capitol ul 5
DESCRIPTION
.....TRANSCRIPT

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
Universitatea din Bucureşti, Facultatea de Biologie Splaiul Independenţei 91-95, 76201 Bucureşti
Bacteriile aparţinând genurilor Bacillus şi Streptomyces sunt cunoscute ca fiind producători ai unor game largi de compuşi biologic activi, dintre care unii au aplicaţii în agricultură şi industria farmaceutică. Dacă în cazul bacililor cunoştinţele sunt mai avansate, iar aplicaţiile practice concentrate mai ales asupra enzimelor, la streptomicete cele mai multe cunoştinţe au fost dobândite în urma studiilor asupra producerii de antibiotice. Scopul acestei lucrări este de a sintetiza datele referitoare la biosinteza de enzime şi de compuşi cu efect inhibitor, utili pentru diferite procese biotehnologice. De asemenea, sunt prezentate unele date referitoare la genele ce codifică biosinteza unor substanţe importante din punct de vedere biotehnologic.
Actinomicetele reprezintă un grup de microorganisme procariote cu numeroase asemănări cu fungii microscopici, alături de care constituie un exemplu clasic de evoluţie convergentă. Relevante în acest sens sunt: morfologia şi organizarea micelială a coloniilor lor ca şi sistemul lor de reproducere. Se poate specula că aceste asemănări rezultă dintr-o adaptare comună în sensul desfăşurării tuturor funcţiilor metabolice în condiţiile absenţei apei în imediata lor vecinătate.
Ciclul de viaţa al streptomicetelor, cel mai important gen din cadrul grupului actinomicetelor, crescute pe mediu solid implică formarea (diferenţierea) a două tipuri diferite de miceliu: miceliul de substrat care creşte în si pe mediul de cultură solid; miceliul aerian care creşte pe primul şi pe care se formează sporii (Figura 1).
In timpul creşterii miceliului de substrat se remarcă o creştere simultană a greutăţii uscate care însă încetineşte semnificativ odată cu formarea miceliului aerian.
Figura 1. Reprezentarea schematică a celor două tipuri de miceliu (după Zarnea, 1982).
123

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
Un aspect interesant este acela că, la multe dintre speciile de streptomicete, încetinirea ratei de creştere este însoţită de producerea de substanţe antibiotice sau cu alte funcţii (metaboliţi secundari), astfel că în timpul formării miceliului aerian se remarcă o foarte intensă activitate antibiotică. Acest fapt vine în sprijinul ipotezei asupra rolului antibioticelor produse de streptomicete şi anume acela de a proteja miceliul de substrat care se lizează împotriva altor bacterii existente în sol care l-ar putea folosi ca sursă de carbon şi azot, păstrându-l astfel pentru creşterea miceliului aerian (Chater şi Merrick, 1979).
Diferenţierea celulară evidenţiată la streptomicete pare a fi un proces controlat de unele substanţe autoreglatoare cum sunt factorii A şi B al căror mecanism de acţiune, cel puţin în cazul unor specii de streptomicete care au fost studiate mai intens, este asemănător mecanismului de acţiune al hormonilor de la eucariote, implicând prezenţa unor receptori specifici (Beppu, 1986). Acest tip de reglaj împreună cu observaţiile legate de asemănările structurale ale factorului B cu AMPc de la eucariote sugerează ideea că anumite substanţe evidenţiate la streptomicete ar putea fi semnale universale sau mesageri pentru funcţii celulare nu numai la procariote ci şi la eucariote.
Ca rezultat al activităţii lor metabolice, atât streptomicetele cât şi bacilii, formează mai multe categorii de compuşi care includ, pe lângă constituenţii structurali ai celulei, enzime intra- şi extracelulare, produşi finali de metabolism precum şi o serie de produşi cu importanţă practică deosebită incluzând metaboliţii primari şi pe cei secundari.
Metaboliţii primari sunt compuşi cu greutate moleculară în general mică, reprezentând produşi intermediari sau finali ai metabolismului intermediar necesari microorganismului producător şi care includ : aminoacizi, acizi organici, nucleotide, vitamine, enzime etc (Zarnea, 1984). Aceşti compuşi sunt sintetizaţi în cursul trofofazei (perioadei de multiplicare rapidă şi acumulare de biomasă celulară), în medii bogate în substanţe nutritive şi condiţii fizice corespunzătoare. După această etapă urmează idiofaza în cursul căreia creşterea şi multiplicarea încetează dar celulele rămân metabolic active sintetizând o categorie specială de substanţe cu structură chimică variată, denumite metaboliţi secundari. Numărul metaboliţilor secundari este foarte mare iar efectele lor sunt de asemenea foarte variate. Diversitatea neobişnuită a efectelor farmacodinamice şi a structurii lor chimice este ilustrată de numeroasele clase de compuşi organici la care pot aparţine şi care includ: glucide aminate, chinone, cumarine, epoxizi, alcaloizi, glicozide, derivaţi indolici, lactione, macrolide, naftalene, nucleozide, terpenoizi şi tetracicline (Berdy, 1974).
Sinteza metaboliţilor secundari se face printr-o mai mare varietate de căi decât aceea a metaboliţilor primari, pe seama compuşilor cu greutate moleculară mică folosiţi în timpul creşterii exponenţiale pentru sinteza constituenţilor celulari normali şi este rezultatul acumulării în mediu a unor substanţe cu rol de inductor. De asemenea, declanşarea sintezei metaboliţilor secundari se poate datora şi epuizării unor substanţe cu funcţie de represor (Vining, 1990). In contrast cu diversitatea căilor de biosinteză şi a produşilor finali, cei mai mulţi dintre metaboliţii secundari sunt asamblaţi pornind de la câţiva metaboliţi cheie. In plus, spre deosebire de metabolismul primar în care procesele de biosinteză se desfăşoară întotdeauna cu mare specificitate (se utilizează un singur substrat şi rezultă un singur produs), în metabolismul secundar proporţia fiecărui component depinde de factori genetici şi de mediu, probabil datorită specificităţii scăzute a enzimelor implicate în desfăşurarea sa.
Majoritatea metaboliţilor secundari au acţiuni antibiotice dar există şi compuşi cu acţiuni diferite. In general ei au mărime redusă chiar dacă nu sunt monomeri ci oligomeri; ei sunt de obicei excretaţi în mediul extern fie sub formă solubilă în apă fie ca produşi insolubili care formează agregate amorfe sau cristaline asociate cu celulele producătoare (Vining, 1990).
124

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
1. Functiile metabolitilor secundari produşi de bacili şi streptomicete
1.1. Activitatea antibiotică a metaboliţilor secundari
Printre bacterii, actinomicetele şi în special cele din genul Streptomyces sunt cunoscute ca fiind principalii producători de antibiotice, sintetizând 3/4 dintre toţi produşii cunoscuţi de acest tip. Antibioticele produse de streptomicete au structură chimică variată putând fi clasificate în diferite tipuri în timp ce compuşii cu funcţii similare elaboraţi de bacteriile din genul Bacillus sau din alte grupe (bacterii lactice, enterobacterii, Pseudomonas sp.) sunt în majoritate peptide sau peptide modificate.
Membrii genului Bacillus (Figura 2) sunt capabili să producă antibiotice ca metaboliţi secundari în faza de creştere logaritmică târzie sau la începutul fazei staţionare. Au fost semnalaţi mai mult de 169 de astfel de metaboliţi. De exemplu, B subtilis poate produce 68 tipuri de antibiotice, în timp ce la B.brevis au fost identificate numai 23 asemenea tipuri de substanţe.(Katz şi Demain 1987).
Figura 2. Aspect electronomicroscopic al celulelor de Bacillus subtilis (după Zarnea, 1984).
Multe dintre antibioticele produse de bacili sunt active împotriva bacteriilor Gram pozitive, dar există şi excepţii. Aşa cum s-a menţionat deja, majoritatea antibioticelor produse de bacili sunt peptide, dar unele aparţin altor clase de compuşi chimici (de exemplu, leutirosin este o amiloglicozidă, iar protacin este o trienă fosforică).
Există o controversă în ceea ce priveşte funcţia acestor antibiotice la bacteriile din genul Bacillus. Ele apar, de obicei, în timpul sporutării, de aceea se crede că antibioticele respective constituie un factor important ce apare la tranziţia de la starea vegetativă la spori (Katz şi Demain,1987).
Multe dintre tulpinile de bacili producătoare de compuşi cu acţiune antimicrobiană au fost supuse unor analize speciale de cartare moleculară, de identificare şi clonare a genelor codificate. Datele obţinute până în prezent sunt prezentate în Tabelul 1. Metaboliţii secundari datorează activitatea antibiotică capacităţii lor de a inhiba procesele metabolice primare. Cei mai mulţi acţionează ca antimetaboliţi deoarece asemănarea lor funcţională cu metaboliţii normali le permite legarea la situsuri ţintă şi să interfere cu activitatea normală. Produşii generaţi de o anumită cale metabolică inhibă de obicei o altă cale metabolică, activitatea lor depinzând în întregime de configuraţia antibioticului şi de distribuţia grupărilor funcţionale la nivelul lor.
125

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
Tabelul 1. Substanţe antibiotice produse de bacteriile genului BacillusSpecie Antibiotic Acţiune inhibitoare Harta
geneticăGene
clonateB.subtilis Subtilin Bacterii Gram + + +
Surfactin + -Bacilysin Bacterii Gram + + -Difficidin Spectru larg de acţiune - -
Oxydificidin Spectru larg de acţiune - -Bacillomycin F Antifungic - -Mycobacillin Antifungic - -
Iturin AntifungicB. brevis Gramicidin S Bacterii Gram + + +
Tyrocidin Bacterii Gram + + +B. licheniformis Bacitracin Bacterii Gram + + +
Proticin - -B. pumilus Pumilin Bacterii Gram + - -
Tetain Bacterii Gram + - -B. mesentericus Esperin Bacterii Gram + - -
B. polymyxa Polymyxin Bacterii Gram - - -Colistin Bacterii Gram - - -
B.tyaminoliticus Octopytin Bacterii Gram + - -Baciphelecin Bacterii Gram + - -
B. circulans Circulin Bacterii Gram - - -Butirosin Bacterii Gram + - -
B.laterosporus LaterospuramineLaterosporin
Bacterii Gram + - -
B. cereus BiorinCerexin
Bacterii Gram + - -
Deoarece nu există o corelaţie strictă între ţintele antibioticului şi căile metabolice în care el este format, este riscant să se spună că toţi metaboliţii secundari manifestă o anumită activitate biologică, mai ales dacă ţinem cont de faptul că de multe ori aceşti compuşi au semnificaţie numai pentru om şi mai puţin pentru organismul producător.
Deşi cercetările asupra utilităţii medicale a antibioticelor s-au referit în special la activitatea antibacteriană, în prezent sunt studiate acţiunile unor asemenea substanţe şi asupra unor alte tipuri de celule, eucariote. Un exemplu în acest sens îl constituie monensinul, identificat iniţial în culturi de Streptomyces cinnamonensis şi care în prezent este utilizat pe scară largă ca şi coccidiostatic sau ca stimulator al creşterii animalelor (Vining, 1990).
O problemă care se pune în cazul producerii de substanţe antibiotice este aceea a modalităţilor în care bacteriile producătoare rezistă la acţiunea acestor substanţe care de obicei inhibă creşterea sau alte procese biochimice esenţiale. Pentru a elimina efectul de inhibare a creşterii, bacteriile producătoare pot prezenta mai multe mecanisme de rezistenţă: modificarea situsului ţintă al antibioticului, modificarea permeabilităţii membranare sau inactivarea metabolitului toxic (Cundliffe, 1989). Deoarece antibioticele sunt rapid eliminate în exteriorul celulei pe măsură ce el este produs, inactivarea celulară nu afectează eficienţa produsului extracelular.Un studiu efectuat asupra rezistenţei la antibiotice a tulpinilor bacteriene sensibile ("ţintă") a evidenţiat faptul că antibioticele excretate în mediu şi care acţionează în vecinătatea organismului producător au semnificaţia unor factori implicaţi în competiţia pentru resurse.
126

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
Rezistenţa la antibioticele obţinute a fost observată la multe tulpini bacteriene de colecţie, izolate înainte de introducerea acestor antibiotice în practica medicală ceea ce sugerează ideea că populatiile bacteriene vin în contact cu asemenea compuşi în mediul lor natural de viaţă. De asemenea, unii metaboliţi toxici chiar dacă nu au o valoare antibiotică mare, ei pot potenţa activitatea altor antibiotice faţă de care s-a instalat rezistenţa la numeroase tulpini bacteriene. Acesta este cazul acidului clavulanic produs de tulpini de S.clavuligerus care are o slabă activitate antibiotică dar care determină o sporire a acţiunii unor antibiotice β-lactamice cum este cefamicinul prin inhibarea β-lactamazelor pro-duse de tulpina bacteriană ţintă.
Antibioticele descoperite până în prezent şi utilizate mai mult sau mai puţin intens în practica medicală pot fi clasificate ţinând cont de diferite criterii: modul de acţiune, natura chimică, spectru de acţiune etc.
O abordare recentă a structurii unor antibiotice consideră că acestea, alături de un număr mare de metaboliţi sintetizaţi atât de către bacterii cât şi de fungi sau plante, pot fi grupate într-o familie de compuşi denumiţi poliketide (Hopwood şi Sherman 1990). Toţi aceşti compuşi au în comun prezenţa unor grupări keto la nivelul unor atomi de carbon foarte variaţi ca poziţie în molecula respectivă. Printre compuşii incluşi în această familie menţionăm: tetraciclinele, antraciclinele, eritromicina, rifamicinele, granaticinul, avermectinul (toate sintetizate de către streptomicete), aurantininul (sintetizat de Bacillus sp.) mupirocin (elaborat de Pseudomonas sp.) precum şi ale substanţe elaborate de celule eucariote: micotoxine, flavonoizi, fitoalexine etc.
1.1.1. Antibiotice care inhibă sinteza peretelui celularDupă descoperirea penicilinei de către Fleming în 1929, un număr mare de antibiotice
înrudite cu aceasta au fost izolate, ele având în comun structura chimică (sunt β-lactami) şi modul de acţiune (inhibarea sintezei peretelui celular bacterian).
Deşi penicilinele au fost descoperite ca fiind produse de către fungi, s-a dovedit că asemenea antibiotice pot fi sintetizate şi de către unele specii de streptomicete cum ar fi: S.clavuligerus, S.lipmanii, S.lactamgens, S.cattleya (Martin şi Liras, 1989; Matsushima şi Baltz, 1989; Srinivasan şi colab.,1991).
Pe lângă antibioticele β-lactamice au mai fost izolaţi şi caracterizaţi şi alţi compuşi care afectează funcţiile membranei plasmatice. Un exemplu îl constituie nistatinul, antifungic produs de unele specii ale genului Streptomyces. Un alt grup este reprezentat de polimixine, bacitracine şi gramicidine, antibiotice polipeptidice produse de bacterii aparţinând genului Bacillus (B.polymixa, B.licheniformis, B.brevis). Aceşti compuşi determină creşterea permeabilităţii membranei plasmatice ceea ce are drept consecinţă ieşirea masivă a unor aminoacizi sau a unor derivaţi purinici şi pirimidinici din bacterie, conducând în final la moartea celulei. De asemenea, bacitracina acţionează asupra sintezei peretelui celular, determinând inhibarea biosintezei peptidoglicanului unor bacterii Gram negative.
1.1.2. Antibiotice care inhibă sinteza proteinelorDintre antibioticele care afectează sinteza proteinelor, un loc important este ocupat de
aminoglicozide, tetracicline şi cloramfenicol, toate cele trei grupe fiind sintetizate de specii ale genului Streptomyces..
Aminoglicozidele reprezintă un grup foarte important de antibiotice care au drept component principal unele glucide aminate. Primul antibiotic din acest grup care a fost descoperit în 1944 este streptomicina produsă de S.griseus, după care au fost izolate numeroase alte aminoglicozide de la bacterii aparţinând în principal genurilor Streptomyces şi Micromonospora: gentamicina produsă de specii de Micromonospora, neomicina produsă de S.fradiae, ribostamicina produsă de S.ribosidificus, kanamicina sintetizată de tulpini de S.kanamyceticus, butirozina produsă de B.circulans etc.(Okuda şi Ito, 1982).
127

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
Aminoglicozidele sunt antibiotice cu spectru larg de acţiune utilizate de obicei pentru tratarea unor infecţii produse de tulpini de E.coli, Klebsiella sp., Proteus sp., Enterobacter sp. Mecanismul prin care aceste antibiotice omoară celulele sensibile nu este pe deplin elucidat. Există numeroase date experimentale care evidenţiază că situsul primar de acţiune al acestor antibiotice ar fi ribosomii, ceea ce determină afectarea sintezei proteinelor. Efectele pleiotropice ale aminoglicozidelor asupra bacteriilor sunt considerate a fi consecinţa interacţiunii lor cu ribosomii. Alături de interferenţa cu sinteza proteică, aminoglicozidele induc distrugeri ale membranei plasmatice, afectează respiraţia celulară, acumularea de ARN şi determină în final moartea celulelor. In ceea ce priveşte mecanismul rezistenţei bacteriilor producătoare la acţiunea acestor antibiotice s-a evidenţiat faptul că acestea prezintă strategii diferite cum ar fi producerea unor enzime ce inactivează antibioticul sau modificarea situsului de acţiune al acestuia (Cundliffe, 1989). Spre exemplu, bacteriile producătoare de aminoglicozide sintetizează în acelaşi timp şi enzime ce modifică antibioticele respective: aminoglicozid-fosfotransferaza (APH) şi acetiltransferaza (AAC). De asemenea, bacteriile respective (S..kanamyceticus, S.tenebrarius) pot produce simultan şi enzime ce modifică unele situsuri de acţiune ale antibioticului (de exemplu, situsurile ribosomale) prin metilarea ARNr.
Tetraciclinele reprezintă un grup destul de heterogen de antibiotice produse în special de bacterii din genul Streptomyces ele având în comun o structură chimică de bază dar variind în funcţie de grupările chimice care se leagă de această structură: oxitetraciclina (produsă de S.rimosus), tetraciclina (produsă de S. viridofaciens), clortetraciclina (sintetizată de S.aureofaciens). Tetraciclinele prezintă un spectru destul de larg de acţiune fiind bacteriostatice pentru diferite bacterii Gram pozitive şi Gram negative (cu excepţia tulpinilor de Salmonella, Proteus şi Pseudomonas) şi bactericide în concentraţii mari. Tinta principală de acţiune a acestor antibiotice este sinteza proteică, ele neavând efecte evidente asupra ADN, ARN sau asupra sintezei peretelui celular. Modalitatea exactă a legării tetraciclinelor la ribosomi nu este încă pe deplin cunoscută. Se pare că ele se leagă puternic de subunitatea 70S a ribosomilor ceea ce determină efectul inhibitor al antibioticelor. De asemenea, tetraciclina se leagă slab de subunitatea 30S ceea ce are drept consecinţă blocarea legării aminoacil-ARNt la această subunitate şi deci a alungirii lanţurilor polipeptidice. Autorezistenţa bacteriilor producătoare poate include modificarea situsurilor ţintă normale, inactivarea intracelulară a antibioticului sau eliminarea sa rapidă din celulă (Cundliffe, 1989).
Cloramfenicolul este un antibiotic cu spectru destul de larg de acţiune, el fiind izolat în 1947 din filtrat de cultură de S.venezuelae. Acţiunea cloramfenicolului constă în împiedicarea ARNm să se lege la ribosomi ceea ce se concretizează prin inhibarea sintezei proteice (a formării legăturilor peptidice) (Pongs, 1979).
1.1.3. Antibiotice care inhibă replicarea şi transcrierea informaţiei geneticeDintre antibioticele care împiedică replicarea ADN şi sinteza ARNm menţionăm
novobiocina, coumermicina şi rifamicinele.Novobiocina este un antibiotic cu o structură complexă produs de S.sphaeroides cu
efect, de obicei, bacteriostatic asupra bacteriilor, mai ales în mediu acid. Similar ca acţiune este coumermicina A, un antibiotic izolat din culturi de S.rishinensis şi S.hazeliensis. La concentraţii mai mici de novobiocină şi cumermicină A este afectată doar replicarea ADN din celulele sensibile, nu şi procesul de transcriere. Cele două antibiotice pot afecta acţiunea ADN girazei, enzimă ce determină suprarăsuciri ale moleculei de ADN, ceea ce explică inhibarea replicării şi transcrierii informaţiei genetice (Ryan, 1979). Rezistenţa la novobiocină a bacteriei producătoare S.sphaeroides este determinată de gena gyrB al cărei produs, ADN giraza B este rezistentă la acţiunea acestui antibiotic (Cundliffe, 1989).
Rifamicinele, având ca reprezentat tipic rifampicina reprezintă un grup de antibiotice macrociclice care acţionează, în special, asupra micobacteriilor şi bacteriilor Gram pozitive.
128

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
Mecanismul lor de acţiune se referă la inhibarea procesului de transcriere a informaţiei genetice prin blocarea acţiunii enzimei ARN-polimeraza.
Un alt antibiotic cu acţiune inhibitoare asupra acizilor nucleici este granaticinul, antibiotic de tip quinonic, izolat din culturi de S.olivaceus şi S.violaceoruber. Acţiunea sa „in vitro” se manifestă, atât la procariote cât şi la eucariote, asupra sintezei de ADN, ARN, dar şi de proteine. De aceea, granaticinul are efect antitumoral, împotriva leucemiei P388 şi efect citotoxic pentru celulele KB (Ogilvie şi Kersten, 1979).
1.2. Acţiuni asupra celulelor animale
Numeroase substanţe antibiotice nu au fost utilizate în practica medicală datorită acţiunii lor toxice asupra celulelor animale. Acest efect toxic rezultă din acţiunea directă asupra uneia dintre căile metabolismului primar, căi care sunt universale, de obicei, asupra producerii de energie sau exprimării genelor. Totuşi, toxicitatea asupra celulelor animale nu rezultă întotdeauna din inhibarea aceleiaşi reacţii care este ţintă la microorganisme. Spre exemplu, amfomicina care interferă cu transportul membranar al intermediarilor mureinei la bacterii, la celulele eucariote determină o blocare a glicozilării.
Unele antibiotice care au evidenţiat o activitate de inhibare a creşterii mai multor tipuri de organisme au început să fie utilizate ca substanţe antitumorale datorită toxicităţii lor asupra celulelor ce proliferează rapid. Printre aceste substanţe, un loc important îl ocupă antraciclinele şi bleomicina substanţe care sunt produse de specii ale genului Streptomyces (Tsukagoshi şi colab.,1986). Descoperite iniţial ca agenţi antibacterieni, s-a dovedit ulterior că cele două familii de substanţe se leagă specific, prin intercalare, la moleculele de ADN iar bleomicina, în plus, poate determina scindarea catenelor de ADN. Citotoxicitatea selectivă a bleomicinei rezultă din scăderea activităţii unei enzime de inactivare din celulele tumorale.
Un alt tip de produşi antimicrobieni sau inhibitori enzimatici cu activitate antitumorală este reprezentat de proteine de dimensiuni mici, cum ar fi: macromicinul, actinoxantinul, forfenicin sau bestatin, ce conţin o grupare prostetică cromoforă ce se intercalează în macromoleculele de ADN, modificându-le structura(Aoyagi şi colab.,1978).
Unii metaboliţi secundari manifestă acţiune farmacologică specificpă asupra animalelor; în acest caz ei nu prezintă o citotoxicitate generală, dar acţiunea lor farmacologică îi poate face uneori extrem de toxici. Un exemplu îl constituie acţiunea vasodilatatoare a unui compus (WS 1228A, B) izolat de la S. aureofaciens care însă nu este însoţită şi de toxicitate sau activitate antibiotică. Un alt compus, MY-336a, izolat din culturi de S. gabonae exercită acţiune antagonică cu receptorii β-adrenergici, avînd deci o acţiune farmacologică selectivă, netoxică (Kase, 1986). In cazul amicourmamicinei A izolate de la B. pumilis s-a evidenţiat iniţial o acţiune antibiotică, după care s-a dovedit a avea şi o activitate antiinflamatorie asupra animalelor testate (Itoh şi colab., 1981).
De asemenea, printre substanţele izolate iniţial în experimentele de screening pentru activitatea antibiotică au fost descoperite unele care au acţiune de modulare a răspunsului imun. Spre exemplu, substanţa IC201 izolată de la S.cirratus şi S fimbriatus ca agent antitumoral, s-a dovedit a avea acţiune activatoare şi asupra macrofagelor. Un alt compus, macrolidul FK 506 izolat de la S.tsukubaensis în cursul unor experimente de selecţie a unor substanţe care să inhibe producerea de interleukină-2 s-a dovedit a avea acţiune imunosupresoare.
1.2.1. Activitate insecticidăS-a evidenţiat că numeroşi metaboliţi secundari produşi de streptomicete în special au
acţiune insecticidă. Unii dintre ei, cum ar fi unele antibiotice macrolidice polienice, blochează respiraţia celulară sau inhibă sinteza proteică la eucariote, fiind astfel agenţi nespecifici. Alţi metaboliţi secundari, de tipul nikamicinei au acţiune specifică inhibând chitin-sintetaza şi deci
129

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
acţionează asupra peretelui celular chitinos al insectelor. Un alt antibiotic, mibelmicina produsă de S. hygroscopicus subsp. aureolacrimosus, nu are efect antibacterian dar acţionează selectiv asupra insectelor având în plus şi efect antihelmintic şi acaricid.
Intr-un studiu recent, Fabre şi colab.(1988) au izolat în urma unui complex proces de selecţie realizat asupra a 6280 de culturi de actinomicete, o nouă substanţă cu acţiune insecticidă. O importantă acţiune insecticidă este datorată biosintezei de către unele tulpini bacteriene a chitinazei care acţionează direct asupra învelişurilor insectelor patogene.
1.3. Activitate asupra celulelor vegetale
Unii metaboliţi secundari produşi de bacteriile din genul Streptomyces au acţiune fitotoxică şi uneori antifungică. Spre exemplu, herbicidinele produse de S.sagonensis sunt nucleozide cu acţiune fitotoxică pentru plantele dicotiledonate. Ele inhibă, de asemenea, şi dezvoltarea fungilor. Alte substanţe, cum ar fi herbimicinele produse de S.hygroscopicus au o foarte slabă activitate antibiotică, în schimb sunt toxice pentru celulele vegetale. In plus, herbimicina A inhibă dezvoltarea virusului mozaicului tutunului şi are acţiune antitumorală pentru celulele animale (Yamashita şi colab., 1989). O altă substanţă, homoalanozina, produsă de S. galileus este un antimetabolit al acidului aspartic şi acidului glutamic, ea exercitând acţiune insecticidă şi herbicidă (Fushimi şi altii,1989).
De asemenea, numeroşi cercetători (Backman 1995, Baker 1985, Bachow 1995) au publicat date referitoare la capacitatea unor specii aparţinând genului Bacillus de a inhiba dezvoltarea unor ciuperci fitopatogene (Tabelul 1). In majoritatea cazurilor combaterea biologică a fungilor fitopatogeni implică folosirea factorilor biotici existenţi în mediul plantelor. Studiile efectuate până în prezent au evidenţiat capacitatea unor tulpini de Bacillus subtilis de a combate diferite ciuperci fitopatogene, cum ar fi : Macrophomina phaseolina – patogen la mazăre (Siddiqui 1995), Rhizoctomia solani - patogen la mazăre (Bochow 1994), fasole şi seminţe de grâu (Lazzareti 1994), Fusarium oxysporum şi Phytyum ultinacere – patogeni la tomate (Sadders 1996).
Cele mai frecvente specii ale genului Bacillus care au prezentat capacitatea de a preveni şi combate evoluţia unor boli determinate de ciuperci fitopatogene sunt: Bacillus subtilis (Figura 3), Bacillus licheniformis, Bacillus polymyxa, Bacillus cereus.
Figura 3. Acţiunea inhibitoare a tulpinii B.subtilis B2 asupra fitopatogenului Sclerotinia sclerotiorum.
Studiile privind oportunitatea utilizării acestor specii bacteriene în combaterea biologică s-au bazat pe proprietatea de a produce spori cu înaltă rezistenţă şi longevitate în condiţii naturale de mediu, acesta determinând posibilitatea obţinerii şi comercializării lor într-o manieră similară cu fungicidele chimice.
In lupta pentru nutrienţi microorganismele au la dispoziţie un întreg arsenal de compuşi chimici care inhibă organismele competitive. Mulţi din aceşti compuşi au origine
130

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
peptidică şi pot fi sintetizaţi ribozomal sau neribozomal. Multe dintre tulpinile de Bacillus subtilis produc o serie de substanţe de natură lipooligopeptidică din gama iturinului, care au activitate antifungică, hemolitică precum şi proprietăţi antibiotice. Structura chimică a acestor substanţe a fost determinată prin diverse metode (spectrometrie de masă (MS), gel cromatografie, cromatografie în strat subţire (TLC), lichid cromatografie (HPLC)). Componenta proteică a acestor substanţe este reprezentată de oligopeptide alcătuite din şapte aminoacizi, iar componenta lipidică conţine acizi graşi cu catena carbonică lungă (C14-C16).
Astfel, iturin AL conţine o heptapeptidă (2 D-Asp, 1 L-Asp, 1 L-Glu, 1 L-Pro, 1 L-Ser, 1 D-Tir) şi un amestec de -amino acizi graşi cu catenă lungă C14-C16. Iturin D şi iturin E conţin o heptapeptidă (3 Asp, 1 Glu, 1 Pro, 1 Ser, 1 Tyr) şi un amestec de -amino acizi graşi cu catenă C14-C16, diferind de iturin A prin prezenţa unei grupări carboxil la iturin D şi o grupare carboximetil la iturin E. Mycosubtilin, o substanţă din gama iturinului, conţine o heptapeptidă ciclică (Asn, Tyr, Asn, Gln, Pro, Ser, Asn) legată de un -amino acid gras. Tot din gama iturinului fac parte surfactinul şi fengycinul care conţin un -hidroxiacid gras şi bacillomycinul care conţine un -amino acid gras.
Studii mai intense au fost efectuate cu o substanţă produsă de mai multe specii de Streptomyces (S. hygroscopicus, S. viridochromogenes) şi care a primit denumirea de bialafos (fosfinotricin-alanil-alanina). Aceasta substanţă a fost iniţial selectată pentru activitatea sa antibiotică după care s-a observat că ea are, de fapt, o acţiune mult mai complexă. Componentul activ al bialafosului este fosfinotricinul care inhibă glutamin-sintetaza din celulele vegetale, ceea ce îi conferă bialafosului o puternică acţiune herbicidă (Lea şi colab., 1984).
Mulţi dintre metaboliţii secundari produşi de diverse microorganisme au fost testaţi pentru eventuala lor acţiune asupra unor enzime. Atunci când sistemul test a inclus enzime cheie în procesele farmacologice de la animale, numeroşi metaboliţi secundari au fost detectaţi ca având activităţi fiziologice "in vivo". Mulţi dintre aceşti compuşi au fost descoperiţi a fi inhibitori proteazici, acţionând asupra pepsinei, papainei, tripsinei, chimiotripsinei, catepsinei, elastazelor, aminopeptidazei B şi leucin-aminopeptidazei (Umezawa, 1982). Printre inhibitorii proteazici izolaţi de la actinomicete unii acţionează asupra reninei sau asupra unei zinc-exopeptidaze care converteşte angiotensina I în angiotensină II.
Cercetările efectuate asupra culturilor microbiene au permis izolarea unor inhibitori şi ai altor enzime. Acesta este cazul inhibitorilor pentru glucozidaze (Muller, 1986), pentru cAMP (reticulol), pentru kinaze etc.
2. Determinismul genetic al biosintezei unor antibiotice si a altor metaboliti secundari
In urma experimentelor de cartare a genelor implicate în metabolismul secundar la procariote s-a evidenţiat faptul că acestea sunt grupate formând "clusteri". Localizarea acestor gene este, în majoritatea cazurilor, cromosomală dar, există cazuri cînd acestea sunt localizate pe plasmide (Hopwood, 1978; Kinashi şi colab.,1987). Spre exemplu, genele implicate în sinteza oxitetraciclinei de către S.rimosus sunt grupate în doi "clusteri" cromosomali (Binnie şi colab., 1989). La fel şi genele care determină sinteza actinorhodinului şi undecilprodigiozinului de către tulpini de S. coelicolor (Rudd şi Hopwood, 1979), a eritromicinei de către Saccharopolyspora erythraea (Weber şi altii, 1985) sau avermectinei de către Streptromyces avermectilis (Ikeda şi colab., 1987). In plus, evidenţierea faptului că aceste grupuri de gene includ şi elemente reglatoare ca şi determinanţi genetici de rezistenţă la produsul antibiotic respectiv sugerează existenţa unui sistem foarte bine organizat, cu funcţionare complexă. Nu este însă stabilit cu exactitate dacă această organizare a genelor există şi în cazul altor metaboliţi secundari, fără
131

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
activitate antibiotică, cum ar fi unii pigmenţi, alcaloizi, reglatori ai celulei vegetale etc. (Martin şi Liras, 1989).
In ceea ce priveşte localizarea plasmidială a unor gene implicate în biosinteza unor antibiotice, datele referitoare la acest aspect sunt mai puţine. Un exemplu îl constituie genele pentru biosinteza metilenomicinei care se găsesc plasate pe plasmida SCP1 de la S. coelicolor (Wright şi Hopwood, 1978; Chater şi Bruton, 1985). De asemenea, de la bacteriile din genul Streptomyces au fost izolate o serie de plasmide lineare cu dimensiuni variate, între 5,4-17 Kb până la 350-520 Kb (Chardon-Loriaux, 1985; Kinashi şi colab., 1987). Evidenţierea plasmidelor lineare uriaşe la şase tulpini de Streptomyces producătoare de antibiotice şi examinarea corelaţiei dintre prezenţa lor şi sinteza de antibiotice a dus la concluzia că, cel puţin în cazul metilenomicinei, genele implicate în sinteza sa sunt plasate pe aceste plasmide care reprezintă aproximativ 5 % din cantitatea totală de ADN celular. Sunt însă necesare studii suplimentare pentru a stabili dacă şi în alte cazuri genele care determină sinteza unor antibiotice sunt localizate pe plasmide similare.
2.1. Organizarea şi exprimarea genelor pentru unii metaboliţi secundari sintetiza şi de specii de Bacillus
Cea mai mare parte a metaboliţilor secundari produşi de bacteriile din genul Bacillus sunt homo- sau heteropeptide. Spre exemplu, gramicidina S este un decapeptid ciclic format din douăă unităţi de D-Phe-L-Pro-L-Val-L-Orn-L-Leu, fiind produsă de tulpini de B. brevis, ca de altfel şi tirocidinele. Familia bacitracinelor cuprinde peptide alcătuite din 12 aminoacizi, produse de tulpini de B. licheniformis. Studiile cu extracte celulare au evidenţiat că aminoacizii constituenţi ai acestor compuşi sunt activaţi şi legaţi în subunităţile respective de către mai multe tipuri de peptid-sintetaze (Kleinkauf şi Van Dohren, 1987).
Experimentele de clonare în E. coli a genelor pentru sinteza gramicidinei şi tirocidinei au evidenţiat faptul că în cazul tirocidinei sunt implicate trei enzime multifuncţionale, în timp ce la producerea gramicidinei S participă cel puţin 2 sintetaze (GS1 şi GS2) codificate de două gene diferite grsA şi grsB.
2.2. Organizarea şi exprimarea genelor pentru unele antibiotice sintetizate de bacteriile din genul Streptomyces
In cazul streptomicetelor au fost evidenţiaţi numeroşi metaboliţi secundari, dar numai puţini au o structură peptidică. Unul dintre aceştia, bialafosul prezintă importanţă practică deosebită, deoarece manifestă activitate herbicidă. Recent, Nakajima şi colaboratorii (1991) au izolat de la S.hygroscopicus un nou compus cu activitate herbicidă, care a primit denumirea de hidantocidin. Bialafosul, care a fost mai intens studiat, este sintetizat pornind de la piruvat sau fosfoenolpiruvat, printr-o cale metabolică care cuprinde cel puţin 13 etape (Thompson şi Anzai, 1987). Experimentele de clonare cu genele implicate în sinteza acestui compus au evidenţiat faptul că în acest proces sunt implicate mai multe gene (cel puţin 7) grupate, alături de care se află şi gena pentru rezistenţa la bialafos (bar). Astfel, au fost obţinute două grupe de clone derivate de la mutante neproducătoare şi care au fost transformate cu plasmida pIJ61 recombinantă (conţinea diferite gene de la S.hygroscopicus). Un grup de asemenea plasmide recombinante restabileşte productivitatea mutantelor care prezintă modificări ale etapelor 5, 6, 12 şi 13 ale căii metabolice, în timp ce un alt grup restabilea producerea de bialafos de către mutantele blocate la nivelul etapelor 1, 3 şi 4. In plus, a fost realizat transferul unui segment de 16 Kb, care restabilea productivitatea tuturor celor 7 tipuri de mutante. Acest segment de ADN conţinea şi gena bar care, de altfel, a permis şi identificarea clonelor transformate. Totuşi, acest fragment nu determină o producţie semnificativă de bialafos la mutantele neproducătoare, ceea
132

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
ce sugerează existenţa unor gene suplimentare aflate în afara segmentului de 16 Kb, necesare exprimării complete a operonului pentru sinteza bialafosului.
De asemenea, în urma experimentelor de clonare a genelor bar (de rezistenţă la bialafos) în diferite celule gazdă, inclusiv în celulele vegetale s-a evidenţiat faptul că pentru funcţionarea normală a genelor pentru biosinteza bialafosului sunt necesare încă trei gene diferite care determină adenilarea la diferite etape ale biosintezei. Aceste noi date conduc la presupunerea că genele implicate în biosinteza acestui compus se găsesc localizate la nivelul unui fragment de ADN de peste 34 Kb.
Experimentele lui Anzai si ale colaboratorilor (1987) au evidenţiat faptul că prin clonarea unui fragment de ADN de 5,9 Kb, care conţine gena brpA are loc activarea transcrierii atât a genei bar cât şi a cel puţin altor 6 gene structurale bap. De asemenea, aceşti autori au arătat că atât la S.viridochromogenes, cât şi la S. hygroscopicus organizarea genetică a genelor implicate în sinteza bialafosului este identică.
Un alt grup de metaboliţi secundari peptidici, actinomicinele (cromopeptide) este sintetizat în urma acţiunii unei enzime, fenoxazinon-sintetaza (PHS). Prin clonarea genei pentru PHS de la S. antibioticus în S. lividans (Jones şi Hopwood, 1984) s-au obţinut mai multe tipuri de clone: unele care conţineau gena structurală pentru PHS, iar altele care prezentau activitate PHS, dar care a rezultat în urma activării unei gene "tăcute" (silent gene) endogene de către fragmentul de ADN exogen transformant. Fenomenul este frecvent întâlnit în cazul streptomicetelor, el fiind semnalat şi în cazul tulpinilor producătoare de alţi metaboliţi secundari. Un exemplu îl constituie clonarea genelor pentru producerea cefamicinei de la S.cattleya la S.lividans, clonele producătoare rezultând în urma activării unor gene endogene silenţioase (Martin şi Liras, 1989).
Mulţi dintre metaboliţii secundari produşi de streptomicete au o structură de tip macrolidic în care inelul macrociclic derivat de la poliketide este închis printr-o legătură lactonică. La acest inel macrociclic sunt, de obicei, ataşate glucide bazice sau neutre. Spre exemplu, eritromicina produsă de Saccharopolyspora erythraea (Streptomyces erythraeus) conţine un inel lactonic (eritronolid) şi 2 glucide specifice, dezozamina şi cladinoza, sinteza sa realizându-se în aproximativ 30 etape enzimatice Prin clonarea în S.lividans a unor fragmente de ADN cromosomal izolate de la tulpini de S.erythraea producătoare de eritromicină s-a stabilit că trei dintre genele implicate în biosinteza antibioticului, eryA, eryB şi eryC sunt grupate şi localizate într-o unică regiune a cromosomului bacteriei producătoare. Utilizând un vector de clonare integrativ (care se poate integra în cromosomul gazdei) s-a stabilit că alături de genele care determină sinteza antibioticului se află şi gena pentru rezistenţa la eritromicină, ermE (genă care determină sinteza unei metilaze pentru ARNr) (Stanzak şi colab., 1986). De asemenea, Mc Alpine şi colab. (1987) au comunicat obţinerea unui antibiotic hibrid care a primit denumirea de 2-noreritromicină, în urma clonării într-o mutantă de S.erythraea, care avea blocată ultima etapă a sintezei eritromicinei a unui fragment de ADN de 30-35 Kb izolat de la S.antibioticus producător de oleandomicină.
Un alt antibiotic, tilozina produs de tulpini de S.fradiae prezintă o structură chimică asemănătoare eritromicinei, având un inel macrociclic, tilanolid şi trei glucide mai puţin răspândite: micaminoza, micaroza şi micinoza. Studiile referitoare la organizarea genelor implicate în sinteza acestui antibiotic au evidenţiat faptul că genele tyl (aproximativ 9 gene) sunt grupate, alături de ele aflându-se şi gena de rezistenţă (tlrB), precum şi o secvenţă de ADN amplificabil flancată de 2 secvenţe repetate direct (Baltz şi Seno, 1988). De asemenea, s-a stabilit că pentru exprimarea genei tylF, care codifică enzima ce controlează ultima etapă a căii metabolice de sinteză a antibioticului este necesară o proteină reglatoare (efector pozitiv) produsă de gena tylG. O situaţie asemănătoare prezintă şi genele care codifică sinteza carbomicinei şi rezistenţa la acest antibiotic (Martin şi Liras, 1989).
133

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
Un alt grup de antibiotice derivate de la poliketide este reprezentat de tetracicline şi antracicline, antibiotice intens studiate datorită importanţei lor practice deosebite. In urma examinării mai multor tipuri de mutante incapabile de a sintetiza oxitetraciclina s-a dovedit că genele care codifică producerea acestui antibiotic se află localizate în două grupe plasate opus pe cromosomul bacterian. Prin clonarea în S.griseus a unor fragmente de ADN izolate de la S.rimosus s-a reuşit identificarea a două gene responsabile de rezistenţa la tetraciclină (Ohnuki şi colab., 1985): gena tetA care codifică un polipeptid care interferă cu sinteza proteinelor la nivel ribosomal şi gena tetB care determină o acumulare intracelulară redusă a antibioticului în celulele producătoare. Nu este, însă, clar dacă aceste gene se află localizate la nivelul celor douăă grupuri de gene sau au o altă dispunere. Au mai fost clonate şi alte gene de rezistenţă la tetraciclină, otcRI şi otcRII, dar nu s-a stabilit încă dacă aceste gene sunt sau nu identice cu cele anterioare. Recent, Butler şi colab. (1989) şi Binnie şi colab. (1989), au realizat o serie de clonări a unor gene implicate în sinteza şi rezistenţa la acest antibiotic şi au sugerat faptul că cel puţin o parte, dacă nu chiar toate genele structurale care codifică sinteza oxitetraciclinei sunt linkate cu genele de rezistenţă otcRI şi otcRII, formând, astfel, un singur "cluster".
O grupare asemănătoare prezintă şi genele implicate în biosinteza/rezistenţa tetracenomicinei C, un antibiotic antraciclinic produs de S.glaucescens.
Aminoglicozidele reprezintă un alt grup de antibiotice intens studiat datorită importanţei practice deosebite. Din acest grup streptomicinei i s-a acordat cea mai mare atenţie. Astfel, la S.griseus, cel mai cunoscut producător al acestui antibiotic, cel puţin 13 gene sunt implicate în biosinteza şi rezistenţa la streptomicină, ele fiind localizate într-un singur "cluster" cromosomal (Mansouri şi altii, 1989) (Figura 4).
Figura 4. Reprezentarea schematică a genelor implicate în biosinteza streptomicinei la două specii de streptomicete: S.griseus şi S.glaucescens.
Ohnuki şi colaboratorii (1985) au arătat că la S.griseus grupul de gene str este format din cel puţin 4 gene: strR, strA, strB şi strC. Gena strA codifică enzima streptomicin-6'-fosfotransferaza care asigură rezistenţa la acest antibiotic; strB complementează o mutaţie la o tulpină care prezenta deficienţă în legarea streptidin-6-fosfatului de dihidrostreptoză; strR este o genă reglatoare care codifică un efector pozitiv necesar exprimării complete a genelor strA şi strB. Rezultate similare au obţinut Distler şi colab.(1987), care au evidenţiat că la S.griseus genele necesare sintezei streptomicinei sunt linkate cu gena aphD care codifică pentru enzima streptomicin 6-fosfotransferaza, genă similară genei sph pentru enzima hidroxi-streptomicin-
134

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
fosfotransferaza de la S.glaucescens. Analizele de restricţie şi studiile de hibridizare a fragmentelor de ADN de la S.griseus şi S.glaucescens au relevat existenţa, în ciuda unei omologii de aproximativ 75 % a genelor implicate în biosinteza streptomicinei, a unor diferenţe majore, ceea ce sugerează faptul că, în cursul evoluţiei a avut loc o separare a genelor de biosinteză în două seturi distincte repartizate doar la speciile de Streptomyces (Martin şi Liras, 1989).
2.3. Organizarea şi exprimarea genelor de biosinteză a unor pigmenţi de către streptomicete
Streptomicetele pot sintetiza pe lângă diferite antibiotice şi alţi metaboliţi secundari, cum ar fi pigmenţii, unii dintre aceştia manifestând şi o uşoară activitate antibiotică. Dintre aceştia cel mai mult studiat este actinorhodinul, un pigment de tip izocromanequinonic, produs de S.coelicolor printr-o cale poliketidică tipică (Floss, 1981). Pigmentul respectiv produs de bacteriile cultivate pe mediul R2YE este roşu şi nedifuzibil la pH neutru dar, după expunere la vapori de amoniac devine albastru şi difuzibil. In urma cercetărilor efectuate pe diferite tulpini mutante de S.coelicolor s-a realizat o grupare a acestora în mai multe clase (7) în funcţie de variaţiile de exprimare a genelor care alcătuiesc "clusterul" act. Rezultatele obţinute au permis elaborarea unei ipoteze referitoare la sinteza unor metaboliţi secundari de tip poliketidic, ţinând cont de marea varietate de asemenea metaboliţi (macrolide, poliene, polieteri, tetracicline, antracicline, ansamicine, isocromaquinone etc). Se pare că la toţi metaboliţii secundari de tip poliketidic, esterii acil-CoA şi -cetoacizii condensează împreună printr-un mecanism de polimerizare de tip cap-coadă, ceea ce conduce la obţinerea unor lanţuri alifatice de diferite dimensiuni. Aceste lanţuri suferă, ulterior, modificări suplimentare, care le asigură specificitatea (Hopwood şi Sherman, 1990). Dacă reacţia de condensare realizată prin intervenţia unei enzime de tip β-ceto-acil-tioester-sintetaza ("enzima de condensare") este comună tuturor metaboliţilor secundari poliketidici, atunci ar însemna că între secvenţele care codifică poliketid-sintetazele la diferite organisme producătoare există o mare omologie care este conservată. Pornind de la o asemenea ipoteză, Malpardita şi colab.(1987) au utilizat genele act de la mutantele din clasele I şi III ca probe de ADN în experimentele de hibridizare şi au observat că între acestea şi ADN de la 14 dintre cei 18 producători de poliketide se produce hibridizare cu intensitate variată, deci există omologie. Probele nu au hibridizat cu ADN izolat de la tulpinile care produc tetraciclină, eritromicină, candicidină şi curamicin. Aceste rezultate demonstrează o corelaţie, incompletă, între producerea de poliketide şi prezenţa de secvenţe de ADN care sunt parţial omoloage cu actinorhodin poliketid-sintetaza. Această strategie de hibridizare poate fi extinsă prin utilizarea unor probe de ADN a unor secvenţe care codifică diferite sintetaze izolate de la tulpini bacteriene variate, ceea ce ar permite recunoaşterea unor producători criptici ai unor clase de metaboliţi secundari particulari. Prin alterarea specificităţii de substrat a unor sintetaze specifice utilizând mutageneza directă sau recombinarea"in vitro" a genelor care codifică pentru aceste enzime pot fi obţinute noi antibiotice cu proprietăţi speciale.
Alte două tipuri de pigmenţi, metilenomicin produs de tulpini de S.coelicolor şi S.violaceoruber sau undecilprodigiozin produs de S.coelicolor A3(2) au fost studiate cu o intensitate mai scăzută, rezultatele obţinute sugerând şi în aceste cazuri o grupare a genelor implicate în biosinteza produşilor respectivi (Malpardita şi colab., 1990).
Gruparea în "clusteri” a genelor care codifică căi ale metabolismului secundar reprezintă un fenomen comun pentru numeroase tipuri de substanţe. In cazul "clusterilor" pentru sinteza tetra-cenomicinei şi actinorhodinului s-a evidenţiat o subgrupare specială a genelor componente care controlează momentul apariţiei fiecărei enzime în calea metabolică respectivă, astfel încât intermediarii apar în anumite etape ale reacţiilor biochimice. Formarea secvenţială a enzimelor este binecunoscută în căile de biosinteză a metaboliţilor secundari la fungi şi streptomicete
135

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
(Martin şi Liras, 1989). Cuplarea strânsă a proceselor de transcriere/traducere la streptomicete şi la alte procariote, precum şi faptul că genele structurale sunt grupate şi ordonate permite ca enzima obţinută în urma acestor fenomene să acţioneze asupra produsului enzimei anterioare (Motamedi şi Hutchinson, 1987).
O acţiune mai eficientă a enzimelor poate rezulta în urma grupării enzimelor în complexe enzimatice, fenomen întâlnit în cazul biosintezei gramicidinei,tirocidinei şi bacitracinei. Poliketidele şi metaboliţii secundari polienici sunt formaţi prin acţiunea unor complexe enzimatice care cuprind sintetaze specifice.
2.4. Coordonarea exprimării genelor de biosinteză şi rezistenţă
Deoarece genele implicate în biosinteza şi rezistenţa la antibiotice sunt grupate este foarte probabil să existe o coordonare a exprimării genelor pentru a nu produce disfuncţii. Dacă un metabolit secundar este activ faţă de tulpina producătoare, mecanismele de protecţie (inactivarea antibioticului sau sporirea eliminării lui) trebuie să fie induse înaintea acumulării metabolitului respectiv.
Există numeroase exemple referitoare la inducerea exprimării genelor de rezistenţă la antibiotice la bacteriile din genul Streptomyces. Astfel, exprimarea genelor de rezistenţă la tetraciclină, tetA şi tetB de la S.rimosus şi S.griseus a fost indusă de adăugarea de tetraciclină în mediul de cultură (Ohnuki şi colab., 1985). Nivelul rezistenţei a crescut de 2 ori atunci când s-a mărit cantitatea de tetraciclină adăugată (de la 5 la 50μg/ml). Aceasta sugerează faptul că inducerea genelor de rezistenţă protejează tulpinile producătoare faţă de creşterea nivelului antibioticului în timpul cultivării.
Gena de rezistenţă la streptomicină (strA sau aphD) de la S.griseus se află plasată alături de genele pentru biosinteza antibioticului şi se găseşte sub un control dublu, pozitiv şi negativ. Ea este indusă de streptomicină, dar nu şi de streptavidin. Exprimarea genei strA se află sub un control negativ exercitat de o proteină represor de 40 KDa codificată de o secvenţă (ORF) plasată în apropierea acestei gene. De asemenea, ea este stimulată de produsul genei strR care activează în acelaşi timp şi gena strB (genă structurală care codifică o amidinotransferază) (Ohnuki şi colab., 1985) (Figura 5).
Figura 5. Model ipotetic al transcrierii genelor implicate în sinteza şi rezistenţa la streptomicină (după Piepersberg şi colab., 1988).
In mod similar, gena de rezistenţă la kanamicină clonată de la bacteria producătoare S.kanamyceticus se exprimă după inducere în noile gazde (S.lividans, S.lavendulae sau S.parvulus) (Nakana şi colab., 1984).
S-a dovedit că multe dintre tulpinile de Streptomyces producătoare de antibiotice prezintă două sau mai multe mecanisme de rezistenţă la antibioticele pe care le produc, probabil ca o modalitate de protecţie faţă de antibiotice înrudite. Spre exemplu, în cazul tulpinilor de
136

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
S.kanamyceticus există două mecanisme de rezistenţă la kanamicină: una este modificarea ribozomilor, iar cealaltă presupune inactivarea antibioticului de către o enzimă specifică (6'-N-acetiltransferaza) (Nakano şi colab., 1988). La S.fradiae, la o tulpină industrială au fost identificate cel puţin trei gene diferite care asigură rezistenţa la tilozin, gene care sunt linkate la genele de biosinteză.
In unele situaţii, enzima codificată de gena de rezistenţă poate îndeplini şi alte funcţii catalitice în cadrul căii metabolice de biosinteză a antibioticului respectiv. Acesta este cazul genelor de rezistenţă la puromicină sau bialafos, care se pare că sunt implicate şi în unele dintre etapele intermediare ale sintezei metabolitului respectiv, ceea ce înseamnă că exprimarea genelor de biosinteză şi rezistenţă se află sub acelaşi mecanism de reglare.
2.5. Reglarea exprimării genelor implicate în metabolismul secundar
Metabolismul secundar este o formă de diferenţiere biochimică a microorganismelor. Reglarea desfăşurării sale este realizată prin diferite mecanisme. Astfel, factorul A este o proteină reglatoare cu efecte pleiotropice care stimulează atât biosinteza şi rezistenţa la streptomicină, cât şi sporularea la S.griseus şi S.bikinensis. Au fost identificate, astfel, mai multe gene ai căror produşi de transcriere controlează diferite procese. Un exemplu este reprezentat de gena afsA ("A factor synthesis"), care a fost izolată de la S.bikinensis, şi apoi clonată în diferite gazde. O altă genă, afsB a fost izolată de la S.coelicolor şi s-a dovedit că ea codifică o proteină reglatoare care stimulează atât producerea de factor A, cât şi a unor pigmenţi cum ar fi actinorhodinul şi undecilprodigiozinul.
O situaţie interesantă o prezintă gena saf ("secondary metabolism-acting factor") izolată de la S.griseus, genă care este implicată într-un mecanism de control comun pentru diferenţierea celulară, pentru producerea a cel puţin patru enzime extracelulare şi pentru formarea de pigment (Martin şi Liras, 1989). Secvenţa aminoacizilor pe care îi conţine produsul genei saf indică faptul că acesta interacţionează cu ADN, secvenţa respectivă conţinînd un domeniu tipic de legare la ADN. Aceasta înseamnă fie că polipeptidele de legare la ADN interacţionează specific cu secvenţe reglatorii ale genelor implicate în metabolismul secundar, fie că ele controlează exprimarea genelor care determină formarea factorului A sau a altor efectori pleiotropici, influenţând, astfel, exprimarea grupurilor de gene care codifică enzimele metabolismului secundar.
Genele implicate în biosinteza metabolismului secundar nu sunt exprimate, de obicei, în culturile cu o rată mare de creştere. Astfel, numeroase sintetaze aparţinând metabolismului secundar sunt fie inhibate fie represate de nivelul surselor de carbon, fosfor, sau azot în timpul fazei de creştere a producătorilor (Martin, 1989).
Un alt mecanism de coordonare a exprimării genelor, evidenţiat iniţial la B.subtilis, E.coli, dar şi la alte bacterii, este producerea unor factori sigma sau a altor proteine care modifică specificitatea ARN polimerazei, conducând la o exprimare selectivă a unor seturi speciale de gene. Si în cazul streptomicetelor a fost evidenţiată o heterogenitate a ARN polimerazelor (Westpheling şi col.ab, 1985). Spre exemplu, două holoenzime de ARN polimerază (E 35 şi E 49) de la S.coelicolor recunosc doi promotori bine definiţi de la B.subtilis. Promotorii de la streptomicete sunt destul de variaţi: unii care prezintă conservate regiunile -10 şi -35 par a fi similari celor de la E.coli, în timp ce alţii sunt complet diferiţi (Jansen şi colab, 1985). Distanţa dintre cele două regiuni conservate, -10 şi -35, este un factor important al determinării tăriei promotorului. Astfel, o distanţă de aproximativ 17 perechi de baze este corelată cu o activitate optimă a promotorului, în timp ce o distanţă mai mică de 15pb sau mai mare de 20pb este asociată cu o scădere a activităţii sale.
Date recente (Buttner şi colab., 1988) indică faptul că gena pentru agarază (dagA) de la S.coelicolor care conţine patru promotori diferiţi este transcrisă de cel puţin trei forme diferite
137

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
(holoenzime) de ARN polimerază. Similar, genele galE şi galK de la S.lividans sunt transcrise de două ARN polimeraze de la nivelul a doi promotori, galP1 şi galP2.
Deşi existenţa mai multor factori sigma este bine cunoscută, semnificaţia fiziologică a diferitelor holoenzime de ARN polimerază rămâne încă neclară. La streptomicete multe dintre secvenţele aflate înaintea regiunii start a transcrierii unor gene conţin doi sau mai mulţi promotori dar nu este clar dacă toţi promotorii funcţionează "in vivo" (Tomich, 1988). Recunoaşterea diferenţiată a promotorilor aranjaţi în tandem de către diferite ARN polimeraze poate explica exprimarea selectivă a anumitor seturi de gene în timpul creşterii şi diferenţierii morfologice şi a altor gene în timpul desfăşurării metabolismului secundar. Aranjarea genelor implicate în biosinteza antibioticelor în grupuri ("clusteri") este deci de mare interes în legătură cu exprimarea diferenţiată a genelor.
2.6. Seminificaţia biologică a metaboliţilor secundari
Semnificaţia biologică a metaboliţilor secundari produşi de diferite microorganisme nu este încă foarte clară. Sinteza lor ar reprezenta rezultatul unor "erori de metabolism" fără importanţă pentru fiziologia microorganismului producător. In legătură cu rolul metaboliţilor secundari au fost avansate numeroase propuneri pornind de la studiul în laborator al activităţii acestor compuşi. Vining (1990) consideră că funcţiile metaboliţilor secundari pot fi grupate în două categorii: unele care privesc direct producătorul (funcţii intrinseci) şi altele care asigură anumite beneficii producătorului prin acţiunea în exterior, în mediu (funcţii extrinseci). In cadrul funcţiilor intrinseci, unii metaboliţi secundari pot fi precursori ai unor componente structurale cum ar fi de exemplu streptomicina care, se sugerează că ar fi un constituent al învelişului mureinic (Szabo şi colab., 1989). De asemenea, alţi metaboliţi secundari joacă un rol important în sporulare şi în germinarea sporilor la actinomicete şi la bacili ca şi în diferenţierea celulară (Chater, 1989; Maplestone şi colab.,1992).
Atunci când activitatea metaboliţilor secundari este direcţionată asupra altor organisme din mediul înconjurător, aceste substanţe pot fi considerate ca "agenţi ecologici" asigurând pro-ducătorilor supravieţuirea în cadrul competiţiei cu alte organisme ce populează aceeaşi nişă ecologică. De asemenea, datele obţinute până în prezent în urma analizei secvenţei de nucleotide sprijină conceptul că metabolismul secundar a rezultat în urma modificărilor căilor existente ale metabolismului primar. Deşi secvenţa de aminoacizi identificată deductiv din secvenţa de nucleotide a genelor corespunzătoare este suficientă pentru a indica o origine comună, pentru ca datele să fie mai concludente ar fi necesară o comparare a genelor de la specii diferite decât de la aceeaşi specie. Informaţiile obţinute în acest fel sugerează că transferul natural de gene între diferite organisme a reprezentat un important factor în evoluţia metaboliţilor secundari. Multe dintre căile metabolismului secundar pot avea o origine foarte veche, modificările care apar la nivelul lor putând rezulta în urma achiziţiilor mai recente de material genetic exogen la nivelul unor microorganisme producătoare.
Pe baza cunoştinţelor acumulate în domeniu, Vining (1992) stabileşte o serie de caracteristici clare ale metabolismului secundar:
1. metaboliţii secundari nu sunt esenţiali pentru creştere şi tind să fie specifici de tulpină;2. aceşti compuşi au o mare varietate de structuri chimice şi activităţi biologice;3. metaboliţii secundari derivă din intermediari ai metabolismului primar prin căi de
biosinteză unice. Aceste căi sunt deseori lungi şi complexe; reacţiile biochimice sunt catalizate de enzime cu specificitate de substrat diferită de cea a enzimelor echivalente ale metabolismului primar;
4. formarea metaboliţilor secundari este determinată de seturi de gene asociate supuse unui mecanism de reglare care se referă atât la nivelul exprimării genelor cât şi la momentul declanşării acesteia;
138

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
5. mecanismele de control este bine integrat cu fiziologia organismului producător.Relativ recent, Davies (1990) a propus ideea că metaboliţii secundari sunt substanţe cu
origine foarte veche şi că mulţi dintre ei exercită activităţi biologice cunoscute (cum sunt antibioticele) datorită interacţiunii cu anumite situsuri vechi, conservate ale diferitelor macromolecule. Aceasta înseamnă că metaboliţii secundari au jucat un rol important în evoluţia reacţiilor biochimice, înainte ca enzimele să fie disponibile. Spre exemplu, reacţiile de biosinteză erau catalizate sau modulate de molecule cu greutate moleculară mică care erau prezente în atmosfera Pământului sau erau produse în "supa primordială". Odată ce sistemul de traducere a evoluat iar proteinele au început să fie produse şi să exercite diferite funcţii, rolul produşilor cu greutate moleculară mică a fost înlocuit de mecanisme mai complexe în care sunt implicate polipeptidele, dar aceştia şi-au menţinut capacitatea de a interacţiona cu anumite situsuri receptoare din macromolecule (acizi nucleici sau proteine).
Conceptul rolului cheie primordial al moleculelor cu greutate moleculară mică ca efectori (stimulatori) ai biosintezelor sugerează o nouă abordare a modalităţilor de screening pentru diferite activităţi farmacologice. In locul urmăririi exclusive a efectelor inhibitorii este necesar să se studieze activitatea stimulatoare fie a unor celule întregi fie a unor extracte celulare (Murakama şi colab.,1989). Numeroase organisme (poate chiar toate) produc compuşi cu greutate moleculară mică cu activitate antibiotică, cum ar fi de exemplu defensinele (Gabai şi colab., 1989). In acest sens, microorganisme de tipul streptomicetelor sunt neobişnuite în sens evolutiv deoarece ele şi-au dezvoltat şi menţinut căile necesare pentru a produce o gamă foarte largă de compuşi cu greutate moleculară mică atunci când sunt cultivate în mod corespunzător. Acţiunea acestor compuşi este foarte variată, ei inhibând de obicei desfăşurarea unor procese biochimice diferite. Studii mai amănunţite asupra activităţii acestor substanţe antibiotice a evidenţiat însă că multe dintre ele au efect stimulator asupra transferului de gene (aminoglicozidele, penicilinele, tetraciclinele), asupra transpoziţiei (tetraciclina), transcrierii (eritromicina, tiostreptonul), creşterii celulare (kanamicina, streptomicina, eritromicina) sau mutagenezei (streptomicina). Pornind de la aceste observaţii, Davies (1990) propune că efectorii cu greutate moleculară mică şi situsurile lor de acţiune datează dinaintea stabilirii rolului proteinelor, deşi acţiunea lor este mai puţin eficientă şi are o specificitate scăzută. Odată ce organismele devin din punct de vedere genetic şi biochimic mai evoluate, rolul efectorilor cu greutate moleculară mică este preluat de proteine care asigură o eficienţă şi specificitate de acţiune ridicată.
3. Enzime produse de bacteriile din genurile Bacillus si Streptomyces
Deşi streptomicetele produc mai mult de 60% dintre antibioticele cunoscute, există relativ puţine date referitoare la echipamentul enzimatic al acestor bacterii. Tinând cont de faptul că streptomicetele sunt microorganisme din sol unde există o mare varietate de posibile substraturi nutritive, este logic de presupus că aceste bacterii prezintă o mare varietate de căi metabolice putând sintetiza unii intermediari prin căi diferite. Căile metabolice alternative fac posibilă utilizarea eficientă a celor mai multe substrate disponibile (Behal, 1986).
In ultimii ani bacteriile din genul Streptomyces constituie un serios subiect de studiu din punctul de vedere al enzimelor pe care le produc, multe dintre ele fiind de importanţă biotehnologică (proteaze, amilaze, glucoz-izomeraze, celulaze, hemicelulaze, endonucleaze, chitinaze etc. Mai mult, s-a dovedit că cel puţin unele dintre aceste enzime sunt produse simultan cu unele antibiotice (tabelul 2) (Pokorny şi Vitale, 1980).
Unele dintre enzimele sintetizate de către streptomicete sunt sau pot fi folosite în practica medicală fie în stabilirea unor diagnostice fie în terapie. Acesta este cazul colesterol-oxidazei produsă de unele specii de Streptomyces şi Nocardia care este utilizată pentru dozările de
139

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
colesterol din ser, sau al urat-oxidazei produsă de S.cyanogenus care poate fi utilizată pe scară largă pentru detectarea enzimatică a acidului uric (Srinivasan şi colab., 1991).
Producerea enzimelor hidrolitice de către streptomicete depinde în mare măsură de substratul pe care acestea sunt cultivate astfel că datele prezentate în Tabelul 2 pot fi completate şi cu alte tipuri de enzime. Uneori însă, datorită scopului în care enzimele ar putea fi utilizate (de exemplu în industria alimentară) se preferă utilizarea unor tulpini bacteriene producătoare care însă să nu sintetizeze substanţe toxice sau potenţial toxice (cum sunt de altfel antibioticele)
Tabelul 2. Enzime entracelulare produse de unele bacterii producătoare de antibioticeSpecia bacteriană Antibioticul Enzimele detectate
S.griseus streptomicina proteaze, elastaze, amilaze, peptidazeS.rimosus oxitetraciclin proteaze, elastaze,
S.aureofaciens tetraciclina proteaze, amilazeS.fradiae neomicina proteaze, elastaze, keratinaze
S.antibioticus oleandomicina proteaze, amilazeS.venezuelae cloramfenicol Proteaze
B.licheniformis Bacitracina ProteazeB.subtilis Bacitracina Proteaze
De asemenea, studii recente au demonstrat posibilitatea utilizării bacteriilor din genul Streptomyces pentru clonarea unor gene cu origine diferită şi pentru producerea proteinelor corespunzătoare (Erpicum şi colab., 1990). In plus, metaboliţii secundari ai acestor bacterii oferă avantajul exprimării (sintezei) prelungite a unui produs şi după faza iniţială de creştere a culturii (Gardner şi Cadman, 1990).
In cazul enzimelor produse de bacteriile din genul Bacillus, aplicaţiile practice sunt bine cunoscute, ele fiind prezentate pe scurt în Tabelul 3.
Tabelul 3. Tipuri de enzime hidrolitice produse de bacili şi principalele aplicaţii ale acestoraEnzima Tipul enzimei Specia producătoare UtilizăriProteaze alcaline Extracelulare B. subtilis
B. pumillusB. amylosacchariticusB. licheniformis
detergenţiindustria alimentarăindustria pielăriei
Proteaze neutre Extracelulare B.subtilisB. amyloliquefaciensB. thermoproteolyticusB. amylosacchariticusB. stearothermophilusB. cereus
Industria beriiIndustria alimentară
α-amilaze Extracelulare B. subtilisB.licheniformisB.coagulansB. stearothermophilusB. cereus var.mycoides
Industria alimentarădegradarea amidonului
Β-amilază Extracelulare B. polymyxa B.circulansB. megatheriumB. cereus var.mycoides
industria alimentarădegradarea amidonului
Celulaze Extracelulare B.subtilisB.polymyxaB. licheniformis B. cereus
Industria alimentarăIndustria textilăIndustria celulozei şi hârtieiZootehnieTratarea deşeurilor agricole
140

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
Datorită acestor considerente o trecere în revistă a principalelor tipuri de enzime (cu posibilă valoare biotehnologică) este nu numai interesantă ci şi necesară.
3.1. Proteaze
Proteazele microbiene constituie un grup complex de enzime hidrolitice, care diferă prin specificitatea de substrat, mecanism de reacţie, dependenţa activităţii enzimatice şi stabilităţii în funcţie de pH şi temperatură. Acţiunea lor poate fi blocată, activată, sau inhibată într-o anumită proporţie de agenţi fizici, chimici sau biologici. Producţia şi caracteristicile enzimelor microbiene pot fi influenţate prin selecţia tulpinii producătoare şi alegerea condiţiilor de cultivare ale acesteia.
Enzimele din categoria proteazelor hidrolizează majoritatea proteinelor comune cum ar fi: caseina, hemoglobina, gelatina, albumina de ou, glutenul, proteina din soia şi alte proteine vegetale şi animale. Condiţiile de hidroliză precum şi randamentul operaţiei în sine depind însă de natura, caracteristicile şi gradul de puritate al preparatului proteolitic utilizat.Numeroasele utilizări ale enzimelor proteolitice, în cele mai diferite domenii de activitate (industria alimentară, farmaceutică, agricultură, industria detergenţilor, medicină) au determinat demararea cercetărilor în vederea obţinerii acestora pe cale biotehnologică ţinând seama în acelaşi timp de avantajele pe care le oferă utilizarea microorganismelor ca sursă biologică, precum şi de proprietăţile enzimelor microbiene.
Proteazele reprezintă un grup de enzime hidrolitice, de obicei extracelulare, care scindează molecula proteică în fragmente polipeptidice formate din câţiva aminoacizi uniţi prin legături peptidice. Aceste polipeptide pot fi în continuare degradate de către peptidaze până la aminoacizii componenţi. Peptidazele, în funcţie de modul lor de acţiune sunt de două tipuri: endopeptidaze şi exopeptidaze. La rândul lor exopeptidazele pot fi: aminopeptidaze care îşi încep acţiunea de la nivelul extremităţii NH2 libere a polipeptidului, activitatea lor depinzând de prezenţa unor ioni metalici; carboxipeptidaze care îşi încep atacul de la capǎtul COOH liber al peptidului (Scriban, 1988).
Clasificarea enzimelor proteolitice se face, conform recomandǎrilor International Unit of Biochemistry în şase familii, diferenţele dintre ele datorându-se secvenţelor de aminoacizi care alcătuiesc situsul activ (Tabelul 4) (Neurath, 1989).
Tabelul 4. Familii de enzime proteolitice (după Neurath, 1989)Familia Proteaze reprezentative Resturi caractiristice de
aminoacizi ale situsului activSerin proteaze I Chimiotripsina, tripsina, elastaza,
kalikreina,pancreatinaAsp102Ser195His57
Serin proteaze II Subtilizina Asp32Ser221His64
Cistein proteaze Papaina, actinidin, catepsina B şi H de la şobolan
Cys25His159Asp158
Aspartic proteaze Penicilopepsin, renina, proteaze acidede la Rhizopus sp.
Asp33Asp213
Metaloproteaze I Carboxipeptidaza A bovină Zn, Glu270 Try248
Metaloproteaze II Termolizina Zn Glu143His231
Membrii fiecărei familii se crede că au rezultat dintr-o formă ancestrală comună, în urma
unui proces evolutiv divergent. Din datele prezentate în tabelul 2 se remarcă faptul că serin proteazele includ două familii distincte: serin-proteazele de la mamifere şi cele de origine bacteriană. Deşi prezintă un mecanism enzimatic comun, cele două familii de serinproteaze diferă ca secvenţă de aminoacizi şi ca structură tridimensională.
141

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
In mod similar, metalopreoteazele includ şi ele tot două familii în funcţie de originea lor şi de structura chimică: de la mamifere şi de la bacterii. Există şi alte enzime proteolitice identificate la diferite grupe de organisme care nu pot fi încadrate încă în nici una dintre grupele de mai sus deoarece mecanismul lor de acţiune şi situsul activ nu este cunoscut cu exactitate. Acesta este cazul colagenazelor şi aminopeptidazelor.
Proteazele bacteriene produse de tulpini de Bacillus subtilis sunt amestecuri de proteaze alcaline (subsubclasa 3.4.21, proteaze serinice) şi proteaze neutre (subsubclasa 3.4.24, metaloproteaze).
Proteazele serinice au în centrul lor activ serina şi histidina. Acest grup a fost divizat de Morihara în alte 4 subgrupe:
proteaze similare tripsinei; proteaze alcaline; proteaze Myxobacter α-litice; proteaze stafilococice.Proteazele similare tripsinei cuprind, în afară de tripsină şi chimotripsină, enzime
produse de specii de Streptomyces, care hidrolizează legăturile Arg22 – Gly23 şi Lys29 – Ala30
din lanţul β-insulinic, această specificitate de substrat fiind trăsătura distinctivă.Aceste enzime acţionează la pH 8,0, au temperatura optimă de cataliză 50 C, sunt
inhibate de diizopropilfluorfosfat (DIFP), p–cloromercurilbenzoic (PCMB), tosyl-L-lizin-clorometilcetonă (TLCK), tripsin-inhibitorul din soia. Activitatea lor nu este influenţată de etilendiaminotetraacetat (EDTA) şi acid iod-acetic (IAA), în plus prezintă şi activitate esterazică şi amidazică.
A doua grupă de proteaze serinice sunt proteazele alcaline; acestea pot fi produse de bacterii, drojdii şi fungi fiind prezente şi în ţesuturile animale. În această grupă se încadrează şi proteazele alcaline produse de unele tulpini de B.subtilis şi cele fungice din Aspergillus oryzae.
Aceste proteaze au acţiune specifică faţă de resturile de aminoacizi aromatici sau hidrofobi care participă cu gruparea carboxil la legătura care se scindează; valoarea optimă a pH este cuprins între pH 9 şi pH 11, dar enzimele rămân active în domeniul de pH 5–9; aceste proteaze sunt inhibate de DIFP, de PMSF fiind însă rezistente şa acţiunea EDTA sau ortofenantrolinei. Din acest grup fac parte proteazele cunoscute sub denumirea de subtilizine sau subtilopeptidaze. S-a constatat că aceste proteaze nu sunt exclusiv produse de B.subtilis ci în general de bacterii ale genului Bacillus astfel încât s-a propus numele generic de bacilopeptidaze. Respectivele enzime pot fi clasificate în două grupe:
Bacilopeptidaze A care cuprind subtilizina Carlsberg (proteaza alcalină produsă de tulpini de B.licheniformis) şi proteaza alcalină produsă de B.pumilis;
Bacilopeptidaze B care includ subtilizina NOVO (protează alcalină sintetizată de B.subtilis NRRL B3411) şi subtilizina BNP (Bacterial Protease Nagarse), protează alcalină din B. subtilis var. amylosacchariticus.Metodele de examinare a amestecurilor de enzime produse de diferiţi bacili au arătat
că bacteriile producătoare de bacilopeptidaze A nu sintetizează proteaze neutre sau amilază, în timp ce bacteriile producătoare de bacilopeptidaze B sintetizează în acelaşi timp şi proteaze neutre şi amilază în acelaşi timp.
Metaloproteazele sunt proteaze activate de ionii metalici legaţi de centrul activ al enzimei (Ca2+, Zn2+, Mg2+, Fe2+). Ele pot fi metaloproteaze neutre şi metaloproteaze alcaline. Proteazele neutre conţin de obicei Zn şi prezintă specificitate faţă de aminoacizii hidrofobi implicaţi cu gruparea amino în legătura peptidică. Ele au un pH optim de acţiune cuprins între pH 7 şi pH 8, sunt inhibate complet de EDTA (care complexează atomul de Zn din structură) fiind sensibile la concentraţii de EDTA sub 10-3M, orto-fenantrolină, nefiind afectate de DIFP şi PCMB.
142

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
Metaloproteazele alcaline conţin, de asemenea, un ion metalic implicat în mecanismul de reacţie catalitică. Ele îşi manifestă activitatea optimă în domeniul de pH 7–9, fiind mai puţin inhibate de EDTA decât proteazele neutre, necesitând concentraţii mai ridicate de 10 -2 M pentru inactivare.
Proteazele neutre produse de Bacillus subtilis var. amylosacchariticus sunt diferite de proteazele neutre produse de Bacillus subtilis NRRL B3441 şi din acest motiv este necesară specificarea exactă a provenienţei proteazei neutre respective. Producerea lor este supusă represiei exercitate de prezenţa în mediul de incubare a unor aminoacizi în cantitate mai mare (Bawden şi colab.,1987). Aceste enzime sunt folosite în prezent la fabricarea detergenţilor la tratarea pieilor în industria pielăriei, frăgezirea cărnii, eliminarea depozitelor proteice din băuturile fermentate (Sasson, 1988). La aceste bacterii s-au realizat şi numeroase studii referitoare la determinismul genetic şi controlul producerii şi excreţiei acestor enzime în scopul obţinerii prin clonare a unor tulpini superproducătoare (Mc Connell şi colab., 1986).
S-a dovedit faptul că producerea de proteaze este supusă unor mecanisme reglatoare complexe, între care un rol cheie este jucat de sistemul DegS-DegU precum şi de factorii de reglare DegR and DegQ (Nagami and Tanaka, 1986; Tanaka et al., 1987; Ogura et al., 1994). Aceşti factori reglatori se pare că sunt implicaţi în superproducţia de proteaze cel puţin în cazul unor tulpini de B.subtilis. De asemenea, relativ recent, Ogura et al (1994) au caracterizat alte două gene: proB (ce codifică sinteza γ-glutamil kinazei) şi proA (codifică sinteza glutamil-γ-semialdehid dehidrogenaza) ale căror produşi par a fi implicaţi în sporirea capacităţii de biosinteză a exoproteazelor la o tulpină de B.subtilis. Astfel, cele două gene au fost clonate într-un vector e clonare (pLC1) şi apoi introduse, împreună cu un alt vector (pNC61) ce conţine gena reglatoare prtR (degR) într-o tulpină de B.subtilis. Transformanţii obţinuţi au prezentat o activitate proteolitică mai mare decât a tulpinii parentale.
Date fiind aplicaţiile practice ale acestor enzime extracelulare, preocupările cercetătorilor vizează sporirea nivelului de biosinteză ca şi mărirea stabilităţii enzimelor în diferite condiţii experimentale. Aceste deziderate se pot realiza pe mai multe căi, printre care cele ce utilizează metodele clasice de ameliorare a microoganismelor (prin tratament cu agenţi mutageni) şi cele moderne, de inginerie genetică (fuziunea de protoplaşti şi tehnologia ADN recombinant).
Conform datelor de literatură, mutagenii cei mai eficienţi sunt nitrosometiluree, nitrosoguanidina, dietilsulfatul, precum şi agenţi fizici de tipul radiaţiilor X sau a radiaţiilor . Astfel, prin mutageneză s-a reuşit ridicarea nivelului de biosinteză a tulpinilor de B.subtilis de aproximativ 8-10 ori.
În cel de-al doilea caz, majoritatea tulpinilor superproducătoare de proteaze care prezintă anumite caracteristici s-au obţinut prin inginerie genetică. Astfel, sunt foarte cunoscute rezultatele obţinute de cercetătorii japonezi care, pornind de la tulpini sălbatice de B.thermoproteolyticus şi B.stearothermophilus au reuşit obţinerea de tulpini producătoare de termolizină, o protează foarte diferită de celelalte proteaze neutre în ceea ce priveşte termostabilitatea şi mecanismul de acţiune. În prezent, cunoscându-se structura termolizinei precum şi genele răspunzătoare de caracteristicile sale deosebite, se încearcă obţinerea de tulpini de B.subtilis, B.stearothermophilus modificate genetic, capabile să producă proteaze termostabile sau stabile la valori diferite ale pH sau la acţiunea unor agenţi inhibitori.
In cazul bacteriilor din genul Streptomyces examinarea lichidelor de cultură a unor tulpini producătoare de antibiotice a dus la evidenţierea prezenţei unui amestec de enzime dintre care, un loc important din punct de vedere cantitativ îl ocupă proteazele (neutre şi alcaline) şi peptidazele (aminopeptidaze) (Pokorny şi Vitale, 1980).
Vitale şi colab.(1980) au reuşit izolarea din filtrat de cultură de S.rimosus a unei serin proteaze care a prezentat un pH optim de acţiune cuprins între 7 şi 9 şi al cărei substrat de acţiune este reprezentat de proteine sau de peptide mari, hidrolizând în general esterii terminali simpli.
143

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
Recent, Renko şi colab.(1989) au prezentat particularităţile unei proteaze de tip tripsinic ("trypsin-like proteinase") izolată tot din filtrat de cultură de S.rimosus evidenţiind faptul că aceasta este o protează alcalină, sensibilă la acţiunea inhibitorului proteazic PMSF (specific pentru serin-proteaze) ca şi la cea a α-antitripsinei. Aceste enzime au activitate arginil-aminopeptidazică atât asupra proteinelor cât şi asupra peptidelor mai mici. Enzime similare au fost izolate şi caracterizate şi de la alte specii de Streptomyces: S.fradiae, S.paromomycinus, S.moderatus, S.hygroscopicus etc. O situaţie oarecum specială o prezintă S.griseus care produce un amestec de proteaze neutre şi alcaline, produsul obţinut fiind denumit pronază, el fiind studiat mai intens în scopul elucidării mecanismului de acţiune şi a omologiei sale cu tripsinele vertebratelor (Read şi colab.,1984).
Aceste studii au importanţă atât teoretică prin contribuţiile aduse la înţelegerea unor aspecte evolutive, a taxonomiei serin-proteazelor dar şi practică, aceste enzime putând înlocui tripsina în anumite reacţii biochimice.
S-a dovedit de asemenea că unele specii de streptomicete, cum ar fi de pildă S.rimosus, pot produce şi o altă categorie de proteaze: metaloproteaze (Vitale şi colab.,1987). Aceste enzime sunt fie bazice fie neutre, fiind activate de ionii de calciu, mangan sau zinc, stabilizate de ionii de calciu şi inhibate de EDTA. Metaloproteazele alcaline acţionează doar asupra proteinelor, nu şi asupra peptidelor în timp ce metaloproteazele neutre degradează în egală măsură proteinele şi peptidele. In cazul acestora din urmă acţiunea enzimatică se manifestă la extremitatea COOH, ceea ce permite încadrarea lor printre dipeptidil-carboxipeptidaze. De asemenea, s-a reuşit izolarea de la mai multe specii de Streptomyces (S.rimosus, S.fradiae, S.peptidofaciens, S.griseus) a unor peptidaze care, pe baza proprietăţilor specifice (specificitatea de substrat, inhibarea de către EDTA, activare de către ionii de calciu, magneziu, prezenţa leucinei la capătul NH2 terminal) au fost identificate ca fiind leucin-amino-peptidaze (Vitale şi colab.,1986).
Tinând cont că în ultima vreme streptomicetele au început să fie utilizate drept gazde pentru clonarea de material genetic heterolog, este interesant de ştiut dacă tulpinile utilizate de obicei drept acceptori elaborează sau nu proteaze. Producerea unor asemenea enzime la aceste tulpini constituie un serios impediment în obţinerea produsului dorit. Pornind de la această idee, Aretz şi colab.(1989) au studiat prezenţa enzimelor proteolitice la tulpina S.lividans TK24, tulpină recombinată obţinută pentru prelucrarea şi secretarea unei proteine de fuziune (între inhibitorul α-amilazei de la S.tendae şi proinsulina de la maimuţa Macacca fascicularis). După purificarea prin cromatografie pe schimbători de ion au fost separate mai multe tipuri de enzime proteolitice: alanin-aminopeptidaze, leucin-aminopeptidaze, proteaze de tip tripsinic şi proteaze de tip chimiotripsinic. Dintre acestea, proteazele de tip chimiotripsinic ("chemo-trypsin-like") sunt cele care degradează proteina de fuziune dar această acţiune poate fi inhibată prin adăugarea în mediul de culturăa unor săruri de zinc sau nichel.
Dacă în cazul bacililor au fost izolate şi clonate genele ce codifică sinteza a două subtilizine şi a unei proteaze neutre (Mc Connell şi colab.,1986), în cazul bacteriilor din genul Streptomyces studiile referitoare la determinismul genetic al producerii de proteaze sunt mult mai puţine. Astfel, Duez şi colab.(1987) au reuşit izolarea, clonarea şi secvenţierea genei pentru o peptidază extracelulară de la Streptomyces sp.R61 la S.lividans, iar Henderson şi colab.(1987) au caracterizat genele pentru proteazele A şi B de la S.griseus.
In cazul unor tulpini de streptomicete au fost evidenţiate unele enzime proteolitice care degradează preferenţial colagenul ("collagenase-like")(Mukhopadhyay şi Chandra, 1987). Deşi nu au fost izolate şi purificate (s-au utilizat filtrate de cultură) enzimele de tip colagenaze de la Streptomyces au evidenţiat o activitate specifică pe colagen din tendon bovin şi piele de viţel, la 280 C şi pH 7,2-7,5, fiind inhibate de EDTA şi azidă de sodiu. Deşi neobişnuită, producerea unor asemenea enzime de către streptomicete nu este surprinzătoare dacă ne gândim la multitudinea de substrate nutritive pe care aceste bacterii le pot găsi în sol.
144

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
3.2. α-Amilazele
α-Amilazele (1,4-α-D-glucan-glucanhidrolaza, EC 3.2.1.1.) sunt larg răspândite în rândul microorganismelor, atât la fungi cât şi la bacterii, mai ales la specii ale genurilor Bacillus şi Streptomyces, acţiunea lor constând în hidroliza legăturilor α-1,4-glicozidice de la nivelul amidonului, eliberând maltoza. Cel mai mult studiate şi utilizate în practică sunt α-amilazele produse de speciile de Bacillus (B.amilolyquefaciens, B.licheniformis, B.stearothermophilus, B.subtilis etc). Aceste bacterii produc două tipuri de α-amilaze: de lichefiere a amidonului (degradarea amidonului până la maltodextrine) şi de zaharificare (convertesc maltoza la glucoză). α-Amilazele produse de tulpinile de B.licheniformis prezintă un interes special datorită rezistenţei lor la temperaturi ridicate (85-1150C). In general, enzimele respective sunt utilizate în practică în lichefierea amidonului în industria distilatelor ca şi în industria textilă (Zarnea şi colab.,1980).
O serie de studii genetice realizate prin experimente de mutageneză sau prin clonări de gene au evidenţiat faptul că gena pentru α-amilază de la B.subtilis se află sub un control genetic complex care este însă puţin cunoscut la nivel molecular (Mc Connell şi colab.,1986). Se pare însă că există mai multe gene reglatoare cu efecte pleiotropice (sacU, sacQ, prtR) care intervin în exprimarea genei pentru α-amilază, levansucrază, serin protează, proteaze neutre şi β-glucanază (Priest, 1987).
In cazul streptomicetelor au fost izolate şi caracterizate α-amilaze de la mai multe specii, acest tip de enzime părând a fi sintetizat practic de majoritatea bacteriilor aparţinând acestui grup. O atenţie specială a fost acordată α-amilazei produsă de S.hygroscopicus (Hoshiko şi colab.,1987), S.limosus (Virolle şi Bibb, 1988) şi S.venezuelae (Virolle şi colab.,1988) la aceste specii fiind studiat şi genele ce codifică sinteza şi reglarea sintezei acestor enzime. De asemenea, s-au efectuat experimente de clonare a genelor respective în diferite gazde (E.coli, S.lividans) în scopul obţinerii unor tulpini superproducătoare de interes industrial. Pe lângă aspectul practic al cercetărilor de acest tip, ele mai pot oferi o serie de informaţii referitoare la modul de exprimare şi reglare a funcţiilor genelor de la streptomicete (în particular a genelor pentru α-amilază). Mai mult, rezultatele obţinute au permis o comparare a enzimelor de tip α-amilază produse de streptomicete cu cele produse de alte organisme, în special cu cele produse de eucarioate (Mac Gregor, 1988).
S-a dovedit că S.hygrosopicus produce o α-amilază care degradează amidonul până la maltoză, acţiunea sa fiind de tip endoglucanolitic (taie legăturile α-1,4 glicozidice din interiorul lanţului polizaharidic). Compararea secvenţei de aminoacizi pe care această enzimă o conţine cu cele ale altor enzime de acest tip, fie de la procariote (de la bacili) fie de la eucariote (de la Aspergillus oryzae, A.nidulans, Hordeum vulgare, Drosophila melanosgaster etc) a evidenţiat faptul că există trei regiuni care sunt conservate la toate aceste enzime, indiferent de origine (Ihara şi colab.,1985). Cu toate acestea, α-amilaza de la S.hygroscopicus este unică deoarece conţine triptofan la nivelul celei de-a treia regiuni conservate (Mac Gregor, 1988).
Experimentele de clonare a genei amy de la S.hygroscopicus în alte gazde cum ar fi E.coli şi S.lividans au evidenţiat faptul că, deşi transferul genei s-a realizat eficient, exprimarea sa variază în funcţie de gazdă şi de numărul de copii ale genei respective (efectul de dozare a genei). Hoshika şi colab. (1987) utilizând cartarea cu o nuclează specifică ("mung bean nuclease") au determinat faptul că gena amy de la S.hygroscopicus prezintă trei situsuri de iniţiere a transcrierii (trei promotori): amyP1, amyP2 şi amyP3, care au asemănări mai mult sau mai puţin extinse cu promotorii altor gene de la Streptomyces (endoH, ermP1, aphP1).
Studii similare au fost făcute şi cu gena pentru α-amilază (aml) de la S.limosus şi S.venezuelae. In cazul tulpinilor de S.venezuelae analizate s-a observat că exprimarea genei aml este represată puternic de glucoză în timp ce asupra exprimării genei similare de la S.limosus
145

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
glucoza nu are efect. Exprimarea acestei gene este însă influenţată (represată) puternic de manitol (Virolle şi colab.,1988). Interesant este că prin clonarea genei aml de la S.limosus în S.lividans exprimarea genei este puternic represată de glucoză în noua gazdă (Virolle şi Bibb 1988). Acesta înseamnă că rezistenţa la acţiunea glucozei evidenţiată la tulpinile de S.limosus analizate este mai degrabă o particularitate a bacteriei decât a genei pentru α-amilază. De asemenea, în urma comparării secvenţei de aminoacizi a acestei proteine cu proteine similare de la eucariote s-a observat o omologie ridicată cu α-amilaza produsă de Drosophila melanogaster (>40%), Mus musculus (36,8%), Aspergillus nidulans (36,1%) şi mai mică cu enzimele similare de la B.subtilis (22,6%) şi B.amylolyquefaciens (16,1%)(Long şi colab.,1987).
3.3. Glucoz-izomeraza
In ultimii ani, de mare interes în Japonia, SUA şi Franţa este obţinerea siropurilor cu conţinut ridicat de fructoză din hidrolizate de amidon în urma acţiunii glucoz-izomerazei (xilo-zoizomeraza, EC 5.3.1.5.). Printre producătorii recunoscuţi de asemenea enzime se numără şi unele specii de streptomicete: S.violaceoniger şi S.olivochromogenes. Suekane şi Iizuka (1982), în cadrul unor cercetări legate de producerea de glucoz-izomerază de către streptomicete au evidenţiat faptul că abilitatea de a converti glucoza la fructoză este larg răspândită între bacteriile acestui gen. Localizarea acestei enzime este atât intracelulară (cea mai mare parte) dar şi extracelulară.
Experimentele lui Szentirmai şi colab.(1986) au evidenţiat faptul că, cel puţin în cazul S.matensis, glucoz-izomeraza intracelulară diferă de cea extracelulară din punctul de vedere al greutăţii moleculare. De asemenea, s-a observat că utilizarea ca sursă de azot a unei combinaţii de glicocol şi sulfat de amoniu şi adăugarea xilozei şi a ionilor de cobalt (de magneziu sau mangan în cazul altor specii bacteriene) duce la stimularea producerii de enzimă la S.olivochromogenes, S.flavogriseus sau S.violaceoniger (Chen şi colab.,1979; Sicard şi colab., 1990).
Park şi Toma (1974) au observat că la unele tulpini de streptomicete există o interrelaţie între producerea de glucoz-izomerază şi cea de xilanază, în funcţie de sursa de carbon utilizată pentru cultivarea bacteriilor respective. Utilizarea unui mediu de cultură ce conţine fie xilan fie glucoză ca sursă de carbon determină producerea de glucoz-izomerază în timp ce xilanaza este produsă doar de bacteriile crescute pe mediu cu xilan, nu şi pe cel ce conţine glucoză.
Datorită importanţei practice deosebite a acestei enzime, în ultimii ani au fost realizate studii referitoare la determinismul genetic al sintezei glucoz-izomerazei ca şi la controlul acestei sinteze, scopul fiind acela al obţinerii prin tehnici de inginerie genetică a unor tulpini capabile de a sintetiza noi tipuri de glucoz-izomeraze cu proprietăţi modificate, cu acţiunea mai eficientă.
Utilizarea D-xilozei ca sursă de carbon de către bacterii se realizează pe o cale metabolică ce presupune transportul prin membrana plasmatică, izomerizarea la D-xiluloză şi apoi fosforilarea acesteia cu ajutorul xiluloz-kinazei cu formarea de D-xiluloz-5-fosfat. Intermediarul fosforilat este apoi metabolizat fie pe calea pentozo-fosfaţilor fie pe calea Embden-Meyerhoff. Pornind de la aceste date s-a determinat faptul că operonul xyl de la streptomicete (de la S.violaceoniger în particular) este alcătuit din cel puţin trei gene: xylA care codifică pentru sinteza xiloz-izomerazei, xylB care codifică xiluloz-kinaza şi xylX care determină sinteza unei proteine reglatoare. In cazul acestor bacterii s-a evidenţiat o situaţie interesantă: genele xylA şi xylB plasate apropiat una de cealaltă pe cromosomul bacterian prezintă o orientare diferită, exprimarea lor datorându-se unor promotori divergenţi plasaţi în regiunea dintre cele două gene (Tiraby şi colab.,1989). Produsul genei xylX pare a fi o proteină reglatoare care controlează atât exprimarea celor două gene cu orientare opusă cât şi propria sa sinteză.
Experimentele de clonare care se realizează în diferite laboratoare încearcă, pe de o parte stabilirea cu precizie a genelor ce alcătuiesc operonul xyl la streptomicete şi modul de reglare a
146

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
exprimării lor atât în tulpina de origine cât şi în diferite tulpini gazdă în care genele respective au fost clonate, şi, pe de altă parte să se construiască "in vitro" tulpini recombinate genetic capabile de a sintetiza glucoz-izomerază cu proprietăţi ameliorate (de exemplu termostabilă) (Sicard şi colab, 1990). Rezultatele obiţute până în prezent sunt promiţătoare, cercetările fiind continuate pe direcţiile menţionate mai sus.
3.4. Celulazele
Biomasa vegetală reprezintă sursa majoră de carbon în ecosistemele terestre astfel că este firească presupunerea că multe dintre microorganismele care populează aceste ecosisteme să fie capabile să degradeze aceste substrate, deci să sintetizeze enzime de timpul celulazelor şi hemicelulazelor. Actinomicetele şi în mod special bacteriile din genul Streptomyces constituie un grup important care intră în alcătuirea populaţiilor microbiene respective (Mc Carthy şi Williams, 1992).
Celuloza este un polimer neramificat de glucoză compus din unităţi de anhidro D-glucoză unite prin legături 1,4-β-D-glucozidice. Aceste legături pot fi hidrolizate de către enzimele celulozolitice. De obicei celuloza este foarte rezistentă la hidroliză datorită gradului ridicat de cristalinitate, rezultatul unei conformaţi lineare (paralele) care permite realizarea unor puternice legături de hidrogen între grupările OH ale lanţurilor paralele vecine. In fibrele de celuloză regiunile cu structură înalt cristalină coexistă cu regiunile cu structură amorfă iar cele cu un raport favorabil zonelor cristaline prezintă o rezistenţă crescută la acţiunea enzimatică. Celuloza, aşa cum se găseşte în natură este un homopolimer cu greutate moleculară mare, insolubil şi adesea în asociere intimă cu lignina, hemiceluloza la nivelul peretelui celular vegetal fapt ce o face puţin accesibilă degradării enzimatice (Robson şi Chambliss, 1989).
Toate organismele care degradează celuloza cristalină secretă sisteme enzimatice mai mult sau mai puţin complexe care sunt alcătuite din enzime cu diferite modalităţi de acţiune, specificitate de substrat şi care acţionează de obicei sinergic pentru a hidroliza celuloza. Cel mai bine studiat sistem celulazic este cel de la fungi (de la Trichoderma reesei), enzimele care îl alcătuiesc fiind grupate în trei categorii:
- exo- -1,4-glucanaze sau celobiohidrolaze (EC 3.2.1.91.)- endo- -1,4-glucanaze sau endocelulaze (endoglucanaze)(EC 3.2.1.4.)- -glucozidaza sau celobiaza (EC 3.2.1.21.)Aceste trei grupe de enzime acţionează sinergic ducând la hidroliza celulozei cristaline.
Endoglucanazele determină ruperea legăturilor interne ale lanţului de celuloză, atacul făcându-se la întâmplare şi ducând la formarea de oligozaharide. Efectul direct al acestui tip de enzime este fragmentarea lanţului de celuloză cuplat cu o creştere slabă a cantităţii de zaharuri reducătoare eliberate. De obicei, în complexele celulozolitice sunt cuprinse mai multe endoglucanaze care pot prezenta afinităţi diferite pentru celo-oligozaharide cu lungimi variate. Spre deosebire de aceste, exoglucanazele (celobiohidrolazele) acţionează la nivelul extremităţilor nereducătoare ale lanţului de celuloză, eliberând glucoză sau celobioză (zaharuri reducătoare) în cantitate crescută, lungimea lanţului celulozic nefiind modificată semnificativ.
Acţionând individual, atât endoglucanazele cât şi celobiohidrolazele pot degrada regiuni restrânse ale lanţului celulozic (cele cu structură amorfă). Pentru a determina hidroliza regiunilor cu structură cristalină este necesar ca în amestecul enzimatic să fie prezente ambele tipuri de enzime.
In ceea ce priveşte β-glucozidazele, acestea determină hidroliza celobiozei în două unităţi de glucoză. Deşi aceste enzime nu sunt propriu-zis celulaze, ele participă la îmbunătăţirea randamentului degradării celulozei prin eliminarea inhibării prin feedback a celulazelor realizată de obicei de celobioză (Robson şi Chambliss, 1989).
147

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
Utilizarea a numeroase substrate şi diverse tehnici de evidenţiere a activităţii celulazice a dus la o comparare dificilă a sistemelor celulozolitice de la diferite organisme. Mai mult, deoarece celulazele aflate în extracte brute prezintă o suprapunere parţială a specificităţii de substrat, un studiu corect trebuie realizat în condiţiile purificării acestor enzime. In ultima vreme au fost obţinute o serie de substrate celulozice care ar putea fi folosite ca substrate mai mult sau mai puţin specifice pentru fiecare dintre componentele sistemului celulazic (Wood şi Bhat, 1988). Cel mai utilizat procedeu de determinare a activităţii enzimatice este măsurarea cantităţii de zaharuri reducătoare eliberate în urma acţiunii enzimelor asupra substratului. Astfel, substratele celulozice cu structură amorfă sau cele modificate chimic cum sunt celuloza-azur, carboximetilceluloza(CMC), celuloza tratată cu acizi ("acid-swollen cellulose") sau trinitrofenil-carboximetilceluloza (TNP-CMC) sunt degradate în mod semnificativ şi specific de către endoglucanaze dar, în unele situaţii şi celobiohidrolazele le pot hidroliza de şi cu o eficienţă mai mică. In cazul utilizării drept substrat a celulozei cu structură cristalină de tipul Avicelului (celuloză microcristalină), bumbacului sau hârtiei de filtru, pentru degradarea lor eficientă este necesar un sistem celulazic complet, care să conţină atât endoglucanază cât şi celobiohidrolază. De obidei, pentru a determina prezenţa într-un preparat enzimatic brut a ambelor tipuri de enzime se determină eliberarea de zaharuri reducătoare în funcţie de substrat. Astfel, endoglucanazele cauzează o creştere rapidă a fluidităţii (reducerea vâscozităţii) carboximetilcelulozei dar o eliberare scăzută de zaharuri reducătoare. In contrast, celobiohidrolazele nu modifică fluiditatea substratului dar conduce la eliberarea unei cantităţi mai mari de zaharuri reducătoare. Cu toate acestea este dificil de precizat dacă o celulază este sau nu celobiohidrolază deoarece nu există un substrat specific pentru aceasta. Deshpande şi colab., (1988) au arătat însă că hidroliza unor compuşi cum ar fi metil-umbeliferil-celobiozid (MUC) sau p-nitrofenil--D-celobiozid (pNPC) ar fi un indiciu al activităţii exo--1,4-glucanazice.
Există în prezent numeroase date referitoare la bacterii celulozolitice dar mulţi autori consideră că bacteriile produc doar endoglucanaze şi nu un sistem celulazic complet. Deşi cea mai acceptată teorie referitoare la modul de degradare a celulozei este cea bazată pe rezultatele cercetărilor efectuate asupra fungilor şi care presupune că hidroliza eficientă a substratelor celulozice se realizează în urma acţiunii sinergice a endo- şi exoglucanazelor, ea nu este aplicată bacteriilor. La acestea, modelul de degradare sugerat presupune că endoglucanazele care au ca substrat specific celo-oligozaharidele cu lungimi variate pot determina hidroliza celulozei cristaline dacă ele se află în concentraţie mare sau dacă se leagă de fibrele de celuloză sau sunt grupate în imediata vecinătate a acestora ("celulosomi") (Robson şi Chambliss, 1989). Reuşita unor laboratoare din lume de a izola şi caracteriza celobiohidrolaze de la bacterii vine să contrazică în cazul speciilor respective teoria enunţată mai sus (Creuzet şi colab.,1983; Mac Kenzie şi colab.,1984; Gardner şi colab., 1987; Beguin, 1990; Ruttersmith şi Daniel, 1991).
Printre bacteriile considerate a avea sisteme celulazice complexe, tipice se numără genurile Clostridium, Cellulomonas, Bacillus, Thermomonospora, Ruminococcus, Bacteroides, Erwinia şi Acetobacter. De asemenea, tulpini celulozilitice au fost identi-ficate şi în cadrul altor genuri de bacterii: Streptomyces, Rhizobium, Cellvibrio, Pseudomonas, Microbispora etc. In studiul de faţă ne vom referi doar la sistemele celulazice de la două genuri de bacterii Gram pozitive, Bacillus şi Streptomyces.
La bacteriile din genul Bacillus s-a evidenţiat faptul că ele produc doar endoglucanaza şi nu sînt capabile de o degradare eficientă a celulozei cristaline. Studiile referitoare la celulazele de la Bacillus au fost sprijinite şi de faptul că aceste bacterii pot fi uşor manipulate genetic (genomurile de la mai multe specii fiind destul de bine caracterizate) şi de caracterul extracelular al acestor enzime. Printre speciile care produc celulaze se numără B.subtilis, B.polymyxa, B.licheniformis, B.cereus etc.
Studiile referitoare la celulazele de la Bacillus se referă atât la caracterizarea biochimică a enzimelor cât şi la identificarea şi secvenţierea genelor implicate în producerea acestor enzime ca
148

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
şi în clonarea şi exprimarea lor în diferite gazde. In prezent se cunoaşte secvenţa de nucleotide a mai multor endoglucanaze de la Bacillus (tabelul 5) şi deci secvenţa de aminoacizi a proteinelor corespunzătoare, realizându-se astfel comparaţii între enzimele secretate de diferite specii de bacili.
Tabelul 5. Genele pentru celulazele de la BacillusTulpina Gena Mărimea proteinei
produse (aminoacizi)B.circulans 378B.subtilis DLG 499 B.subtilis N-24 463B.subtilis PAP 115 499B.lautus celB 536Bacillus N-4 celA
celB celC
384465>800
B.polymyxa 365 Bacillus 1139 celF 770Bacillus sp.KSM 635 912
S-a stabilit astfel că enzimele produse de diferite tulpini de Bacillus prezintă un grad înalt de omologie, porţiunile cu omologie ridicată fiind localizate, în general, în regiunea aminoterminală, ceea ce sugerează că situsul catalitic activ al enzimelor se află plasat în această zonă. Această supoziţie este sprijinită de observaţiile că porţiunea de la capătul carboxiterminal al endoglucanazei respective nu este necesară pentru activitatea celulozolitică, regiunea respectivă putând fi deletată experimental fără pierderea actibvităţii (Nakamura şi colab. 1991). Recent, Henrissat şi colab.(1989) analizând grupările ("clusterii") hidrofobe ale diferitelor enzime au clasificat celulazele, inclusiv pe cele de la Bacillus, în şase familii. Din acest punct de vedere, endoglucanazele de la B.subtilis şi B.ciculans sunt incluse în familia A, subfamiliile A2 şi A4 (Beguin, 1990; Gilkes şi colab.,1991).
De asemenea, rezultatele de până acum referitoare la genele pentru enduglucanazele de la Bacillus sugerează ideea că ele ar deriva dintr-o genă ancestrală comună.
In ceea ce priveşte particularităţile biochimice ale endoglucanazelor de la bacteriile din genul Bacillus acestea au o greutate moleculară ce variază, de obicei, între 35 şi 46 kDa, dar poate atinge, la unele tulpini alcalofile, până la 92 kDa (Fukumori şi colab., 1986), sunt în general termostabile, activitatea optimă realizându-se la 50-60oC.
Reglarea producerii de endoglucanaze de către bacteriile din genul Bacillus variază nu numai de la specie la specie ci chiar şi de la tulpină la tulpină. Spre exemplu, tulpini de B.cereus, Bacillus sp.N-4, B.subtilis AU1, B.licheniformis sintetizează enzima în mod constitutiv în timp ce la alte tulpini, B.polymyxa, B.subtilis DLG, Bacillus sp.1139, aceasta pare a fi inductibilă. De asemenea, nivelul glucozei şi al celobiozei influenţează diferit producerea englucanazei de către tulpinile bacteriene studiate. Dacă în cazul tulpinii B.subtilis AU1 atât glucoza cât şi celobioza inhibă sinteza enzimatică la concentraţii mai mari de 2%, în cazul tulpinii alcalofile Bacillus sp.1139 glucoza inhibă, iar celobioza stimulează producerea de enzimă. La alte tulpini de Bacillus, B.subtilis DGL, B.cereus, B.licheniformis, ambele zaharuri reducătoare stimulează sinteza endoglucanazei.
In urma experimentelor de clonare a genelor pentru endoglucanază în diferite gazde (fie tulpini auxotrofe de E.coli fie alte tulpini de B.subtilis) au fost obţinute informaţii interesante referitoare la exprimarea genelor respective în gazde heterologe ca şi la reglarea exprimării
149

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
acestora. De obicei, celulazele ale căror gene au fost clonate în E.coli sunt sintetizate iniţial sub forma unor compuşi de dimensiuni mari, activi din punct de vedere enzimatic, cu localizare intracelulară. Aceşti compuşi suferă o "prelucrare" în urma căreia dimensiunea enzimei se micşorează, sub această formă ea fiind exportată în spaţiul periplasmic (Cantwell şi Mc Connel, 1983; Robson şi Chambliss, 1989). In cazul utilizării drept gazda pentru clonare a unor tulpini mutante de Bacillus s-a observat un fenomen similar de "procesare intracelulară" a precursorului enzimatic în urma căruia dimensiunia enzimei se apropie de cea a enzimei produsă de tulpina de origine (Robson şi Chambliss, 1989). De asemenea, Nakamura şi colab.(1991) au observat că utilizarea drept gazdă a unei tulpini mutante de B.subtillis care nu produce proteaze a condus la obţinerea unei cantităţi superioare de enzimă de la tulpina recombinată. Această creştere poate fi de cel puţin 9 ori dacă în noua gazdă este clonată suplimentar şi gena sacQ.
Deşi cele mai multe informaţii referitoare la degradarea substratelor celulozice de către actinomicete sunt legate de bacteriile din genul Thermomonospora (T.fusca, T.curvata)(Wilson, 1992), în ultimii ani au fost realizate şi studii referitoare la sistemul celulazic al bacteriilor din genul Streptomyces.
Tulpinile de streptomicete capabile de a degrada celuloza aparţin la mai multe specii cum ar fi: S.antibioticus (Enger şi Mc Coy, 1965), S.thermodiastaticus (Crawford şi Mc Coy, 1972), S.flavogriseus (Ishaque şi Kluepfel, 1980), S.olivochromogenes (Mac Kenzie şi colab.,1987), S.reticuli (Wachinger şi colab., 1989), S.lividans (Theberge şi colab., 1992), S.flavovirens SfG3 (Cornea şi colab., 1993), dar lista poate continua dacă ţinem cont de tulpinile de Streptomyces care nu au fost încadrate până acum într-o specie anume.
Primele lucrări referitoare la sistemul celulazic de la streptomicete au evidenţiat faptul că acesta este complex, enzimele componente fiind capabile de a degrada substrate foarte variate: CMC, hârtie de filtru, bumbac, celuloză microcristalină, celobioză dar şi paie, tărâţe sau alte deşeuri celulozice. Studiind complexul celulazic de la S.flavogriseus, Ishaque şi Kluepfel (1980) au stabilit că aceste bacterii produc enzime extracelulare capabile de a degrada, cu o rată diferită, carboximetilceluloza, hârtia de filtru şi fibrele de bumbac. De asemenea, tulpina respectivă produce şi -glucozidază dar această enzimă rămâne în cea mai mare parte localizată intracelular. Cercetările ulterioare realizate asupra unor tulpini aparţinând aceleiaşi specii (S.flavogriseus) au condus la precizarea că aceste bacterii produc atât endoglucanaze cât şi exoglucanaze (celobio-hibrolaze)(Mac Kenzie şi colab.,1984). Rezultate asemănătoare au fost comunicate şi de alţi autori în cazul unor specii diferite de Streptomyces: S.reticuli (Wachinger şi colab., 1989) sau Streptomyces sp.SD16 (Cornea şi colab.,1992) (Figura 6).
In ceea ce priveşte stabilirea condiţiilor optime de acţiune a acestor enzime, există o oarecare variaţie în funcţie de tulpina bacteriană studiată şi de autor. Astfel, în cazul tulpinilor de S.flavogriseus analizate, testarea acţiunii enzimelor pe CMC se realizează la 500C, în tampon fosfat 50mM pH 6 (Mac Kenzie şi colab.,1984) sau în tampon citrat-fosfat 50mM pH 7 (Ishaque şi Kluepfel, 1980), în timp ce activitatea enzimatică pe hîrtie de filtru sau pe celuloză microcristalină (Avicel) se realizează la 400C fie în tampon fosfat 50mM pH 6 (Mac Kenzie şi colab.,1984) fie în tampon citrat 50mM pH 5,6 (Ishaque şi Kluepfel, 1980). La o altă specie, S.reticuli, testarea activităţii celulazelor s-a realizat la 450C în tampon Tris HCl pH 7,5 (Wachinger şi colab., 1989). Alţi autori (Godden şi colab., 1989) testează activitatea endoglucanazică a filtratului de cultură de la o tulpină de Streptomyces în tampon fosfat de potasiu 100mM, pH 8 la 500C. Recent, Theberge şi colab.(1992) au caracterizat o endoglucanază de la S.lividans 66 care prezenta o activitate optimă la p 5,5 la 500C. O situaţie particulară a fost evidenţiată la o tulpină alcalofilă de Streptomyces tulpină capabilă de a sintetiza trei endoglucanaze diferite, cu pH optim de acţiune ce variază între 6 şi 8,5 (Nakai şi colab.,1988).
150

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
Figura 6. Selecţia unor noi tulpini de Streptomyces sp. producătoare de celulaze.
In cadrul unor exprimente realizate cu o tulpină de Streptomyces sp.SD16 izolată în Institutul de Biologie Bucureşti, Cornea şi colab. (1992) au înregistrat o activitate endoglucanazică maximă la pH 6,4 la 500C în tampon fosfat de potasiu 50mM. In cazul utilizării drept substrat a hârtiei de filtru au fost obţinute valori maxime de degradare (eliberare de zaharuri reducătoare) la 400C în tampon fosfat de potasiu 50mM la pH 5,8 şi pH 8 (Figura 7).
Figura 7. Activitatea enzimatică a unor tulpini de streptomicete izolate din sol (după Cornea şi colab., 1992).
Streptomicetele, ca multe alte bacterii, sitetizează celobiază (β-glucozidază) dar în mod normal nu o excretă în mediul de cultură ci ea rămâne localizată intracelular. Studii mai amănunţite asupra acestui tip de enzime au fost realizate la tulpini de S.granaticolor (Jiresova şi colab.,1987) la care s-a încercat izolarea şi caracterizarea enzimei, ca şi la S.thermodiastaticus (Ozaki şi Yamada, 1991). La această ultimă specie a fost obţinută o tulpină care produce o β-glucozidază a cărei sinteză nu este afectată de glucoză, enzima fiind purificată şi caracterizată. Alături de această enzimă rezistentă la acţiunea glucozei, de la aceeaşi tulpină bacteriană au mai fost izolate încă două enzime de acelaşi tip (β-glucozidaze) dar a căror sinteză este supusă inhibării de către glucoză.
151

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
Un aspect interesant al cercetărilor referitoare la celulazele produse de bacteriile din genul Streptomyces este izolarea şi purificarea componentelor acestui sistem. Spre deosebire de bacteriile din genul Thermomonospora la care au fost bine caracterizate cinci endoglucanaze diferite (E1-E5) dintre care E3 prezintă sinergism atât cu alte endoglucanaze cât şi cu o celobiohidrolază izolată de la Trichoderma reesei (Wilson, 1992), la streptomicete asemenea rezultate sunt mai puţine.
Nakai şi colab.(1987) au purificat şi caracterizat de la o tulpină alcalofilă de Streptomyces (Streptomyces sp.KSM9) trei celulaze diferite care degradează preferenţial CMC şi care au fost notate CMC-aza I, CMC-aza II şi CMC-aza III. Greutăţile moleculare ale acestor enzime variază între 32 kDa (CMC-aza I), 32,5 kDa (CMC-aza II) şi 92 kDa (CMC-aza III). Purificarea şi separarea celor trei enzime s-a realizat prin cromatografie pe schimbători de ioni, prin trecerea pe o coloană de DEAE-Toyopearl 650M a unui preparat celulazic brut, rezultat prin precipitarea cu sulfat de amoniu (70%) a proteinelor din filtratul de cultură şi eluarea lor cu un gradient linear de NaCl (0,05-0,4M). Alţi autori (Wachinger şi colab.,1989) au fracţionat celulazele de la S.reticuli prin gel filtrare pe Superose 12 şi au obţinut cel puţin două CMC-aze precum şi mai multe proteine cu activitate pe celuloza microcristalină. CMC-azele, notate CMC-aza 1 de 40 kDa şi CMC-aza 2 de 48 kDa prezintă un pH optim de acţiune ce variază între 5,5 şi 7,5 la o temperatură de 45-500C. De asemenea ele nu sunt inhibate de tamponul fosfat şi nu sunt stimulate de adăugarea de ioni de calciu în amestecul de reacţie. Spre deosebire de acestea, preparatul enzimatic ce prezintă activitate specifică pe Avicel, supus electroforezei în gel de poliacrilamidă în condiţii de denaturare a evidenţiat prezenţa a mai multor benzi cu greutăţi moleculare ce au variat între 42 şi 120 kDa (42, 56, 70, 120). Preparatul enzimatic obţinut prin gel filtrare a avut o temperatură optimă de acţiune la 45-500C la un pH optim 7. Activitatea celulazică pe substrat de Avicel este redusă semnificativ în tampon fosfat, dar este stimulată de adăugarea de CaCl2 40mM şi de asemenea ea este pierdută rapid, în decursul a câtorva zile prin păstrare la 40C, în timp ce CMC-azele sunt mult mai stabile. Proprietăţile avicelazei de la S.reticuli sunt foarte asemănătoare celor descrise pentru exoglucanaza de la Ruminococcus albus care a fost izolată sub forma unui complex de 230 kDa, alcătuitdin monomeri de 118 kDa (Gardner şi colab., 1987).
Recent, Theberge şi colab.(1992) au purificat şi caracterizat o endoglucanază de la S.lividans 66 după cultivarea bacteriilor pe mediu cu xiloză ca sursă de carbon. Aceasta este o glicoproteină cu greutatea moleculară de 46 kDa, cu pI la 3,3 şi cu o activitate maximă asupra CMC la 500C la pH 5,5. Interesant este că prin cultivarea bacteriilor în mediu cu Avicel sau celobioză ca sursă principală de carbon s-a obţinut un preparat enzimatic alcătuit din mai multe proteine de dimensiuni variate, dar active pe oligozaharide şi care au interacţionat pozitiv cu anticorpii anti-endoglucanază. Fenomenul respectiv a fost observat la celulazele de la Trichoderma reesei şi de la Clostridium thermocellum, el datorându-se în cazul fungilor fie unui grad diferit de glicozilare a proteinelor fie unor modificări proteolitice. In cazul bacteriilor fenomenul s-ar putea explica prin formarea unor complexe enzimatice specifice, celulosomi, doar în prezenţa substratelor microscristaline. Autorii presupun însă că apariţia fragmentelor endoglucanazice la S.lividans 66 s-ar datora mai degrabă unei serii de fenomene proteolitice decât unei glicozilări diferenţiate sau formării celulosomilor. In orice caz, activitatea specifică a endoglucanazei de S.lividans comunicată de autori a fost de 539 IU/mg proteină, ceea ce reprezintă una dintre cele mai mari valori raportate în literatură (Theberge şi colab.,1992).
Rezultate interesante au fost obţinute şi de Mac Kenzie şi colab.(1984) care au reuşit, printre puţinii cercetători de altfel, izolarea şi caracterizarea unei exoglucanaze de la o tulpină de S.flavogriseus. Aceasta s-a realizat prin cromatografie pe DEAE Bio-Gel A a unui preparat brut obşinut prin concentrarea de 100 ori a filtratului de cultură (prin ultrafiltrare). Eluarea enzimelor s-a făcut cu un gradient linear de NaCl (0-0,2M) obţinâdu-se mai multe "peak-uri" cu activitate celulazică. Cinci dintre acestea au prezentat activitate CMC-azică iar unul a evidenţiat o
152
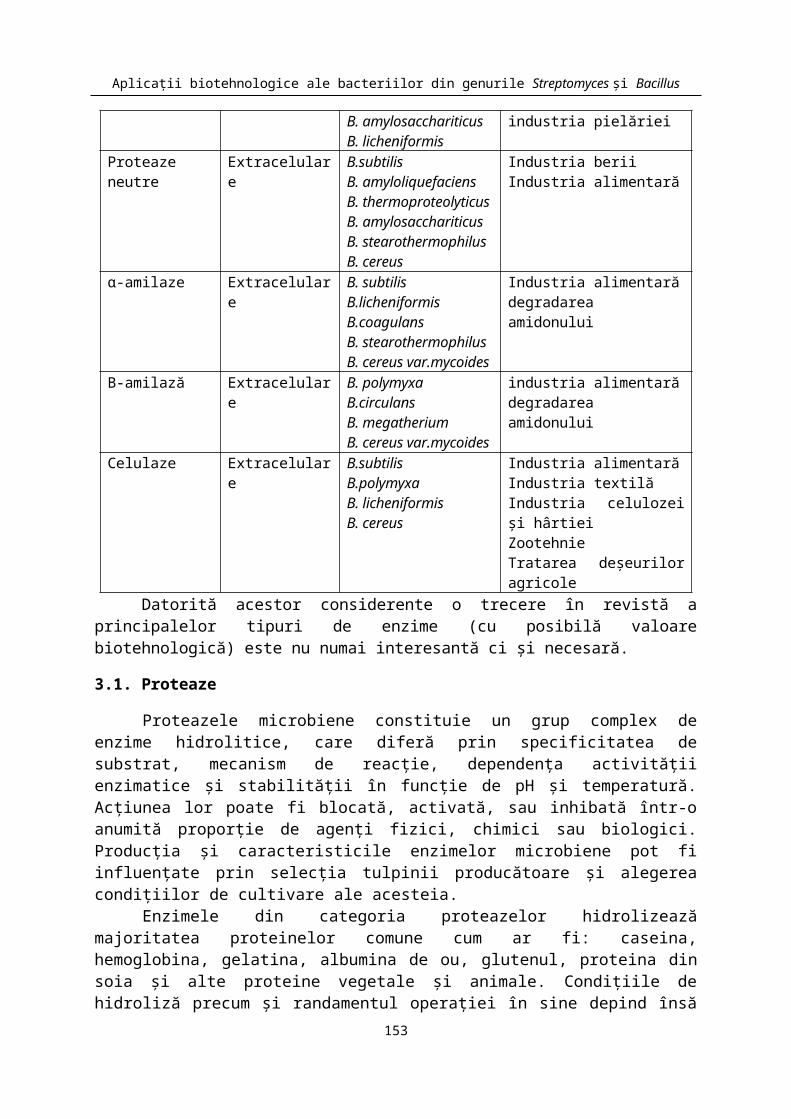
Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
activitate enzimatică complexă (avicelazică, CMC-azică, xilanazică şi proteazică). Fracţiile care au prezentat cea mai mare activitate avicelazică au fost supuse gel filtrării pe Bio-Gel P60 reuşindu-se separarea avicelazei de CMC-ază. Fracţiile active au fost apoi trecute pe o coloană de Concanavalină A-Sefaroză 4B, fiind recuperată o proteină de 46 kDa care nu a prezentat afinitate pentru coloana respectivă (pentru lectina imobilizată). Această proteină prezintă pI la 4,15, determină în special hidroliza celulozei tratate cu acizi ("acid-swollen cellulose"), produşii de hidroliză fiind celobioza şi celotrioza (în cea mai mare parte).
Datele obţinute la streptomicete, în special de la tulpinile de S.flavogriseus evidenţiază faptul că mecanismul de acţiune al celulazelor de la aceste bacterii este mai degrabă asemănător celui descris la fungi la care degradarea substratelor celulozice se datorează acţiunii sinergice a endoglucanazelor, exoglucanazelor şi β-glucozidazei. De asemenea, glicozilarea unora dintre componentele sistemului celulazic de la streptomicete (a endoglucanazelor, de exemplu), fenomen care este destul de puţin răspândit la bacterii dar frecvent la fungi poate sugera un comportament asemănător al celulazelor de la streptomicete cu cel al unor fungi. In ceea ce priveşte semnificaţia glicozilării, deşi unii autori atribuie glucidului un rol în funcţionarea enzimei, ea rămâne încă neclară (Knowles şi colab.,1987).
Cu toate acestea explicarea mecanismului de acţiune a celulazelor de la streptomicete este complicat de faptul că la unele tulpini bacteriene, la S.reticuli de pildă, au fost identificate celulaze legate de miceliu ("mycelium buond cellulase") care amintesc de formarea celulosomilor de la Clostridium thermocellum (Lamed şi Bayer, 1988; Wachinger şi colab.,1989). O organizare a celulazelor în celulosomi similară celei evidenţiate la clostridii a fost descrisă şi la Thermomonospora curvata deşi studiile au fost în special de microscopie electronică (Figura 8).
Figura 8. Aspect electronomicroscopic al suprafeţei celulelor de T.curvata cultivate pe fibre de celuloză (după Bond şi Stutzberger, 1989).
Modificarea aspectului suprafeţei celulelor de streptomicete cultivate în mediu de cultură ce conţine celuloză drept unică sursă de carbon a fost observată la tulpinile Streptomyces sp.SD16 şi Streptomyces sp.P216 (Figura 9) (Cornea, 1996). Aceste rezultate sugerează că fenomenul complexării sistemului celulazic este comun mai multor tipuri de bacterii iar mecanismul de degradare a celulozei este mai complicat decât s-a presupus iniţial (Ljungdahl 1989).
153

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
Figura 9. Aspect electronomicroscopic al celulelor Streptomyces sp.SD16 cultivate în mediu minimal ce conţine celuloză.
In ceea ce priveşte determinismul genetic al producerii enzimelor celulozolitice, datele referitoare la streptomicete sunt destul de puţine şi incomplete. In general, clonarea şi secvenţierea unei gene structurale pentru o enzimă este utilă nu numai pentru determinarea secvenţei de aminoacizi pe care proteina îi conţine ci şi pentru a obţine proteina în cantitate mai mare ca şi pentru studii de mutageneză care să determine rolul anumitor secvenţe de aminoacizi în funcţionarea enzimei. Clonarea prezintă interes mai ales pentru celulaze deoarece prezenţa mai multor gene care codifică aceste enzime la majoritatea microorganismelor celulozolitice face dificilă atât obţinerea de mutante cât şi studiul rolului fiecărui component în parte.
Până în prezent s-a reuşit izolarea, secvenţierea şi clonarea în diferite tulpini mutante de S.lividans (care nu produc celulaze) sau în E.coli a trei gene implicate în sinteza celulazelor la streptomicete: gena casA ce codifică CMC-aza I de la o tulpină alcalofilă de Streptomyces sp. (Nakai şi colab.,1988), gena celA ce codifică o endoglucanază de la S.lividans 66 (Theberge şi colab.,1992) şi gena celA1 ce codifică o endoglucanază de la S.halstedii (Fernandezabalos şi colab.,1992). De asemenea, Shareck şi colab.(1987) au realizat clonarea într-o tulpină mutantă de S.lividans (care nu producea celulaze) a unei secvenţe de ADN care a determinat nu numai restabilirea capacităţii de a degrada celuloza ci şi o sporire de peste 800 ori a activităţii enzimatice comparativ cu tulpina sălbatică. Interpretarea rezultatelor în acest caz rămâne dificilă deoarece genele clonate nu au fost analizate astfel că nu se poate preciza dacă secvenţele de ADN clonate conţin gene ce codifică celulaze (endoglucanaze) sau proteine reglatoare.
Secvenţierea genelor casA şi celA a relevat prezenţa unei secvenţe de 14 pb repetate inversat, plasată înaintea regiunii ce codifică proteinele respective, secvenţă identificată şi la nivelul genelor celE2, celE4 şi celE5 de la Thermomonospora fusca (Wilson, 1992; Theberge şi colab., 1992). De asemenea, din examinarea secvenţelor de aminoacizi deduse din examinarea secvenţelor de nucleotide ale genelor casA şi celA şi compararea lor cu secvenţele de aminoacizi ale altor celulaze au fost evidenţiate elementele structurale ale celulazelor respective, aşa cum au fost ele propuse de Gilkes şi colab.(1991): domeniile catalitice, domeniile de legare la celuloză, linkerii ce unesc asemenea domenii şi secvenţele repetate de aminoacizi. Spre exemplu, endoglucanaza produsă de S.lividans 66 (codificată de gena celA) este alcătuită din două domenii (domeniul catalitic şi domeniul de legare la celuloză) conectate de un linker scurt, extremitatea amino-terminală evidenţiind un grad înalt de omologie cu domeniul de legare la celuloză ("cellulose-binding domain") al endoglucanazei CenA de la Cellulomonas fimi, iar regiunea din
154

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
mijlocul proteinei prezintă omologie extinsă cu celulaza E5 de la T.fusca şi alte endoglucanaze de Bacillus (Theberge şi colab., 1992).
Conform propunerilor lui Henrissat şi colab.(1989) şi Gilkes şi colab.(1991) de grupare a celulazelor în familii pe baza secvenţelor de aminoacizi de la nivelul domeniilor catalitice conservate la mai multe microorganisme şi a analizei regiunilor ("clusteri") hidrofobe, endoglucanazele ale căror gene au fost clonate şi secvenţializate (casA şi celA) pot fi încadrate în familia A (celA), alături de endoglucanaze de la Clostridium thermocellum, B.subtilis, Bacillus sp., Erwinia chrosanthemi, Ruminococcus albus, Clostridium cellulolyticum etc, şi în familia B (casA) alături de endoglucanaze de la Cellulomonas fimi, Microbispora bispora şi o celobiohidrolază de la Trichoderma reesei.
Reglarea activităţii celulazelor reprezintă un alt aspect al cercetărilor cu aceste enzime, în scopul stabilirii condiţiilor optime de producere şi acţiune a acestora. La fel ca şi în cazul mecanismului de degradare a substratelor celulozice, mecanismul de reglare a sintezei şi activităţii celulazelor este destul de neclar. O ipoteză generală susţine că microorganismele celulozolitice produc celulazele la un nivel bazal care este însă suficient pentru a asigura hidroliza celulozei insolubile, rezultând produşi de degradare care pot funcţiona ca inductori determinând sinteza unor cantităţi crescute de celulaze (Robson şi Chambliss, 1989). Ca şi alte enzime hidrolitice extracelulare şi celulazele se sintetizează ca răspuns la prezenţa inductorului, ele fiind represate de obicei de produşii finali de degradare. In cazul actinomicetelor există mai multe date referitoare la reglarea celulazelor la bacteriile din genul Thermomonospora. Astfel, sinteza celulazelor de către T.fusca este reglată atât prin inducere cât şi prin represie (Liu şi Wilson, 1987). Wilson (1992) a arătat că tulpina T.fusca YX cultivată pe celuloză a evidenţiat o activitate celulazică de 100 ori mai mare decât cea obţinută prin cultivarea în mediu cu glucoză ca sursă unică de carbon. Alţi autori (Fennington şi colab.,1984) au obţinut o tulpină mutantă de T.curvata capabilă de a rezista la acţiunea inhibitoare a glucozei. Liu şi Wilson (1988) au reuşit izolarea unei tulpini mutante de T.fusca YX care sintetiza în mod constitutiv celulaze, dar sinteza lor era supusă în continuare represiei produse de glucoză. Spre deosebire de bacteriile din genul Thermomonospora, la streptomicete există foarte puţine date referitoare la reglarea sintezei celulazelor iar cele care există sunt oarecum contradictorii. Astfel, Mac Kenzie şi colab.(1984) au arătat că activitatea celulazelor de la S.flavogriseus este inhibată de concentraţii mai mari de glucoză şi celobioză, în timp ce Ball şi Mc Carthy (1988) susţin că în cazul unor tulpini de Streptomyces izolate de ei glucoza şi xiloza nu exercită efect inhibitor asupra enzimelor implicate în degradarea deşeurilor celulozice de tipul paielor. In orice caz, cultivarea bacteriilor pe mediu ce conţine celuloză pulbere sau CMC are drept consecinţă inducerea sintezei doar a enzimelor celulazice în timp ce prin cultivarea bacteriilor pe medii mai complexe (cu paie, xilan sau tărâţe de grâu) are loc inducerea unei game mai largi de enzime hidrolitice: atât celulaze cât mai ales hemicelulaze (Mac Kenzie şi col., 1987, Godden şi colab., 1989). Spre exemplu, Godden şi colab. (1989) utilizând pentru testări o tulpină de Streptomyces sp. a evidenţiat faptul că endoglucanazele şi β-glucozidazele sunt induse specific prin cultivarea pe mediu cu CMC ca sursă de carbon. Celobioza a evidenţiat o slabă acţiune de inducere a sintezei endoglucanazelor, dar induce -glucozidaza în timp ce celotrioza şi celotetraoza induc slab endoglucanazele. Obţinerea unei activităţi celulazice specifice ridicate în filtratele de cultură rezultate prin cultivarea bacteriilor pe substratele celulozice naturale (de exemplu paie) se datorează probabil unei cooperări dintre diferiţii inductori rezultaţi din acţiunea componentelor complexului lignocelulozic. Mai mult, inducerea sintezei celulazelor este inhibată de adăugarea a 50 g/ml cloramfenicol ceea ce ar sugera că , cel puţin în cazul tulpinii analizate, inducerea enzimelor este mai degrabă o sinteză "de novo" (Godden şi colab. 1989).
3.5. Hemicelulazele155

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
Alături de celuloză şi lignină, hemicelulozele reprezintă componente de bază ale peretelui celular vegetal, degradarea lor în natură realizându-se destul de greoi, prin cooperarea unor sisteme enzimatice de origini foarte diferite. Intre microorganismele producătoare de hemicelulaze un loc important este ocupat de streptomicete datorită sintezei unei game foarte variate de asemenea enzime.
Hemicelulozele sunt polizaharide care conţin diferite tipuri de resturi de zaharuri (xiloză, manoză, galactoză, arabinoză sau glucoză) ce alcătuiesc polimeri lineari sau ramificaţi. Clasificarea acestor substanţe se face în funcţie de conţinutul în zaharuri. Dintre acestea, β-1,4-D-xilanii reprezintă grupul major de hemiceluloze fiind alcătuiţi din resturi de D-xilopiranozil unite prin legături de tip β-1,4. In mulţi xilani resturile de D-xilopiranozil sunt întocmite de arabinozil, glucuronozil sau diferite grupări acetil, în funcţie de planta de origine. Un alt grup de hemiceluloze, mananii sînt constituiţi din resturi de D-manopiranozil unite tot prin legături de tip -1,4. Si în cazul acestor polimeri unităţile specifice de D-manopiranozil pot fi înlocuite de D-galactozil sau grupări acetat (Zimmermann, 1989). Hemicelulozele reprezintă un important component al peretelui celular vegetal, ele putând fi linkate la compuşi fenolici sau pot prezenta legături cu lignina.
Sistemul enzimatic pentru degradarea hemicelulozelor constă în enzime care clivează legăturile din interiorul macromoleculei (endoenzime) şi în exoenzime care taie resturile de zaharuri de la extremităţile nereducătoare ale polimerului. Prezenţa unor substituenţi variaţi influenţează puternic hidroliza enzimatică. De aceea, pentru hidroliza completă a hemicelulozelor sunt necesare enzime care să taie în diferite puncte de ramificaţie. In sistemul xilanolitic, aceste enzime acţionează în cooperare cu xilanazele pentru hidroliza completă a xilanilor (Biely, 1985).
Tinând cont de habitatul natural al streptomicetelor şi de prezenţa în acesta a unei mari cantităţi de materiale celulozice şi lignocelulozice, bacteriile respective pot fi considerate ca o sursă potenţială de enzime hemicelulozolitice (Zimmermann şi colab., 1992).
Sistemul enzimatic de degradare a xilanului de la diferite tulpini de Streptomyces a fost studiat în detaliu. La aceste bacterii au fost identificate mai multe tipuri de hemicelulaze: xilanaze, β-xilozidaze, α-L-arabinofuranozidaze, α-4-O-metil-glucuronidaze, acetil-xilan esteraze, ferulic şi cumaric esteraze, enzime ce degradează mananii (mananaze). Dintre acestea, 1,4-β-D-xilanazele (EC 3.2.1.8.) au fost mai mult studiate, ele fiind detectate la numeroase tulpini de Streptomyces (Ball şi Mc Carthy, 1988). De asemenea, s-a evidenţiat faptul că aceeaşi tulpină poate produce mai multe xilanaze distincte. Spre exemplu, S.ostreogriseus şi S.exfoliatus produc câte 5 asemenea enzime în timp ce S.lividans produce o singură asemenea xilanază (Sreenath şi Joseph, 1982; Morosoli şi colab.,1986). Recent, Lumba şi Penninchx (1992) au caracterizat mai multe forme de β-xilanază izolate de la o tulpină de Streptomyces cultivată pe lignoceluloză. Interesant însă că în cazul acestei tulpini, formele de β-xilanază evidenţiate nu acţionează sinergic pentru hidrolizarea substratului ceea ce ar constitui o redundanţă a cărei semnificaţie nu este clară.
Pentru purificarea şi caracterizarea xilanazelor de la diferite streptomicete (Tabelul 6) s-au utilizat filtrate de cultură din care enzimele au fost precipitate cu sulfat de amoniu şi apoi supuse unor tratamente speciale: ultrafiltrare, cromatrografie pe schimbători de ioni, gel filtrare şi focusare izoelectrică ("isoelectric focusing")(Sreenath şi Joseph, 1982; Morosoli şi colab., 1986; Mac Kenzie şi colab., 1987; Yasui şi colab., 1988).
Cele mai multe dintre aceste enzime au pH între 5 şi 6,5 iar punctul lor izoelectric variază între 5,2 şi 10,3 (Zimmermann, 1989). Xilanazele determină hidroliza xilanilor prin ruperea legăturilor interne β-1,4, determinând eliberarea de xilotrioză, xilobioză sau xiloză (Yasui şi colab., 1988). Un alt tip de hemicelulaze produse de streptomicete, exo-β-1,4-xilozidazele (1,4-
156

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
β-D xylan xylohydrolase, EC 3.2.1.37) hidrolizează xilanii prin atac la extremităţile nereducă toare ale lanţului de polimer.
Tabelul 6. Proprietăţile unor xilanaze izolate din diferite tulpini de streptomicete.
Tulpina pH optim t0 optimă pI greutatemoleculară (kDa)
S.xylophagus 6,5 55-60 - -S.flavogriseus
CD45-26,5 50 - -
S.exfoliatus 5,5 50 - -Streptomyces sp.
KT235,5 55 6,9 43
S.lividans 5,5-6,5 55-65 5,2 -Streptomyces sp.E86 5,5-6,2 55-60 7,3 40
Eliberarea de resturi α-L-arabinoză din arabinoxilani, arabinani şi arabinogalactani se datorează acţiunii specifice a α-L-arabinofuranozidazelor, enzime evidenţiate la unele specii de Streptomyces (S.rubiginosus, S.purpurascens, S.massasporus) (Zimmermann, 1989). Alte enzime, α-4-O-metilglucironidaze, evidenţiate la S.flavogriseus şi S.olivochromogenes (Mac Kenzie şi colab., 1987) determină clivarea legăturilor α-1,2 dintre acidul glucuronic şi resturile de xiloză, acţionând sinergic cu xilanazele în procesul de hidroliză a xilanilor. Pe lângă aceste tipuri de hemicelulaze, unele tulpini de Streptomyces mai pot sintetiza şi alte enzime care hidrolizează alte specii de xilani. Acetil-xilan esterazele produse de S.olivochromogenes şi S.rubiginosus participă alături de xilanaze la hidroliza acetil-xilanilor în timp ce alte tulpini de Streptomyces (S.viridosporus) pot produce ferulic esteraze care eliberează acid ferulic din arabinixilanii aflaţi în cantităţi mai mari în peretele celular al gramineelor (Mac Kenzie şi colab., 1987; Deobald şi Crawford, 1987).
O categorie mai puţin studiată de hemicelulaze produse de streptomicete (ţinând cont de literatura de specialitate) este reprezentată de mananaze (1,4-β-D-manan mananohidrolaza, EC 3.2.1.78.) evidenţiate la unele tulpini de S.olivochromogenes sau la alte specii.
Utilizarea tehnicilor de biologie moleculară şi implicit a celor de inginerie genetică în studiul hemicelulazelor de la streptomicete a permis izolarea şi caracterizarea genelor implicate în producerea unor asemenea enzime. Astfel, Mondou şi colab.,(1986) au raportat clonarea unei gene pentru endoxilanază de la S.lividans într-o tulpină mutantă neproducătoare. Clonele ce conţineau un fragment de ADN de 2kb au evidenţiat o creştere de peste 60 ori a producerii de xilanază comparativ cu tulpina sălbatică . De asemenea, Iwasaki şi colab.(1986) au realizat transferul genei pentru xilanază izolată de la o tulpină de Streptomyces în alte două tulpini de streptomicete - S. lividans şi S. kasugaensis - la care s-a observat o creştere a nivelului de sinteză a enzimei respective prin achiziţionarea unui fragment de ADN de 1,04kb. Alături de clonările de gene, fuziunile de protoplaşti constituie o metodă relativ simplă dar eficientă de a obţine tulpini recombinate genetic, capabile de o mai bună degradare a substratelor complexe. Astfel, Pettey şi Crwford (1984) au obţinut o îmbunătăţire a capacităţii de degradare a hemicelulozelor şi ligninei de către o tulpină recombinată obţinută prin fuziunea interspecifică de protoplaşti de la S.viridospurus T7A şi S.setonii 75 Vi2. Rezultate asemănătoare au for prezentate de Cornea şi colab.(1992) care, în urma fuziunii protoplaştilor de S.flavovirens SfG3 cu protoplaşti izolaţi de la Streptomyces sp.SD16 au obţinut mai multe tulpini recombinate genetic care prezentau o capacitate crescută de degradare a substratelor celulozice şi hemicelulozice comparativ cu tulpinile parentale (Figura 10).
157

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
Uneori, enzimele produse de diferite tulpini de streptomicete (preparate brute sau enzime purificate) au fost studiate în privinţa acţiunii lor asupra peretelui celulei vegetale sau al unor fungi. Astfel, Cornea şi colab.(1992) au evidenţiat faptul că prin utilizarea unui filtrat de cultură de Streptomyces sp.SD16 s-au putut obţine în condiţii experimentale specifice protoplaşti vegetali de Nicotiana tabacum. Acest rezultat sugerează faptul că filtratul de cultură conţine o mare varietate de enzime asemănătoare celulazelor, hemicelulazelor dar şi alte enzime care cooperează în degradarea peretelui celular vegetal.
Figura 10. Comparaţie între activitatea enzimatică (celulozolitică) a unor tulpini de streptomicete de tip sălbatic (P216, SD16 şi SFG3) şi a unor recombinanţi obţinuţi prin fuziune triparentală de protoplaşti.
In ceea ce priveşte acţiunea pe peretele celular fungic, s-a dovedit că printre enzimele sintetizate de unele tulpini de Streptomyces se numără şi chitinazele, enzimele ce degradează componenta chitinică (dominantă) a peretelui celular fungic ducând la formarea protoplaştilor (Tagawa şi Okazaki, 1991). Okazaki şi Tagawa (1991) au izolat şi caracterizat o chitinază produsă de S.cinereoruber după cultivarea pe un mediu ce conţinea ca sursă de carbon pereţi celulari de Aspergillus niger. Enzima respectivă are o greutate moleculară de 19 kDa şi un pI 8,6 iar pH optim se situează în domeniul acid (4,0-5,0) la 500C.
Beyer şi Diekeman (1984) au caracterizat enzime de tip laminarinază de la Streptomyces sp.ATCC11238 ca şi un complex de enzime ce degradează eficient chitina şi laminarina, fiind active asupra a peste 50 specii de ascomicete. Fracţionarea chitinazei prin gel filtrare şi cromatografie pe schimbători de ioni a condus la evidenţierea unei exo-chitinaze şi a unei β-N-acetil-glucozoaminidaze ce eliberează N-acetil glucozamina din crusta chitinoasă de crab (Beyer şi Diekman, 1985). In mod asemănător, Billich şi colab.(1988) au obţinut protoplaşti de Fusarium scirpi prin utilizarea unui preparat enzimatic de S.tsushimaensis.
Datele prezentate în această lucrare reflectă particularităţile deosebite ale bacteriilor din genurile Streptomyces şi Bacillus şi importanţa pe care acestea o au atât din punct de vedere teoretic cât şi practic, fie pentru obţinerea unor compuşi cu aplicaţii in medicină, industria alimentară, a detergenţilor etc. fie în natură, ca verigi importante ale mecanismelor de degradare şi reciclare a unor resturi organice existente de obicei în sol. Cunoaşterea echipamentului lor enzimatic, al potenţialului lor de biosinteză ca şi al determinismului genetic al acestuia corelată cu tehnicile moderne de biologie moleculară şi cu cele de fuziuni celulare asigură, pe de o parte, o mai bună studiere a caracteristicilor acestor bacterii, inclusiv a taxonomiei grupului iar, pe de altă parte, poate conduce la obţinerea unor tulpini recombinate genetic capabile de a sintetiza sau degrada cu o eficienţă crescută anumiţi compuşi cu importanţă practică deosebită.
158

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
Bibliografie selectivă
1. Aretz, W., Koller, K. P., Riess, G., 1989, Proteolytic enzymes from recombinant Streptomyces lividans TK24, FEMS Microbiol. Letts., 65, p.31-36.
2. Ball, A. S., Mc Carthy, A. J., 1988, Saccharification of straw by actinomycetes enzymes, J. Gen. Microbiol., 134, p.2139-2147.
3. Baltz, R. H., Seno, E. T., 1988, Genetics of Streptomyces fradiae and tylosin biosynthesis, Ann. Rev. Microbiol., 42, p. 547-574.
4. Bayer, E. A., Setter, E., Lamed, R., 1985, Organization and distribution of the cellulosome in Clostridium thermocellum, J. Bact., 163, p.552-559.
5. Beguin, P., 1990, Molecular biology of cellulose degradation, Ann. Rev. Microbiol., 44, p. 219-248.
6. Beyer, M., Diekman, H., 1985, The chitinase system of Streptomyces sp. ATCC 11238, Appl. Microbiol. Biotechnol., 23, p. 140-146
7. Billich, A., Keller, U., Kleinkauf, H., Zocher, K., 1988, Production of protoplasts from Fusarium scirpi by lytic enzymes from Streptomyces tsusimaensis, Appl. Microbiol. Biotechnol., 28, p.442-444
8. Binnie, C., Warren, M., Butler, M. J., 1989, Cloning and heterologous expression in S.lividans of S.rimosus genes involved in oxytetracycline biosynthesis, J. Bact., 171, p. 887-895
9. Bond, K., Stutzenberger, F., 1989, A note on the localization of cellulosome formation in Thermomonospora curvata, J. Appl. Bacteriol., 67, p. 605-609.
10. Bonner, M. A., Stutzenberger, F., 1988, Cell surface changes in Thermomonospora curvata during cellulase induction and repression, Letts. Appl. Microbiol., 6, p.59-63.
11. Buttner, M. J., Smith, A. M., Bibb, M. J., 1988, At least three different RNA polymerase holoenzymes direct transcription of the agarase gene (dagA) of S.coelicolor A3(2), Cell, 52, p. 599-607.
12. Chardon-Loriaux, I., Charpentier, M., Percheron, F., 1986, Isolation and characterization of a linear plasmid from Streptomyces rimosus, FEMS Microbiol.Letts., 35, p.151-155.
13. Chater,K., Bruton,C.J., 1985, Resistance, regulatory and production genes for the antibiotic methylenomycin are clustered, EMBO.J., 4, p.1893-1900.
14. Cornea, 1996, Obţinerea unor bacterii cu importanţă biotehnologică prin fuziunea şi transformarea genetică a protoplaştilor, Teză de doctorat, Universitatea Bucureşti.
15. Cornea, C. P., Cogălniceanu, G., Brezeanu, A., Vatafu, I., Savu, L., Popescu, I., Pop, A., 1992, Obţinerea protoplaştilor de tutun prin utilizarea unui preparat enzimatic brut de Streptomyces sp. SD16, St. Cercet. Biol., 44, p. 85-88.
16. Cornea, C. P., Vatafu, I., Pop, A., Brezeanu, A., Savu, L., Dobrotă, S., 1993, Interspecific protoplast fusion in Streptomyces, Rev. Roum. Biol., 38, p. 49-53.
17. Cornea, C. P., Vatafu, I., Savu, L., 1994, Plasmid transfer by protoplast fusion, Microbiol. Ind. Biotehnol., 8, p. 145-148.
18. Coughlan, B. P., 1988, Staining techniques for the detection of the individual components of cellulolytic enzyme system, Methods in Enzymology, 160, p. 135-145.
19. Cundliff, E., 1989, How antibiotic-producing organims avoid suicide, Ann. Rev. Microbiol., 43, p. 207-233.
20. Damude, H. G., Gilkes, N. R., Kilburn, D. G., Miller, R. C., Warren, R. A., 1993, Endoglucanase Cas A from alkalophilic Streptomyces strain KSM-9 is a tipical member of family B of â-1,4-endoglucanase, Gene, 123, p. 105-107.
21. Davies, J., 1990, What are antibiotics? Archaic functions for modern activities, Molec. Microbiol. , 4 (8), p. 1227-1232.
159

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
22. Deobald, L. A., Crawford, D. L., 1987, Activities of cellulase and other extracellular enzymes during lignin solubilization by Streptomyces viridosporus, Appl. Microbiol. Biotechnol., 28, p. 158-163.
23. Dimcev, A., 1988, Protoplast from antibiotic-producing actinomycetes. Application in the biotechnology, Biotechnol. Bioeng., 4, p. 12-19.
24. Doyle, R. J., 1994, Introduction to lectins and their interactions with microorganisms, in Lectin-Microorganism Interaction, Eds. R.J.Doyle si M.Slifkin, Marcel Dekker Inc., p. 1-65.
25. Duitman, E., Hamoen, L., Rembold, M., Vater, L., 1999, The mycosubtilin synthetase of Bacillus subtilis ATCC6633: A multifunctional hybrid between a peptide synthetase, an amino transferase, and a fatty acid synthase, Proc. Natl. Acad. Sci. USA: 96 (23), 13294-13299.
26. Felix, C. R., Ljungdahl, L. G., 1993, The cellulosome: the exocellular organelle of Clostridium, Ann. Rev. Microbiol., 47, p. 791-819.
27. Fernandez-Abalos, J. M., Sanchez, P., Coll, P. M., Villanueva, J. R., Perez, P., Santamaria, R. I., 1992, Cloning and nucleotide sequence of celA1, an endo-β-1,4-glucanase-encoding gene from Streptomyces halstedii JM8, J. Bacteriol., 174, p. 6338-6376.
28. Ghangas, G. S., Wilson, D. B., 1988, Cloning of the Thermomonospora fusca endoglucanase E2 gene in Streptomyceslividans: affinity purification and functional domains of the cloned gene product, Appl. Envir. Microbiol., 54, p. 2521-2526.
29. Gilbert, H. J., Hazlewood, G. P., 1993, Bacterial cellulases and xylanases, J. Gen. Microbiol., 139, p. 187-194.
30. Godden, B., Legon, T., Helvenstein, P., Penninckx, M., 1989, Regulation and production of hemicellulolytic and cellulolytic enzymes by a Streptomyces sp. growing on lignocellulose, J. Gen. Microbiol., 135, p. 285-292.
31. Gokhale, D. V., Deobagkar, D. N., 1989, Differential expression of xylanase and endoglucanase in the hybrid derived from intergeneric protoplast fusion between Cellulomonas sp. and Bacillus subtilis, Appl. Envir. Microbiol., 55, p. 2675-2680.
32. Gormley, E. P., Cantwell, B. A., Barker, P. J., Mc Connell, D. J., 1988, Secretion and processing of the Bacillus subtilis endo-â-1,3-14-glucanase in Escherichia coli, Molec. Microbiol, 2, p. 813-819.
33. Horikoshi, K., Nakao, M., Kurono, Y., Sashihara, N., 1984, Cellulases of an alkalophilic Bacillus strain isolated from soil, Can. J. Microbiol., 30, p. 774-779.
34. Imanaka, T., 1986, Application of recombinant DNA technology to the production of useful biomaterials, Adv. Biochem. Eng., 33, p. 1-27.
35. Ishaque, M., Kluepfel, D., 1980, Cellulase complex of a mesophilic Streptomyces strain, Can. J. Microbiol., 26, p. 183 – 189.
36. Kakoniova, D., Labudova, I, Liskova, D., 1987, Callus culture as substrate for the production of specific cell wall degradind enzymes for derived protoplast isolation, Biotechnol. Letts., 9, p. 721-724.
37. Kinashi, H., 1988, Giant linear plasmids in Streptomyces and antibiotic production, in Biology of Actinomycetes '88, Japan Scientific Societies Press, p. 117-122.
38. Kinashi, H., 1989, Giant linear plasmids in various methylenomycin-producing strains of Streptomyces species, in Genetics and Molecular Biology of Industrial Microorganisms, Ed. C. L. Hershberger, S. W. Queener, G. Hegeman, ASM, p. 141-146.
39. Kirby, R., Hopwood, D. A., 1977, Genetic determination of methylenomycin biosynthesis by the SCP1 plasmid of Streptomyces coelicolor A3(2), J. Gen. Microbiol., 98, p. 239-252.
40. Kluepfel, D., Shareck, F., Mondou, F., Morosoli, R., 1986, Characterization of cellulase and xylanase activities of Streptomyces lividans, Appl. Microbiol. Biotechnol., 24, p. 230-234.
41. Lamed, R., Naimark, J., Morgenstern, E., Bayer, E. A., 1987, Specialized cell surface structures in cellulolytic bacteria, J. Bacteriol., 169, p. 3792-3800.
160

Aplicaţii biotehnologice ale bacteriilor din genurile Streptomyces şi Bacillus
42. Mansouri, K., Pissowotzki, K., Distler, J., Ebert, A., Piepersberg, W., 1989, Genetics of streptomycin production, în Genetics and molecular biology of industrial microorganisms, Ed. Hershberger, C. L., Queener, S. W., Hegeman, G., ASM, p. 61-67.
43. Martin, J. F., 1989, Molecular mechanisms for the control by phosphate of the biosynthesis of antibiotics and other secondary metabolites, în Regulation of secondary metabolism in Actinomycetes, Ed. S. Shapiro, Boca Raton, CRC, p. 230-251.
44. Martin, J. F., Liras, P., 1989, Organization and expression of genes involved in the biosynthesis of antibiotics and other secondary metabolites, Ann. Rev. Microbiol., 43, p. 173-206.
45. Mc Carthy, A. J., Williams, S. T., 1992, Actinomycetes as agents of biodegradation in the environment - a review, Gene, 115, p. 189-192.
46. Nakano, M. N., Butsuya, I., Ogawara, H., 1989, Expression of the kanamycin resistance gene in a kanamycin-producing strain of S. kanamyceticus, J. Antib., 42, p. 423-430.
47. Nakano, M. N., Mashiko, H., Ogawara, H., 1984, Cloning of the kanamycin resistance gene from a kanamycin producing Streptomyces species, J. Bact., 157, p. 79-83.
48. Okanishi, M., Katagiri, K., Furumai, T., Takeda, K., Kawagichi, K., Saitoh, M., Nabeshima, S., 1983, Basic techniques for DNA cloning and conditions required for streptomycetes as host, J. Antib., 36, p. 99-108.
49. Okuda, T., Ito, Y., 1982, Biosynthesis and mutasynthesis of aminoglycoside antibiotics, în Aminoglycoside antibiotics, Ed. Umezawa, H., Hooper, I. R., Springer Verlag., p. 111-203.
50. Omich, P. K., 1988, Streptomyces cloning: useful recombinant DNA systems and a summation of cloned genes, Antim. Ag. Chemoth., 32, p. 1465-1471.
51. Peberdy, J. F., Illing, G. T., 1985, Genetic manipulation of antibiotic producer strains, in The Scientific Basis of Antimicrobial Chemotherapy, Ed. D. Greenwood, F. O' Grady, p.283-322.
52. Pettey, T. M., Crawford, D. L., 1984, Enhancement of lignin degradation in Streptomyces spp. by protoplast fusion, Appl. Envir. Microbiol., 47, p. 439-440.
53. Pokorny, M., Vitale, L., 1980, Enzymes as by-products during biosynthesis of antibiotics, în Industrial and Clinical Enzymology, Ed. Vitale, L., Simeon, V., Pergamon Press Oxford, p. 13-25.
54. Pongs, O., 1979, Cloramphenicol, în Antibiotics, Ed.Hahn, F. E., Springer Verlag, 5, p. 26-42.
55. Ramachandra, M., Crawford, D. L., Pometta, A. L., 1987, Extracellular enzyme activities during lignocellulose degradation by Streptomyces spp.: a comparative study of wild-type and genetically manipulated strains, Appl. Envir. Microbiol., 53, p. 2754-2760.
56. Robson, J. M., Chambliss, G. H., 1989, Cellulases of bacterial origin, Enzyme Microbiol. Technol., 11, p. 626-644.
57. Schlochtermeier, A., Miemeier, F., Schrempf, H., 1992, Biochemical and electron microscopic studies of the Streptomyces reticuli cellulase (avicelase) in its mycelium-associated and extracellular forms, Appl. Environ. Microbiol., 58, p. 3240-3248.
58. Schlochtermeier, A., Walter, S., Schroder, J., Moorman, M., Schrempf, H., 1992, The gene encoding the cellulase (avicelase) Cel1 from Streptomyces reticuli and analysis of protein domains, Molec. Microbiol., 6, p. 3611-362.
59. Srinivasan, M. C., Laxman, R. S., Deshpande, M. V., 1991, Physiology and nutritional aspects of actinomycetes: an overview, World J. Microbiol. Biotechnol., 7, p. 171-184.
60. Tiraby, G., Bejar, S., Reynes, J. P., Sicard, P. J., Farber, G. K., Ringe, D., Petsko, G. A., 1989, Genetic, enzymatic and crystallographic studies of the glucose isomerase of two Streptomyces species, în Genetics and molecular biology of industrial microorganisms, Ed. Hershberger, C. L., Queener, S. W., Hegeman, G., ASM, p. 119-126.
61. Vining, L. C., 1990, Functions of secondary metabolites, Ann. Rev. Microbiol., 44, p. 395-427
161

Rodica Mateescu, Călina Petruţa Cornea, Irina Grebenişan, Gh. Câmpeanu
62. Vining, L. C., 1992, Secondary metabolism, inventive evolution and biochemical diversity: a review, Gene, 115, p. 135-140.
63. Wachinger, G., Bronnenmeier, K., Staudenbauer, W. L., Schrempf, H., 1989, Identification of mycelium-associated cellulase from Streptomyces reticuli, Appl. Environ. Microbiol., 55,
p. 2653-2657.64. Wilson, D. B., 1992, Biochemistry and genetics of actinomycete cellulase, Crit.Rev.
Biotechnol., 12, p. 45-63.65. Winkelmann G., Allgaier H., Lupp R., Jung G., Iturin AL- a new long chain iturin a
possessing an unusual high content of C16--amino acids, J Antibiot (Tokyo) 1987 Apr., 40(4): 437-42.
66. Wood, T. M., Bhat, K. M., 1988, Methods for measuring cellulase activities, Methods Enzymol., 160, p. 87-112.
67. Zarnea, G., 1984, Tratat de microbiologie generală, 1, Ed. Academiei.68. Zimmermann, W., 1989, Hemicellulolytic enzyme systems from actinomycetes, în Enzyme
systems for lignocellulose degradation, Ed. Coughlan, M. P., Elsevier Applied Science, p. 167-181.
69. Zimmermann, W., Davis, B., Winter, B., Shou,W., 1992, Actinomycetes and their enzymes for applications in the pulp and paper industry, în Biotechnology in Pulp and Paper Industry, Ed. Kuwahara, M., Shimada, M., Uni Publishers, Co. LdT, p. 145-150.
162