pharma vision newsbukai.pharm.or.jp/bukai_vision/news/no_10.pdf社団法人 日本薬学会...
TRANSCRIPT

日本薬学会
薬学研究ビジョン部会
Pharma
VISION NEWS
No. 10
巻頭言大学における創薬研究、教育のあり方を
長瀬博(北里大学薬学部教授
ファーマビジョンニュース編集副委員長)
1
薬学研究ビジョン創薬研究への情報計算技法の新しい役割
神沼二眞(広島大学理学部)
2
薬学研究最前線
(1)シナプス可塑性に対するアミロイドβと神経ステロイドの効果-標的分子と作用機序の解析-
曽我部正博1,2,3、陳玲1,2,4(1名古屋大学大学院・医学系研究科・細胞生物物理学、2JST・ICORP/SORST・細胞力覚、3生理学研究所・分子生理、
4南京医大・神経生理)
6
(2)ヒストン脱アセチル化酵素を分子標的とした抗がん剤の探索研究鈴木孝禎、伊藤幸裕、宮田直樹(名古屋市立大学大学院薬学研究科)
12
(3)タンパク質構造変化の新規計測法の開発廣田俊(奈良先端科学技術大学院大学
物質創成科学研究科/科学技術振興機構さきがけ)
17
(4)
GPIアンカー型タンパク質の脂質リモデリング機構の解明梅村真理子、地神芳文((独)産業技術総合研究所セルエンジニアリング研究部門)22
(5)核磁気共鳴法による高分子量タンパク質間相互作用の解析嶋田一夫(東京大学大学院薬学系研究科、産業技術総合研究所生物情報解析研究センター)
27
薬学研究ビジョン部会からのお知らせ
33
編集後記
長洲毅志(エーザイ株式会社)
34
Index( Sep.
2007
)

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 1
巻 頭 言
大学における創薬研究、教育のあり方を
長瀬 博(北里大学薬学部教授 ファーマビジョンニュース編集副委員長)
今年のファルマシアの 4 月号で「創薬の魅力を
伝える」というタイトルで大学と企業の研究者の
座談会を行ったが、その際に大学における創薬教
育について議論が出た。薬学部の教育を受けた学
生が企業に必ずしも製薬企業の研究所にとって
即戦力になっておらず、理学部、工学部、農学部
の学生と比べ、やや薬の知識は多いが、それも入
社数年後には追いつかれてしまう程度であると
いう意見もあった。また、薬学部の教育では個々
の分野について(例えば、有機化学、薬理学、製
剤、ADME、医療薬学)の教育はするが、それぞ
れの分野を通じて創薬という流れに応用できる
教育は余りなされていないということで意見が
一致していた。一方で、6 年制が開始され、益々、
創薬に興味を持つ学生が少なくなる傾向が出て
くることが予想され、企業の求める創薬科学者の
育成が困難になることも恐れられている。欧米の
薬学部(Dept. of Medicinal Chemistry)では実際に
創薬をターゲットにしたテーマで研究をしてお
り、教育者も有機化学者、薬理学者、製剤学者と
いうのでは無く実際に企業で創薬を実施してい
た研究者が大学に移籍して研究教育をしている
者が多い。従って、企業に入社後に創薬の教育を
何年も実施しなくても即戦力として活動できる
研究者を育成できている。この点でも、欧米の製
薬企業と日本とでは大きな違いがある。これから、
益々、探索研究に費用がかかる様になり、製薬企
業は M&A を盛んに行い、多くの研究費を捻出で
きるところでないと競争力が出ないとも言われ
ている。そこで、日本の製薬企業の競争力向上の
ためにも薬学部の創薬教育を充実させ、即戦力の
学生の育成に注力する必要があると感じる。その
ためには、企業と大学の人的交流を盛んにし、企
業の研究経験者が大学で研究教育を行える環境
を作る必要があると思う。さらに、企業は実際に
実施している創薬ターゲットを大学に積極的に
開示することが重要であり、大学では、創薬ター
ゲットから臨床開発に至る長い研究期間の内容
を理解し、ターゲット情報を自分の研究に取り入
れる能力を有する研究者が求められる。日本の創
薬化学者は創薬科学者になっていない場合が多
く、化学は強いが科学は強くない研究者が多い傾
向がみられる。そのような人のために、薬学研究
ビジョン部会が役立つものと思われる。本ニュー
スレターでは創薬研究の死命を制すると言われ
る創薬ターゲットに関する情報が多く、ビジョン
シンポジウム、部会フォーラムでは大学-企業間
の異動の機会にも役立つものと期待される。
◆略 歴◆ 長瀬 博 (Hiroshi NAGASE):北里大学薬学部(薬学研究科)・
教授 1976年名古屋大学大学院理学研究科博士課程修了。東レ基礎研究所入社。主
任研究員、所長を経て2004年より北里大学薬学部教授。 この間、1985年-1987年ミネソタ大学(米国)へ留学。専門はメディシ
ナルケミストリー。すでに安定型 PGI2 誘導体、ベラプロストナトリウム(ドルナ
ー)を上市。さらに現在、ナルフラフィン(TRK-820)を止痒薬として承認申請
中。現在、大学からの創薬を目標に研究を行っている。

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 2
薬学研究ビジョン
創薬研究への情報計算技法の新しい役割
神沼 二眞(広島大学理学部)
はじめに 現在私は、本部会のような専門家のための情報
誌に気の利いたことを書けるだけの仕事をして
いないが、あえてということで、具体性に欠ける
というご批判を覚悟の上で、この数年提唱してい
る「核内受容体と生活習慣病への情報計算からの
アプローチ」という構想1)を下敷きにして、表
題のような Vision を語らせていただきたい。 Vision を語るためには、まず、創薬 Drug
Discovery という関心領域を取り巻く環境の変化
をどう認識しているかを語らねばならない。その
変化を要約すれば、1)進歩が加速度的となる, 2)大量のデータの処理が発生し、情報計算の技
法と専門家への依存度が高まってくる, 3)ヒト
遺伝学が医学の中核知識基盤になってくる, 4)
酵母、線虫、ハエなどのモデル生物を材料とする
比較(進化)ゲノム学が盛んになってくる, 5)
疾病の概念が臓器ごとから遺伝子と経路網を基
礎に理解されるようになってきている, 6)個別
的な医療 Personalized Medicine が求められてい
る, 7)予兆的段階での対策、予防的な対策への
対応が求められてくる, 8)医療の提供者と利用
者との情報知識の共有が必要になってくる、とい
うことになる。 この変化は、Genome、Omics, Pathway/Network
(GOP/N)が生物医学の研究機軸となった時代と
も要約できる。このような変化は、生物医学に関
係する IT およびそれを駆使する専門家の役割を
も大きく変化させている。例えば創薬に関わって
いる計算化学や Bioinformatics などの分野の専門
家も、この現実に適応するには大きな発想の転換
が必要である。そうした発想の転換に参考となる
ような、変化の実相とそれへの対処について、要
点を述べてみたい。 Pathway/Network to Disease and Drug Discovery これは私が関係している CBI 学会(情報計算化
学生物学会)が今年から採択した「標語」である。
Genome-Omics-Pathway/Network からの「疾患の
理解」と「医薬品の開発」という考えは、ポスト・
ゲノム時代の時代精神になっている。実際、いま
や Pathway/Network to Disease, Toxicity, Development,・・・など、“Pathway/Network to ・・・”
は、すっかり生物医学研究の枕詞になってしまっ
た。Drug Discovery も例外ではない。 私が細胞内の信号伝達系のデータベースづく
りに関心をもったのは、1980 年代の中頃からで
あったが、その夢が実現したのは、1990 年代の
初め、WWW 技術が出現した頃である。これが
CSNDB であるが、その後、それを医薬品開発に
応用しようという試みを、当時の山之内製薬と共
同研究として行っていた。 しかし経路網データベース(知識ベース)開発
がブームとなったのは、1990 年代の終わり頃で
ある。そして「経路網のデータベースを整備する
ことが、医薬品開発につながる」という自明のこ
とが、わが国でも広く理解されるようになったの
は、2000 年以後のことである。この間にデータ
ベースの数は増え、数年前には 200 を越えている
(例えば PathGuide(http://www.pathguide.org/)を
参 照 )。 さ ら に Gene Ontology (GO 、
http://www.geneontology.org/)とともに、膨大な
Omics データの解析と解釈の基盤知識であると
みなされるようになった。そして遂に医薬品の研
究開発においても、大切なのは「標的遺伝子」で
はなく「標的経路」だという考えが露に表明され
るようになってきた2)。2006 年には、それを裏
付けるような Review 誌として、Current Signal Transduction Therapy が 創 刊 さ れ て い る
(http://www.bentham.org/cstt/index.htm)。また、NCI主 導 の Pharmacogenetics/ Pharmacogenomics Knowledge Base(http://www.pharmgkb.org/)では、
医薬品情報と経路網情報とを直接結び付ける
Graphical な知識表現が整備され始めている。明
らかに時代は動いているようだ。

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 3
Pathway/Network to Drug Discovery このようにいまや、標的遺伝子に変わって標的
経路網を基盤として創薬を考える時代となって
きたが、その本格的な展開には、当然、「経路網
からの疾患の理解」がなされていなければならな
い。このことは、「いままで臓器別に捉えられて
いた疾患の概念を経路網を基盤としたそれに変
換していく」という作業がなされていなければな
らないことを意味する。そして、このことは、医
学の知識体系の再構築という、医学史上未曾有の
大転換が起きることを意味する。このような目で
注意してみてみると、こうした大転換は実際に起
きていることがわかる。その例として Langmanの発生学3)と Weinberg4)のがんの教科書を上げ
ることができる。例えば Langman の発生学の教
科書は、我が国の医学教科書としてもよく使われ
ているが、私の手にした 2006 年に出版された(原
著)10 版の序文には、「・・・分子生物学と遺伝
学の進歩が発生の理解には欠かせなくなったの
で、最初の章で主要な signal pathway と信号分子
とを説明しておく。・・・」というような記述が
ある。Weinberg の本の序文にもこうある。「・・・
われわれはヒトの細胞の内部で働いている個々
の信号分子については、相当の知識を得てきてい
るが、それらが織り成す複雑な信号回路が、体の
なかの個々の細胞の運命を決める、生死に関わる
決定をどのように行っているかについては、よく
理解していない・・・」。その具体的記述は第6
章にあるが、もちろん、彼の本は全編、
Pathway/Network to Cancer の精神で書かれている。 よく知られているように、発生に関わる遺伝子
や経路網の発見は、ヒトよりは Dorosophila、C.elegans、Zebra fish など、詳細な遺伝的な解析
ができる簡便な多細胞モデル動物を使った研究
で行われてきた。その伝統を引き継ぐように、海
綿、ウニ、ホヤ、ミツバチなどのような、non-model後生(多細胞動物)動物のゲノムが解読されるよ
うになってくると、経路網に関与しているタンパ
ク質が保存されているか否かがしらべられるよ
うになってきた。その結果、後生動物 Metazoa で
発生に関わっている分子や経路網が保存されて
いるということがわかってきた。それらは、
Hedghog、Wnt, Receptor Tyrosin Kinase (RTK), TGF-β, Notch, Jak/STAT, Nuclear Receptor (NR)などの経路網である。これらの経路網が保存されて
いるとは、細胞外からの信号分子、受容体分子、
転写因子、転写因子(とその複合体)が結合する
DNA の(応答)配列、転写因子の標的遺伝子群
などに共通性が見られるということである。進化
と発生に関してはすでに Evo-Devo 問題と呼ばれ
る分野が意識されるようになってきた5)。ここで
中核となるのが経路網であることは容易に理解
されよう。 さらに興味深いことは、これら保存されている
経路網とがん化に関与する経路網の間に共通性
が見られることである6)。このことは、発生やが
んに関係する疾患の研究を、ヒトとは遠く離れて
いるように見えるさまざまな後生動物を材料に
して行うことも可能であることを示唆している。
実際、すでに線虫やハエで肥満やがんの研究を行
うことを売り物にしているベンチャーStart up 会
社が出現しているが、これからは、ウニやカイメ
ン、さらには(もっとも原始的な後生動物あるい
はその元祖と見られている)立襟鞭毛虫なども、
こうした疾患の解明や薬開発の材料に浮上して
くる可能性がある。 後生動物に共通する問題は、遺伝子が如何なる
状況で発現するかであるが、それはそもそも発生
に伴う細胞系譜、空間的な配置、細胞の置かれた
環境によって変化する。それを直接具現する機構
が転写である。ゆえに、この機構は、配列決定後
のゲノム学の目標であり、さまざまな生物を現存
する生物から比較する進化研究の目標であり、発
生学の目標であり、細胞分化の課題であり、おそ
らくがん組織の多様性理解のカギとなる事象で
あり、当然医薬品の研究開発でも重要な課題であ
ると思われる。 物質とエネルギー収支に関わる疾患 考えてみれば発生はヒトの成り立ちに関連し
たもっとも根源的な事象である。がんが発生と分
子レベルで深く関連していることは、この疾患が
ヒトにとって根源的ものであることを意味して
いるような気がする。それでは次に根源的な疾患
とはなんだろうか。それは生命の維持に必要な物
質の摂取とその利用のメカニズムに関わる疾患
であろう。ながらく進化を研究してきた H. J. Morwitz は、比較的最近行った NIH での講演で、
配列などは変化するが、代謝(回路)は進化の過
程に強固に保存されてきたと言っている7)。彼が
もっとも保存されていると言っているのは、TCA cycle (Krebs cycle あるいはクエン酸回路)であ
る。これは単細胞と多細胞生物に共通する代謝回

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 4
路であるが、後生動物の場合は、「生体内の化学
反応の電池のような ATP を産生するミトコンド
リ ア に お け る Oxidative Phosphorylation(OXPHOS)や NR が中核となる物質代謝系では
ないか」、というのが目下の私の作業仮説である。 前者はすでによく知られている。ただし、ATP産生の分子機構とくに、ミトコンドリアの内膜を
隔てたプロトン(H+)の濃度差を利用した分子
モータとして働く ATP 合成酵素の驚くべき機構
が解明されたのは比較的最近のことである。しか
しながら、「NR が中核となる物質代謝系」の全
容とその意義については、まだよく理解されてい
なし、広く知られてもいないように思われる。い
まのところ、NR は後生動物に固有の Superfamilyであり、すでに述べたように発生に関与する経路
網の荷い手の一部として、多細胞動物で広く保存
されていることがゲノム解読の進展で確認され
ている。私の作業仮説というのは、「NR に類縁
の転写因子として、AhR, Nrf2, SREBP, CREB、そ
しておそらくFoxa/Foxoなどを加えた転写因子群
が、後生動物での物質とそれゆえにエネルギーの
収支バランス制御の主役として働いている」ので
はないかという考えである。 疾患との関連 このような仮説を立ててみると、面白いことが
見えてくる。薬との関係で一番重要なのは、これ
らのタンパク質群が、細胞に入ってくる物質のセ
ンサー役を果たし、実際の処理を行うのは、いわ
ゆる(PhaseI, II, III と呼ばれる)薬物代謝酵素で
あるということだ。さらにこのことは、薬だけで
なく、栄養とも関係し、活性酸素除去に関わる
Nrf2 の下流にある酸化ストレス処理系という、
健康や寿命にも関係している。一方、ミトコンド
リアの働きと関係した OXPHOS は、物質という
よりエネルギー収支に関与している。そして
Metabolic Syndrome(MS)は、この双方に関連し
ているのだ。ここにはエネルギー・センサーとし
て AMPK の経路や共役因子としての PGC1も関
連してくる。したがって MS の原因遺伝子として
ミトコンドリアの機能に関わるものが疑われる
のは当然であるが、NR とその親戚のような転写
因子やそれらの共役因子である PGC1 が関与し
てくるのも、また、当然のような気がする。さら
に最近の研究で言えば、食事制限による寿命の延
び、NR と Circadian Rhythm との関連、睡眠と
MS との関連なども、この作業仮説で考えると納
得がいくような気がする。 もう一つ、MS と Inflammation の関係も興味深
い。言うまでもないが Inflammation は免疫系と関
連している。ヒトのような適応免疫 Adaptive immune system が見られるのは脊椎動物なので、
もっとも古い系は、(ヤツメウナギのような)無
顎類だと考えられているが、現在注目されている
自然免疫 Innate immune system ならハエでももっ
ている。上で述べた NR とその親戚分子は、一般
に化合物センターとして(例えば Dioxin のよう
な)環境汚染物質の異物センサーとして働くが、
これが他の生物とかその部品の侵入であると、い
わゆる免疫系が働くことなる。 MS と炎症の関係の研究者として知られる
Hotamisligil は、Drosophila の fat body が、進化的
にヒトの脂肪細胞と肝臓の双方に関連している
ことを指摘している8)。そしてこの2つの組織に
は、免疫を担う血球が侵入しやすいが、Drosophila の fat bodyはまさにこの3つを兼ね備えていると
言っている。つまり、NR を含む物質代謝系と免
疫系は、異物 Sensor として共存し、共通性があ
るのだ。このように MS を見てくると、肥満、糖
尿病、インスリン抵抗性、炎症などが、ヒトの分
子や経路網が語る進化と深く関係していること
が理解でき、また、そうした目で見れば、NR や
MS 研究が、食事、睡眠、寿命などと関連してい
ることもよく理解できる。 おわりに:情報計算の視点から 上で述べたことは、起こりつつある変化のほん
の一例に過ぎない。重要なことは、こうした変化
が情報計算技法の支援で推進されており、それを
さらに加速するには、情報計算技の専門家であり
ながら、生物医学の実験家や臨床家と自由に対話
ができ、対等な立場で(Equal Partner として)共
同研究できるような新しいタイプの研究者の存
在が必要であるということだ。このような研究者
は治験のデザインだけでなく、上市された薬の有
効性や副作用の分析調査や、上市に失敗した薬の
再生の可能性など、Follow up 研究にも活躍でき
る人材である。このような人材を意識的に育成す
ることが、製薬企業にとっても、薬学においても
重要な課題ではないかと考える。

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 5
参考文献 1) T. Kaminuma, Pathways and Networks of Nuclear Receptors and Modeling of Syndrome X, CBI Journal, 3, 130-156 (2003). 2) M. C. Fishman and J. A. Porter, A new grammar for drug discovery, Nature, 437: 491-493, 2005. 3) T. Sadler, Langman‘s Medical, Lippincott Williams & Wilkins, 2005. 4) R. A. Weinberg, The Biology of Cancer, Garland Science, 2006. 5) A. S. Wilkins, The Evolution of Developmental Pathways, Sinauer, 2002. 6) F. C. Kelleher, D. Fennelly, M. Rafferty, Common critical pathways in embryogenesis and cancer, Acta Oncologica, 45: 375-388, 2006. 7) H. J. Morwitz, Evolution and Origin of Life, April 04, 2007 (http://videocast.nih.gov/pastevents.asp?c=64). 8) G. S. Hotamisligil, Inflammation and metabolic disorders, Nature 444: 860-867, 2007.
◆略 歴◆ 神沼 二眞 (Tsuguchika KAMINUMA):1964 年、国際基督教大学教養学部自然科学科物
理学専攻、Yale 大学、Hawaii 大学の大学院を経て、1971 年日立製作所情報システム研究所研究員、1976 年
東京都臨床医学総合研究所研究室長、1989 年国立医薬品食品衛生研究所部長、2001 年退官、2001 年株式会
社バイオダイナミックス設立、2004-2006年広島大学大学院および東京医科歯科大学の人材育成計画の特任
教授。1981 年情報計算化学生物学会(CBI学会)の前身である CBI 研究会設立

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 6
薬学研究最前線(1)
シナプス可塑性に対するアミロイドβと神経ステロイドの効果 -標的分子と作用機序の解析-
曽我部 正博 1,2,3、陳玲 1,2,4(1名古屋大学大学院・医学系研究科・細胞生物物理学、 2JST・ICORP/SORST・細胞力覚、3生理学研究所・分子生理、4南京医大・神経生理)
1.はじめに
アルツハイマー病(AD)の主原因は、アミ
ロイド β ペプチド(Aβ)の過剰産生に基づく神
経傷害であることがほぼ明らかになってきた 1)。
Aβ は、一回膜貫通型蛋白質である前駆蛋白質
βAPP の、β、γ セクレターゼによる2段階の
切り出しによって産生される 2)。家族性 AD の原
因遺伝子として同定されたプレセニリンは γ セ
クレターゼの活性中心サブユニットである。こう
した現状を踏まえると AD の治療、予防の戦略と
して2つの道が考えられる。一つは Aβ の作用機
序を解析する中でその標的分子を探り、Aβ の攻
撃を阻止する道であり、もう一つはプレセニリン
の阻害を通して Aβ の過剰生産を抑制する道で
ある。後者がより根本的であるが、プレセニリン
は多機能で、生命維持に欠かせない機能を併せ持
つので、選択的に Aβ の切り出しを止めることは
難しい。そこで我々は、AD の記憶障害の背後に
予想される海馬シナプス可塑性の障害と Aβ の
関係を詳細に解析する中で、Aβ の標的分子と拮
抗剤の探索を試みた 3)。
2.AD モデルラットの調整とシナプス電気活動
のイメージング 浸透圧ポンプを用いて、7週齢の雄ウィスタ
ーラットの側脳室に 300pM/day の Aβ(1-40)を
11-14 日間慢性投与した。この Aβ 投与ラットは
海馬ニューロンの脱落などは示さないものの、空
間学習障害などの AD 様症状を呈するため、ADの初期段階モデルとして使われてきた 4)。Aβ 投
与終了後に海馬スライス(400μm 厚)を調整し
(図1a)、スチリル系膜電位感受性色素 RH482で染色した。細胞膜に分配した極性色素は、膜電
位のレベルに応じて配向度が変化し、膜電位に比
例した吸光度の変化を生じる。その変化を倒立顕
微鏡を介してダイオードカメラ(128x128 pixel; 25x25 μm2 sample are/pixel with a 4 x objective)で
高速イメージング(0.6msec/frame)して解析した
(図1b)。 a
b
b
図1.ラット海馬スライス標本(右半球水平断)の模式
図と興奮伝搬の様子。a、標本全体を電位感受性色素
(RH482 や RH155 など)で染色した後、顕微鏡下で、
Schaffer 側枝(SC)あるいは貫通線維(PP)を通電刺
激して CA1 領域 CA3/CA1 シナプス、あるいは歯状回
(DG)の PP/顆粒細胞シナプスの電位を解析する。b、
貫通線維(PP)刺激で誘発された歯状回顆粒細胞層に沿
った興奮伝搬のスナップショット。右下の数字は刺激後
の経過時間(ミリ秒)を示す。
社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 7
この手法は通常用いられる細胞外電位(field potential)記録法に比べていくつかの利点を有し
ている。まず、海馬全体の電気活動を同時記録で
きるために、Aβ 投与によってどの部位に変化が
生じたのかを特定できる。さらに pixel(画素)
がカバーする記録領域(視野)は、顕微鏡の倍率
を変えるだけで拡大可能なので、一つの神経細胞
の、樹状突起、細胞体、軸策での電位変化の違い
を特定することも容易にできる(図2左参照)。
また、シナプス部位では、シナプス前細胞とシナ
プス後細胞の電気活動が同時に記録でき、これら
を薬理学的に分離することが可能である(図2)。
特に最後の特徴が重要であり、これによって薬物
がシナプス前膜あるいは後膜の何れに作用した
のかが区別できる。これに薬理学的解析を加えれ
ば薬物の標的分子を同定することも不可能では
ない。
図2.歯状回顆粒細胞層の細胞体とシナプス領域の膜電
位変化。顆粒細胞に被っている格子の各ブロックが pixel(画素)を表す。図1b から抽出した2つの pixel の膜電
位変化で、細胞体の集合スパイク電位(PS、上段のトレ
ース)とシナプス集合電位(下段のトレース)を表す。AP5、CNQX でシナプス後膜の NMDA、AMPA 受容体をブロッ
クすると PS は消失するが、シナプス電位では EPSP 消
失後に、シナプス前膜電位(PSFV))が残り、さらに外
液 Ca2+を除去すれば終末の活動電位のみが残る。
3.シナプス可塑性に対する Aβ の効果
AD に特徴的な初期症状として短期記憶障害
が知られているが、これは記銘-記憶過程(記憶
の書き込みと保存)の障害である。この過程の背
後にある細胞メカニズムはシナプスの長期増強
(Long Term Potentiation, LTP)と呼ばれる機能で
あると信じられている。LTP は、例えば海馬 CA1領域の入力線維である CA3 から伸びる Schaffer側枝を、短時間(1秒)高頻度刺激(100Hz)する
と(図1a 参照)、CA3-CA1 シナプスの伝達効率
が長時間に(1時間以上)に亘って維持される現
象(シナプス後細胞の EPSP の増大として観測さ
れる)を指し、神経の活動(経験)依存的な記憶
のメカニズムを支える機構だという証拠がある。
そこでまず CA3-CA1 シナプスの LTP を調べたと
ころ、Aβ 投与ラットでは LTP の誘導がほぼ完全
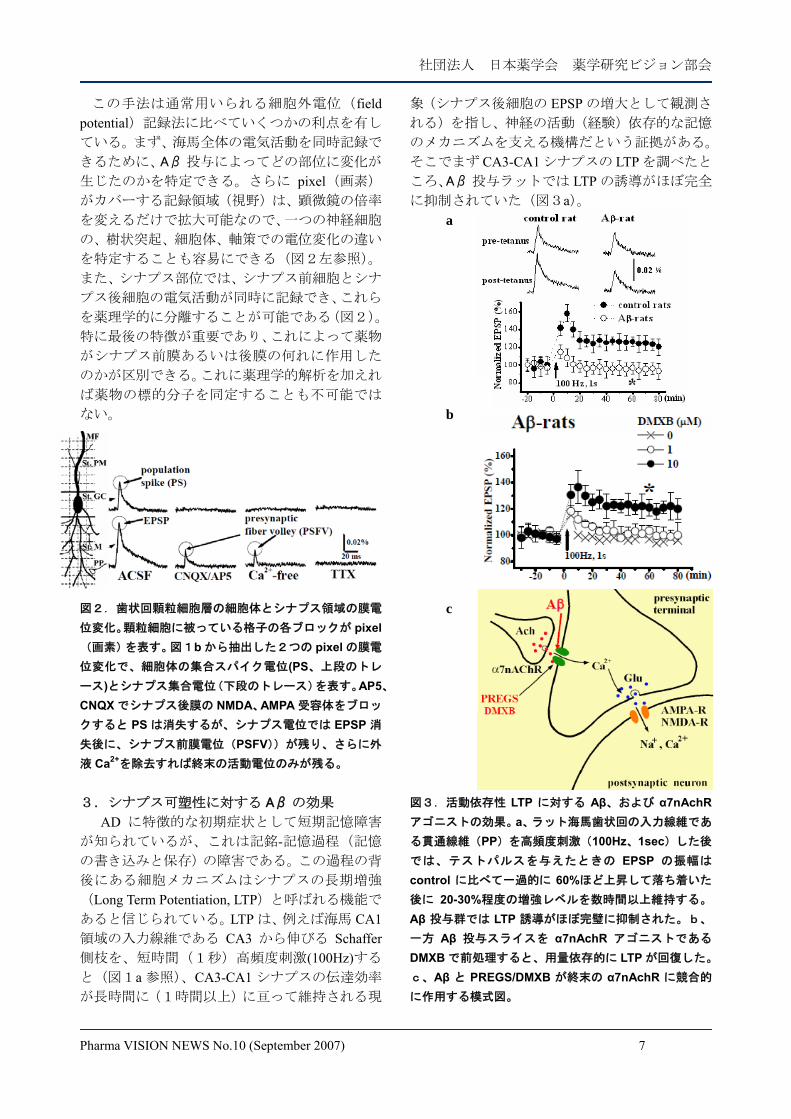
に抑制されていた(図3a)。 a b
c
図3.活動依存性 LTP に対する Aβ、および α7nAchRアゴニストの効果。a、ラット海馬歯状回の入力線維であ
る貫通線維(PP)を高頻度刺激(100Hz、1sec)した後
では、テストパルスを与えたときの EPSP の振幅は
control に比べて一過的に 60%ほど上昇して落ち着いた
後に 20-30%程度の増強レベルを数時間以上維持する。
Aβ 投与群では LTP 誘導がほぼ完璧に抑制された。b、
一方 Aβ 投与スライスを α7nAchR アゴニストである
DMXB で前処理すると、用量依存的に LTP が回復した。 c、Aβ と PREGS/DMXB が終末の α7nAchR に競合的
に作用する模式図。

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 8
4.Aβ の作用部位と標的分子 次の課題は、この LTP 誘導障害の原因がシナ
プス前終末なのか、シナプス後細胞にあるのかを
特定することである。シナプス前終末とシナプス
後細胞の活動(それぞれ PSFV (PreSynaptic Fiber Volley)、EPSP(Excitatory PostSynaptic Potential) と呼ぶ)を分離して解析したところ(図2)、AD投与ラットでは PSFV に若干の障害が見られ
EPSP は正常だったので、障害部位はシナプス前
膜と推定された。ではシナプス前膜のどの分子が
傷害されたのであろうか?これまでの知見から
アセチルコリン受容体(AchR)の可能性が考え
られた。海馬 CA3-CA1 シナプスには中隔核から
のコリン作動性支配(synapse on synapse)があり、
シナプス前終末からのグルタミン酸放出を調節
している。薬理学的解析から、Aβ の分子標的は、
AchR のサブタイプの内、α7 型ニコチン性受容体
(α7nAchR)であることが判明した(図3c)。正
常ラット海馬スライスに α7nAchR の特異的アン
タゴニスト αBTX(αBGT)を作用させると Aβ
投与とほぼ同等な障害が見られたこともこの結
論を支持している。より重要な結果は、Aβ 投与
ラットのスライス標本を α7nAchR の特異的アゴ
ニスト DMXB で処理すると、用量依存的に Aβ
によるLTP障害が改善されたことである(図3b)。ADの治療薬として著名な抗アセチルコリンエス
テラーゼ阻害剤(ドネペシル)は、進行した ADで見られる中隔核やマイネルト核のコリン作動
性ニューロンの脱落を保護するものと解釈され
ているが、本研究の結果はこの薬剤の作用機序が
必ずしもそれだけではなく初期ADのAβによる
α7nAchR の機能障害を補う可能性も示唆してい
る。
5.シナプス伝達に対する神経ステロイドの効果 AD が老化に伴う自然現象の一つであるとす
れば、逆に生体にはこれを防御する内在的仕掛け
があっても不思議ではない。その有力な候補とし
て近年中枢で合成されることが明らかになって
きた種々の神経ステロイドがある。エストラジオ
ールやテストステロンなど末梢器官で合成され
るほとんどのステロイドホルモンは脳内でも合
成されており、様々な生理活性を有することが明
らかになりつつある。とりわけ、認知機能の向上
や神経細胞死の保護など、医学的に極めて興味深
い作用が報告されている。例えば代表的神経ステ
ロイドである硫酸プレグレノロン(PREGS)の脳
内含量の低下と老化ラットの認知障害との相関
が示唆されており 5)、逆にこの障害は PREGS の
海馬注入で改善されることが知られている 6)。最
近我々は PREGS の急性投与自体がシナプス伝達
の効率を高め、顕著な chemLTP(短時間高頻度電
気刺激で誘導される一般的な活動依存的 LTP と
区別して、chemLTP と表記する)を引き起こすこ
と 7)、また活動依存性 LTP 誘導における刺激周波
数の閾値を下げることを見いだした。以下神経ス
テロイド PREGS の作用機序と標的分子について
概説する。 a b
図4.海馬歯状回の興奮(EPSP)に対する神経ステロ
イド PREGS の増強効果。 a、貫通線維刺激後 7.2 ミリ秒での興奮伝搬のスナップ
ショット。b、左図のシナプス領域での EPSP の比較(上)
と EPSP 増強の PREGS 濃度依存性(下)。 6.神経ステロイドの作用部位と標的分子
2節で紹介した手法を使い、海馬シナプス伝
達に及ぼす PREGS の急性効果を調べた。海馬ス
ライスの灌流液に 20μMPREGS を添加して急性
投与(10 分間)すると、歯状回顆粒細胞(CA1錐体細胞でも同様)の EPSP が 20%増強した(図
4)。この急性増強は、細胞外 Ca2+依存的、かつ
NMDA 受容体非依存的であり、増強後には、シ
ナプス終末の性質を表す PPF ( paired-pulse facilitation)の性質が変化したことから、PREGSの作用点はシナプス前膜であると予想された。そ
の最も単純な機序としては、シナプス終末からの
興奮性伝達物質グルタミン酸の放出量の増強が
考えられる。ではこの仮説を証明するにはどうし
たらよいだろうか?シナプス近傍でのグルタミ

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 9
ン酸量を高時間分解能(ミリ秒)で測定すること
は極めて難しいが、膜電位イメージング法を使う
と、容易にこれを実現することができる。 シナプスの周囲は一般的にグリア細胞(アス
トロサイト)で囲まれており、放出されたグルタ
ミン酸の大半はアストロサイトに発現するグル
タミン酸輸送体(GLT-1)で速やかに吸収される。
GLT-1 はグルタミン酸1分子当たり 2-3 個の Na+
をカップルして吸収する起電性(electrogenic)の
2次性能動輸送体なので、吸収したグルタミン酸
の量に比例した脱分極応答を生じる。先に述べた
電位感受性色素 RH482 は神経細胞に選択的でグ
リア細胞を染色しないが、メロシアニン系色素の
RH155 は両者を染色する。そこでこの色素でス
ライス標本を染色し、シナプス後細胞の NMDA型と AMPA 型のグルタミン受容体をそれぞれ
AP5 と CNQX で抑制して EPSP の発生を抑えれ
ば、グリア由来の脱分極(SIGD、synaptically induced glial depolarization)を記録することがで
きる(図5a)8)。これがグリア細胞 GLT-1 由来
の脱分極であることは、その特異的抑制剤である
DHK によって SIGD が完全に抑制されることで
確認された(図5b)。さて PREGS を添加すると
濃度依存的に SIGD が増加し、その増加は
α7nAchR の特異的阻害剤である αBGT あるいは
MLA でほぼ完全に抑制された(図5c)。さらに
α7nAchR の活性剤である DMXB は PREGS と同
様に SIGD を増強する一方で、PREGS の標的と
して可能性のある GABAA受容体、シグマ受容体、
α4nAch 受容体に対する阻害剤は全く影響がなか
った。したがって、PREGS による EPSP の増強
はシナプス前終末の α7nAchR に対する特異的感
作によると結論された(正確にはその下流で L型 Ca2+チャネルが関与している)7)。現在 PREGSがAβのα7nAchRに対する障害効果を抑制できる
か否かを検討中であるが、予備的には顕著な効果
が認められている。もし内因性の生理活性物質で
Aβ の作用を抑えることができれば、より安全な
新しい AD の治療・予防戦略の道が開ける可能性
がある。
a
b
c
図5.グリア細胞のグルタミン酸吸収に伴う膜電位変化
(SIGD)の解析。a、スライスを RH155 で染色して、
CNQX/AP5 で EPSP を除去すると SIGD が抽出できる。
b、SIGD は PREGS で増強し、グルタミン酸トランス
ポーター(GLT-1)の阻害剤 DHK で完全に消失する。c、
PREGS による SIGD の増強分(斜線部)は α7nAchR ア
ンタゴニストである α-BGT で完全に消失する。
社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 10
a
b
図6.PREGS の多様な作用。a、PREGS による chemLTP の
誘導(●)。α7nAchR アンタゴニストの添加で緩やかに
立ち上がる成分(○)のみが残る。b、逆に NMDA チャ
ネル阻害剤の添加でこの遅い成分が消失して、α7nAchR依存的な過渡成分(STP)のみが残る。これは a におけ
る(●-○)にほぼ匹敵する(内挿図)。PREGS はシナ
プス前膜の α7nAchRとシナプス後膜の NMDA受容体に
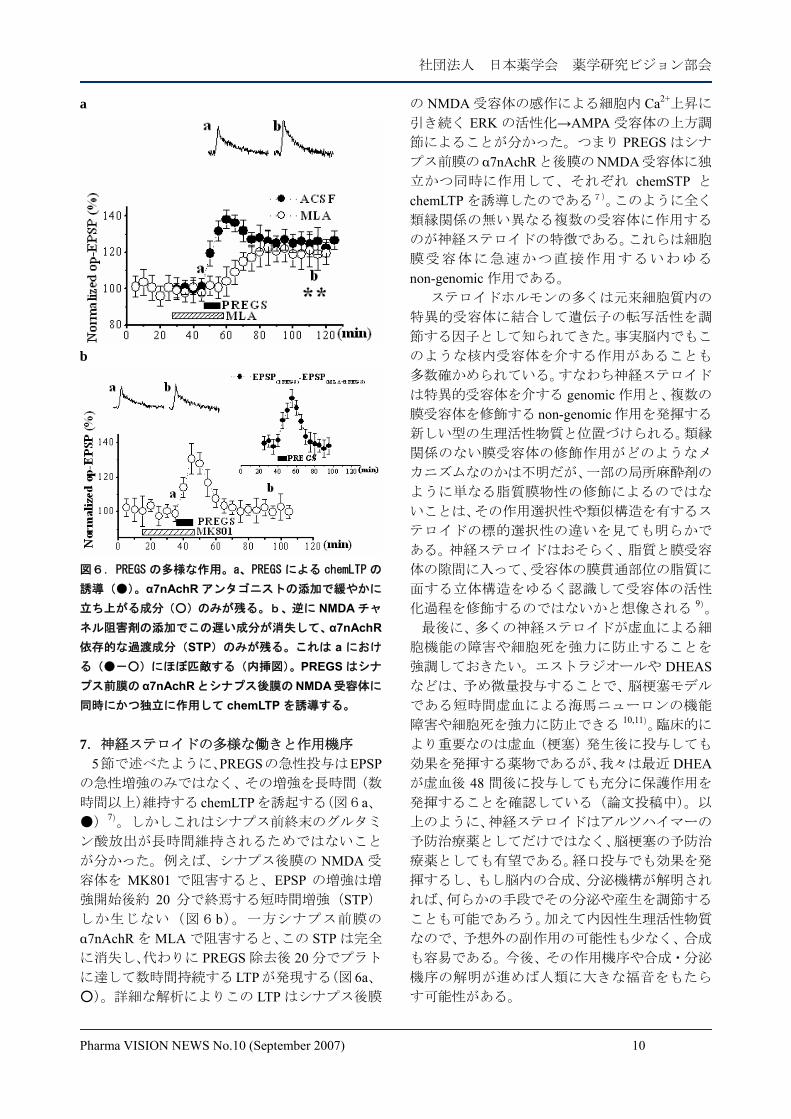
同時にかつ独立に作用して chemLTP を誘導する。 7.神経ステロイドの多様な働きと作用機序
5節で述べたように、PREGSの急性投与はEPSPの急性増強のみではなく、その増強を長時間(数
時間以上)維持する chemLTPを誘起する(図6a、●)7)。しかしこれはシナプス前終末のグルタミ
ン酸放出が長時間維持されるためではないこと
が分かった。例えば、シナプス後膜の NMDA 受
容体を MK801 で阻害すると、EPSP の増強は増
強開始後約 20 分で終焉する短時間増強(STP)しか生じない(図6b)。一方シナプス前膜の
α7nAchR を MLA で阻害すると、この STP は完全
に消失し、代わりに PREGS 除去後 20 分でプラト
に達して数時間持続するLTPが発現する(図 6a、○)。詳細な解析によりこの LTP はシナプス後膜
の NMDA 受容体の感作による細胞内 Ca2+上昇に
引き続く ERK の活性化→AMPA 受容体の上方調
節によることが分かった。つまり PREGS はシナ
プス前膜の α7nAchRと後膜のNMDA受容体に独
立かつ同時に作用して、それぞれ chemSTP と
chemLTP を誘導したのである7)。このように全く
類縁関係の無い異なる複数の受容体に作用する
のが神経ステロイドの特徴である。これらは細胞
膜受容体に急速かつ直接作用するいわゆる
non-genomic 作用である。 ステロイドホルモンの多くは元来細胞質内の
特異的受容体に結合して遺伝子の転写活性を調
節する因子として知られてきた。事実脳内でもこ
のような核内受容体を介する作用があることも
多数確かめられている。すなわち神経ステロイド
は特異的受容体を介する genomic 作用と、複数の
膜受容体を修飾する non-genomic作用を発揮する
新しい型の生理活性物質と位置づけられる。類縁
関係のない膜受容体の修飾作用がどのようなメ
カニズムなのかは不明だが、一部の局所麻酔剤の
ように単なる脂質膜物性の修飾によるのではな
いことは、その作用選択性や類似構造を有するス
テロイドの標的選択性の違いを見ても明らかで
ある。神経ステロイドはおそらく、脂質と膜受容
体の隙間に入って、受容体の膜貫通部位の脂質に
面する立体構造をゆるく認識して受容体の活性
化過程を修飾するのではないかと想像される 9)。 最後に、多くの神経ステロイドが虚血による細
胞機能の障害や細胞死を強力に防止することを
強調しておきたい。エストラジオールや DHEASなどは、予め微量投与することで、脳梗塞モデル
である短時間虚血による海馬ニューロンの機能
障害や細胞死を強力に防止できる 10,11)。臨床的に
より重要なのは虚血(梗塞)発生後に投与しても
効果を発揮する薬物であるが、我々は最近 DHEAが虚血後 48 間後に投与しても充分に保護作用を
発揮することを確認している(論文投稿中)。以
上のように、神経ステロイドはアルツハイマーの
予防治療薬としてだけではなく、脳梗塞の予防治
療薬としても有望である。経口投与でも効果を発
揮するし、もし脳内の合成、分泌機構が解明され
れば、何らかの手段でその分泌や産生を調節する
ことも可能であろう。加えて内因性生理活性物質
なので、予想外の副作用の可能性も少なく、合成
も容易である。今後、その作用機序や合成・分泌
機序の解明が進めば人類に大きな福音をもたら
す可能性がある。

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 11
参考文献 1) Forman MS, Trojanowski JQ, Lee VM: Neurodegenerative diseases: a decade of discoveries paves the way for therapeutic breakthroughs. Nat. Med 10: 1055-1063, 2004 2) Koo EH, Kopan R. Potential role of presenilin-regulated signaling pathways in sporadic neurodegeneration. Nat Med 10: S26-33, 2004 3) Chen L, Yamada K, Nabeshima T, Sokabe M: α7 nicotinic acetylcholine receptor as a target to rescue deficit in hippocampal LTP induction in β-amyloid infused rats. Neuropharmacol. 50(2):254-268, 2006 4) Nitta A, Itoh A, Hasegawa T, Nabeshima T:β-amyloid protein-induced Alzheimer’s disease animal model. Neurosci. Lett. 170: 63-66,1994 5) Weill-Engerer S, David JP, Sazdovitch V, Liere P, Eychenne B, Pianos A, Schumacher M, Delacourte A, Baulieu EE, Akwa Y: Neurosteroid quantification in human brain regions: comparison between Alzheimer’s and nondemented patients. J Clin Endocrinol Metabol. 87:5138-5143, 2002. 6) Flood JF, Morley JE, Roberts E: Pregnenolone sulfate enhances post-training memory processes when injected in very low doses into limbic system structures: the amygdala is by far the most sensitive. PNAS 92:10806-10810, 1995. 7) Chen L, Miyamoto Y, Furuya K, Mori N, Sokabe M: PREGS-induces long-lasing potentiation in the hippocampal dentate gurus of adult rats via NR2B tyrosine phosphorylation that leads to ERK-CREB signaling. J Neuropysiol.(in press) 8) Chen L, Sokabe M:Pregnenolone sulfate enhances presynaptic glutamate releases in rat hippocampal slices as studied by optical recordings. J Neurophysiol. 94(6): 4131-4144, 2005 9) Jiang MK, Mierke DF, Russek SJ, Farb DH: A steroid modulatory domain on NR2B controls N-methyl-D-aspartate receptor proton sensitivity. PNAS 101:8198-8203, 2004 10) Dai S, Chen L, Sokabe M: Neurosteroid Estradiol Rescues Ischemia-Induced Deficit in the Long-Term Potentiation in rat hippocampal CA1 neurons. Neuropharmacol, 52:1124-1138, 2007 11) Li Z, Zou1 R, Cui1 S, Xei1 G, Cai1 W, Sokabe M, Chen L: DHEAS prevents LTP impairment following forebrain ischemia in rat hippocampal CA1 by regulating tyrosine phosphorylation of NMDA receptor. Neuropharmacol, 51: 958-966, 2006
◆略 歴◆ 曽我部 正博 (Masahiro SOKABE):1975年大阪大基礎工・博士課程中退、大阪大人間
科学・助手、1985年名古屋大医・講師、1987年同・助教授、1992年同・教授、2000年JST・ICORP細胞力覚
プロジェクト代表(兼)、2003年生理学研究所・教授(併)、2005年JST・ICORP/SORST細胞力覚プロジェク
ト総括(兼) 陳玲(Ling Chen):1995年筑波大医・博士課程終了、工技院生技研・研究員、1999年名古屋大医・研究員、
2004年南京医大・教授

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 12
薬学研究最前線(2)
ヒストン脱アセチル化酵素を分子標的とした抗がん剤の探索研究
鈴木 孝禎、伊藤 幸裕、宮田 直樹(名古屋市立大学大学院薬学研究科)
1.はじめに 最近の分子生物学研究の急速な進歩により、癌
細胞の増殖あるいは悪性化の分子機構が明らか
にされつつある。癌細胞の増殖に関与する分子の
機能を制御することにより癌治療を行うという
新しいコンセプトが癌の分子標的治療である。癌
の分子標的治療薬は、癌細胞の異常増殖の原因と
なる分子あるいはそのシグナル伝達経路など、癌
の原因により近い分子を標的とするため、従来の
治療薬に比べ副作用が少ないことが期待される。
最近、癌治療における標的分子の一つとして注目
されているのがヒストン脱アセチル化酵素
(HDAC)である 1)。本稿では、新たな癌の分子
標的治療薬の開発を目的として、我々がこれまで
に行った HDAC 阻害薬の探索研究を紹介する。
2. HDAC の機能、構造と触媒メカニズム HDAC は、ヒストンのアセチル化されたリシン
残基からアセチル基を除去する反応を触媒し、遺
伝子発現を調節する重要な酵素である。1999 年
に報告されたヒト HDAC ホモログである HDLP(HDAC-like protein)の結晶構造、2004 年に発表
されたヒト HDAC のアイソフォームの一つであ
る HDAC8 の結晶構造から、HDAC の構造につい
てはある程度の知見が得られている。基質結合部
位の内壁は疎水性残基によって形成され、酵素活
性中心には亜鉛イオンが存在しており、それに水
分子が配位していることが明らかになっている。
また、この結晶構造から HDAC によるヒストン
脱アセチル化のメカニズムは以下のように推定
された。まず、亜鉛及び His140(HDAC1 の番号)
により活性化された水分子が亜鉛に配位したヒ
ストンのアセチル化されたリシン残基のカルボ
ニル基を求核攻撃する。カルボニルの炭素は四面
体構造をとり、最終的に C-N 結合の開裂が起き
て、Asp191-His141 から電荷移動によりプロトン
を受け取り、脱アセチル化が完結する(図 1)。
Zn2+
D264 D176H178
NHO
OHH:
Y303
OH
N
N
H140
O
OD174H
HN
NH141
O
OD191
H
Zn2+
D264 D176H178
NO
Y303
O
HN
N
H140
O
OD174H
N
HN
H141
O
OD191
OH
H
Zn2+
D264 D176H178
NH3
Y303
OH
HN
N
H140
O
OD174H
N
NH141
O
OD191
H
H
O
O
H :
図 1. HDLP の結晶構造より推定された HDAC による脱
アセチル化のメカニズム
3. HDAC 阻害薬 HDAC を阻害すると相対的にヒストンアセチ
ル化酵素(HAT)の働きが強くなり、ヒストンは
過剰にアセチル化された状態となる。このヒスト
ンハイパーアセチレーションにより、がん抑制に
関与する多くの遺伝子の転写活性化が起こる。実
際に HDAC 阻害薬は、臨床試験において抗がん
活性を示すことが明らかにされている。 これまでに報告された HDAC 阻害薬のほとん
どはトリコスタチン A(TSA)やスベロイルアニ
リドヒドロキサム酸(SAHA)(図 2)に代表され
るようなヒドロキサム酸系化合物であり、ヒドロ
キサム酸が酵素活性中心にある亜鉛に配位し、基
質であるアセチル化されたリシンに拮抗して阻
害活性を示すと考えられている。しかしながら、
一般にヒドロキサム酸を zinc-binding group(ZBG)として持つ化合物は体内動態が悪く、毒
性の懸念も多い。それ故に、より体内動態の改善
した抗癌剤、より副作用の少ない抗癌剤になり得
る非ヒドロキサム酸系 HDAC 阻害薬の開発が望
まれている 2)。
Me2N
O O
NH
OH
TSA
HN
O
O
NH
OH
SAHA
図 2. TSA と SAHA の構造
社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 13
4. 非ヒドロキサム酸系 HDAC 阻害薬の創製 3) 我々が本研究を開始するまでに、 ο-アミノア
ニリド 1 やトリフルオロメチルケトン 2、N-ホル
ミルヒドロキシアミン 3 などの低分子化合物が
HDAC 阻害活性を示すことが報告されていたが
(表 1)、それらの化合物は活性面でヒドロキサ
ム酸系 HDAC 阻害薬を凌駕するものではなかっ
た。このような背景を踏まえた上で、我々は、新
たな癌の分子標的治療薬の開発を目的とし、非ヒ
ドロキサム酸系 HDAC 阻害薬の創製を行った。 我々は、以下の 4 つの概念により表 1 に示すよ
うな非ヒドロキサム酸系 HDAC 阻害薬の設計を
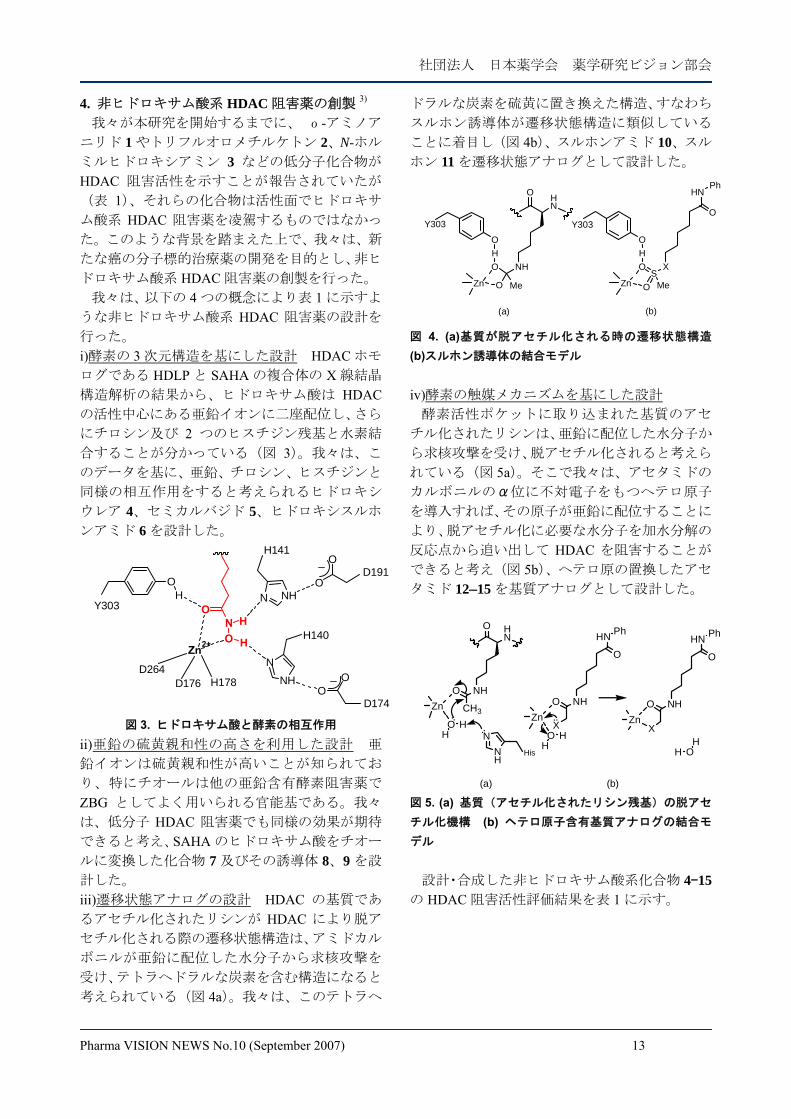
行った。 i)酵素の 3 次元構造を基にした設計 HDAC ホモ
ログである HDLP と SAHA の複合体の X 線結晶
構造解析の結果から、ヒドロキサム酸は HDACの活性中心にある亜鉛イオンに二座配位し、さら
にチロシン及び 2 つのヒスチジン残基と水素結
合することが分かっている(図 3)。我々は、こ
のデータを基に、亜鉛、チロシン、ヒスチジンと
同様の相互作用をすると考えられるヒドロキシ
ウレア 4、セミカルバジド 5、ヒドロキシスルホ
ンアミド 6 を設計した。
NO
O
NHN
Zn
D264D176 H178
OO
D174
N NH
H141
O
OD191
H140
O
2+ H
H
HY303
図 3. ヒドロキサム酸と酵素の相互作用 ii)亜鉛の硫黄親和性の高さを利用した設計 亜
鉛イオンは硫黄親和性が高いことが知られてお
り、特にチオールは他の亜鉛含有酵素阻害薬で
ZBG としてよく用いられる官能基である。我々
は、低分子 HDAC 阻害薬でも同様の効果が期待
できると考え、SAHA のヒドロキサム酸をチオー
ルに変換した化合物 7 及びその誘導体 8、9 を設
計した。 iii)遷移状態アナログの設計 HDAC の基質であ
るアセチル化されたリシンが HDAC により脱ア
セチル化される際の遷移状態構造は、アミドカル
ボニルが亜鉛に配位した水分子から求核攻撃を
受け、テトラヘドラルな炭素を含む構造になると
考えられている(図 4a)。我々は、このテトラヘ
ドラルな炭素を硫黄に置き換えた構造、すなわち
スルホン誘導体が遷移状態構造に類似している
ことに着目し(図 4b)、スルホンアミド 10、スル
ホン 11 を遷移状態アナログとして設計した。
O HN
NH
O
O
Zn
HO
(a)
XS
O
O
Zn
HO
(b)
O
HNPh
Me Me
Y303 Y303
図 4. (a)基質が脱アセチル化される時の遷移状態構造
(b)スルホン誘導体の結合モデル iv)酵素の触媒メカニズムを基にした設計 酵素活性ポケットに取り込まれた基質のアセ
チル化されたリシンは、亜鉛に配位した水分子か
ら求核攻撃を受け、脱アセチル化されると考えら
れている(図 5a)。そこで我々は、アセタミドの
カルボニルのα位に不対電子をもつヘテロ原子
を導入すれば、その原子が亜鉛に配位することに
より、脱アセチル化に必要な水分子を加水分解の
反応点から追い出して HDAC を阻害することが
できると考え(図 5b)、ヘテロ原の置換したアセ
タミド 12–15 を基質アナログとして設計した。
O HN
NH
CH3
OZn
O HH
:
His
NNH
:
(a)
NHOZn
O HH
(b)
X
:
O
HN Ph
NHOZn
O
X
O
HN Ph
HH
図 5. (a) 基質(アセチル化されたリシン残基)の脱アセ
チル化機構 (b) ヘテロ原子含有基質アナログの結合モ
デル 設計・合成した非ヒドロキサム酸系化合物 4–15
の HDAC 阻害活性評価結果を表 1 に示す。
社団法人 日本薬学会 薬学研究ビジョン部会
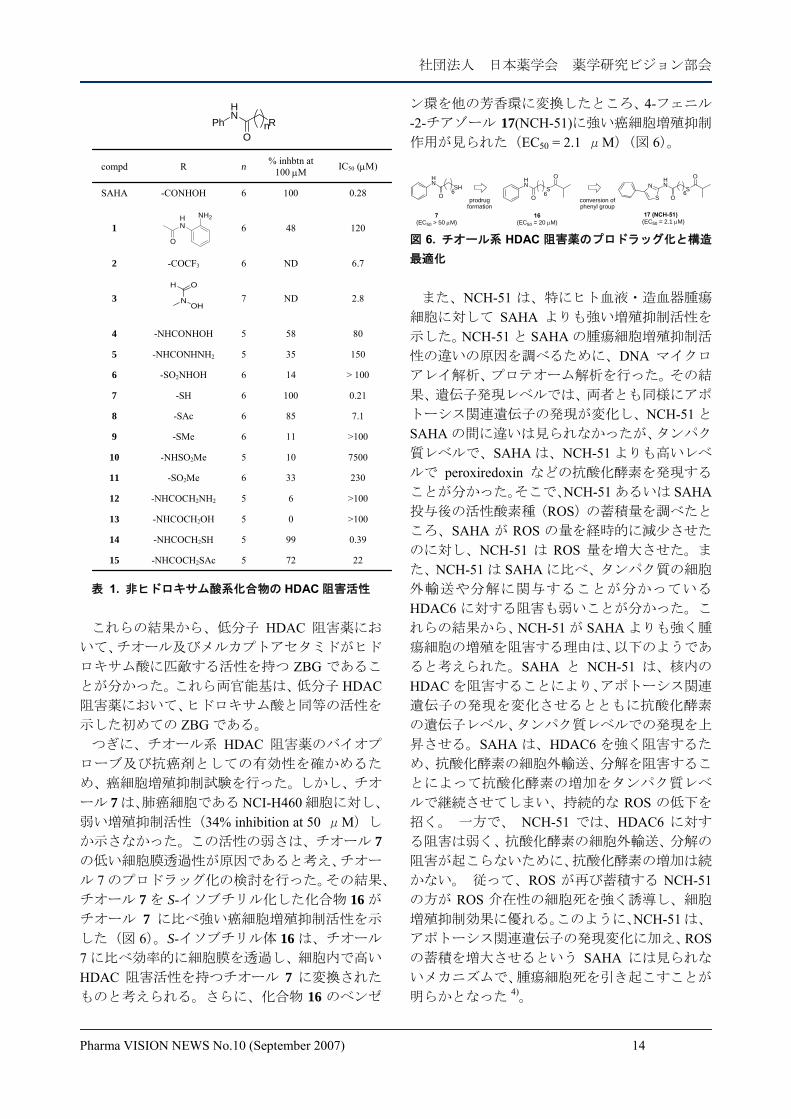
Pharma VISION NEWS No.10 (September 2007) 14
HN
NH2
O
N
OH
OH
PhHN
ORn
22725-NHCOCH2SAc15
0.39995-NHCOCH2SH14
>10005-NHCOCH2OH13
>10065-NHCOCH2NH212
230336-SO2Me11
7500105-NHSO2Me10
>100116-SMe9
7.1856-SAc8
0.211006-SH7
> 100146-SO2NHOH6
150355-NHCONHNH25
80585-NHCONHOH4
2.8ND73
6.7ND6-COCF32
1204861
0.281006-CONHOHSAHA
IC50 (μM)% inhbtn at 100 μMnRcompd
表 1. 非ヒドロキサム酸系化合物の HDAC 阻害活性 これらの結果から、低分子 HDAC 阻害薬にお
いて、チオール及びメルカプトアセタミドがヒド
ロキサム酸に匹敵する活性を持つ ZBG であるこ
とが分かった。これら両官能基は、低分子 HDAC阻害薬において、ヒドロキサム酸と同等の活性を
示した初めての ZBG である。 つぎに、チオール系 HDAC 阻害薬のバイオプ
ローブ及び抗癌剤としての有効性を確かめるた
め、癌細胞増殖抑制試験を行った。しかし、チオ
ール 7 は、肺癌細胞である NCI-H460 細胞に対し、
弱い増殖抑制活性(34% inhibition at 50 μM)し
か示さなかった。この活性の弱さは、チオール 7の低い細胞膜透過性が原因であると考え、チオー
ル 7 のプロドラッグ化の検討を行った。その結果、
チオール 7 を S-イソブチリル化した化合物 16 が
チオール 7 に比べ強い癌細胞増殖抑制活性を示
した(図 6)。S-イソブチリル体 16 は、チオール
7 に比べ効率的に細胞膜を透過し、細胞内で高い
HDAC 阻害活性を持つチオール 7 に変換された
ものと考えられる。さらに、化合物 16 のベンゼ
ン環を他の芳香環に変換したところ、4-フェニル
-2-チアゾール 17(NCH-51)に強い癌細胞増殖抑制
作用が見られた(EC50 = 2.1 μM)(図 6)。
SHHN
O6
prodrugformation
HN
O 6
7(EC50 > 50 μM)
conversion of phenyl group
6
HN
S
17 (NCH-51) (EC50 = 2.1 μM)
S
O
16(EC50 = 20 μM)
O
O
N
S
図 6. チオール系 HDAC 阻害薬のプロドラッグ化と構造
最適化 また、NCH-51 は、特にヒト血液・造血器腫瘍
細胞に対して SAHA よりも強い増殖抑制活性を
示した。NCH-51 と SAHA の腫瘍細胞増殖抑制活
性の違いの原因を調べるために、DNA マイクロ
アレイ解析、プロテオーム解析を行った。その結
果、遺伝子発現レベルでは、両者とも同様にアポ
トーシス関連遺伝子の発現が変化し、NCH-51 と
SAHA の間に違いは見られなかったが、タンパク
質レベルで、SAHA は、NCH-51 よりも高いレベ
ルで peroxiredoxin などの抗酸化酵素を発現する
ことが分かった。そこで、NCH-51 あるいは SAHA投与後の活性酸素種(ROS)の蓄積量を調べたと
ころ、SAHA が ROS の量を経時的に減少させた
のに対し、NCH-51 は ROS 量を増大させた。ま
た、NCH-51 は SAHA に比べ、タンパク質の細胞
外輸送や分解に関与することが分かっている
HDAC6 に対する阻害も弱いことが分かった。こ
れらの結果から、NCH-51 が SAHA よりも強く腫
瘍細胞の増殖を阻害する理由は、以下のようであ
ると考えられた。SAHA と NCH-51 は、核内の
HDAC を阻害することにより、アポトーシス関連
遺伝子の発現を変化させるとともに抗酸化酵素
の遺伝子レベル、タンパク質レベルでの発現を上
昇させる。SAHA は、HDAC6 を強く阻害するた
め、抗酸化酵素の細胞外輸送、分解を阻害するこ
とによって抗酸化酵素の増加をタンパク質レベ
ルで継続させてしまい、持続的な ROS の低下を
招く。 一方で、 NCH-51 では、HDAC6 に対す
る阻害は弱く、抗酸化酵素の細胞外輸送、分解の
阻害が起こらないために、抗酸化酵素の増加は続
かない。 従って、ROS が再び蓄積する NCH-51の方が ROS 介在性の細胞死を強く誘導し、細胞
増殖抑制効果に優れる。このように、NCH-51 は、
アポトーシス関連遺伝子の発現変化に加え、ROSの蓄積を増大させるという SAHA には見られな
いメカニズムで、腫瘍細胞死を引き起こすことが
明らかとなった 4)。

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 15
5. アイソフォーム選択的 HDAC 阻害薬の創製 5) 現在までに HDAC には、18 種類のアイソフォ
ームが知られているが、各々の詳細な機能には不
明な点も多い。アイソフォーム選択的 HDAC 阻
害薬は、各アイソフォームのより詳しい機能の解
明に有用であり、副作用の少ない治療薬としても
期待できる。HDAC のアイソフォームのひとつで
ある HDAC6 は、細胞質に発現しており、α-チューブリン やHSP90の脱アセチル化を行うこと
が知られている。一方、HDAC6 の阻害は、正常
細胞には影響を与えることなく、多発性骨髄腫細
胞の細胞死を誘導することが報告されているこ
とから、HDAC6 選択的阻害薬は、新規の抗がん
剤として期待できる。そこで我々は、HDAC6 選
択的阻害薬の創製を目指し、本研究に着手した。 我々は、化合物 18(図 7)が HDAC6 に選択的
に脱アセチル化されるという報告をもとに、18の N-アセチル部分をチオールに変換した化合物
19 を設計した(図 7)。化合物 18 が HDAC6 選択
的な基質であることから、18 の N-Boc 及びトリ
フルオロメチルクマリルアミド構造は、HDAC6が選択的に認識する構造であると考えられ、その
チオール誘導体 19 は、HDAC6 を選択的に阻害す
ることが期待された(図 7)。
NHBoc
O
HNOO
CF3
NH
O
CH3
18HDAC6選択的な基質
NHBoc
O
HNOO
CF3
SH
19aHDAC6選択的阻害薬 ?
HDAC6に認識される構造
図 7. HDAC6 選択的阻害薬の設計
HDAC6 は、α-チューブリンの脱アセチル化を
行うことが報告されている。したがって、アセチ
ル化α-チューブリンとアセチル化ヒストンを検
出することで間接的に阻害薬のHDAC6選択性を
評価できる。そこで評価方法としてウエスタンブ
ロッティング法を用い、HCT116 細胞にクマリン
体 19 の S-イソブチリルプロドラッグ体を投与後
8時間におけるアセチル化ヒストンとアセチル化
α-チューブリンを検出した。その結果、クマリ
ン体は、NCH-51 に比べると、アセチル化α-チュ
ーブリンがアセチル化ヒストンよりも多く検出
され、わずかではあるが HDAC6 選択的な阻害を
示した。さらに、クマリン環を他の置換基に変換
したところ、かさ高いアルキル基を持つ 20–22
(図 8)で、α-チューブリンが選択的にアセチル
化された。これは、化合物 20–22 が選択的に
HDAC6 を阻害していることを示している。これ
らの化合物は、酵素阻害活性評価においても、
HDAC6 を選択的に阻害し、HDAC1、HDAC4 に
対するその選択性はおよそ 40 倍であった。これ
らの結果から、HDAC6 選択的な阻害には、かさ
高いアルキル基が必要構造であることが明らか
にされた。現在、HDAC6 選択的阻害薬の抗がん
剤としての可能性を探っているところである。
HN
O
HN
RSH
20 : R = cyclopentyl21 : R = cyclohexyl22 : R = cycloheptyl
O
O
図 8. HDAC6 選択的阻害薬
6.おわりに 本稿で紹介したように、我々は、非ヒドロキサ
ム酸系 HDAC 阻害薬の探索研究から高活性な腫
瘍細胞増殖抑制化合物 NCH-51 を見出した。また、
NCH-51 と SAHA を用いた生化学的解析により、
HDAC 阻害薬の作用機序に関して、新たな知見を
得た。また、更なる構造展開により、HDAC6 選
択的な阻害薬を見出すことにも成功した。 現在までに数多くの HDAC 阻害薬が知られて
いるが、我々が本稿で示したようなアイソフォー
ム選択的阻害薬は、数少ない。2004 年に、ヒト
HDACアイソフォームのひとつであるHDAC8の結晶構造が発表され、2007 年には、HDAC7 の結
晶構造が公開された。今後、これらの三次元結晶
構造を基に、アイソフォーム選択的 HDAC 阻害
薬の探索研究が加速化されるものと思われる。さ
らに、それらのアイソフォーム選択的阻害薬によ
り、これまでに知られていなかった各々のアイソ
フォームの機能が明らかになるとともに、副作用
の少ない新しいがん治療への道が開かれること が期待される。

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 16
参考文献 1) Suzuki, T.; Miyata, N. Curr. Med. Chem. 2006, 13, 935–958. 2) Suzuki, T.; Miyata, N. Curr. Med. Chem. 2005, 12, 2867–2880. 3) Suzuki, T. et al. J. Med. Chem. 2005, 48, 1019–1032. 4) Sanda, T. et al. Leukemia 2007, 21, in press. 5) Suzuki, T. et al. J. Med. Chem. 2006, 49, 4809–4812.
◆略 歴◆ 鈴木 孝禎 (Takayoshi SUZUKI):名古屋市立大学大学院薬学研究科・助教
1997 年東京大学大学院薬学系研究科修士課程修了。2005 年薬学博士。1998 年日本たばこ産業(株)医薬総
合研究所入社、2003 年名古屋市立大学大学院薬学研究科・助手。2007 年より現職。 伊藤 幸裕(Yukihiro ITOH):名古屋市立大学大学院薬学研究科・博士前期課程2年 2006年名古屋市立大学薬学部卒業。 宮田 直樹(Naoki MIYATA):名古屋市立大学大学院薬学研究科・教授 1976年東京大学大学院薬学系研究科博士課程修了、1976-82年東京大学薬学部助手、1982年より厚生労働省
国立医薬品食品衛生研究所室長、1991年より同所有機化学部長、2001年より現職。

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 17
薬学研究最前線(3)
タンパク質構造変化の新規計測法の開発
廣田 俊(奈良先端科学技術大学院大学 物質創成科学研究科/科学技術振興機構さきがけ)
1.はじめに 創薬ターゲット分子であるタンパク質の立体
構造やダイナミクス、医薬品との相互作用の解明
は、創薬にとって最も重要な課題の一つである。
タンパク質分子の構造とダイナミクスの解明が
進むにつれ、従来の鍵と鍵穴モデルにかわる、よ
り現実を反映した分子構造やダイナミクス、分子
間相互作用機構の解明が課題となっている。そこ
で筆者らは、タンパク質やその複合体の構造変化、
ダイナミクスを研究するために、これまでの研究
戦略とは異なる新しい計測技術の構築を目指し
ている。 私達の体内では、様々なタンパク質が複雑に機
能しており、タンパク質が機能するためには特定
の高次構造をもつ折れ畳んだ状態を形成するこ
とが必須である。また、プリオンタンパク質やア
ミロイドβタンパク質などのタンパク質は、ある
きっかけで連鎖的に毒性を持った構造へと変化
する。そこで、実験と理論の両面からタンパク質
の構造形成に関する研究が盛んになされており、
ストップトフロー法などにより比較的遅い時間
領域でのタンパク質形成反応の理解が深まった。
しかし、ストップトフロー法では約 1 ms 程度の
不感時間が生じ、タンパク質の構造形成にはいま
だ不明な点も多い。このため、1 ms よりも早い
初期段階での構造形成反応を追跡する良い手法
が必要である。本稿では、まず最初に、種々のタ
ンパク質に広く応用できる方法として、光解離性
修飾基をタンパク質の折れ畳み反応の追跡に利
用する方法を本稿の前半で紹介する。 タンパク質は他のタンパク質との共存下で特
定の反応を行う。よって、タンパク質―タンパク
質相互作用、この相互作用によって引き起こされ
るタンパク質分子の構造変化は、タンパク質の構
造―機能相関を理解するのに重要である。しかし
ながら、有効な測定手法が限られていることもあ
って、他分子との相互作用によって引き起こされ
るタンパク質の構造変化の研究はあまり進んで
いない。そこで、筆者らが行ってきた、モデルオ
リゴペプチドを用いてタンパク質―タンパク質
相互作用、この相互作用によって引き起こされる
構造変化を研究する手法を本稿の後半で紹介す
る。 2.タンパク質の折れ畳み反応の追跡 筆者らは、タンパク質の折れ畳み反応を追跡す
るために、化学修飾により光解離性修飾基をタン
パク質の特定のアミノ酸残基の側鎖に導入し、得
られた修飾タンパク質に光を照射することによ
り、タンパク質の折れ畳み反応を追跡する手法を
提案している。この手法では、不安定化した修飾
タンパク質にパルス光を照射して、タンパク質か
ら修飾基を瞬時に外すことにより、タンパク質の
フォールディング反応を一斉に開始させ、タンパ
ク質の構造形成反応の初期段階を観測する(図
1)。
図1.光解離性修飾基で修飾されたタンパク質の紫外光
照射による折れ畳み反応。
例として、まず発色団としてヘムを有するシト
クロムc(cyt c)の場合について述べる。cyt cは比較的長いαへリックス3本と1巻き半程度
の短いαへリックス1本からなり、変性状態でも
ヘムはタンパク質に結合したままである。光解離
性修飾基の位置選択的な導入のため、ウマ cyt cに 2 個存在するメチオニン残基のオルトニトロ
ベンジル化を行った。cyt c のメチオニン 65(Met65、ヘムに非配位)とメチオニン 80(Met80、ヘムに配位)を修飾し、その後、Met65 のみが修
飾された(NBz-Met65)タンパク質と Met80 のみ
が修飾されたタンパク質を分離した(1-3)。

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 18
未修飾 cyt c と Met65 修飾 cyt c のグアニジウム
塩酸塩による変性の中点濃度はそれぞれ2.8 、1.9 M であり、Met65 修飾 cyt c の立体構造が未修飾
cyt c の立体構造よりも不安定であることが解か
った。2M グアニジウム塩酸塩存在下では、未修
飾 cyt c 分子はほとんど変性しないが、Met65 修
飾 cyt c 分子は約半分が変性した。そこで、2M グ
アニジウム塩酸塩存在下で Met65 修飾 cyt c に光
を照射すると、タンパク質の折れ畳み反応が観測
できた(2)。光照射後のヘムのソーレ帯(401 及び
418.5 nm)の吸光度変化を追跡した結果、時定数、
約 50 μs、10 ms、100 ms の吸収変化の相が観測
された(図2)。特に、時定数が小さい約 50 μsの相の存在を明らかにすることは、タンパク質の
フォールディングが如何にして起こるかを解き
明かすのに大変重要である。約 50 μs の吸収変
化は構造が崩れた状態のタンパク質の疎水性残
基が非特異的に凝集する反応に連動するへムの
環境変化に対応し、10 ms と 100 ms の吸収変化は
へムの軸配位子の変化に対応する。
図2.308-nmパルス光照射による酸化型Met65修飾 cyt c
の吸光度の時間変化:(a)418.5、(b)401 nm (2)。(A)
-200–600 µs と(B) -0.1–0.3 s。挿入図:早い相の吸光
度変化強度の波長依存性。2 M GdnHCl 存在下。
次に、緑色植物などの葉緑体中に存在する可溶
性の電子伝達銅タンパク質であるプラストシア
ニン(PC)から銅原子を取り除いたアポ型タン
パク質(apoPC)の折れ畳み反応を取り上げる。
PC はβシートタンパク質に分類され、8 本から
なる 2つのβシートと 1回転のαへリックスを有
し、タンパク質内の唯一のシステイン残基
(Cys84)は銅原子に配位している。Cys84 の硫
黄原子は銅に配位しているので、PC から銅を外
して得た apoPC の Cys の硫黄原子を、オルトニ
トロベンジル基のジメトキシ誘導体(DMNB)で
修飾した(3,4)。
図3.パルス光照射による DMNB 修飾 apoPC の(A)吸収と
(B)CDスペクトル変化。挿入図:apoPC のCDスペク
トル、(a) pH 7 と(b) pH 2。
修飾 apoPC に 355 nm のパルス光を照射すると、
修飾基に由来する 355 nm の吸収帯が減少したこ
とより、光照射により修飾基が apoPC から遊離
したことが判明した(図3A)。一方、修飾 apoPCと未修飾 apoPCの酸変性状態のCDスペクトルを
比較したところ、修飾 apoPC は変性状態である
ことが解った(図3B)。また、修飾 apoPC への
355 nm の光照射により、変性状態を示していた
CD スペクトルはアポ状態の CD スペクトルに戻
った(図3B)。このことより、光照射によりタ
ンパク質が変性状態からネイティブのβシート
構造の状態に戻ることが確かめられた。 そこで、過渡回折格子法(5)を用いて、修飾タン
パク質に光を照射したときの反応を追跡した。修
飾タンパク質への光照射により得られた回折シ
グナルは、まず、強度が急激に増大し、その後、
ゆっくり増大し、マイクロ秒の時間領域で減少し
た(4)。ゆっくりと増大した成分は修飾基がタンパ
ク質から解離する反応に対応し、400 ns で修飾基
がタンパク質から解離することが判明した(図
4)。詳しい議論は省くが、光照射後、約 270 μs でタンパク質の体積減少が観測され、この体積
変化は変性状態から初期段階での疎水基凝集状
態への変化に帰属できた(4)。さらに、時定数 23 msの相は、タンパク質と水分子との分子間水素結合
社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 19
がタンパク質の分子内水素結合へと変化したこ
とに対応すると考えられた(4)。この他、PC の折
れ畳み反応では、プロリンの異性化に対応する
30-1000 秒の反応が観測されている。
OCH3
CH2S
O2N OCH3
270 μs
23 ms
400 ns
OHC
OCH3
OCH3ON
+
hνhν
30 - 1000 sslow
図4.修飾タンパク質への紫外光照射による apoPC の折
れ畳み反応の模式図。
3.金属タンパク質の分子認識と活性部位の構造
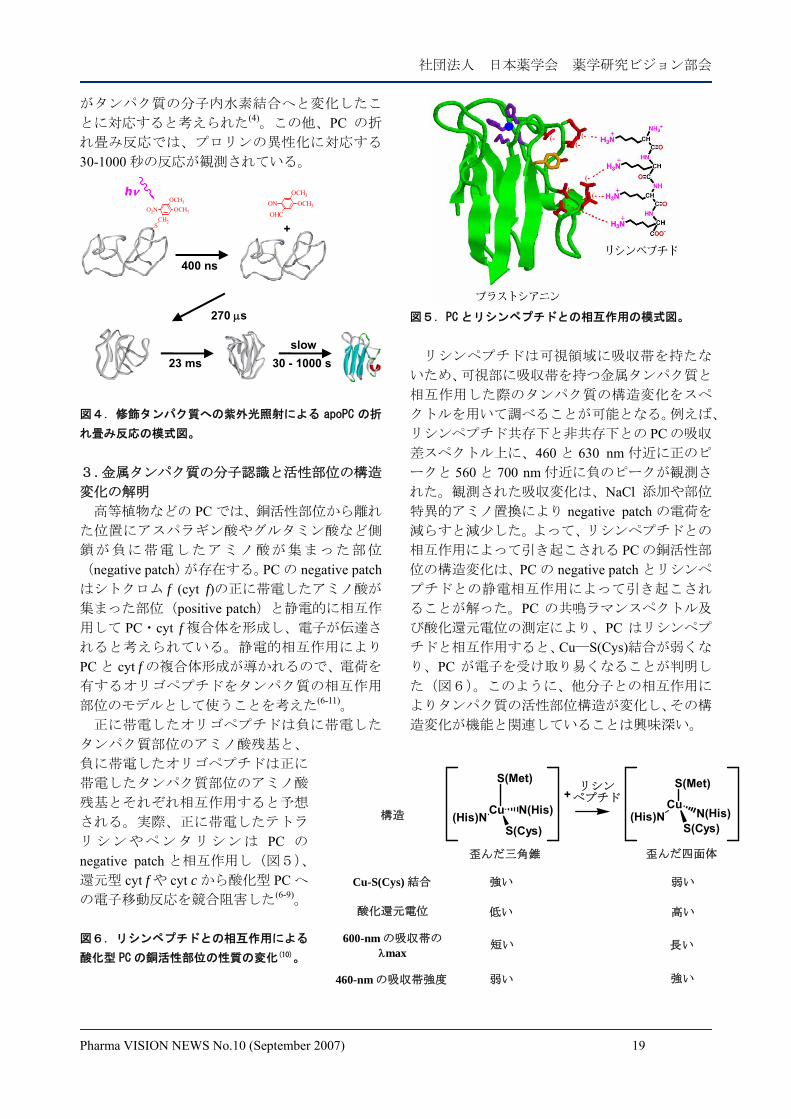
変化の解明 高等植物などの PC では、銅活性部位から離れ
た位置にアスパラギン酸やグルタミン酸など側
鎖が負に帯電したアミノ酸が集まった部位
(negative patch)が存在する。PC の negative patchはシトクロム f (cyt f)の正に帯電したアミノ酸が
集まった部位(positive patch)と静電的に相互作
用して PC・cyt f 複合体を形成し、電子が伝達さ
れると考えられている。静電的相互作用により
PC と cyt f の複合体形成が導かれるので、電荷を
有するオリゴペプチドをタンパク質の相互作用
部位のモデルとして使うことを考えた(6-11)。 正に帯電したオリゴペプチドは負に帯電した
タンパク質部位のアミノ酸残基と、
負に帯電したオリゴペプチドは正に
帯電したタンパク質部位のアミノ酸
残基とそれぞれ相互作用すると予想
される。実際、正に帯電したテトラ
リシンやペンタリシンは PC の
negative patch と相互作用し(図5)、
還元型 cyt f や cyt c から酸化型 PC へ
の電子移動反応を競合阻害した(6-9)。 図6.リシンペプチドとの相互作用による
酸化型 PC の銅活性部位の性質の変化(10)。
図5.PC とリシンペプチドとの相互作用の模式図。
リシンペプチドは可視領域に吸収帯を持たな
いため、可視部に吸収帯を持つ金属タンパク質と
相互作用した際のタンパク質の構造変化をスペ
クトルを用いて調べることが可能となる。例えば、
リシンペプチド共存下と非共存下との PC の吸収
差スペクトル上に、460 と 630 nm 付近に正のピ
ークと 560 と 700 nm 付近に負のピークが観測さ
れた。観測された吸収変化は、NaCl 添加や部位
特異的アミノ置換により negative patch の電荷を
減らすと減少した。よって、リシンペプチドとの
相互作用によって引き起こされる PC の銅活性部
位の構造変化は、PC の negative patch とリシンペ
プチドとの静電相互作用によって引き起こされ
ることが解った。PC の共鳴ラマンスペクトル及
び酸化還元電位の測定により、PC はリシンペプ
チドと相互作用すると、Cu―S(Cys)結合が弱くな
り、PC が電子を受け取り易くなることが判明し
た(図6)。このように、他分子との相互作用に
よりタンパク質の活性部位構造が変化し、その構
造変化が機能と関連していることは興味深い。
Cu
S(Cys)
N(His)(His)N
S(Met)
Cu
S(Cys)N(His)(His)N
S(Met)
歪んだ四面体
Cu-S(Cys) 結合
歪んだ三角錐
460-nm の吸収帯強度
長い
酸化還元電位
リシンペプチド
強い 弱い
高い低い
600-nm の吸収帯の
λmax短い
強い弱い
構造
+

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 20
リシンペプチドは他の金属タンパク質とも相
互作用し、このとき金属タンパク質の活性部位構
造が変化する。例えば、リシンペプチドは cyt cからシトクロム c ペルオキシダーゼへの電子移
動反応を阻害し (12)、リシンペプチドを共存させ
ることにより、シトクロム c ペルオキシダーゼの
ソーレ帯の形が変化した。 一方、負に帯電しているアスパラギン酸ペプチ
ドも正に帯電しているタンパク質と相互作用す
る。例えば、オリゴアスパラギン酸は還元型 cyt c、cyt f から酸化型 PC への電子移動反応を阻害した(6,7,9)。また、アスパラギン酸ペプチドを cyt c 溶
液に加えると、シトクロム c のソーレ帯の吸収極
大波長がシフトし(6,7,9)、共鳴ラマンスペクトルに
よってもアスパラギン酸ペプチドとの相互作用
によって cyt c の状態が変化することが示された。
さらに、cyt c や cyt f の酸化還元電位はアスパラ
ギン酸ペプチドを添加することにより 7~20 mV低い値にシフトし、相互作用によりシトクロム cや cyt f の電子供与能が増すことも解った(9)。 4. おわりに 本稿で紹介した光解離性修飾基を用いた手法
により、cyt c および apoPC のフォールディング
反応が初期段階から観測できた。特に、apoPC で
は、400 ns で修飾基がタンパク質から解離し、270
μs で初期の疎水基の凝集が起こり、23 ms で水
素結合が分子間から分子内に変化する描像が得
られた。ここで紹介した手法は他のタンパク質へ
も応用可能であり、タンパク質の初期段階でのフ
ォールディング反応を追跡するのに有効である。
また、電荷を有するオリゴペプチドが種々の金属
タンパク質の分子認識及び他分子との相互作用
によって引き起こされる構造変化の研究に有用
であることが示された。特に、電荷を有するペプ
チドは可視領域に吸収を持たないため、他分子と
の相互作用による金属タンパク質の活性部位の
構造変化を調べるのに有効である。これらの手法
がタンパク質の構造とタイナミクスの解明に用
いられことにより、タンパク質の構造に関する理
解が深まり、創薬に役立つことを期待する。 謝辞 過渡回折格子法の結果は京都大学大学院理学
研究科化学専攻の寺嶋正秀教授および寺嶋研の
学生さん達との共同研究による成果であり、これ
らの方々に感謝いたします。また、山内脩教授(名
古屋大学名誉教授、関西大学名誉教授)、名古屋
大学大学院理学研究科物質理学専攻の渡辺芳人
教授、藤本ゆかり博士(現大阪大学)、奥野貴士
博士(現京都薬科大学)をはじめとする多くの共
同研究者の方々にも感謝いたします。 参考文献
1) T. Okuno, S. Hirota, O. Yamauchi, Chem. Lett., 290-291 (2000). 2) T. Okuno, S. Hirota, O. Yamauchi, Biochemistry, 39, 7538-7545 (2000). 3) 廣田俊, 生物物理, 45, 207-210 (2005). 4) S. Hirota, Y. Fujimoto, J. Choi, N. Baden, N. Katagiri, M. Akiyama, T. Okajima, T. Takabe, Y. Funasaki, Y. Watanabe, M.
Terazima, J. Am. Chem.. Soc., 128, 7551-7558 (2006). 5) 寺嶋正秀, 分光研究, 47, 230-244 (1998).
6) S. Hirota, O.Yamauchi, Chemical Record (review), 1, 290-29 (2001).
7) S. Hirota, O. Yamauchi, Eur. J. Inorg. Chem. (review), 17-25 (2002).
8) S. Hirota, K. Hayamizu, M. Endo, T. Hibino, T. Takabe, T. Kohzuma, O. Yamauchi, J. Am. Chem. Soc., 120, 8177-8183 (1998).
9) S. Hirota, M. Endo, K. Hayamizu, T. Tsukazaki, T. Takabe, T. Kohzuma, O. Yamauchi, J. Am. Chem. Soc., 121, 849-855 (1999).
10) S. Hirota, K. Hayamizu, T. Okuno, M. Kishi, H. Iwasaki, T. Kondo, T. Hibino, T. Takabe, T. Kohzuma, O. Yamauchi,
Biochemistry, 39, 6357-6364 (2000).
11) S. Hirota, H. Okumura, S. Arie, K. Tanaka, M. Shionoya, T. Takabe, N. Funasaki, Y. Watanabe, J. Inorg. Biochem., 98, 849-855
(2004).
12) S.Hirota, T. Tsukazaki, O. Yamauchi, Biochem. Biophys. Res. Commun., 268, 395-397 (2000).

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 21
◆略 歴◆ 廣田 俊 (Shun HIROTA):1990 年 3 月京都大学工学部石油化学科卒業後、同大学大学院
工学研究科分子工学専攻修士課程、総合研究大学院大学数物科学研究科機能分子科学専攻博士後期課程、日
本学術振興会特別研究員、米国エモリー大学化学科博士研究員、名古屋大学大学院理学研究科物質理学専攻
(化学系)助手、京都薬科大学薬学部助教授を経て、2007 年 4 月より現職。2004 年 10 月より科学技術振興
機構さきがけ研究者を兼任。

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 22
薬学研究最前線(4)
GPI アンカー型タンパク質の脂質リモデリング機構の解明
梅村 真理子、地神 芳文((独)産業技術総合研究所セルエンジニアリング研究部門)
1. GPI アンカー型タンパク質とは グリコシルフォスファチジルイノシトール
(GPI) は真核生物においてタンパク質を細胞膜
に結合させる役割をしている糖脂質である。哺乳
動物の細胞では 150種類以上、酵母Saccharomyces cerevisiae では 60 種類以上のタンパク質が GPI アンカー型タンパク質として同定されている。これ
らの GPI アンカー型タンパク質は細胞間接着や
シグナル伝達に関与しているもの、細胞表層のコ
ートタンパク質として外来からの攻撃を防御し
ているものなど様々な機能を持っている。酵母で
は、細胞壁に局在するタンパク質も GPI アンカー
型であり、細胞壁の生合成や恒常性維持などに重
要な役割を担っている。 通常、GPI アンカー型タンパク質は、細胞膜上
で脂質ラフトと呼ばれるステロールやスフィン
ゴ脂質からなる特別な膜構造に存在している。脂
質ラフトは、シグナル伝達のプラットフォームと
して、またタンパク質の輸送・選別などで重要な
役割を果たしている。 GPIアンカーの基本骨格は真核生物で保存され
ており、フォスファチジルイノシトール (PI)、グ
ルコサミン、マンノース、エタノールアミンリン
酸からなる。小胞体膜上にある PI に、それぞれ
が順次転移されてGPIアンカーが完成する【図 1】。その後トランスアミダーゼ複合体により、完成型
GPI がタンパク質に転移される。この際、タンパ
ク質の C 末端側にある GPI 付加シグナルが除去
される。GPI 生合成に関与する遺伝子の大半は既
に同定されている。GPI の生合成過程は酵母では
細胞機能に必須であり、また哺乳動物では、細胞
レベルでは欠損しても致死ではないが、個体レベ
ルでは胚発生に重要なことが報告されている。 GPI がタンパク質に転移した後に、GPI のイノ
シトール部分に付いたアシル基が除去される (脱アシル化)。哺乳動物細胞では PGAP1 遺伝子
が、酵母では BST1 遺伝子がこれに関与し、それ
ぞれの遺伝子がコードするタンパク質の活性部
位にはリパーゼモチーフが存在する。脱アシル化
された GPI アンカー型タンパク質は、その後、脂
質部分の改変を受ける。小胞体で GPI アンカーが
合成される時の PI はその脂質部分に不飽和脂肪
酸を持っているが、GPI アンカー型タンパク質が
脂質ラフトに効率よく挿入されるためには脂質
部分は飽和脂肪酸に変換される必要がある。この
変換は「脂質リモデリング」と呼ばれている【図
2】。これらのステップに関与する遺伝子は全く解
明されていなかったが、2006 年に哺乳動物細胞
の PGAP2 [1] や酵母のアシルトランスフェラー
ゼをコードするGUP1 [2] が脂質リモデリング反
応に関与する遺伝子として報告されて以来、脂質
リモデリングに関与する遺伝子が次々と同定さ
れた。酵母と哺乳動物細胞を用いた脂質リモデリ
ングに関与する最近の遺伝子同定を中心に報告
する。
図1 酵母の GPI アンカー型タンパク質の合成経路 GPI は小胞体で合成されて、ゴルジ体を経て、細胞膜へ
輸送される。

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 23
PI リゾ型 PI
GUP1 P CWH43 P
ジアシルグリセロール セラミド
PI リゾ型 PI
GUP1 P CWH43 P
ジアシルグリセロール セラミド 図2 酵母の脂質リモデリング機構 小胞体での脂質リモデリング機構を示す。点線で囲んだ
部分は脂質部分の構造の違いを示す。記号と略語は図1
と同様なので省略する。 2. GPI アンカー型タンパク質の脂質リモデリン
グ機構 GPIアンカーの脂質リモデリング反応はタンパ
ク質に完成型 GPI が転移した後に起こる。まず初
めに PI の sn-2 位の飽和脂肪酸がフォスフォリパ
ーゼ A2反応により除去され、リゾ型の GPI がで
きる。次に、アシルトランスフェラーゼにより、
sn-2 位に飽和型の脂肪酸が転移される。哺乳動物
細胞では、主に C18:0 の脂肪酸が転移され、酵母
では C26:0 の長鎖脂肪酸が転移される。さらに、
酵母においては、多くの GPI アンカー型タンパク
質で脂質部分がジアシルグリセロール型からが
セラミド型に変換される【図2】。脂質リモデリ
ングにおける3つのステップに関する最近の知
見を次項で解説する。
3. GPI アンカー型タンパク質の PI 部分の sn-2 位
の脂肪酸除去 (フォスフォリパーゼ A2反応) GPIアンカー型タンパク質の脂質リモデリング
は初めに PI の sn-2 位の不飽和脂肪酸が除去され
る。この反応は、フォスフォリパーゼ A2 反応で
あり、酵母ではこの反応に PER1 遺伝子が関与す
ることを、我々のグループで明らかにした [3]。 我々は既に、GPI アンカー型タンパク質の脱ア
シル化に関与している BST1 が GPI アンカー型タ
ンパク質の品質管理に関与していることを明ら
かにした [4]。さらに BST1 の分子メカニズムを
詳細に検討するために BST1 と遺伝学的関連のあ
るGUP1遺伝子とPER1遺伝子に着目した。GUP1は GPI アンカーの脂質リモデリングにおいて、ア
シルトランスフェラーゼであることが報告され
ている (次項で詳述する)。従って、PER1 遺伝子
も GPI アンカー型タンパク質の生合成や脂質リ
モデリングに関与している可能性が推測された。 per1 破壊株は 37°C での生育に温度感受性を示
す他、細胞壁の構成成分であるキチンに結合する
薬剤 (Calcofluor white) に感受性を示したことか
ら、細胞壁が弱くなっていることが考えられた。
GPIアンカー型タンパク質の一種であるGas1pは、
通常、野生株では細胞膜、特に脂質ラフトに局在
しているが、per1 破壊株ではラフトに局在できな
くなっていた。細胞内の Gas1p 量は野生株よりも
有意に少なく、大半は細胞外へ放出されていた。
従って、GPI アンカー型タンパク質の生合成、脂
質リモデリング、細胞内輸送のいずれかに欠損の
ある可能性が考えられたが、GPI アンカーの生合
成過程には異常は観察されなかった。そこで、
GPI アンカー型タンパク質の脂質部分を解析し
たところ、per1 破壊株において脂質部分が通常の
脂肪酸を持つ PI (すなわち、GPI 生合成の際に用
いられる PI とほぼ同等もの) であることが明ら
かになった。さらに、per1 破壊株から抽出した
Gas1p に小胞体が含まれる膜画分を in vitro で反
応させたところ、野生株の膜画分ではリゾ型 GPIが合成されたが、per1 破壊株の膜画分ではリゾ型
GPI は合成されなかった。従って、Per1p は GPIアンカー型タンパク質の PI 部分をリゾ型に変換
するフォスフォリパーゼ A2 反応に関与している
ことが明らかになった。 哺乳動物細胞では PERLD1 (PGAP3) という酵
母 PER1 の相同遺伝子が存在する。ヒト由来の
PERLD1 は per1 破壊株の機能欠損を回復したこ
とから、PER1 と機能的にも相同であることが判
明した。PGAP3 の機能解明を含めた哺乳動物細
胞での脂質リモデリングの解析も同時に行われ
た [5]。PGAP3 PGAP2 二重欠損株 (PGAP2 につ
いては後述する) が、GPI アンカー型タンパク質
が放出される PGAP2 欠損株の表現型を回復する
変異株として取得された。二重欠損株の GPI アン
カー型タンパク質の脂質部分は不飽和脂肪酸を
持っているのに対し、野生株の脂質部分は飽和脂
肪酸 (C18:0) を持っていた。従って、哺乳動物細
胞の脂質リモデリングは PGAP3 (PERLD1) が不
飽和脂肪酸を除去し、他のアシルトランスフェラ
ーゼが飽和型脂肪酸 (C18:0) を転移するのでは
ないかと考えられている。また、PGAP3 PGAP2二重欠損株では、GPI アンカー型タンパク質が脂
質ラフトに局在できないことが示された。従って、
酵母 PER1 および哺乳類 PERLD1 (PGAP3) の関
与する脂質リモデリングは GPI アンカー型タン

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 24
パク質が脂質ラフトに局在するために必須なも
のと考えられる。
4. PI 部分の sn-2 位へのアシルトランスフェラー
ゼ反応について フォスフォリパーゼ A2 反応の産物であるリゾ
型 GPI には飽和脂肪酸が転移されるが、酵母では、
主に C26 の長鎖脂肪酸が転移される。このアシ
ルトランスフェラーゼ反応に GUP1 遺伝子が関
与することが Conzelmann のグループにより報告
された [2]。Gup1p は膜結合型アシルトランスフ
ェラーゼのモチーフを持つ小胞体局在の膜タン
パク質である。gup1 破壊株において、GPI アンカ
ー型タンパク質 Gas1p は per1 破壊株と同様に脂
質ラフトに局在できず、大半は細胞外に放出され
ていた。さらに、GPI アンカー型タンパク質の脂
質部分を解析したところ、リゾ型 GPI になってい
た。また、活性部位であるアシルトランスフェラ
ーゼモチーフのヒスチジンをアラニンに置換し
た変異株では、gup1 破壊株と同様の欠損を示し
た。従って、GUP1 はリゾ型 GPI に長鎖飽和脂肪
酸を転移する役割を担っていることが示唆され
た。 哺乳動物細胞では、PGPA2 (FRAG1) がこのス
テップに関与していることがTashimaらにより報
告されている [1]。PGAP2 は GPI アンカー型タン
パク質が正常に細胞膜に局在できず、細胞外に放
出されてしまう変異株の原因遺伝子として同定
された。PGAP2 欠損株では細胞内の GPI アンカ
ー型タンパク質はリゾ型 GPI になっており、その
ため細胞膜へ輸送された後にフォスフォリパー
ゼ D などの攻撃を受けやすくなり、細胞外へ放
出されると考えられる。従って、PGAP2 はリゾ
型 GPI に飽和脂肪酸を転移するステップに関与
していることが予想された。しかし、PGAP2 は
アシルトランスフェラーゼモチーフなどを持っ
ていないことから、このステップの触媒サブユニ
ットである可能性は低いと考えられる。
5. PI 部分のジアシルグリセロールからセラミド
への変換 酵母では、GPI アンカー型タンパク質の PI のジ
アシルグリセロールはさらにセラミドに変換さ
れる。この反応に関与している遺伝子は不明であ
ったが、最近、我々は CWH43 遺伝子がこれに関
与していることを明らかにした [6]。 酵母の CWH43 は PGAP2 に相同性を示したが、
相同領域は N 末端側に限られ PGAP2 よりも 700アミノ酸 C 末端側が長くなっていた。さらに、
この部分は哺乳類の機能未知な遺伝子と相同性
があることから、PGAP2 の機能に加えて、別の
新しい機能を持っていることが推測された。そこ
で、cwh43 破壊株の GPI アンカー型タンパク質の
脂質部分を解析したところ、長鎖脂肪酸を持つ
PI が検出されたが、セラミドは検出されなかっ
た。野生株はセラミド型の脂質部分を持っていた
ことから、CWH43 は GPI の脂質部分をセラミド
に変換する反応に関与していることが明らかに
なった。また、この反応には PGAP2 と相同性が
ない C 末端側部分が重要であることも明らかに
なった。先に述べた様に、この領域は哺乳動物細
胞の機能未知な遺伝子と相同性がある。そこで、
その相同遺伝子であるマウスのC130090K23遺伝
子を酵母の cwh43 破壊株にて発現させたところ、
GPI アンカー型タンパク質の脂質部分はセラミ
ド型に変換された。現在までに、哺乳動物細胞に
おいて、セラミドを脂質部分に持つ GPI アンカー
型タンパク質の存在は報告されていない。したが
って、C130090K23 は、酵母内ではセラミド型へ
の変換能を持っているものの、哺乳動物細胞内で
は他の似た様な反応に関与している可能性もあ
り、今後の更なる解析が待たれる。 6. 脂質リモデリングとガンとの関連
PERLD1 が GPIの脂質リモデリングにおけるフ
ォスフォリパーゼ A2 活性に関与していることが
明らかになったが、PERLD1 遺伝子はいくつかの
ガン細胞で、発現上昇していることが報告されて
いる [7]。PERLD1 遺伝子はガン細胞で増幅され
ている 17q12 染色体上に位置し、またガン遺伝子
として有名な c-ERBB-2 遺伝子とともに増幅され
ていた。PERLD1 遺伝子は c-ERBB-2 遺伝子のプ
ロモーター部分から逆向きに転写される。 また、FRAG1 (PGAP2) はもともと骨肉腫細胞
で fibroblast growth factor receptor 2 (FGFR2) の C末端側に染色体再配列によって融合された新規
遺伝子として単離された [8]。ガン細胞で見受け
られる FGFR2 は、N 末端側部分が欠けているこ
とが多く、このためシグナル伝達が異常になるこ
とがガン化の原因であるが、FRAG1 が融合した
FGFR2 タンパク質では、局在が異常になってチ
ロシンリン酸化が加速され、その結果、ガン化し
たものと予想されている。FRAG1 (PGAP2) が脂
質リモデリングに関与していることが明らかに

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 25
なった現在、FRAG1 遺伝子が直接ガン化に関与
しているかは不明であるが、今後の解析が期待さ
れる。 一方、GPI の生合成過程に関与する遺伝子の中
にもガン遺伝子となるものが存在している。ヒト
の完成型 GPI をタンパク質に転移するトランス
アミダーゼ複合体は GPI8、GPAA1、CDC91L1 (PIG-U)、PIG-T、PIG-S の5つのサブユニットか
ら成るが、そのうち、3つの遺伝子 (GPAA1、CDC91L1、PIG-T) についてガン遺伝子であると
の報告がなされている。CDC91L1 (PIG-U)は膀胱
ガン細胞において、特異的に増幅している
20p11~13 の領域に局在しており、その発現がい
くつかの膀胱ガン細胞において増大していた [9]。また、CDC91L1 を高発現させた細胞は増殖
能が高くなり、マウスに導入すると腫瘍を形成し
た。この細胞では GPI アンカー型タンパク質のウ
ロキナーゼレセプターの発現が上昇し、その結果
STAT-3 のリン酸化が亢進していることが報告さ
れている。 GPIアンカーの脂質リモデリングとガン化との
関係は明確ではないが、シグナル伝達に関与する
GPI アンカー型タンパク質が異常になることに
よりガン化が促進されることや、臨床で用いられ
ているガンマーカーの多くが GPI アンカー型タ
ンパク質であることからも、GPI アンカー型タン
パク質の生合成や修飾に関与する遺伝子がガン
に関与する可能性はきわめて高いと考えられる。
7. GPI アンカー型タンパク質と病気 我々は、酵母において Gas1p の変異体 (Gas1*p)
を作成し、これがフォールディング (折りたた
み) が異常な GPI アンカー型タンパク質のモデ
ルタンパク質となることを確認した [4]。この
Gas1*p には GPI は転移されているが、小胞体に
留まり、小胞体関連タンパク質分解系 (ERAD) によりプロテアソームで分解されてしまう。さら
に、Gas1*p の分解や品質管理に GPI の脱アシル
化を担っている BST1 が関与していることを明ら
かにした。このモデル系は、GPI アンカー型タン
パク質の分解や品質管理が原因とされる疾患の
研究に大いに役に立つと期待される。 GPIアンカー型タンパク質が関与する病気とし
て、プリオン病や低フォスファターゼ症などがあ
る。プリオン病は変異型プリオンタンパク質がβ
シート構造をとり凝集して分解されにくくなっ
た結果、脳に異常蓄積することにより発症する。
低フォスファターゼ症は組織非特異的アルカリ
フォスファターゼに変異が入り機能しなくなる
ことにより骨の石灰化異常、てんかん様発作、血
清アルカリ性フォスファターゼ活性の異常低下
などが起こる。いくつかの変異型アルカリフォス
ファターゼはゴルジ体へ送られることなく小胞
体に留まり、その後ユビキチン/プロテアソーム
系で速やかに分解されると報告されている。プリ
オンタンパク質もアルカリフォスファターゼも
GPI アンカー型タンパク質である。これらの病気
の分子メカニズムは少しずつ解明されているも
のの、治療法や薬剤の開発には至っていない。 GPI 生合成過程の異常による病気として、発作
性夜間血色素尿症がある。赤血球膜上にある補体
制御因子である GPI アンカー型タンパク質が欠
損しているために、自己補体により破壊されて、
溶血性貧血を引き起こす。この原因は GPI の生合
成過程において N-アセチルグルコサミンの転移
反応に関与している PIG-A 遺伝子欠損とそのク
ローンの拡大であると考えられている [10]。ま
た、GPI 生合成過程においてマンノース転移に関
与している PIG-M 遺伝子のプロモーター領域の
点変異により転写因子の結合が欠損し、遺伝性
GPI 欠損症が引き起こされることが報告されて
いる [11]。遺伝性 GPI 欠損症は内臓の静脈血栓
とてんかんが主な症状である。 遺伝子の解明やモデル系の確立、分子メカニズ
ムの解明が進みつつある現在、今後の治療法の確
立や阻害剤など薬剤の開発が期待される。
8. おわりに GPIアンカー型タンパク質の脂質リモデリング
機構は今まさに解明されつつある分野である。し
かし、セラミドを含む GPI アンカー型タンパク質
の脂質部分への水酸基の付加など後期のステッ
プに関する知見はまだ得られていない。また、小
胞体内での脂質リモデリングの順序、セラミドへ
の変換の基質やアクセプターの同定はまだ十分
には解明されていない。最近、哺乳動物細胞にお
いて GPI 生合成の途中で、PI の脂質部分がジア
シルグリセロールから、アシル/アルキルグリセ
ロールに変換されることが明らかになった [12]。また、原虫 Trypanosoma brucei では GPI がタンパ
ク質に転移される前に脂質リモデリングが行わ
れる。本稿で紹介した脂質リモデリング機構以外
に、このような GPI がタンパク質に転移される前
の脂質リモデリングが酵母など他の真核生物に

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 26
保存されているかは不明であり、今後の解析が待
たれる。
参考文献 1.) Tashima, Y., Taguchi, R., Murata, C., Ashida, H., Kinoshita, T. & Maeda, Y. (2006) PGAP2 is essential for correct processing
and stable expression of GPI-anchored proteins, Mol Biol Cell. 17, 1410-20.
2.) Bosson, R., Jaquenoud, M. & Conzelmann, A. (2006) GUP1 of Saccharomyces cerevisiae encodes an O-acyltransferase
involved in remodeling of the GPI anchor, Mol Biol Cell. 17, 2636-45.
3.) Fujita, M., Umemura, M., Yoko-o, T. & Jigami, Y. (2006) PER1 is required for GPI-phospholipase A2 activity and involved in
lipid remodeling of GPI-anchored proteins, Mol Biol Cell. 17, 5253-64.
4.) Fujita, M., Yoko-o, T., & Jigami, Y. (2006) Inositol deacylation by Bst1p is required for the quality control of
glycosylphosphatidylinositol-anchored proteins, Mol Biol Cell. 17, 834-50.
5.) Maeda, Y., Tashima, Y., Houjou, T., Fujita, M., Yoko-o, T., Jigami, Y., Taguchi, R. & Kinoshita, T. (2007) Fatty acid remodeling
of GPI-anchored proteins is required for their raft association, Mol Biol Cell. 18, 1497-506.
6.) Umemura, M., Fujita, M., Yoko-o, T., Fukamizu, A., & Jigami, Y. (2007) Saccharomyces cerevisiae CWH43 is involved in the
remodeling of the lipid moiety of GPI anchors to ceramides, Mol Biol Cell. in press.
7.) Nezu, M., Nishigaki, M., Ishizuka, T., Kuwahara, Y., Tanabe, C., Aoyagi, K., Sakamoto, H., Saito, Y., Yoshida, T., Sasaki, H. &
Terada, M. (2002) Identification of the CAB2/hCOS16 gene required for the repair of DNA double-strand breaks on a core
amplified region of the 17q12 locus in breast and gastric cancers, Jpn J Cancer Res. 93, 1183-6.
8.) Lorenzi, M. V., Horii, Y., Yamanaka, R., Sakaguchi, K. & Miki, T. (1996) FRAG1, a gene that potently activates fibroblast
growth factor receptor by C-terminal fusion through chromosomal rearrangement, Proc Natl Acad Sci U S A. 93, 8956-61.
9.) Guo, Z., Linn, J. F., Wu, G., Anzick, S. L., Eisenberger, C. F., Halachmi, S., Cohen, Y., Fomenkov, A., Hoque, M. O., Okami, K.,
Steiner, G., Engles, J. M., Osada, M., Moon, C., Ratovitski, E., Trent, J. M., Meltzer, P. S., Westra, W. H., Kiemeney, L. A.,
Schoenberg, M. P., Sidransky, D. & Trink, B. (2004) CDC91L1 (PIG-U) is a newly discovered oncogene in human bladder
cancer, Nat Med. 10, 374-81.
10.) Takeda, J., Miyata, T., Kawagoe, K., Iida, Y., Endo, Y., Fujita, T., Takahashi, M., Kitani, T. & Kinoshita, T. (1993) Deficiency
of the GPI anchor caused by a somatic mutation of the PIG-A gene in paroxysmal nocturnal hemoglobinuria, Cell. 73, 703-11.
11.) Almeida, A. M., Murakami, Y., Layton, D. M., Hillmen, P., Sellick, G. S., Maeda, Y., Richards, S., Patterson, S., Kotsianidis, I.,
Mollica, L., Crawford, D. H., Baker, A., Ferguson, M., Roberts, I., Houlston, R., Kinoshita, T. & Karadimitris, A. (2006)
Hypomorphic promoter mutation in PIGM causes inherited glycosylphosphatidylinositol deficiency, Nat Med. 12, 846-51.
12.) Houjou, T., Hayakawa, J., Watanabe, R., Tashima, Y., Maeda, Y., Kinoshita, T. & Taguchi, R. (2007) Changes in molecular
species profiles of glycosylphosphatidylinositol anchor precursors in early stages of biosynthesis, J Lipid Res. 48, 1599-606.
略 歴◆ 梅村 真理子 (Mariko UMEMURA):1999 年 筑波大学大学院農学研究科 博士課程前期修了。
同年 工業技術院(現 産総研)契約職員、現在に至る。2006 年 筑波大学大学院生命環境科学研究科博士課程後期に入
学。研究テーマ:GPI アンカー型タンパク質の生合成過程の解明。 地神 芳文(Yoshifumi JIGAMI):1975 年 東京大学大学院農学研究科博士課程修了(農学博士)。1976 年 工業
技術院(現 産総研)入所。生命研室長、次長を経て、2001 年(独)産総研部門長、2002 年 同センター長、2007 年 同
上席研究員、現在に至る。研究テーマ:酵母の糖鎖生物学と糖鎖工学。

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 27
薬学研究最前線(5)
核磁気共鳴法による高分子量タンパク質間相互作用の解析 嶋田 一夫(東京大学大学院薬学系研究科、産業技術総合研究所生物情報解析研究センター)
はじめに
構造生物学的手法の一つである核磁気共鳴法
(NMR)は、我々に蛋白質や核酸など生体高分
子の立体構造や相互作用様式に関する情報を提
供する。しかしながら、NMRで立体構造を求め
ることができる蛋白質は、対象蛋白質の分子量の
点でおよそ30K 以下に制限されている。これは、
高分子量蛋白質になるとNMRシグナルの線幅
が著しく増大し、詳細な解析、特に構造決定プロ
セスに必要な NOE 解析が著しく困難になること
に起因する。 蛋白質複合体における蛋白質-蛋白質相互作
用解析は、その機能を考える上で、また蛋白質の
立体構造をもとに薬物のデザインをする上で重
要なプロセスである。上で述べたNMRの制限は
蛋白質複合体に対する解析にも当然当てはまる。
したがって、高分子量蛋白質複合体の相互作用を
研究する場合、立体構造を直接求めることは行わ
れず、その代わり、複合体形成に伴うNMRシグ
ナルの化学シフト変化(化学シフト摂動法)や主
鎖アミド水素の水素-重水素交換速度変化(H-
D交換法)を指標として、結合部位の同定を行う
ことが多かった。 しかし、それら指標により決定された界面はX
線結晶構造解析により明らかとなったものと必
ずしも一致することはなく、あいまいな点があっ
た。したがって、高分子量蛋白質の相互作用解析
に対して適応可能な新しいNMR測定法が望ま
れていた。 我々が近年開発した交差飽和法および転移交
差飽和法は従来用いられてきた化学シフト摂動
法やH-D交換法に比較して、巨大蛋白質複合体
を取り扱え、精密に界面残基を求めることができ
ることが特徴である。以下に従来法である化学シ
フト摂動法やH-D交換法の解析例と、交差飽和
法の概略とその実例を説明し、これらの方法を比
較する。
抗体結合性蛋白質プロテインAと抗体との相互
作用 ここで取り上げる研究対象は抗体結合性蛋白
質プロテインAと抗体である。プロテインAは黄
色ブドウ菌の細胞壁に存在し、抗体結合性を有す
る 8 個のドメイン(A-E)がつながったビーズ状
構造を持つ。そのうちドメインB(FB)は、こ
れらの 5 個のドメインのうち基本的なものと考
えられている。その立体構造はNMRにより決定
され、図1に示すように 3 本の α-へリックスが
束ねられた構造をとっていることが示された。1)
図1:プロテインA、Bドメイン(FB)の溶液中にお
ける立体構造
FBのどのアミノ酸残基が抗体結合に関与し
ているかを調べるためNMRを用いた解析を行
った。この場合、FB2 分子がFc1 分子と結合
していることがすでに明らかになっているので、
複合体の分子量は 64K となる。したがって、複
合体の全体構造をNMRにより決定することは
困難である。そこで、化学シフト摂動法やH-D
交換法を用いて相互作用部位の同定を行った。
化学シフト摂動法とは 図2はFBおよびFB-Fc複合体の1H-1
5Nシフト相関スペクトルである。ここで用いら
れているFBは、大腸菌により発現された均一安
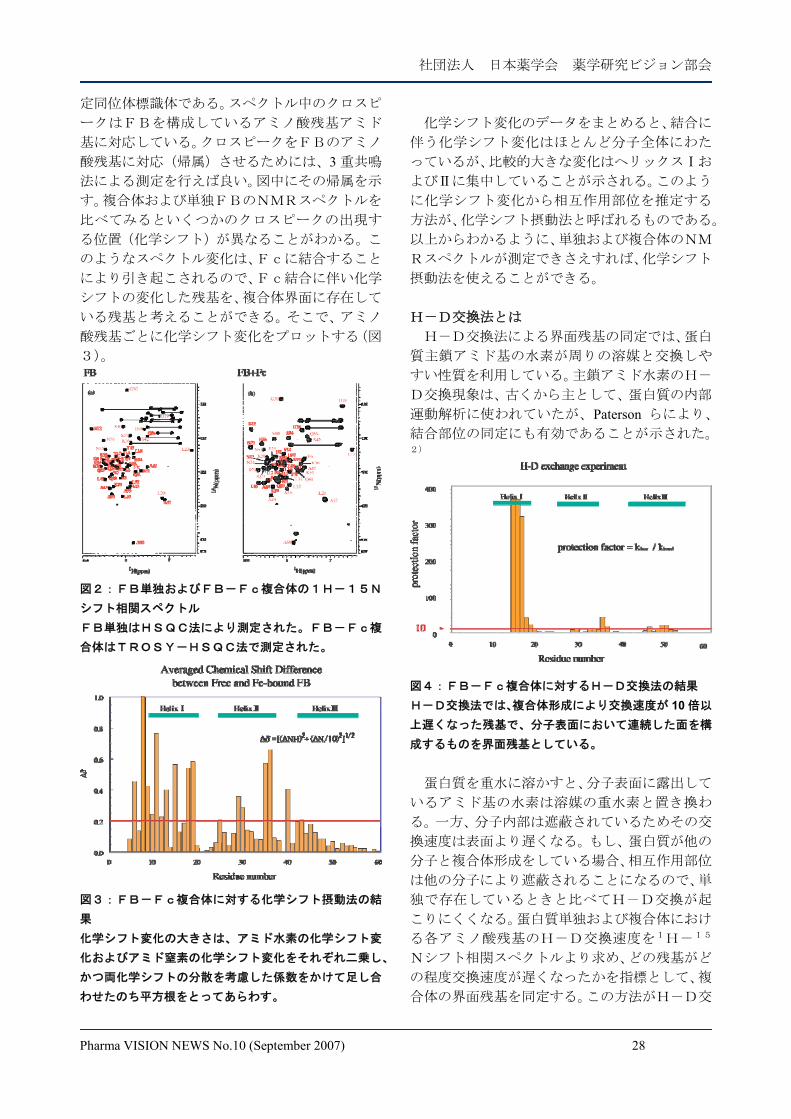
社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 28
定同位体標識体である。スペクトル中のクロスピ
ークはFBを構成しているアミノ酸残基アミド
基に対応している。クロスピークをFBのアミノ
酸残基に対応(帰属)させるためには、3 重共鳴
法による測定を行えば良い。図中にその帰属を示
す。複合体および単独FBのNMRスペクトルを
比べてみるといくつかのクロスピークの出現す
る位置(化学シフト)が異なることがわかる。こ
のようなスペクトル変化は、Fcに結合すること
により引き起こされるので、Fc結合に伴い化学
シフトの変化した残基を、複合体界面に存在して
いる残基と考えることができる。そこで、アミノ
酸残基ごとに化学シフト変化をプロットする(図
3)。
図2:FB単独およびFB-Fc複合体の1H-15N
シフト相関スペクトル FB単独はHSQC法により測定された。FB-Fc複
合体はTROSY-HSQC法で測定された。
図3:FB-Fc複合体に対する化学シフト摂動法の結
果 化学シフト変化の大きさは、アミド水素の化学シフト変
化およびアミド窒素の化学シフト変化をそれぞれ二乗し、
かつ両化学シフトの分散を考慮した係数をかけて足し合
わせたのち平方根をとってあらわす。
化学シフト変化のデータをまとめると、結合に
伴う化学シフト変化はほとんど分子全体にわた
っているが、比較的大きな変化はヘリックスⅠお
よびⅡに集中していることが示される。このよう
に化学シフト変化から相互作用部位を推定する
方法が、化学シフト摂動法と呼ばれるものである。
以上からわかるように、単独および複合体のNM
Rスペクトルが測定できさえすれば、化学シフト
摂動法を使えることができる。
H-D交換法とは H-D交換法による界面残基の同定では、蛋白
質主鎖アミド基の水素が周りの溶媒と交換しや
すい性質を利用している。主鎖アミド水素のH-
D交換現象は、古くから主として、蛋白質の内部
運動解析に使われていたが、Paterson らにより、
結合部位の同定にも有効であることが示された。2)
図4:FB-Fc複合体に対するH-D交換法の結果 H-D交換法では、複合体形成により交換速度が 10 倍以
上遅くなった残基で、分子表面において連続した面を構
成するものを界面残基としている。 蛋白質を重水に溶かすと、分子表面に露出して
いるアミド基の水素は溶媒の重水素と置き換わ
る。一方、分子内部は遮蔽されているためその交
換速度は表面より遅くなる。もし、蛋白質が他の
分子と複合体形成をしている場合、相互作用部位
は他の分子により遮蔽されることになるので、単
独で存在しているときと比べてH-D交換が起
こりにくくなる。蛋白質単独および複合体におけ
る各アミノ酸残基のH-D交換速度を1H-15
Nシフト相関スペクトルより求め、どの残基がど
の程度交換速度が遅くなったかを指標として、複
合体の界面残基を同定する。この方法がH-D交

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 29
換法と呼ばれるものである。図4に、FB-Fc
複合体形成におけるH-D交換速度の減少の程
度を残基ごとに示す。図4に示されるように、先
ほどの化学シフト摂動法から得られた結果と同
様、ヘリックスⅠおよびⅡがFc結合に関与して
いることが示される。3)
X線結晶構造解析法から得られた結果との比較 上記の 2 種類の相互作用解析法は、頻繁に使用
される。特に、化学シフト摂動法は、原理および
解釈も簡単で、また、主鎖アミド水素、窒素だけ
でなく主鎖カルボニル炭素や側鎖炭素、側鎖水素
のNMRシグナルでも、同様なことが行えるため、
一般に用いられている。 さて、FB-Fc複合体に関してはすでにX線
結晶構造解析によりその立体構造が明らかにさ
れている。4)図5に、化学シフト摂動法および
H-D交換法から得られた結果とともにX線の
結果を示す。NMRにより得られた結果はX線の
それと、よく似ているものの、必ずしも一致して
いないことがわかる。これは、それほど驚くこと
ではない。
a b
c
a b
cc
図5:各種手法により求められたFB上のFc結合部位
の比較 a:化学シフト摂動法、b:H-D交換法、c:X線結晶
構造解析法 化学シフトは、さまざまな要因により変化する。
たとえ複合体界面に存在している残基でも、複数
の要因よる化学シフト変化が全体としてキャン
セルアウトされ、結果として化学シフト変化が観
測されないこともある。また、H-D交換現象は、
基本的には-NHから-NDへの化学反応である。
したがって、H-D交換速度に影響を与える要因
は、単なる遮蔽されているかいないかだけではな
い。すなわち、化学シフト変化およびH-D交換
速度ともに、FB-Fc複合体分子間の距離を直
接反映するものではないといえる。もし、FB上
の界面残基を正しく同定したいならば、どうして
もFcからの距離と関連したNMRパラメータ
を用いる必要がある。
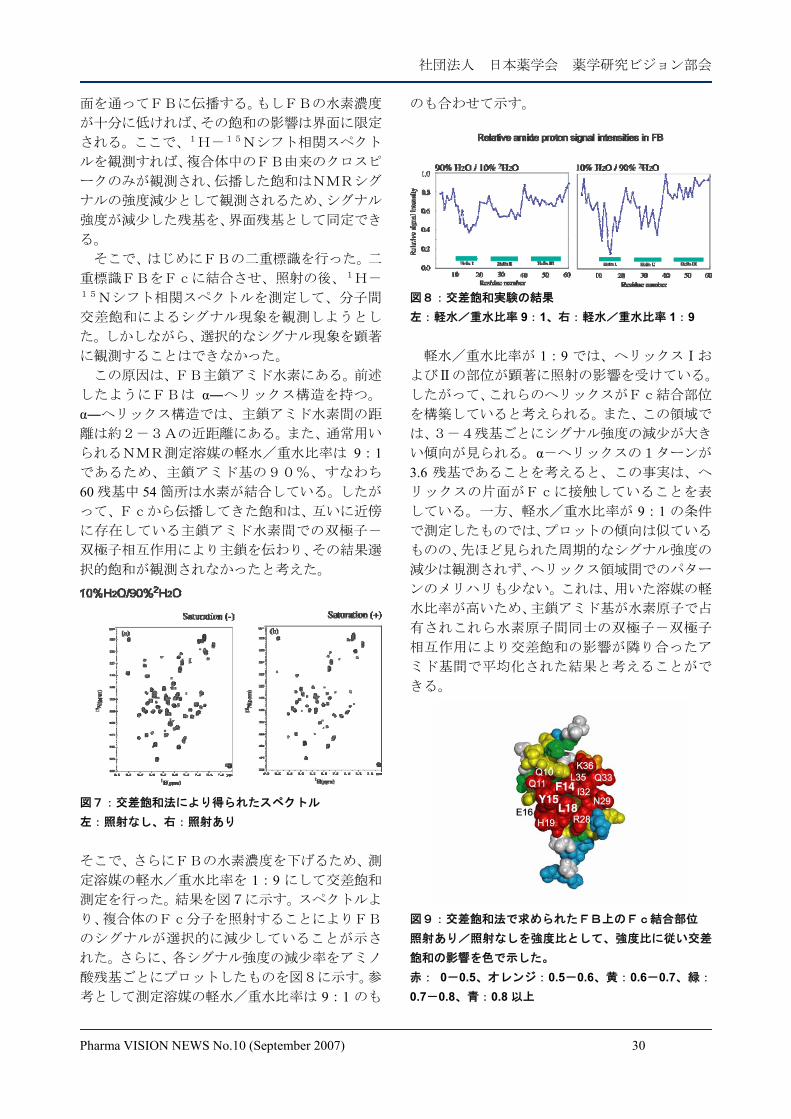
交差飽和法の概略5) そこで、FB-Fcの系を用いて以下の実験を
組み立てた。図6に示すように、2Hおよび15N
で 2 重に均一安定同位体標識したFBを非標識
Fcに結合させる。すると、この複合体は、水素
濃度が高いFc分子と水素濃度が低いFB分子
から構成されることになる。ここで、アリファテ
ィック水素の周波数を持つラジオ波を複合体に
照射すると、複合体のFcのみにラジオ波が作用
し、NMRシグナルの飽和が起こる。なぜならば、
複合体のFBは重水素化されているのでアリフ
ァティック水素が存在しないからである。そして
高分子量蛋白質では、ラジオ波照射により引き起
こされるシグナル飽和が、双極子―双極子相互作
用により近傍のプロトンに伝播するスピン拡散
とよばれる現象が強く起こる。そのため、複合体
のFc分子では、照射したアリファティックシグ
ナルのみならず、アロマティックおよびアミドシ
グナルも飽和する。結果として、分子全体に照射
の影響が伝播することになる。
図6:交差飽和法の原理 照射によって生じた飽和は、複合体のFc分子
内に留まらず、双極子―双極子相互作用により界
社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 30
面を通ってFBに伝播する。もしFBの水素濃度
が十分に低ければ、その飽和の影響は界面に限定
される。ここで、1H-15Nシフト相関スペクト
ルを観測すれば、複合体中のFB由来のクロスピ
ークのみが観測され、伝播した飽和はNMRシグ
ナルの強度減少として観測されるため、シグナル
強度が減少した残基を、界面残基として同定でき
る。 そこで、はじめにFBの二重標識を行った。二
重標識FBをFcに結合させ、照射の後、1H-15Nシフト相関スペクトルを測定して、分子間
交差飽和によるシグナル現象を観測しようとし
た。しかしながら、選択的なシグナル現象を顕著
に観測することはできなかった。 この原因は、FB主鎖アミド水素にある。前述
したようにFBは α―ヘリックス構造を持つ。
α―ヘリックス構造では、主鎖アミド水素間の距
離は約2-3Aの近距離にある。また、通常用い
られるNMR測定溶媒の軽水/重水比率は 9:1であるため、主鎖アミド基の90%、すなわち
60 残基中 54 箇所は水素が結合している。したが
って、Fcから伝播してきた飽和は、互いに近傍
に存在している主鎖アミド水素間での双極子-
双極子相互作用により主鎖を伝わり、その結果選
択的飽和が観測されなかったと考えた。
図7:交差飽和法により得られたスペクトル 左:照射なし、右:照射あり そこで、さらにFBの水素濃度を下げるため、測
定溶媒の軽水/重水比率を 1:9 にして交差飽和
測定を行った。結果を図7に示す。スペクトルよ
り、複合体のFc分子を照射することによりFB
のシグナルが選択的に減少していることが示さ
れた。さらに、各シグナル強度の減少率をアミノ
酸残基ごとにプロットしたものを図8に示す。参
考として測定溶媒の軽水/重水比率は 9:1 のも
のも合わせて示す。
図8:交差飽和実験の結果 左:軽水/重水比率 9:1、右:軽水/重水比率 1:9 軽水/重水比率が 1:9 では、ヘリックスⅠお
よびⅡの部位が顕著に照射の影響を受けている。
したがって、これらのヘリックスがFc結合部位
を構築していると考えられる。また、この領域で
は、3-4残基ごとにシグナル強度の減少が大き
い傾向が見られる。α-ヘリックスの1ターンが
3.6 残基であることを考えると、この事実は、ヘ
リックスの片面がFcに接触していることを表
している。一方、軽水/重水比率が 9:1 の条件
で測定したものでは、プロットの傾向は似ている
ものの、先ほど見られた周期的なシグナル強度の
減少は観測されず、ヘリックス領域間でのパター
ンのメリハリも少ない。これは、用いた溶媒の軽
水比率が高いため、主鎖アミド基が水素原子で占
有されこれら水素原子間同士の双極子-双極子
相互作用により交差飽和の影響が隣り合ったア
ミド基間で平均化された結果と考えることがで
きる。
図9:交差飽和法で求められたFB上のFc結合部位 照射あり/照射なしを強度比として、強度比に従い交差
飽和の影響を色で示した。 赤: 0-0.5、オレンジ:0.5-0.6、黄:0.6-0.7、緑:
0.7-0.8、青:0.8 以上

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 31
図9に、測定溶媒の軽水/重水比率が 1:9 で
測定された結果をFBの立体構造上にマッピン
グしたものを示す。図5に示したX線結晶構造か
ら得られた接触残基と本手法で同定された界面
残基はよく一致していることがわかる。これは、
本手法で用いている交差飽和現象が、距離の情報
を含む双極子―双極子相互作用に基礎を置いて
いるためである。したがって、従来の化学シフト
摂動法やH-D交換法と比較して、交差飽和法で
は厳密に相互作用部位を同定できる。
転移交差飽和法への展開 6) さて、交差飽和法では直接複合体を観測対象と
するため、たとえ高分子量測定を可能とするTR
OSY法を用いたとしても、複合体の分子量制限
が存在する。それを、克服するために、交差飽和
法の改良を行った。
図10:転移交差飽和法の原理
もし、図10に示すように、蛋白質Ⅰ(受容体
蛋白質)と蛋白質Ⅱ(リガンド蛋白質)の間に適
当速度での解離・結合が起こっている場合、結合
状態で観測されたリガンド蛋白質の交差飽和現
象を解離状態のリガンド分子で検出することが
考えられる。このような解離状態のNMRシグナ
ルを用いて結合状態の構造情報を抽出する方法
は、すでに転移NOE法で行われており、これを
交差飽和法に適応することは無理なことではな
いと考えた。 そこで、実際に転移交差飽和法がうまく働くか
どうかを調べるため、FBとインタクト抗体の相
互作用解析を行った。この場合、複合体の分子量
は 164Kに達する。用いた抗体はマウス由来のも
ので、その結合定数は 3.8x104 (M-1)であり、前述
の交差飽和法で用いたヒト抗体との結合定数よ
りも低い。そこで、2 重標識したFBとマウスイ
ンタクト抗体をモル比 1:0.1 にして、NMRス
ペクトルを測定した。 その結果、得られたスペクトルのパターンはF
B単独のものと完全に一致した。これは、大部分
のFBは解離状態で存在していることを示す。そ
こで、この条件下で交差飽和実験を行った。照射
時間が増大するにつれ、選択的にシグナル強度の
減少が見られた。その強度減少した残基をFBの
立体構造上にマッピングしたものが図11であ
る。FB-Fc複合体(分子量64K)を直接観
測して交差飽和法により求められた界面残基と
ほぼ同様な残基が抽出された。したがって、解離
状態のシグナルを用いた転移交差飽和法は予想
どおり界面残基の決定に有効であることが示さ
れた。 いくつかの残基で、交差飽和法の結果と転移交
差飽和法の結果に差異が見られる。これは、用い
ているFcの種(ヒトとマウス)が両実験で異な
るためである。すなわち、Fc領域上のFB結合
部位は、ヒトとマウスの抗体でほぼ保存されてい
るものの、若干のアミノ酸置換が生じている。そ
のため、FB結合部位の表面構造が異なり、その
違いが図9と図11の界面残基に反映している
と思われる。 図11:転移交差飽和法で
求められたFB上のFc結
合部位 照射あり/照射なしを強度
比として、強度比に従い交
差飽和の影響を色で示した。 赤: 0-0.5、オレンジ:0.5-0.6、黄:0.6-0.7、緑:
0.7-0.8、青:0.8 以上
おわりに 以上示したように、交差飽和法で求められた界
面残基は、従来法(化学シフト摂動法、H-D交
換法)で求められたものと比べ、高い精度をもつ。
また、転移交差飽和法は、測定対象が遊離蛋白質
であるため、複合体の分子量の制限はない。実際、
筆者らのグループでは、繊維状コラーゲンとコラ
ーゲン結合蛋白質 7,8)、イオンチャネルとその阻
害蛋白質の相互作用を解析し 9)、界面残基の同定
に成功しつつある。さらに、この手法は、蛋白質
-蛋白質相互作用に限らず、少しの改変を施すこ

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 32
とにより、蛋白質―核酸、蛋白質―脂質など他の
相互作用解析に応用可能である 10)。 多くの生物種の遺伝子配列情報が明らかされ、
蛋白質の立体構造に関しても、X線結晶構造解析、
NMRなどを駆使して網羅的に構造決定する試
みが行われている。近い将来に構築されるであろ
う蛋白質の立体構造データベースを考えると、蛋
白質・蛋白質間相互作用を構造生物学的に解明す
ることの重要性が一層増してきているといえる。
今後、いくつかの蛋白質複合体を研究対象として、
相互作用様式の原子レベルの理解に基礎をおい
た生体内ネットワークの解明をおこなっていき
たい。 謝辞:ここで紹介した研究は、高橋栄夫(産業
総合技術研究所、JBIRC)はじめ東京大学大
学院薬学系研究科生命物理化学教室の研究室員
の協力で行われたものである。また、研究の一部
は新エネルギー・産業技術総合開発機構(NEDO)
の助成金にて行われたものである。ここに謝意を
表します。
参考文献
1)Hiroaki Gouda, Hidetaka Torigoe, Akiko Saito, Moriyuki Sato, Yoji Arata, and Ichio Shimada,
Biochemistry (1992) 31, 9665-9672.
2)Paterson, Y., Englander, S., W., and Roder, H. (1990) Science 249, 755-759.
3)Hiroaki Gouda, Miki Shiraishi, Hideo Takahashi, Koichi Kato, Hidetaka Torigoe, Yoji Arata, and Ichio
Shimada, Biochemistry (1998) 37, 129-136.
4)Deisenhofer, J. Biochemistry (1981) 20, 2361-2370.
5)Hideo Takahashi, Tamiji Nakanishi, Keiichiro Kami, Yoji Arata and Ichio Shimada
Nat. Struct. Biol. (2000) 7, 220-223.
6)Tamiji Nakanishi, Mayumi Miyazawa, Masayoshi Sakakura, Hiroaki Terasawa, Hideo Takahashi, and Ichio
Shimada, J. Mol. Biol. (2002) 318, 245-249.
7)Noritaka Nishida, Hiromi Sumikawa, Masayoshi Sakakura, Nobuhisa Shimba, Hideo Takahashi, Hiroaki
Terasawa, Ei-ichiro Suzuki, Ichio Shimada Nature Struct. Biol. (2003) 10, 53-58.
8)Osamu Ichikawa, Masanori Osawa, Noritaka Nishida, Naoki Goshima, Nobuo Nomura, and Ichio Shimada
EMBO J., in press.
9)Koh Takeuchi, Mariko Yokogawa, Tomoki Matsuda, Mariko Sugai, Seiko Kawano, Toshiyuki Kohno,
Haruki Nakamura, Hideo Takahashi, and Ichio Shimada Structure (2003) 11, 1381-1392.
10)Mitsuhiro Takeda, Shinji Ogino, Ryo Umemoto, Masayoshi Sakakura, Masahiro Kajiwara, Kazuki
Sugahara, Hiroto Kawashima, Masayuki Miyasaka, Hiroaki Terasawa, and Ichio Shimada
J Biol Chem. (2006) 281, 40089-40095.
◆略 歴◆ 嶋田 一夫 (Ichio SHIMADA):
1980年 東京大学大学院理学系研究科生物化学専攻修士課程 修了 1980年 (株)東レリサーチセンター・構造化学研究部 入社 1987年 (財)東京都臨床医学総合研究所・生理活性物質研究部 入所 1989年 東京大学薬学部薬品物理化学講座 1994年より現所属(教授) 2001年 産業技術総合研究所・生物情報解析研究センター・分子認識解析チーム
チームリーダー 併任

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 33
薬学研究ビジョン部会からのお知らせ
第 5 回(平成 19 年度)薬学研究ビジョン部会 部会賞募集
本年度は下記の要領で部会賞候補者を募集しますので、応募・推薦(自薦・他薦)をお願いいたし
ます。薬学のもつ広い研究領域を横断的に活動する本薬学研究ビジョン部会の部会賞は,薬学を担う
研究者を奨励する本部会独自の賞です。本部会賞は,薬学の横断的な分野から選ばれた選考委員が選
考に当たり,真に薬学研究として有望なものに与えられる賞です。多数のご応募をお願い致します。
平成 19 年度部会賞候補者募集要項
1)推薦者 自薦・他薦 いずれも可。
2)受賞の対象 薬学研究に関する明確なビジョンをもち、
創薬を目指して境界領域において独創的な研
究業績をあげつつあり、薬学の将来を担うこ
とが期待される研究者に授与する。日本薬学
会奨励賞の受賞者で、その受賞対象の主要部
分が同一の場合は受賞対象としない。受賞年
度の 4 月 1 日に満 45 歳以下で、原則本部会会
員であること。 3)授賞件数
数件 4)表彰
受賞者には、賞状(楯)を贈る。 5)推薦締切
平成 19 年 10 月 5 日(金)必着 6)推薦方法
推薦理由書(自薦・他薦いずれの場合も、
500-1000 字程度)および業績リストを所定の
書式(部会ホームページ
http://bukai.pharm.or.jp/bukai_vision/
award/award.html からダウンロード)に準じて
作成し、電子メールの添付書類として平成 19年度部会賞選考委員長に送付すること。
7)授賞式 平成 20 年度 1 月 24 日(第 9 回創薬ビジョ
ンシンポジウム時を予定) 8)備考
受賞者には、授賞式時に受賞講演をお願い
するとともに、その研究業績をまとめて部会
ニュース Pharma VISION NEWS の執筆をお願
いすることもある。 9)推薦書類提出先
(電子メールの添付書類としてお送りくださ
い) 薬学研究ビジョン部会 平成19年度部会賞選考委員長 大和田 智彦 E-mail: [email protected]
平成 19 年度部会長 横井 毅 平成 19 年度部会賞選考委員長 大和田 智彦
第 9 回創薬ビジョンシンポジウム
「臨床開発に役立つバイオマーカー選択の科学」
日 時 :平成 20 年 1 月 24 日(木)〜25 日(金) 会 場 :東京大学医学部鉄門記念講堂 Co-Chairs :鈴木 洋史(東京大学医学部附属病院薬剤部) 三橋 晴美(サノフィ・アベンティス) プログラムの詳細が決定しましたら部会ホームページにてお知らせします。 http://bukai.pharm.or.jp/bukai_vision/sympo/sympo.html

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 34
編 集 後 記
長洲 毅志 エーザイ株式会社
Pharma VISION NEWS 編集委員長
ファーマビジョンニュース 10 号をお届けしま
す。今回は巻頭言から、「創薬」についてストレ
ートに語っていただきました。今回のトピックに
なっている計算化学や構造生物学ばかりでなく
「オミクス」も含めて創薬に欠かせぬ技術は飛躍
的に進歩したと感じております。一方で薬に求め
られる資質も安全性、薬剤相互作用など多様な面
から規定され新薬創出の困難性が増しています。
創薬に必要な研究リソースばかりが増加してよ
い薬は一向に増えない、成功確率は上がらない、
という現状なのです。 一つのソリューションが個の医療(いわゆるオ
ーダーメード)の方向とそれを支えるバイオマー
カ(診断)をセットにした創薬といわれています。
CYP 多型は明らかに特定の薬剤の投与量を個別
に規定することが可能でその実現は安全性の面
からも非常に重要です。 しかし一足飛びに、創薬を個の医療に向けると
いう考え方には無理があります。そういうとお叱
りを受けそうですが、個の医療を否定しているの
ではなく、まず疾病をもっと現代科学で再定義・
分類すべきだと思うのです。癌で言えば、大腸癌
や小細胞肺癌という所見だけでなく、ras 活性化、
p53 欠失、APC 欠失の癌・・・という規定をする
ことによって分子標的薬という概念やバイオマ
ーカという概念がサイエンスの場に下りてくる
のではないでしょうか。(分子標的がそのまま適
応症に対応するということになります。)個人的
には、なるべく多くの方に同じ薬剤で効くものな
らそういう薬をつくりたいと考えます。薬物代謝
は既に特定の CYP 多型の影響を受けないデザイ
ンという考え方で研究が進められています。そし
て、疾病が分子生物学的に定義されれば、その疾
病を診断するバイオマーカが導き出されその疾
病をカバーする薬が(分子標的薬として)デザイ
ンできるのではないかと思うのです。考えてみれ
ばこれはまったく当たり前のことで言うまでも
ないことなのかも知れません。 いたずらにゲノム創薬ということはなくなりま
したが、それがバイオマーカ、オーダーメードと
形を変えて言葉だけ先行しているのではないか
とふと感じた次第です。 このビジョン部会では、本当に今後の創薬は
こうあるべきという根本の議論からそれを支え
る新技術まで幅広く語り学べる場であると感じ
ております。皆様のご賛同とご支援をお願い申し
上げます。(長洲・記)

社団法人 日本薬学会 薬学研究ビジョン部会
Pharma VISION NEWS No.10 (September 2007) 35
薬学研究ビジョン部会 常任世話人
大和田 智彦 【部会賞選考委員長】 東京大学大学院薬学系研究科
小澤 正吾 厚生労働省 国立医薬品食品衛生研究所
片倉 晋一 第一製薬株式会社
鈴木 洋史 【副部会長】 東京大学医学部付属病院
辻本 豪三 京都大学大学院薬学研究科
長洲 毅志 【編集委員長】 エーザイ株式会社
長瀬 博 【編集副委員長】 北里大学薬学部
西島 和三 持田製薬株式会社
松崎 勝巳 京都大学大学院薬学研究科
三橋 晴美 【部会賞選考副委員長】 サノフィ・アベンティス株式会社
南野 直人 国立循環器病センター研究所
横井 毅 【部会長】 金沢大学薬学部
編集委員会からのお知らせ
この Pharma VISION NEWS は、本部会が年 2
回の予定で部会員宛にメール発信いたします。
ご希望の方は、薬学研究ビジョン部会事務局宛
にお問合せ下さい。 部会員登録が必要です。部会員登録用紙は、部
会 HP から PDF ファイルをダウンロードして
下さい 部会員の登録には、入会金・年会費は無料です。
日本薬学会の会員でなくても部会委員登録は
できます。 投稿原稿を募集いたします。詳細は、編集事務
局にお問合せ下さい。
発行:薬学研究ビジョン部会【部会長:横井 毅】
編集委員会: 長洲 毅志【委員長】,長瀬 博【副委員長】 鈴木 洋史 ,辻本 豪三 甲斐 俊次 ,曽我 公美子【編集事務局】 編集事務局: 甲斐 俊次 横浜薬科大学 薬品反応学研究室 〒245-0066 神奈川県横浜市戸塚区俣野町 601 TEL:045-859-1300 FAX : 045-859-1301 曽我公美子 エーザイ株式会社 創薬研究本部 〒300-2635 茨城県つくば市東光台 5-1-3 TEL:029-847-5603 FAX:029-847-1006 薬学研究ビジョン部会事務局: ※お問合せ、登録内容変更等のご連絡はこちらへ 金沢大学薬学部 薬物代謝化学研究室内 〒920-1192 金沢市角間町 TEL:076-234-4438 FAX:076-234-4407 E-mail:[email protected]
※本誌全ての記事、図表等の無断複写・転写を禁止いたします。