encimska kinetika govori o hitrosti encimske...
TRANSCRIPT

Encimska kinetika govori o hitrosti
encimske reakcije
• uvod
• encimska kinetika po Michaelisu in Mentenovi
• kinetika večsubstratnih encimskih reakcij
• kinetika alosteričnih encimov
• encimska inhibicija

encimsko katalizirana kemijska reakcija
E + S ↔ ES ↔ EP ↔ E + P
nekatalizirana kemijska reakcija
S ↔ P substrat produkt
Kako encimi delujejo?
E – encim
S – substrat, P – produkt
ES – kompleks encim-substrat
EP – kompleks encim-produkt POZOR, to nista aktivirana kompleksa!
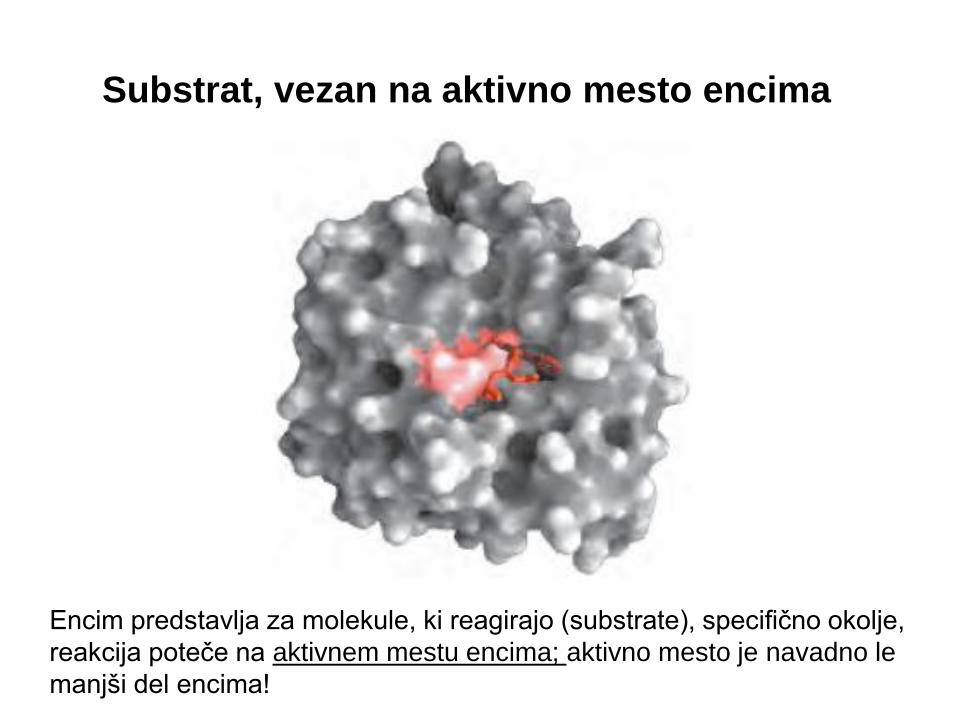
Substrat, vezan na aktivno mesto encima
Encim predstavlja za molekule, ki reagirajo (substrate), specifično okolje,
reakcija poteče na aktivnem mestu encima; aktivno mesto je navadno le
manjši del encima!

Hitrost (v) reakcije S → P določa
hitrostna konstanta k, to pa DG‡
v = k · [S] reakcija S → P
v = k · [S1] · [S2] reakcija S1 + S2 → P
k – Boltzmannova konstanta (n.R/NA; n = št. molov, R = plinska
konstanta, NA = Avogadrova konstanta)
h – Planckova konstanta
DG‡ – aktivacijska energija (aktivacijska prosta entalpija)
R, T – konstanti
k
k

Primerjava nekatalizirane in katalizirane reakcije
Nekatalizirana reakcija
S ↔ P
• Ravnotežje
• Večja aktivacijska energija
• Manjša reakcijska hitrost
• Reakcijski mehanizem preko
prehodnega stanja
Katalizirana z encimom
E + S ↔ ES ↔ EP ↔ E + P
• Ravnotežje
• Manjša aktivacijska energija
• Večja reakcijska hitrost
• Reakcijski mehanizem preko
drugih reakcijskih
intermediatov in prehodnih
stanj
Katalizatorji pospešijo hitrost reakcije (znižajo aktivacijsko
energijo), ne spremenijo pa ravnotežja reakcije!
S
PK
S
PK
Aktivacijska energija ΔG# določa hitrost reakcije

S ↔ P
E + S ↔ ES ↔ EP ↔ E + P
ΔGo = ΔGo = - RT ln K ΔG# < ΔG# v > v
DGo

S ↔ P
E + S ↔ ES ↔ ES1 ↔ ES2 ↔ EP ↔ E + P
- če je reakcija sestavljena iz zaporedja več reakcij,
hitrost celotne reakcije določa najpočasnejša stopnja (največja aktivacijska energija) –
“rate limiting step”
- energijske bariere so ključnega pomena za življenje – če jih ne bi bilo, bi se makromolekule spontano razgrajevale v osnovne gradnike ...
- med evolucijo (zelo dolg čas!) so se razvili encimi, ki selektivno znižujejo aktivacijsko bariero

ES - komplementarnost substrata z vezavnim mestom na encimu:
šibke interakcije so optimizirane v prehodnem stanju
Emil Fischer 1894
E in S - ključ v ključavnico
Inducirano prilagajanje encima
substratu in substrata encimu
Encim je komplementaren
reakcijskemu prehodnemu stanju
Aktivacijska energija se zniža
zaradi sproščene vezavne energije
ob nastanku šibkih interakcij med
encimom in substratom
Primer: encim dihidrofolatna
reduktaza; NADP+ (rdeče) in
tetrahidrofolat (rumeno) sta
substrata.

Imaginarni encim, ki katalizira prelom kovinske palice
S S‡ P

Peptidaza ‘prime’ substrat (dipeptid) z najmanj tremi
šibkimi vezmi, mu vsili novo konformacijo v kateri je
peptidna vez že ‘natrgana’ (prehodno stanje!)
ENCIM
SUBSTRAT (S1)
VODA (S2)
S1 + S2 P1 + P2

Encimska kinetika
• Obravnava hitrost, s katero se spreminjajo reaktanti v
produkte S → P
• Hitrost podaja spremembo koncentracije substrata/produkta
v časovni enoti (mol/s) v = -d(S)/dt = d(P)/dt
• Hitrost encimske reakcije je merilo za aktivnost encima;
enote za aktivnost: U = 1μmol/min ali katal (kat) = 1mol/s
• Specifična aktivnost encimskega pripravka: hitrost na
št. encimskih enot/mg proteina, npr. μmol/min/mg
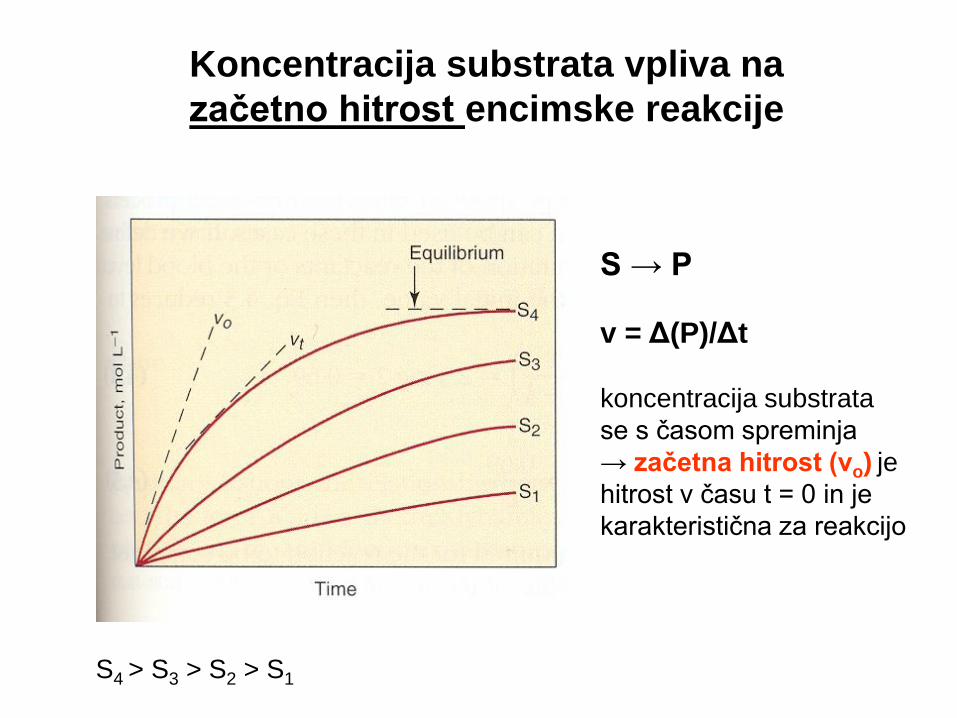
Koncentracija substrata vpliva na
začetno hitrost encimske reakcije
S → P
v = Δ(P)/Δt
koncentracija substrata
se s časom spreminja
→ začetna hitrost (vo) je
hitrost v času t = 0 in je
karakteristična za reakcijo
S4 > S3 > S2 > S1

ODVISNOST ZAČETNE HITROSTI (v0)
OD KONCENTRACIJE SUBSTRATA [S].
POZOR: TO STA DVA RAZLIČNA GRAFA!

Vpliv koncentracije substrata na začetno
hitrost encimske reakcije
maksimalna hitrost Vmax
Michaelisova konstanta Km
Graf ustreza enačbi:
a morajo biti izpolnjeni določeni
pogoji!
Enačba se imenuje po
Michaelisu in Mentenovi, ki sta
jo prva izpeljala (članek 1912).
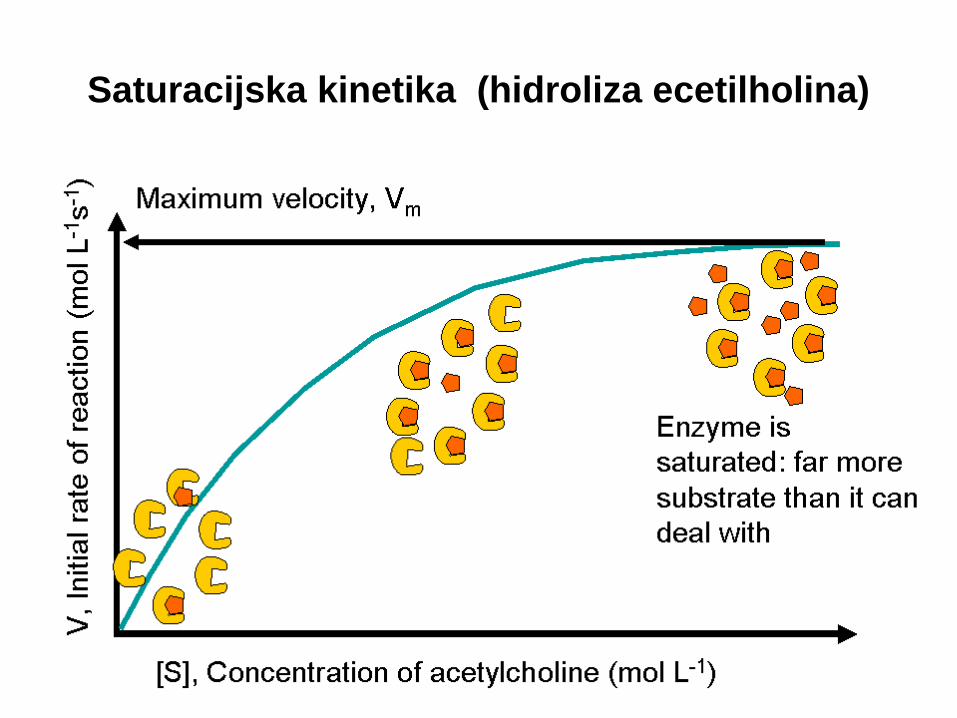
Saturacijska kinetika (hidroliza ecetilholina)

Odvisnost začetne hitrosti v0 od koncentracije
substrata S (v0=k.[S]n)
V0=k[S]1
1. red reakcije
V0=k[S]0
0. red reakcije

Vpliv koncentracije substrata začetno hitrost
encimske reakcije
Enačba po Michaelisu in Mentenovi
Predpostavke pri izpeljavi in namen
- koncept začetne hitrosti: v začetku je koncentracija P
zanemarljivo majhna, ni reakcije P → S (k-2 = 0)
- razgradnja ES določa hitrost celotne reakcije v0 = k2[ES]
- encim ima eno samo aktivno mesto, če pa jih je več, so ta
neodvisna
- (E)o << (S)o
NAMEN: ker je težko izmeriti [ES] (v0 = k2([ES])), določimo Km
in Vmax ter z njima okarakteriziramo encimsko aktivnost

Michaelis-Mentenova kinetika
Ključnega pomena za encimsko katalizo je nastanek
kompleksa ES
L. Michaelis in M. Menten 1912
hitra reverzibilna stopnja
počasna reverzibilna stopnja,
razpad ES določa hitrost celotne reakcije
Z večanjem koncentracije S se vse več encima nahaja v obliki kompleksa
ES, encim postaja nasičen s substratom, zato saturacijska kinetika!
(ES) je konstantna po času, stacionarno stanje – “steady state”
v0 = k2.[ES]
S ↔ P
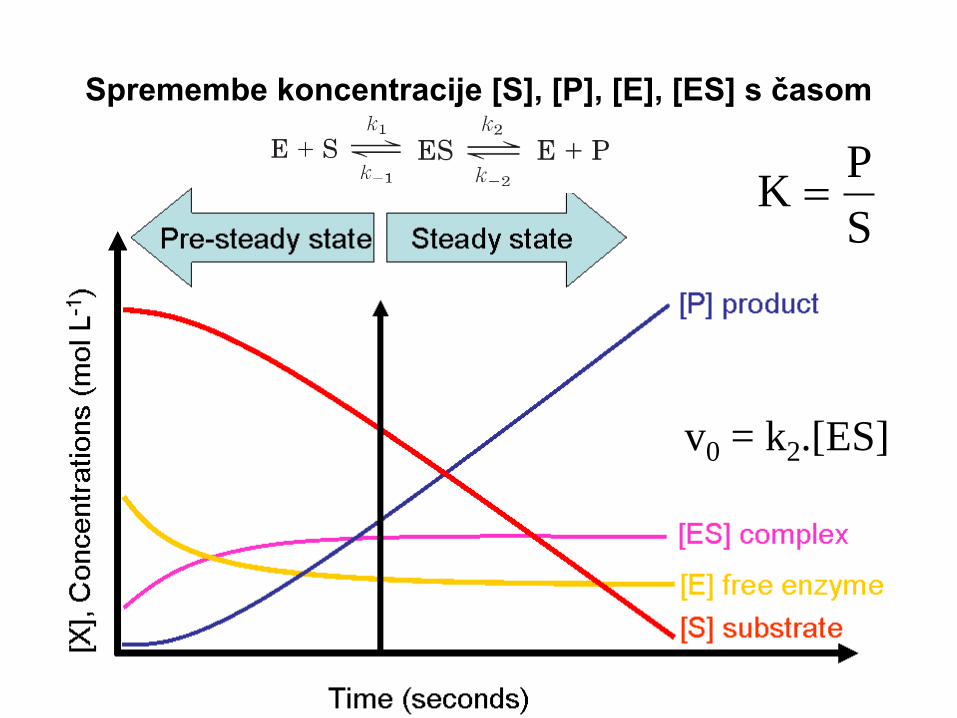
Spremembe koncentracije [S], [P], [E], [ES] s časom
v0 = k2.[ES]
S
PK

hitrost nastanka ES =
hitrost razpada ES =
Michaelisova konstanta Km
v0 = k2.[ES]
V stacionarnem stanju
sta ti dve hitrosti enaki!

Michaelis-Mentenova enačba
Km

Odvisnost začetne hitrosti od koncentracije
substrata – grafična določitev konstant Km in Vmax
[S] << Km →

Transformacija Michaelisove enačbe v dvojno
recipročno enačbo (Lineweaver-Burk)
Dvojni recipročni diagram, 1/V0 proti
1/[S], uporaben za določitev Vmax in Km

Izražanje in primerjava katalitične
učinkovitosti različnih encimov
• Km
• kcat
• kcat/Km

Izražanje in primerjava katalitične učinkovitosti
različnih encimov, Km
• Vmax , Km eksperimentalno določimo
• Km - odraža afiniteto encima do substrata
- odraža fiziološke razmere - za mnoge encime valja:
Km ≈ [S]fiziol.
≈ = Kd
≈
če merimo vo !
VELJA: - velika Km majhna afiniteta encima do substrata
- majhna Km velika afiniteta encima do substrata

Vrednosti Km nekaterih encimov in substratov

Pomen Km v fiziologiji primer: občutljivost Azijcev na alkohol
• Japonci in Kitajci dosežejo isti učinek alkohola (vazodilatacija, pospešen ritem srca...) že pri nižji koncentraciji zaužitega alkohola kot Evropejci
• Reakcije razgradnje alkohola
• CH3CH2OH + NAD+ → CH3CHO + H+ + NADH alkoholna dehidrogenaza
• CH3CHO + NAD+ → CH3COOH + H+ + NADH aldehidna dehidrogenaza
• Azijci imajo aldehidno dehidrogenazo z višjim Km → (aldehid v krvi dolgo kroži po telesu); izoencim!
• Izoemcimi: katalizirajo isto reakcijo (isti substrat), različne pa so molekulske lastnosti encimov: različna encimska aktivnost (kkat in Km), različna molekulska masa (Mr). Genski zapis je drugačen, zato druga sekvenca!

Izražanje in primerjava katalitične učinkovitosti
različnih encimov, kkat
• kkat = k najpočasnejše stopnje! (v smeri, ki daje produkt!)
• kkat - pretvorbeno število: št. molekul substrata,
ki se pretvori v produkt na eni molekuli encima v
enoti časa (encim je nasičen s substratom)
Vmax = k2.[E]t
Vmax = kkat.[E]t ; ESkv kat0

Kinetična učinkovitost encimov: pretvorbeno
število (turnover number), kkat
Slabost izražanje encimske učinkovitosti s Km ali kkat: ni upoštevana
neencimska reakcija, ni informacije za koliko encim pospeši reakcijo

Najboljši način za izražanje encimske
aktivnosti: kkat/Km

Alosterični proteini (encimi)
• Alosterični proteini (encimi) sestojijo iz več podenot (multimerni proteini) in imajo več vezavnih mest (za substrat in/ali modulator)
• Alosterični proteini (podenote) zavzamejo različne konformacije
• Konformacijsko spremembo (prehod v manj ali bolj aktivno obliko) povzroči modulator (ligand, pri encimih tudi S)
• Modulatorji so ali inhibitorji ali aktivatorji
• Če je normalni ligand obenem modulator – homotropična modulacija (O2 na Hb, substrat pri encimu) ; če je modulator druga molekula – heterotropična modulacija
• Mnogi alosterični proteini imajo več modulatorjev – obenem gre za heterotropične in homotropične modulatorje
• Vezava liganda na multimerni protein (O2 na Hb, S na encimu) je kooperativna vezava – vezava enega liganda vpliva na afiniteto drugih vezavnih mest (O2 je ligand in pozitivni homotropični aktivator Hb, S deluje enako pri alosteričnem encimu)
• Za kooperativno vezavo je značilna sigmoidna krivulja
• Sigmoidna krivulja omogoča občutjiv odgovor na majhne spremembe koncentracije liganda v okolju

Hb (2D5Z.pdb)

Prehod Hb iz T (deoksi) v R (oksi) konformacijo
konformacija T konformacija R
vezava O2

(a) Usklajen model: vse podenote so v isti konformaciji, bodisi bodisi
(b) Skupni model: vsaka podenota je lahko ali v eni ali v drugi konformaciji, ali
2 modela za pretvorbo neaktivne v aktivno obliko
proteina po kooperativni vezavi liganda; pri encimih je
ligand kar S (L = S)

Saturacijske krivulje Mb in Hb
Vezava O2 na Mb in Hb Vezava O2 na R, T in naravni Hb
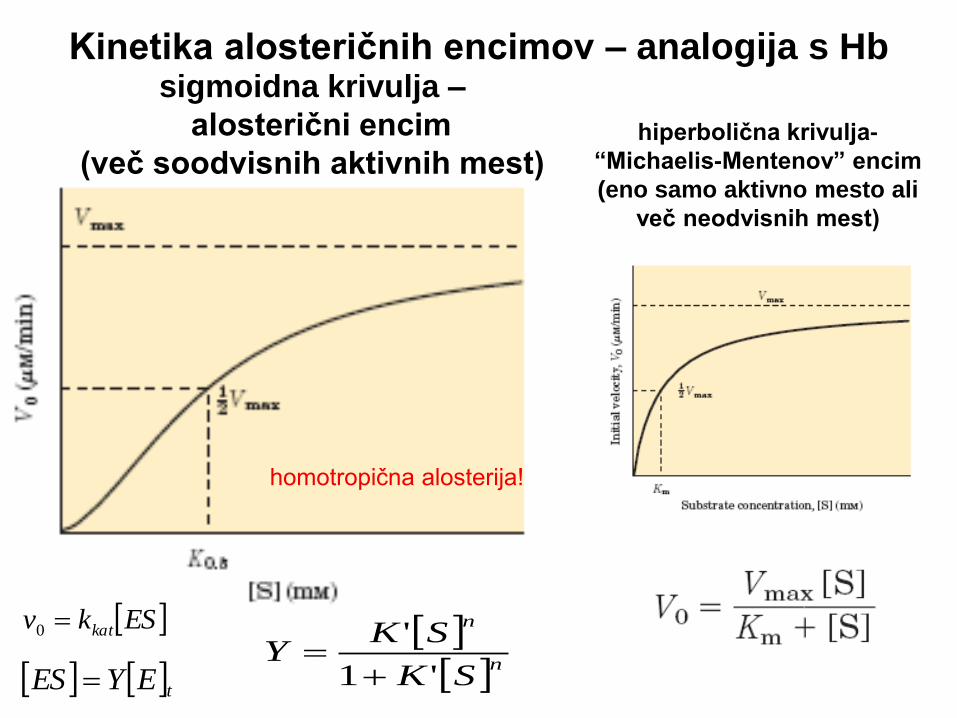
hiperbolična krivulja-
“Michaelis-Mentenov” encim
(eno samo aktivno mesto ali
več neodvisnih mest)
sigmoidna krivulja –
alosterični encim
(več soodvisnih aktivnih mest)
Kinetika alosteričnih encimov – analogija s Hb
homotropična alosterija!
n
n
SK
SKY
'1
'
ESkv kat0
tEYES

Dve vrsti modulatorjev alosteričnih encimov
modulatorji vplivajo na Km modulatorji vplivajo na Vmax
heterotropična alosterija!
S S
M M

Bi(več)substratne reakcije (velikanska večina je takih!)
S1 + S2 ↔ P1 + P2
Primer: prenos funkcionalne skupine (fosforilne, amino...) z enega
substrata na drug substrat
ATP + D-glukoza ↔ ADP + D-glukoza-6-fosfat
ATP + D-fruktoza ↔ ADP + D-fruktoza-6-fosfat
ak1 + ketokislina2 ↔ ketokoslina1 + ak2

Prenos fosforilne skupine z enega na drug
substrat
ATP + D-glukoza ↔ ADP + D-glukoza-6-fosfat
ATP + D-fruktoza ↔ ADP + D-fruktoza-6-fosfat
S1 + S2 ↔ P1 + P2
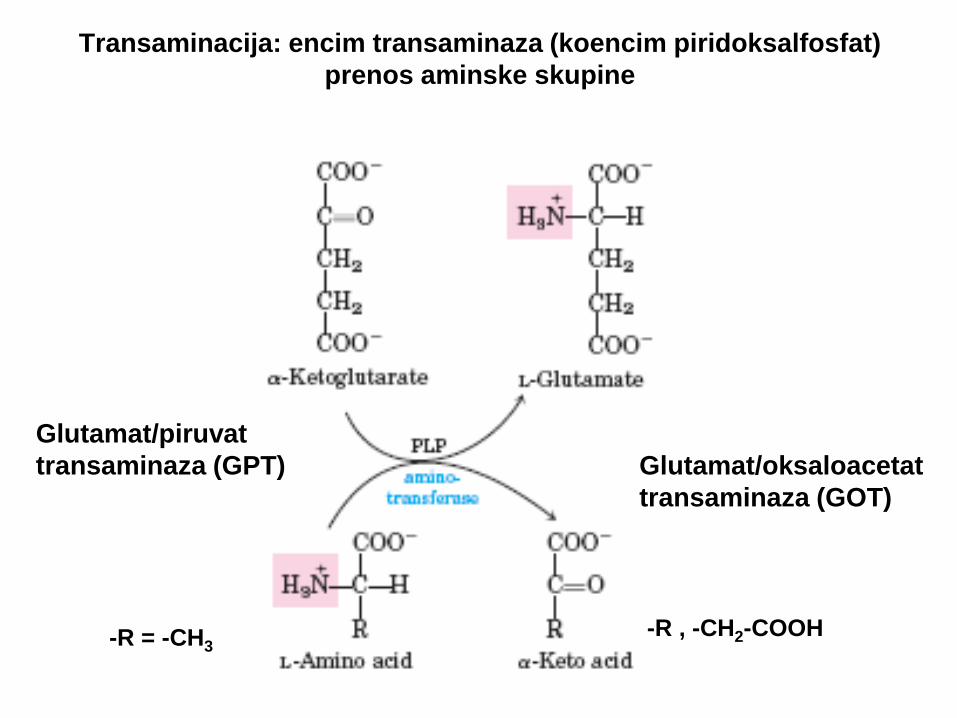
Transaminacija: encim transaminaza (koencim piridoksalfosfat)
prenos aminske skupine
Glutamat/piruvat
transaminaza (GPT)
Glutamat/oksaloacetat
transaminaza (GOT)
-R = -CH3 -R , -CH2-COOH
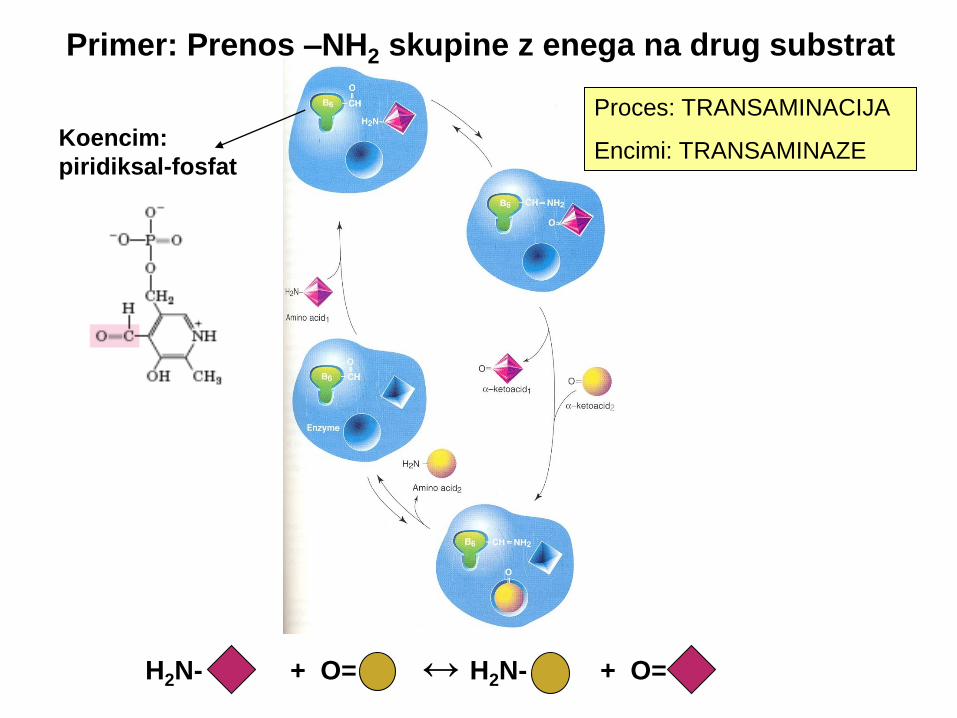
H2N- + O= ↔ H2N- + O=
Koencim:
piridiksal-fosfat
Primer: Prenos –NH2 skupine z enega na drug substrat
Proces: TRANSAMINACIJA
Encimi: TRANSAMINAZE

Mehanizem encimsko kataliziranih
bisubstratnih reakcij
Encimske reakcije, ki vključujejo ternarni kompleks
Encimske reakcije, pri katerih ne nastaja ternarni kompleks
(ping-pong mehanizem)

Kako ugotoviti mehanizem bisubstratnih
reakcij? Kinetična analiza bisubstratnih reakcij
(dvojni recipročni diagram)
potek reakcije preko
ternarnega komleksa ping-pong mehanizem

Na encimsko aktivnost vplivajo zunanje razmere
• Vmax , Km – eksperimentalno določimo
• Vmax , Km – karakteristični za encim
• Vmax , Km – spreminjata se z zunanjimi razmerami:
- pH
- temperatura
- ionska moč
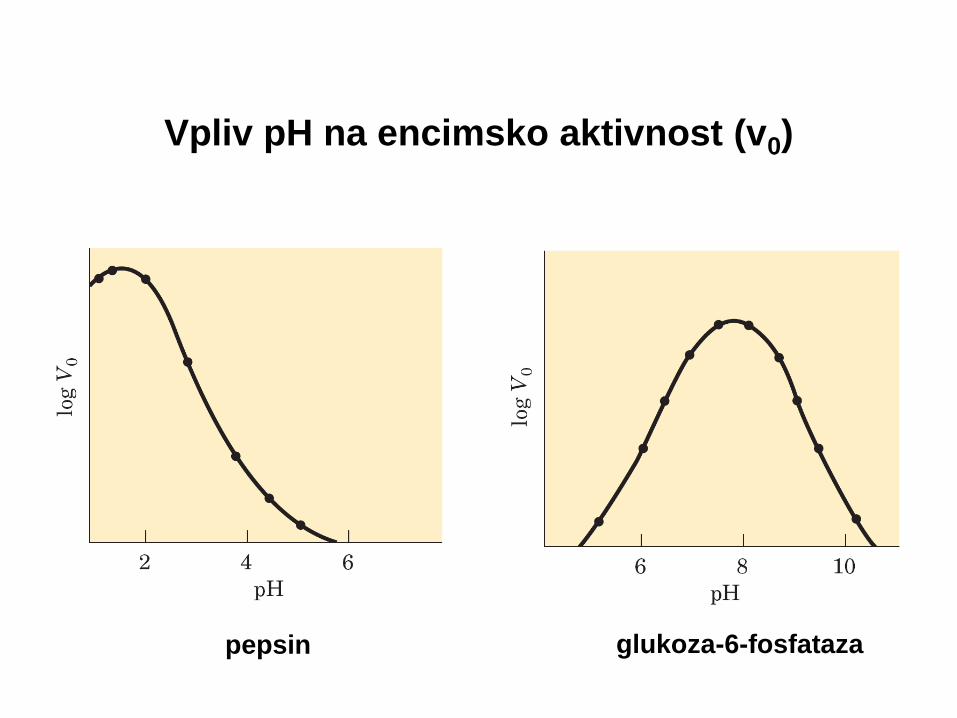
Vpliv pH na encimsko aktivnost (v0)
pepsin glukoza-6-fosfataza
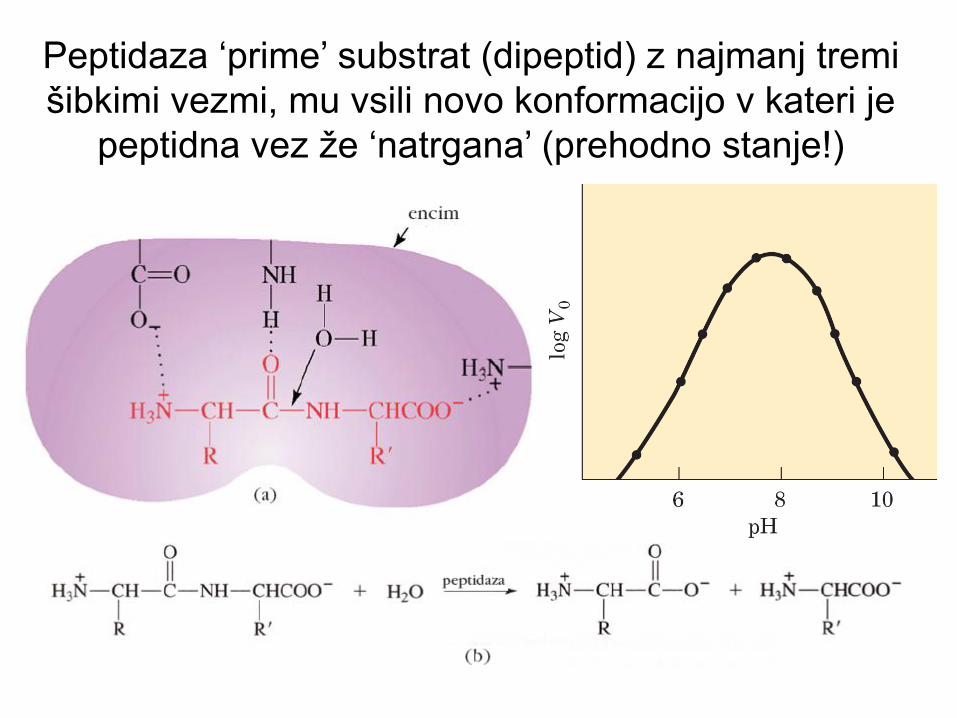
Peptidaza ‘prime’ substrat (dipeptid) z najmanj tremi
šibkimi vezmi, mu vsili novo konformacijo v kateri je
peptidna vez že ‘natrgana’ (prehodno stanje!)
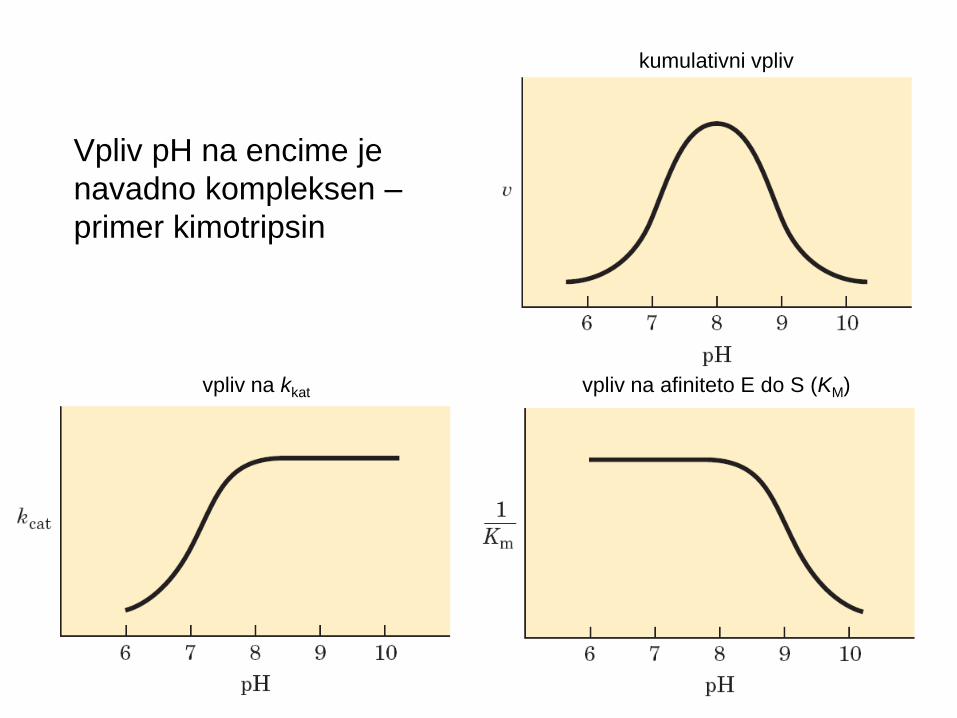
Vpliv pH na encime je
navadno kompleksen –
primer kimotripsin
kumulativni vpliv
vpliv na kkat vpliv na afiniteto E do S (KM)

Vpliv pH in temperature na encimsko
aktivnost (V)