m4:01634 revised gem gtpase and tau: morphological changes
TRANSCRIPT

M4:01634 Revised
Gem GTPase and tau: morphological changes induced by Gem GTPase in CHO cells
are antagonized by tau
Fumitaka Oyama1,3, Svetlana Kotliarova3, Akihiro Harada2,4, Mamoru Ito5, Haruko
Miyazaki3, Yoshito Ueyama5,6, Nobutaka Hirokawa2, Nobuyuki Nukina3, and Yasuo
Ihara1*
1Department of Neuropathology and 2Department of Cell Biology and Anatomy,
Faculty of Medicine, University of Tokyo, Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo,
Japan; 3Laboratory for Structural Neuropathology, Brain Science Institute, RIKEN,
Wako-shi, Saitama, Japan; 4Laboratory of Cellular and Molecular Morphology,
Institute for Molecular and Cellular Regulation, Gunma University, 3-39-15 Showa-
machi, Maebashi, Gunma, Japan; 5Central Institute for Experimental Animals,
Kawasaki, Kanagawa, Japan; 6Department of Pathology, School of Medicine, Tokai
University, Isehara, Kanagawa, Japan
Running Title: Gem GTPase and tau
Correspondence to Yasuo Ihara, M.D.
Department of Neuropathology, Faculty of Medicine, University of Tokyo, 7-3-1
Hongo, Bunkyo-ku, Tokyo 113-0033, Japan
TEL: +81-3-5841-3541; FAX: +81-3-5800-6852; E-mail: [email protected]
A series of observations have indicated that tau, one of the major microtubule-
associated proteins, is involved in neuronal cell morphogenesis and axonal maintenance.
Tau is also the major component of paired helical filaments found in the brain affected
1
JBC Papers in Press. Published on April 15, 2004 as Manuscript M401634200
Copyright 2004 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

by Alzheimer’s disease. To explore an as yet unidentified role of tau in vivo, ~11,000
mRNAs were profiled from tau-deficient mouse brains, and compared with those from
control brains at the same ages. The expression of Gem GTPase, a small GTP-binding
protein of the ras superfamily, was significantly increased in the brain of tau-deficient
mice at eight weeks of age. Because Gem GTPase is a negative regulator of the Rho-
Rho kinase pathway for cytoskeletal organization, this protein was transiently
overexpressed in CHO cells that do not express tau. Overexpression of Gem GTPase
induced a marked elongation of CHO cells, and simultaneous expression of tau
eliminated this effect, although tau did not bind directly to Gem GTPase. This anti-
elongation activity of tau was attributed to its microtubule (MT)-binding domain, and
homologous domains of MAP2 and MAP4 exhibited similar antagonistic activities.
Taken together, the present results indicate that the level of Gem GTPase and its cell
elongation activity are modulated by tau, and suggest that tau may be involved in a Gem
GTPase-mediated signal transduction pathway.
2
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

Tau, a microtubule-associated protein (MAP) that is predominantly expressed in
neurons, promotes tubulin assembly into microtubules (MTs) and stabilizes them in
vitro, and presumably also in vivo. It is mainly localized in axons and forms short
cross-bridges between individual MTs (1-4), and has been thought to play an important
role in maintaining neuronal morphology (5-7). However, the central nervous system of
tau-deficient mice produced by gene targeting appeared to be normal, although the
number of MTs was significantly reduced in some small caliber axons (8). In addition,
according to one report (9), axonal elongation of the neurons cultured from tau-
deficient brains is not affected.
The hyperphosphorylated form of tau is the major constituent of paired helical
filaments (PHFs), unusual fibrils that accumulate in subsets of neurons in brains
affected by Alzheimer’s disease (AD) (10). PHF-like fibrils are also one of the striking
characteristics of frontotemporal dementia and parkinsonism linked to chromosome 17
(FTDP-17), which is caused by a number of missense and intronic mutations in the tau
gene (11-13). Because abundance of the unusual fibrils well correlates with the extent
of neuronal loss, these findings suggest that tau is involved in the cascade to neuronal
death in AD and FTDP-17.
Gem GTPase is a small GTP-binding protein of the ras superfamily, the major
function of which has remained to be determined (14,15). Overexpression of mouse
Gem GTPase induced invasive pseudohyphal growth in Saccharomyces cerevisiae (16).
Two functions of Gem GTPase have recently been reported: the inhibition of voltage-
gated calcium-channel activity and a negative regulation of the Rho-Rho kinase
pathway for cytoskeletal organization. Gem GTPase regulates the expression of α-
subunit of voltage-gated calcium channels at the plasma membrane by binding to ß-
subunit (17). Gem GTPase also regulates the cytoskeletal organization mediated by Rho
kinase (ROK) ß. Overexpression of Gem GTPase inhibits ROK-mediated stress fiber
formation, focal adhesion, and neurite retraction (18). In addition to ROKß, Gem
3
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

GTPase has been shown to bind to KIF-9, a kinesin-like protein (19), and to Gmip, a
RhoGAP-containing protein (20).
To clarify an as yet unidentified role(s) of tau, gene expression was compared
between the brains of wild-type (tau+/+) and tau-deficient (tau-/-) mice, using a high-
density oligonucleotide array. We found that the expression of Gem GTPase is
significantly increased in the brains of eight-week-old tau-deficient mice. Here, we
report that the overexpression of Gem GTPase induces elongation of CHO cells, and
that tau antagonizes this effect, the activity of which is attributable to the MT-binding
domain.
4
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

EXPERIMENTAL PROCEDURES
Animals—Male tau-deficient mice (8) that were backcrossed to C57Bl/6J mice for
seven generations and wild-type (wt) control mice were killed at one week, eight
weeks, and ten months of age. Tau-deficient mice grew normally, but displayed some
muscle weakness at ~10 weeks of age (21).
RNA extraction and array hybridization—RNA was extracted from the cerebrum
using TRIzol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer’s
recommended protocol. RNA from the cerebral cortices of five tau-deficient mice or
five age-matched wt mice was combined into one sample (~250 µg of total RNA) for
each time point. Poly (A)+RNA was isolated from the RNA samples using an Oligotex
mRNA isolation kits (Takara Bio, Kyoto, Japan). Biotinylated cRNAs were prepared
according to the Affymetrix protocol. Mu11K oligonucleotide arrays (Affymetrix, Santa
Clara, CA) were prehybridized, hybridized, washed, and stained as recommended by the
manufacturer, using a GeneChip Fluidics Station 400 (Affymetrix).
Criteria for selection of abnormally expressed genes—Analysis of the array data
was performed essentially as described previously (22). The present study was designed
to minimize potential experimental confounds as follows. First, a pooled sample was
subjected to the GeneChip analysis, and subsequently the result was confirmed by
examining individual variability. Second, each sample was measured in duplicate to
minimize false positives caused by chip hybridization variability. Four comparisons of
control vs. tau-deficient mice were performed using two hybridization replicates for
each of the pooled samples for wt and tau-deficient groups (22). Finally, some of the
findings were confirmed by quantitative RT-PCR. Using the Affymetrix algorithm (23)
and multiple analysis comparison software for assessing gene expression differences,
mRNAs that were increased or decreased in tau-deficient mice relative to wt mice in all
four comparisons (see ref 22; high stringency analysis) were identified. Default setting
5
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

in the GeneChip3.1 software was used to assign a particular mRNA to be increased,
decreased or unchanged.
Verification of altered expression by reverse transcriptase-coupled PCR (RT-
PCR)—Total RNA was extracted from the brains of eight-week-old wt and tau-deficient mice
using the TRIzol reagent. Contaminating genomic DNA was removed with RQ1
RNase-Free DNase I (Promega, Madison, WI). Quantitative RT-PCR was carried out
to confirm the altered gene expression, as described previously (24), except for the use
of [α-32P] dCTP as tracer. Primers were designed to yield ~350-500 bp cDNA
fragments. The nucleotide sequences of the primers are given in Table S1.
Complementary DNA for glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was
simultaneously amplified as a control. Each PCR cycle consisted of 1 min at 94 ÚC, 1
min at 55 ÚC, and 1 min at 72 ÚC. PCR amplification was performed for 22~24 cycles
for the mRNAs suggested as candidates by GeneChip analysis, and 17 cycles for
GAPDH mRNA. These ranges of cycling provided the exponential amplification
required for quantification of each mRNA. After amplification, the products were
separated on a polyacrylamide gel (6%). The gel was then dried and exposed to an
imaging plate, which was subjected to laser image analysis (Fujix BAS 2000; Fuji Film,
Tokyo, Japan). All values obtained were normalized against the levels of GAPDH
mRNA.
Mammalian expression vectors—Total mouse brain RNA was reverse-transcribed
using random hexamers as primers, as described elsewhere (24). Gem GTPase cDNA
was amplified by PCR from cDNAs thus prepared, using oligonucleotide primers
anchored with EcoRI (5’end) and SalI (3’end) restriction sites. The forward primer was
5’-CCCCAGGAATTCCCACTCTGAATAATGTCACCATGCG-3’, which is in frame
with the carboxyl terminal region of GFP, and the reverse primer was 5’-
GCTCGAGTCGACCTAGAGCACAGACAGGTCATGGC. PCR was carried out for
6
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

35 cycles with each consisting of 1 min at 94 ÚC, 1 min at 55 ÚC, and 4 min at 72ÚC.
The PCR products were then digested with EcoRI and SalI, and subcloned into the
expression vector pEGFP-C3 (BD Biosciences, Palo Alto, CA). The entire nucleotide
sequence of Gem GTPase cDNAs was determined by the dideoxynucleotide termination
method.
A tau-red fluorescent protein (RFP) expression construct was generated as follows.
The full-length wt and mutant human tau cDNAs (four-repeat tau cDNAs) lacking
exons 2 and 3 (0N4R), incorporated into the EcoRI and BglII sites in pCXN2, were
prepared as described previously (25). The tau-encoding sequence without stop codon
was amplified by PCR using the forward primer 5’-
GCTTCCATGGCTCTGAATAATGTCACCATG-3’ and the reverse primer 5’-
TTTTGTCGACTAGAGTACAGAGAGGTCA-3’. The PCR product was then digested
with EcoRI and XhoI, and subcloned into the EcoRI and XhoI sites of pDsRed-N1 (BD
Biosciences). Likewise, cDNAs encoding the amino-terminal half (NTH; residues 2-
220 of the 441-residue isoform; GenBank accession number NP_005901) and the
carboxyl-terminal half (CTH; residues 251-419) of tau, the carboxyl-terminal
fragment of MAP2 (CTF; residues 1668-1805 of the 1827-residiue MAP2; GenBank
accession number NP_002365) and the CTF of MAP4 (residues 1001-1136 of the
1152-residue MAP4; GenBank accession number AAA59553) were also prepared by
PCR. These cDNAs were cloned into pDsRed1-N1 in frame with the DsRed1-coding
sequence.
Transient transfection and fluorescence microscopy—CHO cells were maintained at
37 °C in 5% CO2 in Ham’s F-12 medium (Invitrogen) supplemented with 10% fetal
bovine serum. The cells were co-transfected with cDNA encoding EGFP-C3 alone or
EGFP-C3 fused to Gem GTPase, and cDNA encoding Ds-Red-N1 alone or Ds-Red-
N1 fused to tau, CTF of MAP2, or CTF of MAP4. Cells (250 µl) and each plasmid
7
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

DNA (1 µg) encoding GFP-Gem GTPase and tau-RFP were combined with
Lipofectamine (Invitrogen), essentially according to the manufacturer’s instructions. For
standard fluorescence microscopy, cells on coverslips were fixed for 15 min with 4%
paraformaldehyde in phosphate-buffered saline (PBS) and mounted onto a slide.
Labeled cells were examined under a Zeiss Axioskop microscope (Carl Zeiss, Tokyo,
Japan).
Confocal immunofluorescence microscopy—The cells on chamber slides (Nalge
Nunc International, Nperville, IL) were fixed with cold methanol or 4%
paraformaldehyde, followed by permeabilization with 0.1% Triton X-100 in PBS. The
fixed cells were probed with rat anti-tubulin monoclonal antibody (MAB1864, 1 µg/ml,
Chemicon international, Temecula, CA), followed by incubation with goat anti-rat IgG
(H+L) conjugated with Alexa Fluor 647 (1:1000; Molecular Probes, Eugene, OR), as
described previously (25). Specimens were observed under a confocal microscope (TCS
SP2; Leica Microsystems, Wetzlar, Germany).
Brain protein preparation—Control mouse brain at age of 8 weeks was
homogenized in 10 volumes of ice-cold 50 mM Tris-HCl buffer (pH 7.4) containing
150 mM NaCl and protease inhibitors (Complete EDTA-free, Roche Diagnostics,
Mannheim, Germany) using a Teflon/glass homogenizer. The homogenate was
centrifuged at 105,000 x g for 15 min at 4 ÚC, and the supernatant was saved for
biochemical analyses. For preparing heat-stable fraction, the supernatant was heated to
100 ÚC for 10 min and cleared by centrifugation at 105,000 x g for 15 min. Protein
concentrations were determined by the Bradford method (Bio-Rad, Hercules, CA).
Interaction of recombinant Gem GTPase and tauSoluble and heat-stable fractions
prepared from mouse brain homogenates were incubated at 4 ÚC for 1 h with
glutathione-Sepharose beads that were prebound to 50 µg glutathione-S-transferase
(GST) or GST-Gem GTPase as described previously (18). Whether tau binds to GST or
GST-Gem GTPase on glutathione-Sepharose was examined by western blotting using
8
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

tau 1 (Chemicon international).
Results
Gem GTPase is upregulated in the brains of eight-week-old tau-deficient
mice—To identify transcripts with altered expression, the levels of ~11,000 mRNAs in the
brains of tau-deficient mice were compared with those from the brain of control mice of
the same ages. To minimize potential confounding factors, pooled RNA samples from
control and tau-deficient mice were subjected to analysis of expression differences on
Affymetrix GeneChip, as described in Experimental Procedures. This initial microarray
screening suggested that the levels of 74 mRNAs including tau are significantly altered
in the brains of eight-week-old tau-deficient mice (Supplemental Table S2).
Quantitative RT-PCR using five wt and five tau-deficient mice indicated that, of these
transcripts, the levels of 13 mRNAs encoding transcription factors, cytokines, a ras-
related GTP-binding protein, protease inhibitors and a membrane protein were indeed
increased (Supplemental Table S3).
Of those genes with upregulated expression, we selected Gem GTPase for further
analysis, because it was significantly and consistently increased in tau-deficient mice.
The levels of Gem GTPase mRNA were most elevated at eight weeks of age in the
brains of tau-deficient mice (P < 0.001, Student’s t-test; Fig. 1), and did not differ
significantly from those of control mice at one week or eight months of age (Fig. 1).
Overexpression of Gem GTPase induces long cellular processes from CHO
cells—Although the major function of Gem GTPase has not yet been determined, this protein
was recently shown to be a negative regulator of the Rho-Rho kinase pathway for
cytoskeletal organization (18). To gain more insight into the function of Gem GTPase,
we examined the effects of its overexpression on the morphology of CHO cells. The
cells were transfected with GFP-tagged Gem GTPase and/or RFP-tagged tau, and
processed for fluorescence microscopy as described in Experimental Procedures.
9
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

First, we examined the intracellular localization of exogenous Gem GTPase and tau
in CHO cells. When cells were fixed with paraformaldehyde, the fluorescence of GFP
or RFP alone was distributed uniformly in the cytoplasm with occasional aggregation,
suggesting that neither GFP nor RFP is bound to cytoskeletal components, such as MTs
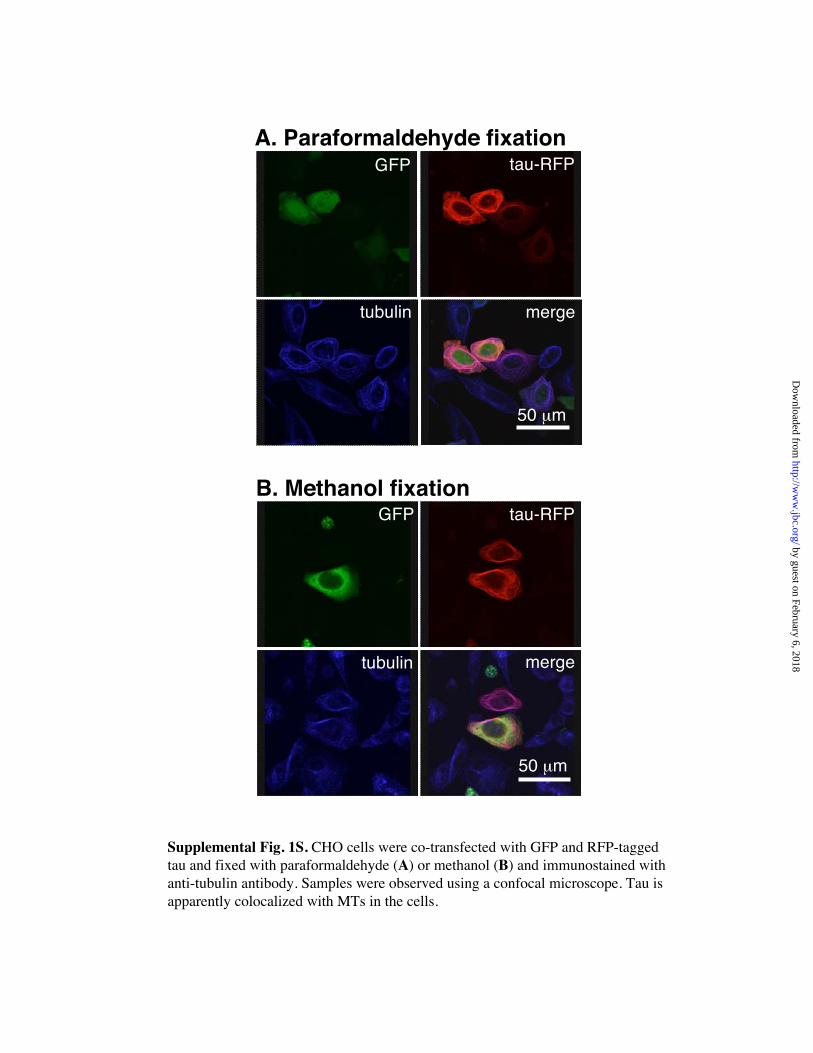
(Fig. 2A). In contrast, tau-RFP mainly colocalized with MTs (Fig. 2B and Fig. 1SA).
To confirm that tau binds to MTs, the cells were fixed with cold methanol to wash away
cytosolic tau-RFP. This fixation protocol showed that most tau-RFP colocalized with
MTs (Fig. 1SB), which suggested that the tau in the cytoplasm represents “overflowing”
tau. In many cells transfected with tau-RFP cDNA, MT bundles were observed (Fig. 2B
and Fig. 1S), but no other distinct morphological features were noted compared with
non-transfectant cells (Fig. 2A and B). Thus, overexpression of GFP, RFP, and
especially tau-RFP, induces no morphological changes in CHO cells.
In contrast, CHO cells overexpressing GFP-Gem GTPase and RFP displayed a
remarkable alteration in their morphology. Each cell became highly elongated and often
had one very long process (Fig. 2C). Because this effect was not observed in the GFP-
or RFP-expressing control cells (Fig. 2A), we concluded that Gem GTPase induced this
morphological change in CHO cells.
Surprisingly, the cell elongation induced by Gem GTPase was completely
suppressed by co-transfection with tau-RFP cDNA (Fig. 2D). To localize the active
region of tau that antagonizes the effect induced by Gem GTPase, either the amino-
terminal half (NTH) or the carboxyl-terminal half (CTH) of tau that is fused to RFP
was co-expressed with Gem GTPase. As shown in Fig. 2E and F, whereas the NTH of
tau did not suppress the Gem-GTPase-induced cell elongation, the CTH of tau
suppressed it completely. Thus, this suggests that the antagonistic effect is attributable
to the MT-binding domain of tau.
One hundred CHO cells expressing both GFP and RFP were chosen in an unselected
way and subjected to the quantification. The ratio of long axis (X)/ short axis (Y) of the
10
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

cell was taken as an index for cell elongation; higher values indicate higher degrees of
cell elongation. As shown in Fig. 3, the mean X values for Gem GTPase-
overexpressing cells were remarkably high, compared with those for the control cells
expressing only GFP or RFP (P < 0.01, Student’s t-test). This effect was completely
abolished by the co-expression of tau (P < 0.01).
A number of isoforms of tau are transcribed from a single gene by alternative
splicing. Each isoform is generated by the following combinations: no insert, inclusion
of exon 2-encoded insert or exons 2- and 3-encoded insert in the amino-terminal half
(referred to as S, M, and L, respectively), and exclusion/inclusion of exon 10-encoded
second repeat in the carboxyl-terminal half [referred to as three-repeat (3R) and four-
repeat (4R) tau, respectively]. This rather complicated splicing is developmentally
regulated, and the species S3R that is expressed exclusively in the fetal brain is rapidly
replaced by 4R isoforms after birth in rodent (26-28). We examined whether the effects
of the Gem GTPase are isoform-dependent. The L and M4R isoforms showed similar
inhibitory activities against the effect induced by Gem GTPase (data not shown). The
three-repeat tau also inhibited Gem-GTPase-induced cell elongation (data not shown).
These results are consistent with the assumption that the MT-binding domain of tau has
the activity that antagonizes Gem-GTPase-induced cell elongation.
Antagonizing activity is attributable to the MT-binding domain of MAPs—The
MT-binding region of tau is composed of highly conserved 18-residue-long binding
elements, each having a distinctive Pro-Gly-Gly-Gly motif (29-31). The MT-binding
motifs are also found in the CTFs of MAP2 and MAP4 (Fig. 4A and B). We examined
whether these two homologous MT-binding regions suppress Gem-GTPase-induced
cell elongation. When GFP-Gem GTPase and MAP2 CTF-RFP were co-expressed,
the elongated cell morphology was no longer observed (Fig. 4C and D). The MT-
binding domain of MAP4 had also a similar inhibitory effect (Fig. 4E). Whereas the C-
terminal extensions of tau and MAP2 are highly homologous, that of MAP4 beyond the
11
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

MT-binding domain is not homologous with the former (Fig. 4B). These results
indicate that the MT-binding domains but not the C-terminal extensions have these
antagonistic activities, and suggest that those MAPs as well as tau may regulate the
function of Gem GTPase in vivo.
To clarify whether the inhibitory effect of tau on cellular elongation is inherent to
CHO cells, other cell lines were examined. HeLa, Neuro2a, or PC12 cells were
transfected with either a GFP-Gem GTPase- or a GFP- (control) expressing construct,
and the cell shape was evaluated 24 h later. Overexpression of Gem GTPase in HeLa
cells caused a morphological change similar to that observed in CHO cells, but no such
phenotype was observed in Neuro2a or PC12 cells (data not shown). We do not yet
know whether this negative effect is due to the presence of low levels of tau in these cell
lines.
Tau does not directly bind to Gem GTPase—Finally, we asked whether there is a
direct molecular interaction between Gem GTPase and tau. We sought to detect a
potential interaction using recombinant glutathione S-transferase (GST)-Gem GTPase
and tau in mouse brain extracts. Further, we used the tau in heat-treated mouse brain
extract, because heat-treated tau must be free of tau-interacting proteins. In either case,
we were unable to find any noticeable interaction between Gem GTPase and tau (Fig.
5A and B). In addition, the intracellular distribution of GFP-Gem GTPase and tau-RFP
in transfected CHO cells was carefully assessed using a confocal microscope. When
fixed with paraformaldehyde, the GFP-Gem GTPase was distributed diffusely in the
cytoplasm, consistent with a previous report (18). In contrast, tau-RFP almost
completely colocalized with MTs (Fig. 5C). Similar results were also obtained in the
methanol-fixed cells (Fig. 5D). All of the above results strongly suggest that there is no
direct molecular interaction between Gem GTPase and tau.
Discussion
12
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

DNA microarray technology allows global analysis of neural responses to tau
deficiency in vivo, and offers comprehensive analysis of the altered or compensatory
gene expression involved in tau deficiency. This screening suggests that the expression
of Gem GTPase, a member of the ras superfamily, is significantly increased in the
brains of eight-week-old tau-deficient mice, which was confirmed by quantitative RT-
PCR.
Because the function of Gem GTPase has not yet been determined, we looked for a
possible cell-biological relationship between Gem GTPase and tau. We first found that
Gem GTPase induces profound changes in the cell morphology in CHO cells. A similar
cellular effect of Gem GTPase was reported in other cells, including human umbilical
vein endothelial cells (HUVEC) and HEK-293 cells (19). In addition, Ges, another
Rad/Gem/Kir (RGK) family member and a human ortholog of mouse Rem1, was
reported to induce endothelial cell sprouting (32). Tau is indeed able to antagonize the
effects induced by Gem GTPase, and this activity is localized to the MT-binding
domain of tau (Fig. 2E and F). The motifs of the domain are composed of highly
conserved 18-amino acid repeat identified in tau (29-31), which are also found in
MAP2 and MAP4 (Fig. 4A and B). Either of these MT-binding regions similarly
antagonized the Gem GTPase–induced effect (Fig. 4). These results indicate that
cellular elongation, a Gem GTPase-induced phenotype, can be modulated by MT-
binding domains of tau, MAP2, and MAP4.
A number of exonic and intronic mutations in the tau gene have been identified in
patients affected by frontotemporal dementia and parkinsonism linked to chromosome
17 (FTDP-17). This indicates that tau is involved in neuronal death in subsets of
neurons in this disease, and presumably this is the case with Alzheimer’s disease.
Because the mutant forms of tau associated with FTDP-17 destabilize MTs to a certain
extent in CHO cells (25), the effects of several mutations on the cell elongation induced
by Gem GTPase were evaluated in CHO cells. Co-expression of GFP-Gem GTPase
13
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

and a tau variant, G272V, P301L, V337M, or R406W completely suppressed the
morphological changes (data not shown). Thus, at least these four mutant tau molecules
have similar degrees of the antagonistic activity.
A syndrome known as hereditary dysphasic disinhibition dementia 2 (HDDD2) is
similar in neuropathology to a certain form of FTDP-17 and maps to 17q21-22, but no
mutation in the tau gene has thus far been identified in the members of this family (33).
Recent studies revealed almost complete loss of tau in several HDDD2 brains, despite
apparently normal levels of tau mRNA (34). These results may suggest that HDDD2 is
a distinct tauopathy, possibly caused by a profound reduction in the protein levels of
tau. If so, it would be of particular interest to examine whether Gem GTPase is
upregulated in HDDD2 brains.
The cytoskeletal changes that alter cellular morphology and motility depend upon a
complicated interplay of the molecules that regulate actin, myosin, and other
cytoskeletal components. The Rho family of GTP-binding proteins is an important
upstream mediator for the cytoskeletal organization. Gem GTPase is a negative
regulator on the Rho-Rho kinase pathway, which is involved in stress fiber formation
and neurite retraction (18). Gem GTPase interacts with the Rho pathway through its
association with the Rho effecter, ROKß. Therefore, Gem GTPase may affect cell
morphology by antagonizing the actions of ROKß. Recently, it was shown that
phosphorylation of serines 261 and 289, located in the carboxyl terminal extension of
Gem GTPase, is required for Gem GTPase-mediated cytoskeletal reorganization (35).
Thus, the phosphorylation of serines 261 and 289, which are outside the GTP-binding
region of Gem GTPase, regulates its inhibition of ROKß and associated changes in the
cytoskeleton. The present results provide evidence that one function of Gem GTPase
related to the cell morphology is modulated by the MT-binding region of tau. This
raises the possibility that tau may exist upstream in the Gem GTPase-mediated signal
transduction pathway. Further studies should provide a better understanding of the
14
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

molecular basis of the tau-modulated, Gem GTPase-mediated signal transduction
pathway.
Acknowledgments: We thank Dr. T. Yamazaki for valuable suggestions on fluorescence
microscopy, and Dr. M. Morishima-Kawashima for critical reading of this manuscript.
We also thank Mr. K. Fukumoto for the help for GeneChip analysis and Dr. Y.V.
Kotliarov for providing us a multiple analysis comparison software.
15
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

References1. Cleveland, D. W., Hwo, S. Y., and Kirschner, M. W. (1977) J Mol Biol 116,
207-2252. Binder, L. I., Frankfurter, A., and Rebhun, L. I. (1985) J Cell Biol 101, 1371-
13783. Matus, A. (1988) Annu Rev Neurosci 11, 29-444. Hirokawa, N., Shiomura, Y., and Okabe, S. (1988) J Cell Biol 107, 1449-14595. Hirokawa, N. (1994) Curr Opin Cell Biol 6, 74-816. Kanai, Y., Takemura, R., Oshima, T., Mori, H., Ihara, Y., Yanagisawa, M.,
Masaki, T., and Hirokawa, N. (1989) J Cell Biol 109, 1173-11847. Chen, J., Kanai, Y., Cowan, N. J., and Hirokawa, N. (1992) Nature 360, 674-
6778. Harada, A., Oguchi, K., Okabe, S., Kuno, J., Terada, S., Ohshima, T., Sato-
Yoshitake, R., Takei, Y., Noda, T., and Hirokawa, N. (1994) Nature 369, 488-491
9. Takei, Y., Teng, J., Harada, A., and Hirokawa, N. (2000) J Cell Biol 150, 989-1000
10. Lee, V. M., Balin, B. J., Otvos, L., Jr., and Trojanowski, J. Q. (1991) Science251, 675-678
11. Hutton, M., Lendon, C. L., Rizzu, P., Baker, M., Froelich, S., Houlden, H., Pickering-Brown, S., Chakraverty, S., Isaacs, A., Grover, A., Hackett, J., Adamson, J., Lincoln, S., Dickson, D., Davies, P., Petersen, R. C., Stevens, M., de Graaff, E., Wauters, E., van Baren, J., Hillebrand, M., Joosse, M., Kwon, J. M., Nowotny, P., Heutink, P., and et al. (1998) Nature 393, 702-705
12. Poorkaj, P., Bird, T. D., Wijsman, E., Nemens, E., Garruto, R. M., Anderson, L., Andreadis, A., Wiederholt, W. C., Raskind, M., and Schellenberg, G. D. (1998)Ann Neurol 43, 815-825
13. Spillantini, M. G., Murrell, J. R., Goedert, M., Farlow, M. R., Klug, A., and Ghetti, B. (1998) Proc Natl Acad Sci U S A 95, 7737-7741
14. Cohen, L., Mohr, R., Chen, Y. Y., Huang, M., Kato, R., Dorin, D., Tamanoi, F., Goga, A., Afar, D., Rosenberg, N., and et al. (1994) Proc Natl Acad Sci U S A91, 12448-12452
15. Maguire, J., Santoro, T., Jensen, P., Siebenlist, U., Yewdell, J., and Kelly, K. (1994) Science 265, 241-244
16. Dorin, D., Cohen, L., Del Villar, K., Poullet, P., Mohr, R., Whiteway, M., Witte, O., and Tamanoi, F. (1995) Oncogene 11, 2267-2271
17. Beguin, P., Nagashima, K., Gonoi, T., Shibasaki, T., Takahashi, K., Kashima, Y., Ozaki, N., Geering, K., Iwanaga, T., and Seino, S. (2001) Nature 411, 701-706
18. Ward, Y., Yap, S. F., Ravichandran, V., Matsumura, F., Ito, M., Spinelli, B., and
16
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

Kelly, K. (2002) J Cell Biol 157, 291-30219. Piddini, E., Schmid, J. A., de Martin, R., and Dotti, C. G. (2001) Embo J 20,
4076-408720. Aresta, S., de Tand-Heim, M. F., Beranger, F., and de Gunzburg, J. (2002)
Biochem J 367, 57-6521. Ikegami, S., Harada, A., and Hirokawa, N. (2000) Neurosci Lett 279, 129-13222. Luthi-Carter, R., Strand, A., Peters, N. L., Solano, S. M., Hollingsworth, Z. R.,
Menon, A. S., Frey, A. S., Spektor, B. S., Penney, E. B., Schilling, G., Ross, C. A., Borchelt, D. R., Tapscott, S. J., Young, A. B., Cha, J. H., and Olson, J. M. (2000) Hum Mol Genet 9, 1259-1271
23. Lipshutz, R. J., Fodor, S. P., Gingeras, T. R., and Lockhart, D. J. (1999) NatGenet 21, 20-24
24. Gu, Y., Oyama, F., and Ihara, Y. (1996) J Neurochem 67, 1235-124425. Matsumura, N., Yamazaki, T., and Ihara, Y. (1999) Am J Pathol 154, 1649-
165626. Goedert, M., Spillantini, M. G., Jakes, R., Rutherford, D., and Crowther, R. A.
(1989) Neuron 3, 519-52627. Himmler, A. (1989) Mol Cell Biol 9, 1389-139628. Kosik, K. S., Orecchio, L. D., Bakalis, S., and Neve, R. L. (1989) Neuron 2,
1389-139729. Himmler, A., Drechsel, D., Kirschner, M. W., and Martin, D. W., Jr. (1989) Mol
Cell Biol 9, 1381-138830. Lee, G., Neve, R. L., and Kosik, K. S. (1989) Neuron 2, 1615-162431. Butner, K. A., and Kirschner, M. W. (1991) J Cell Biol 115, 717-73032. Pan, J. Y., Fieles, W. E., White, A. M., Egerton, M. M., and Silberstein, D. S.
(2000) J Cell Biol 149, 1107-111633. Lendon, C. L., Lynch, T., Norton, J., McKeel, D. W., Jr., Busfield, F., Craddock,
N., Chakraverty, S., Gopalakrishnan, G., Shears, S. D., Grimmett, W., Wilhelmsen, K. C., Hansen, L., Morris, J. C., and Goate, A. M. (1998)Neurology 50, 1546-1555
34. Zhukareva, V., Vogelsberg-Ragaglia, V., Van Deerlin, V. M., Bruce, J., Shuck, T., Grossman, M., Clark, C. M., Arnold, S. E., Masliah, E., Galasko, D., Trojanowski, J. Q., and Lee, V. M. (2001) Ann Neurol 49, 165-175
35. Ward, Y., Spinelli, B., Quon, M. J., Chen, H., Ikeda, S. R., and Kelly, K. (2004)Mol Cell Biol 24, 651-661
36. Kindler, S., and Garner, C. C. (1994) Brain Res Mol Brain Res 26, 218-224
17
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

Footnotes
* This work was supported by Grants-in-Aid for Scientific Research (No. 14580722)
and for Scientific Research on Priority Areas (C), Advanced Brain Science Project,
from the Ministry of Education, Culture, Sports, Science and Technology, Japan.
Abbreviations used: AD, Alzheimer’s disease; CTF, carboxyl-terminal fragment; CTH,
carboxyl-terminal half; FTDP-17, frontotemporal dementia and parkinsonism linked to
chromosome 17; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; GFP, green
fluorescent protein; MAPs, microtubule-associated proteins; MT, microtubule; GST,
glutathione S-transferase; NTF, amino-terminal fragment; NTH, amino-terminal half;
PHFs, paired helical filaments; RFP, red fluorescent protein; ROK, Rho kinase; RT-
PCR, reverse transcriptase-coupled PCR; wt, wild-type
18
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

Figure legends
Fig. 1. Confirmation by RT-PCR of the upregulation of Gem GTPase mRNA in the
tau-deficient mouse brain. Quantitative RT-PCR was performed using individual wild-
type (Wt) and tau-deficient mice (KO) at one week, eight weeks and eight months of
age. Gem GTPase mRNA levels are normalized against those of GAPDH. Values are
given as mean ± SD (n = 5). The levels of Gem GTPase mRNA were significantly
elevated at eight weeks of age in the brains of tau-deficient mice (P < 0.001, Student’s
t-test), but did not differ significantly from those of control mice at one week or eight
months of age.
Fig. 2. Effect of overexpressed Gem GTPase and tau on the morphology of CHO cells.
CHO cells on glass coverslips were co-transfected with cDNAs encoding GFP alone or
GFP fused to Gem GTPase, and RFP alone or RFP fused to tau, and further cultured for
24 h. The cells were then fixed, and the GFP (left) and RFP (right) fluorescence was
observed under a fluorescence microscope. The transfected cDNAs are as follows: A,
GFP and RFP cDNAs; B, GFP and tau-RFP cDNAs; C, GFP-Gem GTPase and RFP
cDNAs; D, GFP-Gem GTPase and tau-RFP cDNAs; E, GFP-Gem GTPase and the
amino-terminal half of tau (tau NTH)-RFP cDNAs; F, GFP-Gem GTPase and the
carboxyl-terminal half of tau (tau CTH)-RFP cDNAs. Bar, 50 µm.
Fig. 3. Quantification of the cellular elongation induced by Gem GTPase. One hundred
transfected cells were unselectedly chosen from each specimen, and the length of the
long axes (X in µm), and short axes (Y in µm), and their ratio (X/Y) are given as means
± SD. The X values for Gem-GTPase-overexpressing cells were markedly shifted to
the right (data not shown), compared with those of control cells expressing only GFP or
RFP. Co-expression of tau or its CTH abolished the effect.
19
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

Fig. 4. The MT-binding domains of MAP2 and MAP4 suppress Gem-GTPase-induced
cell elongation. Sequence similarities between CTH of tau (169 residues) and CTF of
MAP2 (138 residues) or CTF of MAP4 (136 residues) are shown according to NCBI 2
blast algorithm. A, CTF of MAP2 is highly homologous to CTH of tau [Identities =
90/136 (66%), Positives = 110/136 (80%)]. It has been known that, like tau, MAP2
contains either three or four tandem repeats in the MT-binding domain (36). Each
repeat of MAP2 consists of highly conserved 18 residues. Only the CTF from three-
repeat MAP2 is shown here. B, The CTF of MAP4 has also MT-binding motifs
[Identities = 61/120 (50%), Positives = 78/120 (64%), Gaps = 31/120 (25%)]. The 18-
residue-long MT-binding motifs of tau are indicated by thick lines. C, transfection of
GFP-Gem GTPase and RFP cDNAs; D, GFP-Gem GTPase and MAP2 CTF-RFP
cDNAs; and E, GFP-Gem GTPase and MAP4 CTF-RFP cDNAs. The expression of
CTFs of MAP2 and MAP4 abolished the cell elongation (D, E). Bar, 50 µm.
Fig. 5. Lack of direct interaction of Gem GTPase and tau. Mouse brain extract (A) or
heat-treated mouse brain extract (B) was incubated with glutathione-Sepharose beads
alone (lanes 1 and 4), recombinant GST (lanes 2 and 5)- or GST-Gem GTPase (lanes 3
and 6)-bound glutathione-Sepharose beads. The bound and unbound fractions were
subjected to western blot analysis with anti-tau antibody (tau 1). The tau-reactive band
was undetectable in the bound fractions (A, B). CHO cells were co-transfected with
GFP-Gem GTPase and tau-RFP, and fixed with paraformaldehyde (C) or methanol
(D), followed by immunostaining with tubulin. Specimens were observed under a
confocal microscope. Tau apparently colocalized with MTs in the cells. Bar, 50 µm.
20
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

0
200
400
600
800
1000
Gem
GTP
ase
mR
NA
leve
l1 week 8 weeks 8 months
Wt KO Wt KO Wt KO
Fig. 1. Oyama et al.
by guest on February 6, 2018 http://www.jbc.org/ Downloaded from

50 mm
A. GFP / RFP B. GFP / tau-RFP
C. GFP-Gem / RFP D. GFP-Gem / tau-RFP
E. GFP-Gem / tau NTH-RFP F. GFP-Gem / tau CTH-RFP
GFP │ FP
GFP-Gem │ FP
GFP-Gem tau NTH-RFP
GFP
GFP-Gem
GFP-Gem
tau-│ FP
tau-│ FP
tau CTH-RFP
Fig. 2. Oyama et al.
by guest on February 6, 2018 http://www.jbc.org/ Downloaded from

0
10
20
30
40
50
60
70
80
X
0
5
10
15
20
Y
0
5
10
15
20
25
X / Y
GFP
/RFP
GFP
-Gem
/RFP
GFP
/ ta
u-R
FP
GFP
-Gem
/ tau
-RFP
GFP
-Gem
/ tau
CTH
-RFP
Fig. 3. Oyama et al.
GFP
/RFP
GFP
-Gem
/RFP
GFP
/ ta
u-R
FP
GFP
-Gem
/ tau
-RFP
GFP
-Gem
/ tau
CTH
-RFP
GFP
/RFP
GFP
-Gem
/RFP
GFP
/ ta
u-R
FP
GFP
-Gem
/ tau
-RFP
GFP
-Gem
/ tau
CTH
-RFP
by guest on February 6, 2018 http://www.jbc.org/ Downloaded from

C. GFP-Gem / RFP
D. GFP-Gem / MAP2 CTF-RFP
E. GFP-Gem / MAP4 CTF-RFP
50 mm
GFP-Gem RFP
GFP-Gem
GFP-Gem
MAP2 CTF-RFP
MAP4 CTF-RFP
Fig. 4. Oyama et al.
A. Tau-CTH vs. MAP2-CTF
tau-CTH :DLSNVQSKCGSKDNIKHVPGGGSVQIVYKPVDLSKVTSKCDL NV+SK GS DNIK+ P GG VQIV K +DLS VTSKC
MAP2-CTF:DLKNVKSKIGSTDNIKYQPKGGQVQIVTKKIDLSHVTSKC
tau-CTH :GSLGNIHHKPGGGQVEVKSEKLDFKDRVQSKIGSLDNITHGSL NI H+PGGG+V+++S KLDFK++ Q+K+GSLDN H
MAP2-CTF:GSLKNIGHRPGGGRVKIESVKLDFKEKAQAKVGSLDNAHH
tau-CTH :VPGGGNKKIETHKLTFRENAKAKTDHGAEIVYKSPVVSGDVPGGGN KI++ KL FRE+AKA+ DHGAEI+ +SP S
MAP2-CTF:VPGGGNVKIDSQKLNFREHAKARVDHGAEIITQSPGRSSV
tau-CTH :TSPRHLSNVSSTGSIDMSPR LSNVSS+GSI++
MAP2-CTF:ASPRRLSNVSSSGSINL
B. Tau-CTH vs. MAP4-CTF
tau-CTH :LKNVKSKIGSTENLKHQPGGGKVQIINKKLDLSNVQSKCGLKNV+SK+GSTEN+KHQPGGG+VQI NKK+D+S V SKCG
MAP4-CTF:LKNVRSKVGSTENIKHQPGGGRVQIQNKKVDISKVSSKCG
tau-CTH :SKDNIKHVPGGGSVQIVYKPVDLSKVTSKCGSLGNIHHKPSK NIK HKP
MAP4-CTF:SKANIK-------------------------------HKP
tau-CTH :GGGQVEVKSEKLDFKDRVQSKIGSLDNITHVPGGGNKKIEGGG V+++S+KL+FK++ Q+K+GSLDN+ H+P GG K E
MAP4-CTF:GGGDVKIESQKLNFKEKAQAKVGSLDNVGHLPAGGAVKTE
MT-binding motif 2
MT-binding motif 3 MT-binding motif 4
MT-binding motif 1
MT-binding motif 2 MT-binding motif 3
MT-binding motif 4
by guest on February 6, 2018 http://www.jbc.org/ Downloaded from

moc
kG
ST GST
-Gem
moc
kG
ST GST
-Gem
A. Brain extract
1 2 3 4 5 6
BoundUnbound
10075
5037
kDa
moc
kG
STG
ST-G
emm
ock
GST
GST
-Gem
B. Brain heat-stable fraction
1 2 3 4 5 6
BoundUnbound
10075
5037
GFP-Gem tau-RFP
tubulin merge
50 mm
C. Paraformaldehyde fixation D. Methanol fixationGFP-Gem tau-RFP
tubulin merge
50 mm
Fig. 5. Oyama et al.
Glutathione-Sepharose Glutathione-Sepharose
by guest on February 6, 2018 http://www.jbc.org/ Downloaded from

Supplemental Table S1. The nucleotide sequences of the primers for quantitative RT-PCRmRNA name Forward primer Reverse primerFosB protein (fosB) tcaaccagcacaaccaccagtggacctgtg gtctaaagagtttacagagcaagaagggagc-fos(p55) and p15 E reading frames acttgaagatgagaagtctgcgttgcagac cacagggccagcagcgtgggtgagctcagg prostaglandin synthase tgagtcattcaccagacagattgctggccg tcagtagacttttacagctcagttgaacgcsuppressor of cytokine signalling-3 (SOCS-3) aagcgagtaccagctggtggtgaacgccgt gctccttaaagtggagcatcatactgatccmacrophage inflammatory protein (MIP) atgaaggtctccaccactgcccttgctgtt gtctacctagaatagctgtcaccaaacagtinterferon beta (type 1) coding for the c-terminal part gtcactgtttgtggacggatcaggtcctcg gctgctgtctgatagttctttatttcaccamacrophage inflammatory protein (MIP) ccatgaagctctgcgtgtct catcagggctgaggaggcctPTX3 cttcccaatgcgttcgaagaagatttttgg aacacttaagaaacatactgggctcctccgC10-like chemokine agatgaagccttttcatactgccctctcct gtcattgaggtcagcttcagaccttccaggplatelet-derived growth factor-inducible protein (JE) cagctctctcttcctccaccaccatgcagg ggcatcacagtccgagtcacactagttcacctla-2-alpha atggtttctatctgtgaacagaagctgcag ttaatcttacatgaattcaatgttacactatranscription factor LRG-21 tggccgctctctggaccctggccgccccga cctctgcttagctctgcaatgttccttctttumor-associated membrane protein (TMP) ctctactcagaagtcaccaaaaagccaaga acacagtgactcctgcttccctcccaggttMCP-1 related chemokine cctctagctttcatttcgaa tcagcctagacatgaaggttMyD118, a myeloid differentiation primary response gene cagcgtggtcttgtgcctcc tctcagcgttcctctagagaplasminogen activator inhibitor (PAI-1) gaggccagcacccaccagcgcctcttccac tcaaggctccatcacttgccccatgaagagserum amyloid A (SAA) agaagctggtcaagggtctagagacatgtg tgcatttcacaagtatttattcagcacattC/EBP delta caccctagagctgtgccacgacgaactctt gccgcgcgttaccggcagtcggcgccggtgCD14 ctgcaggcgctccgagttgt atgttccttaaacaaagaggtransglutaminase (TGase) ttcgacgtgtttgcccacatcggcaacgac cccttaggccgggccgatgataacattccggly96 gcccctggctccggaggatg cataaatacataaatattaaC3H cytochrome P450 (Cyp1b1) ccaagtggcctaacccagag ccctccggcctctcacttgcspi2 proteinase inhibitor (spi2/eb4) cagcttacaaccagagaccc gtctcatttggggttggctaadipose differentiation related protein (ADFP) gggcttatcaccaggctctcagcagggtta gggtttactgagctttgacctcagactgctbrain-derived neurotrophic factor ggttcgagaggtctgacgacgacatcactg ccactatcttccccttttaatggtcagtgtinterferon beta (type 2) coding for the C-terminal part ttgatgctgcaacacttaaa catcacaaacactttatttgGADD45 protein (gadd45) aatatgactttggaggaatt ttctccagtagcagcagctcN10 gene for a nuclear hormonal binding receptor cagggcctccgccacctcca gcagccctcacaggtgcggaGem GTPase (gem) atgacagcatggacagcgactgtgaggtct gtgcctagagcacagacaggtcatggcaggAPP-S cagaattcggacatgattca gggcttagttctgcatttgcsmall heat shock protein (HSP25) tccctccaggtgtggacccc gtagcttgcaacttttatttgut-enriched Kruppel-like factor GKLF gaggagcccaagccaaagag cgtgggatttaaaagtgcctinterleukin-11 ctccggcatgtacaatggct agtcacagtcgagtctttaagrowth factor-induced protein (zif/268) tccatttaagacagaaggac ccctttagcaaatttcaattheparin-binding EGF-like growth factor precursor gaagaagagagacccatgcc agtgggagctagccacgcccsignal transducer and activator of transcription (Stat3) atgggctataagatcatgga cacatgggggaggtagcacamyelin associated-glycoprotein (MAG) ccgaggatgatggggaatactggtgtgtgg ctcagtgacaatcccgggtagagacctctcmyelin/oligodendrocyte glycoprotein (Mog) ttctattgggtcaaccccgg ccatcactcaaaaggggtttG-alpha-12 protein taacttggaccggattggccagctgaacta gtctctcactgcagcatgatgtctttcaggGAPDH gtggaagggctcatgaccacagtccatgcc tcttactccttggaggccatgtaggccatg
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

Supplemental Table S2. Altered expression of mRNAs in eight weeks old tau-deficient mice by GeneChip analysisProbe Set Fold Change Descriptionx14897_s_at 23.9 Mus musculus FosB protein (fosB) gene, complete cdsv00727_s_at 16.0 Provirus of a replication defective murine sarcoma virus with c-fos(p55) and p15 E reading framesm64291_s_at 11.3 Mus musculus prostaglandin synthase mRNA, complete cds.U88328_at 10.0 Mus musculus suppressor of cytokine signalling-3 (SOCS-3) mRNAm58691-2_s_at 8.5 Mouse TIS11 primary response gene, complete cdsx12531_s_at 8.2 Mouse mRNA for macrophage inflammatory protein (MIP).v00755_s_at 7.4 Messenger RNA fragment for mouse interferon beta (type 1) coding for the c-terminal part.X83601_s_at 7.0 M.musculus PTX3 mRNAu19482_s_at 6.7 Mus musculus C10-like chemokine mRNAw82380_s_at 6.7 Mus musculus cDNA clone 404017 5, mRNA sequencem19681_s_at 6.2 Mouse platelet-derived growth factor-inducible protein (JE) geneX15591_s_at 6.1 Mouse ctla-2-alpha mRNA, homolog. to cysteine protease proregionD67076_s_at 5.9 Mouse mRNA for secretory protein containing thrombospondin motifsm35590_s_at 5.9 Mouse macrophage inflammatory protein 1-beta (MIP-1) mRNAu19118_s_at 5.9 Mus musculus transcription factor LRG-21 mRNAMsa.570.0_g_at 5.5 Mouse connexin (Cx26) geneu25633_s_at 5.4 Mus musculus tumor-associated membrane protein (TMP) mRNAu50712_s_at 5.1 Mus musculus MCP-1 related chemokine mRNAMsa.34555.0_s_at 5.1 Homologous to sp P06685: SODIUM/POTASSIUM-TRANSPORTING ATPASE ALPHA-1 CHAIN M33960_s_at 4.7 Mouse plasminogen activator inhibitor (PAI-1) mRNA, complete cdsX54149_s_at 4.7 Mouse mRNA for MyD118, a myeloid differentiation primary response genex03479_s_at 4.0 Mouse mRNA fragment for serum amyloid A (SAA) 3 proteinX61800_s_at 3.9 M.musculus mRNA for C/EBP deltaC77421_rc_f_at 3.8 Mus musculus cDNA clone J0030G04 3 similar to Mouse B10.VL30LTR geneM55154_s_at 3.6 Mouse transglutaminase (TGase) mRNAx13333_s_at 3.6 Mouse CD14 mRNA for myelid cell-specific leucine-rich glycoprotein.X15592_s_at 3.6 Mouse ctla-2-alpha mRNA, homolog. to cysteine protease proregionMsa.2173.0_at 3.5 M.musculus mRNA for inhibin beta-A subunitaa111276_s_at 3.4 Mus musculus cDNA clone 557308 5 similar to ALPHA-2-MACROGLOBULIN PRECURSOR (HUMAN)X67644_s_at 3.3 M.musculus gly96 mRNAAA289952_at 3.2 Mus musculus cDNA clone 752147 5, mRNA sequence.u03283_s_at 3.2 Mus musculus C3H cytochrome P450 (Cyp1b1) mRNAMsa.31964.0_f_at 3.2 Homologous to sp P05197: ELONGATION FACTOR 2 (EF-2)Msa.7498.0_s_at 2.9 Homologous to sp P24522: GROWTH ARREST AND DNA-DAMAGE-INDUCIBLE PROTEIN GADD45 MIP1-B_at 2.7 #N/AMsa.3437.0_f_at 2.7 Homologous to sp P42930: HEAT SHOCK 27 KD PROTEIN (HSP 27).x51834_f_at 2.7 Murine gene for osteopontinM64086_s_at 2.6 Mouse spi2 proteinase inhibitor (spi2/eb4) mRNAMsa.1376.0_f_at 2.6 Mouse mRNA for early T-lymphocyte activation 1 protein (ETa-1)M93275_s_at 2.5 Mouse adipose differentiation related protein (ADFP) mRNAl28177_s_at 2.4 Mus musculus GADD45 protein (gadd45) genev00756_f_at 2.4 Messenger RNA fragment for mouse interferon beta (type 2) coding for the C-terminal partX55573_s_at 2.4 M.musculus brain-derived neurotrophic factor mRNAx16995_s_at 2.3 Mouse N10 gene for a nuclear hormonal binding receptoraa407278_rc_s_at 2.2 EST02275 Mouse 7.5 dpc embryo ectoplacental cone cDNA library u10551_s_at 2.2 Mus musculus Gem GTPase (gem) mRNAMIP1-B_at 2.2 #N/AMsa.10643.0_s_at 2.2 Homologous to sp P12023: Alzheimer's disease amyloid A4 protein homolog precursorMsa.5082.0_s_at 2.2 Homologous to sp P41220: G0/G1 SWITCH REGULATORY PROTEIN 8.l07577_s_at 2.1 Mus musculus small heat shock protein (HSP25) gene,.aa120653_s_at 2.0 Mus musculus cDNA clone 574724 5 similar to gb:D21261 SM22-ALPHA HOMOLOG (HUMAN)U03421_s_at 2.0 Mus musculus interleukin-11 mRNAu20344_s_at 2.0 Mus musculus gut-enriched Kruppel-like factor GKLF mRNAm22326-2_s_at 1.9 Mouse growth factor-induced protein (zif/268) mRNAx62600_s_at 1.9 Mouse alpha-1-acid glycoprotein (AGP/EBP) mRNAaa184871_s_at 1.8 mu51f10.r1 Soares mouse lymph node NbMLN Mus musculus cDNA clone 642955 5c78546_rc_s_at 1.8 similar to moesin homolog [mice, teratocarcinoma F9 cells, mRNA, mRNA sequence.aa426892_s_at 1.7 Mouse tissue plasminogen activator mRNA, complete cds (MOUSE)l07264_s_at 1.7 Mus musculus (clone lambda mouse 1) heparin-binding EGF-like growth factor precursor mRNAMsa.31166.0_s_at 1.7 Homologous to sp P10644: cAMP-dependent protein kinase type-1 alpha regulatory chainu06922_s_at 1.6 Mus musculus signal transducer and activator of transcription (Stat3) mRNAl21027_s_at -1.6 Mouse A10 mRNAw83858_g_at -1.6 mf26a04.r1 Soares mouse embryo NbME13.5 14.5 Mus musculus cDNA clone 406158 5Msa.24554.0_s_at -1.7 Homologous to sp P28656: BRAIN PROTEIN DN38 Msa.2867.0_s_at -1.9 M.musculus mRNA for astrocytic phosphoprotein, PEA-15m31811_s_at -2.0 Mouse myelin associated-glycoprotein (MAG) mRNAu64572_s_at -2.1 Mus musculus myelin/oligodendrocyte glycoprotein (Mog) mRNAET63463_s_at -2.1 2-HYDROXYACYLSPHINGOSINE 1-BETA-GALACTOSYLTRANSFERASE PRECURSOR aa013647_s_at -2.2 Mus musculus cDNA clone 442176 5, mRNA sequence.Msa.10603.0_at -2.3 Homologous to sp P32076: neuron specific calcium-binding protein hippocalcin (P23K)Msa.22035.0_s_at -2.3 Homologous to sp P48547: POTASSIUM CHANNEL PROTEIN KV3.1/KV4 (NGK2).M63659_s_at -3.0 Mouse G-alpha-12 protein mRNAm18776_s_at -6.8 Mus musculus Him OF1 SPF microtubule-associated protein Tau isoform 23 mRNA
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

Supplemental Table S3. Altered expression of 13 mRNAs confirmed by quantitative RT-PCRAccession No mRNA Fold change Functionv00727 c-fos 2.6 cellular oncogene; regulation of transcriptionv00756 IFN beta tpe 2 1.9 interferon-related developmental regulator u10551 Gem GTPase 1.9 GTP binding protein (gene overexpressed in skeletal muscle)m22326 Zif/268 1.7 growth factor-inducible protein; transcription factoru20344 GKLF 1.7 negative regulator of cell growth; regulation of transcriptionX54149 MyD118 1.6 a novel myeloid differentiation primary response geneM64086 spi2 1.6 serine proteinase inhibitor; serpin; serum protein l28177 GADD45 1.5 activation of MAPKK; protein biosynthesis; cell differentiationX15592 ctla-2 1.4 cysteine protease; proteasex13333 CD14 1.4 CD14 antigen; inflammatory response; immune responseu25633 TMP 1.3 tumor-associated gene; a membrane glycoproteinu06922 Stat3 1.3 signal transducers and activators of transcriptionv00755 IFN beta tpe 1 1.2 interferon-related developmental regulator
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from
50 mm
GFP tau-RFP
tubulin merge
A. Paraformaldehyde fixation
50 mm
GFP tau-RFP
tubulin merge
B. Methanol fixation
Supplemental Fig. 1S. CHO cells were co-transfected with GFP and RFP-taggedtau and fixed with paraformaldehyde (A) or methanol (B) and immunostained withanti-tubulin antibody. Samples were observed using a confocal microscope. Tau is apparently colocalized with MTs in the cells.
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from

Yoshito Ueyama, Nobutaka Hirokawa, Nobuyuki Nukina and Yasuo IharaFumitaka Oyama, Svetlana Kotliarova, Akihiro Harada, Mamoru Ito, Haruko Miyazaki,
are antagonized by tauGem GTPase and tau: morphological changes induced by Gem GTPase in CHO cells
published online April 15, 2004J. Biol. Chem.
10.1074/jbc.M401634200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2004/05/05/M401634200.DC1
by guest on February 6, 2018http://w
ww
.jbc.org/D
ownloaded from