profesor patrocinante dr. pablo cid
TRANSCRIPT

Profesor Patrocinante Dr. Pablo Cid Centro de Estudios Científicos (CECs) Profesor Co-Patrocinante Dr. Alejandro Reyes Instituto de Bioquímica y Microbiología Facultad de Ciencias
ESTUDIO DE LA EXPRESIÓN DEL CANAL DE POTASIO TASK-2 DURANTE EL DESARROLLO Y EN EL RIÑÓN ADULTO: UTILIZACIÓN DE UN RATÓN NULO
PARA EL CANAL QUE EXPRESA β-GALACTOSIDASA COMO GEN REPORTERO.
Tesis de Grado presentada como parte de los requisitos para optar al Título Profesional de Bioquímico
IRMA KAREN LÓPEZ CAYUQUEO
VALDIVIA – CHILE
2013

Dedicado a mis padres Irma Cayuqueo e Iván López, y a mí hermano Jorge.

AGRADECIMIENTOS
Quiero agradecer a todas las personas que han contribuido a mi formación personal
y profesional durante estos años de carrera.
En primer lugar debo agradecer al Centro de Estudios Científicos que fue el lugar
donde se realizó esta tesis, a mi profesor patrocinante Pablo Cid, por la confianza
depositada en mí, por su apoyo y dedicación constante, por darme las herramientas
necesarias para mi formación profesional. También agradezco a Hugo Roa, por su
generosidad en el laboratorio, por estar siempre dispuesto a enseñarme y a
colaborar en mi trabajo. A Francisco Sepúlveda, Carlos Flores y Bredford Kerr por
sus siempre oportunos consejos, que más de alguna vez me ayudaron a resolver los
problemas que se presentaron en este trabajo.
A mis padres por su amor y confianza, a mi hermano quien con su ejemplo me
enseñó la perseverancia. A ellos debo todos lo que soy y todo lo que he logrado es
fruto de sus enseñanzas. A la señora Nelsa Muñoz por su generosa beca de
residencia que me brindo las mejores condiciones para poder estudiar.
Por último agradezco a mis amigos, a los que conocí en la universidad y a los que
están desde antes, a Pamela Garrido por su gran apoyo durante toda la carrera y a
Alejandro por todo su cariño. Finalmente a mis amigos del laboratorio por hacerme
grato cada día de trabajo, en especial a Francisca Julio.
Esta tesis fue realizada por el financiamiento del proyecto FODECYT 11100774.

i
ÍNDICE DE CONTENIDOS
Página
1. RESUMEN 1
1.1 Summary 2
2. INTRODUCCIÓN 3
2.1 Canales de potasio 3
2.2 El canal de potasio TASK-2 6
2.3 Un modelo murino nulo para TASK-2 9
2.4 TASK-2 en el desarrollo embrionario renal 10
2.5 Reabsorción de bicarbonato en los túbulos proximales 11
2.6 Hipótesis 14
2.6.1 Objetivo general 14
2.6.2 Objetivos específicos 15
3. MATERIALES Y MÉTODOS 16
3.1 MATERIALES 16
3.1.1 Reactivos 16
3.1.2 Anticuerpos 17

ii
3.1.3 Soluciones y tampones 17
3.2 METODOLOGÍA 18
3.2.1 Animales de experimentación 18
3.2.2 Generación del ratón nulo para TASK-2 18
3.2.3 Extracción de DNA genómico de ratón 19
3.2.4 Genotipificación de ratones mediante PCR 19
3.2.5 Caracterización del mensajero mutante para TASK-2 21
3.2.6 Tratamiento de embriones a los que se le reveló la actividad
de β-galactosidasa 23
3.2.7 Tratamiento de embriones y postnatales a los que se le reveló
la actividad de β-galactosidasa en cortes histológicos 23
3.2.8 Tratamiento de riñones de animales adultos a los que se le
reveló la actividad de β-galactosidasa en cortes histológicos 24
3.2.9 Tinción con eosina amarillenta 24
3.2.10 Tinción de ácido peryódico de Schiff (PAS) 25
3.2.11 Inmunohistoquímicas 25
3.2.12 Doble Inmunohistoquímica 26
3.2.13 Inmunofluorescencias 27
3.2.14 Extracción de RNA 28

iii
3.2.15 Tratamiento con DNAasa 29
3.2.16 Cuantificación del RNA 29
3.2.17 Retrotranscripción RT-PCR 30
3.2.18 PCR en tiempo real 30
3.2.19 Partidores utilizados para la PCR en tiempo real 32
4. RESULTADOS 34
4.1 Caracterización del mensajero mutante en el ratón nulo para
TASK-2 34
4.2 Expresión de TASK-2 durante el desarrollo embrionario y en
el postnatal 37
4.3 Expresión de TASK-2 durante el desarrollo renal 46
4.4 Expresión de TASK-2 en los túbulos proximales renales 44
4.5 TASK-2 se expresa con NBCe1-A y NHE3 en las células
del túbulo proximal donde estas proteínas participan en la
reabsorción del bicarbonato 56
4.6 Un posible rol compensatorio ante la ausencia de TASK-2 61
5. DISCUSIÓN 66
5.1 TASK-2 en el desarrollo embrionario 66
5.2 TASK-2 en la membrana basolateral de los túbulos proximales 69

iv
5.3 Búsqueda de una actividad compensatoria a la ausencia de
TASK-2 71
5.4 Conclusiones y proyecciones 73
6. REFERENCIAS 75

v
ÍNDICE DE FIGURAS
Página
Figura 1. Representación esquemática de la clasificación
estructural de canales de potasio 4
Figura 2. Topología estructural del canal de potasio TASK-2 8
Figura 3. Caracterización del mensajero mutante para TASK-2
mediante RT-PCR 36
Figura 4. Expresión de TASK-2 a los 9,5 dpc del desarrollo
embrionario 38
Figura 5. Expresión de TASK-2 en el desarrollo embrionario 40
Figura 6. Expresión de TASK-2 en la región cefálica de embriones
de 12,5 y 15,5 dpc 43
Figura 7. Expresión de TASK-2 en el eje dorsal de embriones
de 10,5; 12,5 y 15,5 dpc 44
Figura 8. Expresión de TASK-2 en la región cefálica y en el eje 45
dorsal de postnatales de 3 días.
Figura 9. Expresión de TASK-2 en tejido renal de embriones
de 12,5 dpc, 15,5 dpc y postnatales de 3 días 48
Figura 10. Expresión de TASK-2 en riñones de animales adultos
heterocigotos y silvestre 50

vi
Figura 11. Expresión de TASK-2 en la membrana basolateral
de túbulos renales corticales mediante inmunohistoquímica 54
Figura 12. Inmunohistoquímica antiTASK-2 con tinción de PAS 55
Figura 13. Doble inmunohistoquímica de TASK-2 y NHE3 57
Figura 14. NBCe1-A y TASK-2 se coexpresan en la membrana
basolateral de túbulos proximales 60
Figura 15. Inmunohistoquímica de Kir 7.1 en riñón 62
Figura 16. Expresión relativa de genes candidatos a compensar
la ausencia de TASK-2 64
Figura 17. Modelo de reabsorción del bicarbonato en túbulos
proximales 70

vii
ÍNDICE DE TABLAS
Página
Tabla I. Partidores utilizados para la genotipificación de los
ratones silvestres y TASK-2 nulos 20
Tabla II. Partidores utilizados para la identificación del mensajero
mutante para TASK-2. 22
Tabla III. Partidores utilizados para la PCR en tiempo real 33

viii
LISTA DE ABREVIACIONES
TASK-2: TWIK-related-acid-sensitive-K+
TWIK: Tamdem of P domains in a Weak Inwardly rectifying K+ channel
NHE3: Intercambiador de Sodio e Hidrógeno 3
NBCe1-A: Cotransportador electrogénico de bicarbonato de sodio e1-A
RT-PCR: Reacción en cadena de la polimerasa con transcriptasa inversa
cDNA: Ácido desoxiribonucleico complementario
5’-RACE-PCR: Amplificación rápida de extremos de DNA complementarios

1
1. RESUMEN
TASK-2 (KCNK5 o K2P5.1) es un canal de potasio que pertenece a la familia de canales
K2P, también denominados de tipo “background” porque determinan el potencial de
reposo de la célula. TASK-2 es gatillado por cambios en el volumen celular secundarios
a cambios osmóticos, y por cambios de pH tanto intra como extracelular. El ratón nulo
para TASK-2 presenta un fenotipo de acidosis metabólica tubular proximal leve y se ha
atribuido al canal un papel en la reabsorción renal de bicarbonato. En este trabajo se
utilizó dicho modelo que además de carecer del gen de TASK-2, expresa como gen
reportero la enzima β-galactosidasa bajo el control del promotor endógeno del canal. En
este ratón se estudió la expresión del canal evidenciado por la actividad del reportero
durante el desarrollo renal, encontrándose expresión desde los 12,5 dpc en el
metanefros. Adicionalmente el estudio durante el desarrollo embrionario, reveló nuevos
sitios de expresión para TASK-2, encontrándose que el canal comienza a expresarse
desde los 9,5 dpc en el eje dorsal y la región cefálica del embrión. A los 10,5 dpc
comienza a manifestarse expresión en meninges y desde los 12,5 dpc comienza su
expresión en tejido cartilaginoso. Finalmente utilizando el ratón nulo como el control
óptimo se pudo determinar la localización específica de la proteína del canal en la
membrana basolateral de los túbulos proximales. De esta forma en este trabajo se
demostró que TASK-2 se encuentra en las mismas células que expresan otras
proteínas esenciales para la reabsorción de bicarbonato, tales como el cotranspotador
NBCe1-A y el intercambiador NHE3. El fenotipo leve de acidosis metabólica sugiere una
actividad compensatoria ante la ausencia de TASK-2, por lo que se realizaron
experimentos preliminares para evaluar cambios en la expresión de genes candidatos.

2
1.1 Summary
TASK-2 (KCNK5 or K2P5.1) is a potassium channel that belongs to the family of K2P
channels, also called of "background" type because it determines the resting potential of
the cell. TASK-2 is triggered by changes in cell volume to secondary osmotic changes,
and by changes in intra and extracellular pH. The knock-out mouse for TASK-2 has a
mild phenotype of proximal tubular metabolic acidosis and it has been attributed to this
channel a role in renal reabsorption of bicarbonate. In this work we used this model that,
in addition to the lack of TASK-2 gene, expresses a reporter gene β-galactosidase
driven by the endogenous promoter of the channel. In this mouse the channel
expression was detected by the reporter activity during kidney development, finding
expression at 12.5 dpc in the metanephros. Additionally, the study during embryonic
development revealed new sites of expression of TASK-2, finding that the channel
begins to be expressed at 9.5 dpc in the dorsal axis and cephalic region of the embryo,
and around 10.5 dpc in meninges. On the other hand, TASK-2 also begins to show
expression in cartilage at 12.5 dpc. Finally, using as the optimal control this knock-out
mouse, it was determined that the channel protein is specifically located in the
basolateral membrane of proximal tubules. In this thesis we demonstrated that the
TASK-2 channel is found in the same cells that express other proteins essential for the
reabsorption of bicarbonate such as the NBCe1-A cotransporter and the NHE3
exchanger. The mild phenotype of metabolic acidosis suggests compensatory activity in
the absence of TASK-2, therefore, additional preliminary experiments were performed to
evaluate changes in the expression of candidate genes.

3
2. INTRODUCCIÓN
2.1 Canales de potasio
Los canales de potasio son proteínas integrales de membrana que permiten el flujo de
iones K+ a favor de su gradiente electroquímico. Conforman una familia muy extensa y
diversa cumpliendo funciones esenciales en todos los tipos celulares. En los epitelios,
se localizan tanto en la membrana basolateral como apical, cumpliendo diferentes
funciones relacionadas con el transporte de moléculas. Es así como en las células
epiteliales renales, participan en funciones tales como: generación y mantenimiento del
potencial de membrana celular, repolarización de la membrana necesaria para la
reabsorción sostenida de iones y solutos, reciclaje de potasio a través de la membrana
basolateral y regulación del volumen celular (Hamilton y Devor 2012).
Se han agrupado en 4 familias de acuerdo a su topología estructural, forma de
activación y relación filogenética (Shieh et al. 2000) a saber: canales de potasio
activados por voltaje (Kv), activados por calcio (KCa), rectificadores de entrada (Kir) y
tipo background (K2P).
Los canales de potasio activados por voltaje, están formados por un tetrámero de
subunidades α, donde cada una está formada por 6 segmentos de transmembrana y un
dominio que forma el poro. Estos canales poseen un sensor de voltaje en el cuarto
segmento de transmembrana, compuesto por aminoácidos cargados positivamente
(Figura 1A), capaces de detectar los cambios de voltaje y mediar el cambio
conformacional para abrir el canal cuando la membrana celular se depolariza.

4
Figura 1. Representación esquemática de la clasificación estructural de canales
de potasio. En A se representa una subunidad α de un canal de K+ activado por voltaje
(Kv), este posee 6 segmentos de transmembrana y un dominio formador de poro (P), el
sensor de voltaje se encuentra en el cuarto segmento de transmembrana. B muestra
una subunidad α de un canal de K+ rectificador de entrada (Kir), compuesto por 2
segmentos de transmembrana y un dominio formador de poro. En C está representada
una subunidad α de un canal de K+ K2P, estos poseen 4 segmentos de transmembrana
y 2 dominios formadores de poro.
+
+
+
+

5
Un grupo con características arquitectónicas muy similares a los Kv, son los canales de
potasio activados por calcio (KCa), los que también poseen 6 segmentos de
transmembrana; pero estos en el carboxilo terminal poseen un dominio de unión a
calmodulina, que les confiere la propiedad de ser activados por calcio.
El tercer grupo es de los canales rectificadores de entrada (Kir), estos también son
tetrámeros donde cada subunidad α posee dos dominios de transmembrana y un
dominio formador de poro (Figura 1B), la apertura del canal puede ser regulada tanto
por el carboxilo como por el amino terminal ambos ubicados intracelularmente. Estos
canales se denominan rectificadores de entrada porque poseen una mayor
conductancia de potasio hacia el medio citoplasmático a potenciales de membrana
hiperpolarizantes. A potenciales despolarizantes existe un bloqueo del poro del canal
por Mg2+ y poliaminas intracelulares, lo que disminuye el flujo de salida de los iones de
potasio.
Finalmente tenemos los canales K2P (Figura 1C), los cuales forman homodímeros,
donde cada subunidad es caracterizada por poseer cuatro dominios de transmembrana
y 2 dominios formadores de poro. Estos canales denominados de tipo background son
responsables del potencial de la membrana de prácticamente todas las células ya que
al encontrarse abiertos durante el reposo, el potencial de la membrana tiende al
potencial de equilibrio del potasio; también participan en la regulación del potencial de
acción en células excitables como neuronas y células musculares (Lauritzen et al.
2003). Estos canales son regulados por múltiples factores independientes de voltaje o

6
potencial de la célula, entre ellos destacan el pH, el volumen celular, temperatura, vías
de señalización intracelulares y ácidos grasos (Zúñiga et al. 2011).
2.2 El canal de potasio TASK-2
Dentro de la familia K2P encontramos a la subfamilia TASK (TWIK-related-acid-
sensitive-K+), a la cual pertenecen los canales TASK-1, TASK-3 y TASK-5 que son
inhibidos por acidificación extracelular. Una segunda subfamilia corresponde a los
canales TALK, compuesta por los canales TALK-1, TALK-2 y TASK-2 caracterizados
por ser activados por alcalinización extracelular (Zúñiga et al. 2011).
El canal de potasio TASK-2 (también llamado K2P5.1 o KCNK5) fue clonado por Reyes
et al., 1998 y se demostró la expresión del RNA mensajero en intestino delgado,
páncreas, hígado, riñón y placenta en humanos, y a su vez se encontró expresión en
hígado, páncreas y riñón en ratones. El marco de lectura del cDNA obtenido predice
que el canal posee 499 aminoácidos y una masa molecular estimada de 55 kDa. La
unidad funcional corresponde a un homodímero. Estudios de estructura función por
expresión heteróloga de TASK-2 demostraron inicialmente que el canal se activa a pH
alcalino extracelular y por hinchazón celular debido a hipotonicidad del medio
extracelular (Reyes et al. 1998; Niemeyer et al. 2001; Barriere et al. 2003). Luego se
identificó el aminoácido sensor del pH extracelular correspondiendo a la cadena lateral
del residuo de arginina 224 (R224) ubicado en la región externa cercano al segundo
dominio P (Niemeyer et al., 2007), la deprotonación de la R224 se asocia a un aumento
en la probabilidad de apertura del canal sin afectar su conductancia (Reyes et al. 1998).
Más recientemente se ha demostrado que TASK-2 también es regulado por el pH

7
intracelular, donde la alcalinización aumenta la actividad del canal, el sensor
corresponde a la cadena lateral del residuo de lisina 245 (K245) localizado en el
carboxilo terminal intracelular, como se aprecia en la Figura 2 (Niemeyer et al. 2010).
Recientemente también se ha demostrado que TASK-2 es modulado por la subunidad
Gde proteína G heterotrimérica, la unión de esta subunidad provoca el cierre del
canal. Esta modulación depende de una interacción física entre las subunidades
GGy residuos de lisina (K257-K258) ubicadas en el carboxilo terminal del canal
(Añazco y Peña-Münzenmayer 2013).
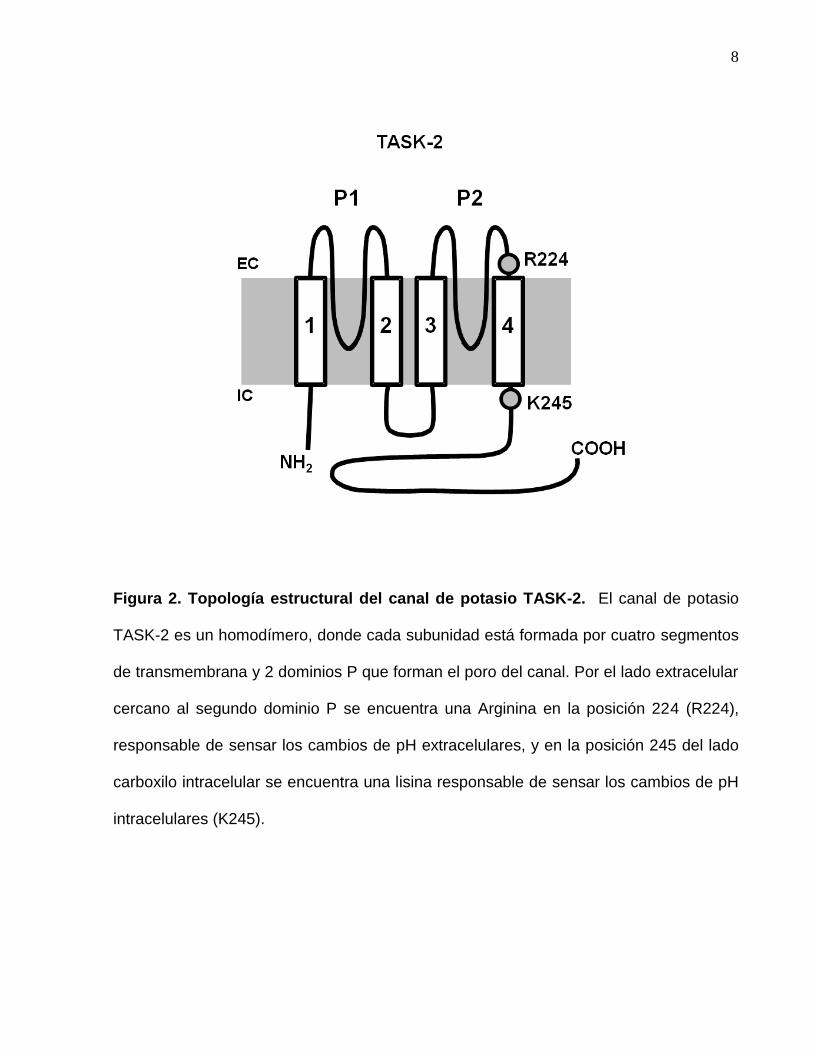
8
Figura 2. Topología estructural del canal de potasio TASK-2. El canal de potasio
TASK-2 es un homodímero, donde cada subunidad está formada por cuatro segmentos
de transmembrana y 2 dominios P que forman el poro del canal. Por el lado extracelular
cercano al segundo dominio P se encuentra una Arginina en la posición 224 (R224),
responsable de sensar los cambios de pH extracelulares, y en la posición 245 del lado
carboxilo intracelular se encuentra una lisina responsable de sensar los cambios de pH
intracelulares (K245).

9
2.3 Un modelo murino nulo para TASK-2
El ratón nulo para TASK-2 fue generado por el método de atrapamiento génico (gene
trap). En este método se interfiere la transcripción normal de un gen mediante la
inserción al azar en el genoma de células totipotentes, de un vector (gene trap vector)
que codifica para una proteína de fusión entre la secuencia del gen reportero β-
galactosidasa y la secuencia de resistencia a neomicina. En el extremo 5´ posee una
secuencia aceptora de empalme, que aprovecha la transcripción endógena e
interrumpe el empalme normal permitiendo de esta manera la transcripción del vector
río abajo. Finalmente en el extremo 3´ el vector posee una señal de poliadenilación que
detiene la transcripción.
Las células totipotentes que presentan la expresión del vector son seleccionadas
mediante la resistencia a neomicina. El método para identificar el gen interferido se
denomina 5´RACE-PCR y consiste en obtener región río arriba del transcrito la
secuencia correspondiente al exón 1 del gen interferido (Scotto-Lavino, Du, y Frohman
2006). Luego estás células son microinyectadas en blastocistos que son transferidos a
ratonas pseudopreñadas. Las crías quiméricas son cruzadas con animales silvestres
obteniéndose heterocigotos que finalmente al cruzarse entre sí darán origen a animales
homocigotos nulos para el gen de TASK-2 (Araki, Araki, y Yamamura 2009).
Los estudios en el ratón nulo para el canal, lograron determinar funciones del canal
previamente insospechadas, por ejemplo, se demostró su presencia en el núcleo
retrotrapezoide del tronco encefálico, que es parte de la red respiratoria del sistema
nervioso central, donde el canal participa en la quimiorrecepción (Gestreau et al. 2010).

10
También se determinó su expresión en el riñón, específicamente en el túbulo proximal y
en el colector papilar. Funcionalmente se demostró que la ausencia de TASK-2 impide
la correcta reabsorción de bicarbonato, originándose un fenotipo de acidosis metabólica
(Warth et al. 2004). En esta tesis se utilizará un ratón nulo para el canal diferente al
utilizado por los autores recién señalados y se determinará inicialmente el mensajero
mutante hasta ahora no caracterizado.
2.4 TASK-2 en el desarrollo embrionario renal
El desarrollo renal en el ratón, se divide en 3 etapas muy definidas, la primera comienza
entre los 8,5 y 9,5 dpc (días post coito) con la formación del cordón nefrogénico del cual
se forma el pronefros, el cordón nefrogénico deriva de la migración en sentido craneal a
caudal de las células del mesodermo intermedio. Entre los 10,5 y los 11,5 dpc se
observa el mesonefros, aquí se evidencian los glomérulos primitivos. Finalmente la
tercera etapa del desarrollo embrionario está determinada por la formación del
metanefros, el cual empieza a manifestarse aproximadamente a los 12,5 dpc y éste es
el que finalmente dará origen al riñón funcional y definitivo (Genito Urinary Molecular
Anatomy Proyect, GUDMAP, www.gudmap.org).
En mamíferos, el embrión es protegido de los cambios osmóticos del ambiente externo
por su desarrollo en el útero y el problema de la excreción de residuos es mantenido
por la alantoides y la placenta. Es por ello que se reducen los requerimientos
funcionales del pronefros y mesonefros, por lo que en mamíferos estas estructuras
involucionan, manteniéndose por un corto tiempo en el desarrollo embrionario
(Davidson 2008). De acuerdo a estos antecedentes es esperable que TASK-2 comience

11
a presentarse desde los 12,5 dpc, cuando la formación del metanefros inicia el
desarrollo del riñón definitivo.
En esta tesis se evaluarán otros sitios de expresión de TASK-2 en el desarrollo
embrionario, para tener una idea general de los sitios de expresión del canal y buscar
posibles nuevas funciones fisiológicas para TASK-2.
2.5 Reabsorción de bicarbonato en los túbulos proximales
En el riñón el 80% del bicarbonato es reabsorbido en el túbulo contorneado proximal, en
el proceso se ven involucradas varias proteínas ubicadas en la membrana apical y
basolateral, como también citosólicas. El proceso ocurre de la siguiente manera: el
bicarbonato del filtrado presente en el lumen tubular se titula con los protones
excretados desde el epitelio del túbulo proximal. Esta reacción es catalizada por la
anhidrasa carbónica IV, la cual se encuentra en la membrana luminal externa; como
producto de la reacción se generan CO2 y agua. El dióxido de carbono difunde hacia el
espacio intracelular donde junto con el agua son sustratos de la anhidrasa carbónica II,
formándose como producto protones y bicarbonato. Los protones activan al
intercambiador luminal NHE3, que transporta protones hacia el lumen contra su
gradiente junto con la entrada Na+ hacia el intracelular, utilizando la gradiente generada
por la bomba de Na+/K+ ATPasa. El bicarbonato producido es transportado hacia el
intersticio mediante el cotrasportador basolateral NBCe1-A, el cual es electrogénico ya
que permite la salida de 1 Na+ y 3 HCO3- , transportándose dos cargas negativas hacia
el espacio extracelular por ciclo. De esta manera se produce la depolarización de la
membrana plasmática, limitándose el transporte de bicarbonato hacia la sangre, el cual

12
debe ser sostenido para evitar de esta manera su pérdida en la orina. La activación del
canal TASK-2 en el túbulo proximal renal es requerida para mantener la fuerza motriz
para la reabsorción del bicarbonato (Warth et al. 2004), ya que permite la repolarización
de la membrana. La ausencia del mecanismo para repolarizar la membrana puede
resultar en un decaimiento de la reabsorción de bicarbonato de sodio en el túbulo
proximal, produciéndose la consecuente pérdida de HCO3-, Na+ y H2O por diuresis
osmótica, desencadenándose un fenotipo de acidosis metabólica en los ratones nulos
para TASK-2.
Sin embargo el grado de acidosis metabólica que se manifiesta en los ratones TASK-2
nulos es leve, comparada con el ratón nulo para NBCe1-A el que presenta un fenotipo
de acidosis metabólica severa con una alta mortalidad, no sobreviviendo más allá de los
24 días (Gawenis et al. 2007). Igual fenotipo presentan los animales knock-in que
expresan la mutación W516X encontrada en pacientes humanos en la secuencia
codificante para NBCe1-A (Lo et al. 2011).
Una posible consecuencia de la eliminación genética de una proteína es la expresión
compensatoria de alguna otra u otras que realicen en parte la función eliminada. Como
es en el caso del modelo murino nulo para NHE3, donde ante la ausencia de este
intercambiador luminal de Na+/H+ se provocó el aumento de la expresión de
aldosterona, renina y del intercambiador Cl-/HCO3- AE1, produciéndose de esta manera
sólo un fenotipo leve de acidosis metabólica (Schultheis et al. 1998). Una situación
análoga podría darse en el caso del ratón nulo para TASK-2 como es un canal activado
por pH e hipotonicidad expresado en el túbulo proximal renal, nos sugiere la búsqueda

13
de otro canal con características similares que podría estar realizando en parte esta
actividad compensatoria. En una revisión reciente se detallan los canales de potasio
expresados en la membrana basolateral del túbulo proximal (Hamilton y Devor 2012).
Se encuentran hasta ahora descritos miembros de las familias Kir y K2P. Entre estos
últimos, aparte de TASK-2 están THIK 1 y 2, el primero es activado por ácido
araquidónico y no es influido por el pH y el segundo no se ha logrado expresar
heterólogamente. Respecto del grupo de los Kir, encontramos los canales Kir5.1 que
forma un canal heterodimerico junto a Kir4.1, este es inhibido por pH intracelular y la
sensibilidad a pH depende de Kir5.1. Por otra parte en túbulos proximales de cobayo y
humanos se ha detectado la presencia del canal Kir7.1 (Derst y Hirsch 2001) que es
inhibido por acidificación extracelular y modulado por pH intracelular de manera bifásica
(Hughes y Swaminathan 2008). Las características expuestas de estos canales los
hacen posibles candidatos para suplir parcialmente la función de TASK-2. Por otra parte
la compensación también podría estar dada por un aumento de la expresión de NHE3 o
NBCe1-A.
Los antecedentes mencionados asumen la localización subcelular de TASK-2 en la
membrana basolateral de los túbulos proximales, sin embargo cabe destacar que hasta
ahora esto no se ha logrado demostrar, por lo que es un objetivo del presente trabajo.

14
2.6 Hipótesis
Considerando la función propuesta en la reabsorción renal proximal de bicarbonato el
canal de potasio TASK-2:
- Se expresa en la membrana basolateral de los túbulos proximales.
- Aparece en el desarrollo embrionario a partir de la formación del metanefros.
- Su ausencia por deleción genética en un modelo murino nulo es parcialmente
compensada.
2.6.1 Objetivo general
Determinar la expresión renal de TASK-2 y en el desarrollo embrionario, ya sea por la
actividad del reportero -galactosidasa o por inmunolocalización de la proteína del
canal. Evaluar una potencial actividad compensatoria a nivel transcripcional de los
canales Kir5.1, 4.1, 7.1 y de los transportadores NHE3 y NBCe1-A en el modelo murino
nulo.

15
2.6.2 Objetivos específicos
1. Caracterizar el mensajero mutante en el ratón nulo de TASK-2.
2. Determinar la expresión de TASK-2 en el desarrollo embrionario y postnatales,
además de la expresión en el desarrollo renal.
3. Determinar la localización subcelular de TASK-2 en los túbulos renales, mediante la
inmunolocalización de la proteína del canal.
4. Estudiar cambios de expresión génica mediante RT-PCR cuantitativa de genes
candidatos a compensar la falta de TASK-2: los canales Kir4.1, 5.1, 7.1, el
intercambiador NHE3 y el cotransportador NBCe1-A.

16
3. MATERIALES Y MÉTODOS
3.1 MATERIALES
3.1.1 Reactivos:
5x Green GoTaq reaction buffer (Promega), 5-bromo-4-chloro-3-indolil-beta-D-
galactosido (X- Gal) (Gibco-invitrogen), 6x DNA Loading Dye (Fermentas), Agarosa
(Lafken), Avertina 2% (Sigma), Ácido Acético glaciar (Merck), Bromuro de Etidio 1
mg/ml (Sigma), BSA (Winkler), dNTPs (Promega), DNA Taq polimeras, Deoxicolato de
sodio 0.01% (Sigma), Dietilpirocarbonato (sigma), Desoxicolato de sodio (sigma), Etanol
(Merck), EDTA (Sigma), Eosina Amarillenta (Winkler), Gene ruler 1 Kb DNA ladder
(Fermentas), Gene ruler 100 pb DNA ladder, Isopropanol (Merck), Igepal (Sigma),
Glycoblue (Fermentas), ImmPRESSTM Reagent Anti-Rabbit IG Peroxidase (Vector
Laboratories), ImmPACTTM DAB Peroxidase Substrate (Vector Laboratories), KCl
(Merck), KH2PO4 (Merck), K4[Fe(Cn)6]x3H20 (Merck), K2Fe(Cn)6 (Sigma), Kit de
retrotranscripción (Invitrogen), Kit de tinción de PAS (millipore), Kapa SYBR® FAST
qPCR Kit Master Mix 2X Universal (Kapabiosystems), MgCl2 (Sigma), NaCl (Merck),
NaOH (Sigma), NGS (sigma), Na2HPO4 (Merck),Paraformaldehído (Sigma), Permount
(Fisher Scientific) Proteinasa K (Bioline), RNAlaterTM (Ambion), Sacarosa (Merck), SDS
(Sigma), SuperScriptTM II Reverese Trancriptase (Sigma), Tris–HCl (Sigma), Tris-
Acetato (Sigma), Tris–Base (Sigma), Trizol® (Invitrogen), Triton X-100 (Sigma), TurboTM
DNAasa (Ambion), Tissue-Tek® O.C.T Compound (Sakura), Tween 20 (Sigma),
Vector®SG Substrate Kit For Peroxidase.

17
3.1.2 Anticuerpos:
Rabbit anti NA+/H+ exchanger 3 (NHE3) (Millipore), Rabbit Anti-Potassium Channel
TASK-2 Affinity Isoleted (Sigma), Anti SLC4A4 (Abcam), Anti-Kir7.1 (extracellular)
(Alomone Labs), Goat Anti Rabbit IgG (H+L) (Jackson Laboratories), Alexa Fluor® 488
goat anti-rabbit IgG (Invitrogen).
3.1.3 Soluciones y tampones:
TEL: Tris-HCl 50 mM pH 8,0, EDTA 50 mM, NaCl 100 mM, SDS 0.5%.
TE: Tris-HCl 10 mM pH 8,0, EDTA 1 mM pH 8.0
TAE 1X: Tris-acetato 40 mM, EDTA 1 mM.
PBS pH 7.4: NaCl 137 mM, KCl 2.7 mM, Na2HPO4 10 mM, KH2PO4 2 mM.
Solución de revelado para β-galactosidasa: K4[Fe(Cn)6]x3H20 5 mM, K2Fe(Cn)6 4.3
mM, MgCl2 2 mM, deoxicolato de sodio 0.01% 2.4 mM, Igepal 0.02%. Ajustar pH a 8,5.
TTBS pH 7.4: Tris-base 20 mM, NaCl 137 mM, Tween 20 0.1%.
Solución de exposición de epítope: Tris-HCl 250 mM, EDTA 10 mM. Ajustar pH a 8,5.
Tampón Rinse: 2 mM MgCl2, 0.02% Igepal, 0.2 mM desoxicolato de sodio, Preparar en
PBS pH 7,4.

18
3.2 METODOLOGÍA
3.2.1 Animales de experimentación.
Los ratones utilizados en esta tesis corresponden a animales TASK-2 nulos y silvestre
de 2 a 3 meses de vida, además de embriones en los estadios embrionarios que van
desde los 9,5 dpc (días post coito) a los 15,5 dpc y postnatales de 3 días, Mus
musculus. Estos fueron criados en las dependencias del Laboratorio de Genómica
Funcional del Centro de Estudios Científicos (CECs), en condiciones de SPF (Specific
Patogen Free), bioterio acreditado por la AAALAC (Association for Assessment and
Acreditation of Laboratory Animal Care intenational). Los animales fueron alimentados
ad libitum con pellet Harlan y sometidos a ciclos de luz/oscuridad de 12 horas. Los
métodos utilizados para la mantención, manipulación y eutanasia de los animales
fueron aprobados por el Comité de Ética del CECs y siguen la línea del Comité Asesor
de Bioética de CONICYT. Las carcasas fueron enviadas a dependencias de la
Universidad Austral de Chile, donde fueron incineradas.
3.2.2 Generación del ratón nulo para TASK-2.
El ratón nulo para TASK-2 con el que se trabajó en esta tesis se adquirió en el Center
for Animal Resources and Development, Kumamoto University Japan. Este se realizó
mediante el método de atrapamiento génico descrito en 2.3.
El gen que codifica para TASK-2 (KCNK5) está formado por 5 exones y entre el exón 1
y 2 se insertó la secuencia codificante para le proteína reportera β-galactosidasa (βgal).

19
De esta manera podemos estudiar de manera indirecta la expresión del canal al revelar
la actividad de β-gal directamente en tejido.
3.2.3 Extracción de DNA genómico de ratón.
El DNA genómico se extrajo a partir de una biopsia de cola de ratón, se digirió el tejido
con 10 μL de Proteinasa K (10 mg/mL) en 500 μL de solución TEL, se incubó por 12
horas a 55ºC en agitación constante a 750 rpm. Posteriormente, se centrifugó a 14.000
rpm por 10 min y se conservó el sobrenadante al que se le agregaron 1 mL de etanol
100%. Se centrifugó a 14.000 rpm por 10 min, se eliminó el sobrenadante y se lavó el
precipitado con etanol 70%, se volvió a centrifugar a 14000 rpm por 10 min y el
precipitado se dejó secar a temperatura ambiente. Luego fue resuspendido con tampón
TE y dejado en agitación constante a 750 rpm por 2 horas a 60ºC. El DNA genómico se
almacenó a 4ºC.
3.2.4 Genotipificación de ratones mediante PCR.
Para identificar los animales que son TASK-2 nulos se realizaron dos reacciones de
PCRs por separado y se utilizaron 3 partidores, de los cuales se forman dos juegos
compartiendo un partidor en cada reacción. Los partidores utilizados se indican en la
Tabla I.

20
Tabla I. Partidores utilizados para la genotipificación de los ratones silvestres y TASK-2
nulos.
For T2-KO-2181-4
5’-GCT GAG AAC AAT AGG GAC AG-3’
Tm: 52,7°C
Rev T2-KO-2181-7
5’-TCA CCC AGC TTT GGG ATT CC-3’
Tm: 57,5°C
For T2-KO-Amp-S
5’-TAC AGG CAT CGT GGT GTC AC-3’
Tm: 57,2°C

21
La mezcla de la reacción contuvo 0,5 μL de DNA genómico como templado; 10 μM de
cada partidor; 5 mM de dNTPs, 25 mM MgCl2 y 0,4 μL de enzima Taq polimerasa en un
volumen final de 20 μL. El programa del termociclador fue el siguiente: denaturación
inicial a 94ºC por 4 min, seguido de 30 ciclos de denaturación a 94ºC por 50 segundos,
alineamiento a 60ºC por 30 segundos, elongación a 72ºC por 1 min y por último una
elongación final a 72ºC por 5 min.
La visualización de los productos de PCR se realizó a través de una electroforesis en
gel de agarosa al 1,2% teñido con bromuro de etidio, el tampón de corrida empleado
fue TAE 1X y el voltaje de corrida 80 V por 50 min. El tamaño del producto de PCR se
determinó por comparación con un patrón de peso molecular de 100 pb. Las fotografías
del gel fueron tomadas en un transiluminador conectado a un computador (Ultraviolet
Transilluminator BioImaging System) y fueron procesadas en el software de imágenes
Labworks (UVP Bioimaging System).
3.2.5 Caracterización del mensajero mutante para TASK-2.
La caracterización del mensajero mutante para TASK-2 se realizó mediante RT-PCR,
utilizando los partidores indicados en la Tabla II.

22
Tabla II. Partidores utilizados para la identificación del mensajero mutante para TASK-2.
TASK2E1 F
5’-TCA CCT CGG CCA TCA TAT TC-3’
Tm: 54,4 °C
TASK2E4 R
5’-GTA GAG GCC CTC AAT GTA GTT CCA-3’
Tm: 58,1°C
TASK2Ex2R
5’- TGA TCC GCA GCA TCA GAC ACC-3’
Tm: 59,7°C
BGal02R
5’-TG TGC AGT TCA ACC ACC GCA C-3’
Tm: 60,9°C

23
3.2.6 Tratamiento de embriones a los que se les reveló la actividad de β-
galactosidasa en tejido completo.
Los embriones fueron removidos del útero materno y lavados con PBS pH 7,4. Luego
estos fueron fijados con PFA (paraformaldehído) 4% pH 7,4, incubando los embriones
de 9,5 y 10,5 dpc 30 min, los de 12,5 y 14,5 dpc por 1 hora.
Luego de ser fijados fueron lavados con tampón Rinse por 30 minutos y sometidos a un
protocolo de aclaramiento que se basa en la incubación de los embriones en soluciones
que contienen concentraciones crecientes de glicerol y decrecientes de KOH, durante 4
días a temperatura ambiente, en agitación. Se utilizó una solución madre de KOH al
1%. El primer día la solución estuvo compuesta por un 20% de glicerol y un 80% de
KOH al 1%, el segundo día un 50% de glicerol, el tercero un 80% y el cuarto día un
100% de glicerol.
3.2.7 Tratamiento de embriones y postnatales a los que se les reveló la actividad
de β-galactosidasa en cortes histológicos.
Desde el útero materno fueron extraídos embriones de 12,5 y 15,5 dpc, estos fueron
genotipificados mediante PCR.
Una vez extraídos del útero materno los animales fueron lavados 3 veces por 5 minutos
con PBS pH 7,4 y fijados posteriormente en PFA 4% pH 7,4 por 1 hora. Luego los
embriones fueron deshidratados en sacarosa al 30% por 12 horas y embebidos en OCT
para posteriormente ser cortados en criostato a 12 micras.

24
Una vez realizados los cortes se procedió a lavarlos con PBS pH 7,4 y a revelar la
actividad de β-gal dejando los tejidos 12 horas a 37ºC en solución de revelado y X-gal
con una concentración final de un 1 mg/mL. Luego los tejidos son nuevamente lavados
y teñidos con eosina
Los postnatales de 3 días fueron tratados de igual manera, a excepción de que estos
fueron decapitados y posteriormente fijados en PFA al 4% por 1 H 30 min.
3.2.8 Tratamiento de riñones de animales adultos a los que se les reveló la
actividad de β-galactosidasa en cortes histológicos.
Los animales heterocigotos y silvestres fueron perfundidos vía ventrículo izquierdo con
PFA 4% pH 7,4. Estos fueron previamente anestesiados con avertina al 2%. Una vez
extraídos los riñones fueron postfijados en PFA 4% pH 7,4 por 2 horas y luego
deshidratados en sacarosa al 30% por 12 horas.
Los riñones deshidratados fueron colocados en criomoldes y embebidos en OCT, para
luego ser cortados en criostato a 12 micras. Finalmente se reveló la actividad de β-
galactosidasa mediante el método anteriormente descrito (3.2.7).
3.2.9 Tinción con eosina amarillenta.
Los tejidos fueron puestos en agua destilada y luego en eosina por 30 segundos,
posteriormente son deshidratados con etanol 70%, 80%, 96% y 100% pasando los
tejidos dos minutos por cada etanol, finalmente se colocaron en xilol por 1 minuto y
nuevamente en xilol por 30 segundos. Las preparaciones fueron montadas con 50 uL
de Permount.

25
3.2.10 Tinción de ácido peryódico de Schiff (PAS).
Los tejidos fueron desparafinados y rehidratados, luego se enjuagó con agua destilada,
las muestras fueron incubadas con ácido peryódico por 5 min y posteriormente lavadas
en agitación constante por 3 min. Se incubaron con reactivo de Schiff por 15 min y
nuevamente se lavó con agua destilada en agitación por 3 minutos, se tiñó con
hematoxilina por 1 min, y se lavó nuevamente por 3 minutos. Los tejidos fueron
deshidratados.
3.2.11 Inmunohistoquímicas.
Se realizaron cortes de riñón en parafina de 4 micras, los que fueron desparafinados
mediante el siguiente procedimiento: Los cortes fueron colocados en bórreles con Xilol
por 3 min, nuevamente en xilol por 3 min, etanol 100%, metanol absoluto, etanol 96% y
etanol 70% por 3 min en cada uno. Las muestras fueron lavadas 3 veces por 5 min con
TTBS pH 7,4, luego se realizó la exposición de epítope con tampón TRIS-EDTA pH 8,5
en microondas por 23 min a máxima potencia (el tiempo debe ser necesario para que
se alcance una temperatura de 97ºC) e inmediatamente se cambió la potencia a 60%
por 6 min para mantener la temperatura. Las muestras se dejaron enfriar por 20 min a
temperatura ambiente y se lavaron. Posteriormente se bloqueó la actividad endógena
de la peroxidasa con peróxido de hidrógeno al 10% por 10 min y se lavó nuevamente.
Adicionalmente se bloqueó con NH4Cl por 30 min, para evitar las uniones inespecíficas
de inmunoglobulinas. Se bloqueó con suero de caballo del KIT Immpress REAGENT
Anti-Rabbit Ig por 1 hora y se lavó. Se aplicó anticuerpo primario (antiTASK-2,
antiNBCe1-A, antiNHE3 o antiKir7.1 en la dilución 1:1000) por 1 hora a temperatura

26
ambiente, el anticuerpo fue diluido en TRIS-HCl pH 8,0 más BSA 1%. Luego se lavó y
se aplicó el polímero del KIT Immpress REAGENT Anti-Rabbit Ig por 30 min, se lavó y
se aplicó ImmPACT DAB peroxidase substrate por 5 min. Nuevamente se lavó y se tiñó
con hematoxilina por 10 segundos y se lavó. Finalmente las muestras fueron
deshidratadas de la siguiente manera: fueron colocadas en etanol 70%, 96% y 100%
por 3 minutos en cada uno, luego las muestras fueron pasadas a xilol por 3 minutos y
nuevamente a xilol por 3 min. Las muestras fueron montadas con Permount y dejadas
secar.
3.2.12 Doble inmunohistoquímica.
Se realizaron cortes de riñón de 4 micras en parafina, luego los cortes fueron
desparafinados como se indica en 3.3.11. Las muestras fueron lavadas 3 veces por 5
min con TTBS pH 7,4, luego se realizó la exposición de epítope con tampón TRIS-
EDTA pH 8.5. Las muestras se dejaron enfriar por 20 min a temperatura ambiente y se
lavaron, posteriormente se bloqueó la actividad endógena de la peroxidasa con
peróxido de hidrógeno al 10% por 10 min y se lavó. Adicionalmente se bloqueó con
NH4Cl por 30 min. Se bloqueó con suero de caballo del KIT Immpress REAGENT Anti-
Rabbit Ig por 1 hora y se lavó, se aplicó anticuerpo primario (antiTASK-2 1:1000) por 1
hora a temperatura ambiente. El anticuerpo fue diluido en TRIS-HCl pH 7.8 más BSA
1%. Luego se lavó 3 veces y se aplicó el polímero del KIT Immpress REAGENT Anti-
Rabbit Ig por 30 min, se lavó y se colocó ImmPACT DAB peroxidase substrate por 5
min. Se aplicó el segundo anticuerpo primario antiNHE3 (dilución 1:1000) por 1 hora a
temperatura ambiente y luego se lavó. Se incubó nuevamente con el polímero del KIT

27
Immpress REAGENT Anti-Rabbit Ig por 30 min, se lavó 3 y se incubó con SG substrate
kit for peroxidase por 10 min. Nuevamente se lavó y se tiñó con hematoxilina por 10
segundos y se lavó. Finalmente las muestras fueron deshidratadas como se indica en
3.2.11. Las muestras fueron montadas con Permount y dejadas secar.
3.2.13 Inmunofluorescencias.
Se realizaron cortes de riñón de 4 micras en parafina, luego los cortes fueron
desparafinados mediante el siguiente procedimiento: Los cortes fueron colocados en
bórreles con Xilol por 5 min, nuevamente en xilol por 5 min, etanol 100%, 96%, 70% por
5 min en cada uno. Las muestras fueron lavadas 3 veces por 5 min con TTBS pH 7,4,
luego se realizó la exposición de epítope, como se describió previamente. Las muestras
se dejaron enfriar por 20 min a temperatura ambiente y se lavaron 3 veces con TTBS
pH 7,4 por 5 min, posteriormente se bloqueó con NH4Cl por 30 min, nuevamente se
lavó 3 veces con TTBS pH 7,4 por 5 min. Se bloqueó con NGS 10% más 3% BSA en
PBS tritón X-100 por 1 hora, se aplicó anticuerpo primario (antiTASK-2 en la dilución
1:400 por 12 horas a 4º C o antiNBCe1-A en la dilución 1:1000 por 1 hora a temperatura
ambiente; el anticuerpo fue diluido en solución de bloqueo), se lavó 6 veces con TTBS
pH 7,4 por 10 minutos en agitación. Finalmente se incubó con anticuerpo secundario
Goat anti rabbit IgG (H+L) de Jackson Laboratories en la dilución 1:500 por 1 hora (Para
TASK-2) o Goat anti rabbit Alexa 448 de Invitrogen en la dilución 1:500 por 30 min. Se
lavó 6 veces con TTBS pH 7,4 por 10 minutos en agitación. Se montaron las muestras
con DAKO.

28
3.2.14 Extracción RNA.
La extracción de RNA se realizó mediante el siguiente procedimiento:
Homogenización del tejido: Se agregó 1 mL de Trizol por cada 50-100 mg de tejido de
corteza renal. Se homogenizó con un homogeneizador impulsado por un taladro hasta
que se desintegró todo el tejido. Luego se incubó por 5 min a temperatura ambiente
para disociar los complejos nucleoproteicos, inmediatamente se centrifugó a 12000 x g
por 10 min a 4ºC. Se tomó el sobrenadante y se pasó a un nuevo tubo.
Separación de fases: Las muestras fueron incubadas a temperatura ambiente por 5
minutos, se añadió 0,2 ml de cloroformo por mL de trizol, se agitó vigorosamente por 15
segundos y se incubó 3 min a temperatura ambiente. Se centrifugó a 12000 x g por 15
min a 4º C y se traspasó la fase acuosa que contenía el RNA a otro tubo.
Precipitación del RNA: Se agregó 1 µL de Glyco Blue a cada muestra (este coprecipita
con el RNA, observándose el RNA de color azul en el fondo del tubo). Se agregó 0,5
mL de isopropanol 100% por cada mL de Trizol y se dejó precipitar por 10 min
temperatura ambiente, se centrifugó 12000 x g por 30 min a 4º C.
Lavado del RNA: Se removió completamente el sobrenadante dejando solo el pellet que
correspondiente al RNA, se lavó con 1 mL de etanol 75% (preparado con agua DEPC)
por mL de trizol. Se agitó suavemente y se centrifugó a 7500 x g por 5 min a 4º C, se
descartó la solución de lavado. Se dejó secar el pellet de RNA al aire, a temperatura
ambiente por un tiempo prudente (5 a 10 min), no dejando secar completamente ya que
se puede perder solubilidad.

29
Resuspensión del RNA: El RNA se resuspendió en 35 µL de agua libre de nucleasas
por pipeteo y se incubó a 55ºC por 15 min. Finalmente fue almacenado a -80ºC
3.2.15 Tratamiento con DNAasa.
Por cada muestra de RNA, se preparó un tubo aparte conteniendo 4 µL de buffer
DNAsa y 1 µL de DNAasa, se agregó la totalidad del RNA (la concentración de RNA no
tiene que ser superior a 200 ng/uL, de lo contrario la muestra se debe diluir) y se incubó
a 37ºC por 30 min en termociclador.
Luego se inactivó la DNAasa, colocando 2 µL de inactivador de DNAasa por tubo, se
resuspendió por pipeteo y se incubó por 5 min a temperatura ambiente. Posteriormente
se centrifugó a 10000 x g por 1,5 min y se transfirió el sobrenadante a un nuevo tubo.
Finalmente el RNA fue almacenado a -80ºC.
3.2.16 Cuantificación del RNA.
Se preparó una dilución de 1:100 de RNA en agua y se midió su absorbancia a 260 nm
en cubeta de cuarzo contra un blanco de agua. Se determinó la razón Abs 260/280
como parámetro de pureza y se calculó la concentración tomando como referencia que
una solución de RNA de 40 µg/mL presenta una Abs260 = 1, a partir de la siguiente
fórmula:
[RNA] (g/mL) = Abs260 * factor de dilución * 40

30
3.2.17 Retrotranscripción RT-PCR.
La retrotranscripción fue realizada con la enzima SuperScriptTM II Reverse
Transcriptasa, colocando por tubo de reacción: 1 µL de Oligo dt, ramdom primer y
dNTPs, 5 µg/µL de RNA, H2O a un volumen final de 12 µL. Las muestras fueron
calentadas a 65ºC por 5 min y se colocaron de inmediato en hielo. Se agregó a cada
tubo de reacción: 4 µL de buffer 5X, 2 µL de DTT 0,1 M y 1 µL de RNAout. Se incubó a
42 ºC por 2 min y se agregó 1 µL de Super Script II por tubo de reacción. Se incubó 45
minutos a 45ºC y se colocó 0,5 µL más de enzima.
Para evaluar el éxito del RT-PCR se realizó un PCR convencional con una TM de 55ºC,
usando partidores para ciclofilina, un gen constitutivo. Los partidores son los siguientes:
F= EX4 GGC AAT GCT GGA CCA AAC ACA A
R: EX5 GTA AAA TGC CCG CAA GTC AAA AG
3.2.18 PCR en tiempo real.
La expresión relativa de cada gen fue determinada según la metodología del 2-ΔΔCT
descrito en Liwak y Schmittgen, 2001. En esta metodología se evalúa la expresión del
gen de interés, respecto a un gen de referencia, el cual debe ser constitutivo y ser
validado previamente para cada experimento, asegurando de esta manera que su
expresión es invariable con el tratamiento experimental aplicado. Las eficiencias de
amplificación del gen de interés y del gen de referencia deben ser similares. Una forma

31
de evaluar esto, es mediante curvas de amplificación con diluciones del templado. Si no
se obtiene la misma eficiencia se debe estandarizar nuevamente el experimento,
diseñando nuevos partidores por ejemplo.
Los cambios de expresión son evaluados por los ΔCT. Donde CT indica el número de
ciclo fraccional al cual la cantidad del blanco amplificado alcanza un umbral fijo, el cual
se obtiene de la intersección entre la curva de amplificación y la línea umbral, ésta es
sugerida por el programa utilizado. El ΔCT es la diferencia entre los ciclos umbrales del
blanco y el referente, es una medida relativa de la concentración del gen analizado.
La metodología es del ΔΔCT porque se evalúa los deltas de cada muestra
individualmente y se normaliza también por los deltas promedio del total de muestras
silvestres analizada como se describe en la siguiente ecuación:
El número que se obtiene de la ecuación se utiliza para obtener la cantidad de blanco
normalizado al referente endógeno, en este caso ciclofilina, de acuerdo a:
Cantidad de blanco = 2-ΔΔCT
Las PCR cuantitativas fueron realizadas con el master mix 2X universal KAPA SYBR
FASTA qPCR Kit, colocando 9 µL de master mix y 1 µL de cDNA en la dilución
apropiada, más 0,5 µL de cada partidor. Se realizaron curvas de calibración para cada
par de partidores utilizando 3 muestras de cDNA de corteza renal de ratones silvestres
ΔΔCT= (CTblanco – CTciclofilina)muestra X - (CTblanco– CTciclofilina)total de muestras silvestres

32
y 3 muestras de cDNA de ratones nulos para TASK-2. Las curvas de calibración
abarcaron las siguientes diluciones, realizadas con el pool de muestras tanto silvestres
como nulas para TASK-2: 1:1, 1:5, 1:10, 1:50 y 1:100. De acuerdo a esta curva de
calibración se evaluó la eficiencia de la PCR en tiempo real y se eligió la dilución
apropiada para cada muestra analizada con su respectivo partidor.
Los resultados fueron evaluados mediante la prueba estadística de Mann-Whitney.
(Livak and Schmittgen 2001)
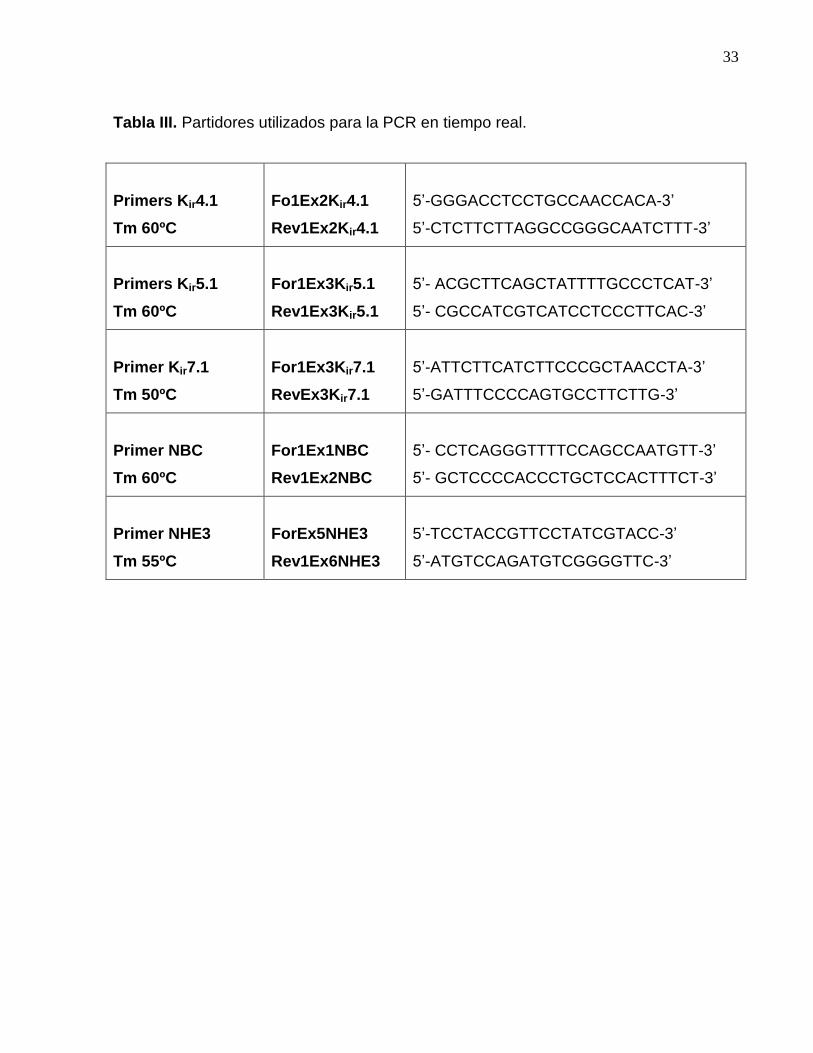
3.2.19 Partidores utilizados para la PCR en tiempo real.
Para realizar la PCR en tiempo real, primero a los partidores seleccionados se les
realizó una gradiente de temperatura, para elegir la Tm más adecuada. Los partidores
usados y sus Tm son presentados en la tabla III.
33
Tabla III. Partidores utilizados para la PCR en tiempo real.
Primers Kir4.1
Tm 60ºC
Fo1Ex2Kir4.1
Rev1Ex2Kir4.1
5’-GGGACCTCCTGCCAACCACA-3’
5’-CTCTTCTTAGGCCGGGCAATCTTT-3’
Primers Kir5.1
Tm 60ºC
For1Ex3Kir5.1
Rev1Ex3Kir5.1
5’- ACGCTTCAGCTATTTTGCCCTCAT-3’
5’- CGCCATCGTCATCCTCCCTTCAC-3’
Primer Kir7.1
Tm 50ºC
For1Ex3Kir7.1
RevEx3Kir7.1
5’-ATTCTTCATCTTCCCGCTAACCTA-3’
5’-GATTTCCCCAGTGCCTTCTTG-3’
Primer NBC
Tm 60ºC
For1Ex1NBC
Rev1Ex2NBC
5’- CCTCAGGGTTTTCCAGCCAATGTT-3’
5’- GCTCCCCACCCTGCTCCACTTTCT-3’
Primer NHE3
Tm 55ºC
ForEx5NHE3
Rev1Ex6NHE3
5’-TCCTACCGTTCCTATCGTACC-3’
5’-ATGTCCAGATGTCGGGGTTC-3’

34
4. Resultados
4.1 Caracterización del mensajero mutante en el ratón nulo para TASK-2.
La secuencia codificante para TASK-2 (KCNK5), cuenta con 5 exones y entre el exón 1
y 2 se espera esté incorporada la secuencia codificante para el gen reportero β-
galactosidasa (β-gal). La caracterización del mensajero mutante para TASK-2 se realizó
mediante RT-PCR, usando partidores dirigidos a reconocer el exón 1 y el exón 2 de la
secuencia codificante para el canal. En la Figura 3A se observa que el PCR con el
cDNA del ratón silvestre presentó una banda de 187 pb, la cual concuerda con el
amplicón esperado y que está ausente en el PCR para el cDNA de los animales nulos.
Lo anterior permite concluir que se ha interrumpido la transcripción del mensajero que
codifica para TASK-2. Adicionalmente se utilizaron partidores dirigidos a reconocer la
existencia del mensajero mutante formado por el exón 1 y la secuencia codificante para
β-gal, en la Figura 3B se observa la presencia del amplicón del tamaño esperado, de
1097 pb sólo en el PCR para el cDNA de los ratones nulos para el canal. Finalmente
para corroborar la información obtenida además se utilizaron los partidores dirigidos al
exón 1 y el exón 4, donde solo se presentó un amplicón de 563 pb en el animal silvestre
(Figura 3C).
Con los resultados expuestos se puede señalar que efectivamente fue insertada la
secuencia codificante para β-gal entre el exón 1 y 2, ya que se demuestra la presencia
de un mensajero híbrido entre el exón 1 y el reportero β-galactosidasa, y que su
inserción fue capaz de truncar la expresión de TASK-2. Lo anterior permite utilizar la

35
expresión de la β-gal como reportero de la presencia del canal en los diferentes tejidos
en condiciones fisiológicas.
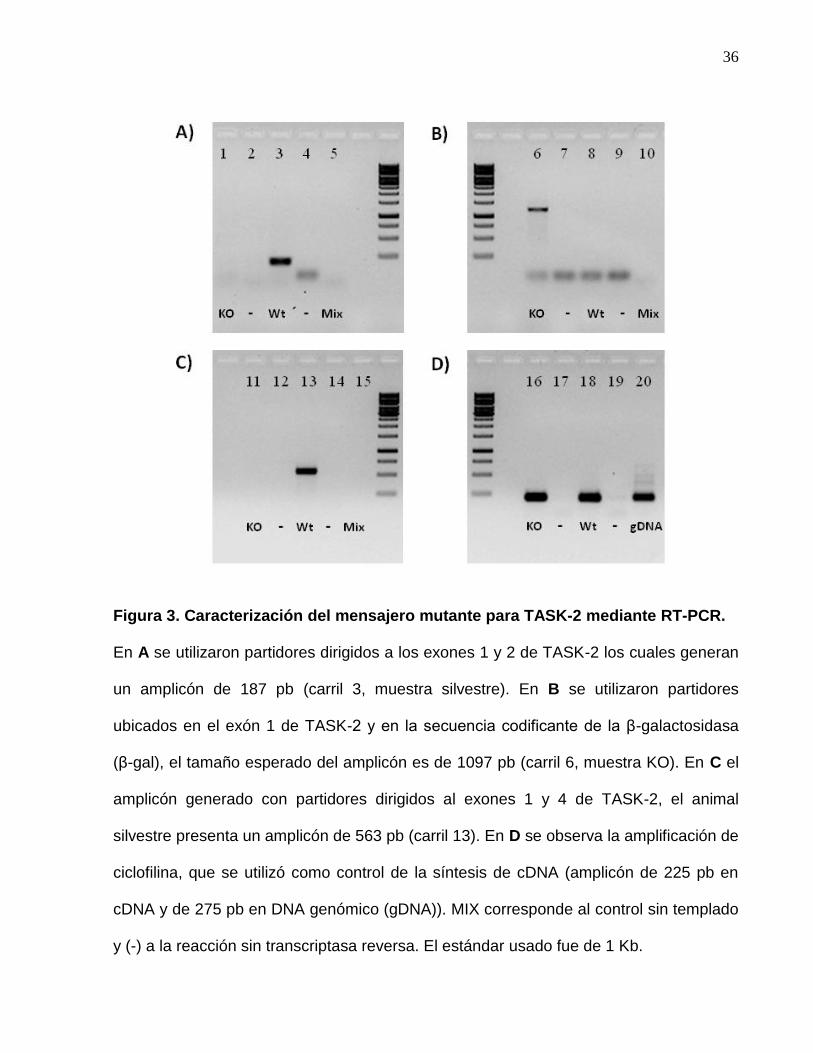
36
Figura 3. Caracterización del mensajero mutante para TASK-2 mediante RT-PCR.
En A se utilizaron partidores dirigidos a los exones 1 y 2 de TASK-2 los cuales generan
un amplicón de 187 pb (carril 3, muestra silvestre). En B se utilizaron partidores
ubicados en el exón 1 de TASK-2 y en la secuencia codificante de la β-galactosidasa
(β-gal), el tamaño esperado del amplicón es de 1097 pb (carril 6, muestra KO). En C el
amplicón generado con partidores dirigidos al exones 1 y 4 de TASK-2, el animal
silvestre presenta un amplicón de 563 pb (carril 13). En D se observa la amplificación de
ciclofilina, que se utilizó como control de la síntesis de cDNA (amplicón de 225 pb en
cDNA y de 275 pb en DNA genómico (gDNA)). MIX corresponde al control sin templado
y (-) a la reacción sin transcriptasa reversa. El estándar usado fue de 1 Kb.

37
4.2 Expresión de TASK-2 durante el desarrollo embrionario y en el postnatal.
Para evaluar la expresión de TASK-2 en el desarrollo embrionario se analizó la
actividad de β-galactosidasa en embriones completos de edades entre 9,5 a 14,5 dpc
(días post coito). Se analizaron también cortes histológicos de embriones de 10,5; 12,5
y 15,5 dpc, así como también ratones postnatales de 3 días.
Se observó que el canal se expresa desde el primer estadio embrionario analizado, a
los 9,5 dpc, donde la señal se restringe al eje dorsal del embrión y a la región cefálica,
zonas que corresponde al sistema nervioso en formación. La señal observada en el
neuroepitelio, en esta etapa del desarrollo es muy tenue, como se observa en la Figura
4.

38
Figura 4. Expresión de TASK-2 a los 9,5 dpc del desarrollo embrionario. Se
muestra la expresión de β-gal en cortes histológicos de 12 micras de espesor,
realizados de forma sagital y en embriones completos en el estadio de 9,5 dpc, tanto
silvestre como nulo para TASK-2, hermanos de camada. La expresión se observa en el
eje dorsal del embrión (imágenes de la izquierda) y en la región cefálica (imágenes de
la derecha). Barra de 10 µm.

39
Para tener una idea general de los sitios de expresión del canal en el desarrollo, se
analizaron embriones de 10,5; 12,5 y 14,5 dpc en tejido completo, como se muestra en
la Figura 5A. Se puede apreciar que a medida que se avanza en los estadios
embrionarios, la expresión se hace más evidente en la región cefálica (estadio 10,5 y
12,5 dpc), específicamente en el telencéfalo y en menor grado en el romboencéfalo. De
igual manera existe expresión en el eje dorsal de los embriones. En la Figura 5B, se
muestra con mayor claridad la expresión en el telencéfalo y se evidencia que la
expresión en el eje dorsal corresponde a las vertebras del tejido cartilaginoso de la
columna vertebral en formación. Luego a los 14,5 dpc el canal también comienza a
manifestarse en el tejido cartilaginoso de las extremidades y se mantienen los sitios de
expresión vistos en los estadios anteriores.
Cabe destacar que a los 12,5 dpc también se manifiesta expresión en la zona
abdominal inferior del embrión, correspondiendo posiblemente está expresión al riñón
en formación.
.

40
A)
B)

41
Figura 5. Expresión de TASK-2 en el desarrollo embrionario. En la Figura A se
muestra la expresión de TASK-2 en los estadio embrionarios 10,5 dpc; 12,5 dpc y 14,5
dpc, donde la expresión se restringe al eje dorsal y a la región cefálica de los embriones
y a los 14,5 dpc además aparece una fuerte expresión en las extremidades. En B se
muestra la vista superior de embriones de 12,5 dpc, donde se observa que la expresión
cefálica corresponde al telencéfalo y que la expresión en el eje dorsal corresponde a las
vertebras del tejido cartilaginoso de la columna vertebral en formación. Como control se
utilizaron animales silvestres hermanos de camada. Fotografías tomadas con lupa.

42
Teniendo en consideración los hallazgos de la expresión de TASK-2 en tejido completo,
se procedió a realizar cortes histológicos en las zonas que evidencian la presencia del
canal. En la Figura 6 se puede apreciar que la señal que se detecta a los 12,5 dpc en la
región cefálica, en el telencéfalo, corresponde a expresión en la zona del palium, que
dará origen a la corteza cerebral y a neuronas de tipo glutamatérgicas. Se destaca que
la expresión es más intensa en la zona de la preplaca, pero de igual manera existe
expresión en la zona ventricular. A los 15,5 dpc se acentúa la expresión en el tejido que
recubre el encéfalo, correspondiendo éste posiblemente a las meninges.
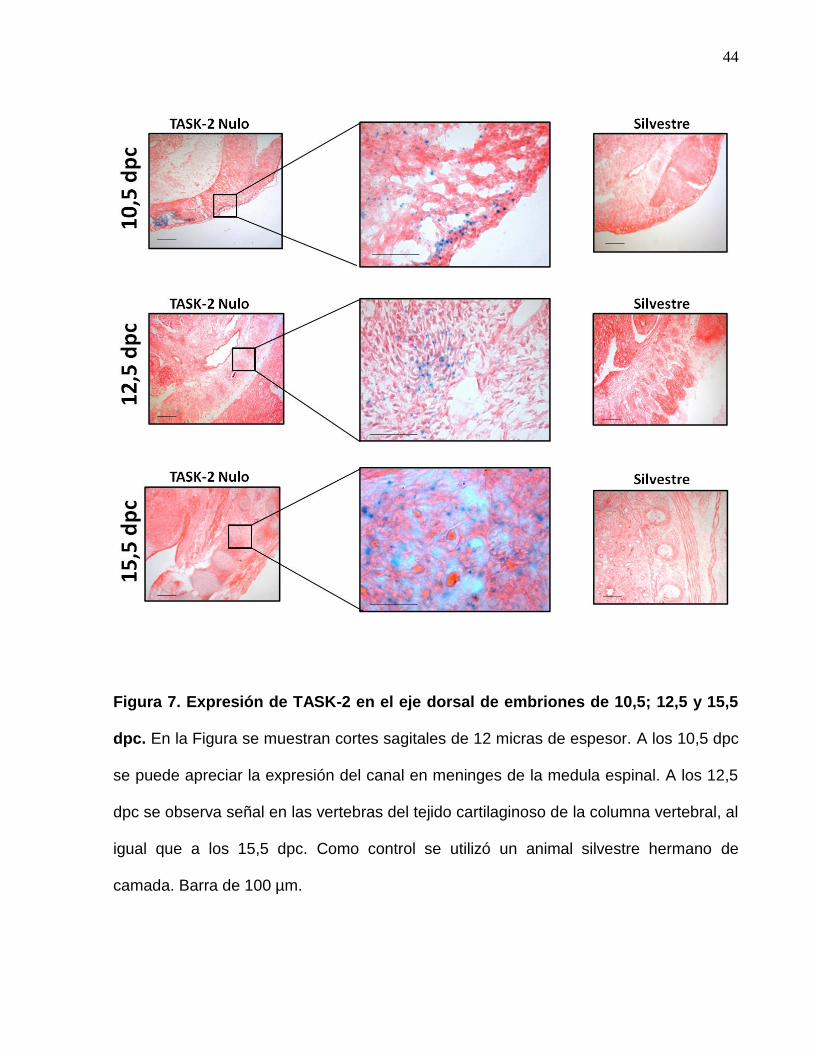
También mediante cortes histológicos se observó la expresión del canal en el eje dorsal
de los embriones analizados (Figura 7). A los 10,5 dpc existe una intensa expresión en
el tejido que rodea la medula espinal en formación, correspondiendo este también
posiblemente a meninges. Luego a los 12,5 dpc se puede apreciar con claridad la
columna vertebral en formación, donde destaca la expresión de TASK-2 en el tejido
cartilaginoso de las vertebras. Este patrón de expresión se mantiene a los 15,5 dpc,
correspondiendo este tipo celular posiblemente a condrocitos, como se discutirá más
adelante.
En postnatales, se hace más evidente que la expresión en la zona cefálica del embrión
corresponde a las meninges (Figura 8A) y a su vez se acentúa la expresión en los
condrocitos de la columna vertebral (Figura 8B).

43
Figura 6. Expresión de TASK-2 en la región cefálica de embriones de 12,5 y 15,5
dpc. En la Figura se muestran cortes sagitales de 12 micras de espesor. A los 12,5 dpc
se puede observar que la expresión del canal se restringe al palium, no presentando
señal el subpalium. La señal es más intensa en la zona de las meninges. A los 15,5 dpc
continua presente la expresión en meninges. Como control se utilizó un animal silvestre
hermano de camada. (C) Caudal, (R) Rostral, (VL) Ventrículo Lateral. Barra de 100 µm.
44
Figura 7. Expresión de TASK-2 en el eje dorsal de embriones de 10,5; 12,5 y 15,5
dpc. En la Figura se muestran cortes sagitales de 12 micras de espesor. A los 10,5 dpc
se puede apreciar la expresión del canal en meninges de la medula espinal. A los 12,5
dpc se observa señal en las vertebras del tejido cartilaginoso de la columna vertebral, al
igual que a los 15,5 dpc. Como control se utilizó un animal silvestre hermano de
camada. Barra de 100 µm.
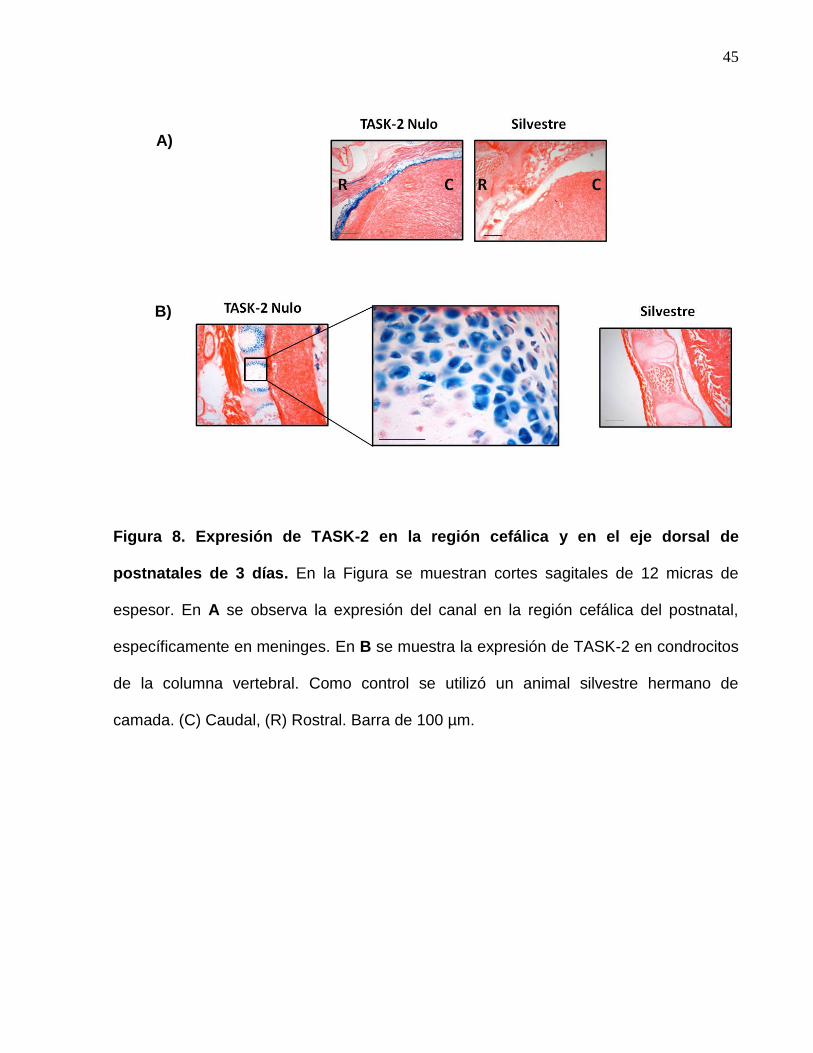
45
Figura 8. Expresión de TASK-2 en la región cefálica y en el eje dorsal de
postnatales de 3 días. En la Figura se muestran cortes sagitales de 12 micras de
espesor. En A se observa la expresión del canal en la región cefálica del postnatal,
específicamente en meninges. En B se muestra la expresión de TASK-2 en condrocitos
de la columna vertebral. Como control se utilizó un animal silvestre hermano de
camada. (C) Caudal, (R) Rostral. Barra de 100 µm.
A)
B)

46
4.3 Expresión de TASK-2 durante el desarrollo renal.
Se evaluó la expresión indirecta de TASK-2 en distintos estadios del desarrollo renal,
encontrándose que el canal se expresa desde tempranas etapas del desarrollo,
evidenciándose en los cortes histológicos expresión desde los 12,5 dpc (Figura 9A).
Esta etapa del desarrollo embrionario coincide con la formación del metanefros, que
finalmente dará origen al riñón de mamíferos definitivo. La expresión se acentúa y se
hace mucho más intensa a medida que se avanza en los estadios embrionarios,
llegando a ser específica a los túbulos renales en formación a los 15,5 dpc,
excluyéndose la presencia del canal en glomérulos (Figura 9B). Estos resultados se
correlacionan con lo visto en postnatales de 3 días (Figura 9C).
Hasta este punto es imposible dilucidar si la expresión del canal es restringida a ciertas
zonas renales, sea medular o cortical, puesto que el riñón aún no presenta su
distribución morfológica definida.
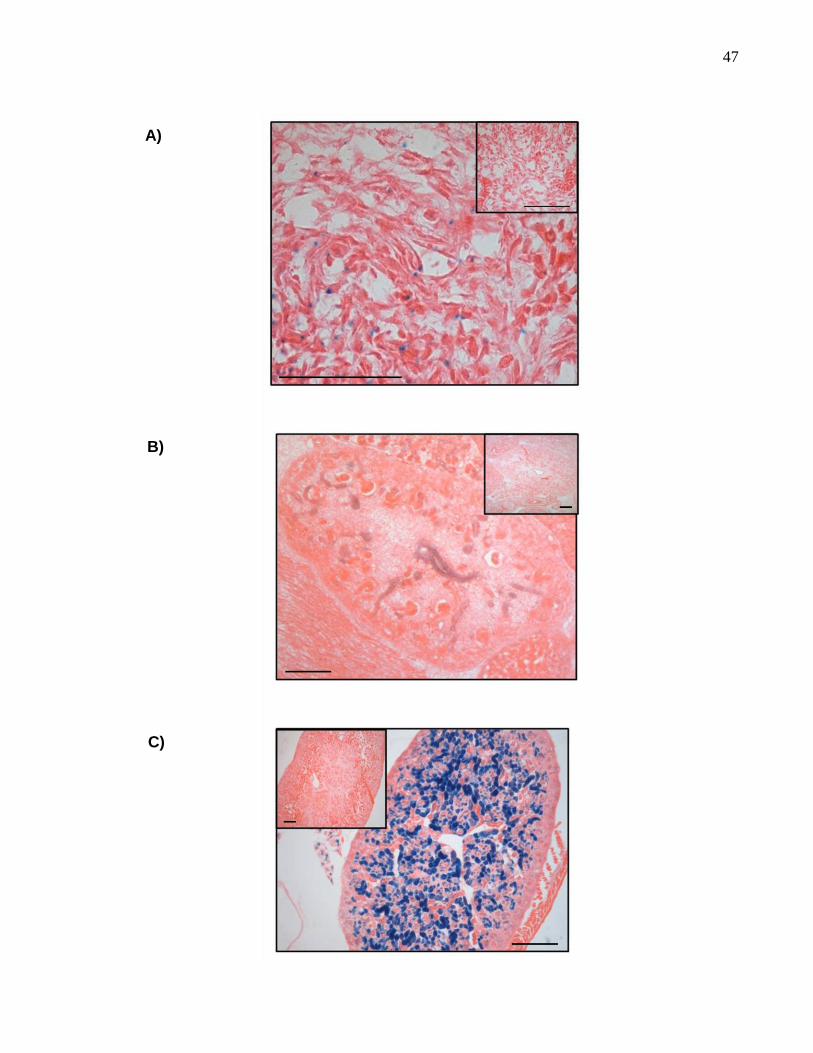
47
A)
B)
C)

48
Figura 9. Expresión de TASK-2 en tejido renal de embriones de 12,5 dpc, 15,5 dpc
y postnatales de 3 días. En A se observa la expresión de TASK-2 mediante el
revelado de β-gal en el tejido renal a los 12,5 dpc del desarrollo embrionario. Como
control en la esquina superior derecha se observa el mismo tejido revelado para un
animal silvestre hermano de camada. En B se muestra la expresión de TASK-2 a los
15,5 dpc. Se observa que la señal se hace específica a los túbulos renales en
formación. En la esquina superior derecha se observa el control. En C se observa la
expresión del canal en postnatales de 3 días; en este estadio del desarrollo la expresión
se acentúa. En la esquina superior izquierda se observa el control. Barra de 100 µm.

49
4.4 Expresión de TASK-2 en los túbulos proximales renales.
Para evaluar la expresión renal de TASK-2 en animales adultos, se procedió a revelar la
actividad de β-gal en un riñón de un animal heterocigoto y de un animal silvestre. En la
Figura 10 se puede observar que la expresión del canal se restringe a ciertos túbulos
renales, preferentemente en la corteza renal. Mediante esta técnica es imposible
dilucidar la localización subcelular del canal (apical o basolateral), ya que la actividad de
la β-gal es citoplasmática, impidiendo de esta manera determinar la localización
específica de TASK-2. Por otra parte se observó una intensa señal en la zona
correspondiente a la pelvis renal, que no pudo ser corroborada mediante la localización
directa de la proteína del canal.

50
Figura 10. Expresión de TASK-2 en riñones de animales adultos heterocigotos y
silvestre. En A se observa la expresión de β-gal en un riñón heterocigoto de un animal
adulto, la expresión es restringida a ciertos túbulos de la corteza renal (fotografía
tomada con lupa). En las imágenes ampliadas de B se muestra el control silvestre y en
C el heterocigoto. Barra 10 µm.
A)
B)
C)

51
La ubicación subcelular específica de TASK-2, se determinó mediante la localización de
la proteína del canal por inmunohistoquímica e inmunofluorescencia en riñones de
animales silvestres utilizándose como controles los hermanos de camada nulos. En la
Figura 11A se muestra la expresión del canal en túbulos ubicados en la corteza renal,
en 11B se observa que la señal se ubica en la membrana basolateral de túbulos
posiblemente proximales y que no existe señal en los glomérulos y otro tipo de túbulos.
La ausencia de señal en riñón del ratón nulo (11A izquierda y 11C) denota la
especificidad de la señal obtenida.
Para corroborar que los túbulos renales que expresan TASK-2 son proximales se
realizó una inmunohistoquímica antiTASK-2 con tinción de PAS (Figura 12). La tinción
de PAS permite la tinción de componentes celulares que contienen hidratos de carbono,
por ejemplo algunas membranas celulares, células caliciformes en la mucosa del
intestino o fibras reticulares que están rodeadas por hidratos de carbono; en esta
técnica, el ácido peryódico oxida a los grupos hidroxilos de dos carbonos cercanos,
formando de esta manera grupos aldehídos, así el ácido peryódico de Schiff puede
reaccionar con estos y dejar una tinción rojiza. En el riñón con esta tinción es posible
detectar el ribete en cepillo de la membrana apical de los túbulos proximales,
facilitándose con ello su identificación. Se pudo corroborar que la expresión basolateral
de TASK-2 es exclusiva de los túbulos proximales.

52
A) Silvestre TASK-2 Nulo

53
C)
B)

54
Figura 11. Expresión de TASK-2 en la membrana basolateral de túbulos renales
corticales mediante inmunohistoquímica. En A se muestra una inmunohistoquímica
de riñones completos, el de la izquierda es silvestre donde se puede ver que la
expresión del canal es exclusiva de la corteza renal y de la capa medular más externa y
la imagen de la derecha muestra el control nulo para TASK-2 que no evidencia de señal
(fotografías tomadas con lupa). En B se observa una imagen obtenida de la zona
cortical y en C el control para la inmunohistoquímica que corresponde a un animal
hermano de camada nulo para el canal donde no se evidencia señal. Demostrandose
con ello la especificidad de la señal detectada. Barra 10 micrometros.

55
Figura 12. Inmunohistoquímica antiTASK-2 con tinción de PAS. Expresión
basolateral de TASK-2 en tubulos proximales, en un animal silvestre, de 3 meses de
edad. Barra 10 micrometros.

56
4.5 TASK-2 se expresa con NBCe1-A y NHE3 en las células del túbulo proximal
donde estas proteínas participan en la reabsorción del bicarbonato.
Por los experimentos anteriores se pudo demostrar que la expresión de TASK-2 es
exclusiva de los túbulos proximales. Con este antecedente se puede esperar que el
canal se exprese en el mismo tipo celular que las otras proteínas que participan en la
reabsorción de bicarbonato, como por ejemplo NBCe1-A y NHE3. Para dilucidar este
punto se realizó una inmunohistoquímica doble dirigida a localizar TASK-2 con NHE3
que se expresa en la membrana apical de los túbulos proximales y una
inmunohistoquímica dirigidas a localizar TASK-2 con NBCe1-A, el cual al igual que
TASK-2 se expresa en la membrana basolateral de túbulos proximales. Por ello en este
último caso se tuvieron que realizar las localizaciones por separado en cortes seriados.
Como se muestra en la Figura 13, todos los túbulos que expresan TASK-2 también
expresan NHE3, no obstante la localización de NHE3 no es exclusiva de los túbulos
proximales, lo cual coincide con los antecedentes bibliográficos existentes donde se ha
descrito que NHE3 también se expresa en la parte apical de la rama ascendente gruesa
del asa de Henle (Bobulescu y Moe 2009).

57
Figura 13. Doble inmunohistoquímica de TASK-2 y NHE3. Expresión de TASK-2 y
NHE3 en túbulos proximales, se puede observar que todos los túbulos que expresan
TASK-2 también expresan NHE3, sin embargo la expresión de NHE3 no es exclusiva a
los túbulos proximales. Barra de 10 µm.

58
Mediante la inmunolocalización en cortes seriados se pudo detectar que TASK-2 y
NBCe1-A se coexpresan en túbulos proximales (Figura 14A y 14B). Además se logró
realizar la inmunolocalización de estas proteínas mediante inmunoflorescencia, donde
se pudo apreciar de mejor manera la localización en la membrana basolateral de estas
proteínas. Empero al ser estos anticuerpos hechos en la misma especie de animal
(conejo) no se pudieron utilizar para realizar una doble inmunolocalización en un mismo
corte de tejido.
Por antecedentes bibliográficos se sabe que el contransportador NBCe1-A se expresa
exclusivamente en la membrana basolateral de los túbulos proximales (Amlal, Chen, y
Greeley 2001), lo cual se corroboró en esta tesis mediante una inmunolocalización de
NBCe1-A con tinción de PAS (Figura 14C).

59

60
Figura 14. NBCe1-A y TASK-2 se coexpresan en la membrana basolateral de
túbulos proximales. En A y B se muestran las inmunolocalizaciones de TASK-2 y
NBCe1-A respectivamente, mediante inmunoshistoquímica en cortes seriados y en los
paneles inferiores se muestran inmunofluorescencias para estas proteínas, donde se
evidencia la clara localización subcelular de estas en la membrana basolateral de las
células de los túbulos proximales. En C se observa una imnunohistoquímica antiNBCe1-
A con tinción de PAS, donde se puede apreciar que la localización de este
contransportador es exclusiva a los túbulos proximales. Barra de 10 µm.

61
4.6 Un posible rol compensatorio ante la ausencia de TASK-2
Cómo se señaló previamente, la ausencia de TASK-2 que provoca una acidosis
metabólica, puede desencadenar una actividad compensatoria capaz de permitir una
reabsorción parcial de bicarbonato y de esta manera evitar una acidosis metabólica
acentuada. Una forma de acercarse a este problema es la evaluación de cambios en la
transcripción del RNA mensajero de posibles candidatos.
Por RT-PCR cuantitativa se evaluaron genes que se postuló podrían compensar la
ausencia de TASK-2, estos son los canales Kir4.1, 5.1, 7.1, el intercambiador NHE3 y el
cotransportador NBCe1-A.
El canal Kir7.1 ha sido inmunolocalizado en la membrana basolateral de los túbulos
distales en ratas (Ookata y Tojo 2000) y en cobayos ha sido detectado en túbulos
proximales (Derst y Hirsch 2001). Para saber la localización renal de este canal en
ratones, se realizó una inmunohistoquímica con tinción de PAS, donde pudimos
detectar la expresión de Kir7.1 en túbulos proximales (Figura 15). Como se señaló en la
introducción este canal es inhibido por acidificación extracelular y modulado por pH
intracelular de manera bifásica. (OOKATA y TOJO 2000)

62
Figura 15. Inmunohistoquímica de Kir7.1 en riñón. Se puede apreciar la
inmunolocalización de Kir7.1 en túbulos proximales mediante tinción de PAS. Barra 10
µm.

63
Por otra parte se ha sugerido que Kir5.1 también se expresa en la membrana
basolateral de los túbulos proximales (Derst et al. 2001), sin embargo la localización de
la proteína hasta ahora no ha sido determinada. Como Kir5.1 y 4.1 forman un canal
heteromérico que es inhibido por pH intracelular, este último también es un buen
candidato a evaluar.
Como se discutió previamente tanto el NBCe1-A como NHE3 tienen una participación
crítica en el proceso de reabsorción de bicarbonato proximal, también podrían variar su
expresión génica en forma compensatoria ante la ausencia de TASK-2.
En esta tesis se realizaron experimentos preliminares de RT-PCR cuantitativas para
todos los genes mencionados. Se utilizaron 3 animales silvestres y 3 animales nulos
para TASK-2, provenientes de diferentes camadas, a los cuales se extrajo RNA sólo de
corteza renal. Al aplicar la prueba estadística de Mann-Whitney. no se encontraron
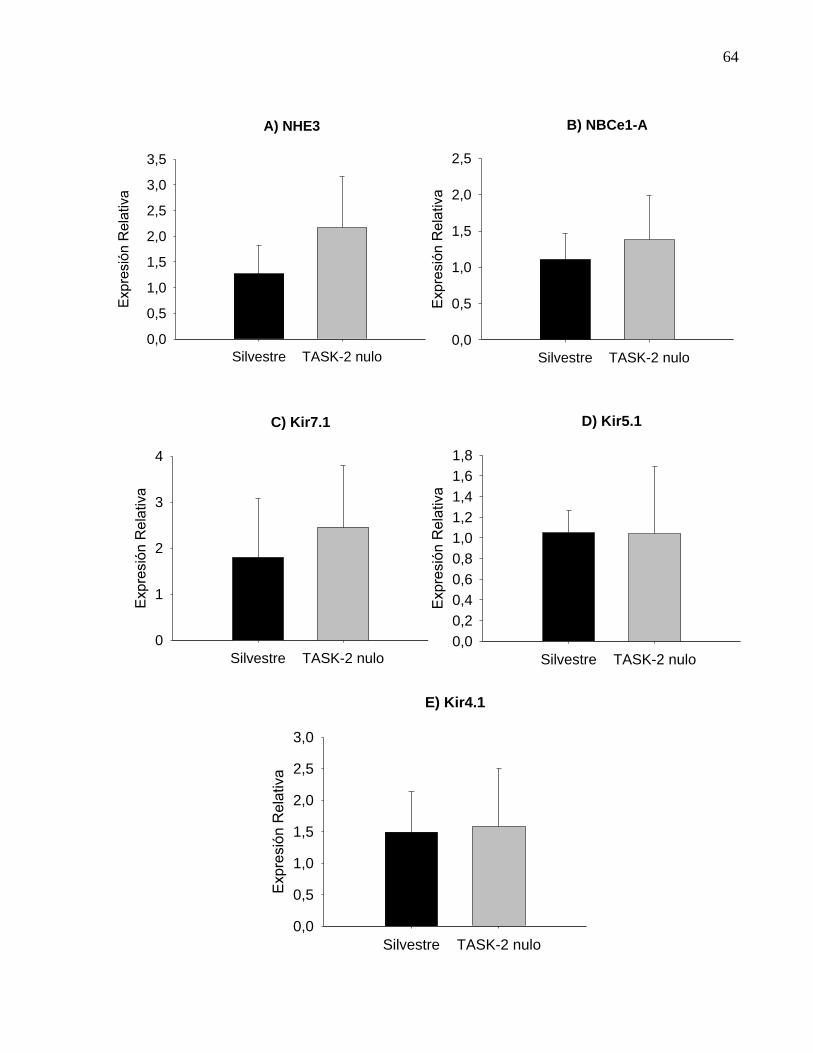
diferencias significativas para ninguno de los genes analizados, como se ve en la Figura
16. Aunque existe gran dispersión de los promedios, tanto en NHE3 (Figura 16 A) como
en el Kir7.1 (Figura 16 C) se vislumbra una tendencia al aumento de la expresión de
ellos en el ratón nulo. Para obtener resultados definitivos y concluyentes, es necesario
repetir el experimento reduciendo al mínimo la variabilidad entre los animales
analizados, por ejemplo estos deben provenir necesariamente de la misma camada, y
por otra parte es necesario aumentar el número de animales de experimentación.
64
A) NHE3
Silvestre TASK-2 nulo
Expre
sió
n R
ela
tiva
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
B) NBCe1-A
Silvestre TASK-2 nulo
Expre
sió
n R
ela
tiva
0,0
0,5
1,0
1,5
2,0
2,5
C) Kir7.1
Silvestre TASK-2 nulo
Exp
resió
n R
ela
tiva
0
1
2
3
4
D) Kir5.1
Silvestre TASK-2 nulo
Expre
sió
n R
ela
tiva
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
E) Kir4.1
Silvestre TASK-2 nulo
Exp
resió
n R
ela
tiva
0,0
0,5
1,0
1,5
2,0
2,5
3,0

65
Figura 16. Expresión relativa de genes candidatos a compensar la ausencia de
TASK-2. La expresión relativa de cada gen fue determinada según la metodología del
ΔΔCT y los datos se presentan como promedios ± error estándar de las medias. En A
se muestra los cambios de expresión de NHE3, en el ratón nulo para TASK-2 respecto
de la expresión que presenta un animal silvestre. En B se presentan la gráfica para
NBCe1-A y en C, D y E de los canales Kir7.1, 5.1 y 4.1 respectivamente.

66
5. DISCUSIÓN
Se ha descrito que los canales de potasio participan en diversas funciones en distintos
tipos celulares, tanto en tejidos excitables como no excitables y juegan un rol crítico en
procesos de señalización celular, regulación de la liberación de neurotransmisores y
excitabilidad neuronal, secreción de insulina, ritmo cardiaco, transporte epitelial de
electrolitos, contracción muscular, entre otros (Shieh et al. 2000).
En esta tesis se trabajó con un modelo murino nulo para el canal de potasio TASK-2, y
a través de él se determino la actividad de la β-galactosidasa bajo la dirección del
promotor endógeno de TASK-2, para estudiar los sitios de expresión del canal. El ratón
nulo fue producido mediante el método de atrapamiento génico, en el cual se trunca la
expresión de TASK-2, debido a la inserción de la secuencia codificante para el gen
reportero β-galactosidasa entre el exón 1 y 2, produciéndose un mensajero hibrido entre
el exón 1 y el reportero (Figura 3).
5.1 TASK-2 en el desarrollo embrionario.
En esta tesis se evaluó la expresión del canal de potasio TASK-2 en distintas etapas del
desarrollo embrionario, encontrándose expresión desde el primer estadio analizado a
los 9,5 dpc (Figura 4), donde la señal se restringe a la región cefálica y al eje dorsal del
embrión. No queda claro en este estadio embrionario el tipo celular especifico que
presenta expresión en el neuroepitelio.
A los 10,5 dpc la expresión del canal se intensifica en la región cefálica del embrión
(Figura 5A), específicamente en el telencéfalo como se puede corroborar a los 12,5 dpc

67
en la expresión del canal en tejido completo (Figura 5B). Además se evidenció
expresión en el romboencéfalo, que dará origen entre otras estructuras al tallo
encefálico, donde en ratones adultos se ha reportado la expresión de TASK-2 en el
núcleo retrotrapezoide (Gestreau et al. 2010).
Como se ve en cortes histológicos a los 12,5 dpc (Figura 6) la expresión es específica a
la zona correspondiente al palium, que dará origen a la corteza cerebral y a neuronas
de tipo glutamatérgicas. La expresión es más intensa en la región de la preplaca, en
tejido que posiblemente corresponde a meninges. Esta señal coincide con los
antecedentes bibliográficos que señalan que la expresión de la primera capa de células
meningeales es identificable desde los 10 dpc, esta capa posteriormente dará origen a
la leptomeniges, compuesta por la aracnoide y la piamadre y esta es inidentificable
desde los 13 dpc (Siegenthaler J y Pleasure S. 2011). Por otra parte hay evidencias
reportadas de la expresión de TASK-1 en células meningiales, donde se postula que el
canal podría ser importante en la contribución de procesos secretorios y en la
regulación del volumen (Kanjhan et al. 2010). Cabe destacar que en este trabajo
también se observó, a los 10,5 dpc, la expresión del canal en meninges de la medula
espinal fetal (Figura 7).
Un estudio que evaluó la expresión de algunos miembros de la familia de canales K2P
en el desarrollo del sistema nervioso central en ratones mediante hibridación in situ,
demostró que los genes analizados manifiestan diferentes patrones de expresión tanto
espacial como temporal y postulan que los genes TREK-1, TREK-2 y TASK-2, podrían
tener un posible rol en el control de la división celular (Aller y Wisden 2008). En este

68
estudio, a diferencia de lo encontrado en esta tesis, solo se detecta la expresión de
TASK-2 en la zona ventricular del encéfalo de embriones desde los 12 a los 19 dpc, y
no evidencian la presencia del canal en animales adultos o postnatales, donde nosotros
sí encontramos expresión. Esta discrepancia puede ser debida a la diferencia de las
técnicas usadas. Por lo tanto podemos señalar que la expresión de TASK-2 no es solo
restringida al desarrollo embrionario del sistema nervioso central, sino que también el
canal sigue estando presente posterior al nacimiento. En animales adultos solo hay
evidencias publicadas de la presencia del canal en una población de neuronas
conocidas por ser esenciales en la quimiosensitividad respiratoria en el núcleo
retrotrapezoide ubicado en el tallo encefálico, donde el canal estaría involucrado en la
regulación de las presiones parciales de CO2 y O2 (Gestreau et al. 2010). Esta particular
localización en el sistema nervioso central fue corroborada en nuestro laboratorio
mediante la expresión de -gal.
La función de TASK-2 en estos nuevos tejidos donde hemos encontrado su expresión,
podría estar relacionada con una contribución del canal en procesos secretorios y/o en
el mantenimiento osmótico, al ser TASK-2 un canal activado por volumen. Por ejemplo
las células de las granulaciones aracnoideas participan en la reabsorción del líquido
cefalorraquídeo (Spassky et al. 2005).
Por otra parte desde los 12,5 dpc de desarrollo embrionario comienza a manifestarse
expresión de TASK-2 en tejido cartilaginoso de las extremidades y columna vertebral,
esta expresión se mantiene a lo largo de todo el desarrollo embrionario y también en
postnatales. Estos resultados concuerdan con los expuestos en una publicación

69
reciente donde se demostró la expresión de TASK-2 en condrocitos humanos, donde el
canal estaría jugando un rol crítico en la estabilización del potencial del membrana de
este tipo celular (Clark et al. 2011).
5.2 TASK-2 en la membrana basolateral de los túbulos proximales.
La expresión de TASK-2 comienza a manifestarse desde tempranos estadios del
desarrollo embrionario renal, observándose la presencia del canal desde los 12,5 dpc,
cuando el riñón se encuentra en la etapa de metanefros, que es la estructura que dará
origen al riñón definitivo. A medida que se avanza en los estadios embrionarios la
expresión se va haciendo más evidente, es así como a los 15,5 dpc podemos ver que el
canal se expresa exclusivamente en los túbulos renales en formación (Figura 9B), al
igual que en postnatales, excluyéndose expresión en glomérulos (Figura 9C).
En animales adultos se pudo corroborar la expresión de TASK-2 en los túbulos renales,
demostrándose que esta es exclusiva de la corteza renal (Figura 10). Específicamente
el canal se expresa en la membrana basolateral de los túbulos proximales (Figura 14).
Estos antecedentes reafirman la función propuesta para TASK-2, que previamente se le
había atribuido mediante experimentos electrofisiológicos en la reabsorción del
bicarbonato (Warth et al. 2004), pero hasta ahora no se había podido determinar la
localización de la proteína y por ello permanecía indefinida la localización subcelular
específica del canal en la células del túbulo proximal. En este trabajo se demostró la
colocalización de la expresión de TASK-2 con NBCe1-A y NHE3 en el epitelio de los
túbulos proximales, permitiéndonos validar el modelo de la Figura 17, planteado en la
introducción.

70
Figura 17. Modelo de reabsorción del bicarbonato en túbulos proximales. El
bicarbonato del filtrado presente en el lumen tubular se titula con los H+ excretados
desde el epitelio del túbulo proximal, esta reacción es catalizada por la anhidrasa
carbónica IV (ACIV), como producto de la reacción se generan CO2 y agua. El dióxido
de carbono difunde hacia el espacio intracelular donde junto con el agua son sustratos
de la AC II, formándose como producto H+ y bicarbonato. Los H+ activan al
intercambiador luminal NHE3, que transporta protones hacia el lumen contra su
gradiente junto con la entrada Na+ hacia el intracelular utilizando la gradiente generada
por la bomba de Na+/K+ ATPasa. El bicarbonato producido es transportado hacia el
intersticio mediante el cotrasportador basolateral NBCe1-A, De esta manera se produce
la depolarización de la membrana plasmática, por ello la activación de TASK-2 es
requerida para mantener la fuerza motriz para la reabsorción del bicarbonato.

71
5.3 Búsqueda de una actividad compensatoria a la ausencia de TASK-2.
La acidosis metabólica presentada en los ratones TASK-2 nulos es leve (Warth et al.
2004), lo cual no se condice con el rol atribuido al canal en la reabsorción de
bicarbonato, planteado en el modelo de la Figura 16. Dado que TASK-2 es requerido
para permitir la repolarización de la membrana celular y con ello dar la fuerza motriz
para el transporte sostenido de bicarbonato. Por ello es esperable que ante la ausencia
del canal se produzca un fenotipo severo de acidosis metabólica, a menos que otra
proteína asuma en parte la función de TASK-2.
Antecedentes que avalan esta hipótesis son por ejemplo el ratón nulo para NBCe1-A,
que presenta un fenotipo severo de acidosis metabólica con alta mortalidad, siendo la
sobrevida inferior a los 24 días. Este ratón no tiene una actividad compensatoria ante la
ausencia de NBCe1-A, (Gawenis et al. 2007). En cambio en el modelo murino nulo para
NHE3, se produce una compensación que sólo desencadena un fenotipo leve de
acidosis metabólica. La ausencia de este intercambiador provoca un aumento de la
expresión de aldosterona, renina y del intercambiador Cl-/HCO3- AE1, (Schultheis et al.
1998).
Al presentar los animales TASK-2 nulos una sobrevida y un desarrollo normal, es
factible proponer que alguna otra proteína este supliendo en parte la ausencia del
canal. Teniendo como antecedentes que TASK-2 es un canal activado por pH e
hipotonicidad y que es expresado en los túbulos proximales, se buscaron otros canales
y proteínas con características similares, como su sensibilidad a pH, como posibles
candidatos. Estos canales fueron: Kir 5.1 que forma un canal heterodimerico junto a

72
Kir4.1, el cual es inhibido por pH intracelular. También se postuló como candidato al
canal Kir7.1 que ha sido descrito en túbulos proximales de cobayo y humanos (Derst y
Hirsch 2001), este es inhibido por acidificación extracelular y modulado por pH
intracelular de manera bifásica (Hughes y Swaminathan 2008). En esta tesis se
demostró la expresión de Kir7.1 en túbulos proximales de ratón (Figura 15).
También se postuló que otra forma de suplir la ausencia de TASK-2, podría estar dada
por un aumento de la expresión de NHE3 o NBCe1-A, proteínas directamente
involucradas en la reabsorción del bicarbonato.
Los cambios de expresión génica fueron evaluados en experimentos piloto mediante
RT-PCR cuantitativa, en animales TASK-2 nulos, comparados con animales silvestres,
de 2 meses de edad. Se usó un número de 3 ratones tanto silvestres como nulos y
debido a la poca disponibilidad de animales, estos fueron de diferentes camadas.
No se encontraron diferencias estadísticamente significativas para ninguno de los
genes analizados. Pero estos resultados solo pueden considerarse como preliminares,
ya que es necesario repetir los análisis reduciendo al mínimo la variabilidad entre los
animales utilizados, por ello estos deben provenir de la misma camada, por otra parte
aumentar el número de animales analizados también es necesario para clarificar los
resultados obtenidos.
Es posible que al repetir estos experimentos se encuentren cambios en la expresión de
alguno de los genes que potencialmente den cuenta de la compensación buscada. Sin
embargo también es probable que exista compensación sin cambios en los niveles de
expresión génica y sea necesario evaluar cambios en la expresión y/o actividad de la

73
proteína. Cambios en los niveles de proteína sin cambios transcripcionales son
posibles, como es el caso reportado de ratas que presentan hipertensión espontánea,
donde ocurrió un aumento de NHE3 sin presentarse cambios en el nivel de su
mensajero, donde cambios post traduccionales podría explicar este fenómeno
(LaPointe et al. 2002).Una redistribución de la proteína desde un compartimento
intracelular a la membrana donde ejerza su acción también podría compensar una
función. Finalmente una última forma de compensación también puede estar dada por
cambios en la actividad sin cambios en los niveles o ubicación intracelular, sino
secundarios a modulación por moléculas u otras proteínas.
5.4 Conclusiones y proyecciones
Mediante la utilización de un modelo murino nulo para TASK-2 que expresa el gen
reportero β-galactosidasa, en esta tesis se logró determinar nuevos sitios de expresión
para TASK-2 en el desarrollo embrionario, encontrándose la expresión del canal desde
los 9,5 dpc en la región cefálica y en el eje dorsal de los embriones. A los 12,5 dpc se
encontró expresión en el telencéfalo, específicamente en el palium, destacando una
fuerte expresión en la preplaca, en tejido que corresponde posiblemente a meninges.
Además destaca la expresión en el romboencéfalo, que dará origen al tallo encefálico,
donde existe expresión de TASK-2 en el núcleo retrotrapezoide. En este mismo estadio
se presentó el primer antecedente de la expresión del canal en tejido cartilaginoso.
Estos sitios de expresión se mantienen durante todo el desarrollo embrionario, al igual
que en postnatales.

74
En el desarrollo renal, se determinó la expresión de TASK-2 desde los 12,5 dpc, estadio
embrionario que coincide con la etapa de formación del metanefros. A medida que se
avanza en los estadios embrionarios, se vio que la expresión del canal se restringe a
los túbulos renales en formación (15,5 dpc). Se logró determinar la localización
basolateral de TASK-2 en túbulos proximales, demostrando de esta manera que el
canal se expresa con NBCe1-A y NHE3. Esta localización lo sitúa en el lugar adecuado
para su participación en la reabsorción del bicarbonato.
Finalmente en experimentos preliminares no se encontró un aumento en la expresión
de genes candidatos que pudiesen compensar la ausencia de TASK-2 en animales
nulos para el canal.
Las proyecciones de este trabajo son por una parte determinar los tipos celulares
específicos donde se evidenció señal en el desarrollo embrionario cerebral. Se postuló
que la señal vista desde los 12,5 dpc en la zona que recubre el cerebro posiblemente
corresponde a meninges, para determinar si este tejido corresponde específicamente a
meninges se puede utilizar como marcador fibronectina, que ha sido reportado tiene
una expresión destacada en células meningiales (Lakatos 2003).
Por otra parte se debe seguir en la búsqueda de una actividad compensatoria ante la
ausencia de TASK2, ya sea a nivel de expresión génica o mediante la evaluación de
cambios en la expresión de proteína. Para validar los resultados obtenidos estos
experimentos deberán ser realizados necesariamente reduciendo al mínimo la
variabilidad entre los animales analizados.

75
6. REFERENCIAS
Aller M.I y Wisden W (2008). Changes in Expression of Some Two-pore Domain
Potassium Channel Genes (KCNK) in Selected Brain Regions of Developing Mice.
Neuroscience 151(4): 1154–72.
Amlal H, Chen Q y Greeley T (2001). Coordinated Down-regulation of NBC-1 and NHE-
3 in Sodium and Bicarbonate Loading. Kidney International 60: 1824–36.
Añazco C y Peña-Münzenmayer G (2013). G Protein Modulation of K2P Potassium
Channel TASK-2. European Journal of Physiology. Versión electronica previa a ser
impresa.
Araki, Masatake, Araki K y Yamamura K (2009). International Gene Trap Project:
Towards Gene-driven Saturation Mutagenesis in Mice. Current Pharmaceutical
Biotechnology 10(2): 221–29.
Barriere, Herve, Belfodil R, Rubera I, Tauc M, Lesage F, Poujeol C, Guy N, Barhanin J,
y Poujeol P (2003). Role of TASK2 Potassium Channels Regarding Volume
Regulation in Primary Cultures of Mouse Proximal Tubules. The Journal of General
physiology 122(2): 177–90.
Bobulescu IA y OW Moe (2009). “Luminal Na+/H+ Exchange in the Proximal Tubule.”
Pflügers Archiv-European Journal of Physiology 458(1): 5–21.

76
Clark, Robert B, Colleen K, Darrell D y Wayne R (2011). Two-pore Domain K+ Channels
Regulate Membrane Potential of Isolated Human Articular Chondrocytes. The
Journal of physiology 589(Pt 21): 5071–89.
Davidson y Alan (2008). Mouse Kidney Development. StemBook: 1–30.
Del Bigio, M. R. (2010). Ependymal cells: biology and pathology. Acta neuropathologica,
119(1), 55–73.
Derst C y Hirsch JR (2001). Cellular Localization of the Potassium Channel Kir7.1 in
Guinea Pig and Human Kidney. Kidney International 59: 2197–2205.
Derst C, Karschin C, Wischmeyer E, Hirsch JR, Preisig-Müller R, Rajan S, Engel H,
Grzeschik K, Daut J y Karschin A (2001). Genetic and Functional Linkage of Kir5.1
and Kir2.1 Channel Subunits. FEBS letters 491(3): 305–11.
Gawenis, Lara R, Bradford E, Prasad V, Lorenz J, Simpson J, Clarke L, Woo A,
Grisham C, Sanford L, Doetschman T, Miller M y Shull G (2007). Colonic Anion
Secretory Defects and Metabolic Acidosis in Mice Lacking the NBC1 Na+/HCO3-
Cotransporter.The Journal of Biological Chemistry 282(12): 9042–52.
Gestreau C, Dirk H, Joerg T, Dubreuil V, Bandulik S, Reichold M, Bendahhou S,
Pierson P, Sterner C, Peyronnet-Roux J, Benfriha C, Tegtmeier I, Ehnes H,
Georgieff M, Lesage F, Brunet J, Goridis C, Warth R and Barhanin J (2010). TASK2
Potassium Channels Set Central Respiratory CO2 and O2 Sensitivity.” Proceedings
of the National Academy of Science of the UU.AA 107(5): 2325–30.

77
Grondona JM, Granados-Durán P, Fernández-Llebrez P y López-Ávalos M (2013). A
Simple Method to Obtain Pure Cultures of Multiciliated Ependymal Cells from Adult
Rodents. Histochemistry and Cell Biology 139(1): 205–20.
Hamilton, Kirk L y Devor D (2012). Basolateral Membrane K+ Channels in Renal
Epithelial Cells. American Journal of Physiology (9): F1069–81.
Hughes BA y Swaminathan A (2008). Modulation of the Kir7.1 Potassium Channel by
Extracellular and Intracellular pH. American Journal of physiology 294(2): C423–31.
Kanjhan, Refik, Pow D, Noakes P y Bellingham M (2010). The Two-pore Domain K+
Channel TASK-1 Is Closely Associated with Brain Barriers and Meninges. Journal
of Molecular Histology 41(6): 315–23.
Lakatos A (2003). Meningeal Cells Enhance Limited CNS Remyelination by
Transplanted Olfactory Ensheathing Cells. Brain 126(3): 598–609.
LaPointe, Michael S, Sodhi C, Sahai A y Batlle D (2002). Na+/H+ Exchange Activity and
NHE-3 Expression in Renal Tubules from the Spontaneously Hypertensive Rat.
Kidney International 62(1): 157–65.
Lauritzen, Inger, Zanzouri M, Honoré E, Duprat F, Ehrengruber M, Lazdunski M y Patel
A (2003). K+-dependent Cerebellar Granule Neuron Apoptosis: Role of TASK Leak
K+ Channels. The Journal of Biological Chemistry 278(34): 32068–76.

78
Livak KJ y Schmittgen T (2001). Analysis of Relative Gene Expression Data Using Real-
time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods San Diego,
Calif 25(4): 402–8.
Lo, Yi-Fen, Sung-Sun Y, Seki G, Yamada H, Horita S, Yamazaki O, Fujita T, Usui T,
Jeng-Daw T, I-Shing Y, Shu-Wha L y Shih-Hua L (2011). Severe Metabolic Acidosis
Causes Early Lethality in NBC1 W516X Knock-in Mice as a Model of Human
Isolated Proximal Renal Tubular Acidosis. Kidney International 79(7): 730–41.
Niemeyer MI, Cid LP, Barros LF y Sepúlveda FV (2001). Modulation of the Two-pore
Domain Acid-sensitive K+ Channel TASK-2 (KCNK5) by Changes in Cell Volume.
The Journal of Biological Chemistry 276(46): 43166–74.
Niemeyer MI, Gonzalez-Nilo FD, Zuniga L, Gonzalez W, Cid LP, Sepulveda FV (2007).
Neutralization of a single arginine residue gates open a two-pore domain, alkali-
activated K+ channel. Proc Natl Acad Sci U S A 104:666-671.
Niemeyer MI, Cid LP, Peña-Münzenmayer G y Sepúlveda FV (2010). Separate Gating
Mechanisms Mediate the Regulation of K2P Potassium Channel TASK-2 by Intra-
and Extracellular pH. The Journal of Biological Chemistry 285(22): 16467–75.
Ookata K y Tojo A (2000). Localization of Inward Rectifier Potassium Channel Kir7. 1 in
the Basolateral Membrane of Distal Nephron and Collecting Duct. Journal of
Physiology 1987–94.

79
Reyes R, Duprat F, Lesage F, Fink M, Salinas M, Farman M y Lazdunski M (1998).
“Cloning and Expression of a Novel pH-sensitive Two Pore Domain K+ Channel
from Human Kidney. The Journal of Biological Chemistry 273(7): 30863-9.
Schultheis PJ, Clarke LL, Meneton P, Miller ML, Soleimani M, Gawenis LR, Riddle TM, J
Duffy JJ, Doetschman T, Wang T, Giebisch G, Aronson PS, Lorenz JN y Shull GE
(1998). Renal and Intestinal Absorptive Defects in Mice Lacking the NHE3 Na+/H+
Exchanger. Nature Genetics 19(3): 282–85.
Scotto-Lavino E, Guangwei D y Frohman M (2006). Amplification of 5’ End cDNA with
‘New RACE’.” Nature protocols 1(6): 3056–61.
Shieh CC, Coghlan M, Sullivan JP y Gopalakrishnan M (2000). Potassium Channels:
Molecular Defects, Diseases, and Therapeutic Opportunities. Pharmacological
reviews 52(4): 557–94.
Siegenthaler J y Pleasure S (2011). We’ve got you “covered”: how the meninges control
brain development. National institute of health 21: 249-245.
Spassky N, Merkle F, Flames N, Tramontin A (2005). Adult Ependymal Cells Are
Postmitotic and Are Derived from Radial Glial Cells during Embryogenesis. The
Journal of Neuroscience 25(1):10 –18.
Warth R, Barrière H, Meneton P, Bloch M, Thomas J, Tauc M, Heitzmann D, Romeo E,
Verrey F, Mengual R, Guy N, Bendahhou S, Lesage F, Poujeol P y Barhanin J
(2004). Proximal Renal Tubular Acidosis in TASK2 K+ Channel-deficient Mice

80
Reveals a Mechanism for Stabilizing Bicarbonate Transport. Proceedings of the
National Academy of Sciences of the UU.EE 101(21): 8215–20.
Zúñiga L, Márquez V, González-Nilo F, Chipot C, Cid LP, Sepúlveda FV y Niemeyer MI
(2011). Gating of a pH-sensitive K(2P) Potassium Channel by an Electrostatic
Effect of Basic Sensor Residues on the Selectivity Filter. PloS one 6(1): e16141.