kvantitatiivinen genetiikka moniste s. 56 - oulu.fi · kühn: grundriss der vererbungslehre, 1950....
TRANSCRIPT

Kvantitatiivinen genetiikka moniste s. 56
- määrällisten ominaisuuksien periytymisen hallinta
- mendelismi oli aluksi vastatuulessa siksi ettädarwinistit, joilla oli paljon valtaa Britanniassa, olivat tottuneet määrällisen genetiikan piirteisiin: jatkuvat jakautumat, vähittäinen muutos jne
- oli myös kehitetty BIOMETRIA (eli tilastotiede), jonka toimialaa määrällisten ominaisuuksien genetiikka on vieläkin (normaali- ym. jakautumat, korrelaatiot, regressiot )
PITUUDEN MUUNTELU JA HERITABILITEETTI IHMISPOPULAATIOSSA

Sulkeisliike “normaalijakautumaan järjesty”

Joitakin määrällisten ominaisuuksien hallinnan juttuja
Keskiarvo
Varianssi
Keskihajonta
Korrelaatio
Regressio

Nilsson-Ehle (ruotsalainen) esitti ensimmäisenämendelistisen mallin määrälliselle ominaisuudelle 1908
Tutkittavana hällä oli vehnänjyvän punertava väri

Kühn: Grundriss der Vererbungslehre, 1950

Kvantitatiivisen periytymisen malli
aa Aa AA|---------------------------|------|---------------------| -a 0 +d +a
q2 2pq p2
m = p2a - q2 a + 2pqd = a (p - q)(p + q) + 2pqd = a(p-q) +2pqd
Jos d = 0, m = a (1 - 2q)jos d = 1, m = a (1 - 2 q2 )

Kahden alleeliparin vaikutuksen yhdistäminen
AA Aa aap2 2pq q2
BB r2 2a a + d 0Bb 2rs a + d 2d d - abb s2 0 d - a -2a

Pyyhitäänpä dominanssi pois:
AA Aa aap2 2pq q2
BB r2 2a a 0Bb 2rs a 0 -abb s2 0 -a -2a
aaBBAABb AAbb Aabb
AABB AaBB AaBb aaBb aabbp2 r2 2rsp2+2pqr2 q2r2 +p2s2 +4pqrs 2rsq2+2pqs2 q2 s2
2a a 0 -a -2a
1 4 6 4 1

Pascalin kolmio eli binomijakautuma -taika
1 1 1
1 2 1 1 alleelipari1 3 3 1
1 4 6 4 1 2 paria1 5 10 10 5 1
1 6 15 20 15 6 1 31 7 21 35 35 21 7 1
1 8 28 56 70 56 28 8 1 4, &c1 9 36 84 126 126 84 36 9 1
Tästä saa johdettua lukusuhteet 1, 2, 3, 4 tai vaikka kuinka monen alleeliparin jakautumille, helposti ja paljoakaan muistamatta. Lopulta jakautuma noudattaa normaalijakatumaa

Esimerkki: vanha kunnon käärmelasku
Risteytetään 200 cm pitkät käärmeetA B C d E A B c d E-------------- x --------------a b C d e a b C d E
Peruspituus 150 cm. Jokainen isolla kirjaimella merkitty alleelilisää pituutta 10 cm. Ennusta jälkeläisen jakautuma pituusluokkiin.
Ratkaisu: heterotsygoottisia lokuksia on kuusi, eli niistä voi tulla jälkeläiselle “pieni tai iso kirjain”. Jokainen jälkeläinen saa varmasti yhden C:n ja yhden E:n, joten pienin poikanen on 170 cm
170 180 190 200 210 220 230 Pituusluokka1 6 15 20 15 6 1 Pascal, rivi 7

Määrällisiin ominaisuuksiin vaikuttaa myös ympäristö, eikäsuinkaan pelkästään 2 tai useampia +/- alleelipareja.
Usein halutaankin tietää, kuinka iso osa varianssista eli yksilöiden välisistä eroista johtuu geneettisistä syistä, ja mikä osa ympäristöstä
Geneettisen osuuden mitan nimi on heritabiliteetti

Jos populaatiossa on sellaisia segregoituvia alleelipareja, jotka vaikuttavat tiettyyn mittaominaisuuteen, siitä koituu tietenkin geneettistä muuntelua
Muuntelua aiheuttaa myös ympäristö

Fenotyyppinen varianssi Vp aiheutuu eri osista:
additiivisesta geneettisestä Va, dominanssin aiheuttamasta geneettisestä Vd, geeni-interaktioiden Vi aiheuttamasta geneettisestä ja vieläympäristön Ve aiheuttamasta, sekä lopulta kaikkien näiden mahdollisista interaktioista eli vuorovaikutuksista.
Vp = Va + Vd + Vi + Ve
varianssi

additiivinen aa / AA dominanssi Aa /0aa / (Aa+AA)

Heritabiliteetti
Yksilöt antavat alleeleistaan näytteen gameettien kautta seuraavaan sukupolveen. Sellainen yksilö, jolla on monta plus-alleelia, antaa tietysti gameetteihinkin useampia plus-alleeleita kuin sellainen, jolla niitä on vähän. Tätä tosiasiaa käytetään hyväksi kvantitatiivisessa genetiikassa. Vanhempien ja jälkeläisten tai sisarusten välillä on tiettyäkorrelaatiota näiden plus-alleelien määrässä.
SUKULAISTEN VÄLINEN KORRELAATIO JOHTUU GEENEISTÄ


Kurssityönä tutkitaan, onko kurssilaisten ja heidän isiensä ja/tai äitiensä välillä korrelaatiota pituuden suhteen, ja esitetään, kuinka tästä kovarianssista voidaan laskea heritabiliteetti.
Heritabiliteetti ahtaassa mielessä h² määritellään additiivisen geneettisen varianssin Va osuudeksi koko fenotyyppisestävarianssista Vp : h² = Va /Vp
Tämä on tärkeä estimaatti koska jalostugenetiikka hyötyy vain ahtaasta heritabiliteetista

Heritabiliteettiestimaatteja

Jalostushyödyn ja heritabiliteetin välillä on seuraava helppo yhteys:
G = h²I
G on jalostushyöty (Gain), h² on additiivinen heritabiliteetti ja I on valinnan intensiteetti, so. se osuus populaatiosta joka karsitaan pois. Tämä johtuu hyvin yksinkertaisesti siitä, ettäjos "suuruus" johtuu tietyistä alleeleista, on niitä alleeleita "suurten" jälkeläisissä sitten enemmän kuin keskinkertaisten jälkeläisissä.

Aina kun ihminen tarvitsee tämän alan kertausta, sen pitääetsiä käsiinsä Falconer, vanha tai uusi

Genetiikan peruskurssi 2003
Descriptives
Descriptive Statistics
44 156 183 165.34 5.51943 150 178 163.37 6.05543 166 194 176.14 5.99814 174 189 180.57 5.18514 155 174 163.79 5.36614 168 190 178.71 6.51013
TYTÄRÄITI(T)ISÄ(T)POIKAÄITI(P)ISÄ(P)Valid N (listwise)
N Minimum Maximum Mean Std. Deviation
Correlations between parents in female (N=43) and male (N=14) populations
Correlations
1 .391**. .009
43 43.391** 1.009 .
43 43
Pearson CorrelationSig. (2-tailed)NPearson CorrelationSig. (2-tailed)N
ÄITI(T)
ISÄ(T)
ÄITI(T) ISÄ(T)
Correlation is significant at the 0.01 level(2 il d)
**.
Correlations
1 .579*. .030
14 14.579* 1.030 .
14 14
Pearson CorrelationSig. (2-tailed)NPearson CorrelationSig. (2-tailed)N
ÄITI(P)
ISÄ(P)
ÄITI(P) ISÄ(P)
Correlation is significant at the 0.05 level (2-tailed).*.
Genetiikan peruskurssi 2003
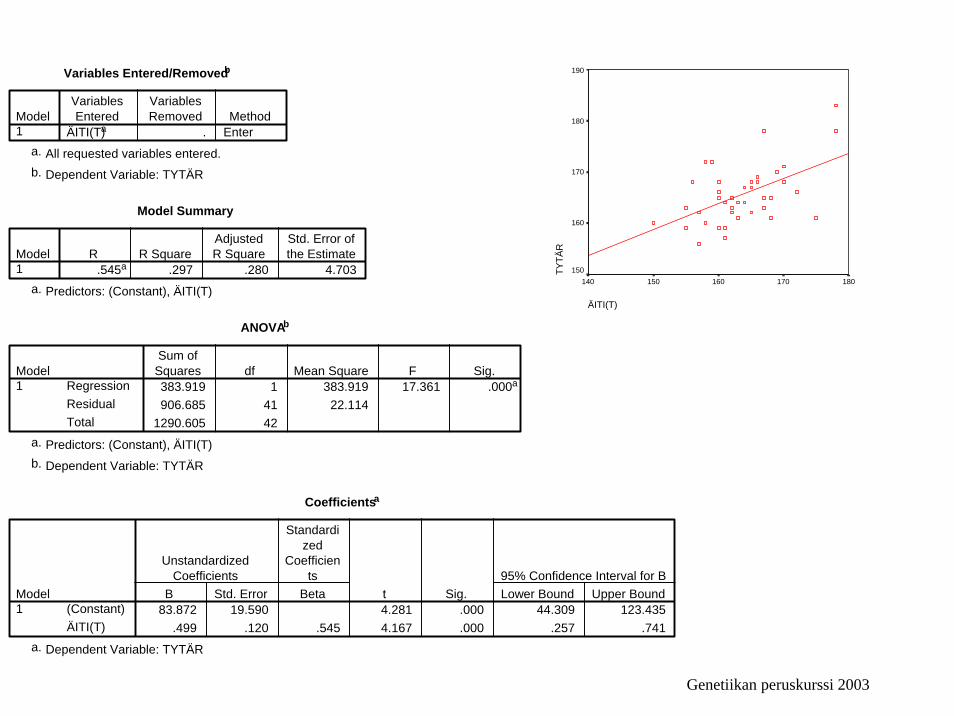
Variables Entered/Removedb
ÄITI(T)a . EnterModel1
VariablesEntered
VariablesRemoved Method
All requested variables entered.a.
Dependent Variable: TYTÄRb.
Model Summary
.545a .297 .280 4.703Model1
R R SquareAdjustedR Square
Std. Error ofthe Estimate
Predictors: (Constant), ÄITI(T)a.
ANOVAb
383.919 1 383.919 17.361 .000a
906.685 41 22.1141290.605 42
RegressionResidualTotal
Model1
Sum ofSquares df Mean Square F Sig.
Predictors: (Constant), ÄITI(T)a.
Dependent Variable: TYTÄRb.
Coefficientsa
83.872 19.590 4.281 .000 44.309 123.435.499 .120 .545 4.167 .000 .257 .741
(Constant)ÄITI(T)
Model1
B Std. Error
UnstandardizedCoefficients
Beta
Standardized
Coefficients
t Sig. Lower Bound Upper Bound95% Confidence Interval for B
Dependent Variable: TYTÄRa.
ÄITI(T)
180170160150140
TYTÄ
R
190
180
170
160
150

Genetiikan peruskurssi 2003
Variables Entered/Removedb
ISÄ(P)a . EnterModel1
VariablesEntered
VariablesRemoved Method
All requested variables entered.a.
Dependent Variable: POIKAb.
Model Summary
.612a .374 .322 4.270Model1
R R SquareAdjustedR Square
Std. Error ofthe Estimate
Predictors: (Constant), ISÄ(P)a.
ANOVAb
130.664 1 130.664 7.167 .020a
218.765 12 18.230349.429 13
RegressionResidualTotal
Model1
Sum ofSquares df Mean Square F Sig.
Predictors: (Constant), ISÄ(P)a.
Dependent Variable: POIKAb.
Coefficientsa
93.532 32.532 2.875 .014 22.651 164.412.487 .182 .612 2.677 .020 .091 .883
(Constant)ISÄ(P)
Model1
B Std. Error
UnstandardizedCoefficients
Beta
Standardized
Coefficients
t Sig. Lower Bound Upper Bound95% Confidence Interval for B
Dependent Variable: POIKAa.
ISÄ(P)
200190180170160
POIK
A
190
188
186
184
182
180
178
176
174
172

Genetiikan peruskurssi 2003
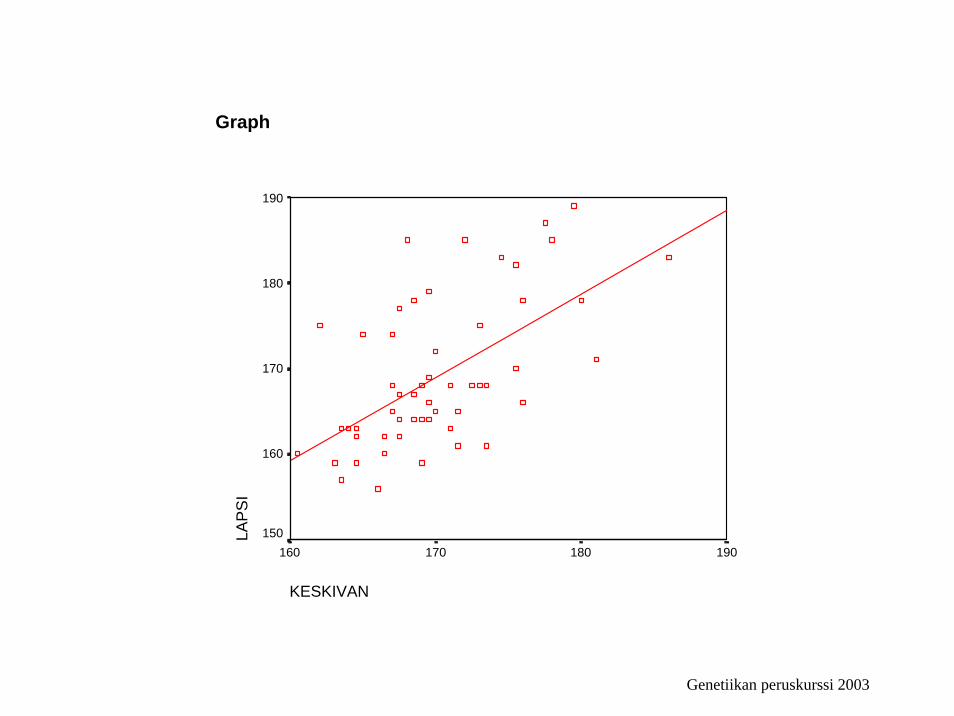
Regression
Variables Entered/Removedb
KESKIVANa . EnterModel1
VariablesEntered
VariablesRemoved Method
All requested variables entered.a.
Dependent Variable: LAPSIb.
Model Summary
.580a .336 .324 6.99892Model1
R R SquareAdjustedR Square
Std. Error ofthe Estimate
Predictors: (Constant), KESKIVANa.
ANOVAb
1363.411 1 1363.411 27.833 .000a
2694.168 55 48.9854057.579 56
RegressionResidualTotal
Model1
Sum ofSquares df Mean Square F Sig.
Predictors: (Constant), KESKIVANa.
Dependent Variable: LAPSIb.
Coefficientsa
4.058 31.308 .130 .897 -58.684 66.801.970 .184 .580 5.276 .000 .602 1.339
(Constant)KESKIVAN
Model1
B Std. Error
UnstandardizedCoefficients
Beta
Standardized
Coefficients
t Sig. Lower Bound Upper Bound95% Confidence Interval for B
Dependent Variable: LAPSIa.
KESKIVAN
190180170160
LAP
SI
190
180
170
160
150

Tulokset vuoden 2001 syksyn peruskurssin oppilaista:pituuden heritabiliteetti
Genetiikan peruskurssi 2001 syksy

Isän pituus160 165 170 175 180 185 190 195
idin
pitu
us
145
150
155
160
165
170
175
180
Isien ja äitien pituus ei ole merkitsevästi korreloitunut (P = 0.08), korrelaatiokerroin on .273 ja sen neliö eli “selityskerroin” vain 7.4 %
Genetiikan peruskurssi 2001 syksy

Genetiikan peruskurssi 2001 syksy
Correlations
.273
.08042
Pearson CorrelationSig. (2-tailed)NPearson CorrelationSig. (2-tailed)N
ISÄ
ÄITI
ISÄ ÄITI

Äidin pituus145 150 155 160 165 170 175 180
Tyttä
ren
pitu
us
150
155
160
165
170
175
180
185
Isän pituus160 165 170 175 180 185 190
Poj
an p
ituus
165
170
175
180
185
190
Genetiikan peruskurssi 2001 syksy
Jälkeläisen ja samaa sukupuolta olevan vanhemman regressiot
Huomaa, että asteikot eivät ole identtiset
Testitulokset seuraavassa slidessä

Genetiikan peruskurssi 2001 syksy
Model Summaryb
.692a .479 .404 4.9889Model1
R R SquareAdjusted R
Square
Std. Errorof the
Estimate
Predictors: (Constant), ISÄa.
Dependent Variable: HENKILÖb.
Coefficientsa
59.103 46.353 1.275 .243.665 .262 .692 2.535 .039
(Constant)ISÄ
Model1
B Std. Error
UnstandardizedCoefficients
Beta
Standardized
Coefficients
t Sig.
Dependent Variable: HENKILÖa.
Model Summary
.662a .438 .420 4.6983Model1
R R SquareAdjusted R
Square
Std. Errorof the
Estimate
Predictors: (Constant), ÄITIa.
Coefficientsa
52.153 22.815 2.286 .029.691 .141 .662 4.915 .000
(Constant)ÄITI
Model1
B Std. Error
UnstandardizedCoefficients
Beta
Standardized
Coefficients
t Sig.
Dependent Variable: HENKILÖa.
Pojat vs. isät
Tyttäret vs. äidit

Vanhempien keskipituus158 160 162 164 166 168 170 172 174 176 178 180
Tyttä
ren
pitu
us
150
155
160
165
170
175
180
185
Vanhempien keskipituus158 160 162 164 166 168 170 172 174 176 178 180
Poj
an p
ituus
165
170
175
180
185
190
Genetiikan peruskurssi 2001 syksy
Jälkeläisen ja vanhempien keskiarvon regressio
Vain Y-akselit ovat erilaiset
Analyysi seuraavalla sivulla

Coefficientsa
2.803 29.088 .096 .924.956 .172 .706 5.551 .000
(Constant)MLENGTH
Model1
B Std. Error
UnstandardizedCoefficients
Beta
Standardized
Coefficients
t Sig.
Dependent Variable: HENKILÖa.
Model Summary
.706a .498 .482 4.4381Model1
R R SquareAdjusted R
Square
Std. Errorof the
Estimate
Predictors: (Constant), MLENGTHa.
Genetiikan peruskurssi 2001 syksy
Model Summary
.768a .590 .532 4.4220Model1
R R SquareAdjusted R
Square
Std. Errorof the
Estimate
Predictors: (Constant), MLENGTHa.
Coefficientsa
19.352 49.506 .391 .707.930 .293 .768 3.177 .016
(Constant)MLENGTH
Model1
B Std. Error
UnstandardizedCoefficients
Beta
Standardized
Coefficients
t Sig.
Dependent Variable: HENKILÖa.
Tytöt vs. vanhempien keskiarvo
Pojat vs. vanhempien keskiarvo

Äidin pituus145 150 155 160 165 170 175 180
Tyttä
ren
pitu
us
150
155
160
165
170
175
180
185
Isän pituus160 165 170 175 180 185 190
Poj
an p
ituus
165
170
175
180
185
190
Vanhempien keskipituus158 160 162 164 166 168 170 172 174 176 178 180
Tyttä
ren
pitu
us
150
155
160
165
170
175
180
185
Vanhempien keskipituus158 160 162 164 166 168 170 172 174 176 178 180
Poj
an p
ituus
165
170
175
180
185
190
Genetiikan peruskurssi 2001 syksy

Genetiikan peruskurssi 2001 syksy
Heritabiliteettiestimaatit olivat aika korkeita!
Yhteisen ympäristön vaikutus on vaikea kitkeä pois

Genetiikan peruskurssi 2003
ÄITI(T)
180170160150140
TYTÄ
R190
180
170
160
150

Genetiikan peruskurssi 2003
Graph
ISÄ(P)
200190180170160
PO
IKA
190
188
186
184
182
180
178
176
174
172
Genetiikan peruskurssi 2003
Graph
KESKIVAN
190180170160
LAP
SI
190
180
170
160
150