סיכום לקורס ביולוגיה מולקולרית וביטוי גנים תשסג
TRANSCRIPT

: הגנום מבנה ותפקיד.1הרצאה
מספר עותקי הכרומוסום: קופי אחד של כל כרומוסום.–הפלואידי הומוזיגוט או הטרוזיגוט(,– שני עותקים )הומולוגים אך לא בהכרח זהים –דיפלואידי
של כל כרומוסום. לדוגמא הפרוקריוטיים וגם שמרים )יוקריוטי(. 2 מספר הסט של כל כרומוסום גדול מ- –פוליפלואידי .
לוקליזציה של הגנום: העובדה שהגנום ביוקריוטים הוא בגרעין גורמת לכך שכל תהליך שקורה בגרעין צריך לעבוריש שעתוק כדי )תוך מצומדים התהליכים בפרוקריוטים זאת לעומת לציטופלסמה אח"כ
תרגום(.
הגנום היוקריוטיהגנום הפרוקריוטידיפלואידי )לרוב(.הפלואידי
ליניארי )יש קצוות התחלה וסוף( אין נקודת התחלה וסוף–מעגלי
:c valueפרדוקס ה- C value –הכמות הסופית של דנ"א לכל תא הפלואידי באורגניזם. והפרדוקס הוא שאין
התאמה בין כמות הדנ"א ביצור לבין מורכבות היצור. למשל ליצורים פשוטים כמו בצל יש יותר דנ"א מיצורים מורכבים כמו הבני אדם, עובדה זו מראה שמה שחשוב זה לא כמות הדנ"א אלא
כמות הגנים ומה הם מקודדים.
:יהגנום המינימאלהמינימאל להתקיים יהסט ממנו אפשר גנים של תרגום, שעתוק,– על: גנים האחראיים
רפליקציה של דנ"א, תיקון דנ"א, חלבונים מלווים )יצירה של מבנה שלישוני לחלבון(, חלבונים האחראיים על מטבוליזם )יצירת אנרגיה(, אנזימים ליצירת מבנה התא, חלבוני ממברנה ועוד
גנים.256 גנים לא ידועים. סה"כ 18 נשים לב שגנים האחראיים על ייצור חומצות אמינו ונוקלאוטידים לא ברשימה, כך שאותם
האורגניזם יוכל לקבל מהסביבה.איך החליטו על הגנים ברשימה:
יכול להתקיים כתוצאה מכך אז הגן ואם החיידק לא פגעו בגן מסוים ע"י מוטציה הכרחי.
אם כן אז הכרחי.–נשווה בין כל החיידקים האם גן מסוים נמצא אצל כולם
תוכן הגנום:ולא מקודדים. האזורים הלא מקודדים הם כל רצפי הבקרה, הגנום ישנם חלקים מקודדים
junk –הפרומוטור והאזורים המבניים. אך ישנם גם אזורים לא מקודדים שלא שייכים לבקרה DNA.
אזורי קריאה פתוחים( : ORFדרכים לחיפוש גנים מקודדים )מציאת וחיפושו בגן או חיפוש החלבון וממנו נמצא את הגן עצמו. שיטות אלו mRNAע"י קריאת ה-
.) RNA )rRNA , tRNA מפספסות את כל הגנים אשר מייצרים ORFאשר מחפשות רק
Definitions:.RNA כל רצף הדנ"א הנחוץ לסינתזה של חלבון או מולקולת –גן

הם95% הם רצפי הבקרה, ו- 4%בגנום האנושי פחות מאחוז אחד מהגנום הם גנים מקודדים, junk DNA.
מפוזר באופן לאjunk DNAאין קורלציה לבין גודל הכרומוסום לבין מספר הגנים שבו, כך שה- אחיד בגנום.
יותר יש שביוקריוטים לב junkנשים DNAלעומתם גדול, הוא המקודדים הגנים ופיזור ופיזור הגנים יותר מצומצם.junkבפרוקריוטים יש פחות
(:transcription unitיחידות שעתוק )בפרוקריוטים הם האופרון גנים מקודדים סידור ביותר של גנים אשר–היחידות הנפוצות
מטרתם הוא מסלול מטבולי אחד ימצאו ביחידה אחת אשר לה אזור בקרה אחד. לדוגמא .trpסינתזת חומצת האמינו
התוצאה של העובדה שבפרוקריוטים הגנום מסודר באופרונים היא שמוטציה בודדת יכולה אופרון תהרוסtrpלשתק את הייצור של כמה חלבונים, למשל מוטציה באזור הבקרה של ה-
הפוליפפטידים האחראיים על יצירת חומצת האמינו טריפופן. )יצירת5את הייצור של כל זו תתרחש ע"י וביצה- לעוברknock outמוטציה זרע גן פגום לתאי של הגן ע"י החדרת
מתאים אלו יהיה את הגן הפגום(.
בגנים יוקריוטים לכל גן אפילו לכאלה אשר קשורים לאותו תהליך יש אזור בקרה משלו ו-mRNA.משלו
Cistron יחידה אשר מקודדת ל- פוליפפטיד בודד. ולכן האופרון נחשב להיות – polycistron. .monocistronic שרשרת פוליפפטידית אחת ולכן הוא נחשב –ביוקריוטים גן אחד
Junk DNA:סיבות לקיומו:
הדנ"א הלא מקודד הוא לא זבל אלא "מסך עשן" כלומר מוטציות מתרחשות כל הזמן ואם כל האזורים בגנון יהיו חיוניים המוטציות עלולות לגרום נזק, אם יש הרבה דנ"א לא מקודד הסיכוי לפגוע באזור מקודד ולגרום נזק יפחת מכיוון שיפגעו גם באזורים לא
מקודדים שם אין נזק. הגישה שאומרת שהחלקים הלא מקודדים הם אכןjunk.ואפשר להוציאם החוצה ה- –תיאורית הגן האנוכי junkהוא פרזיט אשר יש לו יכולת לשרוד בכל חלוקה וכל
עוד הוא לא מפריע ליצור הנושא אותו הוא ישרוד. -ל junk נותן נפח לדנ"א כך– יש תפקיד המיועד לטובת התא, תפקיד מבחינת מבנה
שתהליכים של מידור ביוקריוטים יתרחשו. התיאוריה לכך היא שרפליקציה של הדנ"א לוקחתjunkלמשל במיקרואורגניזמים אין הרבה
דנ"א שפחות כמה להם שיהיה היא מאד קטנים ליצורים שהשאיפה כך אנרגיה הרבה של היצוריםjunkוהתהליך ייקח פחות אנרגיה, לשם כך במהלך האבולוציה התקזז כמות ה-
הקטנים.
.ALU, LINE, SINE הם החזרות הארוכות. חזרות אלו יכולות להיות רצפי junkחלק גדול מה- מרכיבים כ- LINE1רצפי ה- מהגנום יש להם רצפי 25% LTRבקצוות והם מזכירים רטרו
וירוסים )וירוסים היוצרים דנ"א מ- רנ"א(.
חלוקת הגנים ביוקריוטים:הגנים המקודדים מתחלקים לשני סוגים:
גנים אשר מופיעים פעם אחת בסט הפלואידי, אלו הםsolitary gene.או גנים ייחודיים בדרך כלל הם מתבטאים בצורה שונה בין סוגים שונים של תאים, כלומר מבוטאים רק
בתאים מסוימים )למשל בתא כבד ולא בתא עצב(.

גנים אשר ישנם שניים או יותר עותקים שלהם בסט הפלואידי, העותקים לא זהים אך (. סטים של גניםgene duplicationדומים )גנים אלו כנאה מקורם באותו גן קדמון-
כ- גנים. משפחות הגנים בדרך כלל באים house keepingכאלו נקראים משפחות genes.)'שהם גנים אשר בלעדיהם התא לא יכול להתקיים )נשימה, מבנה, שעתוק וכו
גנים אלו מתבטאים בכל תא ותא. גן לא פעיל )כתוצאה ממוטציה או שחלק מהגן חסר(–בתוך משפחות גנים ישנו גם פסאודו גן
היושב בגנום והיה במקור פעיל. הגן לא מפריע ויש עותקים נוספים ממנו ולכן לא יעלם בתהליךהברירה הטבעית.
דוגמא למשפחת גנים הם גנים המקודדים ל-גלובינים, כל הסוגים השונים מקודדים לגלובינים הנושאים חמצן בדם, אך הם מראים שוני ביניהם אשר מתבטא בתפקידים שונים בפיזיולוגיה
של האדם.
Dose repetitions:ל- המקודדים הגנים מהאורגניזמים נמצאיםRNAבחלק מסוימים וחלבונים היסטונים, ,
. כלומר עותקים זהים של הגן יחזרוtandemly repeated arraysבמערכים החוזרים על עצמם אם צריך כמות גדולה של–אחד אחרי השני. זאת בשביל לספק כמות גדולה של תוצר הגן
RNA.לתרגום הגן אז צריך כמות גדולה של עותקי גן זה

ביטוי גנים בפרוקריוטים– 2הרצאה
מודל האופרון: גנים אשר ביטויים מופעל ע"י נוכחות סובסטרט מסוים, למשל הלקטוז מפעיל את ביטוי הגנים
.LACשל ה-אופרון אשר מפרק )למשל אופרון הלקטוז(, או מסלול אנאבולי –מאפשר מסלול קטבולי –
אופרון(.trpאופרון אשר בונה )
Lac Operon:ויוצר אנרגיה בתהליך גליקוליזה. מולקולת הלקטוז היא דו סוכר )חיבור חיידק מפרק גלוקוז גלוקוז וגלקטוז( אשר גם היא משמשת כמקור אנרגיה, חיידק יעדיף גלוקוז מאשר לקטוז )פירוק
הלקטוז לוקח אנרגיה( אך אם אין גלוקוז כלל אז יפרק את הלקטוז.
גנים מבניים וגן בקרה אחד:3אופרון הלקטוז בנוי מ- oZ אשר מקודד את gal β.)אנזים המפרק את הלקטוז לחד סוכר( oY אשר מקודד את permease.)מכניס את הלקטוז דרך הממברנה אל תוך התא( oA.לא ברור מה תפקידו oI -גן הבקרה, נמצא מחוץ לאופרון הלקטוז ומקודד את ה repressor.של האופרון
(, ובנויים מ:Iבנוסף ישנם אזורי בקרה אשר נמצאים לפני האופרון )בין הגנים המבניים לגן oPromoter –.רצך דנ"א אשר מהווה את האתר בו מתחיל השעתוק oOperator – רצף דנ"א אליו נקשר הרפרסור אשר הגן I.מייצר
באופרון זה הוא הלקטוז עצמו.inducerה-
האופרון עובד: איך הגן –כשאין לקטוז בתא Iמייצר את הרפרסור, הרפרסור יהיה פעיל ויתחבר לאופרטור כך
פולימרז אשר נמצא בפרומוטור לא יוכל להמשיך ולתרגם הלאה.RNAשה- גורם לשינוי מרחבי של הרפרסור–כשיש לקטוז בתא זה הלקטוז נקשר לרפרסור, קישור
, רנ"א פולימרז יכול להמשיך בעבודתו ולתרגם אתDNA –וכתוצאה מכך הרפרסור ניתק מה הגנים. מגיעים למקסימום פעילות האופרון ואח"כ יש ירידה הדרגתית בפעילות )ככל שהלקטוז
מפורק(. אך הוא עובד רק אם יהיה בתוך התא, וכדי להכניס את הלקטוז לתאinducerהלקטוז פועל כ-
ייוצר רק אם הלקטוז יהיה בתוך התא.premeaseצריך את הגן אשר שייך לאופרון אשר זו הוא שאין השתקה מוחלטת של האופרון )בערך ואז יש95%הפתרון לבעיה השתקה(
בשביל להכניס את הלקטוז לתא.premeaseמספיק
איך הבינו איך האופרון עובד:עשו ניסויים בהם השתמשו ב-
o גן אשר בנוי כך שאזור הבקרה שלו הוא האזור שרוצים לחקור, והאזור של–גן מדווח הגנים המבניים יהיה של גן אשר קל לראות אם הוא יוצר או לא. למשל חלק הבקרה
יהיה של האופרון עצמו אך הגנים המבניים יהיו של חומר שלאחר יצורו נצבע בכחול.oIPTG –חומר עם מבנה דומה ללקטוז, גם הוא עושה אקטיבציה לאופרון אך הוא
עצמו לא עובר פירוק.

oGal – X -ל סובסטרט מהווה gal βומפרקו, התוצר אותו זה מזהה כך שאנזים , המפורק מייצר צבע כחול כך שקל לזהותו.
אופרון: lac הפנוטיפים האפשריים של ה-oWild type –.האופרון מכובה כשאין לקטוז, האופרון עובד בנוכחות לקטוז oConstitutive –.)האופרון תמיד עובד )בלי או עם לקטוז oNon – inducible –.)האופרון תמיד מכובה )בלי או עם לקטוז
ן- IPTGוהניסוי שנערך: לקחו מושבת חיידקים וגידלו אותם על מצע שמכיל gal- Xרוצים , לראות אם בחיידקים יש מוטציות אשר ישתיקו את האופרון )המושבות יהיו תמיד לבנות כי אין
של לבנות gal –Xפירוק תמיד יהיו )המושבות לעבוד תמיד לו יגרמו או הסובסטרט–( מתפרק(.
התוצאות היו שאכן נוצרו מושבות חיידקים אשר היו רק כחולות, דבר זה יכול לנבוע מהסיבות:o.הרפרסור לא מיוצרo.הרפרסור מיוצר אך הוא בצורה שלא מסוגל להיקשר לדנ"אo אתר הקישור, האופרטור, השתנה.–הרפרסור בסדר אך משהו בבסיסי הדנ"א השתנה
בשביל לדעת האם המוטציה היא באופרטור או ברפרסור נעשה ניסוי בו עבדו על דיפלואיד החלקי )רק האזור הנחקר הוא דיפלואידי וזאת ע"י פלסמיד עם עותק נוסף תקין(.התוצאה היית
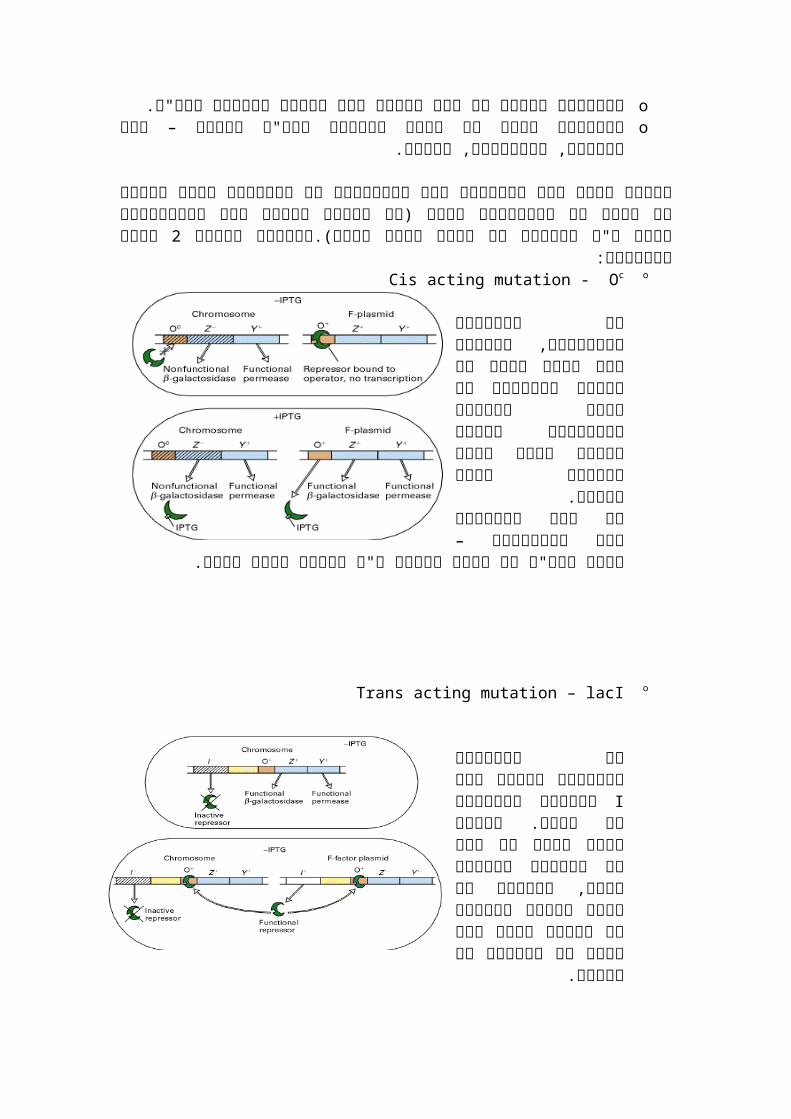
סוגי מוטציות:2oCis acting mutation - Oc
באופרטור, המוטציה פה אך תקין עותק עוד הוספנו יכול לא הרפרסור עדיין הפגום לאופרטור להיקשר המושבה ולכן הישן בעותק
תהיה כחולה.היא המוטציה שאם כך
ברצף הדנ"א אי–באופרטור הוספת ע"י לתקנה אפשר
עותק תקין.
oTrans acting mutation – lacI
ברפרסור המוטציה פה לייצור הרפרסורIכלומר הגן
תקין עותק נוסיף תקין. לא רפרסור שייוצר כך הגן של נמצא זה רפרסור תקין, לעותק גם מספיקה בכמות הישן וגם לחדש כך שתיקנו
את הבעיה.

Cis acting.משפיעה רק על גנים אשר באותם מולקולה. המוטציה היא ברצף הדנ"א :Trans actingמשפיעה על גנים באותה מולקולת דנ"א או במולקולה אחרת. המוטציה היא :
בחלבון כך שניתן לתקן בעזרת חלבונים מבחוץ.
:cAMP –בקרה חיובית נשאלת השאלה כיצד זה כשיש גם לקטוז וגם גלוקוז התא יודע להשתמש בגלוקוז, וכשריכוז
– cAMPהגלוקוז יורד התא עובר לפירוק לקטוז. התשובה מולקולת o-כשהגלוקוז גבוה ה( נמצא בתאים בקורלציה הפוכה לרמת הגלוקוזcAMP.)נמוך o.יוצר קשר בין רמת הגלוקוז לרמת הביטוי של אופרון הלקטוזo מולקולה פשוטה מדי בכדי לעבוד לבד, היא עובדת יחד עם מולקולתCAP.
ו- cAMPהשילוב של המולקולות CAPיוצר הפרומוטור אל קרוב נקשר אשר קומפלקס
ומזרז את הפעילות של רנ"א פולימרז. נמוכה cAMPכשהגלוקוז נוכח בתא כמות ה-
אם אין לקטוז אז הרפרסור מפריע לשעתוק, אך אם יש לקטוז ואין הפרעה של הרפרסור יתרחש שעתוק אך ברמה נמוכה כי אין זירוז
של רנ"א פולימרז. גבוהה ויש זירוז.cAMPכשאין גלוקוז אז רמת
)מדכא בעוד שהלקטוז עושה בקרה שלילית מבצע בקרהcAMPאת פעילות האופרון(, ה-
חוץ בעוד אופרונים גם קיימת חיובית, אשר מאופרון הלקטוז.
משמעות בקרת הגנים:ברירת המחדל של הגנום היא שגנים מבוטאים אלא אם כן הם מושתקים ע"י בקרה.
מהגנים פעילים )מבוטאים( כל הזמן זאת לשם חסכון באנרגיה.5%בחיידקים רק
עקרונות הבקרה:o לחלבון דנ"א בין פולימרז–אינטראקציה רנ"א מ- )מונע רפרסור כמו פקטורים
מלשעתק את הגן( או אקטיביטור )מזרז את פעילות רנ"א פולימרז(, הם אלו אשרנקשרים לדנ"א.
o הפקטורים )רפרסור או אקטיביטור( מגיבים אחד עם–אינטראקציה בין חלבון לחלבון השני ועם רנ"א פולימרז.
o.הסביבה החיצונים גם היא משפיעה
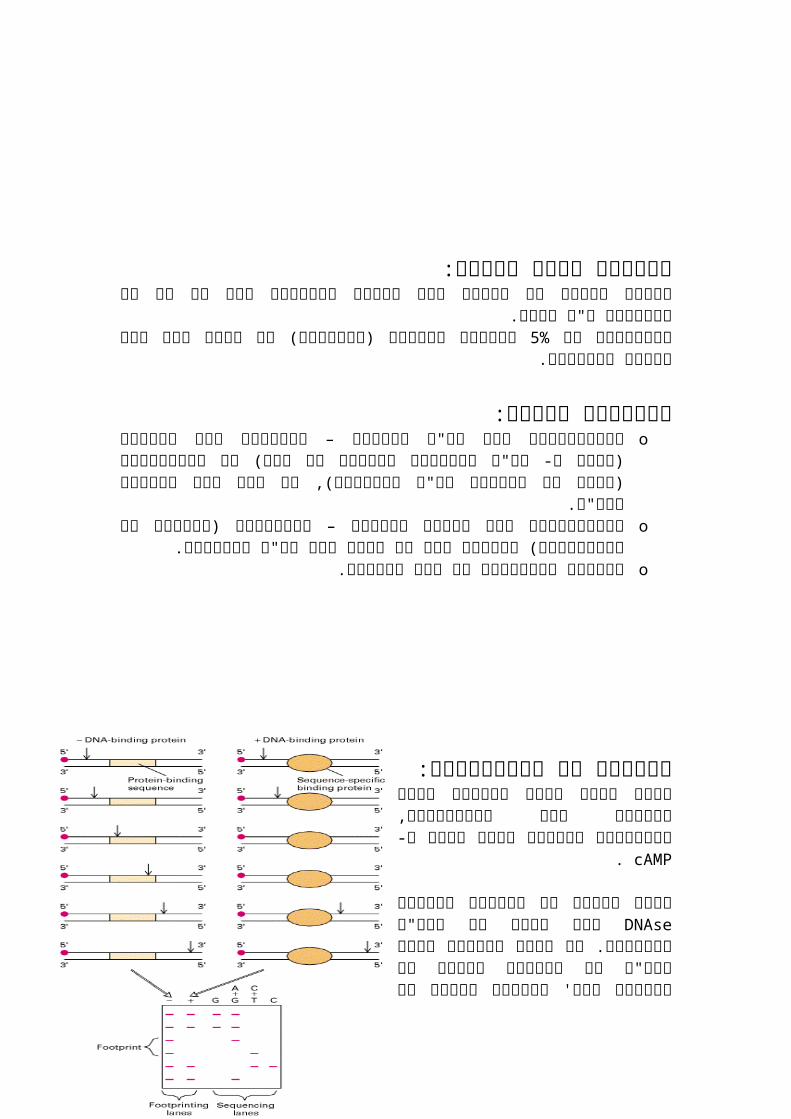
אנליזה של הפרומוטור: נרצה לדעת היכן נמצאים אתרי הקישור כמו הפרומוטור, האופרטור והמקום אליו נקשר ה-
cAMP.
אשרDNAseנעשה ניסוי בו השתמשו באנזים שובר את הדנ"א למקטעים. אם לפני השבירה נשים בדנ"א גם חלבוני שעתוק או רפרסור וכו'לאזורים פרט הדנ"א כל את ישבור האנזים זה הוא אתר לדנ"א מקום בהם חלבון קשור
קישור לחלבון. כתוצאה מניסוי זה נראה כי לאופרון הלקטוז יש
אתרי קישור אשר הם חופפים במקומם והם3– CAP.רפרסור ו- רנ"א פולימרז ,
בשביל היא הבקרה באזורי החפיפה שהרפרסור ימנע מ- רנ"א פולימרז מלהיקשר.
ב- -10-האזורים , קריטיים35 אזורים הם לרנ"א פולימרז ולכן מוגנים.
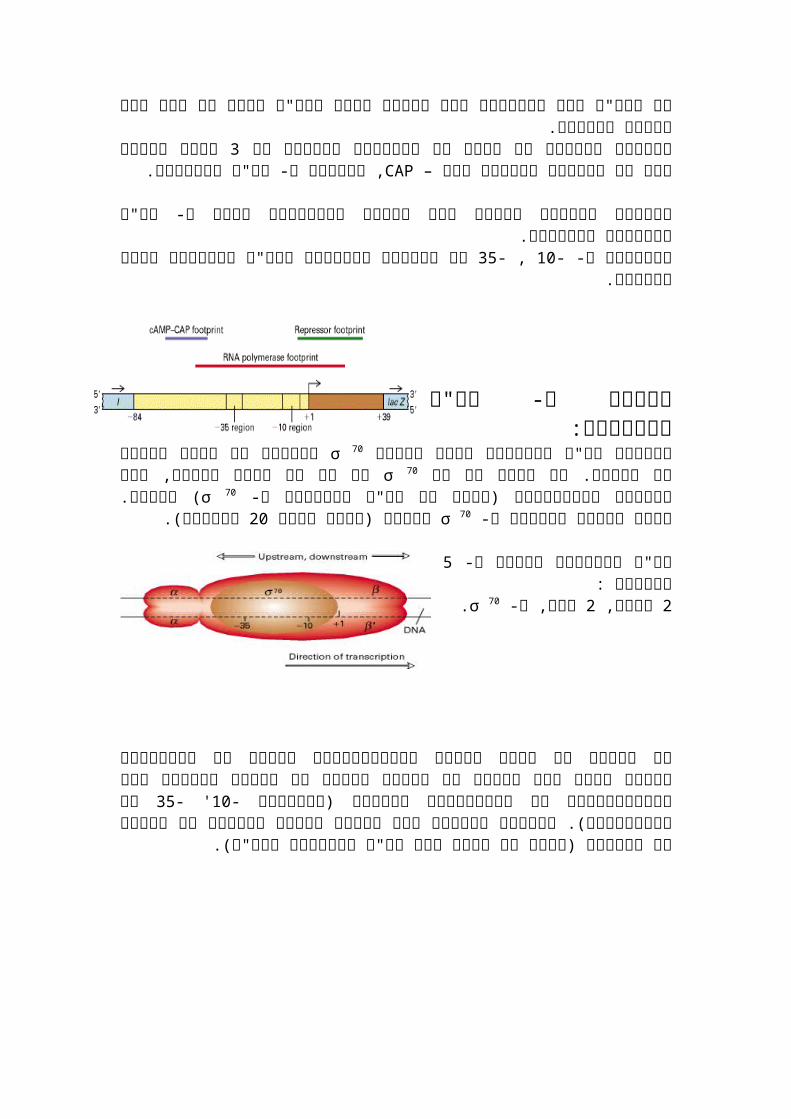
פעולת ה- רנ"א פולימרז: σ 70לפעולת רנ"א פולימרז נחוץ החומר
בלעדיו לא יהיה התחלה של שעתוק. אם גם כן לא יהיה שעתוק, מכך מבינים שהקומפלקס )שלוב של רנ"א פולימרז ו-σ 70נשים רק את
70 σ -70( הכרחי. אחרי התחלת השעתוק ה σ בסיסים(.20 מתנתק )אחרי בערך
יחידות :5רנ"א פולימרז מורכב מ- .σ 70 בטא, ו- 2 אלפא, 2
אם נסתכל על אתרי קישור בפרומוטורים שונים של אופרונים שונים נראה שיש שימור של 35' -10-רצפים כלומר יש רצפים חוזרים בין הפרומוטורים של האופרונים השונים )באזורים

של הפרומוטור(. מוטציה ברצפים אלו יכולה לפגוע בשעתוק או תגביר את השעתוק )יחזק אתהקשר בין רנ"א פולימרז לדנ"א(.
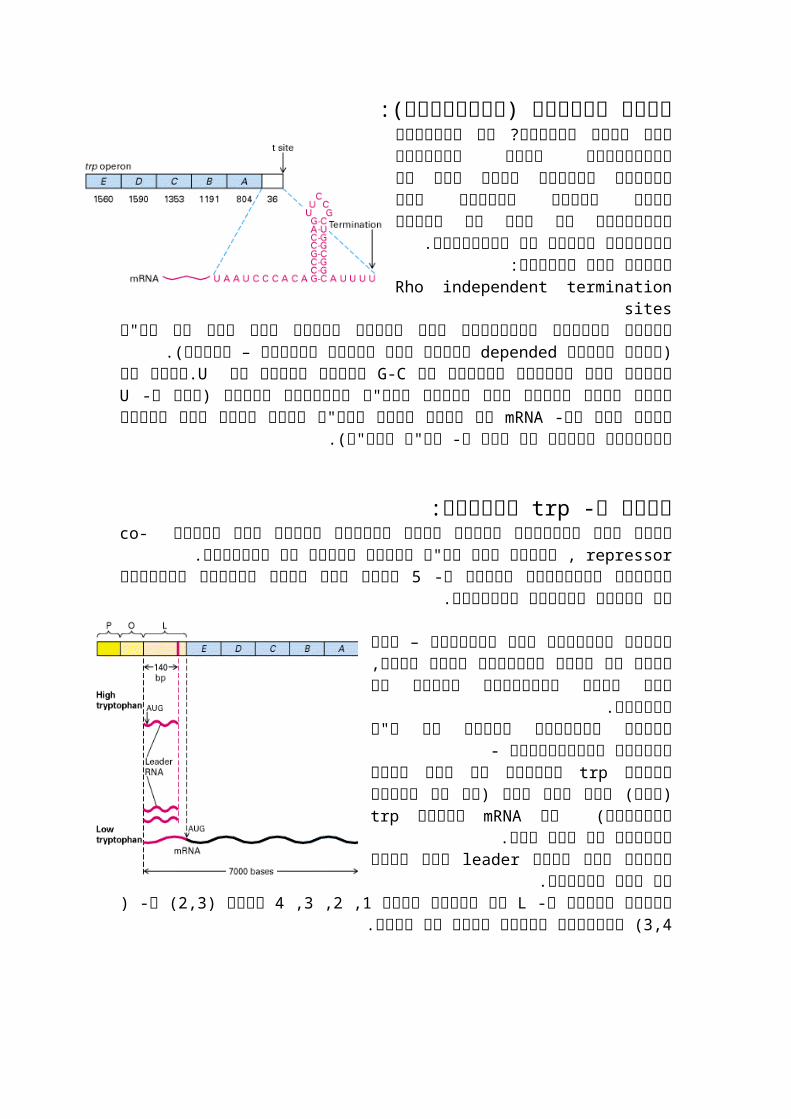
סיום השעתוק )טרמינציה(:אינדיקציה איזושהי יש השעתוק? נעצר מתי בסוף האופרון שאומרת שהגענו לסוף אלו הם אינם רצפים חוזרים בין אופרונים כי אלו הם
רצפים הקשורים למבנה לא לקודונים.רצפים אלו נקראים:
Rho independent termination sites כלומר מנגנון טרמינציה אשר הסימן לסיום הוא
הסימןdependedרצף של דנ"א )בעוד שבסוג אנזים(.–הוא פקטור חלבוני
ובסוף סידרה שלG-Cרצפים אלו עשירים בחזרות של Uמבנה זה יוצר מעין לולאה אשר. גורם לכך שה- Uגורמת לרנ"א פולימרז לעצור )רצף ה- mRNAלא יציב ולכן הרנ"א יורד
ממנו וכן המבנה השניוני מערער את קשר ה- רנ"א לדנ"א(.
אופרון:trpמודל ה- , החומר הוא בד"כco-repressorגנים אשר ביטויים מושתק בגלל נוכחות החומר אשר מהווה
המוצר הסופי של האופרון. גנים אשר כולם תורמים לסינתזה של חומצת האמינו טריפופן. 5אופרון הטריפופן מורכב מ-
היא הרפרסור פעיל רק–הבקרה העיקרית הוא כאשר טריפופן קשור אליו, הוא נקשר לאופרטור
וחוסם את השעתוק. השתקת האופרון נעשית גם ע"י מנגנון האטוניאציה
- משעותק גם רצף ארוך )מלא( וגם רצףtrpכשאין
וכשישmRNAקצר )רק עד הגנים המבניים( של trp.משעותק רק רצף קצר
רמתleaderהתגלה שיש מקטע על אשר שולט השעתוק.
4, 3, 2, 1 יש רצפים ששמם Lהתגלה שברצף ה- ( מסוגלים לבנות מבנה דו סיבי.3,4( ו- )2,3כאשר )
מנגנון האטוניאציה:
מקטע בשעתוק, מתחיל מכיל1הריבוזום . אם ישtrpכמה חזרות של חומצת האמינו
trpבתא הריבוזום ימשיך להתקדם אך בכך למקטעים הזדמנות מבנה3,4יש ליצור
שניוני ובכך נקבל מבנה טרמינציה והשעתוקיפסק.
בתא הריבוזום מתחיל לתרגם אתtrpאם אין ל- 1רצף )מחכה נתקע אך trpכך שיש )
– ליצור מבנה שניוני 2,3הזדמנות למקטעים ממשיך האופרון טרמינציה מבנה נוצר לא
להיות מתורגם.
= מבנה טרמינציה.U נותן מבנה שניוני שבסופו רצפי 3,4רק השילוב של המקטעים בעיה.– )הוספה או הורדה של בסיסים( נאבד את התזמון Leaderאם יהיה מוטציה ברצפי ה-
ביטוי גנים ביוקריוטים– 3הרצאה
בעוד שבפרוקריוטים מנגנון הבקרה של הגנים תלוי מאד בסביבה, ביוקריוטים יש מנגנון בקרה כלומר איזה גן מתבטא באיזו רקמה.– differential gene expressionפנימית וכן
בקרה על ביטוי גנים מתרחשת בכמה מקומות:oTranscription initiation –.האם יש בכלל שעתוק של הגן o אחרי השעתוק ישנם החלטות– splicing, localization, stability.o.) בקרה על שינוי החלבונים לאחר ייצורם )שינוי כימיo -בקרה האם יהיה תרגום שונה של הmRNA.
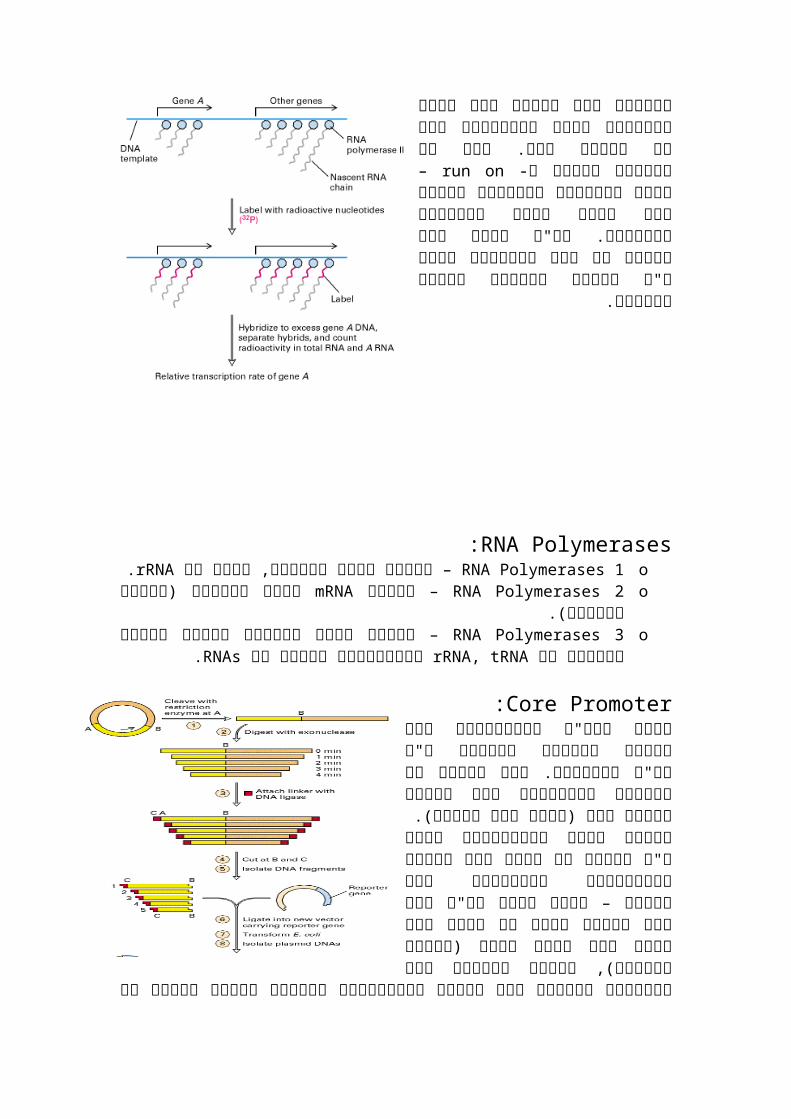
Run on analysis:מסוימים גנים האם לבדוק רצו בתחילה בכלל משעותקים ואם כן באיזה קצב. לשם
ה- לקחו– run onכך השתמשו בשיטת גנים לבטא להם ונתנו מבודדים גרעינים מחומרים מסומנים. אח"כ בדקו מהי הכמות של הגן הספציפי וזאת ע"י בדיקת נוכחות
החומר המסומן.
RNA Polymerases:oRNA Polymerases 1 – שעתוק בתוך הגרעין, יוצר את rRNA.oRNA Polymerases 2 – מייצר mRNA.)בתוך הגרעין )מקודד לחלבון oRNA Polymerases 3 – שעתוק מחוץ לגרעין מייצר סוגים נוספים של rRNA, tRNA
.RNAsומולקולות קטנות של
Core Promoter: אשר חיוני לתחילתימקטע הדנ"א המינימאל
השעתוק ע"י רנ"א פולימרז. רצף מסוים של בסיסים שבלעדיהם אין תחילת שעתוק כלל
)ובכך אין שעתוק(. מציאת הרצף המינימאלי נעשה ע"י ניסוי בו בדקו מהם רכיבי הפרומוטור שבלעדיהם אין
נקצץ מקטע דנ"א בכל פעם בגודל–שעתוק קבוע כך שבכל פעם יהיה לנו מקטע חשוד
המינימאלי שעדיין ונבדוק מהו המקטע לפלסמיד אלו נכניס מקטעים )המקטע המקוצץ(, מאפשר ביטוי או ביטוי חלקי, מזה אפשר ללמוד האם הפרומוטור מהווה אתר קישור אחד או
שהוא בנוי מכמה כאלו.
Core Promoter elements: :core promoterישנם רצפים אשר בד"כ נמצאים ב-
oTSS – transcription start site -אתר שממנו והלאה ייווצר ה mRNA.oTATA box – רצף קונסנזוס של TATAAA)A( -מתקשר ל TFIID בעזרת TBP
. מהווה אינדיקציה לרמ"א פולימרז מהיכן להתחילTTS- מה- 35 ל- 25-ממוקם ביו את השעתוק.
.initiation siteנמצא ברוב המקרים, אם לא קיים אז במקומו יש את ה-
oInitiation site –-כמו ה TATA -ל נקשר כן גם ,TFIIDנקשר הרנ"א וכן אליו מבנה: ישירות. כאשר YY)C(A)+1(N)T/A(YYפולימרז Y ,פירימידינים Nמייצג
.1 נמצא במיקום A- וה- 1 נמצא במיקום C הבסיסים, ה- 4מייצג כל אחד מ- o ישנם אתרים נוספים כמו 35- עד 35במרחקים +TFIIB -ו DPE -ה .DPEמצוי בד"כ
.TFIID ו- initiation site+ הוא חובר עם 32+ עד 28 מיקומו מ- TATAכשאין oCpG islands –לעיתים נמצא כי השעתוק בגנים מסוימים מתחיל באתרים שונים
השונים ביניהם בקצוותmRNAלאורך רצף מסוים של דנ"א, כתוצאה מכך מקבלים גנים אלו לא מכילים 5ה- אוTATA' )התחלת השעתוק כל פעם ממקום אחר(.
initiation site -אלא מכילים אזורים עשירים ב CG פקטור שעתוק בשם ,SP1מזהה .house keeping genesאת אזורים אלו ומשם מתחיל השעתוק. אלו הם לרוב
( ובכך למנועG ל- CH3 יש אפשרות לעשות מתילציה )הוספה של CGלרצפים של את שעתוק הגן, האיים הם גם כנראה לסמן אזורים לא פעילים בגנום.

Additional Promoter Elements: אלמנטים אשר גם מבקרים אתcis actingבנוסף לאלמנטים הקבועים בפרומוטר ישנם עוד
פעולתו של ה- רנ"א פולימרז והם:oProximal elements – בסיסים למעלה בזרם מה 200 – 100 אזורי בקרה השוכנים –
TSSבחלק מהמקרים אלמנטים אלו הם ספציפיים לסוג התא או ספציפיים לשלב , התא. במחזור
( השעתוק פקטורי את קושרים לפקטוריTFהם קישור אתר מהווים שהם כך ) השעתוק.
בניסוי שנערך נראה כי אם החסירו עד–המיקום שלהם בהשוואה לפרומוטור חשוב השפעה, אך כאשר הוסיפוה בסיסים במרחק בין האלמנט לפרומוטור לא היית20ל- ביטול האלמנט לחלוטין.ה התוצאה הייתTATA בסיסים בין האלמנט ל- 50 עד 30
oDistal elements – -גם כן משפיע על התחלת השעתוק אך ממרחק רב מה TSS.
Linker mutation analysis:
נרצה לדעת באיזה חלקיםהשונים הבקרה מאזורי עדיין והגן לפגוע אפשר יבוטא. נעשה ניסוי בו בכלעשו הבקרה באזור חלק גנים ובעזרת מוטציה מדווחים ראו איך הושפע
ביטויו של הגן.
: Additional Promoter Elements המשך oEnhancers –.גם הם אתרי קישור לפקטורי השעתוק ובתוכם יש אתרי קישור רבים
מצויים הם הגן, שעתוק על הגברה אפקט אלו לאתרים אוupstreamיש downstream -מה TSS.יכולים להיות בתוך אינטרון וכן אחרי האקסון האחרון בגן
(.cell type specificבד"כ הם פועלים לסוג תא מסוים ) מכיל כמה אתרי קישור ע"י פגיעה ברצף הדנ"א שלו כשהתוצאהenhancerגילו שה-
היא שאין ביטול של האלמנט ולכן הוא מורכב מכמה אתרים, מוטציה באזור מסויםיכולה לשנות את רמת ביטוי הגן )הגברה או הפחתה(.
ה- יצרו enhancerזיהוי בו ניסוי ע"י היה הראשון ה-2 עם האחד פלסמידים, enhancer-ובשני אין. בדקו האם תהיה הגברה של ייצור גן מסוים )בתא מסוים כי ה enhancerהוא ספציפי לתא( אם יכניסו פלסמידים אלו לתא, התוצאה היא שאכן בתא
היה עוצמה חזקה יותר של ביטוי הגן מהתא ללא ה-enhancerעם הפלסמיד עם ה- enhancer.
נשאלת השאלה איך רצף אשר נמצא רחוק מהפרומוטור משפיע על גן הרחוק ממנו? מאפשר קיפול של מולקולת הדנ"א כךenhancerקישור של החלבונים )פקטורי השעתוק( ל-
שאתרים הרחוקים אחד מן השני יכולים להתקשר. חלבונים אלו עכשיו קרובים לרנ"א פולימרזואינטראקציה זו היא שמזרזת את ייצור הגן.
Transcription Factors: )בקרה בצורת חלבון השולטת בגן אם הוא על כרומוסום אחד אוtrans actingחלבונים מסוג
)בקרה ברצף הדנ"א השולטת בגן רק אם cis actingיותר( אשר נקשרים לאזורי בקרה מסוג הוא על אותו כרומוסום( הם כמו הרפרסור או האקטיבטור בפרוקריוטים.
.enhancers או ל- proximal promoter elementsנקשרים ל- פקטורי השעתוק יכולים להיות אקטיבטורים או רפרסורים.
ניסוי בו נשים בדפנות של מבחנה רצף דנ"א שלו נרצה למצוא מיהם בכדי לזהותם נערוך הפקטורים שנקשרים אליו, נעביר חלבונים במבחנה, החלבון שנקשר זהו הפקטור שעתוק.
באותה צורה אפשר לעשות את ההרצה על ג'ל. או ליצור מוטציות ולראות מה משתנה.
אם נרצה לברר מידע על פקטורי השעתוק כמונעשה הגן ביטויו של מגבירים את בכמה הם המקודד הגן יהיה אחד בפלסמיד בו ניסוי ובפלסמיד אחר אתר הקישור לפקטור שעתוק של הפקטור וגן מדווח בשביל שנוכל לראות את
הפלסמידים לתא ונראה2ההשפעה. נכניס את את כמות הגן המדווח שיוצרה.
מבנה פקטור השעתוק: אתרים החוזרים על עצמם בפקטורי השעתוק:3יש
o.אזור בחלבון אשר נקשר לדנ"אo.אזור בחלבון אזר עובר אינטראקציה עם חלבונים אחריםoאזור אשר מבחינת המבנה המרחבי שלו מסוגל להעניק גמישות )אם היה קשה מבחינה
מרחבית הקישור לדנ"א לא היה יכול להיווצר(.

סוגי פקטורי שעתוק:כל פקטורי השעתוק מתחלקים לשני סוגים חלבונים והורמונים.
דרך לתא להיכנס מסוגל סטרואידי הורמון כל סטרואידים, הורמונים הם ההורמונים רב או להורמונים אחרים לעומתם בתוך התא, יהיו לסטרואידים הרצפטורים ולכן הממברנה
לחלבונים אשר לא מסוגלים להיכנס לתוך התא הרצפטור יהיה על ממברנת התא.
פעולת הסטרואידים: הם חודרים לתא בקלות לכן הרצפטורים בציטופלסמה או בגרעין, כשהסטרואיד מגיע הוא נקשר לרצפטור שעד עכשיו היה קשור אליו חלבון אחר. זאת יוצר שינוי מרחבי ברצפטור כךרפרסור מהווה שהרצפטור היא אחרת דרך לדנ"א. ולהיקשר לגרעין להיכנס יוכל שהוא
וכשהסטרואיד נקשר אליו הוא הופך לאקטיבטור.
פעולת הורמונים אחרים וחלבונים:–מולקולות גדולות אשר לא מסוגלות להיכנס לתא, לכן הם משתמשים במערכת תיווך תאית
signal transduction .מעבירים מסר חיצוני לתא בכדי שתתבצע פעולה מסוימת להיכנס למצב הגנתי ע"י: חלבון האינטרפרון נקשרTלמשל האינטרפרון אמור לסמן לתאי
לקינאז להפוך זה מאפשר דבר נגרם שינוי במבנה המרחבי של הרצפטור, ובכך לרצפטור לפעיל, הקינאז מבצע פוספורילציה של פקטור השעתוק, הפקטור הופך לפעיל ויכול להיכנס
לגרעין להיקשר לאתר הקישור הספציפי לו ולזרז שעתוק של הגנים האנטי ויראלים.

- מבנה כרומטין וביטוי גנים4הרצאה
מבנה הכרומטין:
ושל–כרומטין דנ"א של קומפלקס היסטונים.
קבוצה של חלבונים, משפחה של–היסטון גבוהה. בצורה מתבטאים אשר גנים הנקרא דנ"א של מארז ישנו מסביבם
נוקלאוזום.של–נוקלאוזום קטנה מבנה יחידת
הכרומטין אשר גרעינה הוא ההיסטון. אריזה של נוקלאוזומים אשר דחוסים יחדיו )חלק מהכרומטין(, קשה להביע–הטרו כרומטין
את רצפי הדנ"א באזורים אלו, למשל אזורים סביב הצנטרומרים.
אם בהיסטון אז יש גישה אליו וקל לשעתקו, אם בהטרו–היכן נמצא הגן משפיע את ביטויו כרומטין אז אין גישה אליו ואי אפשר לשעתקו.
Histone acetylation / deacetylation: (, קיומו של זנב זה אוlysine( יכול להיות קשור זנב אצתיל )קבוצות N – terminiלזנב ההיסטון )
לא משפיע על המטען של ההיסטון, הזנב נותן מטען חיובי להיסטון.Deacetylationכך יותר הדוקה הנוקלאוזומים בין האינטראקציה חיובי, המטען כאשר -
שלפקטורי השעתוק אין גישה ולכן אין שעתוק של הגן.Acetylation –המטען שלילי, מבנה הכרומטין פרוש יותר ונגיש, המרחקים בין הנוקלאוזומים
מאפשרים שעתוק.
הוכחה לכך שיש מעבר גן מאזור כרומטין להטרו כרומטין:

שמבוטא גן האם בדקו כן כמו נחפשו כאשר מסוימת, ברקמה ברקמה אחרת בה הוא לא מבוטא
נראה שהוא במבנה אחר.חותך דנ"א, אםDNase 1האנזים
הדנ"א לא נגיש או עטוף בחלבוניםאז הוא לא יחתך.
לקחו תא בו גלובין קיים והאנזים ניסה לעכל את הדנ"א האחראי על גלובין ואכן הוא עוכל, בעוד שבתא בו אין גלובין )לא מבוטא( האנזים לא עיכל את חלקי הדנ"א כלומר הדנ"א היה
במבנה אשר לא אפשר את עיכולו.
סוגים של מבנה דנ"א אשר משפיעים על השעתוק:2יש o כמה הדנ"א ארוז בצפיפות.–הטרו כרומטין o מתילציה תשב–דנ"א ציטוזום גבי על האם
המתילית הקבוצה של קיומה מתילית, קבוצה כלומר פקטורי השעתוק לא גישה לדנ"א מונע
יכולים להתחבר לדנ"א.
כרומוסומי המין: קובעY גבר כרומוסום ה- XY נקבה, XXהמין ביוקריוטים נקבע ע"י כרומוסומי המין. באדם
את המין.(.X0 קובע את המין הזכרי )Xיש מינים שהעדר כרומוסום
אין גןX הם עניים בגנים ביחס לכרומוסומים האחרים, לכרומוסום ה- X, Yכרומוסומי המין סוגי2 יש שילוב של Yהייחודי למין )כל גן שעליו יכול להיות בכל מקום בגנום(, לעומתו ב-
שנוכחותם מכתיבה את קיום המין,sex limit וחלקם גנים שהם Xגנים חלקם זהים לגנים ב- .Xעדיין כרומוסום זה מאד קטן ביחס ל-
אבולוציה של כרומוסומי המין:היונקים של במהלך–התיאוריה אוטוזומלים, כרומוסומים מזוג יצאו המין כרומוסומי
האבולוציה הופיעה על אחד מהם גן דומיננטי אשר קבע את ייחודיות המין. כעת יש התפצלות.sex limit אחד רגיל והשני נושא את התכונה –בין שני הכרומוסומים
בכרומוסום מסוימים באזורים הכרומוסומים שני בין רקומבינציה הגבלת התרחשה אח"כ הכרומוסומים יש רקומבינציה, אך בחלקים השונים אין.2כלומר בחלקים הדומים בין
sexלבסוף התרחשה צבירה של מוטציות ואיבוד מקטעי גנים גדולים על גבי הכרומוסום ה- limit כך שהרבה מהגנים על .Y -הם פסאודו גן, המנגנונים שהכתיבו את איבוד הדנ"א ב Yלא
ידועים אך הם היו מכוונים.
Dosage compensation strategies: עותקים מכל גן ואילו לזכרים עותק אחד. זהו מצב לא תקין ובמהלך2נוצרה בעיה- לנקבות יש
האבולוציה התפתחו מנגנונים לתיקון בעיה זו ע"י השוואת ריכוזים של הגנים בזכר ובנקבה:

o משתיקים את אחד העותקים של הנקבה כלומר הוא נדחס לגופיף–פתרון היונקים .barrהנקרא
o -ה- 2פתרון אשר מוריד בחצי את כמות הביטוי מ X ים כך שכל X.יבטא רק חצי o -פתרון שבו מגבירים את קצת ביטוי הX אצל הזכרים, בנקבות ה- 2 פי Xיהיה פעיל
כרגיל.לא משנה מה כמות הביטוי ובלבד שיהיה זהה ין זכרים לנקבות.
Female mammals X inactivation:נוצרת החלטה בעוברה איזה מה- ים יעבור אינאקטיבציה.Xבשליש הראשון של ההיריון
)פרט לתאים של מעטפות חיצוניות כמו השלייה שםיההחלטה מתרחשת באופן רנדומאל ה- הלאהXחייב זאת יעבירו התאים ההחלטה שנלקחה וברגע להתבטא( האם של
לצאצאיהם. בחתולים מסוימים לעיתים יש לנקבות מבנה
המיקום תלוי–פרווה מנומר )שחור וצהוב( אם למשל האינאקטיבציה התרחש בהיכן
ה- נבחר צהוב.Xבזנב אז הצבע האימהי לזכרים לעומת זאת יהיה צבע פרווה אחיד
הם קיבלו.Xתלוי איזה פגום אשר יגרום למוות התא,Xבמקרה ויש
איזה שהבחירה לאXנראה היא יושתק ה- עם התאים כי הפגוםXרנדומאלית
הפגום מושתק.Xימותו, ישרדו אלה עם ה-
מסוים מתבצעת:Xאיך ההשתקה של שבאמצעותו מתבצעת ההחלטה איזה מה- Xicבכל אחד מכרומוסומי המין יש אתר Xים
להשתיק. יש שני גנים:Xicבתוך oSense – מקודד בכיוון הנכון, מבטא את Xist.oAnti sense – מקודד בכיוון הנגדי, מבטא את Tsix.
אשר נצמד לכרומוסום ממנו נוצר וכך הכרומוסום הופך ללאmRNA נוצר Xistכשמבוטא ה- פעיל.
ישנן כמה פעולות הנדרשות בשביל שהאינאקטיבציה תתרחש:oCounting – סופרים את מספר ההשתקות שצריך לבצע – n-1 -כאלו לפי מספר ה X
השתקות וכך הלאה.XXY = 2 = השתקה אחת, XX השתקות, XY = 0ים. למשל o בחירה איזהX.יושתק o האינאקטיבציה עצמה ע"י מנגנוןXic.

Xist: mRNA אקטיבי בשני הכרומוסומים בצורה לא יציבה כך שה- Xistבהתחלה הגן שמבטא את
שנוצר מהגן השניmRNAהמיוצר משני הכרומוסומים לא יציב. אח"כ אחד מהגנים ייסגר וה- יתייצב.
פעיל הוא יושתק )Xistהכרומוסום שבו הגן נסגר הוא האקטיבי, ואילו הכרומוסום שבו ה- mRNA.)מתיישב על הדנ"א וחוסם אותו
Tsix: אשר בתחילה מבוטא בשני הכרומוסומים ואח"כ עובר לביטוי רק באחדmRNAיוצרת גם כן
מהם.Tsix נקשר אל Xist( כך שהוא לא יהיה פעיל יותר, בעצם ביטוי האנטי סנס Tsixמסייע )
בבחירה מי יהיה הכרומוסום המושתק.
X אקטיבי – Xist -לא פעיל כי ה Tsix.פעיל ונקשר אליו X לא אקטיבי – Xist ,פעיל Tsix.לא פעיל
אז הגן יהיה תמיד אקטיבי.Tsixאם יהיה גן בו יש מוטציה בפרומוטר של
:LINE 1רצפי ה- לאן השאלה ה- mRNAנשאלת של Xist -ה רצפי שאחוז נמצא נקשר? LINE 1
נראה עליהX ל- LINE. אם נוסיף רצפי Xic הוא מאד גבוה בעיקר באזורי ה- Xבכרומוסום ה- באינאקטיביות.
הם רצפים באורך LINEרצפי בסיסים בערך, הם חוזרים בגנום הרבה פעמים במגוון6000 מהגנום.16%מקומות, בבני אדם מהווים
DNA Methylation:ים הוא המושתק, הדנ"אXבעזרת המתילציה אנו מעבירים הלאה את המידע איזה מה-
עותקים מדנ"א זה, כל עותק2המושתק עובר מתילציה כך שבמהלך הרפליקציה הרי נוצרים X סימן לתא שזהו ה- –יהיה מסומן ע"י מתילציה רק בצד אחד שלו )רק בסליל דנ"א אחד(
המושתק ויש לסמן גם את סליל הדנ"א השני שלו. המתילציה האינאקטיבי. כמו רמה באותה לא אך מתילציה יש האקטיבי בכרומוסום גם
, בכרומוסום האקטיבי זה יהיה הפוך.Xistבכרומוסום האינאקטיבי תהיה בכל חוץ מאזור ה-
יש אזורים הומולוגים:Y ול- Xל - יש גנים זהים באזורים מסוימים, אזורים שבמהלך המיוזה יכולים לעבור רקומבינציהX , Yל-
בכרומוסום נקראים Yביניהם, אלו אזורים PAR1, PAR2בקצוות בעיקר מצויים והם הכרומוסום.
של הנקבה אז לגבר יהיה יותרX אם נשתיק את אחת מכרומוסומי ה- –הבעיה שנוצרה היא מנגנונים אפשריים לפתרון:3 לא מושתק, יש Yחומר גנטי כי
o -להשתיק גנים אלו גם בY.o -לבטא גנים אלו בX.המושתק o -להגביר את ביטוי הגנים בX.של הנקבה
ישנם גנים ב- 2המנגנון שנבחר אבולוציונית הוא מספר Xהמושתק שכן מתבטאים ושמם escapes.
Escape from X inactivation:

וגם ב-Xגנים הנמצאים גם ב- Y -בורחים מההשתקה ב Xרוב הגנים כאלו בורחים אך יש . או שהומולוג זה לא פעיל.Y כשאין להם הומולוג ב- X גנים אשר בורחים מ- –חריגות
הגנים הבורחים בד"כ יושבים בקבוצות השאלה היא איך הם בורחים?ישנם כמה מודלים אפשריים לבריחה:
o הרי רב הגנים הבורחים יושבים בקבוצות, כל הקבוצה בורחת יחד ואם–מודל אזורי ישים גן נוסף בקבוצה גם הוא יברח. עדות שמחזקת את מודל זה היא העובדה שרצפי
)אשר קשורים לאינאקטיבציה( הם נמצאים באזורים שבורחים בכמות דומהLINEה- 2לכרומוסום אוטוזומלי ולא בכמות גדולה כמו באזורים שאין בהם בריחה )שם יש פי
(.LINEרצפי oמודל שאומר כי ישנם אתרים בגנים אשר מכתיבים את יכולת הבריחה. עדות התומכת
במודל זה היא ההשוואה בין המינים, אם מודל זה נכון אז נצפה שאותם גנים בכלבורחים מהשתקה ובעכבר באדם גנים רואים שאותם ואכן בריחה יעברו היונקים
)בעכבר הגנים כבר לא בקבוצות אך הבריחהXלמרות שמיקומם שונה על גבי ה- נמשכת( זה מרמז שיש משהו על גבי הגני שגורם להם לברוח.
o הדביקו–המודל המשולב - שילוב של שני המודלים הללו. ניסוי שתומך במודל זה חלק מכרומוסום Xלכרומוסום עוברים בריחה4, נראה כי גם חלקי הכרומוסום 4
X הוא זהה ליחס של הבורחים בכרומוסום 4והיחס של הגנים הבורחים בכרומוסום )30%(.
נרצה לדעת מיהם הגנים הבורחים והאם יש השפעה לסוג רקמה: האינאקטיבי של האדם. עםX נוסיף לתא של עכבר את הכרומוסום –נשתמש בתא היבריד
גלאי המזהה את הגן ההומאני נוכל לבדוק האם יש גנים הומאניים שבוטאו כלומר ברחו. –אם נערוך אותו ניסוי על רקמות עכבר שונות נראה האם יש מאפיינים ספציפיים לרקמה
רק בחלק מהתאים התרחשה–האם יש גנים אשר יברחו רק ברקמה מסוימת, ואכן זה כך בריחה.
Gene imprinting – 5הרצאה
Epigenetic control of gene expression –הכוונה שינוי שעובר בתורשה בפנוטיפ הגן אשר .gene imprintingלא נגרם משינוי ברצף הדנ"א. דוגמא לכך היא ה-
Gene imprinting:
יש גן מהאב, 2לכל והשני מהאם אחד אללים Gene imprintingאחד שרק לכך גורם מהאללים בא לידי ביטוי, והאלל השני מושתק, השאלה היא מי מהאללים מושתק?
, בד"כתההחלטה איזה מהאללים עובר השתקה )מקור אבהי או אימהי( היא לא רנדומאלי הגנים האחראיים על הגדלת העובר )שלד, מסת שרירים( הם ממקור אבהי, והגנים האחראיים
.יעל שינויים במערכת העצבים ממקור אימהלמשל הגן Ifg2פקטור גדילה( פועל בעותק האבהי ומושתק בעותק האימהי, ולעומתו הגן(
Ifg2r.שהוא הרצפטור של הגן הקודם פועל באימהי ומושתק באבהי יצרו עובר שהוא שילוב של :ifg2בניסוי שנערך על הגן
o העובר מת כי לא היה עותק אבהי לפקטור – תאי ביצה 2הפריה של Ifg2.o י עובר נורמאל–תא זרע ותא ביצה.o תא זרע עםIfg2 אין – פגום ותא ביצה( העובר מת Ifg2.)תקין מצד האב
:imprintingמבנה הגנים שעוברים

( והם אלו שמציינים מיהו הגןDMRsבאלל המושתק יש אזורים בפרומוטר העוברים מתילציה ) בעוד שבגן האבהי יש,DMR1 נראה כי אין מתילציה ב- Ifg2rהמושתק. למשל בגן האימהי ל-
בעוד שבאבהי אין, בגן האבהי נוצרת מולקולת אנטי סנסDMR2אך באימהי יש מתילציה ב- .DMR2 שתפקידה לא ברור אך לשם כך אין מתילציה ב- DMR2מ-
בהם יש מתילציה שונה אשר מבקרים את ביטוי הגנים)imprinting centers )Icsמכילים ועוברים בתורשה.
)קבוצות קטנות(.clusters נמצאים ב- imprintingחלק מהגנים העוברים
שינויים ברמות המתילציה בזמן התפתחות תאי זרע או ביצית:
בזמן יצירת הגמטות אנו רואים שינוי ברמת המתילציה על גבי הגנום שלהן )מזהים את השינוי אם קל יותר אין מתילציה וההיפך(. השינוי הוא שבתחילה יש דה מתילציה–ע"י שקילת הגנום
כל הסימונים של המתילציה יורדים )בשביל להוריד את סימוני ההורים(, אח"כ יש מתילציה– בתאיםמחודשת המסמנת את המין )סימון הגנים האבהיים התאי זרע וסימון הגנים האימהיי
ביצית(. בכך המתילציה נותנת לנו דרך להבחין בין שני עותקים זהים של גנים, האם הוא אימהיאו אבהי.
יש שוב גל של דה מתילציה אך הפעם הגנים– ההפריה –ברגע שאוחדו תאי הזרע והביצית לא מושפעים ובכך התא יכול "לזכור" את המוצא ההורי של כל שניimprintingשכבר עברו
גנים זהים. הדה מתילציה השנייה לאחר ההפריה היא כנראה בשביל לקבוע את תפקיד התא)עצב, שריר וכן הלאה(.
רקמות עובר ורקמות חוץ עובריות. יש הבחנה– רקמות שונות 2כשעובר מתפתח הוא מייצר ביניהם בכמות המתילציה.
:imprintingתיאוריות המסבירות את קיום ה- o -בגלל הimprinting.חייבים רבייה מינית o ברגע שגן עבר –בדרך זו אפשר לבדוק האם כל הגנים נמצאים imprintingאז יש רק
עותק אחד ממנו וכך אשפר לבדוק האם כל סט הכרומוסומים בתא, למשל אם גן עברמוטציה והשני מושתק אז חסר עותק.
oParental conflict hypothesis –.משמעות אבולוציונית של מאבק בין זכרים ונקבות ביונקים מסוימים הנקבה יכולה להביא כמה צאצאים בהריון–הקונפליקט הוא כזה
אחד, וכן שיהיה כמה אבות לצאצאים אלו )באותו היריון(, לכן הרצון האבהי של כל אבואילו הרצון האימהי הוא שכל וישרוד, יהיה כמה שיותר חזק הוא שהצאצא שלו
העוברים ישרדו לא משנה מאיזה אב הם. ואילו הגניםimprintingכתוצאה מכך ב- יהיו להקטנת העוברים הגנים האימהיים
האבהיים יהיו לחיזוק והגדלת העובר. בעצם התיאוריה אומרת שצריך אבולוציונית הבדל בין מין מונוגמי )אב יחיד( שאינו
ולכן במין כזה נצפה לראות ירידה ב- imprintingצריך את ה- imprintingכי אין בו .imprintingצורך, לבין מין פוליגמי )הרבה אבות( שכן צריך

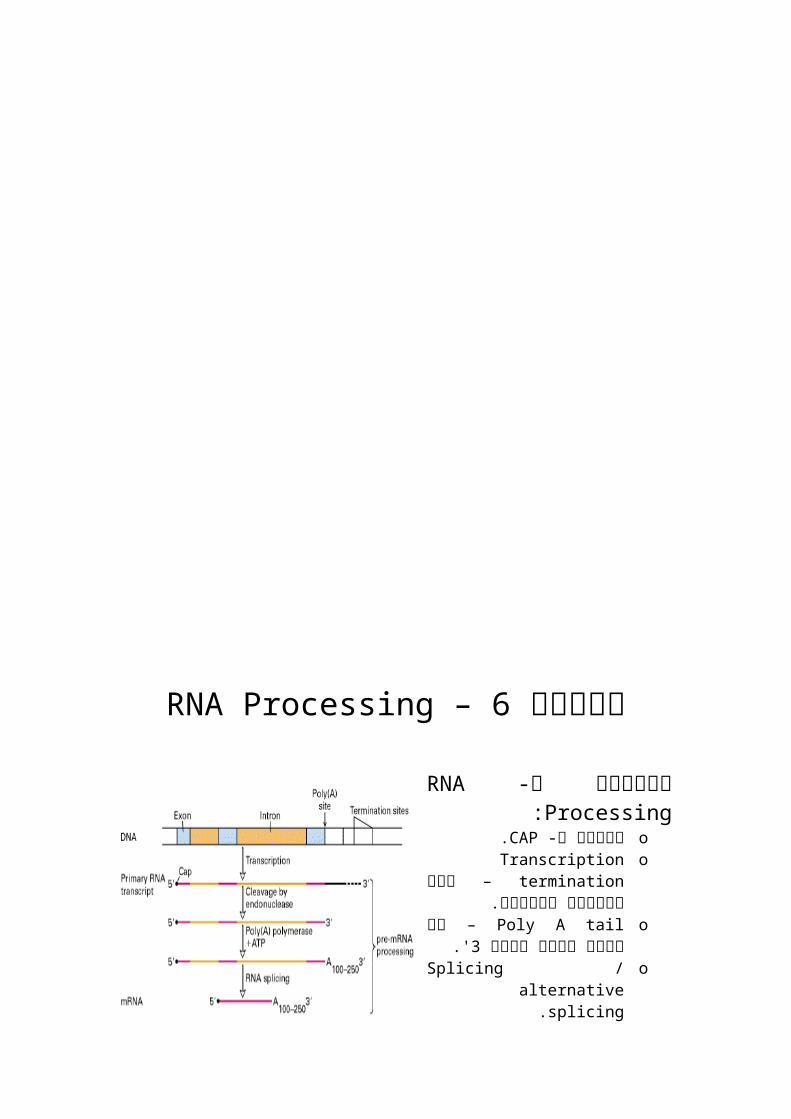
RNA Processing – 6הרצאה
:RNA Processingהשלבים ב- o -הוספת הCAP.oTranscription termination –
איך מסתיים השעתוק.oPoly A tail –לזנב קורה מה
'.3בקצה oSplicing / alternative splicing.oRNA editing.oCytoplasmatic transport.
בגרעין אינן מולקולות חופשיות:mRNAמולקולות ה-

מולקולת בגרעין שלב בשום מוצאים mRNAלא המגנים חלבונים אליה קשורים תמיד לבד, שהיא
נקראים אלו חלבונים חלבונים– hnRNPsעליה. קטנים יחסית אשר נצמדים לכל אורכה של מולקולת
כנראה לצמדhnRNP )כל סוג של חלבון mRNAה- לרצף מסוים במולקולה( , תפקידם הוא:
o -ה מולקולת על זו– mRNAלהגן בצורה המולקולה יציבה מבחינה כימית.
o-הם מונעים היווצרות של מבנים שניוניים ב mRNA.אשר יפריעו לתהליך
o -נראה שישנם תפקידים נוספים לhnRNPs פקטורי ע"י המזוהה למבנה תורמים הם
.mRNAהשעתוק, וכן יש להם תפקיד בהעברת ה-
CAP:ה- מולקולת את לזהות הוא שתפקידו בערך mRNAאנזים , תחילת30 אחרי בסיסים
' קבוצה אשר תגונן עליה.5השעתוק, ולהלביש עליה על קצה
Transcription termination: ' של המולקולה, למעלה הזרם של זנב זה ישנו גם רצף שמור3 בקצה poly A יש זנב mRNAל-
, זנב זה מסמן את סוף השעתוק, במחקרים נוספים התגלה שבנוסף לזנב זהAAUAAAשל למטה בזרם )כ- –ישנו עוד סימן ההכרחי לסיום השעתוק בסיסים( מ- 50 poly Aשם יש
.GUאזורים העשירים ב- מודלים המשערים איך סיום השעתוק מתרחש:2ישנם o -רנ"א פולימרז חש בסיגנלpoly Aמכך ניתקת היחידה החלבונית מהרנ"א פולימרז ,
ומפסיק השעתוק.o נוצרת מולקולתmRNA עם סיגנל של poly Aמרגע שסיגנל זה נחשף יש חיתוך של ,
ה' עד החיתוך והשניי5 מקצה – מולקולות האחת מה שצריך 2 מקבלים mRNAה- ' שלה5מהחיתוך עד איפה שרנ"א פולימרז המשיל לשעתק, מולקולה זו מעוכל מקצה
)הלא יציב( והשעתוק מפסיק.
בכדי שאכן יתבצע החיתוך ויתווסף יש צורך ברצפיםpoly Aזנב ה-
בתהליך:GUהחוזרים של נקשר לרצף השמורCPSFפקטור poly Aברגע שנתקל בו, פקטורים
בשם cleavageנוספים factors ומחברים אתGUנקשרים לרצפי
כתוצאה יחדיו. החלקים שני נוצר ומהאינטראקציה מהקיפול מבנה הזמין לאנזים היודע לחתוך
להוסיףmRNAמולקולת ויודע אדנין )הזנב(.
בניית הזנב

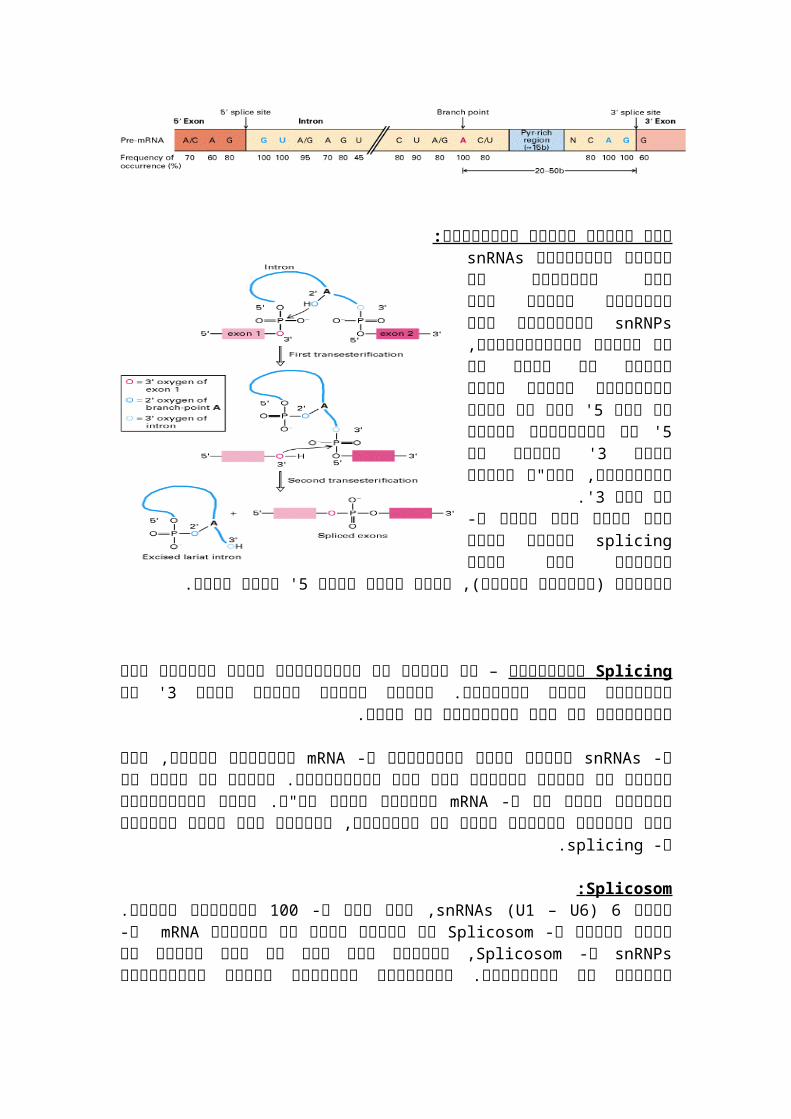
Splicing: הדרך היחידה של התא לדעת שקיים אקסון היא זיהוי האינטרונים, לרצף של אינטרון יש
splice sites 3 שאותם אפשר לזהות: קצה 'AG.......אזור מועשר בפירימידינים.........A.......GU 5 קצה.'
איך מתרחש חיתוך האינטרון:מולקולות אשרsnRNAsבעזרת
קטנים חלבונים עם מתחסרות ומרכיבים יחד אתsnRNPsבשם
יחידת הספלייסוזום, יחידה זו מזההאת קודם חותכת האינטרון את
שקצה 5קצה כך שלו של5' ' ' עדיין עם3האינטרון חופשי וקצה
האינטרון, ואח"כ חותכת את קצה3.'
splicingזאת בגלל שלא תמיד ה- מתרחש אחרי השעתוק אלא בזמןולכן מצומד(, )התהליך השעתוק
' יחתך קודם.5צריך שקצה
Splicingרקורסיבי לא מחכים– עד שהאינטרון כולו משעותק אלא
' של האינטרון הם אלו שמאפשרים3מורידים אותו המקטעים. רצפים דומים מספיק לקצה את הדבר.
ונקשרים אליהם, זהו תהליך של צימוד בסיסיםmRNA מזהים רצפי קונסנזוס ב- snRNAsה- ולגזור אותו אח"כ. רצפיmRNAבין שני המולקולות. צימוד זה נותן את היכולת לקפל את ה-
.splicingהקונסנזוס אלו נמצאים בתחילת וסוף כל אינטרון, מוטציה בהם תפגע בתהליך ה-
Splicosom: ישSplicosom חלבונים קטנים. אחרי יצירת ה- 100, וגם עוד כ- )snRNAs )U1 – U6 6מכיל
, היחידה הזו היא זו אשר מבצעתSplicosom ב- snRNPs ו- mRNAארגון מחדש של הזוגות את החיתוך של האינטרון. האקסונים מודבקים יחדיו והאינטרון משוחרר מהקומפלקס ועוברים
תהליך מחזור )אלא אם כן צריך את האינטרון לתפקיד כלשהו בתא(.

Self splicing introns: לבד. כלsplicing בשביל החיתוך, הם מבצעים Splicosomישנם אינטרונים אשר לא צריכים
האינטרונים הללו יש בתוכם רצף אשר מקודד להתקפלות המולקולה למבנה שניוני הדומה ל-Splicosom.
נוצרו והאינטרונים הקיימים היום כי אלו הם אינטרונים קדומים, מכך הועלתה התיאוריה מאינטרונים אלו ע"י תהליך בו הם איבדו את היכולת להתקפל למבנה שניוני בעצמם, ויצירת
snRNAs.אשר החליפו את יכולת זו
Alternative splicing: במקום להוציא רק את האינטרונים מוצאים גם יחידות של אקסונים, כך שיש כמה קומבינציות
. התהליך יתרחש בשני תנאים:mRNAלמולקולת ה- o( .וכן 1-2-3אי אפשר לשנות את סדר האקסונים אפשרי, אפשרי אך 1-3 לא2-1
אפשרי(.o -שני האקסונים החיצוניים המכילים את בקרות הstop codon , poly Aואת בקרות
ההתחלה חייבים להישאר. רק אם יש רצפים אחרים של אקסונים המכילים מידע לבקרה )עצירה או התחלה של שעתוק(, האקסונים החיצוניים יכולים להיחתך, אחרת
הם חייבים להישאר. Alternative splicingה-
חלבונים ע"י מבוקר )לרצף לרנ"א הנקשרים
ה- ליד splicingמסוים siteחלבונים ישנם .)
גישה לפקטורי המונעים כך שהרצףsplicingה-
וישנם יחתך, לא מזרזים אשר חלבונים ה- פקטורי פעילות את
splicingוהחיתוך יתרחש.
' של האינטרון3בתהליך זה החלבונים המזרזים חיתוך גורמים לכך שיהיה התעלמות מאתר ' של האינטרון השני.5הראשון ומאתר
קשור גם לסוג רקמה, כאשר כל חלבון הנוצר מסוג שונה שלAlternative splicingרואים ש- מאפיין רקמה מסוימת.splicingה-
RNA Editing: מנגנון בקרה המייצר שונות ביולוגית ברמת החלבון כלומר פה ההחלטה היא לא איזה חלקים
נוצר בו לא זהה למידע שבדנ"א. באזוריםmRNAיכנסו לתרגום, אלא רגיל אשר המידע בד"כ ההחלפות הן קבועות:–ספציפיים נוצרת החלפה של בסיסים
oC -מוחלף ל U.oA -מוחלף ל I.
עבר שינוי, הרצף לתרגום שונה כך שהחלבון יהיה שונה.mRNAכך שאחרי שרצף ה- תהליך זה קיים בצורה נפוצה במיטוכונדריה ובפטריות, הוא הופך מסגרת קריאה לא פתוחה
לפתוחה, כלומר מעבר מגן לא מקודד למקודד. הנוצר יהיהmRNA כך שה- stop codon היא החלפה של קודון מסוים ל- editingדוגמא ל-
קצר יותר מהמקורי.

)מכוונים את האנזים האחראי על התחלופה של editingהרבה מהרצפים המכוונים את ה- נמצאים בקצהeditingאתר ספציפי(, נמצאים באינטרונים לכן הרבה מהרצפים העוברים
האקסון )קרובים לאינטרון(. ישנו מנגנון המשמר רצפים אלו בתוך האינטרונים.ה- את המכוונים מסוג editingהמולקולות הן GRNA חלקים משני בנויות אשר חלק–
וחלק אשר יוצר מבנה שניוני עם רצפים כך שבמבנה זה החלקeditingהמתאים לאזור העובר שאין לו זיווג נביא לו בסיס.
RNA Stability Regulation – 7הרצאה
mRNA לציטופלסמה, נראה שיש שונות ביציבות בין mRNAלאחר שעתוק וחיתוך עובר ה- ,mRNA בתא נקבעת ע"י כמה ממנו משועתק וכן ע"י יציבות ה- mRNAשונים. כמות ה-
היציבות או חוסר יציבות יכולה להשפיע על כמה מהר אפשר להפסיק לייצר חלבון מסוים )mRNA תוך דקות נגמר ייצור החלבון(.– לא יציב של הבקטריות הם לא יציבים, גורם זה נותן לתא את היכולת להסתגל מהרmRNAרוב ה-
לשינויים סביבתיים המצריכים שינוי ברמת החלבונים )למשל הפסקת ייצור החלבון כי קייםביצורים רבmRNA של חלבון זה יהיה לא יציב(. רב ה- mRNAבסביבה החיצונית, לכן ה-
תאיים הם יציבים לאורך זמן רב.

לא יציבות נמצאו רצפיmRNAנרצה לדעת מה גורם ליציבות או חוסר יציבות, במולקולות .UTR' של 3 בקצה AUUUAקונסנזוס
היא יציבה למולקולה וכשהוסיפו אותם יציבות במולקולות נמצאו לא אלו מכיוון שרצפים איבדה את היציבות מבינים כי רצפים אלו הם שאחראיים הם חוסר היציבות של המולקולה.
של בטא גלוביןmRNAבניסוי הוסיפו ל- רצפי את יציבה, מולקולה היא אשר ירידה הקונסנזוס הלא רציפים, מקבלים דרסטית ביציבות מולקולת הבטא גלובין.
Iron Response Elements: פיזיולוגיה של ברזל היא דוגמא לכך שאפשר לבקר את יציבות זמן החיים של הגן )זמן מחצית
חיים ארוך או קצר(. אין בשום סיטואציה ברזל חופשי בגוף, הוא תמיד קשור לחלבונים כמו המוגלובין, בכל רגע נתון
צריך בקרה על כמות הברזל בגוף. החלבונים העוזרים לבקרה של כמות הברזל בתא:oTransferring –.מסיע את הברזל ברצף הדם o -ה ו-– Transferringרצפטור ברזל גבי התא, קושר את התרכובת של נמצא על
Transferring.ומכניס את הברזל לתא oFerritin –.אנזים הנמצא בתוך התא ומבקר על אגירת הברזל
בקרה על יציבות מולקולת הרצפטור:של רצפטור ה- mRNAב- Transferring יש בקצה ' רצפי חזרה של אי יציבות, כשרמת3
הברזל בתא נמוכה נראה שמולקולה זו מתייצבת. , אם הם חשופים אז מערכת של התא מזהה אותם ומעכלת אתIRE' נקראים 3הרצפים בקצה
המולקולה. או שהם מוגנים מפני עיכול., ויש לו שני מצבים:IRE-BPהחלבון האחראי על ההגנה נקרא
o הרבה ברזל בתא, החלבון קשור לברזל ולכן לא אקטיבי.–לא אקטיבי o ונקשר לרצפי ה- –אקטיבי ברזל, החלבון משתחרר מהברזל מעט IREומכך מגן
עליהם מעיכול.

בקרה על כמות חלבון האגירה: ', הם לא מקנים חוסר5 יש רצפים של אי יציבות בקצה Ferritin של מולקולת ה- mRNAב-
' היה חוסר יציבות(. אך נוכחות רצפים3' )אם היו בקצה 5יציבות למולקולה מכיוון שהם בקצה תהיה זמינהmRNA אליהם יקבע עד כמה מולקולת ה- IRE-BP' וקישור חלבון 5אלו בקצה
לתרגום.לחלבון הקושר שני מצבים:
o יש הרבה ברזל בתא החלבון יקשר אליו וישאיר את הרצפים חשופים,–לא אקטיבי ולכן המולקולה תתורגם )ובכך יהיה מולקולה שתאגור את הברזל בתא(.
o מעט ברזל בתא, החלבון ניתק מהברזל ונקשר לרצפים כך שה- –אקטיבי mRNAלא יתורגם.
Ala synthas הוא חלבון ferritin' ומבוקר כמו 5חלבון נוסף המכיל רצפים של אי יציבות בקצה בהמוגלובין )קבוצה הקושרת חמצן(. כך שיש הרבה ברזל,HEMהאחראי על יצירת קבוצת ה-
יש יצירת החלבון, מעט ברזל אין יצירת החלבון.
:mRNAחמצן כבקרה ליציבות VEGF –נותן סיגנל לחלוקת תאי אנדותל וגם סיגנל לתזוזה של התאים. במצב של חוסר חמצן
VEGF עובר הגברה בהיפוקסיה )חוסר חמצן(. העובדה ש- VEGFצריך לבנות כלי דם לכן הגן , וכן ייצובVEGF אירועים: הגברת שעתוק גן ה- 2עובר הגברה בחוסר חמצן היא תוצאה של
.VEGF של mRNAשל ה- נצפה לראות חלבונים הקשורים לרצפי האי יציבות–איך נדע שאכן הייצוב הוא בהיפוקסיה
בחוסר חמצן, ואת הרצפים חשופים )חלבונים לא קשורים( במצב רגיל של חמצן )כך שיהיהירידה בביטוי הגן בגלל החוסר יציבות(.
- חשפו תאים למצב רגיל שלmRNAבניסוי שנערך רצו לבדוק איך הייצוב משפיע על רמת על רמת4 עד 3. נראה כי הייצוב משפיע פי mRNAחמצן, וכן לחוסר חמצן ומדדו את רמת ה-
בחוסר חמצן. בניסוי זה מנעו שעתוק לחלוטין בשביל שלא ישבש את התוצאות.mRNAה- מפרידים– mRNA ,רצו לבדוק איך רמת השעתוק משפיע על רמת ה- Run Onבניסוי אחר,
את הגרעין משאר התא, מסמנים רדיואקטיבית את חומרי הגלם לשעתוק, ומסתכלים רק על אשר ישבש את התוצאות.mRNA שנוצרים. בצורה זו מנענו ייצוב של mRNAה-
בחוסר חמצן פיmRNAתוצאות הניסויים היא שהייצוב והשעתוק יחדיו מעלים את רמת ה- בערך מאשר במצב רגיל. כשהורדנו את רצפי החוסר יציבות מהמולקולה, גם במצב של12
)נפטרנו מהחוסר יציבות(.mRNAחמצן רגיל היתה עלייה של רמת ה-
:Cellular Sorting – 8הרצאה
יש חשיבות על מיקום החלבון כי זה משפיע על פונקציה מסוימת, אם התאים רוצים לייחדפעילוצ לאזור מסוים בתא אז הם צריכים למדר את החלבון.
קטגוריות של מיון חלבונים:2יש o.הפרדה לפי זמן - נשלט לפי השעון הביולוגיo.הפרדה מבחינת מיקום
דרכים:2, אשר נעשה ב- sortingאנו נתמקד בהבחנה של המיקום, הפרדה זו נעשית ע"י ה-

oלייצר חלבון במצב לא פעיל ורק כשיגיע למקומו להפעילו, למשל חלבונים גרעיניים המיוצרים בציטופלסמה ופעילים בגרעין ע"י פוספורילציה.
o להעבירmRNA.לאזור בו נרצה חלבון פעיל, ורק שם לתרגמו
RNA export:ומובלות החוצה אל הציטופלסמה, זאת בתהליך אקטיבי מולקולות רנ"א מתורגמות בגרעין
הצורך אנרגיה. הממברנה של הגרעין צריכה לאפשר העברה דו כיוונית של מולקולות גדולות, בחלק הפנימי
של הממברנה קיימת הלמינה אשר תפקידה להגן על מולקולות הדנ"א.mRNAs מובלים מהגרעין החוצה דרך תעלה הנקראת pore complexאלו הם מבנים גדולים ,
. nucleoporinsהמורכבים מחלבונים הנקראים ויציאה, הסיגנל ויצאו בתעלות הממברנה הם צריכים "קוד" כניסה בכדי שהחלבונים יכנסו
NES והסיגנל יציאה, סיגנל הוא NLSלהרבה ומיקום(. )כניסה לגרעין כניסה סיגנל הוא "סוחבים" חלבונים יש את שני הסיגנלים, חלבונים כאלו הם חלבוני מטען אשר חלבונים
קומפלקס של חלבונים והסיגנל נחשף רק כאשר החלבונים מסודרים יחדיו אחרים )לעיתים יש מונע כניסה / יציאה של החלבונים בנפרד(.–
ויצרים קומפלקס אשר הוא זה שנושא אתhnRNRחלבוני המטען חוברים עם חלבונים בשם סיגנלי היציאה או הכניסה.
אשר מכילים את הסיגנל hnRNPsמולקולות הרנ"א מתחברות ל- NESקומפלקס זה נקרא mRNPמוטציה באזור קישור של הרנ"א לחלבון תשאיר את החלבון הגרעין, מוטציה בחלבון ,
עצמו תשאיר את הרנ"א בגרעין.איך ההעברה מתבצעת:
של ה-CAPישנו חלבון הנקשר לאתר mRNA אשר נקרא CAPBPהוא מזהה ,
אשר ההגנה שלהםmRNAרק את ה- )הרנ"א אשר הםCAP' היא 5על קצה
'5רנ"א ריבוזומלי אין להם הגנה על קצה ' של5ולכן לא יוצאו מהגרעין(, קצה ה-
המולקולה יוצא קודם. ברגע שהקומפלקס חוזרים בחזרהhnRNPsיצא מהגרעין ה-
לגרעין למשלוח נוסף.מולקולות אתmRNAרק סיימו אשר
לצאת יכולות והחיתוך השעתוק תהליך מהגרעין.
mRNA localization:ה- את אתmRNAמסיעים שקובעים אזורים אותו, מתרגמים שם ורק מסוים לאתר
, הרצפים יוצרים מבנים שניוניים אשר מזוהיםmRNA' של ה- 3הלוקליזציה נמצאים בקצה עובריםmRNA לאתר המתאים. מרבית ה- mRNAע"י חלבונים שתפקידם להעביר את ה-
)מעכבים תרגום עד שצריךtimingעיכוב תרגום במהלך ההסעה ואפילו באתר, זה כחלק מה- את החלבון(.

mRNA', אז עדיין ה- 3אם נעשה מוטציה אשר לא תפגע במבנה השניוני של הרצפים בקצה ימוקם לאתר המתאים.
Localizer elements: , מבנם יכול ליצורUTR של 3האלמנטים אשר אחראיים על הלוקליזציה נמצאים בד"כ בקצה
מבנה שניוני. ואת השפעתם על הגנים אלפא אקטין בו רצו לראות מיהם רצפי הלוקליזציה ניסוי נערך הנמצא מסביב לגרעין ו- בטא אקטין הנמצא בקצוות התא. ניקח את רצפי הלוקליזציה של
מולקולות אלו ונצמידם לגן מדווח נקבל את התוצאות:o לגן מדווח לא היה שום לוקליזציה היה פיזור אחיד של אלפא5כשהצמדנו את קצה '
ובטא אקטין בכל התא.o נראה צביעה באתרים ספציפיים בתא 3כשהצמדנו את קצה לגן מדווח אלפא–'
מסביב לגרעין ובטא בקצוות התא, אכן הייתה לוקליזציה.
הקשר בין לוקליזציה להתפתחות עוברית: התמיינות התאים מתא מקור אחד לכמה תאים ברקמות שונות.– differentiationתהליך קביעה של צורת האברים. – morphogenesisתהליך
בטן. אם צירים אלו ישובשו נוכל– זנב, גב – צירים: ראש 2כל התפתחות עוברית מוכתבת לפי למשל לקבל יצור דו ראשי.
Positional Information:היכולת של התא לדעת את המיקום שלו ולרכוש לעצמו זהות בהתאם.
כשעובר נמצא בשלבים הראשונים )כמה תאים(, אם ניקח תאים ממקום שאמור להיות רקמת ראש ונעבירם למקום המייצג רקמת גפיים התא אכן יחליף את תפקידו. אך אם נעשה זאת בשלבים מאוחרים יותר התא לא יחליף תפקידים ובאזור הרגלים יצמח ראש. מכך אנו מסיקים
שלמיקום התא יש השפעה על תפקיד התא.מיקומו זהו התא תפקיד את שקובע מה עוברית בהתפתחות מוקדמים בשלבים למעשה
בקומפלקס. פקטורי שעתוק שהתגובה– מורפוגניםהחומרים האחראיים על העברת המידע המיקומי הם
אליהם היא תלוית ריכוז כלומר הגן יופעל או לא לפי ריכוז המורפוגן. לכן לשם יצירת הבדלבמידע בין התאים השונים ניצור הבדל בריכוז המורפוגנים בתאים השונים.
נשאלת השאלה איך יוצרים ריכוזים שונים של המורפוגן בתוך תא אחד? מי שאחראי על האי אשר מושתלים בביציתmaternal mRNA בשם mRNAסימטריה בריכוזים הם קבוצה של
לקודד הוא ותפקידם העוברי, בתא ספציפיים במקומות מושתלים הם ההפריה, לפני עוד למורפוגנים כך שהעובר יתפתח לא יהיה כמות זהה של מורפוגנים בכל התאים.
– תאים תומכים מהם הם 15 תאים שאחד מהם הוא הביצית ו- 16ביצית בעצם מורכבת מ- המטרנלי במקומות מתאימים בציטופלסמה שלmRNAאשר תפקידם הוא לשתול את ה-
הביצית בצורה לא סימטרית.
מטרנלי האחראיות על התפתחות העובר:mRNA קבוצות של 4ישנם o.קבוצה הקובעת את ראש העוברo.קבוצה הקובעת את זנב העוברo.קבוצה הקובעת את הקצוות של בראש ושל הזנבo בטן.–קבוצה הקובעת את ציר זנב
:hunchbackוגן ה- bicoid גן ה-
mRNA המטרנלי הראשון שהתגלה נקרא bicoidהוא עובר לוקליזציה לראש העובר, חוסר בגן של גן זה במרכז נראה סגמנטיםmRNAזה ייתן התפתחות לא תקינה של העובר. אם נשתול
כפולים של ראש זנב. ועלה ריכוזו. bicoid יופעל רק לאחר שהופעל גן ה- hunchback המטרנלי mRNAהגן של ה-
לו hunchback –הפרומוטור של מראה שיש ל- 3 אתרי קישור bicoid,גבוה עם קישור ושלושה כאלו עם קישור נמוך. רמת הקישוריות של האתר קובעת את הריכוז המינימאלי של
bicoid .הנדרש לזרז תרגום הגן bicoidבניסוי שנערך בנו פרומוטורים סינטטיים שבאחד אתרי קישור עם קישוריות נמוכה ל-
עם קישור אתר עם ואחד קישוריות גבוהה.
ע שבגן רואים ואכן הקישוריות עם הפרומוטור יותר ייצר הגבוהה
hunchbackעם זה מאשר הקישוריות הנמוכה.
שיהיה שככל הסיקו בנוסף bicoidיותר אתרי קישור ל-
ה- גבי שלmRNAעל hunchbackיותר ייווצר hunchback.
בתא:hunchback מקורות של 2ישנם oHunchback -אשר מקורו הוא משעתוק זיגוטי אשר מבוקר ע"י ריכוז ה bicoidוהוא
נמצא רק בראש העובר. oHunchback -אשר מקורו מ mRNA.מטרנלי ואשר מפוזר בצורה שווה בכל העובר
בכל העובר ולא רק בראש, אך באמתhunchbackלכן היינו מצפים לראות פיזור של חלבון ה- בשם חלבון שישנו הוא לכך הסבר העובר. בראש רק מצוי זה עוברnanusחלבון אשר
המטרנלי אשר נמצא גם בזנב.hunchbackלוקליזציה לזנב ותפקידו למנוע את תרגום ה- Nanus של ה- 3 נקשר לרצפים בקצה 'hunchback ומוריד את זנב ה- פולי A -כך שה mRNA
לא יוכל לעבור תרגום.נקבל עובר שאין לו זנב כי הגן של nanusאם בעובר יש מוטציה ל- hunchbackהמטרנלי
נבטל את בזנב. אם ה- nanusהתבטא וגם את hunchback-ונשאיר רק את ה המטרנלי hunchback.הזיגוטי העובר יתפתח כרגיל

Hunchback ולעיתים פועל יחד עם גם הוא מורפוגן bicoidגנים שונים וגורם להתבטאות , הוא מופעלhunchback מופעל מריכוזים שונים של kruppelבמקומות שונים בעובר. למשל
אך גם מתחת לריכוז מסוים שלו, כלומר בין ערכיםhunchbackגם מעל ריכוז מסוים של מסוימים של ריכוז.