樟芝短鏈脫氫醇/還原因每之序列分析與蛋白質結構模擬 ·...
TRANSCRIPT

林業研究季刊 32(2): 59-70 , 2010 59
噩噩噩益
樟芝短鏈脫氫醇/還原因每之序列分析與蛋白質結構模擬
許耿豪 1 李宜儒2 曲芳華3
【摘要】樟芝 (Antrodia cinnamomea) 為早期台灣原住民使用來做為消病與保健的傳統中贅,許多
研究報告已經證實樟芝具有許多珍貴醫藥價值的成分,然而目前對於樟芝的細胞生理調控和代謝產
物生合成仍然所知有限。本研究由樟芝表達序列標籤 (expressed sequence tags) 資料庫裡選殖出一
個短鏈脫氫時/還原時 (short-chain dehydrogenase/reductase; SDR)基因,命名為ACSDR , 其全長
為 876 個鹼基對 (base pair) ,可轉錄為 291 個胺基酸,分子量約 29.9回a 。分析ACSDR胺基酸序
列找到SDR家族的保守序列 (conserved residues) TGxxxGxG和預測的酵素活性催化三角 (catalytic
前iad) Ser-乃r-Lys 0 經蛋白質結構模擬計算, ACSDR 以典型的a/ß摺疊組成的Rossmann摺疊呈現,
惟α2 、 ß2 、 ß3 及ß4構成的結構單元 (domain) 與其他 SDR 蛋白質有明顯結構差異,顯見 ACSDR
可能在樟芝特有的受質具有專一選擇性。在親源演化分析結果也發現, ACSDR 雖然與其他真茵
SDR 蛋白質歸屬一群,但演化上仍有距離,顯見此蛋白質卻有可能參與樟芝特有代謝產物之生合
成。因此本研究除了可以幫助了解樟芝 SDR 蛋白質特性與結構,也提供此蛋白質在未來代謝體工
程和酵素產業上的應用價值。
【關鍵詞】樟芝,脫氫醇,還原醇,蛋白質結構
Research oaoer
Sequence analysis and structural modeling of a short-chain dehydrogenase/reductase gene from Antrodia cinnamomea
Keng-Hao Hsu' Yi-Ru Lee2 Fang-Hua Chu'
<Abstract> The basidio血ycetous fungi Antrodia cinnamomea is 0立en used in Traditional Chinese
Medicine and is known to possess a wide range of biological activities. Here, a novel short-chain
dehydrogenase/reductase (SDR) gene was obtained from expressed sequence tags (ESTs) 企omA
cinnamomea named ACSDR. The full-Iength of this gene has an 876 bp open reading 企ame (ORF)
encoding 291 amino acids with a molecular weight of 35.5 kDa. ACSDR contains a SDR family
conserved residues (TGxxxGxG) and a Ser-Tyr-Lys cat叫ytic 甘iad. According to the protein structura1
1 國立中興大學森林學系博士後研究
Postdoc'恤', D呻E個問tofFo扭曲y, Na自由llChung Hs旭g Ui血ve:即可
2 國立臺灣大學森林環境暨資源學系助理Assi蝴醋, Scb∞11offor目甸 and Resour回Con甜vation, N:曲。nal Taiw阻U曲曲i可
3 固立臺灣大學森林環境置費盟軍學系副教授,通訊作者,台北市大安區聶斯稿時四段 1 號
Associa阻 profe目前" School ofForestry and Resource C由lServati凹, Nati阻,a1 τ'aiwan Univers旬" Corresponding Au血。rNo.l, Sec.4, R曲曲ve1t Road, 106 Taipe~ Taiwao (R,Q ,C.)

60 樟芝短鏈脫氫醇/還原醇之序列分析與蛋白質結構模擬
modeli時, the predicted ACSDR protein displays the highly similar a/ß folding oftypical Rossmann
fold.Howev缸., a Iid Iike domaia of ACSDR formed byα2 、 ß2 、 ß3 and ß4 reveals distinct conformation
from other SDR proteins implies it's substrate specificity inA. cinnamomea. The phylogenetic analysis
also showed the evolutional vari祖tofACSDR企omo也er ft血gi SDRs. This is the first SDR identified
and characterized inA. cinnamomea, the study of ACSDR provide the 晶晶,er application for e盟ymatic
industrial and metabolic engineering
<Keywords> Antrodia cinnamomea; dehydrogenase; reductase, protein structure
一、前言
短鏈脫氫醇/還原醇 (short-chaia dehydro
genase/reductase; SDR)組成了一個為數眾多的
蛋白質家族,而這酵素的發現最早可追溯到西
元 70 年代的一個真菌核糖醇脫氫時與果蠅的
酒精脫氫醇 (Jornvall et al, 1981; Schwartz & J ornvall, 1976) 。這兩個脫氫構與當時已知在
動物肝細胞和酵母菌找到的酒精脫氫酪卻有顯
著的不同,鑑於前者胺基酸序列長度較後者
短,因此分類歸屬為短鍊脫氫鸝家族。而SDR
這類酵素直到西元 1990 年才開始受到重視,
原因是在哺乳動物發現一個 15-輝前列腺素脫
氫醇(15-hydroxyprostaglandin dehydrogenase)
會與菸幢胺腺嘿時三核酸 (ni叩Itinamide adenine
dinucleotide; NAD+) 結合(Krook et al, 1990) 。
因此後續將這些依賴菸瞳胺腺嘿時雙核酸
。,AD 冊)和菸瞳胺腺嘿時雙核酸磷酸。可ADP
個))作為輔龍 (cofactor) 的短鏈脫氫酪/還原
蘭歸類為 SDR 家族。 SDR 家族的酵素活性皆
不依賴金屬離子,名為短鏈主因其分子量較其
他脫氫團軍/還原醇小,約在 25 到 35 kDa範圍
之間,而且以兩個單體或四個單體聚合來進行
酵素偕化反應 (Oppermann et al, 2003) 。雖然
此家族蛋白質在胺基酸序列的相間性不高,約
15 %到 30 % '但由其蛋白質立體結構分析,卻有相似的αIß摺疊結構,稱為Rossm阻n核酸
結合模體 (Jornvall et al, 1995; Oppermann et
祉, 2003) 0 SDR家族再由其輔冒種種類、活性區
域與胺基酸序列長短又分為五個E家族,分別
是一般型(classical) 、延伸型 (extended) 、中
間型 (intermediate) 、分歧型 (divergent) 和複
合型 (complex) (Kallberg et al, 2002) 。雖然
SDR家族蛋白質皆進行相同酵素催化反應,但
由其結構與物種間的差異,它們作用的受質種
類涵蓋很大的範圈,主要參與生物體內的多種
代謝,如類固醇、膽酸、前列腺素、類視黃
素、脂肪酸的合成與分解外源性毒素等(Opper
m祖n et al, 2003) 。脂肪酸的合成在所有生物
體內都是很重要的初級代謝反應,丙二盤瞳基
載體蛋白 (malonyl-acyl carrier protein;malonyl
ACP) 與種基-種基載體蛋白 (acyl-ACP) 合成F
﹒間瞳﹒種基載體蛋白(ß-ketoacyl-ACP) 後,需
要:p-閉酪種基載體蛋白還原醇(ß-ketoacyl-ACP
reductase) 還原反應成戶經區基-種基載體蛋白
(ß-hydroxyacyl-ACP) 以進行脂肪酸後績的合
成步膀,其中rp_闡種-種基載體蛋白還原時(又
名 3 被基﹒種基載體蛋白還原醇; 3-oxoacyl
ACP)就是屬於 SDR家族的一員。只是做菌的
脂肪酸合成與脊椎動物一樣屬於第一型的,意
即此型的脂肪酸合成醇為一個具有多酵素功能
的蛋白質,而iP-闢幢﹒種基載體蛋白還原時只是
此多功能蛋白質裡的一個結構單元 (domain) 。
相反的,在植物與大部分細菌所屬的第二型
裡,這些合成脂肪曦的相關酵素則是獨立存在
的。
在哺乳動物體內 SDR 除了參與脂肪酸合
成的重要代謝功能外,也是荷爾蒙生合成的重
要酵素之一,以人類性荷爾蒙為例,雌二醇(自
tradiol) 、畢固圓圓 (t間tos但rone) 和二氫畢固蘭
(dihydrotestosterone)都是需要SDR屬酵素 17ß
.避基類固醇脫氫蘭 (17戶-hydroxysteroid de
hydrog<曲曲目)將無活性的 17闡類固醇 (17 Keto-

林業研究季刊 32(2): 59-70, 2010
steroids)催化為有活性的 17戶-經基型 (17ß-hy
droxy-forms)類固醇 (A,白血ski & Jakob, 2001) 。
在植物方面的研究 'SDR家族也參與很多重要
植物固醇類的生合成,如油菜固醇(brassinos
teroid) 和奮心劑 (Cardenolides) (Finsterbusch
et al, 1999; Seidel et al, 1990)等。在阿拉伯芥
的基因組裡,有 138 個蛋白質被鑑定為屬於
SDR 家族,而其中 41 %屬於需要依賴菸碰胺
腺嘿時三核酸(NADP+-dependent)類型的 σer
sson et al, 2003) 。
相較於動物、植物或細菌,真菌類的SDR
相關研究比較少,對於此酵素家族在生理上扮
演的功能也較不清楚。近兩年的幾篇研究報告
顯示 .SDR除了對真菌生長發育有重要的影響
外,也發現此類酵素參與許多二次代謝產物的
生合成。在引起小麥葉枯病的病盟軍徵菌Stagon
ospora nodorum 裡,發現有一個 SDR家族脫
氫蘭參與此做菌的無性生殖,因此藉由突變抑
制此脫氫時基因不但影響此做茵繁殖,進而影
響此多循環病源的病害發生 (T:阻 et al, 2008) 。
新月彎砲擻 (Curvularia lunata) 是一般植物常
見的病源菌,此菌的兩個屬於 SDR 家族蛋白
質經研究發現其中一個蛋白質具有和動物 17ß
經基類固醇脫氫醇相同的酵素活性,而另一個
蛋白質則參與此儼菌黑色素(血el祖血)生合成
有闕,黑色素在其他動物致病徹菌裡具有抵抗
宿主兔波反應的功能,因此做菌黑色素生合成
的相關蛋白質包括 SDR 在內即成為新的殺菌
和抗生素研發的重要標的但即臨et a1, 2009) 。
在釀酒酵母 (Saccharomyces cerevisiae) 研究
裡,對多種乙陸受質都有具活性的一個SDR家
族乙陸還原醇,可以將在木質纖維素轉化為酒
精過程產生的乙薩受質分解,這些乙陸對大部
分微生物來說都是毒性物質,會抑制和殺死參
與轉化過程的微生物,影響微生物轉化效率,
因此這類 SDR 酵素將可應用於生質酒精生產
(Liu & Moon, 2009) 。另一個重要的研究報告
指出,煙麵菌 (A亮pergillus 戶mlga甜s) 的三菇
類化合物煙麵徹酸 (helvolic acid) 生合成的過
61
程中,一個 SDR酵索和另一個細胞色素 P450
單氧蘭 (cytochrome P450 oxygenase) 共同偕
化煙麵徹酸前過物的C-4 去甲基化(Mitsuguchi
et al. 2009) .由此可見SDR在二次代謝產物的
生合成扮演著重要的角色,這也是微生物SDR
逐漸受到重視的原因之一。
樟芝 (Antrodia cinnamomea) 屬於真菌界
的擔子菌鬥(B阻dio血yco個.) .寄生於台灣特有
之牛樺樹心材內壁,是造成天然林牛樺樹幹中
空腐朽的一種福腐菌。樟芝的保健和療吸除了
在民間廣為流傳外,近年來許多研究報告更以
科學方法證實樟芝的重要醫寮價值,如抗氧化
個seu et al. 2002; Hsiao et al, 2003; Shen et a1,
2006; Song & Yen, 2002; Yang et 叫.2006) 、
血管收縮 (Wang et 叫, 2003) 、抗發炎 (Hseu et
祉, 2005; Shen et 祉, 2004) 、抑制血管增生 (Chen
et 祉, 2005; Cheng et a1, 2005) 、抗瞳瘤(Liu et
叫. 2004; Nakamura et al, 2004) 、抗肝炎 (Lee
et al, 2002; Phuong et al, 2009; Phuong et a1,
2008)與保護肝功能活性扭曲 et a1, 2006) 等。
樟芝這些具有生物嗨性物質已經被分析鐘定,
主要為類固醇化合物、三話類、倍半繭類、聯
笨化合物 (Chenet 叫, 1995; Cherng et a1, 2004;
Hung-Chen Chiang et a1, 1995) .而這些重要化
合物在樟芝進行人工培養時,含量往往遠不如
野生的子實體菌。而台灣牛樟樹過去因為遭到
濫伐而瀕臨絕種,因此以生物技術方法人工培
養能產生與野生樟芝相同成分的技術成為重要
的研究方向。可是目前對於樟芝的細胞生理機
制仍不甚清楚,其中以菌絲體發展為子實體的
過程以及各種重要代謝產物的合成相關機制為
最迫切需要了解的,因此本報告針對樟芝的一
個 SDR 基因就其序列與蛋白質結構進行探討
與研究。
二、研究方法(一)基因還殖與序列分析
樟芝 ACSDR 的全長序列由表現序列標幟
(expressed sequence tags) 文庫裡獲得 (Chu&

62
Ch個.g, 2007; Chu et a1, 2008) 。獲得的ACSDR
基因全長轉錄為胺基酸序列後,由ComputepI/
Mw (http://au.exp晶y.orgltoo1s/pUoo1.h'恤1) 進
行蛋白質分子量仰的與等電點(PI) 的分析。
蛋白質N端細胞內運送的訊號序列 (signa1 pep
tide) 的分析則是由 Signa1P伺服器 3.0 版。句:/
/www.cbs.dtu.dk/servic間/SignaIP/)進行預測。
蛋白質於細胞內位置則是由TargetP伺服器1.1
版(http://www.cbs.d倒也/serviceslTarge佇月進
行預測﹒序列的比對則由國家生技資訊中心
(NCBI) 的 BLAST (Basic Loca1 Alignment
Search Tool) (h社:p://b1ast.ncbi.n1m且ih.gov/) 進
行分析。
(二)蛋白質結構模擬
蛋白質結構模攤採用 SWISS-MODEL 蛋
白質結構模擬系統(http://swissmodel.expasy
恆的進行預測,模擬所得之結構以PROαIECK
軟體驗證,結果為 91.4%通過驗證,由國4蛋
白質結構重疊比較分析可知模擬的 ACSDR結
構在α2 、 ß2 、 ß3 及戶4 結構單元與其它SDR有
很大的差別,導致此部位結構無法正確預測模
擬。結構繪圖與瀏覽採用DeepView (Swiss Pd1>
Viewer) 軟體 4.0.1 版,蛋白質結構以鍛帶圖
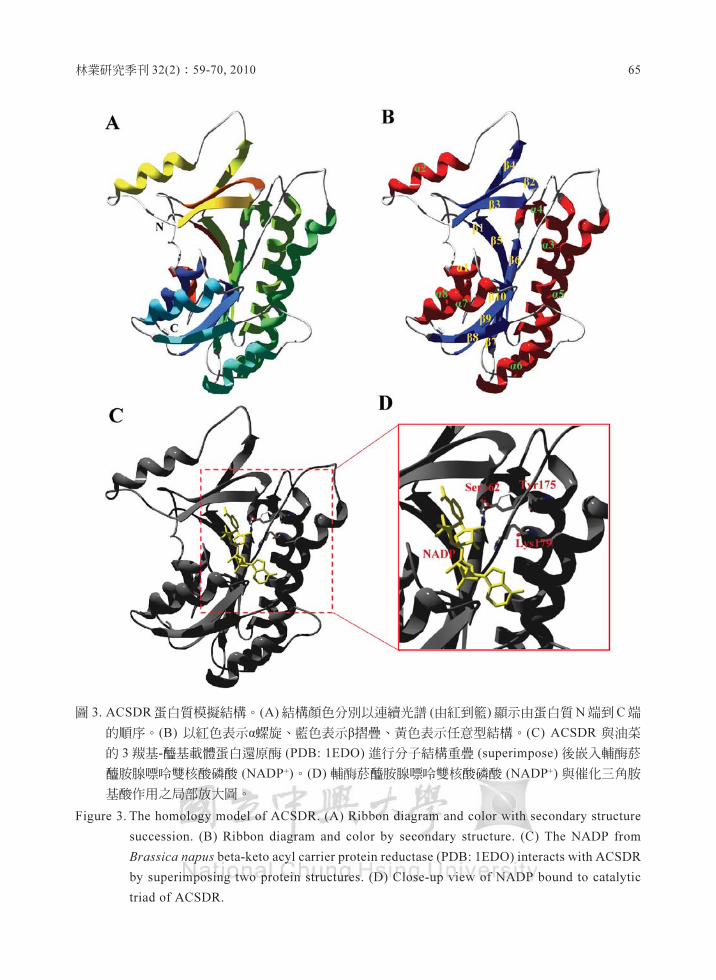
表1. ACSDR序列找到的保守性胺基酸
樟芝短鏈脫氫醇/還原醇之序列分析與蛋白質結構模擬
。ibbon) 顯示其α螺旋、戶摺疊與任意型結構,
結構顏色分別以連續光譜(由紅到籃)顯示由蛋
白質N端到C端的順序或是以不同顏色區別三
級結構差異。活性區域的三個胺基酸(催化三
角)的位置是相對於模飯熱帶念珠菌 (Candida
tropicalis) 的 3R-輝基瞳基輔團軍A脫氫梅σDB
2ET6) 上的結構和胺基酸序列所預測。輔酵菸
碰胺腺嘿時雙核酸磷酸。IADP+)是取自油菜的
3 被基﹒種基載體蛋白還原醇σDB: 1EDO). 進
行分子結構重疊 (superi血.pose) 後散入ACSDR
蛋白質結構。
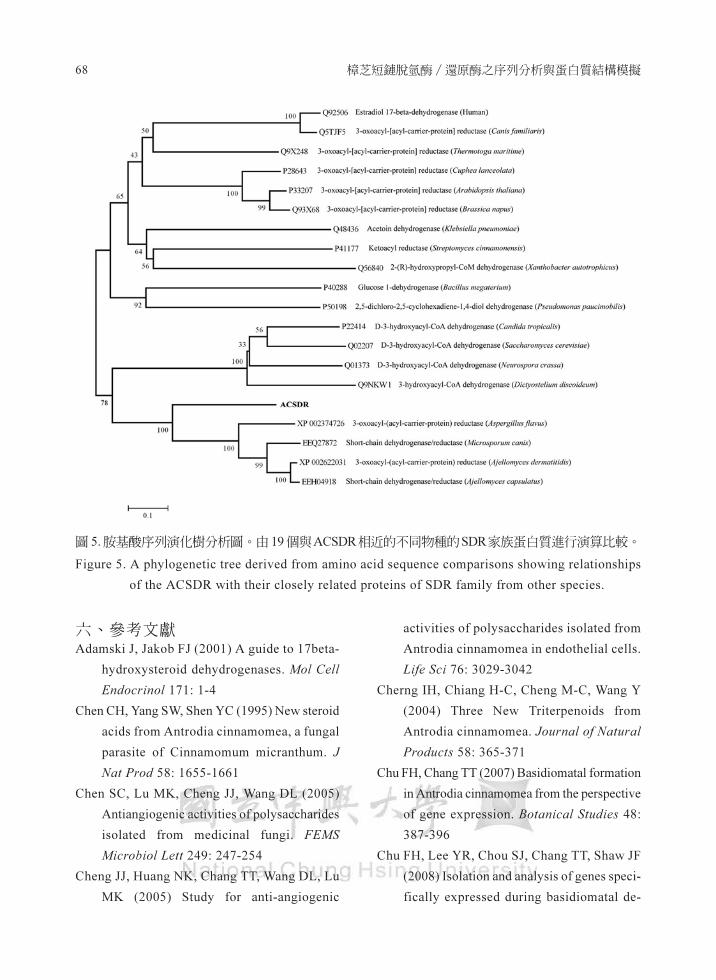
(三)親緣演化分析
親緣演化分析採用MEGA(mo1ec叫ar evo1-
utionary genetics ana1ysis)軟體 4.1 版 (Tarnura
et a1, 2007) .與 ACSDR進行比對分析的序列
由NCBI資料庫搜尋桃選不同物種的SDR家族
蛋白質,共 20個序列由軟體內的CLUSTAL程
式進行比對,比對參數為軟體預設值,親源演
化樹採用Neighbor-Joining法輔以Bootstrap試
算法重複 1000 次進行演算分析,演化樹圓型
以無根樹 (unroot) 型態繪製,區別不同物種和
基因間演化距離遠近。
Tab1e 1. Conserved sequences found in ACSDR
序
TGxxxGxG D NNAG S-Y-K N T
子。 胺基酸位置
29-36 80 106-109 162, 175, 179 201 210
功 能
輔時結合位置,維持中心11-摺板構形
保持腺嘿玲環容納區域的穩定,也幫助輔醇結合
穩定中心11-摺鈑構形活性位置
連結受質結合環與活性位置
以氫鍵結合菸幢胺環的甲瞳胺
註: ACSDR 胺基酸位置是與畢丸闢叢毛單胞菌 (Comamonas testosteroni) 17ß-溼基類固醇脫氫時
最為參考比對
三、結果與討論
(一)序列分析
由樟芝裡選殖出一個短鏈脫氫醇/還原醇
基因 (ACSDR) • 其全長共有 876 個鹼基對(圖
1) 。此基因可轉錄出 291 個胺基酸,預測之分
子量為 29.9四a 、蛋白質等電點為 9.23 。分析

轉聾研究季刊 32(2) ﹒ 59-70.2010
其蛋白質序列 .N端序列並沒有發現控制蛋白
質在個胞內運送的訊服序列 (signa1 peptid吋,
推斷此蛋白質為一個胞內蛋白﹒進一步分析
ACSDR 胺基酸序列也沒有聽現運送到其他胞
器。世幢幢或其他實體)的訊號序列,圖示此蛋
白質應該是存在於細胞質問﹒利用國家生按資
訊中心(NæI)的BLAST扭曲ic Loca1 Aligom甜
Search Tool)生物實訊工具進行ACSDR序列輿
資料庫序列進行分析比對,發現 ACSDR 序列
63
與下列幾個已知序列最為相似,分別是于讀菌
.A.jellomyc目 dermatitidi間的 3 擴基-蘆基載體蛋
白連原龍和垣鱷脫氫酪/還原醋,輿 ACSDR
胺基酸序列相似度 (identities) 分別是 56%和
自%.另外與賣翹菌 (A叩ergillus flavus) 的 3
擻基-種基載體蛋白還原醇有 53 %的相似度,
與犬小芽胞菌 (MicT03porum canis) 的短鍵脫
氫酪/還原蘭也有自%的相似度,這些基因
皆屬於 SDR家旗。
肝GGCAGACCGT口 GTCACAGGTCG口GGGCATCTCACAAAiTCATACGGTCGCGGTCTGC竹GliGGAGAGGTCG且ATCATCACCGGA 90 MADRLSQVAGHLTNSYGRGLLVGEVAIJ T G
GCTGCTCAGGGAATI叫~TAGCGCTGCCATTCTGTTCGCCAAGGAAGGTGCCAAAGTCGTGGTTACTGACπGGATGCGAAAAAGGCC 180 A A Q G 1 G R S A A 1 L P A K E G A K V V V T D L D A K K A
叫AGCAr.TAr.鬥'f,Ar:r.A r.ATT AAr.CAr.r.cAG叮甜;r.r.ATr.cr.crrr.rr,r.鬥仗;r('.f.Tr.AT,盯CA盯r.cCGATr.ArnrlX了TCA r.AA叫行 270
Q A V V D E 1 K Q A G G D A L A V G G 0 V S A 0 0 F P Q K V
GT凹AGGCTACGCTCAAGAAA甘TGGCAAAATCAACCATATAGTGAATAA囚CAGGCTTCACATATGATCGCATGCTTCACACTA囚CCC 360 V E A T V K K F G K I N H I V N N A G F T Y D R M L 11 T T P
GATGATGCtïTTGATATCA1CATTAAAGTGCA臼TTCGCGCGCCGTTCAGACTGATTCGTGCTGCTGCGCCTTATATGCGCATCAAGGGC 450 D D A F 0 1 ! 1 K V I1 V R A P F R L 1 R A A A P Y M R 1 K G
AACATTGAGAACCGCTCAGTCATCAATGT們付TTCCGTGTCCGGTCTGCACGGCAACGTCGGCCAAGCCAATTACGα刮目口AAGTCT 540 N 1 E N R S V 1 N V F S V S G L 11 G N V G Q A N Y A A A K S
GCTGTCGTCGGCCTGACGAAGACTATCTGCAAGGAGTGGGGTGTGTTTGGTGTGAGGGCGAATGT四TCGCGTTCGGCTACGTACAGACT 630 A V V G L T K T 1 C K E W G V F G V R A N V V A F G Y V Q T
CG凹TGACAGCCGCAAAAGAGGAT'品TGCAAOCATOGAGATAGA囚GTAAGAAAGTTGCC竹品GCATTC囚M閏GO臼GA鬥GCGACT 720 R L T A A K E D G A T 1 E [ D G K K V A L G ! P K G Q J A T
GCTGCGGCMG囚GGTCGGGTCCCACGTCTGGTATTCCACTGGGACGGGGTGCMCCCCAGACGAGG囚GCGGCTGGTATACT付TCCTT 810 A A A S G S G P T S G 1 P L G R G A T P D E A A A G 1 L F L
GC什CAC口它付GC鬥'CAmGTGTCA且CCACACGCTCGAAGTCA田間 A品AGC品GCAT口'GAGTT品CTCA服刑ccrCAAT回 A 帥。A S P L A S F V S G H T L E V T G G A G 1 *
T和CACATCATA口OCCCTTATATG'γ鬥CCTTCCTTOGTATCAATAGATACAGCCTTTTGTATCTCCGACATCAATATACTCCCGCATOG 990 CCATAAAAAAAAAAAAAAA 1019
圖 I. A巳百VR序列全長與轉轉之胺基酸。
Figure 1. Nuclcotidc 阻d infcrrcd amino 臨id scqucnccs of ACSDR
(二)保守序列分析
雖然不間物種間 SDR 家農序列的相似度
不高,但是由已知的蛋白質三及結構分析,發
現結構上有很高的相似性,這個在 SDR 家族
共通的呻摺疊結構稱為Rmmnann摺疊σm叫
“凶.1995) ﹒這個相似的摺壘結構大概是由 250
到 350個胺基酸長度所構成的,其中包含了幾
個重要的保守序列。再靠近蛋白質N端有一個
TGxxxGxG的保守序列,還是第鹽胺臨暸時二
核酸的腺哪告結合部位,而此蛋白質家接重要
的酵索個it三角 (catalytic triad) 員Ij由絲胺酸
(Se,)、酪胺酸(可,)和離胺酸(Ly.) 組成,以
上這兩個保守序列是 SDR 家族一定有的螺守
性區域(圍 2) .另外還有幾個重要的保守序列

64
如NNAG是與TG盟xGxG共同種定Rossmann
摺疊中心醉摺桓構形,同時也幫助輔酵的結
合;一個單獨的天門各酸 (A.p) 穩定腺嘿峙的
結脅,一個天各種胺酸 (A.n) 連結受質結會環
與活性位置以及一個蘇胺酸 (Thr) 以氫鍵結合
菸種胺環的甲幢胺﹒經由比對 ACSDR和畢丸
鬧聲毛單胞菌 (Comamonas 翩翩terone) 1713-
邂基顯固神脫氫酵的胺基酸序列和蛋白質二級
結構,可以清楚的預測出這些保守序列,在表
一列出7預測ACSDR的錢個重要保守序列以
及相對應在ACSDR的胺基酸位置﹒ SDR這個
大家族耳依揖某些保守序列的股基酸差異,往
下分額為如前述的四個子家族,根據Kallbe唔
...cIOR XEEFOGZ OE3?4? A 區 H72
...00918 002201 '" m. .. '" QO'''' '"
ACSDR " XEVQ直0023147 9~ n027~72 97 "間自嚕'" " 月E2日EZZO? m
241 . "" QOI J7 J "
樟主題組脫詛.J!Ii/還原龍之序列分析與賣自質結情模擾
等λ於 2002 年所發表的SDR于家族分類方法
(Ka11berg et 剖. 2002) • ACSDR可歸額為一般
型和1.阻1叫}﹒由於半數以上的 SDR屬於一般
型,因此P間.on等λ又於 2003 年將一般型的
SDR再細分7七個于家旅σ間S曲 et al, 2003) •
分別為 cDld 、 cDle 、 cD2、 cD3、 cPl 、 cP2
和c凹,依揖區分的單則, ACSDR屬於 cDld
型, cDld型的SDR在P闖闖n等人的研究申,
人額的頭固醇脫氫醇和前列腺素脫氫時都屬於
此型酵素,這些壘質的結構與真菌中的許多二
次代謝矗物相似,因此可以推論ACSDR可能
參與樟芝的某些重要成分生告戚。
" " " 9 m '" 3 圖。
... '" ." ." ". ... ."
圖 2. ACSDR與其他 SDR胺基酸之多重日序列比對. TGxxxGxG和NNAG兩個保守序列區域分別以
單飽和雙峰標示,活性值化三角以黑色圓形標吉﹒基因代號請書照圖 5 •
Figure 2. Multiple alignmcnt of amino scid scqu聞自CS ofACSDR and other SDR gcncs. The catalytic
triad (Ser-Tyr-Lys), TGxxxGx.G and NNAG co目前ved residues are indicated with black
circles. single u且derline and doublc underli間. respectively. Rcfer 也E 臨閻明ionnumbe四 to
figure 5 for gene functions 個d 血Clr specles
(三)蛋白質結構模擬
透過蛋白質結構的模擬預測可以7解蛋白
質的生化特性、酵素活性和壘賀他化過程等資
訊,幫助後續實驗的芳自輿設計。 ACSDR 胺
基酸序列經由 SWISS-MODEL 這個生物資訊
工作站,比對目前已知蛋白質結構H的資料庫,
找到輿 ACSDR結構最相近的蛋白質,是一個
已經由X光繞射法解出蛋白質結晶結構的熱帶
念珠菌 (Candida tropicalis) 的 3R-擅基種基輔
梅 A 脫氫膺,其蛋白質資料庫 σDB) 的標號
是 2ET6 '以此蛋白質為模缸,模摸出了 AC
SDR的三級結構(圖 3) . 由結構圖可以看出典
型的呻摺疊組成的 Rossmann 摺疊(圖 3A) •
在酵素的作用悟性區域可以看到催化三角的三
個重要跟基酸 Ser162 、 Tyr175 &. 、 Lys179 .利用
分子結構重壘。uperimpose) ,將油菜的 3 嫂
基種基載體蛋白道原關句輔鷺葫蘆歐腺釀時雙
核酸磷酸(N曲.1")最λACSDR的酵素活性區
壩,此區域即酵素進行還原反應的重要位置個
3C和 3D) .另外,將熱帶念珠菌的 3R-糧基瞳
轉業研究季刊 32(2) ﹒ 59-70.2010 6'
D
圍 3. ACSDR蛋白質模擬結構﹒ (A)結構顏色分別以連續光滑(由紅到盤)顯示由蛋白質N端到C端
的順序. (8)以紅色表示a螺龍、藍色表示F摺疊、黃色表示任意型結構﹒(C) ACSDR 興油菜
的 3攤基-種基載體蛋白還原蘭 σDB: lEDO) 進行分子結構重疊 (superimpose) 後骰λ輔酪菸
鹽眩服嘿怖雙核酸磷酸。河ADP') • (D) 輔藺懿鹽眩臨嘿時雙核酸磷酸(NAD的輿偕化三角肢
基餅干用之局部放大圖﹒
Fig四海 3. The homology model of ACSDR. (A) Ribbon diagram 血d color with secondary s當uctu血
succession. (B) Ribbon diagram 阻d ∞lor by secon.也可 struc扭扭(C) Th. NADP from
甜甜s,can申us bcta-kcto acyl carrier protein reductase (pDB: lEDO) interacts w地ACSDR
by superimposing two protcin structures. (0) Close-up view of NADP bound to cata1ytic
triad of ACSDR

66
基輔酪 A 脫氫團軍 (PDB: 2ET6) 和油菜的 3 糢
基-種基載體蛋白還原蘭 (PDB: !EDO) 兩個蛋
白質與ACSDR進行分子結構適重疊 (superim
pose) 分析,在ACDR結構具有酵素活性中心
的結構單元 (domain) 在三級結構排列上較相
近,但在α2 、戶2 、 ß3 及ß4 構成的結構單元有
明顯的差異性(圖 4) ,此結果是否顯示ACSDR
在受質選擇上異於其它 SDR 仍有待更進一步
酵素活性分析來證賞。
(四)演化分析
將樟芝的 ACSDR胺基酸序列與不同物種
的 SDR 家族蛋白質進行序列比對與親緣演化
分析,由所演算出來的演化樹可以發現(圖 5) ,
SDR家族蛋白質在不同物種間的差異,由演化
樹圖可以發現,哺乳動物、植物和原核細菌類
的 SDR各自形成一個群組,而樟芝的ACSDR
分別與同樣真菌類的黃鞠徵菌 (Aspergillus
fla悶。、犬小胞子徹菌 (Micro司porum canis) 和
阿傑羅芽生徵菌 (Ajellomyces dermatitidis) 的
SDR家族蛋白質歸屬間一群組,不過由演化距
離還看, ACSDR與其他真菌的 SDR似乎又獨
自成一屬,顯見在酵素活性與受質選擇性上應
有很大的差異,而這也可能產生樟芝特有的代
謝產物的重要原因。
四、結論
樟芝的藥用與保健價值不管在民間或是科
學研究上都受人重視,因為台灣特有種的牛樺
樹是樟芝唯一寄生樹種,目前由於此保育類牛
樟樹數目稀有,也使得樟芝一時洛陽紙貴,人
工培養的商絲體樟芝成分有別於野生的于實
體,而菌絲體由人工培養成子實體的條件與技
術上仍有瓶頸需要突破,因此造成是市面上販
售的樟芝保健食品品質參差不齊。若要以生物
技術方法來研究樟芝生長與重要成分生合成,
就必須由基因體、蛋白質體與代謝體學等基礎
研究來輔助。樟芝的重要成分大致已經分析,
但是這些代謝產物的生合成路徑與相關基因都
仍不甚清楚。目前短鍵脫氫醇/還原醇 (SDR)
樟芝短鏈脫氫醇/還原醇之序列分析與蛋白質結構模擬
在二次代謝產物生合成的研究會逐漸受到重
視,主要是 SDR 在許多二次代謝產物生合成
中扮演著產物最後修飾 (tailoring) 的重要功能
(Liu & Moon, 2009; Mitsuguchi et a!, 2009) ,
尤其在樟芝的三宿類與類固醇衍生物是一般認
為最具有應用價值的化合物, ACSDR 即有可
能參與這些重要天然產物的生合成過程。因此
本研究針對樟芝的表現序列標幟所選殖出來的
ACSDR 基因進行了蛋白質序列與功能結構上
的探討。由序列比對與保守區域分析來預測酵
素功能,再由蛋白質結構模擬了解酵素活性區
域、輔梅和受質的4僵化立體空間結構。岡時也
由親緣演化的關係,了解 SDR 家族在不同物
種間的差異,而ACSDR與其他真菌的SDR在
演化也有明顯的分歧,顯示此酵素在樟芝特有
的功能,如參與樟芝特有代謝產物的生合成
等。在蛋白質結構分析上, ACSDR 與其他以
之結構 SDR 在蛋白質的酵素活性區域包含催
化三角以及輔醇結合區域都有相似的結構,但
在如圖 4 的α2 、 ß2 、戶3 及ß4 區域卻出現明顯
的差異,而此結構單元有如蓋子般蓋住活性區
域,可以合理推測此結構可能影響不同 SDR的
受質選擇性,而 ACSDR參與樟芝哪些重要謝
產物的生合成則是未來重要的研究方向。本研
究提供了充分資訊幫助 ACSDR後續在進行蛋
白質表現和酵素產業上的發展,以及更進一步
成為運用在代謝體工程學上進行基因表現調控
以增加代謝產物產量時的標的基因。台灣特有
的樟芝具有珍貴的醫藥用途,相信在分子生物
學技衡的輔助下,可以在保育稀有牛樟樹同
時,以人工培養技術或酵素工程生產方式提供
市場所需的樟芝和有效成分,也讓此有台灣森
林中之紅寶石美譽的真菌造福更多人群。
五、誌謝本文承蒙行政院國家科學委員會提供研究
經費。7NSC-23!7-B-002-009) 及台灣林業試
驗所提供樟芝菌絲體和子實體材料。
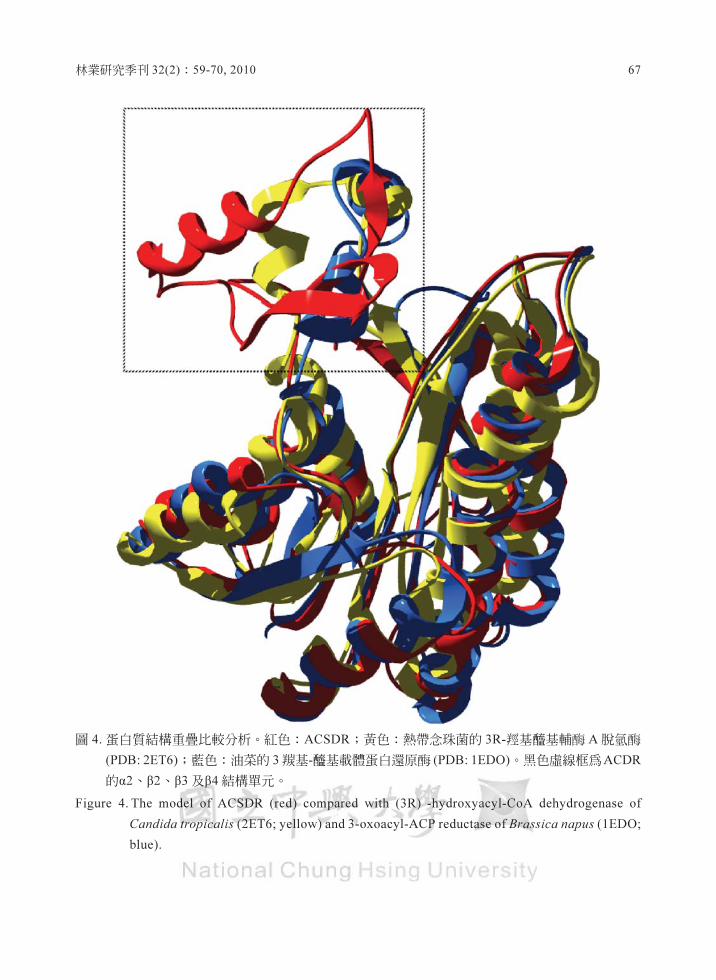
轉業研究季刊 32(2) ﹒ 59-70.2010 67
圖 4 蛋白質結構重疊比較持析﹒紅色: ACSDR ,黃色:熱帶盒珠菌的 3R-糧基種基輔醇A脫氫構
σDB:2ET6) ,藍色:泊蔡的 3皺基-種基載體蛋白還原萬σDB: 1EDO). 黑色虛飽權l14ACDR
的αz 、 p2 、 p3 且p4 結構單元﹒
Fig四海 4. Tl世e model of ACSDR (red) compar吋 with (3R) -hydro:可曲:yl-CoA dehydrogenase of
Condida帥'picalis (2ET6; yellσw)血d3-oxoa.叮l-ACP reductase of Brassica nap間 (IEDO,
blue)
68 樟主題組脫詛.J!Ii/還原龍之序列分析與賣自質結情模擾
'"間。但闢叫 11......叫…"叫
OSTJFS J如“ooy1-1a<y1-.cam ... -proI<吋削岫..(C",,~后""m,..叫
。慘叫 H叫(呵呵叫"一……
'"甜 J-o,.吋{…o;nl…叫阿/1,/叫
" lω
3嘶叫1…..-圳…{叫…
(pJX.ω3呵)1-[詞“聊聊~i "J……叫
“』
un
()IS436 ^叫d膽的…阱l" i<II" fmn'叫
NI In K叫……一
叫 G血哪一……
Q使毒品o 2-(R"hydro均冒-'晶M <kh,-dn滄,-糾W蛤加鯽倆個,.關問時
" 叫 2…一…………
咐
A目""
間"叫叫蚓……C叫呻削的
。0ZW7 D-l 峙的麓)"a<j'I-CoA dchldrogcna叫~~雕"'~Uw)
。酬的……岫……叫詞,
Q9NKW I J…………
XP OO".J747:!6………… 間現n ……岫剛結…'叫
XPOOl叫l ……一……
固樹岫…向…(叫個叫
-" 圖5 眩基酸序列演化樹封析圖﹒由 19個與ACSDR相近的不同物種的SDR家族蛋白質進行演算J:IJ較
Figure S. A phylogenetic tree derived 告。m amino acid 開quence comparisons showing relationships
of the ACSDR with their closely relatcd protcins of SDR family from other species
六、參考文獻Adamski J, Jakob FJ (2001) A guidc to 17beta
hydroxysteroid dehydrogenases. Mol Cell
Endocrino/l71: 1-4
Ch血CH, y"""SW, Sh血YC(1995)N開臨roid
acids from Antrodia cinnamomea. a fungal
p虹asite of Cinnamomum micranthum. J
Nat Prod 58: 1655-1661
Chen SC, Lu MK, Cbl間g JJ, Wang DL (2005)
Anti血giogenic activities ofpolys國charid,個
isolated from medicina1 fungi. FEk甜
Microbiol Lett 249: 247-254
Cheng JJ, Huang NK, Chang TT, WI阻gDL.LuMK (2005) Study r.叮阻ti-祖glOge血C
activities ofpolysaccbarides isolated 金.om
Antrodia cinnamomea in cndo也.clia1 cells
Life Sci 76: 3029-3042
ChemglH, C趾ang H-C, Cheng M-C, WI阻gY
(2004) Three New Triterpenoids from
Antrodia cinnamomea. Journal ofM叫ural
Pro曲ct, 呵。 365-371
ChuFH, C~ TI (20的B回idiomata1 form醋。n
inAn個diacinnamomea企um也叮當咚)OCÜve
of gene exp扭曲ion. Botanical SI個di由 48
387-396
Chu FH, Lee YR, Chou SJ, Chang TT, Shaw JF
(2008) 1so1ation and血alysis of genes speci
fically expressed dur四g basidiomatal de-

林業研究季刊 32(2): 59-70, 2010
ve10pment in Antrodia cionamomea by sub
tractive PCR and cDNA microarray. FEMS
Microbiol Lett 280: 150-159
Finsterbusch A, Lindemann P, Grimm R,
Eckerskom C, Luckner M (1999) Delta
(5) -3beta-hydroxysteroid dehydrogenase
from Digitalis lanata Ehrh.-a multifunctional
enzyme in steroid metabolism? Planta 209:
478-486
Han HF, Nakamura N , Zuo F, Hirakawa A, Yokozawa T, Hattori M (2006) Protective
effects of a neutral polysacch缸ide isolated
from the mycelium of Antrodia cinnamomea
on Propionibacterium acnes 阻d lipopol
ysaccharide induced hepatic inj四ymmlce.
Chem Pharm Bul/ (Tokyo) 54: 496品。
Hseu YC, Chang WC, Hseu YT, Lee CY, Yech
YJ, Chen PC, Chen JY, Yang HL (2002)
Protection of oxidative damage by aqueous
extract from Antrodia camphorata mycelia
in normal human erythrocytes. Lij全 Sci71:
469-482
Hseu YC, Wu FY, Wu 刀, Chen JY, Chang W耳,
Lu FJ, Lai YC, Yang HL (2005) Anti-in
flammatory p。但ntial of Antrodia Camphora旭
through inhibition of iNOS, COX-2 and cy
tokines via the NF -kappaB pathway. 1nt 1m
munopharmacol 5: 1914-1925
Hsiao G, Shen MY, Lin KH, Lan MH, Wu LY, Chou DS, Lin CI宜, Su CH, Sheu JR (2003)
Antioxidative and hepatoprotective effects
of Antrodia camphorata extract. J Agric
Food Chem 51: 3302-3308
Hung-Chen Chiang, De-Peng W, Chemg IW,
Chuen-Her U (1995)A sesq凶t目pene lactone, phenyl and biphenyl compounds from
Antrodia cinnamomea. Phytochemistry 39:
613-616
Jomvall H, Persson B, Krook M, Atrian S,
69
Gonz過lez-Duatte R, Jeffery J, Ghosh D
(1995) Short..,haindehydr甸回扭曲的duc'臨閱
(SDR). Biochemistry 34: 6003-6013
J omvall H, Persson M, J effery J (1981) Alcohol
and polyol dehydrogenases are bo血 divided
into two protein types,阻d s甘uctural
properties cross-relate也e different enzyme
activities within each type. Proc Natl Acad
Sci USA 78: 4226-4230
Kallberg Y, Oppermann U, Jomvall H, Persson
B (2002) Short-chain dehydrogenases/re
ductases (SDRs). Eur J Biochem 269
4409-4417
Kristan K, Brunskole M, Stojan J, Rizner TL
(2009) Two homologous fungal carbonyl
r叫回祖S個WI也 differ凹I阻h甜揖espe地沮cr世es
Chem Biol 1nteract 178: 295-302
Krook M, Marekov L, Jomvall H (1 990)
Purification and.甘uctural characterization
of placental NAD (+) -linked 15-hydro
勾pros個gl個din dehydrogenase. The primary
structure reveals the enzyme to belong to
the short-chain alcohol dehydrogenase
E組IÎly. Biochemistry 29: 738-743
Lee IH, Huang RL, Chen CT, Chen HC, Hsu
WC, Lu MK (2002) Antrodia camphorata
polysaccharides exhibit anti-hepatitis B
virus effe晶• FEMS Microbiol Lett 209: 63-67
Liu JJ, Huang TS, Hsu ML, Chen CC, Lin WS, LuFJ, Chang WH (2004) Antitu血ore扭扭扭
of the pattially purified polysaccharid問
世四nAntrodia間血ph田副a and the mechanism
of its action. Toxicol Appl Pharmacol20 1:
186-193
Liu ZL, Moon J (2009) A novel NADPH
dependent aldehyde reductase gene from
Saccharomyces cerevisiae NRRL 可ι12632
involved in the detoxification of aldehyde
inhibitors derived 企om lignocellulosic

70
biomass conversion. Gene 446: 1-10
Mitsuguchi H, Seshime Y, F吋ii 1, Shibuya M, Ebizuka Y, Kushiro T (2009) Biosynthesis
of steroidal antibiotic fusidanes: functional
analysis of oxidosqua1ene cyclase and sub
sequent tailoring enzymes 企omAspergi11us
fumigatus. J Am Chem Soc 131: 6402-6411
Nak血\Ura N, Hirakawa A, Gao 呵, KakudaH, Shiro M, Komatsu Y, Sheu CC, Hattori M
(2004) Five new ma1e扭曲d succinic acid
derivativ閩台om the myceliu血 ofAn甘odia
camphorata and 也eir cytotoxic effects on
LLCtu血or cellline. J Nat Prod 67: 46-48
Oppermann U, Fi11ing C, Hu1t M, Shafqat N, Wu X, Lindh M, Shafqat J, Nordling E, Kallberg Y, Persson B, Jomva11 H (2003)
Short-chain dehydrogenases/reductases
(SOR): the 2002 update. Chem Biollnteract
143-144: 247-253
Persson B, Ka11berg Y, Oppermann U, Jomva11
H (2003) Coenzyme-based functional as
siguments of short-chain dehydrogenasesl
reductases (SORs). Chem Biol Interact
143-144: 271-278
Phuong OT, Ma C-M, Hattori M, Jin JS (2009)
b由ibitory effects of antrodins A-E from
Antrodia cinnamomea and由eir metabolites
on hepatitis C virus protease. Phytotherapy
Research 23: 582-584
Phuong OT, Ma CM, Hattori M, Jin JS (2008)
k由ibitory effects of antrodins A-E from
Antrodia cim祖momea and their metabolites
on hepatitis C virus prote晶e. Phytother Res
Schwartz MF, Jomvall H (1976) Structural
analyses of mutant and wild-type a1cohol
dehydrogenases 壺四n drosophilamelanog扭扭E
Eur J Biochem 68: 159-168
Seidel S, Kreiss W, Reinhard E (1990) 05-3- /3 -hydroxysteroid dehydrogenaseID5-04-
樟芝短鏈脫氫醇/還原醇之序列分析與蛋白質結構模擬
ketosteroid isomerase (3b-HSO), a possible
enzyme of cardiac glycoside biosynthesis, in cell culture and plants Digitalis lanata
Ehrh. Plant Cell TlSSueα莒酬 Cult38: 337-344
Shen CC, Shen YC, Wang YH, Lin LC, Oon MJ, Liou KT, Wang WY, Hou YC, Chang TT
(2006) New I血。咖nes and naphthoquinone海
isolated from Antrodia s叫血叩開祖d their
祖tioxidative burst activity in hu血血
leukocytes. Planta Med 72: 199品3
Shen YC, Wang YH, Chou YC, Chen CF, Lin
LC, Chang TT, Tien 間, Chou CJ (2004)
Eva1uation of也e血ti-infl血unatory部tivi句
。fzhankuic acids isolated from 吐le fruiting
bodiesofAn甘od祖曲血phorata. Planta Med
70: 310-314
Song TY, Yen GC (2002) Antioxidant properties
of Antrodia camphorata in sub血erged cu1側目.
J Agric Food Chem 50: 3322-3327
T缸nura K, Oudley J, Nei M, Kumar S (2007)
MEGA4: Molecular Evolutionary Genetics
Analysis (MEGA) Softwaτe Version 4.0
Mol Biol Evo124: 1596-1599
Tan KC, Heazlewood JL, Mi11ar AH, Thomson
G, Oliver RP, Solomon PS (2008) A signa1ing
regulated, short-chain dehydrogenase of
Stagonospora nodorum regulates asexua1
development. Eukaryot Cell 7: 1916-1929
Wang 凹, Tseng HW, Chou CJ, Tsai TH, Chen
CT, Lu MK (2003) The vasorelaxation of
Antrodia camphorata mycelia: involvement
of endothelial Ca (2+) -NO-cGMP pathway
Life Sci 73: 2769-2783
Yang HL, Chen CS, Chang WH, Lu FJ, L副 YC,
Chen CC, Hseu TH, Kuo CT, Hseu YC
(2006) Growth inh出版on and induction of
apoptosis in MCF -7 breast cancer cells by
An個diacam抖田間包αmcerLett231:215-227