journal of vietnam agricultural science and technology chi/nam 2018/so 11... · 2019-12-26 · 1...
TRANSCRIPT

1
TẠP CHÍKHOA HỌC CÔNG NGHỆNÔNG NGHIỆP VIỆT NAM
Journal of Vietnam Agricultural Science and Technology
NĂM THỨ MƯỜI BA
SỐ 11 NĂM 2018
TỔNG BIÊN TẬP
Editor in chief
GS.TS. NGUYỄN VĂN TUẤT
PHÓ TỔNG BIÊN TẬP
Deputy EditorGS.TS. BÙI CHÍ BỬU
TS. TRẦN DANH SỬU
TS. NGUYỄN THẾ YÊN
THƯỜNG TRỰC
ThS. PHẠM THỊ XUÂN - THƯ KÝ
TÒA SOẠN - TRỊ SỰ
Ban Thông tinViện Khoa học Nông nghiệp Việt Nam
Vĩnh Quỳnh, Thanh Trì, Hà NộiĐiện thoại: (024) 36490503; (024) 36490504; 0949940399
Fax: (024) 38613937;Website: http://www.vaas.org.vnEmail: [email protected]; [email protected]
ISSN: 1859 - 1558
Giấy phép xuất bản số:1250/GP - BTTTT
Bộ Thông tin và Truyền thông
cấp ngày 08 tháng 8 năm 2011
MỤC LỤC1. Võ Thị Minh Tuyển, Nguyễn Thị Huê, Đoàn Văn Sơn,
Phan Quốc Mỹ. Kết quả chọn tạo và khảo nghiệm giống lúa DT80
2. Doãn Thị Hương Giang, Lưu Minh Cúc, Lê Huy Hàm, Phạm Xuân Hội. Kết quả chọn tạo và khảo nghiệm giống lúa chịu ngập OM351
3. Nguyễn Thị Nhung, Trịnh Văn Mỵ, Ngô Thị Huệ, Nguyễn Mạnh Quy, Nguyễn Thị Thu Hương, Đỗ Thị Bích Nga, Ngô Doãn Đảm, Nguyễn Đạt Thoại, Đỗ Thị Hồng Liễu. Khảo nghiệm giống khoai tây KT4 cho sản xuất vụ Đông ở một số tỉnh Đồng bằng sông Hồng
4. Đinh Thị Dinh, Nguyễn Thị Kim Oanh, Nguyễn Văn Tiến. Kết quả chọn tạo giống hoa lan hồ điệp lai HĐ01
5. Vũ Thị Nhiên, Tạ Kim Nhung, Stefan Jouannic, Lê Hùng Lĩnh, Phạm Xuân Hội, Trần Khánh Vân, Trần Vũ Hằng, Phạm Thị Mai, Lê Thị Như, Khổng Ngân Giang. Tạo quần thể lai F1 làm vật liệu khởi đầu để đánh giá vai trò của QTL9 liên quan đến các tính trạng năng suất của tập đoàn lúa Việt Nam
6. Nguyễn Văn Đồng, Nguyễn Anh Vũ, Lê Thị Mai Hương, Nguyễn Trung Anh. Nghiên cứu chuyển cấu trúc gen 35S::GmNAC004 vào giống đậu tương ĐT22 thông qua chủng khuẩn Agrobacterium tumefaciens EHA101
7. Nguyễn Thị Hồng Hải, Khuất Hữu Trung, Trần Đăng Khánh, Lê Thị Hằng, Phạm Thị Lý Thu, Đặng Tất Thành. Tạo dòng và biểu hiện enzyme endolysin tái tổ hợp trong Escherichia coli từ Staphylococcal bacteriophages NA6
8. Nguyễn Trường Khoa, Nguyễn Thúy Điệp, Nguyễn Thái Dương, Nguyễn Như Toản , Phương Hữu Pha, Đặng Thị Thanh Hà, Kiều Thị Dung, Trần Thị Thúy, Trần Đăng Khánh, Khuất Hữu Trung. Tầm soát và thiết kế marker chức năng xác định candidate gen kháng đạo ôn Pit ở một số giống lúa bản địa của Việt Nam
9. Chu Đưc Hà, Nguyễn Thị Minh Nguyệt, Nguyễn Thị Nhài, Nguyễn Bá Ngoc, Khuất Thị Mai Lương, Phạm Thị Lý Thu, Lê Hùng Lĩnh. Thiết lập phương pháp lấy mâu lúa theo tiêu chuẩn ISO/IEC 17025: 2017 phục vụ thử nghiệm xác định locus gen
10. Khuất Thị Mai Lương, Nguyễn Thị Minh Nguyệt, Chu Đưc Hà, Trần Thị Hoa Mỹ, Đinh Văn Phê, Lê Hùng Lĩnh. Thiết kế và xây dựng bộ chỉ thị phân tử phục vụ kiểm định và nghiên cứu di truyền sâm ngọc linh (Panax vietnamensis)
11. Bùi Thị Thu Hương, Đồng Huy Giới, Trần Thị Thu Thủy, Nguễn Thị Ngoc Quỳnh. Nghiên cứu tác động của nano bạc đến quá trình phát sinh hình thái trong nuôi cấy in vitro hoa đồng tiền
3
7
13
19
25
32
37
41
46
50
55

2
TẠP CHÍKHOA HỌC CÔNG NGHỆNÔNG NGHIỆP VIỆT NAM
Journal of Vietnam Agricultural Science and Technology
NĂM THỨ MƯỜI BA
SỐ 11 NĂM 2018
TỔNG BIÊN TẬP
Editor in chief
GS.TS. NGUYỄN VĂN TUẤT
PHÓ TỔNG BIÊN TẬP
Deputy EditorGS.TS. BÙI CHÍ BỬU
TS. TRẦN DANH SỬU
TS. NGUYỄN THẾ YÊN
THƯỜNG TRỰC
ThS. PHẠM THỊ XUÂN - THƯ KÝ
TÒA SOẠN - TRỊ SỰ
Ban Thông tinViện Khoa học Nông nghiệp Việt Nam
Vĩnh Quỳnh, Thanh Trì, Hà NộiĐiện thoại: (024) 36490503; (024) 36490504; 0949940399
Fax: (024) 38613937;Website: http://www.vaas.org.vnEmail: [email protected]; [email protected]
ISSN: 1859 - 1558
Giấy phép xuất bản số:1250/GP - BTTTT
Bộ Thông tin và Truyền thông
cấp ngày 08 tháng 8 năm 2011
12. Lưu Thúy Hòa, Khuất Hữu Trung, Trần Đăng Khánh, Phạm Thị Lý Thu, Trần Văn Ơn. Nghiên cứu đa dạng di truyền cây bát giác liên thu thập ở miền Bắc Việt Nam bằng chỉ thị RAPD
13. Nguyễn Thị Ngoc Lan, Nguyễn Thị Lan Hoa, Nguyễn Thị Thanh Thủy, Lã Tuấn Nghĩa. Nghiên cứu đa dạng di truyền của đoạn gen RBCL ở một số nguồn gen bưởi Việt Nam
14. Biện Anh Khoa, Bùi Phước Tâm, Nguyễn Thị Hồng Loan, Nguyễn Thị Lang. Đánh giá tính chịu mặn của quần thể lai hồi giao OMCS2000*4/Pokkali ở thế hệ BC3F2
15. Trần Long, Lưu Minh Cúc, Nguyễn Quang Sáng, Phạm Xuân Hội. Phân biệt một số cặp giống lúa giống nhau bằng chỉ thị phân tử để hỗ trợ khảo nghiệm DUS
16. Vũ Ngoc Thắng, Nguyễn Thị Yến, Nông Thảo Diễm, Nguyễn Ngoc Quất, Trần Anh Tuấn. Ảnh hưởng của liều lượng phân lân và che phủ đến sinh trưởng và năng suất của giống lạc L14 ở vụ Xuân tại Gia Lâm - Hà Nội
17. Nguyễn Văn Giang, Trần Thị Đào, Trần Thị Thúy Hà, Nguyễn Thu Trang. Ảnh hưởng của điều kiện nuôi cấy tới khả năng sinh tổng hợp Indole-3-acetic acid của vi khuẩn Bacillus sonorensis LĐ18
18. Tạ Hồng Lĩnh, Nguyễn Văn Tuất, Bùi Quang Đãng, Nguyễn Thị Thanh Mai, Trương Hồng. Nghiên cứu ảnh hưởng của tuổi cây giống và phân bón hữu cơ đến cây cà phê vối trồng tái canh tại Tây Nguyên
19. Nguyễn Đình Chiến, Nguyễn Ngoc Dương, Phạm Xuân Liêm. Nghiên cứu một số biện pháp kỹ thuật trồng bông lai VN01-2 trong mùa khô tại Sơn La
20. Cồ Thị Thuỳ Vân, Trần Liên Hà, Nguyễn Văn Minh. Nghiên cứu xây dựng quy trình công nghệ nuôi trồng nấm đầu khỉ Hericium erinaceus trên bã dong đao
21. Nguyễn Thế Quyết, Nguyễn Đưc Thành, Trịnh Quốc Bình, Bùi Thị Lan Hương, Nguyễn Đưc Huy, Phạm Xuân Hội. Xác định khả năng đối kháng của loài Chaetomium spp. với nấm Neoscytalidium dimidiatum gây bệnh đốm nâu thanh long
22. Đỗ Thi Thu Lai, Nguyễn Thị Thùy Linh, Đinh Trường Sơn, Nguyễn Thị Kim Lý, Phạm Thị Minh Phượng. Đánh giá đa dạng nguồn gen đỗ quyên bằng chỉ thị ISSR
23. Nguyễn Thanh Long, Trần Đắc Định, Mai Viết Văn. Hiện trạng khai thác và quản lý nghề lưới kéo đơn ven bờ ở tỉnh Sóc Trăng và Bến Tre
24. Vũ Thị Thanh Hương, Nguyễn Hồng Đưc, Lê Thị Thu Thảo, Ngô Huỳnh Phương Thảo, Nguyễn Quốc Bình.Hiệu quả của chủng Aeromonas hydrophila nhược độc sử dụng Làm vắc-xin cho ăn trong phòng bệnh xuất huyết cá tra giống
60
65
69
76
83
90
95
101
106
111
115
121
125

3
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
1 Viện Di truyền Nông nghiệp
KẾT QUẢ CHỌN TẠO VÀ KHẢO NGHIỆM GIỐNG LÚA DT80Võ Thị Minh Tuyển1, Nguyễn Thị Huê1,
Đoàn Văn Sơn1, Phan Quốc Mỹ1
TÓM TẮTBài báo trình bày kết quả ứng dụng kỹ thuật chiếu xạ và chỉ thị phân tử trong chọn tạo giống lúa chịu mặn. Vật
liệu sử dụng để chiếu xạ tia gamma (liều chiếu là 250 Gy, nguồn Co60) là hạt giống đã được ngâm nước 48 giờ của dòng lúa chịu mặn TL6.2 (mang gen saltol nhưng năng suất thấp). Sử dụng chỉ thị phân tử và môi trường mặn nhân tạo (6‰ NaCl) để sàng lọc dòng đột biến chịu mặn ở thế hệ M4. Đánh giá các đặc điểm nông, sinh học trên đồng ruộng đã chọn được dòng đột biến triển vọng LT6.2-44, mang saltol QTL và chịu được độ mặn 6‰ trong môi trường mặn nhân tạo, đặt tên là DT80. Giống triển vọng DT80 đã được trồng khảo nghiệm trên các chân đất bị nhiễm mặn và gửi khảo nghiệm VCU 3 vụ, khảo nghiệm DUS (vụ Mùa 2015 và 2016) tại Trung tâm Khảo kiểm nghiệm Giống, Sản phẩm Cây trồng Quốc gia. Kết quả khảo nghiệm cho thấy: Giống có thời gian sinh trưởng ngắn ngày, chất lượng tốt và có năng suất cao, ổn định.
Từ khóa: Tia gamma, giống lúa đột biến, chỉ thị phân tử, chịu mặn
I. ĐẶT VẤN ĐỀHiện nay, biến đổi khí hậu diễn biến ngày một
phức tạp và khó lường, hiện tượng nước biển dâng cao làm cho diện tích đất trồng lúa bị nhiễm mặn ngày một tăng đặc biệt là ở các tỉnh đồng bằng ven biển (Hoàng Ngọc Vệ, 2017). Một số nghiên cứu gần đây cho rằng Việt Nam là 1 trong những nước bị ảnh hưởng nhiều nhất bởi biến đổi khí hậu (Bộ Tài nguyên Môi trường, 2012). Xâm nhập mặn kéo dài có thể dân đến một số tổn hại đáng kể của hệ sinh thái, đe dọa đến đa dạng sinh học và ảnh hưởng tiêu cực đến sinh kế của người dân (Trung and Tri, 2012). Chọn tạo các giống lúa chất lượng cho năng suất cao, có khả năng chịu mặn là rất cần thiết hiện nay. Xuất phát từ thực tế trên, nhóm tác giả đã tiến hành cải tiến dòng lúa chịu mặn TL6.2 bằng phương pháp gây đột biến thực nghiệm kết hợp chỉ thị phân tử.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưuDòng lúa TL6.2 mang QTLs/gen Saltol, do Viện
Di truyền Nông nghiệp chọn lọc từ thế hệ BC2F4 của tổ hợp lai LT6/FL478. Vì còn một số nhược điểm: Năng suất thấp, yếu cây, nên dòng TL6.2 được sử dụng làm nguồn vật liệu gây đột biến với mục đích chọn được dòng đột biến mới cho năng suất cao và khả năng chống chịu tốt hơn.
2.2. Phương pháp nghiên cưu- Chiếu xạ gây đột biến (nguồn chiếu xạ Co60, liều
chiếu 250 Gy, địa điểm chiếu xạ: Bệnh viện 103, Hà Nội, chiếu xạ hạt ướt - ngâm nước 48 giờ).
- Chọn lọc dòng mang gen chịu mặn thông qua phương pháp PCR đối với các chỉ thị SSR liên kết với QTL quy định tính chịu mặn (Saltol).
- Phương pháp Thanh lọc mặn trong phòng theo IRRI (2006).
- Đánh giá đặc điểm nông sinh học của cây lúa theo tiêu chuẩn của Viện Nghiên cứu Lúa Quốc tế (IRRI, 2002).
- Khảo nghiệm DUS và khảo nghiệm VCU do Trung tâm Khảo kiểm nghiệm Giống, Sản phẩm Cây trồng Quốc gia thực hiện.
- Số liệu được xử lý bằng phần mềm Excel và IRRISTAT 4.0.
2.3. Thời gian và địa điểm nghiên cưu Nghiên cứu được thực hiện từ năm 2011 đến
năm 2017 tại Phòng thí nghiệm, nhà lưới và ruộng thí nghiệm của Viện Di truyền Nông nghiệp.
III. KẾT QUẢ NGHIÊN CỨU
3.1. Kết quả chon tạo giống lúa chịu mặn DT80Vụ Mùa 2011, dòng lúa chịu mặn ban đầu TL6.2
được chiếu xạ để gây đột biến và được trồng, đánh giá tại khu ruộng thí nghiệm của Viện Di truyền Nông nghiệp.
Ở thế hệ M1, thu hỗn để phát triển thành quần thể chọn lọc ở thế hệ M2 (vụ Xuân 2012). Các biến dị về thời gian sinh trưởng, các yếu tố cấu thành năng suất, màu sắc hạt, khả năng chống chịu ở thế hệ M2 được thu riêng. 58 cá thể xuất hiện các biến dị được phát triển thành dòng ở thế hệ M3 (vụ Mùa 2012). Qua chọn lọc gen chịu mặn bằng CTPT, thử mặn nhân tạo và đánh giá một số đặc điểm nông sinh học chính đã xác định được 20 dòng đột biến

4
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
chịu mặn ở thế hệ M4 (vụ xuân 2013). Tiếp tục cho tự thụ và đánh giá trên đồng ruộng đã chọn lọc được 16 dòng đột biến ở thế hệ M5 (vụ Mùa 2013) và sàng lọc được 3 dòng đột biến triển vọng ở thế hệ M6 (vụ Xuân 2014). Dòng triển vọng LT6.2-44 được đặt tên DT80. Dòng triển vọng DT80 tiếp tục được khảo
nghiệm tác giả và gửi khảo nghiệm quốc gia (từ vụ Mùa 2015).
Dòng triển vọng DT80 tiếp tục được kiểm tra gen chị mặn bằng CTPT (Hình 1), thử mặn trong môi trường mặn nhân tạo 6‰, ở giai đoạn cây con (Hình 2, 3).
Ảnh điện di trên gel agarose 2,5% (ladder: 1000 bp)ở hình 1 cho thấy các cá thể DT80 kiểm tra đều mang QTLs/gen Saltol. Các vạch băng ADN của các dòng lúa DT80 đều trùng vạch băng của dòng FL478 và dòng TL6.2, mang QTLs/gen Saltol.
Trong môi trường mặn nhân tạo, nồng độ mặn 6‰, và được giữ trong 15 ngày, các dòng lúa DT80 đều thể hiện khả năng chịu mặn điểm từ 3 - 5 (chống chịu TB - chống chịu), tỷ lệ chết từ 0 - 15%. Dòng đối chứng nhiễm IR29, tỷ lệ cây chết là 100%. Tiếp tục tăng độ mặn của môi trường mặn lên độ mặn 9‰, tỷ lệ chết ở tất cả các dòng tăng từ 90 - 100%. Như vậy trong môi trường mặn nhân tạo, giống lúa DT80 có khả năng chịu mặn khoảng 5 - 6‰.
3.2. Kết quả khảo nghiệm giống lúa DT80
3.2.1. Kết quả khảo nghiệm tác giảQua đánh giá một số đặc điểm nông, sinh học
chính và khả năng chống chịu của giống DT80 so với giống gốc ban đầu TL6.2 ở cả 2 vụ Xuân và Mùa năm 2015 tại Hợp tác xã Yên Sở, Hoài Đức, Hà Nội cho thấy giống lúa triển vọng DT80 vân giữ được nhiều tính trạng quý so với giống gốc ban đầu TL6.2 như: Thời gian sinh trưởng ngắn ngày, cao cây trung
bình, thoát cổ bông, lá đòng thẳng, đẻ nhánh chụm, màu sắc vỏ trấu, tỷ lệ lép thấp...
Bên cạnh những đặc tính giống dòng gốc ban đầu, giống DT80 đã được cải tiến rất nhiều về năng suất và các yếu tố cấu thành năng suất như: Số hạt chắc và cấu trúc hạt trên bông, chiều dài lá đòng và số bông hữu hiệu trên khóm.
3.2.2. Kết quả khảo nghiệm quốc giaKết quả khảo nghiệm DUS tại Trạm Khảo kiểm
nghiệm Giống cây trồng Văn Lâm, Hưng Yên, qua 2 vụ khảo nghiệm (vụ Mùa 2015 và vụ Mùa 2016) cho thấy: Giống đã thể hiện tính khác biệt so với giống tương tự TBR45, có tính đồng nhất và tính ổn định.
Giống lúa DT80 đã được khảo nghiệm quốc gia VCU 3 vụ: Vụ Mùa 2015, vụ Xuân 2016 và vụ Mùa 2016.
Đánh giá khả năng chống chịu với một số sâu bệnh hại chính trên đồng ruộng, trong điều kiện có sử dụng thuốc bảo vệ thực vật (Bảng 2) cho thấy giống DT80 có khả năng chống chịu khá (điểm 0 - 1 và 1 - 3), khả năng chống chịu với bệnh bạc lá (1 - 3 và 3 - 5) tốt hơn giống đối chứng Bắc thơm 7 (3 - 5 và 5 - 7).
Hình 1. Ảnh điện di sản phẩm PCR với chỉ thị RM562 và chỉ thị RM3412b liên kết liên kết chặt với QTLs/gen Saltol
Hình 2. Trước khi đưa vào thử mặn Hình 3. 15 ngày sau khi thử trong môi trường mặn 6‰

5
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Kết quả đánh giá năng suất thực thu tại các điểm khảo nghiệm (Bảng 3) cho thấy: Năng suất bình quân tại các điểm khảo nghiệm ở vụ Mùa 2015 và vụ Mùa 2016, giống DT80 cho năng suất đạt 51,8 - 52,6 tạ/ha, tương đương giống Hương thơm số 1 và vượt
đối chứng Bắc thơm 7 khoảng 10 - 12%. Vụ Xuân năm 2016, giống có năng suất cao (64,41 tạ/ha), cao hơn cả 2 giống đối chứng Hương thơm 1 (cao hơn khoảng 6%) và Bắc thơm 7 (cao hơn khoảng 23%).
Bảng 1. Một số đặc điểm nông, sinh học chính của giống lúa DT80 năm 2015 - Hoài Đức, Hà Nội
Bảng 2. Mức độ nhiễm sâu bệnh của các giống khảo nghiệmĐơn vị tính: Điểm
Nguồn: Bộ môn Đột biến & Ưu thế lai - Viện Di truyền Nông nghiệp
Nguồn: Trung tâm Khảo kiểm nghiệm Giống, Sản phẩm Cây trồng Quốc gia
TT Các chỉ tiêuDT80 TL6.2 (Giống ban đầu)
Vụ Xuân Vụ Mùa Vụ Xuân Vụ Mùa
1 TGST (ngày) 135 105 136 105
2 Cao cây (cm) 110 109 111 110
3 Thoát cổ bông (cm) 2,1 2,2 3 2,5
4 Lá đòng (quan sát sớm 70 ngày) Thẳng Thẳng
5 Chiều dài lá đòng 27,5 26,5 23,5 21,5
6 Mầu sắc vỏ trấu Vàng sáng Vàng sang
7 Số bông hữu hiệu trên khóm 5,5 5,2 4,8 4,5
8 Kiểu dáng đẻ nhánh Chụm Chụm
9 Khối lượng 1000 hạt (gam) 21,2 20,8 22,5 22,3
10 Hạt chắc (hạt) 245 216 148 134
11 Hạt lép (hạt) 12,3 13,5 12,5 13,1
12 Chiều dài hạt thóc (mm) 6,4 6,5 6,7 6,8
13 Chiều rộng hạt thóc (mm) 2,2 2,1 2,3 2,1
14 Cấu trúc hạt trên bông Hạt xếp xít Hạt xếp thưa
15 NSTT (tạ/ha) 74,5 63,1 55,6 52,5
Vụ Tên giốngBệnh
đạo ôn hại lá
Bệnh đạo ôn
cổ bông
Bệnh bạc lá
Bệnh khô vằn
Bệnh đốm nâu
Sâu đục thân
Sâu cuốn lá Rầy nâu
Mùa 2015
DT 80 0-1 0-1 1-3 1-3 1-3 0-1 3-5 0-1
HT1 0-1 0-1 1-3 1-3 1-3 0-1 1-3 0-1
TBR225 0-1 0-1 3-5 1-3 1-3 0-1 3-5 1-3
BT7 0-1 0-1 3-5 3-5 0-1 1-3 3-5 1-3
Xuân 2016
DT 80 0-1 0-1 0-1 0-1 0-1 0-1 0-1 0-1
HT1 0-1 0 1-3 3-5 1-3 0-1 1-3 0-1
TBR225 1-2 0 0-1 3-5 0-1 0-1 1-3 1-3
BT7 1-2 0-1 3-5 3-5 1-3 0-1 3-5 0-1
Mùa 2016
DT 80 2-3 0-1 3-5 1-3 1-3 3-5 1-3 0-1
HT1 2-3 0-1 3-5 1-3 1-3 3-5 1-3 0-1
BT7 2-3 0-1 5-7 3-5 0-1 3-5 3-5 1-3

6
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Kết quả đánh giá các chỉ tiêu chất lượng gạo của giống DT80 (Bảng 4) cho thấy: Giống có tỷ lệ gạo lật (80%) tương đương giống Hương thơm 1 và cao hơn giống Bắc thơm 7. Tỷ lệ gạo nguyên/gạo xát của giống DT80 (83,5%) cao hơn giống Hương thơm 1, thấp hơn tỷ lệ này của giống BT7 (95%). Giống DT80 có hạt gạo dài tương đương giống Hương thơm 1. Độ bền gel tương đương các giống đối chứng. Giống DT80 có hàm lượng amylose khoảng 13,4%, cơm ngon, mềm và dẻo.
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luậnGiống lúa DT80 là giống lúa thuần, được chọn
tạo bằng phương pháp gây đột biến dòng lúa chịu mặn TL6.2 kết hợp chỉ thị phân tử. Giống đã được
khảo nghiệm quốc gia và khảo nghiệm sản xuất tại các tỉnh phía Bắc.
Kết quả khảo nghiệm DUS cho thấy giống DT80 thể hiện tính khác biệt, tính đồng nhất và ổn định.
Kết quả khảo nghiệm cơ bản và khảo nghiệm sản xuất ở nhiều địa phương cho thấy:
Giống lúa DT80 có chiều cao cây dao động từ 107 - 109,6 cm, thời gian sinh trưởng thuộc nhóm giống ngắn ngày, vụ Mùa khoảng 105 - 110 ngày và vụ Xuân khoảng 135 - 140 ngày. Giống có dạng hình cây gọn, thân cứng, lá đứng, khả năng đẻ nhánh khá, độ thuần đồng ruộng cao. Tại các tỉnh phía Bắc, giống lúa DT80 cho năng suất cao và ổn định. Trong khảo nghiệm cơ bản năng suất trung bình của giống đạt 51,8 - 64,4 tạ/ha, cao hơn Bắc thơm 7 khoảng 8 - 13%. Giống DT80 có hạt gạo thon dài, trong, cơm dẻo, ngon, đậm cơm, hàm lượng amylose khoảng 13,4%.
Bảng 3. Năng suất thực thu của các giống nhóm ngắn ngày chất lượng Đơn vị tính: tạ/ha
Nguồn: Trung tâm Khảo kiểm nghiệm Giống, Sản phẩm Cây trồng Quốc gia
Bảng 4. Chỉ tiêu chất lượng gạo của các giống khảo nghiệm vụ Mùa 2016
Nguồn: Trung tâm Khảo kiểm nghiệm Giống, Sản phẩm Cây trồng Quốc gia
Vụ Tên giốngĐiểm khảo nghiệm
Bình quânHưng
YênHải
DươngThái Bình
Hòa Bình Yên Bái Thanh
Hóa Nghệ An
Mùa 2015
DT 80 64,80 66,41 61,5 50,00 45,83 41,07 53,37 52,62
HT1 57,51 56,51 61,03 52,00 56,27 47,00 55,67 54,18
BT7 53,29 49,37 43,53 51,33 50,17 43,03 48,03 47,39CV (%) 6,0 5,8 4,3 4,2 6,1 6,2 4,1LSD0,05 5,81 5,41 3,58 3,75 5,51 4,53 3,74
Xuân 2016
DT 80 62,79 70,83 71,27 55,50 66,40 64,97 62,27 64,41
HT1 61,66 63,81 68,40 55,20 59,43 58,00 60,70 60,63
BT7 49,72 53,24 53,93 47,60 56,17 54,17 60,73 52,42CV (%) 7,6 3,9 5,1 4,8 3,2 3,0 3,6LSD0,05 7,58 3,99 5,48 4,58 3,27 3,19 3,63
Mùa 2016
DT 80 53,23 62,58 50,17 55,60 43,73 54,50 54,30 51,76HT1 64,41 58,33 60,27 53,00 52,03 58,00 50,87 56,22BT7 52,73 49,00 39,10 49,30 46,77 47,93 42,03 47,04CV (%) 6,9 4,7 6,7 5,3 6,4 4,4 7,5LSD0,05 7,21 4,43 5,77 4,65 5,62 4,13 6,11
Tên giống Tỷ lệ gạo lật (%)
Tỷ lệ gạo xát (%)
Tỷ lệ gạo nguyên/gạo
xát (%)
Tỷ lệ D/R
Độ bền gel
Tỷ lệ trắng trong
(%)
Hàm lượng Amylose (% CK)
DT 80 80,06 65,30 83,51 2,86 Mềm 47,85 13,39HT1 81,18 69,72 64,29 2,93 Mềm 55,15 16,76BT7 78,72 70,07 95,09 2,75 Mềm 54,85 14,22

7
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Trong điều kiện đồng ruộng, giống ít nhiễm bệnh đạo ôn, rầy nâu, nhiễm nhẹ bệnh bạc lá, khô vằn trong vụ Mùa. Giống có khả năng chịu rét, chống đổ tốt. Giống DT80 có thể phát triển tốt trong điều kiện mặn nhân tạo khoảng 5 - 6‰. Tuy nhiên, do có bản lá to nên trong điều kiện vụ Mùa cần điều chỉnh bón phân hợp lý để tránh nhiễm bệnh bạc lá.
Giống lúa DT80 có thể trồng được cả 2 vụ và phát triển tốt trên chân đất vàn và vàn cao tại các tỉnh phía Bắc.
4.2. Đề nghịTiếp tục sản xuất thử giống lúa mới DT80 tại các
vùng sinh thái khác nhau để sớm công nhận giống lúa mới.
TÀI LIỆU THAM KHẢOBộ Tài nguyên và Môi trường, 2012. Kịch bản biến đổi
khí hậu, nước biển dâng cho Việt Nam. Lần 1. Nhà xuất bản Tài nguyên - Môi trường và Bản đồ Việt Nam. Hà Nội, 84 trang.
Hoàng Ngoc Vệ, Trần Hồng Thái, 2017. Đánh giá tác động của xâm nhập mặn đến sử dụng đất nông nghiệp ở các huyện ven biển của tỉnh Nghệ An trong bối cảnh biến đổi khí hậu. Tạp chí Phát triển Khoa học và Công nghệ, 2017.
International Rice Research Institute (IRRI), 2002. Standard Evaluation System for rice, November, 2002.
International Rice Research Institute (IRRI), 2006. Rice Knowledge Bank, 2006.
Trung, N. H. and Tri, V. P. D., 2012. Possible Impacts of Seawater Intrusion and Strategies for Water Management in Coastal Areas in the Vietnamese Mekong Delta in the Context of Climate Change in Coastal Disasters and Climate Change in Vietnam. Science Direct. 219-349.
Breeding and testing of rice variety DT80Vo Thi Minh Tuyen, Nguyen Thi Hue,
Doan Van Son, Phan Quoc MyAbstractThis article presents the results of application of irradiated techniques and molecular markers for breeding of salt tolerant rice variety. The seeds of salt tolerant rice variety TL6.2 (low yield and carrying saltol QTL controlling the salt tolerance). The M4 progenies were screened by using marker assisted selection (MAS) and salt solution (6‰ NaCl) in green house. The promising mutant line LT6.2-44 named DT80 that carrying saltol QTL and withstand salinity of 6‰ NaCl was selected by evaluating agronomic traits on field. DT80 was tested in salinity fields by the National Testing Center for Crops for VCU in three crop seasons and DUS in 2 similar crop seasons. The tested results showed that mutant DT80 variety had short duration, good quality and high and stable yield. Keywords: Gamma rays, mutant variety, salt tolerant, marker assisted selection
Ngày nhận bài: 18/9/2018Ngày phản biện: 25/9/2018
Người phản biện: TS. Dương Xuân TúNgày duyệt đăng: 15/10/2018
1 Viện Di truyền Nông nghiệp
KẾT QUẢ CHỌN TẠO VÀ KHẢO NGHIỆM GIỐNG LÚA CHỊU NGẬP OM351 Doãn Thị Hương Giang1, Lưu Minh Cúc1, Lê Huy Hàm1, Phạm Xuân Hội1
TÓM TẮTNghiên cứu được tiến hành nhằm đưa gen chịu ngập Sub1 vào giống lúa AS996, mà vân giữ nguyên các đặc tính
cơ bản của giống. Tiến hành khảo nghiệm các yếu tố cấu thành năng suất và năng suất của giống mới được tạo ra tại một số tỉnh thuộc Đồng bằng sông Cửu Long và đánh giá tính chịu ngập của giống lúa. Sử dụng phương pháp MABC đã chọn tạo giống lúa mang gen chịu ngập Sub1 và gần 100% nền gen của giống nhận gen AS996. Đó là các dòng triển vọng C13, C7, C10. Giống lúa OM351 được chọn tạo từ dòng C13. Đã xác định sự có mặt của gen chịu ngập trong giống OM351 sử dụng chỉ thị phân tử SSR liên kết gen chịu ngập. Trong điều kiện nhân tạo ngập chìm hoàn toàn 14 ngày, giống OM351 đạt điểm 3 (60% cây sống). Kết quả khảo nghiệm tại các tỉnh thuộc Đồng bằng sông Cửu Long và miền Đông Nam bộ cho thấy giống lúa OM351 chống chịu sâu bệnh khá, năng suất trung bình đạt cao hơn đối chứng VND95-1 và OMCS2000 từ 10,7 đến 16,1%. Mô hình trình diễn giống lúa OM351 cho thấy giống cho năng suất cao hơn năng suất giống lúa nền gen AS996, khả năng chống chịu sâu bệnh khá, mang gen chịu ngập, có thể phát triển tiếp trong sản xuất.
Từ khóa: Chỉ thị phân tử, gen chịu ngập, giống lúa, khảo nghiệm

8
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
I. ĐẶT VẤN ĐỀ Biến đổi khí hậu là một trong những thách thức
lớn nhất của nhân loại thế kỷ 21; hiện tượng biến đổi khí hậu kéo theo sự gia tăng của nhiệt độ trái đất, sự dâng cao của mực nước biển gây nên ngập lụt và gây nhiễm mặn nguồn nước ảnh hưởng lớn tới sản xuất nông nghiệp (Phạm Khôi Nguyên, 2009). Mỗi năm lũ lụt đã làm mất hàng triệu tấn lúa - chiếm khoảng 25% sản lượng lúa gạo, thất thoát hàng tỷ USD. Trong đó phổ biến nhất là ở châu Á, nơi sản xuất hơn 90% lúa gạo thế giới.
Hiện tượng ngập úng cũng là một vấn đề phổ biến của sản xuất nông nghiệp nước ta. Do biến đổi khí hậu đang xảy ra, vào cuối thế kỷ 21, nếu mực nước biển dâng một mét thì 6,3% diện tích Việt Nam, khoảng 39% diện tích Đồng bằng sông Cửu Long (ĐBSCL), trên 10% diện tích Đồng bằng sông Hồng và Quảng Ninh, trên 2,5% diện tích thuộc các tỉnh ven biển miền Trung và trên 20% diện tích thành phố Hồ Chí Minh có nguy cơ bị ngập (Bộ Tài nguyên và Môi trường, 2012). Ngoài ra, những trận mưa lớn kéo dài có thể gây ngập úng một khu vực sản xuất rộng lớn trong vòng vài ngày làm thất thu mùa màng. Tính chống chịu ngập úng đã được phát hiện từ rất lâu trên các giống lúa địa phương nhưng việc chuyển những tính trạng này vào các giống lúa cao sản gặp rất nhiều khó khăn do sự di truyền phức tạp và sự biến động lớn của chúng. Gần đây, các nhà khoa học đã định vị thành công gen chống chịu ngập Sub1 trên nhiễm sắc thể số 9 của cây lúa (Thomson et al., 2010). Nghiên cứu này nhằm kiểm tra sự có mặt của gen chịu ngập Sub1 trong giống lúa chịu ngập mới được chọn tạo OM351 bằng phương pháp chọn giống nhờ chỉ thị phân tử kết hợp lai hồi giao (MABC). Mục đích của việc chọn tạo là đưa gen chịu ngập Sub1 vào giống lúa AS996, mà vân giữ nguyên các đặc tính cơ bản của giống. Đồng thời tiến hành khảo nghiệm các yếu tố cấu thành năng suất và năng suất của giống mới được tạo ra tại một số tỉnh thuộc ĐBSCL và đánh giá tính chịu ngập của giống lúa OM351.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưu- Dòng lúa triển vọng: C7, C10, C13 mang gen
Sub1 chịu ngập, được chọn tạo từ tổ hợp lai AS996/IR64 Sub1 và lai hồi giao 3 lần, chọn tạo bằng phương pháp chọn giống nhờ chỉ thị phân tử kết hợp lai hồi giao MABC và chọn giống truyền thống.
- Giống lúa đối chứng chọn tạo: Là giống AS996 được tạo ra từ tổ hợp lai IR64/Oryza rufipugon,
được công nhận giống chính thức theo Quyết định số 5310/QĐ/BNN-KHCN ngày 29/11/2002 của Bộ Nông nghiệp và Phát triển nông thôn. Đây là giống lúa ngắn ngày, chất lượng gạo trung bình, năng suất khá cao được trồng phổ biến ở vùng Đồng bằng sông Cửu Long, không có khả năng chịu ngập.
- Giống lúa đối chứng chịu ngập: IR64 Sub1 được nhập nội từ Viện Nghiên cứu Lúa Quốc tế, mang locus gen Sub1, là QTL chính chịu trách nhiệm tới 70% tính chịu ngập chìm trong giống lúa, có khả năng chịu ngập 14 ngày.
- Giống lúa đối chứng mân cảm ngập: IR42, là giống nhập nội từ Viện Nghiên cứu Lúa Quốc tế.
- Các chỉ thị phân tử SSR lúa, các hóa chất sinh học phân tử và vật tư thí nghiệm.
2.2. Phương pháp nghiên cưu- Phương pháp chọn giống MABC: AS996 được
lai với IR64 Sub1 để thu hạt lai F1. Thế hệ F1 được lai trở lại với AS996 để tạo thế hệ BC1F1, BC2F1 và BC3F1. Năm 2012 bắt đầu cho tự thụ và chọn giống truyền thống đánh giá các đặc tính nông sinh học, yếu tố cấu thành năng suất và tính kháng sâu bệnh hại trên đồng ruộng. Kết hợp phương pháp chọn lọc cá thể thông qua công nghệ chỉ thị phân tử kết hợp với chọn giống truyền thống, chọn lọc dòng có kiểu hình đẹp, năng suất cao, chống chịu ngập, thời gian sinh trưởng ngắn để khảo nghiệm.
- Phân tích sự có mặt của gen kháng trong các dòng của giống OM351 bằng các chỉ thị SSR liên kết gen kháng, được nhân bản trong phản ứng PCR và điện di sản phẩm PCR trên gel polyacrylamide 6%, nhuộm bạc để phát hiện băng vạch ADN.
- Thanh lọc ngập nhân tạo theo phương pháp thanh lọc giai đoạn mạ của Pamplona và cộng tác viên (2007): Tiến hành trong bể bằng bê tông của nhà lưới. Giống IR42, IR64-Sub1, các dòng của giống OM351 được ngâm ủ trong đĩa petri với một lớp giấy lọc ẩm ở 30oC để trong tối 3 ngày rồi đem gieo. Bố trí thí nghiệm theo khối hoàn toàn ngâu nhiên 3 lần lặp lại. Cho hạt giống phát triển thêm từ 14 đến 21 ngày khi chiều cao cây khoảng từ 25 đến 35 cm sẽ bắt đầu làm ngập. Không bón phân cho giống. Loại bỏ bớt cây trên mỗi hàng, chỉ để 20 cây/hàng. Bắt đầu làm ngập ở độ sâu 1 m. Quan sát giống IR42 từ 6 ngày sau khi làm ngập bằng cách lấy ra vài cây IR42 khỏi bể và kiểm tra cây có trở nên mềm yếu hay không ở những vùng có chức năng sinh trưởng (đỉnh sinh trưởng của thân hoặc rễ). Mở van thoát nước khi 70% đến 80% giống IR42 bị tổn thương. Đánh giá tính chịu ngập theo công thức của Suprihatno (1980):

9
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
%CS = ˟ 100%S
% sống sót của giống đ/c kháng Trong đó: %S: phần trăm cây sống sót; %CS: tính chống chịu ngập theo thang điểm của Suprihatno (1980).
Bảng 1. Thang điểm đánh giá tính chống chịu ngập (Suprihatno, 1980)
- Đánh giá năng suất và các yếu tố cấu thành năng suất, khả năng chống chịu một số loại sâu bệnh chính theo Quy chuẩn Quốc gia về khảo nghiệm giá trị canh tác và sử dụng giống lúa QCVN 01-55:2011/BNNPTNT.
- Số liệu xử lý theo chương trình IRRISTAT 5.0 và CropStat 7.2.
2.3. Thời gian và địa điểm nghiên cưu- Thời gian nghiên cứu: 2013 - 2016.- Địa điểm nghiên cứu: Vụ Đông Xuân năm 2013
- 2014 và vụ Hè Thu 2014 tại Bạc Liêu; Vụ Đông Xuân
2014 - 2015 tại Đồng Tháp, An Giang, Trà Vinh, Bạc Liêu, Long An, Bến Tre; Vụ Hè Thu 2015 tại Tiền Giang, An Giang, Cờ Đỏ, Thới Lai, Kiên Giang, Tây Ninh và Bình Thuận; Vụ Hè Thu 2016 tại Bạc Liêu; Thí nghiệm thử ngập nhân tạo tại Viện Lúa ĐBSCL; Thí nghiệm kiểm tra gen chống chịu ngập tại Viện Di truyền Nông nghiệp.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Tóm tắt kết quả chon tạo giống lúa OM351 chịu ngập
Vụ Xuân năm 2010 đến năm 2013 lai tạo dòng mang gen chịu ngập bằng phương pháp lai nhân tạo giữa giống nhận gen AS996 và giống cho gen IR64 Sub1. Tạo dòng lai trở lại với giống nhận gen từ BC1F1 đến BC3F1 kết hợp sử dụng chỉ thị phân tử phân tích dòng mang gen chịu ngập và nền gen của giống nhận gen AS996. Đến thế hệ BC3F1 thu được dòng mang gen chịu ngập Sub1 và giữ được 100% nền gen của giống nhận gen trên cơ sở phân tích bằng chỉ thị SSR.
Bằng phương pháp chọn lọc theo phả hệ, đến vụ Đông Xuân năm 2013 - 2014 thu được các dòng triển vọng C13; C7, C10 để đánh giá năng suất và các yếu tố cấu thành năng suất tại Bạc Liêu.
Điểm Tính chống chịu ngập (cây sống)1 100% hoặc hơn3 95 - 99%5 75 - 94%7 50 - 74%9 0 - 49%
Bảng 2. Năng suất và các yếu tố cấu thành năng suất các dòng triển vọng vụ Đông Xuân 2013 - 2014
TT Tên dòng/giống
TGST (ngày)
Cao cây (cm) Bông/ m2 Hạt chắc/
bôngP 1000 hạt
(gr)NSTT
(tấn/ha)% so với
giống ĐC1 C13-1 98 105,2 382,8 128,7 27,2 8,56 28,12 C13-5 97 109,4 349,8 130,2 27,3 8,10 20,73 C7 97 109,0 412,5 125,5 27,0 7,80 16,24 C10 96 113,1 330,0 125,8 26,8 7,24 7,85 C13-4 95 111,2 396,0 115,5 27,0 7,22 7,66 C13-3 95 112,5 379,5 110,0 27,0 7,13 6,27 C13-2 94 102,4 330,1 125,4 27,3 7,02 4,68 C13-6 95 102,2 346,5 102,0 27,4 6,91 2,89 AS996 Đ/c 97 100 373,0 115,0 26,2 6,71 0
CV (%) 2,6LSD0,05 0,33
Kết quả khảo sát 8 dòng triển vọng cho thấy, các dòng đều có thời gian sinh trưởng tương đương với giống AS996, từ 94 - 98 ngày. Chiều cao cây đều cao hơn giống đối chứng từ 2,2 đến 13,1 cm. Số bông/m2 thay đổi từ 330 đến 412,5 bông; số hạt chắc/bông đạt được từ 102 đến 130,2 hạt. Trọng lượng 1000 hạt của tất cả các dòng đều cao hơn giống đối chứng, đạt
từ 26,8 đến 27,3 gam. Từ kết quả năng suất thực thu của các dòng trong vụ Đông Xuân 2013 - 2014, các dòng khảo sát đều có năng suất cao hơn giống đối chứng từ 2,8 đến 28,1%. Từ kết quả trên, đã chọn ra dòng triển vọng C13-1 có năng suất cao nhất 8,56 tấn/ha để tiếp tục đánh giá và chọn lọc trong vụ Hè Thu 2014.

10
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Tiếp tục khảo sát 8 dòng được chọn ra từ dòng C13-1 (Bảng 3) cho thấy, năng suất của các dòng đều dao động trong khoảng 5,5 - 5,75 tấn/ha, vượt so với đối chứng từ 1,9 - 10,3%. Trong đó, dòng C13-1-4 có năng suất vượt trội hơn so với các dòng khác, đạt 5,75 tấn/ha.
Bảng 4. Đánh giá tính chống chịu sâu bệnh của các dòng vụ Hè Thu 2014
Kết quả đánh giá tính chống chịu sâu bệnh trên các dòng ở bảng 4 cho thấy các dòng C13-1 đều có khả năng chống chịu sâu bệnh trên đồng ruộng từ điểm 1 - 3.
3.2. Kết quả đánh giá tính chống chịu ngập của các dòng triển vong
Năm 2014, tổng số 5 dòng triển vọng mang gen Sub1, năng suất cao đã được đánh giá tính chống chịu ngập theo phương pháp đánh giá của IRRI. Kết quả ở bảng 5 cho thấy các dòng triển vọng mang gen Sub1 đã thể hiện tính chống chịu ở mức độ khá cao.
Tỉ lệ cây sống sót cao nhất là dòng C13-1-4 và C13-1-5 với tỉ lệ cây sống sót từ 55 - 60% ở thí nghiệm thử ngập hoàn toàn nhân tạo trong 14 ngày.
Bảng 5. Kết quả đánh giá nhân tạo tính chống chịu ngập các dòng triển vọng
Dòng C13-1-4 cho năng suất cao nhất và thể hiện tính chịu ngập cấp 3 với 60% số cây sống sau 14 ngày ngập hoàn toàn và giống chuẩn mân cảm IR42 chết hết. Dòng C13-1-4 đặt tên là giống OM351 và được gửi đi đánh giá khảo nghiệm trong sản xuất.
3.3. Kết quả xác định sự có mặt của gen Sub1 trong giống OM351 bằng chỉ thị phân tử
Sau khi đã được khảo sát về các yếu tố cấu thành năng suất, năng suất thực thu và đánh giá tính chống chịu các sâu bệnh cơ bản, tính chịu ngập, các dòng triển vọng đã được phân tích để xác định sự có mặt của gen chịu ngập Sub1 sử dụng chỉ thị phân tử SSR liên kết gen Sub1. Hình 1 và hình 2 là các hình ảnh xác định các dòng mang gen chống chịu ngập Sub1 bằng các chỉ thị phân tử liên kết gen trên các dòng triển vọng từ giống OM351 (AS996-Sub1) bằng chỉ thị SC3 và RM23662.
Bảng 3. Đặc tính nông học và năng suất các dòng triển vọng, vụ Hè Thu 2014
TT Tên dòng/giống
TGST (ngày)
Cao cây (cm) Bông/m2 HC/Bông P 1000 hạt NSTT
(T/ha)% so với
giống ĐC1 C13-1-1 100 102,5 280,8 108,7 27,5 5,56 6,72 C13-1-3 96 110,0 259,8 80,5 27,0 5,50 5,53 C13-1-4 100 105,8 296,2 92,5 27,0 5,75 10,34 C13-1-5 105 115,0 286,0 95,8 26,3 5,64 8,25 C13-1-6 100 102,5 280,8 108,7 27,5 5,56 6,76 C13-1-7 105 112,0 273,3 100,5 26,2 5,52 1,97 C13-1-8 105 105,0 270,2 108,6 26,5 5,52 1,98 C13-1-9 102 115,0 268,9 92,5 26,2 5,50 1,99 AS996 Đ/c 102 105,5 263,0 85,0 26,0 5,21 0
CV (%) 2,7LSD0,05 0,26
TTTên
dòng/giống
Đạo ôn
Rầy nâu
Bạc lá
Khô vằn
Sâu cuốn
lá
Chống đổ
1 C13-1-1 1-3 1-3 0-1 1-3 1-3 32 C13-1-3 1-3 1-3 0-1 1-3 1-3 33 C13-1-4 1-3 1-3 0-1 1-3 1-3 34 C13-1-5 1-3 1-3 0-1 1-3 1-3 35 C13-1-6 1-3 1-3 0-1 1-3 1-3 36 C13-1-7 1-3 1-3 1-3 1-3 1-3 37 C13-1-8 1-3 1-3 0-1 1-3 1-3 38 C13-1-9 1-3 1-3 0-1 1-3 1-3 3
9 AS996 Đ/C 1-3 1-3 0-1 1-3 1-3 3
TT Tên dòng /giống
% cây sống
Cấp chống chịu (1-9) Đánh giá
1 C13-1-1 45 3 Chống chịu2 C13-1-3 35 5 Chống chịu3 C13-1-4 60 3 Chống chịu4 C13-1-5 55 3 Chống chịu5 C13-1-6 40 6 Chống chịu6 AS996 Đ/C 10 7 Mân cảm7 IR64-Sub1 50 3 Chống chịu8 IR42 0 9 Mân cảm

11
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Thông qua việc kiểm tra xác định các dòng mang gen chống chịu ngập, chọn ra những dòng có mang gen chống chịu ngập để tiếp tục tiến hành nhân giống và khảo nghiệm sản xuất.
3.4. Kết quả khảo nghiệm sản xuất vụ Đông Xuân 2014 - 2015 và Hè Thu 2015, 2016
Giống lúa OM351 được trồng và khảo nghiệm tại 6 tỉnh thuộc ĐBSCL trong các vụ Đông Xuân
2014 - 2015 và Hè Thu 2015, 2016 để đánh giá về năng suất.
Kết quả khảo nghiệm quốc gia tại 6 điểm khảo nghiệm Đồng Tháp, An Giang, Trà Vinh, Bạc Liêu, Long An, Bến Tre cho thấy, giống OM351 cho năng suất trung bình qua 6 điểm khảo nghiệm tại ĐBSCL đạt 7,16 tấn/ha trong khi giống đối chứng VND95-20 đạt 6,47 tấn/ha. Như vậy, năng suất của giống OM351 cao hơn 10,7% so với đối chứng.
Hình 1. Xác định các dòng mang gen chịu ngập Sub1 bằng chỉ thị phân tử SC3 liên kết gen Sub1 trên các dòng triển vọng từ giống OM351 (AS996-Sub1)
Ghi chú: 1: AS996; 2: IR64 Sub1; 3 - 49: các dòng của giống OM351.
Hình 2. Xác định các dòng mang gen chịu ngập Sub1 bằng chỉ thị phân tử RM23662 liên kết gen Sub1 trên các dòng triển vọng từ giống OM351 (AS996-Sub1)
Ghi chú: L: 25bp ladder; 1: AS996; 2: IR64 Sub1; 3 - 51: các dòng của giống OM351.
Bảng 6. Kết quả khảo nghiệm sản xuất các giống lúa triển vọngvụ Đông Xuân 2014 - 2015, tại 6 tỉnh ĐBSCL
Kết quả khảo nghiệm quốc gia vụ Hè Thu 2015 tại 5 điểm khảo nghiệm Tiền Giang, An Giang, Cờ Đỏ, Thới Lai, Kiên Giang cho thấy, giống OM351 cho năng suất trung bình qua 5 điểm khảo nghiệm tại Đồng bằng sông Cửu Long đạt 4,34 tấn/ha trong khi giống đối chứng OMCS2000 đạt 3,91 tấn/ha. Tại hai tỉnh Đông Nam bộ gồm Bình Thuận, Tây Ninh, giống OM351 cho năng suất 6,05 tấn/ha, trong khi năng suất của giống đối chứng OMCS2000 chỉ đạt 5,25 tấn/ha. Như vậy, năng suất của giống OM351 cao hơn lần lượt là 16,1% và 15,2% so với đối chứng.
Kết quả trình diễn giống triển vọng OM351 tại Bạc Liêu vụ Hè Thu 2016 được thể hiện ở bảng 8 và bảng 9. Kết quả bảng 8 cho thấy: Giống OM351 có thời gian sinh trưởng ngắn (98 ngày); chiều cao cây là 105 cm; chiều dài bông là 22,1 cm; số bông/m2
là 392 bông. Số hạt chắc /bông là 92,4 hạt, tỷ lệ lép 15,5% ,cao hơn IR42 (13,8%) và thấp hơn AS996 (20,5%). Khối lượng 1000 hạt của giống thí nghiệm là 27,0 gam, tương đương giống đối chứng AS996. Năng suất thực tế của giống OM351 là 5,34 tấn/ha, vượt so với đối chứng AS996 là 3,6%.
Tên giống ĐồngTháp
AnGiang
TràVinh
BạcLiêu
Long An
Bến Tre
Trung bình
% so với ĐC
OM351 6,66 8,20 7,12 7,35 6,85 6,80 7,16 10,7
VND95-20 (Đ/c) 6,70 7,00 6,68 6,55 5,89 6,00 6,47 0

12
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Qua kết quả so sánh giống OM351 với giống AS996 đối chứng và IR42 chuẩn nhiễm cho thấy, OM351 là giống có triển vọng, năng suất cao hơn giống nhận gen, ổn định trong các vụ.
Bảng 9. Khả năng kháng sâu bệnh của giống OM351 vụ Hè Thu 2016
Năng suất vụ Đông Xuân 5 - 7 tấn/ha, vụ Hè Thu 4 - 5 tấn/ha tương đương và cao hơn giống lúa IR64 Sub1. Giống có khả năng chống chịu bệnh đạo ôn cấp 1 - 3 và rầy nâu cấp 3, bạc lá, sâu cuốn lá và khô vằn 1 - 3 trong điều kiện tự nhiên ngoài ruộng.
IV. KẾT LUẬN- Sử dụng phương pháp MABC đã chọn tạo được
giống lúa OM351 mang gen chịu ngập Sub1 và gần 100% nền gen của giống nhận gen AS996.
- Đã xác định sự có mặt của gen chịu ngập trong giống OM351 sử dụng chỉ thị phân tử SSR liên kết gen chịu ngập. Giống lúa đạt điểm 3 (60% cây sống) trong điều kiện nhân tạo ngập hoàn toàn 14 ngày.
- Kết quả khảo nghiệm tại các tỉnh thuộc ĐBSCL và miền Đông Nam bộ cho thấy giống lúa OM351 chống chịu sâu bệnh tốt, năng suất trung bình đạt
cao hơn đối chứng VND95-1 và OMCS2000 từ 10,7 đến 16,1%.
- Mô hình trình diễn giống lúa OM351 cho thấy giống cho năng suất cao hơn giống lúa nền gen AS996, khả năng chống chịu sâu bệnh tốt, mang gen chịu ngập, có thể phát triển tiếp trong sản xuất.
TÀI LIỆU THAM KHẢOBộ Nông nghiệp và Phát triển Nông thôn, 2011.
QCVN 01-55:2011/BNNPTNT. Quy chuẩn Quốc gia về khảo nghiệm giá trị canh tác và sử dụng giống lúa.
Bộ Tài nguyên và Môi trường, 2012. Kịch bản biến đổi khí hậu, nước biển dâng cho Việt Nam. Nhà xuất bản Tài nguyên - Môi trường và Bản đồ Việt Nam. Hà Nội, 96 tr.
Phạm Khôi Nguyên, 2009. Diễn đàn “Quan điểm toàn cầu về rừng và biến đổi khí hậu”. Copenhagen, Đan Mạch tháng 12 năm 2009.
Pamplona A., Ella E., Sigh S., Vergara G.V., Ismail A. and Mackill D., 2007. Screening procedures for tolerance of complete Submergence. Sub1 Rice News, Vol.1. No.2, Special Issue December 2007.
Thomson M.J., Marjorie D.O., James E., Rahman M.A., Sajise A.G., Adorada A.L., Raiz E.T., Blumwald E., Seraj Z.I., Singh R.K., Gregorio G.B. & Ismail M.A., 2010. Characterizing the Saltol Quantitative Trait Locus for Salinity Tolerance in Rice. Rice DOI 10.1007/s12284-010-9053-8.
Suprihatno, 1980. Inheritance of submergence tolerance in rice (Oryza sativa L.). PhD. University of the Philippines. LosBanos, Laguna, Philippines.
Bảng 7. Kết quả khảo nghiệm giống lúa OM351 vụ Hè Thu 2015
Bảng 8. Năng suất và các yếu tố cấu thành năng suất vụ Hè Thu 2016 tại Bạc Liêu
Nguồn: Trung tâm Khảo kiểm nghiệm Giống cây trồng và Phân bón Quốc gia vùng Nam bộ năm 2015.
TT Tên giốngĐồng bằng sông Cửu Long Đông Nam bộ
Tiền Giang
An Giang
Cờ Đỏ
Thới Lai
Kiên Giang
Trung bình
Bình Thuận
Tây Ninh
Trung bình
1 OM 351 3,00 3,72 5,20 4,26 5,54 4,34 6,57 5,53 6,05
2 OMCS2000 (Đ/c) 3,19 4,64 4,34 2,27 5,11 3,91 4,97 5,53 5,25
TT Tên giống
TGST (ngày)
Cao cây (cm)
Chiều dài bông
(cm)
Bông/m2
HC/bông
KL.1000 hạt (g) % lép
Năng suất
(T/ha)
% tăng so ĐC
1 OM351 98 105,0 22,1 392 92,4 27,0 15,5 5,34 3,6
2 AS996 105 104,3 21,0 312 88,9 27,0 20,5 5,15
3 IR42 143 92,1 22,0 280 75,2 22,4 13,8 4,10
TTTên
dòng/giống
Đạo ôn
Rầy nâu
Bạc lá
Sâu cuốn
lá
Khô vằn
Chống đổ
1 OM351 1-3 3 1-3 1-3 1-3 3
2 AS996 Đ/c 1-3 3 1-3 1-3 1-3 3
3 IR42 CN 1-3 3 1-3 1-3 1-3 1

13
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Results for breeding and trial production of the OM351 rice variety Doan Thi Huong Giang, Luu Minh Cuc, Le Huy Ham, Pham Xuan HoiAbstractThe study was conducted to introgress the Sub1 gene into AS996 rice variety, which retains the basic characteristics of the variety. After that step, yield and yield components of new rice varieties were tested in some provinces in the Mekong Delta and evaluated submergence tolerance of the rice varieties. Using the MABC method, it was selected the new promising lines having Sub1 gene and almost 100% background of AS996 variety. They were C13, C7, C10 lines. The rice variety OM351 was selected from line C13. The presence of submergence tolerance gene in OM351 was determined by using molecular markers linked to that gene. In the condition of absolute submergence for 14 days, OM 351 was evaluated at level 3 (60% survival plants). The results of the trials in the provinces of the Mekong Delta and the South East showed that the OM351 rice variety was resistant to insect pest and diseases, and the average yield was higher than that of the local check variety VND95-1 and OMCS2000 from 10.7 to 16.1%. The demonstration of the OM351 rice variety showed that the yield was higher than that of the AS996 rice variety, with good resistance to pests, diseases and tolerance to submergence, which could be further developed in production.Keywords: Molecular marker, submergence tolerance gene, rice variety, trial
Ngày nhận bài: 18/9/2018Ngày phản biện: 25/9/2018
Người phản biện: TS. Huỳnh Văn NghiệpNgày duyệt đăng: 15/10/2018
1 Trung tâm Nghiên cứu và Phát triển Cây có củ, Viện Cây lương thực và Cây thực phẩm2 Viện Cây lương thực và Cây thực phẩm
KHẢO NGHIỆM GIỐNG KHOAI TÂY KT4 CHO SẢN XUẤT VỤ ĐÔNG Ở MỘT SỐ TỈNH ĐỒNG BẰNG SÔNG HỒNG
Nguyễn Thị Nhung1, Trịnh Văn Mỵ1, Ngô Thị Huệ1, Nguyễn Mạnh Quy1, Nguyễn Thị Thu Hương1, Đỗ Thị Bích Nga1,
Ngô Doãn Đảm2, Nguyễn Đạt Thoại1, Đỗ Thị Hồng Liễu1
TÓM TẮTKết quả đánh giá và khảo nghiệm giống khoai tây KT4 ở một số tỉnh Đồng bằng sông Hồng từ năm 2014 - 2016
cho thấy giống KT4 đạt năng suất cao (tiềm năng năng suất 25 - 30 tấn/ha); chống chịu được bệnh virus, héo xanh vi khuẩn, mức độ nhiễm bệnh mốc sương nhẹ (điểm 3), bọ trĩ nhẹ (điểm 1). Giống khoai tây KT4 có sức sinh trưởng, phát triển đạt mức tốt (điểm 5), chất lượng tốt. Hàm lượng chất khô đạt 19 - 20%, hàm lượng đường khử đạt 0,42 - 0,51%; hàm lượng tinh bột đạt 16,7%. Củ khoai tây có dạng hình oval, vỏ củ và ruột củ màu vàng.
Từ khóa: Giống khoai tây KT4, năng suất, chất lượng, kháng bệnh
I. ĐẶT VẤN ĐỀTrên thế giới, khoai tây được xếp là cây lương
thực thực phẩm quan trọng thứ ba sau lúa nước và lúa mì. Theo thống kê của tổ chức Nông lương thế giới (FAOSTAT, 2016) diện tích khoai tây trên thế giới là 19,25 triệu ha, năng suất trung bình đạt 19,58 tấn/ha, với tổng sản lượng 376,3 triệu tấn. Trong đó, diện tích khoai tây tại Châu Âu chiếm 29,1% và sản lượng chiếm 31,9%, diện tích Châu Á chiếm 51,9% và sản lượng chiếm 48,8%. Ở Việt Nam diện tích năm 2016 đạt 21,173 ha, năng suất đạt 14,27 tấn/ha và sản lượng đạt 302,229 tấn (FAOSTAT, 2016). Năng suất và sản lượng khoai tây được nâng
cao dần từ những năm 1980 nhờ những chính sách phát triển nông nghiệp của Nhà nước (Trương Văn Hộ, 2010). Diện tích khoai tây vụ Đông ở phía Bắc 2014 và 2015 có xu hướng giảm, năm 2015 đạt 17,10 nghìn ha, giảm gần 3.000 ha so với vụ Đông 2014. Năm 2016, các tỉnh phía Bắc diện tích đạt 18,60 nghìn ha, năng suất đạt 13,24 tấn/ha và sản lượng đạt 246,2 nghìn tấn. Trong đó, vùng Đồng bằng sông Hồng (ĐBSH) có diện tích khoai tây lớn nhất với gần 12.000 ha và ổn định qua các năm (Tổng cục Thống kê, 2016). Tuy nhiên, diện tích và năng suất trồng khoai tây hiện nay của cả nước là không cao do giống trồng không đảm bảo chất lượng và sâu bệnh

14
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Giống Dạng cây Dạng củ Màu vỏ củ Màu ruột củ Độ sâu mắt củ(điểm 1 - 5)
Độ dài tia củ(điểm 1 - 5)
460-1 Nửa đứng Tròn Vàng Vàng 5 3KT4 Nửa đứng Oval Vàng Vàng 3 3
460-18 Nửa đứng Oval Vàng Vàng 5 3466-22 Nửa đứng Tròn dẹt Vàng Vàng 3 3Số 77 Đứng Tròn Vàng Vàng 3 3
Solara (Đ/c) Nửa đứng Oval Vàng Vàng 3 3
gây hại làm giảm năng suất khoai tây đáng kể (Tổng cục Thống kê, 2016). Do đó, giống tốt, có khả năng chống chịu sâu bệnh, cho năng suất và chất lượng cao là một trong những yếu tố quan trọng trong sản xuất khoai tây. Đây là hướng đi đúng và bài bản nhất hiện nay, song cần lượng vốn đầu tư khá lớn. Còn lại chủ yếu phải sử dụng giống nhập từ Trung Quốc mà thực chất đây là khoai tây thương phẩm (Đỗ Kim Chung, 2006). Chính vì vậy, vấn đề giải quyết giống mới, chất lượng củ giống và nhân giống sạch bệnh đủ về lượng đáp ứng cho sản xuất là vấn đề quan trọng trong hệ thống sản xuất khoai tây ở nước ta hiện nay.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưuSáu giống khoai tây khảo nghiệm triển vọng:
460-1; KT4; 466-18; 466-22; Số 77 và giống đối chứng Solara đều cùng đời cấp xác nhận (kích thước củ từ 3-5 cm).
Giống khoai tây KT4 được Trung tâm Nghiên cứu và Phát triển Cây có củ thuộc Viện Cây lương thực và Cây thực phẩm chọn lọc từ tổ hợp lai trong số 36 tổ hợp lai nhập từ CIP (Lima - Peru) năm 2010 mang mã số 302208 (397077.16 ˟ 392639.8).
2.2. Phương pháp nghiên cưu- Thí nghiệm được bố trí theo khối ngâu nhiên
hoàn chỉnh, 3 lần nhắc lại; diện tích ô thí nghiệm: 9 m2 (7,5 m ˟ 1,2 m); luống trồng hàng đôi, mỗi luống trồng 45 củ; mật độ 5 khóm/m2.
- Phân bón: Phân chuồng 15 - 20 tấn + 150 K2O + 150 P2O5 + 150 N. Cách bón: Bón lót: phân chuồng
+ 100% lân + 50% lượng đạm + 50% lượng Kali; Bón thúc: lượng đạm và kali còn lại khi vun lần 1.
- Các chỉ tiêu theo dõi áp dụng theo Quy chuẩn kỹ thuật quốc gia về Khảo nghiệm giá trị canh tác và sử dụng của giống khoai tây QCVN 01-59: 2011/BNNPTNT gồm: Sinh trưởng, phát triển, sâu bệnh hại, năng suất và chất lượng.
- Phương pháp xử lý số liệu bằng Excel và chương trình IRRISTAT 5.0.
2.3. Thời gian và địa điểm nghiên cưuNghiên cứu được thực hiện vào Vụ Đông (từ cuối
tháng 10 đến cuối tháng 1 năm sau) năm 2014, 2015 và năm 2016 tại Bắc Ninh, Nam Định và Hà Nội.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Một số đặc điểm nông sinh hoc của các giống khoai tây
Các giống khoai tây khảo nghiệm đều có dạng cây nửa đứng, riêng giống Số 77 có dạng cây đứng. Dạng củ hình oval có giống KT4 và giống 460-18 tương đương với giống đối chứng Solara. Dạng củ tròn có giống 460-1 và giống Số 77. Còn giống 466-22 có dạng củ hình tròn dẹt và 3 giống cho mắt củ nông đạt điểm 3 là giống KT4; giống 466-22 và Số 77 tương đương so với đối chứng Solara. Độ dài tia củ của các dòng giống ở mức trung bình theo báo cáo kết quả của Nguyễn Thị Nhung và cộng tác viên (2016). Như vậy, cho thấy với đặc điểm hình dạng củ của các giống triển vọng, trong đó giống KT4 đạt được các tiêu chuẩn, hình dạng củ dạng oval, màu vỏ và ruột củ vàng, độ sâu mắt củ nông tương đương với giống đối chứng Solara.
3.2. Sưc sinh trưởng, phát triển của các giống nghiên cưu
Kết quả Bảng 2 cho thấy: Các giống khảo nghiệm tại Hà Nội, Bắc Ninh và Nam Định đều cho sức sinh
trưởng trung bình đạt mức khá đến tốt từ điểm 4,0 - 5,0. Thấp nhất là giống Solara đối chứng đạt trung bình điểm 4, tiếp đến là giống 460 - 18 đạt điểm trung bình 4,3. Còn lại các giống khảo nghiệm khác
Bảng 1. Đặc điểm hình thái của các giống khoai tây
Ghi chú: Độ sâu mắt củ (điểm 1 - 5): 1 - nông; 3 - trung bình; 5 - sâu; Độ dài tia củ (điểm 1 - 5): 1- ngắn; 3: trung bình; 5: dài.

15
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Bảng 2. Đặc điểm sinh trưởng, phát triển của các giống khoai tây
Bảng 3. Số thân chính và động thái tăng chiều cao, thân chính của các giống
Ghi chú: Bảng 2 - 11: BN = Bắc Ninh; NĐ = Nam Định; TT = Thanh Trì; điểm (1 - 5): điểm 1: kém; điểm 3: trung bình; điểm 5: tốt. TB: trung bình 3 năm.
và giống KT4 đạt mức điểm 5. Diện tích tán lá che phủ đất trung bình của các giống khảo nghiệm tại Hà Nội, Nam Định và Thái Bình trong 3 năm 2014 - 2016 đạt tỷ lệ cao từ 92 - 99%, trong đó đạt cao nhất là giống KT4 đạt 99%, tiếp đến là giống 466 - 22 đạt 97%, thấp nhất là giống đối chứng Solara đạt 92%.
Còn lại các giống đạt tương đương nhau 95 - 96%. Như vậy, kết quả sức sinh trưởng, phát triển của các giống KT4, giống 460-1, giống 466-22 và giống số 77 cao hơn giống đối chứng Solara và giống 460-18 ở mức sai số có ý nghĩa LSD0,05 theo báo cáo kết quả của Nguyễn Thị Nhung và cộng tác viên (2016).
GiốngĐặc điểm sinh trưởng (điểm 1 - 5)
TBDiện tích che phủ đất (%)
TBNăm 2014 Năm 2015 Năm 2016 Năm 2014 Năm 2015 Năm 2016HN BN HN NĐ BN NĐ HN BN HN NĐ BN NĐ
460-1 5 5 5 5 5 5 5 100 100 90,0 100 95 95 96,7KT4 5 5 5 5 5 5 5 100 100 100 100 100 95 99,2
460-18 5 5 5 5 3 3 4,3 100 100 90,0 100 95 95 96,7466-22 5 5 5 5 5 5 5 100 100 98,0 100 95 90 97,2Số 77 5 5 5 5 5 5 5 100 100 95,0 95,0 90 90 95,0Solara(đ/c) 5 5 3 5 3 3 4 95,0 100 85,0 95,0 90 90 92,5
CV (%) 13,2 14,7 13,9 13,6 13,8 12,2 12,6 13,3 13,5 12,8 12,6 13,4LSD0,05 0,05 0,09 1,15 0,23 1,05 1,09 1,05 0,07 1,13 1,03 1,35 1,28
Kết quả Bảng 3 cho thấy: Theo báo cáo kết quả của Nguyễn Thị Nhung và cộng tác viên (2016), nhìn chung các giống khảo nghiệm tại Hà Nội, Bắc Ninh và Nam Định trong 3 năm 2014 - 2016 cho thấy số thân chính trung bình đạt từ 2,2 - 3,7 thân. Số thân chính/khóm thấp nhất là giống số 77 đạt 2,2 thân. Cao nhất là giống 460-1 đạt 3,7 thân, tiếp đến là giống KT4 đạt 3,6 thân. Còn lại các giống khảo nghiệm khác đạt tương đương nhau. Chiều cao cây
trung bình của các giống khảo nghiệm đạt từ 59 - 74 cm, giống đối chứng Solara đạt thấp nhất 59,7 cm. Trong đó giống 466 - 22 và giống số 77 đạt cao nhất 74 cm, giống KT4 và các giống khảo nghiệm còn lại đạt tương đương nhau. Như vậy, kết quả sinh trưởng phát triển của giống khoai tây KT4 đạt cao hơn so với giống đối chứng Solara ở mức sai số có ý nghĩa LSD0,05.
GiốngSố thân
TBChiều cao thân chính (cm)
TBNăm 2014 Năm 2015 Năm 2016 Năm 2014 Năm 2015 Năm 2016HN BN HN NĐ BN NĐ HN BN HN NĐ BN NĐ
460-1 3,4 3,4 3,5 3,6 4,1 3,9 3,7 71,6 72,1 73,3 69,9 70,5 68,4 71,0KT4 3,4 3,6 3,5 3,5 3,8 4,0 3,6 67,8 68,6 69,8 66,7 69,7 70,1 68,8
460-18 3,2 3,3 3,7 3,3 3,6 3,7 3,5 69,9 69,5 74,8 70,0 71,5 70,6 71,1466-22 3,0 3,0 3,7 3,4 3,6 3,8 3,4 76,9 75,4 73,6 74,8 72,7 73,1 74,4Số 77 1,3 1,7 3,5 1,7 2,7 2,5 2,2 75,0 77,0 72,4 76,1 71,2 72,5 74,0Solara (Đ/c) 2,7 3,0 3,2 3,1 4,1 3,5 3,3 60,3 61,5 52,3 65,4 53,5 64,9 59,7
CV (%) 12,4 11,3 13,2 12,5 12,1 11,6 11,9 12,7 13,0 12,5 12,8 12,6LSD0,05 1,09 1,02 1,22 1,13 0,04 0,08 1,34 2,23 2,45 0,03 1,28 0,36
16
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
3.3. Mưc độ nhiễm bệnh hại chính của các giống khảo nghiệm
Kết quả Bảng 4 cho thấy: mức độ nhiễm bệnh mốc sương nhẹ (điểm 3) ở tất cả các giống cùng khảo nghiệm tại các điểm khảo nghiệm Hà Nội, Nam Định và Bắc Ninh trong 3 năm 2014 - 2016. Bệnh virus nhiễm trung bình nhẹ từ 0,2 - 0,6%, trong đó giống đối chứng Solara nhiễm trung bình trong 3 năm 2014 - 2016 tại 3 điểm khảo nghiệm Hà Nội, Nam Định và Bắc Ninh. Trong đó, năm 2014
tại Bắc Ninh giống 460-1 nhiễm 1,6%, tại Hà Nội nhiễm 0,7%. Giống 466-22 cả hai điểm Bắc Ninh và Nam Định nhiễm 0,7%, giống đối chứng Solara tại điểm Hà Nội nhiễm 2,2%. Năm 2015 các giống khảo nghiệm không thấy nhiễm bệnh virus, riêng giống đối chứng Solara nhiễm 1,6% tại điểm Hà Nội. Năm 2016 tại cả hai điểm Nam Định và Bắc Ninh không thấy giống nào nhiễm bệnh virus (Nguyễn Thị Nhung và ctv., 2017).
Kết quả bảng 5 cho thấy: Mức độ nhiễm bệnh héo xanh vi khuẩn trung bình của các giống khảo nghiệm tại 3 điểm Bắc Ninh, Hà Nội và Nam Định trong 3 năm 2014 - 2016 cho thấy mức độ nhiễm nhẹ từ 0,2 - 0,9%. Trong đó có 3 giống 460-1, KT4 và giống Số 77 không bị nhiễm bệnh héo xanh, giống đối chứng Solara nhiễm 0,9% và hai giống 460 - 18 và giống 466 - 22 nhiễm 0,2%. Sâu hại chính rệp, nhện và bọ trĩ trong các năm 2014 - 2016 tại 3 điểm khảo
nghiệm Nam Định, Hà Nội và Bắc Ninh đều thấy xuất hiện nhưng mức độ nhiễm nhẹ (điểm 1), theo Nguyễn Thị Nhung và cộng tác viên, 2017. Như vậy, kết quả các giống khoai tây triển vọng khảo nghiệm tại các tỉnh Hà Nội, Bắc Ninh và Nam Định, trong 3 năm 2014 - 2016 nhiễm sâu bệnh hại ở mức không đến nhiễm nhẹ, ít gây ảnh hưởng đến sinh trưởng, phát triển của cây. Như vậy, giống KT4 và giống Số 77 là hai giống nhiễm nhẹ bệnh mốc sương.
Bảng 4. Mức độ nhiễm bệnh hại chính của các giống khảo nghiệm
Bảng 5. Mức độ nhiễm bệnh và sâu hại chính của các giống khảo nghiệm
Ghi chú: Điểm 1: không xuất hiện triệu chứng nhiễm bệnh; điểm 3: nhẹ, < 20% diện tích thân lá nhiễm bệnh; điểm 5: trung bình, 20 - 50% diện tích thân lá nhiễm bệnh; điểm: nặng, > 50 - 75% diện tich thân lá nhiễm bệnh; điểm 9: rất nặng, > 75 - 100% diện tich thân lá nhiễm bệnh.
Ghi chú: Điểm 0: không bị hại; điểm 1: bị hại nhẹ; điểm 3: một số cây có lá bị hại hại; điểm 5: tất cả các cây có lá bị hại, cây sinh trưởng chậm; điểm 7: trên 50% số cây bị chết, số cây còn lại ngừng sinh trưởng; điểm 9: tất cả các cây bị chết.
GiốngBệnh mốc sương (điểm 1 - 9)
TBBệnh virus (%)
TBNăm 2014 Năm 2015 Năm 2016 Năm 2014 Năm 2015 Năm 2016HN BN HN NĐ BN NĐ HN BN HN NĐ BN NĐ
460-1 3 3 3 3 3 3 3 0,7 1,6 0,0 0,0 0,0 0,0 0,4KT4 3 3 3 3 3 3 3 0,0 0,0 0,0 0,0 0,0 0,0 0,0
460-18 3 3 3 3 3 3 3 0,0 0,0 0,0 0,0 0,0 0,0 0,0466-22 3 3 3 3 3 3 3 0,7 0,7 0,0 0,0 0,0 0,0 0,2Số 77 3 3 3 3 3 3 3 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Solara (Đ/c) 3 3 3 3 3 3 3 2,2 0,0 1,6 0,0 0,0 0,0 0,6CV (%) 10,6 11,5 10,2 11,4 10,3 10,1 11,3 10,9 12,1 10,7 11,0 12,1LSD0,05 0,06 0,09 0,10 0,13 0,05 0,07 1,15 1,09 1,17 1,13 1,24 1,22
GiốngBệnh héo xanh vi khuẩn (%)
TBBo trĩ (điểm 0 - 9)
TBNăm 2014 Năm 2015 Năm 2016 Năm 2014 Năm 2015 Năm 2016HN BN HN NĐ BN NĐ HN BN HN NĐ BN NĐ
460-1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 1 1 1 1 1 1 1KT4 0,0 0,0 0,0 0,0 0,0 0,0 0,0 1 1 1 1 1 1 1
460-18 0,0 0,0 0,0 0,0 0,7 0,7 0,2 1 1 1 1 1 1 1466-22 0,0 0,0 0,0 0,0 0,7 0,7 0,2 1 1 1 1 1 1 1Số 77 0,0 0,0 0,0 0,0 0,0 0,0 0,0 1 1 1 1 1 1 1
Solara (Đ/c) 0,7 0,0 0,0 0,0 2,2 2,2 0,9 1 1 1 1 1 1 1CV (%) 11,2 10,5 10,6 10,0 10,3 10,1 11,3 11,9 11,5 11,0 10,6 11,3LSD0,05 1,06 0,09 0,10 0,12 1,02 1,03 0,05 0,06 0,13 0,10 0,14 0,04

17
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Bảng 7. Khối lượng củ trung bình của các giống khoai tây khảo nghiệm
Bảng 8. Năng suất thực thu của các giống khoai tây khảo nghiệm
3.4. Năng suất và các yếu tố cấu thành năng suất của các giống khoai tây
Kết quả Bảng 6 cho thấy: Số củ trung bình/khóm của các giống khảo nghiệm tại Hà Nội, Nam Định và Bắc Ninh trong 3 năm 2014 - 2016 đạt từ 5,7-8,7 củ/khóm, trong đó giống cho số củ trung bình đạt cao nhất là giống 460-1 đạt 8,7 củ, tiếp đến là giống KT4 đạt 8,3 củ cao hơn so với giống đối chứng Solara đạt 6,4 củ và giống số 77 đạt 5,7 củ ở mức sai số có ý nghĩa LSD0,05. Kết quả này tương đương với kết quả của Nguyễn Thị Nhung và cộng tác viên (2017).
Kết quả Bảng 7 cho thấy: Tương tự khối lượng củ/khóm của các giống khoai tây khảo nghiệm tại Hà Nội, Bắc Ninh và Nam Định trong 3 năm 2014 - 2016 đạt trung bình từ 489,5 - 590,8 gam. Cao nhất là giống KT4 đạt 590,8 gam, tiếp đến là giống số 77 đạt 544,6 gam và thấp nhất là giống đối chứng Solara đạt 489,5 gam, tiếp đến là giống số 460 - 22 đạt 531,4 gam. Như vậy, kết quả các yếu tố cấu thành năng suất của các giống khoai tây khảo nghiệm tại Hà Nội, Bắc Ninh và Nam Định trong 3 năm 2014 - 2016,
giống khoai tây KT4 đạt cao nhất, tiếp đến là giống số 77, giống 460-1 cao hơn so với giống đối chứng Solara ở mức sai số có ý nghĩa LSD0,05. Kết quả này tương đương với kết quả của Nguyễn Thị Nhung và cộng tác viên (2017).
Bảng 6. Số củ trung bình của các giống khoai tây khảo nghiệm
GiốngKhối lượng củ/khóm (gam)
Trung bìnhNăm 2014 Năm 2015 Năm 2016HN BN HN NĐ BN NĐ
460-1 564,2 577,0 555,6 545,6 502,6 493,4 539,7KT4 587,8 604,0 581,0 606,2 581,2 584,4 590,8
460-18 553,8 576,2 535,2 557,4 497,0 483,8 533,9466-22 548,8 553,0 528,6 558,6 497,8 501,8 531,4Số 77 536,2 562,6 505,0 573,4 539,6 550,8 544,6
Solara (Đ/c) 495,2 515,8 487,6 516,4 468,8 453,0 489,5CV (%) 13,5 14,0 13,2 13,5 13,6 13,4LSD0,05 1,12 1,15 2,01 1,13 2,10 2,05
GiốngSố củ/khóm (củ)
Trung bìnhNăm 2014 Năm 2015 Năm 2016
HN BN HN NĐ NĐ BN460-1 10,2 9,5 7,0 6,4 12,1 7,0 8,7KT4 7,9 7,6 7,3 7,0 10,9 9,0 8,3
460-18 11,2 10,1 6,8 6,6 5,3 5,0 7,5466-22 8,3 7,7 5,7 6,3 6,2 6,4 6,8Số 77 6,2 5,9 4,7 4,5 6,2 6,5 5,7Solara (Đ/c) 6,6 6,3 6,7 6,2 5,9 6,4 6,4
CV (%) 16,2 16,3 15,5 14,7 13,9 14,8LSD0,05 2,08 2,10 2,14 2,15 2,03 2,11
GiốngNăng suất thực thu (tấn/ha) Năng suất
trung bình (tấn/ha)
Năm 2014 Năm 2015 Năm 2016Hà Nội Bắc Ninh Hà Nội Nam Định Bắc Ninh Nam Định
460-1 28,21 28,85 27,78 27,28 25,13 24,67 26,99KT4 29,39 30,20 29,05 30,31 29,06 29,22 29,54
460-18 27,69 28,81 26,76 27,87 24,85 24,19 26,70466-22 27,44 27,65 26,43 27,93 24,89 25,09 26,57Số 77 26,81 28,13 25,25 28,67 26,98 27,54 27,23
Solara (Đ/c) 24,76 25,79 24,38 25,82 23,44 22,65 24,47CV (%) 13,2 12,7 13,6 12,9 12,5 13,1LSD0,05 1,05 1,09 1,19 1,23 1,20 1,26

18
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Kết quả Bảng 8 cho thấy: Năng suất trung bình của các giống khoai tây khảo nghiệm tại Hà Nội, Bắc Ninh và Nam Định trong 3 năm 2014 - 2016 đạt từ 24,47 - 29,54 tấn/ha, trong đó giống khoai tây KT4 đạt năng suất trung bình cao nhất 29,54 tấn/ha, tiếp đến là giống Số 77 đạt 27,23 tấn/ha, giống 460-1 đạt 26,99 tấn/ha. Đạt năng suất trung bình thấp nhất là giống đối chứng Solara đạt 24,47 tấn/ha, tiếp đến là giống 466-22 đạt 26,57 tấn/ha. Như vậy, giống KT4 năng suất năm 2014 tại Hà Nội đạt 29,39 tấn/ha, giống Solara đạt 24,76 tấn/ha tăng 18,7% . Tại Bắc Ninh đạt 30,20 tấn/ha, giống Solara đạt 25,79 tấn/ha tăng 17,1%. Năm 2015 tại Hà Nội đạt 26,05 tấn/ha, giống Solara đạt 24,38 tấn/ha tăng 16,2%. Tại Nam Định đạt 30,31 tấn/ha, giống Solara đạt 25,82 tấn/ha tăng 17,4%. Năm 2016 tại Bắc Ninh đạt 29,06 tấn/ha, giống Solara đạt 23,44 tấn/ha tăng 23,9%. Tại Nam Định đạt 29,22 tấn/ha, giống Solara đạt 22,65 tấn/ha
tăng 29,0% ở mức sai số có ý nghĩa LSD0,05. Kết quả này tương đương với Nguyễn Thị Nhung và cộng tác viên (2017).
3.5. Một số chỉ tiêu phân tích chất lượng của các giống khảo nghiệm
Kết quả Bảng 9 cho thấy: Tỷ lệ chất khô trung bình của các giống khảo nghiệm được phân tích liền trong 3 năm 2014 - 2016 đạt từ 18 - 20%. Tỷ lệ hàm lượng đường khử trung bình của các giống đạt từ 0,36 - 0,56%. Hàm lượng tinh bột của các giống phân tích cho kết quả trung bình đạt từ 15,3 - 18,1% đạt tương đương nhau. Như vậy, kết quả phân tích chất lượng có giống KT4, giống 460-1, giống số 77 đều đạt tiêu chuẩn chất lượng giống phục vụ nhu cầu cho ăn tươi tương đương giống đối chứng Solara như: hàm lượng chất khô từ 16 - 18%, hàm lượng tinh bột 14 - 16%, đường khử cao > 0,5% (Nguyễn Thị Nhung và ctv., 2016).
Bảng 9. Kết quả phân tích một số chỉ tiêu chất lượng củ của các giống khoai tây
Ghi chú: *, **, ** : Kết quả phân tích do Bộ môn Sinh lý sinh hóa chất lượng nông sản - Viện Cây lương thực và Cây thực phẩm phân tích.
Giống*Chất khô (%)
TB**Hàm lượng đường
khử (% củ tươi) TB***Hàm lượng tinh bột
( % củ tươi) TB2014 2015 2016 2014 2015 2016 2014 2015 2016
460-1 20,2 20,1 19,6 20,0 0,50 0,52 0,67 0,56 16,6 16,8 18,4 17,3KT4 20,7 20,3 19,6 20,2 0,42 0,46 0,51 0,46 16,7 16,6 16,8 16,7
460-18 19,6 19,5 19,6 0,35 0,37 0,36 18,2 18,0 18,1466-22 19,6 19,0 19,3 0,43 0,45 0,44 15,2 15,3 15,3Số 77 20,9 20,5 21,4 20,9 0,52 0,53 0,42 0,49 15,9 15,6 16,6 16,0Solara 18,7 18,5 19,1 18,8 0,54 0,51 0,58 0,54 16,2 16,5 16,5 16,4
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luậnGiống khoai tây KT4 sinh trưởng, phát triển tốt,
thời gian sinh trưởng khoảng 85 - 90 ngày, dạng cây nửa đứng, mức độ nhiễm mốc sương nhẹ, không nhiễm bệnh virus và bệnh héo xanh vi khuẩn. Dạng củ hình oval, mắt củ nông, vỏ củ và ruột củ màu vàng, số lượng củ/khóm đạt 6 - 9 củ. Năng suất thực thu cao và ổn định qua các năm đạt 29 - 30 tấn/ha tăng từ 16,2 - 29,0% so với giống đối chứng Solara đạt 22 - 25 tấn/ha. Giống có chất lượng tốt, tỷ lệ chất khô trung bình đạt 20,2%, hàm lượng đường khử đạt 0,46% và tỷ lệ tinh bột đạt 16,7%.
4.2. Đề nghịTiếp tục mở rộng mô hình giống KT4 ở các vùng
sinh thái khác nhau để phục vụ sản xuất đại trà.
TÀI LIỆU THAM KHẢO Bộ Nông nghiệp và PTNT, 2011. QCVN
01-59:2011-BNNPTNT. Quy chuẩn kỹ thuật quốc gia về khảo nghiệm giá trị canh tác và giá trị sử dụng của giống khoai tây.
Đỗ Kim Chung, 2006. Thị trường khoai tây ở Việt Nam. Dự án thúc đẩy sản xuất khoai tây ở Việt Nam. Nhà xuất bản Thanh Hóa.
Tổng cục Thống kê, 2016. Niên giám thống kê. Nhà xuất bản Thống kê.
Trương Văn Hộ, 2010. Cây khoai tây ở Việt Nam. NXB Nông nghiệp. Hà Nội.
Nguyễn Thị Nhung, Trịnh Văn Mỵ, Đỗ Thị Bích Nga, Nguyễn Thị Thu Hương, Ngô Thị Huệ, Nguyễn Mạnh Quy, Nguyễn Thiên Lương, 2017. Kết quả đánh giá và chọn lọc giống khoai tây 12KT3-1. Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam, số 9 (82)/2017.

19
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Nguyễn Thị Nhung, Hoàng Thị Giang, Nguyễn Quang Thạch, Trịnh Văn Mỵ, Ngô Thị Huệ, Nguyễn Mạnh Quy, Nguyễn Thị Thu Hương, Vũ Thị Hằng, Đỗ Thị Thu Hà, Nguyễn Đưc Mạnh, 2017. Kết quả đánh giá khả năng kháng bệnh virus và mốc sương của các dòng/giống khoai tây bằng phương pháp lây nhiễm nhân tạo. Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam, số 3 (76)/2017.
Nguyễn Thị Nhung, Nguyễn Thế Nhuận, Nguyễn Quang Thạch, Trịnh Văn Mỵ, Ngô Thị Huệ, Hoàng
Thị Giang, Nguyễn Mạnh Quy, Nguyễn Thị Thu Hương, Đinh Thị Hồng Nhung, Vũ Thị Hằng, Đỗ Thị Thu Hà, Nguyễn Đưc Mạnh, 2016. Nghiên cứu chọn tạo giống khoai tây phục vụ chế biến và ăn tươi đạt năng suất cao. Báo cáo đề tài nghiên cứu khoa học cấp Bộ, trang 31-37.
FAO, 2016. Food and Agriculture Organization of the United Nations. The state of food and agriculture, Food and Agriculture Organization of the United Nations, 194pp.
Testing of potato variety KT4 for production in some provinces of the Red River Delta
Nguyen Thi Nhung, Trinh Van My, Ngo Thi Hue, Nguyen Thi Thu Huong, Nguyen Manh Quy, Do Thi Bich Nga,
Ngo Doan Dam, Nguyen Dat Thoai, Do Thi Hong LieuAbstractThe evaluation and testing of potato variety KT4 were conducted during the period of 2014 to 2016 in some provinces of the Red River Delta showed that KT4 variety produced significantly high yield (the potential yield reached 25 to 30 tons/ha) and growth vigor (score 5); high resistance to virus and bacterial wilt disease; moderate resistance to Phytophthora blight (score 3); markedly low infection of Stenchaetothrips biformis (score 1). The dry matter content of tubers was 19 - 20%. The sugar content was considerably low in tuber of this variety (0.42 - 0.51%) and starch content was around 16.7%. The tubers appearance was oval with yellow skin and golden flesh.Keywords: Potato variety KT4, yield, quality, resistance
Ngày nhận bài: 5/8/2018Ngày phản biện: 11/8/2018
Người phản biện: TS. Vũ Thanh HảiNgày duyệt đăng: 18/9/2018
1 Viện Nghiên cứu Rau quả
KẾT QUẢ CHỌN TẠO GIỐNG HOA LAN HỒ ĐIỆP LAI HĐ01Đinh Thị Dinh1, Nguyễn Thị Kim Oanh1, Nguyễn Văn Tiến1
TÓM TẮTKết quả khảo nghiệm sản xuất giống lan Hồ điệp lai HĐ01 ở các địa phương, bao gồm Hà Nội, Bắc Ninh, Quảng
Ninh cho thấy: Dòng lai HĐ01 sinh trưởng, phát triển tốt ở tất cả các điểm trồng. Tỷ lệ ra ngồng và ra hoa đều đạt trên 90%. Chất lượng hoa cao; chiều dài cành hoa đạt 73,8 - 74,5 cm, số hoa 8,5 - 8,6 hoa/cành. Mức độ sâu, bệnh hại thấp (dưới 5%). Hiệu quả kinh tế cao gấp 1,28 - 1,30 lần so với đối chứng. Giống lan Hồ điệp lai HĐ01 được người tiêu dùng và người sản xuất ưa chuộng.
Từ khóa: Chất lượng, giống lai, Hồ điệp HĐ01, khảo nghiệm, phát triển, sản xuất, sinh trưởng
I. ĐẶT VẤN ĐỀ Lan Hồ điệp (Phalaenopsis) là loại hoa đẹp và
sang trọng được ưa chuộng nhất hiện nay không chỉ ở Việt Nam mà còn trên toàn thế giới (Hwang Seung Jae and Jeong Byoung Ryong, 2009). Tuy nhiên, các giống hoa lan Hồ điệp hiện nay vân chủ yếu là các giống nhập nội, chưa chủ động được bộ giống cho sản xuất. Chính vì vậy, việc lai tạo giống mới có hoa đẹp, chất lượng cao là rất cần thiết, trước hết là đáp ứng nhu cầu thị trường trong nước và hướng tới xuất khẩu.
Viện Nghiên cứu Rau quả trong những năm gần đây đã chú trọng cho công tác chọn tạo giống Hồ điệp trong nước và đã có những kết quả nhất định (Đặng Văn Đông và ctv., 2009). Cụ thể, Viện đã tạo ra được nhiều dòng lai lan Hồ điệp có sức sinh trưởng, phát triển tốt, màu sắc hoa đẹp, khác biệt trong đó có dòng lai lan Hồ điệp HĐ01.
Nghiên cứu được thực hiện với mục tiêu chọn lọc và khảo nghiệm được dòng lai lan Hồ điệp có đặc tính tốt, sinh trưởng khỏe, tỷ lệ ra hoa cao (> 90%), chất lượng hoa tốt (8 hoa/cành trở lên,

20
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
chiều dài cành hoa > 70 cm, độ bền hoa > 30 ngày) và có khả năng thích nghi cao trong điều kiện Việt Nam.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưu- Dòng lai lan Hồ điệp HĐ01 (hoa màu phấn
hồng, môi đỏ), là con lai của tổ hợp lai giữa : LVR4 (hoa đỏ mười giờ) ˟ : LVR2 (hoa trắng môi vàng). Giống đã được lai tạo từ năm 2011, qua quá trình đánh giá, chọn lọc và được khảo nghiệm cơ bản tại Gia Lâm, Hà Nội từ tháng 2/2015 đến tháng 2/2017.
- Đối chứng là giống HL3, giống đã được Bộ Nông nghiệp và PTNT công nhận là giống cho sản xuất thử năm 2010. Cây giống là cây nuôi cấy mô sau ra ngôi 12 tháng tuổi cây có 4 - 5 lá, chiều dài lá khoảng 16 - 18 cm, chiều rộng lá khoảng 7 - 9 cm.
- Điều kiện trồng:+ Giá thể rêu ẩm: Rêu khô được ngâm nước và
vắt khô, định lượng 1 kg/40 bầu.+ Chậu trồng: Kích thước: bầu nhỡ (8,3 ˟ 8,3 cm),
sang bầu to (12 ˟ 12 cm), màu trắng, khay để cây tương ứng với loại bầu.
+ Phân bón: Giai đoạn sinh trưởng (tháng 2 - tháng 6) bón phân Plant soul 3: 20% đạm (N) - 20% lân (P2O5) - 20% kali (K2O) - vi lượng. Giai đoạn trước xử lý ra hoa và khi xử lý ra hoa bón phân Plant soul 1: 9% đạm (N) - 45% lân (P2O5) - 15% kali (K2O) - vi lượng. Giai đoạn chăm sóc hoa (sau khi xuất hiện mầm hoa 7 cm), bón phân Plant soul 2: 10% đạm (N) - 20% lân (P2O5) - 20% kali (K2O) - vi lượng. Bón nồng độ 1%, 7 ngày/lần.
+ Thuốc BVTV: Phòng trừ bệnh thối nhũn vi khuẩn (Erwinia sp.) áp dụng phòng trừ tổng hợp IPM, phun phòng bệnh bằng thuốc Steptomicine + Ridomil 68WP 1 tháng 1 lần. Ngừng tưới khi xử lý thuốc bệnh 1 ngày. Phun phòng trừ một số sâu hại, nhện hại bằng một số loại thuốc: Aramite 15%, 15 g/10 lít nước, Kelthane 2% 15 g/10 lít nước, Brightin 10 ml/10 lít nước. Phun 1 tháng 1 lần, phun luân phiên các loại thuốc.
+ Cây được trồng trong nhà lưới hiện đại có mái che mưa, các chế độ nhiệt độ được điều chỉnh tự động thông qua hệ thống thiết bị gồm: Bảng điều khiển tự động, hệ thống lưới đen và bảo ôn 3 lớp, quạt hút, thông gió, tấm liền nước. Nhiệt độ, ánh sáng có thể điều khiển theo từng giai đoạn sinh trưởng, phát triển của cây giai đoạn cây trưởng thành 20 - 30oC. Giai đoạn xử lý ra hoa được đưa
lên vùng núi cao Mộc Châu, Sơn La từ ngày 15/7 - 15/9/2017 cây được đặt trong điều kiện nhà lưới đơn giản, vùng có điều kiện khí hậu ban đêm khoảng 15 - 17 oC, ban ngày khoảng 25 - 27oC, chênh lệch nhiệt độ ngày đêm khoảng 8 - 10oC.
2.2. Phương pháp nghiên cưu
2.2.1. Bố trí thí nghiệm Mỗi địa điểm bố trí thí nghiệm theo kiểu tuần tự
không nhắc lại. Các yếu tố phi thí nghiệm đồng nhất trên mỗi thí nghiệm.
Bảng 1. Quy mô khảo nghiệm sản xuất ở một số địa phương
2.2.2. Quy trình chăm sócCác kỹ thuật chăm sóc áp dụng theo: “Quy trình
sản xuất hoa lan Hồ điệp quy mô công nghiệp tại các tỉnh phía Bắc” của Viện Nghiên cứu Rau quả đã được Bộ Nông nghiệp và PTNT công nhận năm 2010 (Nguyễn Văn Tỉnh và ctv., 2010).
2.2.3. Các chỉ tiêu theo dõi Các chỉ tiêu theo dõi chính được tham khảo theo
tài liệu khảo nghiệm DUS về hoa lan Hồ điệp (Chen Heming et al., 2014). Tỷ lệ sống (%), số lá/cây (lá), chiều dài lá (cm), chiều rộng lá (cm) , tỷ lệ xuất hiện cành hoa (%), chiều dài cành hoa (cm), đường kính cành hoa (cm), số nụ hoa/cành, tỷ lệ hoa nở (%), đường kính hoa (cm), độ bền của cành hoa (ngày).
Mức độ nhiễm sâu bệnh hại: Phương pháp điều tra sâu bệnh hại theo Quy chuẩn Quốc gia về phương pháp điều tra phát hiện dịch hại cây trồng (QCVN 01-38: 2010/BNN PTNT).
2.2.4. Phương pháp xử lý số liệu Số liệu được xử lý bằng chương trình Excel và
IRRISTAT 5.0.
2.3. Thời gian và địa điểm nghiên cưuThí nghiệm được thực hiện tại 3 địa điểm: Viện
Nghiên cứu Rau quả - huyện Gia Lâm, Hà Nội; phường Đông Ngàn, thị xã Từ Sơn, Bắc Ninh và Khu 5, thị trấn Trới, huyện Hoành Bồ, Quảng Ninh trong thời gian từ tháng 2/2017 đến tháng 2/2018.
Địa điểm Số lượng (cây)
Quy mô (m2) Điều kiện trồng
Hà Nội 1.000 30 Nhà lưới hiện đạiBắc Ninh 1.000 30 Nhà lưới hiện đạiQuảng Ninh 1.000 30 Nhà lưới hiện đạiTổng cộng 3.000 90

21
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Khả năng sinh trưởng của giống lan hồ điệp HĐ01
Để thấy được khả năng thích nghi của dòng lai lan Hồ điệp HĐ01 ở các điều kiện sinh thái khác nhau, năm 2017 tiến hành đưa dòng lai lan Hồ điệp HĐ01 khảo nghiệm trên diện rộng tại một số địa
phương khác nhau của Việt Nam. Kết quả về khả năng sinh trưởng được trình bày ở bảng 2.
Kết quả khảo nghiệm ở cả 3 địa điểm cho thấy, dòng lan Hồ điệp HĐ01 có khả năng thích nghi tốt với đều kiện khí hậu miền Bắc Việt Nam. Tỷ lệ sống cao (> 90%), trong đó dòng lai HĐ01 có tỷ lệ sống dao động trong khoảng 92,6% - 93,5% cao hơn so với giống HL3 (91,9% - 92,2%).
Giai đoạn cây sau ra ngôi 12 tháng tuổi dòng lai HĐ01 có khả năng sinh trưởng phát triển tốt hơn so với giống HL3 ở cả 3 điểm khảo nghiệm. Số lá đạt 4,5 - 4,7 lá, dài lá là 16,9 - 17,2 cm, rộng lá 7,7 - 7,8 cm. Sang đến giai đoạn cây 18 tháng tuổi số lá và kích thước lá trên dòng HĐ01 và giống đối chứng HL3 cũng biến động tương tự giai đoạn 12 tháng tuổi, giai đoạn này cây mang đặc trưng riêng của giống, kết quả này phù hợp với kết quả của những nghiên cứu trước đó là lá có hình dạng thuôn dài và cây có số lượng lá, kích thước lá đã đạt gần như tối đa, cây chuẩn bị chuyển sang thời kỳ ra hoa.
Để có được hoa lan Hồ điệp vào đúng dịp mong muốn, tiến hành đưa lan Hồ điệp đi xử lý ra hoa trong điều kiện tự nhiên tại tại Mộc Châu, Sơn La, khi cây ra mầm hoa cao 7 cm , thì đưa trở lại các địa
điểm nghiên cứu để tiếp tục đánh giá đến khi hoa nở và kết quả theo dõi thu được trình bày ở bảng 3.
3.2. Thời gian sinh trưởng, phát triển của giống lan hồ điệp lai HĐ01
Qua kết quả bảng 3 cho thấy: Thời gian từ khi xử lý đến khi xuất hiện cành hoa của các dòng, giống có sự khác nhau khi trồng ở các địa phương. Dòng lai HĐ01 xuất hiện sớm hơn đối chứng, chỉ sau 21 - 22 ngày đã có 50% số cây xuất hiện cành hoa, trong khi giống đối chứng phải mất từ 22 - 24 ngày.
Thời gian phát triển cành hoa và ra nụ của dòng HĐ01 đạt 103 - 109 ngày, thời gian từ khi xuất hiện cành hoa đến khi 3 bông hoa trên cành nở đạt 119 - 124 ngày ở các địa phương, tương đương với giống đối chứng.
Bảng 2. Đặc điểm sinh trưởng của giống lan Hồ điệp HĐ01 trồng tại các địa phương
Ghi chú: Xử lý số liệu: ANOVA tiêu chuẩn t của phân phối Student, α = 0,05. Các chữ cái giống nhau trong cùng một cột thể hiện không có sự sai khác giữa các công thức, các chữ cái khác nhau trong cùng một cột thể hiện sự sai khác giữa các công thức.
Địa điểm Dòng/ giống Tỷ lệ sống (%)
12 tháng tuổi 18 tháng tuổi
Số lá Dài lá (cm)
Rộng lá (cm) Số lá Dài lá
(cm)Rộng lá
(cm)
Bắc Ninh
HL3 (ĐC) 91,9 4,5b 14,4b 6,5b 5,9b 16,6b 7,2b
HĐ01 92,6 4,7a 16,9a 7,8a 6,2a 20,1a 8,5a
Tlt 2,05 2,05 2,05 2,05 2,05 2,05
T tính 2,55 4,30 6,04 2,16 8,49 6,81
Hà Nội
HL3 (ĐC) 92,1 4,4b 14,6b 6,6b 5,5b 16,5b 7,5b
HĐ01 93,5 4,5a 17,2a 7,7a 6,4a 21,0a 8,3a
Tlt 2,05 2,05 2,05 2,05 2,05 2,05
T tính 2,26 7,95 5,59 5,06 8,63 5,57
Quảng Ninh
HL3 (ĐC) 92,2 4,5b 14,7b 6,3b 5,7b 16,3b 7,4b
HĐ01 93,1 4,6a 17,0a 7,7a 6,5a 20,6a 8,4a
Tlt 2,05 2,05 2,05 2,05 2,05 2,05
T tính 2,18 8,12 6,01 4,25 8,96 6,33

22
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
3.3. Chất lượng hoa lan hồ điệp HĐ01Một trong những định hướng chính của nhà tạo
giống hoa nói chung và tạo giống hoa lan hồ điệp nói riêng là tạo ra những giống có khả năng sinh trưởng
tốt, năng suất và chất lượng hoa cao. Chất lượng hoa lan Hồ điệp được đánh giá thông qua các chỉ tiêu như: Chiều dài ngồng hoa, số hoa, tỷ lệ hoa nở và độ bền hoa, kết quả được trình bày ở bảng 4.
Kết quả cho thấy ở cả 3 điểm trồng các chỉ tiêu về tỷ lệ xuất hiện cành hoa, số hoa/cành, chiều dài cành hoa, tỷ lệ hoa nở và độ bền hoa tự nhiên của dòng lai HĐ01 đều đạt cao ở các điểm trồng. Cụ thể, tỷ lệ xuất hiện cành hoa của dòng lai HĐ01 đạt 91,1% - 92,0%, chiều dài cành hoa đạt 73,8 - 74,5 cm cao hơn rõ rệt so với đối chứng đạt 60,4 - 62,1 cm.
Số hoa/cành của dòng lai HĐ01 (8,5 - 8,6 hoa) cao vượt trội so với đối chứng là 8,1 - 8,2 hoa. Tỷ lệ
hoa nở và độ bền hoa nở cũng đạt cao hơn HL3 với tỷ lệ hoa nở 94,3 - 95,9%, độ bền hoa 34 - 36 ngày ở các điểm trồng.
Như vậy, đánh giá về chất lượng hoa của dòng HĐ01 trồng ở các địa phương đều đạt kết quả cao hơn so với đối chứng, điều đó chứng tỏ dòng lai HĐ01 có đặc điểm về tỷ lệ ra hoa và chất lượng hoa đều rất ổn định vân giữ nguyên được đặc tính của chúng như khi khảo nghiệm ban đầu.
Bảng 3. Thời gian qua các giai đoạn phát triển hoa của dòng lai lan Hồ điệp HĐ01 khi trồng ở các địa phương
Bảng 4. Tỷ lệ ra hoa và chất lượng hoa của dòng lai lan Hồ điệp HĐ01 trồng ở các địa phương (cây 22 tháng tuổi)
Ghi chú: Thời gian theo dõi: 9 - 12/2017.
Ghi chú: Thời gian theo dõi: 12/2017 - 2/2018; Xử lý số liệu: ANOVA tiêu chuẩn t của phân phối Student, α = 0,05. Các chữ cái giống nhau trong cùng một cột thể hiện không có sự sai khác giữa các công thức, các chữ cái khác nhau trong cùng một cột thể hiện sự sai khác giữa các công thức.
Địa điểm Dòng/giốngTG từ khi xử lý đến
50% số cây xuất hiện cành hoa (ngày)
TG xuất hiện cành hoa đến 10% số cây nở bông
hoa đầu tiên (ngày)
Tổng TG xuất hiện mầm hoa đến 3 bông
hoa nở (ngày)
Bắc NinhHL3 (ĐC) 24 105 120HĐ01 22 109 124
Hà NộiHL3 (ĐC) 22 107 115HĐ01 21 103 119
Quảng NinhHL3 (ĐC) 23 114 119HĐ01 22 108 121
Địa điểm Dòng/giốngTỷ lệ xuất hiện cành hoa (%)
Tỷ lệ hoa nở (%)
Chiều dài cành hoa
(cm)
Số hoa/cành (hoa)
Độ bền cành hoa
(ngày)
Bắc Ninh
HL3 (ĐC) 90,8 92,8 60,4b 8,1b 32HĐ01 91,1 94,4 73,8a 8,5a 34Tlt 2,05 2,05T tính 9,16 2,18
Hà Nội
HL3 (ĐC) 91,3 92,6 62,1b 8,1b 33HĐ01 92,0 95,9 74,5a 8,6a 35Tlt 2,05 2,05T tính 9,15 2,48
Quảng Ninh
HL3 (ĐC) 91,1 91,7 61,9b 8,2b 33HĐ01 91,8 94,3 74,3a 8,5a 36Tlt 2,05 2,05T tính 9,7 2,39
23
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Bảng 6. Hiệu quả kinh tế của dòng lai hoa lan Hồ điệp HĐ01 trồng ở các địa phương năm 2018 (Tính cho 30.000 cây/1.000 m2 /24 tháng)
Bảng 5. Mức độ bị sâu bệnh hại chính trên dòng lai lan Hồ điệp HĐ01 khi trồng ở các địa phương
Ghi chú: Thời gian theo dõi: tháng 2/2018. Tổng chi ở các địa điểm trồng và trên các dòng, giống coi là như nhau 1.580.000.000 đồng.
3.4. Mưc độ sâu bệnh hại chính trên giống hoa lan hồ điệp HĐ01
Kết quả đánh giá cho thấy ở cả 3 địa điểm dòng lai HĐ01 có tỷ lệ cây bị bệnh thối nhũn thấp dao động trong khoảng 0,9 - 3,9%, bệnh đốm lá và rệp gây hại đều ở mức độ nhẹ, khả năng kháng bệnh tốt hơn so với giống HL3. Điều này thể hiện khả năng thích nghi tốt của dòng lai HĐ01 tại các địa phương khác nhau của miền Bắc Việt Nam.
Qua kết quả khảo nghiệm sản xuất tại các địa phương khác nhau của Việt Nam cho thấy dòng Hồ điệp lai HĐ01 có khả năng thích nghi tốt, sinh trưởng khỏe, chất lượng hoa cao và ổn định, hoa màu hồng phấn có kẻ, đẹp và sắc nét, mức độ nhiễm một số sâu bệnh hại nhẹ, có khả năng kháng bệnh thối nhũn tốt, có thể phát triển thành giống ngoài sản xuất.
Địa điểm Dòng/giống
Thối nhũn (Pseudomonas gadioli)
Đốm lá (Cercospora sojina)
Rệp hại (Chrysomphalus ficus)
Cấp TL (%) Cấp TL (%) Cấp
Bắc NinhHL3 (ĐC) Cấp3 4,8 Cấp 3 3,7 +
HĐ01 Cấp 1 0,9 Cấp 1 0,5 +
Hà NộiHL3 (ĐC) Cấp 3 4,3 Cấp 3 2,9 +
HĐ01 Cấp 1 0,9 Cấp 1 0,5 +
Quảng NinhHL3 (ĐC) Cấp 3 4,6 Cấp 3 4,1 +
HĐ01 Cấp 3 3,9 Cấp 3 3,4 +
3.5. Hiệu quả kinh tế của giống hoa lan Hồ điệp HĐ01Mục đích cuối cùng của người sản xuất là hiệu
quả kinh tế, hiệu quả kinh tế không những phụ thuộc vào năng suất, chất lượng hoa và chi phí đầu tư, mà còn phụ thuộc vào giá trị thương phẩm của
từng loại. Khả năng sinh trưởng của cây tốt thì năng suất hoa cao, chất lượng tốt. Chất lượng hoa tốt thì giá trị thương phẩm cao.
Để đánh giá hiệu quả kinh tế thu được từ việc đầu tư trồng lan Hồ điệp được trình bày ở bảng 6.
Đia điểm Dòng/ giốngTỷ lệ cây
xuất vườn (%)
Giá bán (1.000 đ/
cây)
Tổng thu (1.000 đ)/ 1000 m2
Phần chi (1000 đ)/ 1000 m2
Lãi thuần (1.000đ)
Lợi nhuận (lần)
Bắc NinhHL3 (ĐC) 85,0 85 2.167.500 1.580.000 587.500 1,00
HĐ01 88,5 88 2.336.400 1.580.000 756.400 1,29
Hà NộiHL3 (ĐC) 85,5 85 2.180.250 1.580.000 600.250 1,00
HĐ01 89,0 88 2.349.600 1.580.000 769.600 1,28
Quảng Ninh
HL3 (ĐC) 84,0 85 2.142.000 1.580.000 562.000 1,00
HĐ01 87,5 88 2.310.000 1.580.000 730.000 1,30
Kết quả cho thấy: Tỷ lệ cây xuất vườn của dòng lai HĐ01 ở các địa điểm khảo nghiệm dao động trong khoảng 87,5 - 89,0% cao hơn so với giống đối chứng. Bên cạnh đó, giá bán của dòng lai HĐ01 cũng cao hơn so với giống đối chứng HL3 do dòng lai HĐ01
có màu hoa hồng phấn, màu sắc tươi, cành hoa dài, nhiều bông, ít bị sâu bệnh hại thích hợp trưng bày trong dịp Tết Nguyên Đán hơn màu hoa trắng của giống HL3.

24
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Phần lãi thu được từ các giống đều ở mức cao và có sự chênh lệch do tỷ lệ xuất vườn ở các địa phương là khác nhau. Khi đưa ra ngoài sản xuất, giống nào cho hiệu quả kinh tế cao thì sẽ chọn giống đó để phát triển sản xuất. Tại cả 3 địa điểm khảo nghiệm sản xuất, dòng lai HĐ01 cho lãi cao hơn giống HL3, chúng sinh trưởng tốt, phát triển ổn định qua các năm khảo nghiệm, giữ được ưu điểm như khi khảo nghiệm cơ bản và được thị trường chấp nhận cao.
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luận Kết quả khảo nghiệm sản xuất dòng lan Hồ điệp
lai HĐ01 ở các địa phương cho thấy dòng lai HĐ01 sinh trưởng, phát triển tốt hơn so với đối chứng, khả năng ra hoa ổn định (tỷ lệ ra ngồng và ra hoa đều đạt trên 90%), chất lượng hoa cao (chiều dài cành hoa 73,8 - 74,5 cm, số hoa 8,5 - 8,6 hoa/cành), khả năng kháng sâu bệnh hại tốt, hiệu quả kinh tế cao (gấp 1,28 - 1,30 lần so với đối chứng). Các đặc điểm ưu việt trên không có sự sai khác khi trồng ở các vùng sinh thái khác nhau và không có sự khác biệt so với giai đoạn khảo nghiệm cơ bản. Điều đó thể hiện, năng suất, chất lượng hoa của giống ổn định ở không gian và thời gian khác nhau. Giống được người tiêu dùng và người sản xuất ưa chuộng.
4.2. Đề nghị- Đưa giống ra sản xuất thử ở một số địa phương
thuộc miền Bắc Việt Nam.
- Nghiên cứu một số biện pháp kỹ thuật xây dựng quy trình sản xuất lan Hồ điệp HĐ01 phục vụ sản xuất.
TÀI LIỆU THAM KHẢOĐặng Văn Đông, Bùi Thị Hồng, Mai Thị Ngoan, Trịnh
Khắc Quang, 2009. Kết quả tuyển chọn giống hoa lan Hồ Điệp LVR2, LVR4. Báo cáo nghiệm thu dự án giống giai đoạn 2006 - 2010. Viện Nghiên cứu Rau quả.
QCVN 01-38:2010/BNNPTNT. Quy chuẩn kỹ thuật quốc gia về phương pháp điều tra phát hiện dịch hại cây trồng do Bộ Nông nghiệp và PTNT ban hành tại Thông tư số 71/2010/TT-BNNPTNT ngày 10 tháng 12 năm 2010.
Nguyễn Văn Tỉnh, Đặng Văn Đông, Trịnh Khắc Quang, 2010. Nghiên cứu hoàn thiên quy trình sản xuất hoa lan Hồ Điệp quy mô công nghiệp. Tạp chí Khoa học và Công nghệ Nông nghiệp Việt Nam (ISSN 1859-1558), 5, tr 56-62.
Chen Heming , Zhu Genfa , Lv Fubing , Xiao Wenfang , You Yi1, Li Zuo1,2 , Li Dongmei, 2014. Study on Test Guideline of Distinctness, Uniformity and Stability for Phalaenopsis. Environmental Horticulture Research Institute, Guangdong Academy of Agricultural Sciences, Guangzhou 510640.
Hwang Seung Jae, Jeong Byoung Ryong, 2009. Growth of Phalaenopsis Plants as Affected by Nutrient Solution Composition. Korean Journal of Horticultural Science and Technology, 27 (8): 73-80.
Breeding of Phalaenopsis hybrid variety HD01 Dinh Thi Dinh, Nguyen Thi Kim Oanh, Nguyen Van Tien
AbstractThe testing of Phalaenopsis hybrid variety HD01 in Hanoi, Bac Ninh and Quang Ninh showed that the HD01 hybrid was well grown at all sites. The ratio of outflow and flowering were over 90%. The quality of flower was high; the flower length reached 73.8 - 74.5 cm and the number of flower was 8.5 - 8.6 flowers/spike. The levels of pests and diseases were below 5% and the economic efficiency was 1.28 - 1.30 times higher than that of the control. This variety is preferred by consumers and producers.Keywords: Development, evaluate, growth, hybrid, HĐ01 phalaenopsis, production, quality
Ngày nhận bài: 12/10/2018Ngày phản biện: 19/10/2018
Người phản biện: TS. Đặng Văn ĐôngNgày duyệt đăng: 15/11/2018

25
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
TẠO QUẦN THỂ LAI F1 LÀM VẬT LIỆU KHỞI ĐẦU ĐỂ ĐÁNH GIÁ VAI TRÒ CỦA QTL9 LIÊN QUAN ĐẾN CÁC TÍNH TRẠNG NĂNG SUẤT
CỦA TẬP ĐOÀN LÚA VIỆT NAMVũ Thị Nhiên², Tạ Kim Nhung1,5, Stefan Jouannic4
Lê Hùng Lĩnh¹, Phạm Xuân Hội¹, Trần Khánh Vân², Trần Vũ Hằng¹, Phạm Thị Mai¹, Lê Thị Như1,3, Khổng Ngân Giang¹
TÓM TẮTNăng suất hạt là một trong những chỉ số quan trọng nhất trong chọn tạo lúa, được điều khiển bởi các locus tính
trạng định lượng (QTLs). Các quần thể lai tái tổ hợp RILs (Recombinant Inbred Lines) được sử dụng rộng rãi để nghiên cứu QTL trong hệ gen lúa, với hàng trăm QTL liên quan đến năng suất đã được phát hiện. Nghiên cứu liên kết trên toàn hệ gen (GWAS) các tính trạng liên quan đến năng suất đã được tiến hành trên tập đoàn lúa địa phương Việt Nam và đã xác định được QTL9 mới tiềm năng, liên quan đến số gié thứ cấp/bông và số hạt/bông. Trong nghiên cứu này, các quần thể lai F1 đã được tạo ra từ các cặp lai giữa 2 nhóm lúa có kiểu hình bông trái ngược nhau (bông to và bông nhỏ) để làm vật liệu khởi đầu cho việc đánh giá vai trò của QTL9 thông qua các quần thể lai tái tổ hợp. Mười hai chỉ thị phân tử SSR được sử dụng để kiểm tra cây F1, trong đó 7 chỉ thị cho sự đa hình chiều dài ADN giữa các giống bố mẹ. Như vậy 51 dòng lai F1 đã được chọn lọc bằng 7 chỉ thị phân tử SSR, là nguồn vật liệu để tạo các quần thể lai tái tổ hợp F2, F3.
Từ khóa: QTL, quần thể tái tổ hợp, quần thể lai F1, ADN, SSR
1 Phòng thí nghiệm Việt - Pháp (LMI-RICE2), Phòng thí nghiệm trọng điểm Công nghệ Tế bào thực vật, Viện Di truyền Nông nghiệp, Viện Khoa học Nông nghiệp Việt Nam; 2 Trường Đại học Sư phạm Hà Nội3 Trường Đại học Khoa học Tự nhiên, Đại học Quốc gia Hà Nội; 4 Viện Nghiên cứu và Phát triển Pháp (IRD)5 Plant Genetics Laboratory, National Institute of Genetics, Japan
I. ĐẶT VẤN ĐỀLúa gạo (Oryza sativa L.) là một trong những cây
trồng quan trọng hàng đầu cung cấp lương thực cho hơn 50% dân số thế giới. Để đáp ứng nhu cầu tiêu thụ ngày càng cao, dự đoán sản lượng lúa gạo đến năm 2025 cần tăng thêm 30%. Không những thế, trong bối cảnh bùng nổ dân số, mở rộng đô thị hóa và biến đổi khí hậu, cải tiến và ổn định năng suất lúa gạo trở thành một thách thức lớn đối với các nhà chọn tạo giống.
Các nghiên cứu từ những năm 1960 cho thấy sự đa dạng của các tính trạng nông học quy định năng suất lúa, đây là nguồn tài nguyên di truyền lớn phục vụ cho các chương trình chọn tạo giống lúa cao sản. Theo đó, nhiều công trình nghiên cứu nhằm cải thiện và tạo ra những giống mới có năng suất cao hơn đã được công bố (Xing and Zhang, 2010). Trong số đó, xác định các QTL ảnh hưởng tích cực đến tính trạng năng suất có ý nghĩa quan trọng (Bai et al., 2012; Ikeda et al., 2013; Xing and Zhang, 2010). Gần đây, phương pháp phân tích GWAS ra đời cùng với các công nghệ giải trình tự thế hệ mới đã trở thành một công cụ mạnh mẽ để nghiên cứu sự đa dạng của quần thể và phát hiện thêm nhiều loci quan trọng, đặc biệt là các loci liên kết với các tính trạng nông học phức tạp như tính trạng năng suất (Huang et al., 2012).
Nghiên cứu GWAS các tính trạng liên quan đến năng suất đã được tiến hành trên tập đoàn lúa địa
phương Việt Nam và đã xác định được 29 QTLs tiềm năng. Trong số đó, đáng chú ý là QTL9 chứa 9 chỉ thị SNP (Single Nucleotide Polymorphism), liên kết chặt với 2 tính trạng số gié thứ cấp/bông và số hạt/bông. Tuy nhiên, kết quả của GWAS dựa trên các phân tích thống kê, do vậy các QTL mới tìm được cần được nghiên cứu và chứng minh trong quần thể con lai trước khi đưa vào sử dụng trong các chương trình chọn giống. Vì vậy, trong nghiên cứu này, các quần thể lai F1 đã được tạo ra bằng cách lai 2 nhóm giống lúa bản địa Việt Nam có kiểu hình bông trái ngược nhau (bông to và bông nhỏ), nhằm tạo vật liệu khởi đầu cho việc đánh giá vai trò của QTL9 trong các quần thể lai tái tổ hợp (F2, F3).
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưu- Bốn giống lúa địa phương chứa 9 SNPs nằm
trong vùng QTL9 liên kết với số gié thứ cấp/bông và số hạt/bông, có kiểu hình cấu trúc bông trái ngược nhau (bông to và bông nhỏ) được sử dụng làm bố mẹ để tạo quần thể lai. Nhóm bông nhỏ gồm 2 giống: Sớm Giai Hưng Yên (G6), Ỏn (G19), nhóm bông to: Khẩu Nam Rinh (G189), Blé Blâu Cho (G205). Thông tin chi tiết về các giống được trình bày trong bảng 1.
- 12 cặp mồi SSR (Microsatellite Marker) đã được công bố cho sự đa hình giữa các giống lúa được sử dụng để chọn lọc các cây F1 (Bảng 2).

26
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Bảng 1. Một số chỉ số về cấu trúc bông và năng suất của các giống lúa sử dụng làm bố mẹ để tạo các quần thể lai F1
Ghi chú: Các giống lúa được trồng vào vụ Mùa tại ruộng thí nghiệm của Trung tâm Tài nguyên Di truyền thực vật (2014) và Trạm Nghiên cứu Nông nghiệp Văn Giang (2015).
Tên giống
Địa điểm thu
thập
Năm
Chiều dài
bông (cm)
Số gié sơ cấp/ bông
Chiều dài
gié sơ cấp
(cm)
Khoảng cách
giữa các gié sơ
cấp
Số gié thư cấp/ bông
Chiều dài gié
thư cấp
Khoảng cách
giữa các gié thư
cấp
Số gié tam cấp/ bông
Số hạt/
bông
Thời gian
ra hoa (ngày)
Sớm Giai Hưng Yên
Không xác định
2014 18,21 11,22 11,88 1,81 23,67 2,93 1,12 0,06 130,22 96,50
Ỏn Hà Nội 2014 19,61 11,39 12,10 1,90 24,22 2,58 1,16 0,00 133,94 95,50Khẩu Nam Rinh
Điện Biên 2014 20,41 11,83 14,29 1,91 48,67 3,30 0,72 0,44 250,50 94,50
Blé Blâu Cho Sơn La 2014 22,09 11,78 14,58 2,05 44,44 3,15 0,99 0,00 220,33 99,00
Sớm Giai Hưng Yên
Không xác định
2015 25,04 13,61 16,38 2,03 34,83 4,01 1,40 0,06 187,33 101,00
Ỏn Hà Nội 2015 27,58 13,83 17,02 2,18 31,11 3,53 1,69 0,44 176,83 100,50Khẩu Nam Rinh
Điện Biên 2015 29,85 12,72 20,59 2,57 45,22 4,76 1,19 0,89 228,22 79,00
Blé Blâu Cho Sơn La 2015 28,97 13,33 18,30 2,35 47,39 3,97 1,38 0,50 244,22 108,00
Bảng 2. Các chỉ thị phân tử SSR sử dụng để chọn lọc cây F1Tên chỉ
thị Số hiệu NST Trình tự lặp
Kích cỡ (bp) Trình tự mồi Tác giả
RM151 L37528 1 (TA)23 205 - 317 F-ggctgctcatcagctgcatgcg Temnykh et al., 2000; Akagi et al., 1996R-tcggcagtggtagagtttgatctgc
RM180 D63901 7 (ATT)10 107 - 204 F-ctacatcggcttaggtgtagcaacacg Temnykh et al., 2000R-acttgctctacttgtggtgagggactg
RM204 AF344025 6 (CT)44 106 - 194 F-gtgactgacttggtcataggg Chen et al., 1997R-gctagccatgctctcgtacc
RM289 AF344115 5 G11(GA)16 88 - 180 F-ttccatggcacacaagcc Temnykh et al., 2000R-ctgtgcacgaacttccaaag
RM320 AF344145 7 (AT)11G-TAT(GT)13 153 - 254 F-caacgtgatcgaggatagatc Temnykh et al., 2000
R-ggatttgcttaccacagctc
RM400 AQ051253 6 (ATA)63 195 - 321 F-acaccaggctacccaaactc Temnykh et al., 2001R-cggagagatctgacatgtgg
RM410 AQ156440 9 (TA)13 173 - 269 F-gctcaacgtttcgttcctg Temnykh et al., 2001R-gaagatgcgtaaagtgaacgg
RM491 AQ510175 12 (AT)14 263 - 400 F-acatgatgcgtagcgagttg Temnykh et al., 2001R-ctctcccttcccaattcctc
RM532 AQ843286 3 (CA)9 166 - 180 F-tctataatgtagcccccccc Temnykh et al., 2001R-tttcaggggcttctaccaac
RM535 AQ857127 2 (AG)11 138 - 410 F-actacatacacggcccttgc Temnykh et al., 2001R-ctacgtggacaccgtcacac
RM577 AP000816 1 (TA)9(-CA)8 191 - 270 F-gctttccctctaacccctct Temnykh et al., 2001
McCouch et al., 2001R-ggatgtaccgctgacatgaa
RM592 AC016779 5 (ATT)20 210 - 400 F-tctttggtatgaggaacacc Temnykh et al., 2001R-agagatccggtttgttgtaa

27
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
2.2. Phương pháp nghiên cưu
2.2.1. Xác định các giống lúa bố mẹ để tạo quần thể lai F1
Dựa vào sự phân tích đa hình của 9 SNP nằm trong vùng QTL9 và kiểu hình cấu trúc bông cũng như các tính trạng khác liên quan đến năng suất giữa các giống lúa trong tập đoàn nghiên cứu GWAS để xác định các giống lúa chứa đa hình của 9 SNPs nằm trong vùng QTL9 và có kiểu hình bông trái ngược nhau (bông to và bông nhỏ), dùng làm bố mẹ để tạo quần thể lai F1.
2.2.2. Tạo quần thể lai F1
Quần thể F1 được lai tạo bằng phương pháp lai truyền thống. Các giống lúa được chọn làm bố mẹ để lai được gieo trồng tại nhà lưới Viện Di truyền Nông nghiệp, mỗi giống 5 cây, trồng thành 3 đợt, mỗi đợt cách nhau 1 tuần. Cây được trồng trong xô nhựa có dung tích 5 L chứa 50% đất thịt + 50% giá thể chuyên dùng trồng lúa. Vì có sự chênh lệch về thời gian ra hoa nên các giống có thời gian ra hoa dài hơn được trồng trước (G6, G205) 1 - 2 tuần so với các giống có thời gian ra hoa sớm (G19, G189) nhằm đảm bảo các giống ra hoa cùng thời điểm để tiến hành lai. Việc xử lý thời gian chiếu sáng trong ngày nhằm mục đích điều khiển sự ra hoa đã được tiến hành. Các giống lúa bố mẹ được xử lý theo quang chu kỳ 9 giờ chiếu sáng/ngày (từ 8 h sáng đến 17 h chiều) và 15 h trong buồng tối sau 2 tháng gieo trồng cho đến khi cây lúa trổ bông.
2.2.3. Chọn lọc cây F1 bằng chỉ thị phân tử SSR
a) Thu thập mẫu và tách chiết ADN
20 hạt F1 của mỗi cặp lai được gieo trồng trong nhà lưới để đánh giá bằng chỉ thị phân tử SSR. Cây lúa sau khi gieo trồng từ 20 ngày, lấy lá thứ 2 từ trên xuống làm mâu phân tích ADN. ADN được tách chiết bằng phương pháp sử dụng CTAB của Doyle và Doyle có cải tiến (Doyle, 1991).
b) Phản ứng PCR
Phản ứng PCR được thực hiện trong thể tích 20 µl, gồm 20 ng ADN, 0,1 mM dNTPs (#R0182, Fermentas), 0,1 µM mồi xuôi và ngược, 2 µl 10X Dream Taq Buffer (#R0971, Fermentas), 0,2 µl Dream
Taq DNA polymerase 5u/µl (#EP0702, Fermentas) và nước Milli Q khử trùng. Chu trình nhiệt: 950C trong 5 phút, và 30 chu kỳ: 950C - 30 giây, Ta - 30 giây (nhiệt độ gắn mồi), 720C - 30 giây và giữ 5 phút ở 720C. Sản phẩm PCR được kiểm tra bằng điện di trên gel agarose 2,5% rồi được soi trong buồng UV (Uvltec, Cambridge), phần mềm Fire Reader. Các thí nghiệm điện di sử dụng thang chuẩn ADN Low Gene Ruler 100 bp (#SM1163, Fermentas).
2.3. Thời gian và địa điểm nghiên cưu
Các giống lúa bố mẹ được gieo trồng vào vụ Xuân năm 2017 và được lai với nhau tại nhà lưới Viện Di truyền Nông nghiệp. Các dòng F1 được trồng và đánh giá vào vụ Mùa năm 2017, cũng tại nhà lưới Viện Di truyền Nông nghiệp.
Các thí nghiệm sinh học phân tử nhằm chọn lọc cây F1 được tiến hành tại Phòng thí nghiệm LMI-RICE2,Viện Di truyền Nông nghiệp.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Xác định các giống lúa bố mẹ để tiến hành lai tạo quần thể lai F1
Vùng QTL9 chứa 9 SNP (gagagcgaa/atataaatt) liên kết chặt với nhau và liên kết với 2 tính trạng số gié thứ cấp/bông và số hạt/bông. Các giống lúa bố mẹ được xác định dựa vào phân tích sự đa hình của 9 SNP và kiểu hình cấu trúc bông cũng như thời gian ra hoa của các giống lúa trong tập đoàn. Bốn giống lúa cho sự đa hình SNP và có kiểu hình cấu trúc bông khác biệt tương đối lớn đã được chọn ra. Hai giống G6 và G19 chứa 9 SNP (gagagcgaa) có kiểu hình bông nhỏ, số gié thứ cấp/bông dao động từ 23 - 34 gié đối với giống G6 và 24 - 31 gié đối với giống G19, số hạt/bông của giống G6 là từ 130 - 187 hạt trong khi giống G19 đạt 133 - 177 hạt. Hai giống có kiểu hình bông to G189 và G205 chứa 9 SNP (ata-taaatt) có số gié thứ cấp/bông khoảng 45 - 48 gié và số hạt/bông từ 220 - 250 hạt (Bảng 1, hình 1). Như vậy bốn giống lúa (G6, G19, G189, G205) được chọn để lai với nhau để tạo các quần thể lai F1, trong đó nhóm lúa bông nhỏ (G6, G19) được chọn làm cây mẹ, nhóm lúa bông to (G189, G205) được chọn làm cây bố.

28
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
3.2. Tạo quần thể lai F1Các giống lúa bố mẹ (G6, G19 và G189, G205)
được trồng trong chậu vại tại nhà lưới của Viện Di truyền Nông nghiệp vào vụ Xuân năm 2017. Đến thời kỳ ra hoa, các cặp lai được tiến hành lai với nhau theo phương pháp truyền thống. Năm ngày sau khi lai, hạt lai F1 đã xuất hiện trên bông ở tất cả các cây mẹ của 2 giống G6, G19.
3.3. Chon loc cây F1 bằng chỉ thị phân tử SSR
3.3.1. Khảo sát đa hình giữa hai giống bông bố mẹ để xác định những chỉ thị SSR cho đa hình
Mười hai chỉ thị phân tử SSR đã được công bố cho sự đa hình giữa các giống lúa (Bảng 2) đã được sử dụng để khảo sát đa hình giữa các cặp giống bông bố mẹ. Phân tích kết quả các phản ứng PCR với ADN khuôn là ADN của các giống bố mẹ với 12 cặp mồi tương ứng với 12 chỉ thị phân tử cho thấy có 7 chỉ thị cho đa hình giữa các giống bố mẹ. Trong đó, 2 chỉ thị RM320 và RM180 chỉ cho duy nhất sự đa hình về chiều dài ADN giữa giống G6 và 3 giống còn lại (G19, G189, G205) (Hình 3B, 3C), do đó nó có thể được sử dụng để phân tích quần thể F1 của 2 cặp lai G6 ˟ G189 và G6 ˟ G205. Ngược lại, 2 chỉ thị phân tử RM204 và RM289 cho đa hình về chiều dài ADN giữa giống G19 và 3 giống còn lại (G6, G189, G205)
và có thể sử dụng để phân tích quần thể lai F1 của 2 cặp G19 ˟ G189 cũng như G19 và G205 (Hình 3C). Chỉ thị phân tử RM491 mặc dù không đặc hiệu ở 2 giống G6 và G19 nhưng vân cho sự đa hình giữa 2 giống so với giống 189 (Hình 3A) nên có thể sử dụng để chọn lọc cây F1 của 2 cặp lai G6 ˟ G189 và G19 ˟ G189. Hai chỉ thị phân tử RM592 và RM410 cho sự đa hình giữa 2 nhóm giống bố, mẹ (Hình 3A và 3C) nên có thể sử dụng để xác định cây F1 của cả 4 cặp lai. Năm chỉ thị RM151, RM 400, RM532, RM535 và RM577 không cho đa hình giữa các giống bố mẹ.
3.3.2. Chọn lọc các dòng F1 bằng chỉ thị phân tửTừ kết quả thu được của phần khảo sát đa hình
các chỉ thị phân tử ở các giống bố mẹ, các dòng lai F1 thu được từ 4 cặp lai khác nhau đã được đánh giá với 7 chỉ thị phân tử cho sự đa hình tương ứng với từng cặp lai như trong bảng 3. a) Quần thể lai F1 của cặp lai G6 ˟ G189
Quần thể lai F1 của cặp lai G6xG189 được phân tích với 3 chỉ thị phân tử RM320, RM180, RM491. Chỉ thị RM320 cho phép xác định 11 dòng F1 mang cả 2 locus của bố và mẹ (Hình 4A), còn với chỉ thị RM180 và RM491 số dòng F1 được xác định lần lượt là 16 và 10 dòng (Hình 4B và 4C). Tổng hợp kết quả phân tích với 3 chỉ thị phân tử, 11 dòng F1 được xác định ít nhất với 2 chỉ thị phân tử (Hình 4D).
A BSố
giéthứcấp/
bông
Sốhạ
t/bô
ng
G189, G205
G6
G19
G189
G205
C
G6, G19G189, G205G6, G19
Hình 1. Hình thái cấu trúc bông của 4 giống lúa sử dụng làm bố, mẹ để tạo quần thể lai F1Ghi chú: A: số gié thứ cấp/bông của 2 nhóm lúa bông nhỏ G6, G19 (màu ghi) và bông to G189, G205 (màu vàng);
B: số hạt/bông của 2 nhóm lúa bông nhỏ G6, G19 (màu ghi) và bông to G189, G205 (màu vàng); C: ảnh bông lúa của 4 giống G6, G19, G189 và G205.
D

29
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
H20 G6 G19 G189 G205 M G6 G19 G189 G205 H20
H20 G6 G19 G189 G205 M G6 G19 G189 G205 H20
RM 180 RM204
RM289 RM410
107 bp204 bp
88 bp180 bp
106 bp194 bp
173 bp269 bp
C
Hình 3. Khảo sát sự đa hình của các chỉ thị phân tử SSR ở các giống bố mẹ (G6, G19, G189, G205)Ghi chú: A: khảo sát sự đa hình của 2 chỉ thị phân tử RM491 và RM592; B: khảo sát sự đa hình của 2 chỉ thị phân
tử RM320 và RM400; C: khảo sát sự đa hình của 4 chỉ thị phân tử RM180, RM204, RM289, RM410.
Hình 4. Kết quả chọn lọc dòng F1 của cặp lai G6 ˟ G189 với 3 chỉ thị phân tử RM320 (A), RM180 (B) và RM491 (C). D, các dòng F1 được chọn bằng ít nhất 2 chỉ thị SSR
Ghi chú: M: thang chuẩn ADN 100 bp (Promega).
D
A B
C
C
B
A
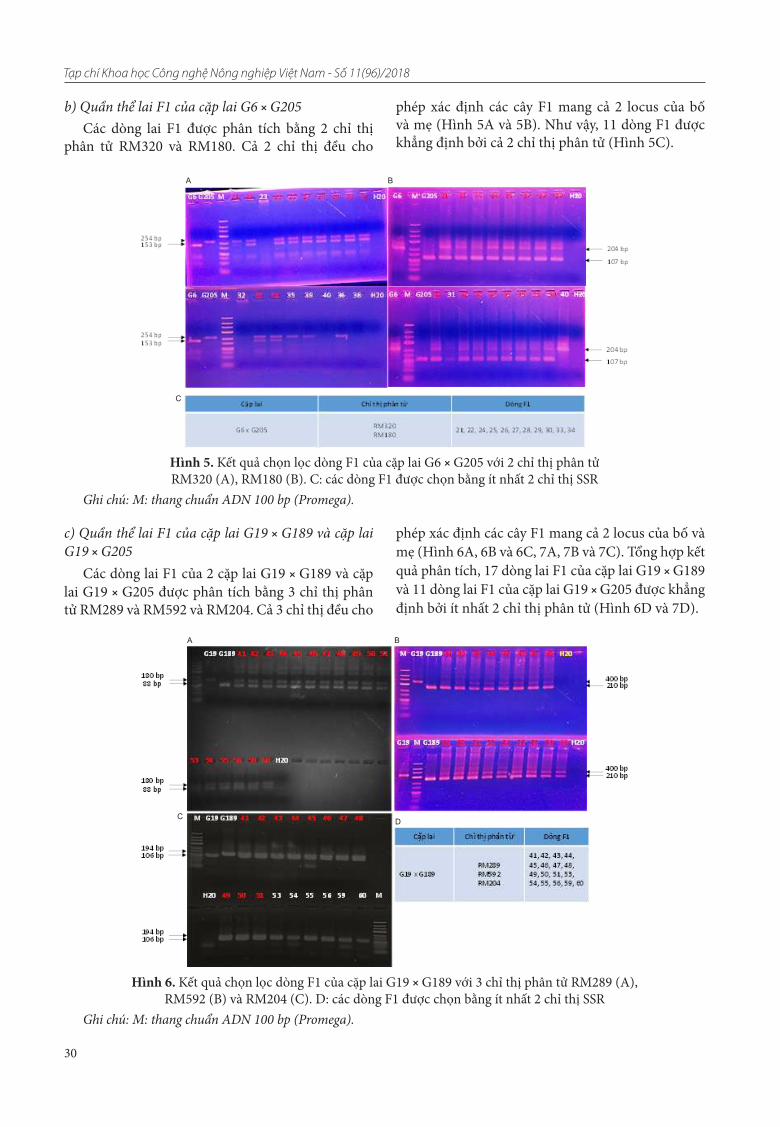
30
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
b) Quần thể lai F1 của cặp lai G6 ˟ G205Các dòng lai F1 được phân tích bằng 2 chỉ thị
phân tử RM320 và RM180. Cả 2 chỉ thị đều cho
phép xác định các cây F1 mang cả 2 locus của bố và mẹ (Hình 5A và 5B). Như vậy, 11 dòng F1 được khẳng định bởi cả 2 chỉ thị phân tử (Hình 5C).
c) Quần thể lai F1 của cặp lai G19 ˟ G189 và cặp lai G19 ˟ G205
Các dòng lai F1 của 2 cặp lai G19 ˟ G189 và cặp lai G19 ˟ G205 được phân tích bằng 3 chỉ thị phân tử RM289 và RM592 và RM204. Cả 3 chỉ thị đều cho
phép xác định các cây F1 mang cả 2 locus của bố và mẹ (Hình 6A, 6B và 6C, 7A, 7B và 7C). Tổng hợp kết quả phân tích, 17 dòng lai F1 của cặp lai G19 ˟ G189 và 11 dòng lai F1 của cặp lai G19 ˟ G205 được khẳng định bởi ít nhất 2 chỉ thị phân tử (Hình 6D và 7D).
Hình 5. Kết quả chọn lọc dòng F1 của cặp lai G6 ˟ G205 với 2 chỉ thị phân tử RM320 (A), RM180 (B). C: các dòng F1 được chọn bằng ít nhất 2 chỉ thị SSR
Ghi chú: M: thang chuẩn ADN 100 bp (Promega).
Hình 6. Kết quả chọn lọc dòng F1 của cặp lai G19 ˟ G189 với 3 chỉ thị phân tử RM289 (A), RM592 (B) và RM204 (C). D: các dòng F1 được chọn bằng ít nhất 2 chỉ thị SSR
Ghi chú: M: thang chuẩn ADN 100 bp (Promega).
C
D
A B
C
A B

31
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Hình 7. Kết quả chọn lọc dòng F1 của cặp lai G19 ˟ G205 với 3 chỉ thị phân tử RM289 (A), RM592 (B) và RM204 (C). D, các dòng F1 được chọn bằng ít nhất 2 chỉ thị SSR
Ghi chú: M: thang chuẩn ADN 100 bp (Promega).
IV. KẾT LUẬNBốn quần thể lai F1 đã được tạo ra từ 4 giống
lúa bố mẹ đại diện cho 2 nhóm lúa có cấu trúc bông to và nhóm có cấu trúc bông nhỏ, mang QTL9 liên quan đến các tính tạng năng suất. Đánh giá độ đa hình của mười hai chỉ thị phân tử SSR giữa các cây bố, mẹ đã thu được 7 chỉ thị có sự đa hình và 5 chỉ thị không cho sự đa hình hoặc không đặc hiệu. Phân tích 80 cây F1 bằng 7 chỉ thị cho sự đa hình đã chọn ra được 51 cây F1 mang cả 2 locus của bố và mẹ, là nguồn vật liệu khởi đầu để tạo các quần thể tái tổ hợp F2 và F3.
TÀI LIỆU THAM KHẢOAkagi Y. Yokozeki A. Inagaki T. Fujimura, 1996.
Microsatellite DNA markers for rice chromosomes. Theoretical and Applied Genetics, 93 (7): 1071-1077.
Bai, Xufeng, Bi Wu, and Yongzhong Xing, 2012. Yield-Related QTLs and Their Applications in Rice Genetic ImprovementF. Journal of Integrative Plant Biology, 54 (5): 300-311.
Chen S. Temnykh Y. Xu Y. G. Cho S. R. McCouch, 1997. Development of a microsatellite framework map providing genome-wide coverage in rice (Oryza sativa L.). Theoretical and Applied Genetics, 95 (4): 553-567.
Doyle J., 1991. DNA Protocols for Plants. In: Hewitt G.M., Johnston A.W.B., Young J.P.W. (eds) Molecular Techniques in Taxonomy. NATO ASI Series (Series H: Cell Biology), vol 57. Springer, Berlin, Heidelberg.
Huang, Xuehui, Yan Zhao, Xinghua Wei, Canyang Li, Ahong Wang, Qiang Zhao, Wenjun, 2012. Genome-Wide Association Study of Flowering Time and Grain Yield Traits in a Worldwide Collection of Rice Germplasm. Nature Genetics, 44 (1): 32-39.
Ikeda, Mayuko, Kotaro Miura, Koichiro Aya, Hidemi Kitano, and Makoto Matsuoka, 2013. Genes Offering the Potential for Designing Yield-Related Traits in Rice. Current Opinion in Plant Biology, March.
McCouch, S. Temnykh, A. Lukashova, J. Coburn, G. DeClerck, S. Cartinhour, S. Harrington, M. Thomson, E. Septiningsih, M. Semon, P. Moncada and Jiming Li, 2001. Microsatellite markers in rice: abundance, diversity, and applications. Rice Genetics Collection Rice Genetics IV: 117-135.
Temnykh S., William D. Park, Nicola Ayres, Sam Cartinhour, N. Hauck, L.Lipovich, Y. G. Cho, T. Ishii, S. R. McCouch, 2000. Mapping and genome organization of microsatellite sequences in rice (Oryza sativa L.). Theoretical and Applied Genetics, 100 (5): 697-712.
Temnykh Svetlana, Genevieve DeClerck, Angelika Lukashova, Leonard Lipovich, Samuel Cartinhour1, and Susan McCouch, 2001. Computational and Experimental Analysis of Microsatellites in Rice (Oryza sativa L.): Frequency, Length Variation, Transposon Associations, and Genetic Marker Potential. Genome Res., 11: 1441-1452.
Xing, Yongzhong, and Qifa Zhang, 2010. Genetic and Molecular Bases of Rice Yield. Annual Review of Plant Biology 61 (1): 421-442.
D
C
A B

32
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Creation of F1 populations as prebreeding materials to characterize the role of QTL9 for yield-related traits in Vietnamese local rice collection
Vu Thi Nhien, Tạ Kim Nhung , Stefan Jouannic, Le Hung Linh, Pham Xuan Hoi, Tran Khanh Van,
Tran Vu Hang, Pham Thi Mai, Le Thi Nhu, Khong Ngan GiangAbstractGrain yield is one of the most important indexes in rice breeding, which is controlled by quantitative trait loci (QTLs). Recombinant inbred line populations (RILs) have been widely used to discover QTLs in rice genome-wide, with hundreds of yield-related QTLs. In this research, F1 populations were created as prebreeding materials to develop RILs populations for validation of new QTL deriving from GWAS analysis for yield-related traits in Vietnamese local varieties. Four F1 populations were obtained from 4 crossings between 2 groups of rice displaying contrasted panicle structures, low branched versus high branched. Twelve SSR markers were tested for checking F1 lines, among them 7 SSR markers gave polymorphisms between parent lines. Therefore, 51 F1 lines selected by 7 SSR markers can be used as prebreeding materials to evaluate the role of the candidate QTL.Keywords: QTL, F1 populations, RILs, DNA, SSR
Ngày nhận bài: 18/9/2018Ngày phản biện: 22/9/2018
Người phản biện: TS. Trần Danh SửuNgày duyệt đăng: 15/10/2018
1 Phòng Thí nghiệm Trọng điểm Công nghệ Tế bào thực vật, Viện Di truyền Nông nghiệp
NGHIÊN CỨU CHUYỂN CẤU TRÚC GEN 35S::GmNAC004 VÀO GIỐNG ĐẬU TƯƠNG ĐT22 THÔNG QUA CHỦNG KHUẨN
Agrobacterium tumefaciens EHA101Nguyễn Văn Đồng1, Nguyễn Anh Vũ1,
Lê Thị Mai Hương1, Nguyễn Trung Anh1
TÓM TẮTTrong nghiên cứu này, gen GmNAC004 được sử dụng để biến nạp vào 3029 mâu nửa lá mầm của giống đậu tương
ĐT22 thông qua chủng khuẩn A. tumefaciens EHA101 mang vector pZY101::35S::GmNAC004. Kết quả biến nạp cho thấy, tỷ lệ mâu tạo đa chồi đạt 69,56% và tỷ lệ mâu sống sót sau chọn lọc đạt 3,84%. Phân tích cây đậu tương tái sinh sau quá trình chuyển gen đã thu được 18 dòng, vừa kháng thuốc trừ cỏ đồng thời cũng dương tính với phân tích PCR, đạt hiệu suất chuyển gen 0,59%, 18 dòng này đều có biểu hiện gen ở thế hệ T1.
Từ khóa: Agrobacterium tumefaciens, chuyển gen, 35S::GmNAC004
I. ĐẶT VẤN ĐỀCác yếu tố phiên mã NAC (NAM, ATAF và CUC)
đã được báo cáo là tăng cường khả năng chống chịu của cây trồng đối với các điều kiện bất thuận như hạn, mặn và lạnh (Tran et al., 2010). Các nghiên cứu sâu về yếu tố NAC trên một số cây trồng đã đưa ra giả thuyết là ít nhất có 105 yếu tố phiên mã NAC ở cây Arabidopsis, 140 ở cây lúa, 205 ở cây đậu tương và 152 ở cây thuốc lá (Fang et al., 2008; Mochida et al., 2009; Ooka et al., 2003; Rushton et al., 2008).
Trong nhóm gen GmNAC, gen GmNAC004 là một trong những gen điều khiển liên quan đến khả năng chịu hạn, mặn và lạnh có khả năng biểu hiện mạnh (Tran et al., 2009). Lần đầu tiên, nghiên cứu về siêu
biểu hiện của 2 cấu trúc pGreen-P35SGmNAC003 và pGreen-P35S-GmNAC004 trên cây Arabidopsis được công bố (Truyen et al., 2014). Các cây Arabidopsis biểu hiện gen GmNAC004 cho thấy sự gia tăng số lượng và chiều dài rễ trong điều kiện không hạn và duy trì số lượng và chiều dài rễ dưới điều kiện hạn nhẹ so với cây đối chứng, trong khi cây biến đổi gen GmNAC003 không cho thấy bất kỳ phản ứng nào.
Cho tới nay, tất cả các công trình biến nạp gen vào đậu tương mới chỉ thành công trên giống mô hình như G. max ‘Jack’, Williams. Do có sự khác biệt về nguồn gốc nên hầu hết các giống mô hình khó ra hoa kết quả tại Việt Nam. Nguyễn Văn Đồng và cộng tác viên (2017) đã nghiên cứu chuyển gen

33
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
GmNAC004 vào giống đậu tương ĐT22 sử dụng promoter RD29A và xác định được 8 dòng có kết quả PCR dương tính với gen đích ở thế hệ T0 với hiệu suất chuyển gen đạt 0,28%. RD29A được xác định là một promoter cảm ứng với các điều kiện môi trường bất lợi như hạn, mặn và lạnh, trong khi 35S là promoter cơ định tham gia điều khiển quá trình biểu hiện gen ở hầu hết các loại mô khác nhau trong nhiều giai đoạn phát triển của sinh vật. Promoter 35Slà một promoter cấu trúc mạnh, gây ra mức độ cao trong sự biểu hiện gen ở thực vật và có khả năng kích hoạt sự hoạt động của các gen thực vật hoặc gen nội sinh.
Nhằm tăng cường mức độ biểu hiện gen và nâng cao hiệu suất chuyển gen GmNAC004 ở đậu tương, nghiên cứu chuyển cấu trúc promoter::gen 35S::GmNAC004 vào giống đậu tương ĐT22 thông qua vi khuẩn A. tumefaciens EHA101 được tiến hành.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưu- Vật liệu thực vật: Giống đậu tương ĐT22 (Trung
tâm Nghiên cứu và Phát triển Đậu đỗ - Viện Cây lương thực và Cây thực phẩm chọn tạo).
- Vật liệu di truyền: Chủng vi khuẩn A. tumefaciens EHA101 mang vector pZY101::35S::GmNAC004 chứa gen chịu hạn GmNAC004 và gen chỉ thị chọn lọc thực vật bar - kháng glufosinate (Hình 1), hiện đang được lưu giữ tại Phòng Thí nghiệm Trọng điểm Công nghệ Tế bào thực vật (Nguyễn Văn Đồng và ctv., 2012).
Hình 1. Sơ đồ vector pZY101::35S::GmNAC004
2.2. Phương pháp nghiên cưu
2.2.1. Phương pháp biến nạpPhương pháp tạo vật liệu vô trùng và phương
pháp chuyển gen vào cây đậu tương theo quy trình của Zhang và cộng tác viên (2004) có cải tiến (Nguyễn Văn Đồng và ctv., 2012).
2.2.2. Phân tích cây chuyển gen- Sàng lọc cây chuyển gen T0 và T1 thông qua xử lý
với thuốc trừ cỏ basta: Nhằm lựa chọn những dòng đậu tương đồng hợp tử cho các thí nghiệm đánh giá tiếp theo ngoài đồng ruộng, cây con T0 và 30 - 35 cây T1 của mỗi một dòng T0 tương ứng sau khi trồng trong bầu đất được 2 tuần tuổi đã phát sinh 3 - 5 lá thật được sử dụng trong thí nghiệm phun basta (chứa 2% glufosinate). Nồng độ basta sử dụng để chọn lọc là 100 mg/l. Thí nghiệm phun basta được tiến hành lặp lại 2 lần, mỗi lần cách nhau 3 ngày. Những cây con sống sót sau 2 lần phun basta được tiếp tục chăm sóc làm vật liệu cho những phân tích đánh giá tiếp theo.
- Phân tích PCR: Tách chiết ADN tổng số theo phương pháp của Doyle và cộng tác viên (1987) có cải tiến. Tiến hành phân tích PCR với cặp mồi BAR-F78/ BAR-R506 đặc hiệu cho gen bar và cặp mồi 2X35S-F/ RB-R đặc hiệu cho gen GmNAC004 (Bảng 1). Chương trình phản ứng chung của 3 cặp mồi là 950C/5 phút, 950C/30 giây, 570C/30 giây, 720C/90 giây, lặp lại trong 35 chu kỳ, 720C/10 phút, kết thúc 40C. Sản phẩm PCR được kiểm tra bằng điện di trên gel agarose 1%.
Bảng 1. Trình tự các đoạn mồi sử dụng trong nghiên cứu
- Các chỉ tiêu theo dõi và đánh giá:Số mâu phát sinh đa chồi Tổng số mâu lây nhiễm
Tỷ lệ mâu đậu tương đa chồi (%)
˟ 100=
Số mâu sống sót sau chọn lọc Số mâu đưa vào chọn lọc
= ˟ 100Tỷ lệ sống sótsau chọn lọc (%)
Số cây dương tính với PCRTổng số mâu lây nhiễm ban đầu
= ˟ 100Hiệu suất biến nạp ở thế hệ T0 (%)
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Chuyển cấu trúc 35S::GmNAC004 vào giống đậu tương ĐT22
Kết quả biến nạp cấu trúc 35S::GmNAC004 vào 3029 mâu nửa lá mầm cho thấy, tỉ lệ mâu biến nạp tạo đa chồi đạt 69,56 %, tỉ lệ mâu sống sót sau giai đoạn chọn lọc đạt 3,84 %, số cây chuyển gen thế hệ T0 sống sót khi đưa ra môi trường tự nhiên thu
TT Tên mồi Trình tự mồi 1 BAR-R506 5’-CTGAAGTCCAGCTGCCAGA-3’2 BAR-F78 5’-CCACTACATCGAGACCAGCA-3’3 RB-R CTTTTCTCTTAGGTTTACCCGCCA4 2X35S-F TGCTCCACCATGTTGACCTGCA
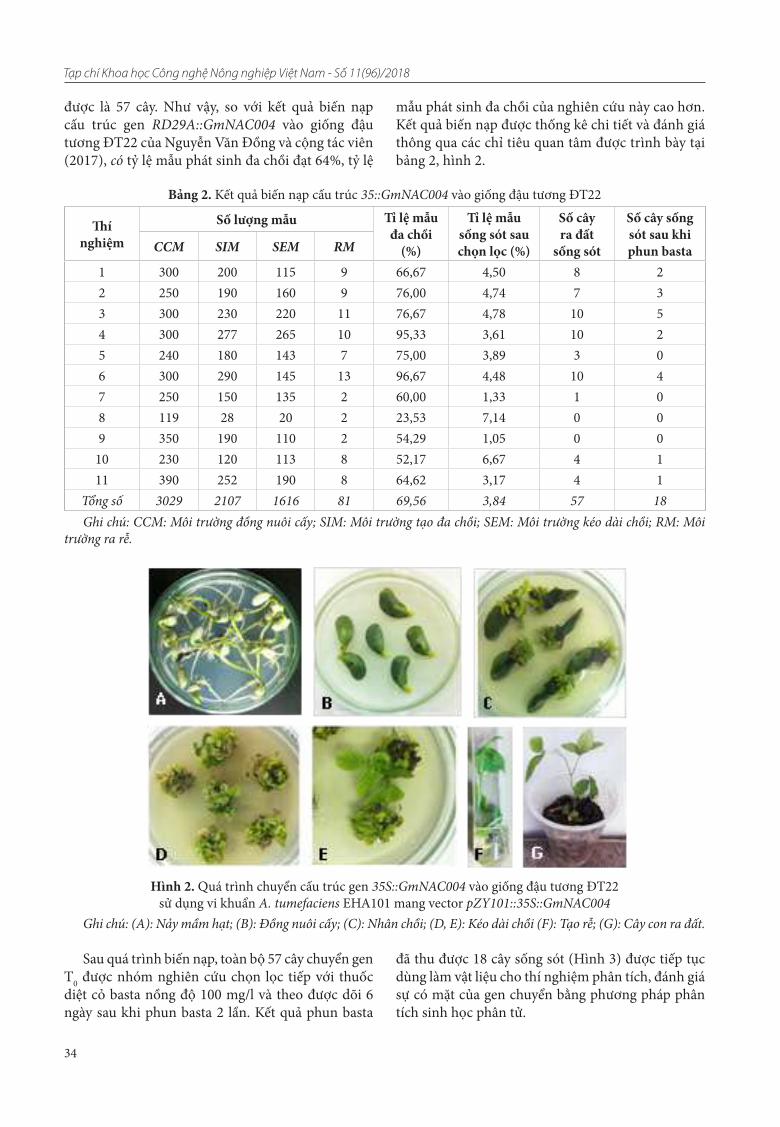
34
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
được là 57 cây. Như vậy, so với kết quả biến nạp cấu trúc gen RD29A::GmNAC004 vào giống đậu tương ĐT22 của Nguyễn Văn Đồng và cộng tác viên (2017), có tỷ lệ mâu phát sinh đa chồi đạt 64%, tỷ lệ
mâu phát sinh đa chồi của nghiên cứu này cao hơn. Kết quả biến nạp được thống kê chi tiết và đánh giá thông qua các chỉ tiêu quan tâm được trình bày tại bảng 2, hình 2.
Sau quá trình biến nạp, toàn bộ 57 cây chuyển gen T0 được nhóm nghiên cứu chọn lọc tiếp với thuốc diệt cỏ basta nồng độ 100 mg/l và theo được dõi 6 ngày sau khi phun basta 2 lần. Kết quả phun basta
đã thu được 18 cây sống sót (Hình 3) được tiếp tục dùng làm vật liệu cho thí nghiệm phân tích, đánh giá sự có mặt của gen chuyển bằng phương pháp phân tích sinh học phân tử.
Bảng 2. Kết quả biến nạp cấu trúc 35::GmNAC004 vào giống đậu tương ĐT22
Ghi chú: CCM: Môi trường đồng nuôi cấy; SIM: Môi trường tạo đa chồi; SEM: Môi trường kéo dài chồi; RM: Môi trường ra rễ.
Hình 2. Quá trình chuyển cấu trúc gen 35S::GmNAC004 vào giống đậu tương ĐT22 sử dụng vi khuẩn A. tumefaciens EHA101 mang vector pZY101::35S::GmNAC004
Ghi chú: (A): Nảy mầm hạt; (B): Đồng nuôi cấy; (C): Nhân chồi; (D, E): Kéo dài chồi (F): Tạo rễ; (G): Cây con ra đất.
Thí nghiệm
Số lượng mẫu Tỉ lệ mẫu đa chồi
(%)
Tỉ lệ mẫu sống sót sau chon loc (%)
Số cây ra đất
sống sót
Số cây sống sót sau khi phun bastaCCM SIM SEM RM
1 300 200 115 9 66,67 4,50 8 22 250 190 160 9 76,00 4,74 7 33 300 230 220 11 76,67 4,78 10 54 300 277 265 10 95,33 3,61 10 25 240 180 143 7 75,00 3,89 3 06 300 290 145 13 96,67 4,48 10 47 250 150 135 2 60,00 1,33 1 08 119 28 20 2 23,53 7,14 0 09 350 190 110 2 54,29 1,05 0 0
10 230 120 113 8 52,17 6,67 4 111 390 252 190 8 64,62 3,17 4 1
Tổng số 3029 2107 1616 81 69,56 3,84 57 18

35
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Bảng 3. Kết quả phân tích PCR các cây đậu tương chuyển cấu trúc gen 35S::GmNAC004 thế hệ T0
Hình 4. Kết quả điện di sản phẩm PCR sử dụng cặp mồi BAR-F78/ BAR- R506Ghi chú: (M): Marker 1kb plus genruler; (-): H2O; (P): Plasmid 35S::GmNAC004; (1-14): Cây chuyển gen
35S::GmNAC004 thế hệ T0.
Hình 3. Kết quả chọn lọc bằng basta sau 6 ngày của các cây đậu tương chuyển cấu trúc gen 35S::GmNAC004 thế hệ T0
Ghi chú: (A): Cây đậu tương ĐT22 không chuyển gen; (B, C): Cây chuyển gen T0.
3.2. Phân tích sự có mặt của cấu trúc gen 35S::GmNAC004 trong genome của dòng đậu tương chuyển gen
Tiến hành thu mâu lá của 18 cây con chuyển gen T0 sống sót sau khi phun basta để tách chiết ADN tổng số phục vụ cho thí nghiệm phân tích PCR. Kết quả phân tích được trình bày trong bảng 3.
Phân tích PCR sự có mặt gen bar bằng cặp mồi BAR-F78/ BAR-R506, kết quả cho thấy: đối chứng âm và cây ĐT22 không cho băng, đối chứng dương cho băng nét và rõ ràng, các cây chuyển gen cho băng có kích thước bằng với kích thước plasmid. Tổng số 18 cây chuyển gen T0 đều dương tính với gen bar. Kết quả điện di kiểm tra sản phẩm PCR gen bar được thể hiện đại diện như hình 4.
Tiếp tục phân tích PCR sự có mặt của gen GmNAC004 bằng cặp mồi 2X35S-F/ RB-R trong 18 cây chuyển gen T0 có kết quả PCR gen bar dương tính, kết quả 18 cây đều dương tính với gen quan tâm đạt hiệu suất chuyển gen 0,59 %. Các băng thu nhận được có kích thước bằng với kích thước đối chứng dương plasmid 35S::GmNAC004 (1400 bp). Đối chứng âm nước và ĐT22 không xuất hiện băng chứng tỏ phản ứng không bị nhiễm và cặp mồi sử dụng đặc hiệu không xuất hiện băng nội sinh. Kết quả điện di kiểm tra sản phẩm PCR gen GmNAC004 được thể hiện đại diện như hình 5.
Hình 5. Kết quả điện di sản phẩm PCR sử dụng cặp mồi 2X35S-F/ RB-R
Ghi chú: (M): Marker 1kb plus genruler; (P): Plasmid 35S::GmNAC004; (-): H2O; (đc): Cây ĐT22 không chuyển gen; (1-5): Cây chuyển gen 35S::GmNAC004 thế hệ T0.
Số lượng mẫu biến nạp
Cây đậu tương chuyển gen T0 sống sót sau
phun basta
Kết quả phân tích PCRdương tính Hiệu suất chuyển
gen (%)Gen bar Gen 35S::GmNAC004
3029 18 18 18 0,59

36
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Nhóm nghiên cứu đã tiến hành gieo trồng hạt T1 của 18 dòng T0 dương tính PCR để sàng lọc bằng phương pháp phun thuốc diệt cỏ basta. Kết quả ban đầu cho thấy cả 18 dòng đều có sự phân ly về kiểu gen. Sau khi phân tích PCR cây sống sót sau khi phun basta, kết quả cho thấy 18 dòng đều có sự biểu hiện của gen chuyển ở thế hệ T1 (Hình 6, 7).
Hình 6. Kết quả điện di sản phẩm PCR gen bar của các cây đậu tương chuyển cấu trúc gen
35S::GmNAC004 thế hệ T1
Ghi chú: (M): Marker 1kb plus genruler; (-): H2O; (P4): Plasmid 35S/ GmNAC004; (1-9): Cây chuyển gen 35S::GmNAC004 thế hệ T1; (đc): Cây không chuyển gen ĐT22.
Hình 7. Kết quả điện di sản phẩm PCR gen 35S::GmNAC004 của các cây đậu tương
chuyển gen thế hệ T1
Ghi chú: (M): Marker 1kb plus gen ruler;(P4): Plasmid 35S/ GmNAC004; (-): H2O; (1-6): Cây chuyển gen 35S::GmNAC004 thế hệ T1.
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luậnKết quả nghiên cứu biến nạp cấu trúc gen
35S::GmNAC004 vào giống đậu tương ĐT22 thông qua chủng khuẩn A. tumefaciens EHA101 mang vector pZY101::25S::GmNAC004 đã thu được 57 cây đậu tương chuyển gen thế hệ T0 sống sót sau quá trình chọn lọc và đưa ra môi trường tự nhiên. Sau khi sàng lọc bằng phun basta, thu được 18 cây sống
sót. Kết quả phân tích sinh học phân tử đã xác định được 18 dòng có kết quả PCR dương tính với gen đích ở thế hệ T0 với hiệu suất chuyển gen đạt 0,59%, 18 dòng này đều có biểu hiện gen ở thế hệ T1.
4.2. Đề nghịTiếp tục thực hiện các thí nghiệm phân tích sinh
học phân tử, chọn lọc dòng chuyển gen đồng hợp để tiến tới đánh giá khả năng chịu hạn ở các thế hệ tiếp theo của 18 dòng chuyển gen thu được.
LỜI CẢM ƠNNghiên cứu này được hỗ trợ kinh phí từ Chương
trình Khoa học và Công nghệ Độc lập cấp Nhà nước của Bộ Khoa học và Công nghệ theo Hợp đồng số 03/2012/HĐ-ĐTĐL.
TÀI LIỆU THAM KHẢO Nguyễn Văn Đồng, Nguyễn Mai Hương và Nguyễn
Hữu Kiên, 2012. Nghiên cứu quy trình biến nạp gen vào giống đậu tương ĐT22 thông qua Agrobacterium tumefaciens. Tạp chí Khoa học và Công nghệ Việt Nam, 9 (39), tr. 119-124.
Nguyễn Văn Đồng, Nguyễn Anh Vũ, Lê Thị Mai Hương, Nguyễn Trung Anh, 2017. Nghiên cứu chuyển gen GmNAC004 vào giống đậu tương ĐT22 thông qua vi khuẩn Agrobacterium. Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam, 3 (76), tr. 13-17.
Doyle J.J, Doyle J.L, 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin, 19:11-15.
Tran L. S., T. N. Quach, S. K. Guttikonda, D. L. Aldrich, R. Kumar, A. Neelakandan, B. Valliyodan and H. T Nguyen, 2009. Molecular characterization of stress-inducible GmNAC genes in soybean. Division ofPlant Sciences. Mol Genet Genomics, 281(6): 647-664.
Tran L.S., Nishiyama R., Yamaguchi-Shinozaki K., Shinozaki K, 2010. Potential utilization of NAC transcription factors to enhance abiotic stress tolerance in plants by biotechnological approach. GM Crops, 1(1): 32-39.
Truyen N. Quach, Lam-Son Phan Tran, Babu Valliyodan, Hanh TM. Nguyen, Rajesh Kumar, Anjanasree K. Neelakandan, Satish Kumar Guttikonda, Robert E. Sharp, Henry T. Nguyen, 2014. Functional Analysis of Water Stress-Responsive Soybean GmNAC003 and GmNAC004 Transcription Factors in Lateral Root Development in Arabidopsis, PLOS ONE, 9(1): e84886.

37
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Research of 35S::GmNAC004 gene structure transfer into DT22 soybean variety using A. tumefaciens EHA101
Nguyen Van Dong, Nguyen Anh Vu, Le Thi Mai Huong, Nguyen Trung Anh
AbstractIn this study, the full coding sequence of GmNAC004 gene was amplified and cloned into pZY101 vector under the control of the 35S promoter. The 35S:GmNAC004 construct was transformed into DT22 soybean variety via A. tumefaciens EHA101. The results revealed that the rate of multiplication of shoots was 69.56% and the survival rate was 3.84% after selecting on shoot elongation medium with glufosinate. Additionally, the result showed that 18 transgenic lines were herbicide resistant. Taken together, all the transgenic lines were also positive with specific primer by PCR analysis.Keywords: Agrobacterium tumefaciens, transformation, 35S::GmNAC004
Ngày nhận bài: 15/9/2018Ngày phản biện: 21/9/2018
Người phản biện: PGS.TS. Nguyễn Thanh HảiNgày duyệt đăng: 15/10/2018
1 Viện Di truyền Nông nghiệp; 2 Vụ Khoa học và Công nghệ, Bộ Công thương
TẠO DÒNG VÀ BIỂU HIỆN ENZYME ENDOLYSIN TÁI TỔ HỢP TRONG Escherichia coli TỪ STAPHYLOCOCCAL BACTERIOPHAGES NA6
Nguyễn Thị Hồng Hải1, Khuất Hữu Trung1, Trần Đăng Khánh1, Lê Thị Hằng1, Phạm Thị Lý Thu1, Đặng Tất Thành2
TÓM TẮTEndolysins (tên gọi khác là lysins) là các enzyme thủy phân có nguồn gốc từ thực khuẩn thể (bacteriophage) có
tác dụng phá hủy peptidoglycan của thành tế bào vi khuẩn trong giai đoạn cuối chu trình sinh sản của thực khuẩn thể. Dòng thực khuẩn thể vi khuẩn Staphylococcus NA6 được phân lập từ mâu sữa tươi nguyên liệu cho thấy khả năng ức chế vi khuẩn gây bệnh Staphylococcus aureus cao. Gen (LysSA) mã hóa enzyme endolysin của thực khuẩn thể NA6 được tạo dòng và tối ưu hóa mã bộ ba biểu hiện enzyme endolysin trong vi khuẩn E.coli. Sau đó toàn bộ đoạn DNA được chuyển vào vector biểu hiện pET21b(+) trong vi khuẩn E. coli BL21. Gen LysSA mã hóa protein endolysin (LysSA) có khối lượng phân tử được tính toán khoảng 50 kDa. Endolysin tái tổ hợp tách chiết được có hoạt tính kháng cao với vi khuẩn gây bệnh S. aureus.
Từ khóa: Escherichia coli, Endolysin, Staphylococcus aureus
I. ĐẶT VẤN ĐỀStaphylococcus aureus là tụ cầu khuẩn có khả
năng sản sinh ra các độc tố gây nên sự ngộ độc thực phẩm, một trong những nguyên nhân phổ biến nhất gây ra bệnh viêm dạ dày, ruột trên toàn thế giới. Staphylococcus aureus cũng là tác nhân thường xuyên gây nhiễm trùng cận lâm sàng ở bò sữa và đặc biệt là bệnh viêm vú - bệnh gây tử vong và phổ biến nhất ở bò sữa. Vì vậy, sự có mặt của S. aureus là mối nguy cơ lớn cho sự an toàn và chất lượng của các loại sữa, phomai truyền thống.
Endolysin có nguồn gốc từ thực khuẩn thể có tác dụng phá hủy peptidoglycan của thành tế bào vi khuẩn trong giai đoạn cuối chu trình sinh sản của thực khuẩn thể. Các enzyme này làm giảm độ mạnh của thành tế bào dân đến dung giải tế bào và
giải phóng ra các thực khuẩn thể thế hệ tiếp theo. Vì vậy, khi có mặt endolysin chúng có thể phân hủy lớp peptidoglycan của thành tế bào của mọi vi sinh vật khi tiếp xúc, dân đến các vi sinh vật sẽ bị chết. Bên cạnh sự phân giải từ bên trong, các endolysin từ thực khuẩn thể của các vật chủ Gram dương vân có thể phân giải nhanh chóng vi khuẩn khi chúng ở ngoại bào (Loessner, 2005). Giống như chất kháng vi khuẩn tiềm năng, endolysin có phương thức hoạt động riêng biệt, có tính chuyên biệt cao, hoạt động chống lại các vi khuẩn kể cả các loại nhạy cảm với thuốc kháng sinh (Borysowski et al., 2006). Ngoài ra, các endolysin tái tổ hợp được báo cáo là ức chế nhiều loại vi khuẩn gây bệnh và được khẳng định như một nguồn kháng vi sinh vật thay thế để điều trị các bệnh nhiễm khuẩn do vi khuẩn Gram dương gây ra (Loessner, 2005). Nhờ đó mà endolysin được

38
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
xem như một chất kháng sinh thực phẩm lý tưởng có tính tác động chọn lọc. Trong công nghiệp lên men, endolysin như Ply511 có thể được sử dụng để kiểm soát vi khuẩn Listeria monocytogenes làm hỏng sữa (Gaeng et al., 2000) hoặc sử dụng lysine CTP1 để kiểm soát vi khuẩn clostridium trong quá trình chế biến phomat (Mayer et al., 2010). Trong chế biến sữa, endolysin LysH5 từ thực khuẩn thể của vi khuẩn Staphylococcus LysH5 có thể tiêu diệt vi khuẩn S. aureus trong sữa tươi nguyên liệu sau 4 giờ ở 37oC (Obeso et al., 2008). Ngoài ra, endolysin có thể kiểm soát vi khuẩn gây bệnh viêm vú ở tuyến vú của gia súc lấy sữa, phòng trừ nhiễm chéo thực phẩm trong quá trình lấy sữa và giết mổ gia súc như endolysin Ply700 và LysH5 kiểm soát vi khuẩn gây bệnh Streptococcus uberis và S. aureus (Celia et al., 2008; Obeso et al., 2008).
Được phân lập từ mâu sữa tươi nguyên liệu thu thập từ Nghệ An, dòng thực khuẩn thể vi khuẩn Staphylococcus NA6 có khả năng ức chế vi khuẩn gây bệnh S. aureus cao nhất. Sau khi giải trình tự gen, đã tiến hành nối và kiểm tra biểu hiện đa hình gen endolysin LysSA của thực khuẩn thể vi khuẩn Staphylococcus NA6 vào Escherichia coli. Đồng thời, nghiên cứu biểu hiện protein endolysin tái tổ hợp và đánh giá hoạt tính kháng với S. aureus.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưuGen mã hóa endolysin của thực khuẩn thể S.
aureus LysSA được tách dòng trong vector tách dòng pCRTM4–TOPO (Invitrogen). Plasmid pET21b(+) của hãng Novogen được dùng làm vector biểu hiện gen mã hóa endolysin. Tế bào E. coli DH5α được dùng cho mục đích tách dòng gen. Tế bào E. coli BL21 dùng để biểu hiện protein.
Enzyme cắt giới hạn NdeI, XhoI được dùng để xử lí vector tách dòng và vector biểu hiện. Cặp mồi T7 Terminator (GCTAGTTATTGCTCAGCGG) và T7 Promoter (TAATACGACTCACTATAGGG) được dùng trong PCR kiểm tra các sản phẩm ghép nối và vi khuẩn mang gen tái tổ hợp sau biến nạp.
2.2. Phương pháp nghiên cưu
2.2.1. Thiết kế vector biểu hiện pET21b – LysSA Trình tự gen LysSA mã hóa enzyme Endolysin
(được phân lập từ thực khuẩn thể NA6) được tách dòng trong vector pCRTM4–TOPO (Invitrogen) và được gửi sang Canada để tối ưu hóa mã bộ ba biểu hiện protein Endolysin trong vi khuẩn E.coli. Sau đó, vector tái tổ hợp và vector biểu hiện pET21b(+)
được xử lý đồng thời bằng 2 enzyme cắt giới hạn là NdeI và XhoI và được nối lại với nhau bằng enzyme T4 DNA ligase (Sigma) để tạo thành tổ hợp pLysSA. Sản phẩm ghép nối được biến nạp vào tế bào khả biến E. coli BL21 và nuôi trên môi trường LB agar bổ sung kháng sinh ampicillin ở nồng độ 100 µg/ml.
2.2.2. Biểu hiện protein tái tổ hợpTế bào E. coli BL21 mang plasmid tái tổ hợp
pET21b(+) – LysSA được nuôi lắc trên môi trường Luria Broth có bổ sung ampicillin nồng độ 100 µg/ml, lắc 200 vòng/phút qua đêm ở 37oC. Sau đó 0,5 ml dịch nuôi cấy được chuyển qua 50 ml môi trường LB chứa ampicillin 100 µg/ml và được nuôi lắc ở 37oC đến khi mật độ quang (OD 600 nm) đạt 0,6 - 0,8. Chất cảm ứng IPTG được thêm vào đến nồng độ cuối cùng đạt 1 mM, dịch nuôi cấy được tiếp tục nuôi lắc trong 4 h. Tế bào vi khuẩn được thu bằng ly tâm 6.000 vòng /phút trong 6 phút tại 4oC và rửa trong Lysis buffer (Tris 50 mM pH = 7,5; NaCl 100 mM; MgSO4 8 mM) sau đó được phá vỡ bằng sóng siêu âm. Protein được thu bằng ly tâm 13.000 vòng/phút trong 30 phút tại 4oC và loại bỏ cặn tế bào (dịch enzyme endolysin). Sau đó protein được biến tính và phân tích bằng SDS-PAGE 12%.
2.2.3. Hoạt tính của enzyme endolysinVi khuẩn chỉ thị S. aureus được nuôi cấy trên môi
trường LB lỏng ở 37oC cho đến khi OD.600nm = 0,05 - 0,1 (103 - 106 cfu/ml). Chuẩn bị môi trường LB bán lỏng có chứa vi khuẩn với nồng độ 103 cfu/ml. Đục thạch thành các lỗ nhỏ, đều nhau. Nhỏ 100 µl dịch có chứa enzyme endolysin đã được pha loãng theo cấp số 2 vào mỗi giếng. Để đĩa trong tủ lạnh trong 1 giờ, sau đó chuyển sang tủ ấm 30oC nuôi qua đêm và quan sát vòng vô khuẩn. Hoạt tính của endolysin được biểu thị bằng đơn vị units trên 1 ml (U/ml) và được tính theo công thức: (1000 ˟ 25-1) ˟ (D-1). Trong đó: D là độ pha loãng cao nhất mà ở đó không có sự phát triển của chủng chỉ thị S. aureus.
2.3. Thời gian và địa điểm nghiên cưu Nghiên cứu được thực hiện từ năm 2016 -
2018 tại Viện Di truyền Nông nghiệp và Tập đoàn SyntBioLab, Canada.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Thiết kế vector biểu hiện pET21b – LysSAGen mã hóa endolysin LysSa có kích thước 1443
bp. Để thuận lợi cho việc đưa trình tự gen vào vector biểu hiện, trình tự nhận biết của 2 enzyme XhoI và NdeI được thêm vào 2 đầu đoạn gen đích, kích thước gen tăng lên là 1452 bp. Vector tách dòng

39
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
pCR-LysSa và vector pET21b (+) sau khi xử lí bằng XhoI và NdeI được phân tách trên gel agarose 1%. Kích thước sản phẩm cắt giới hạn lần lượt là 1444 bp (gen LysSa), khoảng 3,9 kb (vector tách dòng) và 5366 bp (vector pET21 (+)). Sản phẩm cắt của gen LysSa và pET21b(+) được tinh sạch bằng bộ kit Gel
Extraction Kit (Fermentas) và được nối với nhau bằng enzyme T4 ligase để tạo thành vector tái tổ hợp pLysSa. Sản phẩm nối được kiểm tra bằng PCR sử dụng cặp mồi T7 promoter/T7 terminator. Kết quả thu được vạch băng mong muốn với kích thước 1668 bp (Hình 2).
Để tìm được dòng vi khuẩn E. coli biểu hiện Endolysin hiệu quả, plasmid tái tổ hợp pLysSA được biến nạp vào 3 dòng E. coli khác nhau là Tuner, Rosetta (Novagen) và BL21-DE3 bằng kĩ thuật xung điện. Các khuẩn lạc mọc trên môi trường LB agar/ampicillin được chọn ngâu nhiên để kiểm tra sự có mặt của vector tái tổ hợp bằng PCR với cặp mồi T7 promoter/T7 terminator (Hình 3).
Kết quả cho thấy tất cả các giếng đều xuất hiện vạch băng tương ứng với kích thước của vector pLysSA là 1688 bp. Như vậy, nghiên cứu này đã biến nạp thành công vector pLysSA vào cả ba dòng vi khuẩn nhận. Sau đó, chúng tôi dùng dòng E.coli BL21 DE3 để tiến hành biểu hiện protein Endolysin.
Hình 1. Kết quả cắt vector tách dòng pCR-LysSA bằng enzyme cắt giới hạn XhoI và NdeI
Ghi chú: M: Marker 1 kb DNA plus (Invitrogen); 1: Sản phẩm cắt của vector tách dòng pCR-LysSA bằng enzyme cắt giới hạn XhoI và NdeI.
Hình 3. Kết quả điện di sản phẩm PCR khuẩn lạc của 3 dòng E.coli sau khi được biến nạp vector pLysSA,
bằng cặp mồi T7 prom/termGhi chú: M. marker 1kb plus DNA (Invitrogen); 1, 2 -
Tuner + pLysSa; 3, 4 - Rosetta + pLysSa; 5, 6 - BL21DE3 + pLysSa; 7 - vector pLysSa (đối chứng).
Hình 2. Kết quả kiểm tra sản phẩm nối vector pET21b(+) và gen LysSA
bằng cặp mồi T7prom/T7 termGhi chú: M: Marker 1 kb DNA plus (Invitrogen);
1: pET21b(+) / LysSA.
Hình 4. Kiểm tra sự biểu hiện protein Endolysin từ dòng E.coli BL21 mang plasmide tái tổ hợp pLysSA
bằng điện di SDS-PAGE (12%)Ghi chú: M: thang protein chuẩn (Biorad); 1: chủng đối
chứng E.coli BL21/không cảm ứng; 2: E. coli BL21/pLysSA/không cảm ứng; 3: E. coli BL21/pLysSA/cảm ứng IPTG.

40
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
3.2. Biểu hiện protein tái tổ hợpTế bào E. coli BL21 mang vector tái tổ hợp pLysSA
được nuôi lắc trong môi trường LB broth có bổ sung kháng sinh Ampicillin ở 37oC và cảm ứng bằng IPTG 1mM (khi OD600 nm đạt 0,6, sinh khối được thu sau 4 h. Protein được tách ra khỏi tế bào E. coli bằng sóng siêu âm và được kiểm tra trên gel polyacrylamide 12% (Hình 4). Kết quả điện di cho thấy E. coli BL21 mang vector tái tổ hợp khi cảm ứng bằng IPTG, protein tái tổ hợp được thấy rõ và có kích thước khoảng hơn 50 kDa (giếng 3) trong khi trường hợp không bổ sung chất cảm ứng thì không xuất hiện protein tái tổ hợp (giếng 1, 2).
3.3. Kiểm tra khả năng kháng vi khuẩn S. aureus của endolysin tái tổ hợp
Sau khi thu được enzyme endolysin như mô tả ở mục 2.2.2, tiến hành đánh giá hoạt tính kháng với vi khuẩn S. aureus bằng phương pháp đục lỗ thạch. Hoạt tính của endolysin được đánh giá thông qua chỉ số U/ml (unit per milliliter) bằng nghịch đảo của độ pha loãng lớn nhất mà tại đó, 100 µl dịch endolysin vân thể hiện khả năng ức chế vi sinh vật chỉ thị.
Theo kết quả trên hình 5, endolysin có khả năng kháng vi khuẩn S. aureus và độ hòa loãng cao nhất mà tại đó còn xuất hiện vòng kháng khuẩn là 1:16, khi đó hoạt độ của endolysin tái tổ hợp ước tính khoảng 15 U/ml.
Hình 5. Kiểm tra hoạt tính của endolysin tái tổ hợp thu được sau biểu hiện
Ghi chú: 1: 50 µl dung dịch TFA 0,1% (Đối chứng); 2 - 7: 50 µl dịch endolysin sau khi tinh sạch một phần tại các độ pha loãng 1:2, 1:4, 1:8, 1:16, 1:32, 1:64.
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luậnĐã thiết kế thành công vector tái tổ hợp mang
gen LysSA mã hóa enzyme Endolysin được phân lập từ thực khuẩn thể NA6 (pLysSA) và được biểu hiện trong tế bào vi khuẩn E. coli BL21. Kết quả này sẽ là tiền đề cho những nghiên cứu tiếp theo nhằm sản xuất enzyme endolysin tái tổ hợp ứng dụng thực tế trong bảo quản sữa.
4.2. Đề nghịTiếp tục nghiên cứu các điều kiện nhân nuôi
chủng tái tổ hợp E. coli BL21 mang gen LysSA, tách chiết và tinh sạch enzyme endolysin để ứng dụng trong bảo quản sữa tươi nguyên liệu và sữa tươi thanh trùng.
LỜI CẢM ƠNNghiên cứu này được hỗ trợ kinh phí từ Đề án
phát triển và ứng dụng công nghệ sinh học trong lĩnh vực công nghệ chế biến đến năm 2020 theo Hợp đồng số 06/HĐ-ĐT.06.16/CNSHCB.
TÀI LIỆU THAM KHẢO Borysowski J., Weber-Dabrowska, B., Gorski, A.,
2006. Bacteriophage endolysins as a novel class of antibacterial agents. Experimental Biology and Medicine, 231, 366-377.
Celia, L. K., Nelson, D., & Kerr, D. E., 2008. Characterization of a bacteriophage lysin (Ply700) from Streptococcus uberis. Veterinary Microbiology, 130, 107-117.
Gaeng, S., Scherer, S., Neve, H., & Loessner, M. J., 2000. Gene cloning and expression and secretion of Listeria monocytogenes bacteriophage-lytic enzymes in Lactococcus lactis. Applied and Environmental Microbiology, 66, 2951-2958.
Loessner, M.J., 2005. Bacteriophage endolysins--current state of research and applications. Current Opinion in Microbiology, 8, 480-487.
Mayer, M. J., Payne, J., Gasson, M. J., & Narbad, A., 2010. Genomic sequence and characterization of the virulent bacteriophage phiCTP1 from Clostridium tyrobutyricum and heterologous expression of its endolysin. Applied and Environmental Microbiology, 76, 5415-5422.
Obeso, J. M., Martinez, B., Rodriguez, A., & Garcia, P., 2008. Lytic activity of the recombinant staphylococcal bacteriophage PhiH5 endolysin active against Staphylococcus aureus in milk. International Journal of Food Microbiology, 128, 212-218.

41
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Cloning and expression in Escherichia coli of the recombinant endolysin from Staphylococcal bacteriophages NA6
Nguyen Thị Hong Hai, Khuat Huu Trung, Tran Dang Khanh,Le Thi Hang, Pham Thi Ly Thu and Dang Tat Thanh
AbstractEndolysins (or lysins) are bacteriophage-encoded lytic enzymes that break down the peptidoglycan of the bacterial cell wall during the terminal stage of the bacteriophage reproduction cycle. Bacteriophage NA6 was isolated from raw milk showed good capacity against bacteria Staphylococcus aureus. The endolysin gene (LysSA) from genome of Staphylococcus aureus NA6 was cloned, codon-usage optimized for expression in E.coli and characterized. Then optimized sequence LysSA was moved on vector pET21b (+) in order to get enzyme expression induction in E.coli BL21 with protein molecular mass of 50 kDa. Testing enzyme lytic activity showed that recombinant endolysin had antibacterial activity against Staphylococcus aureus.Keywords: Escherichia coli, Endolysin, Staphylococcus aureu
Ngày nhận bài: 22/9/2018Ngày phản biện: 27/9/2018
Người phản biện: TS. Huỳnh Ngọc Thanh TâmNgày duyệt đăng: 15/10/2018
1 Viện Di truyền Nông nghiệp; 2 Đại học Thủ đô; 3 Học viện Nông nghiệp Việt Nam
TẦM SOÁT VÀ THIẾT KẾ MARKER CHỨC NĂNG XÁC ĐỊNH CANDIDATE GEN KHÁNG ĐẠO ÔN PIT Ở MỘT SỐ GIỐNG LÚA BẢN ĐỊA CỦA VIỆT NAM
Nguyễn Trường Khoa1, Nguyễn Thúy Điệp1, Nguyễn Thái Dương1, Nguyễn Như Toản2 , Phương Hữu Pha3, Đặng Thị Thanh Hà1,
Kiều Thị Dung1, Trần Thị Thúy1, Trần Đăng Khánh1, Khuất Hữu Trung1
TÓM TẮTGen kháng đạo ôn Pit đã được xác định có trong giống lúa Indica (Tjahaja) của Indonesia, có phổ kháng rộng
(Kiyosawa et al.,1972). Gen Pit được phân lập và xác định thuộc họ CC-NBS-LRR (Hayashi et al., 2009). Nhiều nghiên cứu cho thấy rằng chỉ thị phân tử cho phép xác định các gen kháng trong nguồn gen lúa và có nhiều ưu thế trong chọn tạo giống lúa kháng do nó có thể đánh giá được sự tồn tại của các gen mà không cần lây nhiễm nhân tạo. Các marker chức năng được thiết kế từ các đa hình trong trình tự gen và không bị ảnh hưởng bởi sự thay đổi alen phi chức năng, đặc biệt có thể xác định các gen riêng lẻ. Dựa vào trình tự hệ gen của 17 giống lúa bản địa được giải mã, trong nghiên cứu này đã tầm soát và xác định được các candidate gen Pit ở 17 giống lúa nghiên cứu. Trong đó, có 9 giống có tổng số nucleotide ít hơn 3 nucleotide và 8 giống có tổng số nucleotide ít hơn 20 nucleotide so với gen tham chiếu LOC_Os01g05620. Đồng thời, đã thiết kế được marker Ins_Pit_17 dựa trên sự thêm/bớt đoạn ADN của các candidate gen Pit trong các giống lúa bản địa. Kết quả cho thấy: Có thể sử dụng marker chức năng đã thiết kế để nghiên cứu dự đoán chức năng gen Pit và ứng dụng trong các chương trình chọn tạo giống lúa.
Từ khóa: Candidate gen, Pit, kháng đạo ôn, lúa, marker
I. ĐẶT VẤN ĐỀ Bệnh đạo ôn do nấm Magnaporthe oryzae gây ra
là một trong những bệnh phổ biến nhất và tàn phá ảnh hưởng lớn đến năng suất lúa gạo. Quản lý bệnh đạo ôn lúa thông qua sức đề kháng của vật chủ là một hướng đầy hứa hẹn của chương trình Quản lý bệnh hại tổng hợp (IDM). Cho đến nay có khoảng 102 gen kháng đạo ôn đã được xác định (Wu et al., 2016; Vasudevan et al., 2015). Trong số đó 27 gen đã được nhân bản (clonning) đó là: Pib, Pb1, Pita, Pi9, Pi2, Pizt, Pid2, Pi33, Pii, Pi36, Pi37, Pikm, Pit, Pi5, Pid3, Pid3-A4, Pi54, Pish, Pik, Pikp, Pia, PiCO39,
Pi25, Pi1, Pi21, P50 và Pi65 (t) (Sharma et al., 2012; Devanna et al., 2014; Xiao et al., 2016). Nhiều gen kháng được xác định và đã được phân loại thành 8 nhóm khác nhau (Sharma et al., 2012). Đa số locus liên quan đến khả năng kháng bệnh đạo ôn đã được tìm thấy nằm trên nhiễm sắc thể số 11 của cây lúa dựa trên các nghiên cứu tương quan toàn bộ nhiễm sắc thể (Wang et al., 2013). Mặc dù một số locus kháng bệnh đạo ôn đã được xác định nhưng chỉ có một vài trong số chúng được ứng dụng trong lai tạo giống để quản lý bệnh đạo ôn ở một số quốc gia (Singh et al., 2014).

42
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Hiện nay, các nghiên cứu khám phá các biến thể di truyền từ các nguồn gen kháng (lúa hoang dại và giống lúa canh tác) ở nhiều loài cây trồng để tìm ra điểm chung nhất đang được các nhà khoa học chú trọng. Một trong những phương pháp được sử dụng rộng rãi nhất để xác định các biến thể là sử dụng kỹ thuật dựa trên phản ứng chuỗi polymerase (PCR) để khuếch đại các alen khả thi (homolog) từ nhóm gen được gọi là khai thác alen. Gần đây dựa vào các alen có tính kháng bệnh đạo ôn được báo cáo, phân lập tách dòng từ các giống lúa hoang dại và giống lúa trồng đã giúp các nhà chọn giống chọn tạo các giống lúa mới có khả năng kháng bệnh đạo ôn cao. (Geng et al., 2008; Huang et al., 2008).
Gen Pit kháng đạo ôn ở lúa được xác định trong giống lúa Tjahaja của Indonesian, giống lúa indica này có phổ kháng bệnh rộng (Kiyosawa et al., 1972). Trong 1 nghiên cứu trước đây, nhóm tác giả đã phân lập Pit và chỉ ra rằng gen Pit thuộc họ CC-NBS-LRR (Hayashi et al., 2009). So sánh trình tự của alen Pit giữa giống kháng K59 và giống mân cảm Nippon-bare cho thấy alen liên quan đến tính kháng đạo ôn có chứa 4 amino acid thay thế, 1 DNA nhảy dDart và 1 đoạn lặp dài cuối (LTR)-retrotransposon, tên là Renovator, chèn vào vùng promoter.
Trong nghiên cứu này, dựa vào kết quả tầm soát candidate gen Pit có trong một số giống lúa bản địa của Việt Nam từ đó phát triển marker chức năng để xác định candidate gen Pit. Sử dụng kết quả này trong nghiên cứu chức năng gen và để sàng lọc các dòng/giống lúa phục vụ cho công tác chọn tạo giống kháng bệnh đạo ôn.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưuCơ sở dữ liệu trình tự genome 17 giống lúa bản
địa của Việt Nam đã được giải mã.
Bảng 1. Bảng vật liệu nghiên cứu
2.2. Phương pháp nghiên cưu
2.2.1. Phương pháp tầm soát gen Pit và thiết kế mồi đặc hiệu dựa trên trình tự genome
- Lắp ráp và gọi SNP sử dụng phần mềm NextGENe_V2.4.1.1.
- Trình tự các nucleotide được so sánh và phân tích sử dụng phần mềm ClustalW2 (http://www.ebi.ac.uk/Tools/msa/clustalw2/); Cặp mồi đặc hiệu được thiết kế bằng phần mềm Primer 3,0 (http://primer3plus.com/web_3.0.0/primer3web_input.htm) và phần mềm xây dựng cây phả hệ MEGA6.0 cho hệ điều hành Windows (http://www.megasoftware.net/mega.php).
2.2.2. Phương pháp kiểm tra candidate gen kháng Pit- Mâu lá lúa được thu thập và tách chiết ADN
tổng số theo phương pháp CTAB của (Obara et al., 1998) có cải tiến.
- Phản ứng PCR được tiến hành trên máy Veriti 96 well Thermal cycler. Tổng thể tích phản ứng là 15 µl, bao gồm: 5 µl ADN (nông độ 10 ng); 0,15 µM mồi; 0,2 mM dNTPs; 1X Buffer PCR; 2,5 mM MgCl2 và 0,25 đơn vị Taq polymerase.
- Sản phẩm PCR được điện di trên gel polyacrylamide 6,0% hoặc trên gel agarose và được phát hiện dưới tia cực tím bằng phương pháp nhuộm ethidium bromide.
2.3. Thời gian và địa điểm nghiên cưu Nghiên cứu được thực hiện từ tháng 3 đến tháng
8/2018 tại Viện Di truyền Nông nghiệp.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Kết quả tầm soát gen Pit kháng đạo ôn dựa trên dữ liệu trình tự genome của các giống lúa bản địa đã được giải mã
Trình tự gen của gen Pit được lấy từ cơ sở dữ liệu của NCBI (https: //www.ncbi.nlm. nih.gov) và chuỗi tham chiếu được lấy từ cơ sở dữ liệu công bố (http://rice.plantbiology.msu.edu/cgi-bin/sequence_display.cgi?orf=LOC_Os01g05620) là LOC_Os01g05620,vùng CDS (Coding DNA Sequence) có 2970 nucleotide mã hóa cho 989 amino acid. Dựa vào trình tự genome của 17 giống lúa bản địa đã giải mã, tiến hành tìm trình tự tương đồng của 17 giống lúa đã giải mã và so sánh với locus gen Pit đã được công bố.
TT Tên giống TT Tên giống1 Khấu điển lư 10 Lúa gốc đỏ2 Thơm Lài 11 Tám xoan Hải Hậu3 OM6377 12 Hom râu4 Nếp mặn 13 Khẩu Liến5 Tốc lùn 14 Tám xoan Bắc Ninh6 Ble te lo 15 Chiêm nhỡ Bắc Ninh 27 Khấu mặc buộc 16 Nếp lùn8 Nếp mèo nương 17 Nàng quớt biển9 Tép Thái Bình
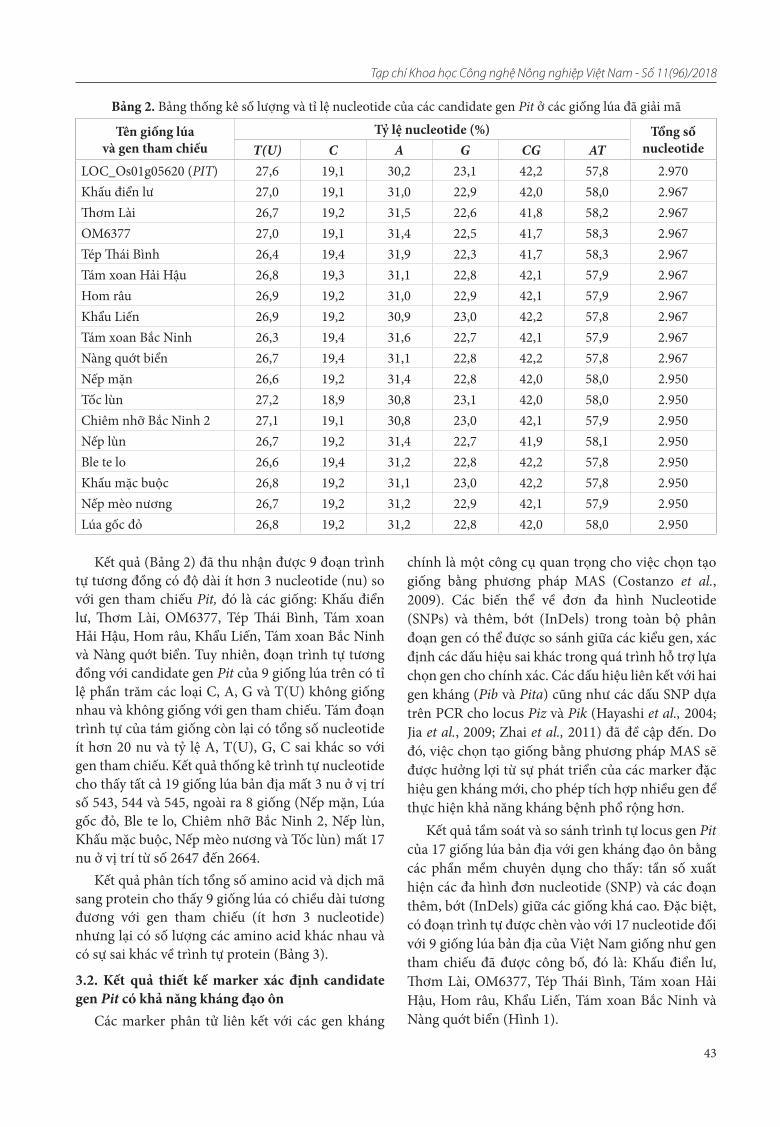
43
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Bảng 2. Bảng thống kê số lượng và tỉ lệ nucleotide của các candidate gen Pit ở các giống lúa đã giải mã
Kết quả (Bảng 2) đã thu nhận được 9 đoạn trình tự tương đồng có độ dài ít hơn 3 nucleotide (nu) so với gen tham chiếu Pit, đó là các giống: Khấu điển lư, Thơm Lài, OM6377, Tép Thái Bình, Tám xoan Hải Hậu, Hom râu, Khẩu Liến, Tám xoan Bắc Ninh và Nàng quớt biển. Tuy nhiên, đoạn trình tự tương đồng với candidate gen Pit của 9 giống lúa trên có tỉ lệ phần trăm các loại C, A, G và T(U) không giống nhau và không giống với gen tham chiếu. Tám đoạn trình tự của tám giống còn lại có tổng số nucleotide ít hơn 20 nu và tỷ lệ A, T(U), G, C sai khác so với gen tham chiếu. Kết quả thống kê trình tự nucleotide cho thấy tất cả 19 giống lúa bản địa mất 3 nu ở vị trí số 543, 544 và 545, ngoài ra 8 giống (Nếp mặn, Lúa gốc đỏ, Ble te lo, Chiêm nhỡ Bắc Ninh 2, Nếp lùn, Khấu mặc buộc, Nếp mèo nương và Tốc lùn) mất 17 nu ở vị trí từ số 2647 đến 2664.
Kết quả phân tích tổng số amino acid và dịch mã sang protein cho thấy 9 giống lúa có chiều dài tương đương với gen tham chiếu (ít hơn 3 nucleotide) nhưng lại có số lượng các amino acid khác nhau và có sự sai khác về trình tự protein (Bảng 3).
3.2. Kết quả thiết kế marker xác định candidate gen Pit có khả năng kháng đạo ôn
Các marker phân tử liên kết với các gen kháng
chính là một công cụ quan trọng cho việc chọn tạo giống bằng phương pháp MAS (Costanzo et al.,2009). Các biến thể về đơn đa hình Nucleotide (SNPs) và thêm, bớt (InDels) trong toàn bộ phân đoạn gen có thể được so sánh giữa các kiểu gen, xác định các dấu hiệu sai khác trong quá trình hỗ trợ lựa chọn gen cho chính xác. Các dấu hiệu liên kết với hai gen kháng (Pib và Pita) cũng như các dấu SNP dựa trên PCR cho locus Piz và Pik (Hayashi et al., 2004; Jia et al., 2009; Zhai et al., 2011) đã đề cập đến. Do đó, việc chọn tạo giống bằng phương pháp MAS sẽ được hưởng lợi từ sự phát triển của các marker đặc hiệu gen kháng mới, cho phép tích hợp nhiều gen để thực hiện khả năng kháng bệnh phổ rộng hơn.
Kết quả tầm soát và so sánh trình tự locus gen Pit của 17 giống lúa bản địa với gen kháng đạo ôn bằng các phần mềm chuyên dụng cho thấy: tần số xuất hiện các đa hình đơn nucleotide (SNP) và các đoạn thêm, bớt (InDels) giữa các giống khá cao. Đặc biệt, có đoạn trình tự được chèn vào với 17 nucleotide đối với 9 giống lúa bản địa của Việt Nam giống như gen tham chiếu đã được công bố, đó là: Khấu điển lư, Thơm Lài, OM6377, Tép Thái Bình, Tám xoan Hải Hậu, Hom râu, Khẩu Liến, Tám xoan Bắc Ninh và Nàng quớt biển (Hình 1).
Tên giống lúa và gen tham chiếu
Tỷ lệ nucleotide (%) Tổng số nucleotideT(U) C A G CG AT
LOC_Os01g05620 (PIT) 27,6 19,1 30,2 23,1 42,2 57,8 2.970Khấu điển lư 27,0 19,1 31,0 22,9 42,0 58,0 2.967Thơm Lài 26,7 19,2 31,5 22,6 41,8 58,2 2.967OM6377 27,0 19,1 31,4 22,5 41,7 58,3 2.967Tép Thái Bình 26,4 19,4 31,9 22,3 41,7 58,3 2.967Tám xoan Hải Hậu 26,8 19,3 31,1 22,8 42,1 57,9 2.967Hom râu 26,9 19,2 31,0 22,9 42,1 57,9 2.967Khẩu Liến 26,9 19,2 30,9 23,0 42,2 57,8 2.967Tám xoan Bắc Ninh 26,3 19,4 31,6 22,7 42,1 57,9 2.967Nàng quớt biển 26,7 19,4 31,1 22,8 42,2 57,8 2.967Nếp mặn 26,6 19,2 31,4 22,8 42,0 58,0 2.950Tốc lùn 27,2 18,9 30,8 23,1 42,0 58,0 2.950Chiêm nhỡ Bắc Ninh 2 27,1 19,1 30,8 23,0 42,1 57,9 2.950Nếp lùn 26,7 19,2 31,4 22,7 41,9 58,1 2.950Ble te lo 26,6 19,4 31,2 22,8 42,2 57,8 2.950Khấu mặc buộc 26,8 19,2 31,1 23,0 42,2 57,8 2.950Nếp mèo nương 26,7 19,2 31,2 22,9 42,1 57,9 2.950Lúa gốc đỏ 26,8 19,2 31,2 22,8 42,0 58,0 2.950

44
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Trong nghiên cứu này nhờ có sự khác biệt của gen Pit giữa các giống lúa đã giải trình tự mà đã phát triển được marker SSLP dựa vào sự thêm/bớt ADN của các alen Pit với marker thiết kế có tên Ins_Pit_17 (Ins_Pit_17F: TCTTAACGACTGCCCAAAACT/Ins_Pit_17R: AGATTA CAGAGATTGGAAATTCTC). Theo tính toán thiết kế và so sánh với gen tham chiếu của thế giới đã công bố, cặp mồi Ins_Pit_17 sẽ nhân lên băng kích thước 180 bp (ở các giống mang candidate gen Pit giống với trình tự thế giới công bố) và băng 163 bp (ở các giống mang candidate gen Pit khác với trình tự gen Pit thế giới công bố).
Sử dụng cặp mồi thiết kế Ins_Pit_17 để kiểm tra
sự đa hình của locus gen kháng Pit với 17 giống lúa trong tập đoàn lúa bản địa (Hình 2). Kết quả thu nhận được thông qua các số liệu và hình ảnh cho thấy 9/17 giống lúa bản địa đã giải mã có sự xuất hiện của băng kích thước 180 bp, đây là 9 giống có đoạn chèn thêm 17 nu và chiều dài gần tương đương như gen tham chiếu. Tám giống còn lại xuất hiện băng với kích thước 163 bp, là các giống không có đoạn chèn thêm 17 nucleotide. Như vậy, cặp mồi này có thể xác định được sự đa hình của candidate gen Pit và có thể sử dụng để xác định sự có mặt của các candidate gen này ở bố, mẹ và các thế hệ con lai phục vụ cho công tác chọn tạo giống lúa có khả năng kháng đạo ôn.
Bảng 3. Bảng thống kê số lượng amino acid của các gen ứng viên Pit ở các giống lúa đã giải mã
Hình 1. Ảnh dóng hàng, dóng cột so sánh trình tự một đoạn gen kháng đạo ôn của một số giống lúa bản địa
Tên giống/ gen tham chiếu Ala Cys Asp Glu Phe Gly His Ile Lys Leu Met Asn Pro Gln Arg Ser Thr Val Trp Tyr Tổng
LOC Os-01g05620(PIT) 41 24 55 67 38 54 28 67 61 143 29 49 40 38 52 65 35 64 16 23 989
Khấu điển lư 40 23 55 68 37 56 29 66 63 143 30 51 40 40 52 62 35 59 16 22 987Thơm Lài 46 23 54 67 37 53 27 70 64 143 30 56 40 38 54 58 36 55 15 21 987OM6377 45 24 54 67 38 51 28 69 65 143 31 55 40 38 53 58 35 57 15 21 987Tép Thái Bình 43 22 53 68 37 55 29 70 67 144 29 57 41 38 51 57 37 53 15 21 987Tám xoan Hải Hậu 41 22 55 67 37 57 29 66 64 144 30 51 40 38 53 61 36 59 15 22 987Hom râu 40 23 54 69 37 56 29 68 62 143 28 52 40 40 52 62 35 59 16 22 987Khẩu Liến 41 23 56 67 37 56 29 67 62 143 29 51 41 40 52 61 34 60 16 22 987Tám xoan Bắc Ninh 42 22 54 69 36 56 29 67 69 142 29 52 41 37 50 63 36 56 15 22 987Nàng quớt biển 42 23 56 67 36 55 28 67 65 142 31 51 41 39 52 63 34 57 16 22 987Nếp mặn 40 22 55 69 36 56 28 65 68 141 30 53 41 37 49 63 34 58 15 21 981Chiêm nhỡ Bắc Ninh 2 41 23 55 67 37 55 27 64 63 142 30 52 41 39 51 62 34 60 16 22 981
Nếp lùn 41 22 55 66 37 56 27 65 69 142 29 53 41 37 50 62 36 57 15 21 981Ble te lo 41 23 56 67 36 55 27 64 65 140 30 51 42 38 51 63 35 59 16 22 981Khấu mặc buộc 40 22 55 68 37 57 28 64 63 142 30 52 41 39 51 62 34 59 15 22 981Nếp mèo nương 41 23 54 70 36 55 27 65 66 140 29 53 41 38 50 64 33 58 16 22 981Tốc lùn 40 22 56 66 37 55 26 63 64 143 29 52 40 38 52 62 35 63 16 21 980Lúa gốc đỏ 41 22 55 67 37 55 28 64 66 142 30 52 41 38 49 62 35 59 16 21 980

45
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
IV. KẾT LUẬNĐã tầm soát, xác định được candidate gen Pit
trong 17 giống lúa đã giải mã, thu nhận được 9 đoạn trình tự tương đồng có độ dài ít hơn 3 nucleotide và được 8 đoạn trình tự tương đồng có độ dài ít hơn 20 nucleotide so với gen tham chiếu đã công bố.
Đã thiết kế được cặp mồi Ins_Pit_17 để nhận dạng candidate gen Pit có trong 17 giống lúa bản địa của Việt Nam phục vụ cho công tác chọn tạo giống.
TÀI LIỆU THAM KHẢOCostanzo, S. and Y. Jia, 2009. Alternatively spliced
transcripts of Pi-ta blast resistance gene in Oryza sativa. Plant Sci., 177 (5): 468-478.
Devanna, N.B., Vijayan J. and Sharma T.R., 2014. The blast resistance gene Pi54of cloned from Oryza officinalis interacts with Avr-Pi54 through its novel non-LRR domains. PLoS One 9:e104840.
Geng X.S., Yang M.Z., Huang X.Q., Cheng Z.Q., Fu J., Sun T. and Li J., 2008. Cloning and analyzing of rice blast resistance gene Pi-ta+ allele from Jinghong erect type of common wild rice (Oryza rufipogon Griff ) in Yunnan. Yi Chuan 30, 109-114. doi: 10.3724/SP.J.1005.2008.00109.
Hayashi K. and H. Yoshida, 2009. Refunctionalization of the ancient rice blast disease resistance gene Pit by the recruitment of a retrotrans-poson as a promoter. Plant J., 57 (3): 413-425.
Hayashi, K., N. Hashimoto, M. Daigen and I. Ashikawa, 2004. Development of PCR-based SNP markers for rice blast resistance genes at the Piz locus. Theor. Appl. Genet., 108, 1212-1220. doi: 10.1007/s00122-003-1553-0.
Huang, C.L., S.Y. Hwang, Y.C. Chiang and T.P. Lin, 2008. Molecular evolution of the Pi-ta gene resistant to rice blast in wild rice (Oryza rufipogon). Genetics, 179, 1527-1538. doi: 10.1534/genetics.108.089805.
Jia, Y., G. Liu, S. Costanzo, S. Lee and Y. Dai, 2009. Current progress on genetic interactions of rice with rice blast and sheath blight fungi. Front. Agric. China 3 (3): 231-239. doi: 10.1007/s11703-009-0062-6.
Kiyosawa, S., 1972. The inheritance of blast resistance transferred from some indica varieties in rice. Bull. Nat. Inst. Agric. Sci., D23:69-96.
Obara-Okeyo, P. and S. Kako, 1998. Genetic diversity and identification of Cymbidium cultivars as measured by random amplified polymorphic DNA (RAPD) markers. Euphytica, 99 (2): 95-101
Sharma T.R., A.K. Rai, S.K. Gupta, J. Vijayan, B.N. Devanna and S. Ray, 2012. Rice blast management through host-plant resistance: retrospect and prospects. Agric. Res., 1 (1): 37-52.
Singh P.K., S. Thakur, R. Rathour, M. Variar, S.K. Prashanthi, A.K. Singh, U.D. Singh, V. Sharma, N.K. Singh and T.R. Sharma, 2014. Transposon-based high sequence diversity in Avr-Pita alleles increases the potential for pathogenicity of Magnaporthe oryzaepopula-tions. Funct. Integr. Genom., 14 (2): 419-429.
Vasudevan K., W. Gruissem and N.K. Bhullar, 2015. Identification of novel alleles of the rice blast resistance genePi54. Sci. Rep. 5:15678.
Wang, X., Lee, S., Wang, J., Ma, J., Bianco, T., Jia, Y., 2013. Current Advances on Genetic Resistance to Rice Blast Disease. In: Rice - Germplasm, Gene and Improvement, pages 195-217.
Wu, Y.Y., He, J.B., Li, A.H., Fang, N.Y., He, W.W., Dang, L.L., Zeng, G.Y., Huang, J., Bao Y.M. and Zhang, H.S., 2016. Population structure analysis and association mapping of blast resistance in indicarice (Oryza sativa L.) landraces. Genet. Mol. Res. Online publication. doi:10.4238/gmr.15038254.
Xiao N., Y. Wu, C. Pan, L. Yu, Y. Chen, Y. Li, Z. Dai, C. Liang and A. Li, 2016. Improving of rice blast resistances in japonica by pyramiding major R genes. Front Plant Sci, 7:1918.
Hình 2. Ảnh điện di sản phẩm PCR đối với cặp mồi Ins_Pit_17 nhận biết candidate gen Pit ở các giống lúa bản địa của Việt Nam (thứ tự các mâu theo bảng 1)

46
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Predicting, identifying and designing functional marker to identify blast resistance candidate gene (Pit) in Vietnamese rice varieties
Nguyen Truong Khoa, Nguyen Thuy Diep, Nguyen Thai Duong, Nguyen Nhu Toan , Phuong Huu Pha, Dang Thi Thanh Ha,
Kieu Thi Dung, Tran Thi Thuy, Tran Dang Khanh, Khuat Huu TrungAbstractThe rice blast resistance gene Pit, which was identified in an Indonesian indica rice variety, Tjahaja, confers broad-spectrum resistance (Kiyosawa, 1972). Pit gene was isolated and revealed that Pit belongs to the CC-NBS-LRR family of resistance genes (Hayashi and Yoshida, 2009). DNA markers that allow for identification of resistance genes in rice germplasm have a great advantage in resistance breeding because they can assess the existence of the genes without laborious inoculation tests. Functional markers, which are designed from functional polymorphisms within the sequence of genes, are unaffected by nonfunctional allelic variation and make it possible to identify an individual gene. In this study, based on the genome sequence databases of 17 Vietnamese rice varieties, we have predicted and identified the candidate gene Pit that involved in blast resistance ability of 17 local rice varieties. In which, 9 homologous sequences had total nucleotides of less than 3 nucleotides and 8 homologous sequences had total nucleotides of less than 20 nucleotides compared to the reference gene LOC_Os01g05620. Simultaneously, we developed Ins_Pit_17 marker derived from InDels segments of candidate gene Pit in local rice varieties. These results indicate that our functional marker should enhance prediction of Pit function and be applicable in rice breeding programs.Keywords: Candidate gene, Pit, blast resistance, rice, marker
Ngày nhận bài: 18/9/2018Ngày phản biện: 23/9/2018
Người phản biện: TS. Phạm Thiên ThànhNgày duyệt đăng: 15/10/2018
THIẾT LẬP PHƯƠNG PHÁP LẤY MÂU LÚA THEO TIÊU CHUẨN ISO/IEC 17025: 2017 PHỤC VỤ THỬ NGHIỆM XÁC ĐỊNH LOCUS GEN
Chu Đức Hà1, Nguyễn Thị Minh Nguyệt1, Nguyễn Thị Nhài1, Nguyễn Bá Ngọc1, Khuất Thị Mai Lương1, Phạm Thị Lý Thu1, Lê Hùng Lĩnh1
TÓM TẮTTiêu chuẩn ISO/IEC 17025 (International Organization for Standardization/ International Electrotechnical
Commission) là một trong những thước đo quan trọng chứng nhận cho phòng thí nghiệm đạt tiêu chuẩn quốc tế. Trong nghiên cứu này, các phương pháp lấy mâu lúa phục vụ thử nghiệm xác định locus gen mục tiêu đã được thiết lập. Cụ thể, hai phương pháp lấy mâu thử nghiệm ngoài đồng ruộng và trong phòng thí nghiệm đã được xây dựng thành công. Trong đó, các bước thu, bảo quản mâu lá lúa trên đồng ruộng và tiếp nhận, bảo quản, chuẩn bị mâu thử nghiệm từ lô mâu hạt giống nhận tại phòng thử nghiệm cũng đã được chú ý. Kết quả của nghiên cứu này có ý nghĩa trong đưa ra quy trình lấy mâu theo tiêu chuẩn ISO/IEC 17025: 2017, từ đó thành lập và vận hành phòng sinh học phân tử giám định gen thực vật đạt chuẩn ISO/IEC 17025:2017.
Từ khóa: ISO, lúa, thử nghiệm, quy trình, lấy mâu
1 Viện Di truyền Nông nghiệp, VAAS
I. ĐẶT VẤN ĐỀISO/IEC 17025 (International Organization for
Standardization/ International Electrotechnical Commission) là tiêu chuẩn về hệ thống quản lý chất lượng, được áp dụng đặc thù cho các lĩnh vực thử nghiệm và hiệu chuẩn do Tổ chức Quốc tế về Tiêu chuẩn hóa thiết lập và ban hành (Honsa and McIntyre, 2003). Được thiết kế nhằm hợp nhất với tiêu chuẩn về hệ thống quản lý chất lượng ISO 9001, tiêu chuẩn ISO/IEC 17025 đưa ra một cách
nghiêm ngặt các quy định để đảm bảo năng lực kỹ thuật, mang lại kết quả đo lường/thử nghiệm đạt độ tin cậy cao và được quốc tế thừa nhận (Yamamoto et al., 2010). Vì vậy, đạt chuẩn ISO/IEC 17025 được xem là một tiêu chuẩn “cứng” để đánh giá khả năng hoạt động của phòng thử nghiệm và hiệu chuẩn kiểu mâu.
Hiện nay, nhu cầu xác định sự có mặt của một số locus gen mục tiêu trong các giống lúa (Oryza sativa) được xem là quan tâm hàng đầu trong các

47
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
chương trình chọn tạo giống nhằm nâng cao tính chống chịu điều kiện bất lợi. Nhiều locus gen kháng với bất lợi sinh học, như bệnh đạo ôn, bạc lá và rầy nâu, và bất lợi phi sinh học, như ngập úng, mặn có thể phát hiện một cách chính xác bằng kỹ thuật sinh học phân tử dựa trên các chỉ thị liên kết chặt với gen kháng. Do vậy, thiết lập quy trình xác định chính xác sự có mặt của gen kháng mục tiêu trong các giống lúa được xem là rất cần thiết. Tuy nhiên, cho đến nay chưa có ghi nhận về phòng thử nghiệm đạt chuẩn ISO/IEC 17025: 2017 đảm nhận chức năng xác định sự có mặt của các locus gen mục tiêu trên các giống lúa ở Việt Nam.
Trong nghiên cứu này, phương pháp lấy mâu thử nghiệm đã được thiết lập làm cơ sở quan trọng cho ban hành Quy trình lấy mâu và quản lý mâu thử theo tiêu chuẩn ISO/IEC 17025: 2017. Mục đích của nghiên cứu này nhằm triển khai một phương pháp lấy mâu tiêu chuẩn đảm bảo tính đại diện của mâu, đồng thời giảm thiểu tối đa lỗi trong quá trình xác định gen mục tiêu.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưuMâu giống lúa cần thử nghiệm sự có mặt/ vắng
mặt của locus gen mục tiêu, gồm lá lúa thu thập trên đồng ruộng hoặc hạt giống lúa do khách hàng cung cấp.
2.2. Phương pháp nghiên cưu
2.2.1. Chuân bị trang thiết bị và dụng cụTúi ziplock, kéo cắt mâu, găng tay y tế, panh kẹp,
thùng lạnh bảo quản mâu và một số đồ dùng cần thiết khác.
2.2.2. Phương pháp tiếp nhận mâuMâu giống có thể được thu thập trên đồng ruộng
hoặc được khách hàng mang đến phòng thí nghiệm. Đối với mâu giống trên đồng ruộng, tiến hành đánh số cây theo thứ tự hàng 1 - 10 - 20 - 30 - 40 - 50 - 60 - 70 - 80 - 90 - 100. Nếu trong hàng có khóm bị thiếu thì đeo thẻ tag đánh dấu và đếm số tiếp tục từ khóm tiếp theo. Đối với lô hạt giống giao nhận tại phòng thử nghiệm, tiến hành kiểm tra tem nhãn và phiếu giao nhận mâu giống trước khi lấy mâu.
2.2.3. Phương pháp thu mâu và chuân bị mâuĐối với mâu lá thu trực tiếp trên đồng ruộng: Thu
100 mâu lá đại diện cho 100 cây của giống lúa cần thử nghiệm. Cây được đánh số và ghi nhãn theo thứ tự. Mâu lá non, sạch bệnh đượ c cắt bằng kéo và cất
trong túi ziplock, ghi nhãn và bảo quản trong thùng lạnh. Mâu được vận chuyển về phòng thí nghiệm và bảo quản ở 4oC.
Đối với lô hạt giống tại phòng thí nghiệm: Phương pháp thu mâu được bên giao thực hiện theo hướng dân của Tiêu chuẩn Việt Nam về Hạt giống lúa nước - Phương pháp thử TCVN 1700:1976 (Bộ Khoa học và Công nghệ, 1986). Mâu giống được kiểm tra tem nhãn, phiếu giao nhận trước khi bảo quản trong tủ mát 4oC. Hạt giống đảm bảo chất lượng được ngâm ủ nảy mầm trên đĩa petri trong điều kiện ẩm, nhiệt độ 37oC (Bộ Khoa học và Công nghệ, 1986).
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Xây dựng phương pháp thu và bảo quản mẫu lá lúa trên đồng ruộng
Phòng Sinh học phân tử Giám định gen thực vật được xây dựng nhằm thử nghiệm các chỉ tiêu đăng ký công nhận theo tiêu chuẩn ISO/IEC 17025: 2017. Các chỉ tiêu thử nghiệm là các locus gen mục tiêu (kháng sâu bệnh, chống chịu với các điều kiện bất lợi của ngoại cảnh, các gen năng suất, chất lượng... trong các giống lúa). Để đạt được mục tiêu trên, việc xây dựng và ban hành phương pháp tiêu chuẩn đối với việc thu mâu và bảo quản mâu là cần thiết nhằm xây dựng được Quy trình Lấy mâu và quản lý mâu thử của phòng thử nghiệm.
Để cung cấp mâu đại diện cho các phân tích thử nghiệm và giảm tối đa lỗi xảy ra trong quá trình xác định, quy trình lấy mâu phải phụ thuộc vào kế hoạch lấy mâu của phòng thử nghiệm. Mặc dù các bước lấy mâu lá trên đồng ruộng khá đơn giản, tuy nhiên quy trình thu mâu cần phải trở thành một tài liệu tiêu chuẩn, từ đó có thể sử dụng cho phòng thử nghiệm một cách chính quy (Bùi Hữu Điền, 2018).
Cụ thể, phương pháp thu và bảo quản mâu lá lúa trên đồng ruộng phục vụ cho Phòng Sinh học phân tử giám định gen thực vật theo tiêu chuẩn ISO/IEC 17025 được xây dựng gồm năm bước chính (Hình 1).
- Bước 1. Chuẩn bị dụng cụ lấy mâu, vật chứa mâu: Các dụng cụ lấy mâu, chia mâu, chứa mâu được chuẩn bị theo nguyên tắc chung: phải sạch trung tính và không làm nhiễm bẩn, nhiễm chéo cho mâu lấy, đồng thời hình dáng, vật liệu chế tạo và kích thước của dụng cụ lấy mâu và chứa mâu phải dựa vào các tiêu chuẩn phù hợp cho mâu lá lúa (Bùi Hữu Điền, 2018). Trong nghiên cứu này, dụng cụ được chuẩn bị bao gồm: kéo y tế, túi ziplock, bút viết kính, găng tay y tế, thùng lạnh bảo quản mâu.

48
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
- Bước 2. Xác định và kiểm tra khu ruộng thu mâu lá: Trước khi lấy mâu giống lúa thử nghiệm, cần xác định và kiểm tra khu ruộng thu mâu lá lúa do khách hàng yêu cầu. Ruộng thu mâu lá là ruộng cấy theo hàng, lúa được cấy 1 dảnh, đảm bảo sạch bệnh, tiện lợi cho việc ghi sơ đồ và đánh dấu khu thu mâu.
- Bước 3. Đánh số và treo nhãn cho cây lấy mâu lá theo hàng: Đối với mỗi mâu giống cần thử nghiệm sẽ tiến hành thu mâu lá trên 100 cây, do đó hàng thu mâu phải được ghi sơ đồ và đánh số số cây theo thứ tự hàng 1 - 10 - 20 - 30 - 40 - 50 - 60 - 70 - 80 - 90, trong đó đánh dấu lại những khóm lúa trong hàng không đạt tiêu chuẩn, như biểu hiện nhiễm bệnh, hình thái không đồng đều để tránh sai sót trong quá trình thu mâu.
- Bước 4. Thu mâu và ghi nhãn mâu lá vừa thu: Mỗi mâu giống lúa cần thử nghiệm sẽ được thu 100 cá thể, tương ứng với 100 cây lúa. Mỗi cây lúa được thu 2 mâu lá non, không nhiễm bệnh, đảm bảo đủ khối lượng mâu cho phân tích thử nghiệm và lưu mâu. Sau mỗi lần cắt cần phải vệ sinh kéo cắt mâu bằng cồn 70o để tránh nhiễm bẩn, nhiễm chéo mâu trong quá trình lấy mâu, chia mâu. Các mâu lá được lưu giữ trong túi ziplock đã ghi nhãn và bảo quản trong thùng bảo quản lạnh đảm bảo mâu lá tươi cho bước tách chiết ADN.
- Bước 5. Lưu, bảo quản mâu trong vật chứa mâu và vận chuyển mâu về phòng thử nghiệm: Để tránh việc xảy ra mâu thuân về tính đại diện của mâu được thu (Bùi Hữu Điền, 2018). Trong nghiên cứu này, 100 cây của mỗi mâu giống lúa cần thử nghiệm được thu mâu lá, mỗi cây thu hai lá (một lá cho phân tích
thử nghiệm, một lá để lưu mâu). Trong đó, người lấy mâu cần đả m bảo các điều kiện bảo quản ngoài đồng ruộng để tránh các tác nhân như nhiệt độ, ánh sáng, độ ẩm có thể ảnh hưởng đến độ tươi của lá (Semagn, 2014). Cuối cùng, mâu lá được bảo quản trong thùng bảo quản lạnh và được vận chuyển về phòng thử nghiệm để chuẩn bị cho các bước tiếp theo của quy trình thử nghiệm.
Đối với phương pháp thu mâu lá từ ruộng thí nghiệm, thời gian phù hợp nhất để tiến hành là sáng sớm để đảm bảo mâu lá tươi được vận chuyển về phòng thử nghiệm và tách chiết ADN ngay trong ngày. Mâu lưu được bảo quản trong tủ lạnh sâu -20oC trong thời hạn 1 tháng kể từ ngày thu mâu. Như vậy, với năm bước chính và các điểm cần chú ý nêu trên, phương pháp thu mâu lá lúa trên đồng ruộng đã đảm bảo những tiêu chuẩn cần thiết cho Phòng Sinh học phân tử giám định gen thực vật thiết lập Quy trình Lấy mâu và Quản lý mâu thử phù hợp với tiêu chuẩn ISO/IEC 17025: 2017.
3.2. Xây dựng phương pháp thu nhận, bảo quản và chuẩn bị mẫu từ lô hạt giống
Quy trình nhận lô mâu hạt giống và xử lý mâu do khách hàng cung cấp phục vụ cho Phòng Sinh học phân tử giám định gen thực vật được triển khai tùy thuộc vào tình trạng lô mâu giống. Cán bộ tiếp nhận sẽ kiểm tra sơ bộ tính đồng nhất của lô mâu đối chiếu với giấy tờ kèm theo, mô tả và xác định hình dạng, sự nguyên vẹn, độ sạch và mức độ nhiễm tạp của bao gói. Các lô mâu giống không đạt yêu cầu về sự đồng nhất, bị lân hạt mốc, mối mọt sẽ không được tiếp nhận. Lô mâu hạt giống đạt yêu cầu sẽ
Hình 1. Các bước tiến hành khi thu mâu lúa trên đồng ruộng phục vụ cho phòng thí nghiệm giám định gen thực vật chuẩn ISO/IEC 17025

49
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
được cán bộ của phòng thử nghiệm tiếp nhận và hai bên cùng ký nhận vào phiếu giao nhận mâu giống. Phiếu giao nhận sẽ được photo gửi khách hàng 1 bản, bản gốc được lưu trong hồ sơ của phòng thử nghiệm. Mâu hạt giống đạt yêu cầu sẽ được lấy mâu và chuẩn bị mâu thử nghiệm như sau: Đối với mỗi lô mâu cần thử nghiệm, 20 hạt được lấy ngâu nhiên tại mỗi 10 vị trí khác nhau trong lô. Tổng số 200 hạt lúa sẽ được ngâm ủ và cho nảy mầm.
Hạt mâu được xử lý ngâm ủ theo hướng dân trong TCVN 1700:1976 (Bộ Khoa học và Công nghệ, 1986). Trong đó, hạt được nảy mầm trên đĩa petri có giấy ẩm, đặt ở điều kiện nhiệt độ ấm 37 ÷ 40oC trong 2 ÷ 3 ngày. Cần chú ý phải luôn duy trì độ ẩm cho đĩa petri bằng cách bổ sung nước vào giấy. Mâu lá mầm của từng hạt giống được cắt ở giai đoạn 7 ÷ 10 ngày sau khi ngâm ủ, chiều dài mâu cắt 2,0 ÷ 2,5 cm, chuyển vào các ống eppendorf 2 ml đã khử trùng và
giữ lạnh để tiến hành bước tách chiết ADN tiếp theo. Như vậy, với các bước và các điểm cần chú ý nêu trên, phương pháp thu nhận, bảo quản và chuẩn bị mâu thử nghiệm từ lô mâu hạt giống nhận tại phòng thử nghiệm đã đảm bảo những tiêu chuẩn cần thiết cho Phòng Sinh học phân tử giám định gen thực vật thiết lập Quy trình Lấy mâu và Quản lý mâu thử phù hợp với tiêu chuẩn ISO/IEC 17025: 2017.
3.3. Hoàn thiện quy trình quản lý lô hàng và bảo quản mẫu
Việc kiểm soát quá trình bảo quản sau lấy mâu và thông tin về mâu giống thử nghiệm rất quan trọng, mâu sau khi lấy phải bao gói bằng các bao bì thích hợp và phải được ghi nhãn với đầy đủ các thông tin để truy xuất nguồn gốc nếu cần. Để thống nhất theo quy chuẩn chung, nhãn mác thông tin về mâu thử nghiệm đã được thiết kế với mục đích cung cấp đầy đủ thông tin về lô mâu giống (Hình 2).
Hình 2. Mâu nhãn mác lưu trữ và bảo quản mâu thử nghiệm của Phòng Sinh học phân tử giám định gen thực vật theo tiêu chuẩn ISO/IEC 17025: 2017
Cụ thể, nhãn mác lưu trữ và bảo quản mâu thử nghiệm của Phòng Sinh học phân tử giám định gen thực vật có đầy đủ các thông tin về tên phòng thử nghiệm, số thứ tự đợt lấy mâu trong năm, đối tượng thu mâu (lúa hoặc loại cây trồng khác), mã số mâu thử nghiệm. Nhãn mác cũng cung cấp thông tin về người có trách nhiệm lấy mâu, thời gian và địa điểm lấy mâu và loại mâu (lá, hạt...). Các thông tin trên nhãn mác cần được thống nhất với biên bản tiếp nhận lô mâu giống và khách hàng để tránh sai sót cũng như xác thực tính đại diện của phép thử nghiệm.
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luậnĐã thiết lập được phương pháp lấy mâu thử
nghiệm phục vụ xác định locus gen mục tiêu. Phương pháp lấy mâu thử nghiệm được chia thành
phương pháp áp dụng cho việc thu, bảo quản mâu lá lúa trên đồng ruộng và phương pháp áp dụng cho việc thu nhận, bảo quản, chuẩn bị mâu thử nghiệm từ lô mâu hạt giống nhận tại phòng thử nghiệm.
Việc xây dựng phương pháp tiêu chuẩn cho việc lấy mâu thử nghiệm được xem là những bước đi đầu tiên cho việc ban hành Quy trình lấy mâu và quản lý mâu thử theo tiêu chuẩn ISO/IEC 17025: 2017, làm cơ sở cho việc xây dựng và vận hành Phòng Sinh học phân tử giám định gen thực vật đạt chuẩn ISO/IEC 17025:2017.
4.2. Đề nghịNghiên cứu sẽ được tiếp tục nhằm thiết lập quy
trình đánh giá 10 gen chỉ tiêu ở lúa, từ đó đưa vào áp dụng cho Phòng Sinh học phân tử giám định gen thực vật đạt chuẩn ISO/IEC 17025:2017.

50
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
LỜI CẢM ƠNCông trình nghiên cứu nằm trong hoạt động xây
dựng Phòng Sinh học phân tử giám định gen thực vật đạt chuẩn ISO/IEC 17025:2017, thuộc Tiểu dự án FIRST-AGI “Nâng cao năng lực nghiên cứu, làm chủ công nghệ genom học (Genomics-assisted breeding - GAB) và công nghệ chọn giống ứng dụng chỉ thị phân tử (Marker-assisted backcrossing - MABC) để chọn tạo các giống lúa kháng đa yếu tố ứng phó với biến đổi khí hậu”.
TÀI LIỆU THAM KHẢOBộ Khoa hoc và Công nghệ, 1986. Tiêu chuẩn Việt
Nam về Hạt giống lúa nước - Phương pháp thử: TCVN 1700:1976.
Bùi Hữu Điền, 2018. Lấy mâu cho phân tích thử nghiệm. Tạp chí Thử nghiệm Ngày nay, 12: 52-54.
Helliwell, E. E., Yang, Y., 2013. Molecular strategies to improve rice disease resistance. Methods Mol Biol, 956: 285-309.
Honsa, J. D., McIntyre, D. A., 2003. ISO 17025: Practical benefits of implementing a quality system. J AOAC Int, 86(5): 1038-1044.
Semagn, K., 2014. Leaf tissue sampling and DNA extraction protocols. In: Besse P (ed) Molecular plant taxonomy: Methods and Protocols. Humana Press, Totowa, NJ: 53-67.
Yamamoto, S., Asakura, H., Machii, K., Igimi, S., 2010. Approval of ISO/IEC 17025 and quality control of laboratory testing. Kokuritsu Iyakuhin Shokuhin Eisei Kenkyusho Hokoku, 128: 78-80.
THIẾT KẾ VÀ XÂY DỰNG BỘ CHỈ THỊ PHÂN TỬ PHỤC VỤ KIỂM ĐỊNH VÀ NGHIÊN CỨU DI TRUYÊN SÂM NGỌC LINH (Panax vietnamensis)
Khuất Thị Mai Lương1, Nguyễn Thị Minh Nguyệt1, Chu Đức Hà1, Trần Thị Hoa Mỹ1, Đinh Văn Phê2, Lê Hùng Lĩnh1
TÓM TẮTSử dụng chỉ thị phân tử phục vụ kiểm định và phân tích di truyền các loài sâm, đặc biệt là sâm Ngọc Linh
(Panax vietnamensis) là một trong những vấn đề được quan tâm hiện nay. Trong nghiên cứu này, các chỉ thị phân tử trình tự lặp đơn giản (SSR) phục vụ kiểm định và phân tích di truyền kiểu gen sâm Ngọc Linh đã được thiết kế dựa trên cơ sở dữ liệu tham chiếu hệ gen nhân và lục lạp của sâm Hàn Quốc (Panax ginseng). Kết quả đã thiết kế được 600 chỉ thị phân tử, 200 chỉ thị thiết kế từ trình tự SSR có độ lặp cao được chọn lọc để chạy khảo sát. Trong đó, ba motif lặp, bao gồm -AT-, -AA- và -TT- được đánh giá là các trình tự lặp phổ biến nhất được thiết kế cho các chỉ thị SSR. Kết quả điện di cho thấy, 134 chỉ thị SSR có khả năng khuếch đại đoạn trình tự mục tiêu của P. ginseng và P. vietnamensis với kích thước băng từ 100 ÷ 300 bp như thiết kế. Kết quả của nghiên cứu này sẽ cung cấp những thông tin cơ bản về các chỉ thị phân tử phục vụ công tác kiểm định và phân tích di truyền các loài sâm Việt trong đó có sâm Ngọc Linh.
Từ khóa: Chỉ thị phân tử, PCR, sâm Ngọc Linh, thiết kế
Establishment of rice sampling protocols with the ISO/IEC 17025: 2017 standard for the identification of locus for target genes
Chu Duc Ha, Nguyen Thi Minh Nguyet, Nguyen Thi Nhai, Nguyen Ba Ngoc, Khuat Thi Mai Luong, Pham Thi Ly Thu, Le Hung Linh
AbstractISO/IEC 17025 Standard (International Organization for Standardization/ International Electrotechnical Commission) is considered as one of the most critical criteria for the international laboratory certification. In this study, the rice sampling protocols have been successfully established for the testing of the identification of target genes. Particularly, two methods in the field and laboratory have been classified. Among them, the steps of sample collecting and storage in the field were highly recommended as much care as the process of sample receiving, storage and preparation from the seed package in the laboratory. Our results could provide the rice sampling protocols according to the ISO/IEC 17025: 2017 standard, therefore, contribute to the establishment and operation of ISO/IEC 17025: 2017 certified laboratory of the molecular biology for the plant gene(s) detection. Keywords: ISO, rice, test, protocol, sampling
Ngày nhận bài: 24/10/2087Ngày phản biện: 29/10/2018
Người phản biện: TS. Huỳnh Văn NghiệpNgày duyệt đăng: 15/11/2018
1 Viện Di truyền Nông nghiệp, VAAS; 2 Viện Khoa học kỹ thuật Nông lâm nghiệp Tây Nguyên (WASI)

51
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
I. ĐẶT VẤN ĐỀỞ Việt Nam, chi Panax được ghi nhận gồm
4 loài chính là sâm Vũ Diệp (P. bipinnatifidus), tam thất hoang (P. stipuleanatus), sâm Lai Châu (P. vietnamensis var. fuscidiscus) và sâm Ngọc Linh (P. vietnamensis) (Nguyễn Tập, 2005). Trong đó, sâm Ngọc Linh được xác định là cây thuốc quý về giá trị sử dụng cũng như giá trị nguồn gen. Phần thân rễ của cây sâm Ngọc Linh có chứa ít nhất 50 saponin có tác dụng dược lý quan trọng (Duc et al., 1994). Đặc biệt, saponin tách chiết từ sâm Ngọc Linh đã được chứng minh có tác dụng tăng cường khả năng miễn dịch và ngăn ngừa ung thư (Chu Đức Hà và ctv., 2018). Với tác dụng dược lý và giá trị kinh tế kinh tế cao như vậy, sâm Ngọc Linh hiện nay đã và đang bị làm giả mạo. Vì vậy, phân loại, kiểm định và nghiên cứu di truyền chi Panax nói chung và sâm Ngọc Linh nói riêng được xem là bài toán cấp thiết hiện nay.
Bên cạnh phương pháp xác định và đánh giá các loài thực vật bằng nhận dạng hình thái và phân tích hóa sinh, kỹ thuật chẩn đoán bằng chỉ thị phân tử được xem là một trong những cách tiếp cận hiện đại, nhanh chóng và chính xác nhất (Jo et al., 2017). Trong đó, dựa trên sự phát triển của công nghệ giải trình tự thế hệ mới, thông tin di truyền (hệ gen nhân và hệ gen tế bào chất) của rất nhiều loài trong chi Panax đã được xác định một cách cụ thể. Đây được xem là một hệ tham chiếu rất quan trọng để thiết kế và xác định các chỉ thị phân tử đặc trưng nhằm phân loại và phân tích chính xác các loài sâm (Jo et al., 2017).
Trong nghiên cứu này, các chỉ thị phân tử trình tự lặp đơn giản (Simple sequence repeat, SSR) sử dụng trong kiểm định sâm Ngọc Linh đã được thiết kế dựa trên hệ gen nhân và lục lạp tham chiếu của P. ginseng (Jayakodi et al., 2018; Kim et al., 2012; Um et al., 2016). Sau đó, khả năng bắt cặp để khuếch đại sản phẩm của các chỉ thị được kiểm tra bằng kỹ thuật PCR với mâu P. ginseng và P. vietnamensis. Kết quả của nghiên cứu này là bước khởi đầu quan trọng trong việc thiết lập bộ chỉ thị nhằm kiểm định và phân tích di truyền của sâm Ngọc Linh.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưuMâu lá của cây P. vietnamensis được thu thập tại
vườn ươm của Trung tâm nghiên cứu và phát triển sâm Ngọc Linh thuộc Trung tâm Ươm tạo và Hỗ trợ doanh nghiệp KHCN, tỉnh Kon Tum và mâu củ P. ginseng được thu thập tại cơ quan hợp tác về công nghệ nhân sâm Chungnam, Hàn Quốc.
2.2. Phương pháp nghiên cưu
2.2.1. Phương pháp tách chiết ADN Tách chiết ADN tổng số được thực hiện theo
phương pháp CTAB mô tả trong nghiên cứu gần đây (Li et al., 2013). Nồng độ và độ tinh sạch của mâu ADN được kiểm tra bằng máy NanoDrop2000 (ThermoFisher, Hoa Kỳ).
2.2.2. Phương pháp thiết kế chỉ thị phân tửThông tin về hệ gen nhân của P. ginseng (Jayakodi
et al., 2018) được khai thác trên các cơ sở dữ liệu chứa trình tự đơn lặp lại (microsatellite), trình tự Genome survey sequence (GSS), Expressed sequence tag (EST) và trình tự lục lạp trên NCBI (Wheeler et al., 2008) được sử dụng làm hệ tham chiếu cho sâm Ngọc Linh. Các chỉ thị phân tử sau đó được thiết kế bằng chương trình Primer3 với các thông số mặc định (Rozen and Skaletsky, 2000). Trình tự mồi xuôi và môi ngược được gửi sang công ty IDT (Intergrated DNA Technologies, Hoa Kỳ) tổng hợp.
2.2.3. Ky thuật PCR và kiểm tra sản phâm PCR bằng gel agarose
Phản ứng PCR được tiến hành trên máy chu kỳ nhiệt Mastercycler Pro (Eppendorf, Đức). Tổng dung dịch phản ứng là 15 µl, bao gồm 50 ng ADN tổng số, 0,4 µM mồi, 0,2 mM dNTP, 1X dịch đệm PCR chứa 10 mM Tris-HCl, 50 mM KCl và 1,5 mM MgCl2, và 1,0 đơn vị Taq TaKaRa. Điều kiện phản ứng PCR được minh họa tại bảng 1. Sản phẩm PCR được kiểm tra trên gel agarose 2,0% chứa ethidium bromide với thang ADN chuẩn 1 kb Plus (Thermo Scientific, Anh) ở hiệu điện thế 150V trong 90 phút trong dung dịch đệm Tris-boric acid-EDTA 0,5X (Morgante et al., 1998).
Bảng 1. Điều kiện phản ứng PCR
2.3. Thời gian và địa điểm nghiên cưu Nghiên cứu được thực hiện tại Bộ môn Sinh
học phân tử - Viện Di truyền Nông nghiệp từ tháng 5/2017 đến tháng 6/2018.
STT Giai đoạn phản ưng Điều kiện phản ưng
1 Khởi đầu 94°C - 5 phút2 Biến tính 94oC - 30 giây
35 ch
u kỳ3 Gắn mồi (52 ÷ 60oC) - 45 giây
4 Kéo dài 72oC - 2 phút5 Kết thúc kéo dài 72oC - 7 phút6 Bảo quản 4oC - ∞

52
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Kết quả thiết kế chỉ thị phân tử cho phân tích P. vietnamensis dựa vào hệ gen tham chiếu của P. ginseng
Để thiết kế chỉ thị phân tử SSR cho kiểm định và phân tích di truyền sâm Ngọc Linh, dữ liệu từ P. ginseng, bao gồm ngân hàng microsatellite, GSS và EST (Wheeler et al., 2008) và trình tự hệ gen lục
lạp được sử dụng làm hệ tham chiếu. Kết quả đã tìm kiếm được 935 đoạn lặp trên các trình tự gen của P. ginseng, từ đó thiết kế ra tổng số 600 chỉ thị phân tử SSR. Trong đó, 200 cặp mồi được sử dụng chạy khảo sát bao gồm 43 chỉ thị dựa trên EST (đặt tên là PVE) (Bảng 2), 70 chỉ thị dựa trên GSS (đặt tên là PVG) (Hình 1A) và 40 chỉ thị dựa trên cơ sở dữ liệu microsatellite (đặt tên là PVM) (Hình 1B) và 47 chỉ thị dựa trên trình tự lục lạp.
Kết quả thu được cho thấy, 60,5% PVE được thiết kế dựa trên kiểu lặp 2 nucleotide (19 chỉ thị) và 3 nucleotide (7 chỉ thị) với độ lặp cao (Bảng 2). Tương tự, 57,1% PVG, tương ứng 40 chỉ thị, được thiết lập từ kiểu lặp 2 nucleotide, 11,3% PVG (8 chỉ thị) được xác định từ kiểu lặp 3 nucleotide và 31,6% PVG (22
chỉ thị) được thu thập từ kiểu lặp 4 nucleotide trở lên (Hình 1A). Trong khi đó, 40 chỉ thị PVM được thiết kế dựa trên kiểu lặp 3 nucleotide (16 chỉ thị, chiếm 40%), kiểu lặp đa nucleotide (14 chỉ thị, chiếm 35%) và motif lặp 2 nucleotide (10 chỉ thị, chiếm 25%) (Hình 1B).
Bảng 2. Danh sách chỉ thị phân tử cho phân tích P. vietnamensis được thiết kế tư cơ sở dữ liệu EST của P. ginseng
Ghi chú: [PCR]E: kích thước dự kiến.
Hình 1. Thiết kế chỉ thị phân tử SSR từ cơ sở dữ liệu (A) GSS và (B) microsatellite
STT Tên chỉ thị Kiểu lặp [PCR]E STT Tên
chỉ thị Kiểu lặp [PCR]E STT Tên chỉ thị Kiểu lặp [PCR]E
1 PVE01 (TTA)7 288 16 PVE17 (TA)31 182 31 PVE32 (CACTG)5 1992 PVE02 (CCT)9 169 17 PVE18 (AAACAG)4 202 32 PVE33 (GCCCCT)3 1733 PVE03 (TTTGT)3 240 18 PVE19 (AT)32 191 33 PVE34 (TCAGCA)3 1824 PVE04 (AATTTA)3 240 19 PVE20 (AT)21 212 34 PVE35 (GAG)7 1605 PVE05 (GGGGA)3 250 20 PVE21 (TCTCAAC)4 171 35 PVE36 (TC)20 2466 PVE07 (TTA)9 245 21 PVE22 (AT)22 218 36 PVE37 (GGTGAG)5 1997 PVE08 (TCTAAA)5 154 22 PVE23 (AT)20 298 37 PVE38 (TC)10 1898 PVE09 (AG)14 151 23 PVE24 (AT)21 209 38 PVE39 (CCATCT)5 1509 PVE10 (TATG)6 231 24 PVE25 (AT)28 196 39 PVE40 (AT)12 200
10 PVE11 (AT)12 200 25 PVE26 (AAG)7 235 40 PVE41 (TA)14 21711 PVE12 (TA)29 159 26 PVE27 (TAT)6 186 41 PVE42 (TGATGC)3 24912 PVE13 (CAAAAT)5 179 27 PVE28 (AAAAAT)4 207 42 PVE43 (AT)15 16613 PVE14 (AT)30 224 28 PVE29 (TA)11 235 43 PVE44 (GAG)8 15914 PVE15 (AT)15 238 29 PVE30 (TGTTCA)3 21415 PVE16 (AT)22 237 30 PVE31 (GAAAGA)4 246

53
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Kiểu lặp được xác định nhiều nhất là 2 nucleotide -AT- với số lần lặp lớn hơn 10 (Bảng 2, Hình 1) và dạng -AA- hoặc -TT- tồn tại trong kiểu lặp lớn hơn 4 nucleotide (Bảng 2, Hình 1). Các kiểu lặp này thường được lặp với tần suất rất cao, ví dụ như (AT)n với n ≥ 10 ở các mồi PVE (Bảng 2). Đây là các nucleotide lặp xuất hiện phổ biến nhất được ghi nhận ở hệ gen thực vật (Lagercrantz et al., 1993). Nghiên cứu trước đây cũng cho thấy chỉ thị phân tử được thiết kế dựa trên những motif -AT-, -AA- hoặc -TT- thường được sử dụng để phân biệt quan hệ giữa các loài (Lagercrantz et al., 1993).
3.2. Kết quả khảo sát khả năng phản ưng của chỉ thị phân tử thiết kế cho P. vietnamensis
Trong nghiên cứu này, bộ chỉ thị gồm 200 cặp mồi SSR đã được thiết kế và tổng hợp. Nhiệt độ gắn mồi của các chỉ thị dao động từ 52 ÷ 60oC (Bảng 1).Kích thước dự kiến đoạn khuếch đại từ các mồi PVE từ 150 ÷ 298 bp (Bảng 2), trong khi mồi PVG và PVM có thể nhân được các đoạn có kích thước
lần lượt từ 150 ÷ 319 bp (Hình 1A) và 100 ÷ 250 bp (Hình 1B). Tiếp theo, khả năng khuếch đại của chỉ thị SSR thiết kế dựa trên hệ gen P. ginseng phục vụ cho kiểm định và phân tích di truyền P. vietnamensis được kiểm tra thông qua phản ứng PCR và điện di trên gel agarose 2%.
Kết quả khảo sát cho thấy 145 trên tổng số 200 chỉ thị SSR, chiếm tỉ lệ 72,5%, có khả năng khuếch đại đoạn nhân từ cả 2 mâu ADN P. ginseng và P. vietnamensis. Cụ thể, 30 chỉ thị PVM (trên tổng số 40), 51 chỉ thị PVG (trên tổng số 70) và 34 chỉ thị PVE (trên tổng số 43) và 30 chỉ thị lục lạp (trên tổng số 47) (Hình 2). Có thể nhận thấy rằng, hầu hết các chỉ thị SSR được thiết kế dựa trên cơ sở dữ liệu microsatellite và EST đều có khả năng khuếch đại đoạn trình tự của P. vietnamensis rất cao, so với chỉ thị SSR được xây dựng từ GSS và hệ gen lục lạp. Bên cạnh đó, sản phẩm được nhân lên từ 134 trên tổng số 145 (92,4%) chỉ thị SSR có kích thước như dự kiến. Phần lớn sản phẩm PCR có kích thước từ 100 ÷ 300 bp (Hình 2).
Những kết quả này cho thấy hệ gen nhân của P. vietnamensis tương đối tương đồng so với loài tham chiếu P. ginseng. Nghiên cứu ứng dụng trình tự microsatellite và EST trên hệ gen nhân của P. ginseng để thiết kế chỉ thị phân tử phục vụ kiểm định và phân tích di truyền P. vietnamensis là hướng đi mới và có khả thi. Nghiên cứu này sẽ được tiếp tục nhằm sàng lọc những chỉ thị cho kết quả đa hình giữa P. ginseng và P. vietnamensis nhằm đề xuất bộ chỉ thị cho kiểm định và phân tích di truyền các loài sâm Việt.
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luậnĐã thiết kế và chạy khảo sát được 200 chỉ thị
phân tử SSR có độ lặp cao cho P. vietnamensis. Trong đó, 43 chỉ thị PVE được thiết kế từ cơ sở dữ liệu EST, 70 chỉ thị PVG và 40 chỉ thị PVM lần lượt được xây dựng từ dữ liệu GSS và microsatellite từ hệ gen nhân tham chiếu của P. ginseng và 47 chỉ thị dựa trên hệ gen lục lạp. Dạng -AT-, -AA- và -TT- được xác định là các kiểu lặp phổ biến nhất được thiết kế cho các chỉ thị SSR.
Hình 2. Kết quả điện di 48 chỉ thị phân tử trên gel agarose 2%với thang ADN chuẩn. Pg: ADN tách chiết từ củ P. ginseng; Pv: ADN tách chiết từ mâu lá P. vietnamensis

54
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Kiểm chứng bằng kỹ thuật PCR cho thấy 134 trên tổng số 145 chỉ thị SSR có khả năng khuếch đại đoạn ADN của P. ginseng và P. vietnamensis với kích thước băng từ 100÷300 bp như thiết kế. Hầu hết các chỉ thị kiểm chứng đều là chỉ thị PVM và PVE được xây dựng trên cơ sở dữ liệu microsatellite và EST.
4.2. Đề nghịNghiên cứu này sẽ được tiếp tục nhằm tiến hành
định danh phân tử và bước đầu phân loại sâm Ngọc Linh bằng chỉ thị SSR.
LỜI CẢM ƠNNghiên cứu này được thực hiện với sự tài trợ từ
đề tài “Nghiên cứu xây dựng bộ chỉ thị phân tử phục vụ giám định, khai thác và phát triển sâm Ngọc Linh (Panax vietnamensis)” (mã số: ĐTĐL.CN-29/16) thuộc Bộ Khoa học và Công nghệ.
TÀI LIỆU THAM KHẢOChu Đưc Hà, Lê Hùng Lĩnh, Nguyễn Văn Kết, Lê
Tiến Dũng, Đỗ Mạnh Cường, Hoàng Thanh Tùng, Dương Tấn Nhựt, 2018. Sâm Ngọc Linh: Cây dược liệu quý mang thương hiệu quốc gia. Tạp chí Khoa học & Công nghệ Việt Nam, 1(706): 32-35.
Nguyễn Tập, 2005. Các loài thuộc chi Panax L. ở Việt Nam. Tạp chí Dược liệu, 10(3): 71-76.
Duc, N. M., Kasai, R., Ohtani, K., Aiko, I., Nguyen, T. N., Yamasaki, K. and Tanaka, O., 1994. Saponins from Vietnamese ginseng, Panax vietnamensis Ha et Grushv. collected in Central Vietnam. III. Chem Pharm Bull, 42(3): 634-640.
Jayakodi, M., Choi, B.-S., Lee, S.-C., Kim, N.-H., Park, J. Y., Jang, W., Lakshmanan, M., Mohan, S. V. G., Lee, D.-Y., Yang, T.-J., 2018. Ginseng Genome Database: An open-access platform for genomics of Panax ginseng. BMC Plant Biol, 18(1): 62.
Jo, I. H., Kim, Y. C., Kim, D. H., Kim, K. H., Hyun, T. K., Ryu, H., Bang, K. H., 2017. Applications of molecular markers in the discrimination of Panax species and Korean ginseng cultivars (Panax ginseng). J Ginseng Res, 41(4): 444-449.
Kim, N. H.; Choi, H. I.; Ahn, I. O.; Yang, T. J., 2012. EST-SSR marker sets for practical authentication of all nine registered ginseng cultivars in Korea. J Ginseng Res, 36(3): 298-307.
Lagercrantz, U., Ellegren, H., Andersson, L., 1993. The abundance of various polymorphic microsatellite motifs differs between plants and vertebrates. Nucleic Acid Res, 21(5): 1111-1115.
Li, J., Wang, S., Jing, Y., Wang, L., Zhou, S., 2013. A modified CTAB protocol for plant DNA extraction. Chinese Bull Bot, 48: 1-7.
Morgante, M., Pfeiffer, A., Jurman, I., Paglia, G., Olivieri, A. M., 1998. PCR analysis of SSR polymorphisms in plants using agarose gels. In: Karp A, Isaac PG, Ingram DS (eds) Molecular tools for screening biodiversity: Plants and animals. Springer Netherlands, 206-207.
Rozen, S., Skaletsky, H., 2000. Primer3 on the WWW for general users and for biologist programmers. Methods Mol Biol, 132: 365-386.
Wheeler, D. L., Barrett, T., Benson, D. A., Bryant, S. H., Canese, K., Chetvernin, V., Church, D. M., DiCuccio, M., Edgar, R., Federhen, S., Feolo, M., Geer, L. Y., Helmberg, W., Kapustin, Y., Khovayko, O., Landsman, D., Lipman, D. J., Madden, T. L., Maglott, D. R., Miller, V., Ostell, J., Pruitt, K. D., Schuler, G. D., 2008. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res, 36(Database issue): D13-D21.
Um Yurry, Jin M.L., Kim O.T., Kim Y.C., Kim S.C., Cha S.W., Chung K.W., 2016. Identification of Korean ginseng (Panax ginseng) cultivars using simple sequence repeat markers. Plant Breed Biotech, 4(1): 71-78.
Design and validation of the molecular markers for genetic analysis of Vietnamese ginseng (Panax vietnamensis)
Khuat Thi Mai Luong, Nguyen Thi Minh Nguyet, Chu Duc Ha, Tran Thi Hoa My, Dinh Van Phe, Le Hung Linh
AbstractGenetic analysis of medical plants, especially of Vietnamese ginseng (Panax vietnamensis) is considered to be one of the most fascinating unsolved issues. In this study, a set of the simple sequence repeats (SSRs) molecular markers for P. vietnamensis was established based on the reference nuclear and chloroplast genome of the Korean ginseng (P. ginseng). As a result, a total of 200 SSRs (out of 600 designed markers) was used to amplify DNA of P. vietnamensis and P. ginseng. Among them, three di-nucleotide motifs, -AT-, -AA- and -TT- were identified to be the most common repeats in the design of SSR markers for P. vietnamensis. PCR-validation confirmed that 134 SSR markers were successfully amplified the approximately 100÷300-bp-sequences of both P. ginseng and P. vietnamensis. Our study would provide an initial background for the genetic analysis of the Panax species in Vietnam. Keywords: Molecular marker, PCR, Vietnamese ginseng, design
Ngày nhận bài: 18/9/2018Ngày phản biện: 28/9/2018
Người phản biện: Trần Thanh MếnNgày duyệt đăng: 15/10/2018

55
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
NGHIÊN CỨU TÁC ĐỘNG CỦA NANO BẠC ĐẾN QUÁ TRÌNH PHÁT SINH HÌNH THÁI TRONG NUÔI CẤY IN VITRO HOA ĐỒNG TIÊN
Bùi Thị Thu Hương1, Đồng Huy Giới1, Trần Thị Thu Thủy1, Nguễn Thị Ngọc Quỳnh1
TÓM TẮTNghiên cứu được tiến hành với mục đích đánh giá tác động của nano bạc đến quá trình phát sinh hình thái của
hoa đồng tiền nuôi cấy mô từ nguồn vật liệu là các mâu lá in vitro. Kết quả nghiên cứu đã chỉ ra rằng: (i) Môi trường thích hợp nhất cho việc tạo mô sẹo từ mâu lá in vitro hoa Đồng tiền là môi trường MS có bổ sung 30 g/l saccharose; 6,5 g/l agar; 1,5 mg/l 2,4D; pH 5,7 và 10 ppm nano bạc, với tỉ lệ tạo mô sẹo cao nhất đạt 95,56%, mô sẹo mềm và có màu vàng sáng; (ii) Môi trường MS có bổ sung 0,7 mg/l BA; 0,1 mg/l IAA và 4 ppm nano bạc cho hiệu quả tái sinh chồi từ mô sẹo tốt nhất, tỉ lệ tái sinh chồi đạt 84,45%, số chồi trung bình đạt 3,29 chồi/mô sẹo sau 6 tuần nuôi cấy; (iii) Môi trường MS có bổ sung 3 mg/l BAP; 0,1 mg/l α- NAA và 2 ppm nano bạc cho hiệu quả nhân nhanh chồi hoa Đồng tiền tốt nhất, hệ số nhân chồi đạt 8,22 lần, chiều cao chồi trung bình đạt 5,75 cm sau 6 tuần nuôi cấy; (iv) Chồi in vitro hoa Đồng tiền ra rễ hiệu quả nhất trong môi trường MS có bổ sung 50 g/l saccharose; 6,5 g/l agar; 1,0 mg/l α-NAA và 4 ppm nano bạc, tỉ lệ chồi ra rễ đạt 100%, số rễ trung bình là 5,73 rễ/chồi và chiều dài rễ trung bình đạt 5,93 cm sau 4 tuần nuôi cấy.
Từ khóa: Hoa đồng tiền, lá in vitro, môi trường MS, nano bạc
1 Khoa Công nghệ sinh học, Học viện Nông nghiệp Việt Nam
I. ĐẶT VẤN ĐỀ Hoa Đồng tiền (Gerbera jamesonii Bolus ex Hook. F.)
là một trong những loài hoa đẹp, có giá trị kinh tế cao, được trồng phổ biến ở Việt Nam và nhiều quốc gia trên thế giới, tuy nhiên hiện nay nguồn giống tốt cung cấp cho sản xuất còn thiếu, người trồng hoa thường phải mua giống không rõ nguồn gốc, có khi mua phải giống bị thoái hóa, giống bị nhiễm bệnh (Đỗ Năng Vịnh, 2003).
Phương pháp nhân giống bằng nuôi cấy mô tế bào thực vật đã và đang được áp dụng khá phổ biến trên nhiều đối tượng cây trồng khác nhau. Một trong những ưu điểm nổi bật của nhân giống bằng phương pháp nuôi cấy mô là hệ số nhân cao, trong một thời gian ngắn có thể tạo ra một số lượng lớn cây giống tương đối đồng nhất, cây giống sạch bệnh, giá thành thấp. Hiện nay đã có một số nghiên cứu thành công trong việc nhân giống hoa đồng tiền bằng nuôi cấy mô từ các nguồn vật liệu khác nhau như nụ hoa non, cuống lá, phiến lá (Bhavya Bhargava et al., 2013; Shylaja et al., 2014; Nguyễn Thị Mỹ Duyên và Trương Thị Hằng, 2014). Tuy nhiên một số nghiên cứu trên cho kết quả chưa thực sự tốt, bên cạnh đó việc sử dụng đế hoa non làm nguyên liệu cho nuôi cấy mô sẽ làm thất thoát đi một số lượng lớn các bông hoa đẹp, vì vậy rất cần thiết tìm kiếm một loại vật liệu khác phù hợp hơn để thay thế.
Vật liệu nano nói chung và nano bạc nói riêng đã và đang được ứng dụng rộng rãi trong đời sống những năm gần đây, tạo nên những bước đột phá trong nhiều lĩnh vực. Nano bạc đang được chú ý sử
dụng trong nuôi cấy mô tế bào thực vật do đặc điểm kháng khuẩn tuyệt vời của nó, bên cạnh đó nano bạc còn có vai trò tích cực tới sự phát sinh hình thái của cây in vitro (Rostami and Shahsavar , 2009; Shokri et al., 2015).
Xuất phát từ những lí do nêu trên, nghiên cứu này được thực hiện nhằm nhân giống hoa Đồng tiền bằng nuôi cấy mô từ nguồn vật liệu là lá in vitro, đồng thời bước đầu sử dụng nano bạc nhằm nâng cao hiệu quả của quá trình nuôi cấy.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưuChồi in vitro giống hoa Đồng tiền ĐTH125 có
nguồn gốc từ Hà Lan. Dung dịch nano bạc với kích thước hạt dao động 15 - 20 nm được điều chế tại Bộ môn Sinh học, Khoa Công nghệ sinh học, Học viện Nông nghiệp Việt Nam; môi trường MS (T. Murashige& F. Skoog, 1962) và một số loại hóa chất cần thiết khác sử dụng trong nuôi cấy mô tế bào thực vật.
2.2. Phương pháp nghiên cưu
2.2.1. Điều kiện thí nghiệmMôi trường nuôi cấy được điều chỉnh pH là 5,7 và
được hấp khử trùng ở 1210C, áp suất 1,1 atm trong 15 phút. Các thí nghiệm được bố trí hoàn toàn ngâu nhiên (RCB), mỗi công thức 3 lần nhắc lại, mỗi lần nhắc lại 15 mâu.
Điều kiện phòng nuôi: Nhiệt độ 250C - 270C; Cường độ ánh sáng: 2400 lux - 2600 lux; Độ ẩm: 70%; Thời gian chiếu sáng: 16 h chiếu sáng/ngày.

56
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
2.2.2. Nghiên cứu ảnh hưởng của nano bạc đến khả năng tạo mô sẹo từ mâu lá in vitro
Các mâu lá in vitro sạch bệnh được cắt thành các mảnh có kích thước 1 cm ˟ 1 cm, sau đó cấy vào môi trường tạo mô sẹo: Môi trường MS có bổ sung 30 g/l saccharose; 6,5 g/l agar; 1,5 mg/l 2,4D (Nguyễn Thị Mỹ Duyên và ctv., 2014) và bổ sung thêm nano bạc với các nồng độ khác nhau (0, 6, 8, 10 hoặc 12 ppm). Tiến hành theo dõi tỉ lệ tạo mô sẹo và đặc điểm mô sẹo sau 4 tuần nuôi cấy.
2.2.3. Nghiên cứu ảnh hưởng của nano bạc đến khả năng tái sinh chồi từ mô sẹo
Các mô sẹo có màu vàng sáng, mềm được chuyển sang môi trường tái sinh chồi: Môi trường MS có bổ sung 0,7 mg/l BA; 0,1 mg/l IAA (Phạm Thị Bích Ngọc và ctv., 2009) và bổ sung thêm nano bạc với các nồng độ là 0, 2, 4, hoặc 6 ppm. Tiến hành theo dõi các chỉ tiêu tỉ lệ mô sẹo tái sinh chồi, số chồi/mô sẹo sau 6 tuần nuôi cấy.
2.2.4. Nghiên cứu ảnh hưởng của nano bạc đến khả năng nhân nhanh của chồi hoa Đồng tiền từ chồi in vitro
Các chồi sinh trưởng bình thường có đầy đủ thân và lá (không bị dị dạng), cao từ 5 - 6 cm được sử dụng làm vật liệu cấy chuyển sang môi trường nhân nhanh: Môi trường MS có bổ sung 30 g/l saccharose; 3 mg/l BAP; 0,1 mg/l α-NAA; 6,5 g/l agar (Son et al. 2011; Shylaja et al., 2014) và bổ sung thêm nano bạc với các nồng độ là 0, 2, 4, hoặc 6 ppm. Tiến hành theo dõi các chỉ tiêu hệ số nhân chồi, chiều cao chồi, đặc điểm chồi sau 2 tuần, 4 tuần và 6 tuần nuôi cấy.
2.2.5. Nghiên cứu ảnh hưởng của nano bạc đến khả năng ra rễ của chồi hoa Đồng tiền
Chồi hoa Đồng tiền khỏe mạnh có từ 3 - 5 lá được nuôi cấy trong môi trường ra rễ: Môi trường MS có bổ sung 30 g/l saccharose, 1,0 mg/l α-NAA, 6,5 g/l agar (Nguyễn Văn Hồng, 2009; Paduchuri et al., 2010) và bổ sung thêm nano bạc với các nồng độ là 0, 2, 4, hoặc 6 ppm. Tiến hành theo dõi các chỉ tiêu tỉ lệ ra rễ, số rễ/chồi, chiều dài rễ, màu sắc rễ sau 4 tuần nuôi cấy.
2.2.6. Phương pháp xử lý số liệuSố liệu được xử lý theo chương trình Microsoft
Excel và IRRISTART 5.0 .Các công thức so sánh được tiến hành theo phương pháp kiểm tra sự sai khác giữa các giá trị trung bình bằng phép ước lượng và sử dụng tiêu chuẩn LSD (Least Significant Different) ở độ tin cậy 95%.
2.3. Thời gian và địa điểm nghiên cưuNghiên cứu được thực hiện tại phòng thí nghiệm
Khoa Công nghệ sinh học, Học viện Nông nghiệp Việt Nam từ tháng 8/2017 đến tháng 8/2018.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Ảnh hưởng của nồng độ nano bạc đến khả năng tạo mô sẹo từ mẫu lá in vitro
Nhằm đánh giá tác động của các nồng độ nano bạc khác nhau đến quá trình hình thành mô sẹo, trong thí nghiệm này các mâu lá in vitro hoa Đồng tiền được nuôi cấy trong môi trường có bổ sung nano bạc với 4 nồng độ khác nhau là 6 ppm, 8 ppm, 10 ppm và 12 ppm (Emad et al., 2015). Kết quả thu được sau 4 tuần nuôi cấy được thể hiện ở bảng 1.
Bảng 1. Kết quả tạo mô sẹo từ mâu lá in vitro hoa Đồng tiền sau 4 tuần nuôi cấy
Ghi chú: Bảng 1, 2, 3, 4: Trong cùng một cột, các giá trị mang các chữ cái khác nhau thì khác nhau có ý nghĩa ở mức α = 0,05.
Kết quả nghiên cứu thể hiện ở bảng 1 cho thấy, tỷ lệ tạo thành mô sẹo của các công thức thí nghiệm là khá cao, dao động từ 66,67% đến 95,56%. Ở hầu hết các công thức có bổ sung nano bạc đều cho tỉ lệ tạo mô sẹo cao hơn so với công thức đối chứng (không bổ sung nano bạc). Khi tăng nồng độ nano bạc từ 6 ppm - 10 ppm thì tỉ lệ tạo mô sẹo cũng tăng lên theo, trong đó công thức bổ sung 10 ppm nano bạc cho tỉ lệ tạo mô sẹo cao nhất đạt 95,56%, cao hơn có ý nghĩa thống kê so với tất cả các công thức còn lại, mô sẹo mềm và có màu vàng sáng. Tuy nhiên, khi tăng nồng độ nano bạc lên 12 ppm thì tỉ lệ tạo mô sẹo lại giảm xuống đáng kể, chỉ còn 66,67%, mô sẹo rắn và có màu vàng sậm. Theo kết quả nghiên cứu của
Nồng độ nano bạc
(ppm)
Tổng số
mẫu
Số mẫu tạo mô
sẹo
Tỷ lệ tạo mô sẹo
(%)
Đặc điểmMô sẹo
0 45 30 66,67d Mềm, màu vàng sáng
6 45 34 75,56c Mềm, màu vàng sáng
8 45 38 84,45b Mềm, màu vàng sáng
10 45 43 95,56a Mềm, màu vàng sáng
12 45 30 66,67d Rắn, màu hơi sậm
CV (%) 4,6LSD0,05 0,77

57
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Zainab M. Almutairi (2016), hạt cây cà chua xử lý bằng nano bạc với nồng độ lớn hơn 2 mg/l gây tăng chỉ số phân bào, và có sự xuất hiện hình dạng sai của nhiễm sắc thể và làm giảm tỷ lệ nảy mầm. Điều đó
lý giải vì sao khi ở nồng độ thích hợp thì nano bạc kích thích tạo mô sẹo mạnh, nhưng khi quá ngưỡng thì lại gây ức chế sự phân chia của tế bào và gây ra những biến đổi về màu sắc của mô sẹo.
3.2. Ảnh hưởng của nồng độ nano bạc đến khả năng tái sinh chồi từ mô sẹo
Kết quả đánh giá ảnh hưởng của nồng độ nano bạc đến khả năng tái sinh chồi từ mô sẹo của hoa Đồng tiền sau 6 tuần nuôi cấy được trình bày ở Bảng 2.
Bảng 2. Kết quả tái sinh chồi từ mô sẹo sau 6 tuần nuôi cấy
Kết quả thu được ở bảng 2 cho thấy, tất cả các công thức bổ sung nano bạc đều cho tỉ lệ mô sẹo tái sinh chồi và số chồi trung bình cao hơn so với công thức không bổ sung nano bạc ở mức có ý nghĩa, điều đó chứng tỏ nano bạc có tác dụng thúc đẩy sự tái sinh chồi từ mô sẹo của hoa Đồng tiền. Cụ thể, ở chỉ tiêu tỉ lệ tái sinh chồi, công thức không bổ sung nano bạc cho tỉ lệ tái sinh chồi là 42,22%, trong khi đó các công thức bổ sung nano bạc tỉ lệ tái sinh chồi dao động từ 48,89% (bổ sung 6 ppm) đến 84,45% (bổ sung 4 ppm). Đối với chỉ tiêu số chồi trung bình, ở công thức không bổ sung nano bạc thu được 1,10 chồi/mô sẹo, trong khi đó các công thức bổ sung nano bạc dao động từ 2,14 đến 3,29 chồi/mô sẹo. Trong các công thức thí nghiệm, công thức bổ sung 4 ppm nano bạc cho kết quả tốt nhất với tỉ lệ tái sinh chồi đạt 84,45% và hệ số tái sinh chồi đạt 3,29 chồi/mô sẹo.
3.3. Ảnh hưởng của nano bạc đến khả năng nhân nhanh từ chồi in vitro hoa Đồng tiền
Nhân nhanh chồi là giai đoạn không thể thiếu với một quy trình sản xuất giống có tính chất công nghiệp bằng phương pháp nhân giống in vitro. Trong giai đoạn này sự sinh trưởng và phát triển của mô nuôi cấy phụ thuộc vào điều kiện nuôi cấy (nhiệt độ, ánh sáng), môi trường nuôi cấy (thành phần dinh dưỡng, chất điều tiết sinh trưởng và tỷ lệ giữa chúng). Tùy từng loại cây mà người ta có thể bổ sung vào môi trường nuôi cấy các chất điều tiết sinh trưởng với nồng độ và tỷ lệ khác nhau để thu được hệ số nhân chồi và chất lượng chồi tốt nhất. Kết quả nghiên cứu ảnh hưởng của nano bạc đến khả năng nhân nhanh từ chồi in vitro hoa Đồng tiền được thể hiện ở Bảng 3 và Hình 2.
Bảng 3. Ảnh hưởng của nano bạc đến khả năng nhân nhanh chồi hoa Đồng tiền
0 ppm 6 ppm 8 ppm 10 ppm 12 ppm
Hình 1. Ảnh mô sẹo hoa Đồng tiền sau 4 tuần nuôi cấy trong môi trường có bổ sung nano bạc với các nồng độ khác nhau
Nồng độ nano
bạc (ppm)
Số mẫu
Số mô sẹo
tái sinh chồi
Tỷ lệ tái sinh
chồi (%)
Tổng số
chồi
Số chồi trung bình (chồi/mô
sẹo)0 45 19 42,22d 21 1,10d
2 45 29 64,45b 62 2,14c
4 45 38 84,45a 125 3,29a
6 45 22 48,89c 56 2,55b
CV (%) 11,2 4,6LSD0,05 3,8 0,2
Nồng độ
nano bạc
Hệ số nhân chồi Chiều cao trung bình sau 6 tuần (cm)
Đặc điểm chồi sau 6 tuần nuôi
cấy2
tuần4
tuần6
tuần
0 ppm 1,17c 3,38c 5,71c 4,22bChồi xanh, phát triển
bình thường
2 ppm 2,42a 6,62a 8,22a 5,75a Chồi xanh, phát triển tốt
4 ppm 1,62b 3,78b 6,55b 2,86cChồi xanh, phát triển không đều
6 ppm 0,62d 2,35d 3,44d 2,13dChồi có màu
tím, phát triển kém
CV (%) 1,2 2,5 2,9 2,3
LSD0,05 0,14 0,19 0,33 0,21

58
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Từ kết quả thu được cho thấy nồng độ nano bạc khác nhau có ảnh hưởng khác nhau đến quá trình nhân nhanh chồi hoa Đồng tiền. Cụ thể, ở công thức bổ sung 2 ppm và 4 ppm nano bạc cho hệ số nhân chồi cao hơn có ý nghĩa so với công thức đối chứng không bổ sung nano bạc ở cả 3 thời gian theo dõi. Đặc biệt ở công thức bổ sung 2 ppm nano bạc cho kết quả tốt nhất với hệ số nhân chồi là 8,22 lần sau 6 tuần nuôi cấy, chiều cao trung bình chồi đạt 5,75 cm, các chồi xanh, phát triển tốt và đồng đều. Tuy nhiên khi tăng nồng độ nano bạc lên 4 ppm và 6 ppm thì hệ số nhân chồi, chiều cao trung bình chồi và chất lượng chồi đều giảm đi đáng kể. Ở công thức bổ sung 4 ppm nano bạc, hệ số nhân chồi sau 6 tuần nuôi cấy đạt 6,55 lần, cao hơn so với công thức không bổ sung nano bạc (5,71), tuy nhiên ở công thức này chồi được tạo ra có cuống lá kém phát triển nên chiều cao trung bình chồi chỉ đạt 2,36 cm, thấp hơn so với công thức không bổ sung nano bạc (4,22 cm). Ở công thức bổ sung 6 ppm nano bạc thì cả 3 chỉ tiêu là hệ số nhân chồi, chiều cao trung bình chồi và chất lượng chồi đều cho kết quả thấp nhất, xuất hiện nhiều chồi dị dạng, chồi có nhiều lá chuyển sang màu tím (Hình 2).
Theo kết quả nghiên cứu của Nguyễn Thị Mỹ Duyên và cộng tác viên (2014), khi sử dụng các chất điều tiết sinh trưởng khác nhau để nhân chồi hoa Đồng tiền thì kết quả thu được tốt nhất sau 6 tuần nuôi cấy là hệ số nhân chồi đạt 5,4 lần, chiều cao chồi đạt 1,87 cm; Son và cộng tác viên (2011) thu được hệ số nhân chồi tốt nhất khi nhân nhanh chồi 3 giống hoa đồng tiền là là 7,4 lần đối với giống Arianna, 4,6 lần đối với giống Bonnie và 5,4 lần đối với giống Tobia. Như vậy có thể nhận thấy, bổ sung 2 ppm nano bạc vào môi trường nhân nhanh chồi hoa đồng tiền cho hiệu quả tốt hơn so với kết quả của các nghiên cứu trước đã công bố.
3.4. Ảnh hưởng của nano bạc đến khả năng ra rễ của chồi hoa Đồng tiền
Kích thích cho chồi ra rễ là công đoạn cuối cùng
trong quy trình nhân giống bằng phương pháp nuôi cấy in vitro. Kết thúc giai đoạn này chúng ta sẽ thu được những cây hoàn chỉnh (đầy đủ rễ và thân, lá) cung cấp cho giai đoạn luyện cây trong vườn ươm. Kết quả nghiên cứu ảnh hưởng của nano bạc đến khả năng ra rễ của chồi hoa Đồng tiền sau 4 tuần nuôi cấy được thể hiện ở Bảng 4.
Bảng 4. Ảnh hưởng của nồng độ nano bạc đến khả năng ra rễ của chồi hoa Đồng tiền sau 4 tuần nuôi cấy
Kết quả thu được trong bảng 4 cho thấy nano bạc có ảnh hưởng tích cực đến khả năng ra rễ của chồi hoa Đồng tiền in vitro, ở tất cả các công thức bổ sung nano bạc đều cho số rễ/chồi và chiều dài rễ cao hơn so với công thức không bổ sung nano bạc. Đối với chỉ tiêu số rễ/chồi, ở công thức không bổ sung nano bạc là 1,73 rễ/chồi, trong khi đó chỉ tiêu này ở các công thức bổ sung nano bạc dao động từ 2,53 (ở công thức bổ sung 6 ppm nano bạc) đến 5,73 (ở công thức bổ sung 4 ppm nano bạc). Đối với chỉ tiêu chiều dài rễ, ở công thức không bổ sung nano bạc chiều dài trung bình rễ là 2,50 cm, trong khi đó chỉ tiêu này ở các công thức bổ sung nano bạc dao động từ 3,33 cm (ở công thức bổ sung 2 ppm nano bạc) đến 5,93 cm (ở công thức bổ sung 4 ppm nano bạc). Công thức cho hiệu quả ra rễ tốt nhất là công thức bổ sung 4 ppm nano bạc với 5,73 rễ/chồi và chiều dài rễ trung bình đạt 5,93 cm.
0 ppm 2 ppm 4 ppm 6 ppm
Hình 2. Chồi hoa Đồng tiền in vitro sau 6 tuần nuôi cấy trong môi trường có bổ sung các nồng độ nano bạc khác nhau
Nồng độ nano bạc
(ppm)
Tỷ lệ ra rễ
(%)
Số rễ/chồi(rễ)
Chiều dài rễ (cm)
Màu sắc rễ
0 100 1,73d 2,50d Vàng nhạt2 100 3,47b 3,33c Vàng nhạt4 100 5,73a 5,93a Vàng nhạt6 100 2,53c 4,23b Vàng nhạt
CV (%) 1,9 4,0LSD0,05 0,12 0,30

59
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
IV. KẾT LUẬN- Môi trường thích hợp nhất cho việc tạo mô sẹo
từ mâu lá in vitro hoa Đồng tiền là môi trường MS có bổ sung 30 g/l saccharose; 6,5 g/l agar; 1,5 mg/l 2,4D; pH 5,7 và 10 ppm nano bạc. Sau 4 tuần nuôi cấy, tỉ lệ tạo mô sẹo đạt 95,56%, mô sẹo mềm và có màu sáng.
- Khả năng tái sinh chồi từ mô sẹo đạt hiệu quả cao nhất trong môi trường MS có bổ sung 0,7 mg/l BA;0,1 mg/l IAA và 4 ppm nano bạc, tỉ lệ tái sinh chồi đạt 84,45%, số chồi trung bình đạt 3,29 chồi/mô sẹo sau 6 tuần nuôi cấy.
- Hiệu quả nhân nhanh chồi hoa Đồng tiền tốt nhất trong môi trường MS có bổ sung 3 mg/l BAP; 0,1 mg/l α- NAA và 2 ppm nano bạc, hệ số nhân chồi đạt 8,22 lần, chiều cao chồi đạt 5,75 cm sau 6 tuần nuôi cấy.
- Chồi in vitro hoa Đồng tiền ra rễ hiệu quả nhất trong môi trường MS có bổ sung 50 g/l saccharose; 6,5 g/l agar; 1,0 mg/l α-NAA và 4 ppm nano bạc, với 5,73 rễ/chồi và chiều dài rễ trung bình đạt 5,93 cm sau 4 tuần nuôi cấy.
LỜI CẢM ƠNNghiên cứu này được hỗ trợ kinh phí từ đề tài
“Nghiên cứu sử dụng nano bạc trong nuôi cấy in vitro hoa Đồng tiền (Gerbera jamesonii Bolus ex Hook. F.)”, mã số: SV 2017 - 12 - 14 MST.
TÀI LIỆU THAM KHẢONguyễn Thị Mỹ Duyên, Trương Thị Hằng, 2014. Khảo
sát khả năng tạo mô sẹo từ cuống lá, phiến lá và nụ
hoa non phục vụ cho việc vi nhân giống hoa Đồng tiền (Gerbera jamesonii Bolus). Tạp chí Khoa học, Trường Đại học An Giang, 4 (3): 114-120.
Nguyễn Văn Hồng, 2009. Nghiên cứu biện pháp kỹ thuật nhân giống hoa Đồng tiền bằng phương pháp nuôi cấy mô tại Thái Nguyên. Luận văn Thạc sĩ Nông nghiệp. Đại học Nông Lâm - Đại học Thái Nguyên.
Phạm Thị Bích Ngoc, Phan Ngô Hoàng, 2009. Tìm hiểu sự phát sinh chồi từ mô sẹo lá cây dây chiều (Tetracera scandens L.). Tạp chí Phát triển KH&CN, 12 (7): 79-85.
Đỗ Năng Vịnh, 2003. Nghiên cứu hoàn thiện quy trình nhânh nhanh các giống hoa Đồng tiền nhập nội bằng công nghệ in vitro. Tạp chí Nông nghiệp và Phát triển nông thôn, 8: 1012-1014.
Bhavya Bhargava, B. S. Dilta, Y.C. Gupta, S.R. Dhiman & Manju Modgil, 2013. Studies on micropropagation of gerbera (Gerbera jamesonii Bolus). Indian Journal of Applied Research, 3 (11): 8-11.
Emad A. Ewais, Said A. Desouky, Ezzat H. Elshazly, 2015. Evaluation of Mô sẹo Responses of Solanum nigrum L. Exposed to Biologically Synthesized Silver Nanoparticles. Nanoscience and Nanotechnology, 5 (3): 45-56.
Murashige T. & F. Skoog, 1962. A revised medium for rapid growth and bioassay with tobacco tissue cultures. Physiol Plant, 15: 473-497.
Son N.V., A.N. Mokashi, R. Hegde, V.S. Patil & S. Lingaraju, 2011. Response of gerbera (Gerbera jamesonii Bolus) varieties to micropropagation. Karnataka J. Agric. Sci., 24 (3): 354-357.
Hình 3. Rễ hoa Đồng tiền trong môi trường có bổ sung các nồng độ nano bạc khác nhau sau 2 tuần nuôi cấyGhi chú: CT1: 0 ppm; CT2: 2 ppm; CT3: 4 ppm; CT4: 6 ppm.
Hình 4. Rễ hoa Đồng tiền trong môi trường có bổ sung các nồng độ nano bạc khác nhau sau 4 tuần nuôi cấy0 ppm 2 ppm 4 ppm 6 ppm

60
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Paduchuri, V.Y., G.V. Deogirkar, S.R. Kamdi, M.C. Kale & M.D. Rajurkar, 2010. In-vitro Callus Induction and Root Regeneration Studies in Gerbera jamesonii. International Journal of Advanced Biotechnology and Research, 1 (2): 87-90.
Rostami A.A. and A. Shahsavar, 2009. Nano-Silver Particles Eliminate the in vitro contaminations of Olive ‘Mission’ Explants. Journal of Plant Sciences, 8 (7): 505-509.
Shokri S., A. Babaei, M. Ahmadian, M.M. Arab and S. Hessami, 2015. The effects of different concentrations of nano - silver on elimination of
bacterial contaminations and phenolic exudation of rose (Rosa hybrida L.) in vitro culture. International Society for Horticultural Science, 3 (1): 50-54.
Shylaja M.R., P. Sashna, V. Chinjusha and P.A. Nazeem, 2014. An efficient micropropagation protocol for Gerbera jamesonii Bolus from flower buds. International Journal of Plant, Animal and Environmental Sciences, 4 (3): 641-643.
Zainab M. Almutairi, 2016. Influence of Silver Nano-particles on the Salt Resistance of Tomato (Solanum lycopersicum) during Germination. Int. J. Agric. Biol., 18(2): 449-457.
NGHIÊN CỨU ĐA DẠNG DI TRUYÊN CÂY BÁT GIÁC LIÊN THU THẬP Ở MIÊN BẮC VIỆT NAM BẰNG CHỈ THỊ RAPD
Lưu Thúy Hòa1, Khuất Hữu Trung2, Trần Đăng Khánh2, Phạm Thị Lý Thu2, Trần Văn Ơn3
TÓM TẮTBát giác liên (Dysosma Woodson) là chi thuộc họ Berberidaceae có hoạt chất berberine với hàm lượng cao và
được sử dụng rộng rãi trong y học cổ truyền. Trong nghiên cứu này, 10 mâu Bát giác được phân tích bằng 25 chỉ thị RAPD. Kết quả thu được 17 chỉ thị cho đa hình và 816 băng ADN gồm 111 loại băng khác nhau (trung bình là 6,5 loại băng/mồi), trong đó có 62 băng đa hình (55,86%) và 49 băng đơn hình (44,14%). Các băng có kích thước dao động từ khoảng 250 đến 2000bp. Giá trị PIC của 17 mồi trong khoảng 0,5 đến 0,88. Giá trị trung bình của hệ số PIC là 0,78 phản ánh các băng ADN được nhân lên là khá đa dạng. Kết quả phân tích hệ số tương đồng di truyền của 10 mâu nghiên cứu liên dao động từ 0,57 đến 0,91. Ở mức tương đồng di truyền khoảng 0,67 có thể chia 10 mâu nghiên cứu thành 2 nhóm cách biệt di truyền. Kết quả nghiên cứu cũng chỉ ra 9 trong số 17 mồi RAPD có thể nhận biết chính xác 5 mâu giống nghiên cứu là PO1, PO2, PO9, PO10 và PO11. Kết quả này rất có ý nghĩa trong việc nghiên cứu tính đúng giống phục vụ công tác chọn tạo giống, nhân nhanh, bảo tồn và kiểm soát cây giống có chất lượng cao.
Từ khóa: Bát giác liên, Dysosma Woodson, RAPD, đa dạng di truyền
Effects of nanosilver on morphogenesis of tissue cultured gerbera (Gerbera jamesonii)
Bui Thi Thu Huong, Dong Huy Gioi, Tran Thi Thu Thuy, Nguyen Thi Ngoc Quynh
AbstractThe study aimed to evaluate the effects of silver nanoparticles on in vitro propagation of gerbera by in vitro leaf material. The results showed that: (i) the optimal medium for formation of callus from the in vitro leaf piece of gerbera was MS medium containing 30 g/l saccharose, 6.5 g/l agar, 1.5 mg/l 2.4D, 10 ppm of silvernano, the rate of callus formation was 92.53%; (ii) 85.45% of callus formed shoot after 6 weeks culturing on the MS medium supplemented with 0.7 mg/l BA, 0.1 mg/l IAA and 4 ppm silvernano; (iii) the maximum number of shoots was obtained on MS medium containing 3 mg/l BAP, 0.1 mg/l α- NAA, 2 ppm of silvernano and the multiplication coefficient reached 8.22 times and shoot height was 5.75 cm after 6 weeks culturing; (iv) The optimum medium for in vitro rooting was the MS medium supplemented with 50 g/l saccharose, 6.5 g/l agar, 1.0 mg/l α-NAA, 4 ppm of silvernano; 100% of the shoots produced roots, with an average of 5.73 roots/shoot and the average length of roots was 5.93 cm after 4 weeks culturing.Keywords: Gerbera, in vitro leaf, MS medium, nanosilver
Ngày nhận bài: 16/9/2018Ngày phản biện: 24/9/2018
Người phản biện: TS. Nguyễn Thị PhaNgày duyệt đăng: 15/10/2018
1 Đại học Hải Phòng; 2 Viện Di truyền Nông nghiệp, 3 Đại học Dược Hà Nội.

61
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
I. ĐẶT VẤN ĐỀ Bát giác liên (Dysosma Woodson) là chi thuộc họ
Berberidaceae. Chi Dysosma Woodson được đánh giá là một trong số các chi có hoạt chất berberine với hàm lượng cao. Berberin là alcaloid có đặc tính kháng khuẩn, nấm, đơn bào… được ứng dụng từ lâu đời trong các nền y học truyền thống trên thế giới. Đến nay hoạt chất này còn được đánh giá có tiềm năng rất lớn trong điều trị các bệnh đang phát triển như tiểu đường, ung thư, tăng mỡ máu (Wang et al., 2010).
Ở nước ta, chi Dysosma Woodson được biết đến chỉ có một loài duy nhất với tên thường gọi Bát giác liên (Dysosma difformis). Mặc dù vậy Bát giác liên phân bố rộng ở nhiều vùng sinh thái khác nhau, đặc biệt là các tỉnh miền núi phía bắc như Lai Châu, Lào Cai, Ba Vì, và Hòa Bình (Nguyễn Kim Cẩn, 2002).
Sự phân bố của Bát giác liên phản ánh sự thích nghi đa dạng của loài, chi ở các khu vực địa lý khác nhau, do vậy đánh giá đa dạng di truyền và xác định mối tương quan với hàm lượng berberin của loài, chi là cần thiết để bảo tồn và phát triển các nguồn gen Dysosma Woodson bản địa. Đặc biệt là hiện nay nguồn gen Bát giác liên đang được khuyến cáo dần dần cạn kiệt (Nguyễn Tiến Bân, 2007).
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưuGồm 10 mâu Bát Giác liên (Dysosma Woodson)
được thu thập ở độ cao từ 800 - 1.300 m so với mặt nước biển, ở nơi ẩm, dưới tán cây rừng. Địa điểm thu mâu được định vị GPS và mâu được lưu tiêu bản tại Bộ môn Thực vật, Trường Đại học Dược Hà Nội (Bảng 1).
Hai mươi lăm mồi khác nhau sử dụng trong các phản ứng PCR-RAPD có độ dài 9 - 10 nucleotide thuộc các nhóm BIO, OPA, OPN, OPO, S và UBC của hãng Bioneer.
2.2. Phương pháp nghiên cưu
2.2.1. Tách chiết ADN tổng số và RAPD-PCRADN tổng số được tách chiết từ mâu lá theo
phương pháp CTAB của Obara và Kako (Obara and Kako, 1998) có một số cải tiến. Xác định nồng độ và chất lượng ADN bằng máy quang phổ Jenway - Model 6715. Các mâu ADN tổng số được pha loãng với đệm TE vô trùng đến nồng độ 50 ng/µl để thực hiện phản ứng PCR.
2.2.2. Phương pháp PCR và điện di sản phâm PCRTổng thể tích phản ứng PCR là 15µl gồm: 1,5µl
Buffer MgCl2 25mM; 0,5µl dNTPs 10mM; 1,2µl Taq ADN polymerase 5U/µl; 1,5µl Mồi 10µM; 2µl ADN khuôn nồng độ 50 ng và 8,3µl nước cất hai lần khử ion.
Phản ứng PCR được tiến hành trên máy PCR Mastercycler epgradient S theo chu trình nhiệt sau: 950C trong 5 phút, 38 - 40 chu kỳ (940C trong 1 phút, 33 - 350C trong 1 phút 10 giây, 720C trong 1 phút 55 giây); 720C trong thời gian 7 phút.
Sản phẩm PCR được điện di trên gel agarose 1% và được phát hiện dưới tia UV bằng phương pháp nhuộm ethidium bromide.
2.2.3. Xử lý số liệuSố liệu thu thập được xử lý theo phương pháp
thống kê sinh học trên nền Excel 2014. Hệ số PIC
Bảng 1. Danh sách ký hiệu các mâu Bát giác liên (Dysosma Woodson)
TT MẫuVị trí Thời gian
thu mẫu Địa điểm thu mẫuTọa độ X Tọa độ Y
1 PO1 21o4’44.80”N 105o21’54.90”E 2/2016 Vườn Quốc gia Ba Vì2 PO2 22o17’44.40”N 105o28’11.40”E 3/2016 Bản Thi, Chợ Đồn, Bắc Kạn 3 PO3 21o4’44.80”N 105o21’54.90”E 3/2016 Vườn Quốc gia Ba Vì4 PO5 21o4’33.40”N 105o21’45.20”E 3/2016 Vườn Quốc gia Ba Vì5 PO6 21o4’23.00”N 105o21’43.20”E 3/2016 Vườn Quốc gia Ba Vì6 PO7 21o4’21.70”N 105o21’43.30”E 3/2016 Vườn Quốc gia Ba Vì7 PO8 21o4’33.10”N 105o21’58.70”E 3/2016 Vườn Quốc gia Ba Vì
8 PO9 23o5’6.00”N 104o56’45.50”E 3/2016 Thanh Long, Thanh Vân, Quản Bạ, Hà Giang
9 PO10 21o36’39.25”N 104o35’22.38”E 4/2016 Văn Chấn, Yên Bái 10 PO11 22o16’29.03”N 105o31’13.14”E 4/2016 Bản Thi, Chợ Đồn, Bắc Kạn
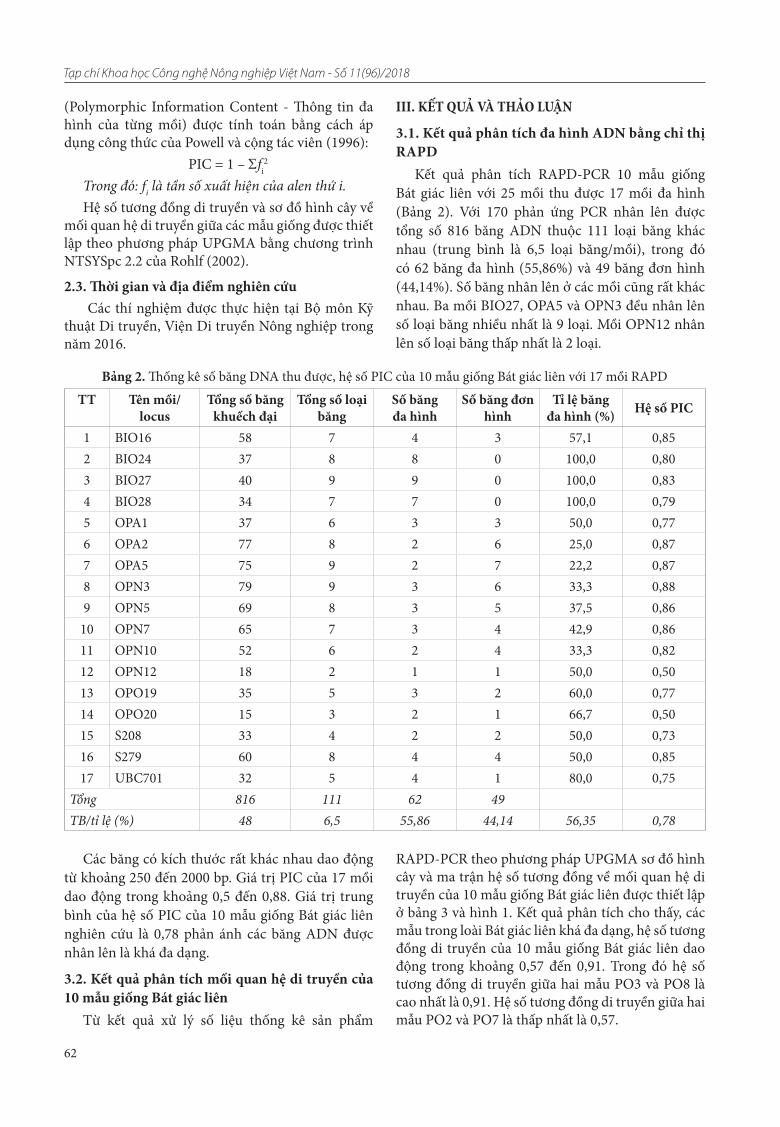
62
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
(Polymorphic Information Content - Thông tin đa hình của từng mồi) được tính toán bằng cách áp dụng công thức của Powell và cộng tác viên (1996):
PIC = 1 – Sfi2
Trong đó: fi là tần số xuất hiện của alen thứ i.Hệ số tương đồng di truyền và sơ đồ hình cây về
mối quan hệ di truyền giữa các mâu giống được thiết lập theo phương pháp UPGMA bằng chương trình NTSYSpc 2.2 của Rohlf (2002).
2.3. Thời gian và địa điểm nghiên cưu Các thí nghiệm được thực hiện tại Bộ môn Kỹ
thuật Di truyền, Viện Di truyền Nông nghiệp trong năm 2016.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Kết quả phân tích đa hình ADN bằng chỉ thị RAPD
Kết quả phân tích RAPD-PCR 10 mâu giống Bát giác liên với 25 mồi thu được 17 mồi đa hình (Bảng 2). Với 170 phản ứng PCR nhân lên được tổng số 816 băng ADN thuộc 111 loại băng khác nhau (trung bình là 6,5 loại băng/mồi), trong đó có 62 băng đa hình (55,86%) và 49 băng đơn hình (44,14%). Số băng nhân lên ở các mồi cũng rất khác nhau. Ba mồi BIO27, OPA5 và OPN3 đều nhân lên số loại băng nhiều nhất là 9 loại. Mồi OPN12 nhân lên số loại băng thấp nhất là 2 loại.
Các băng có kích thước rất khác nhau dao động từ khoảng 250 đến 2000 bp. Giá trị PIC của 17 mồi dao động trong khoảng 0,5 đến 0,88. Giá trị trung bình của hệ số PIC của 10 mâu giống Bát giác liên nghiên cứu là 0,78 phản ánh các băng ADN được nhân lên là khá đa dạng.
3.2. Kết quả phân tích mối quan hệ di truyền của 10 mẫu giống Bát giác liên
Từ kết quả xử lý số liệu thống kê sản phẩm
RAPD-PCR theo phương pháp UPGMA sơ đồ hình cây và ma trận hệ số tương đồng về mối quan hệ di truyền của 10 mâu giống Bát giác liên được thiết lập ở bảng 3 và hình 1. Kết quả phân tích cho thấy, các mâu trong loài Bát giác liên khá đa dạng, hệ số tương đồng di truyền của 10 mâu giống Bát giác liên dao động trong khoảng 0,57 đến 0,91. Trong đó hệ số tương đồng di truyền giữa hai mâu PO3 và PO8 là cao nhất là 0,91. Hệ số tương đồng di truyền giữa hai mâu PO2 và PO7 là thấp nhất là 0,57.
Bảng 2. Thống kê số băng DNA thu được, hệ số PIC của 10 mâu giống Bát giác liên với 17 mồi RAPD
TT Tên mồi/locus
Tổng số băng khuếch đại
Tổng số loại băng
Số băng đa hình
Số băng đơn hình
Tỉ lệ băng đa hình (%) Hệ số PIC
1 BIO16 58 7 4 3 57,1 0,852 BIO24 37 8 8 0 100,0 0,803 BIO27 40 9 9 0 100,0 0,834 BIO28 34 7 7 0 100,0 0,795 OPA1 37 6 3 3 50,0 0,776 OPA2 77 8 2 6 25,0 0,877 OPA5 75 9 2 7 22,2 0,878 OPN3 79 9 3 6 33,3 0,889 OPN5 69 8 3 5 37,5 0,86
10 OPN7 65 7 3 4 42,9 0,8611 OPN10 52 6 2 4 33,3 0,8212 OPN12 18 2 1 1 50,0 0,5013 OPO19 35 5 3 2 60,0 0,7714 OPO20 15 3 2 1 66,7 0,5015 S208 33 4 2 2 50,0 0,7316 S279 60 8 4 4 50,0 0,8517 UBC701 32 5 4 1 80,0 0,75
Tổng 816 111 62 49TB/tỉ lệ (%) 48 6,5 55,86 44,14 56,35 0,78

63
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Bảng 3. Hệ số tương đồng di truyền giữa 10 mâu Bát giác liên
Hình 2. Kết quả điện di sản phẩm RAPD-PCR của 10 mâu nghiên cứu với mồi S208 và OPA5 (M: GenRulerTM 1kb DNA Ladder)
Ở mức tương đồng di truyền khoảng 0,67 có thể chia 10 mâu nghiên cứu thành 2 nhóm cách biệt di truyền.
Nhóm 1 gồm các mâu giống: PO1, PO3, PO7, PO8 và PO11 được chia thành hai nhóm phụ, nhóm phụ 1.1 và nhóm phụ 1.2. Mâu PO1 được thu thập từ Vườn Quốc gia Ba Vì tách riêng thành nhóm phụ 1.1. Bốn mâu còn lại thuộc nhóm phụ 1.2. Trong số đó 3 mâu PO3, PO7 và PO8 đều thu thập từ Vườn Quốc gia Ba Vì, còn PO11 thu thập từ Bản Thi, Bắc Kạn.
Hình 1. Sơ đồ hình cây về mối quan hệ di truyền giữa các mâu giống nghiên cứu bằng chỉ thị RAPD
Nhóm 2 gồm các mâu giống: PO2, PO6, PO5, PO9 và PO10 được chia thành hai nhóm phụ, nhóm
phụ 2.1 và nhóm phụ 2.2. Nhóm phụ 2.1 gồm mâu giống PO2 thu thập từ Bản Thi, Chợ Đồn, Bắc Kạn và PO6 thu thập từ Vườn Quốc gia Ba Vì. Nhóm phụ 2.2 gồm 3 mâu giống là PO5, PO9 và PO10. Mâu giống PO5 được thu thập từ Vườn Quốc gia Ba Vì, PO9 được thu thập từ Thanh Vân, Quản Bạ, Hà Giang và PO10 được thu thập từ Văn Chấn, Yên Bái.
Như vậy, kết quả phân tích đa dạng di truyền dựa vào chỉ thị RAPD đối với 10 mâu Bát giác liên nghiên cứu cho thấy khác biệt di truyền các mâu không bị ảnh hưởng bởi các vùng phân bố. Sự đa dạng về cấu trúc của loài Bát giác liên ở Việt Nam khá cao (hệ số tương đồng giữa các mâu giống trong loài thấp) có thể do sự di thực, chuyển vùng địa lý hoặc do sinh sản thấp. Chính vì vậy, việc bảo tồn, chọn tạo giống và phát triển nguồn gen cây thuốc trong loài Bát giác liên là việc làm cần thiết.
3.3. Kết quả xác định các chỉ thị đặc trưng nhận dạng một số nguồn gen Bát giác liên
Kết quả phân tích đa hình ADN bằng 17 mồi RAPD với 10 mâu Bát giác liên đã thu được 6 băng cá biệt (băng chỉ xuất hiện ở một mâu duy nhất và không xuất hiện ở các mâu còn lại đối với một mồi) và 3 băng khuyết cá biệt (băng không xuất hiện ở một mâu tại vị trí tương ứng có băng của các mâu còn lại) (Hình 2, Bảng 4).
PO1 PO2 PO3 PO5 PO6 PO7 PO8 PO9 PO10 PO11PO1 1,00PO2 0,64 1,00PO3 0,65 0,66 1,00PO5 0,66 0,78 0,67 1,00PO6 0,58 0,83 0,76 0,70 1,00PO7 0,74 0,57 0,73 0,59 0,69 1,00PO8 0,67 0,62 0,91 0,65 0,73 0,77 1,00PO9 0,60 0,65 0,67 0,76 0,72 0,65 0,73 1,00PO10 0,58 0,60 0,60 0,74 0,64 0,60 0,65 0,82 1,00PO11 0,66 0,62 0,72 0,62 0,70 0,69 0,73 0,70 0,68 1,00

64
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Bảng 4. Thống kê băng DNA cá biệt của các mâu giống với các mồi RAPD
TT Tên mồi/locus
Mẫu có băng cá biệt
Băng cá biệt
Có băng Kích thướcbăng cá biệt (bp) Băng khuyết Vị trí băng khuyết
cá biệt (bp)1 BIO24 PO2 ˟ 10002 BIO28 PO10 ˟ 7303 OPA2 PO11 ˟ 12504 OPA5 PO11 ˟ 12005 OPN5 PO9 ˟ 3006 OPN7 PO2 ˟ 4007 OPN10 PO1 ˟ 18008 S208 PO10 ˟ 3709 UBC701 PO1 ˟ 500
Qua bảng 4 và hình 2 cho thấy: Mồi BIO24 xuất hiện 1 băng cá biệt với kích thước khoảng 1000 bp ở mâu PO2. Mồi BIO28 xuất hiện 1 băng cá biệt với kích thước khoảng 730 bp ở mâu PO10. Mồi OPA2 xuất hiện băng khuyết cá biệt với kích thước 1.250 bp ở PO11. Mồi OPA5 xuất hiện băng khuyết cá biệt với kích thước 1.200 bp ở PO11. Mồi OPN5 xuất hiện 1 băng cá biệt với kích thước khoảng 300 bp ở mâu PO9. Mồi OPN7 xuất hiện băng cá biệt có kích thước khoảng 400 bp ở mâu PO2. Mồi OPN10 xuất hiện băng cá biệt có kích thước khoảng 1800 bp ở mâu PO1. Mồi S208 xuất hiện 1 băng cá biệt với kích thước khoảng 370 bp ở mâu PO10. Mồi UBC701 xuất hiện băng khuyết cá biệt có kích thước khoảng 500 bp ở mâu PO1. Kết quả nghiên cứu cho thấy 9 trong số 17 mồi thí nghiệm có thể nhận biết chính xác 5 mâu nghiên cứu là PO1, PO2, PO9, PO10 và PO11. Kết quả này rất có ý nghĩa trong việc nghiên cứu tính đúng giống phục vụ nhân nhanh, bảo tồn và kiểm soát cây giống có chất lượng cao.
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luậnCác mâu giống trong loài Bát giác liên thu thập
tại một số tỉnh miền núi phía Bắc của Việt Nam khá đa dạng. Kết quả phân tích RAPD-PCR 10 mâu giống Bát giác liên với 25 mồi thu được 17 mồi đa hình. Giá trị PIC của 17 mồi dao động trong khoảng 0,5 đến 0,88. Giá trị trung bình của hệ số PIC của 10 mâu giống Bát giác liên nghiên cứu là 0,78 phản ánh các băng ADN được nhân lên là khá đa dạng. Kết quả phân tích cho thấy, hệ số tương đồng di truyền của 10 mâu giống Bát giác liên dao động trong khoảng 0,57 đến 0,91. Ở mức tương đồng di truyền khoảng 0,67 có thể chia 10 mâu nghiên cứu
thành 2 nhóm cách biệt di truyền. Kết quả nghiên cứu cũng chỉ ra 9 trong số 17 mồi thí nghiệm có thể nhận biết chính xác 5 mâu giống là PO1, PO2, PO9, PO10 và PO11. Kết quả này rất có ý nghĩa trong việc xác định tính đúng giống phục vụ công tác chọn tạo giống, nhân nhanh, bảo tồn và kiểm soát cây giống có chất lượng cao.
4.2. Đề nghịCần xác định hàm lượng hoạt chất berberine
trong các mâu thu thập nhằm phục vụ công tác bảo tồn và phát triển các loài dược liệu quý.
TÀI LIỆU THAM KHẢONguyễn Tiến Bân, chủ biên, 2007. Sách đỏ Việt Nam,
Tập 2, Phần Thực vật. NXB KHTN&CN.Nguyễn Kim Cẩn, 2002. Nghiên cứu những cây chứa
Berberin trên thế giới và trong nước. Tạp chí Dược liệu, Tập 7, số 1, 2, 3, 4.
Obara, O.P., Kako, S., 1998. Genetic diversity and identification of Cymbidium cultivars as measured by amplified polymorphic RAPD markers. Euphytica, 99: 95-101.
Powell W, Morgante M, Andre C, Hanafey M, Vogel J, Tingey S and Rafalski A, 1996. The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for gemplasm analysis. Molecular Breeding, 2: 225-238.
Rohlf F, 2002, Estimation of the coancestry coefficient: Basic for a short term genetic distance. Genetics, 105:767-779.
Wang Y, Campbell T, Perry B, Beaurepaire C, Qin L., 2010. Hypoglycemic and insulin-sensitizing effects of berberine in high-fat diet- and streptozotocin-induced diabetic rats. Metabolism, 60 (2): 298-305.

65
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Study on genetic diversity of Dysosma by RAPD markersLuu Thuy Hoa, Khuat Huu Trung, Tran Dang Khanh, Pham Thi Ly Thu, Tran Van On
Abstract Dysosma (Woodson) is a genus, belonging to Berberidaceae family. This plant contains high berberine constituent and widely used as the medicinal treatments in many countries. Ten samples of Dysosma Woodson collected from some different geographical areas in northern Vietnam, were evaluated for their genetic diversity by applying 25 different RAPD markers, of which 17 markers showed polymorphism. Total 170 PCR reactions obtained a total of 816 DNA bands that belonged to 111 different patterns (average 6.5 patterns/primer). Of which, 62 bands were polymorphic bands (55.86%) and 49 monomorphic bands (44.14%). The obtained bands sized from 250 bp to 2000 bp. PIC value of 17 primers changed from 0.5 to 0.88 with a mean of 0.78 which showed amplified DNA bands are very diversified. The analyzed result showed that samples of Dysosma in our research were very diverse. Genetic similarity coefficients of 10 samples of Dysosma ranged from 0.57 to 0.91. At genetic similarity coefficient of 0.67, total 10 studied samples was divided into 2 different genetic groups. The results showed that 9 out of 17 used RAPD primers defined exactly 5 studied samples (PO1, PO9, PO10 and PO11). This results may be useful for the classification and identification of the genus indigenous nomadic dysentery (Dysosma) for conservation and control of high quality seedlings. Keywords: Diphtheria, Dysosma, RAPD, genetic diversity
Ngày nhận bài: 18/9/2018Ngày phản biện: 25/9/2018
Người phản biện: PGS. TS. Đồng Huy GiớiNgày duyệt đăng: 15/10/2018
1 Viện Di truyền Nông nghiệp; 2 Trung tâm Tài nguyên thực vật; 3 Bộ Nông nghiệp và PTNT
NGHIÊN CỨU ĐA DẠNG DI TRUYÊN CỦA ĐOẠN GEN rbcL Ở MỘT SỐ NGUỒN GEN BƯỞI VIỆT NAM
Nguyễn Thị Ngọc Lan1, Nguyễn Thị Lan Hoa2, Nguyễn Thị Thanh Thủy3, Lã Tuấn Nghĩa2
TÓM TẮTBưởi là một trong những cây ăn quả nhiệt đới quan trọng có giá trị kinh tế ở nhiều nước trên thế giới. Nghiên
cứu đa dạng di truyền đoạn gen rbcL của tập đoàn 25 nguồn gen bưởi Việt Nam đã xác định được 2 trình tự nucleotit đặc trưng cho giống bưởi Thanh Trà (G2) và bưởi Da Xanh (G27). Đột biến điểm được tìm thấy là đột biến đồng hoán (C>T) tại vị trí 595 của gen ở cả 2 giống bưởi Thanh Trà (G2) và Da Xanh (G27), đột biến này có ý nghĩa trong nhận dạng các nguồn gen bưởi Thanh Trà và bưởi Da Xanh của nước ta. Hai trình tự này đã được đăng ký NCBI với số đăng ký lần lượt là KR073282 và KR073281. Phân tích cây phả hệ bằng phương pháp Neighbor Joining dựa trên trình tự của đoạn gen rbcL 600 nucleotid đã nhóm thành công được các trình tự của chi Citrus thuộc họ Rutaceae, tách biệt rõ ràng được hai giống bưởi Thanh Trà và Da Xanh của Việt Nam.
Từ khóa: Bưởi, giải trình tự, ADN mã vạch, rbcL
I. ĐẶT VẤN ĐỀBưởi là một trong những loại cây ăn quả nhiệt
đới quan trọng và có giá trị kinh tế ở nhiều nước trên thế giới. Bưởi được trồng rộng rãi ở Việt Nam, tuy nhiên mỗi vùng có một số giống bưởi khác nhau do kết quả của quá trình chọn lọc cũng như ảnh hưởng của điều kiện của khí hậu và thổ nhưỡng. Ở nước ta, từ lâu đã hình thành những vùng trồng bưởi với những giống bưởi nổi tiếng như Đoan Hùng (Phú Thọ), bưởi Đường (Hương Sơn), bưởi Phúc Trạch (Hương Khê), bưởi Thanh Trà (Huế)… Các giống bưởi này về mặt hình thái khác nhau không đáng kể
từ lá, hoa cho đến hình dạng trái. Đây chính là một trở ngại trong công tác phân loại, quản lý và chọn tạo nguồn gen.
Ngày nay, với sự phát triển của sinh học phân tử và phương pháp phân tích hệ gen sinh vật đã mở ra hướng tiếp cận mới có vai trò đầy hứa hẹn cải thiện hiệu quả được cách nhận dạng giống cây trồng. Những tiến bộ trong kỹ thuật giải trình tự ADN bằng các đoạn gen ngắn đặc biệt là đoạn gen rbcL trong hệ gen lục lạp. Đây là một trong những đoạn gen phổ biến để phân loại cây trồng (Kellogg E. and Juliano ND., 1997). Mặc dù, gen rbcL không

66
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
- Bộ mồi khuếch đại các vùng gen rbcL:
Bảng 2. Danh sách mồi khuếch đại vùng gen rbcL
được tính là chỉ thị để phân biệt mức độ loài và dưới loài nhưng các nhà nghiên cứu vân sử dụng rbcL như là một mã vạch để nhận dạng cây trồng. Ưu điểm của đoạn gen này là dễ dàng khuếch đại và giải trình tự được hầu hết các loại cây trồng. RbcL được coi là một trong những đoạn gen chuẩn để xây dựng phát sinh loài (W. John Kress and David L. Erickson, 2007). Trong phân loại, rbcL là đoạn gen ưa thích để xây dựng cây phả hệ và là đoạn gen được sử dụng nhiều nhất của hệ gen lục lạp để giải trình tự (Suyama et al., 2000; Tshering Penjor et al., 2010).
Chính vì vậy, việc lựa chọn đoạn gen rbcL để tiến hành khuếch đại và xác định trình tự nucleotit là cần thiết, từ đó làm cơ sở cho việc nhận dạng các giống bưởi, góp phần nâng cao hiệu quả quản lý, bảo tồn và chọn tạo các nguồn gen quý.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưu- 25 giống bưởi được lưu giữ tại các vườn bảo tồn
của các đơn vị như Trung tâm Tài nguyên thực vật, Trung tâm ăn quả Phủ Quỳ, Viện Nghiên cứu Rau quả, Viện Cây ăn quả miền Nam.
Bảng 1. Danh sách 12 giống chuối sử dụng trong nghiên cứu
Ghi chú: ID1: Số đăng ký cơ quan mạng lưới, ngân hàng gen cây trồng Quốc gia (GBVNML); ID2: Ký hiệu giống.
ID1 ID2 Tên giống ID1 ID2 Tên giốngGBVNML 1.177 G1 Phúc Trạch GBVNML 18.57 G16 Bưởi lông cổ còGBVNML1.180 G2 Thanh Trà GBVNML 18.8 G18 Bưởi đường Hooc MônGBVNML 1.185 G3 Phú Diễn GBVNML 18.46 G19 Bưởi hồng đườngGBVNML11.36 G4 Bưởi đỏ TEMPDA9002 G20 Bưởi Bằng LuânTEMPDA9000 G6 Bưởi đường An Phú Đông TEMPDA9003 G21 Bưởi đắngTEMPDA5173 G7 Bưởi đường Hiệp Thuận GBVNML 18.50 G22 Bưởi xiêm taTEMPDA5146 G8 Bưởi Quế Dương GBVNML18.7 G24 Bưởi đường bánh xeGBVNML18.49 G10 Bưởi ổi GBVNML 18.29 G25 Bưởi mật ongGBVNML 18.6 G12 Bưởi đường lá cam GBVNML 18.41 G26 Bưởi Năm RoiTEMPDA9001 G13 Bưởi Trụ GBVNML18.2 G27 Bưởi da xanhTEMPDA0283 G14 Bưởi Luận văn GBVNML 18.51 G28 Bưởi thanh da lángTEMPDA1975 G15 Bưởi Đoan Hùng GBVNML 18.12 G29 Bưởi xiêm vang
GBVNML 18.4 G30 Bưởi đường da láng
2.2. Phương pháp nghiên cưuADN tổng số được tách chiết và tinh sạch theo
phương pháp dùng công nghệ màng lọc silica và bằng cột lọc của QUIAGEN Dnaeasy Plan Kit.
Phản ứng PCR khuếch đại gen và rbcL được thực hiện với thành phần: Taq KodFx (1 đơn vị), hỗn hợp dNTPs (200 mM), mồi xuôi - ngược (0,5 µl mỗi mồi), DMSO (3%) và ADN tổng số (25 ng) trong đệm HF/GC 1x; thể tích phản ứng PCR là 25 µl; ở điều kiện nhiệt: biến tính ở 98oC trong 30 giây, 1 chu trình; 98oC trong 10 giây, 55 - 60oC trong 30 giây,
72oC trong 1 phút, 30 chu trình; 72oC trong 10 phút, 1 chu trình, bảo quản tại 16oC. Sản phẩm PCR được điện di trên gel agarose 1% trong đệm TBE, nhuộm bằng EtBr. Kiểm tra sản phẩm PCR bằng máy đo quang phổ nanodrop 2000. Những mâu có nồng độ sản phẩm khoảng 100 ng/µl được dùng để giải trình tự.
Chu trình và phản ứng cho giải trình tự: Mâu được khuếch đại với từng mồi, mỗi mâu lặp lại 5 lần với thành phần như sau: 100 ng sản phẩm PCR tinh sạch; 3,2 pmol mồi xuôi hoặc ngược và 8 µl hỗn hợp
Gen Mồi Trình tự 5’-3’ Tham khảo
rbcLrbcLa-F CGCGCATGGTGGATTCACAATCC W. John Kress and
David L. Erickson, 2007rbcLa-724R GTAAAATCAAGTCCACCRCG

67
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
BigDye Terminator Ready Reaction; tổng thể tích 20 µl; ở điều kiện 960C trong 1 phút, 1 chu trình; 960C trong 10 giây, 500C trong 5 giây, 600C trong 1 phút, 25 chu trình; bảo quản tại 40C. Sản phẩm được đưa vào hệ thống giải trình tự bằng máy ABI PRISM 3100 Genetic Analyzer.
Trình tự của các mâu thu được có QV>20 sẽ được xử lý, hiệu chỉnh bằng phần mềm BioEdit 4.9, sắp xếp thẳng hàng trình tự bằng công cụ ClustalW và so sánh trên ngân hàng gen bằng công cụ NCBI/BLAST được tích hợp trong phần mềm phân tích Geneious 7.11.
2.3. Thời gian và địa điểm nghiên cưuNghiên cứu được tiến hành năm từ 2014 đến
năm 2016 tại Trung tâm Tài nguyên Thực vật (An Khánh - Hoài Đức - Hà Nội).
III. KẾT QUẢ VÀ THẢO LUẬNKết quả tách chiết mâu lá của giống bưởi cho thấy
ADN có sự đồng đều về kích thước và không còn lân tạp ARN, đủ tiêu chuẩn để thực hiện các bước khuếch đại để giải trình tự tiếp theo.
Kết quả giải trình tự đoạn gen rbcL từ 25 giống bưởi được so sánh với trình tự của toàn bộ hệ gen lục lạp (BLAST với dữ liệu trình tự NCBI). Kết quả BLAST hit cho thấy trình tự 600bp nucleotide của các giống bưởi nằm trong vùng cấu trúc gen của gen rbcL. Vì vậy, các trình tự này đã được phân tích để tìm khung đọc ORF để dịch mã cho các axit amin tương ứng.
Kết quả phân tích so sánh các trình tự đoạn gen rbcL thu được từ 25 giống bưởi thu được các trình tự của đoạn gen rbcL gần như hoàn toàn tương đồng. Tuy nhiên, có sự khác biệt của 2 giống bưởi Thanh Trà (G2) và bưởi Da Xanh (G27) với cả tập đoàn bưởi nghiên cứu tại vị trí 583 (T>C) và 595 C>T (vị trí của gen rbcL, tham chiếu từ trình tự có số đăng ký NCBI NC_034290, gene ID: 32229015).
Hình 1. Khuếch đại vùng gen rbcL bằng cặp mồi rbcLa-F/724R
Hình 2. So sánh trình tự gen rbcL của các giống bưởi nghiên cứu
Hình 3. Kết quả BLAST của mâu bưởi ở vùng gen rbcL
Tiến hành BLAST các trình tự này với ngân hàng dữ liệu trình tự của NCBI và CBOLD, kết quả thu được như sau:
- Mức độ tương đồng của trình tự đoạn gen rbcL giữa các loài bưởi trong chi Citrus biến thiên từ 98,9% lên đến 100%.

68
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
- Tuy có trình tự tương đồng 100% nhưng độ bao phủ không đạt 100% nên đột biến đồng hoán (C>T) tại vị trí 595 của đoạn trình tự ở cả 2 giống bưởi Thanh Trà và Da Xanh khác biệt hoàn toàn với toàn bộ các đoạn trình tự trên ngân hàng gen. Hai trình tự đặc biệt này đã được đăng ký NCBI với số đăng ký lần lượt là KR073282 và KR073281.
Các trình tự của đoạn gen rbcL gồm 600 nucleotid và một số trình tự tham chiếu từ NCBI
tương ứng đại diện cho Citrus được sử dụng để phân tích tương quan di truyền giữa các nguồn gen trong tập đoàn nghiên cứu, bằng phương pháp lập sơ đồ hình cây NJ (Saitou N & Nei M., 1987), với outgroup là trình tự của loài Oryza sativa - đại diện nhóm cây 1 lá mầm. Các trình tự tham chiếu được chọn với tiêu chí là các trình tự có tương đồng từ gần nhất đến xa nhất trong danh sách BLAST-hit bao gồm các trình tự Citrus aurantium, Citrus maxima, Pierreodendron africanum.
Kết quả cho thấy cây phả hệ đã nhóm được các trình tự thuộc Bộ Sapindales. Trong Bộ Sapindales, các trình tự thuộc chi Citrus được nhóm thành 1 nhóm lớn, tách biệt với trình tự thuộc chi Citropsis, Triphasia. Kết quả phân nhóm dựa vào đoạn 600 nucleotid này phù hợp với phân loại từ Bộ, Họ, Chi theo khóa phân loại thông thường tham khảo từ NCBI. Kết quả này cho thấy đoạn gen rbcL có ý nghĩa trong phân loại đến mức nhận dạng Chi.
Các trình tự bưởi nghiên cứu đều nằm trong nhóm chi Citrus. Hai trình tự bưởi Thanh Trà (G2) và Da Xanh (G27) đứng tách biệt với các trình tự của bưởi Việt Nam và các nguồn gen đại diện khác (Hình 4).
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luậnKết quả nghiên cứu đa dạng trình tự đoạn gen
rbcL gồm 600 nucleotid của tập đoàn 25 nguồn gen bưởi nghiên cứu đã xác định được đột biến đồng hoán (C>T) tại vị trí 595 của đoạn trình tự ở cả 2 giống bưởi Thanh Trà (G2) và Da Xanh (G27). Các đột biến này khác biệt với toàn bộ các đoạn trình tự trên ngân hàng gen và được đăng ký trên NCBI với số đăng ký lần lượt là KR073282 và KR073281.
Phân tích cây phả hệ cho thấy, trình tự của đoạn gen rbcL đã nhóm được các trình tự chi Citrus trong cùng họ Rutaceae. Thông tin cả đoạn với 600 nucleotit đã không phân biệt chính xác các loài bưởi nghiên cứu và tham chiếu. Tuy nhiên đoạn gen rbcL đã tách biệt được giống bưởi Thanh Trà và Da Xanh của Việt Nam.
4.2. Đề nghịKết quả giải trình tự các giống chuối này có thể
được sử dụng trong nhận dạng nguồn gen và các chương trình chọn tạo giống bưởi trong tương lai.
Hình 4. Cây phân loại dựa trên đoạn gen rbcL của các giống bưởi nghiên cứu

69
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Công trình là kết quả của đề tài “Xây dựng mã vạch ADN (DNA barcode) cho các giống cây trồng đặc hữu có giá trị kinh tế của Việt Nam” thuộc chương trình Công nghệ Sinh học Nông nghiệp và Thủy sản.
TÀI LIỆU THAM KHẢOKellogg E., Juliano ND, 1997. The structure and
function of RuBisCo and their implications for systematic studies. Am J Bot, 84: 413-428.
Saitou N, Nei M, 1987. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, volume 4, issue 4, 406-425.
Suyama Y., Yoshimaru H., Tsmura Y, 2000. Molecular phylogenetic position of Japanese Abies (Pinaceae) based on chloroplast sequences. Mol Phylogenet Evol, 16(2):271-277, doi: 10.1006/mpev.2000.0795.
Tshering Penjor, Toyoaki Anai, Yukio Nagano, Ryoji Matsumoto, Masashi Yamamoto, 2010, Phylogenetic relationships of Citrus and its relatives based on rbcL gene sequences. Tree Genetics & Genomes (2010) 6: 931-939, doi: 10.1007/s11295-010-0302-1.
John Kress W. and David L. Erickson, 2007. A two - locus global DNA barcode for land plants: the coding rbcL gene complements the non-coding trnH-psbA spacer region. PLoS One, 2(6): e508, doi: 10.1371/journal.pone.0000508.
Genetic diversity of rbcL gene in Vietnam’s grapefruit germplasmsNguyen Thi Ngoc Lan, Nguyen Thi Lan Hoa,
Nguyen Thi Thanh Thuy, La Tuan NghiaAbstractGrapefruit is one of the most important tropical fruit trees and has high economic value in many countries. Research on genetic diversity in rbcL gene of 25 Vietnam’s grapefruit germplasms has identified two specific nucleotide sequences of Thanh Tra (G2) and Da Xanh (G27). This mutation was transition mutation (C>T) at 595 downstream position of sequence in both Thanh Tra and Da Xanh and it can be used for distinguishing Thanh Tra and Da Xanh varieties from others. These two sequences have been registered with NCBI codes as KR073282 and KR073281, respectively. The phylogenetic tree analysis by Neighbour Joining method based on 600 nucleotides of rbcL gene exactly grouped all surveying sequences. In addition, this analysis has clearly separated the Citrus in Rutaceae and discriminated two grapefruits of Vietnam (Thanh Tra and Da Xanh).Keywords: Grapefruit, sequencing, DNA barcode, rbcL
Ngày nhận bài: 16/9/2018Ngày phản biện: 21/9/2018
Người phản biện: TS. Trần Danh SửuNgày duyệt đăng: 15/10/2018
1 Viện Nghiên cứu Nông nghiệp công nghệ cao Đồng bằng sông Cửu Long2 Viện Lúa Đồng bằng sông Cửu Long
ĐÁNH GIÁ TÍNH CHỊU MẶN CỦA QUẦN THỂ LAI HỒI GIAO OMCS2000*4/POKKALI Ở THẾ HỆ BC3F2
Biện Anh Khoa1, Bùi Phước Tâm2, Nguyễn Thị Hồng Loan1, Nguyễn Thị Lang1
TÓM TẮTCây lúa mân cảm với điều kiện mặn và cho phản ứng khác nhau tùy thuộc vào loại môi trường cũng như bản
chất di truyền của từng cá thể. Phân tích tính chịu mặn ở cây lúa đòi hỏi phải kết hợp giữa đánh giá kiểu hình và kiểu gen. Chín mươi chín (99) dòng BC3F2 của quần thể lai hồi giao OMCS2000*4/ Pokkali được thanh lọc khả năng chịu mặn với ba nồng độ muối khác nhau (EC = 0, 6 và 12 dS/m) ở giai đoạn mạ sau hai và bốn tuần thanh lọc. Kết quả ghi nhận 21/99 dòng biểu hiện tính chịu khá tốt ở cả 6 và 12 dS/m. Các dòng này tiếp tục được kiểm tra kiểu gen liên quan tính chịu mặn (Saltol) trên NST1 với hai chỉ thị phân tử RM3412b và RM8094. Kết quả cho thấy 100% dòng chịu mặn (21 dòng) đều có alen mặn trên NST1. Qua đánh giá kiểu hình và kiểu gen, các dòng lúa xác định có tiềm năng chịu mặn tốt là BC3F2-65, BC3F2-66, BC3F2-70, BC3F2-71, BC3F2-76, BC3F2-77, BC3F2-81 và BC3F2-83.
Từ khóa: Mặn, chịu mặn, quần thể lai hồi giao, thanh lọc, giai đoạn mạ

70
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
I. ĐẶT VẤN ĐỀChiến lược tạo chọn giống chịu mặn được xem
như là cách làm kinh tế và có hiệu quả nhất để gia tăng sản lượng lúa ở vùng nhiễm mặn (Buu et al., 1995). Tính chịu mặn ở cây lúa là một tính trạng đa gen, chịu ảnh hưởng nhiều của các điều kiện môi trường. Trong công tác chọn lọc dòng lúa chịu mặn, bên cạnh kiểu hình, kiểu gen cũng đóng vai trò quan trọng. Nếu như lai hồi giao giúp chuyển một gen mục tiêu từ giống cho sang giống nhận gen trong khi vân giữ lại các đặc tính quan trọng của giống nhận thì việc sử dụng các chỉ thị phân tử cho phép giải mã di truyền của con lai ở mỗi thế hệ, làm tăng tốc độ của quá trình chọn tạo, do đó tăng hiệu quả chọn lọc gen trên một đơn vị thời gian (Hospital, 2003). Nhiều nghiên cứu tạo chọn giống lúa chịu mặn nhờ chỉ thị phân tử đã được thực hiện trước đây (Mondal và Borromeo, 2016; Huyen et al., 2013; Thomson et al., 2010).
Vì vậy, nghiên cứu này được thực hiện nhằm đánh giá tính chịu mặn của quần thể hồi giao BC3F2 (OMCS2000*4/Pokkali) trên cơ sở thanh lọc kiểu hình ở giai đoạn mạ và đánh giá kiểu gen. Qua đó chọn lọc các dòng mang gen mục tiêu và chịu mặn tốt.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưu Giống lúa Pokkali có nguồn gốc từ Ấn Độ, được
dùng làm bố và là giống chịu mặn đến 15dS/m (Zeng, 2005). Giống lúa OMCS2000 được dùng làm mẹ, đây là giống có năng suất ổn định và phẩm chất tốt, được công nhận giống quốc gia vào năm 2002 (theo Ngân hàng kiến thức cây lúa). Giống chuẩn nhiễm IR29 có nguồn gốc từ Viện lúa quốc tế (IRRI), thường được dùng làm đối chứng nhiễm (khả năng chịu thấp) trong các nghiên cứu về gen mặn trên cây lúa (Bonilla et al., 2002). Quần thể lúa hồi giao BC3F2 của tổ hợp lai OMCS2000*4/Pokkali bao gồm 99 dòng được kế thừa từ nghiên cứu của Nguyễn Thị Lang và cộng tác viên (2016).
2.2. Phương pháp nghiên cưu- Thanh lọc mặn giai đoạn mạ trong nhà lưới:
Thanh lọc mặn ở giai đoạn mạ trong dung dịch Yoshida chứa muối NaCl theo phương pháp của IRRI (1997). Các nồng độ mặn lần lượt là 0, 6 và 12 dS/m. Các thí nghiệm được thiết kế theo khối ngâu nhiên với 3 lần lặp lại. Kết quả thanh lọc được ghi nhận ở 2 tuần và 4 tuần sau khi cho cây lúa nhiễm mặn. Các chỉ tiêu theo dõi: cấp chịu mặn (cấp 1, 3, 5, 7, 9), chiều dài thân (cm), chiều dài rễ (cm), sinh khối tươi (mg) và sinh khối khô (mg). Cấp chịu mặn được phân loại theo hệ thống SES của IRRI (1997) (Bảng 1).
- Đánh giá kiểu gen liên quan đến tính chịu mặn: Ly trích DNA, đo nồng độ DNA, phản ứng PCR, điện di kiểm tra kiểu gen trên gel agarose 2,5% được thực hiện theo phương pháp của IRRI (2011).
- Phân tích thống kê: Nhập, xử lý và phân tích số liệu dựa trên phần mềm Microsoft Office Excel, STAR và R-studio.
2.3. Thời gian và địa điểm nghiên cưuThí nghiệm được thực hiện tại Viện nghiên cứu
Nông nghiệp công nghệ cao Đồng bằng sông Cửu Long từ tháng 1/2018 đến tháng 6/2018.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Đánh giá cấp chịu mặn (SES)Dựa vào thang điểm đánh giá chuẩn (SES), nếu
qui ước nhóm chịu của cây lúa từ cấp 1 đến cấp 5 và nhóm nhiễm từ trên cấp 5 đến cấp 9 thì thứ tự tính chịu theo môi trường lần lượt là 0 dS/m > 6 dS/m > 12 dS/m và theo thời gian là 2 tuần STL > 4 tuần STL. Các thứ tự này cho thấy có tính chịu mặn của cây lúa biến thiên theo nồng độ mặn và thời gian mà cây lúa tiếp xúc với môi trường mặn tương ứng.
Bảng 1. Đánh giá tính chịu mặn ở giai đoạn mạ trên cây lúa
(Nguồn: IRRI, 1997).
Cấp Mô tả Mưc chịu mặn1 Tăng trưởng bình thường, không có vết cháy lá trên lá non Chịu tốt3 Gần như bình thường, nhưng đầu lá hoặc vài lá bị cháy và cuộn lại Chịu khá5 Tăng trưởng chậm, hầu hết lá bị khô, một vài chồi bị chết Chịu trung bình7 Ngừng tăng trưởng, hầu hết lá bị khô, một vài chồi bị chết Nhiễm9 Tất cả các cây bị chết hoặc khô Rất nhiễm

71
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Hình 1. Sự khác biệt về cấp chịu của các dòng lúa ở thời điểm 4 tuần sau thanh lọc
Hình 2. So sánh tính chịu mặn của quần thể lúa với các nồng độ mặn khác nhau sau 4 tuần thanh lọcChú thích: a) EC = 0 dS/m; b) EC = 6 dS/m; c) EC = 12 dS/m.
(a) (b) (c)
Về mức chịu mặn cụ thể ở từng nồng độ mặn, 100% số dòng ở môi trường 0 dS/m đạt mức độ chịu tốt (1 ≤ SES ≤ 3). Ở môi trường EC = 6 dS/m, các dòng có khả năng mức chịu tốt (1 ≤ SES ≤ 3) chiếm tỉ lệ thấp nhất (1,0 %) và chịu trung bình (3 < SES ≤ 5)chiếm tỉ lệ 22,5%. Ở môi trường EC = 12 dS/m, mức chịu của các dòng giảm nhanh và hầu hết đều biểu hiện từ hơi nhiễm đến nhiễm (SES > 5) sau 4 tuần thanh lọc (Hình 2).
Kết quả thí nghiệm ghi nhận mức độ chịu mặn của cây lúa không những bị ảnh hưởng của môi trường mà còn phụ thuộc vào thời gian mà cây lúa tiếp xúc với điều kiện mặn (Hình 3). Ở môi trường 6 dS/m, sau 2 tuần thanh lọc, tính chịu của các dòng biểu hiện khác nhau từ chịu tốt đến nhiễm, trong đó, số lượng các dòng chịu tốt và trung bình (1 ≤ SES ≤ 5) chiếm 25%. Tuy nhiên, sau 4 tuần thanh lọc
ở nồng độ này, cấp độ chịu mặn 1< SES ≤ 3 giảm đi khoảng 4%, khoảng 3% cho cấp 3< SES ≤ 5 và 13% cho cấp 5 < SES ≤ 7 nhưng tăng khoảng 21% các dòng nhiễm ở mức 7 < SES ≤ 9.
Tương tự, cây lúa ở môi trường 12 dS/m sau 2 tuần thanh lọc, có mức độ chịu mặn thấp hơn ở môi trường 6 dS/m. Không có dòng nào chịu tốt, có 12,7% dòng chịu trung bình, còn lại là mức hơi nhiễm đến nhiễm. Tuy nhiên, so với cấp độ chịu ở 6dS/m sau 4 tuần thanh lọc, tỷ lệ dòng chịu mặn là tương đương nhau. Ở thời điểm 4 tuần sau thanh lọc ở môi trường 12 dS/m, không có dòng nào chịu, đa số các dòng đều thể hiện tính nhiễm (7 < SES ≤ 9).
Như vậy, trong số 99 dòng lúa ban đầu, có 21 dòng thể hiện khả năng chịu mặn (Bảng 2). Các dòng này chịu tốt ở 6 dS/m trong 4 tuần và 12 dS/m trong 2 tuần.

72
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Bảng 2. Khả năng chịu mặn của các dòng con lai ưu tú
Ghi chú: F: mức ý nghĩa; **: mức ý nghĩa 99%; CV: hệ số biến thiên; SD: Độ lệch chuẩn; C: Chịu tốt (1≤ SES ≤3); HC: Chịu trung bình (3 < SES ≤ 5); HN: Hơi nhiễm (5 < SES ≤ 7); N: Nhiễm (7 < SES ≤ 9).
3.2. Các đặc tính nông hoc của các dòng lúa thông qua thanh loc ở các nồng độ mặn khác nhau
Chiều cao cây (cm): ở môi trường có EC là 0, 6 và 12 dS/m, chiều cao cây trung bình theo thứ tự giảm dần là 15,7 cm, 13,9 cm và 12,4 cm. Điều này cho thấy ở nồng độ mặn càng cao, chiều cao của cây lúa càng giảm. Sự giảm của chiều cao trong điều kiện mặn phản ánh ảnh hưởng của muối đối với sự phát triển của cây, tương tự như kết quả của các nghiên cứu trước đây (Hasanuzzaman et al., 2009; Hakim et al., 2014). Các dòng có khả năng chịu mặn có xu hướng phát triển nhanh để thoát khỏi giai đoạn mân cảm để chống chịu với điều kiện bất lợi. Ngược lại, các dòng mân cảm sẽ phát triển chậm lại, sinh trưởng kém và chết (Yeo và Flower, 1984) (Hình 4).
Chiều dài rễ (cm): Ở 0 dS/m, giá trị về chiều dài rễ dao động từ 5,7 - 12,9 cm, đạt trung bình 8,8 cm. Ở môi trường 6 dS/m, chiều dài rễ trung bình giảm (9,0 cm) và dao động từ 5,6 - 14,0 cm. Ở nồng độ 12 dS/m, chiều dài rễ của các dòng lại nhỏ hơn so với môi trường 0 và 6 dS/m, giá trị thấp nhất là 3,8 cm, cao nhất là 12,9 cm và trung bình là 8,8 cm. Điều này cho thấy khi nồng độ muối của môi trường tăng lên sẽ gây ức chế sự sinh trưởng ở các dòng lúa, nhất là các dòng mân cảm hay có khả năng thích nghi kém.
Sinh khối tích lũy trong cây (mg): Khối lượng sinh khối tươi của các dòng đạt cao nhất ở nồng độ 0 dS/m, trung bình là 111,9 mg, cao nhất đạt 160,0 mg (Pokkali) và thấp nhất đạt 86,7 mg. Ở nồng độ 6 dS/m, khối lượng sinh khối tươi giảm hơn so với môi trường đối chứng, đạt trung bình 70,0 mg. Trong khi đó, ở nồng độ 12 dS/m, giá trị trung bình này giảm xuống còn 49,8 cm. Kết quả khối lượng sinh khối khô được ghi nhận tương tự như khối lượng sinh khối tươi. Khối lượng sinh khối khô có giá trị cao nhất ở 0 dS/m, kế đến là 6 dS/m và thấp
Hình 3. Tính chịu mặn của quần thể lúa ở 3 nồng độ mặn 0, 6 và 12 dS/m trong các khoảng thời gian thanh lọc khác nhau
Ghi chú: MT: Môi trường; SES: Hệ thống đánh giá tiêu chuẩn; STL: sau thanh lọc.
TT Tên dòng/giống
Cấp chịu mặn (SES)
0 dS/m
6 dS/m 12 dS/m2 tuần –STL
4 tuần –STL
2 tuần –STL
4 tuần –STL
1 BC3F2-2 C C HC HC HN2 BC3F2-15 C HC HC HC N3 BC3F2-18 C HC HC HN N4 BC3F2-19 C HC HC HN N5 BC3F2-22 C HC HN HN HN6 BC3F2-59 C HC HC HC HN7 BC3F2-64 C C HC HC HN8 BC3F2-65 C C HC C HN9 BC3F2-66 C C HC HN HN
10 BC3F2-70 C C HC HC HN11 BC3F2-71 C HC HC HC HN12 BC3F2-72 C C HC HC HN13 BC3F2-73 C HC HC HC HN14 BC3F2-76 C C HC HC HN15 BC3F2-77 C C HC HC HN16 BC3F2-78 C C HC HC HN17 BC3F2-79 C HC HN HN HN18 BC3F2-81 C HC HC HC HN19 BC3F2-83 C C C C HN20 BC3F2-100 C HC HC HC N21 BC3F2-101 C HC HN HC HN22 Pokkali C C HC HC HN23 IR29 C N N N N24 OMCS2000 C N N N N
F - ** ** ** **CV (%) - 16,68 15,55 18,82 9,46SD - 1,82 1,94 1,85 1,18

73
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
nhất ở 12 dS/m. Như vậy, qua phân tích kết quả khối lượng sinh khối tươi và khô của các dòng lúa ở 3 môi trường ghi nhận rằng khi độ mặn càng tăng thì sinh khối tích lũy trong cây lúa càng giảm. Haq và cộng tác viên (2009) cũng báo cáo rằng sinh khối tích lũy
của cây lúa giảm đi trong điều kiện mặn. Điều này xảy ra là do mặn kìm hãm sự sinh trưởng và phát triển của cây lúa bởi các stress thẩm thấu và sinh lý (Shani and Ben-Gal, 2005).
Hình 4. Sự biến động của chiều cao cây, chiều dài rễ, sinh khối tươi và khô của cây lúa qua ba môi trường mặn (0, 6 và 12 dS/m)
3.3. Đánh giá kiểu gen liên quan tính chịu mặn trên các dòng lúa
Trong thí nghiệm này, các alen mặn được đánh giá thông qua chỉ thị phân tử liên kết với gen mặn trên vùng gen chính (Saltol) (nhiễm sắc thể số 1)
được đánh dấu bởi chỉ thị RM8904 ở vị trí 44,9 cM và RM3412b ở vị trí 46,3 cM (Thomson et al., 2010). Các (21) dòng có kiểu hình chịu mặn tốt nhất và các đối chứng (bố Pokkali, mẹ OMCS2000, IR29) được chọn để tìm alen mặn với hai chỉ thị này.

74
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Chỉ thị RM3412b cho sản phẩm PCR ở vị trí 110 và 120 bp. Giống bố cũng là giống chuẩn kháng Pokkali có vị trí băng ở 110 bp, trong khi đó, giống mẹ OMCS2000 và giống chuẩn nhiễm IR29 cùng thể hiện băng hình ở vị trí 120 bp. Do đó, vị trí 110 bpcho biết cá thể mang gen mặn, ngược lại, vị trí không mang gen là 120bp. Quần thể OMCS2000*4/Pokkali có 8/21 dòng mang gen mặn đồng hợp và 13/21 dòng biểu hiện dị hợp tử. Tương tự, đối với chỉ thị RM8094, Pokkali thể hiện băng ở vị trí 150bp, đây là vị trí của gen mục tiêu. Xét trên tổ hợp OMCS2000*4/Pokkali, 8/21 dòng mang gen mặn đồng hợp, 12/21 dòng dị hợp tử và một dòng (BC3F2-100) không có băng. Như vậy, 100% dòng chịu mặn thuộc quần thể
OMCS2000*4/Pokkali đều tìm thấy được alen mặn (Saltol) trên NST1.
3.4. Sự tương đồng giữa kiểu gen và kiểu hình của tính trạng chịu mặn
Qua đánh giá sự tương đồng giữa kiểu hình và kiểu gen của 21 dòng chịu mặn, kết quả ghi nhận 100% các dòng này đều mang gen mục tiêu ở thể đồng hợp hoặc dị hợp. Trong đó, 8 dòng (BC3F2-65, BC3F2-66, BC3F2-70, BC3F2-71, BC3F2-76, BC3F2-77, BC3F2-81 và BC3F2-83) mang gen mặn đồng hợp, các dòng còn lại mang alen mặn nhưng vân còn ở thể dị hợp tử. Kết quả này có thể giải thích do sự phân ly của các dòng lúa, vì vậy, các dòng này cần tiếp tục được chọn lọc trong các thế hệ tiếp theo.
Hình 5. Kết quả đánh giá kiểu gen các dòng lúa chịu mặn trên gel agarose 2,5% với chỉ thị RM3412b và RM8094 đánh dấu gen mặn định vị trên NST1
Ghi chú: M (thang chuẩn 1 Kb+); NST1: Nhiễm sắc thể số 1.
Bảng 3. Kết quả kiểu gen quy định tính chịu mặn của các dòng lúa có kiểu hình chịu mặn trên quần thể OMCS2000*4/Pokkali
Ghi chú: ĐHT: đồng hợp tử; DHT: dị hợp tử.
TT TênKiểu gen (bp) Gen
Saltol TT TênKiểu gen (bp) Gen
SaltolRM3412b RM8094 RM3412b RM80941 BC3F2-2 110-120 150-180 DHT 13 BC3F2-73 110-120 150-180 DHT2 BC3F2-15 110-120 150-180 DHT 14 BC3F2-76 110 150 ĐHT3 BC3F2-18 110-120 150-180 DHT 15 BC3F2-77 110 150 ĐHT4 BC3F2-19 110-120 150-180 DHT 16 BC3F2-78 110-120 150-180 DHT5 BC3F2-22 110-120 150-180 DHT 17 BC3F2-79 110-120 150-180 DHT6 BC3F2-59 110-120 150-180 DHT 18 BC3F2-81 110 150 ĐHT7 BC3F2-64 110-120 150-180 DHT 19 BC3F2-83 110 150 ĐHT8 BC3F2-65 110 150 ĐHT 20 BC3F2-100 110-120 - DHT9 BC3F2-66 110 150 ĐHT 21 BC3F2-101 110-120 150-180 DHT
10 BC3F2-70 110 150 ĐHT 22 Pokkali 110 150 ĐHT11 BC3F2-71 110 150 ĐHT 23 IR29 120 180 không12 BC3F2-72 110-120 150-180 DHT 24 OMCS2000 120 180 không

75
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
IV. KẾT LUẬNCác dòng của quần thể OMCS2000*4/Pokkali thể
hiện nhiều mức độ mân cảm với từng môi trường mặn khác nhau. Môi trường có độ mặn càng cao, thời gian tiếp xúc của cây lúa với môi trường càng dài, mức chống chịu và sự sinh trưởng của cây lúa càng giảm. Quần thể OMCS2000*4/Pokkali chịu tốt ở 6 dS/m trong 4 tuần và ở 12 dS/m trong 2 tuần sau khi cây lúa tiếp xúc với điều kiện mặn. Kết quả xác định 8 dòng thể hiện tính chịu khá tốt và mang gen mặn là BC3F2-65, BC3F2-66, BC3F2-70, BC3F2-71,BC3F2-76, BC3F2-77, BC3F2-81 và BC3F2-83. Các dòng này cần được tiếp tục chọn lọc và phát triển cho các vùng bị ảnh hưởng bởi mặn.
TÀI LIỆU THAM KHẢONguyễn Thị Lang, Russell R, Ismail AM, Nguyễn
Văn Hiếu, Bùi Phước Tâm, Phạm Thị Thu Hà, Nguyễn Hoàng Thái Bình, Phạm Công Trư, Trần Thị Nhiên, Nguyễn Trong Phước, Bùi Chí Bửu, Wassmann R, 2016. Chọn giống lúa ngập và mặn phục vụ Đồng bằng sông Cửu Long. Dự án CLUES, Cần Thơ, 2012-2016.
Bonilla P, Dvorak J, Mackill D, Deal K, GregorioG, 2002. RFLP and SSLP mapping of salinity tolerance genes in chromosome 1 of rice (Oryza sativa L.) using recombinant inbred lines. Philipp Agricultural Scientist, 85: 68-76.
Buu BC, Lang NT, Tao PB, Bay ND, 1995. Rice breeding research strategy in the Mekong Delta. Proc. Of the Int. Rice Res. Conf. “Fragile Lives in Fragile Ecosystems, IRRI”, page: 739-755.
Hakim MA, Juraimi AS, Hanafi MM, Ismail MR, Rafii MY, Islam MM, Selamat A, 2014. The effect of salinity on growth, ion accumulation and yield of rice varieties. The Journal of Animal & Plant Sciences, 24(3): page: 874-885.
Haq T, Akhtar J, Nawaz S, Ahmad R, 2009. Morpho- physiological response of rice (Oryza sativa L.) varieties
to salinity stress. Pak. J.Bot., 41 (6): 2943-2956.Hasanuzzaman M, Fujita M, Islam MN, Ahamed KU,
Nahar K, 2009. Performance of four irrigated rice varieties under different levels of salinity stress. Int J Integ Biol., 6: 85 - 90.
Hospital F, 2003. Marker-assisted breeding. In: Newbury HJ, editor. Plant molecular breeding. Oxford: Blackwell; pp. 30-59.
Huyen LTN, Cuc LM, Ham LH, Khanh TD, 2013. Introgression the Saltol QTL into Q5BD, the elite variety of Vietnam using marker assisted selection (MAS). Am J Biosci., 1: 80-84.
IRRI, 1997. Screening rice for salinity tolerance. International Rice Research Institute. Discussion paper series. No.22.
IRRI, 2011. Laboratory Handbook of Molecular Marker Application for Rice Breeding. International Rice Research Institute. Rice Breeding Course 19 August 2011.
Mondal S, Borromeo TH, 2016. Screening of salinity tolerance of rice at early seedling stage. J. Biosci. Agric. Res., 10 (01): 843-847.
Shani U, Ben-Gal A, 2005. Long-term response of grapevines to salinity: Osmotic effects and ion toxicity. Am. J. Enol. Vitic., 56: 148-154.
Thomson MJ, de Ocampo M, Egdane J, Rahman MA, Sajise AG, Adorada DL, Tumimbang-Raiz E, Blumwald E, Seraj ZI, Singh RK, Gregorio GB, Ismail AM, 2010. Characterizing the Saltol quantitative trait locus for salinity tolerance in rice. Rice., 3: 148-160.
Yeo AR, Flowers TJ, 1984. Mechanisms of salinity resistance in rice and their role as physiological criteria in plant breeding. In: Salinity Tolerance in Plants: Strategies for Crop Improvement, Wiley Interscience, New York, pp.151-170.
Zeng LH, 2005. Exploration of relationships between physiological parameters and growth performance of rice (Oryza sativa L.) seedlings under salinity stress usingmultivariate analysis. Plant Soil, 268: 51-59.
Evaluation of salinity tolerance of rice backcross population (OMCS2000*4/Pokkali) in BC3F2 generation
Bien Anh Khoa, Bui Phuoc Tam, Nguyen Thi Hong Loan, Nguyen Thi Lang
AbstractRice plants are susceptible to salinity conditions and give different responses depending on the type of environment as well as the genetic nature of the individual. Analysis of salinity tolerance in rice requires a combination of phenotypic and genotypic assessment. Ninety nine (99) BC3F2 lines of OMCS2000*4/Pokkali hybrid population were screened for salinity tolerance at three different concentrations (EC = 0, 6 and 12 dS/m) after two and four weeks at seedling stage. Of these, 21 lines were tolerant to both 6 and 12 dS/m. These lines were further tested for Saltol gene expression on the chromosome 1 with two tightly linked markers, RM3412b and RM8094. The results showed that 100% of identified lines (21 lines) carrying Saltol allele. With the phenotypic and genotypic analysis, the study identified 8 of promising salinity tolerant lines that were BC3F2-65, BC3F2-66, BC3F2-70, BC3F2-71, BC3F2-76, BC3F2-77, BC3F2-81, and BC3F2-83.Keywords: Salinity, tolerance, backcross population, screen, seedling stage
Ngày nhận bài: 12/7/2018 Ngày phản biện: 19/7/2018
Người phản biện: TS. Dương Hoàng SơnNgày duyệt đăng: 15/10/2018

76
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
PHÂN BIỆT MỘT SỐ CẶP GIỐNG LÚA GIỐNG NHAU BẰNG CHỈ THỊ PHÂN TỬ ĐỂ HỖ TRỢ KHẢO NGHIỆM DUS
Trần Long2, Lưu Minh Cúc1, Nguyễn Quang Sáng2, Phạm Xuân Hội1
TÓM TẮTĐặc trưng ADN có thể trở thành chỉ tiêu quan trọng hỗ trợ trong khảo nghiệm DUS vì nó giúp đánh giá chính
xác cho việc giám định và phân biệt một giống cây trồng mới. Nghiên cứu tiến hành phân tích một số cặp giống lúa có đặc điểm hình thái giống nhau bằng phương pháp sử dụng chỉ thị phân tử ADN. Phân tích bằng phần mềm POWER MARKER cho thấy với tần suất 4,9 - 7,2 alen, thì độ tương đồng chung của 62 tính trạng hình thái và các chỉ thị SSR là từ 0,36 - 0,64, trong khi hệ số đa dạng di truyền sẽ từ 0,33 - 0,59. Sử dụng bộ 30 chỉ thị SSR để phân tích 19 giống lúa giống nhau theo cặp trong khảo nghiệm DUS, kết quả cho thấy, cặp giống Nếp Triều Tiên cũ - Nếp Triều Tiên mới và cặp QX22 - X33 có hệ số tương đồng di truyền là 1.00. Các cặp có hệ số tương đồng di truyền xa nhau nhất là DB15 - NH3 (0,63), Thịnh dụ 8 - Thịnh dụ 4 (0,87). Còn lại gồm có cặp giống Nếp NĐ 1 - NĐ 2; cặp giống Nhiệt đới 1 - CL10; ba giống HT1, HT6, HT9, cặp giống TBR36 - TQ08, cặp giống Nếp Lang Liêu - Nếp GRQ10 có sự khác biệt không rõ ràng, chưa đủ để công nhận đó là các giống khác nhau theo cặp.
Từ khóa: Chỉ thị phân tử, giống lúa, khảo nghiệm DUS
1 Viện Di truyền Nông nghiệp; 2 Trường Đại học Khoa học Tự nhiên
I. ĐẶT VẤN ĐỀTheo tiêu chuẩn của Hiệp hội Quốc tế về Bảo
hộ giống cây trồng mới (UPOV), các cây trồng mới được chọn tạo phải qua được khâu kiểm nghiệm theo tiêu chí DUS mới được đưa vào sản xuất. Khảo nghiệm DUS bao gồm khảo nghiệm tính Khác biệt - Distinctness, tính Đồng nhất - Uniformity, và tính Ổn định - Stability. Trong 15 năm gần đây, mỗi năm Trung tâm Khảo kiểm nghiệm Giống, Sản phẩm Cây trồng Quốc gia (TTKKNG) tiến hành khảo nghiệm khoảng 100 giống lúa. Khi khảo nghiệm, TTKKNG thường gặp phải 5 - 10 trường hợp mỗi năm các giống đưa ra khảo nghiệm có các tính trạng hình thái, đôi khi cả tính trạng hóa sinh giống nhau. Trong một số trường hợp, giống của các tác giả khác nhau nghiên cứu ra hoặc được nhập nội vào Việt Nam, tên gọi khác nhau, nhưng về mặt hình thái lại hoàn toàn giống nhau gây ra những tranh cãi khó giải quyết. Câu hỏi đặt ra là thực chất chúng thuộc cùng một giống hay chúng thuộc các giống khác nhau? Các tiêu chí khảo nghiệm DUS truyền thống dựa trên cơ sở 62 - 65 tính trạng hình thái và hóa sinh đôi khi chưa đủ để phân biệt các giống, làm dấy lên sự tranh cãi về bản quyền giống. Để giải quyết vấn đề đó, nghiên cứu này đã tiến hành phân tích một số cặp giống lúa giống nhau bằng phương pháp sử dụng chỉ thị phân tử ADN. Đặc trưng ADN có thể trở thành chỉ tiêu quan trọng hỗ trợ trong khảo nghiệm DUS vì nó giúp đánh giá chính xác cho việc giám định và phân biệt một giống cây trồng mới. Kết quả “vân tay ADN - DNA fingerprinting” nhận được thông qua phân tích PCR cùng với thông tin về phả hệ, kết hợp đánh giá các tính trạng đa gen quan trọng có ý nghĩa nông học được thừa nhận như là cơ sở khoa học và thực tiễn khách quan nhất để mô tả và đăng ký giống. Các nhà khoa học Trung Quốc đã đi tiên
phong trong lĩnh vực sử dụng chỉ thị phân tử SSR cho đánh giá và khảo nghiệm lúa lai. Họ đã chọn được 2 bộ chỉ thị chuẩn (1 bộ gồm 10 chỉ thị, bộ kia gồm 12 chỉ thị) để sử dụng (Xiao et al., 2006). Tiếp theo, các nhà khoa học ở Viện Nghiên cứu Lúa Quốc gia (Hàng Châu) phối hợp với Trường Đại học Nông nghiệp Hoa Nam và Viện Nghiên cứu Lúa Quốc tế (IRRI) đã sử dụng bộ chỉ thị chuẩn gồm 24 chỉ thị SSR (2 chỉ thị SSR trên mỗi NST lúa) để đánh giá 63 dòng bố mẹ lúa lai cùng các con lai (Ying et al., 2007). Theo các tác giả này, chỉ cần sử dụng bộ gồm 12 chỉ thị chính để đánh giá. Mark và cộng tác viên đã phân biệt các giống lúa mang tên Basmati trong thị trường Anh bằng 9 chỉ thị (Mark et al., 2004).
II.VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưu- 19 giống lúa giống nhau theo cặp trong khảo
nghiệm DUS năm 2010 - 2012: Cặp số 1 (1.Nếp triều tiên cũ, 2.Nếp triều tiên mới); Cặp số 2 (3.Nếp NĐ1, 4.Nếp NĐ2); Cặp số 3 (5.Nhiệt đới 1, 6.CL10); Bộ ba (7.HT1, 8.HT6, 9.HT9); Cặp số 4 (10.TBR36; 11.TQ08); Cặp số 5 (12.Thịnh dụ 8, 13.Thịnh dụ 4); Cặp số 6 (14.Nếp Lang Liêu, 15.Nếp GRQ10); Cặp số 7 (16.ĐB15, 17.NH3); Cặp số 8 (18.QX2, 19.X33).
- Các hóa chất sinh học phân tử và vật tư thí nghiệm.- Bộ 30 chỉ thị SSR: RM11, RM21, RM163,
RM481, RM3412, RM1, RM5, RM6, RM17, RM19, RM25, RM206, RM215, RM223, RM333, RM341, RM3252, RM3843, RM6318, RM3486, RM5758, RM10825, RM17954, RM26063, R4M13, MADS3, MADS8, SO1160, S11033, EST20.
2.2. Phương pháp nghiên cưu- Thí nghiệm được tiến hành theo quy phạm

77
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
khảo nghiệm tính khác biệt, tính đồng nhất và tính ổn định của giống lúa 10TCN 554 2002.
- Phương pháp đánh giá 62 tính trạng: Các quan sát đánh giá được tiến hành trên 20 cây hoặc 20 bộ phận của cây trong mỗi lần nhắc. Số liệu được xử lý trên phần mềm khảo nghiệm DUS-T (Trung tâm Khảo nghiệm Giống). Mỗi một tính trạng có phương pháp cụ thể và được theo dõi ở những giai đoạn nhất định mà tính trạng đó biểu hiện rõ nhất.
- DNA tổng số được tách chiết tinh sạch bằng phương pháp CTAB cải tiến (Zheng et al., 1995). Phản ứng PCR được thực hiện với tổng thể tích phản ứng 15 μl, trong đó gồm 2 μl DNA khuôn mâu (10 - 20 ng/ μl); 1,5 μl PCR buffer; 1 μl dNTPs (2 mM); 0,5 μlmồi xuôi và ngược; 8,5 μl Q-water và 1 μl Taq polymerase (Fermentas, California, USA). Hỗn hợp phản ứng PCR được chạy trên máy Eppendoft Thermocycler (Mastercycler Pro S, Germany) loại 96 giếng. Chương trình PCR: 940C trong 5 phút, tiếp theo là 35 chu kỳ với các bước: biến tính 940C trong 1 phút, gắn nối ở 550C C trong 1 phút, kéo dài ở 720C trong 1 phút, và bước hoàn thiện ở 720C trong 5 phút. Sản phẩm PCR được trộn với hóa chất nhuộm bromophenol blue và phân tích trên gel 8% polyacrylamide ở 100 volts (Dual Trip-Wide Mini-Vertical System, C.B.S. Scientific, CA, USA), nhuộm SYBR-Safe staining (Invitrogen) để phát hiện băng DNA trên gel.
- Số liệu kiểu gen được phân tích trên phần mềm NTSYS 2.1. Sơ đồ hình cây được thiết lập theo phương pháp UPGMA. Chương trình Excel và Power Marker 3.25 tính toán các chỉ số tần suất alen, số alen, độ tương đồng di truyền, hệ số đa dạng di truyền.
- Độ tương đồng di truyền theo công thức của Nei và Li (1972):
Sij = Sij= 2a/(2a + b + c)Trong đó, Sij: độ tương đồng giữa hai mẫu i và j; aij
là số băng có mặt ở cả hai mẫu i và j; bij là số băng có mặt ở mẫu i nhưng không có ở mẫu j ; cij là số băng có mặt ở mẫu j nhưng không có ở mẫu i.
- Hệ số đa dạng di truyền hay giá trị thông tin đa hình giữa các marker (PIC, Polymorphism Information Content) được tính toán theo Saal and Wricke (1999):
PIC = 1 – SPij2
Trong đó, Pij là tần suất của alen thứ i đối với marker thứ j.
2.3. Thời gian và địa điểm nghiên cưuThí nghiệm được tiến hành trong năm 2016 -
2017 tại Trung tâm Khảo kiểm nghiệm Giống và Viện Di truyền Nông nghiệp.
III. KẾT QUẢ VÀ THẢO LUẬNKết hợp cùng với Trung tâm Khảo kiểm nghiệm
Giống, tiến hành đánh giá DUS của các giống trong các năm 2010, 2011, 2012. Mục đích của công việc là tìm kiếm sự liên quan giữa các tính trạng hình thái và các chỉ tiêu ADN. Sau khi khảo nghiệm, sẽ tiến hành so sánh mức độ tương đồng giữa khảo nghiệm hình thái và đánh giá chỉ tiêu ADN bằng các chỉ thị của bộ chỉ thị tham chiếu SSR sử dụng. Trên cơ sở đó, kết luận về sự liên quan của hai phương pháp với nhau. Số liệu phân tích 62 tính trạng hình thái DUS và phân tích ADN với các chỉ thị được chuyển đổi thành định dạng file dữ liệu đầu vào của chương trình Excel. Số liệu đầu ra được thể hiện trong bảng 1, bảng 2.
Bảng 1. Phân tích 30 chỉ thị của bộ chỉ thị SSR trong chương trình PowerMarker
Chỉ thịTần suất alen
Số alen
Độ tương đồng di truyền
Hệ số đa dạng
di truyềnRM11 0,5405 5 0,6355 0,5906RM21 0,2162 6 0,8225 0,7974RM163 0,2703 5 0,7772 0,7409RM481 0,3243 7 0,7889 0,7596RM3412 0,3243 7 0,8167 0,7952RM1 0,5676 5 0,6048 0,5550RM5 0,6216 4 0,5581 0,5135RM6 0,3784 4 0,7071 0,6539RM17 0,5135 4 0,6574 0,6117RM19 0,6757 6 0,5113 0,4814RM206 0,3514 7 0,7421 0,7023RM333 0,4865 6 0,6939 0,6580RM3252 0,3243 5 0,7626 0,7242RM3843 0,3784 5 0,6998 0,6461RM7097 0,6486 3 0,5142 0,4578R4M13 0,5405 5 0,6384 0,5951SO1160 0,4865 3 0,5698 0,4757S11033 0,6757 5 0,5113 0,4814RM223 0,4595 4 0,6311 0,5603RM341 0,5946 5 0,5873 0,5442RM3468 0,7568 2 0,3682 0,3004RM10825 0,3784 4 0,7144 0,6627RM17954 0,5405 7 0,6472 0,6103RM26063 0,3784 4 0,7275 0,6795MADS3 0,4865 4 0,6662 0,6151MADS8 0,8649 2 0,2337 0,2064EST20 0,4595 3 0,6297 0,5536RM215 0,5135 4 0,6443 0,5915RM25 0,7838 5 0,3696 0,3498RM5758 0,6757 4 0,5026 0,4644Trung bình 0,4882 5,6563 0,6374 0,5931

78
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Đối với 62 tính trạng hình thái, số liệu khảo nghiệm DUS được ký hiệu hóa theo nguyên tắc: sự khác biệt đối với từng tính trạng được coi như là các
alen khác nhau trong cùng một locut. Sau khi phân tích trong chương trình PowerMarker, các kết quả được liệt kê ở bảng 2.
Ở cả bảng 1 và 2 cho thấy với tần suất 4,9 - 7,2 alen, thì độ tương đồng chung của các tính trạng hình thái và các chỉ thị SSR là từ 0,36 - 0,64, trong khi hệ số đa dạng di truyền sẽ thay đổi từ 0,33 - 0,59. Dựa trên kết quả phân tích tìm sự liên quan trong phân nhóm các giống giữa các tính trạng hình thái trong khảo nghiệm DUS, và các chỉ thị ADN trong bộ chỉ thị tham chiếu cho thấy, không có sự liên quan mật thiết khi phân tích cả tổng thể chung. Như
vậy, biểu hiện kiểu hình của 62 tính trạng và kết quả phân tích ADN đối với các locut của bộ chỉ thị tham chiếu là độc lập nhau. Điều này cho thấy sự cần thiết khi phân tích chỉ tiêu ADN như là một trong các yếu tố hỗ trợ để đánh giá, phân biệt các giống lúa trong khảo nghiệm DUS, đặc biệt trong những trường hợp sự khác biệt về hình thái không rõ ràng như ở trong nghiên cứu này.
Bảng 2. Phân tích 62 tính trạng hình thái trong chương trình PowerMarker
Tính trạng
Tần suất alen
Số alen
Độ tương đồng di truyền
Hệ số đa dạng di truyền
Tính trạng
Tần suất alen
Số alen
Độ tương đồng di truyền
Hệ số đa dạng di truyền
1 1,0000 1 0,0000 0,0000 34 0,6486 3 0,4733 0,38412 1,0000 1 0,0000 0,0000 35 0,7297 2 0,3944 0,31673 0,5135 5 0,6355 0,5782 36 0,8378 5 0,2907 0,28044 1,0000 1 0,0000 0,0000 37 0,2973 6 0,7801 0,74576 1,0000 1 0,0000 0,0000 38 0,7297 2 0,3944 0,31678 0,2973 8 0,8108 0,7868 39 0,7297 3 0,4076 0,34379 1,0000 1 0,0000 0,0000 40 0,7297 5 0,4412 0,4129
10 1,0000 1 0,0000 0,0000 41 1,0000 1 0,0000 0,000011 1,0000 1 0,0000 0,0000 42 0,5946 2 0,4821 0,365912 1,0000 1 0,0000 0,0000 43 0,7297 3 0,4076 0,343713 1,0000 1 0,0000 0,0000 44 0,8378 4 0,2849 0,266414 1,0000 1 0,0000 0,0000 45 0,3784 4 0,7071 0,653915 1,0000 1 0,0000 0,0000 46 0,9459 3 0,1037 0,101116 0,6216 5 0,5654 0,5268 48 1,0000 1 0,0000 0,000017 0,9730 2 0,0526 0,0512 49 0,4054 4 0,6998 0,645918 0,9730 2 0,0526 0,0512 50 0,5946 4 0,5683 0,510819 0,2162 10 0,8619 0,8468 52 0,6757 3 0,4777 0,416921 0,4054 5 0,7392 0,7012 53 0,4865 4 0,6457 0,584722 0,4865 5 0,6560 0,6016 54 0,5135 4 0,6209 0,555223 0,8919 2 0,1928 0,1742 55 0,6486 3 0,4733 0,384124 0,7568 4 0,3974 0,3629 56 0,3784 4 0,7188 0,668025 0,8919 3 0,1987 0,1894 57 0,8378 3 0,2849 0,266326 1,0000 1 0,0000 0,0000 58 0,7027 3 0,4617 0,416727 0,7568 3 0,3886 0,3441 59 0,2973 7 0,8093 0,783528 0,4324 5 0,6896 0,6357 60 0,3514 5 0,7480 0,706429 1,0000 1 0,0000 0,0000 61 0,5946 4 0,5741 0,521431 1,0000 1 0,0000 0,0000 62 0,7568 2 0,3682 0,300432 0,4595 4 0,6384 0,5682 Trung
bình 0,7202 3,7018 0,3626 0,331133 0,8919 4 0,2001 0,1932

79
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Hình 1. Tính trạng “Trạng thái lá đòng” của các cặp giống: Nếp GRQ10 và Nếp Lang Liêu; Nếp Triều Tiên cũ và Nếp Triều Tiên mới
Hình 2. Tính trạng “hạt gạo lật” và “hạt thóc” của 3 giống HT1, HT6, HT9;2 giống Nếp Lang Liêu, Nếp GQR10 giống nhau trong khảo nghiệm DUS
Tổng số 19 giống lúa giống nhau theo cặp trong khảo nghiệm DUS năm 2010 - 2012 đã nêu trong phần vật liệu được song song đánh giá các tính trạng hình thái theo quy định của khảo nghiệm DUS và đánh giá phân tích DNA. Hình 1 và 2 là hình ảnh đánh giá tính trạng “trạng thái lá đòng” (Hình 1)
của giống Nếp GRQ10 và Nếp Lang Liêu; Nếp Triều Tiên cũ và Nếp Triều Tiên mới; “hạt gạo lật và hạt thóc” (hình 2) của 3 giống HT1, HT6, HT9, 2 giống Nếp Lang Liêu, Nếp GQR10 giống nhau trong khảo nghiệm DUS. Qua đó có thể thấy, các cặp giống này giống hệt nhau ở các tính trạng hình thái.
Các cặp giống lúa giống nhau theo 62 tính trạng DUS hình thái đã được liệt kê trong phần vật liệu được thu mâu lá lúa và tách chiết DNA tổng số. Tiến hành phản ứng PCR để với điện di trên gel polyacrylamide và so sánh với kích thước alen chuẩn của bộ giống tham chiếu, thang chuẩn 25bp. Trên hình 3 là hình ảnh điện di trên gel polyacrylamide 8% của một số chỉ thị a.MADS8, b.RM223, c.RM19, d.RM21 cho thấy một số cặp giống cho các băng giống nhau như cặp giống Nếp Triều Tiên cũ và Nếp Triều Tiên mới tương ứng với giếng số thứ tự 1 và 2 trên hình.
Ba giống HT1, HT6, HT9 (giếng số thứ tự 7, 8, 9 trên hình) cho băng giống hệt nhau trên các chỉ thị phân tử được dùng để khảo sát. Chỉ có cặp giống
Nếp NĐ1, Nếp NĐ2 và cặp giống ĐB15, NH3 cho alen khác biệt với chỉ thị RM223. Trên hình 4 cho thấy, cặp giống ĐB15, NH3 cho alen khác biệt với chỉ thị RM6318.
Sau khi điện di sản phẩm PCR trên gel polyacrylamide, tiến hành ghi nhận số liệu, đọc kích thước, xử lý số liệu bằng phần mềm NTSYS, kết quả được thể hiện ở bảng 3 và hình 5.
Qua bảng 3 và hình 5 cho thấy các cặp giống giống nhau có quan hệ di truyền rất gần nhau. Cụ thể cặp giống TTC (Triều Tiên cũ), TTM (Triều Tiên mới) và cặp QX22, X33 có hệ số tương đồng di truyền là 1,00; các alen giống hệt nhau đối với cả 30 chỉ thị SSR được sử dụng. Hai cặp giống có quan hệ di truyền xa nhau nhất là DB15 và NH3 có hệ số

80
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
tương đồng di truyền là 0,63 tức là đã khá xa nhau về mặt phân tích ADN. Tuy nhiên, khi nhìn trên ảnh gel điện di đối với bộ chỉ thị, rất nhiều locut cho kết quả hai băng ADN, nhưng cặp giống này vân cho sự khác biệt hoàn toàn với 4/30 chỉ thị khảo sát, do
đó có thể thấy rằng DB15 và NH3 là hai giống có sự khác biệt rõ ràng. Tiếp tục phân tích cặp giống Thịnh dụ 8 với Thịnh dụ 4 cho thấy, cặp giống này có mức độ tương đồng 0,857 thể hiện bằng sự khác biệt hoàn toàn của 3/30 chỉ thị khảo sát.
Hình 3. Hình ảnh điện di trên gel polyacrylamide của 19 giống với các chỉ thịa. MADS8, b. RM223, c. RM19, d. RM21.
Giếng số 1: Nếp Triều Tiên cũ, 2: Nếp Triều Tiên mới, 3: Nếp NĐ1, 4: Nếp NĐ2, 5: Nhiệt đới1, 6: CL10, 7: HT1, 8: HT6, 9: HT9, 10: TBR36; 11: TQ08, 12: Thịnh dụ 8, 13: Thịnh dụ 4, 14: Nếp Lang Liêu, 15: Nếp GRQ10, 16: ĐB15, 17: NH3, 18: QX2, 19: X33.
Hình 5. Sơ đồ hình cây thể hiện quan hệ 19 giống lúa khi phân tích bằng 30 chỉ thị SSR, xử lý trong chương trình NTSYS
Hình 4. Hình ảnh điện di trên gel polyacrylamide của 19 giống với chỉ thị RM63181: Nếp Triều Tiên cũ, 2: Nếp Triều Tiên mới, 3: Nếp NĐ1, 4: Nếp NĐ2, 5: Nhiệt đới 1, 6: CL10, 7: HT1,
8: HT6, 9: HT9, 10: TBR36; 11: TQ08, 12: Thịnh dụ 8, 13: Thịnh dụ 4, 14: Nếp Lang Liêu, 15: Nếp GRQ10, 16: ĐB15, 17: NH3, 18: QX2, 19: X33.
a
c
b
d
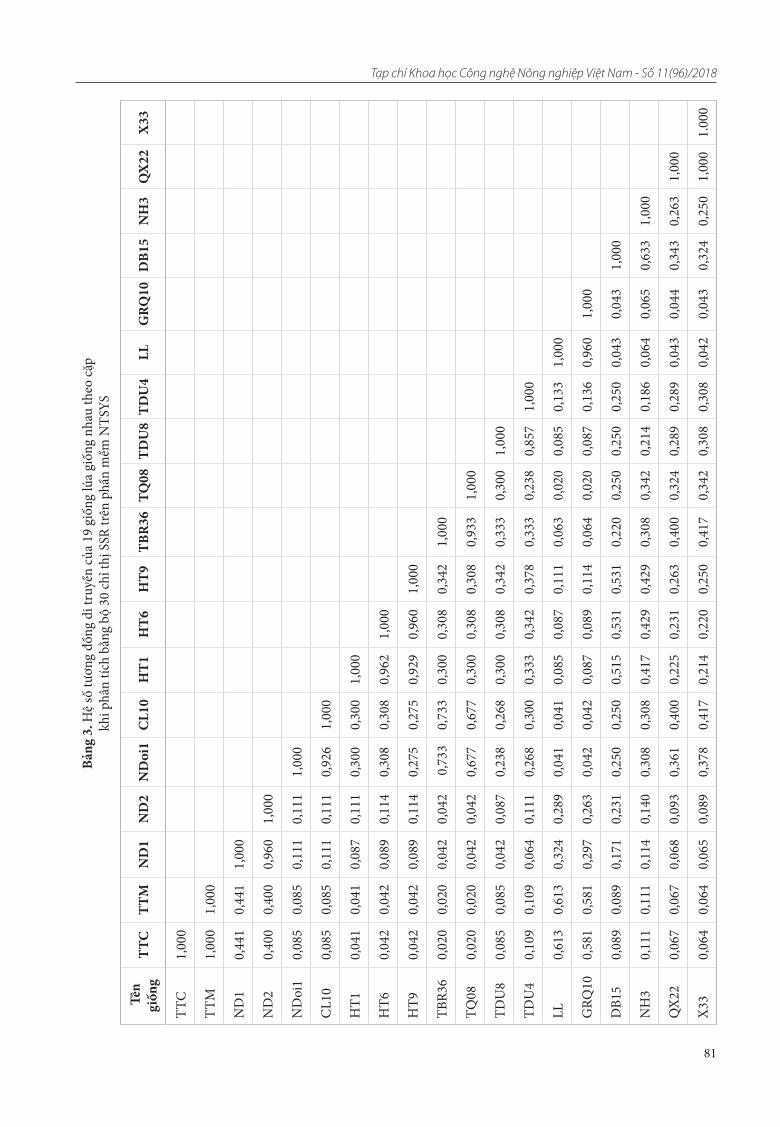
81
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018Bả
ng 3
. Hệ
số tư
ơng
đồng
di t
ruyề
n củ
a 19
giố
ng lú
a gi
ống
nhau
theo
cặp
kh
i phâ
n tíc
h bằ
ng b
ộ 30
chỉ t
hị S
SR tr
ên p
hần
mềm
NTS
YS
Tên
gi
ống
TTC
TTM
ND
1N
D2
ND
oi1
CL1
0H
T1H
T6H
T9TB
R36
TQ08
TDU
8TD
U4
LLG
RQ10
DB1
5N
H3
QX
22X
33
TTC
1,00
0
TTM
1,00
01,
000
ND
10,
441
0,44
11,
000
ND
20,
400
0,40
00,
960
1,00
0
ND
oi1
0,08
50,
085
0,11
10,
111
1,00
0
CL1
00,
085
0,08
50,
111
0,11
10,
926
1,00
0
HT1
0,04
10,
041
0,08
70,
111
0,30
00,
300
1,00
0
HT6
0,04
20,
042
0,08
90,
114
0,30
80,
308
0,96
21,
000
HT9
0,04
20,
042
0,08
90,
114
0,27
50,
275
0,92
90,
960
1,00
0
TBR3
60,
020
0,02
00,
042
0,04
2 0
,733
0,73
30,
300
0,30
80,
342
1,00
0
TQ08
0,02
00,
020
0,04
20,
042
0,67
70,
677
0,30
00,
308
0,30
80,
933
1,00
0
TDU
80,
085
0,08
50,
042
0,08
70,
238
0,26
80,
300
0,30
80,
342
0,33
30,
300
1,00
0
TDU
40,
109
0,10
90,
064
0,11
10,
268
0,30
00,
333
0,34
20,
378
0,33
30,
238
0,85
71,
000
LL0,
613
0,61
30,
324
0,28
90,
041
0,04
10,
085
0,08
70,
111
0,06
30,
020
0,08
50,
133
1,00
0
GRQ
100,
581
0,58
10,
297
0,26
30,
042
0,04
20,
087
0,08
90,
114
0,06
40,
020
0,08
70,
136
0,96
01,
000
DB1
50,
089
0,08
90,
171
0,23
10,
250
0,25
00,
515
0,53
10,
531
0,22
00,
250
0,25
00,
250
0,04
30,
043
1,00
0
NH
30,
111
0,11
10,
114
0,14
00,
308
0,30
80,
417
0,42
90,
429
0,30
80,
342
0,21
40,
186
0,06
40,
065
0,63
31,
000
QX
220,
067
0,06
70,
068
0,09
30,
361
0,40
00,
225
0,23
10,
263
0,40
00,
324
0,28
90,
289
0,04
30,
044
0,34
30,
263
1,00
0
X33
0,06
40,
064
0,06
50,
089
0,37
80,
417
0,21
40,
220
0,25
00,
417
0,34
20,
308
0,30
80,
042
0,04
30,
324
0,25
01,
000
1.00
0

82
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Như vậy, các cặp giống ĐB15-NH13, TD8-TD4 này có sự khác biệt rõ ràng đã được chứng minh. Cả hai cặp giống này đã được Trung tâm Khảo nghiệm Giống đánh giá DUS trong vụ thứ hai và đã tìm thấy các sự khác biệt về thời gian sinh trưởng và chất lượng gạo rõ ràng đủ để công nhận giống. Ngoài ra, các cặp giống còn lại có sự tương đồng di truyền từ 92,6 - 100% đối với 30 chỉ thị SSR khảo sát; nhưng khi phân tích từng alen trên mỗi locut, một số locut cho 2 băng ADN. Số locut có sự khác biệt hoàn toàn là 0 - 2 trên tổng số 30 chỉ thị khảo sát. Như vậy, các cặp giống còn lại gồm có cặp giống Nếp NĐ1 - NĐ 2; cặp giống Nhiệt đới 1 - CL10; ba giống HT1, HT6, HT9, cặp giống TBR36 - TQ08, cặp giống Nếp Lang Liêu - Nếp GRQ10 có sự khác biệt không rõ ràng, chưa đủ để công nhận đó là các giống khác nhau theo cặp.
IV. KẾT LUẬN
4.1. Kết luận Phân tích bằng phần mềm POWERMARKER
cho thấy với tần suất 4,9 - 7,2 alen, thì độ tương đồng chung của 62 tính trạng hình thái và các chỉ thị SSR là từ 0,36 - 0,64, trong khi hệ số đa dạng di truyền sẽ thay đổi từ 0,33 - 0,59.
Đã sử dụng bộ 30 chỉ thị SSR để phân tích 19 giống lúa giống nhau theo cặp trong khảo nghiệm DUS: Cặp giống TTC (Nếp Triều Tiên cũ), TTM (Nếp Triều Tiên mới) và cặp QX22, X33 có hệ số tương đồng di truyền là 1.00.
Các cặp hệ số tương đồng di truyền xa nhau nhất là DB15 và NH3 (0,63), Thịnh dụ 8 - Thịnh dụ 4 (0,87) có sự khác biệt rõ ràng trong chỉ tiêu DNA.
Các cặp giống còn lại gồm cặp giống Nếp NĐ1 - NĐ2; cặp giống Nhiệt đới 1 - CL10; ba giống HT1,
HT6, HT9, cặp giống TBR36 - TQ08, cặp giống Nếp Lang Liêu - Nếp GRQ10 có sự khác biệt không rõ ràng (0,926 - 0,962) chưa đủ để công nhận đó là các giống khác nhau theo cặp.
4.2. Đề nghịCần tiếp tục sử dụng thêm các chỉ thị SSR để
đánh giá các cặp có hệ số tương đồng di truyền là 1 phục vụ cho việc xác định các giống trùng lặp.
TÀI LIỆU THAM KHẢO Bộ Nông nghiệp và Phát triển nông thôn, 2011. QCVN
01-65:2011/BNNPTNT. Quy chuẩn Quốc gia về khảo nghiệm giống cây trồng.
Nei M. and Li T., 1972. Genetic distance between populations. Am. Nat., 106: 283-292.
PowerMarker 3.25 - Statistical Software for Genetic Marker data analysis. www.mybiosoftware.com/powermarker-3-25-statistical-software-genetics.
Saal B., Wricke G., 1999. Devolopment of simple sequence repeat makers in rye (Secale cereale L.). Genome, 42(5): 964-972.
Zheng K.L. , Huang N., Bennett J., Khush G.S., 1995. PCR - Based Marker Assisted Selection in Rice Breeding. International Rice Research Institute, Manila, the Philippines. 300p.
Mark W., Simon K., Marie LJ., and Nigel B., 2004. Varietal and geographic origin of five commercial USA rice samples labelled and Basmati rice. Working party on food authenticity methodology report. 20 pages.
Xiao X.Y., Wang Y.P., Zhang J.Y., Li S.G., Rong T.Z., 2006. SSR marker-based genetic diversity fingerprinting of hybrid rice in Sichuan, China. Chinese J. Rice Sci., 20(1): 1-7.
Ying J.Z., Shi Y.F., E Z.G, Zeng R.Z., Chen J., Zhu Z.W., Zhuang J.Y., 2007. Construction and testing of a primary microsatellite database of major rice varieties in China. Rice Science, 14(4): 247-255.
Distinction of the similar rice varieties using molecular markers to support for DUS test
Tran Long, Luu Minh Cuc, Nguyen Quang Sang, Pham Xuan HoiAbstractDNA profile can become an important indicator of support in the DUS test as it provides an accurate assessment of the inspection and identification of a new plant variety. The research was carried out to analyze some rice varieties with similar morphological characteristics by using DNA markers, combining with 62 morphological traits. The analysis by the POWER MARKER software showed that the frequency of alleles was 4.9 - 7.2; the general similarity of 62 morphological traits and SSR markers ranged from 0.36 to 0.64, while the number of genetic similarity coefficient was from 0.33 to 0.59. By using a set of 30 SSR markers for analysing of 19 rice varieties similar in pairs tested by DUS, showed that: The pairs TTC (Nep Trieu Tien cu) - TTM (Nep Trieu Tien moi) and QX22 - X33 had a genetic similarity coefficient of 1.00. The most different pair in genetic correlation coefficient were DB15 and NH3 (0.63), Thinh du 8 - Thinh du 4 (0.87). The remaining pairs (ND1 - ND 2; Nhiet đoi 1 - CL10; HT1 - HT6 - HT9, TBR36 - TQ08, Nep Lang Lieu - Nep GRQ10) were unclear differences.Keywords: DUS test, molecular marker, rice variety
Ngày nhận bài: 18/9/2018Ngày phản biện: 24/9/2018
Người phản biện: TS. Trần Danh SửuNgày duyệt đăng: 15/10/2018

83
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
ẢNH HƯỞNG CỦA LIÊU LƯỢNG PHÂN LÂN VÀ CHE PHỦ ĐẾN SINH TRƯỞNG VÀ NĂNG SUẤT CỦA GIỐNG LẠC L14 Ở VỤ XUÂN TẠI GIA LÂM - HÀ NỘI
Vũ Ngọc Thắng1, Nguyễn Thị Yến1, Nông Thảo Diễm1, Nguyễn Ngọc Quất2, Trần Anh Tuấn1
TÓM TẮTThí nghiệm được tiến hành nhằm đánh giá ảnh hưởng của 3 mức lân bón (60, 90, 120 kg P2O5/ha) đến sinh
trưởng và năng suất của giống lạc L14 trong các điều kiện che phủ khác nhau (không che phủ, che phủ bằng trấu, che phủ bằng nilon). Kết quả thí nghiệm cho thấy giống lạc L14 được trồng trong điều kiện có che phủ có thời gian sinh trưởng ngắn hơn trong khi các chỉ tiêu sinh trưởng và năng suất lại cao hơn so với trồng trong điều kiện không được che phủ. Bên cạnh đó, trong cùng một điều kiện che phủ, khi tăng lượng lân bón thì các chỉ tiêu này cũng có xu hướng tăng lên. Các chỉ tiêu sinh trưởng và năng suất của giống lạc L14 đạt giá trị cao nhất được quan sát tại mức bón 120 kg P2O5/ha trong điều kiện che phủ nilon. Đánh giá hiệu quả kinh tế kết quả cho thấy lãi thuần đạt giá trị cao nhất trong điều kiện che phủ trấu với mức lân bón là 120 kg P2O5/ha.
Từ khóa: Vật liệu che phủ, lạc, phân lân, sinh trưởng, năng suất
1 Khoa Nông học, Học viện Nông nghiệp Việt Nam; 2 Viện Cây lương thực và Cây thực phẩm
I. ĐẶT VẤN ĐỀBên cạnh công tác chọn tạo giống thì áp dụng các
biện pháp kỹ thuật tổng hợp cũng góp phần mang lại nhiều thành công trong sản xuất lạc. Với khả năng ưu việt của kỹ thuật che phủ nilon như làm tăng nhiệt độ đất, duy trì độ ẩm, cải thiện kết cấu đất, tạo môi trường thuận lợi cho vi sinh vật hoạt động, hạn chế sự thoát hơi nước và dinh dưỡng, tăng khả năng phát triển của hệ thống rễ giúp cho cho cây lạc sinh trưởng và phát triển tốt cho năng suất cao (Duan Shufen, 1999; Trần Đình Long và ctv., 1999). Tuy nhiên, áp dụng kỹ thuật che phủ nilon dễ gây ô nhiễm và không thân thiện với môi trường. Do vậy, nhằm tăng năng suất cho lạc ngoài biện pháp che phủ nilon trong những năm gầy đây đã có nhiều công trình công bố áp dụng các biện pháp kỹ thuật che phủ bằng các vật liệu khác như trấu, xác thực vật đã mang lại hiệu quả kinh tế cao tương đương với kỹ thuật che phủ nilon đặc biệt tránh được ô nhiễm môi trường (Ramakrishna et al., 2006; Vũ Ngọc Thắng và Vũ Đình Chính, 2007; Vũ Văn Liết và ctv., 2010). Mặc dù các kỹ thuật che phủ và lượng phân bón tối thích cho lạc đã được các nhà khoa học trong và ngoài nước đầu tư nghiên cứu tuy nhiên đánh giá hiệu quả của các mức lân bón khác nhau cho lạc trong các điều kiện che phủ khác nhau đến nay chưa được quan tâm đúng mức.
Mục đích của nghiên cứu này nhằm đánh giá tác động của các mức lân bón cho giống lạc L14 trong các điều kiện che phủ khác nhau góp phần xây dựng quy trình thâm canh tăng năng suất cho lạc.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưu Giống lạc L14 được Trung tâm Nghiên cứu và
Phát triển Đậu đỗ - Viện Cây lương thực và Cây thực phẩm chọn lọc theo phương pháp chọn lọc quần thể từ dòng lạc QĐ5 từ tập đoàn lạc nhập nội của Trung Quốc. Giống được công nhận chính thức là giống tiến bộ kỹ thuật năm 2002.
2.2. Phương pháp nghiên cưuThí nghiệm được bố trí theo phương pháp ô lớn
ô nhỏ (Slip - plot) với 2 nhân tố. Nền là: 30 kg N + 60 kg K2O + 1.000 kg phân vi sinh Sông Gianh cho 1 ha. Nhân tố chính là 3 mức lân bón (60; 90; 120 kg P2O5/ha). Nhân tố phụ là các điều kiện che phủ khác nhau (Không che phủ; Che phủ bằng trấu; Che phủ bằng nilon). Diện tích mỗi ô thí nghiệm là 10 m2.
Các chỉ tiêu theo dõi theo quy chuẩn kỹ thuật Quốc gia về khảo nghiệm giá trị canh tác và sử dụng giống lạc QCVN 01-57: 2011/BNNPTNN của Bộ Nông nghiệp và Phát triển nông thôn (2011).
Số liệu được xử lý theo phương pháp phân tích phương sai (ANOVA) bằng phần mềm IRISTAT 5.0 và Excel.
2.3. Thời gian và địa điểm nghiên cưuThí nghiệm được thực hiện trên nền đất phù sa
trong đê sông Hồng, tại khu đất thí nghiệm Khoa Nông học - Học viện Nông nghiệp Việt Nam. Thời gian triển khai thí nghiệm từ tháng 2 đến tháng 6 năm 2018.

84
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Ảnh hưởng của các mưc bón lân đến thời gian sinh trưởng của giống lạc L14 trong các điều kiện che phủ khác nhau trong vụ Xuân
Theo dõi ảnh hưởng của các mức bón lân đến thời gian sinh trưởng của giống lạc L14 trong các điều kiện che phủ khác nhau kết quả thu được trình bày tại bảng 1. Số liệu bảng 1 cho thấy không có sự sai khác giữa các mức lân bón đến thời gian từ gieo đến mọc, từ gieo đến ra hoa và tổng thời gian sinh trưởng của giống lạc L14 trong cùng một điều kiện che phủ ngoại trừ mức bón 120 kg P2O5/ha trong điều kiện không che phủ. Tuy nhiên có sự sai khác giữa các công thức che phủ (Bảng 1). Sử dụng vật liệu che phủ giúp rút ngắn thời gian từ gieo đến mọc, tổng thời gian sinh trưởng và đặc biệt nâng cao tỉ lệ nảy mầm cho giống lạc L14. Kết quả nghiên cứu này cũng tương đồng với các kết quả nghiên cứu của các tác giả Ramakrishna và cộng tác viên (2006); Vũ Ngọc Thắng và Vũ Đình Chính (2007); Vũ Văn Liết và cộng tác viên (2010).
Bảng 1. Ảnh hưởng của các mức bón lân đến thời gian sinh trưởng của giống lạc L14
trong các điều kiện che phủ khác nhau
3.2. Ảnh hưởng của các mưc lân bón đến một số chỉ tiêu sinh trưởng của giống lạc L14 trong các điều kiện che phủ khác nhau trong vụ Xuân
Theo dõi ảnh hưởng của các mức lân bón đến một số chỉ tiêu sinh trưởng của giống lạc L14 trong các điều kiện che phủ khác nhau kết quả thu được trình bày tại bảng 2.
Bảng 2. Ảnh hưởng của các mức lân bón đến một số chỉ tiêu sinh trưởng của giống lạc L14
trong các điều kiện che phủ khác nhau
Ghi chú: CT: công thức; MB: mức bón.
Số liệu bảng 2 cho thấy có sự sai khác có ý nghĩa giữa các mức lân bón về chiều cao cây, chiều dài cành cấp 1 thứ nhất của giống L14 trong cùng một điều kiện che phủ. Khi tăng hàm lượng lân bón thì các chỉ tiêu trên cũng có xu hướng tăng lên. Tuy nhiên số lá trên thân chính không có sự sai khác có ý nghĩa ở mức bón 90 kg P2O5/ha và 120 kg P2O5/ha nhưng lại có sự sai khác có ý nghĩa giữa mức bón 60 kg P2O5/ha và mức bón 90 kg P2O5/ha hoặc 120 kgP2O5/ha. So sánh giữa các công thức che phủ kết quả cho thấy có sựa sai khác có ý nghĩa giữa công
Công thưc
che phủ
Mưc lân bón
(kg/ha)
Gieo đến moc
(ngày)
Tỷ lệ moc mầm (%)
Gieo đến ra
hoa(ngày)
TGST (ngày)
Không che phủ
60 7 88,31 35 12890 7 88,23 35 128
120 7 88,81 35 127
Che phủ trấu
60 5 92,48 34 12390 5 91,87 34 123
120 5 91,94 34 123
Che phủ
nilon
60 5 93,23 33 12090 5 93,09 33 120
120 5 93,54 33 120
TB côngthức
che phủ
Không che phủ 7 88,45 35 128
Che phủ trấu 5 92,10 34 123
Che phủ nilon 5 93,29 33 120
TB mức lân bón
60 6 91,34 34 12490 6 91,06 34 124
120 6 91,43 34 123
Công thưc che
phủ
Mưc lân bón
(kg/ha)
Chiều cao cây
cuối cùng (cm)
Chiều dài cành
cấp 1 thư nhất
(cm)
Số lá trên thân
chính (lá)
Không che phủ
60 43,55 52,00 19,8290 48,26 57,13 20,87
120 51,86 61,61 21,31
Che phủ trấu
60 56,75 65,09 21,2690 59,08 67,42 22,41
120 60,29 69,15 22,53
Che phủ nilon
60 59,10 66,59 21,6890 62,24 68,09 22,81
120 63,52 69,48 22,94CV (%) 10,4 9,1 9,4
LSD(CT ˟ MB)0,05 2,39 2,35 0,92
TB công thức che
phủ
Không che phủ 47,89 56,91 20,67
Che phủ trấu 58,71 67,22 22,07
Che phủ nilon 61,62 68,05 22,48
LSD(CT)0,05 0,32 1,70 0,98
TB mức lân bón
60 53,13 61,23 20,9290 56,53 64,21 22,03
120 58,56 66,75 22,26LSD(MB)0,05 1,38 1,36 0,53

85
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
thức che phủ và công thức không che phủ tuy nhiên không có sự sai khác giữa công thức che phủ trấu và công thức che phủ nilon về chiều dài cành cấp 1 thứ nhất và số lá trên thân chính. Kết quả nghiên cứu này cũng tương đồng với kết quả nghiên cứu của tác giả Vũ Ngọc Thắng và Vũ Đình Chính (2007). Bên cạnh đó tác giả Ramakrishna và cộng tác viên (2006) thực hiện nghiên cứu trong vụ Xuân ở miền Bắc Việt Nam và theo nghiên cứu dài hơi (từ năm 1992 đến năm 1999) của tác giả Ghosh và cộng tác viên (2006) trong vụ Hè ở Ấn Độ cũng có đánh giá chung khi so sánh giữa che phủ bằng vật liệu nilon chuyên dụng và rơm thì cả hai loại vật liệu cơ bản đều có ảnh hưởng tốt đến sinh trưởng, phát triển của lạc hơn so với không che phủ.
3.3. Ảnh hưởng của các mưc lân bón đến diện tích lá và chỉ số diện tích lá của giống lạc L14 trong các điều kiện che phủ khác nhau trong vụ Xuân
Diện tích lá và chỉ số diện tích lá là hai chỉ tiêu quan trọng có liên quan chặt chẽ đến sinh trưởng, phát triển và năng suất của cây trồng. Kết quả ở bảng 3 cho thấy diện tích lá và chỉ số diện tích lá của giống lạc L14 có xu hướng tăng dần từ thời kỳ cây
con và đạt giá trị cao vào thời kỳ quả chắc. Có sự sai khác giữa các mức lân bón về diện tích lá và chỉ số diện tích lá của giống L14 trong cùng một điều kiện che phủ. Khi tăng hàm lượng lân bón thì diện tích lá và chỉ số diện tích lá cũng có xu hướng tăng lên. Kết quả nghiên cứu này cũng tương tự như kết quả nghiên cứu trên cây đậu tương của tác giả Đoàn Văn Lưu và cộng tác viên (2017): khi tăng hàm lượng lân bón từ 60 kg/ha lên 90 kg/ha và 120 kg/ha thì diện tích lá và chỉ số diện tích lá cũng có xu hướng tăng lên và đạt giá trị cao nhất ở mức bón 120 kg/ha. Tuy nhiên, trong kết quả nghiên cứu trình bày trong bài báo này, tại thời kỳ ra hoa và thời kỳ quả chắc diện tích lá và chỉ số diện tích lá không có sự sai khác có ý nghĩa ở mức bón 90 kg P2O5/ha và 120 kg P2O5/ha nhưng lại có sự sai khác có ý nghĩa giữa mức bón 60 kg P2O5/ha và mức bón 90 kg P2O5/ha hoặc 120 kg P2O5/ha. So sánh giữa các công thức che phủ kết quả cho thấy có sựa sai khác có ý nghĩa giữa công thức che phủ và công thức không che phủ tuy nhiên không có sự sai khác giữa công thức che phủ trấu và công thức che phủ nilon về diện tích lá và chỉ số diện tích lá.
Bảng 3. Ảnh hưởng của các mức lân bón đến diện tích lá và chỉ số diện tích lá của giống lạc L14 trong các điều kiện che phủ khác nhau
Ghi chú: CT: công thức; MB: mức bón.
Công thưc che phủ
Mưc lân bón
(kg/ha)
Cây con Thời kì ra hoa Thời kì quả chắcDiện tích lá (dm2/cây)
LAI (m2 lá/m2 đất)
Diện tích lá (dm2/cây)
LAI (m2 lá/m2 đất)
Diện tích lá (dm2/cây)
LAI (m2 lá/m2 đất)
Không che phủ
60 2,11 0,63 6,57 1,97 11,42 3,4390 2,40 0,72 7,25 2,18 12,36 3,71
120 2,52 0,76 7,92 2,38 12,65 3,80
Che phủ trấu
60 2,61 0,78 8,09 2,43 13,01 3,9090 2,81 0,84 8,86 2,66 13,31 3,99
120 3,01 0,90 9,11 2,73 13,73 4,12
Che phủ nilon
60 2,87 0,86 8,13 2,44 13,14 3,9490 2,98 0,89 9,04 2,71 13,35 4,01
120 3,09 0,93 9,08 2,72 13,39 4,02CV (%) 4,5 4,0 4,5
LSD(CT ˟ MB)0,05 0,62 0,94 0,16
TB công thức che phủ
Không che phủ 2,34 0,70 7,25 2,17 12,14 3,64
Che phủ trấu 2,81 0,84 8,69 2,61 13,35 4,01
Che phủ nilon 2,98 0,89 8,75 2,63 13,29 3,99
LSD(CT)0,05 0,29 0,12 0,31
TB mức lân bón
60 2,53 0,76 7,60 2,28 12,52 3,7690 2,73 0,82 8,38 2,52 13,01 3,90
120 2,87 0,86 8,70 2,62 13,26 3,98LSD(MB)0,05 0,26 0,54 0,32

86
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
3.4. Ảnh hưởng của các mưc lân bón đến khả năng tích lũy chất khô trên cây của giống lạc L14 trong các điều kiện che phủ khác nhau trong vụ Xuân
Khả năng tích lũy chất khô của giống lạc L14 có xu hướng tăng dần từ thời kỳ cây con và đạt giá trị cao vào thời kỳ quả chắc. Có sự sai khác có ý thống kê giữa các mức lân bón về khả năng tích lũy chất khô của giống L14 trong cùng một điều kiện che phủ. Khi tăng hàm lượng lân bón thì khả năng tích lũy chất khô cũng có xu hướng tăng lên. So sánh giữa các công thức che phủ kết quả cho thấy có sựa sai khác có ý nghĩa giữa các công thức che phủ và công thức không được che phủ. Điều này cũng đã được kết luận trong kết quả nghiên cứu của tác giả Vũ Văn Liết và cộng tác viên (2010) khi nghiên cứu tuyển chọn giống và vật liệu che phủ thích hợp cho lạc xuân tại Bắc Giang.
Bảng 4. Ảnh hưởng của các mức lân bón đến khả năng tích lũy chất khô của giống lạc L14
trong các điều kiện che phủ khác nhau
Ghi chú: CT: công thức; MB: mức bón.
Trong nghiên cứu ở đây cũng cho thấy giống lạc L14 trong điều kiện không che phủ có khả năng tích lũy chất khô đạt giá trị thấp nhất và che phủ bằng nilon có khả năng tích lũy chất khô đạt giá trị cao nhất tiếp đến là công thức che phủ bằng trấu. Kết quả nghiên cứu này cũng tương đồng với kết quả nghiên cứu trước đây về ảnh hưởng của một số vật liệu che phủ đến sinh trưởng, phát triển và năng suất của giống lạc L14 trong điều kiện vụ Thu trên đất Gia Lâm Hà Nội với kết luận là trồng trong điều kiện che phủ nilon cây luôn có độ ẩm đất cao hơn so với không che phủ và che phủ trấu; đây cũng là một trong những nhân tố làm tăng khả năng tích lũy sinh khối của giống lạc L14 (Vũ Ngọc Thắng và Vũ Đình Chính, 2007).
3.5. Ảnh hưởng của các mưc lân bón đến khả năng hình thành nốt sần của giống lạc L14 trong các điều kiện che phủ khác nhau trong vụ Xuân
Khả năng hình thành nốt sần của giống lạc L14 cũng có xu hướng tăng dần từ thời kỳ cây con và đạt giá trị cao vào thời kỳ quả chắc. Có sự sai khác giữa các mức lân bón về khả năng hình thành nốt sần của giống L14 trong cùng một điều kiện che phủ. Khi tăng hàm lượng lân bón thì số lượng và khối lượng nốt sần cũng có xu hướng tăng. Kết quả nghiên cứu này cũng tương tự như kết quả nghiên cứu trên cây đậu tương của tác giả Đoàn Văn Lưu và cộng tác viên (2017), trong nghiên cứu tác giả cũng cho thấy khi tăng hàm lượng lân bón từ 60 lên 90 kg/ha và 120 kg/ha thì số lượng và khối lượng nốt sần cũng có xu hướng tăng lên và đạt giá trị cao nhất ở mức bón 120 kg/ha. Tuy nhiên nếu tiếp tục tăng mức bón lên 150 kg/ha thì số lượng và khối lượng nốt sần lại có xu hướng giảm xuống. So sánh giữa các công thức che phủ kết quả cho thấy có sự sai khác giữa các công thức che phủ và công thức che phủ bằng nilon có số lượng và khối lượng nốt sần đạt giá trị cao nhất tiếp đến là công thức che phủ bằng trấu (Bảng 5).
3.6. Ảnh hưởng của các mưc lân bón đến một số chỉ tiêu sinh lý của giống lạc L14 trong các điều kiện che phủ khác nhau trong vụ Xuân
Từ kết quả nghiên cứu thu được ở bảng 6 cho thấy có sự sai khác có ý nghĩa thống kê giữa các mức lân bón đến chỉ số SPAD của giống lạc L14 ở cả 3 thời kì theo dõi trong cùng điều kiện che phủ. Chỉ số SPAD tăng dần khi tăng dần mức lân bón và đạt giá trị cao nhất ở mức bón 120 kg P2O5/ha. Bên cạnh đó chỉ số SPAD có xu hướng tăng dần từ thời kỳ cây con và đạt giá trị cao vào thời kỳ ra hoa. Tuy nhiên bước vào thời kì quả chắc chỉ số SPAD có xu hướng giảm
Công thưc che
phủ
Mưc lân bón
(kg/ha)
Thời kỳ cây con (g/cây)
Thời kỳ ra hoa(g/cây)
Thời kỳ quả chắc (g/cây)
Không che phủ
60 1,78 10,18 18,0590 2,15 12,19 21,21
120 3,01 12,66 24,02
Che phủ trấu
60 2,17 11,70 22,4290 3,72 12,98 24,26
120 4,26 13,64 25,25
Che phủ nilon
60 3,25 12,55 23,0390 4,01 13,27 24,78
120 4,42 13,92 25,42CV (%) 7,2 7,1 6,4
LSD(CT ˟ MB)0,05 0,13 0,47 0,58
TB công thức che
phủ
Không che phủ 2,31 11,68 21,09
Che phủ trấu 3,38 12,77 23,98
Che phủ nilon 3,89 13,25 24,41
LSD(CT)0,05 0,42 0,46 0,3360 2,40 11,48 21,17
TB mức lân bón 90 3,29 12,81 23,42
120 3,90 13,41 24,90
LSD(MB)0,05 0,13 0,47 0,58

87
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
xuống. So sánh giữa các công thức che phủ kết quả cho thấy có sự sai khác có ý nghĩa giữa các công thức che phủ ở 2 thời kỳ cây con và ra hoa. Tuy nhiên, bước sang thời kỳ quả chắc không có sự sai khác có ý nghĩa ở mức bón 90 kg P2O5/ha và 120 kg P2O5/ha
nhưng lại có sự sai khác có ý nghĩa giữa mức bón 60 kg P2O5/ha và mức bón 90 kg P2O5/ha hoặc 120 kg P2O5/ha. Bên cạnh đó công thức che phủ bằng nilon cũng luôn có chỉ số SPAD đạt giá trị cao nhất tiếp đến là công thức che phủ bằng trấu (Bảng 6).
Bảng 5. Ảnh hưởng của mức lân bón đến khả năng hình thành nốt sần của giống lạc L14 trong các điều kiện che phủ khác nhau
Ghi chú: CT: công thức; MB: mức bón.
Công thưc che phủ
Mưc lân bón(kg/ha)
Thời kỳ cây con Thời kỳ ra hoa Thời kỳ quả chắcTổng số nốt sần
(nốt/cây)
Khối lượng nốt sần(g/cây)
Tổng số nốt sần
(nốt/cây)
Khối lượng nốt sần(g/cây)
Tổng số nốt sần
(nốt/cây)
Khối lượng nốt sần(g/cây)
Không che phủ
60 14,51 0,03 44,12 0,15 77,54 0,3690 17,47 0,05 50,05 0,17 99,69 0,49
120 19,80 0,06 55,12 0,19 108,65 0,53
Che phủ trấu
60 18,17 0,06 53,91 0,18 103,91 0,5290 20,85 0,07 61,46 0,21 109,16 0,53
120 23,80 0,08 69,57 0,24 115,14 0,55
Che phủ nilon
60 20,78 0,07 58,46 0,26 109,80 0,5390 23,61 0,08 64,90 0,28 111,95 0,56
120 24,87 0,09 71,25 0,29 121,69 0,58CV (%) 5,9 7,2 4,9 9,0 9,1 6,9
LSD(CT ˟ MB)0,05 0,71 0,84 2,94 0,12 5,78 0,18
TB công thức che phủ
Không che phủ 17,26 0,47 49,76 0,17 95,29 0,46Che phủ trấu 20,94 0,70 61,65 0,21 109,43 0,53
Che phủ nilon 23,09 0,80 64,87 0,28 114,48 0,56LSD(CT)0,05 4,42 0,075 7,90 0,087 4,34 0,013
TB mức lân bón
60 17,82 0,53 52,16 0,20 97,08 0,4790 20,64 0,67 58,8 0,22 106,93 0,53
120 22,82 0,77 65,31 0,24 115,16 0,55LSD(MB)0,05 4,10 0,048 5,40 0,068 3,34 0,011
3.7. Ảnh hưởng của các mưc lân bón đến các yếu tố cấu thành năng suất của giống lạc L14 trong các điều kiện che phủ khác nhau trong vụ Xuân
Trong sản xuất nông nghiệp thì mục đích cuối cùng và quan trọng nhất là đạt được hiệu quả kinh tế cao, thu được sản phẩm với năng suất cao, chất lượng tốt. Năng suất cây trồng thể hiện kết quả tác động tổng hợp của các yếu tố nội tại với điều kiện môi trường và các biện pháp kỹ thuật tác động. Theo dõi ảnh hưởng của các mức lân bón đến một số chỉ tiêu liên quan đến năng suất của giống lạc L14 trên các điều kiện che phủ khác nhau kết quả được trình bày tại bảng 7. Số liệu bảng 7 cho thấy có sự sai khác giữa các mức lân bón đến tổng số hoa nở trên cây, tổng số quả trên cây, tỉ lệ quả chắc trên cây, khối lượng 100 quả, 100 hạt trong cùng điều kiện che phủ của giống lạc L14. Tăng lượng lân bón các chỉ tiêu liên quan đến năng suất của giống lạc L14 cũng có
xu hướng tăng lên. Ở mức bón 120 kg P2O5/ha trong điều kiện che phủ nilon cho các chỉ tiêu này đạt giá trị cao nhất. So sánh giữa các công thức che phủ kết quả cho thấy giống lạc L14 ở công thức được che phủ luôn có các chỉ tiêu trên cao hơn so với công thức không được che phủ và công thức che phủ nilon có giá trị cao nhất. Kết quả nghiên cứu này cũng được ghi nhận như kết quả nghiên cứu trước đây của tác giả Vũ Ngọc Thắng và Vũ Đình Chính (2007) về ảnh hưởng của một số vật liệu che phủ đến sinh trưởng, phát triển và năng suất của giống lạc L14 trong điều kiện vụ Thu trên đất Gia Lâm Hà Nội cũng. Nghiên cứu của các tác giả này cho thấy trồng trong điều kiện che phủ nilon các yếu tố cấu thành năng suất và năng suất của giống lạc L14 cũng luôn đạt giá trị cao nhất. Tuy nhiên, trong nghiên cứu này của nhóm tác giả trình này trong bài báo này tỷ lệ nhân và tổng số quả/cây không có sự sai khác giữa công thức che phủ
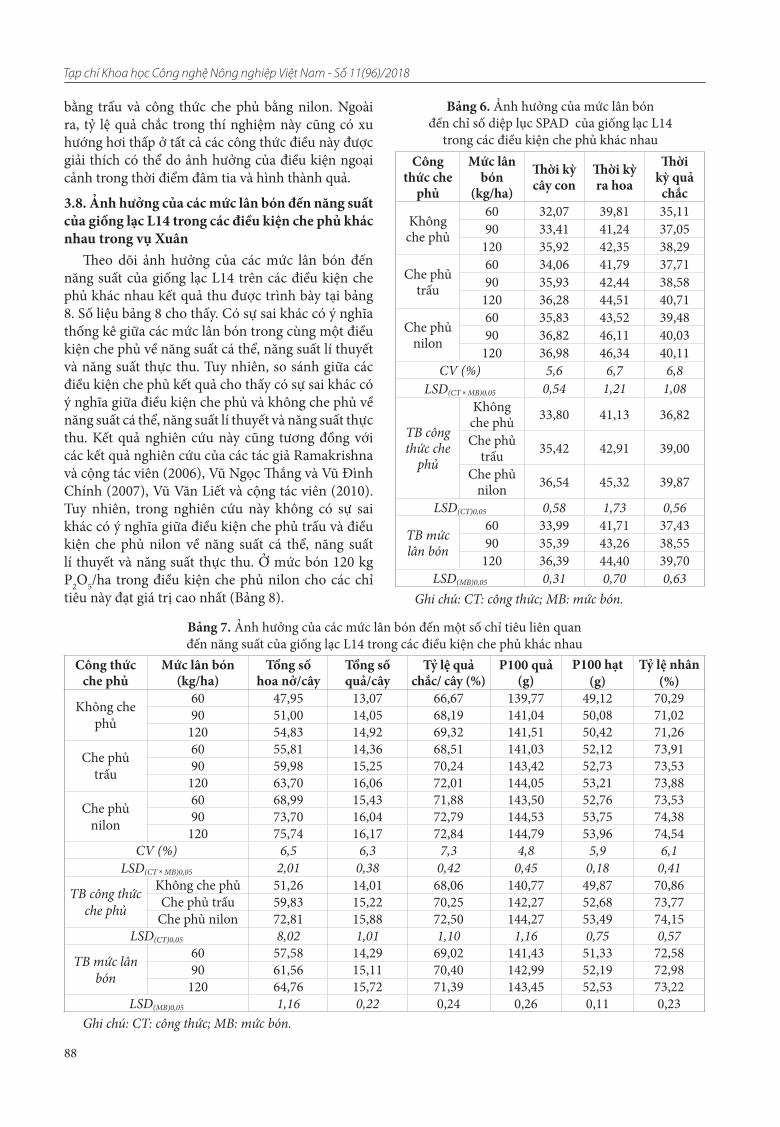
88
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
bằng trấu và công thức che phủ bằng nilon. Ngoài ra, tỷ lệ quả chắc trong thí nghiệm này cũng có xu hướng hơi thấp ở tất cả các công thức điều này được giải thích có thể do ảnh hưởng của điều kiện ngoại cảnh trong thời điểm đâm tia và hình thành quả.
3.8. Ảnh hưởng của các mưc lân bón đến năng suất của giống lạc L14 trong các điều kiện che phủ khác nhau trong vụ Xuân
Theo dõi ảnh hưởng của các mức lân bón đến năng suất của giống lạc L14 trên các điều kiện che phủ khác nhau kết quả thu được trình bày tại bảng 8. Số liệu bảng 8 cho thấy. Có sự sai khác có ý nghĩa thống kê giữa các mức lân bón trong cùng một điều kiện che phủ về năng suất cá thể, năng suất lí thuyết và năng suất thực thu. Tuy nhiên, so sánh giữa các điều kiện che phủ kết quả cho thấy có sự sai khác có ý nghĩa giữa điều kiện che phủ và không che phủ về năng suất cá thể, năng suất lí thuyết và năng suất thực thu. Kết quả nghiên cứu này cũng tương đồng với các kết quả nghiên cứu của các tác giả Ramakrishna và cộng tác viên (2006), Vũ Ngọc Thắng và Vũ Đình Chính (2007), Vũ Văn Liết và cộng tác viên (2010). Tuy nhiên, trong nghiên cứu này không có sự sai khác có ý nghĩa giữa điều kiện che phủ trấu và điều kiện che phủ nilon về năng suất cá thể, năng suất lí thuyết và năng suất thực thu. Ở mức bón 120 kg P2O5/ha trong điều kiện che phủ nilon cho các chỉ tiêu này đạt giá trị cao nhất (Bảng 8).
Bảng 6. Ảnh hưởng của mức lân bón đến chỉ số diệp lục SPAD của giống lạc L14
trong các điều kiện che phủ khác nhau
Ghi chú: CT: công thức; MB: mức bón.
Công thưc che
phủ
Mưc lân bón
(kg/ha)
Thời kỳ cây con
Thời kỳ ra hoa
Thời kỳ quả
chắc
Không che phủ
60 32,07 39,81 35,1190 33,41 41,24 37,05
120 35,92 42,35 38,29
Che phủ trấu
60 34,06 41,79 37,7190 35,93 42,44 38,58
120 36,28 44,51 40,71
Che phủ nilon
60 35,83 43,52 39,4890 36,82 46,11 40,03
120 36,98 46,34 40,11CV (%) 5,6 6,7 6,8
LSD(CT ˟ MB)0,05 0,54 1,21 1,08
TB công thức che
phủ
Không che phủ 33,80 41,13 36,82
Che phủ trấu 35,42 42,91 39,00
Che phủ nilon 36,54 45,32 39,87
LSD(CT)0,05 0,58 1,73 0,56
TB mức lân bón
60 33,99 41,71 37,4390 35,39 43,26 38,55
120 36,39 44,40 39,70LSD(MB)0,05 0,31 0,70 0,63
Bảng 7. Ảnh hưởng của các mức lân bón đến một số chỉ tiêu liên quan đến năng suất của giống lạc L14 trong các điều kiện che phủ khác nhau
Ghi chú: CT: công thức; MB: mức bón.
Công thưc che phủ
Mưc lân bón (kg/ha)
Tổng số hoa nở/cây
Tổng số quả/cây
Tỷ lệ quả chắc/ cây (%)
P100 quả (g)
P100 hạt (g)
Tỷ lệ nhân (%)
Không che phủ
60 47,95 13,07 66,67 139,77 49,12 70,2990 51,00 14,05 68,19 141,04 50,08 71,02
120 54,83 14,92 69,32 141,51 50,42 71,26
Che phủ trấu
60 55,81 14,36 68,51 141,03 52,12 73,9190 59,98 15,25 70,24 143,42 52,73 73,53
120 63,70 16,06 72,01 144,05 53,21 73,88
Che phủ nilon
60 68,99 15,43 71,88 143,50 52,76 73,5390 73,70 16,04 72,79 144,53 53,75 74,38
120 75,74 16,17 72,84 144,79 53,96 74,54CV (%) 6,5 6,3 7,3 4,8 5,9 6,1
LSD(CT ˟ MB)0,05 2,01 0,38 0,42 0,45 0,18 0,41
TB công thức che phủ
Không che phủ 51,26 14,01 68,06 140,77 49,87 70,86Che phủ trấu 59,83 15,22 70,25 142,27 52,68 73,77
Che phủ nilon 72,81 15,88 72,50 144,27 53,49 74,15LSD(CT)0,05 8,02 1,01 1,10 1,16 0,75 0,57
TB mức lân bón
60 57,58 14,29 69,02 141,43 51,33 72,5890 61,56 15,11 70,40 142,99 52,19 72,98
120 64,76 15,72 71,39 143,45 52,53 73,22LSD(MB)0,05 1,16 0,22 0,24 0,26 0,11 0,23
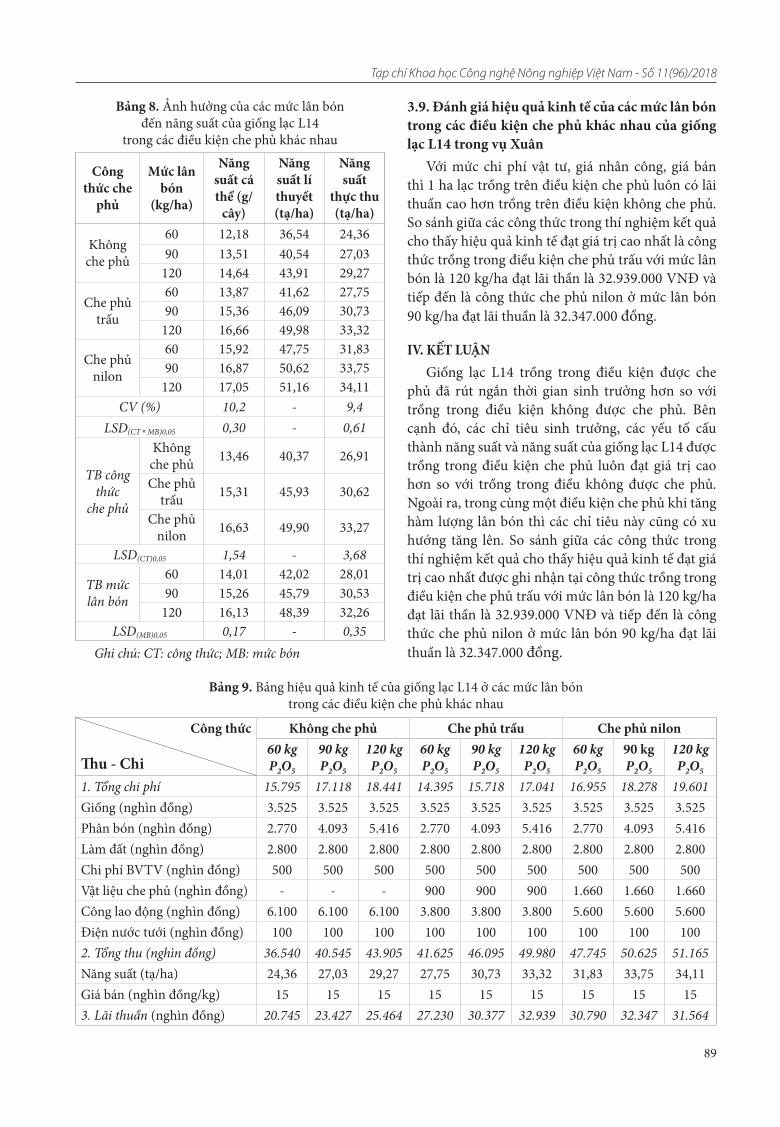
89
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Bảng 8. Ảnh hưởng của các mức lân bón đến năng suất của giống lạc L14
trong các điều kiện che phủ khác nhau
Ghi chú: CT: công thức; MB: mức bón
3.9. Đánh giá hiệu quả kinh tế của các mưc lân bón trong các điều kiện che phủ khác nhau của giống lạc L14 trong vụ Xuân
Với mức chi phí vật tư, giá nhân công, giá bán thì 1 ha lạc trồng trên điều kiện che phủ luôn có lãi thuần cao hơn trồng trên điều kiện không che phủ. So sánh giữa các công thức trong thí nghiệm kết quả cho thấy hiệu quả kinh tế đạt giá trị cao nhất là công thức trồng trong điều kiện che phủ trấu với mức lân bón là 120 kg/ha đạt lãi thần là 32.939.000 VNĐ và tiếp đến là công thức che phủ nilon ở mức lân bón 90 kg/ha đạt lãi thuần là 32.347.000 đồng.
IV. KẾT LUẬNGiống lạc L14 trồng trong điều kiện được che
phủ đã rút ngắn thời gian sinh trưởng hơn so với trồng trong điều kiện không được che phủ. Bên cạnh đó, các chỉ tiêu sinh trưởng, các yếu tố cấu thành năng suất và năng suất của giống lạc L14 được trồng trong điều kiện che phủ luôn đạt giá trị cao hơn so với trồng trong điều không được che phủ. Ngoài ra, trong cùng một điều kiện che phủ khi tăng hàm lượng lân bón thì các chỉ tiêu này cũng có xu hướng tăng lên. So sánh giữa các công thức trong thí nghiệm kết quả cho thấy hiệu quả kinh tế đạt giá trị cao nhất được ghi nhận tại công thức trồng trong điều kiện che phủ trấu với mức lân bón là 120 kg/ha đạt lãi thần là 32.939.000 VNĐ và tiếp đến là công thức che phủ nilon ở mức lân bón 90 kg/ha đạt lãi thuần là 32.347.000 đồng.
Bảng 9. Bảng hiệu quả kinh tế của giống lạc L14 ở các mức lân bón trong các điều kiện che phủ khác nhau
Công thưc
Thu - Chi
Không che phủ Che phủ trấu Che phủ nilon60 kg P2O5
90 kg P2O5
120 kg P2O5
60 kg P2O5
90 kg P2O5
120 kg P2O5
60 kg P2O5
90 kg P2O5
120 kg P2O5
1. Tổng chi phí 15.795 17.118 18.441 14.395 15.718 17.041 16.955 18.278 19.601Giống (nghìn đồng) 3.525 3.525 3.525 3.525 3.525 3.525 3.525 3.525 3.525Phân bón (nghìn đồng) 2.770 4.093 5.416 2.770 4.093 5.416 2.770 4.093 5.416Làm đất (nghìn đồng) 2.800 2.800 2.800 2.800 2.800 2.800 2.800 2.800 2.800Chi phí BVTV (nghìn đồng) 500 500 500 500 500 500 500 500 500Vật liệu che phủ (nghìn đồng) - - - 900 900 900 1.660 1.660 1.660Công lao động (nghìn đồng) 6.100 6.100 6.100 3.800 3.800 3.800 5.600 5.600 5.600Điện nước tưới (nghìn đồng) 100 100 100 100 100 100 100 100 1002. Tổng thu (nghìn đồng) 36.540 40.545 43.905 41.625 46.095 49.980 47.745 50.625 51.165Năng suất (tạ/ha) 24,36 27,03 29,27 27,75 30,73 33,32 31,83 33,75 34,11Giá bán (nghìn đồng/kg) 15 15 15 15 15 15 15 15 153. Lãi thuần (nghìn đồng) 20.745 23.427 25.464 27.230 30.377 32.939 30.790 32.347 31.564
Công thưc che
phủ
Mưc lân bón
(kg/ha)
Năng suất cá thể (g/
cây)
Năng suất lí thuyết (tạ/ha)
Năng suất
thực thu (tạ/ha)
Không che phủ
60 12,18 36,54 24,3690 13,51 40,54 27,03
120 14,64 43,91 29,27
Che phủ trấu
60 13,87 41,62 27,7590 15,36 46,09 30,73
120 16,66 49,98 33,32
Che phủ nilon
60 15,92 47,75 31,8390 16,87 50,62 33,75
120 17,05 51,16 34,11CV (%) 10,2 - 9,4
LSD(CT ˟ MB)0,05 0,30 - 0,61
TB công thức
che phủ
Không che phủ 13,46 40,37 26,91
Che phủ trấu 15,31 45,93 30,62
Che phủ nilon 16,63 49,90 33,27
LSD(CT)0,05 1,54 - 3,68
TB mức lân bón
60 14,01 42,02 28,0190 15,26 45,79 30,53
120 16,13 48,39 32,26LSD(MB)0,05 0,17 - 0,35

90
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
TÀI LIỆU THAM KHẢOBộ Nông nghiệp và Phát triển nông thôn, 2011. QCVN
01-57:2011/BNNPTNT. Quy chuẩn kỹ thuật Quốc gia về khảo nghiệm giá trị canh tác và sử dụng của giống lạc.
Vũ Văn Liết, Nguyễn Mai Thơm, Ninh Thị Phíp và Lê Thị Minh Thảo, 2010. Nghiên cứu tuyển chọn giống và vật liệu che phủ thích hợp cho lạc xuân tại xã Lệ Viễn - huyện Sơn Động - tỉnh Bắc Giang. Tạp chí Khoa học và Phát triển, 8 (1): 33-39.
Trần Đình Long, Nguyễn Thị Chinh, Nguyễn Văn Thắng và Hoàng Minh Tâm, 1999. Tổng quan tình hình nghiên cứu phát triển kỹ thuật tiến bộ trồng lạc ở Việt Nam trong thời gian qua và phương hướng trong những năm tới. Hội thảo về kỹ thuật trồng lạc toàn quốc. Thanh Hóa, ngày 2 - 4/6/1999.
Đoàn Văn Lưu, Vũ Đình Chính, Vũ Quang Sáng, 2017. Ảnh hưởng của liều lượng phân bón cho đậu tương đông trên đất phù sà huyện Triệu Sơn, tỉnh
Thanh Hóa. Tạp chí Khoa học Nông nghiệp Việt Nam, 15 (12): 1690-1698.
Vũ Ngoc Thắng và Vũ Đình Chính, 2007. Ảnh hưởng của một số vật liệu che phủ đến sinh trưởng, phát triển và năng suất của giống lạc L14 trong điều kiện vụ thu trên đất Gia Lâm - Hà Nội. Tạp chí KHKT Nông nghiệp, 5 (3): 23-31.
Duan Shufen, 1999. Cây lạc ở Trung Quốc - những bí quyết thành công. Tài liệu dịch của Ngô Thế Dân và Phạm Thị Vương. Nhà xuất bản Nông nghiệp.
Ghosh P. K., K.K. Bandyopadhyay, Devi Daya and M. Mohanty, 2006. Evaluation of straw and polythene mulch for enhancing productivity of irrigated summer groundnut. Field Crops Research, 99 (2-3): 76-86.
Ramakrishna A., Hoang Minh Tam, Suhas P. Wani and Tran Dinh Long, 2006. Effect of mulching on soil temperature, moisture, weed infestation and yield of groundnut in northern Vietnam. Field Crops Research, 95 (2-3): 115-125.
Effect of phosphorus fertilizer dose and different types of mulching on growth, development and yield of L14 groundnut variety in Spring season at Gialam - Hanoi
Vu Ngoc Thang, Nguyen Thi Yen, Nong Thao Diem, Nguyen Ngoc Quat, Tran Anh Tuan
AbstractAn experiment was conducted to study effect of 3 phosphorus fertilizer doses (60, 90 and 120 kg P2O5/ha) on growth, development and yield of groundnut variety L14 under different types of mulching (Non-mulching, rice straws mulching, plastic mulching). The result showed that growth and yield of groundnut variety L14 under mulching condition were higher than that without mulching. However, the growth duration of groundnut variety L14 under mulching condition was lower than that without mulching. The growth traits and the yield of groundnut variety L14 increased with increasing phosphorus fertilizer doses under the same mulching or non-mulching condition. The growth duration and the yield of groundnut variety L14 were recored highest at 120 kg P2O5/ha under plastic mulching condition. The highest economic efficiency was recorded at 120 kg P2O5/ha under rice husk mulching condition in comperasion to other treatments.Keywords: Mulching material, groundnut, growth, phosphorus, yield
Ngày nhận bài: 19/9/2018Ngày phản biện: 25/9/2018
Người phản biện: PGS. TS. Ninh Thị PhípNgày duyệt đăng: 15/10/2018
ẢNH HƯỞNG CỦA ĐIÊU KIỆN NUÔI CẤY TỚI KHẢ NĂNG SINH TỔNG HỢP INDOLE-3-ACETIC ACID CỦA VI KHUẨN Bacillus sonorensis LĐ18
Nguyễn Văn Giang1, Trần Thị Đào1, Trần Thị Thúy Hà2, Nguyễn Thu Trang1
TÓM TẮTHiện nay, khai thác và sử dụng các chủng vi sinh vật hữu ích trong kiểm soát sinh học đã và đang được triển khai
mạnh mẽ trong lĩnh vực trồng trọt, bảo vệ thực vật. Các chủng vi sinh vật hữu ích trước khi được đưa vào sản xuất chế phẩm sinh học cần được xác định các điều kiện nuôi thích hợp để đảm bảo mật độ và hoạt tính tốt nhất. Trong thí nghiệm này, ảnh hưởng của nhiệt độ, pH môi trường nuôi cấy và một số nguồn carbon, nitơ tới khả năng tổng 1 Khoa Công nghệ sinh học, Học viện Nông nghiệp Việt Nam2 Trung tâm Công nghệ sinh học Thủy sản - Viện Nghiên cứu Nuôi trồng thủy sản 1

91
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
hợp IAA của chủng vi khuẩn B. sonorensis LĐ18 được đánh giá. Chủng vi khuẩn B. sonorensis LĐ18 tổng hợp IAA tại các giá trị pH 4 - 8. Nhiệt độ thích hợp cho chủng B.sonorensis LĐ18 tổng hợp IAA là 300C. Nguồn carbon thích hợp để tổng hợp IAA là sucrose và sorbitol. Chủng B.sonorensis LĐ18 tổng hợp IAA mạnh nhất khi nguồn nitơ là cao nấm men.
Từ khóa: Bacillus sp., hoạt lực đối kháng nấm, Phytophthora capsici
I. ĐẶT VẤN ĐỀPhytophthora gây bệnh chết nhanh trên hồ tiêu
ảnh hưởng lớn đến năng suất và nguy hiểm hơn nữa là có thể làm chết hết cả vườn tiêu trong thời gian ngắn. Để phòng trừ bệnh này, người trồng hồ tiêu kết hợp các biện pháp dùng thuốc hóa học và các chế phẩm sinh học được sản xuất từ các chủng vi sinh vật hữu ích. Trước khi sản xuất chế phẩm sinh học từ các chủng vi sinh vật hữu ích với khối lượng lớn cần khảo sát ảnh hưởng của các thành phần môi trường như nguồn carbon, nitơ, pH môi trường và nhiệt độ nuôi cấy tới chủng vi sinh vật. Môi trường nuôi cấy là một trong các nhân tố quan trọng ảnh hưởng tới sinh trưởng, tổng hợp các hoạt chất sinh học, khả năng đối kháng với các tác nhân gây bệnh của các chủng vi sinh vật hữu ích như B. subtilis, Pseudomonas fluorescens, Trichoderma harzianum. Sự kết hợp giữa các thành phần môi trường nuôi cấy chủng vi sinh vật có thể tăng sinh khối của chủng vi sinh vật, tăng hoặc giảm hiệu lực đối kháng với các tác nhân gây bệnh. Thí nghiệm này được tiến hành với mục đích khảo sát ảnh hưởng của các yếu tố môi trường tới khả năng sinh tổng hợp Indol 3-acetic acid (IAA), một hormone kích thích sinh trưởng thực vật thuộc nhóm auxin và ức chế nấm Phytophthora capsici của chủng vi khuẩn B. sonorensis LĐ18.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưu Vi khuẩn Bacillus sonorensis LĐ18 được lưu
giữ tại phòng thí nghiệm của Bộ môn Công nghệ vi sinh, Khoa Công nghệ sinh học, Học viện Nông nghiệp Việt Nam.
2.2. Phương pháp nghiên cưu
2.2.1. Khảo sát ảnh hưởng của nhiệt độ, pH đến khả năng sản sinh IAA
Chủng vi khuẩn B. sonorensis LĐ18 được nuôi cấy trong môi trường Nutrient Broth (NB) bổ sung L-tryptopan (100 mg/l) trong 10 ngày ở điều kiện tối, ở các giá trị nhiệt độ (25°, 30°, 37º, 40°, 50) và pH (4, 5, 6, 7, 8) sau khi đã biết nhiệt độ thích hợp (Apine and Jadhav, 2011).
2.2.2. Khảo sát ảnh hưởng của nguồn carbon và nitơ đến khả năng sinh IAA
Vi khuẩn B. sonorensis LĐ18 được nuôi trong
môi trường muối khoáng cơ bản được bổ sung thêm 1% các loại đường là D- Glucose, lactose, dextrin, sucrose, D- sorbitol, mannitol, fructose, xylose vào các công thức thí nghiệm để xác định ảnh hưởng của nguồn carbon; bổ sung thêm bổ sung 0,1% nguồn nitrogen gồm nitơ hữu cơ: pepton, cao nấm men và nitơ vô cơ: NH4Cl, KNO3, NH4NO3, NaNO3, NH4Cl, (NH4)2SO4, (NH4)2SO4, (NH4)2HPO4 sau khi đã xác định được nguồn carbon thích hợp.
2.2.3. Xác định hàm lượng IAA được tổng hợpSau 10 ngày nuôi chủng vi khuẩn B.sonorensis
LĐ18 ở các điều kiện thí nghiệm, thu dịch nuôi cấy đem ly tâm ở 5.500 vòng/phút, trong 5 phút ở 4. Hút 1 ml phần dịch trong sau khi ly tâm dịch nuôi vi khuẩn cho vào các ống nghiệm và bổ sung 2 ml thuốc thử Salkowski (300 ml H2SO4 98%, 15 ml FeCl3 0,5M). Ủ hỗn hợp trên trong tối 30 phút để phản ứng xảy ra hoàn toàn, sau đó đo OD ở bước sóng λ = 530 nm.Kết quả đo OD của các chủng phân lập được thay vào phương trình đồ thị đường chuẩn với giá trị OD của các nồng độ IAA đã biết, từ đó suy ra được nồng độ IAA của các chủng vi khuẩn (Glickmann and Dessaux, 1995).
2.2.4. Khảo sát ảnh hưởng thời gian nuôi đến khả năng đối kháng nấm P. capsici của B. sonorensis LĐ18
Khả năng kháng với nấm P. capsici của vi khuẩn B. sonorensis LĐ18 được xác định bằng cách đồng nuôi cấy trên môi trường PDA (g/l: glucose 15, agar 15, dịch chiết từ 200 g khoai tây) theo mô tả của Nascimento và cộng tác viên (2015). Dùng dụng cụ đục lỗ lấy thỏi thạch (Ø = 8 mm) chứa nấm và đặt thỏi thạch vào trung tâm đĩa petri chứa môi trường PDA. Vi khuẩn B. sonorensis LĐ18 được nuôi trong môi trường NB lỏng ở nhiệt độ 300C, điều kiện lắc 200 vòng/ phút. Sau 24, 36, 48, 60 và 72 h nuôi cấy, hút 100 µl dịch nuôi vi khuẩn nhỏ vào các giếng thạch đã chuẩn bị sẵn trên đĩa Petri chứa nấm P. capsici ở trung tâm đĩa. Đĩa đối chứng chứa thỉ thạch nấm P. capsici, không có vi khuẩn. Đĩa thí nghiệm và đối chứng được nuôi ở 30ºC trong 4 ngày. Hoạt lực đối kháng được tính bằng cách đo bán kính hệ sợi nấm P. capsici trên đĩa đối chứng và đĩa thí nghiệm. Hoạt lực kháng nấm P. capsici được tính toán theo công thức sau:

92
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
RI (%) = (R – r)/R ˟ 100%Trong đó: R: bán kính của nấm P. capsici ở đĩa
đối chứng, r: bán kính của nấm P. capsici ở đĩa có vi khuẩn đối kháng
Hình 1. Sơ đồ bố trí thí nghiệm khảo sát ảnh hưởng của thời gian nuôi cấy tới khả năng kháng nấm
P. capsici của vi khuẩn B.sornorensis LĐ18 (các số từ 1-5: các giếng chứa vi khuẩn B. sonorensis ở các thời gian nuôi cấy khác nhau, 6: nấm P. capsici)
2.2.4. Môi trườngMôi trường khoáng cơ bản để khảo sát ảnh hưởng
của nguồn carbon (g/l): (NH4)2SO4 2; MgSO4.7H2O 0,2; NaH2PO4. H2O 0,5; CaCl2.2H2O 0,1; KH2PO4 0,5; H2O 1 l. Môi trường khoáng cơ bản để khảo sát ảnh hưởng của nguồn nitơ (g/l): KH2PO4 1,36; CaCl2 0,03; NaH2PO4 2,13; MgSO4.7H2O 0,2; FeSO4.7H2O 0,01; Glucose 10; H2O 1 l. Môi trường thạch NA (g/l): Pepton 5; NaCl 5; cao thịt 1; cao nấm men 2; agar 18.Môi trường nutrient broth: Pepton 5; NaCl 5; cao thịt 2; cao nấm men 3, nước cất 1000 ml.
2.2.5. Phương pháp xử lý số liệuKết quả thí nghiệm được xử lý thống kê ANOVA
bằng Excel 2013 và sử dụng hàm số STDEV.
2.3. Thời gian và địa điểm nghiên cưuNghiên cứu được thực hiện từ tháng 5 đến tháng
12/2017 tại phòng thí nghiệm Khoa Công nghệ sinh học - Học viện Nông nghiệp Việt Nam.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Ảnh hưởng của pH môi trường đến khả năng sinh IAA của B. sonorensis LĐ18
Vi khuẩn B. sonorensis LĐ18 có khả năng sinh tổng hợp IAA ở cả 5 mức pH thí nghiệm (Hình 2). Khi phân tích thống kê ANOVA, (với độ tin cậy α = 0,05, Ftn = 5,16 > Fcrit = 3,47) nhận thấy, lượng IAA do vi khuẩn này tổng hợp rất khác nhau tại các giá trị pH khác nhau, lượng IAA cao nhất (74,06 mg/l) được vi khuẩn B. sonorensis LĐ18 tổng hợp tại pH 4. Tại pH 6 và 7, hàm lượng IAA được tổng hợp tương đương nhau và giảm khi pH tăng tới 8. Patil và cộng tác viên (2011) đã công bố hàm lượng IAA cao nhất
được tổng hợp bởi chủng Acetobacter diazotrophicus phân lập từ cây mía khi chủng này được nuôi trong môi trường cơ sở với pH6, pH kiềm và pH acid đều không thích hợp cho quá trình tổng hợp IAA. Trong nghiên cứu của Jasim và cộng tác viên (2014) các chủng vi khuẩn nội sinh phân lập được từ cây hồ tiêu (Piper nigrum) tổng hợp IAA cao nhất (71,66 mg/l) khi pH môi trường là 4, khi tăng giá trị pH thì hàm lượng IAA giảm. Các chủng vi khuẩn tổng hợp IAA nhiều nhất tại pH4 có thể do môi trường acid nhẹ của các mô trong thân, rễ cây hồ tiêu, từ các phần này của cây hồ tiêu các chủng vi khuẩn được phân lập. Khamna và cộng tác viên (2010) kết luận tổng hợp IAA bởi vi sinh vật diễn ra mạnh nhất tại pH trung tính hay acid nhẹ. Như vậy, tùy thuộc vào nguồn phân lập, các chủng vi khuẩn sẽ biểu hiện tốt nhất khả năng tổng hợp IAA ở giá trị pH cụ thể.
Hình 2. Ảnh hưởng pH môi trường tới khả năng sinh IAA của vi khuẩn B. sonorensis LĐ18
3.2. Ảnh hưởng của nhiệt độ môi trường đến khả năng sinh IAA B. sonorensis LĐ18
Trong thí nghiệm này, chủng B. sonorensis LĐ18 có khả năng sinh IAA khi nuôi trong môi trường có dải nhiệt độ rộng từ 25oC đến 50oC; Nồng độ IAA được chủng vi khuẩn B. sonorensis LĐ18 tổng hợp có sự chệnh lệch khá lớn tại các nhiệt độ thí nghiệm (độ tin cậy α = 0,05; Ftn = 14,17 > Fcrit = 3,47 ). Hàm lượng IAA đạt cao nhất 59,29 µg/ml khi nuôi chủng này ở 30oC, khi nuôi chủng B. sonorensis LĐ18 ở nhiệt độ cao hơn hay thấp hơn 30oC, hàm lượng IAA giảm (Hình 3). Ambreen và cộng tác viên (2010); Monita và cộng tác viên (2014) ghi nhận nồng độ IAA cao nhất được tổng hợp bởi Bacillus sp. khi nuôi chủng này tại 37oC. Jasim và cộng tác viên (2014) đã thông báo rằng các chủng vi khuẩn nội sinh phân lập từ cây hồ tiêu (Peper nigrum) tổng hợp IAA mạnh nhất khi nhiệt độ môi trường nuôi là 37oC và 28oC, thậm chí các chủng này còn duy trì khả năng tổng hợp IAA ở 4oC. Các chủng vi khuẩn nghiên cứu có khuynh hướng tổng hợp IAA nhiều nhất ở ngưỡng nhiệt độ

93
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
phòng có thể do bản chất sống nội sinh của chúng. Một số nghiên cứu khác như nghiên cứu Apine và cộng tác viên (2011) và của Mohite (2013) cũng đã cho thấy các chủng vi khuẩn tổng IAA nhiều nhất tại 30oC. Anjhana và cộng tác viên (2017) khi khảo sát khả năng đối kháng của chủng Bacillus subtilis MS21 đã công bố kết quả, chủng vi khuẩn này sinh trưởng tốt tại nhiệt độ 35oC. Một số tác giả khác lại có kết luận khác về nhiệt độ sinh trưởng tối ưu của vi khuẩn Bacilus sp. như Okanlawon và cộng tác viên (2010) đã công bố B. cereus sinh trưởng tốt nhất tại 37oC, trong khi đó Johnson và cộng tác viên (1974) lại thông báo rằng B.cereus sinh trưởng tốt nhất trong ngưỡng nhiệt độ 30oC - 37oC và một số chủng có thể sinh trưởng ở nhiệt độ 4,5oC hay cao tới 55oC.
Hình 3. Ảnh hưởng nhiệt độ môi trường nuôi tới khả năng sinh IAA của vi khuẩn B. sonorensis LĐ18
3.3. Ảnh hưởng của nguồn carbon và nitơ đến khả năng sinh IAA của B. sonorensis LĐ18
Các chủng vi khuẩn khác nhau sẽ có nhu cầu về chất dinh dưỡng khác nhau để đáp ứng nhu cầu sinh trưởng, phát triển và tổng hợp các hợp chất trao đổi chất của chúng. Thí nghiệm này được tiến hành với mục đích khảo sát ảnh hưởng của các nguồn carbon và nitơ khác với nguồn carbon và nitơ trong môi trường NB tới khả năng sinh tổng hợp IAA của chủng B. sonorensis LĐ18, từ đó có thể chọn được nguồn carbon và nitơ thích hợp trong duy trì khả năng tổng hợp IAA của chủng này.
Chủng B. sonorensis LĐ18 được nuôi trong các môi trường đã nêu tại mục 2.2.4, kết quả thí nghiệm thu được được phân tích ANOVA. Kết quả cho thấy, với độ tin cậy (độ tin cậy α = 0,05; Ftn
= 3,33 > Fcrit = 2,65), vi khuẩn B. sonorensis LĐ18 sử dụng D-sobitol hiệu quả nhất, lượng IAA được tổng hợp là 22,67 µg/ml, tiếp theo là sucrose với lượng IAA được tổng hợp là 7,39 µg/ml. các nguồn đường khác đều
không làm tăng hoặc tăng không nhiều nồng độ IAA trong môi trường (Hình 4). Namita Bhutani và cộng tác viên (2018) đã báo cáo rằng các chủng vi khuẩn nội sinh Bacillus spp. phân lập từ Vigna radiata tổng hợp IAA với hàm lượng rất khác nhau khi sử dụng các nguồn carbon là glucose, mannitol và sucrose. Chủng Bacillus MBN3 tổng hợp IAA nhiều nhất khi nguồn carbon trong môi trường nuôi là mannitol, với chủng Bacillus MJHN1 thì nguồn carbon thích hợp nhất là sucrose, trong khi đó chủng Bacillus MJHN10 ưa thích nguồn carbon là đường glucose. Chủng vi khuẩn nội sinh Acetobacter diazotrophicus L1 sinh tổng hợp IAA nhiều nhất khi trong môi trường nuôi cấy có sucrose.
Hình 4. Ảnh hưởng của nguồn carbon tới khả năng sinh IAA của vi khuẩn B. sonorensis LĐ18.
Chủng vi khuẩn B. sonorensis LĐ18 được nuôi trong môi trường khoáng cơ bản với nguồn nitrogen là NH4Cl, KNO3, NH4NO3, NaNO3, NH4Cl, (NH4)2SO4, (NH4)2SO4, (NH4)2HPO4, cao nấm men, pepton. Sau 10 ngày nuôi tiến hành kiểm tra hàm lượng IAA có trong dịch nuôi.
Hình 5. Ảnh hưởng nguồn nitơ tới khả năng sinh IAA của vi khuẩn B. sonorensis LĐ18
Ghi chú: CNM: cao nấm men.
Suc

94
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Sau khi phân tích các kết quả thí nghiệm bằng chương trình ANOVA, với độ tin cậy α = 0,05, ghi nhận được: nguồn nitơ vô cơ không phải là nguồn nitơ thích hợp với chủng vi khuẩn B. sonorensis LĐ18 vì hàm lượng IAA được vi khuẩn này tổng hợp rất ít (5,03 µg/ml khi nguồn nitơ là NH4Cl), thậm chí không có (Hình 5). Ngược lại, nguồn nitơ hữu cơ tác động tích cực tới quá trình tổng hợp IAA của chủng vi khuẩn B. sonorensis LĐ18. Khi trong môi trường có cao nấm men, B. sonorensis LĐ18 đã tổng hợp được 33,3 µg/ml IAA (Hình 5). Trong thí nghiệm của Namita và cộng tác viên (2018), nguồn nitơ thích hợp cho các chủng vi khuẩn Baillus spp. tổng hợp IAA là cao nấm men. Patil và cộng tác viên (2011) thông báo chủng vi khuẩn nội sinh Acetobacter diazotrophicus L1 tổng hợp IAA nhiều nhất khi trong môi trường nuôi có cao nấm men và NH4Cl.
3.4. Ảnh hưởng của thời gian nuôi cấy đến khả năng đối kháng nấm P. capsici của vi khuẩn B. sonorensis LĐ18
Sau 24 và 36 h đồng nuôi cấy với nấm P. capsici trên môi trường PDA, vi khuẩn B. sonorensis LĐ18 chưa biểu hiện khả năng kháng, có thể do chủng vi khuẩn cần có thời gian thích nghi với môi trường, tốc độ sinh trưởng và tổng hợp các hợp chất thứ cấp diễn ra chậm.
Hình 5. Ảnh hưởng của thời gian nuôi tới hoạt lực đối kháng nấm P. capsici của vi khuẩn B. sonorensis LĐ18
Kết thúc 48 h nuôi, vi khuẩn B.sonorensis LĐ18 đã thể hiện khả năng đối kháng, hoạt lực đối kháng đạt khoảng 50%. Hiệu lực đối kháng mạnh nhất (66,7%) của chủng B. sonorensis LĐ18 đạt được sau 3 ngày nuôi cấy (Hình 5).
IV. KẾT LUẬNChủng vi khuẩn Bacilus soronensis LĐ18 có khả
năng tổng hợp IAA ở dải pH từ 4 đến 8, IAA được tổng hợp nhiều nhất tại pH 4 và nhiệt độ 30oC. Nguồn carbon thích hợp cho chủng B. sonorensis LĐ18 tổng hợp IAA là sorbitol, sucrose. B. sonorensis LĐ18 tổng hợp IAA mạnh khi nguồn nitơ trong môi trường nuôi là cao nấm men. Hoạt lực đối kháng với nấm P. capsici của chủng vi khuẩn B. sonorensis LĐ18 bắt đầu thể hiện sau 48 giờ nuôi cấy.
TÀI LIỆU THAM KHẢOAmbreen, A. and Hasnain, S., 2010. Auxin producing
Bacillus sp.: Auxin quantification and effect on the growth of Solanum tuberosum. Pure Appl. Chem., 82 (1): 313-319.
Anjhana, V.R. and Sasikala, S.L., 2017. Isolation, screening and growth optimization of antagonistic Bacillus subtilis MS21 from Thengapattanam estuary against plant fungal pathogens. Int. J. Adv. Res. Biol. Sci., 4 (12): 15-26.
Apine, O.A. and Jadhav, J.P., 2011. Optimization of medium for indole-3-acetic acid production using Pantoea agglomerans strain PVM. J. of Applied Microbiology, 110: 1235-1244.
Glickmann, E. and Dessaux, Y., 1995. A critical examination of the specificity of the salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl Environ Microbiol, 61 (2): 793-796.
Jasim, B., John, C. J., Shimil, V., Jyothis, M. and Radhakrishnan, R.K., 2014. Studies on the factors modulating indole-3-acetic acid production in endophytic bacterial isolates from Piper nigrum and molecular analysis of ipdc gene. J Appl Microbiol. 117 (3): 786-799.
Johnson, U. and Snygg, B.G., 1974. Lipase production and activity as a function of incubation, time, pH and temperature of four lipolytic organisms. J. Appl. Bacteriol., 37: 571-581.
Khamna, S., Yokota, A., Peberdy, J.F. and Lumyong, S., 2010. Indole-3-acetic acid production by Streptomyces sp. isolated from some Thai medicinal plant rhizosphere soils. Eur Asia J. Bio Sci., 4: 23-32.
Mohite, B., 2013. Isolation and characterization of indole acetic acid (IAA) producing bacteria from rhizospheric soil and its effect on plant growth. J Soil Sci Plant Nutr, 13, 638-649.

95
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Monita, V.P. and Patel, R.K., 2014. Indole-3-acetic aicd (IAA) production by Endophytic bacteria isolated from saline dessert, the little runn of Kutch. An online international Journal, available at http://www.cibtech.org/cjm.htm 2014 Vol. 3 (2) April-June, pp. 17-28/Patel and Patel.
Namita, B., Maheshwari, R., Negi, M. and Suneja, P., 2018. Optimization of IAA production by endophytic Bacillus spp. from Vigna radiate for their potential use as plant growth promoters. Israel Journal of Plant Sciences, http://booksandjournals.brillonline.com/content/journals/10.1163/22238980-00001025.
Nascimento, S.B., Lima, A.M., Borges, B.N. and de Souza, C.R.B., 2015. Endophytic bacteria from Piper tuberculatum Jacq.: isolation, molecular
characterization, and in vitro screening for the control of Fusarium solani f. sp piperis, the causal agent of root rot disease in black pepper (Piper nigrum L.). Genetics and Molecular Research, 14 (3): 7567-7577.
Okanlawon, B.M., Ogunbanwo, S.T. and Okunlola, A.O., 2010. Growth of Bacillus cereus isolated from some traditional condiments under different regimens. Afr. J. Biotechnol., 8 (14): 2129-2135.
Patil, N.B., Gajbhiye, M., Sangita, S. A., Aparna, B., Balasaheb, P., 2011. Optimization of Indole 3-acetic acid (IAA) production by Acetobacter diazotrophicus L1 isolated from Sugarcane. Int. J. of Env. Sci., 2 (1): 307-314.
Effects of culture condition on IAA biosynthesis of Bacillus sonorensis LD18
Nguyen Van Giang, Tran Thi Dao, Tran Thi Thuy Ha, Nguyen Thu Trang
AbstractThis experiment was carried out with the aim to evaluate the influence of temperature, culture medium, pH and carbon and nitrogen sources on the ability of IAA synthesis of B.sonorensis LD18. B. sonorensis LĐ18 was able to synthesize IAA at 4 - 8 pH values, exhibited the highest antifungal activity at pH 6. Temperature suitable for IAA synthesis of B. sonorensis LD18 was at 300C; the strongest antifungal activity of this strain was exhibited at 370C. Carbon sources suitable for the IAA synthesis were sucrose and sorbitol; the most potent antagonist of P.capsici when the carbon source in the medium was sorbitol and dextrose. B.sonorensis LD18 synthesized IAA; the maximum when the nitrogen source in culture medium was yeast extract; the most potent antagonist when nitrogen sources were peptone, KNO3 and NH4Cl.Keywords: Bacillus sp., antifungal activity, Phytophthora capsici
Ngày nhận bài: 25/7/2018Ngày phản biện: 6/8/2018
Người phản biện: PGS. TS. Hồ Phú HàNgày duyệt đăng: 15/10/2018
1 Viện Khoa học Nông nghiệp Việt Nam; 2 Viện Khoa học Kỹ thuật Nông Lâm nghiệp Tây Nguyên
NGHIÊN CỨU ẢNH HƯỞNG CỦA TUỔI CÂY GIỐNG VÀ PHÂN BÓN HỮU CƠ ĐẾN CÂY CÀ PHÊ VỐI TRỒNG TÁI CANH TẠI TÂY NGUYÊN
Tạ Hồng Lĩnh1, Nguyễn Văn Tuất1, Bùi Quang Đãng1, Nguyễn Thị Thanh Mai2, Trương Hồng2
TÓM TẮTNghiên cứu về ảnh hưởng của tuổi cây giống và các công thức bón phân hữu cơ khác nhau trên cây cà phê vối
trồng tái canh được thực hiện tại huyện Krông Pắk, tỉnh Đắk Lắk từ năm 2015 - 2017. Kết quả nghiên cứu đã chỉ ra rằng sử dụng giống cây cà phê ghép 18 tháng tuổi trong giai đoạn kiến thiết cơ bản kết hợp với quy trình kỹ thuật canh tác hợp lý đảm bảo cây sinh trưởng và phát triển đồng đều, năng suất đạt 2,31 tấn nhân/ha sau 30 tháng trồng. Kết hợp giữa cây giống ghép 18 tháng tuổi và bón phân hữu cơ 26 kg/hố (bón liên tục 3 năm) đã làm giảm mật độ tuyến trùng và nấm bệnh trong đất từ đó giảm tỷ lệ bệnh vàng lá, thối rễ, năng suất tối đa có thể đạt 2,79 tấn nhân/ha sau 30 tháng trồng.
Từ khóa: Cây giống ghép, nấm, tuyến trùng, bệnh vàng lá, thối rễ

96
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
I. ĐẶT VẤN ĐỀHiện nay, việc tái canh cây cà phê ở Tây Nguyên
nói chung, đặc biệt là việc trồng ngay trên đất cà phê già cỗi đang là vấn đề nan giải và vô cùng khó khăn cho người trồng cà phê cũng như đối với ngành cà phê Việt Nam khi mà diện tích cà phê cần tái canh ngày càng gia tăng. Theo thống kê của Cục Trồng trọt (2016), toàn vùng Tây Nguyên có diện tích cà phê là 551.670 ha và theo đề án tái canh các tỉnh Tây Nguyên từ 2014 - 2020, diện tích cà phê cần tái canh giai đoạn này khoảng 120.000 ha, trong đó diện tích tái canh 90.000 ha và diện tích ghép cải tạo 30.000 ha.
Trong thực tiễn, các diện tích cà phê trồng lại trên nền đất cũ thường bị chết, nguyên nhân chủ yếu là do tuyến trùng trong đất tấn công và nấm bệnh xâm nhập làm thối nhanh rễ cà phê hoặc cây sinh trưởng kém do vấn đề quản lý cây giống và một số biện pháp kỹ thuật chưa tuân thủ nghiêm ngặt quy trình tái canh cà phê do Bộ Nông nghiệp và Phát triển nông thôn ban hành năm 2013 và 2016 dân đến việc tái canh tác cà phê thường không có hiệu quả, tỷ lệ thành công thấp (Lê Ngọc Báu và Chế Thị Đa, 2012). Đây là vấn đề đang tồn tại cần có các biện pháp giải quyết để ngành cà phê phát triển ổn định, bền vững.
Do vậy việc nghiên cứu xác định tuổi cây giống và phân bón hữu cơ thích hợp cho cây cà phê tái canh tại Tây Nguyên là một trong những giải pháp mang lại hiệu quả cao trong việc hạn chế bệnh vàng lá, thối rễ phục vụ yêu cầu phát triển cà phê bền vững của Việt Nam.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưu- Cây cà phê vối thực sinh trồng từ hạt lai đa dòng
TRS1 6 tháng tuổi và 18 tháng tuổi.- Cây cà phê ghép 6 tháng tuổi và 18 tháng tuổi.- Phân hữu cơ (phân chuồng).- Cây cà phê thực sinh và cây ghép được gieo
ươm tại Viện KHKT Nông Lâm nghiệp Tây Nguyên.
2.2. Phương pháp nghiên cưu
2.2.1. Phương pháp bố trí thí nghiệm- Thí nghiệm về tuổi cây giống: Thí nghiệm được bố trí theo kiểu khối đầy đủ
ngâu nhiên gồm 4 công thức, 3 lần lặp, mỗi ô cơ sở 30 cây, tổng số cây theo dõi 360 cây, quy mô thí nghiệm 0,5 ha.
Trồng và chăm sóc theo quy trình tái canh năm 2013.
Các công thức (CT) thí nghiệm: CT1: Trồng cây
cà phê ghép 6 tháng tuổi (bầu nhỏ, kích thước bầu 12 ˟ 22 cm); CT2: Trồng cây cà phê ghép 18 tháng tuổi (bầu to, kích thước bầu 20 ˟ 30 cm); CT3: Trồng cây cà phê thực sinh TRS1 6 tháng tuổi (bầu nhỏ, kích thước bầu 12 ˟ 22 cm); CT4: Trồng cây thực sinh TRS1 18 tháng tuổi (bầu to, kích thước bầu 20 ˟ 30 cm).
- Thí nghiệm về bón phân:Thí nghiệm được bố trí theo kiểu lô chính, lô phụ
gồm 2 chu kỳ bón và 3 mức phân hữu cơ (10 kg, 18 kg, 26 kg) với 3 lần lặp lại, gồm 18 ô cơ sở, mỗi ô cơ sở 30 cây, tổng số cây thí nghiệm là 540 cây, tổng diện tích 0,5 ha. Cây cà phê ghép 18 tháng tuổi.
Các công thức bón phân: C1M1: Bón liên tục 3 năm KTCB, 10 kg phân chuồng; C1M2: Bón liên tục 3 năm KTCB, 18 kg phân chuồng; C1M3: Bón liên tục 3 năm KTCB, 26 kg phân chuồng; C2M1: 2 năm bón 1 lần, 10 kg phân chuồng; C2M2: 2 năm bón 1 lần, 18 kg phân chuồng; C2M3: 2 năm bón 1 lần, 26 kg phân chuồng.
2.2.2. Phương pháp phân tích tuyến trùng và nấm - Phân tích tuyến trùng theo phương pháp lọc
(Maceration - sieving method) và ly trích tuyến trùng từ đất sử dụng phễu Baermann (Baermann funnel techniques) (Hooper, 1986).
- Phân lập các loài nấm trong đất theo phương pháp pha loăng đất (soil dilution plate technique) của Lester W. Burgess và cộng tác viên (2009).
2.2.3. Chỉ tiêu theo dõi - Thành phần, mật độ nấm và tuyến trùng trong
đất (trước thí nghiệm), trong đất và rễ (sau thí nghiệm) 12, 24, 30 tháng trồng.
- Tình hình sinh trưởng, phát triển, tỷ lệ cây vàng lá, cây chết sau 6 tháng, 12 tháng, 18 tháng, 24 tháng, 30 tháng trồng.
- Năng suất cà phê sau 30 tháng trồng.- Tỷ lệ cây vàng lá (%), chỉ số vàng lá (%).- Tỷ lệ vàng lá được tính theo công thức: TLVL
(%) = (A/B) ˟ 100Trong đó, TLVL: tỷ lệ cây bị vàng lá; A: tổng số cây
bị vàng lá; B: tổng số cây điều tra.
2.2.4. Phương pháp xử lý số liệuSố liệu được xử lý theo chương trình Statistic 8.2,
Excel 2010 và SPSS 16.0. Các giá trị trung bình của các nghiệm thức được so sánh bằng trắc nghiệm F, t, Duncan ở mức xác suất p ≤ 95%. Các giá trị a, b... được ghi kế bên các giá trị trung bình, ký hiệu chữ giống nhau thì có giá trị giống nhau về ý nghĩa thống kê.
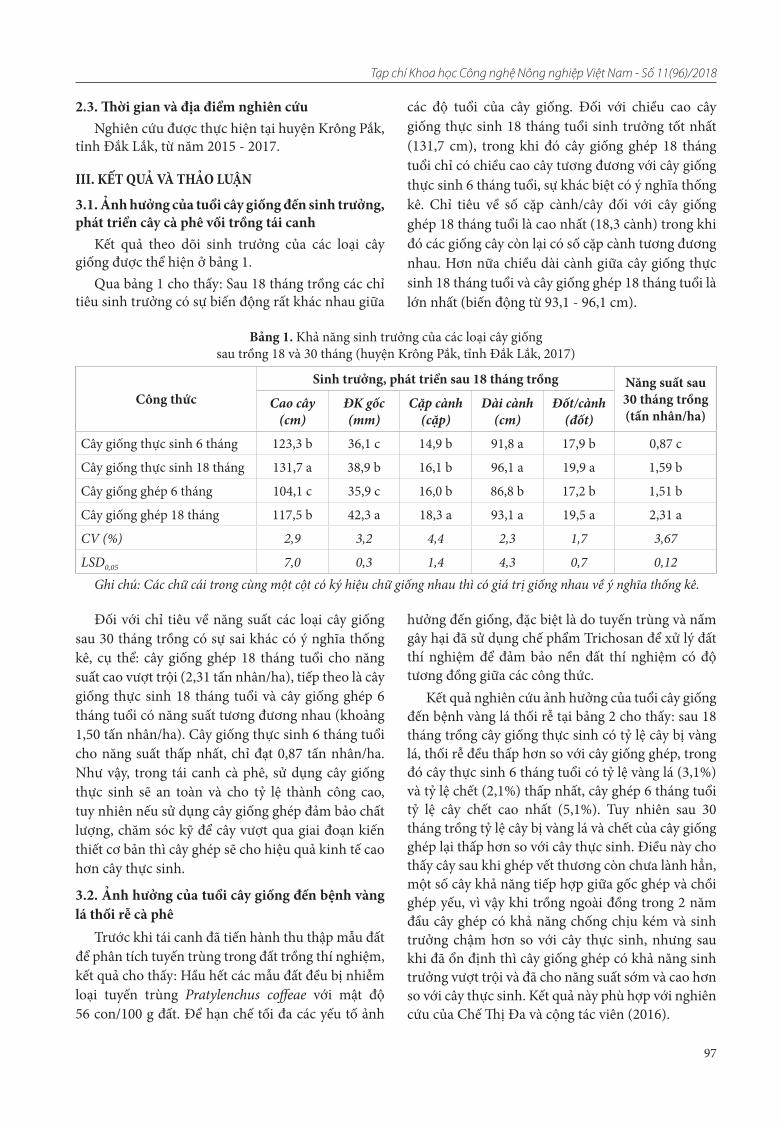
97
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
2.3. Thời gian và địa điểm nghiên cưu Nghiên cứu được thực hiện tại huyện Krông Pắk,
tỉnh Đắk Lắk, từ năm 2015 - 2017.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Ảnh hưởng của tuổi cây giống đến sinh trưởng, phát triển cây cà phê vối trồng tái canh
Kết quả theo dõi sinh trưởng của các loại cây giống được thể hiện ở bảng 1.
Qua bảng 1 cho thấy: Sau 18 tháng trồng các chỉ tiêu sinh trưởng có sự biến động rất khác nhau giữa
các độ tuổi của cây giống. Đối với chiều cao cây giống thực sinh 18 tháng tuổi sinh trưởng tốt nhất (131,7 cm), trong khi đó cây giống ghép 18 tháng tuổi chỉ có chiều cao cây tương đương với cây giống thực sinh 6 tháng tuổi, sự khác biệt có ý nghĩa thống kê. Chỉ tiêu về số cặp cành/cây đối với cây giống ghép 18 tháng tuổi là cao nhất (18,3 cành) trong khi đó các giống cây còn lại có số cặp cành tương đương nhau. Hơn nữa chiều dài cành giữa cây giống thực sinh 18 tháng tuổi và cây giống ghép 18 tháng tuổi là lớn nhất (biến động từ 93,1 - 96,1 cm).
Đối với chỉ tiêu về năng suất các loại cây giống sau 30 tháng trồng có sự sai khác có ý nghĩa thống kê, cụ thể: cây giống ghép 18 tháng tuổi cho năng suất cao vượt trội (2,31 tấn nhân/ha), tiếp theo là cây giống thực sinh 18 tháng tuổi và cây giống ghép 6 tháng tuổi có năng suất tương đương nhau (khoảng 1,50 tấn nhân/ha). Cây giống thực sinh 6 tháng tuổi cho năng suất thấp nhất, chỉ đạt 0,87 tấn nhân/ha. Như vậy, trong tái canh cà phê, sử dụng cây giống thực sinh sẽ an toàn và cho tỷ lệ thành công cao, tuy nhiên nếu sử dụng cây giống ghép đảm bảo chất lượng, chăm sóc kỹ để cây vượt qua giai đoạn kiến thiết cơ bản thì cây ghép sẽ cho hiệu quả kinh tế cao hơn cây thực sinh.
3.2. Ảnh hưởng của tuổi cây giống đến bệnh vàng lá thối rễ cà phê
Trước khi tái canh đã tiến hành thu thập mâu đất để phân tích tuyến trùng trong đất trồng thí nghiệm, kết quả cho thấy: Hầu hết các mâu đất đều bị nhiễm loại tuyến trùng Pratylenchus coffeae với mật độ 56 con/100 g đất. Để hạn chế tối đa các yếu tố ảnh
hưởng đến giống, đặc biệt là do tuyến trùng và nấm gây hại đã sử dụng chế phẩm Trichosan để xử lý đất thí nghiệm để đảm bảo nền đất thí nghiệm có độ tương đồng giữa các công thức.
Kết quả nghiên cứu ảnh hưởng của tuổi cây giống đến bệnh vàng lá thối rễ tại bảng 2 cho thấy: sau 18 tháng trồng cây giống thực sinh có tỷ lệ cây bị vàng lá, thối rễ đều thấp hơn so với cây giống ghép, trong đó cây thực sinh 6 tháng tuổi có tỷ lệ vàng lá (3,1%) và tỷ lệ chết (2,1%) thấp nhất, cây ghép 6 tháng tuổi tỷ lệ cây chết cao nhất (5,1%). Tuy nhiên sau 30 tháng trồng tỷ lệ cây bị vàng lá và chết của cây giống ghép lại thấp hơn so với cây thực sinh. Điều này cho thấy cây sau khi ghép vết thương còn chưa lành hẳn, một số cây khả năng tiếp hợp giữa gốc ghép và chồi ghép yếu, vì vậy khi trồng ngoài đồng trong 2 năm đầu cây ghép có khả năng chống chịu kém và sinh trưởng chậm hơn so với cây thực sinh, nhưng sau khi đã ổn định thì cây giống ghép có khả năng sinh trưởng vượt trội và đã cho năng suất sớm và cao hơn so với cây thực sinh. Kết quả này phù hợp với nghiên cứu của Chế Thị Đa và cộng tác viên (2016).
Bảng 1. Khả năng sinh trưởng của các loại cây giống sau trồng 18 và 30 tháng (huyện Krông Pắk, tỉnh Đắk Lắk, 2017)
Ghi chú: Các chữ cái trong cùng một cột có ký hiệu chữ giống nhau thì có giá trị giống nhau về ý nghĩa thống kê.
Công thưcSinh trưởng, phát triển sau 18 tháng trồng Năng suất sau
30 tháng trồng (tấn nhân/ha)
Cao cây (cm)
ĐK gốc (mm)
Cặp cành (cặp)
Dài cành (cm)
Đốt/cành (đốt)
Cây giống thực sinh 6 tháng 123,3 b 36,1 c 14,9 b 91,8 a 17,9 b 0,87 c
Cây giống thực sinh 18 tháng 131,7 a 38,9 b 16,1 b 96,1 a 19,9 a 1,59 b
Cây giống ghép 6 tháng 104,1 c 35,9 c 16,0 b 86,8 b 17,2 b 1,51 b
Cây giống ghép 18 tháng 117,5 b 42,3 a 18,3 a 93,1 a 19,5 a 2,31 a
CV (%) 2,9 3,2 4,4 2,3 1,7 3,67
LSD0,05 7,0 0,3 1,4 4,3 0,7 0,12
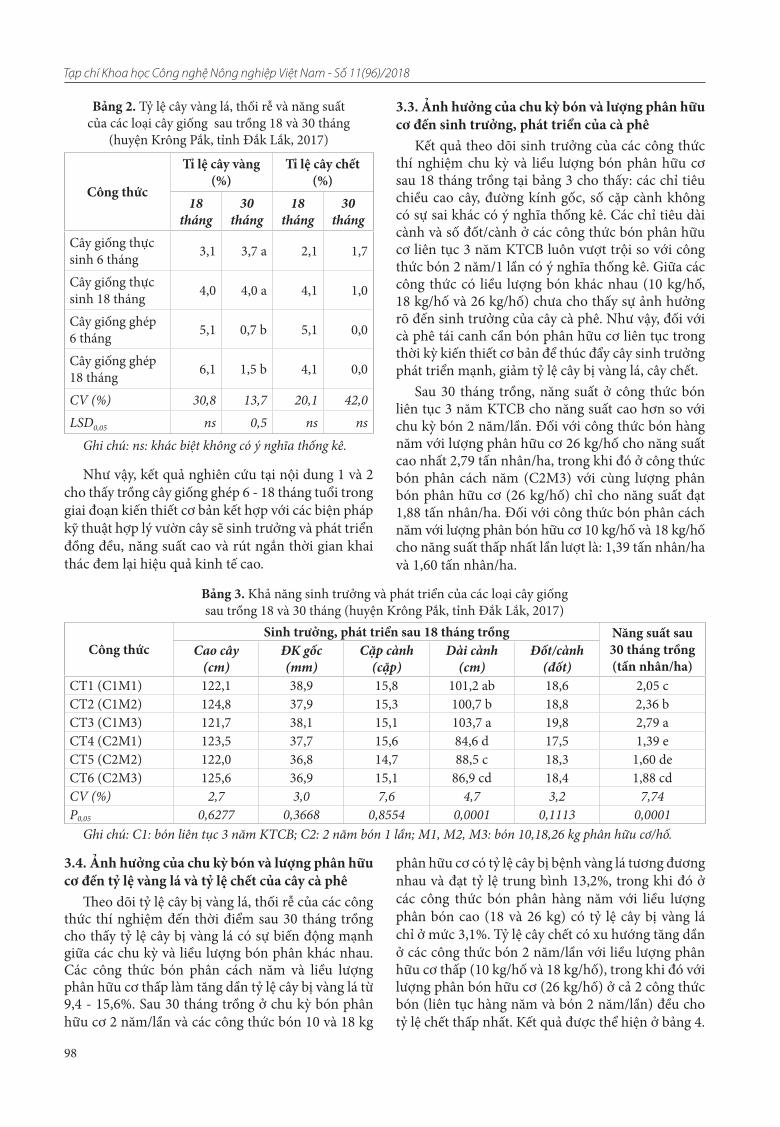
98
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Bảng 2. Tỷ lệ cây vàng lá, thối rễ và năng suất của các loại cây giống sau trồng 18 và 30 tháng
(huyện Krông Pắk, tỉnh Đắk Lắk, 2017)
Ghi chú: ns: khác biệt không có ý nghĩa thống kê.
Như vậy, kết quả nghiên cứu tại nội dung 1 và 2 cho thấy trồng cây giống ghép 6 - 18 tháng tuổi trong giai đoạn kiến thiết cơ bản kết hợp với các biện pháp kỹ thuật hợp lý vườn cây sẽ sinh trưởng và phát triển đồng đều, năng suất cao và rút ngắn thời gian khai thác đem lại hiệu quả kinh tế cao.
3.3. Ảnh hưởng của chu kỳ bón và lượng phân hữu cơ đến sinh trưởng, phát triển của cà phê
Kết quả theo dõi sinh trưởng của các công thức thí nghiệm chu kỳ và liều lượng bón phân hữu cơ sau 18 tháng trồng tại bảng 3 cho thấy: các chỉ tiêu chiều cao cây, đường kính gốc, số cặp cành không có sự sai khác có ý nghĩa thống kê. Các chỉ tiêu dài cành và số đốt/cành ở các công thức bón phân hữu cơ liên tục 3 năm KTCB luôn vượt trội so với công thức bón 2 năm/1 lần có ý nghĩa thống kê. Giữa các công thức có liều lượng bón khác nhau (10 kg/hố, 18 kg/hố và 26 kg/hố) chưa cho thấy sự ảnh hưởng rõ đến sinh trưởng của cây cà phê. Như vậy, đối với cà phê tái canh cần bón phân hữu cơ liên tục trong thời kỳ kiến thiết cơ bản để thúc đẩy cây sinh trưởng phát triển mạnh, giảm tỷ lệ cây bị vàng lá, cây chết.
Sau 30 tháng trồng, năng suất ở công thức bón liên tục 3 năm KTCB cho năng suất cao hơn so với chu kỳ bón 2 năm/lần. Đối với công thức bón hàng năm với lượng phân hữu cơ 26 kg/hố cho năng suất cao nhất 2,79 tấn nhân/ha, trong khi đó ở công thức bón phân cách năm (C2M3) với cùng lượng phân bón phân hữu cơ (26 kg/hố) chỉ cho năng suất đạt 1,88 tấn nhân/ha. Đối với công thức bón phân cách năm với lượng phân bón hữu cơ 10 kg/hố và 18 kg/hốcho năng suất thấp nhất lần lượt là: 1,39 tấn nhân/ha và 1,60 tấn nhân/ha.
Công thưc
Tỉ lệ cây vàng (%)
Tỉ lệ cây chết (%)
18 tháng
30 tháng
18 tháng
30 tháng
Cây giống thực sinh 6 tháng 3,1 3,7 a 2,1 1,7
Cây giống thực sinh 18 tháng 4,0 4,0 a 4,1 1,0
Cây giống ghép 6 tháng 5,1 0,7 b 5,1 0,0
Cây giống ghép 18 tháng 6,1 1,5 b 4,1 0,0
CV (%) 30,8 13,7 20,1 42,0LSD0,05 ns 0,5 ns ns
Bảng 3. Khả năng sinh trưởng và phát triển của các loại cây giống sau trồng 18 và 30 tháng (huyện Krông Pắk, tỉnh Đắk Lắk, 2017)
Ghi chú: C1: bón liên tục 3 năm KTCB; C2: 2 năm bón 1 lần; M1, M2, M3: bón 10,18,26 kg phân hữu cơ/hố.
Công thưcSinh trưởng, phát triển sau 18 tháng trồng Năng suất sau
30 tháng trồng (tấn nhân/ha)
Cao cây (cm)
ĐK gốc(mm)
Cặp cành(cặp)
Dài cành (cm)
Đốt/cành (đốt)
CT1 (C1M1) 122,1 38,9 15,8 101,2 ab 18,6 2,05 cCT2 (C1M2) 124,8 37,9 15,3 100,7 b 18,8 2,36 bCT3 (C1M3) 121,7 38,1 15,1 103,7 a 19,8 2,79 aCT4 (C2M1) 123,5 37,7 15,6 84,6 d 17,5 1,39 eCT5 (C2M2) 122,0 36,8 14,7 88,5 c 18,3 1,60 deCT6 (C2M3) 125,6 36,9 15,1 86,9 cd 18,4 1,88 cdCV (%) 2,7 3,0 7,6 4,7 3,2 7,74P0,05 0,6277 0,3668 0,8554 0,0001 0,1113 0,0001
3.4. Ảnh hưởng của chu kỳ bón và lượng phân hữu cơ đến tỷ lệ vàng lá và tỷ lệ chết của cây cà phê
Theo dõi tỷ lệ cây bị vàng lá, thối rễ của các công thức thí nghiệm đến thời điểm sau 30 tháng trồng cho thấy tỷ lệ cây bị vàng lá có sự biến động mạnh giữa các chu kỳ và liều lượng bón phân khác nhau. Các công thức bón phân cách năm và liều lượng phân hữu cơ thấp làm tăng dần tỷ lệ cây bị vàng lá từ 9,4 - 15,6%. Sau 30 tháng trồng ở chu kỳ bón phân hữu cơ 2 năm/lần và các công thức bón 10 và 18 kg
phân hữu cơ có tỷ lệ cây bị bệnh vàng lá tương đương nhau và đạt tỷ lệ trung bình 13,2%, trong khi đó ở các công thức bón phân hàng năm với liều lượng phân bón cao (18 và 26 kg) có tỷ lệ cây bị vàng lá chỉ ở mức 3,1%. Tỷ lệ cây chết có xu hướng tăng dần ở các công thức bón 2 năm/lần với liều lượng phân hữu cơ thấp (10 kg/hố và 18 kg/hố), trong khi đó với lượng phân bón hữu cơ (26 kg/hố) ở cả 2 công thức bón (liên tục hàng năm và bón 2 năm/lần) đều cho tỷ lệ chết thấp nhất. Kết quả được thể hiện ở bảng 4.
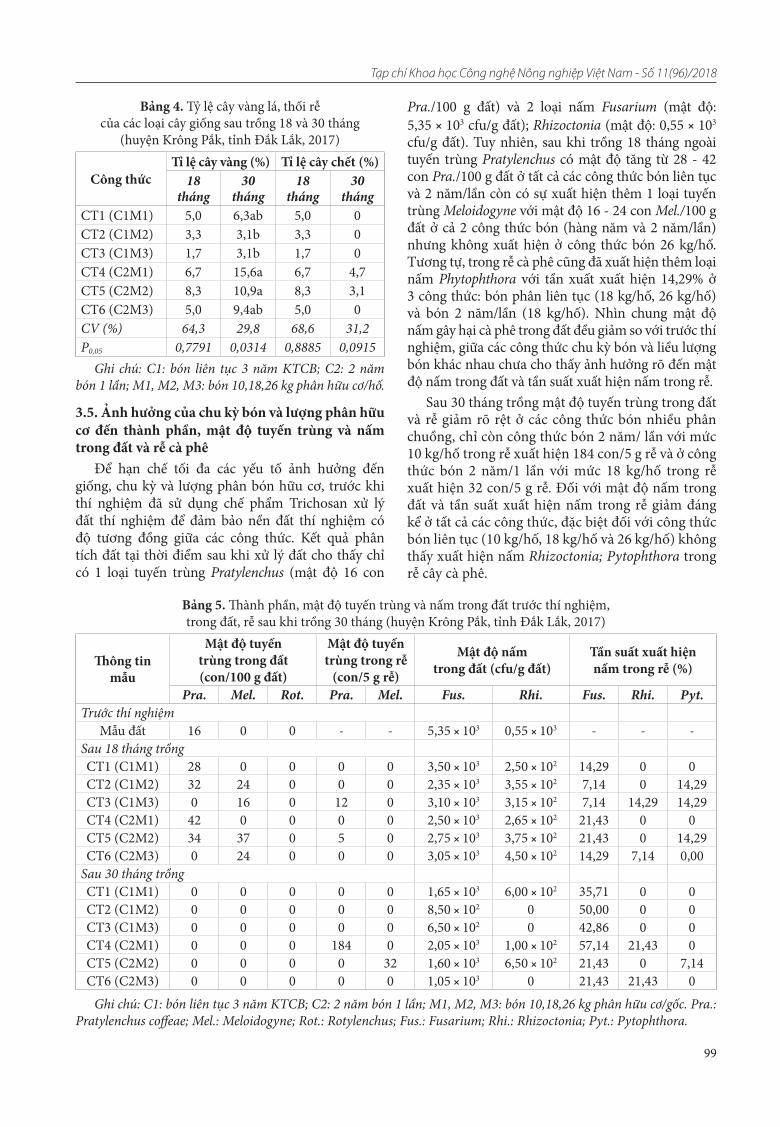
99
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Thông tin mẫu
Mật độ tuyến trùng trong đất (con/100 g đất)
Mật độ tuyến trùng trong rễ
(con/5 g rễ)
Mật độ nấm trong đất (cfu/g đất)
Tần suất xuất hiện nấm trong rễ (%)
Pra. Mel. Rot. Pra. Mel. Fus. Rhi. Fus. Rhi. Pyt.Trước thí nghiệm
Mâu đất 16 0 0 - - 5,35 ˟ 103 0,55 ˟ 103 - - -Sau 18 tháng trồngCT1 (C1M1) 28 0 0 0 0 3,50 ˟ 103 2,50 ˟ 102 14,29 0 0CT2 (C1M2) 32 24 0 0 0 2,35 ˟ 103 3,55 ˟ 102 7,14 0 14,29CT3 (C1M3) 0 16 0 12 0 3,10 ˟ 103 3,15 ˟ 102 7,14 14,29 14,29CT4 (C2M1) 42 0 0 0 0 2,50 ˟ 103 2,65 ˟ 102 21,43 0 0CT5 (C2M2) 34 37 0 5 0 2,75 ˟ 103 3,75 ˟ 102 21,43 0 14,29CT6 (C2M3) 0 24 0 0 0 3,05 ˟ 103 4,50 ˟ 102 14,29 7,14 0,00
Sau 30 tháng trồngCT1 (C1M1) 0 0 0 0 0 1,65 ˟ 103 6,00 ˟ 102 35,71 0 0CT2 (C1M2) 0 0 0 0 0 8,50 ˟ 102 0 50,00 0 0CT3 (C1M3) 0 0 0 0 0 6,50 ˟ 102 0 42,86 0 0CT4 (C2M1) 0 0 0 184 0 2,05 ˟ 103 1,00 ˟ 102 57,14 21,43 0CT5 (C2M2) 0 0 0 0 32 1,60 ˟ 103 6,50 ˟ 102 21,43 0 7,14CT6 (C2M3) 0 0 0 0 0 1,05 ˟ 103 0 21,43 21,43 0
Bảng 4. Tỷ lệ cây vàng lá, thối rễ của các loại cây giống sau trồng 18 và 30 tháng
(huyện Krông Pắk, tỉnh Đắk Lắk, 2017)
Ghi chú: C1: bón liên tục 3 năm KTCB; C2: 2 năm bón 1 lần; M1, M2, M3: bón 10,18,26 kg phân hữu cơ/hố.
3.5. Ảnh hưởng của chu kỳ bón và lượng phân hữu cơ đến thành phần, mật độ tuyến trùng và nấm trong đất và rễ cà phê
Để hạn chế tối đa các yếu tố ảnh hưởng đến giống, chu kỳ và lượng phân bón hữu cơ, trước khi thí nghiệm đã sử dụng chế phẩm Trichosan xử lý đất thí nghiệm để đảm bảo nền đất thí nghiệm có độ tương đồng giữa các công thức. Kết quả phân tích đất tại thời điểm sau khi xử lý đất cho thấy chỉ có 1 loại tuyến trùng Pratylenchus (mật độ 16 con
Pra./100 g đất) và 2 loại nấm Fusarium (mật độ: 5,35 ˟ 103 cfu/g đất); Rhizoctonia (mật độ: 0,55 ˟ 103 cfu/g đất). Tuy nhiên, sau khi trồng 18 tháng ngoài tuyến trùng Pratylenchus có mật độ tăng từ 28 - 42 con Pra./100 g đất ở tất cả các công thức bón liên tục và 2 năm/lần còn có sự xuất hiện thêm 1 loại tuyến trùng Meloidogyne với mật độ 16 - 24 con Mel./100 gđất ở cả 2 công thức bón (hàng năm và 2 năm/lần) nhưng không xuất hiện ở công thức bón 26 kg/hố. Tương tự, trong rễ cà phê cũng đã xuất hiện thêm loại nấm Phytophthora với tần xuất xuất hiện 14,29% ở 3 công thức: bón phân liên tục (18 kg/hố, 26 kg/hố)và bón 2 năm/lần (18 kg/hố). Nhìn chung mật độ nấm gây hại cà phê trong đất đều giảm so với trước thí nghiệm, giữa các công thức chu kỳ bón và liều lượng bón khác nhau chưa cho thấy ảnh hưởng rõ đến mật độ nấm trong đất và tần suất xuất hiện nấm trong rễ.
Sau 30 tháng trồng mật độ tuyến trùng trong đất và rễ giảm rõ rệt ở các công thức bón nhiều phân chuồng, chỉ còn công thức bón 2 năm/ lần với mức 10 kg/hố trong rễ xuất hiện 184 con/5 g rễ và ở công thức bón 2 năm/1 lần với mức 18 kg/hố trong rễ xuất hiện 32 con/5 g rễ. Đối với mật độ nấm trong đất và tần suất xuất hiện nấm trong rễ giảm đáng kể ở tất cả các công thức, đặc biệt đối với công thức bón liên tục (10 kg/hố, 18 kg/hố và 26 kg/hố) không thấy xuất hiện nấm Rhizoctonia; Pytophthora trong rễ cây cà phê.
Bảng 5. Thành phần, mật độ tuyến trùng và nấm trong đất trước thí nghiệm, trong đất, rễ sau khi trồng 30 tháng (huyện Krông Pắk, tỉnh Đắk Lắk, 2017)
Ghi chú: C1: bón liên tục 3 năm KTCB; C2: 2 năm bón 1 lần; M1, M2, M3: bón 10,18,26 kg phân hữu cơ/gốc. Pra.: Pratylenchus coffeae; Mel.: Meloidogyne; Rot.: Rotylenchus; Fus.: Fusarium; Rhi.: Rhizoctonia; Pyt.: Pytophthora.
Công thưcTỉ lệ cây vàng (%) Tỉ lệ cây chết (%)
18 tháng
30 tháng
18 tháng
30 tháng
CT1 (C1M1) 5,0 6,3ab 5,0 0CT2 (C1M2) 3,3 3,1b 3,3 0CT3 (C1M3) 1,7 3,1b 1,7 0CT4 (C2M1) 6,7 15,6a 6,7 4,7CT5 (C2M2) 8,3 10,9a 8,3 3,1CT6 (C2M3) 5,0 9,4ab 5,0 0CV (%) 64,3 29,8 68,6 31,2P0,05 0,7791 0,0314 0,8885 0,0915

100
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Như vậy, chu kỳ và liều lượng phân chuồng có ảnh hưởng đến tuyến trùng và nấm trong đất, rễ cà phê, lượng bón càng cao và chu kỳ liên tục hàng năm đã giảm mật độ tuyến trùng và nấm trên cây cà phê tái canh. Kết quả thể hiện ở Bảng 5.
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luậnSử dụng cây giống cà phê ghép 6 - 18 tháng tuổi
trong giai đoạn kiến thiết cơ bản kết hợp với các biện pháp kỹ thuật tuân thủ theo quy trình tái canh cà phê vối năm 2013 của Bộ Nông nghiệp và Phát triển nông thôn đảm bảo cây sinh trưởng và phát triển đồng đều, năng suất cao (2,31 tấn nhân/ha sau 30 tháng trồng), giảm tỷ lệ vàng lá, thối rễ.
Áp dụng mức độ bón phân hữu cơ liên tục trong giai đoạn kiến thiết cơ bản từ 18 - 26 kg/hố sẽ giảm mật độ tuyến trùng và nấm trên cây cà phê tái canh từ đó giảm rõ rệt bệnh vàng lá, thối rễ và có thể đạt năng suất tối đa 2,79 tấn nhân/ha sau 30 tháng trồng.
4.2. Đề nghịSử dụng kết quả nghiên cứu trên để khuyến cáo
trong công tác chỉ đạo tái canh cà phê cho vùng Tây Nguyên.
LỜI CẢM ƠNKết quả nghiên cứu này được hoàn thành trong
khuôn khổ đề tài: “Nghiên cứu nguyên nhân chính gây chết cà phê tái canh và đề xuất giải pháp khắc phục” do Bộ Nông nghiệp và Phát triển nông thôn cấp kinh phí. Nhóm tác giả xin chân thành cảm
ơn Viện Khoa học Kỹ thuật Nông Lâm nghiệp Tây Nguyên và các cộng tác viên đã hỗ trợ và tạo điều kiện thuận lợi để nhóm thực hiện nội dung nghiên cứu này.
TÀI LIỆU THAM KHẢOLê Ngoc Báu, Chế Thị Đa, 2012. Hiện trạng tái canh cà
phê ở Tây Nguyên và giải pháp để tái canh cà phê chu kỳ hai đạt hiệu quả. Hội nghị đánh giá chương trình tái canh cà phê đến năm 2012, phương hướng và giải pháp trong thời gian tới. Lâm Đồng, tháng 10/2012. Trang 28-31.
Cục Trồng trot - Bộ Nông nghiệp và PTNT, 2013. Quyết định số: 273/QĐ-TT-CCN ngày 03/7/2013 về việc ban hành quy trình tái canh cà phê vối. Hà Nội, tháng 7/2013.
Cục Trồng trot - Bộ Nông nghiệp và PTNT, 2016. Báo cáo một số khuyến cáo, định hướng và giải pháp thực hiện chương trình tái canh cà phê giai đoạn 2016 - 2020. Hà Nội, tháng 5/2016.
Chế Thị Đa, Nguyễn Thị Thanh Mai, Lê Đăng Khoa, Nông Khánh Nương, 2016. Nghiên cứu biện pháp kỹ thuật tổng hợp trong tái canh cà phê vối tại Tây Nguyên. Hội thảo Quốc gia về Khoa học cây trồng lần thứ nhất (trang 923-930).
Hooper, D J., 1986. Extraction of free living stages from soil. In Laboratory methods for work with plant and soil nematodes. Ministry of Agriculture, Fisheries and Food J. F. Southey, ed., London., pp.5-30.
Lester W. Burgess, Timothy E. Knight, Len Tesoriero, Phan Thúy Hiền, 2009. Cẩm nang đoán bệnh cây trồng ở Việt Nam. Trung tâm Nghiên cứu Nông nghiệp Quốc tế Australia.
Effects of seedling age and organic fertilizer on growth and development of Robusta coffee in Central Highland
Ta Hong Linh, Nguyen Van Tuat, Bui Quang Dang, Nguyen Thi Thanh Mai, Truong Hong
AbstractThe effect of seedling age and different doses of organic fertilizers on growth and development of replanted Robusta coffee was conducted during the period of 2015 to 2017 in Krong Pak district, Dak Lak province. The results showed that using 18-month grafted-plantlets at the basic establishment stage combined with reasonable cultivation techniques made the plants grown and developed equally, with the yield of 2.31 tons/ha at 30 months after transplanting. The combination of 18-month grafted-plantlets and manure application of 26 kg/pit (continuous application for 3 years) reduced nematode and fungal density, thereby reducing the risk of yellow leaf disease, root rot disease and the maximum yield reached 2.79 tons/ha at 30 months after transplanting.Keywords: Grafted-seedling, nematode, fungus, yellow leaf disease, root rot disease
Ngày nhận bài: 20/7/2018Ngày phản biện: 25/7/2018
Người phản biện: TS. Trần VinhNgày duyệt đăng: 15/10/2018

101
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
NGHIÊN CỨU MỘT SỐ BIỆN PHÁP KỸ THUẬT TRỒNG BÔNG LAI VN01-2 TRONG MÙA KHÔ TẠI SƠN LA
Nguyễn Đình Chiến1, Nguyễn Ngọc Dương1 , Phạm Xuân Liêm2
TÓM TẮTKết quả bước đầu nghiên cứu một số biện pháp kỹ thuật trồng bông mùa khô có tưới bổ sung tại Sơn La trong
năm 2017 cho thấy: Giống bông VN01-2 gieo với mật độ 6,5 vạn cây/ha kết hợp với bấm ngọn vào thời điểm từ 10 đến 12 cành quả cho năng suất bông hạt cao (26,7 tạ/ha); Bón phân với liều lượng phân bón 150 N + 75 P2 O5 + 75 K2O/ha kết hợp với mật độ gieo 5,0 vạn cây/ha cho năng suất cao nhất (27,6 tạ/ha); Sử dụng chất điều hòa sinh trưởng phun lên lá đều làm tăng năng suất bông hạt trong vụ trồng mùa khô, phun Pix (Mepiquat chloride)một lần vào 60 ngày sau gieo, liều lượng 100 ml/ha đã đạt năng suất bông 27,7 tạ/ha, phun α- NAA (α- Naphtalen acetic axit) ba lần (lần 1 vào 30 ngày sau gieo, nồng độ 60 ppm; lần 2 vào 45 ngày sau gieo, nồng độ 80ppm; và lần 3 vào 60 ngày sau gieo, nồng độ 100 ppm) đã đạt năng suất bông 27,13 tạ/ha.
Từ khóa: Giống bông lai VN01-2, mùa khô, biện pháp kỹ thuật, năng suất
1 Công ty Cổ phần Bông miền Bắc; 2 Hiệp Hội Thương mại Giống cây trồng Việt Nam
I. ĐẶT VẤN ĐỀVùng sản xuất bông ở các tỉnh miền núi phía Bắc
nói chung và Sơn La nói riêng là vùng trồng bông bán thâm canh phụ thuộc nước trời. Vụ bông truyền thống bắt đầu gieo hạt vào đầu mùa mưa tháng 6 và thu hoạch vào tháng 10 - 11, không tưới nước trong suốt cả vụ, năng suất đạt khoảng 2 tấn/ha (Công ty Cổ phần Bông miền Bắc, 2015). Hiện nay, sản xuất bông đang phải cạnh tranh với các cây trồng ngắn ngày khác về hiệu quả kinh tế nên diện tích bông đang dần bị thu hẹp. Để mở rộng diện tích sản xuất, cây bông đang được trồng thử nghiệm mùa khô, trên diện tích đất chỉ gieo trồng 1 vụ mùa mưa, bỏ hoang trong mùa khô từ tháng 11 đến tháng 6 và đất hoang ven các triền sông suối ở các tỉnh miền núi phía Bắc. Bài viết này giới thiệu kết quả bước đầu nghiên cứu một số biện pháp kỹ thuật cho giống bông lai VN01-2 trồng mùa khô có tưới bổ sung tại Sơn La để hoàn thiện quy trình kỹ thuật canh tác nhằm nâng cao năng suất, chất lượng bông xơ và phục vụ cho kế hoạch phát triển sản xuất bông vùng miền núi Tây Bắc Việt Nam.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưuGiống bông: Giống bông lai kháng sâu VN01-2,
được công nhận giống quốc gia năm 2004, là giống chủ lực tại vùng sản xuất bông miền núi phía Bắc. Phân bón các loại: phân đạm Urê (46% N), NPK Lâm Thao (5N:10P205:K20), Super lân Lâm Thao (16% P205 ), Kaliclorua (58% K20); Thuốc bảo vệ thực vật: thuốc trừ cỏ Bravo 480 SL phun trước khi làm đất gieo bông, thuốc trừ bệnh Carbenzim 500 FL và thuốc trừ rầy Conphai 15 WP.
2.2. Phương pháp nghiên cưu
2.2.1. Bố trí thí nghiệma) Thí nghiệm 1: Ảnh hưởng của thời điểm bấm ngọn ở các mật độ gieo trồng đến sinh trưởng và năng suất giống bông VN01-2 trong mùa khô có tưới bổ sung tại Sơn La
Thí nghiệm 2 nhân tố bố trí theo kiểu ô chính, ô phụ (split-plot design) trong đó nhân tố chính là mật độ, nhân tố phụ là thời điểm bấm ngọn, diện tích mỗi ô là 50 m2 , 3 lần nhắc lại. Phân bón: 120 kg N + 60 kg P205 + 60 kg K2O/ha. Tưới rãnh đủ ẩm. Ngày gieo 20/1/2017.
- Mật độ gieo: M1: 3,5 vạn cây/ha (khoảng cách 80 cm ˟ 36 cm); M2: 5,0 vạn cây/ha, (khoảng cách 80 cm ˟ 25 cm); M3: 6,5 vạn cây/ha (khoảng cách 80 cm ˟ 19 cm); và M4: 8,0 vạn cây/ha (khoảng cách 80 cm ˟ 16 cm).
- Thời điểm bấm ngọn: T0 - Không bấm ngọn; T1 - Bấm ngọn khi cây bông có 8 - 10 cành quả; T2 - Bấm ngọn khi cây bông có 10 - 12 cành quả; và T3 - Bấm ngọn khi cây bông có 12 - 14 cành quả.b) Thí nghiệm 2: Ảnh hưởng của liều lượng phân bón ở các mật độ gieo trồng đến sinh trưởng và năng suất giống bông VN01-2 trong mùa khô có tưới bổ sung tại Sơn La
Thí nghiệm hai nhân tố bố trí theo kiểu ô chính, ô phụ (split-plot design) trong đó nhân tố chính là lượng phân bón, nhân tố phụ là mật độ cây, diện tích mỗi ô là 50 m2, 3 lần nhắc lại. Tưới rãnh đủ ẩm. Ngày gieo 20/1/2017.
- Lượng bón phân: P1: (90 kg N + 45 kg P2O5 + 45 kg K2O)/ha; P2: (120 kg N + 60 kg P2O5 + 60 kg K2O)/ha; và P3: (150 kg N + 75 kg P2O5 + 75 kg K2O)/ha.

102
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
- Mật độ gieo bông: 3,5 vạn cây; 5,0 vạn cây và 6,5 vạn cây/ha. c) Thí nghiệm 3: Ảnh hưởng của chất điều hòa sinh trưởng Pix (Mepiquat chloride) đến sinh trưởng, phát triển của giống bông VN01-2 trong mùa khô có tưới bổ sung tại Sơn La
- Công thức : CT 1: Phun nước lã (Đ/c) 3 lần vào thời điểm 30 ngày, 45 ngày và 60 ngày sau gieo; CT2: Phun Pix 1 lần vào thời điểm 60 ngày sau gieo, liều lượng 100 ml/ha; CT3: Phun Pix 2 lần; lần 1 vào 45 ngày sau gieo, liều lượng 50 ml/ha; lần 2 vào 60 ngày sau gieo, liều lượng 70 ml/ha; và CT4 - Phun Pix 3 lần; lần 1 vào 30 ngày sau gieo, liều lượng 30 ml/ha; lần 2 vào 45 ngày sau gieo, liều lượng 50 ml/ha và lần 3 vào 60 ngày sau gieo, liều lượng 70 ml/ha.
- Lượng dung dịch phun mỗi lần cho cả 4 công thức là 160 lít/ha. Phân bón: 120 kg N + 60 kg P2O 5 + 60 kg K2O/ha. Mật độ gieo: 5 vạn cây/ha. Không bấm ngọn. Tưới rãnh đủ ẩm. Ngày gieo 25/1/2017. Thí nghiệm bố trí theo kiểu khối ngâu nhiên đầy đủ (RCBD), diện tích ô 50 m2 , 3 lần nhắc lại.d) Thí nghiệm 4: Ảnh hưởng của chất điều hòa sinh trưởng α- NAA (α- Naphtalen acetic axit) đến sinh trưởng, phát triển của giống bông VN01-2 trong mùa khô có tưới bổ sung tại Sơn La
- Công thức : CT 1 - Phun nước lã (Đ/c) 3 lần vào thời điểm 30 ngày, 45 ngày và 60 ngày sau gieo; CT2: Phun α- NAA 1 lần vào 60 ngày sau gieo, nồng độ 100 ppm; CT3 - Phun α- NAA 2 lần; Lần 1vào 45 ngày sau gieo, nồng độ 80 ppm; Lần 2 vào 60 ngày sau gieo, nồng độ 100 ppm; và CT 4 - Phun α- NAA 3 lần; Lần 1 vào 30 ngày sau gieo, nồng độ 60 ppm; Lần 2 vào 45 ngày sau gieo, nồng độ 80 ppm; Lần 3 vào 60 ngày sau gieo, nồng độ 100 ppm.
- Lượng dung dịch phun, mật độ cây, phân bón và các điều kiện khác như thí nghiệm 3.
2.2.2. Chỉ tiêu theo dõiÁp dụng Tiêu chuẩn ngành “Quy phạm khảo
nghiệm giá trị canh tác và giá trị sử dụng của giống bông” - 10TCN 911:2006 (Bộ Nông nghiệp và PTNT, 2006).
2.2.3. Phương pháp xử lý số liệu Số liệu thí nghiệm được xử lý thống kê trên máy
vi tính với phần mềm chuyên dụng IRRISTAT 5.0 và ch ương trình Microsoft Excel.
2.3. Thời gian và địa điểm nghiên cưuNghiên cứu được thực hiện từ tháng 1 đến tháng
6/2017 tại xã Tú Nang, huyện Yên Châu, tỉnh Sơn La.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Ảnh hưởng của thời điểm bấm ngon ở các mật độ gieo trồng đến sinh trưởng và năng suất giống bông VN01-2 trong mùa khô có tưới bổ sung tại Sơn La
3.1.1. Ảnh hưởng của thời điểm bấm ngọn ở các mật độ gieo trồng đến sinh trưởng của cây bông
Kết quả trên bảng 1 cho thấy: Bấm ngọn sớm, hay tăng mật độ có xu hướng rút ngắn thời gian sinh trưởng và giảm chiều cao cây bông; Bấm ngọn muộn thì số cành quả/cây tăng, nhưng tăng mật độ trồng thì số cành quả/cây giảm; Bấm ngọn và tăng mật độ trồng có ảnh hưởng đến số cành đực/cây nhưng không có quy luật (Bảng 1).
3.1.2. Ảnh hưởng của mật độ và bấm ngọn đến năng suất của giống bông VN01-2
Kết quả trên bảng 2 cho thấy: Khi tăng mật độ từ 3,5 vạn cây/ha lên 5,0 vạn cây và 6,5 vạn cây thì số quả/m2 tăng nhưng tăng đến 8,0 vạn cây thì số quả/m2 giảm; Bấm ngọn ở các thời điểm khác nhau không ảnh hưởng rõ rệt đến số quả/m2; Bấm ngọn và tăng mật độ hầu như không ảnh hưởng đến khối lượng quả. Năng suất bông đạt cao (25,86 - 26,73 tạ/ha) ở các mật độ 5,0 vạn, 6,5 vạn cây/ha kết hợp với bấm ngọn ở các thời điểm từ 8 đến 14 cành quả. Kết quả này gần như giống với nghiên cứu của Dương Xuân Diêu (2003) trên giống bông VN01-2tại Duyên hải Nam Trung bộ. Năng suất cao nhất (26,73 tạ/ha) ở mật độ 6,5 vạn cây/ha và bấm ngọn vào thời điểm cây có 10 - 12 cành quả (Bảng 2).
3.2. Ảnh hưởng của liều lượng phân bón ở các mật độ gieo trồng đến sinh trưởng và năng suất giống bông VN01-2 trong mùa khô có tưới bổ sung tại Sơn La
3.2.1. Ảnh hưởng của liều lượng phân bón ở các mật độ gieo trồng đến sinh trưởng của cây bông
Kết quả trên bảng 3 cho thấy: Thời gian sinh trưởng và chiều cao cây bông có xu hướng tăng khi lượng phân bón tăng, hoặc mật độ gieo tăng từ 3,5 vạn cây đến 6,5 vạn cây/ha. Số cành đực và số cành quả/cây cũng có xu hướng tăng khi lượng phân bón tăng, nhưng có xu hướng giảm khi mật độ cây tăng (Bảng 3).
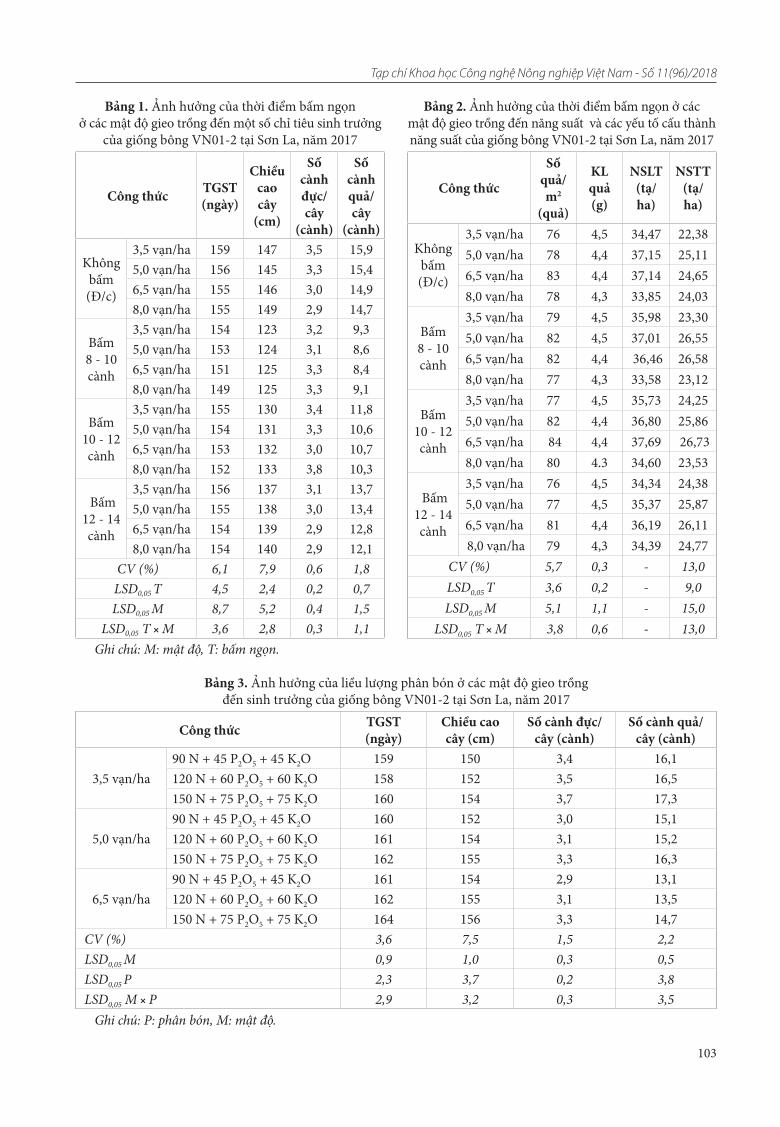
103
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Ghi chú: M: mật độ, T: bấm ngọn.
Bảng 1. Ảnh hưởng của thời điểm bấm ngọn ở các mật độ gieo trồng đến một số chỉ tiêu sinh trưởng
của giống bông VN01-2 tại Sơn La, năm 2017
Công thưc TGST(ngày)
Chiều caocây
(cm)
Số cành đực/cây
(cành)
Số cành quả/cây
(cành)
Không bấm (Đ/c)
3,5 vạn/ha 159 147 3,5 15,95,0 vạn/ha 156 145 3,3 15,46,5 vạn/ha 155 146 3,0 14,98,0 vạn/ha 155 149 2,9 14,7
Bấm 8 - 10 cành
3,5 vạn/ha 154 123 3,2 9,35,0 vạn/ha 153 124 3,1 8,66,5 vạn/ha 151 125 3,3 8,48,0 vạn/ha 149 125 3,3 9,1
Bấm 10 - 12 cành
3,5 vạn/ha 155 130 3,4 11,85,0 vạn/ha 154 131 3,3 10,66,5 vạn/ha 153 132 3,0 10,78,0 vạn/ha 152 133 3,8 10,3
Bấm 12 - 14 cành
3,5 vạn/ha 156 137 3,1 13,75,0 vạn/ha 155 138 3,0 13,46,5 vạn/ha 154 139 2,9 12,88,0 vạn/ha 154 140 2,9 12,1
CV (%) 6,1 7,9 0,6 1,8 LSD0,05 T 4,5 2,4 0,2 0,7 LSD0,05 M 8,7 5,2 0,4 1,5
LSD0,05 T ˟ M 3,6 2,8 0,3 1,1
Bảng 2. Ảnh hưởng của thời điểm bấm ngọn ở các mật độ gieo trồng đến năng suất và các yếu tố cấu thành năng suất của giống bông VN01-2 tại Sơn La, năm 2017
Công thưc
Số quả/m2
(quả)
KL quả(g)
NSLT(tạ/ha)
NSTT(tạ/ha)
Không bấm (Đ/c)
3,5 vạn/ha 76 4,5 34,47 22,385,0 vạn/ha 78 4,4 37,15 25,116,5 vạn/ha 83 4,4 37,14 24,658,0 vạn/ha 78 4,3 33,85 24,03
Bấm 8 - 10 cành
3,5 vạn/ha 79 4,5 35,98 23,305,0 vạn/ha 82 4,5 37,01 26,556,5 vạn/ha 82 4,4 36,46 26,588,0 vạn/ha 77 4,3 33,58 23,12
Bấm 10 - 12 cành
3,5 vạn/ha 77 4,5 35,73 24,255,0 vạn/ha 82 4,4 36,80 25,866,5 vạn/ha 84 4,4 37,69 26,738,0 vạn/ha 80 4.3 34,60 23,53
Bấm 12 - 14 cành
3,5 vạn/ha 76 4,5 34,34 24,385,0 vạn/ha 77 4,5 35,37 25,876,5 vạn/ha 81 4,4 36,19 26,11 8,0 vạn/ha 79 4,3 34,39 24,77
CV (%) 5,7 0,3 - 13,0 LSD0,05 T 3,6 0,2 - 9,0 LSD0,05 M 5,1 1,1 - 15,0
LSD0,05 T ˟ M 3,8 0,6 - 13,0
Bảng 3. Ảnh hưởng của liều lượng phân bón ở các mật độ gieo trồng đến sinh trưởng của giống bông VN01-2 tại Sơn La, năm 2017
Ghi chú: P: phân bón, M: mật độ.
Công thưc TGST(ngày)
Chiều caocây (cm)
Số cành đực/cây (cành)
Số cành quả/cây (cành)
3,5 vạn/ha90 N + 45 P2 O5 + 45 K2 O 159 150 3,4 16,1120 N + 60 P2 O5 + 60 K2 O 158 152 3,5 16,5150 N + 75 P2 O5 + 75 K2 O 160 154 3,7 17,3
5,0 vạn/ha90 N + 45 P2 O5 + 45 K2 O 160 152 3,0 15,1120 N + 60 P2 O5 + 60 K2 O 161 154 3,1 15,2150 N + 75 P2 O5 + 75 K2 O 162 155 3,3 16,3
6,5 vạn/ha90 N + 45 P2 O5 + 45 K2 O 161 154 2,9 13,1120 N + 60 P2 O5 + 60 K2 O 162 155 3,1 13,5150 N + 75 P2 O5 + 75 K2 O 164 156 3,3 14,7
CV (%) 3,6 7,5 1,5 2,2 LSD0,05 M 0,9 1,0 0,3 0,5 LSD0,05 P 2,3 3,7 0,2 3,8 LSD0,05 M ˟ P 2,9 3,2 0,3 3,5
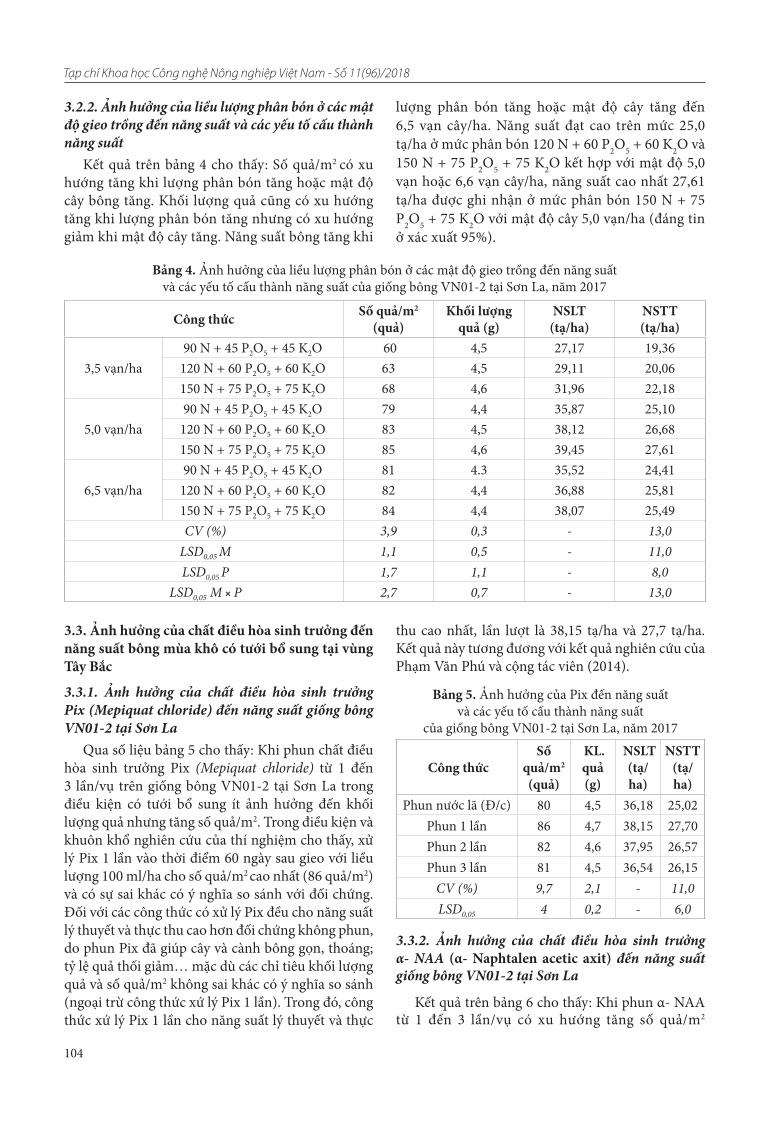
104
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
3.2.2. Ảnh hưởng của liều lượng phân bón ở các mật độ gieo trồng đến năng suất và các yếu tố cấu thành năng suất
Kết quả trên bảng 4 cho thấy: Số quả/m2 có xu hướng tăng khi lượng phân bón tăng hoặc mật độ cây bông tăng. Khối lượng quả cũng có xu hướng tăng khi lượng phân bón tăng nhưng có xu hướng giảm khi mật độ cây tăng. Năng suất bông tăng khi
lượng phân bón tăng hoặc mật độ cây tăng đến 6,5 vạn cây/ha. Năng suất đạt cao trên mức 25,0 tạ/ha ở mức phân bón 120 N + 60 P2 O5 + 60 K2 O và 150 N + 75 P2 O5 + 75 K2O kết hợp với mật độ 5,0 vạn hoặc 6,6 vạn cây/ha, năng suất cao nhất 27,61 tạ/ha được ghi nhận ở mức phân bón 150 N + 75 P2 O5 + 75 K2O với mật độ cây 5,0 vạn/ha (đáng tin ở xác xuất 95%).
3.3. Ảnh hưởng của chất điều hòa sinh trưởng đến năng suất bông mùa khô có tưới bổ sung tại vùng Tây Bắc
3.3.1. Ảnh hưởng của chất điều hòa sinh trưởng Pix (Mepiquat chloride) đến năng suất giống bông VN01-2 tại Sơn La
Qua số liệu bảng 5 cho thấy: Khi phun chất điều hòa sinh trưởng Pix (Mepiquat chloride) từ 1 đến 3 lần/vụ trên giống bông VN01-2 tại Sơn La trong điều kiện có tưới bổ sung ít ảnh hưởng đến khối lượng quả nhưng tăng số quả/m2. Trong điều kiện và khuôn khổ nghiên cứu của thí nghiệm cho thấy, xử lý Pix 1 lần vào thời điểm 60 ngày sau gieo với liều lượng 100 ml/ha cho số quả/m2 cao nhất (86 quả/m2) và có sự sai khác có ý nghĩa so sánh với đối chứng. Đối với các công thức có xử lý Pix đều cho năng suất lý thuyết và thực thu cao hơn đối chứng không phun, do phun Pix đã giúp cây và cành bông gọn, thoáng; tỷ lệ quả thối giảm… mặc dù các chỉ tiêu khối lượng quả và số quả/m2 không sai khác có ý nghĩa so sánh (ngoại trừ công thức xứ lý Pix 1 lần). Trong đó, công thức xứ lý Pix 1 lần cho năng suất lý thuyết và thực
thu cao nhất, lần lượt là 38,15 tạ/ha và 27,7 tạ/ha. Kết quả này tương đương với kết quả nghiên cứu của Phạm Văn Phú và cộng tác viên (2014).
Bảng 5. Ảnh hưởng của Pix đến năng suất và các yếu tố cấu thành năng suất
của giống bông VN01-2 tại Sơn La, năm 2017
3.3.2. Ảnh hưởng của chất điều hòa sinh trưởng α- NAA (α- Naphtalen acetic axit) đến năng suất giống bông VN01-2 tại Sơn La
Kết quả trên bảng 6 cho thấy: Khi phun α- NAA từ 1 đến 3 lần/vụ có xu hướng tăng số quả/m2
Bảng 4. Ảnh hưởng của liều lượng phân bón ở các mật độ gieo trồng đến năng suất và các yếu tố cấu thành năng suất của giống bông VN01-2 tại Sơn La, năm 2017
Công thưc Số quả/m2
(quả)Khối lượng
quả (g)NSLT
(tạ/ha)NSTT(tạ/ha)
3,5 vạn/ha90 N + 45 P2 O5 + 45 K2 O 60 4,5 27,17 19,36
120 N + 60 P2 O5 + 60 K2 O 63 4,5 29,11 20,06150 N + 75 P2 O5 + 75 K2 O 68 4,6 31,96 22,18
5,0 vạn/ha90 N + 45 P2 O5 + 45 K2 O 79 4,4 35,87 25,10
120 N + 60 P2 O5 + 60 K2 O 83 4,5 38,12 26,68150 N + 75 P2 O5 + 75 K2 O 85 4,6 39,45 27,61
6,5 vạn/ha90 N + 45 P2 O5 + 45 K2 O 81 4.3 35,52 24,41
120 N + 60 P2 O5 + 60 K2 O 82 4,4 36,88 25,81150 N + 75 P2 O5 + 75 K2 O 84 4,4 38,07 25,49 CV (%) 3,9 0,3 - 13,0
LSD0,05 M 1,1 0,5 - 11,0 LSD0,05 P 1,7 1,1 - 8,0
LSD0,05 M ˟ P 2,7 0,7 - 13,0
Công thưcSố
quả/m2
(quả)
KL. quả (g)
NSLT(tạ/ha)
NSTT(tạ/ha)
Phun nước lã (Đ/c) 80 4,5 36,18 25,02Phun 1 lần 86 4,7 38,15 27,70Phun 2 lần 82 4,6 37,95 26,57Phun 3 lần 81 4,5 36,54 26,15
CV (%) 9,7 2,1 - 11,0LSD0,05 4 0,2 - 6,0

105
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Công thưc Số quả/m2 (quả)
KL quả(g)
NSLT(tạ/ha)
NSTT(tạ/ha)
Phun nước lã (Đ/c) 81 4,5 36,40 24,68
Phun 1 lần 81 4,6 37,44 26,21 Phun 2 lần 82 4,7 38,63 27,04Phun 3 lần 85 4,8 41,14 27,13
CV (%) 8,4 0,9 - 16,0LSD0,05 5 0,3 - 8,0
và khối lượng quả nhưng không có sự sai khác có ý nghĩa so sánh. Tuy nhiên, năng suất lý thuyết và năng suất thực thu trên các công thức có phun α- NAA đều cao hơn có ý nghĩa so với đối chứng; công phun α- NAA ba lần (vào 30, 45, 60 ngày sau gieo với nồng độ lần lượt 60, 80, 100 ppm) cho năng suất thực thu cao nhất 27,13 tạ/ha.
Bảng 6. Ảnh hưởng của α- NAA đến năng suất và các yếu tố cấu thành năng suất
của giống bông VN01-2 tại Sơn La, năm 2017
IV. KẾT LUẬN Kết quả nghiên cứu một số biện pháp kỹ thuật
trồng giống bông lai VN01-2 trong mùa khô năm 2017, có tưới bổ sung tại Sơn La cho thấy:
- Giống bông lai VN01-2 tại Sơn La trồng trong vụ khô có tưới bổ sung với mật độ 6,5 vạn cây/ha kết hợp với bấm ngọn vào thời điểm 10 đến 12 cành quả cho năng suất cao nhất (26,73 tạ/ha).
- Trong điều kiện vụ khô có tưới bổ sung tại Sơn La, giống bông lai VN01-2 trồng mật độ 5,0 vạn cây/ha với liều lượng phân bón 150 N + 75 P2 O5 + 75 K2O/ha cho năng suất cao nhất (27,61 tạ/ha).
- Giống bông lai VN01-2 trồng trong vụ khô có tưới bổ sung tại Sơn La, sử dụng chất điều hòa sinh
trưởng phun lên lá đều tăng năng suất bông hạt. Phun Pix (Mepiquat chloride) một lần vào 60 ngày sau gieo, liều lượng 100 ml đã đạt năng suất bông 27,7 tạ/ha, hoặc phun α- NAA (α- Naphtalen acetic axit) ba lần (Lần 1 vào 30 ngày sau gieo, nồng độ 60 ppm; Lần 2 vào 45 ngày sau gieo, nồng độ 80 ppm; và lần 3 vào 60 ngày sau gieo, nồng độ 100 ppm) đã đạt năng suất bông 27,13 tạ/ha.
LỜI CẢM ƠN Nghiên cứu được hoàn thành với nguồn kinh phí
của nhiệm vụ khoa học công nghệ cấp Bộ “Xây dựng mô hình sản xuất bông mùa khô có tưới bổ sung tại vùng Tây Bắc”, 2016 - 2018. Nhóm tác giả xin trân trọng cảm ơn sự hỗ trợ của Bộ Công thương.
TÀI LIỆU THAM KHẢOBộ Nông nghiệp và PTNT, 2006. Tiêu chuẩn ngành
10TCN 911:2006, Quy phạm khảo nghiệm giá trị canh tác và giá trị sử dụng của giống bông.
Công ty Cổ phần Bông miền Bắc, 2015. Báo cáo tổng kết sản xuất bông 2013 - 2015.
Dương Xuân Diêu, 2003. Nghiên cứu ảnh hưởng của mật độ và chất điều hoà sinh trưởng PIX đến một số chỉ tiêu sinh lý và năng suất bông giống VN01-2 tại Duyên hải Nam Trung bộ. Luận văn Thạc sỹ Khoa học Nông nghiệp, Trường Đại học Nông nghiệp I, Hà Nội.
Dương Xuân Diêu và ctv, 2008. “Xây dựng mô hình phát triển bông vụ khô có tưới”. Báo cáo nghiệm thu tại Hội đồng Khoa học Tập đoàn Dệt May Việt Nam 2008.
Phạm Văn Phú, Phạm Đình Ổn, Nguyễn Ngoc Dương và Phan Quốc Hiển, 2014. Nghiên cứu sử dụng chất điều tiết sinh trưởng cho bông chín tập trung, giảm công thu hoạch nhằm khuyến khích đồng bào miền núi trồng bông. Báo cáo nghiệm thu tại Hội đồng Khoa học Bộ Công thương.
Study on technical measures for growing cotton hybrid variety VN01-2 in dry season in Sonla province
Nguyen Dinh Chien, Nguyen Ngoc Duong and Pham Xuan LiemAbstractThe study result of technical measures for growing cotton hybrid variety VN01-2 in dry season of 2017 in combination with irrigating in Sonla province showed that: The cotton variety VN01-2 had high yield (26.7 quintals/ha) with growing density of 65 thousand of seedlings/ha combining with sprouting when plants had 10 -12 fruit branches; The yield was highest when applying 150 N + 75 P2 O5 + 75 K2O/ha combining with sowing density of 50 thousand of seedlings/ha; The yield reached 27.7 quintals/ha when spraying regulatory hormone Pix (Mepiquat chloride) with the dose of 100 ml/ha one time at the age of 60 days affter sowing; and reached 27.13 quintals/ha when spraying α- NAA (α- Naphtalen acetic axit) three times (at the age of 30; 40 and 60 days affter sowing).Keywords: Cotton hybrid variety VN01-2, dry season, technical measures, yield
Ngày nhận bài: 14/8/2018Ngày phản biện: 27/8/2018
Người phản biện: TS. Phan Công KiênNgày duyệt đăng: 18/9/2018

106
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
NGHIÊN CỨU XÂY DỰNG QUY TRÌNH CÔNG NGHỆ NUÔI TRỒNG NẤM ĐẦU KHỈ Hericium erinaceus TRÊN BÃ DONG ĐAO
Cồ Thị Thuỳ Vân1, Trần Liên Hà2, Nguyễn Văn Minh2
TÓM TẮTNấm Đầu khỉ (hay còn gọi là nấm Hầu thủ) có tên khoa học là Hericium erinaceus. Nấm Đầu khỉ là loại nấm dược
liệu quý chứa đầy đủ các chất dinh dưỡng (axit amin, đường, lipit, nguyên tố khoáng, vitamin) và các chất có hoạt tính sinh học. Về dược lý, nấm Đầu khỉ được chứng minh có tác dụng nâng cao khả năng miễn dịch, phục hồi niêm mạc dạ dày, chữa loét thủng ruột, nâng cao năng lực đề kháng với tình trạng thiếu oxy, chống mệt mỏi, chống oxy hóa, chống đột biến, làm giảm mỡ máu, xúc tiến tuần hoàn máu, chống lão hóa, ức chế sinh trưởng của tế bào ung thư. Các nội dung được tập trung nghiên cứu trong bài này là xác định các điều kiện thích hợp để nuôi trồng nấm Đầu khỉ Hericium erinaceus trên nguồn bã dong đao: Nấm Đầu khỉ sinh trưởng và phát triển tốt trên môi trường 85% bã dong đao + 15% dinh dưỡng (7,5% cám ngô + 7,5% cám gạo). Trên môi trường 50% bã dong + 35% lõi ngô + 15% (cám gạo + bột ngô) thu được nấm đầu khỉ có năng suất thu lần thứ nhất là cao nhất (3060 g nấm tươi). Bã dong đao hoàn toàn phù hợp làm cơ chất nuôi trồng nấm Đầu khỉ.
Từ khóa: Nấm Đầu khỉ, Hericium erinaceus, giống nấm dược liệu, nấm đảm, nấm ăn - nấm dược liệu
1 Trung tâm Nghiên cứu và Phát triển Nấm - Viện Di truyền Nông nghiệp 2 Viện Công nghệ sinh học và Công nghệ thực phẩm - Trường ĐH Bách khoa Hà Nội
I. ĐẶT VẤN ĐỀNấm Đầu khỉ có tên khoa học là Hericium
erinaceus (Bull.; Fr.) (Lê Xuân Thám, 2004). Ở Trung Quốc nấm Đầu khỉ được gọi là Shishigashida và ở Nhật Bản nó được gọi là Yamabushi-take. Đây là loại nấm được biết đến như một vị thuốc dân gian có tác dụng giải nhiệt, điều hòa huyết áp, kích thích tiêu hóa, tăng cường miễn dịch (Nguyễn Lân Dũng, 2001; Khuất Hữu Trung, 2003; Trung tâm Công nghệ sinh học thực vật, 2012). ß-(1-3)-D-glucan được tách chiết từ loại nấm này đã chứng tỏ khả năng ức chế sự hoạt động của tế bào ung thư và sự hoạt động của u bướu, có tác dụng tốt đối với hệ tim mạch, hệ thần kinh (Han Gyu Ko, et al., 2005; Han ZH, Ye JM, Wang GF, 2012; Md Asaduzzaman Khan, et al. 2009).
Nấm Đầu khỉ đã được nuôi trồng khá phổ biến trên thế giới với nhiều phương pháp khác nhau, trên nhiều nguồn cơ chất giàu cellulo. Giống nấm Đầu khỉ Hericium erinaceus ký hiệu He1 được lưu giữ tại Trung tâm Nghiên cứu và Phát triển Nấm - Viện Di truyền Nông nghiệp. Đặc tính nông sinh học của giống He1: Quả thể nấm có màu trắng muốt, hình dạng giống như bộ óc khỉ, gồm các múi thịt nấm ghép lại với nhau. Giai đoạn nhân giống: nhiệt độ nuôi sợi thích hợp 23 - 25oC; Giai đoạn nuôi trồng: thời gian ươm sợi từ 25 - 26 ngày, nhiệt độ nuôi sợi thích hợp 23 - 25oC; Nhiệt độ ra quả thể 20 - 27oC.
Các nghiên cứu trong phạm vi bài báo này được thực hiện nhằm xác định các điều kiện để nuôi trồng nấm Đầu khỉ He1 trên nguồn bã dong đao được lấy từ làng nghề làm miến dong ở Việt Nam.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưu - Giống nấm Đầu khỉ Hericium erinaceus ký hiệu
He1 được lưu giữ tại Trung tâm Nghiên cứu và Phát triển Nấm - Viện Di truyền Nông nghiệp. Giống He1 đã được Bộ Nông nghiệp và Phát triển nông thôn công nhận là giống sản xuất thử.
- Giống đưa vào sử dụng là giống dạng hạt nhân trên thóc luộc: Thực hiện theo qui trình nhân giống đã được công nhận tiến bộ khoa học của Trung tâm Nghiên cứu và Phát triển Nấm - Viện Di truyền Nông nghiệp.
- Bã dong đao lấy từ làng So, xã Cộng Hòa, huyện Quốc Oai, Hà Nội.
2.2. Phương pháp nghiên cưu- Sử dụng các phương pháp nấm học trong
nghiên cứu hình thái quả thể, sự sinh trưởng của hệ sợi (Trịnh Tam Kiệt, 2013; Lê Xuân Thám, 2004).
- Phương pháp nuôi trồng nấm trên nguồn cơ chất tổng hợp theo Nguyễn Hữu Đống, Đinh Xuân Linh, Nguyễn Lân Dũng, Lê Xuân Thám (Lê Xuân Thám, 2004; Nguyễn Lân Dũng, 2001; Khuất Hữu Trung, 2003; Trung tâm Công nghệ sinh học thực vật, 2012).
- Phương pháp khảo sát ảnh hưởng của độ ẩm cơ chất phối trộn đến sự sinh trưởng, phát triển và khả năng nhiễm bệnh của nấm Đầu khỉ trong giai đoạn nuôi trồng thu quả thể được tiến hành thí nghiệm như sau: nguyên liệu 100% bã dong đao, được điều chỉnh độ ẩm ở các ngưỡng 40%, 45%, 50%, 55%,

107
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
60%, 65%, 70%, 75%, 80%, 85%. Thực hiện trên ống nghiệm, mỗi ngưỡng độ ẩm 5 ống nghiệm; Hấp khử trùng ống nghiệm ở 1210C trong 120 phút sau đó làm nguội nhanh; Cấy giống; nuôi sợi ở nhiệt độ 22 - 25oC, độ ẩm không khí 65 - 70%; Sau 3 ngày cấy giống kiểm và quan sát đặc điểm hệ sợi, tiến hành đo độ dài sợi ăn, ghi chép số liệu, kiểm tra ống nhiễm để loại bỏ.
- Phương pháp khảo sát ảnh hưởng của nguồn dinh dưỡng đến sự snh trưởng và phát triển của nấm Đầu khỉ: Các công thức dinh dưỡng sử dụng trong nuôi trồng thử nghiệm nấm Đầu khỉ bao gồm: CT 1: 85% bã dong đao + 15% (cám gạo + bột ngô); CT 2: 90% bã dong + 10% (cám gạo + bột ngô); CT 3: 50% bã dong + 35% lõi ngô + 15% (cám gạo + bột ngô); CT 4: 45% bông + 40% lõi ngô + 15% (cám gạo + bột ngô); CT 5: 45% mùn cưa + 40% bông + 15% (cám gạo + bột ngô); CT6: 95% bã dong đao, 5% cám ngô cám gạo. Mỗi bịch nặng 800 g.
- Các cơ chế khử trùng nguyên liệu:+ Khử trùng bằng lò hấp thủ công: Nhiệt độ
nguyên liệu đạt 100oC/7 giờ liên tục.+ Khử trùng bằng nồi hấp áp lực áp suất 1,3 - 1,4
atm trong 3,0 giờ.- Bố trí thí nghiệm lặp lại 3 lần.- Các chỉ tiêu theo dõi:+ Thời gian bung sợi; Đặc điểm hệ sợi.+ Tốc độ phát triển của hệ sợi được tính theo
công thức: V = D/T1.Trong đó: V là tốc độ phát triển của hệ sợi (mm/ngày);
D là chiều cao bịch nuôi trồng (mm); T1 là thời gian ươm sợi (ngày), được tính từ ngày cấy giống đến ngày hệ sợi ăn kín bịch nuôi trồng.
+ Tổng thời gian nuôi trồng, kí hiệu T (ngày): T = T1 + T2 + T3 + T4.
Trong đó: T1 là thời gian ươm sợi (ngày), được tính từ ngày cấy giống đến ngày hệ sợi ăn kín bịch nuôi trồng; T2: Thời gian hình thành mầm quả thể nấm, tính từ khi hệ sợi mọc kín bịch nguyên liệu đến khi thấy xuất hiện mầm quả thể ở vết rạch hay trên cổ nút bịch nguyên liệu; T3: Thời gian quả thể trưởng thành, tính từ khi xuất hiện mầm quả thể đến khi quả trưởng thành có thể thu hoạch; T4: Thời gian thu quả thể lần 2, tính từ khi thu quả thể lần 1 đến khi quả thể lần 2 được thu hoạch.
+ Hiệu suất sinh học (%) được tính theo công thức: Tổng khối lượng nấm tươi
Khối lượng nguyên liệu khôHiệu suất sinh học (%) = ˟ 100%
+ Năng suất nấm tươi được tính như sau:
Năng suất nấm tươi (%) =M1 - M2
Khối lượng nguyên liệu khô ˟ 100%
Trong đó: M1 là tổng khối lượng nấm thu được;
M2 là khối lượng phần nấm không có giá trị sử dụng (chân nấm, cuống nấm).
Đối với nấm Đầu khỉ năng suất nấm thường bằng hiệu suất sinh học do nấm Đầu khỉ không có phần cuống và gốc nấm.
+ Năng suất nấm khô được tính như sau:
Năng suất nấm khô (%) =Tổng khối lượng nấm khô
Khối lượng nguyên liệu khô ˟ 100%
Năng suất nấm tươi trung bình (%) =Năng suất nấm tươi
Tổng số bịch thí nghiệm
Năng suất nấm khô trung bình =Năng suất nấm tươi
Tổng số bịch thí nghiệm + Tỉ lệ nhiễm bệnh, được tính theo công thức:
Tỷ lệ nhiễm (%) = ˟ 100%Tổng số mâu nhiễm
Tổng số mâu thí nghiệm
2.3. Thời gian và địa điểm nghiên cưuNghiên cứu được thực hiện từ tháng 1/2017 đến
tháng 6/2018 tại Trung tâm Nghiên cứu và Phát triển Nấm - Viện Di truyền Nông nghiệp và Viện Công nghệ sinh học và Công nghệ thực phẩm - Trường Đại học Bách khoa Hà Nội.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Ảnh hưởng của độ ẩm cơ chất phối trộn đến sự sinh trưởng, phát triển của hệ sợi và khả năng nhiễm bệnh trong giai đoạn nuôi trồng thu quả thể
Kết quả được ghi nhận ở Hình 1 và Hình 2.
Hình 1. Ảnh hưởng của độ ẩm cơ chất phối trộn đến tốc độ phát triển của hệ sợi nấm Đầu khỉ
Nhận xét: Kết quả nghiên cứu cho thấy độ ẩm của nguyên liệu có ảnh hưởng rõ rệt tới sự sinh trưởng và phát triển của hệ sợi nấm Đầu khỉ; sau 15 ngày cấy trên cơ chất bã dong đao hệ sợi nấm Đầu khỉ

108
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
có thể phát triển ở các độ ẩm 50%, 55%, 60%, 65%, 70%, 75%, trong đó hệ sợi phát triển tốt ở độ ẩm 60% và 65%, tốt nhất là 65% với độ vượt là 2,96 mm/ngày, hệ sợi khỏe mạnh, mật độ dày. Ở các độ ẩm 40%, 45% không có dấu hiệu bung sợi, do độ ẩm quá thấp nấm không có nước để phân giải cơ chất, không hòa tan được các chất dinh dưỡng, hệ sợi không phát triển. Độ ẩm 80%, 85%, 3 ngày đầu thì giống bung sợi tốt, nhưng do độ ẩm quá cao làm cho sợi không ăn vào bã dong đao, mà hệ sợi chùn lại có dấu hiệu ăn ngược lên.
Kết luận: lựa chọn độ ẩm nguyên liệu 65% để tiếp tục tiến hành các thí nghiệm tiếp theo.
Hình 2. Đặc điểm hệ sợi nấm Đầu khỉ trên các độ ẩm nguyên liệu khác nhau
3.2. Ảnh hưởng của thành phần dinh dưỡng phối trộn đến sự sinh trưởng của hệ sợi nấm Đầu khỉ
Kết quả khảo sát ảnh hưởng của thành phần cơ chất phối trộn đến tốc sự sinh trưởng của hệ sợi nấm đầu khỉ được ghi ở hình 3.
Nhận xét: Kết quả nghiên cứu sau 38 - 40 ngày hệ sợi trên các công thức nuôi trồng sẽ lan kín bịch. Hai công thức đối chứng là CT4 và CT5, đây là hai công thức nuôi trồng của Trung tâm Phát triển Nấm
thuộc Viện Di truyền Nông nghiệp đã tìm ra trong quá trình nuôi trồng sản xuất nấm Đầu khỉ, đây là hai công thức cho năng suất cao nhất. Nổi trội là công thức CT4 có tốc độ lan sợi nhanh nhất độ vượt là 3,33 mm/ngày. CT4 chứa thành phần là bông mà lõi ngô, CT5 thành phần là mùn cưa, bông hai công thức chỉ khác nhau về thành phần mùn và lõi ngô nhưng tốc độ lan sợi lại khác nhau rất nhiều. Có thể thấy lõi ngô là nguyên liệu tốt cho hệ sợi phát triển do có độ xốp cao hơn so với mùn cưa. Các CT1, CT2, CT3 là các công thức có thành phần là bã dong đao đều phát triển tốt, đặc biệt CT3 có thành phần 50% bã dong đao tốc độ lan sợi nhanh 2,92 mm/ngày, hệ sợi dày và khỏe, giúp cho năng suất quả thể thu được sau này sẽ cao. CT3 cơ chất chính là bã dong và được bổ sung thêm 35% lõi ngô bổ sung thêm lượng lớn cellulose và làm tăng thêm độ xốp cho cơ chất. Riêng đối với CT6 do có thành phần dinh dưỡng thấp do bổ sung 5% cám ngô và cám gạo nên hệ sợi rất mỏng, tốc độ lan sợi rất chậm.
Hình 3. Ảnh hưởng của nguyên liệu nuôi trồng đến tốc độ sinh trưởng của hệ sợi nấm Đầu khỉ
Tiếp tục theo dõi năng suất nấm Đầu khỉ thương phẩm ở các công thức môi trường dinh dưỡng khác nhau thu được kết quả ở Bảng 1.
Bảng 1. Ảnh hưởng của công thức phối trộn nguyên liệu nuôi trồng đến năng suất nấm thương phẩm
CT Số bịchThời gian ra mầm quả thể
(ngày)
Thời gian quả thể trưởng
thành (ngày)
Năng suất tươi (g)
Năng suất tươi TB (g)
Năng suất khô (g)
Năng suất khô TB (g)
CT1 47 40 14 2630,00 ± 5 56,00 410,00 ± 5 8,72
CT2 42 40 14 2140,00 ± 5 51,00 335,00 ± 5 7,98
CT3 40 40 14 3060,00 ± 5 76,50 420,00 ± 5 10,50
CT4 38 39 14 2800,00 ± 5 73,68 400,00 ± 5 10,52
CT5 40 40 14 2850,00 ± 5 71,25 415,00 ± 5 10,38

109
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Kết quả thu quả thể đợt một sau 54 ngày nuôi trồng. CT3 cho năng suất tươi là cao nhất, khối lượng tươi là 3060 g, mặc dù tốc độ sợi lan không nhanh nhất. Qủa thể to, mập, trắng đều. CT4 thu được quả thể sau 39 ngày sớm hơn một ngày so với các công thức khác năng suất tươi đạt 2800 g. Năng
suất khô các công thức có bã dong tương đối cao, đặc biệt CT1 có tổng năng suất khô 410 g rất là cao. CT6 không cho quả thể, mầm quả thể mọc sau đó chuyển màu vàng và thối.
Sau 18 ngày tiếp tục thu được quả thể lần thứ hai. Kết quả thể hiện qua Bảng 2.
Năng suất thu quả thể lần hai ở tất cả các công thức đã giảm đi khá nhiều do nguồn dinh dưỡng đã cạn dần. Quả thể không còn to, đây như lần thứ nhất mà bé, xốp và nhẹ. CT4 có năng suất cao nhất 2090 g.
- Tổng năng suất nấm Đầu khỉ thu được sau hai lần thu quả thể được tổng hợp trong bảng 3.
Sau 72 ngày nghiên cứu, kết quả cho thấy: CT4
là công thức có năng suất tươi và hiệu suất sinh học cao nhất lần lượt là 4890 g và 12,23%. Tiếp theo là CT5 có năng suất tươi là 4550 g và hiệu suất sinh học là 11,38%. CT3 có 50% cơ chất bã dong đao, năng suất tươi đạt 4260 g, hiệu suất sinh học 10,65% không chênh lệch quá nhiều so với hai công thức CT4, CT5. CT1 có tỉ lệ hàm lượng chất khô cao nhất 14,55%.
Bảng 2. Năng suất quả thể nấm Đầu khỉ thu lần thứ hai
Bảng 3. Tổng năng suất quả thể thu được
CT Thời gian thu quả thể (ngày) Số bịch NS tươi
(g)NS tươi TB
(g)NS khô
(g)NS khô TB
(g)CT1 18 47 1220,00 ± 5 25,96 150,00 ± 5 3,20CT2 18 42 1360,00 ± 5 32,38 165,00 ± 5 3,93CT3 18 40 1200,00 ± 5 30,00 150,00 ± 5 3,75CT4 18 38 2090,00 ± 5 55,00 250,00 ± 5 6,58CT5 18 40 1700,00 ± 5 42,50 200,00 ± 5 5,00
CTThời gian
thu quả thể (ngày)
NS tươi (g)
NS nấm tươi trung bình 1
bịch (g)
NS khô (g)
Tỉ lệ khối NS khô/NS tươi
(%)
Hiệu suất sinh hoc
(%)CT1 72 3850,00 81,91 560,00 14,55 9,63CT2 72 3500,00 83,33 500,00 14,30 8,75CT3 72 4260,00 106,50 570,00 13,38 10,65CT4 71 4890,00 128,68 650,00 13,29 2,23CT5 72 4550,00 113,75 615,00 13,52 11,38
3.3. Tỉ lệ nhiễm của quá trình nuôi trồngTrong sản xuất tỉ lệ nhiểm ảnh hưởng đến giá
trị kinh tế rất lớn, cần giảm thiểu tỉ lệ nhiễm đến mức thấp nhất. Trong quá trình nuôi trồng nấm Đầu khỉ không tránh khỏi việc các vi sinh vật có hại như nấm dại, nấm mốc... gây hại cho nấm. Nhiễm có thể do nhiều nguyên nhân: giống gốc không thuần bị nhiễm nấm dại, quá trình hấp khử trùng không đạt yêu cầu, kĩ thuật cấy không tốt, phòng cấy không đảm bảo vệ sinh, do bịch bị rách...
Tỉ lệ nhiễm của toàn bộ quá trình nuôi trồng được thể hiện dưới hình 4.
Hình 4. Tỉ lệ nhiễm của quá trình nuôi trồng

110
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Tất cả các công thức nuôi trồng đều được thực hiện trong cùng một điều kiện. Các công thức có thành phần bã dong đao CT1, CT2, CT3 có tỉ lệ nhiễm thấp, đặc biệt CT1 tỉ lệ nhiễm chiếm 6% rất thấp so với các công thức đối chứng. Có thể do nguyên liệu bã dong đao là nguyên liệu mới lấy về còn mới nên nhiễm ít hơn, còn các nguyên liệu mùn
cưa, lõi ngô, bông phế liệu là những nguyên liệu đã cũ và qua sử dụng nên chứa nhiều vi sinh vật gây hại.
Sau khi đã khảo sát một số yếu tố thích hợp cho nuôi trồng nấm Đầu khỉ trên bã dong đao; quy trình công nghệ nuôi trồng nấm Đầu khỉ đề xuất được tóm tắt như sau:
IV. KẾT LUẬN- Bã dong đao hoàn toàn phù hợp làm cơ chất
nuôi trồng nấm Đầu khỉ.- Độ ẩm cơ chất phù hợp cho nấm Đầu khỉ sinh
trưởng và phát triển là 65%.- Sau 72 ngày kết thúc nuôi trồng thử nghiệm
trên 6 công thức dinh dưỡng, kết quả về năng suất nấm tươi được ghi nhận như sau: (1) CT4 là công thức có tốc độ sợi ăn nhanh (3,3 mm/ngày) và năng suất quả thể cao nhất (4890 g nấm tươi). Tỉ lệ nhiễm cao nhất 24%; (2) CT1, CT2, quả thể có tỉ lệ hàm lượng chất khô cao lần lượt là 14,55% , 14,3%, có tỉ
lệ nhiễm thấp lần lượt là 6% và 16%; (3) CT3 có năng suất thu lần thứ nhất là cao nhất (3060 g nấm tươi).
- Đã xác định được các điều kiện thích hợp để nuôi trồng nấm Đầu khỉ trên bã dong đao, từ đó đề xuất được quy trình công nghệ nuôi trồng nấm đầu khỉ Hericium erinaceus trên bã dong đao.
LỜI CẢM ƠNCông trình nghiên cứu được hỗ trợ kinh phí từ
Đề tài Nghiên cứu ứng dụng công nghệ sinh học để chế biến và nâng cao hiệu quả kinh tế các sản phẩm từ dong đao làng nghề ĐT.08.17/CNSHCB thuộc Đề

111
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
án Phát triển và ứng dụng công nghệ sinh học trong lĩnh vực công nghiệp chế đến năm 2020.
TÀI LIỆU THAM KHẢONguyễn Lân Dũng, 2001. Công nghệ nuôi trồng nấm.
Tập 1, NXB Nông nghiệp.Trịnh Tam Kiệt, 2013. Nấm lớn ở Việt Nam. Tập 3, NXB
Khoa học tự nhiên và Công nghệ. Hà Nội.Lê Xuân Thám, 2004. Nấm trong công nghệ và chuyển
hóa môi trường. NXB Khoa học và kỹ thuật - Thành phố Hồ Chí Minh.
Khuất Hữu Trung, 2003. Kết quả bước đầu trong nghiên cứu đánh giá, tuyển chọn và thử nghiệm nuôi trồng nấm Hầu thủ. Hội nghị Công nghệ sinh học toàn quốc, Hà nội, tr. 136-140.
Trung tâm Công nghệ sinh hoc thực vật, 2012. Quy
trình công nghệ nuôi trồng nấm Đầu khỉ trên nguồn cơ chất tổng hợp. QTCN được Bộ Nông nghiệp và PTNT công nhận là tiến bộ khoa học.
Han Gyu Ko, Hyuk Gu Park, Sang Ho Park, Chang Won Choi, Seong Hwan Kim, Won Mok Park, 2005. Comparative study of mycelial growth and basidiomata formation in seven different species of the edible mushroom genus Hericium. Bioresource Technology 96, 1439-1444.
Han ZH, Ye JM, Wang GF, 2012. Sep 19, 2012. Evaluation of in vivo antioxidant activity of Hericium erinaceus polysaccharides. Int J Biol Macromol.
Md Asaduzzaman Khan, Mousumi Tania, Rui Liu và Mohammad Mijanur Rahman, 2013. Hericium erinaceus: an edible mushroom with medicinal values. Microbiol technology, 2013; 10 (1): 1-6.
Building a technological process of monkey head mushroom (Hericium erinaceus) cultivation by using Canna edulis dreg
Co Thi Thuy Van, Tran Lien Ha, Nguyen Van MinhAbstract“Monkey head” mushroom (Hericium erinaceus) is a valuable medicine with nutritous properties such as amino acids, polysacharides, lipid, minerals, vitamin as well as bioactivity compounds. Medical wise, “monkey head” mushroom is proved to amend the immune system, recover gastric ulcer, helps with inflammatory responses, antioxidant, anticarcinogenic, lower blood cholesterol, aid blood circulation. In this article, we focused on the conditions for cultivation of “Monkey head” mushroom Hericium erinaceus by using Canna edulis dreg. The best conditions for “Monkey head” mushroom growth was medium containing 85% Canna edulis dreg + 7.5% rice bran + 7.5% corn bran). The best conditions for “Monkey head” mushroom production (3060 g mushroom) was medium containing 50% Canna edulis dreg + 35% corncob + 7.5% rice bran + 7.5% corn bran. The canna edulis dreg was suitable for “Monkey head” mushroom production.Keywords: Monkey head mushroom, medicinal mushrooms, mushroom cultivation technology, Canna edulis dreg
Ngày nhận bài: 18/9/2018Ngày phản biện: 23/9/2018
Người phản biện: PGS.TS. Nguyễn Thị Minh TúNgày duyệt đăng: 15/10/2018
1 Viện Di truyền Nông nghiệp; 2 Viện Môi trường Nông nghiệp; 3 Khoa Nông học, Học viện Nông nghiệp Việt Nam
XÁC ĐỊNH KHẢ NĂNG ĐỐI KHÁNG CỦA LOÀI Chaetomium spp. VỚI NẤM Neoscytalidium dimidiatum GÂY BỆNH ĐỐM NÂU THANH LONG
Nguyễn Thế Quyết1, Nguyễn Đức Thành1, Trịnh Quốc Bình1,Bùi Thị Lan Hương2, Nguyễn Đức Huy3 và Phạm Xuân Hội1
TÓM TẮTBệnh đốm nâu thanh long do nấm Neoscytalidium dimidiatum gây ra là một trong những bệnh hại nghiêm trọng
nhất trên cây thanh long Hylocereus undatus, gây thiệt hại kinh tế rất lớn cho người trồng thanh long. Nghiên cứu này được tiến hành nhằm xác định khả năng đối kháng của loài nấm Chaetomium spp. đối với nấm N. dimidiatum gây bệnh đốm nâu thanh long bằng phương pháp đồng nuôi cấy trên môi trường thạch-dextrose-khoai tây (PDA). Kết quả nghiên cứu cho thấy, các chủng nấm Chaetomium spp. đều có hiệu lực ức chế đường kính tản nấm và sinh bào tử của nấm N. dimidiatum gây bệnh đốm nâu thanh long. Trong đó, loài Arcopilus cupreus có hiệu lực ức chế đường kính tản nấm bệnh cao nhất (77,67%) và loài C. globosum có hiệu lực ức chế sinh bào tử của nấm N. dimidiatum (79,75%) sau 14 ngày nuôi cấy.
Từ khóa: Bệnh đốm nâu, Chaetomium, đồng nuôi cấy, đối kháng, thanh long

112
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
I. ĐẶT VẤN ĐỀ Ở Việt Nam, cây thanh long (Hydrocereus
undatus) được người Pháp đưa vào trồng đã trên 100 năm (Luders and McMahon, 2006), cây thanh long đã góp phần to lớn trong việc chuyển đổi cơ cấu kinh tế, nâng cao thu nhập, phát triển nông nghiệp nông thôn với giá trị hàng hóa cao. Những năm gần đây, bệnh đốm nâu trên cây thanh long do nấm Neoscytalidium dimidiatum thuộc họ Botryosphaeriaceae, chi Neoscytalidium đã gây thiệt hại nghiêm trọng và có xu hướng ngày càng lan rộng. Bệnh này đã được ghi nhận xuất hiện rải rác đầu tiên vào năm 2008 tại Bình Thuận và Tiền Giang và đến năm 2011 trở lại đây thì bệnh tấn công mạnh và lây lan nhanh hơn. Mức độ bệnh ở các vườn, địa phương khác nhau, dao động từ 30-70%,có những vườn mất trắng năng suất do quả bị nhiễm bệnh không thể thu hoạch được, thiệt hại rất lớn cho nhà vườn trồng thanh long (Nguyễn Thành Hiếu và ctv., 2014).
Hiện nay, sử dụng các vi sinh vật đối kháng đang là một biện pháp đầy tiềm năng nhằm kiểm soát nhóm tác nhân nấm gây bệnh cây. Đã có một số nghiên cứu sử dụng vi sinh vật đối kháng với nấm Neoscytalidium dimidiatum gây bệnh cây thanh long, lan ngọc điểm (Hà Thị Thúy và ctv., 2016; Hien et al., 2017; Đỗ Thị Huỳnh Mai và Nguyễn Thị Liên, 2018). Tuy nhiên, chưa có nghiên cứu công bố sử dụng nấm Chaetomium spp. kiểm soát nấm N. dimidiatum gây bệnh đốm nâu trên thanh long.
Nấm Chaetomium spp. là một trong những loại nấm túi lớn nhất với trên 300 loài đã được mô tả (von Arx et al., 1986), đã được chứng minh sản sinh nhiều loại hợp chất có hoạt tính sinh học, có ít nhất hơn 200 loại hoạt chất được tách chiết từ các loài nấm Chaetomium spp. Các hoạt chất này có hiệu lực phòng trừ nhiều loại tác nhân gây bệnh cây (Zhang et al., 2012). Việc nghiên cứu các chủng vi sinh vật có lợi, có khả năng kiểm soát phòng ngừa bệnh đốm nâu trên cây thanh long, phục vụ canh tác thanh long theo hướng bền vững là điều cần thiết.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưuNấm Neoscytalidium dimidiatum gây bệnh đốm
nâu trên cây thanh long từ nguồn lưu trữ của Bộ môn Công nghệ vi sinh, Viện Di truyền Nông nghiệp.
Nấm đối kháng Chaetomium spp. (gồm các loài Arcopilus aureus, Arcopilus cupreus, Chaetomium cochliodes và Chaetomium globosum) được phân
lập trên đất trồng cây thanh long thu thập tại huyện Châu Thành, tỉnh Long An và huyện Chợ Gạo, tỉnh Tiền Giang.
Môi trường PDA (potato dextrose agar): Khoai tây (200 g), dextrose (20 g), agar (20 g) và nước cất (1.000 ml). Môi trường đều được hấp vô trùng ở điều kiện áp suất 1,4 atm ở điều kiện nhiệt độ 121oC trong thời gian 30 phút.
Hóa chất dùng trong thí nghiệm có nguồn gốc từ Việt Nam, Trung Quốc như agar, dextrose, NaOH,...
2.2. Phương pháp nghiên cưu
2.2.1. Đánh giá khả năng đối kháng của 4 loài nấm nghiên cứu đối với nấm N. dimidiatum bằng phương pháp đồng nuôi cấy
Khả năng đối kháng của 4 loài nấm Arcopilus aureus, Arcopilus cupreus, Chaetomium cochliodes và Chaetomium globosum với nấm N. dimidiatum được đánh giá trên môi trường PDA trong đĩa petri (loại 9 cm) theo phương pháp của Soytong (2009), các công thức thí nghiệm gồm: Công thức 1: Đối chứng nấm đối kháng; Công thức 2: Cấy nấm đối kháng + N. dimidiatum; Công thức 3: Đối chứng nấm gây bệnh. Mỗi công thức lặp lại 3 lần, mỗi lần 3 đĩa petri/lần nhắc lại.
Mỗi một đĩa cấy 2 điểm ở vị trí cách mép đĩa petri 1 cm. Hàng ngày theo dõi tốc độ phát triển của nấm đối kháng và nấm gây bệnh.
Chỉ tiêu theo dõi: Tính hiệu lực (%) ức chế phát triển (percent inhibition of radical growth, PIRG) của nấm đối kháng đối với nấm gây bệnh về đường kính tản nấm (cm) và mật số bào tử (CFU/ml) theo tài liệu của Soytong (2009).
Cách xác định số lượng bào tử: Được áp dụng phương pháp đếm bào tử trên thước đếm hồng cầu.
Cách thực hiện như sau: Cắt lấy tản nấm có kích thước 1 ˟ 1 cm2 cho vào tuýp nhựa trắng đã hấp khử trùng (loại 1,5 ml; eppendorf) có chứa 1,0 ml nước cất vô trùng và để trong thời gian 60 phút ở điều kiện nhiệt độ phòng. Sau đó, lắc nhẹ bằng tay để bào tử rời khỏi tản nấm và phát tán đều vào trong nước. Tiến hành đếm nồng độ bào tử trên buồng đếm hồng cầu, mỗi tản nấm đếm 3 lần và lấy giá trị trung bình. Nồng độ bào tử/ml chính là lượng bào tử sinh ra từ tản nấm thu được trên môi trường nuôi cấy.
2.2.2. Phương pháp tính và xử lý số liệuHiệu lực ức chế phát triển nấm (PIRG) của nấm
đối kháng đối với nấm gây bệnh được tính theo công thức:

113
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
PIRG (%) = ˟ 100R1 − R2
R1
Trong đó: R1: Đường kính/số bào tử nấm N. dimidiatum; R2: Đường kính/số bào tử nấm Chaetomium spp.
Số liệu phần trăm (%) được chuyển đổi sang dạng arcsin (x)½ trước khi xử lý thống kê. Các số liệu được phân tích thống kê dùng phầm mềm IRRISTAT 4.0.
2.3. Thời gian và địa điểm nghiên cưuNghiên cứu được thực hiện trong năm 2017 tại Bộ
môn Công nghệ vi sinh, Viện Di truyền Nông nghiệp.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Khả năng đối kháng của 4 loài nấm nghiên cưu đối với N. dimidiatum bằng phương pháp đồng nuôi cấy
Nấm Arcopilus aureus, Arcopilus cupreus, Chaetomium cochliodes và Chaetomium globosum đã xác định trước đó (Quyet et al., 2018) được đánh giá khả năng đối kháng với nấm N. dimidiatum trong điều kiện in vitro (Bảng 1).
Bảng 1. Hiệu lực ức chế đường kính tản nấm N. dimidiatum của nấm đối kháng Chaetomium spp.
bằng phương pháp đồng nuôi cấy
Ghi chú: Giá trị trung bình trong cùng một cột mang chữ cái khác nhau thì sai khác có ý nghĩa thống kê ở mức α = 0,05 với mức xác suất p < 0,05.
Kết quả thí nghiệm cho thấy, các loài nấm Chaetomium spp. đều có hiệu lực ức chế đường kính tản nấm N. dimidiatum từ 51,11% (mâu nấm A. aureus)đến 77,67% (mâu nấm A. cupreus) sau 14 ngày nuôi cấy. So sánh với kết quả nghiên cứu của một số tác giả khác cho thấy rằng, Hà Thị Thúy và cộng tác viên (2016) đã xác định được chủng B7 là loài Bacillus polyfermenticus và chủng A3 là loài Streptomyces fradiae có đường kính ức chế nấm N. dimidiatum gây bệnh đốm nâu thanh long đạt cao nhất, lần lượt là 2,10 ± 3 cm (74,44%) và 2,30 ± 3 cm (76,67%).
Hiệu lực ức chế mật số bào tử nấm N. dimidiatum của 4 loài nấm đối kháng Chaetomium spp. được trình bày ở bảng 2.
Bảng 2. Hiệu lực ức chế bào tử nấm N. dimidiatum của các dòng nấm đối kháng Chaetomium spp.
bằng phương pháp đồng nuôi cấy
Ghi chú: Giá trị trung bình trong cùng một cột mang chữ cái khác nhau thì sai khác có ý nghĩa thống kê ở mức α = 0,05 với mức xác suất p < 0,05.
Kết quả thí nghiệm cho thấy, các loài nấm Chaetomium spp. có hiệu lực ức chế khả năng sinh bào tử nấm gây bệnh N. dimidiatum từ 44,30% (mâu nấm C. cochliodes) đến 79,75% (mâu nấm C. globosum) sau 14 ngày nuôi cấy. So sánh với kết quả nghiên cứu của một số tác giả khác cho thấy rằng, nghiên cứu của tác giả Đỗ Thị Huỳnh Mai và Nguyễn Thị Liên (2018) đã xác định loài Bacillus amyloliquefaciens có hiệu lực đối kháng với nấm Neoscytalidium sp. là cao nhất (79,2%). Tác giả Hien và cộng tác viên (2017) về các dòng vi khuẩn Bacillus sp.được phân lập từ đất vùng rễ trồng thanh long có khả năng đối kháng với nấm N. dimidiatum gây bệnh đốm nâu cho kết quả cao hơn với khả năng sản sinh siderophore từ 1,83 - 0,06 cm.
Các loài nấm Chaetomium spp. đã được chứng minh là có hiệu lực ức chế đường kính tản nấm và sinh bào tử của nhiều loài nấm gây bệnh cây như Phytophthora palmivora, Colletotrichum gloeosporioides, C. lunata, R. microporus, Fusarium sp.,… gây hại trên nho, chè, cà phê, cà chua, khoai tây, lúa bằng phương pháp đồng nuôi cấy trong điều kiện in vitro (Lê Thị Ánh Hồng và ctv., 2005; Soytong, 2009; Tathan et al., 2012; Thiep and Soytong, 2015; Phong et al., 2016). Tuy nhiên, chưa có bất kỳ công bố nào trên thế giới hay ở Việt Nam sử dụng nấm Chaetomium spp. phòng chống nấm N. dimidiatum gây bệnh đốm nâu hại cây thanh long.
TT Công thưc
Sau cấy 14 ngàyĐường
kính tản nấm gây
bệnh (cm)
Giá trị PIRG (%)
1 N. dimidiatum + A. aureus 4,40 51,11c
2 N. dimidiatum + A. cupreus 2,01 77,67a
3 N. dimidiatum + C. globosum 2,13 76,33a
4 N. dimidiatum + C. cochliodes 3,11 65,44b
5 Đối chứng nấm bệnh 9,00 -LSD0,05 4,53
TT Công thưc
Sau cấy 14 ngàySố lượng
bào tử nấm gây bệnh
(˟ 107 CFU/ml)
Giá trịPIRG (%)
1 N. dimidiatum + A. aureus 19,33 64,22b
2 N. dimidiatum + A. cupreus 11,60 78,53a
3 N. dimidiatum + C. globosum 10,94 79,75a
4 N. dimidiatum + C. cochliodes 21,51 60,19c
5 N. dimidiatum 54,03 -LSD0,05 4,10

114
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luậnCác dòng nấm đối kháng Chaetomium spp. đều
có hiệu lực ức chế đường kính tản nấm và sinh bào tử của nấm N. dimidiatum gây bệnh đốm nâu thanh long. Trong đó, loài A. cupreus có hiệu lực ức chế đường kính tản nấm bệnh cao nhất (77,67%) và loài C. globosum có hiệu lực ức chế sinh bào tử của nấm (79,75%) sau 14 ngày nuôi cấy.
4.2. Đề nghịĐánh giá các đặc điểm sinh học và nghiên cứu
tách chiết các hoạt tính sinh học từ các loài nấm đối kháng Chaetomium spp. trong các thí nghiệm tiếp theo.
TÀI LIỆU THAM KHẢONguyễn Thành Hiếu, Nguyễn Ngoc Anh Thư và Nguyễn
Văn Hòa, 2014. Nghiên cứu xác định tác nhân, đặc điểm hình thái và sinh học của nấm Neoscytalium dimidiatum gây bệnh đốm trắng trên thanh long (Hylocereus undatus). Trong Hội thảo Quốc gia bệnh hại thực vật Việt Nam lần thứ 13, tổ chức tại Đại học Nông Lâm TP. Hồ Chí Minh, tr. 114-120.
Lê Thị Ánh Hồng, Nguyễn Thanh Hà, Nguyễn Thị Hằng Phương, Nguyễn Thị Thanh Nga, Nguyễn Thế Quyết, Nhữ Viết Cường, Nguyễn Thuý Mùi và Kasem Soytong, 2005. Nghiên cứu ứng dụng nấm
Chaetomium spp. trong sản xuất các chế phẩm vi sinh bảo vệ thực vật phòng chống các bệnh nấm hại. Trong Báo cáo tổng kết khoa học kỹ thuật - Đề tài cấp Nhà nước, 111 tr.
Đỗ Thị Huỳnh Mai và Nguyễn Thị Liên, 2018. Phân lập và tuyển chọn vi khuẩn đối kháng nấm Neoscytalidium sp. gây bệnh đốm trắng trên cây lan Ngọc điểm. Tạp chí Khoa học & Công nghệ Nông nghiệp, 2(1): 499-508.
Hà Thị Thúy, Lương Hữu Thành, Vũ Thúy Nga, Hưa Thị Sơn và Tống Hải Vân, 2016. Tuyển chọn chủng vi sinh vật có khả năng ức chế nấm Neoscytalidium dimitiatum gây bệnh đốm nâu thanh long. Trong Hội thảo Quốc gia về khoa học cây trồng lần thứ hai, tổ chức tại tại TP. Cần Thơ, ngày 11-12/8/2016, tr. 1167-1172.
Hien, O.T.M., Huyen, N.T.M., Anh, T.D., and Lien, N.T., 2017. Isolation and identification of antagonisticbacteria against the causative fungus of white spot disease (Neoscytalidium dimidiatum (Penz.) on dragon fruits. Journal of Science, 49: 28-41.
Luders, L. and McMahon, G., 2006. The pitaya or dragon fruit. Department of Primary Industries and Fisheries. Northern Territory, Australia, Agnote D, 42.
Quyet, N.T., Thanh, N.D., Cuong, H.V., Huong, B.T.L. and Soytong, K., 2018. Molecular identification of Chaetomium species from soil in Vietnam. International Journal of Agricultural Technology, 14(2): 225-232.
A-Nấm A. cupreusB- Nấm bệnh N. dimidiatum + A. cupreus
C-Nấm bệnh N. dimidiatum
A-Nấm C. globosumB- Nấm bệnh N. dimidiatum + C. globosum
C- Nấm bệnh N. dimidiatum
A-Nấm A. cupreusB- Nấm bệnh N. dimidiatum + A. cupreus
C-Nấm bệnh N. dimidiatum
A-Nấm C. cochliodesB- Nấm bệnh N. dimidiatum + C. cochliodes
C-Nấm bệnh N. dimidiatum
Hình 1. Đánh giá hiệu lực đối kháng của nấm Chaetomium spp. đối với nấm N. dimidiatum bằng phương pháp đồng nuôi cấy

115
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Phong, N.H., Wattanachai, P., Kasem, S. and Luu, N.T., 2014. Antimicrobial substances from Chaetomium spp. against Pestalotia spp. causing grey blight disease of tea. Journal of Agricultural Technology, 10(4): 863-874.
Soytong, K., 2009. Evaluation of Chaetomium biological fungicide to control Phytophthora stem and root rot of durian. Kasetsart J. (Nat. Sci.), 3(2): 115-124.
Tathan, S., Sibounnavong, P., Soytong, K. and Toanun C., 2012. Biological metabolites from Chaetomium
spp. to inhibit Drechslera oryzae causing leaf spot of rice. Journal of Agricultural Technology, 8(4): 1691-1701.
Thiep, N.V. and Soytong, K., 2015. Chaetomium spp. as biocontrol potential to control tea and coffee pathogens in Vietnam. International Journal of Agricultural Technology, 11(6): 1381-1392.
Von Arx J.A., Guarro, J. and Figueras, M.J., 1986. Ascomycete genus Chaetomium. Beih. Nova Hedwigia, 84: 1-162.
Identification of Chaetomium spp. antagonistic activity against Neoscytalidium dimidiatum causing brown spot disease of dragon fruit in Vietnam
Nguyen The Quyet, Nguyen Duc Thanh, Trinh Quoc Binh, Bui Thi Lan Huong, Nguyen Duc Huy and Pham Xuan Hoi
AbstractThe brown spot disease caused by Neoscytalidium dimidiatum is one of the most serious diseases of Hylocereus undatus in Vietnam, causing great economic losses for dragon fruit growers. This study was conducted to determine the antagonistic activity of Chaetomium species against N. dimidiatum causing brown spot disease on dragon fruit by using bi-culture technique on potato dextrose agar medium. The results showed that Chaetomium species had the effect of inhibiting diameter and spore formation of N. dimidiatum fungus. Among them, Arcopilus cupreus had the highest inhibition of colony growth (77.67%) and C. globosum had the highest inhibition of spore formation (79.75%) after 14 days of culture. Keywords: Antagonistic, bi-culture, brown spot, Chaetomium, dragon fruit
Ngày nhận bài: 18/9/2018Ngày phản biện: 27/9/2018
Người phản biện: TS. Trần Thị Mỹ HạnhNgày duyệt đăng: 15/10/2018
1 Ban Quản lý Lăng Chủ tịch Hồ Chí Minh; 2 Học viện Nông nghiệp Việt Nam3 Viện Di truyền Nông nghiệp
ĐÁNH GIÁ ĐA DẠNG NGUỒN GEN ĐỖ QUYÊN BẰNG CHỈ THỊ ISSRĐỗ Thị Thu Lai1, Nguyễn Thị Thùy Linh2, Đinh Trường Sơn2,
Nguyễn Thị Kim Lý3, Phạm Thị Minh Phượng2
TÓM TẮTSự đa dạng nguồn gen của 8 mâu đỗ quyên thu thập được từ các tỉnh Lào Cai, Vĩnh Phúc, Nam Định đã được
phân tích dựa trên 22 chỉ thị phân tử ISSR; nhân bản được 954 sản phẩm PCR thuộc 200 locus từ 8 mâu giống đỗ quyên. Hệ số tương đồng di truyền của 8 mâu đỗ quyên dao động từ 49,0 - 86,2%. Mức độ đa dạng của 8 mâu đỗ quyên ở mức trung bình với giá trị PIC là 0,24. Mối quan hệ di truyền của 8 mâu đỗ quyên được phân tích dựa trên Hệ số tương đồng Sokal and Michener. Kết quả phân nhóm bằng thuật toán UPGMA sử dụng phần mềm NTSYS 2.1 cho thấy: ở mức độ tương đồng di truyền là 75%, 8 mâu đỗ quyên được phân tách thành 4 nhóm khác nhau trong đó có nhóm 1 bao gồm 4 mâu Q1, Q2, Q4 và Q5, nhóm 2 bao gồm hai mâu Q3 và Q6, nhóm 3 chỉ có mâu Q7 và nhóm 4 chỉ có mâu Q8. Sự tương đồng di truyền của mâu Q8 so với 7 mâu còn lại là khá thấp. Chính vì vậy, có thể sử dụng mâu Q8 (là mâu hiện đang được ưa chuộng trên thị trường) để lai với 7 mâu còn lại nhằm phát triển nguồn gen phục vụ công tác chọn tạo giống hoa đỗ quyên mới ở Việt Nam.
Từ khóa: Đỗ quyên, Rhododendron, đa dạng nguồn gen, chỉ thị phân tử, ISSR
I. ĐẶT VẤN ĐỀChi đỗ quyên (Rhododendron) là một chi lớn với
khoảng 1.025 loài và hầu hết các loài đều cho hoa đẹp rực rỡ (Chamberlain et al., 1996; Furbee, 2009).
Trên thế giới, chi đỗ quyên phân bố rất rộng, xuất hiện ở hầu khắp Bắc bán cầu tới Nam bán cầu, vùng Đông Nam Á, các quần đảo và lãnh thổ và phổ biến rộng từ Nam Himalaya tới Tây Nam Trung Quốc.

116
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Rất nhiều loài xuất hiện ở vùng núi của Thái Lan, Việt Nam và Malaysia (Irving and Hebda, 1993).
Ở Việt Nam, chi Rhododendron có 44 loài (Nguyễn Thị Thanh Hương và ctv., 2009). Trong tự nhiên, có thể tìm thấy một số loài thuộc chi này ở các vùng núi như Bạch Mã, Nam Đông, đầu nguồn sông Bồ, rừng Hương Nguyên, sông A Sáp, Sa Pa (Lào Cai), Tam Đảo (Vĩnh Phúc) và Đà Lạt (Lâm Đồng). Ở nước ta, loài được sử dụng nhiều là Rhododendron simsii vì hoa đẹp, màu sắc rực rỡ (Mai Văn Phô, 2009). Mặc dù là cây hoa quý và phổ biến, cho đến nay, có rất ít tài liệu trong nước công bố về các dữ liệu phân tích đa dạng nguồn gen đỗ quyên phân bố ở nước ta.
Chỉ thị DNA có khả năng ứng dụng rất lớn trong phân tích đa dạng di truyền cũng như ứng dụng trong chọn tạo giống. Cho đến nay, hàng loạt chỉ thị phân tử đã được phát minh và ứng dụng rộng rãi trong đó có chỉ thị ISSR (Collard and Mackill, 2008). Chỉ thị ISSR dựa trên phản ứng PCR nhằm khuếch đại ngâu nhiên các đoạn DNA ở các vị trí khác nhau trong bộ DNA genome. Do sự đa hình về trình tự DNA giữa các cá thể dân tới vị trí bắt mồi là khác
nhau và do vậy các locus được khuếch đại của các cá thể có thể cũng khác nhau. Chính vì vậy, ISSR là chỉ thị có mức độ đa hình cao và được sử dụng nhiều trong nghiên cứu đa dạng di truyền giữa các loài thực vật (Collard and Mackill, 2008; Charrier et al., 2014; Xu et al., 2017).
Việc khảo sát, đánh giá mức độ đa hình của các mâu đỗ quyên là cần thiết. Với mục đích phân tích mối quan hệ di truyền của các mâu đỗ quyên thu thập được, công trình này đã sử dụng chỉ thị phân tử ISSR nhằm phát hiện mức độ đa hình của 8 mâu đỗ quyên thu thập được. Kết quả nghiên cứu sẽ góp phần phân loại, nhận dạng, bảo tồn, khai thác hiệu quả nguồn gen cũng như chọn tạo giống hoa đỗ quyên mới tại Việt Nam.
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưuVật liệu nghiên cứu gồm 08 loài đỗ quyên bản địa
được thể hiện trong Bảng 1. Bên cạnh đó, hình ảnh các vật liệu nghiên cứu địa điểm thu thập và ký hiệu mâu được trình bày trên Hình 1.
2.2. Phương pháp nghiên cưu- Phương pháp đánh giá đa dạng di truyền bằng
chỉ thị ISSR.Tách chiết DNA: Khoảng 100 mg lá của 8 mâu
đỗ quyên được tách chiết theo hướng dân của bộ kit NucleoSpin Plant II, Macherey-Nagel. Hàm lượng và độ nguyên vẹn của các mâu DNA sau khi tách chiết được kiểm tra bằng phương pháp điện di trên agarose 1,0%. Các mâu DNA sau đó được bảo quản trong tủ lạnh sâu –200 C.
Thực hiện phản ứng PCR: Phản ứng PCR được thực hiện trên máy chu trình nhiệt (Eppendorf) với
thể tích 20 µL chứa 50 ng DNA, 0,2 mM mỗi loại dNTP, 0,5 µM mồi, 1,5 mM MgCl2 và 0,5U Taq DNA Polymerase (D1086-Sigma). Chu trình của phản ứng PCR được thực hiện với điều kiện như sau: giai đoạn biến tính ban đầu 94oC trong 5 phút; 40 chu kỳ gồm 1 phút biến tính ở 94oC, 1 phút gắn mồi ở nhiệt độ Tm (Bảng 1) và 2 phút kéo dài ở 72oC. Phản ứng PCR kết thúc với một chu kì kéo dài trong 7 phút ở 72oC và bảo quản ở 4oC (Barcaccia et al., 2003). Thông tin cụ thể của các mồi được liệt kê trên Bảng 2.Sản phẩm PCR được kiểm tra trên gel agarose 1,5% và quan sát dưới đèn chiếu UV.
Hình 1. Các C. ﷽﷽﷽﷽﷽﷽﷽﷽﷽﷽﷽﷽ cuHmâu hoa đỗ quyên (ký hiệu từ Q1 - Q8) sử dụng trong nghiên cứu
Bảng 1. Các mâu đỗ quyên sử dụng trong nghiên cứu
Ký hiệu Tên Khoa hoc Tên địa phương Nơi thu thậpQ1 Rhododendron sp. Đỗ quyên trắng Sa Pa, Lào CaiQ2 Rhododendron lyi Le Đỗ quyên trắng xanh Sa Pa, Lào CaiQ3 Rhododendron sp. Đỗ quyên hồng đậm Tam Đảo, Vĩnh PhúcQ4 Rhododendron sp. Đỗ quyên hồng nhạt Tam Đảo, Vĩnh PhúcQ5 Rhododendron chpaensis P. Dop Đỗ quyên tím đậm Tam Đảo, Vĩnh PhúcQ6 Rhododendron sp. Đỗ quyên tím nhạt Nam Điền, Nam ĐịnhQ7 Rhododendron sp. Đỗ quyên đỏ Sa Pa, Lào CaiQ8 Rhododendron simsii Planch Đỗ quyên cà rốt Tam Đảo, Vĩnh Phúc

117
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Hình 1. Các mâu hoa đỗ quyên (ký hiệu từ Q1 - Q8) sử dụng trong nghiên cứu

118
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Bảng 2. Tên, trình tự và nhiệt độ bắt mồi của các chỉ thị ISSR sử dụng trong nghiên cứu
- Xử lý số liệu: Sử dụng “Hệ số tương đồng Sokal and Michener (1958)” và phương pháp phân nhóm UPGMA trong NTSYS 2.1. Chỉ số đa hình PIC (Polymorphic Information content) của mỗi locus được tính theo công thức: PIC (i) = 1 – Σ Pij2
(Sokal and Michener, 1958; Botstein et al., 1980). Trong đó, i là số locus ISSR tương ứng; j là số thứ tự allen của locus ISSR và Pij là tần suất allen thứ j với locus ISSR thứ i. Chỉ số sai khác giữa các cặp mồi Rp (resolving power) được tính theo công thức: Rp = ∑Ib. Trong đó Ib là giá trị đại diện cho thông tin của đoạn. Ib = 1 – [2 ˟ (0,5 – p)], p là tỷ lệ các mẫu xuất hiện băng vạch (Prevost and Wilkinson, 1999).
2.3. Thời gian và địa điểm nghiên cưu Nghiên cứu được thực hiện năm 2015 - 2016 tại
Bộ môn Công nghệ sinh học thực vật, Khoa Công nghệ sinh học, Học viện Nông nghiệp Việt Nam.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Kết quả phân tích đa dạng di truyền bằng chỉ thị ISSR
Kết quả ở Bảng 3 cho thấy: 22 chỉ thị ISSR đã
phát hiện được tổng số 200 locus với tỷ lệ đa hình của các locus trung bình đạt 74,1%. Với 8 mâu đỗ quyên thu thập được, 22 chỉ thị ISSR đã nhân được tổng số là 954 băng vạch DNA, trung bình đạt 43,36 băng vạch/mồi, 119,25 băng vạch DNA/mâu, trung bình đạt 5,42 băng vạch/mồi/mâu. Nhóm các chỉ thị UBC_808, UBC_876, ISSR-T1, ISSR-T3, UBC_811 cho số băng DNA trung bình tính trên các mâu đỗ quyên đạt cao nhất, tương ứng 8,00 - 10,13 băng/mâu. Nhóm chỉ thị ISSR cho số băng DNA trung bình trên mâu thấp nhất là UBC_873, UBC_862, UBC_824, cho số băng DNA trung bình thấp nhất tương ứng là 1,63 - 2,00 băng/mâu. Chỉ số đa hình PIC trung bình đạt 0,24 và chỉ số sai khác giữa các cặp mồi (Rp) là 10,84. Hệ số tương đồng Sokal and Michener (1958) đã được sử dụng để xác định mối tương đồng di truyền giữa 8 mâu đỗ quyên. Kết quả được thể hiện trên Bảng 4.
Từ kết quả phân tích hệ số tương đồng di truyền của 8 mâu đỗ quyên, sử dụng phương pháp phân nhóm UPGMA trong NTSYS 2.1, mối quan hệ di truyền của 8 mâu đỗ quyên được trình bày ở Hình 2.
Kết quả ở Bảng 3 và Hình 2 cho thấy: Khi được phân tích bởi 22 chỉ thị ISSR, ở mức độ tương đồng di truyền là 75% thì 8 mâu đỗ quyên được phân tách thành 4 nhóm khác nhau trong đó có nhóm 1 bao gồm 4 mâu Q1, Q2, Q4 và Q5, nhóm 2 bao gồm hai mâu Q3 và Q6, hai mâu còn lại là Q7 và Q8 được tách biệt thành 2 nhóm độc lập nhau là nhóm 3 và nhóm 4. Sự tương đồng di truyền của các mâu đỗ quyên có sự tương quan với màu sắc. Rõ ràng, các mâu Q1 và Q2 có màu sắc gần giống nhau nhất trong số các mâu (đều sáng màu) có hệ số tương đồng di truyền tới 86,2%, mâu Q8 có sự khác biệt rõ rệt về màu sắc với các mâu còn lại nên có hệ số tương đồng di truyền là nhỏ nhất với 7 mâu còn lại, trong đó mức độ tương đồng di truyền của mâu Q7 và Q8 chỉ đạt 49,0% (Bảng 2).
Kết quả phân tích cho hệ số PIC của chỉ thị chỉ đạt 0,24 chứng tỏ mức độ đa dạng của 8 mâu đỗ quyên là ở mức trung bình. Thực tế thì ngoài hai mâu Q7 và Q8 có sự khác biệt lớn thì các mâu còn lại có sự khác biệt khá nhỏ. Cụ thể, sự khác biệt di truyền giữa mâu Q3 và Q6 là 10,5%, Q2 và Q4 là 11,2% và Q1 và Q2 là 11,4%, Q1 và Q4 là 14,4%. Do mâu Q8 (Rhododendron simsii Planch) là mâu có kiểu hình khá đặc biệt (lá rụng khi ra hoa nên trên cây chỉ còn hoa do vậy được thị trường rất ưa chuộng) nên có thể sử dụng Q8 để lai tạo với 7 mâu còn lại nhằm tạo nguồn vật liệu, phục vụ công tác chọn tạo giống hoa đỗ quyên mới.
Tên mồi Trình tự mồi (5’ - 3’)Nhiệt độ bắt mồi
(oC)UBC_807 AGAGAGAGAGAGAGAGT 49UBC_808 AGAGAGAGAGAGAGAGC 50UBC_811 GAGAGAGAGAGAGAGAC 50UBC_812 GAGAGAGAGAGAGAGAA 50UBC_813 CTCTCTCTCTCTCTCTT 49UBC_817 CACACACACACACACAA 49UBC_818 CACACACACACACACAAG 56UBC_819 GTGTGTGTGTGTGTGTA 50UBC_823 TCTCTCTCTCTCTCTCC 50UBC_824 TCTCTCTCTCTCTCTCG 50UBC_827 ACACACACACACACACG 56UBC_847 CACACACACACACACAGC 56UBC_862 AGCAGCAGCAGCAGCAGC 56UBC_864 ATGATGATGATGATGATG 45UBC_866 CTCCTCCTCCTCCTCCTC 56UBC_868 GAAGAAGAAGAAGAAGAA 45UBC_869 GTTGTTGTTGTTGTTGTT 45UBC_872 GATAGATAGATAGATA 35UBC_873 GACAGACAGACAGACA 45UBC_876 GATAGATAGACAGACA 42ISSR-T1 GTGTGTGTGTGTCC 54ISSR-T3 ACACACACACACCG 42
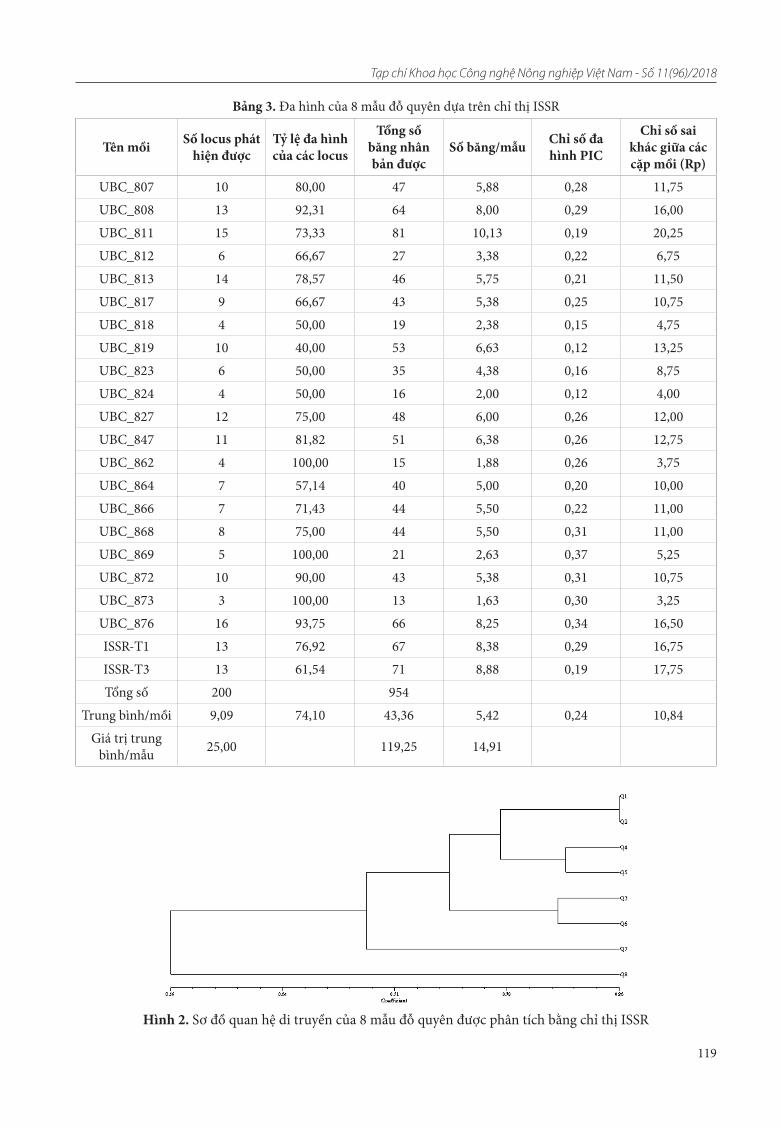
119
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Bảng 3. Đa hình của 8 mâu đỗ quyên dựa trên chỉ thị ISSR
Tên mồi Số locus phát hiện được
Tỷ lệ đa hình của các locus
Tổng số băng nhân bản được
Số băng/mẫu Chỉ số đa hình PIC
Chỉ số sai khác giữa các cặp mồi (Rp)
UBC_807 10 80,00 47 5,88 0,28 11,75
UBC_808 13 92,31 64 8,00 0,29 16,00
UBC_811 15 73,33 81 10,13 0,19 20,25
UBC_812 6 66,67 27 3,38 0,22 6,75
UBC_813 14 78,57 46 5,75 0,21 11,50
UBC_817 9 66,67 43 5,38 0,25 10,75
UBC_818 4 50,00 19 2,38 0,15 4,75
UBC_819 10 40,00 53 6,63 0,12 13,25
UBC_823 6 50,00 35 4,38 0,16 8,75
UBC_824 4 50,00 16 2,00 0,12 4,00
UBC_827 12 75,00 48 6,00 0,26 12,00
UBC_847 11 81,82 51 6,38 0,26 12,75
UBC_862 4 100,00 15 1,88 0,26 3,75
UBC_864 7 57,14 40 5,00 0,20 10,00
UBC_866 7 71,43 44 5,50 0,22 11,00
UBC_868 8 75,00 44 5,50 0,31 11,00
UBC_869 5 100,00 21 2,63 0,37 5,25
UBC_872 10 90,00 43 5,38 0,31 10,75
UBC_873 3 100,00 13 1,63 0,30 3,25
UBC_876 16 93,75 66 8,25 0,34 16,50
ISSR-T1 13 76,92 67 8,38 0,29 16,75
ISSR-T3 13 61,54 71 8,88 0,19 17,75
Tổng số 200 954
Trung bình/mồi 9,09 74,10 43,36 5,42 0,24 10,84
Giá trị trung bình/mâu 25,00 119,25 14,91
Hình 2. Sơ đồ quan hệ di truyền của 8 mâu đỗ quyên được phân tích bằng chỉ thị ISSR

120
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Bảng 4. Hệ số tương đồng di truyền của 8 mâu đỗ quyên
Trong một số năm gần đây, đã có một số công trình nghiên cứu nhằm phát triển chỉ thị phân tử đặc hiệu cho chi đỗ quyên đặc biệt là loài Rhododendron simsii. Li và cộng tác viên (2016) đã phát triển được 7 cặp mồi EST-SSR và ứng dụng trong phân tích đa dạng nguồn gen của 32 mâu đỗ quyên thuộc loài R. simsii thu thập tại tỉnh Hồ Bắc, Trung Quốc (Li et al., 2016). Shuzhen và cộng tác viên (2017) cũng phát triển được 14 chỉ thị microsatellite cho loài Rhododendron simsii.
Việc phân tích đa dạng di truyền của một số mâu đỗ quyên thuộc loài Rhododendron simsii phân bố ở Việt Nam đã được tác giả Nguyễn Thị Thu Hằng (2011) thực hiện. Tác giả cũng so sánh sự đa dạng di truyền của các mâu đỗ quyên Việt Nam với các cá thể đỗ quyên cùng loài của Nhật Bản và nghiên cứu khả năng lai tạo giữa các mâu hoa đỗ quyên Việt Nam và Nhật Bản nhằm phát triển giống hoa đỗ quyên mới (Nguyễn Thị Thu Hằng, 2011). Kết quả nghiên cứu này được thực hiện trên 8 mâu đỗ quyên thuộc nhiều loài khác nhau (mâu Q2 thuộc loài Rhododendron lyi Le, mâu Q5 thuộc loài Rhododendron chapaensis P.Dop, mâu Q8 thuộc loài Rhododendron simsii, các mâu Q1, Q3, Q4, Q6 và Q7 chưa được định danh loài) và sẽ là những dân liệu quý, góp phần vào mục tiêu bảo tồn, phát triển nguồn gen, phục vụ công tác lai tạo giống đỗ quyên mới tại Việt Nam.
IV. KẾT LUẬNMối quan hệ di truyền của 8 mâu đỗ quyên được
thu thập tại Lào Cai, Vĩnh Phúc và Nam Định đã được phân tích bởi 22 chỉ thị ISSR. Đã nhân bản được 954 sản phẩm PCR thuộc 200 locus. Hệ số tương đồng di truyền của 8 mâu đỗ quyên dao động từ 49,0 - 86,2%. Mức độ đa dạng của 8 mâu đỗ quyên ở mức trung bình với giá trị PIC là 0,24. Mối quan
hệ di truyền của 8 mâu đỗ quyên được phân tích dưa trên Hệ số tương đồng Sokal and Michener (1958). Kết quả phân nhóm bằng thuật toán UPGMA trong phần mềm NTSYS 2.1 cho thấy: ở mức độ tương đồng di truyền là 75%, 8 mâu đỗ quyên được phân tách thành 4 nhóm khác nhau trong đó có nhóm 1 bao gồm 4 mâu Q1, Q2, Q4 và Q5, nhóm 2 bao gồm hai mâu Q3 và Q6, nhóm 3 chỉ có mâu Q7 và nhóm 4 chỉ có mâu Q8. Sự tương đồng di truyền của mâu Q8 là mâu hiện đang được ưa chuộng trên thị trường so với 7 mâu còn lại là khá thấp. Chính vì vậy, có thể sử dụng mâu Q8 để lai với 7 mâu còn lại nhằm phát triển nguồn gen phục vụ công tác chọn tạo giống hoa đỗ quyên mới ở Việt Nam.
TÀI LIỆU THAM KHẢONguyễn Thị Thu Hằng, 2011. Studies on genetic diversity
and utilization of Rhododendron simsii distributed in Vietnam. Doctoral degree, Kyushu University, Fukuoka.
Nguyễn Thị Thanh Hương, Trần Minh Hợi, Nguyễn Tiến Hiệp, 2009. Một số loài có giá trị làm cảnh trong Chi Đỗ quyên (Rhododendron L.) thuộc họ Đỗ quyên (Ericaceae Juss.) ở Việt Nam. Báo cáo Hội nghị Khoa học toàn quốc về sinh thái và tài nguyên sinh vật lần thứ ba, Hà Nội.
Mai Văn Phô, 2009. Hoa đỗ quyên ở Việt Nam. Tạp chí Nghiên cứu và Phát triển, số 2.2009.
Barcaccia G, Lucchin M and Parrini P, 2003. Characterization of a flint maize (Zea mays var. indurata) Italian landrace, II. Genetic diversity and relatedness assessed by SSR and Inter-SSR molecular markers. Genetic Resources and Crop Evolution, 50: 253-271.
Botstein D, White RL, Skolnick M, Davis RW, 1980. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. American Journal of Human Genetics, 32: 314-331.
Chamberlain D, Hyam R, Argent G, Fairweather G, Walter KS, Royal Botanic Garden E, 1996. The genus Rhododendron: It’s classification & synonymy. Royal Botanic Gardens, Kew.
Charrier O, Dupont P, Pornon A, Escaravage N, 2014. Microsatellite marker analysis reveals the complex phylogeographic history of Rhododendron ferrugineum (Ericaceae) in the Pyrenees. PLoS One 9: e92976.
Collard BC, Mackill DJ, 2008. Marker-assisted selection: an approach for precision plant breeding in the twenty-first century. Philos Trans R Soc Lond B Biol Sci, 363: 557-572.
Mẫu Q1 Q2 Q3 Q4 Q5 Q6 Q7 Q8Q1 1,000Q2 0,862 1,000Q3 0,760 0,750 1,000Q4 0,821 0,816 0,776 1,000Q5 0,750 0,745 0,755 0,827 1,000Q6 0,704 0,684 0,821 0,801 0,760 1,000Q7 0,730 0,663 0,643 0,724 0,689 0,709 1,000Q8 0,577 0,582 0,571 0,582 0,597 0,536 0,490 1,000

121
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Furbee B, 2009. CHAPTER 47 - Neurotoxic Plants. In MR Dobbs, ed, Clinical Neurotoxicology. W.B. Saunders, Philadelphia, pp 523-542.
Irving E, Hebda R, 1993. Concerning the origin and distribution of Rhododendrons. The American Rhododendron Society 47.
Li Z, Cheng C, Zhang G, Fang Y, Jin W, Wang S, 2016. EST-SSR marker-based genetic diversity analysis of Rhododendron simsii germplasm in Guifeng mountain. Agricultural Science & Technology, 17: 1073-1076.
Prevost A, Wilkinson MJ, 1999. A new system of comparing PCR primers applied to ISSR fingerprinting of potato cultivars. Theoretical and
Applied Genetics, 98: 107-112.Shuzhen W, Zhiliang L, Weibin J, Fu X, Jun
X, Yuanping F, 2017. Development and characterization of polymorphic microsatellite markers in Rhododendron simsii (Ericaceae). Plant Species Biology, 32: 100-103.
Sokal RR, Michener CD, 1958. A statistical method for evaluating systematic relationships. Univ. Kans. Sci. Bull, 28: 1409-1438.
Xu JJ, Zhang LY, Zhao B, Shen HF, 2017. Assessment of genetic diversity among six populations of Rhododendron triflorum in Tibet using ISSR and AFLP markers. South African Journal of Botany, 108: 175-183.
Evaluation of genetic diversity of Rhododendron by ISSR markersDo Thi Thu Lai, Nguyen Thi Thuy Linh, Dinh Truong Son,
Nguyen Thi Kim Ly, Pham Thi Minh PhuongAbstractThe genetic diversity of eight Rhododendron individuals collected from Lao Cai, Vinh Phuc and Nam Dinh provinces was evaluated by 22 ISSR markers. 22 ISSR markers detected a total of 200 loci with 954 DNA bands. The genetic similarity of 8 individuals varied from 49.0 - 86.2% with average PIC value of 0.24 and the diversity of eight individuals was at medium level. A dendrogram obtained by using UPGMA cluster analysis showed that 8 individuals were separated into 3 distinct clusters at the genetic similarity of 75%; group 1 consisted of 6 individuals (Q1, Q2, Q3, Q4, Q6 and Q7), group 2 and group 3 each had only 1 individual (Q5 or Q8). Q8 individual had the lowest genetic similarity with the other seven; therefore, it is possible to use Q8 for crossing with 7 other individuals for new cultivar development in Vietnam.Keywords: Rhododendron, genetic diversity, molecular marker, Inter Simple Sequence Repeat
Ngày nhận bài: 16/10/2018Ngày phản biện: 24/10/2018
Người phản biện: TS. Trần Thị Thu HoàiNgày duyệt đăng: 15/11/2018
1 Khoa Thủy sản, Đại học Cần Thơ
HIỆN TRẠNG KHAI THÁC VÀ QUẢN LÝ NGHÊ LƯỚI KÉO ĐƠN VEN BỜ Ở TỈNH SÓC TRĂNG VÀ BẾN TRE
Nguyễn Thanh Long1, Trần Đắc Định1 và Mai Viết Văn1
TÓM TẮTNghiên cứu hiện trạng khai thác và quản lý nghề lưới kéo đơn ven bờ được thực hiện từ tháng 10/2017 đến tháng
5/2018 ở tỉnh Sóc Trăng và Bến Tre thông qua phỏng vấn trực tiếp 90 hộ làm nghề đánh bắt bằng lưới kéo về khía cạnh kỹ thuật và tài chính. Kết quả cho thấy, nghề lưới kéo là nghề có số lượng tàu nhiều nhất ở hai tỉnh. Nghề lưới kéo có thể khai thác quanh năm. Công suất tàu lưới kéo ở tỉnh Sóc Trăng (39,20 CV) và ở tỉnh Bến Tre (34,87 CV). Sản lượng và tỉ lệ cá tạp của nghề lưới kéo đơn ở tỉnh Sóc Trăng (16,7 tấn/năm, 39,7%) cao hơn ở tỉnh Bến Tre (15,5 tấn/năm, 33,3%) và lợi nhuận và tỉ suất lợi nhuận của nghề lưới kéo đơn ở tỉnh Sóc Trăng (6,28 triệu đồng/chuyến; 0,44 lần) cũng cao hơn ở tỉnh Bến Tre (3,38 triệu đồng/chuyến; 0,38 lần). Để nghề lưới kéo đơn ven bờ phát triển ổn định cần đẩy mạnh công tác quản lý và phát triển nguồn lợi thủy sản, tạo điều kiện cho ngư dân tiếp cận vốn với lãi suất thấp để đầu tư sản xuất và tập huấn ngư dân biết sử dụng các thiết bị khai thác nhằm tăng hiệu quả khai thác của họ.
Từ khóa: Bến Tre, khai thác thủy sản, lưới kéo, quản lý thủy sản, Sóc Trăng

122
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
I. ĐẶT VẤN ĐỀViệt Nam là quốc gia có tiềm năng phát triển về
kinh tế biển, đặc biệt là ngành thủy sản ven biển. Việt Nam có 3.260 km bờ biển, 12 đầm phá, eo biển và vịnh, 112 cửa sông và hàng ngàn đảo lớn nhỏ trải dài dọc theo bờ biển; cùng với hệ thống sông ngòi chằng chịt và các hồ chứa tạo nên tiềm năng to lớn về khai thác thủy sản và diện tích mặt nước cho nuôi trồng thủy sản (Lê Trần Nguyên Hùng, 2009).
Đồng bằng sông Cửu Long (ĐBSCL) với bờ biển dài trên 780 km chiếm 23% chiều dài bờ biển cả nước, vùng kinh tế đặc quyền khoảng 297.000 km2, giáp biển Đông và biển Tây Nam Bộ, vùng thềm lục địa có thế mạnh về thủy sản với trữ lượng thủy sản ước tính trên 2 triệu tấn và khả năng khai thác khoảng 830.000 tấn/năm, rất thuận lợi cho phát triển kinh tế biển (Lê Văn Ninh, 2006). ĐBSCL có những đóng góp rất lớn đối với cả nước với sản lượng khai thác hàng năm dân đầu cả nước và sản lượng năm 2016 là 1,29 triệu tấn chiếm 40,7% sản lượng thủy sản khai thác của cả nước (Tổng cục Thống kê, 2017).
Tỉnh Sóc Trăng và Bến Tre là hai trong bốn tỉnh ven biển vùng cửa sông Cửu Long có các hoạt động khai thác thủy sản đa dạng và biến đổi phức tạp, phần lớn là tự phát nhằm đáp ứng theo nhu cầu kiếm sống của người dân trong vùng. Để quản lý tốt các hoạt động thủy sản ven biển và quyết định những định hướng phát triển ổn định cho vùng ven biển thì cần phải nghiên cứu các giải pháp quản lý cho vùng này. Chính vì vậy việc nghiên cứu hiện trạng khai thác và quản lý nghề lưới kéo ở hai tỉnh này là cần thiết cho việc quản lý và phát triển ổn định các hoạt động khai thác thủy sản vùng cửa sông Cửu Long.
II. ĐỐI TƯỢNG VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Đối tượng nghiên cưuĐối tượng nghiên cứu là ngư dân làm nghề khai
thác lưới kéo đơn (tàu < 60 CV) ở vùng ven biển tỉnh Sóc Trăng và Bến Tre.
Các nghề khai thác ven bờ chủ yếu ở các tỉnh ven biển ở vùng cửa sông Cửu Long là nghề lưới kéo, nghề lưới rê và cá nghề khác như nghề lưới đáy, nghề rập xếp, nghề cào khung (trong sông), nghề câu và nghề xiệp… Trong nghiên cứu này tập trung khảo sát hoạt động khai thác của các nghề khai thác ven bờ có số lượng tàu và sản lượng cao nhất đó là nghề lưới kéo.
2.2. Phương pháp nghiên cưu
2.2.1. Thông tin thứ cấp Các số liệu thứ cấp được thu thập từ các nghiên
cứu, báo cáo của các cơ quan địa phương, sách báo, tạp chí và các website có liên quan… về các thông tin gồm: Điều kiện tự nhiên, tình hình phát triển khai thác thủy sản, số tàu theo từng loại nghề, sản lượng khai thác thủy sản của các tỉnh…
2.2.2. Thông tin sơ cấpSố liệu sơ cấp được thu thập từ phỏng vấn trực
tiếp 90 ngư dân làm nghề khai thác lưới kéo đơn ở tỉnh Sóc Trăng và Bến Tre (mỗi tỉnh 45 mâu) theo bảng câu hỏi đã soạn sẵn với những nội dung chủ yếu gồm: (i) Hiện trạng khai thác thủy sản: Ngư trường và mùa vụ khai thác thủy sản; kích thước tàu khai thác, ngư cụ, sản lượng khai thác, hiện trạng quản lý nghề khai thác thủy sản,… và (ii) Hiệu quả tài chính: Chi phí cố định, chi phí biến đổi, lợi nhuận, tỉ suất lợi nhuận.
2.2.3. Phương pháp xử lý số liệuSố liệu phỏng vấn được sử dụng phần mềm SPSS
for Windows để nhập số liệu và phân tích. Các kết quả được thể hiện qua thống kê mô tả như: tần số xuất hiện, giá trị trung bình và độ lệch chuẩn.
2.3. Thời gian và địa điểm nghiên cưuNghiên cứu được thực hiện từ tháng 10/2017 đến
tháng 5/2018 tại các huyện ven biển 2 tỉnh Sóc Trăng và Bến Tre.
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Các loại nghề khai thác ven bờ vùng nghiên cưu Các nghề khai thác thủy sản chủ yếu ở các tỉnh
Sóc trăng và Bến Tre là nghề lưới kéo (57,1%), nghề lưới rê (25,7%) và các nghề khác (17,2%). Các nghề khai thác khác bao gồm các nghề như nghề lưới đáy, nghề rập xếp, nghề cào khung (trong sông), nghề câu và nghề xiệp. Qua đây cho thấy nghề lưới kéo có vai trò quan trọng trong lĩnh vực khai thác thủy sản ở hai tỉnh Sóc Trăng và Bến Tre vì nó có tỉ lệ tàu cao nhất.
Bảng 1. Các nghề khai thác thủy sản ở tỉnh Sóc Trăng và Bến Tre
Nghề khai thác thủy sản
Bến Tre
Sóc Trăng Tổng Tỉ lệ
(%)Nghề lưới kéo 1.003 415 1418 57,1Nghề lưới rê 439 200 639 25,7Nghề khác (rập xếp, nghề đáy, cào khung, nghề câu, xiệp…)
190 237 427 17,2
Tổng 1.632 852 2.484 100

123
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
3.2. Hiện trạng khai thác của nghề lưới kéo
3.2.1. Khía cạnh ky thuật của nghề lưới kéoNghề lưới kéo đơn ven bờ tỉnh tỉnh Sóc Trăng và
Bến Tre khai thác quanh năm, những tháng có sản lượng cao tập trung vào 2 mùa từ tháng 2 đến tháng 5 và từ tháng 10 đến tháng 12.
Thời gian khai thác một chuyến biển của nghề lưới kéo đơn ở vùng này tương đối ngắn vì tàu có tải trọng và công suất tàu nhỏ (Bảng 3). Thời gian một chuyến biển từ 4 - 5 ngày tùy thuộc tàu lớn hay nhỏ và tàu khai thác xa hay gần bờ. Tàu lưới kéo đơn ở Bến Tre có công suất nhỏ hơn tàu ở Sóc Trăng nên có thời gian chuyến biển ngắn, nhưng số chuyến biển trong tháng lại cao hơn (Bảng 2). Nhìn chung, tàu lưới kéo đơn ở hai tỉnh này có thể khai thác quanh năm, trừ thời tiết xấu tàu không thể ra khơi để đánh cá được. Trung bình một năm nghề lưới kéo đơn ở tỉnh Sóc Trăng có thể khai thác 8,33 tháng và ở tỉnh Bến Tre là 8,78 tháng (Bảng 2).
Bảng 2. Thời gian khai thác của nghề lưới kéo đơn
Bảng 3. Công suất và tải trọng của tàu lưới kéo đơn
Trung bình mỗi tàu lưới kéo đơn cần 3,67 ± 0,80 lao động (Sóc Trăng) và 3,33 ± 0,90 lao động (Bến Tre). Số lao động của tàu lưới kéo đơn ở Bến Tre thấp hơn tàu ở tỉnh Sóc Trăng là do qui mô của tàu và lưới nhỏ hơn. Mặc dù vậy, nghề lưới kéo đơn không những tạo việc làm cho gia đình mà còn tạo việc làm cho người dân sống vùng cửa sông Cửu Long. Số lao động thuê mướn của tàu lưới kéo đơn ở tỉnh Bến tre là 1,56 ± 0,97 lao động và tỉnh Sóc Trăng là 1,33 ± 0,66 lao động (Bảng 4).
Bảng 4. Lực lượng lao động của tàu lưới kéo đơn
Do loại hình khai thác của nghề lưới kéo là qui mô nhỏ nên kích thước lưới nhỏ. Kích thước mắt lưới 2a ở đụt lưới (nơi chứa cá) là 22,3 mm (Sóc Trăng) và 24,71 mm (Bến Tre). Với kết quả này thì kích thước mắt lưới ở đụt lưới ở tỉnh Bến Tre lớn hơn ở Sóc Trăng. Nhưng cả hai đều nhỏ hơn kích thước mắt lưới qui định là tối thiểu phải là từ 28 mm trở lên (Bộ Thủy sản, 2006). Chính vì vậy, để hoạt động của nghề lưới kéo không ảnh hưởng đến nguồn lợi thủy sản thì cần phải tuyên truyền và kiểm tra thực hiện nghiêm kích thước mắt lưới theo quy định của ngư dân.
Bảng 5. Các thông số của lưới kéo đơn
Sản lượng một năm của nghề lưới kéo đơn ở tỉnh Sóc Trăng và Bến Tre lần lượt là 16.721 ± 4.400 kg/nămvà 15.565 ± 1.890 kg/năm và tỉ lệ cá tạp lần lượt là 33,3% và 39,7%. Tỉ lệ các tạp của tàu lưới kéo đơn ở tỉnh Bến Tre lớn hơn ở tỉnh Sóc Trăng là do tàu này có qui mô nhỏ nên chỉ khai thác gần bờ nên nhiều các tạp hơn. Kết quả cho thấy sản lượng khai thác của nghề lưới kéo đơn ở hai tỉnh này rất thấp so với sản lượng khai thác của nghề lưới kéo ở tỉnh Bạc Liêu là 104 tấn/tàu/năm và tỉ lệ cá tạp là 42,6% (Nguyễn Thanh Long, 2016). Đây là do qui mô tàu khai thác ở hai tỉnh này nhỏ hơn tàu ở Bạc Liêu (57,7 CV) nên sản lượng không cao (Nguyễn Thanh Long, 2016).
Bảng 6. Sản lượng của nghề lưới kéo đơn
3.2.2. Khía cạnh tài chính của nghề lưới kéo đơnKết quả khảo sát cho thấy để đầu tư trung bình
cho một tàu lưới kéo đơn ở tỉnh Sóc Trăng và Bến
Nội dung Sóc Trăng Bến TreThời gian trung bình một mẻ lưới (giờ) 3,03 ± 0,49 2,14 ± 0,54
Số mẻ lưới khai thác trong một ngày (mẻ) 3,60 ± 0,62 3,60 ± 0,65
Số ngày khai thác trong một chuyến biển (ngày) 4,90 ± 0,40 4,11 ± 1,19
Số chuyến biển trong tháng (chuyến) 3,93 ± 0,78 5,22 ± 1,55
Số tháng khai thác trong một năm (tháng) 8,33 ± 0,96 8,78 ± 0,97
Nội dung Sóc Trăng Bến TreTải trọng của tàu (tấn) 6,73 ± 0,98 5,64 ± 0,74Công suất của máy tàu (CV) 39,20 ± 5,06 34,87 ± 7,21
Nội dung Sóc Trăng Bến TreTổng số lao động trên tàu (người/tàu) 3,67 ± 0,80 3,33 ± 0,90
Số lao động thuê mướn thêm trên tàu (người) 1,33 ± 0,66 1,56 ± 0,97
Nội dung Sóc Trăng Bến TreKích thước mắt lưới 2a lớn nhất (mm) 40,67 ± 9,24 38,56 ± 8,23
Kích thước mắt lưới 2a nhỏ (mm) 22,30 ± 3,21 24,71 ± 4,15
Nội dung Sóc Trăng Bến TreSản lượng (kg/năm) 16.721 ± 4.400 15.565 ± 1.890Năng suất (kg/CV/năm) 430 ± 113 466 ± 113
Tỉ lệ cá tạp (%) 33,3 ± 3,3 39,7 ± 7,4

124
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
tre lần lượt là 132 triệu đồng và 103 triệu đồng. Qua đây cũng thấy qui mô tàu lưới kéo ở tỉnh Bến Tre nhỏ hơn tàu ở tỉnh Sóc Trăng nên có tiền đầu tư thấp hơn. Chi phí một chuyến biển cho tàu lưới kéo đơn ở tỉnh Bến Tre (10,45 ± 4,29 triệu đồng/chuyến) cũng thấp hơn chi phí ở tỉnh Sóc Trăng (16,97 ± 3,58 triệu đồng/ chuyến), chủ yếu là tiền dầu (57,05% - Bến Tre và 64,18% - Sóc Trăng) và nhân công (27,76% - Bến Tre và 21,65% - Sóc Trăng). Tỉ suất lợi nhuận của tàu lưới kéo đơn ở hai tỉnh này đạt 0,44 ± 0,19 lần (Tỉnh Sóc Trăng) và 0,38 ± 0,35 lần (Tỉnh Bến Tre). So với kết quả nghiên cứu của Nguyễn Thanh Long và Nguyễn Thanh Phương (2010) tỉ suất lợi nhuận của nghề lưới kéo ven bờ ở tỉnh Sóc Trăng năm 2010 là 0,51 lần. Qua đây cho thấy nghề khai thác lưới kéo đơn qui mô nhỏ ngày càng không hiệu quả. Đây là do nguồn lợi thủy sản ven bờ ngày càng cạn kiệt. Chi phí sản xuất 1 kg cá đạt khá cao 33.178 ± 4.791 đồng/kg (tỉnh Sóc Trăng) và 27.996 ± 6.727 đồng/kg (Tỉnh Bến Tre). Trong khi giá bán trung bình ở Tỉnh Sóc Trăng chỉ là 42.350 ± 5.321 đồng/kg và ở Bến Tre là 38.215 ± 4.369 đồng/kg.
Bảng 7. Hiệu quả tài chính của nghề lưới kéo đơn
3.3. Hiện trạng quản lý nghề khai thác thủy sản Ở hai tỉnh Sóc Trăng và Bến Tre đều có tàu tuần
tra khai thác thủy sản nhưng các tàu này cũ và lạc hậu. Lực lượng thanh tra, kiểm ngư hiện tại rất mỏng, thiếu thốn về phương tiện, thiết bị và đào tạo nên hoạt động kiểm tra, giám sát, tuần tra trên biển hiện nay không hiệu quả. Về đăng kiểm, hiện nay chỉ đăng kiểm được khoảng 77% số tàu trong tỉnh và còn rất nhiều tàu thuyền khai thác không đăng ký. Mặc dù cấp tỉnh đã có chủ trương không cho
đăng ký tàu có công suất nhỏ (< 20 CV), giảm dần các phương tiện khai thác hủy diệt (lưới cào, te v.v.), tuy nhiên trên thực tế ở nhiều nơi số lượng tàu nhỏ vân tiếp tục tăng trong những năm gần đây và các phương tiện khai thác trái phép vân còn tiếp diễn. Để tăng cường công tác quản lý Bộ NN&PTNT chỉ đạo tạm dừng việc cấp văn bản chấp thuận cho phép đóng mới tàu cá làm nghề lưới kéo dưới mọi hình thức kể cả tàu hư hỏng, mục nát, tai nạn (Tổng cục Thủy sản, 2017).
Ngoài ra, Thủ tướng Chính phủ đã có một số chính sách hổ trợ ngư dân hoạt động trên các vùng biển xa, từ năm 2010 đến 2017 các tỉnh đã thực hiện chính sách hỗ trợ chi phí nhiên liệu, trang bị máy thông tin liên lạc, kinh phí mua bảo hiểm thân tàu và tai nạn thuyền viên cho các tàu có đăng ký hoạt động trên vùng biển xa. Qua đây phần nào tạo điều kiện cho ngư dân khai thác xa bờ, nhằm giảm áp lực khai thác ven bờ và làm suy giảm nguồn lợi thủy sản.
Qua đây cho thấy cần hỗ trợ nhiều hơn nữa cho lực lượng thanh tra, kiểm ngư về nhân lực, phương tiện và kinh phí hoạt động để kiểm tra thường xuyên các tàu khai thác trái phép làm ảnh hưởng đến nguồn lợi thủy sản.
IV. KẾT LUẬNNghề lưới kéo đơn ven bờ là nghề khai thác thủy
sản chủ lực của tỉnh Sóc Trăng và Bến Tre. Nghề này có thể khai thác quanh năm. Qui mô tàu lưới kéo ở tỉnh Bến Tre nhỏ hơn tàu ở Sóc Trăng nên có sản lượng thấp hơn và tỉ lệ các tạp cao hơn tàu ở tỉnh Sóc Trăng. Nghề lưới kéo đơn ven bờ ở cả hai tỉnh đều có tỉ suất lợi nhuận thấp hơn những nghiên cứu trước đây, chứng tỏ nghề lười kéo qui mô nhỏ khai thác ngày càng khó khăn và không hiệu quả. Cần có giải pháp quản lý và hỗ trợ để giảm tác động ảnh hưởng đến nguồn lợi thủy sản và tạo điều kiện để ngư dân tiếp cận vốn vay để cải hoán tàu của họ có công suất lớn, có thể khai thác xa bờ và đạt hiệu quả hơn.
TÀI LIỆU THAM KHẢOBộ Thủy sản, 2006. Thông tư số 02/2006/TT-BTS ngày
20 tháng 3 năm 2006 về việc “Hướng dân thực hiện Nghị định của Chính phủ số 59/2005/NĐ-CP ngày 4 tháng 5 năm 2005 về điều kiện sản xuất, kinh doanh một số ngành thủy sản”.
Lê Trần Nguyên Hùng, 2009. Tổng quan mô hình đồng quản lý nghề cá ở Việt Nam. Trong Hội nghị Đồng quản lý nghề cá quy mô nhỏ tại Việt Nam, Đà Nẵng từ 26 - 27 tháng 10/2009, 22 trang.
Nguyễn Thanh Long, 2016. Nghiên cứu hoạt động khai thác của nghề lưới kéo đơn ven bờ và xa bờ ở tỉnh
Hiệu quả tài chính Sóc Trăng Bến TreChi phí cố định (Triệu đồng) 132 ± 43 103 ± 15
Chi phí khấu hao (Triệu đồng/chuyến) 0,46 ± 0,14 0,55 ± 0,22
Chi phí biến đổi (Triệu đồng/chuyến) 16,51 ± 3,53 9,91 ± 4,14
Tổng chi phí (Triệu đồng/chuyến) 16,97 ± 3,58 10,45 ± 4,29
Doanh thu (Triệu đồng/chuyến) 23,25 ± 4,93 14,28 ± 4,97
Lợi nhuận (Triệu đồng/chuyến) 6,28 ± 3,32 3,83 ± 3,09
Tỉ suất lợi nhuận (lần) 0,44 ± 0,19 0,38 ± 0,35Chi phí cho 1 kg sản phẩm (đồng/kg) 33.178 ± 4.791 27.996 ± 6.727

125
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
Bạc Liêu. Tạp chí Khoa học, Trường Đại học Cần Thơ, 45b: 112 - 118.
Nguyễn Thanh Long và Nguyễn Thanh Phương, 2010. Phân tích khía cạnh tài chính và kỹ thuật của các nghề khai thác thủy sản chủ yếu ở tỉnh Sóc Trăng. Tạp chí Khoa học, Trường Đại học Cần Thơ, 14b: 354-366.
Lê Văn Ninh, 2006. Hiện trạng nghề khai thác hải sản tỉnh Bà Rịa - Vũng Tàu và một số định hướng
phát triển trong thời gian tới. Tạp chí Thủy sản, số 11/2006: 29-30.
Tổng cục Thống kê, 2017. Niên giám Thống kê 2016. Nhà Xuất bản Thống kê. 946 trang.
Tổng cục Thủy sản, 2017. Chỉ thị số 3727/CT-BNN-TCTS ngày 05/05/2017 về việc “Tăng cường công tác quản lý khai thác thủy sản, đảm bảo cho người, tàu cá hoạt động trên biển”.
Status of capture and management of single trawl fishery in coastal areas of Soc Trang and Ben Tre provinces
Nguyen Thanh Long, Tran Dac Dinh and Mai Viet Van AbstractThe study on the status of capture and management of trawl-net fishery was conducted from October 2017 to May 2018 in coastal areas of Soc Trang and Ben Tre provinces. 90 households conducting trawl-net were interviewed on main contents such as technical and financial aspects . The results showed that the highest number of fishing boats with the trawl-net in two provinces of Mekong delta was recorded. The fishing season of single trawling was from January to December. The capacity of trawl-net boats in Soc Trang province was 37.5 CV and in Ben Tre was 38.9 CV. The fishing yield and ratio of trash-fish of trawl-net fishery in Soc Trang province (16.7 tons/year; 39.7%) were higher than that in Ben Tre province (15.5 tons/year; 33.32%); the profits and benefit ratios of trawling in Soc Trang (6.28 million VND/trip; 0.44 times) were also higher than that in Ben Tre province (3.38 million VND/trip; 0.38 times). For the sustainable fisheries by trawl-net, the development and management of fishery resources should be promoted, supporting fishermen to apply loan with low interest rates, and training fishermen to use fishing equipment to increase their fishing efficiency.Keywords: Fishery, trawl-net, fishery management, Soc Trang, Ben Tre
Ngày nhận bài: 23/7/2018Ngày phản biện: 3/8/2018
Người phản biện: TS. Võ Thành ToànNgày duyệt đăng: 18/9/2018
1 Phòng Công nghệ sinh học Thủy sản, Trung tâm Công nghệ sinh học TP. Hồ Chí Minh2 Trường Đại học Quốc tế - Đại học Quốc gia TP. Hồ Chí Minh
HIỆU QUẢ CỦA CHỦNG AEROMONAS HYDROPHILA NHƯỢC ĐỘC SỬ DỤNG LÀM VẮC-XIN CHO ĂN TRONG PHÒNG BỆNH XUẤT HUYẾT CÁ TRA GIỐNG
Vũ Thị Thanh Hương1, Nguyễn Hồng Đức2, Lê Thị Thu Thảo1, Ngô Huỳnh Phương Thảo1, Nguyễn Quốc Bình1
TÓM TẮTNghề nuôi cá tra (Pangasianodon hypophthalmus) ở Việt Nam hiện đang bị đe dọa bởi các đợt bùng phát dịch
bệnh, bệnh xuất huyết do Aeromonas hydrophila là một trong những bệnh phổ biến nhất với tỷ lệ thất thoát cao. Trong nghiên cứu này, chủng A. hydrophila đột biến nhược độc (M14) tạo ra bằng phương pháp knock-out gen aroA được sử dụng dưới dạng vắc-xin cho ăn để khảo sát hiệu quả bảo vệ trước chủng hoang dại ở điều kiện phòng thí nghiệm trên cá tra giống (trọng lượng 8 ± 2 g). Kết quả cho thấy hiệu quả bảo vệ (RPS) đạt 64%. Xử lý vắc xin bằng phương pháp cho ăn cần được hoàn thiện hơn nữa để sản xuất vắc xin phòng bệnh xuất huyết trên cá tra.
Từ khóa: Aeromonas hydrophila, bệnh xuất huyết, cá tra, chủng đột biến nhược độc, vắc xin cho ăn
I. ĐẶT VẤN ĐỀNgành thuỷ sản Việt Nam đóng vai trò quan
trọng trong sự phát triển kinh tế đất nước với quy mô ngày càng mở rộng trong nền kinh tế quốc dân. Trong số các mặt hàng thủy sản, sản phẩm từ
cá tra mang lại hàm lượng dinh dưỡng cao, tốt cho sức khỏe, được nhiều người trong và ngoài nước ưa dùng. Theo Hiệp hội Chế biến và Xuất khẩu Thủy sản Việt Nam (VASEP) (2017), kim ngạch xuất khẩu cá tra trong năm 2017 đạt 1,78 tỷ USD, diện tích mặt

126
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
nước thả nuôi cá tra đạt 5.230 ha, sản lượng đạt 1.250 nghìn tấn. Tuy nhiên, bệnh do vi khuẩn trên cá tra vân đang là vấn đề lớn nhất cần kiểm soát, đặc biệt bệnh xuất huyết do vi khuẩn Aeromonas hydrophila là bệnh phổ biến, xảy ra quanh năm, tập trung vào đầu mùa khô, đặc biệt là khi cá bị stress như sau khi trời mưa, thiệt hại nghiêm trọng cho người nuôi (Từ Thanh Dung, 2015). Một trong những phương pháp ngừa bệnh hữu hiệu, an toàn và bền vững trong nuôi cá là sử dụng vắc xin.
Năm 2016, vắc xin ALPHA JECT ® Panga 2 sản xuất bởi Pharmaq đã được cấp phép lưu hành. Đây là một loại vắc xin chết, sử dụng bằng phương pháp tiêm giúp phòng bệnh xuất huyết do A. hydrophila và bệnh gan thận mủ do Edwardsiella ictaluri (VASEP, 2016).
Mặc dù có tỷ lệ bảo hộ cao, phương pháp tiêm vắc xin vân còn nhiều hạn chế về mặt kinh phí, nhân lực và rủi ro thương tích cao cho cá (Mutoloki et al., 2008). Phương pháp ngâm có hiệu quả và thực tế khi tiến hành xác định đáp ứng miễn dịch cho cá với số lượng lớn. Tuy nhiên, về mặt thực tiễn, vắc xin cho ăn dễ dàng sử dụng đối với người nuôi và thuận tiện cho giai đoạn cá bắt đầu ăn thức ăn công nghiệp. Cho đến nay, ở Việt Nam chưa có vắc xin sống phòng bệnh cho cá tra dạng cho ăn được thương mại.
Vắc xin sống nhược độc dạng cho ăn đang được phát triển mạnh mẽ trên thế giới để ứng dụng trong thủy sản. Gần đây nhất là nghiên cứu của Nagaraj Chatakondi và cộng sự cho thấy vắc xin sống nhược độc E. ictaluri dạng cho ăn khi chủng ngừa cho hai dòng cá nheo channel catfish (Ictalurus punctatus) và hybrid catfish (Ictalurus punctatus) ở 4 liều từ 4 ˟ 106 - 3,2 ˟ 107 tế bào vắc xin sống/g thức ăncó tỉ lệ chết sau công độc trong khoảng 10,4 - 19,4%. Đối với cá không được chủng ngừa vắc xin, tỉ lệ chết là 73 - 85% (Nagaraj Chatakondi, 2018).
II. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Vật liệu nghiên cưuCá tra giống (Pangasianodon hypophthalmus)
từ 2,5 - 3 tháng tuổi có trọng lượng 8 ± 2 g, chủng Aeromonas hydrophila hoang dại và chủng đột biến nhược độc M14 sử dụng trong nghiên cứu được cung cấp từ Trung tâm Công nghệ sinh học thành phố Hồ Chí Minh.
Chủng vi khuẩn Aeromonas hydrophila M14 đột biến nhược độc sử dụng như một vắc xin sống
nhược độc được tạo ra bằng phương pháp knock out gen aroA đã được tạo ra bởi chính nhóm tác giả, kết quả cho thấy hiệu quả bảo vệ (RPS) bằng phương pháp tiêm và ngâm trên 95% (Vũ Thị Thanh Hương, 2016).
2.2. Phương pháp nghiên cưu
2.2.1. Chuân bị cá thí nghiệmCá tra bột mới nở trong vòng 24 h được cung cấp
bởi các trại sản xuất cá tra giống được nuôi trong bể composite 500 L tại phòng thí nghiệm của Trung tâm Công nghệ sinh học thành phố Hồ Chí Minh. Ở giai đoạn cá bột (trước 15 ngày tuổi), cá được cho ăn moina và artemia. Giai đoạn 16 - 20 ngày tuổi, cá sử dụng thức ăn là trùn chỉ. Đến giai đoạn 21 - 45 ngày tuổi cho cá ăn thức ăn công nghiệp 30 độ đạm xay nhỏ phù hợp với kích cỡ miệng của cá. Trong quá trình nuôi, các bể nuôi thường xuyên được xi phông nhằm loại trừ thức ăn thừa, tránh tình trạng ô nhiễm môi trường nước, định kì 1 - 2 ngày thay nước một lần (mỗi lần thay từ 30% - 40% lượng nước) ở tất cả các bể, hệ thống sục khí hoạt động 24/24 giờ. Cá nuôi đến giai đoạn 2,5 - 3 tháng tuổi được dùng cho các thử nghiệm.
2.2.2. Chuân bị chủng Aeromonas hydrophila đột biến và hoang dại
Các chủng sử dụng trong nghiên cứu bao gồm chủng A. hydrophila hoang dại (ký hiệu A. hydrophila AGII) được phân lập tại thành phố Long Xuyên, tỉnh An Giang và chủng A. hydrophila đột biến nhược độc (ký hiệu M14) được tạo ra bằng phương pháp knock-out gen aroA. a) Chuẩn bị chủng A. hydrophila đột biến nhược độc M14
Để chuẩn bị dịch vi khuẩn A. hydrophila đột biến, lấy một ống giống được giữ ở –80oC tăng sinh trong 5 mL Luria-Bertani (LB) lỏng (Kay et al., 1985) có bổ sung kháng sinh ampicillin và kanamycin (50 µg/mL), nuôi cấy qua đêm ở điều kiện lắc 250 vòng/phút ở 28oC, trong 48 giờ. Sau đó, 2 mL dịch nuôi cấy được cấy chuyền vào 100 mL LB lỏng có bổ sung kháng sinh ampicillin và kanamycin (50 µg/mL)và nuôi cấy 250 vòng/phút ở 28oC. Khi OD600 đạt khoảng từ 1,3 - 1,4 (tương đương với mật độ 2 ˟ 109 CFU/mL) dịch nuôi cấy được ly tâm ở 2.000 ˟ g/20 phút ở 4oC để loại bỏ dịch nổi thu sinh khối, sau đó sinh khối được huyền phù lại trong dung dịch NaCL 0,65%. Mật độ vi khuẩn được kiểm tra bằng cách cấy trải trên môi trường LB có bổ sung kháng sinh ampicillin và kanamycin (50 µg/mL).

127
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
b) Chuẩn bị chủng A. hydrophila hoang dại AGIITương tự, đối với chuẩn bị dịch vi khuẩn
A. hydrophila hoang dại, lấy một ống giống được giữ ở –80oC tăng sinh trong 5 mL LB lỏng (Kay et al., 1985) có bổ sung kháng sinh ampicilin (50 µg/mL), nuôi cấy qua đêm ở điều kiện lắc 250 vòng/phút ở 28oC, trong 16 h. Sau đó, 2 mL dịch nuôi cấy được cấy chuyền vào 100 mL LB lỏng có bổ sung kháng sinh ampicilin (50 µg/mL) và nuôi cấy 250 vòng/phútở 28oC. Dịch nuôi cấy được ly tâm và thu sinh khối, sau đó huyền phù và pha loãng đến OD600~1,0 (tương đương với mật độ 109 CFU/mL) bằng dung dịch NaCl 0,65%. Mật độ vi khuẩn được kiểm tra bằng cách cấy trải trên môi trường LB có bổ sung kháng sinh ampicilin (50 µg/mL).
2.2.3. Khảo sát liều LD50 của chủng A. hydrophila hoang dại AGII trên cá tra giống
Phương pháp xác định liều gây chết 50% (Lethal Dose 50% - LD50) được tiến hành dựa theo phương pháp của Reed and Muench (1938). Thí nghiệm bao gồm 3 nghiệm thức, mỗi nghiệm thức 10 cá tra giống 2,5 - 3 tháng tuổi. Cá được ngâm trong xô 5 L chứa dịch vi khuẩn A. hydrophila AGII nồng độ 105, 106 và 107 CFU/mL với thời gian ngâm 30 phút, sau đó cá được rửa sạch và chuyển vào bể nuôi 100 L, hàng ngày tiến hành ghi nhận số cá chết. Giá trị LD50 được tính theo công thức sau:
𝐿𝐷50 = 10(𝑎+𝑥)
Trong đó: 10a là nồng độ tại đó số lượng cá sống và cá chết sau thí nghiệm là 50%; x được tính theo công thức:
x = (Pa – 50)/(Pa – Pu)Trong đó Pa, Pu là tỉ lệ cận trên và cận dưới của
nồng độ gây chết 50% (Reed, L., & Muench, H., 1938).Đồng thời, sự hiện diện của vi khuẩn A. hydrophila
trong các mâu cá chết được kiểm tra bằng phương pháp PCR với cặp mồi Aer đặc hiệu cho A. hydrophila được thiết kế dựa trên trình tự gen AHA-0438 (gen ID: 4490401) mã hóa cho protein Hemolysin cho kích thước 191bp (Vũ Thị Thanh Hương, 2016). Khuẩn lạc được xác định là A. hydrophila được tăng sinh lại theo phương pháp tương tự như trong mục 2.2.2 (chuẩn bị chủng A. hydrophila hoang dại AGII) để dùng trong các thử nghiệm công độc tiếp theo.
2.2.4. Khảo sát hiệu quả bảo vệ của chủng A. hydrophila đột biến M14 trên cá tra giống
Bố trí thí nghiệm: Dịch vi khuẩn M14 sau tăng sinh được cô đặc thành nồng độ 2 ˟ 1010 CFU/mL và pha loãng bậc 10 thành các nồng độ: 2 ˟ 109, 2 ˟ 108,
2 ˟ 107 CFU/mL. Mỗi nồng độ được trộn với thức ăn thủy sản DT04 (công ty TNHH TAGS Lái Thiêu) theo tỉ lệ 10 mL dịch vi khuẩn và 5 g thức ăn. Như vậy, thức ăn sau khi trộn với M14 có 4 nồng độ 1010, 109, 108, 107 CFU/g, tương ứng với 4 nghiệm thức 1, 2, 3, 4. Hỗn hợp này sau đó được áo dầu mực theo tỉ lệ 10 g thức ăn và 1 mL dầu mực nhằm hạn chế sự thất thoát vi khuẩn và tăng sự hấp dân mùi vị đối với cá. Để tăng hiệu suất ăn của cá, cá tra giống (8 ± 2 g)bị nhịn ăn một ngày trước khi cho ăn thức ăn có vi khuẩn M14. Cá (20 con/nghiệm thức) được cho ăn liên tục trong năm ngày, mỗi ngày ăn 2 lần. Ở nghiệm thức 5, cá được ngâm với chủng M14 nồng độ 107 CFU/mL trong bể 5 L, thời gian ngâm 30 phút, sau đó rửa cá sạch và đưa trở lại bể nuôi. Nghiệm thức 6 là đối chứng dương không xử lý M14, nhưng công độc với chủng hoang dại AGII. Nghiệm thức 7là đối chứng âm, không xử lý M14 và không công độc. Cá ở các nghiệm thức 5, 6, 7 được cho ăn thức ăn không chứa chủng A. hydrophila đột biến M14. Mỗi nghiệm thức lặp lại 3 lần, tương ứng với ba bể, mỗi bể bố trí 20 cá.
Bảng 1. Các nghiệm thức khảo sát hiệu quả bảo vệ của chủng A. hydrophila đột biến M14 trên cá Tra giống
14 ngày sau khi cho ăn hoặc ngâm với chủng A. hydrophila đột biến M14, cá ở các nghiệm thức được công độc với chủng A. hydrophila AGII ở nồng độ LD50 theo phương pháp được mô tả ở trên. Số liệu cá sống sót sau 2 tuần công độc được xử lý thống kê bằng phần mềm IBM SPSS 20. Với mỗi nghiệm thức, hiệu quả bảo vệ (RPS) được tính theo công thức (Amend, 1981):
RPS (%) = 1 − ˟ 100tỉ lệ chết của nghiệm thức
tỉ lệ chết của đối chứng
2.3. Thời gian và địa điểm nghiên cưuNghiên cứu được thực hiện năm 2018 tại phòng
Công nghệ sinh học Thủy sản, Trung tâm Công nghệ sinh học thành phố Hồ Chí Minh.
Nghiệm thưc Liều dùng Phương
pháp dùngSố lượng
cá/bểNT1 1 ˟ 1010 CFU/g Cho ăn 20NT2 1 ˟ 109 CFU/g Cho ăn 20NT3 1 ˟ 108 CFU/g Cho ăn 20NT4 1 ˟ 107 CFU/g Cho ăn 20NT5 1 ˟ 107 CFU/mL Ngâm 20NT6 ĐC (+) 20NT7 ĐC (-) 20

128
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
III. KẾT QUẢ VÀ THẢO LUẬN
3.1. Khảo sát liều LD50 của chủng A. hydrophila hoang dại AGII trên cá Tra giống
Sau 2,5 - 3 tháng nuôi, cá tra nuôi đạt trọng lượng 8 ± 2 g khỏe mạnh, có kết quả âm tính khi kiểm tra sự hiện diện của A. hydrophila và E. ictaluri bằng PCR, sẵn sàng cho thử nghiệm. Chủng A.hydrophila M14 và AGII sau tăng sinh có nồng độ 1 ˟ 109 CFU/mL- 2 ˟ 109 CFU/mL, được ly tâm, thu sinh khối và huyền phù lại trong NaCl 0,65%.
Sau công độc bằng phương pháp ngâm chủng AGII, ở nghiệm thức ngâm nồng độ 107 CFU/mL cá chết liền sau 12 h và ngưng chết ở ngày thứ 3, tuy nhiên ở nồng độ 105 CFU/mL và 106 CFU/mL cá chết bắt đầu từ ngày thứ 3 và ngưng chết sau 7 ngày. Những cá chết liền sau ngâm chủng AGII không có dấu hiệu bệnh, cá chết sau 2 ngày có biệu hiện xuất huyết gốc vây bụng, vây lưng, vây đuôi, và phù nề xung quanh hốc mắt. Các cá chết được phân lập vi khuẩn và kiểm tra sự hiện diện A. hydrophila bằng cặp mồi định danh A. hydrophila cho thấy tất cả các kết quả đều dương tính.
Bảng 2. Kết quả khảo sát liều LD50
Hình 1. Kết quả điện di trên gel agarose 1,2% các sản phẩm PCR khuẩn lạc phân lập từ cá chết sau cảm nhiễm ngược. Giếng 1, 2, 3, 4: Sản phẩm PCR khuẩn lạc phân lập từ cá chết; Giếng 5: Đối chứng âm; Giếng 6 là thang 1kb
3.2. Khảo sát hiệu quả bảo vệ của chủng A. hydrophila đột biến nhược độc M14 trên cá tra giống
Ở các nghiệm thức (NT) cho ăn dịch vi khuẩn M14 với nồng độ 1010 (NT1) 109 (NT2), 108 (NT3) 107 CFU/g (NT4), tỉ lệ cá sống sau công độc lần lượt là 5%,7%, 2%, 15% so với nghiệm thức ngâm ở nồng độ 107 CFU/mL (NT5) là 10%. Trong khi đó, ở ng-hiệm thức đối chứng dương (NT6) (cá không được
xử lý với dịch vi khuẩn M14) sau công độc, tỉ lệ cá chết chỉ là 42% (Hình 2).
Hình 2. Biểu đồ tỉ lệ cá chết sau 14 ngày công độc với chủng A. hydrophila AGII
Từ số liệu tỷ lệ chết của cá, hiệu quả bảo vệ (RPS) của các nghiệm thức lần lượt được tính là 88%, 83%, 95%, 64% và 76%. Tỷ lệ chết của các nghiệm thức có cùng kí hiệu a không có khác biệt về mặt thống kê, ngược lại nghiệm thức có sự khác biệt về thống kê khi có kí hiệu khác nhau a và b.
Theo kết quả thống kê ANOVA với Duncan post-hoc test, có sự khác biệt về mặt thống kê trong tỉ lệ sống của cá ở tất cả các nghiệm thức cho ăn và ngâm so với nghiệm thức đối chứng dương (p < 0,05).Tuy nhiên, giữa nghiệm thức cho ăn và nghiệm thức ngâm không có sự khác biệt có ý nghĩa, và tương tự với các nghiệm thức cho ăn (p > 0,05). Điều này cho thấy phương pháp cho ăn có thể thay thế phương pháp ngâm để đưa chủng đột biến nhược độc vào cơ thể cá; và do không có sự khác biệt về thống kê giữa tỉ lệ sống của các nghiệm thức cho ăn với nhau, nên nồng độ cho ăn 107 CFU/g với hiệu quả bảo vệ 64% là sự lựa chọn hiệu quả nhất về mặt kinh tế và đáp ứng được tiêu chuẩn vắc xin thủy sản là hiệu quả bảo vệ cần đạt trên 60%. Trong nghiên cứu về vắc xin sống nhược độc E. ictaluri cho cá nheo (chanel catfish) 7 - 9 cm với liều lượng 1 lần ăn duy nhất ở mật độ 4,6 - 6 ˟ 106 CFU/g và 4,6 - 6 ˟ 107CFU/g đạt hiệu quả bảo vệ lần lượt 92 - 100% và 97 - 100% (Embregts và Forlenza, 2016). Để tăng hiệu quả bảo vệ của chủng A. hydrophila đột biến nhược độc M14 trên cá tra, cần thêm các nghiên cứu về cách thức phối trộn thức ăn với chủng vi khuẩn, số lần cho ăn, giai đoạn phát triển phù hợp của cá để cho ăn vắc xin,... Nghiên cứu này có ý nghĩa quan trọng trong việc đánh giá hiệu quả bảo vệ của chủng đột biến nhược độc M14 trên cá bằng phương pháp cho ăn, có độ an toàn và hiệu quả về mặt kinh tế cao hơn so với phương pháp tiêm và ngâm.
Nồng độ Số lượng cá chết Tỉ lệ chết105 CFU/mL 4 40%106 CFU/mL 5 50%107 CFU/mL 8 80%

129
Tạp chí Khoa học Công nghệ Nông nghiệp Việt Nam - Số 11(96)/2018
IV. KẾT LUẬN VÀ ĐỀ NGHỊ
4.1. Kết luậnChủng A. hydrophila đột biến nhược độc M14
khi đưa vào cá bằng phương pháp trộn với thức ăn ở nồng độ 107 CFU/g, sau 14 ngày công độc với chủng A. hydrophila có hiệu quả bảo vệ là 64% và có tiềm năng phát triển thành vắc xin cho ăn để phòng ngừa bệnh xuất huyết do A. hydrophila trên cá tra.
4.2. Đề nghịNghiên cứu này là tiền đề để phát triển vắc xin
phòng bệnh xuất huyết trên cá tra bằng phương pháp cho ăn từ đó cần có những nghiên cứu thêm về phương pháp trộn thức ăn, con đường đi của thức ăn trong ruột cá để giảm sự thất thoát vi khuẩn M14, nhằm tăng sự kích thích miễn dịch và hiệu quả bảo vệ cao hơn.
TÀI LIỆU THAM KHẢOTừ Thanh Dung, Đặng Thị Hoàng Oanh và Phạm
Minh Đưc, 2015. Bệnh và quản lý dịch bệnh trong nuôi cá tra (Pangasianodon hypophthalmus). Trong: Nguyễn Thanh Phương và Nguyễn Anh Tuấn (chủ biên). Nuôi cá tra (Pangasianodon hypophthalmus) ở Đồng bằng sông Cửu Long: Thành công và thách thức trong phát triển bền vững. Nhà xuất bản Đại học Cần Thơ, trang 156-189.
Vũ Thị Thanh Hương, 2016. Tạo chủng Aeromonas hydrophila đột biến và thử nghiệm đánh giá khả năng đáp ứng miễn dịch của cá Tra giống. Báo cáo nghiệm thu đề tài Sở Khoa học và Công nghệ TP. HCM.
Amend., 1981. Potency testing of fish vaccines. Dev Biol Stand, 447.
Embregts, C. W. E. and Forlenza, M., 2016. Oral vaccination of fish: Lessons from humans and
veterinary species, Developmental and Comparative Immunology. Elsevier Ltd, 64, pp. 118-137. doi: 10.1016/j.dci.2016.03.024.
Kay, B. A., Guerrero, C. E. and Sack, R. B., 1985. Media for the isolation of Aeromonas hydrophila. Journal of clinical microbiology, 22 (5), pp. 888-890. Available from: http://www.pubmedcentral.nih.gov/articler-ender.fcgi?artid=268556&tool=pmcentrez&render-type=abstract (Accessed: 3 May 2013).
Mutoloki, S., Alexandersen, S., Gravningen, K., Evensen, Ø., 2008. Time-course study of injection site inflammatory reactions following intraperitoneal injection of Atlantic cod (Gadmus morthua L.) with oil-adjuvanted vaccines. Fish shellfish Immunol, 386 - 393.
Nagaraj Chatakondi et al ., 2018. Efficacy of a Live-at-tenuated Edwardsiella ictaluri Oral Vaccine in Channel and Hybrid Catfish. Journal of the world aquaculture society, Volume 49, issue 4, 686-691.
Reed, L., & Muench, H., 1938. A simple method of estimating fifty percent endpoints. The American Journal of Hygiene, 493-497.
Tobar J.A., Jerez S., Caruffo M., Bravo C., Contreras F., Bucarey S.A., Harel M., 2011. Oral vaccination of Atlantic salmon (Salmo salar) against salmonid rickettsial septicaemia. Vaccine, 2336-2340.
Vietnam Association of Seafood Exporters and Producers (VASEP), 2017. Statistical Data: Total Export by Year: 2017. Retrieved 15/03/2018 from vasep.com.vn.
Vietnam Association of Seafood Exporters and Producers (VASEP), 2016. http://vasep.com.vn/Tin-Tuc/1206_46412/Vac-xin-cho-ca-tra-ALPHA-JECT-Panga-2-cua-PHARMAQ-da-duoc-cap-phep-luu-hanh.htm.
Efficacy of live-attenuated Aeromonas hydrophila oral vaccine against Hemorrahagic septicemia in Tra catfish fingerling (Pangasianodon hypophthalmus)
Vu Thi Thanh Huong, Nguyen Hong Duc,Le Thi Thu Thao, Ngo Huynh Phuong Thao, Nguyen Quoc Binh
AbstractTra catfish production in Vietnam is currently being threatened by disease outbreaks, among them Hemorrhagic septicemia disease caused by Aeromonas hydrophila is one of the most prominent, resulting in high mortality. A mutant strain developed (M14) by knocking out the aroA gene, a reduced virulent strain, was used in this study as an orally delivered vaccine. The protective efficacy against the virulent wild type in Tra catfish fingerlings (8 ± 2g) in laboratory experiments with the relative percent survival (RPS) of 64%, showed the potential of the strain as a live attenuated vaccine candidate on average. More studies need to be done for developing commercialized oral vaccine against hemorrhagic disease on Tra catfish.Keywords: Aeromonas hydrophila, hemorrhagic, Tra catfish (Pangasianodon hypophthalmus), attenuated mutant Aeromonas hydrophila, oral vaccine
Ngày nhận bài: 14/10/2018Ngày phản biện: 21/10/2018
Người phản biện: TS. Nguyễn Ngọc DuNgày duyệt đăng: 15/11/2018
