第十三章 程序性细胞死亡与细胞衰老
DESCRIPTION
第十三章 程序性细胞死亡与细胞衰老. ● 程序性细胞死亡 ● 细胞衰老. 第一节 程序性细胞死亡. ● 程序性细胞死亡的涵义 ● 动物细胞的程序性死亡 ● 植物细胞与酵母细胞的 程序性死亡. 一、动物细胞的程序性死亡. ● 细胞凋亡 (apoptosis) ● 细胞坏死 (necrosis) ● 细胞自噬 (autophagy). 第二节 细胞衰老 (cell aging or cell senescence). ● 细胞衰老的概念 ● 细胞衰老的研究 ● 衰老细胞形态结构的变化 ● 细胞衰老的分子机制 ● 细胞在体内条件下的衰老 - PowerPoint PPT PresentationTRANSCRIPT

第十三章程序性细胞死亡与细胞衰老
● 程序性细胞死亡
● 细胞衰老

第一节 程序性细胞死亡
● 程序性细胞死亡的涵义● 动物细胞的程序性死亡● 植物细胞与酵母细胞的
程序性死亡

一、动物细胞的程序性死亡
● 细胞凋亡 (apoptosis)
● 细胞坏死 (necrosis)
● 细胞自噬 (autophag
y)

第二节 细胞衰老(cell aging or cell senescence)
● 细胞衰老的概念
● 细胞衰老的研究● 衰老细胞形态结构的变化
● 细胞衰老的分子机制
● 细胞在体内条件下的衰老
● 细胞衰老与个体衰老的关系

二、细胞衰老的研究
● 细胞不死说● Hayflick 界限

程序性细胞死亡的涵义
不论是单细胞生物还是多细胞生物,其细胞死亡往往受到细胞内某种由遗传机制决定的“死亡程序”所控制,称为程序性细胞死亡。 ( programmed cell death,PCD )

一、细胞衰老的概念 细胞衰老( cell aging )一般的含义是复制衰老 (replicative senescene , RS) 。 指体外培养的正常细胞经过有限次数的分裂后,停止分裂 , 细胞形态和生理代谢活动发生显著改变的现象。

细胞不死说◆1892 年,魏斯曼提出 : 种质不死而体质会衰老和死亡的学说 原生动物细胞“不死性”
◆ 证据 原生动物某些无性系可以长期保持很高的分裂
速度
◆ 证据不足 被废弃。某些无性系的稳定性的观点
细胞不死说◆1912 年 , Alexis Carrel (Nobel Laureate)
细胞“不死性”的观点 卷土重来
◆ 证据 在实验室培养鸡心脏细胞可以无限制地生长和分裂。(连续培养了 34年)

细胞不死说◆20 世纪 40-50 年代, L 系小鼠和 HeLa 细胞系
的建立。(可以一直分裂、存活下去)
◆使细胞“不死性”的观点的统治地位更加巩 固了。

细胞不死说
细胞本身并没有衰老和死亡,衰老只是一种多细胞现象,多细胞生物体内细胞的衰老,起因不在细胞本身,而是由于体内、体外环境的影响。

细胞不死说
谁动摇了这种学说?

◆Hayflick 认为Carrel 的实验不能被重复 , 可能是由于混入了新鲜的细胞。
HeLa 癌细胞和培养的 L 细胞系是不正常细胞,其染色体数目或形态已经不同于原先的细胞。
◆Hayflick 提出
细胞增殖能力和寿命是有限的观点

一、 Hayflick 界限 (Hayflick Limitation )
◆Hayflick 界限的概念 细胞,至少是培养的二倍体细胞,不是不死的,而
是有一定的寿命;它们的增殖能力不是无限的,而是有一定的界限,这就是 Hayflick界限。
◆Hayflick 界限的证据 细细细细细细细细细细细细细细
细细细细细细细细细细细细细细细细细细细细细

一、 Hayflick 界限(Hayflick Limitation )
◆决定细胞衰老的因素 二倍体细胞的衰老是由细胞自身决定的
细细细细细细细细细细细细细细细细细细细细 细细细
细细细细细细细细细细细细细细细细细

细胞衰老概念被普遍接受的一些经典实验
( 细胞体外生命跨度与供体年龄的正相关性 ) 过程: 年轻女性细胞 (间期可见巴氏小体 ) + 年老男性细胞 (间期无巴氏小体 )
混合培养 (一样的外部环境 ) 结果:年轻女性细胞倍增次数多于年老男性细胞
结论:细胞衰老的过程是由细胞自身决定的 , 而非环境因素
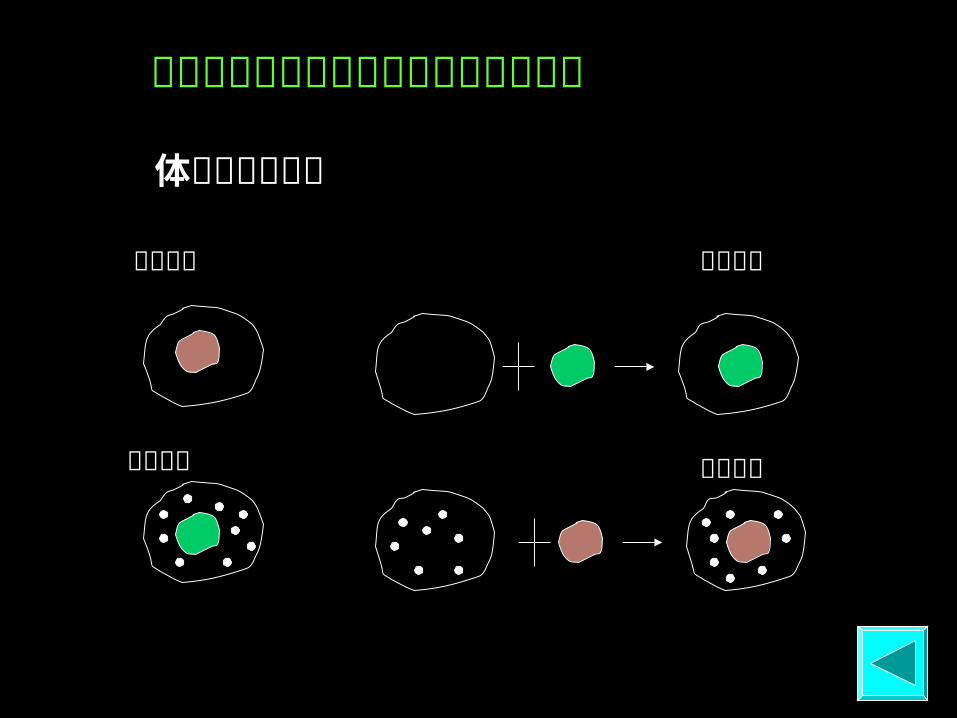
细胞衰老是由细胞核而非细胞质决定的
体细胞杂交实验
年老细胞
年轻细胞
年轻细胞
年老细胞

细胞来源 人胚肺成纤维 细 胞
中年人成 纤维细胞
老年人成 纤维细胞
可 增 殖代 数 40 ~ 60 20 2 ~ 4
不同年龄来源的人成纤维细胞的增殖代数
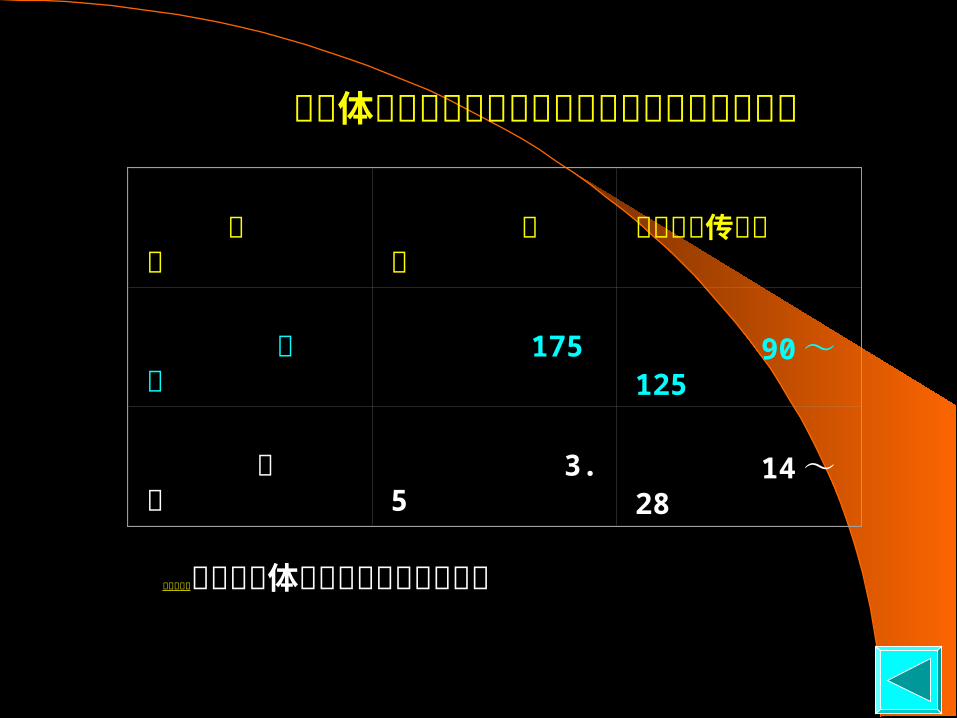
1. 细胞体外生命跨度与物种平均生命跨度的正相关性
物 种 寿 命 培养细胞传代数
海 龟 175 90 ~ 125
小 鼠 3. 5 14 ~ 28
早老症病人的细胞在体外只有较短的生命跨度

六、细胞衰老与个体衰老的关系 呈正相关性 :
◆成人的细胞能在体外传代的次数比胎儿 的细胞少 ; ◆早老症病人的细胞能在体外传代的次数 比正常人少 ;◆寿命长的物种的细胞能在体外传代的次 数比寿命短的物种的细胞多。

五、细胞在体内条件下的衰老
● 在机体内,细胞的衰老和死亡是常见的现象,甚至在个体发育的早期也会发生 ;
● 正常情况下终生保持分裂的细胞,其分裂能力是否随着有机体年龄的增高而下降?它们会不会衰老?
◆ 衰老动物体内,细胞分裂速度显著减慢,其原因主要是 G1期明显延长 ;
◆衰老个体内的环境因素影响了细胞的增殖和衰老 ;◆骨髓干细胞移植实验说明随着年龄的增加,干细胞增殖速度
也趋缓慢 .

三、衰老细胞形态结构的变化● 细胞核的变化
● 内质网的变化 :
衰老动物内质网成分弥散性地分散于核周胞质中,粗面内质网的总量似乎是减少了
● 线粒体的变化 :
通常,细胞中线粒体的数量随龄减少,而其体积则随龄增大
● 致密体的生成● 膜系统的变化

细胞核的变化◆ 体外培养的二倍体细胞,细胞核
随着细胞分裂次数的增加不断增大
◆ 细胞核的核膜内折( invagination )、
染色质固缩化

膜系统的变化
◆衰老的细胞,其膜流动性降低、韧性减小
◆衰老细胞间间隙连接减少 ;
细胞膜内( P 面)颗粒的分布也发生变化细

四、细胞衰老的分子机制
● 复制衰老的机制 端粒与衰老● 胁迫诱导的早熟性衰老 氧化性损伤学说● 单细胞生物的衰老

结 论细胞衰老◆ 可能是以上综合因素的集合,或某
几种因素的集合,其中有一种起主导作用。
◆ 对于不同的生物体可能有不同的导致衰老的诱因和相应的机制。

端粒与衰老◆1990 年 Harley 发现端粒长度确实与衰老有着密切的关系,
提出细胞衰老的“有丝分裂钟”学说。
◆“ 有丝分裂钟”学说认为,随着细胞的分裂,染色体末端的
(端粒)不断缩短,当其长度缩短到一个阈值时,细胞进
入衰老。
细胞每分裂一次,端粒缩短 50~ 100bp 端粒缩短到一定程度,细胞不再分裂 端粒酶过量表达能延长细胞的生命跨度

(一) 细胞凋亡 (Apoptosis)
● 细胞凋亡的概念● 细胞凋亡的生理意义● 细胞凋亡的特征● 细胞凋亡的检测方法● 诱导细胞凋亡的因子● 细胞凋亡的分子机制

1.细胞凋亡的概念
细胞凋亡是一种程序性的细胞死亡方式,受基因调控的主动的生理性细胞自杀行为,凋亡细胞最后以凋亡小体的形式被吞噬消化。

2.细胞凋亡的生物学意义● 细胞凋亡对于多细胞动物个体的正常发 育,自稳态的维持,免疫忍受的形成,肿 瘤的监控具有重要意义。
◆ 蝌蚪尾的消失,骨髓和肠的细胞凋亡 ◆ 发育过程中手和足的成形过程 ◆ 脊椎动物的神经系统的发育
● 细胞凋亡失衡会引发多种疾病。 ◆ 细胞凋亡不足 ◆ 细胞凋亡过度

3.细胞凋亡的特征
● 细胞凋亡的形态学特征
● 细胞凋亡的生化特征

3.1 细胞凋亡的形态学特征 动物细胞凋亡的过程可以分为两个阶段:
◆激活期( activation phase) 在这一阶段,细胞对要求它进行自我毁灭的 “ 死亡信号”作出反应。
◆执行期( execution phase) 细胞死刑此时付诸实施。我们先来看比较容易理解的执行期,由凋亡引起的细胞死亡是一个干净利落的有序过程其特征表现为:

3.1 细胞凋亡的形态学特征◆凋亡的起始:细胞表面的特化结构如微绒毛消失,细胞间接触的消失,但细胞膜依然完整;线粒体大体完整,但核糖体逐渐从内质网上脱离,内质网囊腔膨胀,并逐渐与质膜融合;染色质固缩,形成新月形帽状结构等形态,沿着核膜分布
◆凋亡小体的形成:核染色质断裂为大小不等的片段,与某些细胞器如线粒体一起聚集,为反折的细胞质膜所包围。细胞表面产生了许多泡状或芽状突起,逐渐
形成单个的凋亡小体
◆凋亡小体逐渐为邻近的细胞吞噬并消化

3.2 细胞凋亡的生化特征
◆ 细胞凋亡的主要特征是形成大小为
180 ~ 200bp特征性的 DNA ladders。
◆ 凋亡细胞组织转谷氨酰胺酶 tTG
( tissueTransglutaminase )积累并达到
较高水平。

5. 诱导细胞凋亡的因子◆ 物理性因子
包括射线(紫外线, 射线等),较温和的温度
刺 激(如热激,冷激)等。
◆ 化学及生物因子
包括活性氧基团和分子, DNA 和蛋白质合成的抑
制剂,激素,细胞生长因子,肿瘤坏死因子( TN
F ),抗 Fas/Apo-1/CD95抗体等。

4.4. 细胞凋亡的检测方法细胞凋亡的检测方法◆ 形态学观测:染色法、透射和扫描电镜观察
◆DNA电泳: DNA片段就呈现出梯状条带
◆TUNEL测定法 :即 DNA 断裂的原位末端标记法
◆彗星电泳法( comet assay )
◆流式细胞分析 根据凋亡细胞 DNA 断裂和丢失,采用碘化丙啶使 DNA
产生激发荧光,用流式细胞仪检出凋亡的亚二倍体细
胞,同时又能观察细胞的周期状态。

6.细胞凋亡的分子机制
● 线虫(C.elegans)
●Caspase家族与凋亡
●Bcl-2家族与细胞凋亡
● 细胞凋亡的分子机理
细胞凋亡受基因控制。线虫是研究细胞凋亡分子基础的理想材料。 每条线虫具有 1090 个细胞,其中在正常情况下有 131 个细胞在发育过程中以凋亡的方式死亡。从线虫中找到若干控制细胞凋亡的基因。
6.1 线虫 (C.elegans)
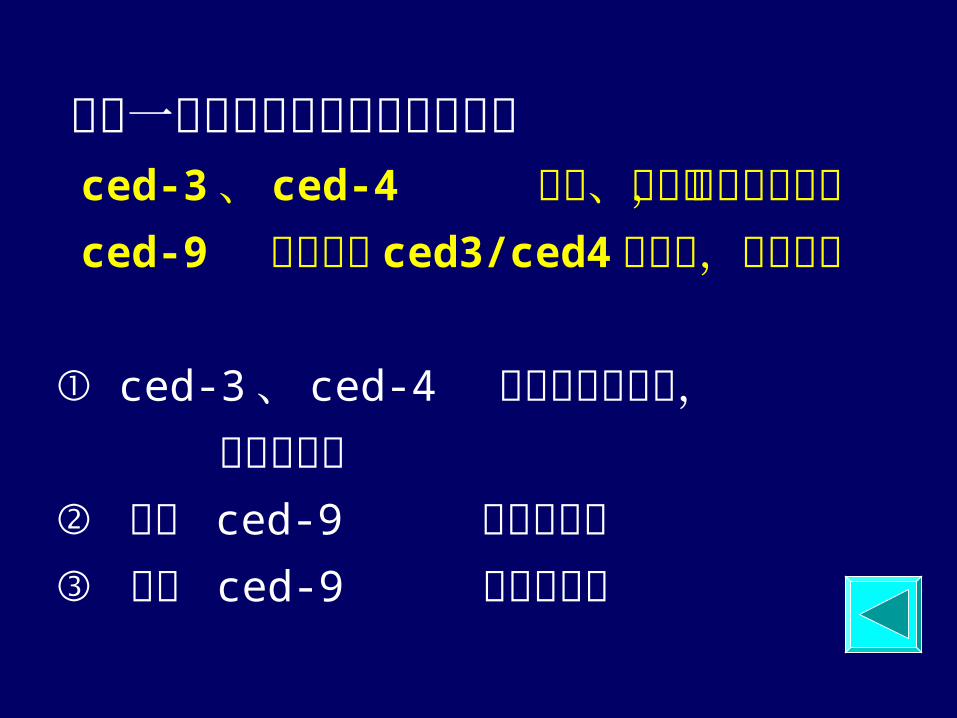
找到一系列与细胞凋亡有关的基因 ced-3、 ced-4 诱发、启动,促进细胞凋亡 ced-9 基因阻止 ced3/ced4的激活,抑制凋亡
① ced-3 、 ced-4 基因突变或缺失, 使凋亡受阻 ② 移入 ced-9 使凋亡受阻 ③ 失去 ced-9 使细胞凋亡

6.2 Caspase家族与凋亡
◆ Caspase家族
◆ Caspase激活
◆ Caspase水解的靶蛋白
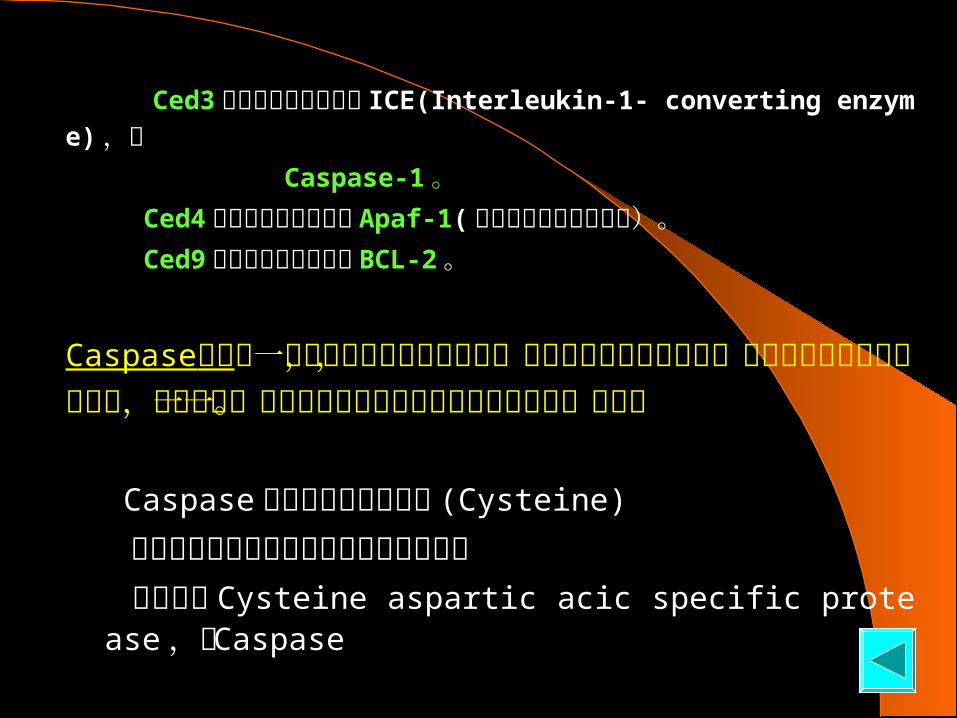
Ced3 的哺乳类同源基因是 ICE(Interleukin-1- converting enzyme) ,即 Caspase-1 。 Ced4 的哺乳类同源基因是 Apaf-1( 细胞凋亡蛋白活化因子)。 Ced9 的哺乳类同源基因是 BCL-2 。
Caspase家族是一组与众不同的蛋白水解酶,在凋亡早期阶段被激活,并负责引发随后的凋亡变化,通过切割一组高度选择性的必须蛋白质来实现这一功能。
Caspase 活性位点是半胱氨酸 (Cysteine)
裂解靶蛋白位点是天冬氨酸残基后的肽键 因此称为 Cysteine aspartic acic specific protease ,即 Caspase

Caspase激活
细胞中合成的 Caspase 自身以无活性的酶原状态( Procaspase )存在,经激活后方能执行其功能。 其激活依赖于其他的 Caspase 在它的天冬氨酸位点水解活化或自身活化

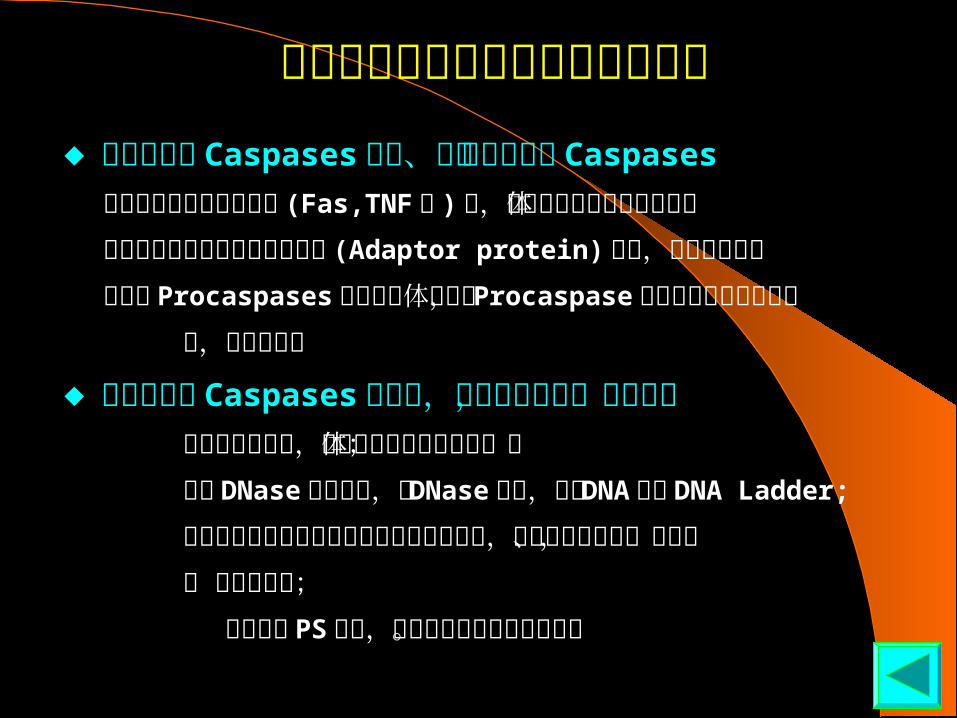
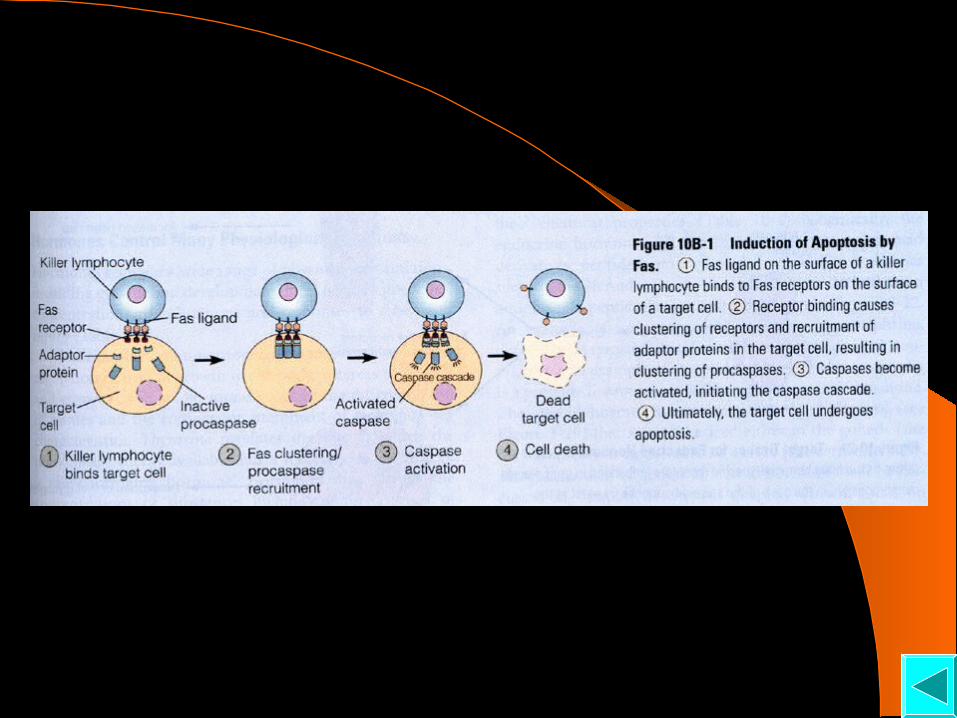
胞外信号分子诱导的细胞凋亡途径
◆上游启始者 Caspases 切割、活化下游执行者 Caspases
当细胞接受凋亡信号分子 (Fas,TNF 等 ) 后,凋亡细胞表面信号分子受体 相互聚集并与细胞内的衔接蛋白 (Adaptor protein)结合,这些衔接蛋白 又募集 Procaspases聚集在受体部位, Procaspase 相互活化并产生级联反 应,使细胞凋亡
◆下游执行者 Caspases 活化后,水解蛋白底物,启动凋亡 裂解核纤层蛋白,导致细胞核形成凋亡小体; 裂解 DNase结合蛋白,使 DNase释放,降解 DNA 形成 DNA Ladder;
裂解参与细胞连接或附着的骨架和其他蛋白,使凋亡细胞皱缩、脱落, 便 于细胞吞噬;
导致膜脂 PS 重排,便于吞噬细胞识别并吞噬。

6.3 Bcl-2家族与细胞凋亡
◆Bcl-2
是一种原癌基因,是 ced-9 在哺乳类中的同源物,能抑制细胞凋
亡;与线粒体及内质网膜相结合; Bcl-2蛋白的羧基末端有一穿
膜的结构域; Bcl-2家族成员的基因中,常常含有三个保守的
Bcl-2同源区,即 BH1 , BH2 和 BH3
◆Bcl-2蛋白质家族成员以不同的方式调节细胞凋亡 促进凋亡 : Bad 、 Bid 、 Bax
阻止凋亡: Bel-X 、 Bel-w 、 Bel-2
◆哺乳动物细胞中发现的 Apaf2即是 CytC
6.4 细胞凋亡的分子机理
◆ 胞外信号分子诱导的细胞凋亡途径
◆ 胞内信号刺激诱导的细胞凋亡途径

Bcl-2家族凋亡成员与细胞凋亡



(二)细胞坏死 (necrosis)
● 细胞坏死的涵义及特点● 细胞凋亡与坏死的区别

(二)细胞凋亡与坏死的区别◆细胞凋亡过程中,细胞质膜反折,包裹断裂的染 色质片段或细胞器,然后逐渐分离,形成众多的凋亡小体( apoptotic bodies ),凋亡小体则为邻近的细胞所吞噬。整个过程中,细胞质膜的整合性保持良好,死亡细胞的内容物不会逸散到胞外环境中去,因而不引发炎症反应。◆相反,在细胞坏死时,细胞质膜发生渗漏,细胞内容物,包括膨大和破碎的细胞器以及染色质片段,释放到胞外,导致炎症反应。

(三)细胞自噬 (autophagy)
● 细胞自噬的涵义及特点● 细胞自噬的分子机制

二、植物细胞与酵母细胞 的程序性死亡
●植物细胞的程序性死亡●酵母细胞的程序性死亡




名称及其别名 底物
caspase-1(ICE) Pro-IL ; pro-caspase 3,7
caspase-2(Nedd-2/ICH1)
caspase-3(apopain/CPP32/Yama) PARP ; SREBP ; DFF ; DNA-PK
caspase-4(Tx/ICH2/ICE rel-II)
caspase-5(ICE rel-III/TY)
caspase-6(Mch2) Lamin A ; keratin 18
caspase-7(ICE-LAP3/Mch3/CMH-1) PARP ; pro-caspase 6 ; DFF
caspase-8(FLICE/MACH/Mch5)
caspase-9(ICE-LAP6/Mch6)
caspase-10(Mch4/FLICE2) PARP
caspase-11(ICH3)
caspase 超家族成员及其相应底物

BCL-2家族成员


坏死细胞 凋亡细胞


细胞色素 c 诱导的凋亡细胞 DNA电泳图
1 .细胞色素 c 诱导 0 h2 .细胞色素 c 诱导 1 h3 .细胞色素 c 诱导 2 h4 .细胞色素 c 诱导 3 h5 .细胞色素 c 诱导 4 h6 .阴性对照7 . Marker ( 自赵允、翟中和)

自由基理论
1956 年 Harman 提出来的
生物氧化中产生自由基,自由基破坏生物大分子——蛋白质、核酸、脂类等。使得细胞结构破坏,基因突变,导致细胞衰老。

自由基
自由基是一类瞬时形成的含不成对电子的原子
或功能基团,普遍存在于生物系统。
主要包括:氧自由基(如羟自由基 ·OH )、氢
自由基( ·H )、碳自由基、脂自由基等,其中 ·O
H
的化学性质最活泼。

自由基产生的原因 人体内自由基的产生有两方面: 一是环境中的高温、辐射、光解、化学物质等引起
的外源性自由基。 二是体内各种代谢反应产生的内源性自由基。内源
性自由基是人体自由基的主要来源,其产生的主要途径有:①由线粒体呼吸链电子泄漏产生;②由经过氧化物酶体的多功能氧化酶(MFO)等催化底物羟化产生。

自由基对细胞的危害
对许多生物功能是非常重要的,有些学者认为没有自由基的生物不能生存。

自由基对细胞的危害如果细胞中的自由基过多,就会对许
多细胞组分造成损伤。
自由基含有未配对电子,具有高度反应活性,可引发链式自由基反应,引起 DNA 、蛋白质和脂类,尤其是多不饱和脂肪酸等大分子物质变性和交联,损伤 DNA 、生物膜、重要的结构蛋白和功能蛋白,从而引起衰老各种现象的发生。

细胞内清除自由基的机制细胞内部隔离化使产生自由基的位点与
细胞的其他组分分开酶系统——保护性的酶 超氧化物歧化酶( SOD ) 过氧化氢酶( CAT )非酶系统——其他抗氧化物分子 VE 、 VC 、醌类物质等电子受体

两种不同观点支持者 细胞衰老的 90%的原因归咎于自由基
反对者 大多数自由基存在的时间很短,对细胞危害不大。 细胞内有抗氧化剂,来对抗自由基的危机。 增加食物中的 VE ,并不能抵抗自由基的有害作
用,反而会抑制细胞中其他抗氧化剂的产生。