吃軟不吃硬:警戒色昆蟲的神祕...
TRANSCRIPT

國立臺灣師範大學生命科學系碩士論文
吃軟不吃硬:警戒色昆蟲的神祕
次級防禦機制
Too hard to swallow: A secret secondary
defense of an aposematic insect
研究生:王露翊Lu-Yi Wang
指導教授:林仲平博士 Dr. Chung-Ping Lin
黃文山 博士 Dr. Wen-San Huang
中華民國 106 年 11 月 01 日

i
致謝
長長兩年多的碩士生涯,短短的精采的過完了。回首過程中的一切辛苦與
挑戰,我何其幸運的能夠沉浸其中,享受著與這個有趣的題目相伴的甜蜜時光。
感謝我的指導老師林仲平博士,他總是能夠適時的為我打氣,每次與他談話後
總覺得實驗又充滿希望!也謝謝共同指導我的黃文山博士,和口試委員顏聖紘博
士,點出論文的問題與建議。
實驗室與師大共同努力的夥伴們(惠芸、若凡、祐薰學姊、Wataru、小 A、
緊張哥、一休哥、小哈、震邑、Pincess、Leo、Ian、劉班、俊佑、阿薇、中中),
感謝他們的陪伴與時間,陪我聊天放鬆;謝謝馥慈師母,替我處理出國與出差的
各種繁瑣行政工作。謝謝溫泉扮演著野外工作與生活中不可或缺的要角;也謝謝
昆蟲館唐欣潔館長與春捲龍學長,協助我飼養實驗所需的象鼻蟲與提供食草;還
有為實驗犧牲的象鼻蟲們,辛苦你們了。感謝阿磐學長在統計與 R上提供許多
幫助;謝謝Dr.HamedRajabi 在昆蟲結構與物理名詞上給了許多建議。
最後感謝我的家人,義無反顧的支持我想做的事情。我很幸運能擁有你們!

ii
中文摘要
1. 抵禦掠食者攻擊是獵物適應上重要的一環。獵物的次級防禦機制 (例如毒
性) 常伴隨著警戒色的出現,並可藉由避免被捕食讓掠食者與獵物彼此獲
利。因此警戒色獵物被預期具有和警戒色彩一起演化的有效次級防禦機制。
2. 早在將近一百五十年前,華萊士就已經提出球背象鼻蟲 (Pachyrhynchus) 以
鮮豔的花紋來警告掠食者,並以堅硬的外骨骼作為防禦的假說;然而這項假
說至今依然無實驗證據驗證。本研究檢驗球背象鼻蟲的次級防禦機制假說,
並探討堅硬外骨骼在球背象鼻蟲身上所扮演的生態功能。
3. 我們以斯文豪氏攀木蜥蜴(Japalura swinhonis)作為掠食者,實驗設計四組
象鼻蟲處理(軟/硬;有足鉤/無足鉤)進行操控性行為實驗測試其存活率。
並以比較破壞球背象鼻蟲的外骨骼,和掠食者共域昆蟲獵物所需的力量 (牛
頓),與掠食者的咬合力大小,來了解球背象鼻蟲的硬度在生態上的重要性。
最後用氣相色譜質譜法 (GC-MS) 分析球背象鼻蟲體內是否有毒性或驅避
性的化合物。
4. 所有「硬」球背象鼻蟲被蜥蜴攻擊後都立即被吐出且存活。相較之下「軟」
球背象鼻蟲則在被多次咀嚼後吞下。此外不論足鉤存在與否,蜥蜴對球背象
鼻蟲的捕食結果都與上述相同。
5. 「硬」球背象鼻蟲的硬度顯著高於共域昆蟲的平均硬度和當地蜥蜴族群的平
均咬合力。
6. GC-MS分析出球背象鼻蟲體內四種可能的化合物(ethyl propyl ether、
diethylene glycol、butylated hydroxy toluene以及 cadalene) 均無毒性或對脊

iii
椎動物有驅避的功能。
7. 本研究指出具警戒色物種的硬度是一項有效抵禦捕食者的次級防禦機制,亦
提供ㄧ個探討脊椎動物掠食者與具有警戒色獵物之間時空互動的研究模
式。
關鍵字:防禦機制、警戒色、硬度、斯文豪氏攀木蜥蜴、球背象鼻蟲、捕食-
被捕食交互作用、次級防禦、台灣

iv
Abstract
1. Anti-predator strategies are significant components of adaptation for prey species.
The advance of the secondary defenses like toxins have led to the evolution of
warning colours that benefit both prey and predators by mutual avoidance.
Therefore aposematic prey are expected to possess effective secondary defenses
evolved together with their warning colours.
2. This study tested the hypothesis of secondary defensive function and ecological
significance of the hard body in aposematic Pachyrhynchus weevils, which was
pioneered by Alfred Russel Wallace nearly 150 years.
3. We used predation trials of Japalura tree lizards to assess the survivorship of
‘hard’ (mature) vs. ‘soft’ (teneral) and ‘claw’ (intact) vs. ‘clawless’ (surgically
removed) weevils. The ecological significance of the weevil’s hard body was
evaluated by assessing the hardness of the weevils, local insect prey, and the bite
forces of lizard populations. The existence of toxins or deterrents of the weevil
was examined by Gas Chromatography-Mass Spectrometry (GC-MS).
4. All ‘hard’ weevils were spat out and survived the attack by the lizards. By contrast,
the ‘soft’ weevils were chewed and subsequently swallowed. The results were the
same regardless of presence or absence of the weevil’s tarsal claws.
5. The hardness of ‘hard’ Pachyrhynchus weevil was significantly higher than the
average hardness of insect prey in the same habitat and mean bite forces of local
lizards.

v
6. The four candidate compounds of the weevil identified by GC-MS (ethyl propyl
ether, diethylene glycol, butylated hydroxy toluene and cadalene) had no known
toxic or repellent functions against the vertebrates.
7. These results reveal that the hardness of aposematic prey functions as effective
secondary defense, and provide a framework for understanding spatio-temporal
interactions between vertebrate predators and aposematic insect prey.
Key-words: Anti-predator strategy, aposematic colour, hardness, Japalura lizard,
Pachyrhynchus weevil, predator-prey dynamics, secondary defense, Taiwan

vi
Contents
致謝 ................................................................................................................................. i
中文摘要 ....................................................................................................................... ii
Abstract ......................................................................................................................... iv
Introduction .................................................................................................................... 1
Materials and methods ................................................................................................... 5 Ethics statement ..................................................................................................................... 5
Weevil and lizard ................................................................................................................... 5
Colour and mobility of ‘hard’ and ‘soft’ weevils .................................................................. 7
Behavioral trials .................................................................................................................... 9
Insect prey and hardness measurement ............................................................................... 10
Chemical analysis ................................................................................................................ 12
Statistical analyses .............................................................................................................. 13
Results .......................................................................................................................... 13 Effects of the hardness and claws ........................................................................................ 13
Hardness of prey and lizard’s bite force ............................................................................. 15
Biochemical compounds of P. sarcitis kotoensis ................................................................. 16
Discussion .................................................................................................................... 17 Effective secondary defense ................................................................................................. 17
Multiple secondary defenses ............................................................................................... 19
Mechanical mechanism of hardness .................................................................................... 20
Adaptive defense against lizard’s predation ....................................................................... 21
References .................................................................................................................... 24
Figures………………………………………………………………………………..31
Supplementary tables………………………………………………………………...39
Supplementary movies……………………………………………………………….56

1
“… tropical weevils which have the elytra and the whole covering of the body so hard
as to be a great annoyance to the entomologist, …, they cannot be pinned without first
drilling a hole to receive the pin, and it is probable that all such find a protection in
this excessive hardness.”
“The insects which others imitate always have a special protection, which leads them
to be avoided as dangerous or uneatable by small insectivorous animals; some have a
disgusting taste …; others have such a hard and stony covering that they cannot be
crushed or digested;…”
Alfred Russel Wallace, 1867
Introduction
Predation is one of the most visible selective forces driving the ecology and evolution
of organisms in nature (Abrams 2000). Therefore, evolving effective anti-predator
strategies is a significant component of adaptation for many prey species. Diverse
defensive strategies have evolved at a range of specific stages of the encounters
between predators and preys (Stevens 2013). These anti-predator strategies can be
classified as the ‘primary’ and ‘secondary’ defenses depending on the timing when
they are performed (Ruxton, Sherratt & Speed 2004). The primary defenses serve to
avoid detection (crypsis) by operating before predators initiate prey-catching
behaviour, or to prevent pursuit by advertising themselves unprofitable to predators
(mimicry, warning signals) (Robinson 1969; Edmunds 1974). The secondary defenses

2
are post-detection defenses that function to increase the chances of surviving
prey-capturing process (deflection or startle), or to make an encounter unprofitable to
predators (e.g. spines, stings and toxins) (Robinson 1969; Edmunds 1974). The
development of the secondary defenses using chemical or physical deterrents have led
to the evolution of warning signals in prey that benefit both well-defended prey and
their potential predators by mutual avoidance (Summers & Clough 2001; Sherratt &
Beatty 2003; Ruxton et al. 2004). Well-known examples of co-evolution between
warning signals and prey defenses include the generation of aposematic colouration
and toxicity in the poison frogs (Summers & Clough 2001; Maan & Cummings 2011)
and ladybird beetles (Blount et al. 2012).
Pachyrhynchus weevils (Germar, 1824) (Coleoptera: Curculionidae: Entiminae:
Pachyrhynchini) are a group of brilliant metallic-coloured weevils distributing in the
Old World tropics (Fig. 1a) (Wallace 1895; Schultze 1923; Tseng et al. 2017). The
observations of diverse colours of more abundant Pachyrhynchus weevils (as models)
being mimicked by several relatively rare species of longhorn beetles (Cerambycidae:
Doliops) suggested the aposematic function of their colouration (Wallace 1867;
Dickerson et al. 1928; Barševskis 2013). A recent manipulative experiment showed
the first empirical evidences that the conspicuous colouration of Pachyrhynchus
weevils could function as effective warning signals (the primary defense) to prevent
predacious pursuit by Japalura lizards (Fig. 1c) (Tseng et al. 2014). In those
behavioural trials, the lizards attacked weevils without conspicuous colours at higher
rates than weevils with intact colours. During those trials, the lizards were observed to
display irritating behavioral response (spitting out & throwing off) after biting the

3
weevils, indicating the unpalatability of the prey. Moreover, the weevils frequently
survived the lizard’s attack without visible harm to their bodies, suggesting the
existence of an effective secondary defense, which is expected to have evolved in
concert with their warning colouration.
Alfred Russel Wallace first hypothesized that the excessive hardness of
aposematic Pachyrhynchus weevils serves as a defense mechanism against small
insectivorous animals (morphological defense hypothesis) (Wallace 1867). Despite
that the defensive function appears to be a charming and conceivable explanation for
these weevils with completely fused and rigid elytra, approximately 150 years after
Wallace’s proposal we still do not understand the ecological and adaptive significance
of the hard body in these weevils, and have no solid empirical evidences to evaluate
this hypothesis. Nevertheless, our earlier observations of only extremely large lizards
(thus higher biting force; ~ 3% of the trials) can consume the weevils pointed to the
feasibility of Wallace’s hypothesis (Tseng et al. 2014). Alternatively, other external
rigid morphologies such as sharp claws and spines of insect’s legs can often be used
as effective weapons upon being swallowed by small vertebrate predators (Eisner,
Eisner & Siegler 2005; Ruxton et al. 2004). Pachyrhynchus weevils have three pairs
of elongated legs with strong grip on the objects (Fig. 1e) (Schultze 1923; Starr &
Wang 1992). The sharp tarsal claws at the tips of these legs combined with strong grip
could potentially cause the irritating behavioral response of the lizards, which have
soft tongues and mouth cavities.
In addition to morphological (mechanical) defense, the secondary defense of
animals can often take the form of toxic or distasteful chemicals (chemical defense

4
hypothesis) (Ruxton et al. 2004). Although chemical defense is the most common
secondary defensive strategies in aposematic prey, so far no specialized secretory
glands or storage for toxic chemicals have been found in Pachyrhynchus weevils
(Pasteels, Grégoire & Rowell-Rahier 1983). Therefore, these weevils are less likely to
be chemically defended. However, a few Pachyrhynchus weevils are known to feed
on poisonous plants (e.g., sea poison tree, Barringtonia asiatica, Lecythidaceae in P.
sonani) (Chen et al. 2017), thus their bodies may potentially contain distasteful
chemicals or plant-derived toxins.
This study investigated the secondary defensive function and ecological
significance of the hard body in Pachyrhynchus weevils. First, the morphological
defense hypothesis was tested using manipulative experiments of lizard predation, by
comparing the survivorship between the ‘hard’ (mature) and ‘soft’ (teneral) weevils
(Figs 1a & b), and between the ‘claw’ (intact) and ‘clawless’ (surgically removed)
weevils (Figs 1e & f). If the weevils used the hard bodies or tarsal claws as the
secondary defenses against the lizards, the prediction was that after the encounters
with lizards, either the ‘hard’ weevils would have a higher survival rate than the ‘soft’
ones, or that the ‘claw’ weevils would survive better than the ‘clawless’ counterparts.
Next, the ecological relevance and adaptive significance of the weevil’s hard body
was assessed by measuring and comparing the hardness of the weevils and other
insect prey species found in the lizard’s natural habitat. Because the bite capacity is an
important functional trait and ecological indicator for the width of diet niche in the
lizards (Herrel et al. 2001; Meyers & Irschick 2015), the hardness (measured as force
at failure) was compared to the bite forces of the local lizard populations. If the hard

5
body of the weevils represented an important adaptive defense mechanism within the
potential prey spectrum of the lizards, the prediction was that the hardness of the
weevils would be higher than that of the other potential preys in lizard’s habitat. The
hardness of the weevils was also expected to be higher than the average bite force of
the local lizard populations. Finally, the chemical defense hypothesis was excluded by
analyzing biochemical content of the weevils for the existence of toxins or deterrents
of the vertebrates. The prediction was to find no toxic or distasteful chemicals in the
weevils.
Materials and methods
Ethics statement
The permission of using the protected Pachyrhynchus weevils was granted by the
Forestry Bureau, Council of Agriculture, Taiwan (permit no. 1031700770 &
1041700842). The Institutional Animal Care and Use Committee of the National
Taiwan Normal University approved the behavioural experiments using Japalura
lizards (No. 105012). The animal ethics protocols of the Wildlife Conservation Act of
Taiwan were followed throughout the experiments and no harmful effect to the lizards
was observed. All lizards were released to their capturing locations after the
experiments and the weevils were fed and kept in the laboratory until they died.
Weevil and lizard
Six adults (3 males and 3 females) of endemic Taiwanese Pachyrhynchus sarcitis

6
kotoensis Kôno, 1930 (Kôno 1930) (Fig. 1a) were collected by hands from Orchid
Island in August of 2015. This Pachyrhynchus species was chosen for the study
because it is relatively larger and easier to rear in the laboratory. The collected
weevils were reared individually in cylinder plastic containers (8cm diameter, 6cm
height) in the laboratory at 25°C and under 12-day/12-night cycles. Once a week, they
were supplied with fresh leaves of the known host plant, Leea guineensis (Leeaceae).
These weevils were later placed together in the containers and allowed to mate freely.
The eggs and the first instar larvae produced by these individuals were reared inside
the stems of their host plants. The plant stems were individually kept in transparent
plastic boxes (5.4cm x 4.3cm x 4.0cm) at 25°C with 12-day/12-night cycles. Once
eclosed, the teneral adults stayed in their pupal chambers for approximately five days.
The cuticles of the teneral adults remained soft (unsclerotized) for at least seven days
after they emerged from the chambers. For the following behavioral trials, the ‘soft’
weevils were defined as the individuals being within the first five days of emergence
from the pupal chambers (Fig. 1b); the ‘hard’ weevils were defined as those having
emerged for more than two months (Fig. 1a). The ‘clawless’ weevils (Fig. 1f) were
made by removing all of their tarsal claws with scissors.
The diurnal tree lizards, Japalura swinhonis (Agamidae) (Fig. 1c) are common
arboreal predators in lowland forests of Taiwan, Orchid and Green Islands. They are
sit-and-wait predators (Huang 2007) and agamid lizards use mainly visual cues for
prey detection (Cooper Jr 1989), and feed on the arthropods including weevils (Huang
2007). Therefore, J. swinhonis was selected as the testing predator for Pachyrhynchus
weevils in this study. 78 adult lizards (male: n = 60, snout-vent length (SVL) = 73.16

7
± 4.79 mm; female: n = 18, SVL = 66.57 ± 4.13 mm) were collected using a noose
between May and October of 2016 in the forests of Taipei Zoo (24°59'36.4" N,
121°34'51.9" E). The allopatrically distributed (thus likely naïve) Taipei lizard
population was used to increase the attacking rate of weevils in the behavioural trials
because the sympatric lizards attack weevils at lower frequencies (Tseng et al. 2014).
However, the male body size of Taipei’s lizards were smaller than that of Green
Island (SVL, male: 76.594 ± 4.440 mm, n = 92; female: 63.403 ± 5.094 mm, n = 80)
and Orchid Island populations (SVL, male: 75.927 ± 4.865, n = 80; female: 69.596 ±
4.061 mm, n = 89), which were used for measuring bite forces (Wang et al.
unpublished data). The captured lizards were kept individually in plastic containers
(34cm x 17cm x 24cm) at 25-30°C with water ad libitum in the laboratory for three
days before the experiments. The three-day deprivation of food prior to the
behavioural trials was used to increase the level of hunger in lizards. Each lizard was
used only once in the behavioural trials. The fifth toe of right hind legs of the lizards
was clipped and used as a marker before releasing to prevent recapture.
Colour and mobility of ‘hard’ and ‘soft’ weevils
Before conducting the behavioural trials, we measured the colouration and daily
rhythm of ‘hard’ and ‘soft’ weevils to examine the level of their resemblance in
appearance and mobility. Reflectance spectra of the colouration were measured with a
spectrometer (detection range: 250-800 nm, Jaz spectrometer, Ocean Optics, Inc.,
Dunedin, Florida) connected to a reflection probe (ZFQ-13101) illuminated by a
deuterium-tungsten halogen light source (DH-2000-BAL). The spectrometer was

8
calibrated with a reflectance standard (WS-1-SL). For each individual, we measured
the three blue spots (Fig. 1d, red circles) and three black background areas (Fig. 1d,
white circles). The light source was positioned 1mm perpendicularly above the focal
area of the weevil body surface (diameter of the illuminated area: ~ 2mm). The three
measurements of the blue spots and black areas from each individual were averaged.
The hue (wavelength of the apex of the spectrum), brightness (reflectance ratio of the
apex of the spectrum), and saturation (area under the spectral curve between the
visible wavelengths of diurnal lizards, 440 - 625nm; Yewers et al. 2015) of the
measured spectrum were calculated. The ultraviolet-sensitive (UVS) (364 - 383 nm)
spectrum was excluded from the calculation because of the low UV reflectance (<
40%) of the colouration (Fig. 2a). No significant difference between the colour of
‘hard’ and ‘soft’ weevils was detected (Fig. 2a) (two-sample t test, n = 4; hue, ‘hard’:
555.36 ± 19.76 nm, ‘soft’: 547.15 ± 8.73 nm, t = 0.761, p = 0.476; brightness, ‘hard’:
128.97 ± 4.79%, ‘soft’: 125.00 ± 3.99%, t = 1.274, p = 0.250; saturation, ‘hard’: 66.40
± 2.99%, ‘soft’: 67.23 ± 1.90%, t = -0.454, p = 0.666).
The daily rhythm of the weevils (n: ‘hard’ = 10, ‘soft’ = 10) were recorded at
25°C for six consecutive days using a video monitor (DS-VR7160H, Der Shuenn,
Taipei, Taiwan). The weevils were fed once on the third day. Each weevil was placed
in a petri dish (diameter: 90mm, height: 15mm) divided into four equal quadrants. We
recorded the number of weevil’s movements between the quadrants in the first 10
minutes of every two hours. The total numbers of movements for each individual
during the six days were summed up as a proxy of its mobility. The daily rhythm of
the two weevil groups was similar (Fig. 2b) (number of movements per day, ‘hard’:

9
42.5 ± 15.9, ‘soft’: 44.6 ± 15.9; two-sample t test of daytime scores, n = 10, t = -0.376,
p = 0.7113). These results suggested that the ‘hard’ and ‘soft’ weevils represent prey
items with equal appearance and mobility for the visually oriented lizard predator.
Behavioral trials
To test the effectiveness of weevil’s hard exoskeleton and tarsal claws against
predation by lizards, the weevils were randomly divided into four treatments (n = 78):
(1) ‘hard’ weevils with claws, (2) ‘hard’ weevils without claws, (3) ‘soft’ weevils
with claws, (4) ‘soft’ weevils without claws. The trial arena was a transparent plastic
container (58cm x 30cm x 40cm) with the bottom padded with soil (Fig. 3a). The
three vertical planes of the container were covered with light brown cardboard paper
while one side was left transparent for recording by a camcorder (HDR-XR200,
SONY, Tokyo, Japan). The operator and the camcorder were located behind a
cardboard (75cm x 100cm) with an opening (diameters, 10cm) to minimizing the
disturbance. The trials were carried out between 10 am and 6 pm and at 26-30°C
when the lizards were most active. The lizards were placed into the arena three
minutes prior to the trials for accommodation. The lizards appeared to settle down
within one minute of introducing by not running around or climbing out of the
container. The weevils were tied with a black cotton thread, and the opposite side of
the thread was tied to the center of a transparent plastic bar (58cm x 3cm x 0.3cm).
The length of the thread was ~ 43cm to confine the weevil’s movement to the center
of the arena for video recording. Three minutes after placing the lizard, the weevil
was introduced into the arena by hanging the plastic bar above the arena. The weevil

10
was placed at the center of the arena (about 30cm diameter) and approximately
15-30cm away from the lizard that often positioned itself at the periphery. The trial
started when the lizard was aware of the weevil by turning its eyes or head toward the
prey and ended after the lizard have stopped response to the prey. The survival rate,
number of bites, and prey handling time (when the lizard had the first physical contact
with the weevil until the lizard stopped responding to it) were calculated from the
videos.
Insect prey and hardness measurement
To assess the level and distribution of hardness in the lizard’s prey community,
the insects occurring within the habitats of lizards were sampled using hands,
sweeping nets, light traps, and pitfalls in Orchid and Green Islands in September and
October of 2015. The insects were collected along the forest trails where most lizards
could be found (eight sites in Orchid Island for five days and four sites in Green
Island for four days). The light traps (high-intensity discharge lamps, 35W, 6000K,
Cnight, Guangdong, China) were set up in Orchid Island for two hours/nights. The
lepidopterans and spiders were not sampled because they are considered to be ‘soft’
preys (Verwaijen, Van Damme & Herrel 2002). One to two individuals of each insect
species were obtained and later stored at a -20°C freezer in the laboratory until the
measurement. All voucher insect specimens of this study were archived in the insect
collection of the laboratory (Table S1).
The hardness of the insects (field-collected insect prey and the ‘hard’ and ‘soft’
weevils) was measured following Herrel et al. 2001. The ‘force at failure’ of the elytra

11
(abdomen) or the thorax of the insects was measured to represent the ‘hardness’ of the
subjects. The ‘failure’ was defined as the completely loss of load-carrying capacity of
the structure (Collins 1993). The definition of ‘hardness’ in biological studies are
debatable and this term often refers to the phenomena of combining effects of
material and structural properties (e.g., for alternative ‘intractability’ in Evans &
Sanson 2005). Here we use ‘hardness’ to indicate the maximum force at failure for
crushing an insect’s exoskeleton. The upper and lower jaws of the lizard were
dissected, cleaned and then embedded in resin (model 40200029, Struers, Ballerup,
Denmark; diameter: 25mm) with the teeth rows exposed (H in Fig. 3b) to build the
biting platform (F & G in Fig. 3b). The upper part of the platform (F in Fig. 3b) was
connected to a force transducer (model 9203, Kistler Inc., Wintherthur, Swizerland)
(D in Fig. 3b), which was attached to the lower end of a camera mount on a copy
stand (E in Fig. 3b) and connected to the charge amplifier (model 5995). The lower
part of the platform was placed below the upper part at a position where the two jaws
can match completely. These insect specimens was measured at 25°C after they were
fully thawed in the laboratory. The specimen was positioned perpendicularly with
respect to the tooth rows. The upper platform was slowly moved down until the
exoskeleton of the insect was broken or crashed, and the amount of forces exerted on
the transducer was recorded simultaneously. The breakdown of the exoskeleton of the
specimen was recognized by a structural failure on the exoskeleton. The maximum
force needed to break the specimen’s exoskeleton was recorded as a proxy for the
hardness of the insect. These measurements were compared with the bite forces of the
J. swinhonis populations from Orchid and Green Islands (Wang et al. unpublished

12
data).
Chemical analysis
The existence of potential deterrent chemicals within the body of P. sarcitis
kotoensis was examined using the Gas Chromatograph-Mass Spectrometer (GC-MS).
The thorax and abdomen of three reared P. sarcitis kotoensis adults were separated
and immersed in 3ml of ether for 10 minutes of chemical extraction. 0.5µl of the
extraction was used for the GC-MS analysis. The GC-MS system consisted a gas
chromatograph (model: TRACE™ 1300), a mass selective detector (model: ISQ™,
Thermo Fisher Scientific Inc., Waltham, Massachusetts, USA) and the GC column
(inside diameter: 0.25mm, length: 30m, film thickness: 0.25um) (model: Rxi-5MS,
13423, Restek Corporation, Bellefonte, Pennsylvania, USA). The inlet temperature
was 280°C and helium was the carrier gas with the constant flow at 1 ml/min. The
initial temperature was 50°C and rose with a rate of 10°C/min to 300°C. This
temperature was held for 10 minutes without delay of the solvent. The peaks with
higher relative abundance were identified and compared with the spectrum libraries
(NIST, Gaithersburg, Maryland, USA, and WILEY 10R, Hoboken, New Jersey,
USA). If the SI (similarity index) and RSI (reverse similarity index) value of the
candidate were both higher than 800, it was considered as a possible chemical
compound extracted from the weevils (Tian et al. 2014). Then this compound was
identified and determined if it represents a potential repellent for vertebrate predators
in the online Pherobase (El-Sayed 2014).

13
Statistical analyses
The two-sample t tests were used to compare the reflectance spectra and daily
rhythm (daytime) between ‘hard’ and ‘soft’ weevils, and Mann-Whitney U test was
used to compare the daily rhythm of day and nighttime. Fisher’s exact test, negative
binomial regression, and survival analysis were employed to compare the survival rate,
number of bites, and prey handling duration in behavioral trials, respectively. In
negative binomial regression and survival analysis, we used the elytra length of the
weevils and the snout-vent length of the lizards as covariates. These statistics were
followed by Tukey post hoc tests to examine the differences between treatment
groups. Welch two sample t-test was used to compare the hardness between ‘hard’
and ‘soft’ weevils, and linear regression analyses were conducted to assess the
relationship between prey hardness and body size. All statistical analyses were carried
out in R 3.3.1 (R Development Core Team 2016).
Results
Effects of the hardness and claws
Immediately after introducing the weevils into the trial arena, the lizards
perceived the presence of the prey by turning their eyes and heads toward them
(movie 1 & 2). Then the lizards approached and bit the weevils (often the abdomen)
using their lateral teeth rows. The weevils were attacked by the lizards within an
average of 18.8 seconds (18.8 ± 22.9 sec, n = 59) after the introduction of the prey.
Approximately one half of the lizards (51%, n = 76) showed no predatory response

14
toward the weevils after more than 3 minutes, suggesting that they either were not
interested, neophobia, or avoided attacking aposematic preys. After the lizard’s first
attack, the weevils were either spat out (movie 1, 00:50) or consumed (movie 2, 00:40)
by the lizards. All ‘hard’ weevils were spat out immediately by the lizards and
survived the first attack. All rejected weevils have no visible physical damage to the
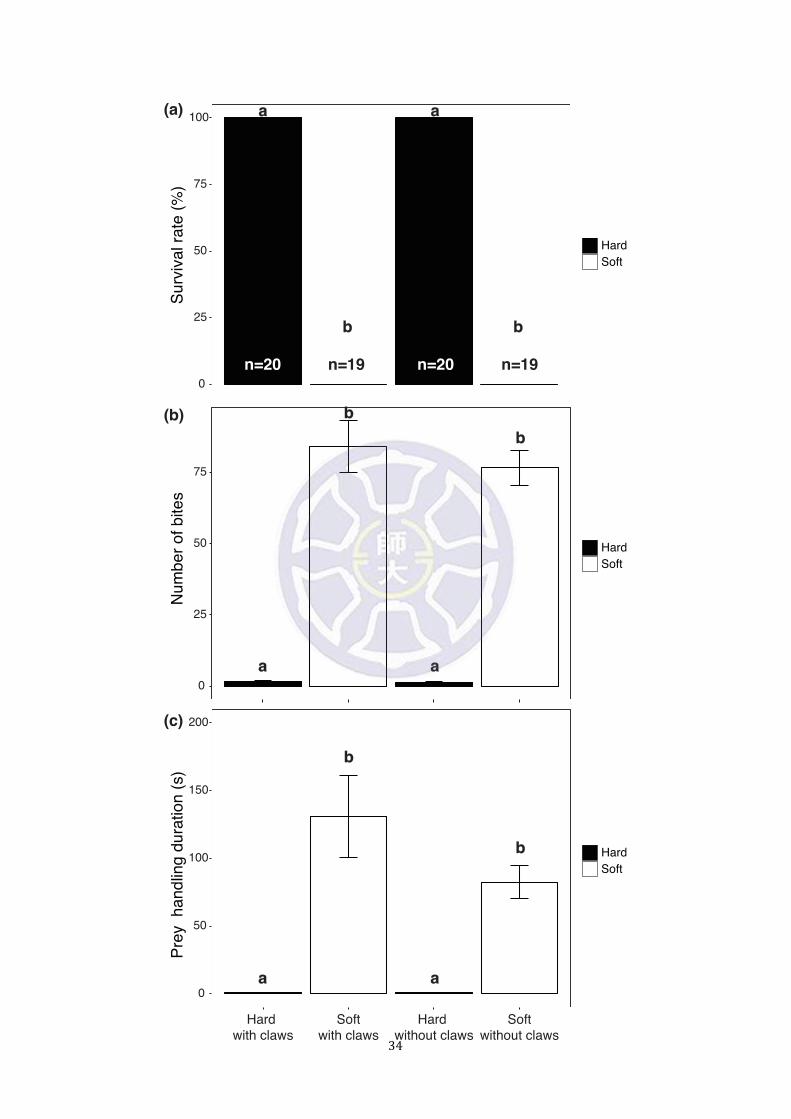
body and lived for more than two months after the trials (survival rate = 100%, n = 40)
(Fig. 4a), except that two individuals were injured and suffered a small depression on
the surface of lateral abdomen. The lizards made no more predatory attempt after
spitting out the weevils (movie 1). By contrast, all ‘soft’ weevils were chewed
continuously and subsequently swallowed by the lizards (movie 2) (Fig. 4a) (survival
rate = 0%; Fisher’s exact test: n = 38; ‘hard’ with claws vs. ‘soft’ with claws: p <
0.001; ‘hard’ without claws vs. ‘soft’ with claws: p < 0.001; ‘hard’ with claws vs.
‘soft’ without claws: p < 0.001; ‘hard’ without claws vs. ‘soft’ without claws: p <
0.001). The results were the same regardless of tarsal claws (Fig. 4a) (‘hard’ with
claws vs. ‘hard’ without claws: p = 1; ‘soft’ with claws vs. ‘soft’ without claws: p =
1).
The lizards bit ‘soft’ weevils significantly more times than ‘hard’ weevils (Fig.
4b) (mean number of bites = 1.5 ± 1.0 ‘hard’, 80.3 ± 34.0 ‘soft’; negative binomial
regression: χ2 = 227.8264, p < 0.001) (‘hard’ with claws vs. ‘soft’ with claws: Z =
18.991, p < 0.001; ‘hard’ without claws vs. ‘soft’ with claws: Z = -19.014, p < 0.001;
‘hard’ with claws vs. ‘soft’ without claws: Z = 19.146, p < 0.001; ‘hard’ without
claws vs. ‘soft’ without claws: Z = 19.044, p < 0.001), with no effect of tarsal claws
(‘hard’ with claws vs. ‘hard’ without claws: Z = -0.476, p = 1; ‘soft’ with claws vs.

15
‘soft’ without claws: Z = -0.060, p = 1). The lizards spent significantly more time
handling prey in ‘soft’ than in ‘hard’ weevils (Fig. 4c) (mean handling time = 0.7 ±
0.5 sec ‘hard’, 106.5 ± 102.4 sec ‘soft’; survival analysis: n = 78, χ2 = 202.98, p <
0.001) (‘hard’ with claws vs. ‘soft’ with claws: Z = 22.992, p < 0.001; ‘hard’ without
claws vs. ‘soft’ with claws: Z = -22.937, p < 0.001; ‘hard’ with claws vs. ‘soft’
without claws: Z = 21.637, p < 0.001; ‘hard’ without claws vs. ‘soft’ without claws: Z
= 22.049, p < 0.001). Again, there was no significant difference between treatments
with and without tarsal claws (Fig. 4c) (‘hard’ with claws vs. ‘hard’ without claws: Z
= -0.109, p = 0.914; ‘soft’ with claws vs. ‘soft’ without claws: Z = -1.954, p = 0.102).
Hardness of prey and lizard’s bite force
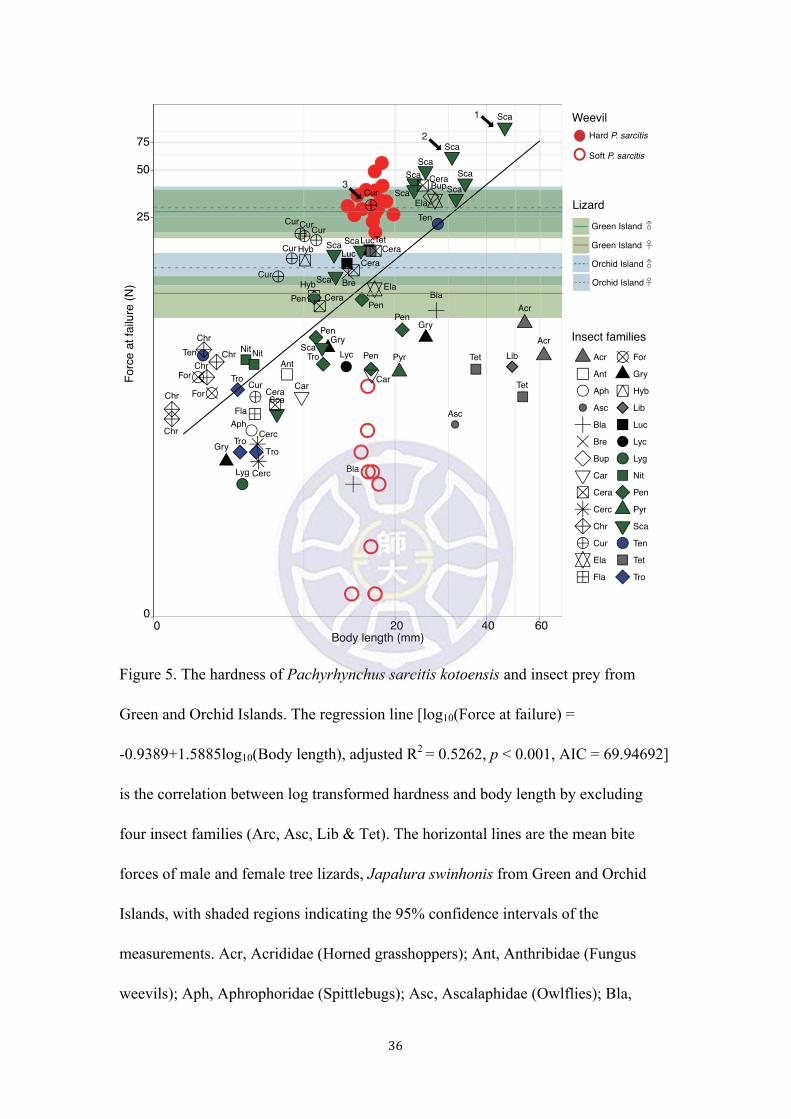
The prey hardness increased allometrically with size for most prey species
measured in the habitats (Fig. 5) [log10(Force at failure) = -0.9389 + 1.5885
log10(Body length), adjusted R2 = 0.5262, p < 0.001, AIC = 69.94692; Force at failure
= -9.2195 + 1.5678 Body length, adjusted R2 = 0.6494, p < 0.001, AIC = 503.2208],
with the exception that grasshoppers (Acr, Acrididae), owlflies (Asc, Ascalaphidae),
dragonflies (Lib, Libellulidae) and katydids (Tet, Tettigoniidae) were larger but softer
insect prey. The ‘hard’ and ‘soft’ weevils deviated from the regression of hardness
versus body length by being harder and softer than the prey of similar sizes,
respectively (Fig. 5). The ‘hard’ P. sarcitis kotoensis (32.6 ± 9.1 N, n = 17) were
significantly harder than their ‘soft’ counterparts (0.6 ± 0.6 N, n = 10) (Welch two
sample t-test, t = 14.46, p < 0.001) and the average hardness of insect prey in the same
habitat (Fig. 5) (9.95 ± 11.98 N, n = 72), except for the two rhinoceros beetles,

16
Xylotrupes philippinensis, 93.5 N and 60.8 N, black arrow 1 & 2, Fig. 5). The ‘hard’
P. sarcitis kotoensis were also harder than most other weevil species (Cur) in the
habitat, except for Aclees hirayamai (black arrow 3, Fig. 5). The average hardness of
‘hard’ P. sarcitis kotoensis was higher than the mean bite force of J. swinhonis from
Green Island (males: 27.97 ± 9.55 N, n = 91; females: 8.14 ± 2.42 N, n = 80) and
Orchid Island (males: 29.66 ± 9.62 N, n = 80; females: 12.03 ± 2.80 N, n = 89) (Fig.
5), indicating that most lizards from these two islands lack sufficient bite force
capable of penetrating the sclerotized exoskeletons of P. sarcitis kotoensis, especially
for females and smaller males. As what we would expect from insects mimicking
Pachyrhynchus, the hardness of a longhorn beetle, Doliops similis from Orchid Island
was merely 11.5 N (Table S1), which was much lower than that of ‘hard’ P. sarcitis
kotoensis.
Biochemical compounds of P. sarcitis kotoensis
We selected twenty peaks with the highest relative abundance from the master
chromatogram of the extraction of P. sarcitis kotoensis to compare with the spectrum
library (Fig. 6). Five of them, including the solvent (ethane; RT: 1.72, ST: 935, RST:
936, CAS number: 60-29-7), had both SI and RSI values higher than 800. The four
candidate compounds in P. sarcitis kotoensis were identified as ethyl propyl ether,
diethylene glycol, butylated hydroxy toluene and cadalene. These chemicals can be
found in other animals or plants but have no toxic or repellent functions, and do not
fall into the 13 categories of known insect toxins and repellents against vertebrate
predators (Blum 2012).

17
Discussion
Effective secondary defense
The study provides the first empirical evidences to support the morphological
defense hypothesis pioneered by A. R. Wallace almost 150 years ago. Our findings
clearly indicate that the hardness of aposematic Pachyrhynchus functions as a
secondary defense against the lizards, whereas the tarsal claw serves no defense, and
no toxin or repellent is stored in the weevil. This secondary defense of hardness is
highly effective in at least three intriguing features. First, the hardness of exoskeleton
significantly increased the survival of the weevils under lizard’s attack. In fact, the
survival rate was 100% for the ‘hard’ weevils. Almost all rejected weevils by the
lizard were undamaged and resumed walking immediately after the attack. By
contrast, the ‘soft’ weevils were all consumed by the lizards during the staged
encounters. These results suggest that the hardness of exoskeletion is an important
fitness component of morphological adaptation for aposematism in Pachyrhynchus.
Given that physical attacks by the predators are often costly to the prey in terms of
injury, escaping time or survival (Ruxton et al. 2004), it was very surprising to
observe that Pachyrhynchus with such highly effective defense as hardness that they
practically suffered no cost in terms of injury and survival. The high survival of
Pachyrhynchus under lizard’s attack can provide an empirical evidence for
evolutionary origin of aposematic colours. Because one of the conundrum of evolving
aposematic colours is that a novel conspicuous individual will be detected easily in

18
the population, and its rarity will make it challenging for predators to learn to avoid
the colour signal in future encounter (Fisher 1958). However, at the beginning of
evolving warning signals, if a rare novel conspicuous prey such as the case in
Pachyrhynchus can survive the initial attack, then the survived aposematic individual
will make it easier for predators to recognize the colour signal, thus the warming
colour can increase its frequency in the prey population (Ruxton et al. 2004).
Secondly, the hardness of weevil’s exoskeleton immediately triggered strong
aversive responses of the lizards after biting the ‘hard’ weevils (such as instantly
shaking heads and spitting out the prey). Similar to some of colourful Eupholus and
Trigonopterus species (Seago et al. 2009; Riedel et al. 2013), the Pachyrhynchus are
unique among those highly sclerotized flightless weevils to possess the most diverse
aposematic colours (Wallace 1895; Schultze 1923). Our findings indicate that this
anti-predation strategy is very effective in strengthening the aposematism in colourful
Pachyrhynchus weevils, by developing a robust association between warning displays
and prey unprofitability. Our earlier study found that after only a single encounter the
naïve lizard predators rapidly learned to avoid weevils for at least more than 23 days
(Tseng et al. 2014). The lizards can instantly learn from the weevil’s unprofitability
and reduce future attacks when they encounter prey items with similar warning
signals. Therefore, the inclusive fitness of aposematic weevils that confer such
defense may subsequently be raised.
Finally, the secondary defense using the hardness is a ‘secret weapon’ (sensu
lato Eisner, Eisner & Siegler 2005) in the sense that, similar to internally stored
chemical defenses in toxic prey, it is invisible and unexpected by the naïve predators.

19
Aposematic prey species that use internally stored toxic chemicals as the source of
defenses are well known in the nature. However, other invisible defense strategies
such as heavy armature of a harvestman (da Silva Souza & Willemart 2011) and hard
body used by aposematic weevils in this study are poorly studied (Caro 2017). Our
results suggest that, in additional to surviving the attack, the ‘secret weapon’ of
weevils simultanously strengthens lizard’s ‘unexpectancy’ via their outstanding
hardness compared to most insect prey with similar and even larger sizes in the
habitat. This high level of hardness largely explain the instant and strong aversive
responses displayed by the lizards after biting weevils with their teeth.
Multiple secondary defenses
This study showed that the hardness of Pachyrhynchus weevils is the main
secondary defenses against lizard predators. However, like other flightless weevils
such as Trigonopterus (Van De Kamp et al. 2014), Pachyrhynchus employs a series
of secondary defensive strategies when encounters human predators, by firstly turning
itself around the tree branches or leaves to hide, then performing a sudden drop-off
from the trees, and finally a death-feigning on the ground (Wang et al. personal
observations). This series of secondary defensive strategies may be essential in
evading predators at initial stage of prey recognition and pursuing, especially when
weevils are soft (i.e., newly emerged without a ‘hard’ defense) or encountering
predators with greater bite forces. Nevertheless, it is still unknown whether ‘soft’
weevils could perform drop-off and death-feigning equally well as that of ‘hard’
individuals.

20
Mechanical mechanism of hardness
The mechanical mechanism of achieving the outstanding hardness for a small
sized insect such as Pachyrhynchus remains elusive. The mechanical properties of
insect’s cuticle like strength and stiffness depend on many physical and chemical
factors, including arrangement of cuticle microfibers, protein and water content, and
process of pigmentation and sclerotization (Andersen 2010; Klocke & Schimitz 2011).
A recent study indicated that an endosymbiont Nardonella specialized in cuticle
formation and hardening of Pachyrhynchus host by tyrosine provisioning, suggesting
the important role of the bacterial symbiosis for the weevil’s hardness (Anbutsu et al.
2017). The microstructure of elytral cuticle in weevils is phylogenetically conserved
to have a ‘weevil-specific’ combination of interlocking of exo- and endocuticle, an
endocuticle with distinct ovoid macrofibers embedded in a matrix, and small angles
between successive endocuticular layers (Van de Kamp, Riedel & Greven 2016).
These microstructural attributes or their modifications may have contributed to the
hardness of Pachyrhynchus. However, the cuticular microstructures in other hard
flightless weevils such as Trigonopterus do not deviate from the weevil’s uniform
ground plan but instead possess thicker elytra (Van de Kamp et al. 2016). Besides
cuticular microstructures and thickness, sclerotization (Hopkins & Kramer 1992),
heavy metals and halogens (Schofield, Nesson & Richardson 2002) can enhance
mechanic strength of insect cuticles. The mechanical behaviour of an insect body is
the result of combining material properties and geometric characteristics. Thus the
geometry of an organism can modify the stress distribution and reduce the stress

21
concentration to prevent mechanical failure (e.g., coiling mollusk shells in Rajabi et al.
2014). Because the Pachyrhynchus has a peculiar dome-shaped thorax, abdomen and
densely interlocked elytra, these oval architectures may build these weevils more
robust bodies as a defense strategy against predators.
Adaptive defense against lizard’s predation
This study provided convincing evidences to indicate that the hardness of
aposematic Pachyrhynchus weevils is ecologically important and represents an
adaptive defense mechanism against the predation of tree lizards. The results
suggested that the hardness of the weevils was among the highest in the prey spectrum
of the tree lizards and higher than the average bite force of the lizards from Green and
Orchid Islands. Therefore, the predation of small vertebrate predators like tree lizards
may have acted as primary driving/maintaining forces for the evolution of hardness in
Pachyrhynchus weevils. At present, Japalura swinhonis is the only known natural
predator of Pachyrhynchus weevils in these two islands (Huang 2007). The other
eight lizard species of the islands are less likely to be major predators of the weevils
due to their habitat and prey preference (Chen et al. 2008; Li et al. 2010) (Table S2).
Pachyrhynchus’s hard body likely has been originated in the common ancestors of
relates like Eupyrgops, Metapocyrtus and Polycatus in the Philippine Archipelago and
neighboring islands (Schultze 1923), therefore the arboreal lizards of Southeast Asia
such as Gekko spp. might also be important in promoting the origin of weevil’s hard
exoskeletons.
Multiple selective forces of diverse predator community may each play an

22
important role in driving and maintaining the evolution of aposematic colours and
associated defense strategies (Willink et al. 2014). In addition to lizard’s (reptilian)
predation, avian and mammalian predations constitute the other two major selective
forces for tropical insects with aposematic or cryptic colouration (Mappes et al. 2014;
Roslin et al. 2017). Avian predation is likely to be essential for the evolution of
aposematism and defense strategies in Pachyrhynchus weevils because birds in
general possess a wide range of colour spectrum (Bennett & Théry 2007), and they
are primary predators for aposematic insects (Iniesta, Ratton & Guerra 2017). In
Green and Orchid Islands, there are about 62 bird species that eat insects as parts of
their diet (Chen et al. 2008; Li et al. 2010; Severinghaus et al. 2012) (Table S3).
Among them, 12 species [emerald dove (Chalcophaps indica), large hawk-cuckoo
(Cuculus sparverioides), Himalayan cuckoo (Cuculus saturatus), lesser coucal
(Centropus bengalensis), common kingfisher (Alcedo atthis), brown shrike (Lanius
cristatus), Japanese paradise-flycatcher (Terpsiphone atrocaudata), brown-eared
bulbul (Microscelis amaurotis), lowland white-eye (Zosterops meyeni),
white-shouldered starling (Sturnus sinensis), eyebrowed thrush (Turdus obscurus),
and blue rock thrush (Monticola solitarius)] inhibit the same habitat and are probable
avian predators of Pachyrhynchus (Table S3). At present no bite forces of avian fauna
from Green and Orchid islands is available for comparison. However, the bite forces
of the vertebrates are often strongly correlated with head sizes and shapes (Anderson,
McBrayer & Herrel 2008). The average bite force of small to median size passerines
ranges was 9.3 ± 10.5 N (2.9-38.4 N, n = 18, Estrildidae & Fringillidae, Van der Meij
& Bout 2006), suggesting that the hardness of Pachyrhynchus weevils (32.6 ± 9.1 N)

23
may be effective against predation from most passerine birds of similar sizes.
Nevertheless, the shape beaks and pecking behavior of most birds alone may be
proven effective to penetrate Pachyrhynchus’s exoskeleton. Our field observations
suggested that Pachyrhynchus weevils were defenseless and could be easily pecked
and swallowed by adult domestic chickens. However, smaller domestic chicks were
observed to spend considerable amount of time pecking the weevils, but could not
successfully swallow them. These preliminary observations and the bite force
measurement of birds together suggest that the hardness of Pachyrhynchus weevils
may only be effective against smaller avian predators in their habitats.
There are 14 mammal species found in Green and Orchid Islands that may
consume insects (Chen et al. 2008; Li et al. 2010), in which 10 species [masked palm
civet (Paguma larvata), Chinese white-toothed shrew (Crocidura rapax tadae & C. r.
lutaoensis), Pallas's squirrel (Callosciurus erythraeus thaiwanensis), house mouse
(Mus musculus), lesser rice field rat (R. losea), Tanezumi rat (R. tanezumi), greater
bandicoot rat (Bandicota indica), brown rat (R. norvegicus) and black rat (R. rattus)]
occur in Pachyrhynchus’s habitats. Tanezumi rats were observed to forage on tree
trunks of weevil’s habitat during the day and nighttime, and are likely to be one of
their mammal predators, with capable bite force of penetrating their exoskeleton
(24.7-76.8 N, brown rat, R. norvegicus, Cox et al. 2012). Future studies on identifying
the major predators and assessing spatio-temporal variation of predation in
Pachyrhynchus weevils will help us to better understand the ecology and evolution of
the hardness in aposematic prey.

24
References
Abrams, P.A. (2000) The evolution of predator-prey interactions: theory and evidence.
Annual Review of Ecology and Systematics, 31, 79-105.
Anbutsu, H., Moriyama, M., Nikoh, N., Hosokawa, T., Futahashi, R., Tanahashi, M.,
Meng, X.Y., Kuriwada, T., Mori., N., Oshima., K., Hattori, M., Fujie, M., Satoh,
N., Maeda, T., Shigenobu, S., Koga, R. & Fukatsu, T. (2017) Small genome
symbiont underlies cuticle hardness in beetles. Proceedings of the National
Academy of Sciences of the United States of America, 201712857.
Andersen, S.O. (2010) Insect cuticular sclerotization: a review. Insect Biochemistry
and Molecular Biology, 40, 166-178.
Anderson, R.A., McBrayer, L.D., & Herrel, A. (2008) Bite force in vertebrates:
opportunities and caveats for use of a nonpareil whole‐animal performance
measure. Biological Journal of the Linnean Society, 93, 709-720.
Barševskis, A. (2013) Contribution to the knowledge of the genus Doliops
Waterhouse, 1841 (Coleoptera: Cerambycidae). Baltic Journal of Coleopterology,
13, 73-89.
Bennett, A.T. & Théry, M. (2007) Avian color vision and coloration:
multidisciplinary evolutionary biology. The American Naturalist, 169, S1-S6.
Blount, J.D., Rowland, H.M., Drijfhout, F.P., Endler, J.A., Inger, R., Sloggett, J.J.,
Hurst, G.D.D., Hodgson, D.J. & Speed, M.P. (2012) How the ladybird got its
spots: Effects of resource limitation on the honesty of aposematic signals.
Functional Ecology, 26, 334-342.

25
Blum, M. (2012) Chemical Defenses of Arthropods. Elsevier.
Caro, T. (2017) Wallace on coloration: contemporary perspective and unresolved
insights. Trends in Ecology & Evolution, 32, 23-30.
Chen, P.C., Chang, M.H., Chen, S.F., Li, Z.L., Chen, S.L., Lin, H.C. & Chu, C.W.
(2008) The Research of Terrestrial Vertebrate Fauna on the Green Island.
Marine National Park Headquarters.
Chen, Y.T., Tseng, H.Y., Jeng, M.L., Su, Y.C., Huang, W.S. & Lin, C.P. (2017)
Integrated species delimitation and conservation implications of an endangered
weevil Pachyrhynchus sonani (Coleoptera: Curculionidae) in Green and Orchid
Islands of Taiwan. Systematic Entomology, 42, 796-813.
Collins, J.A. (1993) Failure of materials in mechanical design: analysis, prediction,
prevention. John Wiley & Sons.
Cooper Jr, W.E. (1989) Absence of prey odor discrimination by iguanid and agamid
lizards in applicator tests. Copeia, 472-478.
Cox, P.G., Rayfield, E.J., Fagan, M.J., Herrel, A., Pataky, T.C. & Jeffery, N. (2012)
Functional evolution of the feeding system in rodents. PLoS ONE, 7, e36299.
Dickerson, R.E., Merrill, E.D., McGregor, R.C., Schultze, W., Taylor, E.H. & Herre,
A.W. (1928) Distribution of Life in the Philippines. Bureau of Printing.
Edmunds, M. (1974) Defence in Animals: A Survey of Anti-Predator Defences.
Longman Publishing Group.
Eisner, T., Eisner, M., & Siegler, M. (2005) Secret weapons: defenses of insects,
spiders, scorpions, and other many-legged creatures. Harvard University Press.
El-Sayed, A.M. (2014) The Pherobase: Database of pheromones and semiochemicals.

26
The Pherobase.
Evans, A.R., & Sanson, G.D. (2005) Biomechanical properties of insects in relation to
insectivory: cuticle thickness as an indicator of insect ‘hardness’ and
‘intractability’. Australian Journal of Zoology, 53, 9-19.
Fisher, R.A. (1958) The genetic theory of natural selection. Dover.
Herrel, A., Damme, R.V., Vanhooydonck, B., & Vree, F.D. (2001) The implications
of bite performance for diet in two species of lacertid lizards. Canadian Journal
of Zoology, 79, 662-670.
Hopkins, T.L. & Kramer, K.J. (1992) Insect cuticle sclerotization. Annual Review of
Entomology, 37, 273-302.
Huang, W.S. (2007) Ecology and reproductive patterns of the agamid lizard Japalura
swinhonis on an east Asian island, with comments on the small clutch sizes of
island lizards. Zoological Science, 24, 181-188.
Iniesta, L.F.M., Ratton, P. & Guerra, T.J. (2017) Avian predators avoid attacking
artificial aposematic millipedes in Brazilian Atlantic Forest. Journal of Tropical
Ecology, 33, 89-93.
Klocke, D., & Schmitz, H. (2011) Water as a major modulator of the mechanical
properties of insect cuticle. Acta Biomaterialia, 7, 2935-2942.
Li, Z.D., Su, Y., Li, Z.M., Guo, S.J., Wu, T.Y., Lin, Y.J., Yang, D.Q., Liang, Y.Z., Yu,
H.Y., Qiu, B.H., Hsu, S.T., Lin, Y.Z., Liu, Z.H., Weng, R.H., Ye, R.K., Ye, C.B.,
Wu, Y.X., Tseng, Y.H., Pan, T.Z., Tsai, Z.H. Su, X.Q., Chen, Y.J. & Tsai, J.H.
(2010) The Inventory of Natural Systems in Orchid Island. Marine National Park
Headquarters.

27
Maan, M.E. & Cummings, M.E. (2011) Poison frog colors are honest signals of
toxicity, particularly for bird predators. The American Naturalist, 179, E1-E14.
Mappes, J., Kokko, H., Ojala, K., & Lindström, L. (2014) Seasonal changes in
predator community switch the direction of selection for prey defences. Nature
Communications, 5, 5016.
Meyers, J.J. & Irschick, D.J. (2015) Does whole-organism performance constrain
resource use? A community test with desert lizards. Biological Journal of the
Linnean Society, 115, 859-868.
Pasteels, J.M., Grégoire, J.C. & Rowell-Rahier, M. (1983) The chemical ecology of
defense in arthropods. Annual Review of Entomology, 28, 263-289.
R Development Core Team. (2016) R: A Language and Environment for Statistical
Computing. Vienna, Austria. ISBN 3-900051-07-0. URL
http://www.R-project.org.
Rajabi, H., Darvizeh, A., Shafiei, A., Eshghi, S. & Khaheshi, A. (2014) Experimental
and numerical investigations of Otala lactea’s shell–I. Quasi-static analysis.
Journal of the Mechanical Behavior of Biomedical Materials, 32, 8-16.
Riedel, A., Sagata, K., Surbakti, S., Tänzler, R., & Balke, M. (2013) One hundred and
one new species of Trigonopterus weevils from New Guinea. ZooKeys, 280, 1.
Robinson, M.H. (1969) Defenses against visually hunting predators.Evolutionary
Biology, 3, 5-59.
Roslin, T., Hardwick, B., Novotny, V., Petry, W.K., Andrew, N.R., Asmus, A., ... &
Cameron, E.K. (2017) Higher predation risk for insect prey at low latitudes and
elevations. Science, 356, 742-744.

28
Ruxton, G.D., Sherratt, T.N. & Speed, M.P. (2004) Avoiding Attack: The Evolutionary
Ecology of Crypsis, Warning Signals and Mimicry. Oxford University Press.
Schofield, R.M.S., Nesson, M.H. & Richardson, K.A. (2002) Tooth hardness
increases with zinc-content in mandibles of young adult leaf-cutter ants.
Naturwissenschaften, 89, 579-583.
Schultze, W. (1923) A monograph of the pachyrrhynchid group of the Brachyderinae,
Curculionidae: Part I. The genus Pachyrhynchus Germar. The Philippine Journal
of Science, 23, 609-673.
Seago, A.E., Brady, P., Vigneron, J.P., & Schultz, T.D. (2009) Gold bugs and beyond:
a review of iridescence and structural colour mechanisms in beetles (Coleoptera).
Journal of the Royal Society Interface, 6, S165-S184.
Severinghaus, L.L., Ding, T.S., Fang, W.H., Lin, W.H., Tsai, M.C., & Yen, C.W.
(2012) The avifauna of Taiwan 2nd edition. Taipei, Taiwan: Forestry Bureau of
Council of Agriculture of Executive Yuan.
Sherratt, T.N. & Beatty, C.D. (2003) The evolution of warning signals as reliable
indicators of prey defense. The American Naturalist, 162, 377-389.
da Silva Souza, E. & Willemart, R.H. (2011) Harvest-ironman: heavy armature, and
not its defensive secretions, protects a harvestman against a spider. Animal
Behaviour, 81, 127-133.
Starr, C.K. & Wang, H.Y. (1992) Pachyrhynchine weevils (Coleoptera: Curculionidae)
of the islands fringing Taiwan. Journal of Taiwan Museum, 45, 5-14.
Stevens, M. (2013) Sensory Ecology, Behaviour, and Evolution. Oxford University
Press.

29
Summers, K. & Clough, M.E. (2001) The evolution of coloration and toxicity in the
poison frog family (Dendrobatidae). Proceedings of the National Academy of
Sciences of the USA, 98, 6227-6232.
Tian, H., Zhan, P., Deng, Z., Yan, H. & Zhu, X. (2014) Development of a flavour
fingerprint by GC‐MS and GC‐O combined with chemometric methods for the
quality control of Korla pear (Pyrus serotina Reld). International Journal of
Food Science & Technology, 49, 2546-2552.
Tseng, H.Y., Lin, C.P., Hsu, J.Y., Pike, D.A. & Huang, W.S. (2014) The functional
significance of aposematic signals: geographic variation in the responses of
widespread lizard predators to colourful invertebrate prey. PLoS ONE, 9,
e91777.
Tseng, H.Y., W.S. Huang, M.L. Jeng, R.J.T. Villanueva, O.M. Nuñeza & Lin, C.P.
(2017) Complex inter-island colonization and peripatric founder speciation
promote diversification of flightless Pachyrhynchus weevils in the
Taiwan-Luzon volcanic belt. Journal of Biogeography, in press.
Van De Kamp, T., dos Santos Rolo, T., Vagovič, P., Baumbach, T., & Riedel, A.
(2014) Three-dimensional reconstructions come to life-interactive 3D PDF
animations in functional morphology. PLoS ONE, 9, e102355.
Van de Kamp, T., Riedel, A. & Greven, H. (2016) Micromorphology of the elytral
cuticle of beetles, with an emphasis on weevils (Coleoptera: Curculionoidea).
Arthropod Structure and Development, 45, 14-22.
Van der Meij, M.A.A., & Bout, R.G. (2006) Seed husking time and maximal bite
force in finches. Journal of Experimental Biology, 209, 3329-3335.

30
Verwaijen, D., Van Damme, R., & Herrel, A. (2002) Relationships between head size,
bite force, prey handling efficiency and diet in two sympatric lacertid lizards.
Functional Ecology, 16, 842-850.
Wallace, A.R. (1867) Mimicry and other protective resemblance among animals.
Westminster and Foreign Quarterly Review, 32, 1-43.
Wallace, A.R. (1895) Natural Selection and Tropical Nature: Essays on Descriptive
and Theoretical Biology. Macmillan, London, UK.
Willink, B., Garcia-Rodriguez, A., Bolaños, F., & Pröhl, H. (2014). The interplay
between multiple predators and prey colour divergence. Biological Journal of the
Linnean Society, 113, 580-589.
Yewers, M.S., McLean, C.A., Moussalli, A., Stuart-Fox, D., Bennett, A.T.D. & Knott,
B. (2015) Spectral sensitivity of cone photoreceptors and opsin expression in two
colour-divergent lineages of the lizard Ctenophorus decresii. Journal of
Experimental Biology, 218, 1556-1563.

31
Figure 1. Pachyrhynchus sarcitis kotoensis and experimental manipulation. (a) ‘Hard’
and (b) ‘soft’ P. sarcitis kotoensis. (c) The predator, Japalura swinhonis lizard. (d)
The locality for measuring the colour spectrum of the blue spots (red circles) and the
black background (white circles) of P. sarcitis kotoensis. (e) ‘Claw’ (intact) and (f)
‘clawless’ (surgically removed) tarsi of P. sarcitis kotoensis. Images by L.Y. Wang (a
& b) and F.C. Hsu (c).

32
Figure 2. The reflectance spectra and daily rhythm of ‘hard’ and ‘soft’ Pachyrhynchus
sarcitis kotoensis in the behavioural experiments. (a) The reflectance spectra of blue
spot and dark background colours on the cuticles of the weevils (‘hard’ n = 4, ‘soft’ n
= 4) were measured using a spectrometer (Jaz spectrometer, Ocean Optics, Inc.,
Dunedin, Florida). The colour and grey shaded regions are the 95% confidence
intervals of the measurements. (b) The daily rhythm of the weevils (‘hard’ n = 10,
‘soft’ n = 10) were recorded for six consecutive days using a video monitor
(DS-VR7160H, Der Shuenn, Taipei, Taiwan). The weevils were placed in petri dishes
divided into four equal quadrants. The number of the weevil’s movements between
the quadrants in the first 10 minutes of every two hours was calculated. The shaded
regions are the standard deviation of the number of movements.
0
50
100
150
300 400 500 600 700
Reflc
etan
ce ra
tio (%
)
Wavelength (nm)
Hard weevil / blue spot
Hard weevil /black background
Soft weevil / blue spot
Soft weevil /black background 0
25
50
75
Num
ber o
f mov
emen
ts
SoftHard
7:00 A
M
7:00 P
M
Time
7:00 A
M
7:00 P
M
7:00 A
M
7:00 P
M
7:00 A
M
7:00 P
M
7:00 A
M
7:00 P
M
7:00 A
M
7:00 P
M
7:00 A
M
Day 1 Day 2 Day 3 Day 4 Day 5 Day 6
(a) (b)

33
Figure 3. The arena and recording set up for behavioral trials of Japalura swinhonis’s
predation on Pachyrhynchus sarcitis kotoensis (a). A: The bottom of the plastic
container was padded with soil. The three inner vertical planes of the plastic container
were covered with cardboard paper while left one side transparent for video recording.
B: The cardboard was used to hide the operator and video camera from disturbing the
lizards. C: The video camera (HDR-XR200, SONY, Tokyo, Japan). The weevil was
introduced to the arena using a black cotton thread tied between its thorax and
abdomen, and attached to the center of a transparent plastic bar. Experimental set-up
for measuring the hardness of insect prey (b). The force transducer (D) was attached
to the lower end of the camera mount on the copy stand (E). The upper and lower
jaws of the lizard were embedded in resin platforms (F) and (G), respectively. The
upper jaw was attached to the force transducer and the lower jaw was placed directly
below the upper jaw to match the upper one. The prey was oriented perpendicularly
with respect to the lower tooth rows (H).
D EF
G
(a) (b)
AB
C 5cm
15cm
10cm
58cm
3cm
43cm
30cm
40cm
H
GH
25mm
34
0
25
50
75
100
Surv
ival r
ate
(%)
0
25
50
75
Num
ber o
f bite
s
0
50
100
150
200
Prey
han
dling
dur
ation
(s)
HardSoft
HardSoft
HardSoft
Hard with claws
Softwith claws
Hard without claws
Softwithout claws
a a
b b
bb
a a
a a
b
b
(a)
(b)
(c)
n=20 n=20n=19 n=19

35
Figure 4. The survivorship of the weevil, Pachyrhynchus sarcitis kotoensis and
predatory response of the tree lizard, Japalura swinhonis in the behavioral trials. The
‘hard’ treatments were significantly different from ‘soft’ ones in (a) survival rate, (b)
number of bites, and (c) prey handling duration of the lizards. a and b indicate groups
with significant differences (Tukey post hoc tests, p < 0.001).
36
Figure 5. The hardness of Pachyrhynchus sarcitis kotoensis and insect prey from
Green and Orchid Islands. The regression line [log10(Force at failure) =
-0.9389+1.5885log10(Body length), adjusted R2 = 0.5262, p < 0.001, AIC = 69.94692]
is the correlation between log transformed hardness and body length by excluding
four insect families (Arc, Asc, Lib & Tet). The horizontal lines are the mean bite
forces of male and female tree lizards, Japalura swinhonis from Green and Orchid
Islands, with shaded regions indicating the 95% confidence intervals of the
measurements. Acr, Acrididae (Horned grasshoppers); Ant, Anthribidae (Fungus
weevils); Aph, Aphrophoridae (Spittlebugs); Asc, Ascalaphidae (Owlflies); Bla,
Acr
Acr
AphAsc
Bla
Bla
Bup
CarCar
CeraCera
Cera
Cera
Cera
Cerc
Cerc
Chr
Chr
ChrChr
Chr
Cur
Cur
CurCur
Cur
Ant
Bre
CurCur
Ela
Ela
For
ForFla
Tro
Tr oTro
Tro
Gry
GryGry
Hyb
Hyb
Lib
LucLuc
Lyc
Lyg
NitNit
Pen
PenPen
Pen
Pen Pyr
Sca
Sca
Sca
Sca
Sca
Sca
Sca
Sca
Sca
Sca
Sca
Sca
Ten
Ten
Tet
Tet
Tet
25
50
75
20 40 60Body length (mm)
Forc
e at
failu
re (N
)
Insect families
WeevilHard P. sarcitis
Soft P. sarcitis
Lizard
Orchid Island
Green Island
Orchid Island
Green Island
♀
♂
♂
♀
1
2
3
AcrAntAphAscBlaBreBupCarCeraCercChrCurElaFla
ForGryHybLibLucLycLygNitPenPyrScaTenTetTro
00

37
Blaberidae (Cockroaches); Bre, Brentidae (Straight-snouted weevils); Bup,
Buprestidae (Jewel beetles); Car, Carabidae (Ground beetles); Cera, Cerambycidae
(Long-horned beetles); Cerc, Cercopidae (Froghoppers); Chr, Chrysomelidae (Leaf
Beetles); Cur, Curculionoidea (Weevils); Ela, Elateridae (Click beetles); Fla, Flatidae
(Flatid planthoppers); For, Formicidae (Ants); Gry, Gryllidae (Crickets); Hyb,
Hybosoridae (Scavenger scarab beetles); Lib, Libellulidae (Dragonflies); Luc,
Lucanidae (Stag beetles); Lyc, Lycidae (Net-winged beetles); Lyg, Lygaeoidea
(Chinch bugs); Nit, Nitidulidae (Sap beetles); Pen, Pentatomoidea (Stink bugs); Pyr,
Pyrgomorphidae (Gaudy grasshoppers); Sca, Scarabaeidae (Scarab beetles), Ten,
Tenebrionidae (Darkling beetles); Tet, Tettigoniidae (Katydids); Tro, Tropiduchidae
(Tropiduchid planthoppers).

38
Figure 6. The master chromatogram and identified candidate compounds from the
extraction of Pachyrhynchus sarcitis kotoensis. The compounds that have SI
(similarity index) and RSI (relative similarity index) higher than 800 are shown in the
figure with retention time (RT). The chemical profile of these compounds are: Ethane
(solvent), RT: 1.72, SI: 935, RSI: 936, Chemical Abstract Service (CAS) registry
number: 60-29-7; Ethyl propyl ether, RT: 1.85, SI: 867, RSI: 890, CAS#: 628-32-0;
Diethylene glycol, RT: 5.30, SI: 967, RSI: 967, CAS#: 111-46-6; Butylated hydroxy
toluene, RT: 13.17, SI: 941, RSI: 945, CAS#: 128-37-0; Cadalene, RT: 15.65, SI: 947,
RSI: 956, CAS#: 483-78-3
5 10 15 20 25 30 35Time (min)
10
20
30
40
50
60
70
80
90
100R
elat
ive
abun
danc
e (%
)
00
RT: 1.72Ethane
RT: 1.85Ethyl propyl ether
RT: 5.30Diethylene glycol
RT: 13.17Butylated hydroxy toluene
RT: 15.65Cadalene

39
Table S1. List of insect prey species and hardness measurements from Green and Orchid Islands.
No. Order Family Species Common name Chinese name ID Abb. Body
part a
Hardn
ess (N)
Body
weight
(g)
Body
lengt
h
(mm)
Body
width
(mm)
Site b
36 Odonata Libellulidae/
蜻蜓科 Pantala flavescen Dragonfly 薄翅蜻蜓 Lu-Yi Wang Lib AN 2.8 0.302 48.62 8.40 OI
32 Orthoptera Acrididae/蝗科
Horned
Grasshopper Lu-Yi Wang Acr TX 5.3 0.279 53.49 5.12 OI
64 Orthoptera Acrididae
Horned
Grasshopper Lu-Yi Wang Acr TX 3.3 1.458 61.87 7.83 GI
7 Orthoptera Gryllidae/蟋蟀科
Cricket
Lu-Yi Wang Gry AN 0.7 0.052 5.55 1.91 OI
39 Orthoptera Gryllidae
Cricket
Lu-Yi Wang Gry AN 4.3 1.100 25.24 8.62 OI
57 Orthoptera Gryllidae
Cricket
Lu-Yi Wang Gry AN 3.7 0.068 12.01 3.44 GI
21 Orthoptera Pyrgomorphidae/
錐頭蝗科
Gaudy
grasshopper Lu-Yi Wang Pyr TX 2.6 0.103 20.65 3.48 OI
38 Orthoptera Tettigoniidae/
螽斯科
Katydid
Lu-Yi Wang Tet TX 1.8 4.472 52.70 9.40 OI
65 Orthoptera Tettigoniidae
Katydid
Lu-Yi Wang Tet AN 2.7 0.243 36.92 4.82 GI
30 Orthoptera Tettigoniidae Mecopoda elongata Katydid 臺灣騷斯 Feng-Chuan Hsu Tet AN 15.2 2.337 16.66 8.48 OI
40
49 Blattodea Blaberidae/
匍蜚蠊科
Cockroach
Lu-Yi Wang Bla TX 0.5 0.070 14.54 6.52 OI
62 Blattodea Blaberidae
Cockroach
Lu-Yi Wang Bla TX 6.4 0.283 27.33 11.55 GI
80 Hemiptera Aphrophoridae/
尖胸沫蟬科 Ariptyelus sp. Spittlebug
Shun-Chern
Tsaur Aph AN 1.1 0.011 6.72 2.81 GI
9 Hemiptera Cercopidae/
沫蟬科 Eoscarta botelensis Froghopper 蘭嶼曙沫蟬
Hsien-Tzung
Shih Cerc AN 0.7 0.009 7.08 3.48 OI
17 Hemiptera Cercopidae Eoscarta botelensis Froghopper 蘭嶼曙沫蟬 Hsien-Tzung
Shih Cerc AN 0.9 0.008 7.04 3.07 OI
68 Hemiptera Flatidae/
青翅飛蝨科 Mimophantia maritima
Flatid
planthopper 禾草褐蛾蠟禪
Shun-Chern
Tsaur Fla AN 1.4 0.016 6.89 3.09 GI
23 Hemiptera Lygaeoidea/
長蝽科 Neoletheus sp. Chinch bug
Jing-Fu Tsai Lyg AN 0.5 0.007 6.27 2.25 OI
26 Hemiptera Pentatomoidea/
蝽科 Lampromicra myakona Stink bug
Jing-Fu Tsai Pen AN 4.3 0.078 10.96 5.44 OI
52 Hemiptera Pentatomoidea Canto ocellatus Stink bug 角盾蝽 Jing-Fu Tsai Pen AN 4.8 0.206 21.12 10.00 OI
34 Hemiptera Pentatomoidea Glaucia sp. Stink bug
Jing-Fu Tsai Pen AN 7.5 0.128 15.54 8.37 OI
61 Hemiptera Pentatomoidea Leptocorisa acuta Stink bug 禾蛛緣椿象 Jing-Fu Tsai Pen AN 2.7 0.156 16.65 2.99 GI
45 Hemiptera Pentatomoidea Plautia sp. Stink bug
Jing-Fu Tsai Pen AN 7.7 0.082 10.82 6.74 OI
78 Hemiptera Tropiduchidae/ Kallitaxila sinica Tropiduchid Shun-Chern Tro AN 0.8 0.010 6.16 2.74 GI

41
軍配飛蝨科 planthopper Tsaur
79 Hemiptera Tropiduchidae Kallitaxila sinica Tropiduchid
planthopper
Shun-Chern
Tsaur Tro AN 0.8 0.018 6.97 3.09 GI
76 Hemiptera Tropiduchidae Sogana hopponis Tropiduchid
planthopper 軍配飛蝨
Shun-Chern
Tsaur Tro AN 2.0 0.017 6.05 2.52 GI
81 Hemiptera Tropiduchidae Sogana hopponis Tropiduchid
planthopper
Shun-Chern
Tsaur Tro AN 2.9 0.023 11.58 3.96 GI
53(2) Coleoptera Anthribidae/
長角象鼻蟲科 Anthribidae sp. Fungus weevil
Chen-Fu Hsu Ant AN 2.5 0.023 8.80 2.95 OI
53 Coleoptera Brentidae/
三錐象鼻蟲科 Baryrrhynchus sp.
Straight-snoute
d weevil Chen-Fu Hsu Bre AN 11.0 0.025 13.94 3.28 OI
83 Coleoptera Buprestidae/
吉丁蟲科 Chrysodema dalmanni Jewel beetle 粉彩吉丁蟲 Feng-Chuan Hsu Bup AN 33.9 0.636 26.23 9.00 GI
67 Coleoptera Carabidae/
步行蟲科 Calleida sp. Ground beetle
Lan-Wei Yeh Car AN 1.8 0.022 9.83 3.02 GI
44 Coleoptera Carabidae Cosmodela batesi Ground beetle 臺灣八星虎甲蟲 Lu-Yi Wang Car AN 2.5 0.128 16.71 5.43 OI
22 Coleoptera Cerambycidae/
天牛科 Ceresium longicorne
Long-horned
beetle 長角姬天牛 Sin-Yan Shih Cera AN 1.6 0.009 8.04 1.85 OI
10 Coleoptera Cerambycidae Doliops similis Long-horned
beetle 擬硬象天牛 Sin-Yan Shih Cera AN 11.5 0.243 14.61 5.84 OI

42
31 Coleoptera Cerambycidae Epepeotes ambigenus Long-horned
beetle 蘭嶼縱紋長角天牛 Sin-Yan Shih Cera AN 39.9 0.623 24.69 8.00 OI
3 Coleoptera Cerambycidae Olenecamptus bilobus Long-horned
beetle 五星白天牛 Sin-Yan Shih Cera AN 15.3 0.118 17.25 4.22 OI
48 Coleoptera Cerambycidae Sybra sp. Long-horned
beetle Sin-Yan Shih Cera AN 6.8 0.042 11.28 2.78 OI
77 Coleoptera Chrysomelidae/
金花蟲科 Cleorina janthina Leaf beetle 小溝腳猿金花蟲 Chi-Feng Lee Chr AN 1.3 0.006 3.67 2.28 GI
18 Coleoptera Chrysomelidae Pyrrhalta sp. Leaf beetle
Chi-Feng Lee Chr AN 3.6 0.010 4.68 2.43 OI
28 Coleoptera Chrysomelidae Monolepta ongi Leaf beetle 蘭嶼長腳螢金花蟲 Chi-Feng Lee Chr AN 1.5 <0.001 3.67 2.25 OI
47 Coleoptera Chrysomelidae Rhyparida sakisimensis Leaf beetle 巨島嶼猿金花蟲 Chi-Feng Lee Chr AN 2.4 0.010 4.79 2.91 OI
75 Coleoptera Chrysomelidae Rhyparida sakisimensis Leaf beetle 巨島嶼猿金花蟲 Chi-Feng Lee Chr AN 3.0 0.022 5.15 3.12 GI
6 Coleoptera Curculionidae/
象鼻蟲科 Aclees hirayamai Weevil 白波粉象鼻蟲 Chen-Fu Hsu Cur AN 29.9 0.149 16.65 5.86 OI
37 Coleoptera Curculionidae Mechistocerus sp. Weevil
Chen-Fu Hsu Cur AN 19.7 0.067 9.75 4.23 OI
13 Coleoptera Curculionidae Metapocyrtus immeritus Weevil 紅足鏽象鼻 Chen-Fu Hsu Cur AN 10.5 0.040 8.23 3.36 OI
14 Coleoptera Curculionidae Metapocyrtus immeritus Weevil 紅足鏽象鼻 Chen-Fu Hsu Cur AN 19.0 0.061 10.04 3.83 OI
73 Coleoptera Curculionidae Metapocyrtus sp. Weevil
Chen-Fu Hsu Cur AN 13.7 0.048 9.13 3.87 GI
74 Coleoptera Curculionidae Metapocyrtus sp. Weevil
Chen-Fu Hsu Cur AN 17.9 0.095 10.99 4.36 GI
p1 Coleoptera Curculionidae Pachyrhynchus sarcitis Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 29.5 0.250 13.94 3.28 OI
43
(hard)
160514 Coleoptera Curculionidae Pachyrhynchus sarcitis
(hard) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 49.0 0.306 17.12 6.43 OI
160505
D Coleoptera Curculionidae
Pachyrhynchus sarcitis
(hard) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 55.5 0.364 18.07 6.83 OI
160515 Coleoptera Curculionidae Pachyrhynchus sarcitis
(hard) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 26.1 0.410 19.74 7.29 OI
p5 Coleoptera Curculionidae Pachyrhynchus sarcitis
(hard) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 20.1 0.240 17.22 6.32 OI
p6 Coleoptera Curculionidae Pachyrhynchus sarcitis
(hard) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 28.2 0.241 17.54 6.38 OI
p7 Coleoptera Curculionidae Pachyrhynchus sarcitis
(hard) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 28.2 0.190 15.54 5.85 OI
p8 Coleoptera Curculionidae Pachyrhynchus sarcitis
(hard) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 31.6 0.297 18.68 6.91 OI
p9 Coleoptera Curculionidae Pachyrhynchus sarcitis
(hard) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 37.4 0.189 16.03 5.95 OI
p10 Coleoptera Curculionidae Pachyrhynchus sarcitis
(hard) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 34.5 0.182 15.03 5.44 OI
p11 Coleoptera Curculionidae Pachyrhynchus sarcitis Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 37.1 0.255 17.22 6.45 OI
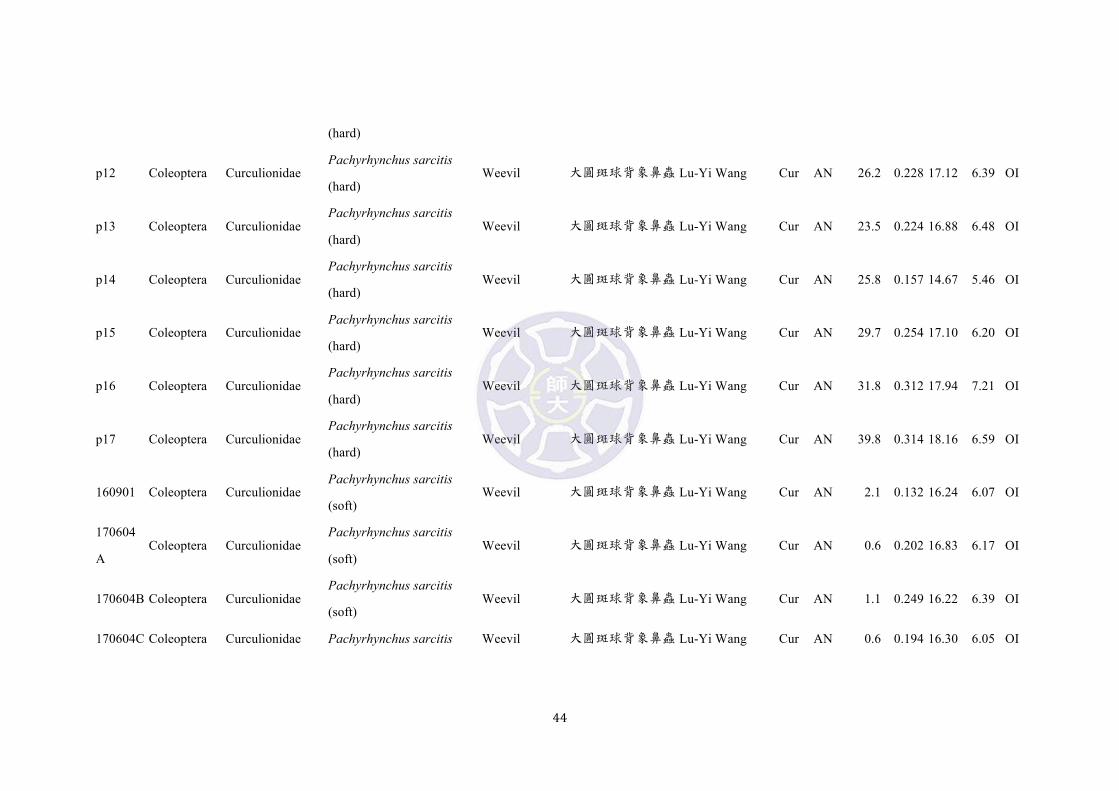
44
(hard)
p12 Coleoptera Curculionidae Pachyrhynchus sarcitis
(hard) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 26.2 0.228 17.12 6.39 OI
p13 Coleoptera Curculionidae Pachyrhynchus sarcitis
(hard) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 23.5 0.224 16.88 6.48 OI
p14 Coleoptera Curculionidae Pachyrhynchus sarcitis
(hard) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 25.8 0.157 14.67 5.46 OI
p15 Coleoptera Curculionidae Pachyrhynchus sarcitis
(hard) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 29.7 0.254 17.10 6.20 OI
p16 Coleoptera Curculionidae Pachyrhynchus sarcitis
(hard) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 31.8 0.312 17.94 7.21 OI
p17 Coleoptera Curculionidae Pachyrhynchus sarcitis
(hard) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 39.8 0.314 18.16 6.59 OI
160901 Coleoptera Curculionidae Pachyrhynchus sarcitis
(soft) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 2.1 0.132 16.24 6.07 OI
170604
A Coleoptera Curculionidae
Pachyrhynchus sarcitis
(soft) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 0.6 0.202 16.83 6.17 OI
170604B Coleoptera Curculionidae Pachyrhynchus sarcitis
(soft) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 1.1 0.249 16.22 6.39 OI
170604C Coleoptera Curculionidae Pachyrhynchus sarcitis Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 0.6 0.194 16.30 6.05 OI
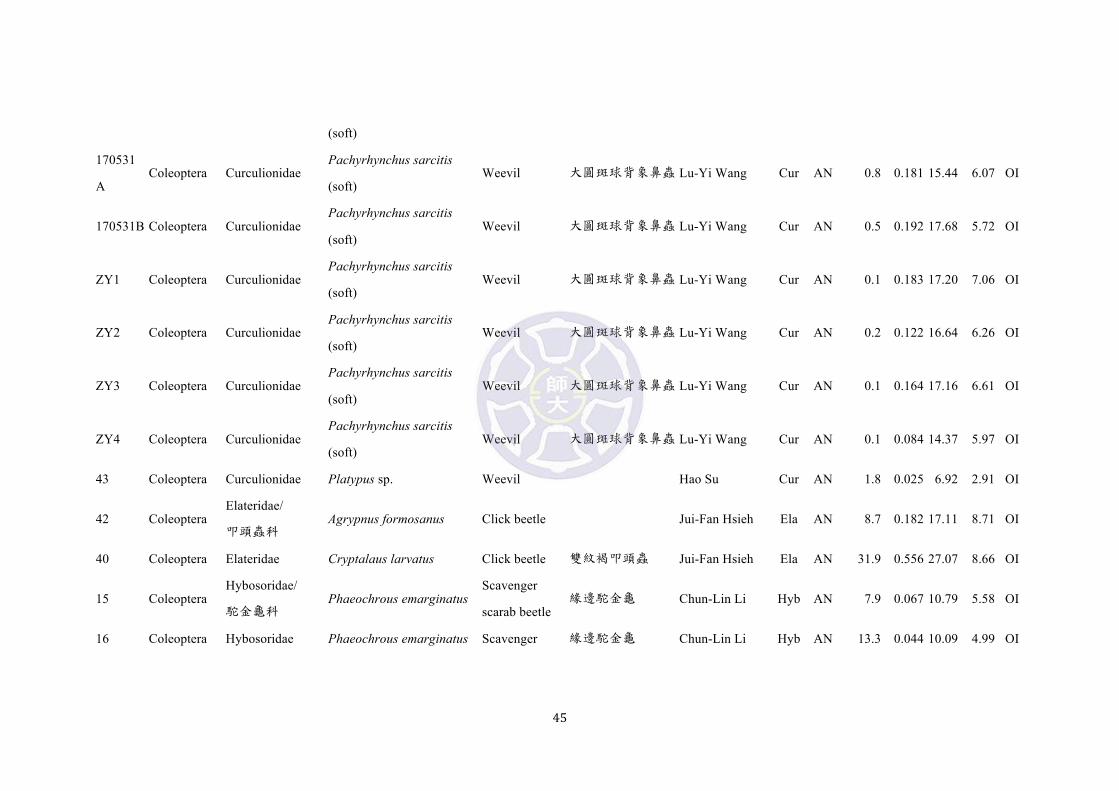
45
(soft)
170531
A Coleoptera Curculionidae
Pachyrhynchus sarcitis
(soft) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 0.8 0.181 15.44 6.07 OI
170531B Coleoptera Curculionidae Pachyrhynchus sarcitis
(soft) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 0.5 0.192 17.68 5.72 OI
ZY1 Coleoptera Curculionidae Pachyrhynchus sarcitis
(soft) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 0.1 0.183 17.20 7.06 OI
ZY2 Coleoptera Curculionidae Pachyrhynchus sarcitis
(soft) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 0.2 0.122 16.64 6.26 OI
ZY3 Coleoptera Curculionidae Pachyrhynchus sarcitis
(soft) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 0.1 0.164 17.16 6.61 OI
ZY4 Coleoptera Curculionidae Pachyrhynchus sarcitis
(soft) Weevil 大圓斑球背象鼻蟲 Lu-Yi Wang Cur AN 0.1 0.084 14.37 5.97 OI
43 Coleoptera Curculionidae Platypus sp. Weevil
Hao Su Cur AN 1.8 0.025 6.92 2.91 OI
42 Coleoptera Elateridae/
叩頭蟲科 Agrypnus formosanus Click beetle
Jui-Fan Hsieh Ela AN 8.7 0.182 17.11 8.71 OI
40 Coleoptera Elateridae Cryptalaus larvatus Click beetle 雙紋褐叩頭蟲 Jui-Fan Hsieh Ela AN 31.9 0.556 27.07 8.66 OI
15 Coleoptera Hybosoridae/
駝金龜科 Phaeochrous emarginatus
Scavenger
scarab beetle 緣邊駝金龜 Chun-Lin Li Hyb AN 7.9 0.067 10.79 5.58 OI
16 Coleoptera Hybosoridae Phaeochrous emarginatus Scavenger 緣邊駝金龜 Chun-Lin Li Hyb AN 13.3 0.044 10.09 4.99 OI

46
scarab beetle
8 Coleoptera Lucanidae/
鍬形蟲科 Metallactulus parvulus Stag beetle 姬扁鍬形蟲 Chun-Lin Li Luc AN 12.7 0.132 13.87 6.34 OI
20 Coleoptera Lucanidae Metallactulus parvulus Stag beetle 姬扁鍬形蟲 Chun-Lin Li Luc AN 15.5 0.219 16.39 7.84 OI
82 Coleoptera Lycidae/紅螢科
Net-winged
beetle Lu-Yi Wang Lyc AN 2.8 0.050 13.80 5.28 GI
19 Coleoptera Nitidulidae/
出尾蟲科
Sap beetle
Wenbe Hwang Nit AN 3.1 0.016 6.44 3.59 OI
46 Coleoptera Nitidulidae
Sap beetle
Wenbe Hwang Nit AN 2.9 0.020 6.85 3.60 OI
56 Coleoptera Scarabaeidae/
金龜子科
Anomala expansa
lutaoensis Scarab beetle 綠島青銅金龜 Chun-Lin Li Sca AN 37.3 1.111 23.09 14.16 GI
63 Coleoptera Scarabaeidae Anomala expansa
lutaoensis Scarab beetle 綠島青銅金龜 Chun-Lin Li Sca AN 49.4 1.213 25.18 14.18 GI
4 Coleoptera Scarabaeidae Anomala siniopyga Scarab beetle 櫻花青銅金龜 Chun-Lin Li Sca AN 10.5 0.157 12.72 6.59 OI
5 Coleoptera Scarabaeidae Anomala siniopyga Scarab beetle 櫻花青銅金龜 Chun-Lin Li Sca AN 14.5 0.205 12.57 7.09 OI
66 Coleoptera Scarabaeidae Anomala siniopyga Scarab beetle 櫻花青銅金龜 Chun-Lin Li Sca AN 15.4 0.421 15.41 9.34 GI
33 Coleoptera Scarabaeidae Maladera hongkongica Scarab beetle 香港絨毛金龜 Chun-Lin Li Sca AN 3.7 0.122 11.49 5.89 OI
58 Coleoptera Scarabaeidae Maladera annamensis Scarab beetle 安南絨毛金龜 Chun-Lin Li Sca AN 1.4 0.015 8.15 4.71 GI
50 Coleoptera Scarabaeidae Papuana philippinica Scarab beetle 菲律賓圓金龜 Chun-Lin Li Sca AN 41.3 14.393 23.13 11.26 OI
1 Coleoptera Scarabaeidae Xylotrupes philippinensis Scarab beetle 蘭嶼姬兜蟲 Chun-Lin Li Sca AN 60.8 18.590 30.80 16.68 OI
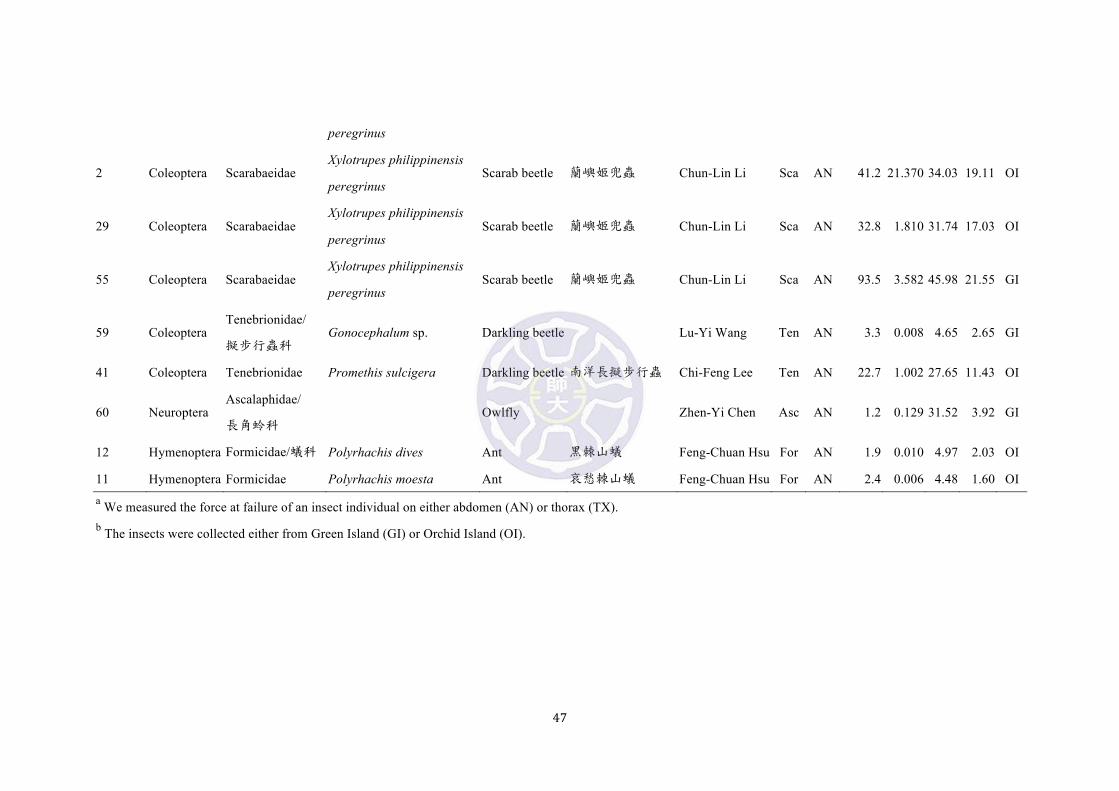
47
peregrinus
2 Coleoptera Scarabaeidae Xylotrupes philippinensis
peregrinus Scarab beetle 蘭嶼姬兜蟲 Chun-Lin Li Sca AN 41.2 21.370 34.03 19.11 OI
29 Coleoptera Scarabaeidae Xylotrupes philippinensis
peregrinus Scarab beetle 蘭嶼姬兜蟲 Chun-Lin Li Sca AN 32.8 1.810 31.74 17.03 OI
55 Coleoptera Scarabaeidae Xylotrupes philippinensis
peregrinus Scarab beetle 蘭嶼姬兜蟲 Chun-Lin Li Sca AN 93.5 3.582 45.98 21.55 GI
59 Coleoptera Tenebrionidae/
擬步行蟲科 Gonocephalum sp. Darkling beetle
Lu-Yi Wang Ten AN 3.3 0.008 4.65 2.65 GI
41 Coleoptera Tenebrionidae Promethis sulcigera Darkling beetle 南洋長擬步行蟲 Chi-Feng Lee Ten AN 22.7 1.002 27.65 11.43 OI
60 Neuroptera Ascalaphidae/
長角蛉科 Owlfly
Zhen-Yi Chen Asc AN 1.2 0.129 31.52 3.92 GI
12 Hymenoptera Formicidae/蟻科 Polyrhachis dives Ant 黑棘山蟻 Feng-Chuan Hsu For AN 1.9 0.010 4.97 2.03 OI
11 Hymenoptera Formicidae Polyrhachis moesta Ant 哀愁棘山蟻 Feng-Chuan Hsu For AN 2.4 0.006 4.48 1.60 OI a We measured the force at failure of an insect individual on either abdomen (AN) or thorax (TX). b The insects were collected either from Green Island (GI) or Orchid Island (OI).

48
Table S2. List of candidate mammal and reptile predators of Pachyrhynchus weevils from Green and Orchid Islands (Chen et al. 2008; Li et al.
2010). We selected the candidate predators (*) based on their diet and distribution. Class Family Species Common name Chinese name Location
Mammalia Soricidae Crocidura rapax tadae Chinese white-toothed shrew 蘭嶼長尾麝鼩 Orchid Island
Soricidae Crocidura rapax lutaoensis Chinese white-toothed shrew 綠島長尾麝鼩 Green Island
Rhinolophidae Hipposideros armiger terasensis Formosan leaf-nosed bat 台灣葉鼻蝠 Green Island
Rhinolophidae Rhinolophus monoceros Formosan lesser horseshoe bat 台灣小蹄鼻蝠 Orchid Island
Molossidae Tadarida insignis European free-tailed bat 游離尾蝠 Orchid Island
Vespertilionidae Pipistrellus abramus Japanese house bat 東亞家蝠 Green Island
Viverridae *Paguma larvata Masked palm civet 白鼻心 Green Island; Orchid Island
Sciuridae *Callosciurus erythraeus thaiwanensis Pallas's squirrel 赤腹松鼠 Green Island
Muridae *Mus musculus House mouse 家鼷鼠 Green Island
Muridae *Rattus losea Lesser ricefield rat 小黃腹鼠 Green Island; Orchid Island
Muridae *Rattus tanezumi Tanezumi rat 亞洲家鼠 Green Island; Orchid Island
Muridae *Bandicota indica Greater bandicoot rat 鬼鼠 Orchid Island
Muridae *Rattus norvegicus Brown rat 溝鼠 Green Island; Orchid Island
Muridae *Rattus rattus Black rat 玄鼠 Orchid Island
Reptilia Agamidae *Japalura swinhonis Swinhoe's tree lizard 斯文豪氏攀蜥 Green Island; Orchid Island
Lacertidae Takydromus luyeanus
台灣草蜥 Green Island
Lacertidae Takydromus sauteri Koshun grass lizard 梭德氏草蜥 Green Island; Orchid Island

49
Scincidae Emoia atrocostata Mangrove skink 岩岸島蜥 Green Island; Orchid Island
Scincidae *Eutropis longicaudata Long-tailed sun skink 長尾南蜥 Green Island; Orchid Island
Scincidae *Eutropis multifasciata East Indian brown mabuya 多線南蜥 Green Island; Orchid Island
Scincidae *Plestiodon leucostictus
白斑石龍子 Green Island
Scincidae *Sphenomorphus incognitus Brown forest skink 股鱗蜓蜥 Green Island; Orchid Island
Scincidae *Eutropis cumingi Cuming's mabuya 庫氏南蜥 Orchid Island
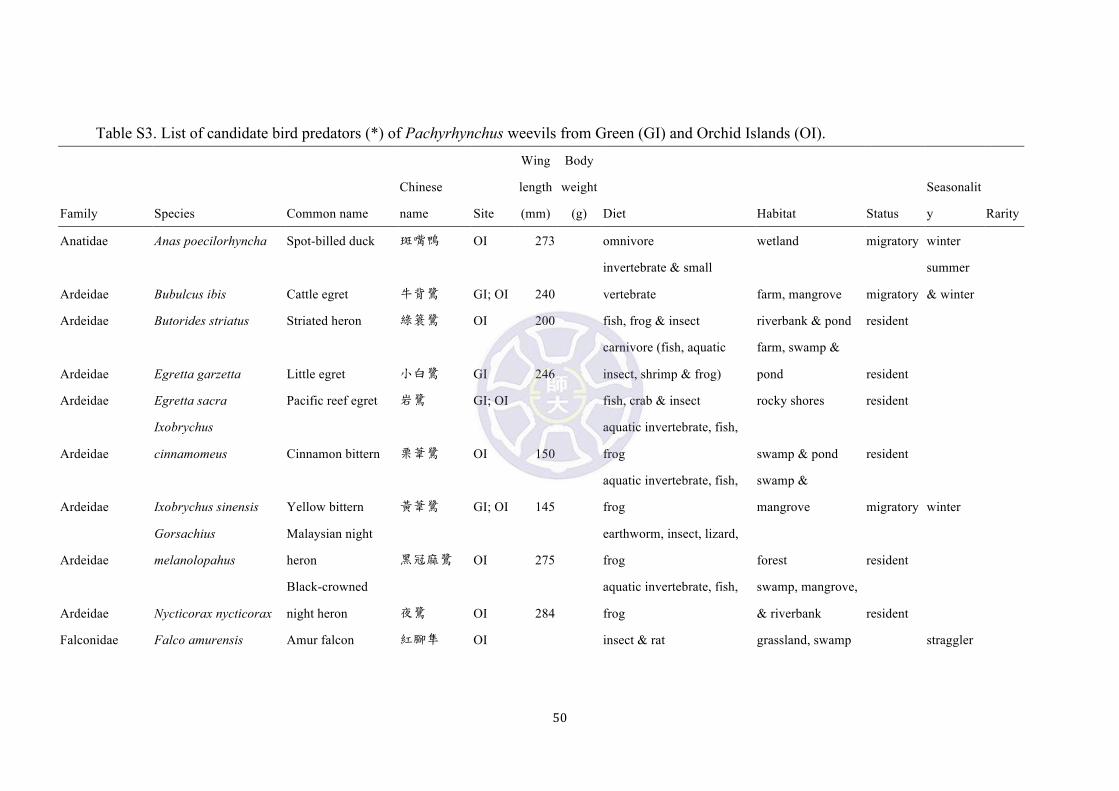
50
Table S3. List of candidate bird predators (*) of Pachyrhynchus weevils from Green (GI) and Orchid Islands (OI).
Family Species Common name
Chinese
name Site
Wing
length
(mm)
Body
weight
(g) Diet Habitat Status
Seasonalit
y Rarity
Anatidae Anas poecilorhyncha Spot-billed duck 斑嘴鴨 OI 273
omnivore wetland migratory winter
Ardeidae Bubulcus ibis Cattle egret 牛背鷺 GI; OI 240
invertebrate & small
vertebrate farm, mangrove migratory
summer
& winter
Ardeidae Butorides striatus Striated heron 綠簑鷺 OI 200
fish, frog & insect riverbank & pond resident
Ardeidae Egretta garzetta Little egret 小白鷺 GI 246
carnivore (fish, aquatic
insect, shrimp & frog)
farm, swamp &
pond resident
Ardeidae Egretta sacra Pacific reef egret 岩鷺 GI; OI
fish, crab & insect rocky shores resident
Ardeidae
Ixobrychus
cinnamomeus Cinnamon bittern 栗葦鷺 OI 150
aquatic invertebrate, fish,
frog swamp & pond resident
Ardeidae Ixobrychus sinensis Yellow bittern 黃葦鷺 GI; OI 145
aquatic invertebrate, fish,
frog
swamp &
mangrove migratory winter
Ardeidae
Gorsachius
melanolopahus
Malaysian night
heron 黑冠麻鷺 OI 275
earthworm, insect, lizard,
frog forest resident
Ardeidae Nycticorax nycticorax
Black-crowned
night heron 夜鷺 OI 284
aquatic invertebrate, fish,
frog
swamp, mangrove,
& riverbank resident
Falconidae Falco amurensis Amur falcon 紅腳隼 OI
insect & rat grassland, swamp straggler
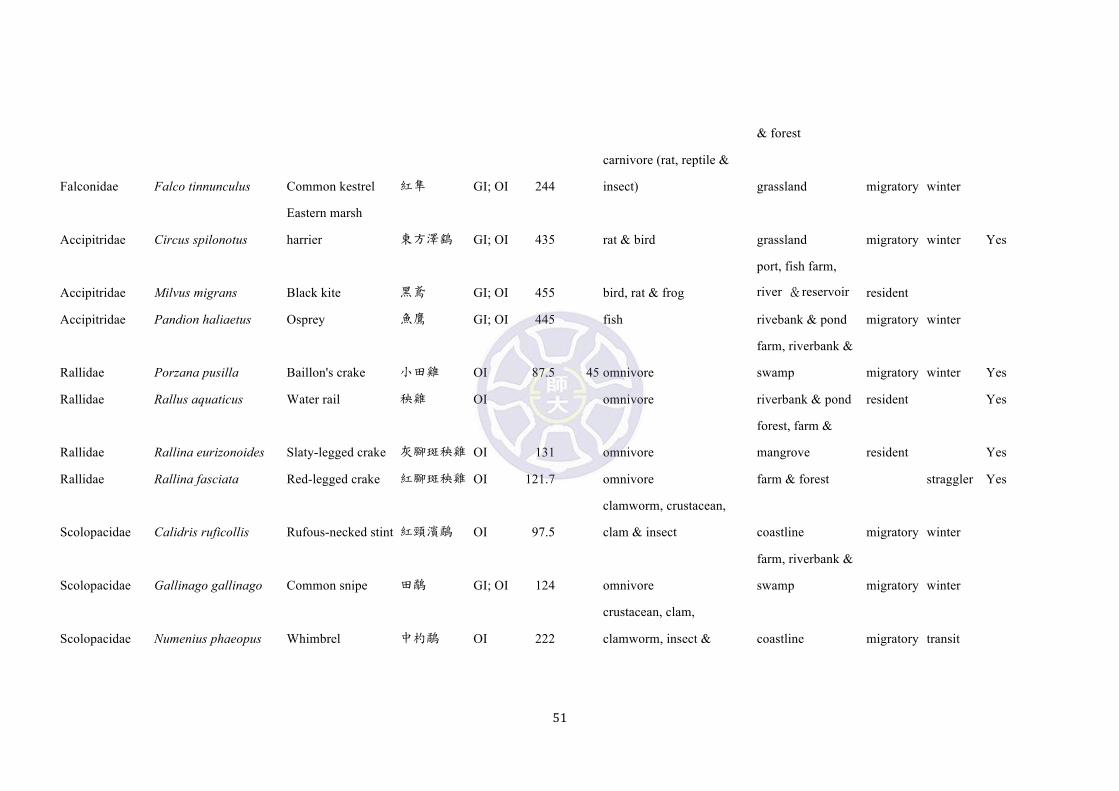
51
& forest
Falconidae Falco tinnunculus Common kestrel 紅隼 GI; OI 244
carnivore (rat, reptile &
insect) grassland migratory winter
Accipitridae Circus spilonotus
Eastern marsh
harrier 東方澤鷂 GI; OI 435
rat & bird grassland migratory winter Yes
Accipitridae Milvus migrans Black kite 黑鳶 GI; OI 455
bird, rat & frog
port, fish farm,
river &reservoir resident
Accipitridae Pandion haliaetus Osprey 魚鷹 GI; OI 445
fish rivebank & pond migratory winter
Rallidae Porzana pusilla Baillon's crake 小田雞 OI 87.5 45 omnivore
farm, riverbank &
swamp migratory winter Yes
Rallidae Rallus aquaticus Water rail 秧雞 OI
omnivore riverbank & pond resident
Yes
Rallidae Rallina eurizonoides Slaty-legged crake 灰腳斑秧雞 OI 131
omnivore
forest, farm &
mangrove resident
Yes
Rallidae Rallina fasciata Red-legged crake 紅腳斑秧雞 OI 121.7
omnivore farm & forest
straggler Yes
Scolopacidae Calidris ruficollis Rufous-necked stint 紅頸濱鷸 OI 97.5
clamworm, crustacean,
clam & insect coastline migratory winter
Scolopacidae Gallinago gallinago Common snipe 田鷸 GI; OI 124
omnivore
farm, riverbank &
swamp migratory winter
Scolopacidae Numenius phaeopus Whimbrel 中杓鷸 OI 222
crustacean, clam,
clamworm, insect & coastline migratory transit

52
earthworm
Laridae Chlidonias leucopterus White-winged tern 白翅黑浮鷗 OI 201
aquatic insect, small fish &
tadpole
coastline, estuary,
wetland & farm migratory
spring &
fall
Laridae Stercorarius pomarinus Pomarine skua 中賊鷗 OI
small mammal, fish,
crustacean & insect coastline & estuary migratory transit Yes
Laridae Sterna albifrons Little tern 小燕鷗 GI; OI 184
fish, shrimp & insect
coastline &
wetland migratory summer
Columbidae *Chalcophaps indica Emerald dove 翠翼鳩 GI; OI 144.5
omnivore forest resident
Cuculidae *Centropus bengalensis Lesser coucal 小鴉鵑 OI 156
omnivore open area & bush resident
Cuculidae *Cuculus sparverioides Large hawk-cuckoo 鷹鵑 OI 203
omnivore forest migratory summer
Cuculidae *Cuculus saturatus Himalayan cuckoo 中杜鵑 GI; OI 185
omnivore forest migratory summer
Strigidae Otus elegans Elegant scops owl 優雅角鴞 OI 169 115 insectivore forest; nocturnal resident
Strigidae Otus sunia Oriental scops owl 東方角鴞 OI 149
carnivore
savanna, forest in
the wetland & city
park; nocturnal migratory transit
Strigidae Asio otus Long-eared owl 長耳鴞 OI 284
carnivore forest migratory winter Yes
Strigidae Ninox scutulata Brown hawk-owl 褐鷹鴞 GI; OI 238
insectivore, also small
mammal forest; nocturnal resident
Caprimulgidae Caprimulgus affinis Savanna nightjar 南亞夜鷹 OI 187
insectivore river bed resident
Apodidae Apus nipalensis House swift 家雨燕 GI; OI 134
insectivore (only flying farm & city resident
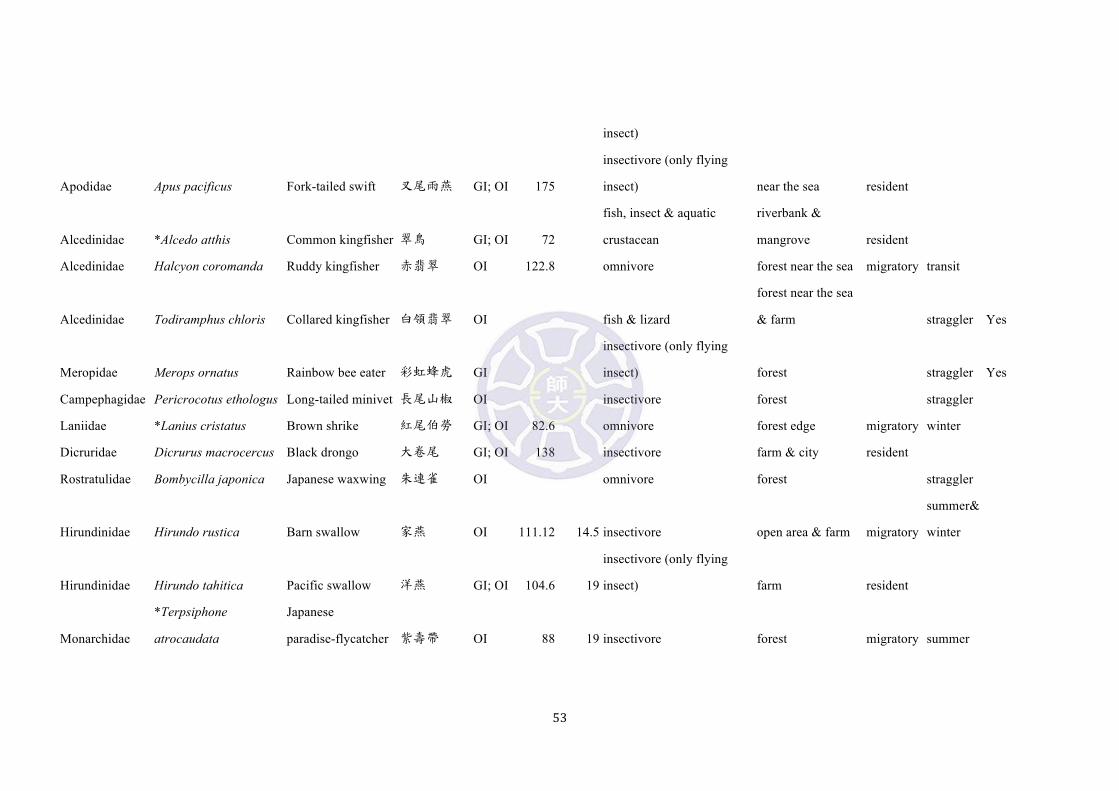
53
insect)
Apodidae Apus pacificus Fork-tailed swift 叉尾雨燕 GI; OI 175
insectivore (only flying
insect) near the sea resident
Alcedinidae *Alcedo atthis Common kingfisher 翠鳥 GI; OI 72
fish, insect & aquatic
crustacean
riverbank &
mangrove resident
Alcedinidae Halcyon coromanda Ruddy kingfisher 赤翡翠 OI 122.8
omnivore forest near the sea migratory transit
Alcedinidae Todiramphus chloris Collared kingfisher 白領翡翠 OI
fish & lizard
forest near the sea
& farm straggler Yes
Meropidae Merops ornatus Rainbow bee eater 彩虹蜂虎 GI
insectivore (only flying
insect) forest
straggler Yes
Campephagidae Pericrocotus ethologus Long-tailed minivet 長尾山椒 OI
insectivore forest
straggler
Laniidae *Lanius cristatus Brown shrike 紅尾伯勞 GI; OI 82.6
omnivore forest edge migratory winter
Dicruridae Dicrurus macrocercus Black drongo 大卷尾 GI; OI 138
insectivore farm & city resident
Rostratulidae Bombycilla japonica Japanese waxwing 朱連雀 OI
omnivore forest
straggler
Hirundinidae Hirundo rustica Barn swallow 家燕 OI 111.12 14.5 insectivore open area & farm migratory
summer&
winter
Hirundinidae Hirundo tahitica Pacific swallow 洋燕 GI; OI 104.6 19
insectivore (only flying
insect) farm resident
Monarchidae
*Terpsiphone
atrocaudata
Japanese
paradise-flycatcher 紫壽帶 OI 88 19 insectivore forest migratory summer

54
Corvidae Corvus dauuricus Daurian jackdaw 東方寒鴉 OI
omnivore forest & farm
straggler
Corvidae Corvus splendens House crow 家烏鴉 OI
omnivore human community
straggler
Alaudidae Alauda gulgula Oriental skylark 小雲雀 OI 98 21.7 omnivore open area & farm resident
Pycnonotidae *Microscelis amaurotis Brown-eared bulbul 棕耳鵯 GI; OI 128 73 omnivore forest resident
Zosteropidae *Zosterops meyeni Lowland white-eye 低地繡眼 GI; OI 41.1
omnivore forest & bush resident
Sturnidae Sturnus cineraceus
White-cheeked
starling 灰椋鳥 GI; OI
omnivore farm migratory winter
Sturnidae Sturnus philippensis
Chestnut-cheeked
starling 紫背椋鳥 OI
omnivore farm migratory transit Yes
Sturnidae Sturnus sericeus Red-billed starling 絲光椋鳥 OI 118
omnivore farm migratory winter
Sturnidae *Sturnus sinensis
White-shouldered
starling 灰背椋鳥 GI; OI
omnivore savanna & park migratory winter
Sturnidae Sturnus vulgaris European starling 歐洲椋鳥 OI
omnivore open area & farm migratory winter Yes
Turdidae *Turdus obscurus Eyebrowed thrush 白眉鶇 GI 127.7 63 omnivore forest migratory winter
Muscicapidae *Monticola solitarius Blue rock thrush 藍磯鶇 GI; OI 112.7 53 omnivore open area migratory winter
Fringillidae Carpodacus erythrinus Common rosefinch 普通朱雀 OI
omnivore bush & forest
straggler Yes
Fringillidae Carduelis flammea Common redpoll 白腰朱頂雀 OI
omnivore
forest along the
coastline &
grassland straggler Yes
Emberizidae Emberiza Pine bunting 白頭鵐 OI
omnivore open area & farm
straggler

55
leucocephalos
Emberizidae Emberiza pallasi Pallas's bunting 葦鵐 OI
omnivore swamp & bush
straggler

56
Movie 1. ‘Hard’ weevil. A Pachyrhynchus sarcitis kotoensis weevil was introduced in
the arena (00:12), and a Japalura swinhonis lizard noticed the weevil by shifting its
body (00:20) and turning its head toward the weevil (00:27). The lizard approached
the weevil (00:48) when it resumed moving, and then spitted it out immediately
(00:51) after giving it a bite (00:50) without any visible damage to the weevil.

57
Movie 2: ‘Soft’ weevil. A Pachyrhynchus sarcitis kotoensis weevil was introduced in
the arena (00:12), and a Japalura swinhonis lizard noticed the weevil by turning its
head (00:30) and quickly approached the weevil (00:35). The lizard jumped onto the
weevil (00:39), bit on its abdomen (00:40) and started chewing it continuously
(00:41). After ~ 40 seconds, the weevil was completely consumed (01:19).