informe tecnico final análisis de algunos mecanismos de la...
TRANSCRIPT

INFORME TECNICO FINAL
Análisis de algunos mecanismos de la inmunidad innata involucrados en la destrucción de Entamoeba histolytica
Registro SIP: 20060359
Responsable: Dr Rafael Campos Rodriguez
RESUMEN
Actualmente conocemos muy poco sobre la inmunidad protectora en amibiasis,
pero aparentemente tanto la inmunidad innata como adquirida evitan la
infeccion amibiana. Los trofozoítos de Entamoeba histolytica deben
sobrepasar, evadir y sobrevivir a las respuestas inmunitarias del hospedero
para alcanzar y dividirse en el hígado. Es decir, la amibiasis invasora
solamente ocurre cuando Entamoeba histolytica tiene éxito en evadir las
respuestas inmunitarias naturales o cuando estas respuestas son ineficientes.
Aunque los mecanismos de la inmunidad natural implicados en la eliminación
de Entamoeba histolytica son mal entendidos, algunos resultados demuestran
que la inmunidad natural es suficiente para eliminar al parásito.
El objetivo de este trabajo es identificar cuales son las moléculas o células con
actividad citotóxica en contra de los trofozoítos de E. histolytica. Los resultados
mas relevantes son que moléculas con peso molecular de 250 y 50 KDa tienen
efecto citotóxico sobre trofozoitos de Entamoeba histolytica . Por otro lado, aun
cuando existe correlación entre el número de macrofagos y eosinofilos en la
protección in vivo contra la invasión amibiana. In vitro, los trofozoitos destruyen
a macrófagos y eosinófilos. La inmunidad innata en contra de Entamoeba
histolytica puede residir básicamente en mecanismos humorales citotóxicos.
ANTECEDENTES
El problema que se pretende estudiar es ¿cuáles son los mecanismos
humorales y celulares de la inmunidad natural involucrados en la eliminación de
Entamoeba histolytica ?
Actualmente conocemos muy poco sobre la inmunidad protectora en
amibiasis, pero aparentemente tanto la inmunidad innata como adquirida evitan
la infección amibiana (Haque et al, 2002). La inmunidad innata se refiere a la
1

primera linea de defensa del huésped en contra de la infección, la cual sirve
para controlar la infección durante las primeras horas de exposición al
microorganismo patogeno (Hoffman et al, 1999). Los trofozoítos de Eh deben
sobrepasar, evadir y sobrevivir a las respuestas inmunitarias del hospedero
para alcanzar y dividirse en el hígado. Es decir, la amibiasis invasora
solamente ocurre cuando Entamoeba histolytica tiene éxito en evadir las
respuestas inmunitarias naturales o cuando estas respuestas son ineficientes.
Aunque los mecanismos de la inmunidad natural implicados en la eliminación
de Entamoeba histolytica son mal entendidos, algunos resultados demuestran
que la inmunidad natural es suficiente para eliminar E. histolytica.
Los resultados de un trabajo previo que realizamos (Shibayama y otros,
2000) apoyan la importancia de la inmunidad natural en amibiasis. En este
modelo, el estímulo no específico (adyuvante incompleto de Freund, adyuvante
completo de Freund, BCG o aceite mineral) de células peritoneales,
especialmente las células inflamatorias mononucleares, parece ser suficiente
para eliminar el parásito. Por lo tanto, en ausencia de las células de T y de B
específicas para los antígenos de E. histolytica, el hámster infectado puede
montar una respuesta protectora contra E. histolytica. Sin embargo en ese
estudio no se analizaron cuales son los mecanismos humorales o celulares
involucrados en la eliminación de las amibas, por lo tanto, uno de los objetivos
generales de este trabajo es identificar cuales son las moléculas o células con
actividad citotoxica en contra de los trofozoítos de E. histolytica
Por otro lado, el sistema innato reconoce a los patógenos por medio de
receptores, en la membrana o solubles, que se unen a estructuras que se
repiten en patrones regulares característicos. Uno de estos es la lectina fijadora
de manosa (MBL). La activación del complemento por la vía de la MBL (MB-
lectin pathway) se inicia por la unión de la lectina a carbohidratos que contienen
manosa y algunos otros azucares, que se encuentran expuestos y dispuestos
en un patrón que es característico de los microorganismos. Por lo tanto, el
reconocimiento del patógeno y su discriminación de lo propio se debe al
reconocimiento de la orientación particular de ciertos restos de azucares así
como de su distribución en el espacio (o separación que es propio de microbios
y no de las células del hospedero. En consecuencia se propone como hipótesis
2

que los trofozoítos de Entamoeba histolytica no son reconocidos como
extraños porque la MBL no reconoce y se une al patrón de azúcares en su
membrana.
Finalmente, otra molécula que puede participar en la eliminación de las
amibas es la proteína C reactiva (CRP). CRP se une a fosfocolina,
fosfoetanolamina, fibronectina y otros ligandos (Black et al, 2004, 2003; Szalai
et al, 1999). Cuando estos ligandos estan expuestos (Hack et al,) la CRP se
une a ellos y activa eficientemente la via clasica del complemento a traves de
interacción directa con C1q. Tambien CRP puede interactuar con los
receptores para inmunoglobulinas Fc RI y Fc RII iniciando respuestas desde
las células fagocíticas. Consecuentemente, CRP es un importante componente
de la primera linea de defensa del huésped por su habilidad de reconocer
patógenos con el subsiguiente reclutamiento y activación del complemento y de
células fagocíticas (Szalai, 2002). Por lo consiguiente, es posible que la CRP
no se fije a fosfolipidos de membrana de Entamoeba histolytica porque éstos
no se encuentran expuestos o están cubiertos por proteínas propias o del
hospedero. Por el contrario, en amibas no patogenas, como Entamoeba dispar,
la proteína C reactiva fija y activa al complemento lo cual conduce a la lisis de
los trofozoítos.
Objetivo general
Identificar cuales son las moléculas o células con actividad citotoxica en
contra de los trofozoítos de E. histolytica en el peritoneo de hámsteres tratados
con agentes pro-inflamatorios no especificos (adyuvante incompleto de Freund
y aceite mineral) o preparaciones que inducen eosinofilia.
3

MATERIAL Y MÉTODOS Cultivo de trofozoítos de Eh en medio axénico. Los trofozoítos de
Entamoeba histolytica de la cepa HM-1: IMSS se cultivan en medio axénico de
acuerdo al procedimiento de Diamond (Diamond y cols, 1978).
Animales. Se utilizaron hámsteres adultos (Mesocricetus auratus), machos, de
90-110 g de peso y 2 meses de edad, en dos condiciones de estudio: grupo a)
hámsteres estimulados con aceite mineral (A/M) y b) grupo de hámsteres no
estimulados.
Inducción de exudado peritoneal. A cada hámster se les estimula con aceite
mineral (5 ml) por vía intraperitoneal (única dosis) con el objeto de producir
exudado peritoneal 7 días antes de la colocación de la bolsa de diálisis.
Procedimiento para la preparación y colocación de la bolsa de diálisis. La
bolsa de diálisis (Spectrum/Por 12 mm) de 25 kDa de poro, se corta a una
longitud de 5 cm, posteriormente se anuda uno de sus extremos con hilo seda
2-0 y se esteriliza a 15 libras de presión por 15 min, colocando 4x106
trofozoítos/1.0 ml de medio. Posteriormente los hámsteres se dejan en ayunas
por 12-24 h, se anestesiaron por vía intraperitoneal con 1.5 ml de Anestesal en
dosis de 4.72 mg/Kg de peso y bajo condiciones de esterilidad se procede a
realizar una laparotomía abdominal con una incisión media de 1.5 cm
aproximadamente. Posteriormente se colocaron las bolsas de diálisis y se
sutura por planos con un hilo de seda 2-0. Al término de cada tiempo (3, 6, 12 y
24 h) se realiza un examen macroscópico de los órganos internos y se
recuperaran las bolsas de diálisis. Se utilizaron tres animales por tiempo como
mínimo.
Viabilidad amibiana de los trofozoítos recuperados de la bolsa de diálisis de la cavidad peritoneal. Se sacrifican los hámsteres colocándolos en una
cámara de cristal con éter o cloroformo durante 3 a 5 min, posteriormente se
recuperaron las bolsas de diálisis en condiciones de esterilidad a los tiempos
de 3, 6, 12 y 24 h, se cuantifica el volumen final por bolsa y se determina la
viabilidad amibina. La viabilidad de los trofozoítos de E. histolytica a los
tiempos estudiados se mide empleando el colorante vital azul tripano entre
los 5-10 minutos después de recuperar la bolsa de diálisis, manteniendo las
4

muestras en hielo hasta realizarse todos los conteos. El número total de
trofozoítos en cada experimento se determinó usando el hematocitómetro de
Neubauer, en el microscopio de luz.
Obtención del sobrenadante de la bolsa de diálisis. Una vez obtenido el
sobrenadante de la bolsa de diálisis, se centrifuga a 10,000 rpm por 10 min y
se filtra en Millipore de 22 µM para asegurarse de que no existan células o
restos celulares. Las muestras se guardan a –70°C hasta su posterior
utilización.
Obtención del líquido peritoneal. En la cavidad abdominal se inyectan 15 ml
de solución salina al 0.9% o PBS amortiguador, se realiza un masaje suave en
el abdomen por 10 minutos y se obtiene el líquido peritoneal con una pipeta
Pasteur. Las muestras se guardan a –70°C hasta su posterior utilización.
Inducción de exudado peritoneal rico en eosinófilos. Para realizar la
inducción de eosinofilia peritoneal contamos con tres modelos en donde se
utilizara sephadex G-100, aceite mineral, Ascaris lumbricoides y Ascaris
suum,con el objetivo de obtener el modelo más adecuado.
En el caso de inoculación con sephadex G-100, se aplica una sola dosis por vía
intraperitoneal de 100mgs, y se realizan varios ensayos con recuento de
eosinofilos cada semana por cinco semanas.
Para la inoculación de aceite mineral se realizaron ensayos previos para poder
determinar la cantidad de inoculo, para obtener el mayor beneficio donde se
probaron dosis de 0.5, 2.0, 4.0 y 8.0ml, así como el tiempo más adecuado de
espera entre el inoculo y el sacrificio. De este ensayo se concluyo que un
inoculo de 4.0ml de aceite mineral y un periodo de espera de 7 días es el más
efectivo. Al realizar la inoculación con aceite mineral se utiliza una dosis de
1ml intraperitoneales y como en el caso de la inoculación con sephadex G-100
se realizan varios ensayos uno cada semana, por 5 semanas, cada una con su
respectiva cuenta de eosinòfilos.
Para la inducción de eosinofilia por medio de Ascaris lumbricoides y Ascaris
suum, se realiza un homogeneizado de las mismas al cual se le realiza
determinación de proteínas por el método de Bradford. Se inocula al hamster
por vía intraperitoneal una dosis de 19mg en un mililitro, y se realizan ensayos
en periodos de una a cinco semanas.
5

Posteriormente a encontrar el modelo más adecuado se procede a esperar un
periodo de 7 días para inocular al hamster, ahora, con trofozoitos de
Entamoeba Histolytica a dosis de 2X10° para la formación de absceso
hepático.
Cuenta diferencial de células del exudado peritoneal y tinción específica para eosinófilos. La cuenta diferencial se realiza mediante el empleo de
colorante de Wright-Giemsa modificado para el cuál se preparó solución stock
de Giemsa mezclando 66ml de glicerina y 1gr de Giemsa en polvo. Esta
mezcla se colocó a 60°C en un horno por dos horas. Finalmente se agrega a
esta mezcla 66 mL de alcohol metílico. Para preparar la solución de trabajo se
tomó una gota de la solución stock de Giemsa y se mezclo con un mlilitro de
solución salina 0.85%. Se añadió la solución de trabajo al frotis fijado y se tiño
durante 30 min. Se cubrió el frotis con agua corriente durante 5 min.
Posteriormente se lavo con agua destilada y se deja secar. Finalmente se
observa al microscopio, con aceite de imersión a 100x.. Los eosinófilos se
identificarán por el empleo de eritocina B. Los macrófagos se identificarán por
el empleo de la técnica de esterasas.
Tinción histoquímica por medio de eritrocina B. Se fijan los frotis con
formalina-etanol al 10%. Posteriormente se tiñen con hematoxilina de Harris
diluida durante 5 min. Se diferencian en agua corriente y se enjuagan en agua
destilada. A continuación se tiñen con eritrocina B al 0.05% en buffer de glicina
a pH 10, 30 min. Por último se diferencian en etanol al 70%. Deshidratar,
aclarar y montar en resina sintética. Los gránulos eosinofilicos se tiñeron de un
color rojo brilante. Se presume que este colorante se une preferentemente a la
MBP de los gránulos eosinófilos. .
Tinción histoquímica para macrófagos en frotis de células (Esterasas no específicas): Los frotis se fijarán con formaldehido al 4% en 0.15 M de buffer
sodio/fosfato (ph 7.2). Agregar colorante para esterasas incubar por 3 a 40 min
a 230C (temperatura ambiente). Lavar con agua destilada en tres tiempos,
contrateñir con hematoxilina de Harris. Por último montar en glicerol gelatina.
Técnica de Griess. La técnica de Griess para determinar nitritos se realiza de
la siguiente manera (Pacheco y cols. 2001): Las muestras de suero se
desproteinizadas previamente con sulfato de zinc al 30%. A 250 µl de la
6

muestra se le adicionan 12.5 µl de sulfato de zinc, se centrifuga a 9000 rpm x
15 min en una microcentrífuga (Fisher Scientific). A las muestras de líquido
peritoneal y sobrenadante de la bolsa de diálisis no es necesario desproteinizar
con sulfato de zinc. Posteriormente, las muestras de suero, líquido peritoneal y
sobrenadante de la bolsa de diálisis se diluyen dos formas (dilución 1:100 y
1:500 con agua bidestilada), con el objeto de que las absorbancias obtenidas
entraran en el rango de la curva estándar. Posteriormente se toman 150 µl de
cada muestra y se colocan en cada pozo (placas de 96 pozos) por duplicado.
En cada pozo se colocan dos limaduras de cadmio para reducir nitratos a
nitritos, se incuba por 45 min a temperatura ambiente, se retiran las limaduras
de cadmio y se agregan 50µl del reactivo de Griess
(Sulfanilamida/naftilendiamina 1:1), incubando por 10 min a temperatura
ambiente. La absorbancia se lee a 570 nm y el estándar de NaNO2 se prepara
a una concentración de 2.5 a 100 µM.
Ensayo de viabilidad de amibas por metodo espectrometrico empleando cristal violeta. Ajustar suspensión de amibas a 5 x 105 células/mL. Diluir la
solución por probar en medio de cultivo. Añadir 100 ul de cada solución por
triplicado (cuadriplicado). En las dos primeras columnas, añadir diferentes
diluciones de la suspensión de amibas para elaborar curva estándar (50,000,
25,000, 12,500, 6250, etc). Añadir medio de cultivo en cuatro pozos de control.
A cada pozo con las muestras por probar se agregan las células (p, ej., 50,000
celulas/100ul). Sellar las placas con parafilm e incubar (2-4 horas) a 37 oC.
Eliminar el medio sacudiendo la placa (flick out). Lavar suavemente con salina
o PBS tibio (dos veces), y las restantes celulas se fijan con 100 ul, o llenando
los pozos con metanol por 30 segundos. Retirar el resto de metanol y añadir
100 ul de la solución acuosa de cristal violeta al 1 %, en cada pozo y dejar
incubando por 5-20 min. Eliminar el colorante sacudiendo la placa y lavar con
agua destilada. Invertir la placa sobre papel secante por uno minutos hasta
secar. Las células teñidas se solubilizan en SDS 1 % en 50 % de etanol. La
intensidad de la tinción se lee a una longitud de 570 -620 nm.
Separación de células de cavidad peritoneal por medio de gradientes de Percoll. Se recomienda preparar los gradientes cada día. Los gradientes se
preparan a partir de Percoll al 100%, con volúmenes de Percoll puro y 1
7

volumen de PBS 10x. Esta solución se diluye con Hanks para preparar
soluciones al 30, 50 y 80% que posteriormente se depositan en un tubo de
vidrio de 13x100 mm, quedando al fondo, el gradiente más denso.
Las células peritoneales se concentran por centrifugación, lavan en PBS y
resuspenden a 2-4 x108 en 3 ml de PBS. Se depositan sobre los gradientes
discontinuos de Percoll. Estos se forman en tubos cónicos de 15 ml. de
polestireno con 3 ml. de las siguientes concentraciones: 45%, 55%, 65% y
75%. Centrifuga a 400 x g por 15 min. a temperatura ambiente. Las diferentes
bandas se colectan con pipeta y se lavan con PBS. Lavar con solución de
Hanks una vez a 400 x g a 40C. La viabilidad se determina por exclusión de
tripano azul y la pureza (mayor de 90%) por Wright-Giemsa modificado.
Interacción amibas y células peritoneales. Las amibas y las células del
exudado peritoneal (eosinófilos, macrófagos) se combinan en medio TYI
suplementado con 5.7 mM de cisteína a diferentes relaciones células/amibas.
Las interacciones se realizan con tubos de polipropileno de 12 x 75 mm.
Centrifuga a 150 g. X 5 min. para formar un botón que permita el contacto
célula a célula. Los controles se incuban solos en su propio medio de cultivo.
Se incuba a 37 grados y toman muestras a 0, 60, 90, 180 min. y 6 hrs.. Se
retira 0.5 ml del sobrenadante dejando el paquete intacto. Se agrega 0.5 ml de
tripano azúl 1% en NaCl 0.9%. El paquete se disgrega por vortex e incuba por
5 min., se cuenta el número de amibas, células vivas y muertas, se cuentan en
el hemocitómetro por tinción con tripano azúl. Los datos se expresan: a) como
el porcentaje de la viabilidad de los testigos de amibas incubadas solas por 6
hrs., b) como por ciento de destrucción de amibas, c) como por ciento de
sobrevida
Células viables / ml al min. “n”
% de sobrevida= -------------------------------------------
Células viables / ml al min. 0
Columnas
Se pasa este mL de bilis en una columna de Sepharosa 4B, obteniendo
dos picos (1 y 2) de IgA, detectados por el procedimiento de doble
inmunodifusión en gel de agarosa al 2%. Se corrobora la presencia de IgA de
bilis de rata, mediante el procedimiento de DOT-Elisa (ELISA en mancha), en el
8

pico 1 y 2. Se concentra la IgA de bilis de rata del pico 1 y 2, mediante
liofilización y se procede a dializarlos en solución salina. En ambos picos se
procede a cuantificar la concentración de IgA mediante el procedimiento de
Bradford.
Purificación de fracciones proteicas en liquido peritoneal mediante cromatografía. El liquido peritoneal concentrado previamente por liofilización
se depositó sobre una columna de separación de Separosa 4B de 2.5 x 100
cm, previamente equilibrada con regulador de Tris –HCl, EDTA 2mM, pH 7.3.
Se colectan partes alicuotas de 2 ml cada una hasta un total de 140 fracciones,
en las cuales se determina la concentración de proteínas por el método de
Bradford, utilizando un equipo comercial. Con los datos obtenidos se
elaboraron las graficas de los perfiles de elución. El siguiente paso fue liofilizar
las fracciones obtenidas. Después se reúnen y diluyen en un mínimo de
solvente, se separa el exceso de sales por diálisis en regulador de fosfato y se
separan en una columna de DEAE sepharosa.
Electroforesis en SDS-PAGE. El liquido peritoneal concentrado se mezcla
volumen a volumen con regulador de muestra (2 % de SDS en regulador de
Tris 0.5 M pH 6.8, con glicerol al 10%) y se somete a ebullición por 5 minutos.
La mezcla (20 ug de proteínas amibianas por carril) se separa por
electroforesis en geles de poliacrilamida-SDS (Laemmli, 1970), al 7.5 % de
acrilamida en el gel separador, por dos horas a 75 V y no más de 20 mA.
Analisis estadístico. Para la evaluación estadística de los resultados
obtenidos se utiliza la prueba de análisis de varianza (ANOVA). .
9

RESULTADOS
I. MECANISMOS HUMORALES
Transferencia pasiva de inmunidad protectora. Grupos de 10-12
animales se inocularon por vía intraperitoneal con 8 ml de medio de aceite
mineral (Sigma). A los tres días se obtuvieron de un grupo donador las
suspensiones celulares del exudado peritoneal y transfirieron 5 ml de la
suspensión (con un promedio de 12.5 x 106 células) a un grupo de animales
denominados receptores. Inmediatamente después se aplicaron por
intraperitoneal 2 x 106 trofozoítos y a los siete días se sacrificaron. En los
animales que recibieron aceite mineral (grupo P) no hubo lesiones (Figura 1),
tampoco en los donadores (grupo D) y en los animales receptores (grupo R)
de células del exudado, excepto en un caso, pero esta fue tamaño pequeño.
Esto indica que la protección puede transferirse mediante células del exudado
inflamatorio sin especificidad para Eh.
0
5
10
15
20
25
30
35
40
45
50
T P D R
Tam
año
del a
bsce
so (%
)
10

Figura 1. Transferencia de células de exudado peritoneal (inmunidad pasiva): Aceite Mineral. Grupos de 10-12 animales se inocularon por vía intraperitoneal con 9 ml de medio de aceite mineral (Sigma). A los tres días se obtuvieron de un grupo donador las suspensiones célulares del exudado peritoneal y transfieron 5 ml de la suspension (con un promedio de 12.5 x 106 células) a un grupo de animales denominados receptores. Inmediatamente después se aplicaron por intraperitoneal 2 x 106 trofozoitos y a los siete días se sacrificaron.
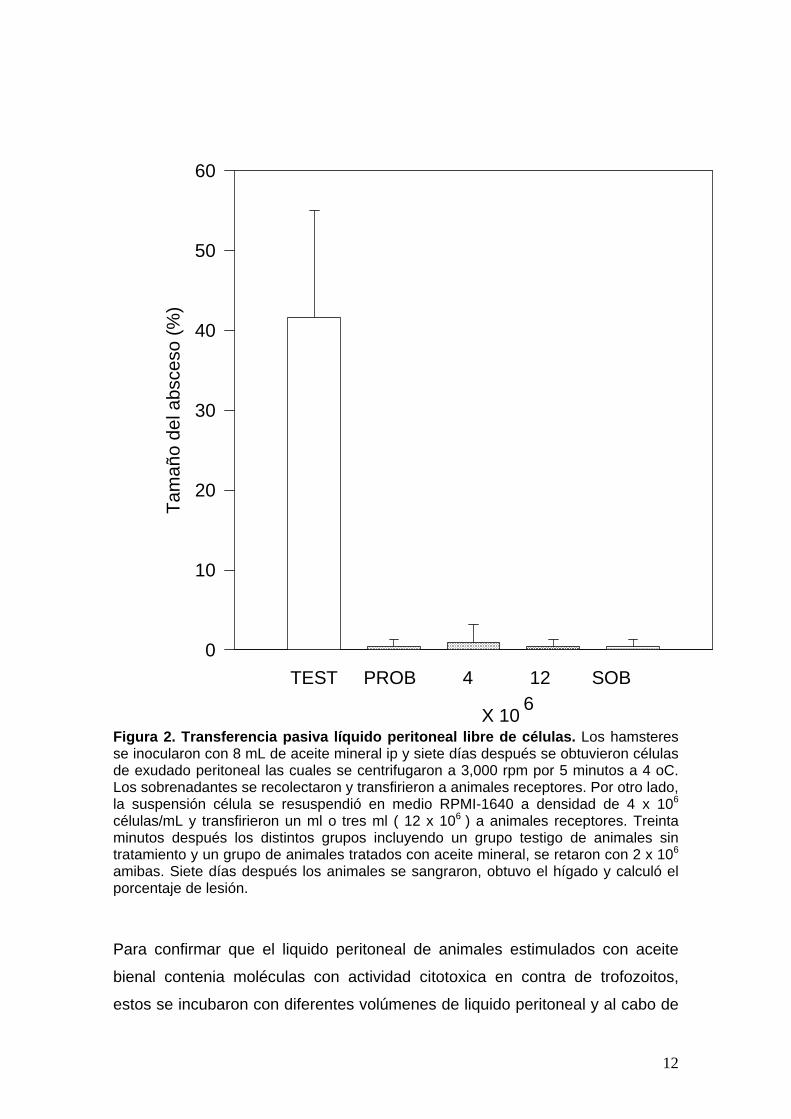
La transferencia de protección puede hacerse con células y líquido peritoneal libre de células.. Los hámsteres se inocularon con 8 mL de aceite
mineral ip y siete días después se obtuvieron células de exudado peritoneal las
cuales se centrifugaron a 3,000 rpm por 5 minutos a 4 oC. Los sobrenadantes
se recolectaron y transfirieron a animales receptores. Por otro lado, la
suspension celular se resuspendió en medio RPMI-1640 a densidad de 4 x 106
células/mL y transfirieron un ml o tres ml ( 12 x 106 ) a animales receptores.
Treinta minutos después los distintos grupos incluyendo un grupo testigo de
animales sin tratamiento y un grupo de animales tratados con aceite mineral, se
retaron con 2 x 106 amibas. Siete días después los animales se sangraron,
obtuvo el hígado el cual se pesó así como las lesiones hepáticas. De acuerdo a
los resultados (Figura 2) los animales que recibieron pasivamente células de
exudado peritoneal no tuvieron lesiones hepáticas. Lo mismo sucedió con
animales receptores que únicamente recibieron sobrenadante libre de células.
Los resultados sugieren que en el líquido peritoneal, libre de células, de
animales tratados con aceite mineral, existe algún componente soluble capaz
de destruir a los trofozoítos de Eh. Es poco probable que sea óxido nítrico
puesto que tiene vida media muy corta.
11
Tam
año
del a
bsce
so (%
)
0
10
20
30
40
50
60
TEST PROB 4 12 SOB
X 10 6
Figura 2. Transferencia pasiva líquido peritoneal libre de células. Los hamsteres se inocularon con 8 mL de aceite mineral ip y siete días después se obtuvieron células de exudado peritoneal las cuales se centrifugaron a 3,000 rpm por 5 minutos a 4 oC. Los sobrenadantes se recolectaron y transfirieron a animales receptores. Por otro lado, la suspensión célula se resuspendió en medio RPMI-1640 a densidad de 4 x 106
células/mL y transfirieron un ml o tres ml ( 12 x 106 ) a animales receptores. Treinta minutos después los distintos grupos incluyendo un grupo testigo de animales sin tratamiento y un grupo de animales tratados con aceite mineral, se retaron con 2 x 106
amibas. Siete días después los animales se sangraron, obtuvo el hígado y calculó el porcentaje de lesión.
Para confirmar que el liquido peritoneal de animales estimulados con aceite
bienal contenia moléculas con actividad citotoxica en contra de trofozoitos,
estos se incubaron con diferentes volúmenes de liquido peritoneal y al cabo de
12
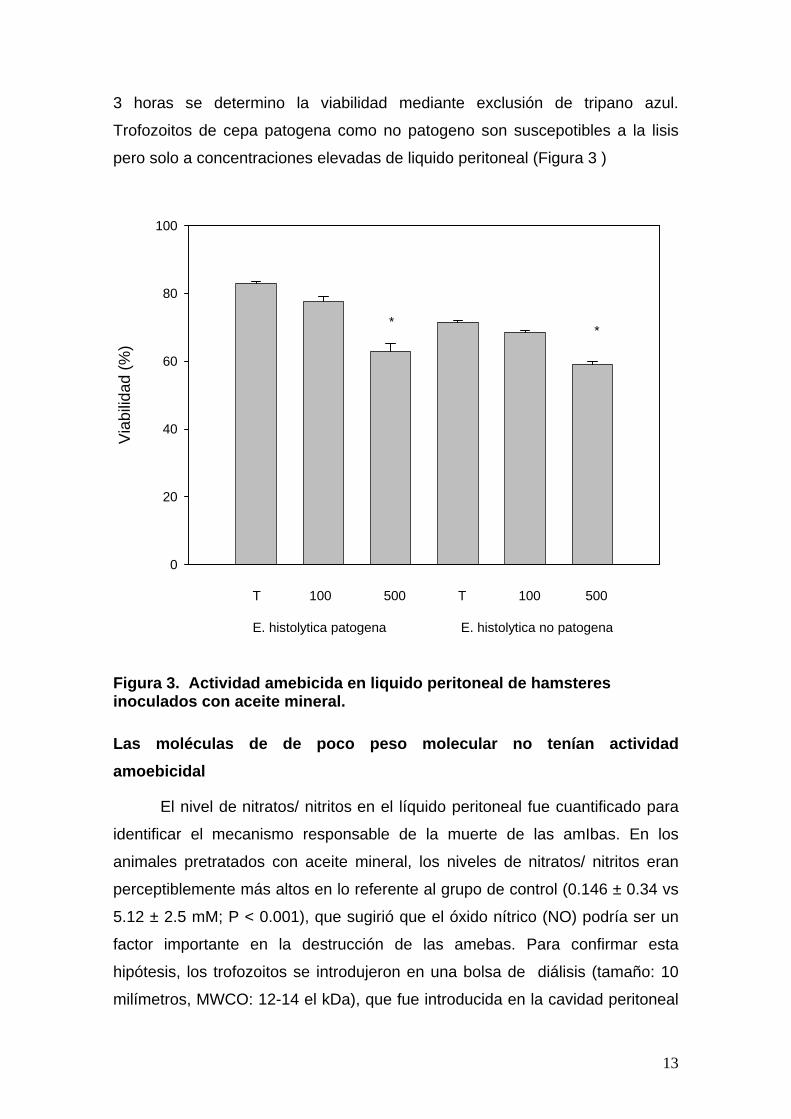
3 horas se determino la viabilidad mediante exclusión de tripano azul.
Trofozoitos de cepa patogena como no patogeno son suscepotibles a la lisis
pero solo a concentraciones elevadas de liquido peritoneal (Figura 3 )
Viab
ilida
d (%
)
0
20
40
60
80
100
T 100 500 T 100 500 E. histolytica patogena E. histolytica no patogena
**
Figura 3. Actividad amebicida en liquido peritoneal de hamsteres inoculados con aceite mineral.
Las moléculas de de poco peso molecular no tenían actividad amoebicidal
El nivel de nitratos/ nitritos en el líquido peritoneal fue cuantificado para
identificar el mecanismo responsable de la muerte de las amIbas. En los
animales pretratados con aceite mineral, los niveles de nitratos/ nitritos eran
perceptiblemente más altos en lo referente al grupo de control (0.146 ± 0.34 vs
5.12 ± 2.5 mM; P < 0.001), que sugirió que el óxido nítrico (NO) podría ser un
factor importante en la destrucción de las amebas. Para confirmar esta
hipótesis, los trofozoitos se introdujeron en una bolsa de diálisis (tamaño: 10
milímetros, MWCO: 12-14 el kDa), que fue introducida en la cavidad peritoneal
13
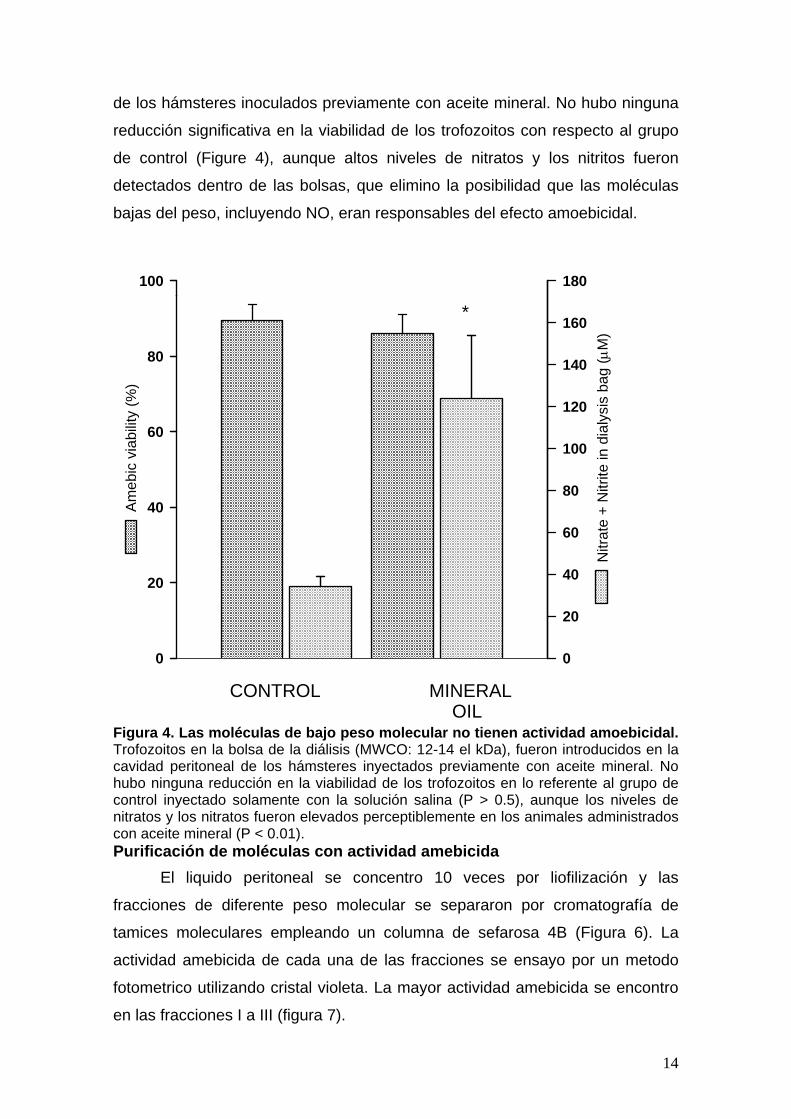
de los hámsteres inoculados previamente con aceite mineral. No hubo ninguna
reducción significativa en la viabilidad de los trofozoitos con respecto al grupo
de control (Figure 4), aunque altos niveles de nitratos y los nitritos fueron
detectados dentro de las bolsas, que elimino la posibilidad que las moléculas
bajas del peso, incluyendo NO, eran responsables del efecto amoebicidal.
Am
ebic
via
bilit
y (%
)
0
20
40
60
80
100
Nitr
ate
+ N
itrite
in d
ialy
sis
bag
(µM
)
0
20
40
60
80
100
120
140
160
180
CONTROL MINERAL OIL
*
Figura 4. Las moléculas de bajo peso molecular no tienen actividad amoebicidal. Trofozoitos en la bolsa de la diálisis (MWCO: 12-14 el kDa), fueron introducidos en la cavidad peritoneal de los hámsteres inyectados previamente con aceite mineral. No hubo ninguna reducción en la viabilidad de los trofozoitos en lo referente al grupo de control inyectado solamente con la solución salina (P > 0.5), aunque los niveles de nitratos y los nitratos fueron elevados perceptiblemente en los animales administrados con aceite mineral (P < 0.01). Purificación de moléculas con actividad amebicida
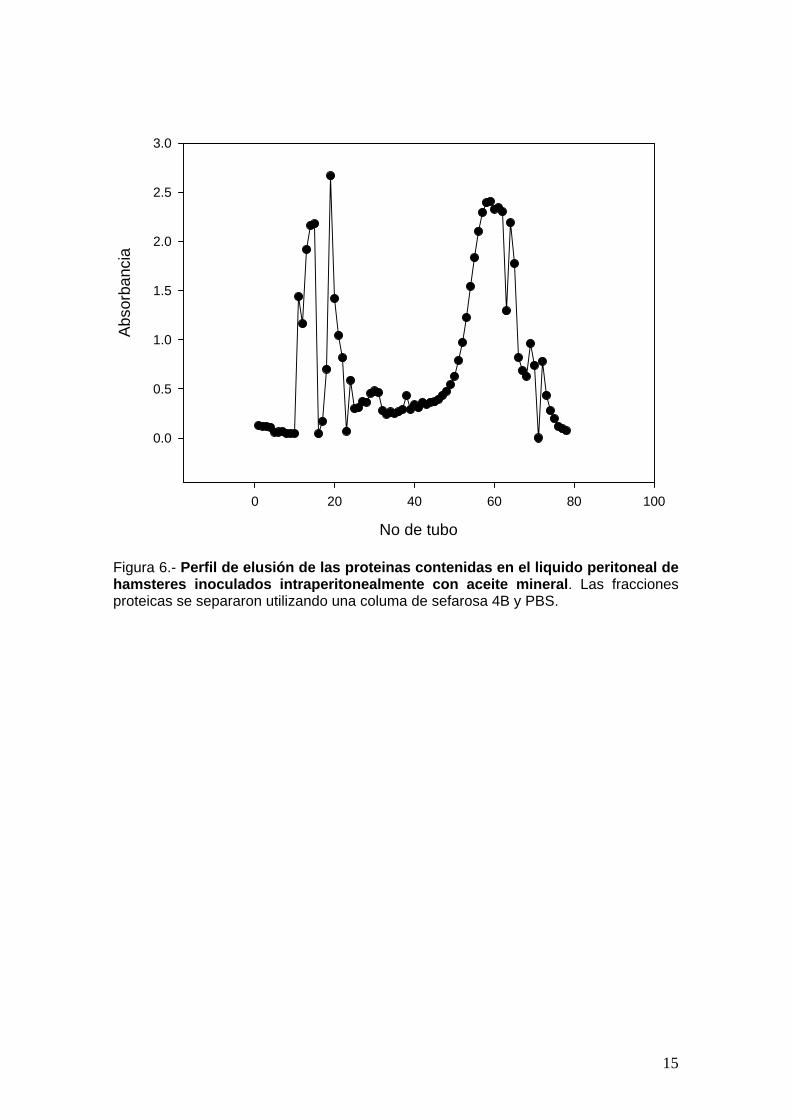
El liquido peritoneal se concentro 10 veces por liofilización y las
fracciones de diferente peso molecular se separaron por cromatografía de
tamices moleculares empleando un columna de sefarosa 4B (Figura 6). La
actividad amebicida de cada una de las fracciones se ensayo por un metodo
fotometrico utilizando cristal violeta. La mayor actividad amebicida se encontro
en las fracciones I a III (figura 7).
14
No de tubo
0 20 40 60 80 100
Abso
rban
cia
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Figura 6.- Perfil de elusión de las proteinas contenidas en el liquido peritoneal de hamsteres inoculados intraperitonealmente con aceite mineral. Las fracciones proteicas se separaron utilizando una columa de sefarosa 4B y PBS.
15

Via
bilid
ad (N
o am
ibas
)
0.0
5.0e+4
1.0e+5
1.5e+5
2.0e+5
2.5e+5
3.0e+5
3.5e+5
FraccionesT I II III IV
Figura 7 Actividad amebicida de fracciones separadas por cromatografía de tamices moleculares. La lisis de trofozoitos se cuantifico mediante ensayo fotometrico que emplea cristal violeta. Hubo reducción significativa en el número de amibas en las fracciones I a IV.
Las fracciones I a III se reunieron, liofilizaron, resuspendieron en
regulador de fosfatos y pasaron por columna de intercambio iónico (DEAE-
sefarosa) . Empleando diferentes fgradientes de concertación o fuerza iónica se
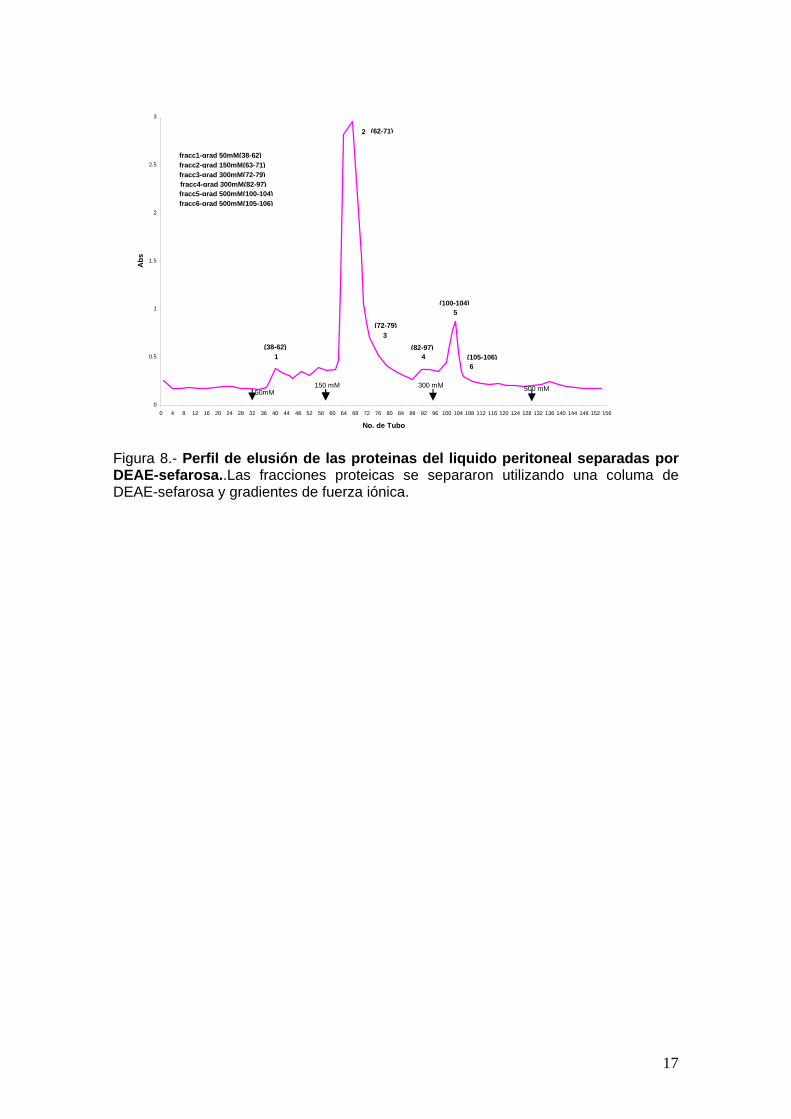
separaron varias fracciones (Figura 8) cuya actividad citotoxica se probó
utilizando el ensayo de viabilidad mencionado (Figura 9). La mayor actividad
citotoxico se encontro en las fracciones eluidas con regulador de fuerza ionica
elevada (fraciones IV a VI).
16
0
0.5
1
1.5
2
2.5
3
0 4 8 12 16 20 24 28 32 36 40 44 48 52 56 60 64 68 72 76 80 84 88 92 96 100 104 108 112 116 120 124 128 132 136 140 144 148 152 156
No. de Tubo
Abs
fracc1-grad 50mM(38-62)fracc2-grad 150mM(63-71)fracc3-grad 300mM(72-79) fracc4-grad 300mM(82-97)fracc5-grad 500mM(100-104)fracc6-grad 500mM(105-106)
1
2
3
4
5
6
(38-62)
(62-71)
(72-79)
(82-97)
(100-104)
(105-106)
50mM150 mM 300 mM 500 mM
Figura 8.- Perfil de elusión de las proteinas del liquido peritoneal separadas por DEAE-sefarosa..Las fracciones proteicas se separaron utilizando una columa de DEAE-sefarosa y gradientes de fuerza iónica.
17

Viab
ilidad
(No
amib
as)
0
10000
20000
30000
40000
50000
60000
T I II III III IV V VI
Figura 9.- Actividad amebicida de fracciones separadas por cromatografía de intercambio iónico. La lisis de trofozoitos se cuantifico mediante ensayo fotometrico que emplea cristal violeta. Hubo reducción significativa en el número de amibas en las fracciones IV a VI..
Las fracciones se analizaron mediante SDS-PAGE y se muestran en la figura
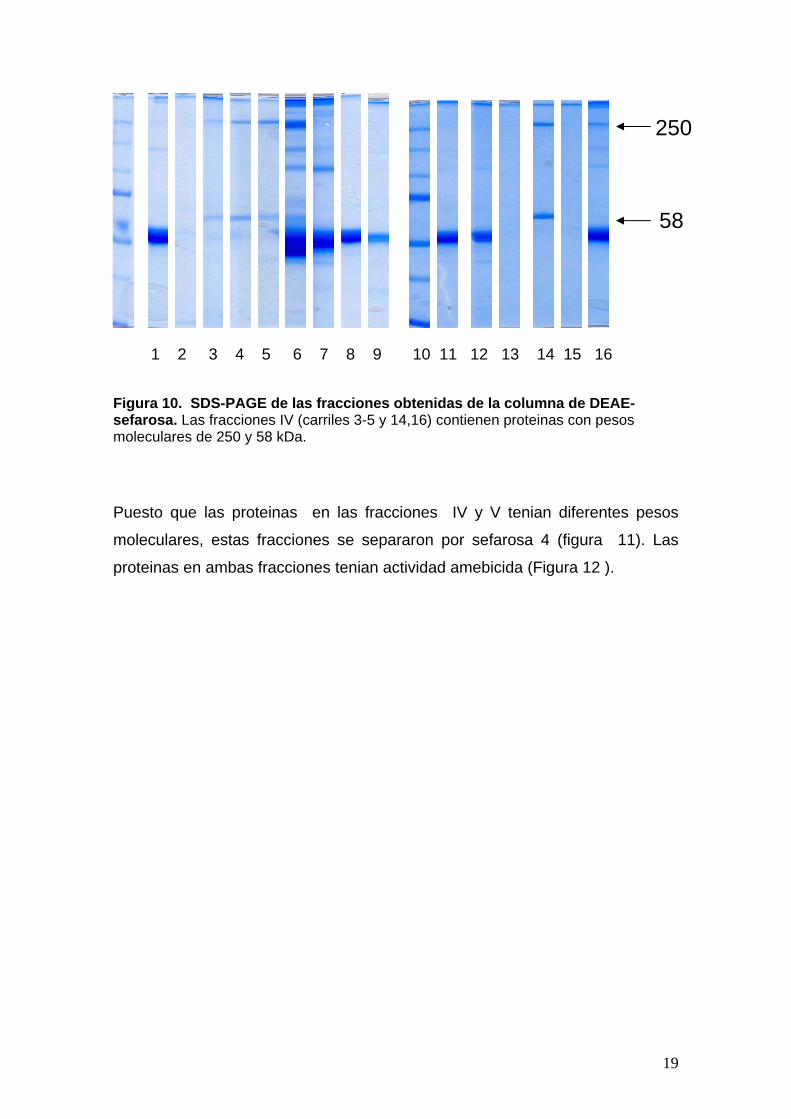
10. Las fracciones con mayor actividad amebicidad contienen proteinas con
pesos moleculares de 250 y 57 kDa.
18
250
58
6 Figusefamole
Pue
mole
prote
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 1
ra 10. SDS-PAGE de las fracciones obtenidas de la columna de DEAE-rosa. Las fracciones IV (carriles 3-5 y 14,16) contienen proteinas con pesos culares de 250 y 58 kDa.
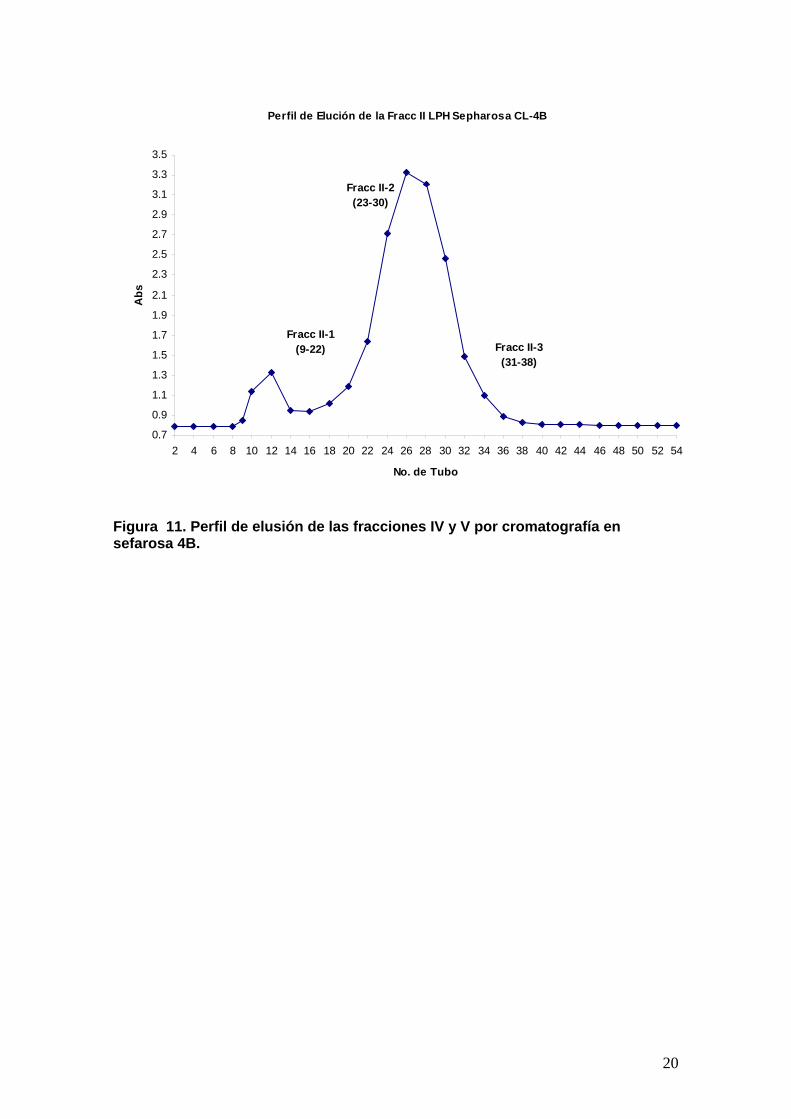
sto que las proteinas en las fracciones IV y V tenian diferentes pesos
culares, estas fracciones se separaron por sefarosa 4 (figura 11). Las
inas en ambas fracciones tenian actividad amebicida (Figura 12 ).
19
Perfil de Elución de la Fracc II LPH Sepharosa CL-4B
0.7
0.9
1.1
1.3
1.5
1.7
1.9
2.1
2.3
2.5
2.7
2.9
3.1
3.3
3.5
2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40 42 44 46 48 50 52 54
No. de Tubo
Abs
Fracc II-1(9-22)
Fracc II-2(23-30)
Fracc II-3(31-38)
Figura 11. Perfil de elusión de las fracciones IV y V por cromatografía en sefarosa 4B.
20
Via
bilid
ad (%
)
0
20
40
60
80
100
T II-1 II-2 FRACCIONES
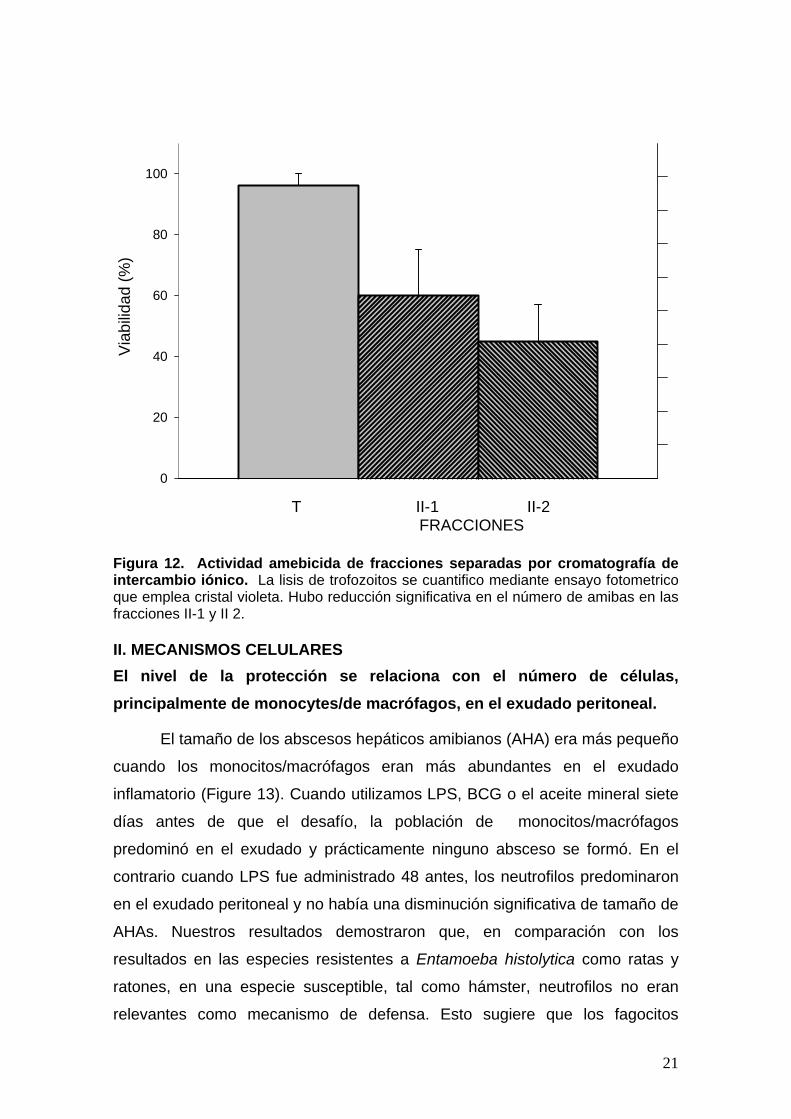
Figura 12. Actividad amebicida de fracciones separadas por cromatografía de intercambio iónico. La lisis de trofozoitos se cuantifico mediante ensayo fotometrico que emplea cristal violeta. Hubo reducción significativa en el número de amibas en las fracciones II-1 y II 2. II. MECANISMOS CELULARES El nivel de la protección se relaciona con el número de células, principalmente de monocytes/de macrófagos, en el exudado peritoneal.
El tamaño de los abscesos hepáticos amibianos (AHA) era más pequeño
cuando los monocitos/macrófagos eran más abundantes en el exudado
inflamatorio (Figure 13). Cuando utilizamos LPS, BCG o el aceite mineral siete
días antes de que el desafío, la población de monocitos/macrófagos
predominó en el exudado y prácticamente ninguno absceso se formó. En el
contrario cuando LPS fue administrado 48 antes, los neutrofilos predominaron
en el exudado peritoneal y no había una disminución significativa de tamaño de
AHAs. Nuestros resultados demostraron que, en comparación con los
resultados en las especies resistentes a Entamoeba histolytica como ratas y
ratones, en una especie susceptible, tal como hámster, neutrofilos no eran
relevantes como mecanismo de defensa. Esto sugiere que los fagocitos
21
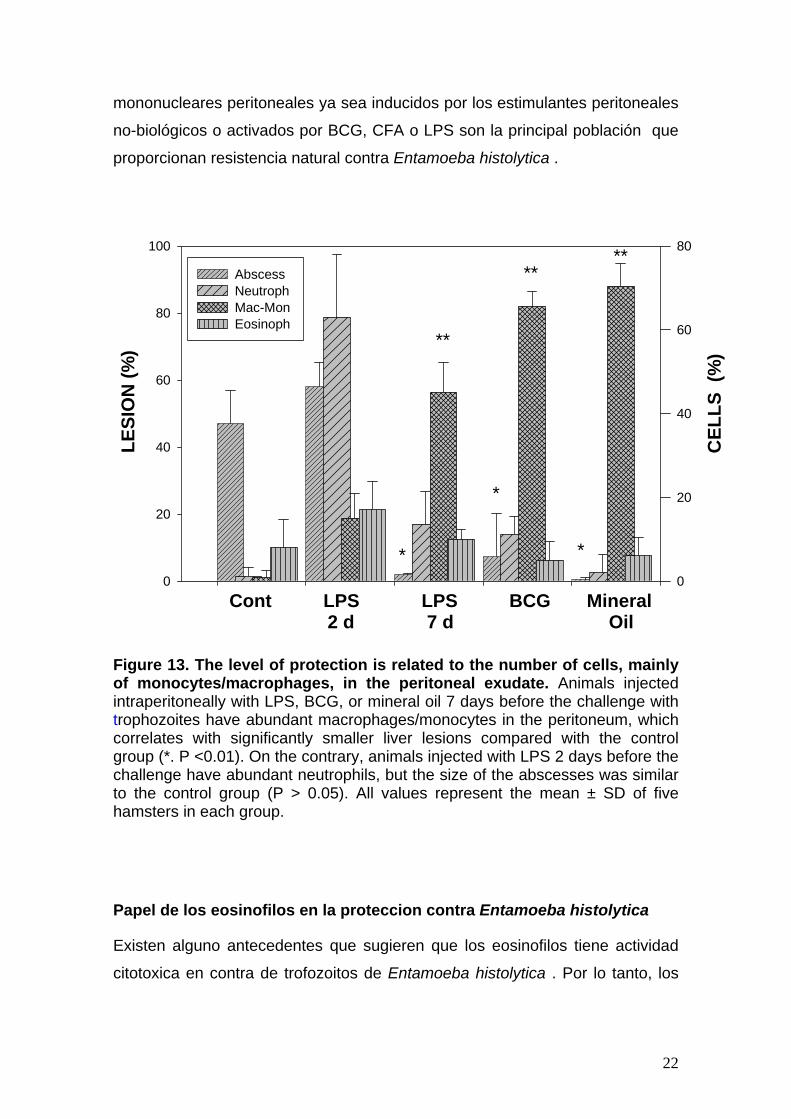
mononucleares peritoneales ya sea inducidos por los estimulantes peritoneales
no-biológicos o activados por BCG, CFA o LPS son la principal población que
proporcionan resistencia natural contra Entamoeba histolytica .
LESI
ON
(%)
0
20
40
60
80
100
CEL
LS (
%)
0
20
40
60
80
AbscessNeutroph Mac-Mon Eosinoph
Cont LPS LPS BCG Mineral 2 d 7 d Oil
*
*
*
**
****
Figure 13. The level of protection is related to the number of cells, mainly of monocytes/macrophages, in the peritoneal exudate. Animals injected intraperitoneally with LPS, BCG, or mineral oil 7 days before the challenge with trophozoites have abundant macrophages/monocytes in the peritoneum, which correlates with significantly smaller liver lesions compared with the control group (*. P <0.01). On the contrary, animals injected with LPS 2 days before the challenge have abundant neutrophils, but the size of the abscesses was similar to the control group (P > 0.05). All values represent the mean ± SD of five hamsters in each group.
Papel de los eosinofilos en la proteccion contra Entamoeba histolytica
Existen alguno antecedentes que sugieren que los eosinofilos tiene actividad
citotoxica en contra de trofozoitos de Entamoeba histolytica . Por lo tanto, los
22
siguientes procedimientos tuvieron como finalidad analizar la participación del
eosinofilo en la inmunidad innata en la amibiasis.
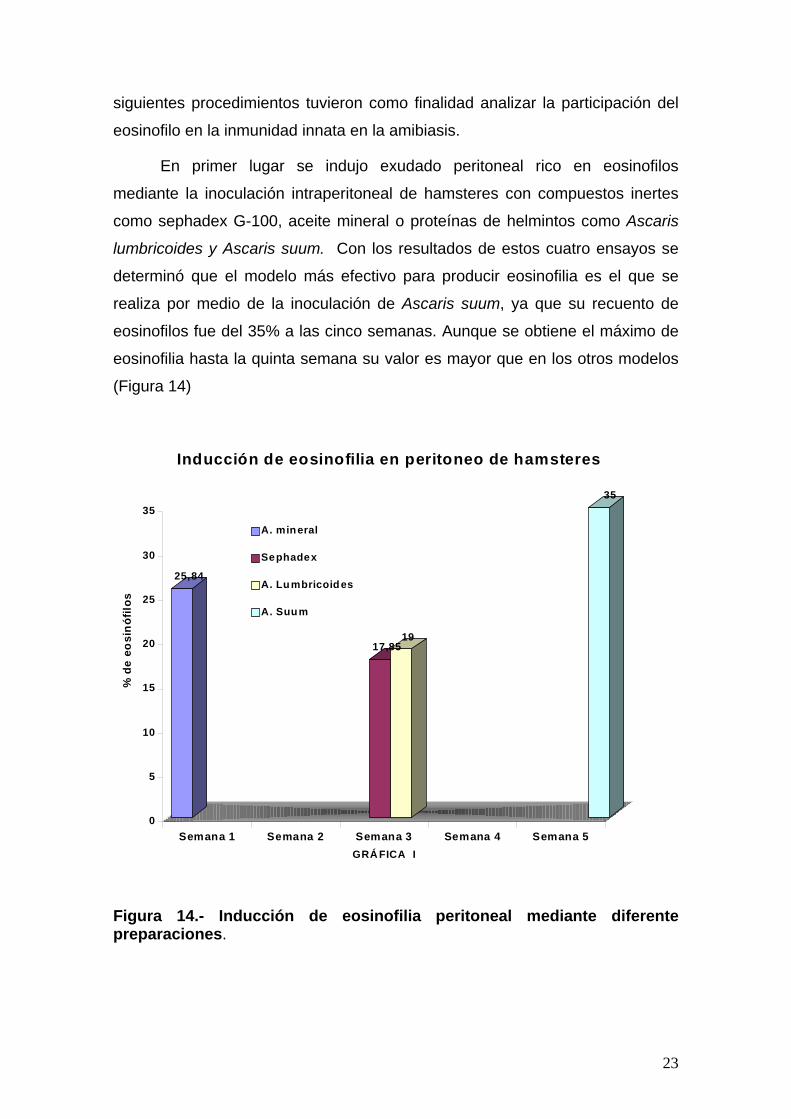
En primer lugar se indujo exudado peritoneal rico en eosinofilos
mediante la inoculación intraperitoneal de hamsteres con compuestos inertes
como sephadex G-100, aceite mineral o proteínas de helmintos como Ascaris
lumbricoides y Ascaris suum. Con los resultados de estos cuatro ensayos se
determinó que el modelo más efectivo para producir eosinofilia es el que se
realiza por medio de la inoculación de Ascaris suum, ya que su recuento de
eosinofilos fue del 35% a las cinco semanas. Aunque se obtiene el máximo de
eosinofilia hasta la quinta semana su valor es mayor que en los otros modelos
(Figura 14)
25,84
17,8519
35
0
5
10
15
20
25
30
35
% d
e eo
sin
ófilo
s
Semana 1 Semana 2 Semana 3 Semana 4 Semana 5GRÁ FICA I
Inducción de eosinofilia en peritoneo de hamsteres
A. mineral
Sephadex
A. Lumbricoides
A. Suum
Figura 14.- Inducción de eosinofilia peritoneal mediante diferente preparaciones.
23

Para identificar los eosinófilos y otras células del exudado peritoneal mediante
observaciones al microscopio óptico se realizaron las siguientes tinciones
específicas: Técnica de eritrosina B para identificar eosinófilos; esterasa no
específica para macrófagos y azul de toluidina para mastocitos (Figuras 15, 16
). y Wright Giemsa para otras células.
l
FiguLa figa un prote
Fipeobob
contro
ra 15: Eosinofilos. Se observan fura del lado izquierdo pertenece ahamster estimulado con aceite minínas de scaris suum. ( 100X )
A
gura 16.-Macrofagos y mastocritoneal de hamsteres previamenservamos macrofagos teñidos coservamos mastocitos teñidos con
Aceitemineral
rotis de exudado peritoneal, teñ un hamster no estimulado la deral y la del lado derecho a unoB
itos. Fotomicrografía de celulte inmunoestimulados con Ascn esterasa no especifica a 40Xazul de toluidina a 100X(flecha
A suum
idos con eritrosina B; e en medio pertenece estimulado con
as de exudado aris suum. En A
(flechas). En B s).
24

Después de establecer que el mejor modelo de inducción de eosinofilia
peritoneal fue en el que se utilizo proteínas de Ascaris suum, se inocularon 2 x
106 trofozoitos patógenos de Eh en hamsteres testigo y en aquellos inoculados
con. Ascaris suum y a los siete dìas se sacrificaron y se cuantifico el tamaño
del absceso. Los abscesos hepáticos amibianos en hamsteres inoculados con
proteínas de Ascaris suum fueron considerablemente menores (4.3%) en
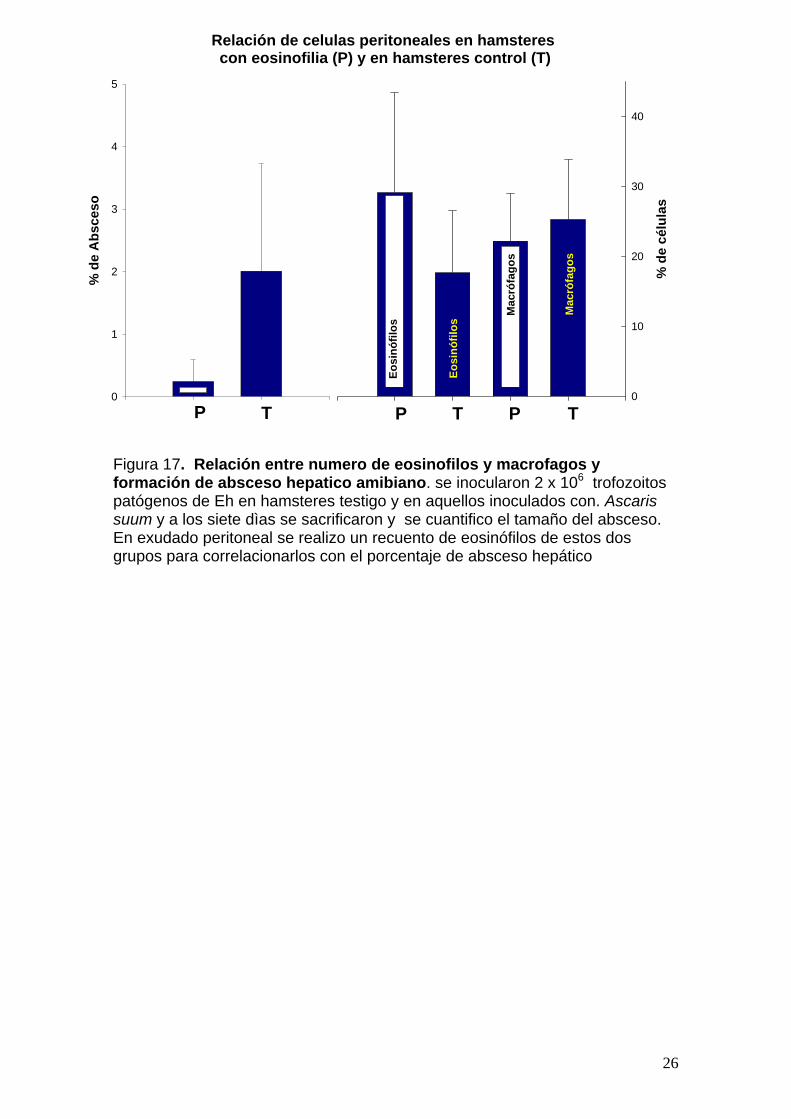
comparación con los formados en el grupo testigo (27.8%). (Figura 17). Se
realizo un recuento de eosinófilos de estos dos grupos para correlacionarlos
con el porcentaje de absceso hepático. El grupo testigo presento un porcentaje
de absceso del 27.8% y un recuento de eosinofilos del 17.6%, comparado con
un 4.3% de absceso hepático y un 29% de eosinófilos en el grupo problema.
Con estos resultados podemos decir que los eosinófilos probablemente
juegan un papel importante en la protección contra el absceso hepático
amibiano, ya que en los animales que se les provoco eosinofilia presentaron un
porcentaje de lesión considerablemente menor a los animales del grupo testigo.
Es decir a mayor cantidad de eosinófilos, menor tamaño de absceso hepático y
a menor número de eosinófilos, mayor tamaño de absceso hepático. Cabe
mencionar que también se cuantificaron otras células del exudado peritoneal
para correlacionarlas con el absceso hepático amibiano y observar si tienen
alguna participación; mediante la técnica de azul de toluidina se identificaron
mastocitos; con el método de esterasa no especifica se identificaron
macrófagos, estos últimos son los de importancia por el porcentaje con que se
observaron siendo del 22.1% para el grupo de hamsteres con eosinofilia y del
25.2% para el grupo testigo.
El examen macroscópico nos revela que el absceso hepático en hamsteres con
eosinófilia al termino de 7 días es significativamente menor (Figura 18-A) que
en el grupo de hamsteres testigo (Figura 18-B).
25
% d
e A
bsce
so
0
1
2
3
4
5
Relación de celulas peritoneales en hamsteres con eosinofilia (P) y en hamsteres control (T)
% d
e cé
lula
s
0
10
20
30
40
P T P TEo
sinó
filos
Eosi
nófil
os
Mac
rófa
gos
Mac
rófa
gos
P T
Figura 17. Relación entre numero de eosinofilos y macrofagos y formación de absceso hepatico amibiano. se inocularon 2 x 106 trofozoitos patógenos de Eh en hamsteres testigo y en aquellos inoculados con. Ascaris suum y a los siete dìas se sacrificaron y se cuantifico el tamaño del absceso. En exudado peritoneal se realizo un recuento de eosinófilos de estos dos grupos para correlacionarlos con el porcentaje de absceso hepático
26

B A
Figura 18.- Absceso hepático y eosinofilia.-En A observamos un hígado que pesa
7.8 g con un absceso de 2 cm aproximadamente, corresponde a un hamster que se le inoculo intraperitonealmente 2,000,000 de trofozoitos de Entamoeba histolytica de la cepa H-1IMSS. En B observamos un hígado que pesa 6.1g practicamente sin absceso, corresponde a un hamster que previo al inoculo de trofozoitos de Entamoeba histolytica se inmunoestimulo intraperitonealmente con proteínas de Ascaris summ.
Purificación de macrofagos y eosinofilos a partir de celulas de exudado peritoneal empleando gradientes de Percoll. Las poblaciones del exudado peritoneal se separaron células mediante
gradientes de Percoll. A partir de las células de exudado peritoneal de animales
inoculados con aceite mineral se obtuvieron dos poblaciones la de la interfase
30/60 (G I) y la de la interfase 60/80% (G II) En la primera población (G I) la
viabilidad celular fue de 85.8 %; los porcentajes de eosinófilos y macrofagos
fueron de 30.3 y 30.6 % respectivamente; en la segunda población (G II) la
viabilidad celular fue del 85.6 %, los porcentajes de eosinòfilos y macrofagos
27

fueron de 35.5 y 21.9 respectivamente; en el grupo testigo el porcentaje de
eosinòfilos y de macrófagos fue 8 % y 20.5 % respectivamente.
Tambien a partir de las células de exudado peritoneal de animales inoculados
con Ascaris suum se obtuvieron dos poblaciones la de la interfase 30/60 (G I) y
la de la interfase 60/80% (G II). En la primera población (G I) la viabilidad
celular fue de 88.25 % , los porcentajes de eosinófilos y macrofagos fueron de
41.5 y 36.2 % respectivamente. En la segunda población (G II) la viabilidad
celular fue del 90 % y los porcentajes de eosinòfilos y macrofagos fueron de
44.5 y 25.5 % respectivamente. En el grupo testigo el porcentaje de eosinòfilos
y de macrófagos fue 5.5 y 20.5 % respectivamente.
En resumen, al realizar la técnica de gradientes de Percoll, se obtuvieron dos
poblaciones celulares en las interfases 30-60% ( GI ) y a 60–80% ( G II ), en las
cuales no se encontró diferencia significativa en el numero de eosinòfilos y
macrofagos como se esperaba, ya que se menciona que la mayor cantidad de
eosinòfilos debería de localizarse en la población celular del gradiente de
Percoll II correspondiente a 60-80%. Es importante mencionar que no se logro
obtener una población pura de eosinòfilos, por lo que los resultados obtenidos
no podemos asegurar que correspondan en su totalidad a la acción de los
mismos.
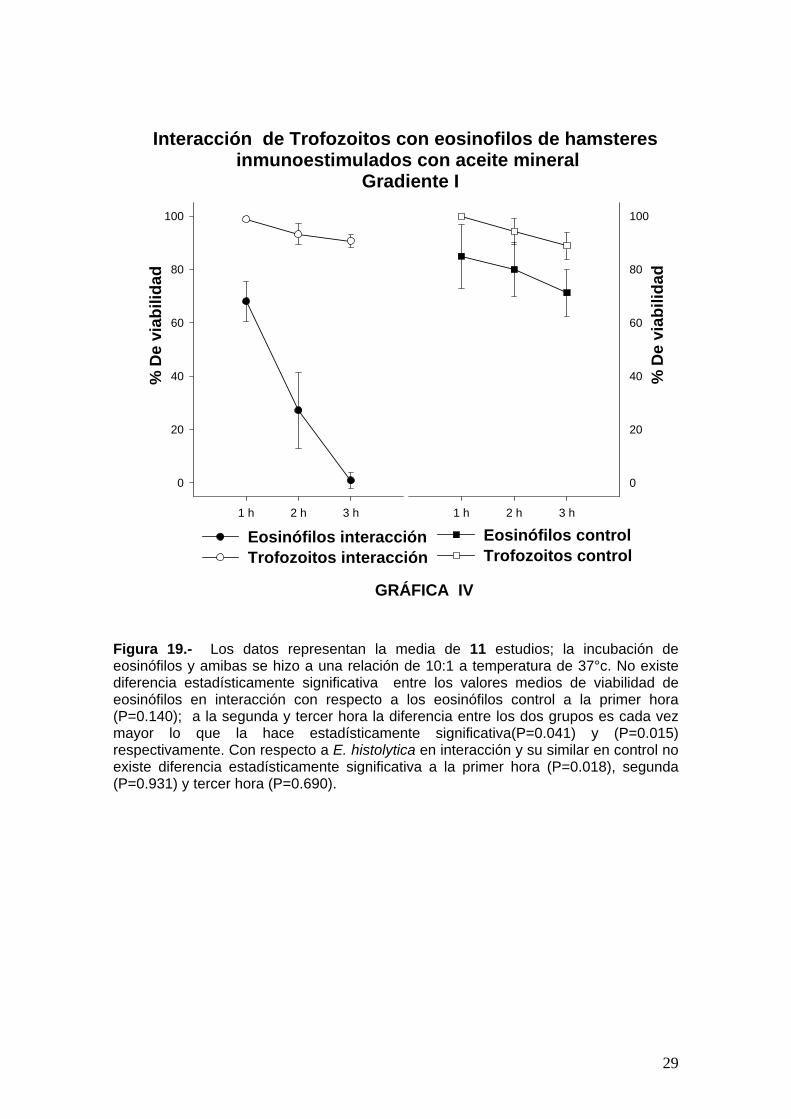
Interacciones in vitro de trofozoitos de Entamoeba histolytica células del exudado peritoneal (eosinófilos y macrófagos). Al interactuar in vitro trofozoitos patógenos de Entamoeba Histolytica con las
dos poblaciones celulares de Percoll, formadas principalmente de eosinòfilos y
macrofagos, estimuladas una con aceite mineral y otra con Ascaris summ, se
pudo observar que la viabilidad de los trofozoitos de Entamoeba histolytica
prácticamente no se modifica en comparación con la del grupo testigo, mientras
que la viabilidad de las células de las poblaciones de GI y GII, va disminuyendo
progresivamente en el tiempo (Figuras ). A travès del microscopio óptico se
observa que los trofozoitos van fagocitando a las mismas. Por lo anterior se
infiere que in vitro los eosinòfilos no poseen capacidad por si mismos para
lesionar o eliminar a Eh.
28
Interacción de Trofozoitos con eosinofilos de hamsteres inmunoestimulados con aceite mineral
Gradiente I
GRÁFICA IV
1 h 2 h 3 h
% D
e vi
abili
dad
0
20
40
60
80
100
Eosinófilos interacción Trofozoitos interacción
1 h 2 h 3 h
% D
e vi
abili
dad
0
20
40
60
80
100
Eosinófilos control Trofozoitos control
Figura 19.- Los datos representan la media de 11 estudios; la incubación de eosinófilos y amibas se hizo a una relación de 10:1 a temperatura de 37°c. No existe diferencia estadísticamente significativa entre los valores medios de viabilidad de eosinófilos en interacción con respecto a los eosinófilos control a la primer hora (P=0.140); a la segunda y tercer hora la diferencia entre los dos grupos es cada vez mayor lo que la hace estadísticamente significativa(P=0.041) y (P=0.015) respectivamente. Con respecto a E. histolytica en interacción y su similar en control no existe diferencia estadísticamente significativa a la primer hora (P=0.018), segunda (P=0.931) y tercer hora (P=0.690).
29
Interacción trofozoitos con eosinófilos de hamsteresinmunoestimulados con aceite mineral
Gradiente II
GRÁFICA V
1 h 2 h 3 h
% D
e vi
abili
dad
0
20
40
60
80
100
Eosinófilos interacción Trofozoitos interacción
1 h 2 h 3 h
% D
e vi
abili
dad
0
20
40
60
80
100
Eosinófilos control Trofozoitos control
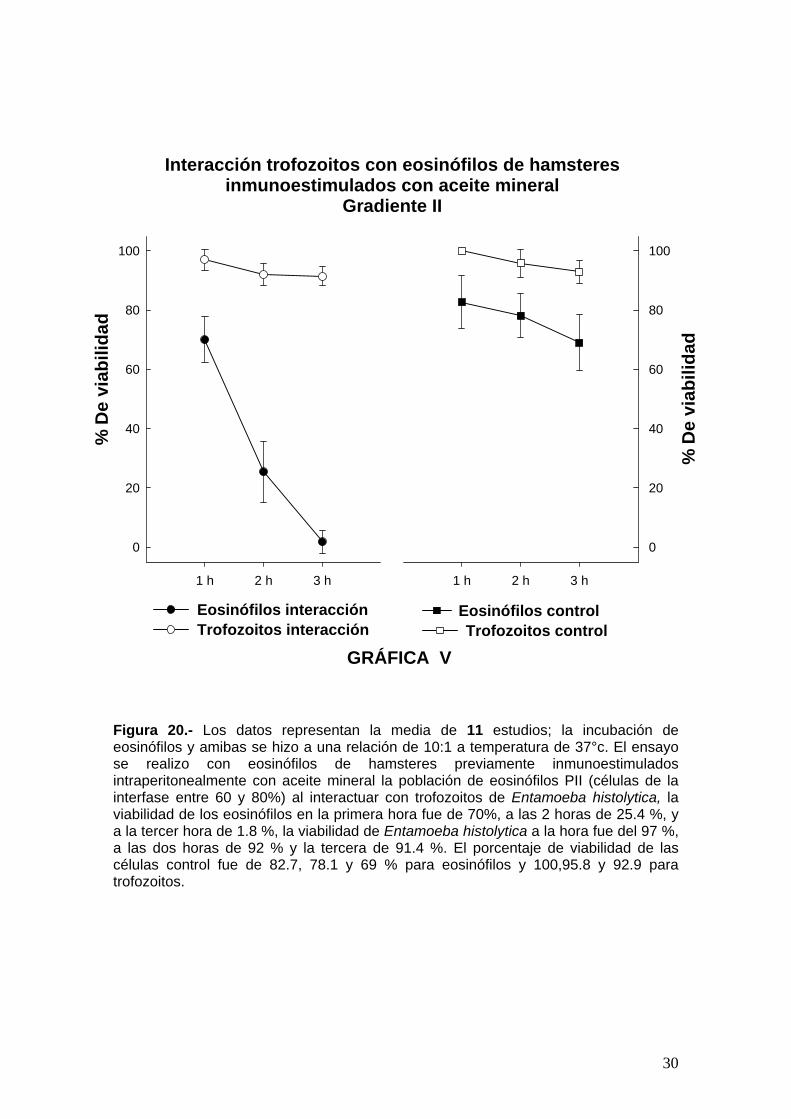
Figura 20.- Los datos representan la media de 11 estudios; la incubación de eosinófilos y amibas se hizo a una relación de 10:1 a temperatura de 37°c. El ensayo se realizo con eosinófilos de hamsteres previamente inmunoestimulados intraperitonealmente con aceite mineral la población de eosinófilos PII (células de la interfase entre 60 y 80%) al interactuar con trofozoitos de Entamoeba histolytica, la viabilidad de los eosinófilos en la primera hora fue de 70%, a las 2 horas de 25.4 %, y a la tercer hora de 1.8 %, la viabilidad de Entamoeba histolytica a la hora fue del 97 %, a las dos horas de 92 % y la tercera de 91.4 %. El porcentaje de viabilidad de las células control fue de 82.7, 78.1 y 69 % para eosinófilos y 100,95.8 y 92.9 para trofozoitos.
30
Interacción in vitro de trofozoitos con eosinófilos de hamsteres inmunoestimulados con A suum
Gradiente II
GRÁFICA VII
1 h 2 h 3 h
% D
e vi
abili
dad
0
20
40
60
80
100
Eosinófilos interacción Trofozoitos interacción
1 h 2 h 3 h
% D
e vi
abili
dad
0
20
40
60
80
100
Eosinófilos control Trofozoitos control
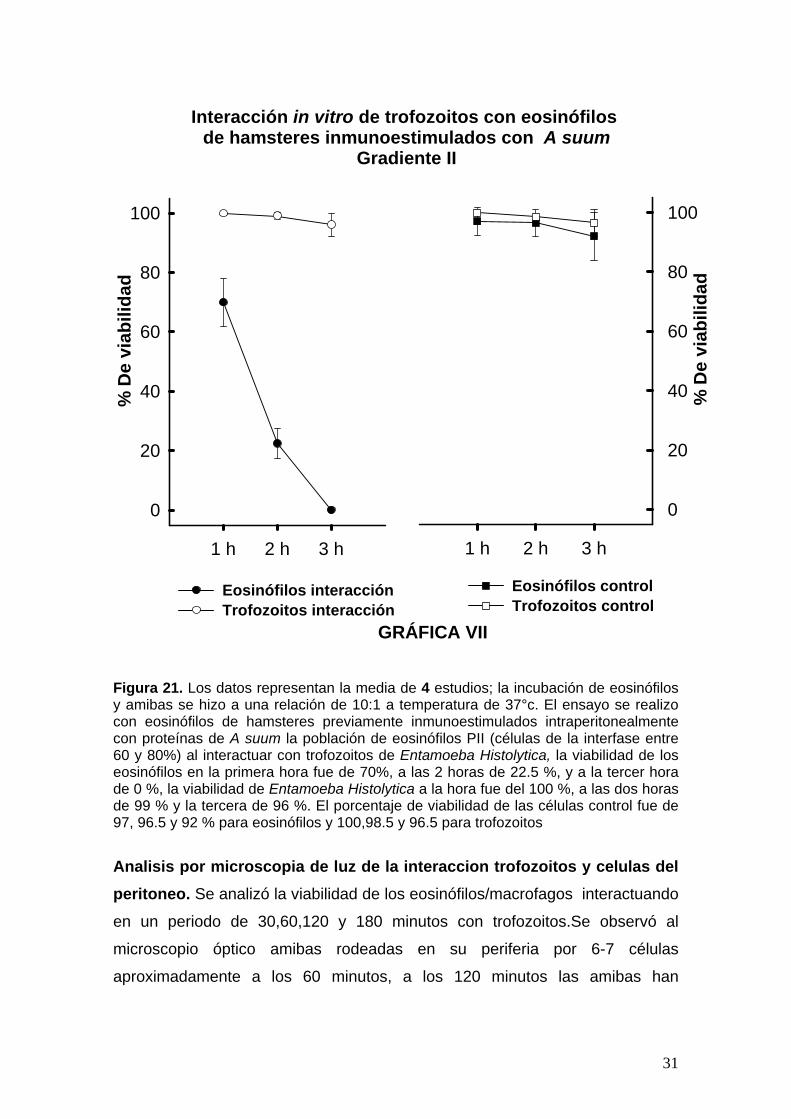
Figura 21. Los datos representan la media de 4 estudios; la incubación de eosinófilos y amibas se hizo a una relación de 10:1 a temperatura de 37°c. El ensayo se realizo con eosinófilos de hamsteres previamente inmunoestimulados intraperitonealmente con proteínas de A suum la población de eosinófilos PII (células de la interfase entre 60 y 80%) al interactuar con trofozoitos de Entamoeba Histolytica, la viabilidad de los eosinófilos en la primera hora fue de 70%, a las 2 horas de 22.5 %, y a la tercer hora de 0 %, la viabilidad de Entamoeba Histolytica a la hora fue del 100 %, a las dos horas de 99 % y la tercera de 96 %. El porcentaje de viabilidad de las células control fue de 97, 96.5 y 92 % para eosinófilos y 100,98.5 y 96.5 para trofozoitos
Analisis por microscopia de luz de la interaccion trofozoitos y celulas del peritoneo. Se analizó la viabilidad de los eosinófilos/macrofagos interactuando
en un periodo de 30,60,120 y 180 minutos con trofozoitos.Se observó al
microscopio óptico amibas rodeadas en su periferia por 6-7 células
aproximadamente a los 60 minutos, a los 120 minutos las amibas han
31
fagocitado aproximadamente de 6-9 eosinófilos de 10 y a los 180 minutos todos
los eosinófilos han sido fagocitados. (Figuras 22, 23, 24, 25, 26, 27).
Interacción Eh / Eosinófilos Minuto 0 ( 10 X )
Trofozoitos de Entamoeba h l ( )
BA
Interacción Eh / Eosinófilos Minuto 30 ( 100 X )
DInteracción Eh / Eosinófilos
Minuto 0 ( 40 X )
C
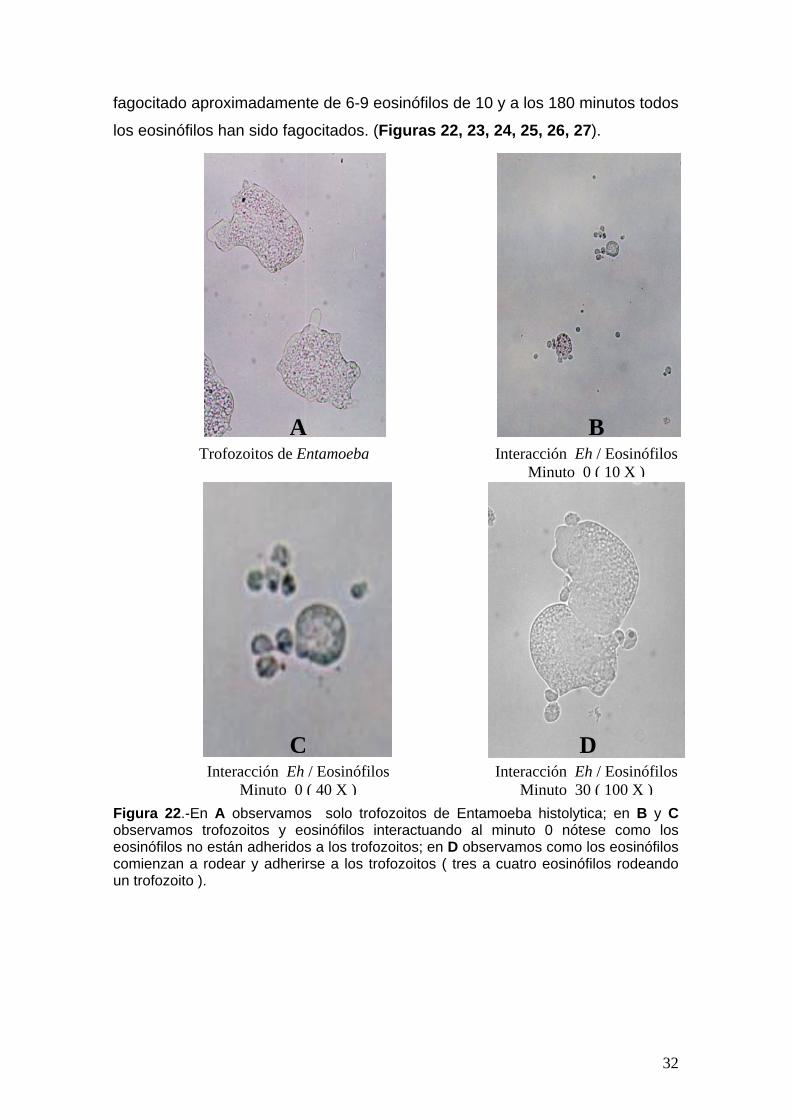
Figura 22.-En A observamos solo trofozoitos de Entamoeba histolytica; en B y C observamos trofozoitos y eosinófilos interactuando al minuto 0 nótese como los eosinófilos no están adheridos a los trofozoitos; en D observamos como los eosinófilos comienzan a rodear y adherirse a los trofozoitos ( tres a cuatro eosinófilos rodeando un trofozoito ).
32
A B
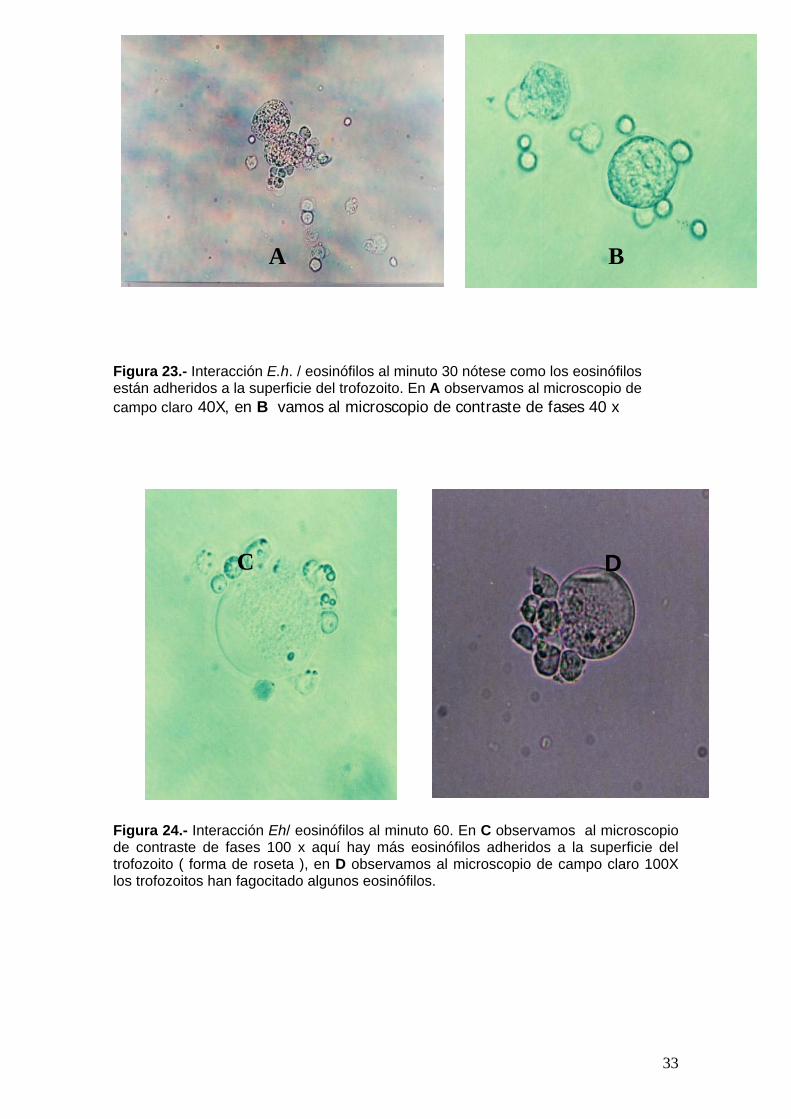
Figura 23.- Interacción E.h. / eosinófilos al minuto 30 nótese como los eosinófilos están adheridos a la superficie del trofozoito. En A observamos al microscopio de campo claro 40X, en B vamos al microscopio de contraste de fases 40 x
D
Figura 24.- Interade contraste de ftrofozoito ( forma los trofozoitos han
C
cción Eh/ eosinófilos al minuto 60. En C observamos alases 100 x aquí hay más eosinófilos adheridos a la sde roseta ), en D observamos al microscopio de campo fagocitado algunos eosinófilos.
microscopio uperficie del claro 100X
33

A B
Figura 25.-Fotomicrografías observadas al microscopio de campo claro 40 X de la interacción Eh/eosinófilos al minuto 90 , en A observamos un trofozoito que ha fagocitado varios eosinófilos; en B se aplico la tinción con cristal violeta para corroborar la presencia de los eosinófilos interactuando con los trofozoitos.
E C D Figura 26.- Fotomicrografías observadas al microscopio de campo claro 40 X de la interacción Eh/eosinófilos al minuto 120 , en C,D y E observamos un trofozoito que ha fagocitado casi el total de los eosinófilos. C se tiño con cristal violeta y vemos como el colorante entro al trofozoito probablemente en el momento en que esta fagocitando.
34
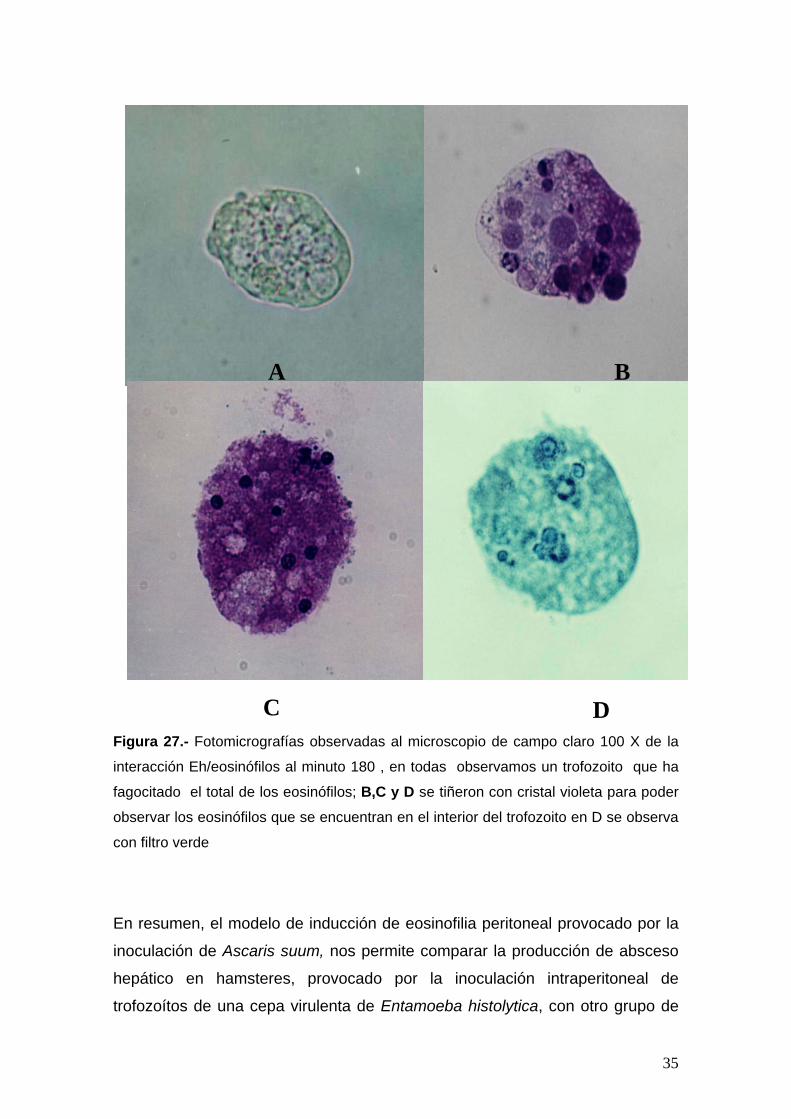
B
Figura 27.- Fotomicrog
interacción Eh/eosinófi
fagocitado el total de
observar los eosinófilo
con filtro verde
En resumen, el mode
inoculación de Ascar
hepático en hamst
trofozoítos de una ce
A
rafías observadas al microscopio de campo claro 1
los al minuto 180 , en todas observamos un trofozo
los eosinófilos; B,C y D se tiñeron con cristal violeta
s que se encuentran en el interior del trofozoito en D
lo de inducción de eosinofilia peritoneal provoc
is suum, nos permite comparar la producción d
eres, provocado por la inoculación intraperi
pa virulenta de Entamoeba histolytica, con otro
C D 00 X de laito que ha
para poder
se observa
ado por la
e absceso
toneal de
grupo de
35

hamsteres sin inducción de eosinofilia, lo cual revelo que, probablemente los
eosinofilos jueguen un papel importante en la destrucción de Eh, ya que
aquellos animales con eosinofilia peritoneal presentaron disminución
considerable en la fomaciòn de absceso en comparación con los no inducidos.
Cabe mencionar que al ser un ensayo in vivo, no se descarta la participación e
interacción de otras células y substancias liberadas por el organismo.
Si comparamos el resultado de los ensayos in vivo e in vitro, podemos decir
que en los primeros si existe protección contra la invasión hepática amibiana,
pero en la segunda no se presenta este proceso en contra de Eh, por lo que
podemos referir que existen factores externos al eosinòfilo que actúan
directamente contra Eh o que de alguna manera estimulan la producción de
substancias del mismo eosinòfilo tales como, la proteína básica mayor, la
neurotoxina derivada de eosinòfilo, la proteína catiònica eosinofìlica,la proteína
formadora de cristales de Charcot-Leyden y la peroxidasa eosinofilica, las
cuales actùan a distancia y son potentes toxinas para larvas de helmintos,
protozoarios, células humanas y que pudieran provocar la muerte de los
trofozoitos y con esto impedir la formación del absceso.
36

Bibliografía
Black S, Agrawal A, Samols D. The phosphocholine and the polycation-binding sites on rabbit C-reactive protein are structurally and functionally distinct. Mol Immunol. 2003 39:1045-1054
Black S, Kushner I, Samols D. C-reactive Protein. J Biol Chem. 2004 279: 48487-48490
Coghlan VM. Analysis of protein:protein interactions by gel electrophoresis. En Gel Electrophoresis of proteins. Ed por BD Hames. 3 Ed. Oxford University Pres. Oxford, pp 293-311, 1998
Diamond, L. S., Harlow, D. R. and Cunnick, C. C. A new medium for axenic cultivation of Entamoeba histolytica and other Entamoeba. Transactions of the Royal Society of Tropical Medicine and Hygiene, 1978, 72, 431-432.
Hack CE, Wolbink GJ, Schalkwijk C, Speijer H, Hermens WT, van den Bosch H.. A role for secretory phospholipase A2 and C-reactive protein in the removal of injured cells Immunol Today. 1997 18:111-115. Haque R, Duggal P, Ali IM, Hossain MB, Mondal D, Sack RB, Farr BM, Beaty TH, Petri WA Jr. Innate and acquired resistance to amebiasis in bangladeshi children. J Infect Dis, 2002; 186: 547-552. Hoffmann JA, Kafatos FC, Janeway CA, Ezekowitz RA. Phylogenetic
perspectives in innate immunity. Science, 1999; 284:1313-1318.
Laemmli, U K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriopahge T4, Nature, 1970, 227, 680-685.
Pacheco-Yépez J, Campos–Rodríguez R, Shibayama M, Ventura-Juárez J, Serrano-Luna J, Tsutsumi V Entamoeba histolytica: production of nitric oxide and in situ activity of NADPH diaphorase in amebic liver abscess of hamsters. Parasitol Res, 2001, 87: 49-56. Shibayama M, Campos-Rodriguez R, Ramirez C, Pacheco-Yepez J, Tsutsumi V. Studies on the natural immunity in hamsters using the intraperitoneal model of amebic liver abscess. Arch Med Res, 2000; 31(4 Suppl):S78-80.
Szalai AJ, Agrawal A, Greenhough TJ, Volanakis JE. C-reactive protein: structural biology and host defense function. Clin Chem Lab Med. 1999 37: 265-270.
Szalai AJ. The antimicrobial activity of C-reactive protein. Microbes Infect. 2002 4: 201-205
Towbin, H., Staehelin, T. & Gordon, J. Electrophoretic transfer of proteinns from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proceedings of the National Academy of Sciences of the USA, 1979, 76, 4350-4354
37