wk11 glycogen metabolism
TRANSCRIPT

Pentose phosphate PathwayPentose phosphate Pathway
Chapter 20.4Chapter 20.4


Glycolysis Glycolysis 中間代謝物中間代謝物

Transketolase Transketolase 轉酮醇酶轉酮醇酶
TPP 為 transketolase 的輔基 接收 aldose
得到重組的 ketose

Transaldolase Transaldolase 轉醛醇酶轉醛醇酶
Transaldolase Transaldolase 上的 上的 LysLys
脫水脫水 質子化質子化 去質子化去質子化

Transketolase Transketolase 轉酮醇酶轉酮醇酶
Transaldolase Transaldolase 轉醛醇酶轉醛醇酶

磷酸五碳糖的四個代謝模式磷酸五碳糖的四個代謝模式取決於 取決於 NADPHNADPH 、、 Ribose-5-PRibose-5-P 、、 ATPATP

GycolysisGycolysis
當 當 NADPH NADPH 需要量 需要量 < Ribose-5-P < Ribose-5-P 需要量需要量
5 G-6-P + ATP 5 G-6-P + ATP 6 Ribose-5-P + ADP +2H 6 Ribose-5-P + ADP +2H++

Glycolysis Glycolysis 中間代謝物中間代謝物

當 當 NADPH NADPH 需要量 需要量 = Ribose-5-P = Ribose-5-P 需要量需要量
6 G-6-P + 2 NADP6 G-6-P + 2 NADP++ + H + H22OO 5 Ribose-5-P + 2 NADPH+ 5 Ribose-5-P + 2 NADPH+ 2 H2 H++ + CO + CO22

Ribulose-5-P Ribulose-5-P 完全氧化產生 完全氧化產生 NADPH NADPH 進行糖質新生進行糖質新生6 G-6-P + 12 NADP6 G-6-P + 12 NADP++ +7 H +7 H22OO 6 CO 6 CO2 2 + +
12 NADPH + Pi+ 12 H12 NADPH + Pi+ 12 H++

當 當 NADPH NADPH 需要量 需要量 = ATP = ATP 需要量需要量3 G-6-P + 6 NADP3 G-6-P + 6 NADP++ + 5 NAD + 5 NAD++ + 5 Pi +8 ATP + 5 Pi +8 ATP 5 5
pyruvatepyruvate + + 6 NADPH + 5 NADH + 8 ATP6 NADPH + 5 NADH + 8 ATP + 8 H + 8 H22O + 8 HO + 8 H++


體內重要的抗氧化酵素需要 體內重要的抗氧化酵素需要 NADPH NADPH 還原成還原態還原成還原態
2 GSH + ROOH 2 GSH + ROOH GSSG + ROH + H GSSG + ROH + H22OOGlutathione peroxidaseGlutathione peroxidase
Flavoprotein Glutathione reductase Flavoprotein Glutathione reductase (( 需要 需要 NADPH NADPH 使 使 –– S-S- S-S- 還原成 還原成 -SH)-SH)

缺乏 缺乏 G-6-P dehydrogenase G-6-P dehydrogenase 導致溶血性貧血導致溶血性貧血

BiochemistrySixth Edition
Chapter 21:Glycogen Metabolism
Berg • Tymoczko • Stryer

身體需要的能量時,迅速轉換成 glucose ,並
產生能量
Glycogen 的功用在維持血糖的
恒定

Structure of Glycogen
直鏈— α-1,4 l inkage支鏈— α-1,6 l inkage
非還原端

肝細胞中細胞質電子顯微鏡圖

肝糖的三個代謝路徑肝糖的三個代謝路徑
需要能量時
提供組織利用,維持血糖恆定
DNA 、 RNA、 β-
oxidation 等

Glycogen phosphorylase—Glycogen phosphorylase—剪切後以磷酸根取代 剪切後以磷酸根取代 1st 1st 碳的殘基,碳的殘基,
切下一個 切下一個 glucose glucose 分子。分子。

由非環原端逐次切下 glucose 分
子,切下的 glucose 轉換成
G-1-P 。 支鏈的殘基轉移至直鏈部分
切除支鏈的剩下的一個殘基完全直鏈狀
態
G-6-P

(Debranching enzyme)(Debranching enzyme)去分支水解酵素去分支水解酵素
物是 產物是 產 glucoseglucose ,不是 ,不是 G-1-PG-1-P
αα -1,6 glucosidase-1,6 glucosidase

Epimer ( 表異構物 ) : Gal Glc
G-6-P
Glycolysispathway
Phosphoglucomutase
Gal-1-P
G-1-P轉移帶有磷酸根的 uridine 到 Gal-1-P 原本的 UDP-glucose 轉變成
G-1-P
與醣類代謝路徑接軌的酵素

PhosphoglucomutasePhosphoglucomutase
與醣類代謝路徑接軌的酵素

G-6-P
Glucose
Glycolysis
糖質新生,維持血糖恆定
糖解代謝,產生能量
Pentose phosphate pathway
PhosphoglucomutasePhosphoglucomutase
G-6-PhosphaseG-6-Phosphase只有 只有 Liver Liver 才具有的酵素才具有的酵素
內質網內質網Glucose
主要提供腦細胞與主要提供腦細胞與骨骼肌利用骨骼肌利用

Glycogen phosphorylase
HomodimerHomodimer同分子量同分子量
可與磷酸鹽受可與磷酸鹽受質結合的部位質結合的部位

Glycogen phosphorylase
所需要的輔酶
Pyridoxine (Vit B6) 的衍生物

GlucoseGlucose G-1-PG-1-P
Glycogen phosphorylase
作用機制

Glycogen phosphorylase
的活性調控
Insulin胰島素 Epinephri
ne腎上腺素
Glucagon昇糖激素
Regulation Glycogen MetabolismRegulation Glycogen Metabolism

Muscle Liver
Glycogen phosphorylase
的活性調控
產生能量供 Muscle 利用 維持全身組織 glucose 的恒定

Muscle Glycogen Phosphorylase
Phosphorylase aRelaxed state
不受 AMP 、 ATP 與 G-6-P 影響,一直具有活性
Phosphorylase b常以 Tense state
High AMP conc. 時有活性 R stateHigh ATP conc. 低活性 T stateHigh G-6-P 抑制活性 T state Epinephrine ↑
興奮、恐懼、運動
Phosphorylase kinase

平衡時以 平衡時以 Tense state Tense state
存在,不具有活存在,不具有活性。性。
AMP↑AMP↑ ATP↑ATP↑
Phosphorylase kinase
Phosphorylase kinase
具有活性
具有活性
Epinephrine ↑興奮、恐懼、運動
G-6-P↑G-6-P↑

缺乏能量時
能量充足時
不具有活不具有活性性
具有活性具有活性

LiverLiver 中 Phosphorylase 的調控方式— 當血糖濃度低時活性增加。
不具有活不具有活性性
具有活性具有活性
Glucose Glucose 充充足,足, Glycogen Glycogen 沒有沒有
必要分解產生 必要分解產生 glucoseglucose
不受 不受 AMP AMP 影響,影響,因為 因為 liver l iver 中不如中不如
肌肉中會有劇烈的能肌肉中會有劇烈的能量變化。量變化。

增加 Ca2+ 含量
磷酸化
β-subunit 被磷酸化
δ-subunit 被鈣離子化
得到最大活性
Phosphorylase kinase

Epinephrine 與 Glucagon 對於肝醣代謝之調控

胰臟
腎髓質
ReceptorReceptor
ReceptorReceptor

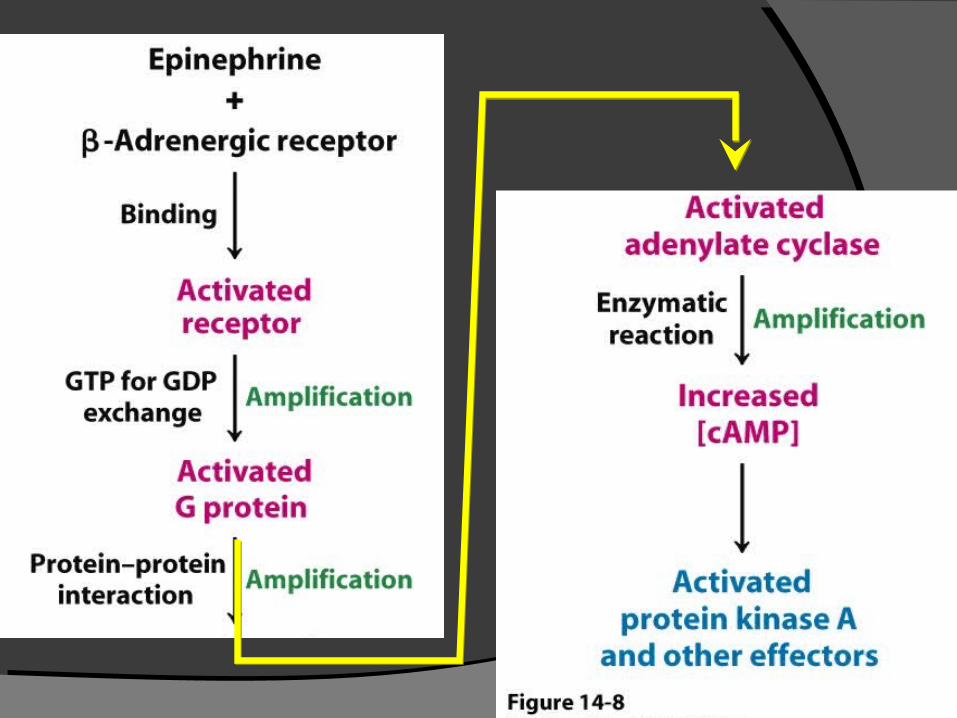

1. Epinephrine 與 Glucagon 進入 transmenbrane 與特定 receptor 結合 。
2. 結合後活化 Gs protein ,然後活化 G-protein 。
3. GTP Gs subunit 活 化 adenylate cyclase 。
1. Epinephrine 與 Glucagon 進入 transmenbrane 與特定 receptor 結合 。
2. 結合後活化 Gs protein ,然後活化 G-protein 。
3. GTP Gs subunit 活 化 adenylate cyclase 。
GTPase
終止肝醣分解

Regulatory Cascade for Glycogen Breakdown.
1.Glycogen degradation is stimulated by hormone
binding to 7TM receptors.
2.Hormone binding initiates a G-protein-dependent G-protein-dependent
signal-transduction pathway signal-transduction pathway
3.that results in the phosphorylation and activation activation
of glycogen phosphorylase.of glycogen phosphorylase.

Glycogen synthesis


(Uridine triphosphate)
(UDP-Glucose pyrophosphoryla
se)
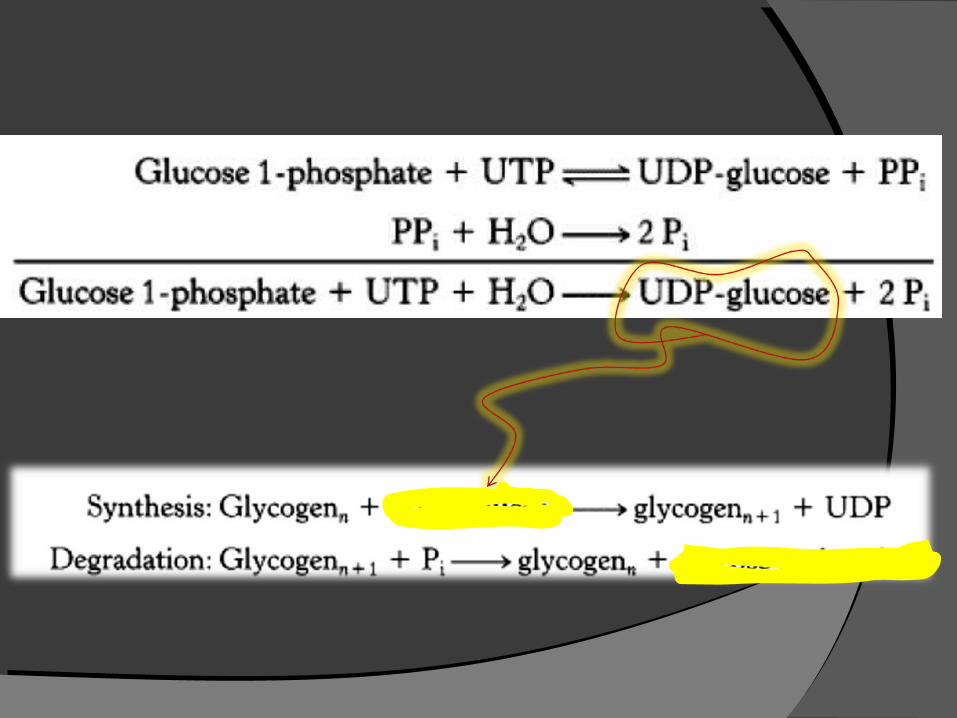
UDP-glucose, the glucose donor in the biosynthesis of glycogenglucose donor in the biosynthesis of glycogen, is an activated form of glucoseactivated form of glucose, just as ATP and acetyl CoA are activated forms of orthophosphate and acetate, respectively.
hydroxyl group is esterified to the hydroxyl group is esterified to the diphosphate moiety of UDP.diphosphate moiety of UDP.

Glycogen synthase
Glycogen 合成的關鍵酵素—只能作用在超過 4 個殘基的多醣鏈
上需要依賴 Glycogenin ( 肝醣合成蛋
白 )

Glycogenin ( 肝醣合成蛋白 )
分支 α-1,6 由 branching enzyme 轉移合成
A Branching Enzyme Forms α-1,6 Linkages

Branching is important because it increases
the solubility of glycogen.
Furthermore, branching creates a large number
of terminal residues, the sites of action of
glycogen phosphorylase and synthase
Thus, branching increases the branching increases the rate of glycogen
synthesis and degradation.

Glycogen branching requires a
single transferase activity.
Glycogen debranching requires
two enzyme activities: a transferase and
an α -1,6 glucosidase.

Synthase a
不受 G-6-P 影響,一直具有活性
Synthase b
不具活性Glycogen
sythatase kinase/
Protein kinase A
High G-6-P concentration
合成肝醣

G-6-P G-1-P UTP
UDP-glucosePPi
H2OPi
Glycogen (n)
Glycogen (n+1)UDP
ATP
ADP
Glycogen synthesis pathway

抑制肝醣合成
運動時需要肝醣分解以提高血糖濃度與能量

bbaa
具有活性不具活性
PP1

使 GM 磷酸化 與 PP1
分離
肌肉中肝醣結合位置
肌肉中的小分子 protein
運動過程中,使 Protein Phosphotase 1 (PP1) 失去活性以抑制肝醣生成之作用機
制

Insulin 對於 Glycogen synthesis 的調控—Glycogen synthase
kinase



(Insulin receptor substrate)(Insulin receptor substrate)
Insulin 活化 Glycogen sythase kinase 活性以
促進 Glycogen synthesis
作用在使 作用在使 Glycogen Glycogen synthesis synthesis 維持在不維持在不
活化的狀態活化的狀態不活化狀態不活化狀態
活化狀態活化狀態
活化狀態活化狀態 不活化狀態不活化狀態

