국 립 암 센 터 원 장 귀 하 (첨부서류) 출간논문...
TRANSCRIPT

- 1 -
<붙임 4>
기관고유연구사업 최종보고서
연구분야(코드) T-3 과제번호 1110110 지원프로그램
창의(일반연구)과제
과제성격(기초,응용,개발) 기초 실용화 대상여부 비실용화 공개가능여부 공개
연구과제명 (국문) RNA수준에서 특이적 전사후 조절에 의한 발암기전 연구: RNA결합-수식단백질을 중심으로 (영문) Post-transcriptional Regulation of Tumorigeneis on the RNA Level: RNA-Binding Proteins and RNA-Modifying Enzymes
과제책임자 소 속 분자종양학연구과 직 위 선임연구원성 명 김종헌 전 공 분자생물학
세부과제구분 세부과제명 세부과제책임자
성명 소속(직위) 전 공123
총 연구기간 2011년 1월 ~ 2013년 12월 (총 3년)
참여연구원수(단위: 명, MY) 3.2
연구기간 및연구비
(단위:천원)
구분 연 구 기 간 계 국 립암센터
기 업 부 담 금소계 현금 현물
계 2011년 1월 1일~ 2013년 12월 31일 3 395,000
제1차 2011년 1월 1일~ 2011년 12월 31일 1 130,000
제2차 2011년 1월 1일~ 2012년 12월 31일 1 145,000
제3차 2011년 1월 1일~ 2013년 12월 31일 1 120,000
참 여 기 업 명 칭 전화 FAX기관고유연구사업관리규칙에 따라 본 연구개발사업을 성실히 수행하였으며 아래와 같이 최종보고서를 제출합니다.
2013년 10월 30일
과제책임자 김 종 헌 (서명)국 립 암 센 터 원 장 귀 하(첨부서류) 출간논문 초록목록

- 2 -
목 차
< 요 약 문 >
(한글) ---------------------------------------- 3
(영문) ---------------------------------------- 4
1. 연구의 최종목표 ------------------------------ 5
2. 연구의 내용 및 결과 -------------------------- 5
3. 연구결과 고찰 및 결론 ------------------------ 26
4. 연구성과 및 목표달성도 ----------------------- 28
5. 연구결과의 활용계획 -------------------------- 31
6. 참고문헌 ----------------------------------- 32
7. 첨부서류 ----------------------------------- 33

- 3 -
연구목표<최종목표>종양 마이크로RNA의 turnover 기전 연구
<당해연도목표>PARN에 의한 종양억제 마이크로RNA let-7의 deuridylation 기전연구
연구내용 및 방법
1. PARN에 의한 종양억제 마이크로RNA let-7의 deuridylation 기전 규명- RNA결합단백질 HuR이 Lin28B-TuT4 oligo-uridylation 복합체의 구성분자로동정됨.- HuR은 마이크로RNA let-7의 가공을 가속시키는 조절자로 작용함.- HuR의 oligo-uridylation된 전구체 let-7 마이크로RNA에 특이적으로 결합함.- HuR은 핵심 마이크로RNA 가공기구인 Dicer 복합체와 결합하며 3′-5′deadenylase PARN이 그 복합체 상에 존재함.- 3′-5′ deadenylase PARN은 uridylation된 전구체 let-7 마이크로RNA에 대해 3′-5′exo-deuridylase activity를 지니고 있음.- PARN의 과발현은 전구체 및 성숙 let-7 마이크로RNA의 발현을 증대시킴.- Lin28B의 S221 인산화는 PARN을 oligouridylation complex에서 해리시킴.
2. TRIM71-Lin28B-let-7 negative regulatory axis에 의한 종양억제기전규명- Lin28B분자는 세포 내에서 유비퀴틴화됨.- TRIM71은 Lin28B C-말단에 ~50 아미노산의 serine/lysine이 풍부한 서열에특이적으로 결합하며 이러한 부위가 없는 Lin28A는 결합을 하지 못함.- TRIM71은 post-Drosha 과정에서 Lin28B protein의 발현저해와 이에 따른let-7 가공-생성과정을 조절함.- TRIM71의 RING finger motif가 Lin28B의 유비퀴틴화, 결합, 그리고 let-7 조절에 매우 중요함.- TRIM71의 Lin28B에 의해서 유도되는 cellular transforming 잠재력을 억제함.
3. 뇌신경교종에서 DDX23 RNA헬리케이즈에 의한 종양 마이크로RNA-21의가공조절 연구- 종양억제유전자 PTEN의 tandem affinity purification 및 LC/MS/MS 기법을통해 종양 마이크로RNA-21 가공 전사후 조절인자로서 DDX23의 분리동정- 뇌신경교종환자의 조직에서의 DDX23의 발현 및 생존율이 매우 유의적임.- DDX23은 뇌신경교종세포주의 침윤과 증식을 증대시킴.- DDX23은 뇌신경교종세포주에서 종양 miR-21의 발현을 조절함.- DDX23은 종양 miR-21 발현을 Drosha 단계에서 조절함.- DDX23은 Drosha microprocessor와 pri-miR-21과 특이적으로 결합함.- DDX23의 RNA헬리케이즈 활성이 종양 miR-21 특이적 가공조절에 중요함.
연구개발에 따른
기대성과
<정량적 성과>
구분 달성치/목표치 달성도(%)
SCI 논문 편수 7/2 350%
IF 합 20.8/8 260%
<정성적 성과>1) 종양마이크로RNA발현 및 조절에 관여하는 RNA수준에서의 특이적 전사후조절인자의 조직적 분석시도2) 발굴된 RNA수준에서의 특이적 전사후 조절인자에 의한 종양마이크로RNA발현 및 분자기구의 총체적인 연구가능
색인어국문
전사후 조절 암화과정 RNA결합단백질
RNA수식단백질 마이크로RNA
영문
Post-transcriptional control Tumorigenesis RNA-Binding Proteins
RNA-Modifiers microRNA
< 요 약 문 >

- 4 -
Project Summary
Title of ProjectPost-transcriptional Regulation of Tumorigeneis on the RNA Level:
RNA-Binding Proteins and RNA-Modifying Enzymes
Key WordsPost-transcriptional Control, Tumorigenesis, RNA-Binding Proteins,
RNA-Modifiers, MicroRNA
Project Leader Jong Heon Kim
Associated Company
The expression of let-7 is majorly regulated on the posttranscriptional level. RNA-bindingprotein Lin28 and 3′ terminal uridyltransferase TuT4 have essential role for this process, whichdestabilize precursor let-7 (pre-let-7) through anchoring (Lin28) and addition of oligomeric uridine (U) tail(TuT4) at the 3′ end. To identify additional component(s) which potentially modulates oligo-uridylationof pre-let-7, tandem affinity purification was assessed with the Lin28B. The RNA-binding protein HuRwas identified as one of the novel interaction partner. Overexpression of HuR significantly enhances thelevel of precursor and mature let-7 but not primary form. Moreover, knockdown of HuR by specificsiRNA shows reduced level of let-7. Surprisingly, well known poly(A)-specific ribonuclease PARN whichis associated with the Lin28B-TuT4-HuR oligo-uridylaton complex has unconventional 3′ to 5′exo-deuridylase activity toward oligouridylated pre-let-7. HuR recruited all the components for theoligo-uridylation (Lin28-TuT4), dicing (Dicer), and deuridylation (PARN). More interestingly, specificphopshprylation of S221 residue of Lin28B expel PARN from this complex. These data suggest thatpost-transcriptional regulation of let-7 biogenesis is not a simple and unidirectional mechanism. It issurprising to find that 3′ terminal uridyltransferase TuT4, and 3′ to 5′ exoribonuclease PARN areboth present in the same RNP complex and that their opposing activities modulate oligomeric U taillength of pre-let-7. Taken together these data suggest newly identified HuR and 3′ to 5′exoribonuclease PARN potentially act as critical regulators of let-7 biogenesis including tumorigenesisstep. Moreover, we showed that the Lin28B protein level is regulated via ubiquitin-mediated proteasomaldegradation, and identified the ubiquitin ligase as human TRIM-NHL domain-containing TRIM71. In cells,TRIM71 negatively regulates Lin28B protein stability by catalyzing polyubiquitination. Compared with itsparalog, Lin28A, a C-terminal unique ~50 amino acid stretch of Lin28B is essential for TRIM71interactions and subsequent polyubiquitination. Moreover, the N-terminal RING finger motif of TRIM71 iscritical for protein-protein interactions and polyubiquitination of Lin28B, and consequent let-7 expression.Consistent with the let-7 stimulatory role of TRIM71 via Lin28B polyubiquitination, specific knockdown ofTRIM71 led to downregulation of let-7 expression. Expression of one of the known let-7 targets,HMGA2, was derepressed after knockdown of TRIM71. Moreover, overexpression of TRIM71 suppressedcellular transforming potential of Lin28B. We additionally showed that enhanced expression of let-7 ispart of a feedback loop that targets TRIM71 3'UTR, which contains two conserved let-7 target sites. Ourfindings collectively reveal critical aspects of regulatory complexity of let-7 biogenesis at theposttranscriptional level.
Aberrant miR-21 expression is closely associated with cell proliferation, anti-apoptosis,migration, invasion, and metastasis in various cancers. However, the regulatory mechanism of miR-21biogenesis is largely unknown. Here, we demonstrated that the tumor suppressor PTEN negativelyregulates the expression of oncogenic miR-21 at the post-transcriptional level. Moreover, our resultssuggest that PTEN plays such a role through the indirect interaction with the Drosha complex. Toelucidate how PTEN regulates pri- to pre-miR-21 processing, we attempted to find PTEN-interactingproteins and identified an RNA-regulatory protein, RNH1. Using the sensor to monitor pri-miR-21processing, we demonstrated that RNH1 is necessary and sufficient for pri-miR-21 processing. Moreover,our results propose that the nuclear localization of RNH1 is important for this function. Further analysisshowed that RNH1 directly interacts with the Drosha complex and that PTEN blocks this interaction.Taken together, these results suggest that the PTEN-mediated miR-21 regulation is achieved byinhibiting the interaction between the Drosha complex and RNH1, revealing the previously unidentifiedrole of PTEN in the oncogenic miR-21 biogenesis. we also identified novel RNA processing moleculessuch as DDX23 and hnRNP L by tandemn affinity purification. Dead box RNA helicase DDX23expression is up-regulated and essential for the survival pattern of glioma patients. DDX23 promotedinvasion and proliferation of glioma cell lines. More interestingly DDX23 regulates miR-21 biogenesis atthe post-transcriptional level. DDX23 interacts with core microprocessor Drosha and promotes pri-miR-21processing and the helicase activity of DDX23 is essential for the invasion and migration of glioma celllines and together with miR-21 processing. These results suggest the possibility of microRNA processingby DDX23 implicates clinical importance of microRNA processing in cancer progression.

- 5 -
1. 연구의 최종목표
<최종목표>
종양 마이크로RNA의 turnover 기전 연구
<2012년도 목표>
PARN에 의한 종양억제 마이크로RNA let-7의 deuridylation 기전연구
2. 연구의 내용 및 결과
가. PARN에 의한 종양억제 마이크로RNA let-7의 deuridylation 기전 규명
- 선행 연구를 통해 암의 발생, 진행, 전이, 억제에 중요한 마이크로RNA의 가공과정에 있어서 다
양한 RNA결합-수식단백질들 역할 조사하기 위해서 in vitro 상에서 마이크로RNA의 가공과정을
모방할 수 있는 기법을 확립 조직적 분석을 시도하였음.
- 가공조절 분자기전이 다소 알려져 있는 Lin28B-TuT4 oligo-uridylation 복합체의 추가 조절분자 동
정을 위해 FLAG-tagging한 Lin28B분자를 발현시킨 뒤 affinity purification과 MALDI-TOF 분석을 통
해 HuR 분자를 조절후보자로 동정하였음.
- 두 가지 동정방법을 통해 선별-확인된 HuR [(embryonic lethal, abnormal vision, Drosophila)-like 1
= ELAVL1 (human antigen R)]은 mRNA의 AU-rich 서열에 결합하는 분자로 잘 알려져 있으며 다양
한 종간 (인간에서 zebra fish까지 비교하였을 경우)에 그 아미노산 서열이 매우 높게 보존되어있으며
이를 근거로 생체 내에서 매우 중요한 역할을 수행하는 것으로 예상 할 수 있음.
- FLAG-tagging한 Lin28B분자의 affinity purification과 MALDI-TOF 결과를 재확인하기위해 다시
FLAG-Lin28B의 co-immunoprecipitation(IP)후 HuR과의 결합을 그리고 FLAG-HuR의 co-IP후
Lin28B와 결합을 immunoblot으로 재확인 할 수 있었음.
그림 1. RNA결합단백질 HuR이 Lin28B-TuT4 oligo-uridylation 복합체의 구성분자로 동정됨.
(A) FLAG-Lin28B의 co-immunoprecipitation(IP)후 coomassie brilliant blue 염색을 통한
단백질체의 확인. (B) FLAG-Lin28B의 co-IP후 HuR과의 결합을 immunoblot으로 재확인. (C)
FLAG-HuR의 co-IP후 Lin28B와 결합을 immunoblot으로 재확인.

- 6 -
- 다음으로 HuR과 함께 pri-let-7a-1 발현시켜서 Northern blot 분석을 통해 마이크로RNA의 가공과정
을 검증하였을 때 HuR의 경우 마이크로RNA의 가공과정에서 전구체와 성숙형태 모두 공히 증대시키는
것을 관찰할 수 있었음 (그림 2).
그림 2. HuR은 마이크로RNA let-7의 가공을 가속시키는 조절자로 작용함. (A) 293T 세포주에
서 pri-let-7a-1과 HuR의 과발현에 의한 마이크로RNA let-7의 생합성 영향에 대한 Northern
blot 결과 (좌). Northern blot의 정량화 (중). 각각의 RNA결합-수식단백질들의 발현양상에 대
한 immunoblot 분석결과 (우). (B) HuR의 knock down은 let-7a의 발현을 특이적으로 감소시킴 .
(C) HuR은 let-7 표적인 암유전자 HMGA2의 발현을 조절함.
- HuR의 마이크로RNA의 가공과정을 활성화 시키는 능력이 이미 보고된 KSRP 분자보다도 강력한 것
을 알 수 있었으며 HuR이 마이크로RNA의 가공을 촉진시키는 활성자로서 결론 내릴 수 있었음.
- HuR을 세포에서 특이적으로 제거했을 때 마이크로RNA의 가공과정에서의 촉진현상에 어떠한 영향
을 미치는지 조사하였음. HuR을 성공적으로 knockdown 시킬 수 있었으며 이에 따라서 성숙한 let-7a
의 발현도 감소함을 확인할 수 있었음. 흥미로운 점은 대표적인 종양 마이크로RNA인 miR-21의 경우
그 발현이 HuR의 knockdown영향을 받지 않는 것을 볼 때 HuR은 let-7 마이크로RNA를 특이적으
로 조절함을 일차적으로 파악할 수 있었음 (그림 2).
- 다음으로는 마이크로RNA let-7의 세포 내 표적분자로 알려진 high-mobility group AT-hook 2
(HMGA2)의 3′UTR을 Renilla luciferase의 뒤쪽에 cloning을 진행하여서 실제 HuR의 세포 내 표적에
대한 영향을 조사하였음.
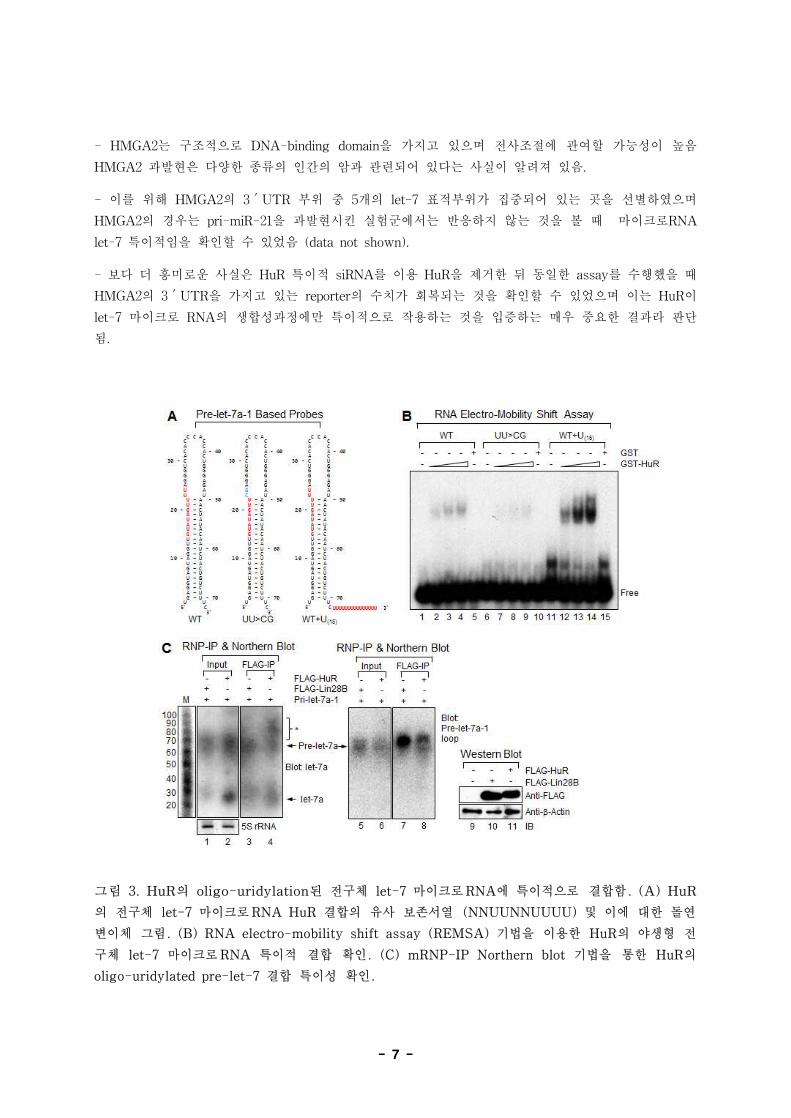
- 7 -
- HMGA2는 구조적으로 DNA-binding domain을 가지고 있으며 전사조절에 관여할 가능성이 높음
HMGA2 과발현은 다양한 종류의 인간의 암과 관련되어 있다는 사실이 알려져 있음.
- 이를 위해 HMGA2의 3′UTR 부위 중 5개의 let-7 표적부위가 집중되어 있는 곳을 선별하였으며
HMGA2의 경우는 pri-miR-21을 과발현시킨 실험군에서는 반응하지 않는 것을 볼 때 마이크로RNA
let-7 특이적임을 확인할 수 있었음 (data not shown).
- 보다 더 흥미로운 사실은 HuR 특이적 siRNA를 이용 HuR을 제거한 뒤 동일한 assay를 수행했을 때
HMGA2의 3′UTR을 가지고 있는 reporter의 수치가 회복되는 것을 확인할 수 있었으며 이는 HuR이
let-7 마이크로 RNA의 생합성과정에만 특이적으로 작용하는 것을 입증하는 매우 중요한 결과라 판단
됨.
그림 3. HuR의 oligo-uridylation된 전구체 let-7 마이크로RNA에 특이적으로 결합함. (A) HuR
의 전구체 let-7 마이크로RNA HuR 결합의 유사 보존서열 (NNUUNNUUUU) 및 이에 대한 돌연
변이체 그림. (B) RNA electro-mobility shift assay (REMSA) 기법을 이용한 HuR의 야생형 전
구체 let-7 마이크로RNA 특이적 결합 확인. (C) mRNP-IP Northern blot 기법을 통한 HuR의
oligo-uridylated pre-let-7 결합 특이성 확인.
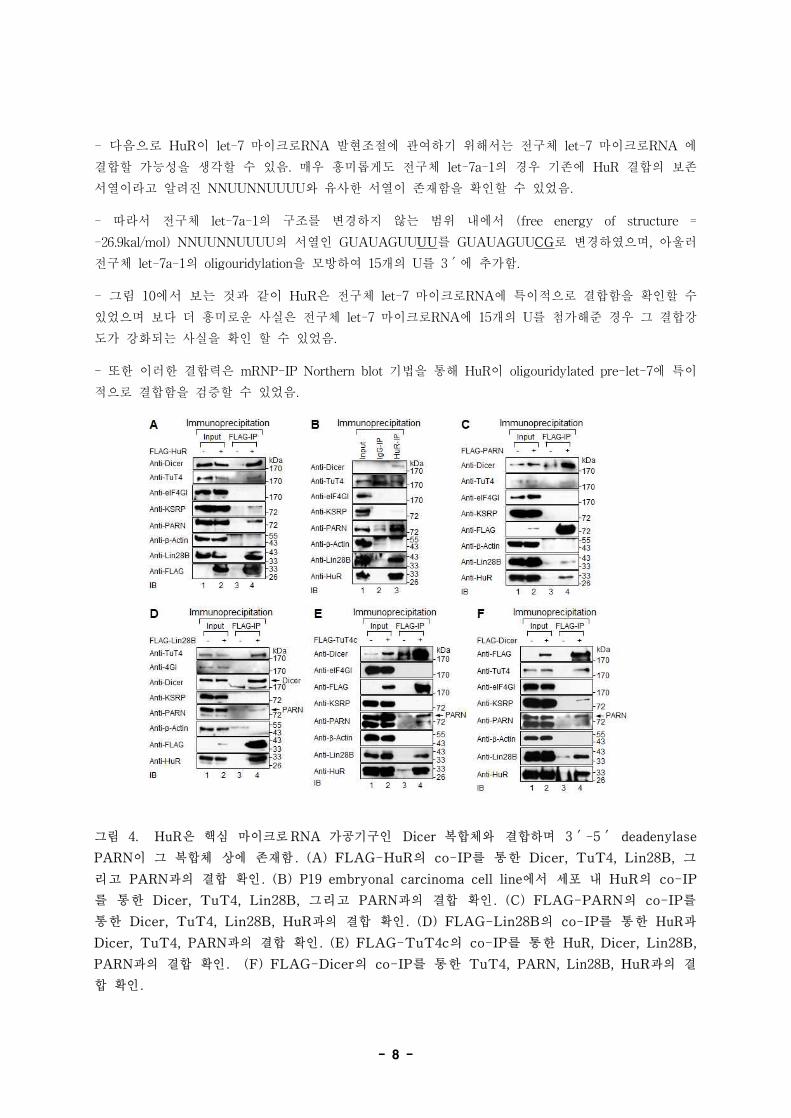
- 8 -
- 다음으로 HuR이 let-7 마이크로RNA 발현조절에 관여하기 위해서는 전구체 let-7 마이크로RNA 에
결합할 가능성을 생각할 수 있음. 매우 흥미롭게도 전구체 let-7a-1의 경우 기존에 HuR 결합의 보존
서열이라고 알려진 NNUUNNUUUU와 유사한 서열이 존재함을 확인할 수 있었음.
- 따라서 전구체 let-7a-1의 구조를 변경하지 않는 범위 내에서 (free energy of structure =
-26.9kal/mol) NNUUNNUUUU의 서열인 GUAUAGUUUU를 GUAUAGUUCG로 변경하였으며, 아울러
전구체 let-7a-1의 oligouridylation을 모방하여 15개의 U를 3′에 추가함.
- 그림 10에서 보는 것과 같이 HuR은 전구체 let-7 마이크로RNA에 특이적으로 결합함을 확인할 수
있었으며 보다 더 흥미로운 사실은 전구체 let-7 마이크로RNA에 15개의 U를 첨가해준 경우 그 결합강
도가 강화되는 사실을 확인 할 수 있었음.
- 또한 이러한 결합력은 mRNP-IP Northern blot 기법을 통해 HuR이 oligouridylated pre-let-7에 특이
적으로 결합함을 검증할 수 있었음.
그림 4. HuR은 핵심 마이크로RNA 가공기구인 Dicer 복합체와 결합하며 3′-5′ deadenylase
PARN이 그 복합체 상에 존재함. (A) FLAG-HuR의 co-IP를 통한 Dicer, TuT4, Lin28B, 그
리고 PARN과의 결합 확인. (B) P19 embryonal carcinoma cell line에서 세포 내 HuR의 co-IP
를 통한 Dicer, TuT4, Lin28B, 그리고 PARN과의 결합 확인. (C) FLAG-PARN의 co-IP를
통한 Dicer, TuT4, Lin28B, HuR과의 결합 확인. (D) FLAG-Lin28B의 co-IP를 통한 HuR과
Dicer, TuT4, PARN과의 결합 확인. (E) FLAG-TuT4c의 co-IP를 통한 HuR, Dicer, Lin28B,
PARN과의 결합 확인. (F) FLAG-Dicer의 co-IP를 통한 TuT4, PARN, Lin28B, HuR과의 결
합 확인.

- 9 -
- 따라서 HuR은 전구체 let-7 마이크로RNA 말단 oligo-uridylation 전후 Lin28B, TuT4와 같은 분자들
과 특이적 관련성을 가지고 let-7마이크로RNA의 생합성과정에 관여함을 예상할 수 있음.
- HuR이 전구체 let-7 마이크로RNA와 성숙 let-7 마이크로RNA를 증대시키는 방식이 Lin28B와
TuT4와 같은 분자들을 기능적으로 저해하거나 아니면 아직까지 알려지지 않은 3′-5′
exo-deuridylase를 끌고 와서 U-tail을 제거하여 pre-let-7 마이크로RNA를 안정화시켜 성숙 let-7
마이크로RNA의 발현을 증대시키는 가정을 할 수 있음.
- Trypanosoma brucei 와 같은 수면병을 유도하는 원충류의 경우 RNA editosome의 구성체로서
3′-5′exo-deuridylase가 존재하나 척추동물 이상에서는 그 존재가 아직 확인되어있지 않음.
- 매우 흥미롭게도 기존에 연구에 따르면 3′-5′ deadenylase인 PARN이 uridine 잔기에 대해서
adenine 잔기 못지않은 특이성을 가지고 있는 것이 보고되어 있으나 이는 인위적인
homo-polymeric U에 대해서만 실험을 진행하였고 실질적으로 3′uridylation 되어있는 let-7 마이
크로RNA에 대해서는 그 작용이 알려진 바가 전혀 없음.
- 일차적으로 3′-5′ deadenylase인 PARN이 결합가능성을 검증하기 위해서 FLAG-tag을 포함
하는 HuR, Lin28B, Dicer, TuT4, PARN을 제작하였으며 매우 흥미롭게도 모든 분자들이 동일
complex로 상에 3′-5′ deadenylase인 PARN을 끌고옴을 확인할 수 있었음 (그림 4).
- 따라서 HuR이 전구체 let-7 마이크로RNA와 성숙 let-7 마이크로RNA를 증대시키는 방식이
일차적으로 3′-5′exo-deuridylase PARN을 끌고 와서 oligo-uridylation 된 let-7 마이크로RNA
의 deuridylation을 유도하여 전구체 let-7 마이크로RNA을 안정화시키고 결국 전구체 let-7 마이
크로RNA와 성숙 let-7 마이크로RNA를 증대시키는 기전을 생각할 수 있음.
- 이를 입증하기 위해서 gamma-32
P-UTP를 이용 uridylation된 전구체 let-7 마이크로RNA의 5′에
방사성동위원소로 표지한 뒤 세포내에서 분리정제한 FLAG-PARN 또는 E. coli에서 순수분리한 재
조합 PARN을 이용 본 연구진이 고안한 in vitro PARN processing assay를 수행함.
- 매우 놀랍게도 3′-5′ deadenylase인 PARN은 uridylation된 전구체 let-7 마이크로RNA에 대해
3′-5′exo-deuridylase activity를 지니고 있음을 확인할 수 있었음.
- 이러한 3′-5′exo-deuridylase activity가 PARN 자체의 기능이 아닌 PARN과 결합하고 있는 아직
알려지지 않은 3′-5′exo-deuridylase activity를 가진 분자에 의해 매개될 가능성을 배제하기 위해서
PARN의 야생형과 촉매활성이 결여된 D28A 돌연변이체에 대한 in vitro PARN processing assay를 수
행하였으며 촉매활성이 결여된 D28A 돌연변이체에의 경우 3′-5′exo-deuridylase activity가 관찰되지
않는 결과를 얻었으며 이는 3′-5′exo-deuridylase activity가 PARN의 고유 성질임을 입증하는 결과
임.
- 결론적으로 HuR에 의해 Dicer 복합체로 recruit된 PARN은 uridylation된 전구체 let-7 마이크로
RNA의 deuridylation을 매개해서 안정화를 유도하며 전구체 및 성숙 let-7 마이크로RNA의 발현
을 증대시키는 것으로 파악할 수 있음. 이러한 결과는 Northern blot 및 let-7 sensor를 이용한
PARN 과발현 실험을 통해서 검증 할 수 있었음.

- 10 -
그림 5. 3′-5′ deadenylase PARN은 uridylation된 전구체 let-7 마이크로RNA에 대해 3′-
5′exo-deuridylase activity를 지니고 있음. (A) human PARN에 대한 in vitro PARN
processing assay 결과. gamma-32P-UTP를 이용 생합성된 uridylation된 전구체 let-7 마이크로
RNA의 방사성동위원소로 5′표지후 in vitro PARN processing를 수행함 (B) human PARN
의 야생형과 촉매활성이 결여된 D28A 돌연변이체에 대한 in vitro PARN processing assay 결과.
gamma-32P-ATP를 이용 uridylation된 전구체 let-7 마이크로RNA의 5′끝부분만 인산기를
방사성동위원소로 표지함. (C) 재조합 3′-5′ deadenylase PARN은 uridylation된 전구체
let-7 마이크로RNA에 대해 3′-5′exo-deuridylase activity를 지니고 있음.
- 다음으로 PARN의 과발현이 pre-let-7과 성숙 let-7의 발현에 어떠한 영향을 미치는지 여부를
조사하기 위해 PARN과 pri-let-7a-1을 과발현하였을 때 발현이 증대되는 상황을 Northern blot을
통해서 확인할 수 있었으며 또한 D28A 돌연변이체는 이러한 특징을 가지지 않음을 8개의 let-7
표적 site을 중에서 5개 포함하고 있는 HMGA2의 3′UTR 연결된 리포터 시스템을 이용해서 그
효과를 검증할 수 있었음.
- 실질적으로 이러한 PARN에 의한 pre-let-7 family의 deuridylation 매개와 이를 통한 효과가
특이적으로 pre-let-7 family의 안정화를 유도하고 이에 따른 mature let-7의 발현을 증대시키는
지 여부를 파악하기 위해서 Lin28B의 발현이 매우 높은 Huh-7 hepatocellular carcinoma cell에서
특이적으로 PARN을 knock down 시킴.
- 이때 RNA 수준에서 let-7의 변화가 어떻게 일어나는지 알기 위해서 splinted ligation 방법을
통해 정량적으로 let-7 마이크로 RNA의 변화를 보았을 때 그 수준이 현저히 감소함을 통해 앞선
결과들을 다시 재확인할 수 있었음.

- 11 -
그림 6. PARN의 과발현은 전구체 및 성숙 let-7 마이크로RNA의 발현을 증대시킴. (A)
293T 세포주에서 pri-let-7a-1과 PARN의 과발현에 의한 마이크로RNA let-7의 생합성 영향
에 대한 Northern blot 결과. (B) let-7 마이크로RNA의 발현변화를 monitoring 할 수 있는
5개의 반복된 let-7 표적 부위를 지니고 있는 Renilla luciferase-HMGA2 3′UTR를 이용한
human PARN의 야생형과 촉매활성이 결여된 D28A 돌연변이체를 과발현시킨 dual lucifierase
assay 결과. (C) Splinted ligation을 통한 정량적인 let-7 마이크로RNA의 변화관찰. 특이적
으로 PARN을 knock down 시켰을 때 RNA 수준에서 let-7의 변화를 관찰함. (D) PARN을
knock down 시켰을 때 Renilla luciferase-HMGA2 3′UTR sensor 변화를 관찰 (E) PARN
의 knockdown은 let-7a의 발현을 특이적으로 감소시키며 따라서 Ras와 HMGA2의 발현을 증대
시킴.
- 이러한 PARN 특이적 knockdown에 의한 let-7의 발현 변화 효과는 동일하게 HMGA2 sensor
와 그리고 기존에 let-7의 표적으로 알려진 Ras와 세포내 HMGA2의 다시 derepression되는 것을
통해서 실질적으로 PARN이 전구체 let-7의 발현조절을 deuridylation을 통해 매개하고 있음을 시
사하는 매우 중요한 결과라 판단할 수 있음.
- 흥미로운 사실은 PARN에 의한 전구체 let-7의 발현조절의 정확한 분자기전을 파악하기 위해서
조사하던 중 Lin28B분자가 특이적으로 인산화되는 사실을 파악하게 되었음.
- 매우 흥미롭게도 Lin28B는 기존의 보고처럼 hepatocellular carcinoma에서 그 발현정도가 높게
관찰되었으나 특별한 점은 lung carcinoma인 A549나 NCI-H1299 세포주에서도 그 발현이 높을
것을 확인할 수 있었음.
- 기존의 보고에 따르면 lung tumor에서 let-7의 발현이 낮은 것으로 알려져 있으며 이러한 현상
은 Lin28B의 과발현이 전구체 let-7을 불안정화를 유도하기 때문이 아닌가 판단됨.

- 12 -
그림 7. (A) 다양한 암세포주와 embryonal carcioma 세포주에서의 Lin28B의 발현 양상.
Lin28B*; 전사후조절이 일어났을 것으로 예상되는 Lin28B, ΔN-Lin28B; Lin28B의 splicing
variant. 그림 32. 종양억제 마이크로RNA let-7 분해촉진인자 RNA결합단백질 Lin28의 S221 잔기
가 특이적으로 인산화됨. (B) Lin28A 와 Lin28B의 구조 비교. Lin28B의 경우 C-terminal로 50개
의 아미노산이 확장되어 있음. (C) peptide 분석결과를 통한 Lin28B 상의 예상되는 인산화 잔기들
과 종간의 보존 정도 분석 (D) Lin28B의 S221 잔기를 alanine으로 치환하였을 때 특이적인 인산
화정도가 완전히 사라짐.
- Immunoblot 결과 일관적으로 야생형의 Lin28B의 보다 1-2kDa정도 크기의 band retardation이
관찰되고 있다는 점임 (Lin28B*). 이는 splicing variant롤 생각할 수 있으나 그 보고가 없고 sumo
modification 또는 ubiquitination과 같은 번역후 조절으로 판단하기에는 크기가 작은 것으로 사료
됨.
- 그림 14에서 보는 것처럼 LC-MS/MS기법을 이용한 완전한 서열분석에는 실패하였으나 일차
적으로 앞서 언급한 C-terminal의 50개의 확장되어있는 아미노산 서열 영역에서 인산화 가능성이
있는 peptide가 검출되었음.
- 본 연구진은 peptide 분석 결과를 근거로 peptide상에 존재하는 인산화 가능성이 가장 높은
seriene, threonine 서열에 대해서 모두 site-directed mutagenesis를 수행함.
- 앞서 Lin28B과 HuR은 기능적으로 3′-5′exo-deuridylase activity를 지니고 있는 PARN을 끌고 와서
결국 pre-let-7 family의 안정성을 조절한다는 사실을 알 수 있었는데 실질적으로 이러한 인산화와
PARN과의 생체 특이적 연관성을 조사하지 못했음. 한 가지 가능성은 인산화의 상태에 따라 세포질
polyadenylation과 마찬가지로 PARN과 oligo-uridylation 복합체간의 결합이 변화될 가능성을 상정해 볼
수 있었음.
- 이를 입증하기 위해 인산화를 막거나 아니면 계속적으로 인산화를 모방할 수 있도록 Lin28B의 S221
부분을 alanine 또는 aspartate로 변경시킨 뒤 PARN과의 결합을 구체적으로 조사하였음.
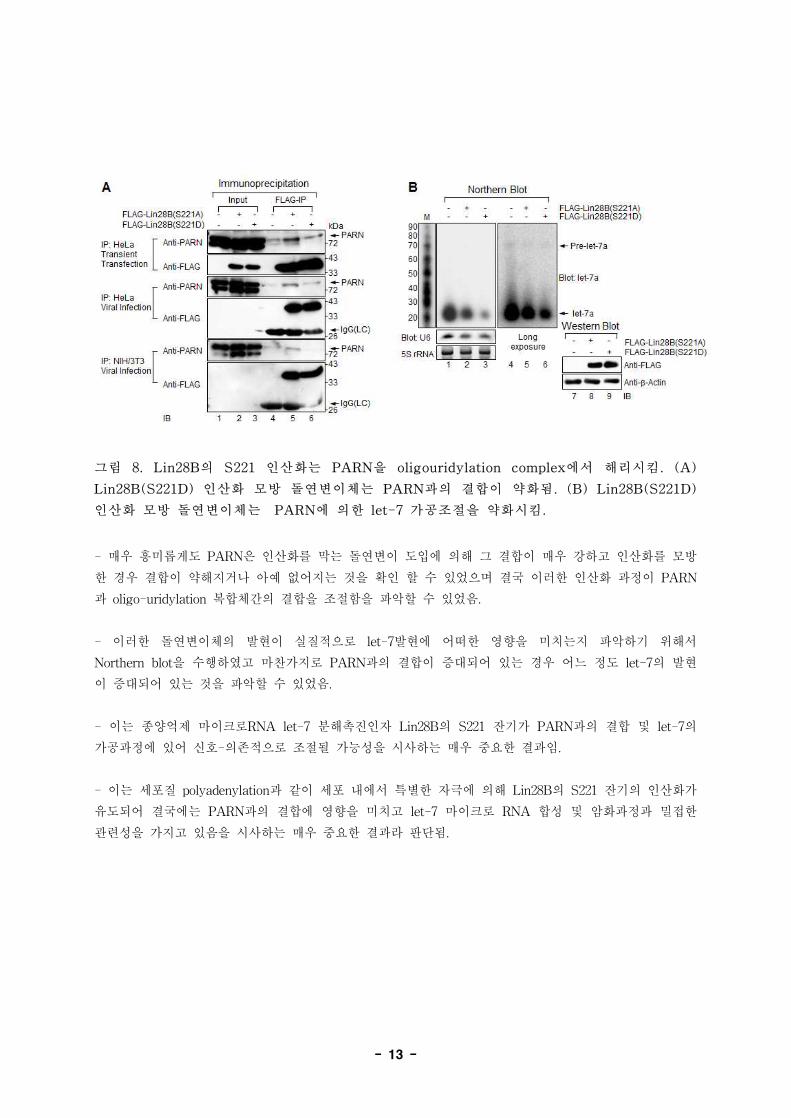
- 13 -
그림 8. Lin28B의 S221 인산화는 PARN을 oligouridylation complex에서 해리시킴. (A)
Lin28B(S221D) 인산화 모방 돌연변이체는 PARN과의 결합이 약화됨. (B) Lin28B(S221D)
인산화 모방 돌연변이체는 PARN에 의한 let-7 가공조절을 약화시킴.
- 매우 흥미롭게도 PARN은 인산화를 막는 돌연변이 도입에 의해 그 결합이 매우 강하고 인산화를 모방
한 경우 결합이 약해지거나 아예 없어지는 것을 확인 할 수 있었으며 결국 이러한 인산화 과정이 PARN
과 oligo-uridylation 복합체간의 결합을 조절함을 파악할 수 있었음.
- 이러한 돌연변이체의 발현이 실질적으로 let-7발현에 어떠한 영향을 미치는지 파악하기 위해서
Northern blot을 수행하였고 마찬가지로 PARN과의 결합이 증대되어 있는 경우 어느 정도 let-7의 발현
이 증대되어 있는 것을 파악할 수 있었음.
- 이는 종양억제 마이크로RNA let-7 분해촉진인자 Lin28B의 S221 잔기가 PARN과의 결합 및 let-7의
가공과정에 있어 신호-의존적으로 조절될 가능성을 시사하는 매우 중요한 결과임.
- 이는 세포질 polyadenylation과 같이 세포 내에서 특별한 자극에 의해 Lin28B의 S221 잔기의 인산화가
유도되어 결국에는 PARN과의 결합에 영향을 미치고 let-7 마이크로 RNA 합성 및 암화과정과 밀접한
관련성을 가지고 있음을 시사하는 매우 중요한 결과라 판단됨.
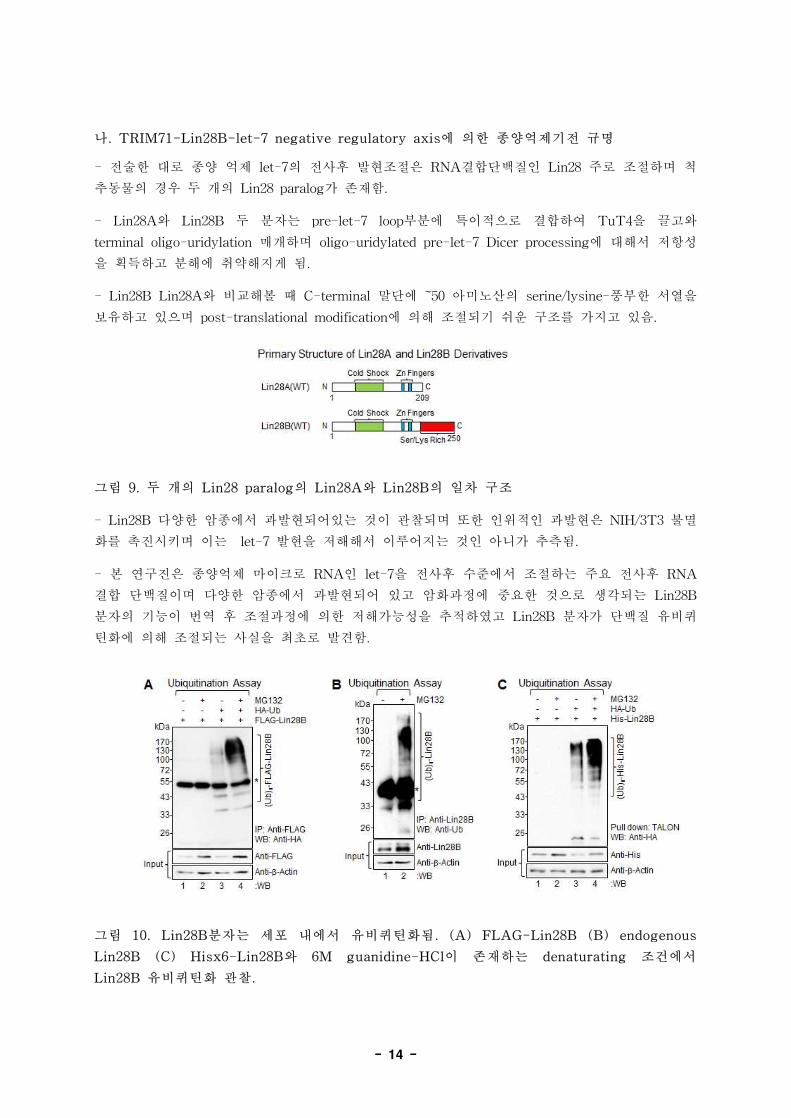
- 14 -
나. TRIM71-Lin28B-let-7 negative regulatory axis에 의한 종양억제기전 규명
- 전술한 대로 종양 억제 let-7의 전사후 발현조절은 RNA결합단백질인 Lin28 주로 조절하며 척
추동물의 경우 두 개의 Lin28 paralog가 존재함.
- Lin28A와 Lin28B 두 분자는 pre-let-7 loop부분에 특이적으로 결합하여 TuT4을 끌고와
terminal oligo-uridylation 매개하며 oligo-uridylated pre-let-7 Dicer processing에 대해서 저항성
을 획득하고 분해에 취약해지게 됨.
- Lin28B Lin28A와 비교해볼 때 C-terminal 말단에 ~50 아미노산의 serine/lysine-풍부한 서열을
보유하고 있으며 post-translational modification에 의해 조절되기 쉬운 구조를 가지고 있음.
그림 9. 두 개의 Lin28 paralog의 Lin28A와 Lin28B의 일차 구조
- Lin28B 다양한 암종에서 과발현되어있는 것이 관찰되며 또한 인위적인 과발현은 NIH/3T3 불멸
화를 촉진시키며 이는 let-7 발현을 저해해서 이루어지는 것인 아니가 추측됨.
- 본 연구진은 종양억제 마이크로 RNA인 let-7을 전사후 수준에서 조절하는 주요 전사후 RNA
결합 단백질이며 다양한 암종에서 과발현되어 있고 암화과정에 중요한 것으로 생각되는 Lin28B
분자의 기능이 번역 후 조절과정에 의한 저해가능성을 추적하였고 Lin28B 분자가 단백질 유비퀴
틴화에 의해 조절되는 사실을 최초로 발견함.
그림 10. Lin28B분자는 세포 내에서 유비퀴틴화됨. (A) FLAG-Lin28B (B) endogenous
Lin28B (C) Hisx6-Lin28B와 6M guanidine-HCl이 존재하는 denaturating 조건에서
Lin28B 유비퀴틴화 관찰.

- 15 -
- 그림 17에서 보이듯이 다양한 조건에서 Lin28B는 세포 내에서 유비퀴틴화되는 것을 확인할 수
있으며 유비퀴틴화를 통한 단백질의 분해과정은 특이적인 E3 유비퀴틴 ligase가 매개를 하게 되며
따라서 어떠한 분자가 Lin28B 단백질의 안정성을 조절하는지를 알게 된다면 생체 내에서 발암과
정을 통제할 수 있을 것으로 사료됨.
- 본 연구진은 기존에 Ago2의 유비퀴틴화를 유도하는 것으로 알려진 TRIM71이라는 분자가
Lin28B 분자를 특이적으로 유비퀴틴화 시킬 가능성을 가정하고 연구를 진행함.
- TRIM71 분자는 N-terminus에 RING-type zinc finger, B‑box domains, 그리고 coiled coil
domain으로 구성되어있고 다수의 TRIM 단백질들은 유비퀴틴 ligase 활성을 지니고 있는 것으로
알려져 있음. 또한 최근에 TRIM71은 유비퀴틴 ligase 활성을 지니고 있는 RNA결합단백질로서
분류되었음.
그림 11. TRIM71의 일차구조 (좌상). TRIM71과 Lin28B는 colocalize함 (좌하). Ago2분자가
TRIM71에 의해 특이적으로 유비퀴틴화 돼서 분해되는 분자기전 모식도(우).
- 다양한 생화학적 분자생물학 기법을 이용 TRIM71분자가 Lin28B분자가 특이적인 결합을 수행
하며 TRIM71은 Lin28B의 분해를 가속화시킴을 알 수 있었음 (data not shown).
- 전술했듯이 Lin28B Lin28A와 비교해볼때 C-말단에 ~50 아미노산의 serine/lysine-풍부한 서열
을 보유하고 있으며 post-translational modification에 의해 조절되기 쉬운 구조를 가지고 있음.
Lin28A와 그 구조적 차이를 볼 때 이 부위를 통해서 TRIM71에 의한 유비퀴틴화와 단백질-단백
질 간의 결합이 이루어질 가능성이 높음.
- 그림 19에서 보이듯이 TRIM71과 Lin28B의 야생형, Lin28A, 그리고 Lin28B의 C-말단부위를 제
거한 돌연변이체를 이용하여 co-IP 실험을 진행하였을 때 매우 흥미롭게도 Lin28B의 야생형만 그
결합이 관찰됨을 알 수 있었음. 따라서 Lin28B C-말단에 ~50 아미노산의 serine/lysine이 풍부한
서열 TRIM71에 의한 유비퀴틴화와 단백질-단백질 간의 결합에 매우 중요하다는 사실을 시사하는
매우 중요한 결과임.
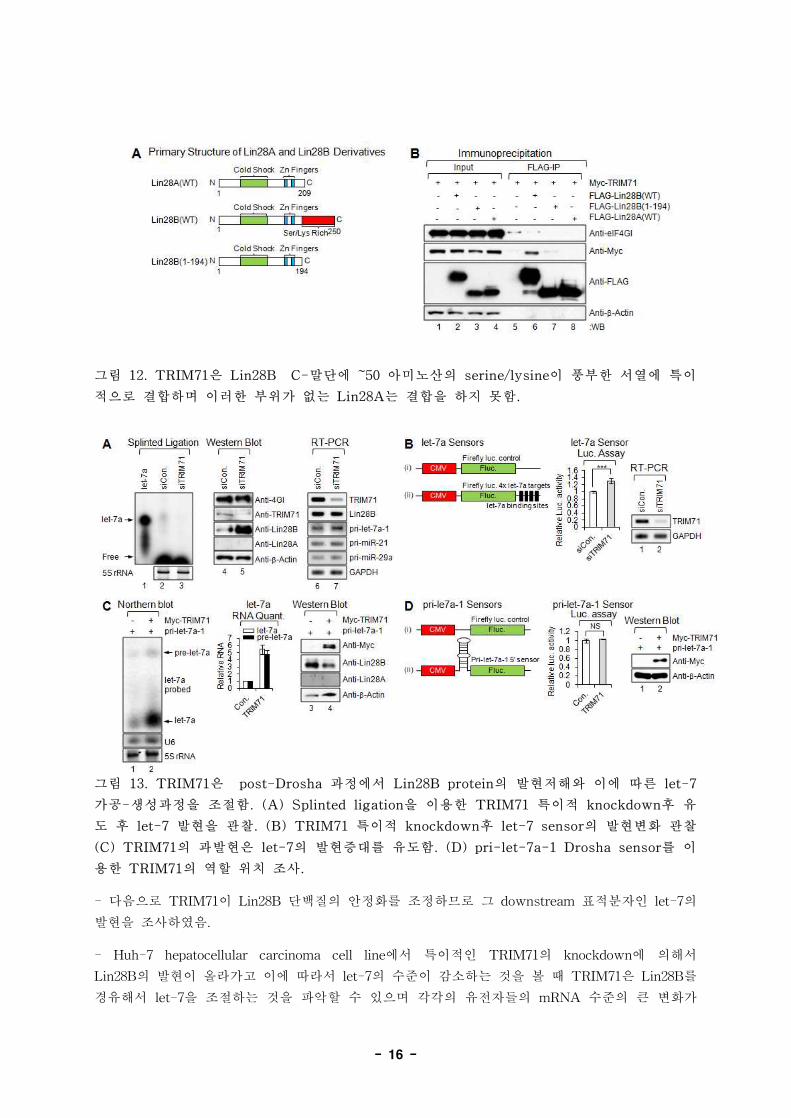
- 16 -
그림 12. TRIM71은 Lin28B C-말단에 ~50 아미노산의 serine/lysine이 풍부한 서열에 특이
적으로 결합하며 이러한 부위가 없는 Lin28A는 결합을 하지 못함.
그림 13. TRIM71은 post-Drosha 과정에서 Lin28B protein의 발현저해와 이에 따른 let-7
가공-생성과정을 조절함. (A) Splinted ligation을 이용한 TRIM71 특이적 knockdown후 유
도 후 let-7 발현을 관찰. (B) TRIM71 특이적 knockdown후 let-7 sensor의 발현변화 관찰
(C) TRIM71의 과발현은 let-7의 발현증대를 유도함. (D) pri-let-7a-1 Drosha sensor를 이
용한 TRIM71의 역할 위치 조사.
- 다음으로 TRIM71이 Lin28B 단백질의 안정화를 조정하므로 그 downstream 표적분자인 let-7의
발현을 조사하였음.
- Huh-7 hepatocellular carcinoma cell line에서 특이적인 TRIM71의 knockdown에 의해서
Lin28B의 발현이 올라가고 이에 따라서 let-7의 수준이 감소하는 것을 볼 때 TRIM71은 Lin28B를
경유해서 let-7을 조절하는 것을 파악할 수 있으며 각각의 유전자들의 mRNA 수준의 큰 변화가

- 17 -
없는 것으로 볼 때 이는 전사후 수준에서의 조절이라는 것을 알 수 있음.
- 다양한 sensor system을 이용 TRIM71이 let-7을 전사후 단계에서 조절하는 것을 알 수 있었으
며 pri-let-7a-1을 이용한 Drosha sensor를 이용하여 이러한 과정이 post-Drosha step에서 일어나
고 있음을 검증함.
그림 14. TRIM71의 RING finger motif가 Lin28B의 유비퀴틴화, 결합, 그리고 let-7 조절에
매우 중요함. (A) TRIM71의 1차구조. (B) TRIM71의 RING finger 돌연변이체는 Lin28B를
유비퀴틴화 시키지 못함. (C) let-7 sensor를 이용한 TRIM71의 RING finger 돌연변이체의
기능 검증. (D) (B) TRIM71의 RING finger 돌연변이체는 let-7의 발현을 증대시키지 못함.
(E) TRIM71의 RING finger motif는 Lin28B와의 protein-protein interaction에 매우 중요
함. (F) TRIM71의 RING finger motif 돌연변이 유도는 pre-let-7의 uridyl화를 촉진시키지
못함.
- 다음으로 TRIM71의 어느 부위가 Lin28B의 유비퀴틴화, 결합, 그리고 let-7 조절에 중요한지 파
악하기위해서 TRIM71에서 유비퀴틴 전달에 매우 중요한 것으로 알려진 RING finger motif의 두
개의 cysteine 잔기를 alanine으로 치환한 돌연변이체를 제작하였고 매우 놀랍게도 Lin28B의 유비
퀴틴화뿐만아니라 Lin28B와의 결합 그리고 이에 따른 let-7의 합성과정에 증가시키는 능력을 상
실함을 알 수 있었음.
- 이는 TRIM71의 유비퀴틴화 기능에 중요한 RING finger motif이 Lin28B-let-7 axis에 중요한
기능을 가지는 것을 시사하는 매우 중요한 발견이라 판단됨.

- 18 -
그림 15. TRIM71의 Lin28B에 의해서 유도되는 cellular transforming 잠재력을 억제함. (A)
soft agar colony formation assay를 통한 TRIM71의 Lin28B colony 형성능력억제 계측.
(D) TRIM71의 Lin28B에 의해서 유도되는 cellular transforming 잠재력의 억제는 nude
mouse의 tumorigenicity assay를 통해서도 재확인됨.
- 다음으로는 기존에 알려진 Lin28B의 cellular transformation potential을 TRIM71이 어떻게 조절
하는지 파악하기 위해서 각각의 유전자들을 과발현 시킬 수 있는 retrovirus system을 이용해서
soft agar colony formation assay를 수행하였음.
- 기존의 보고와 일치하여 Lin28B는 강력한 cellular transforming potential을 가지고 있음이
NIH/3T3 세포주에서 확인할 수 있었으며 매우 놀랍게도 여기에 TRIM71 분자를 공동발현시킨 경
우 이러한 Lin28B에 의한 cellular transforming 잠재력을 저해하는 결과를 얻을 수 있었음.
- 이러한 TRIM71의 Lin28B에 의해서 유도되는 cellular transforming 잠재력의 억제는 nude
mouse의 tumorigenicity assay를 통해서도 재확인 할 수 있었으며 이는 실질적으로 TRIM71이
Lin28B-let-7 axis에서 상위조절자로 작용할 가능성을 시사하는 매우 중요한 결과라 판단됨.

- 19 -
다. 뇌신경교종에서 DDX23 RNA헬리케이즈에 의한 종양 마이크로RNA-21의 가공조절 연구
- 본 연구진은 선행 연구논문발표를 통해 종양 miR-21이 전사후 조절단계에서 PTEN이라는 종양
억제인자에 의해 조절된다는 사실을 밝혔으며 PTEN의 소실은 마이크로 RNA 가공기구에 영향을
미치는 것을 밝힘 (Kim et al., 2011).
- 본 연구진은 선행연구를 통해 proteomics 접근법을 통해 (PTEN에 대한 tandem affinity
purification과 LC/MS/MS 분석) 종양 miR-21의 가공과정에 관여할 가능성이 높은 다수의 전사후
조절분자들을 발굴-분석하였음. 또한 선행 연구발표에서 본 연구진은 종양억제유전자인 PTEN이
RNH1-Drosha간의 결합을 방해해서 종양 miR-21의 가공과정에 관여함을 규명함 (Kim et al.,
2011).
- 매우 흥미롭게도 다양한 RNA헬리케이즈, RNA결합단백질, RNase inhibitor등 전사후 조절에 관
여하는 다수의 분자들이 검출되어 나옴.
그림 16. 종양억제유전자 PTEN의 tandem affinity purification 및 LC/MS/MS 기법을 통해
잠재적인 종양 miR-21 가공 전사후 조절인자로서 DDX23의 분리동정 (A-C).
- 분리-동정된 분자들중 DDX23 RNA헬리케이즈의 경우 놀랍게도 National Cancer Institute의
TCGA에서 데이터를 추출하여 분석을 진행하였을 때 그 발현은 정상조직에 비해 악성뇌신경교종
환자에서 그 발현이 현저히 높은 것을 관찰할 수 있으며 종양 miR-21의 발현과 유사한 패턴을
보여주고 있음 (그림 17).
- DDX23은 DEAD (Asp-Glu-Ala-Asp) box 계열의 RNA헬리케이즈로서, DEAD box RNA헬리
케이즈들은 현재까지 50여 종으로 알려져 있으며 대다수의 분자들이 pre-mRNA의 가공, splicing
등의 과정에 관련되어 있는 것으로 밝혀져 있음.

- 20 -
- 본 연구진은 이러한 모든 50여종의 DEAD box RNA헬리케이즈들에 대해서 악성뇌신경교종에
서 환자의 예후 및 생존도를 REMBRANDT에서 분석을 시도하였으며, 매우 놀랍게도
Kaplan-Meier survival plot의 양상이 오직 DDX23만이 악성뇌신경교종 환자군에서 유의적으로
생존률이 낮은 것을 확인할 수 있었음.
- 본 연구진은 종양 miR-21의 가공과정과 악성뇌신경교종환자의 생존율에 대해서 DDX23 RNA
헬리케이즈가 매우 중요한 역할을 하는 것으로 판단할 수 있었으며 연구의 중심방향을 DDX23로
설정 (그림 17).
그림 17. 악성뇌신경교종환자의 암조직과 정상조직에서의 DDX23의 발현 및 생존예후 분석.
(A) NCI REMBRANDT database 추출결과 (Kim et al., 미발표 결과). (B) 악성뇌신경교종
에서 DDX23의발현에 따른 Kaplan-Meier survival plot. (C) 본 연구진이 54명의 뇌신경교
종환자의 조직을 이용 Western blot을 이용한 DDX23의 발현을 분석한 결과 (Yin et al., 미
발표 결과).
- 뇌신경교종은 크게 4단계로 나눠지며 1-2등급의 저등급의 성상세포종(low grade astrocytoma)
은 비교적 정상적인 세포와 덜 악성화된 세포를 포함하고 있으며 4단계의 악성뇌신경교종
(glioblastoma)은 매우 빨리 자라며 조직근처로 침범하면서 악성도가 아주 높은 세포들을 포함하
며 가장 예후가 좋지 않음.
- 본 연구진이 TCGA에서 추출해낸 결과를 재확인하기 위해서 54명의 뇌신경교종환자의 조직을
이용 분석한 결과 높은 단계의 뇌종양에서 DDX23이 높은 발현을 보이는 것이 관찰됨 (그림 17).
-모든 임상적인 분석결과를 종합해볼 때 DDX23 RNA헬리케이즈가 뇌신경교종의 증식, 침윤, 악
성화에 있어서 매우 중요한 역할을 수행할 것으로 판단되며 이는 종양 miR-21의 특이적 전사후
가공기구와 밀접한 연관성을 지니고 있는 것을 암시하는 매우 중요한 결과들로 판단됨.

- 21 -
1) DDX23은 뇌신경교종세포주의 침윤(invasion)과 증식(proliferation)을 증대시킴.
- DDX23의 뇌신경교종에서의 역할을 규명하기 위해서 렌티바이러스 시스템을 이용 DDX23분자
를 특이적으로 knockdown을 유도하거나 과발현을 유도해서 뇌신경교종세포주(U87-MG)의 침윤
과 증식에 어떠한 영향을 미치는지 조사해봄.
- 흥미롭게도 본 연구진이 기존에 분석한 환자시료와 TCGA 분석결과와 일맥상통하게 DDX23의
제거는 뇌신경교종세포주의 침윤과 증식을 억제하는 결과를 얻을 수 있었으며, 이와 반대로
DDX23의 과발현은 침윤과 증식을 증대시키는 결과를 얻을 수 있었음. 이를 통해 DDX23이 뇌신
경교종세포주의 침윤과 증식에 매우 중요한 분자라는 결론을 얻을 수 있었음 (그림 18).
그림 18. DDX23은 뇌신경교종세포주의 침윤과 증식을 증대시킴. (A) pGIPZ-shDDX23 렌티
바이러스를 U87MG 뇌신경교종세포주에 감염시킨 뒤 DDX23분자의 특이적 knockdown을 유
도 침윤 assay(A)와 세포증식 assay(B)를 수행. HRST-DDX23 렌티바이러스 시스템을 이용
DDX23분자를 특이적으로 과발현을 유도해 침윤 assay(C)와 세포증식 assay(D)를 수행.
2) DDX23은 종양 miR-21의 발현을 조절함.
- 다음으로 본 연구진이 분석한 환자시료와 TCGA 분석결과에 근거하여 실질적으로 DDX23이 암
의 발생, 증식, 침윤에 중요한 종양 miR-21의 발현 조절에 관여하는지 여부를 조사해봄.

- 22 -
- DDX23 특이적인 siRNA를 이용 뇌신경교종세포주인 U87MG 세포주에서 DDX23의 특이적
knockdown을 유도하였을 때 종양 miR-21의 발현이 그 전구체(precursor)와 성숙 형태에서 ~70%
정도 감소하는 결과를 얻을 수 있었음 (그림 12). 이와 반대로 대표적인 종양억제 miRNA로 알려
진 let-7의 발현은 전구체와 성숙형태에 큰 변화를 보이고 있지 않으며 이 결과는 DDX23의
miRNA 가공과정 조절작용이 종양 miR-21 특이적임을 시사하는 매우 중요한 결과라 판단됨,
- 본 연구진은 종양 miRNA들의 변화를 보다 정량적이고 대단위 스크리닝이 용이한 종양
miR-21“센서” (sensor)를 개발하였고 knockdown과 과발현에서 앞선 Northern blotting과 유사한
결과를 얻을 수 있었음. 결론적으로 DDX23은 종양 miR-21 특이적으로 조절하는 분자임을 암시
하는 매우 중요한 결과라 판단됨.
그림 19. DDX23은 뇌신경교종세포주에서 종양 miR-21의 발현을 조절함. (A) DDX23 특이
적 siRNA를 U87MG 뇌신경교종세포주에 transfection한 뒤 Northern blotting을 이용 종양
miR-21의 발현을 관찰. let-7 종양억제 miRNA는 비교군으로 사용됨. (B) DDX23 특이적
siRNA를 U87MG 뇌신경교종세포주에 transfection한 뒤 splinted ligation 방법을 이용종양
miR-21의 발현을 관찰. (C) 종양 miR-21의 표적 site을 4개 가지고 있는 센서의 개발. (D)
DDX23 특이적 siRNA와 센서를 co-transfection한 뒤 센서의 변화를 감지함. (E) DDX23
과발현체와 센서를 co-transfection한 뒤 센서의 변화를 감지함.

- 23 -
3) DDX23은 종양 miR-21 발현을 Drosha단계에서 전사후 조절함.
- 다음으로 DDX23이 종양 miR-21을 miRNA 가공과정에서 어느 단계에서 조절하는지 여부를 확
인하기 위해서 DDX23의 과발현을 유도하였으며 DDX23의 과발현은 종양 miR-21을 전구체
(precursor)와 성숙체 (mature) 형태을 모두 증대 시키는 것을 알 수 있었음 (그림 20). 이는 특이
적으로 DDX23을 knockdown을 유도하였을 때 각종 1차 (primary) 전구체 miRNA들이 발현의 변
화가 없는 것을 봤을 때 전사수준에서의 작용이 아닌 전사후 조절 과정임을 알 수 있음 (그림
20).
- DDX23이 전사후 조절과정의 어느 과정에서 종양 miR-21의 가공에 관여하는지를 명확하게 하
기위해서 Drosha에 의한 가공과정을 쉽게 검출할 수 있는 종양 pri-miR-21센서를 제작 DDX23의
영향을 검증함.
- 그림 20에서 보이듯 DDX23의 과발현을 유도하였을 때 Drosha에 의한 가공반응성이 증대되어
서 센서의 활성이 감소됨을 확인할 수 있었으며 반대로 특이적으로 knockdown을 유도하였을 때
센서의 활성이 증대되는 결과를 볼 때 DDX23은 Drosha에 의한 가공과정에 있어서 특이적인 역
할을 수행하는 것으로 볼 수수 있는 매우 중요한 결과라 판단됨.
그림 20. DDX23은 종양 miR-21 발현을 Drosha 단계에서 조절함. (A) DDX23 특이적
siRNA를 U87MG 뇌신경교종세포주에 transfection한 뒤 RT-PCR을 통한 각종 pri-miRNA
발현수준 관찰. (B) DDX23 과발현체 transfection한 뒤 Northern blotting을 이용 전구체
(precursor)와 성숙 형태 종양 miR-21의 발현을 관찰. (C) Drosha에 의한 종양 pri-miR-21
발현을 확인할 수 있는 센서의 고안. (D) & (E) DDX23 과발현체 또는 특이적 siRNA를 센
서를 co-transfection한 뒤 센서의 변화를 감지함.

- 24 -
4) DDX23은 Drosha microprocessor 및 pri-miR-21과 특이적으로 결합함.
- 이제까지의 결과를 통해 DDX23은 Drosha miRNA microprocessor의 관계를 통해서 종양
miR-21의 가공과정에 특이적으로 관여함을 추정 할 수 있음. 실제적으로 DDX23이 종양 miR-21
의 가공기구의 일원임을 확인하기 위해 각각 FLAG-DDX23 또는 Drosha-FLAG을 과발현 시켜
세포 내에서 각 주요 분자 간의 결합을 immunoprecipitaion (IP) 기법을 통해 확인함.
- DDX23이 종양 pri-miR-21과 특이적으로 결합하는지 여부를 확인하기 위해서 RNP-IP
RT-PCR을 수행하였으며 이를 통해서 DDX23-Drosha복합체상에 종양 pri-miR-21이 특이적으로
존재하는 것을 확인할 수 있었음. 이 모든 결과는 DDX23이 Drosha microprocessor와 연합해서
특이적으로 종양 miR-21 가공과정에 관여함을 시사하는 매우 중요한 결과라고 판단됨.
5) DDX23의 RNA헬리케이즈 활성이 종양 miR-21 특이적 가공조절에 중요함.
- DDX23은 RNA헬리케이즈 활성을 지니기 위해서 Walker A motif에 존재하는 ATP 결합에 관
여하는 보존된 아미노산 잔기가 매우 중요한 것으로 알려져 있음.
그림 21. DDX23은 Drosha microprocessor와 pri-miR-21과 특이적으로 결합함. (A) & (B)
FLAG-DDX23 또는 Drosha-FLAG을 과발현-immunoprecipitation을 수행 각 분자 간의 특
이적 결합을 확인. (C) RNP-IP RT-PCR을 통한 DDX23-Drosha 복합체 상의 종양
pri-miR-21이 존재 확인.
- 매우 흥미롭게도 DDX23의 보존된 Walker A motif상의 보존된 한 개의 아미노산을 활성을 없
애는 잔기로 치환하였을 때 (K441N) DDX23은 종양 miR-21의 가공조절을 증대시키는 역할을 상
실하고 오히려 dominant negative mutant와 같은 특징을 보이는 것을 확인 할 수 있었음 (그림

- 25 -
22).
- 이러한 DDX23 K441N 돌연변이체에 대한 특성은 Northern blotting, 성숙 혹은 pri-miR-21 센
서, 뇌신경교종세포주에서의 침윤 assay등을 통해서 확인할 수 있었으며 이는 DDX23가 다른 특
징이 아닌 RNA헬리케이즈 활성을 통해 종양 miR-21의 가공을 전사후 수준에서 조절하고 있음을
시사하는 매우 중요한 결과라 판단됨.
그림 22. DDX23의 RNA헬리케이즈 활성이 종양 miR-21 특이적 가공조절에 중요함. (A)
DDX23의 활성관련 주요 motif, 주요 아미노산잔기의 모식도. (B) DDX23의 야생형과
K441N 돌연변이의 과발현체와 pri-miR-21을 co-transfection한 뒤 Northern blotting을 이
용 전구체(precursor)와 성숙형태 종양 miR-21의 발현을 관찰. (C) 성숙 혹은 pri-miR-21
센서를 DDX23의 야생형과 K441N 돌연변이의 과발현체와 pri-miR-21을 co-transfection한
뒤 그 활성을 관찰함. (D) 렌티바이러스 HRST-DDX23의 야생형과 K441N 돌연변이체를 이
용 과발현을 유도 침윤 assay를 수행.

- 26 -
3. 연구결과 고찰 및 결론
<국내외 연구동향>
- 최근 마이크로RNA연구의 가장 큰 화두는 이들이 세포 외부환경 혹은 유전적 요인에 의해 일
어나는 변이를 관찰하거는 그 분자기전을 밝히는 일임. 하지만, 아직까지 암세포에서의 특이적인
조절기전에 관한 연구는 아직 많이 미진한 상황임.
- 최근에 마이크로RNA 연구에 있어서 단순한 가공조절뿐만 아니라 중간단계 (precusor) 최종단
계 (mature) 산물의 3′ 또는 5′의 추가적인 수식과정에 많은 관심을 기울이고 있는 형편임 이
는 이러한 전사후 수식과정들이 각 과정에서 생성되는 산물의 안정화와 그리고 표적분자에 대한
특이성을 결정짓는데 있어서 매우 중요한 역할을 수행하기 때문임.
<연구결과가 국내․외 기술개발 분야에서 차지하는 위치>
- 현재 국내 연구진과 외국연구진의 마이크로RNA turnover 기작을 규명하기위한 연구가 활발히
진행되고 있음. 본 연구진은 기존 연구진들의 연구방법 및 연구재료를 모두 확보하고 있고 이들보
다 한 단계 앞선 암화과정에서의 구체적인 연구를 통해 새로운 패러다임을 제시하고자 함.
- 본 연구 결과는 이제까지 예상되지 않았던 새로운 종류의 마이크로RNA turnover 기전을 구체
적으로 규명한 사례로 암화과정과 연결지어 매우 중요한 위치를 차지한다고 판단내릴 수 있음.
- 본 연구를 통해 확보된 정량적 프로테오믹스, microarray, RNA-seq 기법과 다양한 분자생물학-생
화학적인 방법은 향후 RNA수준에서 특이적 전사후 조절에 의한 질병의 가능성을 다양한 환자의 암
조직으로부터 확인 동물모델에 적용하여 기능적 중요성을 검증하는데 있어 매우 중요할 것으로 판단됨.
<연구결과 해석>
1. PARN에 의한 종양억제 마이크로RNA let-7의 deuridylation 기전 규명
- 중요 세포 내 RNA결합단백질 HuR이 Lin28B-TuT4 oligo-uridylation 복합체 구성분자로 동정
하고 마이크로RNA let-7의 가공을 가속시키는 조절자로 작용함을 밝혔으며 핵심 마이크로RNA
가공기구인 Dicer 쪽으로 3′-5′ deadenylase PARN이 그 복합체 상에 존재함을 밝힘.
- 3′-5′ deadenylase PARN은 uridylation된 전구체 let-7 마이크로RNA에 대해 3′-5′
exo-deuridylase activity를 지니고 있음을 밝힌 부분은 기대이상의 성과라고 판단할 수 있겠음.
- 종양억제 마이크로RNA let-7 분해촉진인자 Lin28B의 S221 잔기가 특이적으로 인산화됨을 밝
혔으며 Lin28B의 S221 인산화는 PARN을 oligouridylation complex에서 해리시키고 PARN에 의
한 let-7 가공조절을 약화시킴을 밝힘. 이는 암화과정의 Lin28B의 인산화과정이 매우 중요함을 시
사하는 중요한 발견임.

- 27 -
2. TRIM71-Lin28B-let-7 negative regulatory axis에 의한 종양억제기전 규명
- Lin28B분자가 세포 내에서 유비퀴틴화됨을 발혔으며 TRIM71은 Lin28B C-말단에 ~50 아미노
산의 serine/lysine이 풍부한 서열에 특이적으로 결합하며 이러한 부위가 없는 Lin28A는 결합을
하지 못함을 밝힘. 이는 마이크로RNA 가공조절이 번역후 조절에 의해서 발생한다는 새로운 증거
로서 매우 중요함.
- TRIM71은 post-Drosha 과정에서 Lin28B protein의 발현저해와 RING finger motif가 Lin28B
의 유비퀴틴화, 결합, 그리고 let-7 조절에 매우 중요함을 밝힘.
- TRIM71의 Lin28B에 의해서 유도되는 cellular transforming 잠재력을 억제함을 밝힘. 이는 암
화과정에서 TRIM71의 역할을 새롭게 조망할 수 있는 중요한 관찰임.
3. 뇌신경교종에서 DDX23 RNA헬리케이즈에 의한 종양 마이크로RNA-21의 가공조절 기전
규명
- 종양 마이크로RNA-21의 특이적 가공조절 조절 세포 내 분자로서 DDX23 RNA헬리케이즈를
최초 분리-동정하였으며 DDX23은 뇌신경교종세포주의 침윤과 증식을 증대시킴을 밝히며 종양 마
이크로RNA-21의 발현을 조절함을 밝힘.
- DDX23은 Drosha microprocessor 및 pri-마이크로RNA-21과 특이적으로 결합함을 밝혔으며
RNA헬리케이즈 활성이 종양 마이크로RNA-21 특이적 가공조절에 중요함을 밝힘. 이는 종양 마
이크로RNA-21 가공과정과 이에 따른 RNA 수식단백질간의 관련성이 뇌신경교종의 발생, 침윤,
악성화와의 연관성을 보여준 중요한 관찰이라 판단됨.

- 28 -
논문명 저자 학술대회명 지역 지원과제번호The ubiquitin ligase human TRIM71 regulates
let-7 microRNA biogenesis through themodulation of Lin28B protein level
김종헌 한국분자세포생물학회(2013)
국내 1110110
let-7 microRNA turnover: Role of novelRNA-binding protein and 3′ to 5′
exoribonuclease for the let-7 microRNAbiogenesis
김종헌 분자암학회(2012)
국내 1110110
let-7 microRNA turnover: Role of novelRNA-binding protein and 3′ to 5′
exoribonuclease for the let-7 microRNAbiogenesis
김종헌 연세대학교 SRC 세미나(2012)
국내 1110110
let-7 microRNA turnover: Role of novelRNA-binding protein and 3′ to 5′
exoribonuclease for the let-7 microRNAbiogenesis
김종헌 한국유전학회-정기학술대회
(2012)
국내 1110110
논문명저자
(저자구분)저널명(I.F.)
Year;Vol(No):Page
구분지원과제번호
Adenosine dialdehyde suppressesMMP-9-mediated invasion of cancer
cells by blocking theRas/Raf-1/ERK/AP-1 signaling pathway
제 1저자Biochem
Pharmacol.2013
1285~1300
4.576(국외SCI)
없음
AP-1/IRF-3 Targeted Anti-InflammatoryActivity of Andrographolide Isolated
from Andrographis paniculata.교신저자
Evid BasedComplement
Alternat Med.
20131~16
1.722(국외SCIE)
1110110
Bidirectional control of mRNA translationand synaptic plasticity by the
cytoplasmic polyadenylation complex.공동저자 Mol. Cell 2012
253~266
14.178(국외SCI)
없음
Silencing of MicroRNA-21 ConfersRadio-Sensitivity through Inhibition of
the PI3K/AKT Pathway and EnhancingAutophagy in Malignant Glioma Cell
Lines.
공동저자 PLoS ONE2012
e47449~e47449
4.092(국외SCIE)
1110110
PTEN Modulates miR-21 Processing viaRNA-Regulatory Protein RNH1
교신저자 PLoS ONE2011
6(12):e28308
4.411(국외SCIE)
1110110
Translation-competent 48S complexformation on HCV IRES requires the
RNA-binding protein NSAP1.공동저자
Nucleic AcidsRes.
20117791~7802
7.836(국외SCI)
없음
Downregulation of Spry2 by miR-21triggers malignancy in human
gliomaschemotherapy-naive patients withgood performance status and
adenocarcinoma histology: Retrospectiveanalysis of 575 Korean patients
공동저자 Oncogene2011
2433~2442
7.414(국외SCI)
1110110
4. 연구성과 및 목표달성도
(1) 연구성과
가. 국내 및 국제 전문학술지 논문 게재 및 신청
나. 국내 및 국제 학술대회 논문 발표

- 29 -
구분1) 특허명 출원인 출원국 출원번호
발명특허 METHOD OF REGULATINGMAMMALIAN
TARGET-OF-RAPAMYCIN ACTIVITYBY INTERACTION BETWEEN
PHOSPHOLIPASE D AND RAPTOR.
Sung Ho Ryu,Pann-Ghill SuhSang Hoon Ha,Do-Hyung Kim,
Il Shin Kim,Jung Hwan Kim,
Mi Nam Lee,Hyun Ju Lee,Jong Heon Kim,Sung Key Jang,Tae-Hoon Lee.
미국 PCT/KR07/03753.
072010-07-22.
다. 산업재산권

- 30 -
최종목표 연차별목표 달성내용달성도(%)연차 최종
종양마이크로RN
A의 turnover기전 연구
1차년도
1. PARN에 의한 종양억제마이크로RNA let-7의deuridylation 기전 규명
- RNA결합단백질 HuR이 Lin28B-TuT4oligo-uridylation 복합체의 구성분자로 동정함.- HuR이 마이크로RNA let-7의 가공을 가속시키는 조절자로 작용함을 밝힘.- HuR의 oligo-uridylation된 전구체 let-7 마이크로RNA에 특이적으로 결합함을 밝힘.
1 100%
2.TRIM71-Lin28B-let-7negative regulatory axis에 의한 종양억제기전 규명
- Lin28B분자는 세포 내에서 유비퀴틴화됨을발힘.- TRIM71은 Lin28B C-말단에 ~50 아미노산의 serine/lysine이 풍부한 서열에 특이적으로 결합하며 이러한 부위가 없는 Lin28A는결합을 하지 못함을 밝힘.- TRIM71은 post-Drosha 과정에서 Lin28Bprotein의 발현저해와 이에 따른 let-7 가공-생성과정을 조절함을 밝힘.
2차년도
1. PARN에 의한 종양억제마이크로RNA let-7의deuridylation 기전 규명
- HuR은 핵심 마이크로RNA 가공기구인Dicer 복합체와 결합하며 3′-5′deadenylase PARN이 그 복합체 상에 존재함을 밝힘.- 3′-5′ deadenylase PARN은 uridylation된 전구체 let-7 마이크로RNA에 대해 3′-5′exo-deuridylase activity를 지니고 있음을밝힘.- PARN의 과발현은 전구체 및 성숙 let-7마이크로RNA의 발현을 증대시킴을 밝힘.
2 100%2.TRIM71-Lin28B-let-7negative regulatory axis에 의한 종양억제기전 규명
- TRIM71의 RING finger motif가 Lin28B의유비퀴틴화, 결합, 그리고 let-7 조절에 매우중요함을 밝힘.
3. 뇌신경교종에서 DDX23RNA헬리케이즈에 의한종양 마이크로RNA-21의가공조절 연구
- 뇌신경교종환자에서 종양 miR-21의 발현이 전사후 단계에서 조절됨을 밝힘.- 종양 miR-21의 특이적 가공조절 조절 세포 내 분자로서 DDX23 RNA헬리케이즈를최초 분리동정함.- DDX23은 뇌신경교종세포주의 침윤과 증식을 증대시킴을 밝힘.
3차년도
1. PARN에 의한 종양억제마이크로RNA let-7의deuridylation 기전 규명
- 종양억제 마이크로RNA let-7 분해촉진인자 Lin28B의 S221 잔기가 특이적으로 인산화됨을 밝힘.- Lin28B의 S221A 돌연변이체 마이크로RNA let-7의 가공과정을 저해하는 기능을상실함을 밝힘.- Lin28B의 S221 인산화는 PARN을oligouridylation complex에서 해리시키고PARN에 의한 let-7 가공조절을 약화시킴을밝힘.
3 100%2.TRIM71-Lin28B-let-7negative regulatory axis에 의한 종양억제기전 규명
- TRIM71의 Lin28B에 의해서 유도되는cellular transforming 잠재력을 억제함을 밝힘.
3. 뇌신경교종에서 DDX23RNA헬리케이즈에 의한종양 마이크로RNA-21의가공조절 연구
- DDX23은 종양 miR-21의 발현을 조절함을밝힘.- DDX23은 종양 miR-21 발현을 Drosha단계에서 전사후 조절함을 밝힘.- DDX23은 Drosha microprocessor 및pri-miR-21과 특이적으로 결합함을 밝힘.- DDX23의 RNA헬리케이즈 활성이 종양miR-21 특이적 가공조절에 중요함을 밝힘.
(2) 목표달성도
가. 연구목표의 달성도

- 31 -
평가의 착안점 자 체 평 가
1. PARN에 의한 종양억제 마이크로RNA let-7의 deuridylation 기전 규명여부
- 중요 세포 내 RNA결합단백질 HuR이 Lin28B-TuT4oligo-uridylation 복합체 구성분자로 동정하고 마이크로RNAlet-7의 가공을 가속시키는 조절자로 작용함을 밝혔으며 핵심 마이크로RNA 가공기구인 Dicer 쪽으로 3′-5′ deadenylasePARN이 그 복합체 상에 존재함을 밝힘.
- 3′-5′ deadenylase PARN은 uridylation된 전구체 let-7 마이크로RNA에 대해 3′-5′exo-deuridylase activity를 지니고 있음을 밝힌 부분은 기대이상의 성과라고 판단할 수 있겠음.
- 종양억제 마이크로RNA let-7 분해촉진인자 Lin28B의 S221 잔기가 특이적으로 인산화됨을 밝혔으며 Lin28B의 S221 인산화는PARN을 oligouridylation complex에서 해리시키고 PARN에 의한let-7 가공조절을 약화시킴을 밝힘. 이는 암화과정의 Lin28B의 인산화과정이 매우 중요함을 시사하는 중요한 발견임.
2. TRIM71-Lin28B-let-7 negativeregulatory axis에 의한 종양억제기전규명 여부
- Lin28B분자가 세포 내에서 유비퀴틴화됨을 발혔으며 TRIM71은 Lin28B C-말단에 ~50 아미노산의 serine/lysine이 풍부한 서열에 특이적으로 결합하며 이러한 부위가 없는 Lin28A는 결합을하지 못함을 밝힘. 이는 마이크로RNA 가공조절이 번역후 조절에의해서 발생한다는 새로운 증거로서 매우 중요함.
- TRIM71은 post-Drosha 과정에서 Lin28B protein의 발현저해와 RING finger motif가 Lin28B의 유비퀴틴화, 결합, 그리고let-7 조절에 매우 중요함을 밝힘.
- TRIM71의 Lin28B에 의해서 유도되는 cellular transforming 잠재력을 억제함을 밝힘. 이는 암화과정에서 TRIM71의 역할을 새롭게 조망할 수 있는 중요한 관찰임.
3. 뇌신경교종에서 DDX23 RNA헬리케이즈에 의한 종양 마이크로RNA-21의가공조절 기전 규명 여부
- 종양 마이크로RNA-21의 특이적 가공조절 조절 세포 내 분자로서 DDX23 RNA헬리케이즈를 최초 분리-동정하였으며 DDX23은 뇌신경교종세포주의 침윤과 증식을 증대시킴을 밝히며 종양마이크로RNA-21의 발현을 조절함을 밝힘.
- DDX23은 Drosha microprocessor 및 pri-마이크로RNA-21과특이적으로 결합함을 밝혔으며 RNA헬리케이즈 활성이 종양 마이크로RNA-21 특이적 가공조절에 중요함을 밝힘. 이는 종양 마이크로RNA-21 가공과정과 이에 따른 RNA 수식단백질간의 관련성이 뇌신경교종의 발생, 침윤, 악성화와의 연관성을 보여준 중요한관찰이라 판단됨.
나. 평가의 착안점에 따른 목표달성도에 대한 자체평가
5. 연구결과의 활용계획
(1) 연구종료 2년후 예상 연구성과
구 분 건 수 비 고
학술지 논문 게재 3
Molecular Cell
(SCI-IF 14.178)BBA-Gene Regulatory Mechanism
(SCI-IF 5.456)
Journal of Clinical Investigation(SCI-IF 12.812)

- 32 -
6. 참고문헌
1. Heo I, Joo C, Cho J, Ha M, Han J, Kim VN.Lin28 mediates the terminal uridylation of let-7
precursor microRNA.Mol Cell. 2008;32(2):276-84.
2. Heo I, Joo C, Kim YK, Ha M, Yoon MJ, Cho J, Yeom KH, Han J, Kim VN.TUT4 in
concert with Lin28 suppresses microRNA biogenesis through pre-microRNA uridylation. Cell.
2009;138(4):696-708.
3. Martello G, Rosato A, Ferrari F, Manfrin A, Cordenonsi M, Dupont S, Enzo E, Guzzardo V,
Rondina M, Spruce T, Parenti AR, Daidone MG, Bicciato S, Piccolo S. A. MicroRNA targeting
dicer for metastasis control. Cell. 2010;141(7):1195-207.
4. Rybak A, Fuchs H, Hadian K, Smirnova L, Wulczyn EA, Michel G, Nitsch R, Krappmann
D, Wulczyn FG. The let-7 target gene mouse lin-41 is a stem cell specific E3 ubiquitin ligase
for the miRNA pathway protein Ago2. Nat Cell Biol. 2009;11(12):1411-20.
5. Su X, Chakravarti D, Cho MS, Liu L, Gi YJ, Lin YL, Leung ML, El-Naggar A, Creighton
CJ, Suraokar MB, Wistuba I, Flores ER. TAp63 suppresses metastasis through coordinate
regulation of Dicer and miRNAs. Nature. 2010;467(7318):986-90.
6. Trabucchi M, Briata P, Garcia-Mayoral M, Haase AD, Filipowicz W, Ramos A, Gherzi R,
Rosenfeld MG.The RNA-binding protein KSRP promotes the biogenesis of a subset of
microRNAs. Nature. 2009;459(7249):1010-4.
7. Viswanathan SR, Daley GQ, Gregory RI. Selective blockade of microRNA processing by
Lin28. Science. 2008;320(5872):97-100.
8. Viswanathan SR, Powers JT, Einhorn W, Hoshida Y, Ng TL, Toffanin S, O'Sullivan M, Lu
J, Phillips LA, Lockhart VL, Shah SP, Tanwar PS, Mermel CH, Beroukhim R, Azam M,
Teixeira J, Meyerson M, Hughes TP, Llovet JM, Radich J, Mullighan CG, Golub TR, Sorensen
PH, Daley GQ. Lin28 promotes transformation and is associated with advanced human
malignancies. Nat Genet. 2009;41(7):843-8.
9. West JA, Viswanathan SR, Yabuuchi A, Cunniff K, Takeuchi A, Park IH, Sero JE, Zhu H,
Perez-Atayde A, Frazier AL, Surani MA, Daley GQ.A role for Lin28 in primordial germ-cell
development and germ-cell malignancy. Nature. 2009;460(7257):909-13.
10. H.S. Gwak, T.H. Kim, G. H. Jo, Y.J. Kim, H.J. Kwak, J. H. Kim, J. Yin, H. Yoo, S. H. Lee, J. B.
Park. 2012. Silencing of MicroRNA-21 Confers Radio-Sensitivity through Inhibition of the PI3K/AKT
Pathway and Enhancing Autophagy in Malignant Glioma Cell Lines. PLoS ONE 7: e47449.
11. Y.J. Kim, S.J. Park, E.Y. Choi, S. Kim, H.J. Kwak, B.C. Yoo, H. Yoo, S.H. Lee, D. Kim, J.B.
Park, J.H. Kim, 2011, PTEN modulates miR-21 processing via RNA-regulatory protein RNH1, PLoS
ONE 6: e28308.
12. Kwak HJ, Kim YJ, Chun KR, Woo YM, Park SJ, Jeong JA, Jo SH, Kim TH, Min HS, Chae JS,
Choi EJ, Kim G, Shin SH, Gwak HS, Kim SK, Hong EK, Lee GK, Choi KH, Kim JH, Yoo H, Park JB,
Lee SH. 2011. Downregulation of Spry2 by miR-21 triggers malignancy in human gliomas.
Oncogene 30: 2433-2442.
13. J. Lu, G. Getz, E.A. Miska, E. Alvarez-Saavedra, J. Lamb, D. Peck, A. Sweet-Cordero, B.L.
Ebert, R.H. Mak, A.A. Ferrando, J.R. Downing, T. Jacks, H.R. Horvitz, T.R. Golub, 2005. MicroRNA
expression profiles classify human cancers, Nature 435: 834-838.
14. J.M. Thomson, M. Newman, J.S. Parker, E.M. Morin-Kensicki, T. Wright, S.M. Hammond, 2006.
Extensive post-transcriptional regulation of microRNAs and its implications for cancer. Genes Dev.
20 2202-2207.

- 33 -
7. 첨부서류

- 34 -

- 35 -

- 36 -

- 37 -

- 38 -

- 39 -