tp biologie des membranes - irifaahee.free.fririfaahee.free.fr/cours/nerfsciatiqgre.pdf ·...
TRANSCRIPT

Nom PrénomDateGroupeEnseignant
TP Biologie des membranes :Potentiel d'action du nerf sciatique de grenouille.

Nous voulons étudier les réponses biologiques (potentiel d'action) d'un nerf face à des stimulations électriques de nombres, d'espacements et d'intensités différentes.
Pour ce faire nous disposons d'un dispositif capable d'émettre un (des) signal (signaux) électrique(s) d'intensité (et de durée entre ces signaux) réglable à l'extrémité du nerf et de visionner et mesurer la réponse de ce nerf à l'autre extrémité.
Afin que le nerf possède encore un potentiel de réponse et ne soit pas « mort » nous devons le prélever sur une grenouille tuée juste avant. La première étape de la manipulation est donc la dissection de cette dernière afin de récupérer le nerf sciatique. Il est recommandé toutefois d'effectuer le montage électrique, expliqué en deuxième étape, avant pour ne pas défraichir notre nerf.a - La dissection :Pincez la peau de la grenouille et découpez tout autour au niveau du ventre, puis saississez la dans une main et avec l'autre main tirez sur la peau vers le bas afn de lui découvrir le bas du ventre et les jambes. Puis découpez la grenouille en deux en coupant sous les bras, videz la et épinglez la aux pieds et haut du dos. Il faut ensuite effectuer une petit section sur le bord intérieur du cartilage blanc qui apparaît sur le dos et remontant vers le haut, épinglez le bord que vous avez découper vers l'extérieur libérant le champs sur la colonne dorsale et votre nerf sciatique en dessous. Sans jamais pincer, toucher ou encore couper le nerf, dégagez le en délicatesse en tirant sur les tissus qui le retiennent jusqu'au niveau du genou de la grenouille. Afin de ne pas dessécher notre nerf il est important de l'enduire de liquide de Ringer régulièrement. Lorsque le nerf est suffisament dégagé ligaturez le à chaque extrémité à l'aide d'un petit fil et coupez le nerf juste derrière la ligature. Enfin déposez votre nerf dans la boîte d'étude avec quelques gouttes de liquide de Ringer pour garder une humidité ambiante favorable dans la boîte. Nous pouvons passer à la deuxième étape qui est le dispositif d'expérimentation.b – Le dispositif électrique :Effectuez le circuit électrique comme montré dans le schéma ci dessous.
Boite d'étude avec votre nerf
Oscilloscope
Stimulateur
Sens
du
cour
ant
Amplificateur

Vérifiez bien les réglages au niveau de l'oscilloscope et du stimulateur afin de ne pas faire d'erreurs ou de risquer de griller votre nerf.
Ce premier graphe nous permet d'observer l'artefact de stimulation qui apparait dés le début de la stimulation suivi d'une phase de latence et enfin le potentiel d'action du nerf qui est la somme de tous les potentiels d'actions des fibres de ce nerf. Pour ce premier test nous avons pris une amplitude de 0,4 V, une durée de 0,1 ms et un délai de 1 ms. Même si nous le verrons un peu plus loin nous pouvons observer le recrutement des fibres α et une partie des fibres β comme indiqué sur le graphe 1.
Graphe 1 : Potentiel d'action en réponse à un stimuli d'amplitude 0,4 V

On peut identifier comme précédemment l'artefact de stimulation ainsi qu'une période de latence équivalente à 2,5 carreaux soit : 2,5 ×0,5 =1,25 ms . La période de latence est de 1,25 ms. Après cela on voit l'apparition d'une légère bosse qui correspond au fibres α : V seuil α=0,13 V . Le V seuil α correspond à l'amplitude minimale nécessaire pour obtenir une réponse des fibres α, qui sont les plus grosses dans le nerf et par conséquent plus facile à dépolariser et donc à émettre un potentiel d'action en réponse à un stimuli.
On a cherché à déterminé le V max α qui correspond au potentiel d'action émit par le maximum de fibres α. Celui ci est atteint pour V max α=0,22 V . Au delà de cette amplitude on ne peut pas
Graphe 2 : V seuil de α
Graphe 3 : V max de α

recruter plus de fibres α, on commence le recrutement des fibres β dont nous expliquerons les causes plus loin.
Comme nous l'avons dit dans le graphe précédent si on dépasse le V max α on commence à recruter peu à peu des fibres β. Mais il faut attendre le V seuil β=0,33 V pour enfin apercevoir sur notre oscilloscope le début du pic correspondant au potentiel d'action des fibres β.
Ici nous avons envoyé un signal de 0,8 ms et d'amplitude 0,4 V afin de voir apparaître un petit pic correspondant au début du recrutement des fibres γ. Ce sont les dernières a être recruté en cas de fort signal.
Graphe 4 : V seuil de β
Graphe 5 : Recrutement des fibres γ

Nous pouvons calculer la vitesse de conduction du nerf, c'est à dire la vitesse à laquelle le « message » circule d'une extrémité du nerf à l'autre :
On mesure la distance d entre les capteurs de la boîte : d =4,7 cm=4,7 ⋅10−2 m .Ainsi que la durée de la phase de latence : L=2,2 ×0,5 ms=1,1 ⋅10−3 s .
D'où : V =4,7 ⋅10−21,1 ⋅10−3
=42,73 m⋅s−1 .
Ce qui signifie que le « message » traverse le nerf à une vitesse de 42,73 m⋅s−1 .
Nous abordons maintenant deux méthodes d'études de la période réfractaire relative et de la période réfractaire absolue. La première est la méthode par amplitude qui consiste à fixer la valeur d'amplitude et de faire varier le délai entre les deux signaux. La seconde consiste cette fois à fixer le second potentiel d'action petit et faire varier la durée entre les deux stimuli ainsi que l'amplitude du deuxième afin de garder notre second potentiel d'action petit. Nous verrons les causes de ces périodes dans la partie discussion.
Methode des amplitudes :
Nous effectuons d'abord une première mesure avec une amplitude assez élevée (0,5 V) et un délai de séparation elle aussi élevée afin de définir un « début » qui équivaut à l'obtention de deux potentiel d'action strictement identiques. Ceci nous permettra de comparer les autres courbes afin de bien voir la période réfractaire relative que nous abordons ci dessous.
Graphe 6 : Méthode des amplitudes : début

En réduisant le délai on diminue peu à peu l'amplitude du second potentiel d'action, ici un exemple d'un moment durant la période réfractaire relative dont nous verrons les causes un peu plus bas.
Nous avons continué à diminuer peu à peu le délai entre les deux signaux jusqu'à ce que le second potentiel d'action n'existe plus. Cette absence de réaction correspond à la période réfractaire absolue dont le système sera détaillé plus loin.
Méthode des seuils :Nous allons voir maintenant les résultats de la méthode des seuils.
Graphe 7 : Méthode des amplitudes : période réfractaire relative
Graphe 8 : Méthode des amplitudes : période réfractaire absolue

Ici nous avons réalisé un début à une amplitude de 0,18 V, aux alentours de notre V seuil α , et un délai de 8 ms.
On diminue le délai entre les deux stimulations il faut donc augmenter l'amplitude afin de garder un potentiel d'action aussi élevé que précédemment au début. Ici le délai est de 4 ms et l'amplitude de 0,21 V.
Graphe 9 : Méthode des seuils : début
Graphe 10 : Méthode des seuils : période réfractaire relative
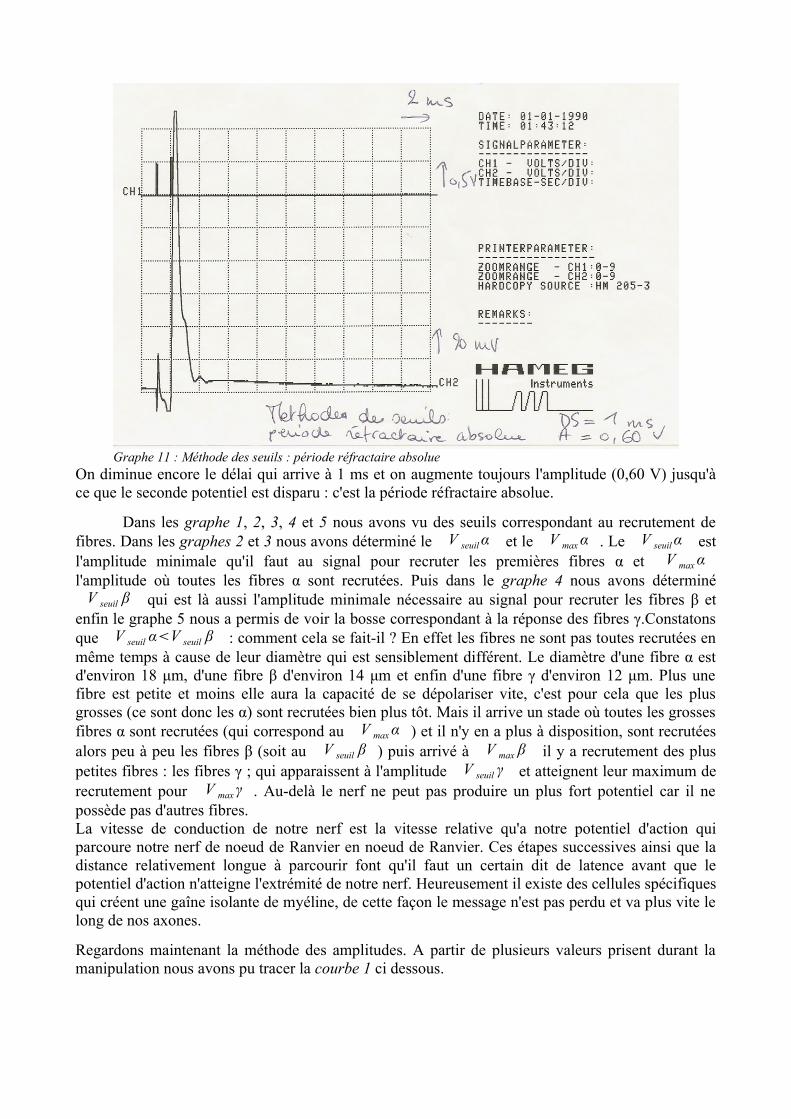
On diminue encore le délai qui arrive à 1 ms et on augmente toujours l'amplitude (0,60 V) jusqu'à ce que le seconde potentiel est disparu : c'est la période réfractaire absolue.
Dans les graphe 1, 2, 3, 4 et 5 nous avons vu des seuils correspondant au recrutement de fibres. Dans les graphes 2 et 3 nous avons déterminé le V seuil α et le V max α . Le V seuil α est l'amplitude minimale qu'il faut au signal pour recruter les premières fibres α et V max α l'amplitude où toutes les fibres α sont recrutées. Puis dans le graphe 4 nous avons déterminé
V seuil β qui est là aussi l'amplitude minimale nécessaire au signal pour recruter les fibres β et enfin le graphe 5 nous a permis de voir la bosse correspondant à la réponse des fibres γ.Constatons que V seuil αV seuil β : comment cela se fait-il ? En effet les fibres ne sont pas toutes recrutées en même temps à cause de leur diamètre qui est sensiblement différent. Le diamètre d'une fibre α est d'environ 18 μm, d'une fibre β d'environ 14 μm et enfin d'une fibre γ d'environ 12 μm. Plus une fibre est petite et moins elle aura la capacité de se dépolariser vite, c'est pour cela que les plus grosses (ce sont donc les α) sont recrutées bien plus tôt. Mais il arrive un stade où toutes les grosses fibres α sont recrutées (qui correspond au V max α ) et il n'y en a plus à disposition, sont recrutées alors peu à peu les fibres β (soit au V seuil β ) puis arrivé à V max β il y a recrutement des plus petites fibres : les fibres γ ; qui apparaissent à l'amplitude V seuil γ et atteignent leur maximum de recrutement pour V max γ . Au-delà le nerf ne peut pas produire un plus fort potentiel car il ne possède pas d'autres fibres.La vitesse de conduction de notre nerf est la vitesse relative qu'a notre potentiel d'action qui parcoure notre nerf de noeud de Ranvier en noeud de Ranvier. Ces étapes successives ainsi que la distance relativement longue à parcourir font qu'il faut un certain dit de latence avant que le potentiel d'action n'atteigne l'extrémité de notre nerf. Heureusement il existe des cellules spécifiques qui créent une gaîne isolante de myéline, de cette façon le message n'est pas perdu et va plus vite le long de nos axones.
Regardons maintenant la méthode des amplitudes. A partir de plusieurs valeurs prisent durant la manipulation nous avons pu tracer la courbe 1 ci dessous.
Graphe 11 : Méthode des seuils : période réfractaire absolue

Cette courbe nous montre que plus on augmente le délai entre les deux signaux et plus l'amplitude de réponse est élevée. Cette croissance atteint un maximum qui correspond au maximum de recrutement soit toutes les fibres ont été recruté. Ceci est dû aux raisons que nous avons expliqué précédemment mais si nous approfondissons un peu, nous pouvons aussi dire que puisque toutes les fibres sont recrutées cela signifie que tous les canaux échangeurs d'ions, responsables de la dépolarisation puis de l'hyperpolarisation, sont mobilisés eux aussi. La phase décroissante se situe durant la période réfractaire relative : c'est à dire que les canaux Na+ ont été ouvert puis refermé et ont une période d'inhibition durant laquelle ils ne peuvent rien faire même si les conditions à leur réouverture sont favorables. C'est cette durée d'inhibition que l'on approche au fur et à mesure que la courbe descend et que l'on atteint la période relative absolue : ici tous les canaux sodiques se sont ouverts puis ont été inhibés par le passage du premier signal mais comme le second arrive trop vite après le celui-ci, la période d'inhibition n'est pas fini et il n'y a plus assez de canaux pour dépolariser la cellule et émettre un potentiel d'action.
Sur cette courbe 2 correspondant à la méthode des seuils on remarque que plus on augmente le délai et plus on diminue l'amplitude du signal. Ici, contrairement à la courbe 1, on a un maximum qui correspond à l'amplitude maximale de stimulation atteinte durant la période réfractaire pour les
Courbe 11 1,5 2 2,5 3 3,5 4 4,5 5 5,5 6 6,5 7 7,5 8 8,5 9 9,5 10
0102030405060708090
100110120130140150160
Amplitude de réponse (mV) en fonction du temps de séparation (ms)
Courbe 21 1,5 2 2,5 3 3,5 4 4,5 5 5,5 6 6,5 7 7,5 8 8,5 9 9,5 10
0,15
0,2
0,25
0,3
0,35
0,4
0,45
0,5
0,55
0,6
0,65
Amplitude de stimulation (V) en fonction du délai (ms)

mêmes raisons que précédemment.
Nous pouvons nous questionner sur laquelle de ces deux méthodes se rapproche le plus de ce qui se passe dans l'organisme. Il est peu probable qu'il y est un succession de très fort signaux qui créent un potentiel d'action maximum mais plutot une multitude de petits signaux plus ou moins espacés qui seront traités par le nerf. La méthode des seuils est peut-être physiologiquement plus vraie que la méthode des amplitudes.
Nous avons donc vu que le nerf réagit de façon différente selon l'amplitude, le délai et la durée du ou des signaux. Ces différentes réactions permettent de traiter les signaux qui lui sont envoyés afin d'inihiber, atténuer, accentuer ceux ci. C'est par ces mécanismes que l'information est traitée puis transmise autour de lui grâce à ces axones.
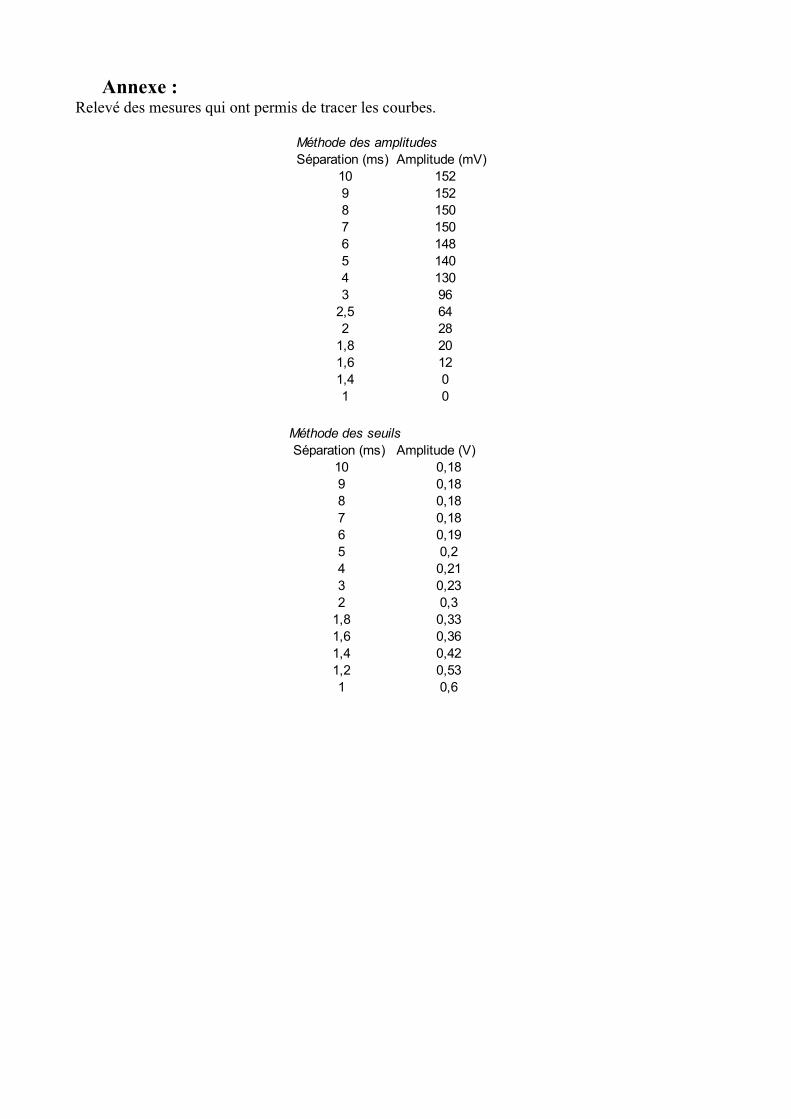
Annexe :Relevé des mesures qui ont permis de tracer les courbes.
Méthode des amplitudesSéparation (ms) Amplitude (mV)
10 1529 1528 1507 1506 1485 1404 1303 96
2,5 642 28
1,8 201,6 121,4 01 0
Méthode des seuils Séparation (ms) Amplitude (V)
10 0,189 0,188 0,187 0,186 0,195 0,24 0,213 0,232 0,3
1,8 0,331,6 0,361,4 0,421,2 0,531 0,6