on β-catenin-mediated activation of the BAMBI promoter. COS-1
cells were transfected with a luciferase reporter plasmid (0.3 µg, pTOP-tk-luciferase or -
586+82-luc) along with indicated plasmids (0.6 µg for β-cat S33Y, 1.2 µg for others),
and luciferase activity was measured. In all of the luciferase-reporter assays in this
figure, the total amount of plasmid DNA was adjusted with pcDNA3.1(+) empty
plasmid. The pRL-tk Renilla luciferase reporter (0.07 µg) was co-transfected to
normalize transfection efficiency. (Right, upper panel) Detection of exogenously
expressed FLAG-tagged TCF4-∆C, TCF4-∆N, and ICAT. Lysates prepared from
COS-1 cells transfected with the indicated plasmids were subjected to immunoblotting
with anti-FLAG antibody. (Right, lower panel) Nuclear localization of TCF4-∆C.
COS-1 cells were transfected with FLAG-tagged TCF4-∆C and stained with ant-FLAG
antibody (left). The nucleus was stained with TOTO-3 (right).
Fig. 3. Expression of BAMBI in colorectal and hepatocellular carcinomas. (A)
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

19
BAMBI expression was examined by semi-quantitative RT-PCR analysis. AXIN2 was
used as a marker of β-catenin-TCF-mediated activation. β-ACTIN was used as an RT-
PCR control to verify both the quality and quantity of template. N, non-cancerous
tissue; T, tumor. (B) Human colon tumor tissues and adjacent non-cancerous tissues
were double-stained with anti-β-catenin antibody (A, B, C, D) and anti-BAMBI-N-ter
(E, G) or –C-ter (F, H) antibody.
Fig. 4. BAMBI expression interferes with TGF-β signaling. (A) Effect of BAMBI
on colony formation. DU-145 cells were transfected with the indicated plasmids, and
cultured with 400 µg/ml of geneticin in the presence or absence of TGF-β1 for 3 weeks.
(B) Colonies generated in A were cloned and propagated. Lysates from these cells
were subjected to immunoblotting with anti-GFP or -α-tubulin antibody. Arrowheads
indicate BAMBI-GFP. BAMBI was detected as two bands presumably because of N-
glycosylation, since BAMBI was detected as one band in cells treated with tunicamycin
(our unpublished observation). The asterisk indicates a non-specific protein. (C)
BAMBI represses TGF-β-mediated transactivation. Cells expressing BAMBI obtained
in B were transfected with p3TP-lux, cultured in the presence or absence of TGF-β, and
luciferase activity was measured.
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Kuhara, Susumu Ohwada and Tetsu AkiyamaYoichi Furukawa, Yusuke Nakamura, Tsutomu Nakamura, Kousuke Tashiro, Satoru
Takashi Sekiya, Shungo Adachi, Kazuyoshi Kohu, Tatsuya Yamada, Osamu Higuchi,b-catenin pathway in colorectal tumor cellsS
Identification of BAMBI, an inhibitor of TGF-b signaling, as a target of the
published online December 1, 2003J. Biol. Chem.
10.1074/jbc.M310876200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on March 5, 2020
http://ww
w.jbc.org/
Dow
nloaded from