훼손된 자연 생태계 복원기술webbook.me.go.kr/dli-file/075/172704.pdf · 서식처의...
TRANSCRIPT

- 1 -
과제 번호: 051-041-013
훼손된 자연 생태계 복원기술Restoration technology for damaged ecosystem
낙동강 하구역 습지 생태계 훼손지역 복원 및 관리 기술
Restoration and management for degraded wetland
ecosystem in Nak-Dong estuary
주관연구기관 : 부산대학교
환 경 부

- 1 -
제 출 문
환경부장관 귀하
본 보고서를 “낙동강 하구역 습지 생태계 훼손지역 복원 및 관리기
술”의 보고서로 제출합니다.
2007 년 3 월 31 일
주관연구기관명 : 부산대학교
연구책임자 : 안순모
연구원 : 강창근
연구원 : 정익교
연구원 : 이근섭
연구원 : 이동섭
- 2 -
보고서 초록
과제번호051-041
-013
해당단계
연구기간
2006.04.01
~
2007.03.31
구분 3차년도 / 총 3년
연구사업명 차세대 핵심환경기술개발사업대분야명 생태계 복원.관리중분야명 훼손된 자연생태계 복원기술
연구개발
과제명
국문명 낙동강 하구역 습지 생태계 훼손지역 복원 및 관리기술
영문명Restoration and management for degraded wetland
ecosystem in Nak-Dong estuary
연구책임자 안 순 모
해당단계
참여연구원
수
총 : 16 명
내부 : 5 명
외부 : 11 명
해당단계
연구개발
비
정부: 235,000 천원
기업: 천원
계 : 235,000 천원
총연구기간
참여연구원
수
총 : 28 명
내부 : 5 명
외부 : 23 명
총
연구개발
비
정부: 626,000 천원
기업: 천원
계 : 626,000 천원연구기관명
및
소속부서명
부산대학교 해양학과 참여기업명 해당사항 없음
국제공동연
구 상대국명 : 해당사항 없음 상대국연구기관명 : 해당사항 없음
위 탁 연 구 연구기관명 : 해당사항 없음 연구책임자 : 해당사항 없음
요약(연구개발결과를 중심으로 개조식 500자 이내)보고서
면수 305
연구의 목표는 낙동강 하구 갯벌 생태계의 구조와 기능을 파악하여 환경/저서생태계
/물질순환에 관련된 복원 및 관리 파라미터를 도출하는 것이다. 2004년부터 3년 동안
낙동강 하구 갯벌 퇴적물 및 수층 특성변화를 조사하였고, 생산자, 소비자, 분해자의
상대적인 중요성과 생태기능을 파악하였는데, 갈대, 세모고랭이, 잘피 등의 대형일차생
산자가 낙동강 생태계의 물질순환, 자정능력에서 핵심적인 역할을 함이 밝혀졌다. 안
정동위원소 추적자 실험 결과 저서미세조류는 소비자의 먹이로 중요하였다. 이 지역에
서 거의 사라진 잘피는 보호구역을 설정한다면 복원가능성이 높았다. 실험용 갯벌을
이용한 복원 실험에서 저서동물군집은 1년 정도 지난 후 자연 갯벌과 유사한 종 다양
성을 나타냈다. 이외에 실험용 갯벌의 유기물분해와 탈질소화 속도, 저서미세조류 종
조성 등은 6개월 이내에 자연 상태와 유사해졌다. 낙동강 하구에서 퇴적물은 육상기원
유기물을 제거 하는데 큰 역할을 하였으며, 대형일차생산자도 육상기원 물질의 저장
및 제거에 중요하였다.
색 인 어
(각 5개
이상)
한 글 낙동강 하구, 연안습지복원, 갯벌생태계, 잘피복원, 연안물질순환
영 어 Nakdong estuary, Coastal wet land, Tidal flat, Seagrass,
Material cycling

- 3 -
요 약 문 I
Ⅰ. 제 목 : 낙동강 하구역 습지 생태계 훼손지역 복원 및 관리기술
Ⅱ. 연구개발의 목적 및 필요성
낙동강하구 생태계는 하구 습지의 소실이 극심한 지역으로 철새를 비롯한 생물들이
서식처의 단편화, 생태계의 구조와 기능상의 변화를 겪고 있다. 이는 낙동강 하구만의
문제는 아니며, 시화호와 새만금 매립에서 보듯이 연안의 개발과 보존에 대한 갈등이
심각해지고 있으며, 국론이 분열되고 막대한 사회 비용이 치러지고 있다. 인구밀도가
높고, 해안을 따라 U-자형 개발이 이루어질 수밖에 없는 국내 현실에서 연안개발은
피할 수 없으며, 해안습지의 보전과 복원, 관리를 위한 기술과 기준을 마련하는 것이
시급하나, 국내의 연안 하구역 습지의 복원과 관리기술에 관한 지식 및 기술 수준은
일천하다.
본 연구의 목표는 낙동강 하구에 위치한 자연 상태의 연안 하구 습지 (갯벌 및 조
하대)의 물리 및 생태 환경 구조, 물질순환 구조, 건강한 습지를 유지하는 요인들을 파
악함과 동시에, 실험용 복원습지 (갯벌 및 조하대 잘피림)를 조성하여 안정화 과정을
살피며 자연습지와의 비교를 통하여 환경/저서생태계/물질순환에 관련된 복원 및 관리
파라미터를 도출하는 것이다.
Ⅲ. 연구개발의 내용 및 범위
- 낙동강 하구 습지의 환경특성 파악
건강한 연안 습지를 유지시키는 물리학적, 퇴적학적, 수질 요인을 찾아내고 이들 요인간의
인과관계를 구명하여 최적 습지 복원/관리 방안을 도출.
- 낙동강 하구 습지의 저서생태계 특성 파악
저서 생태계의 생물학적 구성요소 (1. 일차생산자: 미세조류, 잘피, 세모고랭이, 갈대
; 2: 소비자; 저서동물; 3: 분해자; 미생물)와 무생물학적 구성요소 (환경요인, 수질, 퇴
적물, 부유물, 유기물, 무기물 등)의 구조와 기능을 파악.
- 낙동강 하구 습지의 물질순환 특성 파악 및 복원/관리 파라미터 개발
서로 다른 환경 조건에서 먹이망을 통한 물질순환경로 및 갯벌의 자연 정화능력을
파악하여, 생태계 구성요소의 변동에 대한 전체 생태계의 반응을 예측하고, 연안 습지
복원 및 관리에 필요한 환경/생물학적 파라미터 개발.

- 4 -
요 약 문 II
Ⅳ. 연구개발결과
제 1절 낙동강 하구의 환경 특성1. 갯벌의 조위 및 퇴적물 특성
낙동강 하구지역은 하구둑에서 바깥쪽에 위치한 갯벌 지역으로서 평균 해수면 위
112.5-189.1cm 에 걸쳐 위치하였다. 낙동강 갯벌의 경사도는 갯벌의 발달이 적은 진우
도 인근에서 0.11~0.23 % 였고 갯벌이 잘 발달한 을숙도지역의 경우 0.016 % 로 낮았
다. 갯벌의 조석 분석결과 평균 해수면 위 70, 90, 110 cm 를 기준으로 노출 특성이
달라졌으며 이를 통해 노출지역이 구분되었다. 12시간이상의 연속노출이 나타나는 노
출 지역I은 서해 새만금에서 전체면적은 40 % 정도이나 낙동강 갯벌은 90 %이상으로
낙동강 갯벌은 연속노출이 많아 이에 적응한 생물이 서식할 것으로 예상된다. 퇴적물
타입은 전체적으로 모래 퇴적물이 90%이상 우세하였으며 일부지역 (명지주거단지 등)
실트-점토 퇴적물이 상대적으로 높았다. 표층퇴적물의 평균입도는 2.6 Φ~3.9 Φ 로
slightly muddy sand였다. 사질 퇴적물이 우세하여, 유기탄소(C) 와 유기질소(N)는
0~1.4 % 로 매우 낮게 나타났다. 강열감량으로 측정된 유기물 농도는 가덕도 지역의
퇴적물 내 유기물 농도는 평균 4.73 % 였고, 을숙도 지역은 평균 2.13 % 였다. 퇴적물
공극수내 암모니아 농도는 40-160 μM 이였고, 질산과 아질산 농도는 4 μM 이하였다
. 2. 낙동강 하구 수층 특성
낙동강 하구지역 수온은 계절변화를 잘 보였는데, 을숙도 지역 (4.5-31 ℃)이 가덕
도 지역(7.8-28.4 ℃)보다 수온 변동 폭이 컸다. 가덕도 지역의 염도는 바다의 영향으
로 평균 28.8 ppt, 을숙도는 담수의 영향으로 평균 13 ppt정도 였으며, 특히, 여름철 장
마로 인한 담수 유입의 증가는 염도의 급격한 감소를 유발하였다. 조사기간 동안 해수
내 엽록소의 양은 10 ㎍/L 내외였으나 2006년 5-8월까지 급격히 증가하여 최대 46 ㎍
/L 까지 관찰되었다. 가덕도 지역과 진우도 지역에서 부유물질농도는 뚜렷한 계절적인
양상을 보이지는 않았으나, 22-60 ㎎/L로 비교적 높은 값을 나타내었다. 수층의 영양
염 농도는 을숙도 지역이 가덕도 지역보다 높았는데, 특히 질산과 아질산의 농도가
매우 높았다 (평균 43 μM). 수중 광량은 뚜렷한 계절적인 변이를 나타내지 않았다 (일
일 최대 광량: 45 ㏖ m-2
d-1
, 최소 일일 최소 광량: 0.1 ㏖ m-2
d-1
). Light attenuation
coefficient는 봄에 낮은 값을 보였으며, 여름에 높은 값을 보였고 평균적으로 높은 수
치를 보였으며, 이런 현상은 부유물질 또는 부유생물에 의한 빛의 감소현상으로 추측
되어진다.

- 5 -
요 약 문 III
제 2절 낙동강 하구의 생태 구성 요소1. 일차 생산자
가. 갈대
을숙도 지역의 갈대는 4월부터 싹을 틔기 시작하여 7월에 거의 최고점에 도달, 10월부
터 시들기 시작하여 이듬해 1-2월에 거의 없어진다. 전체길이는 최고 230㎝에 달하며,
최대 생육밀도는 360 shoots/㎡ 이고 갈대의 전체 생체량은 생육밀도와 유사한 경향을 나
타내었으며, 가을철에 최고점(2004년-2,400g, 2005년-2160g, 2006년-3590g)에 도달하였다.
잎 조직 내 탄소 함량은 약 48% 이고 조직 내 질소 함량은 3 % 정도였다. 갈대 잎의
일일 순 생산량은 6.68-9.84g DW ㎡ day-1, 단위면적당 연간 순 생산량은 최고 3,590g
DW ㎡ year-1 이고, 낙동강 하구 전체에서 갈대가 생산하는 순 생산량은 약 4371t DW
year-1로 매우 높았다.
나. 세모고랭이
을숙도 지역의 세모고랭이의 전체길이는 최고 100 ㎝ 에 달하며 맹금머리도 지역은
최고 53 ㎝ 였다. 을숙도 지역의 세모고랭이는 맹금머리도 지역의 세모고랭이보다 한
달 정도 빠르게 가입되었고 한 달 먼저 시들어 사그러지는 것으로 관찰되었다. 세모고랭
이는 을숙도 지역이 맹금머리도 지역보다 더 높은 생체량을 보였다. 을숙도 지역의 세모
고랭이 잎 조직 내 탄소 함량은 평균 42 % 이였고, 맹금머리도 지역은 평균 39 % 로
약간 낮았다. 세모고랭이 잎의 일일 순 생산량은 을숙도 지역을 기준으로 0.97-1.03 g
DW ㎡ day-1이였으며, 단위면적당 연간 순 생산량은 최고 376 g DW ㎡ year
-1이였다.
또한 낙동강 지역에서 세모고랭이가 생산하는 순 생산량은 약 1835 t DW year-1였다.
다. 잘피
거머리말(Zostera marina)의 형태학적 특성은 계절 변이를 나타냈는데, 잎 조직 내 엽
록소의 함량은 4.9-11 mg DW g-1 였고, 단위면적당(㎡) 생육 밀도는 가덕도 70-270, 진
우도 40-220 개체 정도 였다. 생체량은 가덕도 지역(지상부: 40-32 g, 지하부: 12-164 g,
전체: 49-484 g)이 진우도 지역(지상부: 13-94 g, 지하부: 5-32 g, 전체- 19-116 g)보다
훨씬 높은 값을 나타내었다. 거머리말 잎의 생산성은 가덕도 지역(6.7-45.8 ㎎ DW sht-1
d-1
, 0.77-11.27 g DW m-2d
-1 이 진우도 지역(7.2-38.6 ㎎ DW sht-1 d
-1, 0.34-7.16 g DW
m-2
d-1)보다 높았다. 거머리말의 잎 질소 함량은 겨울철에 높고 여름철에 낮은 뚜렷한 계
절적 양상을 보였으며 (2.13-4.07 %), 탄소 함량은 35% 정도 였다. 거머리말 잎의 일일
순 생산량은 가덕도 지역을 기준으로 2.80-4.72 g DW ㎡ day-1이였으며, 연구 대상 종
중 갈대 다음으로 높았다. 단위면적당 연간 순 생산량은 1,019-1,722 g DW ㎡ year-1, 거
머리말이 낙동강 지역에서 생산하는 순 생산량은 약 923 t DW year-1이였다.

- 6 -
요 약 문 IV
라. 저서미세규조류 (MPB; micro phyto benthos)
낙동강 명지지역 저서미세조류 생물량(평균 클로로필; 23.2 ㎍‧ ㎝-3
)은 국내 타지역
갯벌과 유사한 수준이었으며, 계절에 따라 그 변화가 심하였는데, 겨울철-초봄에 특히
높은 값을 보였다. 연변화가 계절변화보다 적었고, 퇴적상에 따라 생물량의 차이가 컸
다. 낙동강 명지지역 저서미세조류는 총 35종이 출현하였다. 계절 및 퇴적상에 상관없
이 우점하는 종은 약 3종으로 확인되었다 (Amphora coffeaeformis, Navicula
digitoconvergence, Navicula sp.2). 지역적 차이를 보면 진우도, 장자도 정점에서 상
대적으로 엽록소 a 농도가 높았으며, 백합등 정점에서는 낮았다.
2. 소비자
가. 조간대 갯벌 저서동물의 군집구조
총 7개 정선의 28개 정점에서 출현한 저서동물은 64종/5.6㎡, 14,230개체/28㎡와
3,335.03gWWt/28㎡이었다. 분류군별 출현종 수를 보면, 절지동물이 32종으로 전체의
50%를 점유하였고, 연체동물과 환형동물의 다모류는 각각 20종, 31.25%와 11종,
17.19%를 차지하였다. 개체수는 다모류가 6,008개체가 채집되어 42.22%를 점유하여 가
장 우점 하였다. 다음으로 절지동물과 연체동물이 각각 4,666개체, 32.79%와 3,530개체,
24.81%를 차지하였고, 기타동물군은 26개체로 0.18%를 점유하였다. 갯벌에 서식하는
저서동물 군집의 다양도와 균등도를 보면, 총 28개 정점에서 다양도는 0.05~2.42의 범
위에 평균 1.39를 나타내 다양도가 비교적 낮은 해역으로 판단할 수 있었다. 반면 균등
도는 최소 0.08에서 최대 0.82를 나타내었고, 평균값은 0.59이었다.
나. 낙동강 하구역 조하대 저서동물의 군집구조
총 36개 정점에서 출현한 저서동물은 145종/7.2㎡, 35,554개체/36㎡와 2,036
gWWt/36㎡ 이었다. 분류군별 출현종 수를 보면, 다모류가 65종으로 44.83%를 점유하
였고, 연체동물과 절지동물이 각각 38종, 26.21%와 37종, 25.52%를 차지하였다. 개체수
는 출현종 수와 동일하게 다모류가 28,444개체가 채집되어 전체의 80%를 점유하여 가
장 우점하였다. 다음으로 절지동물과 연체동물이 각각 4,220개체, 11.87%와 2,810개체,
7.9%를 차지하였고, 기타동물군은 80개체로 0.23%의 낮은 점유율을 보였다. 생체량은
단일개체의 생체량이 높은 연체동물이 1,795.2gWWt으로 88.17%의 높은 점유율을 보
였다. 다음으로 다모류는 185.09gWWt으로 9.09%를, 절지동물과 기타동물군은 각각
39.17gWWt, 1.92%와 16.56gWWt, 0.46%로 생체량이 낮았다.

- 7 -
요 약 문 V
제 3절 낙동강 하구의 생태 기능 요소1. 생태 과정 (ecological processes)
가. 저서미세조류 (MPB) 의 일차생산
낙동강 명지지역 저서미세조류 일차생산량은 국내 타지역 갯벌의 일차생산량과 유
사한 수준이었는데, 평균 15 mmole m-2
d-1 정도였다. 여름에 비해 겨울철 생산량이 높
았는데 이는 겨울철 높은 저서미세조류 현존량을 반영하는 것이다.
나. 유기물 분해 속도의 변화
2005년 실험용 갯벌의 SOD 값의 범위는 -0.7 ~ 35.3 mmole m-2
d-1 였고, 하절기에
가장 높은 SOD 값(mean: 30 mmole m-2
d-1 )을 보였으며, 동절기에는 산소가 소비되
기 보다는 광합성에 의해 산소가 생성되는 것이 관찰되었다(mean :-0.5 mmole m-2
d-1
). 2006년 인공 갯벌의 SOD 값의 범위는 -0.5 ~ 38.2 mmole m-2
d-1 였고, 2005년 인
공 갯벌과 유사하게 하절기 (mean :37mmole • mmole m-2
d-1 ), 동절기 (mean
:0.1mmole m-2
d-1 )의 SOD 값을 나타냈다. 이는 미생물 활동을 조절하는 큰 요인은
계절적인 수온 변화임을 시사한다.
다. 탈질소화의 계절변화
2005년 실험용 갯벌의 탈질소화 값의 범위는 평균 5.9 ~ 2652 μmole m-2
d-1 였고, 하
절기에 높았으며(4564.6 μmole m-2
d-1
) 동절기에는 거의 탈질소화가 발생하지 않았다
(1.6 μmole m-2
d-1
). 2006년 인공 갯벌의 탈질소화 값의 범위는 3.9 ~ 2082 μmole
m-2
d-1 였고, 2005년 인공 갯벌과 유사하게 하절기 (2626.5 μmole m
-2d
-1), 동절기 (0.01
μmole m-2
d-1
)의 탈질소화 값을 나타냈다. 탈질소화 역시 SOD와 마찬가지로 미생물
활동을 조절하는 큰 요인은 계절적인 수온 변화였다.
라. 각 과정들 사이의 상관관계
유기물분해속도 (SOD)와 암모니움 플럭스는 양의 상관관계를 보였다. 여름철 강한
유기물 분해에 의해 퇴적물내 NH4+이 증가함에 따라 퇴적물로부터 용출되는 결과가
나타났다. 질산 영양염 (NO2-+NO3
-) flux는 퇴적층으로 소비되는 경향이 나타났다.
이는 낙동강 하구로 유입되는 강물의 NO2-+NO3
-의 농도가 매우 높음을 나타내는 것이
며, NO2-+NO3
- 를 전자 수용체로 이용하는 혐기성 유기물 분해과정인 탈질소화가 잘
일어남을 시사하는 것이다. 탈질소화도 SOD와 양의 상관관계를 보였다.

- 8 -
요 약 문 VI
2 먹이망 분석
가 갈대(Phragmites australis) 생태계 먹이
갈대 서식지의 주요 기초생산자와 소비자 동물군의 기여를 모식화한 결과 여과섭식
자인 두 종의 이매패류Laternula marilina 와 Corbicula japonica 는 식물플랑크톤의
기여가 51-55, 70-74%로 각각 높았고 저서미세조류의 기여가 높은 종으로는 I lyoplax
pusila가 99-100%가 가장 높았고, Amphipods가 81-95%, Assiminea japonica가
82-88%로 상대적으로 높은 기여를 나타내었다. 세모고랭이와 갈대서식지에서 얻어진
소비자 동물군들의 탄소 및 질소 안정동위원소 비값들은 낙동강 하구역에서 대단히 넓
은 서식지와 그에 따른 높은 생물량을 이들 염습지 식물군들이 가지지만 이들이 하구
역에 서식하고 있는 동물들에게 거의 이용되지 못한다는 것을 시사해 주었다.
나 갯벌생태계와 조하대
갯벌과 조하대에 서식하는 저서생물 중 Batillaria cimingii 는 탄소 및 질소 안정동
위원소 비 값은 각각 -9.1 ± 0.5, 14.9 ± 0.6 ‰, Bullacta exarata 탄소 및 질소 안정동
위원소 비 값은 각각 -7.9 ± 1.1, 13.7 ± 0.7 ‰로 매우 낮은 탄소안정동위원소비 값을
가지는 제 3의 source의 기여가 상당히 높음을 알 수 있다. 낙동강 하구역에 서식하는
어류의 탄소안정동위원소와 질소안정동위원소의 비 값을 보면, 탄소안정동위원소의 평
균값은 -18.5 ± 2.9‰이고, 질소안정동위원소의 경우 평균 -14.5 ± 2.5‰이었다. 낙동강
하구역에 서식하는 문절망둑의 경우 높은 평균 14.5‰의 탄소안정동위원소를 보여 하
구역에서 생산되는 자체유기물에 상당히 의존함을 알 수 있으나, 대부분의 어류는 식
물플랑크톤과 저서미세조류 같은 조류기원의 유기물을 섭식함을 알 수 있다.
3. 물질수지
낙동강 하구에서는 물질 유입이 집중되는 7~9월동안 하구 퇴적물에서 일어나는 유기
물 분해과정도 최댓값을 보이며, 세모고랭이와 갈대와 같은 대형일차생산자의 생체량
도 최댓값을 보이고 있다. 우기동안 낙동강을 통해 하구로 유입된 유기물은 일간 평방
미터당 평균적으로 237 mmole-C 정도였는데, 갈대가 평방미터당 하루에 320
mmole-C, 세모고랭이가 32 mmole-C, 잘피가 35 mmole-C, MPB가 15 mmole-C 정도
의 생산성을 보여 유사한 범위에 있었다. 이외에 퇴적물 미생물에 의한 유기물 분해
량은 평방미터당 하루에 25 mmole-C, 탈질소화로 인한 탄소제거 효과는 40 mmole-C
에 달한다.

- 9 -
요 약 문 VII
제 4절. 낙동강 하구의 생태 구성 요소의 복원1. 잘피
진우도 부근에서 측정된 수중광량은 잘피가 생육하는데 충분한 것으로 판단된다. 2004
년 11월 16일과 2005월 12월 16일에 수행된 두 번의 이식 실험은 이식 개체의 생존율이
50%이상으로 성공적이었으나, 어민들에 의한 배의 운항으로 인해 상당부분 피해가 발생
하였다. 따라서 이 지역에서 인공 잘피림 조성은 어민들의 협조가 절대적이며, 이식 개
체가 이식 지역에 안정화 될 때까지 보호구역을 설정할 필요성이 대두되었다. 이식한 지
한 달 후, 인공 잘피림의 개체는 원생육지에 적응한 생리적 특성을 잃어버리고 이식 장
소의 환경적 특성에 잘 적응하고 있었다.
2 세모고랭이
2006년 8월 중갯벌의 mixed 퇴적물에서 111개체/m2으로 가장 높은 세모고랭이 밀도가
나타났으며 sand 퇴적물에서, 26개체/m2로 낮았고, mud 퇴적물에서는 전혀 나타나지 않
았다. 중갯벌의 조고는 평균 해수면 위 142.9 cm 에 위치하고 있는데, 이 위치에서는
6~12시간 연속노출이 전체 노출에 50%정도를 차지하며, 48시간 이상의 연속노출이 25%,
그리고 12~24시간 연속노출이 15%정도 나타나는 지역이다 세모고랭이가 가장 높게 나타
난 mixed 퇴적물은 니질 비율이 24~33 %정도였으며, 전혀 나타나지 않은 mud 퇴적물의
경우 니질 비율이 57~61 %였다.
3. 실험용 갯벌의 환경과 기능변화
가. 퇴적물 특성변화
실험용 갯벌 내 퇴적물 내 유기물의 함량은 0.7∼17.4% 내외였다. 시간에 따라 표층
퇴적물은 세립화되었고, 각 실험용 갯벌에서 표층입도가 유사해 지는 경향을 보였다. 조
고가 가장 높은 upper station은 실험용 갯벌 조성 당시의 퇴적물 특성을 그대로 유지하
였으나, lower station의 경우는 유기물 함량이 높아졌다. 유기물 농도와 마찬가지로 퇴
적물 입도 분석에서도 실험용 갯벌 조성 이후 세립한 퇴적물이 쌓임을 알 수 있었다.
나. SOD를 통해본 유기물 분해 속도의 변화
조고에 따른 SOD의 값을 보면 2005년 station의 경우 중갯벌 가장 높은 SOD(15.7
mmoleㆍm-1
d-1
)를 보였고, 상(12.9 mmoleㆍm-1
d-1
), 하(12.8 mmole m-1
d-1
)의 순서였다.
퇴적물 입자 차이에 따른 SOD의 값을 보면 2005년 station의 경우 mixed에서 가장 높
은 SOD(14.9 mmole m-1
d-1
)를 보였고, 사질(SAND; 13.9 mmole m-1
d-1
), 니질(MUD;
12.6 mmole m-1
d-1
)의 순서로 낮아졌다. 퇴적물내 유기물 농도(I.L.)와 SOD는 크게 상관
관계가 나타나지 않았다.

- 10 -
요 약 문 VIII
다. 탈질소화의 변화
조고에 따른 탈질소화는 05년 station의 경우 하갯벌에서 가장 높은 탈질소화(949.2 μ
mole m-1
d-1
)를 보였고, 중(932.7 μmole m-1d
-11), 상(438.2 μmole m
-1d
-11)의 순서였다. 퇴
적물 차이에 따라서는 니질(921 μmole m-1
d-11)에서 높은 탈질소화를 보였고, 사질(743.5
μmole m-1d
-11), mixed퇴적물(655.8 μmole m-1
d-1
1) 순서로 낮아졌다.
라. 실험용 갯벌의 저서동물 군집 변화
조사 기간 중 출현한 저서동물의 종수는 1년정도 안정화된 05년 갯벌이 22~23종으로
06년 갯벌의 12~22종보다 높은 경향을 보였다. 출현 개체수에 있어서도 05년 갯벌의 경우
8400~8800 개체 m-2 으로 06년 갯벌의 평균 출현개체수 (~4500 개체 m
-2) 보다 높았다.
05년도 갯벌에서 종수와 서식밀도의 차이를 살펴보면, 2006년 4월과 9월 모두 상갯벌(U)
에서 낮은 종수를 보였고, 하갯벌 (L)에서 출현종수가 높은 경향이 나타났다.
제 5절 낙동강 하구의 자정 능력1. 인공 갯벌의 유기물 분해와 탈질소화를 통한 유기물 제거
낙동강 하구에서 퇴적물에 의한 유기물 분해는 탄소량으로 년간 평균 7.1 x 106 ton
이 된다. 이는 년간 낙동강 하구로 유입되는 유기물 총량인 38.3 x 106 ton (화학적산소
요구량 COD에서 환산)의 약 19%에 해당하는 값이다. 탈질소화에 의해 제거되는 탄소량
은 낙동강 전체에서 3.9 x 106 ton이 된다. 이는 COD 유입량의 약 10%에 달하는 양이
다.
2. 잘피에 의한 영양염 제거
낙동강 하구지역에 생육하고 있는 잘피 지상부는 암모니아 10μM의 해수를 연간 1억
8900만톤을 정화할 수 있는 능력을 가지고 있었다.
3. 세모고랭이에 의한 영양염 저장
세모고랭이의 성장이 활발한 시기에 퇴적물 공극수 내 암모니아 농도는 감소하여, 세
모고랭이는 낙동강 지역에서 영양염 조절에 기여하는 것으로 추측된다. 낙동강 하구지역
에서 세모고랭이 지상부의 생산성은 연간 1890 ton DW, 탄소 저장량은 연간 752 ton
DW이고 질소 저장량은 53 ton DW 이었다.
4. 갈대에 의한 영양염 저장
낙동강 하구지역에서 가장 높은 생산량을 나타내는 갈대는 연간 4371 ton DW, 탄소
저장량은 연간 1740 ton DW이고 질소 저장량은 122 ton DW 정도 였으며, 낙동강 하구
지역의 영양염류 순환에 중요한 역할을 할 것으로 판단되어진다.

- 11 -
요 약 문 IX
Ⅴ. 연구개발결과의 활용계획
본 연구 결과는 최근에 정부나 민간에서 습지복원/관리가 주요 이슈로 주목받는 상
황에서 다음과 같은 분야에 활용될 수 있다.
대형 복원공사에서 각 지역의 여건에 맞는 복원 목표를 설정, 경제적인 시공 및 관
리 기법을 선택.
- 해양환경의 기준설정이나 해양환경 영향평가에 구체적인 평가 자료를 제공.
- 연안 수산자원관리 조성사업에 대한 정책설정의 기본적인 방향을 자연과학적 측면
에서 평가할 수 있는 틀 제시.
-현재 대형 연안개발에서 나타나는 논쟁에서 “개발” 혹은 “보존” 이외의 제 3의 방
안으로 복원이 검토되어 막대한 사회비용을 절감할 수며 복원 산업이 활성화.
-조하대 잘피림을 조성하면 1) 수산어족자원 및 해양생물의 다양성 보존 2) 짧은 기
간 내 환경친화적으로 수산생산성 향상을 꾀할 수 있음. 3) 조성된 잘피밭에 의한 무
기영양염류 및 오염물질의 흡수제거에 따른 연안 생태계 정화 등에 기여.
- 연안생태계의 물질순환 모델을 통하여 얻어지는 결과들로부터 인위적 연안환경의
변동이 연안생태계의 생물다양성과 생물생산 특히 수산자원생물의 생산 변동에 미치는
직접적인 효과에 대한 구체적인 정보를 제공.
- 연안생태계의 물질순환 모델을 통한 각 연안생태계 구성요소들의 역할에 대한 정
확한 평가를 바탕으로 합리적인 연안역의 보호와 관리를 위한 해양환경 보전정책수립.

- 12 -
S U M M A R Y I
I. Title: Restoration and management plan for a degraded wetland ecosystem in the Nak-Dong estuary
II Purpose of the study and necessity
The Nak-dong estuary is suffering from habitat loss and both structural and
functional changes of the ecosystem are occurring. The problem is pertinent not
only to the Nak-dong estuary, but also to other estuaries in Korea such as the
Saamangeum and Shihwa reclamation areas. Coastal development is unavoidable in
Korea due to the high population density and large available coastal line. However,
although management and restoration plans are necessary for coastal wetlands the
technology level relating to the area at present under-developed. The purpose of
this study is to identify both the physical and environmental characteristics of the
Nak-dong estuarine ecosystem and the associated ecosystem structure and
functioning. Material cycling patterns and factors contributing to the maintenance
of healthy wet land ecosystems were determined. Experimental wetlands were built
and the temporal change in the environment and biota such as microphytobenthos
and macro benthos was studied.
III Content and scope
The environmental characteristics of the Nak-dong estuaries ecosystem were
monitored. The physical features, water quality and depositional factors responsible
for the maintenance of healthy wet land were identified and interactions between
the factors studied. Benthic ecosystem characteristics of the Nak-dong estuaries
ecosystem were also recorded. The structure and functioning of biological (primary
producers, consumers, decomposers) and abiotic components (environmental factors,
water quality, sediment, suspended matter, organic matter, inorganic matter) of the
ecosystem were studied. Material cycling patterns and the temporal development of
restoration parameters observed. The natural purification capacity of the tidal flat
sediment was identified and the interactions between ecological components were
studied through food web analysis. Skills were developed which involved the
identification of environmental/biological parameters necessary for the restoration of
wetland ecosystems.

- 13 -
S U M M A R Y II
IV. Results
Section 1. Characteristics of the Nak-dong estuary
1. Sediment characteristics and tidal height of the tidal flat.
The Nak-dong tidal flat is located outside the Nak-dong estuary dam. The
height of the tidal flat is approximately 112.5-189.1 cm above mean sea level. The
tidal height of the Nak-dong can be considered to be between the height of the
Kimje tidal flat (-50~100 cm) located in the sea outside the Semankum and the
tidal flat (150~280 cm) nearer the river.
The slope of the Nak-dong tidal flat is low at 0.11~0.23 % near Jinwoo Island
and 0.016 % at the Eulsuk Island region. The degree of atmospheric exposure of
certain levels of the Nak-dong tidalflat was used to characterize the exposure
regime of the system. The continuous exposure range were divided into the tidal
heights of 70, 90, 110 cm above sea level. Approximately 40 % of the whole
Semankum area is subject to complete exposure over a continuous 12-hour period.
But in the Nak-dong tidal area this is over 90 %, and it is predicted that the
organisms present have adapted to current environmental conditions resulting from
the long continual exposure.
Sandy sediment is the dominant sediment type in the area representing over 90%
of the current area. In one specific area (Myungji), silty-mud is relatively
dominant. As a result of the low mean particle size (2.6 Φ-3.9 Φ) of the surface
sediment it can be classed as slightly muddy sand. As a result of being composed
of mainly sandy sediment, the values of organic carbon and organic nitrogen are
extremely low (0-1.4 %). Concentrations of organic matter measured in the
sediment from the Gadeok Island area and the Eulsuk Island area by ignition
experiments were on average 4.73 % and 4.73 % respectively. Concentration of
ammonia in the sediment pore water was between 40-160 μM, while the
concentrations of nitrate and nitrite were 4 μM.

- 14 -
S U M M A R Y III
2. Water characteristics of the Nak-dong estuary
The temperature of the Nak-dong estuary varies seasonally. The temperature
variation at Eulsuk Island (4.5-31 ℃) is however higher than at Gadeok Island
(7.8-28.4 ℃). The salinity of overlying waters at the Gadeok Island site is on
average 28.8 PSU, while the salinity at Eulsuk Island is on average 13 PSU. The
increase in freshwater inflow in the summer rainy season causes the decrease in
salinity seen in Eulsuk Island.
The typical concentration of Chl-a during the research period was about 10 ㎍/L
but a sudden increase was observed from May to August 2006 (maximum of 46 ㎍
/L). The concentration of suspended matter in Gadeok Island and Jinwoo Island
showed no obvious seasonal variation, values were however relatively high and
ranged between 22-60 ㎎/L.
Nutrient concentrations at the Eulsuk Island site were higher than at the Gadeok
Island site. Nitrate and nitrite were especially high (average: 43 μM). Seasonal
variations in the intensity of radiation in the water column were not evidenced (the
daytime maximum intensity was 45 ㏖ m-2d-1, while the daytime minimum
intensity was 0.1 ㏖ m-2d-1). The light attenuation coefficient was low in spring
and high in summer. The reason behind this phenomenon is suspected to be the
light decrease caused by suspended matter and increased organisms in the water.
Section 2. Structure of ecological components of the Nak-dong estuarine ecosystem
1. Primary producers
Reed beds
New reed shoots can be observed from April on Eulsuk Island, these reach a
maximum in July, and begin to wither from October, disappearing completely
between the January and February periods. The length of shoots can reach a
maximum of 230㎝, while the maximum density was observed to be 360 shoots/㎡.
The temporal evolution of the total biomass of reeds is similar to that of the
density, showing maximum values in fall (2400 g in 2004, 2160 g in 2005, and 3590
g in 2006). The carbon and nitrogen content in the leaf tissue is approximately 48
% and 3 % respectively. The net reed production per unit area per day is 6.68-9.84
gDW㎡day-1
.The reed net production per unit area per year is 3,590 g DW㎡year-1
.

- 15 -
S U M M A R Y IV
The annual net production is extremely high at approximately 4371 tDWyear-1
Scripus trgueter
The length of Scripus trgueter was measured to be maximum on Eulsuk Island
(100 ㎝) and was lower on Maenggeummury Island (53 ㎝). However it should be
noted that Scripus trgueter on Eulsuk Island sprout and wither a month earlier
than populations observed on Maenggeummury Island.
The biomass of Scripus trgueter on Eulsuk Island was higher than on
Maenggeummury Island. The average carbon content of leaf tissue was 42 % on
Eulsuk Island, and 39 % on Maenggeummury Island. Gross leaf production of
Scripus trgueter per unit area per day was 0.97-1.03 g DW ㎡ day-1
, and gross
production per unit area per year was maximum on Eulsuk Island at 376 g DW ㎡
year-1
. Gross production was estimated at approximately 1835 t DW year-1 in the
Nak-dong estuary.
Seagrass
The morphological characteristics of Zostera marina were found to have seasonal
variations, Chl-a content in the leaf tissues was 4.9 - 11 mg DW g-1
, rearing
densities per unit area (m3) was 70-270 on Gadeok Island and 40-220 on Jinwoo
Island. The biomass in Gadeok Island (ground: 40-32 g, underground: 12-164 g,
whole: 19-116 g) was higher than was on Jinwwo Island (ground: 13-94 g,
underground: 5-32 g, whole: 19-116 g). Production of Zostera marina leaves in
Gadeok Island (6.7-45.8 ㎎ DW sht-1d-1, 0.77-11.27 g DW m-2
d-1
) was estimated
to be higher than in Jinwoo Island (7.2-38.6 ㎎ DW sht-1
d-1
, 0.34-7.16 g DW
m-2
d-1
).
The nitrogen content of Zostera marina leaves has a clear seasonal variation
with higher values measured in winter and a lower content observed in summer
(2.13-4.07 %); the carbon content was approximately 35 %. Net production of
Zostera marina leaves per day was 2.80-4.72 g DW ㎡day-1 in Gadeok Island, this
was higher then other reed species researched. Net production per unit area per
year was 1,019-1,722 g DW ㎡ year-1
, net production of Zostera marina in the
Nak-dong estuary was about 923 t DW year-1
.
MPB; Microphytobenthos (MPB)
The biomass of MPB in Myungji within the Nak-dong estuary (average Chl;
23.2 ㎍‧ ?-3) was similar to other tidal flats in Korea. The seasonal variation in

- 16 -
S U M M A R Y V
biomass was large and was especially high from winter to early spring. The
annual variation was lower than the seasonal variation; dissimilarity in the biomass
with sediment type was high. MPB biomass tended to decrease with sediment
depth, the MPB biomass at 1 cm below the surface was over 30-50 % of the
entire biomass. The specific number of MPB species of Myungji in Nak-dong
estuary was 35 in total. Irrespective of season and sediment type, the dominant
species were Amphora coffeaeformis, Navicula digitoconvergence, and Navicula
sp.2. An annual variation in specific composition and the dominant species was in
evidence. The Chl-a concentration was relatively high in JInwoo Island and Jangja
Island, but it was low in Baekhapdeung.
Amphora coffeaformis and Navicula sp. were confirmed to be in abundance in
JInwoo Island and Jangja Island, Amphora coffeaformis was in abundance and other
species such as Navicula were found to be distributed in large quantities. In
Baekhabdeung the seasonal specific variation was the highest.
2. Consumers
Community structure of benthos in the intertidal flat
The benthos was sampled along 7 transect lines each encompassing 4 stations,
making up a total of 28 stations sampled and showed an overall result of 64
species/5.6㎡, 14,230 individuals/28㎡ and 3,335.03gWWt/28㎡. According to the
specific number of groups, 32 species of Arthropods were present and made up
50% of the total, 20 species of Mollusca and 11 species of Polychaete were
observed, making up 31.25 % and 17.19 %, respectively. In individual numbers,
6,008 individuals of Polychaete were found and made up 42.22% of the total fauna
and hence were extremely dominant. The overall number of Arthropods and
Mollusca were 4,666 and 3,530 individuals, making up 32.79 % and 24.81 %
respectively. The overall diversity in the 28 stations was on average 1.39 in the
range of 0.05~2.42, it was estimated that diversity was relatively low in the
region. On the other hand species diversity was from minimum 0.08 to maximum
0.82, and was 0.59 on average.
Community structure of benthos in subtidal of Nakdong estuary
In a total of 36 stations sampled the benthos was found to contain 145
species/7.2㎡, 35,554 individuals/36㎡ and 2,036 gWWt/36㎡. According to the
specific number of groups, 65 Polychaete species were found, and were dominant at

- 17 -
S U M M A R Y VI
42.22 %, 38 and 37 species Mollusca and Arthropod were found, making up 26.21
% and 25.52 % respectively. Individuals were similar in specific number, 28,444
Polychaete individuals were counted, and dominated the benthos at 80 % of the
total fauna. There were 4,220 and 2,810 Arthropod and Mollusca individuals
present, making up 11.87 % and 7.9 %, while other groups were found in low
quantities of solely 80 species, and thus 0.23 %. In terms of biomass, the Moluska
was highly dominant with 1,795.2 gWWt, and made up 88.17 % of the overall
biomass present. Polychaete amounted to 185.09 gWWt and therefore 9.09 %, while
Anthropod and others had a low biomass at 39.17 gWWt, 1.92% and 16.56 gWWt,
0.46 % respectively.
Section 3. Functions of ecological components of the Nak-dong estuarine ecosystem
1. Ecological processes
Primary production of MPB
Primary production of MPB at the Myungji in the Nak-dong was similar to
other tidal flats in Korea, with an average of 15 mmole m-2
d-1
. Production in
winter was higher than in summer, this reflects the higher biomass of MPB found
in winter.
Organic matter remineralization
In 2005 SOD (sediment oxygen demand) value' s of the artificial tidal flat ranged
from -0.7 to 35.3 mmole m-2
d-1
, the mean maximum SOD value was: 30 mmole
m-2
d-1 in summer, more oxygen was created by photosynthesis than oxygen was
consumed in winter (mean:-0.5 mmole m-2
d-1
). In 2006 the SOD value's of the
artificial tidal flat ranged from -0.5 to 38.2 mmole m-2
d-1
, SOD values in winter
were on average 0.1 mmole m-2
d-1 and in summer were 37 mmole m
-2d
-1 and were
similar to the results found for the artificial tidal flat in 2005. The main controlling
factor for the variations of SOD seems to be the seasonal temperature change.
Seasonal change in denitrification
Denitrification ranges from 5.9 to 2652 μmole m-2
d-1 in the artificial tidal flat in
2005, and was highest in summer (4564.6 μmole m-2
d-1
) and was barely present in
winter (1.6 μmole m-2
d-1
). The mean denitrification value's ranged from 3.9 to 2082
μmole m-2
d-1 in artificial tidal flat in 2006, and was 2626.5 μmole m
-2d
-1 in summer
and 0.01 μmole m-2
d-1 in winter, similar to values in 2005. As with the μmole m-2
d-1

- 18 -
S U M M A R Y VII
in artificial tidal flat in 2006, and was 2626.5 μmole m-2
d-1 in summer and 0.01 μ
mole m-2
d-1 in winter, similar to values in 2005. As with the SOD, the main factor
regulating denitirification activity was the seasonal change of temperature.
Relationships between each process
The organism matter remineralization activity (SOD) and ammonium (NH4+) flux
showed a linear relationship. NH4+ concentration in sediment is increased by the
large SOD in summer, resulting in high efflux from the sediment. The nitrate
(NO2-+NO3
-) flux mostly showed consumption by the sediment (flux into the
sediment). This fact suggest that the concentration of NO2-+NO3
- in fresh water
flowing into the Nak-dong estuary is high compared to the porewater concentration
and that the denitrification process is responsible for the consumption in the
sediment. The SOD and denitrification also showed a linear relationship.
2. Food web analysis
Phragmites australis as an ecosystem food
The contribution of phytoplankton to the food source of two shellfish species,
Laternula marilina and Corbicula japonica was high at 51-55 % and 70-74 %,
respectively. The contribution of MPB to the I lyoplax pusila diet was the highest
at 99-100 % and was relatively high for the Amphipods and also Assiminea
japonica at 81-95 % and 82-88 %, respectively. The relative contribution of
Phragmites australis to the diet of Erimacrus isenbeckii was 66-69 % and the
relative contribution of Phragmites australis to the diet Tylorrhychus heterochaeus
was 43-44 %. Therefore, consumers were inclined to use Phragmites australis as a
food source. Although salt marsh plants have very wide habitats and high biomass
in the Phragmites australis and Scirpus triqueter habitats, according to the carbon
and nitrogen stable isotope value of consumers in the Nak-dong estuary, salt
marsh plants are not of dietary use to the animals in the estuary.
Tidal flat ecosystem and subtidal zone
In the benthos of the tidal flat and the subtidal zone, Batillaria cimingii carbon
and nitrogen stable isotope ratios are -9.1 ± 0.5, 14.9 ± 0.6 ‰ respectively.
Bullacta exarata carbon and nitrogen stable isotope ratios are -7.9 ± 1.1, 13.7 ± 0.7
‰ respectively. According to this value, a third source of food with a very low
carbon stable isotope ratio contributes significantly to their diet. The two species
of shellfish mainly use phytoplankton organic matter that constitutes the particle

- 19 -
S U M M A R Y VIII
organic matter in the estuary. However, Calliansa japonica carbon and nitrogen
stable isotope ratios are -15.2 ± 2.4, 12.2 ± 0.7 ‰ respectively which suggests that
they are highly depend on microphytobenthos. In the Nak-dong estuary, fish
carbon and nitrogen stable isotope ratios are -18.5 ± 2.9 ‰ and -14.5 ± 2.5 ‰
respectively. In the Nak-dong estuary, the carbon stable isotope of Acanthogobius
flavimanus is 14.5 ‰. This is very high value suggesting that Acanthogobius
flavimanus depend highly on the organic matter contained in the Nak-dong estuary.
However, fish mostly eat organic matter of algae source such as phytoplankton
and microphytobenthos.
3. Material budget
In the sediment of the Nak-dong estuary, organic matter remineralization
processes was high from July to September (rainy season). The biomass of
Phragmites australis and Scirpus triqueter are also the highest at this period.
During the wet season, organic matter input to the Nak-dong estuary is on
average 237 mmole-C/m2/day. Production of Phragmites australis is 320
mmole-C/m2/day, Scirpus triqueter is 32 mmole-C/m
2/day, Zostera marina is 35
mmole-C/m2/day and MPB is 15 mmole-C/m
2/day. In the sediment the
decomposition of organic matter by bacteria is 25 mmole-C/m2/day, while carbon
removal as a result of denitrification is 40 mmole-C m2/day.
Section 4. The restoration of the ecological components in the Nak-dong estuary
1. Zostera M arina
Near Jinwoo Island, the measured intensity of radiation in the water column is
sufficient for the growth of Zostera marina. The water temperature is also suitable
for the growth of Zostera marina.. Transplantation tests were carried out twice, on
the 16 of November 2004 and the 16 of December 2005. These tests were
successful with the viability of transplanted plants being over 50%. However
damage did occur due to the passage of small fishing boats. To ensure the
maintenance of artificial Zostera marina. beds the collaboration of local fishermen
and the creation of a reserve in this area would be necessary. After a one month
period, artificial Zostera marina. beds are well adapted to the transplantation area.

- 20 -
S U M M A R Y IX
2 Scirpus triqueter
At the artificial tidal flat, Scirpus triqueter' s density was the highest on the
middle tidal flat which contained a mixed sediment type in August 2006 (111
individual/m2). The sandy sediment contained a low concentration of individuals (26
individual/m2) while in the muddy sediment no individuals were detected. As the
middle tidal flat height is located on average 142.9 cm above sea level, the "6~12
hour continuous exposure" is over 50 %, and 25 % of the tidal flat has "48 hour
continuous exposure", while 15 % has "12~24 hour continuous exposure". In the
mixed type sediment, the silt & clay content ranges between 24~33 %, in the mud
type sediment, the silt & clay content ranges between 57~61 %.
3. Environmental and functional change of the artificial tidal flat
Changes in the sediment characteristics
In the artificial tidal flat, the organic matter content is 0.7 ∼ 17.4% of the
sediment. The surface sediments have become finer over time and the surface
sediments of each treatment in the artificial tidal flats are becoming more similar
over time. The sediment grain composition in the upper tidal flat remained similar
to the initial conditions set, however that of the lower tidal flat become finer with
time.
Change of the organic matter reminrealization activity
The SOD varied according to the height of the tidal flat with the middle tidal
flat having the highest value (15.7 mmole m-1
d-1
), followed by the upper (12.9
mmole m-1
d-1
), and lower (12.8 mmole m-1
d-1
) flats.
The SOD also varied according to grain size in 2005 with the highest value
being observed in mixed sediment type (14.9 mmole m-1
d-1
) followed by the sand
type (13.9 mmole m-1
d-1
), and then mud type (12.6 mmole m-1
d-1
). The organic
matter concentration was not related SOD.
Temporal variations in denitrification activity
In 2005 the lower tidal flat had the highest denitrification rate (949.2 μmole
m-1
d-1
) followed by the middle (932.7 μmole m-1
d-1
), and then the upper (438.2 μ
mole m-1
d-1
) tidal flats.

- 21 -
S U M M A R Y X
In 2006 the denitrification was the highest in the middle tidal flat (881 μmole
m-1
d-1
), followed by the lower (760.3 μmole m-1
d-1
), and then the upper (591.9μmole
m-1
d-1
) tidal flats. In 2005 the station containing the muddy sediment had the
highest denitrification rate (921 μmole m-1
d-1
) followed by the sand (743.5 μmole
m-1
d-1
), and the mixed (655.8 μmole m-1
d-1
) sediment types.
Benthic community change on the artificial tidal flat
The station constructed in 2005 had 22~23 species and this amount was higher
then the station constructed in 2006, which contained 12~22 species. The stability
of the 2005 station seems to be higher than of the 2006 station. The amount of
individuals in the 2005 station was higher at 8400~8800 individual m-2 while for the
2006 station the number was ~4500 individual m-2.
In the 2005 station differences in species number and individual density were due
to tidal height and sediment type and lower species number were found at the
upper tidal flat in April and September 2006. The lower tidal flat had a higher
species number. Individual density was different for each sediment type. The upper
tidal flat had the lowest density with an increased gradient observed towards the
lowest stations. The maximum density was found for the mixed sediment type.
Section 5. Natural cleaning capacity of the Nak-dong estuary
1. Organic matter decomposition and organic matter removal through
denitrification
The annual organic matter decomposition in the sediment was approximately 7.1
x 106 tons-C. This value amounts to 19 % of the total amount of organic matter
(COD equivalent, 38.3 x 106 ton/yr) flowing into the Nak-dong estuarine system.
Organic matter removed by denitrification is 3.9 x 106 tons-C in the Nak-dong
estuary, this value is 10% of incoming COD.
2. Nutrient removal by Zostera marina
Zostera marina shoots can clean 60 ton of seawater containing 10 μM of
ammonia and can clean 270 ton annually. Zostera marina shoots living specifically
in the Nak-dong estuary can clean 189 million tons of seawater annually
containing 10μM ammonia.

- 22 -
S U M M A R Y XI
3. Nutrient storage by Scirpus triqueter
When Scirpus triqueter growth is high the ammonia concentration in the
sediment porewater declines and Scirpus triqueter is thought to contribute to
nutrient control in the Nak-dong estuary. The production of Scirpus triqueter
shoots is 1890 ton DW/yr, carbon storage is 752 ton DW/yr, and nitrogen storage
is 53 ton DW/yr, in the Nak-dong estuary.
4. Nutrient storage by Phragmites australis
Phragmites australis has the highest production with 4371 ton DW/yr, carbon
storage is 1740 ton DW/yr, nitrogen store is 122 ton DW/yr, in the Nak-dong
estuary. We can suppose that Phragmites australis has an important role to play
in the cycling of nutrients in the estuary.
V. Applications
The project will aid in the:
- Choosing of optimal building strategies during large scale restoration projects
- Setting of marine environmental quality standards for the future
- Improvement of fishery resource management in coastal region
- Enhancement of fishery resources through sea grass bed restoration
-of a management plan for coastal regions

- 23 -
C O N T E N T S
Chapter I. Introduction ············································································································ 25
Section 1. Purpose of the study and necessity ···································································· 27
Section 2. Content and scope ····································································································· 33
Chapter 2. International and domestic trends in related technology ·············· 37
Section 1. International trends ··································································································· 39
Section 2. Domestic trends ········································································································· 40
Section 3. Importance of the research ····················································································· 41
Chapter 3. Results of the research ······················································································ 43
Section 1. Characteristics of the Nak-dong estuary ············································ 45
1. Sediment characteristics and tidal height of the tidal flat ········································ 45
2. Water characteristics of the Nak-dong estuary ··························································· 53
Section 2. Structure of the ecological components of the Nak-dong
estuarine ecosystem ······················································································································· 58
1. Primary producers ················································································································· 58
2. Consumers ····························································································································· 103
Section 3. Functions of the ecological components of the Nak-dong
estuarine ecosystem ····················································································································· 156
1. Ecological processes ············································································································ 156
2. Food web analysis ·············································································································· 193
3. Material budget ···················································································································· 212
Section 4. The restoration of the ecological components in the Nak-dong
estuary ················································································································································· 216
1. Zostera Marina ····················································································································· 216
2. Scirpus triqueter ·················································································································· 222
3. Environmental and functional change of the artificial tidal flat ···························· 236
Section 5. Natural cleaning capacity of the Nak-dong estuary ·················· 275
1. Organic matter decomposition and organic matter removal through denitrification
······················································································································································ 2275
2. Nutrient removal by Zostera marina ············································································· 277
3. Nutrient storage by Scirpus triqueter ··········································································· 279
4. Nutrient storage by Phragmites australis ···································································· 280
Chapter 4. Contribution to related fields ······································································· 281
Chapter 5. Applications of the results ············································································· 285
Chapter 6. Information gathered during the research ············································ 289
Chapter 7. References ················································································································· 293

- 24 -
목 차
제 1장 서론 ········································································································································ 25
제 1절 연구개발의 필요성 ············································································································· 27
제 2절 연구 목표 및 범위 ············································································································· 33
제 2장 국내외 기술개발 현황 ··································································································· 37
제 1절 국외의 하구 습지 연구 ····································································································· 39
제 2절 국내의 연안 하구 습지 연구 ························································································· 40
제 3절 국내외 기술과의 차별성 ··································································································· 41
제 3장 연구개발 수행 내용 및 결과 ······················································································ 43
제 1절 낙동강 하구의 환경 특성 ······························································································· 45
1. 퇴적물 특성 ····························································································································· 45
2. 수층 특성 ································································································································· 53
제 2절 낙동강 하구의 생태 구성 요소 ···················································································· 58
1. 일차 생산자 ····························································································································· 58
2. 소비자 ····································································································································· 103
제 3절 낙동강 하구의 생태 기능 요소 ·················································································· 156
1. 생태 과정 (ecological processes) ····················································································· 156
2. 먹이망 분석 ··························································································································· 193
3. 물질수지 ································································································································· 212
제 4절 낙동강 하구의 생태 구성 요소의 복원 ···································································· 216
1. 잘피 ········································································································································· 216
2. 세모고랭이 ····························································································································· 222
3. 실험용 갯벌의 환경과 기능변화 ······················································································· 236
제 5절 낙동강 하구의 자정 능력 ····························································································· 275
1. 인공 갯벌의 유기물 분해와 탈질소화를 통한 유기물 제거 ······································· 275
2. 잘피에 의한 영양염 제거 ··································································································· 277
3. 세모고랭이에 의한 영양염 저장 ······················································································· 279
4. 갈대에 의한 영양염 저장 ··································································································· 280
제 4장 목표 달성도 및 관련분야에의 기여도 ··························································· 281
제 5장 연구개발결과의 활용계획 ·················································································· 285
제 6장 연구개발과정에서 수집한 해외과학기술정보 ·············································· 289
제 7장 참고문헌 ····························································································································· 293

- 25 -
제 1장 서론

- 26 -

- 27 -
제 1장 서론
제 1절 연구개발의 필요성
낙동강 하구는 부산광역시에 접하여 있고, 김해국제공항이 위치하고 있어 국내 연안
중에서도 이용이 가장 활발한 곳이다. 이곳은 한반도에서 유일한 하구 삼각주 보전구역
으로 부산광역시 사하구의 34.2㎢가 생태계 보전지역으로 지정되었으며, 생태계의 중요
성으로 문화재 보호구역으로 중복 지정되어 있다. 이곳의 생태 환경을 살펴보면, 낙동
강 하류에서 철새도래지로서 기능은 유지되고 있으나, 면허 허가어업과 생계유지형 어
업이 생태계 균형을 위협하고 지형이 급격히 변하고 있을 뿐만 아니라 관리인원, 지원,
체계 미비로 적절한 관리가 이루어지지 않고 있는 실정이다. 낙동강하구의 자연생태계
보전구역 생물상 및 서식환경을 요약해보면 장림 하수처리장 인근의 수질오염이 심각
하며 하구둑 상부의 담수역의 부영양화 정도가 심각한 수준이다.
낙동강하구 생태계 변화를 살펴보면 가장 뚜렷한 것이 하구습지의 소실을 들 수 있
는데, 1980년대 말부터 하구둑 건설, 공단 및 주거단지 조성 등으로 약 500만평의 하구
습지가 매립 소실되었다. 이러한 습지소실은 서식처의 단편화(fragmentation), 생태계의
구조와 기능상의 변화를 초래하였으며 철새 도래에도 큰 영향을 미쳐 이미 그 수가 크
게 감소한 상태이다. 기타 습지 및 갯벌 소실원인은 을숙도내 노천분뇨 처리장, 하구둑
건설과 동시에 을숙도 및 하단 쪽 수변부의 콘크리트제방 설치, 을숙도내 쓰레기 매립
장 건설, 문화재 지정구역의 무분별한 해제 등을 들 수 있다. 한편, 낙동강하구 지형은
급격하게 변하고 있다. 특히 생태계보전지역 내에는 새로운 모래톱이 계속 생성되고 있
으며 이런 새로운 서식처에는 식물의 천이에 따라 토양미생물, 소형동물, 조류 등의 서
식환경이 조성된다. 이러한 낙동강하구 서식처의 환경변화는 지형분야에 제한적으로 연
구가 되고 있을뿐 생태연구의 대상으로 다루어진 경우는 극히 적다.
생태계보전지역 및 인근 해역의 생태계 변화를 일으키는 주 요인들로는 우선 가덕
신항만 조성에 따라 자연생태계 보전지역의 해류정체가 가속화할 가능성이 높으며, 명
지 주거단지 및 신호, 녹산공단의 오폐수 발류, 낙동강하류에 도달하는 유량의 감소, 어
로행위, 을숙도 하단부의 개발(명자대교 건설), 그리고 장림 하수처리장들의 운영(염색
및 피혁 포함)을 들 수 있다. 그러나 낙동강하구에는 다양한 보호구역 명목으로 법적,
제도적 장치가 마련되어 있음에도 불구하고 저서생물, 어류, 철새 등 생물 서식환경은
악화되고 실질적인 생태계 관리가 이루어지지 않고 있다. 따라서 생물다양성 증대 및
지속적인 유지와 관리를 위해서는 하구습지의 복원과 같은 실질적인 방안이 절실히 요
구된다.
대규모 간척사업이나 토사채취 등과 같은 연안역의 무분별한 개발과 이용으로 해안
습지 특히 갯벌과 같은 연안 생태계가 심각하게 위협받는 것은 단지 낙동강 하구만의
문제는 아니다. 시화호와 새만금 매립에서 보듯이 연안의 개발과 보존에 대한 갈등이

- 28 -
심각해지고 있으며, 국론이 분열되고 막대한 사회 비용이 치러지고 있다. 연안환경보존
에 대한 일반인의 인식이 높아지고 공유수면 매립법, 습지 보전법, 연안역통합관리법
등이 제정되면서 연안습지의 개발 힘들어진 것은 사실이나, 인구밀도가 높고, 해안을
따라 U-자형 개발이 이루어질 수밖에 없는 국내 현실에서 연안개발은 피할 수 없을 것
이다. 따라서 해안습지의 보전과 복원, 관리를 위한 기술과 기준을 마련하는 것이 시급
하나, 국내의 연안 하구역 습지의 복원과 관리기술에 관한 지식 및 기술 수준은 일천하
다. 선진국의 경우 일찍이 연안역통합관리(integrated coastal management)를 통해 연
안 이용과 지속가능한 개발에 힘쓰고 있으며, 단순한 보존을 넘어 친환경적인 개발을
통해 경제적인 이익창출을 도모하고 있고 관련 기술이 잘 발달되어 있다. 그러나 국내
와 같이 집중적 연안개발이 드물고, 조석 조건, 연안의 이용 등이 달라 단순한 기술 차
용에는 한계가 있다. 따라서 본 연구의 목표는 국내 현실에 맞는 연안 습지(갯벌 및
조하대) 생태계의 복원 및 관리에 필요한 환경/생태학적 파라미터들을 개발하여 향후
실제 복원공사에서 지속가능하고 경제적인 복원 및 관리에 고려해야할 사항들을 제공
하는데 있다.
가. 연안하구 습지 생태계의 특성을 고려한 습지 복원
생태학적으로 연안 하구 지역은 육상기원 물질이 해양으로 유입되는 통로이다. 주로
강을 통해 무기물 (토양, 중금속, 영양염 등)이나 유기물 (식물체나 동물체의 잔재물,
생활 쓰레기 등) 이 연안지역으로 유입되며, 해안을 통해 직접 유입되는 양도 많으며
기타 지하수를 통해 혹은 대기 중으로 물질 유입이 이루어진다. 적당한 양의 물질유입
은 연안 해역의 생산성을 높이는데 기여 하지만, 너무 많은 양의 유입이나, 독성 물질
의 유입 물질 유입은 국내 여러 하구 연안에서 보듯이 오염, 적조, 무산소층형성 등 연
안생태계에 부정적인 영향을 끼치게 된다.
연안 하구 습지의 주요 환경중 하나인 갯벌은 지리적인 위치의 뿐만 아니라, 생태 기
능 측면에서도 매우 중요한 지역이다. 갯벌은 육상 환경과 해양 수환경과의 경계에 있
어 여러 환경요인의 변화 경사 (gradient)가 큰 곳이며, 자체 생산된 유기물과 유입된
유기물이 풍부하여, 많은 생물이 살고 있는 곳이다. 갯벌에 살고 있는 다양한 생물과
풍부한 생물량은 이곳의 생산성이 매우 높다는 것을 증명하고 있다. 이곳에서는 저서
생태계의 중요 기능 중의 하나인 물질의 분해과정이 활발하게 진행되어 갯벌은 자연의
자정 작용이 일어나는 곳이기도 하다. 갯벌에서 일어나는 일차생산과 분해과정은 갯벌
과 주변 해역의 영양염 순환과 수지를 조절하는 기본 과정이다. 갯벌이 갖는 가치는 이
미 교과서 수준에서 다루어지고 있으며 최근 생태계 서비스(ecosystem service) 측면에
서 보면 인간 생활은 물론 주변 환경과의 연계된 파급 효과가 매우 크다(Nybakken
2001). 최근 연안 습지 개발 사업은 이러한 갯벌의 생태계 서비스를 간과하여 진행되고
있는 경향이 있다. 특히 매립과 둑 건설로 비롯된 서식처 훼손과 손실을 포함한 연안생
태계의 변화가 예상보다는 훨씬 나쁜 양상으로 나타나 기존의 시설을 다시 사업 전의

- 29 -
상태로 회복시키는 복원 사업도 진행되고 있다.
갯벌의 기능과 중요성은 지역에 따라 많은 차이가 있으며 각 갯벌 생태계의 구성 및
기능 특성을 파악하여 갯벌 자원을 효율적이고 보존가능하게 이용하기 위해 연구가 필
요하다. 특히 일차생산을 주도하는 퇴적물내의 미세조류(microphytobenthos: 이후 MPB
로 약칭)에 대한 연구는 갯벌의 저서생태계를 먹여 살리는 유기물을 생산하는 중요한
과정이다. 갯벌의 일차 생산은 크기가 작은 미세조류에 속하는 저서 돌말류, 유글레나
를 포함하는 편모조류와 다른 단세포 조류를 포함하는 총칭 MPB의 광합성에서 비롯된
다. 또한 생산된 유기물은 먹이 그물 속에서의 이동과 분해 과정을 거쳐 순환되고 있
다. 특히 퇴적물에서 일어나는 미생물먹이망(mirobial food web)의 구실은 매우 큰 것
으로 추정하고 있다. 또한 유입된 유기물에 의한 과정도 간과할 수 없지만, 갯벌 생태
계의 일차생산의 기여도를 파악하는 것이 기본적으로 파악되어야 한다.
대기중에 노출되는 해양환경인 갯벌과 수심이 낮은 조하대에서는 MPB 이외에 잘피
가 높은 기초생산을 담당하고 있다. 잘피 (seagrass)는 해수에 적응되어 연안이나 하구
생태계에 분포하고 있는 고등현화식물을 통칭하며, 해조류 (algae)와는 구별되어진다.
약 60여종의 잘피가 전 세계 거의 모든 해안에 분포하고 있고, 우리나라 연안에는 약 8
종의 잘피가 분포하고 있는 것으로 알려져 있다. 잘피는 많은 수산어족 자원들에게 중
요한 서식지, 산란장 및 치어 생육지를 제공해주며, 이들이 만든 유기물질은 많은 어류
의 직접적인 먹이원으로, 또는 미생물에 의해 해양 및 연안생태계의 먹이사슬로 편입되
어져 수산생산성과 밀접하게 관련되어 있다. 잘피의 밀생한 지하경 및 뿌리조직은 해저
토양을 안정화시키며, 또한 해저토양내의 중금속과 같은 오염물질을 흡수제거 할 수 있
고, 잘피의 잎은 하구로부터 과도하게 유입되는 질소나 인과 같은 영양물질을 빠른 속
도로 흡수해 제거함으로써 적조현상과 같은 환경재해를 줄여줄 수 있어, 연안의 수질을
향상시키는데 중요한 역할을 하고 있다 (Lee and Dunton 1999). 이처럼 잘피는 경제적
가치와 생태적 가치를 함께 지니고 있어, 수산생산성 향상과 연안 생태계 정화를 위해
보호해야할 중요한 우리의 자원이다.
연안 및 하구 생태계에 중요한 역할을 하고 있는 잘피가 최근에 전 세계 서식지의
약 50%가 인간에 의한 환경 파괴로 사라졌다고 보고되고 있으며, 우리나라에서는 1970
년대 이후 산업화의 영향으로 대량의 오염물질이 연안 및 하구생태계로 유입되어 연안
과 내만의 수질과 저질을 심하게 오염 시켰으며, 이러한 오염에 의해 연안의 잘피서식
지가 많이 훼손되었다. 또한 농지나 공장 부지를 위한 매립은 수산생산성과 밀접하게
관련되어있는 잘피서식지를 크게 감소시켰다. 이러한 연안의 잘피서식지 감소는 우리나
라 주요 어족자원들의 치어 및 성어의 생육지 감소를 야기하여, 수산생산성이 저하와,
연안 생물다양성 감소로 이어졌다. 따라서 연안 생태계의 정화와 수산생산성의 향상을
위하여 잘피서식지의 보존 및 복원이 시급한 실정이다. 잘피 이식을 통한 훼손된 잘피
서식지의 복원이나 새로운 잘피밭의 조성은 우리의 연안환경을 환경친화적으로 정화하
고 수산경제적 가치가 높은 어족자원들에게 적합한 생육지를 제공하여 매우 효과적으
로 수산생산성의 향상을 꾀할 수 있다. 본 연구에서는 수산생산성향상과 연안환경정화

- 30 -
에 중요한 역할을 하고 있는 잘피 서식지의 복원을 위하여 효과적인 잘피 이식방법을
개발하고 낙동강 하구 습지에 적용할 것이다.
하구역 갯벌의 경우 조석운동과 담수 유입에 의한 물의 유동에 노출되어 있어 퇴적
물의 퇴적과 침식이 작용이 아주 활발하게 일어나고 있는 지역이다. 과거에는 갯벌과
같은 습지는 모래로 된 사구와는 달리 계절 변동을 보이지 않는다고 생각해 왔으나 최
근 연구 결과를 보면 놀랄만한 정도로 큰 계절 변동을 보이는 것으로 보고하고 있다
(해양연구원, 2004). 특히 연안 개발에 따른 피해를 보상하기 위해 인공적으로 갯벌을
조성할 경우에는 물리적인 환경요인 (조고, 바람, 담수 유입, 연안해류)과 퇴적 환경특
성(퇴적입자의 조성, 노출시간 및 경사도)이 잘 어우러질 경우에만 유지가 가능할 것으
로 점쳐진다. 따라서 최적의 환경 조건의 조합을 찾아내서 조성하는 것이 갯벌이란 공
간 환경의 유지뿐만 아니라 건강한 저서생태계를 부양하고 유지하는데 필수적이다.
그 다음으로 중요한 조건은 깨끗한 환경이다. 훼손된 습지는 흔히 산소를 요구하는
오염물 부하가 소위 자정능력이라고 부르는 능력에 비해 과다하게 걸렸을 때 잘 나타
난다(Jørgenson and Richardson, 1996). 산소 공급이 원활하지 못해서 분해 되지 못하
고 떠다니는 각종 입자와 유기물로 인해 바닷물 색이 탁해지고, 그 결과 광합성이 저해
되면서 먹이 피라미드의 근간이 무너지게 되어 우리에게 친숙한 대형 동식물이 사라진
다. 이것이 더 진행되면 빈산소화를 거쳐 무산소화로 진행되면서, 산소 이외의 산화제
를 사용하면서 나오는 황화수소나 메탄이 내는 고약한 냄새와 독성으로 인해 수산물
생산이나, 관광과 여흥을 제공하는 공간 자원으로서의 가치를 상실하게 된다. 따라서
훼손된 습지를 복원하고자 할 때 최우선적으로 살펴보아야 할 사항 가운데 하나가 습
지의 유기물 부하와 퇴적물의 산소요구량을 파악하는 일이다. 이러한 관계를 파악하기
위해서 본 연구에서는 아직까지 제대로 보존되어 있는 낙동강 하구 일대의 자연연안습
지를 대상으로 퇴적물 특성과 환경 조건을 연구하여 이상적인 습지를 이루는 환경 요
인의 조합을 파악하고자 한다. 여기서 얻은 지식은 곧바로 인공습지를 조성하고 관찰하
는 구체적인 실험에 적용될 수 있다. 낙동강 하구역 인근에는 부산진해 신항만 조성과
같은 대규모 토목 공사가 진행되고 있기 때문에 자연습지의 훼손이 불가피한 실정이다.
공사 후반기에 또는 공사 뒤에는 인공습지를 조성해야할 필요가 아주 높은 곳이다. 그
래서 식생이나 생태계가 비슷한 인근 지역인 낙동강 하구에서 습지 보전과 복원에 대
한 연구를 하는 것이 이상적이다. 이 연구의 결과를 바탕으로 해서 다양한 퇴적물 특성
과 지형을 가진 실험용 인공 습지를 조성해서 비교 연구를 통해 “유지될 수 있으며, 건
강한 저서생태계를 가질 수 있는 조건”을 제시하고자 한다.
나. 연안하구 습지의 기능과 자연정화
연안역의 무분별한 개발과 이용으로 야기되는 인위적인 환경 변화가 해안습지 특히
갯벌과 같은 연안 생태계를 파괴시키는 정도는 이미 매우 심각한 수준에 이르렀다. 이
에 따른 연안생태계의 가장 뚜렷한 반응에는 생물종의 구성(생물다양성)과 생물량(생물

- 31 -
생산)의 변동이 있다. 매립, 간척, 골재 및 규사용 모래채취 등에 따른 갯벌생태계의 변
화는 현재 가장 사회적인 논란거리가 되고 있는 대표적인 연안생태계 변화의 한 예가
된다. 이와 같은 생태계 변화는 기존의 생태학적 주요 기능들을 수행하던 생물군들을
사라지게하고 다른 기능을 가진 생물군들로 대체되는 현상을 가져올 뿐만 아니라 생물
생산 구조와 관련된 물질순환 구조를 변화시켜 궁극적으로 연안생태계 전체의 변동을
수반할 것으로 예상된다. 따라서 훼손된 생태계의 복원을 통해서 생물종 다양성과 생물
생산의 증대를 꾀하기 위한 노력들이 절실히 요구된다. 이와 같은 요구에 따라 최근 생
태계의 복원에 대한 넓은 공감대를 형성해 가고 있어 대규모 갯벌생태계 복원이 예상
되지만 실제 복원의 효과를 증명하기는 쉽지 않은 실정이다.
생태계 먹이원 분석에 의한 먹이망 구조를 밝히기 위한 전통적인 접근방법으로는 주
로 소화관 내용물을 분석하는 방법들을 이용해 왔다. 그러나 이 방법들은 많은 시료들
을 분석해야 하기 때문에 많은 시간과 경제성 및 노력이 요구될 뿐만 아니라 먹이생물
로 다양한 생물종을 대상으로 연구를 수행해야 하기 때문에 생물 분류에 대한 어려움
에 직면하게 된다. 또한 현미경으로 관찰하는 시료는 동물이 선호하는 먹이가 이미 소
화되고 난 후의 찌꺼기를 대상으로 할 수도 있다는 연구방법상의 오류를 가지고 있다.
그에 대한 대안의 방법으로 탄소, 질소 혹은 황화합물의 안정동위체 조성 분석을 이용
하는 방법이 효과적으로 이용될 수 있다. 이 방법은 동물이 먹이를 섭취하게 되면 이화
와 동화 과정을 거치면서 기원 유기물이 가지는 본래의 특성을 잃어버리는 반면 안정
동위원소 조성은 변하지 않고 보존된다는 특성에 근거한다. 따라서 어떤 연구대상 동물
체의 안정동위원소 조성을 알게 되면 이들이 이용하는 유기물의 기원을 평가할 수 있
게 된다. 이 분석 방법은 최소한의 시료로 짧은 시간 내에 비교적 적은 노력으로 동물
들이 상당기간 동안 실제로 이용해 온 먹이의 기원을 해석하게 해 준다.
본 연구에서는 전통적인 먹이 연구방법인 소화관 내용물 분석법과 함께 안정동위원
소 응용기법을 적용한 연안생태계 먹이망 분석을 통하여 연안생태계 내의 정확한 물질
순환 구조를 파악하고자 한다. 이로부터 자연갯벌과 복원갯벌 생태계 내 먹이연쇄를 통
한 물질순환을 이해함으로써 1) 연안역 개발과 환경변동에 의한 갯벌이나 해초장 및
하구생태계를 구성하는 기능군별 생물군집변동과 그에 따른 생물생산 및 물질순환구조
변동을 분석하고, 2) 연안의 생물생산에 대한 이들 생태계 각각의 역할과 기여를 평가
할 수 있게 될 것이다. 연안 하구 습지 (갯벌 및 조하대)의 생태 구조와 물질순환 구조
에 대한 이해와 실험 갯벌 조성이나 잘피 서식지 복원을 통한 연안 천해 생태계 안정
화과정의 파악은 연안의 생물종 다양성과 생물생산의 증대를 위해 필연적으로 요구되
는 기본정보라 할 수 있다.
새만금 매립 갯벌의 자연 정화 능력에 관한 논란이 있었는데 이는 갯벌로 유입된 유
기물이 미생물에 의해 분해되는, 갯벌의 유기물분해 능력이 얼마나 큰 것인가에 관한
것이었다. 갯벌은 노출과 침수로 인한 수층과 표면 퇴적물의 교란과 저서동물의 활동에
의한 퇴적층의 생교란(bioturbation) 등으로 인하여 퇴적층 내로의 산소 공급이 원활하
며, 많은 유기물의 유입에도 불구하고, 혐기성 분해과정보다 유기물 분해에 있어서 좀

- 32 -
더 효과적인 호기성 분해과정의 비중이 높을 것으로 예상되며 따라서 갯벌에서는 육상
에서 기원한 유기물의 분해가 매우 활발할 것으로 예상된다 (Howe et al., 1984).
그러나 갯벌에서 미생물에 의한 유기물의 분해 능력 전체를 유기오염 자연정화능력
으로 간주하는데는 어려움이 있다. 현재 갯벌의 자연정화능력에 대한 논란의 일부는
“자연정화의 개념”을 유기물 분해 능력으로 잘못 이해한데서 비롯된다. 유기물은 탄소
성분과 질소성분으로 이루어져 있는데, 미생물이 유기물을 분해할 경우 탄소성분은 이
산화탄소로서 시스템 내에서 제거되나 질소성분은 무기질소로 변하여 남아 있게 되며,
이러한 무기질소는 일차 생산을 제한하는 제한 요소로서, 광합성에 의한 유기물 생산에
이용될 것이다(Jorgensen 1977). 결국 탄소성분이 제거되었다 하더라도 질소성분은 남
아 다시 유기물을 생산하게 될 것이며, 미생물에 의한 유기물의 분해 능력 전체를 시스
템 내에서 유기물의 제거능력으로 간주하는 것은 잘못이다.
갯벌의 유기정화 능력을 파악하기 위해서는 미생물에 의한 유기물의 분해 능력과 더
불어 일차생산에 의한 유기물의 합성과정, 합성된 유기물의 소비과정을 파악하는 것이
중요할 것이다. 갯벌에서 일차 생산에 의해 생산된 유기물은 먹이망에서 상위 단계에
있는 생물들에게 소비되거나, 입자성 유기물 (POM; particulated organic matter)로서
미생물에 의한 무기화과정에 이용될 것이다. 어류나 기타 대형 저서동물들이 일차 생산
에서 생산된 유기물을 소비하여 생체량 (biomass) 을 만들고, 인간이나 조류 등이 이러
한 어류나 대형 저서동물을 소비하면, 갯벌 내에서 유통되는 유기물의 양은 감소할 것
이고 따라서 갯벌의 유기정화능력을 증가시키는데 기여할 것이다. 갯벌에 의한 유기물
의 분해와 미세조류에 의한 재생산과정은 유용하지 않은 유기물 혹은 생태계에 부정적
인 영향을 주는 유기물 (오염유기물; 생활폐수 등) 이 인간이나 조류 등이 이용할 수
있는 자원(자원유기물; 저서동물, 어패류)으로 변할 수 있는 기작을 보여준다. 이 기작
이 중요하다면 갯벌은 유기오염 자연 정화와 자원생성이 동시에 이루어지는 곳이 될
것이다.
먹이망에서 상위 단계에 있는 생물들이 수확되면서 갯벌 시스템에서 유통되는 유기
물의 양이 감소되는 것과 더불어 유기 정화능력에 중요한 과정은 탈질소화이다. 해양에
서 무기질소성분은 일차생산을 제한하는 영양분이므로 (Capone 1988), 무기질소성분의
변환과정의 속도 및 생성물의 성격은 일차생산에 의한 유기물 생산에 직접적으로 영향
을 미칠 것이다 (Rowe et al. 1975). 탈질소화(denitification)는 혐기성 미생물에 의해
질산이 질소가스로 변화되는 과정으로써, 탈질소화의 산물인 질소가스는 일차생산에 이
용되기 힘들다(Seitzinger 1990). 따라서 탈질소화는 시스템 내에서 총질소량을 감소시
키고 결국 유기물을 제거하는 기작이다. 탈질소화가 상세히 연구된 연안이나 하구에서
는 이 과정이 플랑크톤에 의한 일차생산성을 제한하거나 부영양화를 감소시키는데 중
요하다는 것이 밝혀졌다 (An and Joye 2001). 일반적으로 갯벌은 염습지와 마찬가지로
유기물 공급이 높아 탈질소화가 높을 가능성이 있으나 현재 세계적으로 갯벌에서 탈질
소화를 측정 자료는 매우 희귀한 실정이다 .
연안 습지 복원 의 목표는 생태계 구성요소의 서식처를 보호하여 종다양성을 확보하

- 33 -
여 건강한 생태계를 유지시키며, 자연습지가 가진 정화능력을 극대화 하는 것이 되어야
할 것이다. 이를 달성하기 위해서는 이 생태계의 생물학적 구성요소 (1. 일차생산자: 미
세조류, 잘피 등; 2. 소비자: 저서동물, 어류; 3. 분해자: 미생물)와 무생물학적 구성요소
(환경요인, 수질, 퇴적물, 부유물, 유기물, 무기물 등)의 상호관계를 면밀하게 밝히는 것
이 필요할 것이다. 본 연구의 목적은 연안하구 습지생태계의 구조와 기능을 살펴 환경
변화에 대한 생태계변화를 예측할 수 있는 일련의 환경/생태학적 파라미터를 개발하는
데 있다. 낙동강 하구의 습지 훼손지역에 실험용 복원습지를 습지를 조성한 후 종다양
성, 생물량 등의 생물학적 요소와 지형, 퇴적물 특성 등의 변화과정을 살펴봄으로서 1.
유지가능하고, 2. 종다양성이 확보되며, 3. 습지 본래의 기능을 다하는 습지를 만드는데
필요한 환경/생물학적 파라미터를 찾는 것이 목표이다.
제 2절 연구 목표 및 범위
가. 연구 목표
본 연구의 연구 목표는 낙동강 하구에 위치한 자연 상태의 연안 하구 습지 (갯벌 및
조하대)의 물리 및 생태 환경 구조, 물질순환 구조, 건강한 습지를 유지하는 요인들을
파악함과 동시에, 실험용 복원습지 (갯벌 및 조하대 잘피림)를 조성하여 환경 및 생물
상 변화를 통해 안정화 과정을 살펴 환경/저서생태계/물질순환에 관련된 복원 및 관리
파라미터를 도출하는 것이다.
- 낙동강 하구 습지의 환경특성 파악
건강한 연안 습지를 유지시키는 물리학적, 퇴적학적, 수질 요인을 찾아내고 이들 요
인간의 인과관계를 구명하여 최적 습지 복원/관리 방안을 도출.
- 낙동강 하구 습지의 저서생태계 특성 파악
저서 생태계의 생물학적 구성요소 (1. 일차생산자: 미세조류, 잘피, 세모고랭이, 갈대;
2. 소비자: 저서동물; 3. 분해자: 미생물)와 무생물학적 구성요소 (환경요인, 수질, 퇴적
물, 부유물, 유기물, 무기물 등)의 구조와 기능을 파악.
- 낙동강 하구 습지의 물질순환 특성 파악 및 복원/관리 파라미터 개발
서로 다른 환경 조건에서 먹이망을 통한 물질 순환 경로 및 갯벌의 자연 정화능력을
파악하여, 생태계 구성요소의 변동에 대한 전체 생태계의 반응을 예측하고, 연안 습지

- 34 -
복원 및 관리에 필요한 환경/생물학적 파라미터 개발.
나. 연구 범위
Table 가. 낙동강 하구역 습지 생태계 연구 범위 및 내용
구분연구 범위 및 내용
자연 및 실험용 습지 퇴적물의 지화학적
특성 변화 (계절변화 및 지형변화)
퇴적물 입도, 영양염 농도, 유기물농도, 조고 및
지형변화.
자연 및 실험습지의 퇴적물에서 미세조류
의 종다양성, 일차생산자 밀도와 생물량
수직 이동 등의 변동, 광합성 효율 분석
종조성, 다양성, 서식밀도, 클로로필량, 광합성효
율, 형광수율 측정
미세 전극을 이용한 일차생산성 측정
자연 및 실험습지의 퇴적물의 저서동물 종
조성, 생물량의 계절변화와 환경요인
종조성, 다양성, 서식밀도, 습중량, 건중량,
탄소, 질소량 파악
주요 일차생산자의 현황 및 생산성 조사,
인공 잘피림과 자연 잘피림 비교 평가, 잘
피의 생존조건
주요 일차생산자의 현황 및 분포도 작성, 인
공 잘피림 서식지 내 물리 화학적 환경요인,
인공잘피림과 자연 잘피림의 일일 광량 및
온도 측정
자연 및 실험습지 퇴적물내 미생물의 유기
물 무기화 과정, 영양염 제거 및 순환형태
파악, 각 과정의 상관관계
퇴적물산소요구량 (SOD), 탈질소화, 영양염 flux.
자연 및 실험습지의 퇴적물의 생태계 생물
-무생물간의 물질순환 파악
입자성 및 용존성 부유물질, 생체의 탄소 및 질소
안정동위원소 분석을 통한 물질순환 파악
실험용 갯벌 조성을 통해, 시간에 따른 환
경변화, 일차생산자, 소비자, 분해자의 종
조성, 밀도 및 활동 변화 파악
안정화에 따른 조고, 입도, 유기물 농도,
클로로필, 대형저서동물, SOD, 탈질소화 변화
파악을 위한 실험용 갯벌 조성 (1 x 1 m x 9개 x
2조)
시스템 budget 을 통한 각 생태 요인의 기
능 파악
담수 및 영양염 유입량 파악, 각 생태 요인의
현존량 비교 (환산인자 개발), 생지화학적 변환
process 속도 파악
(1) 자연 및 실험 습지 퇴적물의 지화학적 특성
자연 및 실험 습지 환경 특성 파악: 여기에 해당하는 인자는 크게 퇴적환경, 수질과
조석으로 대별되며, 이들에 관련된 인자들의 시공간적 변동에 대해 조사하였다.
- 퇴적환경: 퇴적입자의 입도, 함수율, 유기물 함량
- 수질: 수온, 염분, 용존산소부유물질 (유, 무기), 엽록소-a
- 조석 및 지형: 조석주기, 조고, 노출시간 및 노출면적

- 35 -
(2) 저서미세조류(MPB) 의 동태
갯벌의 MPB 군집의 일차생산과 관련된 생태 연구는 일차생산자를 구성하는 종의 확
인과, 색소체를 분석하여 추정한 생물량의 변동과 일차생산성의 측정으로 진행되어 왔
다(참고: 고, 2001). 기존의 국내 연구에서 간과되어온 저서미세조류의 수직 이동과 조
석주기에 따른 시계열 변동에 대한 연구를 시도하였다. 또한 생 시료의 관찰을 병행하
여 단세포 미세조류의 다양성도 확인하였다. 현장에서 수집한 갯벌의 MPB 군집의 종
구성을 분석하기 위해 생 시료 또는 고정된 시료를 광학현미경(Zeiss, Olympus; 형광,
DIC 부착), 전자현미경(SEM)으로 관찰하여 종을 동정한다. 갯벌의 MPB 군집과 표영
계 식물플랑크톤 군집이 갖고 있는 엽록소-a와 보조 색소체 함량의 분석은 아세톤으로
추출한 엽록소와 다른 색소체의 함량을 분광분석과 HPLC 법으로 정량 분석하였다
(Jeffrey et al. 1997; Brotas and Plante-Cuny 1998).
(3) 저서동물의 동태
자연 습지 및 실험용 습지에서 저서동물의 군집천이과정 살펴 보기위하여 현장조사
를 수행하였다. 각 조사정점에서는 소형 저서생물채집기인 grab (채집면적이 0.025m2)
이나 박스 코어를 사용하여 4회씩 채집한다. 현장에서 채집된 퇴적물은 0.5mm 체에 넣
고 해수로 걸러 남은 시료를 10% 포르말린으로 고정한다. 각 시료는 별도 보관하여 조
사정점 당 평균 현존량의 산출에 사용한다. 실험실에서는 고정된 시료를 주요 분류군별
로 선별하여 습중생물량과 개체수를 산출하고, 가능한 종 수준까지 동정하여 현존량 자
료를 산출한다. 이렇게 산출된 현존량 자료를 이용하여 각 출현종의 시기별 변동을 추
적하고, 다음과 같은 군집지수를 구한다. 저서생물군집의 구조적인 특성을 설명하는 생
태지수로는 종다양성지수 (H' ), 종풍부도지수 (R), 균등도지수 (J), 우점도지수 (D)가
있으며, 이들 군집지수들은 생태계 구성원들의 변동을 지수라는 요약된 형태로 제시해
주기 때문에 환경요인에 반응하는 시공간적인 생물군집의 변화를 알아낼 수 있다.
(4) 잘피 분포와 생존조건 판별, 복원기술개발 및 인공 잘피림 조성
낙동강 하구에 아직까지 현존하고 있는 잘피 서식지의 분포를 SCUBA, 수중 카메라
및 GPS 장치를 통하여 파악하고 분포도를 작성하였다. 이외에 지상부와 지하부 식물조
직 내 탄소 및 질소 함량 측정, 저장 탄소 (carbon reserve)의 계절적 및 공간적 변화
측정, 잘피의 형태학적 특성 및 엽록소 함량 측정, 잘피 서식지의 물리 화학적 환경요
인 측정 등을 실시하였으며, 잘피 이식 방법을 개발 하여 실제 복원을 시행하였다.
(5) 퇴적물내 미생물의 유기물 무기화 과정, 영양염 제거 및 순환형태 파악

- 36 -
미생물에 의한 유기물 분해 과정은 갯벌이 가진 유기물 정화능력을 파악하는데 중요
하다. 이중에서도 탈질소화는 일차생산을 제한하는 영양분인 무기질소 성분을 제거하는
과정으로서 일차생산에 의한 유기물 생산량에 영향을 미칠 것이며 결과적으로 정화능
력을 결정하는 한 부분이 될 것이다. 본 조사에서는 퇴적물 배양 실험을 통하여, 낙동
강 하구 자연 갯벌과 실험용 습지에서 미생물에 의한 유기물의 무기화 과정, 질소 영양
염의 변환과정 특히 탈질소화를 통한 유기물의 제거 과정을 살폈다. 자연 갯벌과 실험
용 갯벌에서 각각의 과정을 측정하여, 이들 미생물 과정에 영향을 끼치는 환경 및 생태
적 요인을 알아보았다.
(6) 연안 습지 생태계 생물-무생물간의 물질순환 파악
연안생태계로 유기물을 공급하는 주요 경로로는 인근의 바다와 강을 통한 외래기원
(타지성) 유입과, 갯벌, 해조장, 해초장 및 하구생태계에 유래를 둔 연안 자체생산(원지
성)을 들 수가 있는데, 본 연구에서는 이들 각각의 공급원들로부터 유래되는 유기물의
연안생태계 물질순환에서의 역할과 기여를 정확하게 평가하고 연안환경의 변화에 따른
전체 연안생태계 변화에 미치는 효과를 구체적으로 제시하고, 생물종의 보존과 생물생
산을 유지하는데 필요한 연안생태계 복원의 필요성을 증명하여 과학적 연안생태계 관
리방안을 제시한다. 낙동강 하구 주요 생태계 구성요소들 각각의 안정동위원소 조성을
분석하여, 먹이망을 통한 물질순환 경로와 연결고리들을 모식화 하였다.
(7) 실험용 갯벌 조성
실험용 갯벌은 자연 상태에서 나타나는 갯벌의 다양한 퇴적물 입자(니질, 사니질, 사
질)로 조성하였다. 퇴적물 입자와 함께 중요한 사항은 조고이다. 조고는 특정 갯벌 지
역이 대기중에 노출되는 시간을 결정하는데, 일반적으로 갯벌의 경사도는 매우 완만하
여 (1/100 이하) 인접 지역간에 노출시간의 차이는 크지 않으나 저서생물의 분포는 매
우 명확한 대상분포 (zonation)를 보이는 경우가 흔하다. 이러한 양상은 갯벌에 서식하
는 노출 시간보다는 연속노출시간이 중요함을 시사한다. 즉 임계수심 (critical tidal
level) 전후로 조고의 차는 크지 않으나 연속노출은 뚜렷한 구분을 보이게 되며 각 연
속노출에 적응할 수 있는 생물만이 각 지역에 서식하는 것이다. 본 조사에서는 DGPS
를 이용한 조고 산출, 장기간의 조위 변동 분석을 통해 임계수심을 파악하고, 각 연속
노출 범위에 해당하는 조고에 실험용 습지를 설치하였다. 1차년도에 1m2 (1m x 1m)
면적의 9의 실험용 습지를 조성하여, 운용하고 있으며, 2차년도인 2006년 2월에 4m2
(2m x 2m) 면적을 가지는 습지를 조성하여, 이곳에서 시간에 따른 환경변화, 일차생산
자, 소비자, 분해자의 종조성, 밀도 및 활동 변화 파악하였다.

- 37 -
제 2장 국내외 기술개발 현황

- 38 -

- 39 -
제 2장 국내외 기술개발 현황
제 1절 국외의 하구 습지 연구
미국, 일본 유럽 등 선진국에서는 연안역통합관리의 개념을 도입하여, 연안을 보존
복원하는 제도 및 기술이 정착되어 있다. 미국의 경우 1990년부터 8년 동안 연안 습지
계획, 보호, 복원법(Coastal Wetland Planing, Protection and Restoration Act)을 통해
루이지에나 주 연안의 습지를 복원하였다. 식물의 식재 (Vegitative technique), 구조물
(Structural technique), 퇴적물 (Sedimentary technique), 수리기법 (Hydrolgic
technique) 등을 통해 성공적으로 습지를 조성하였다. 일본의 경우 대표적인 습지 복원
사례로서는 1990년에 완성된 히로시마현의 이츠카이치 인공습지로서 42억엔을 투자하
여 24ha 의 인공습지를 조성하였다. 유럽 연합의 경우도 1989년부터 MAST(Marine
Science and Technology) 프로그램을 운영하여, 연안역 수리학 및 퇴적역학을 연구하
여, 연안습지 관리 및 인공습지 조성을 위한 기초 기술을 가지고 있다. 그러나 이 들
선진국의 국내와 다른 여건을 가지고 있어 단순한 기술 차용에는 한계가 있다.
미국의 경우, 연안 인구 집중 정도가 낮아 국내처럼 집중적 연안개발이 드물고, 갯벌
의 발달이 미미하여 연안 하구 “복원”보다는 “보존” 에 중점을 두고 있다. 조하대 잘피
림의 경우 생리생태학적 특성에 관한 많은 정보를 가지고 있으며, 이들 정보가 실제 잘
피서식지의 복원 수행에 이용되어 수산 생산성 향상 및 연안생태계 정화를 꿰하고 있
다 (Davis and Short, 1997, Short et al, 2000). 이러한 이식기술을 우리나라의 잘피서
식지 복원에 적용할 수도 있으나, 우리나라 연안환경에 적절한 이식기술과 이식 비용을
줄일 수 있는 기술개발이 필수적이다.
국내와 여건이 유사한 일본의 경우 파괴된 연안 습지를 대체하는 대체 습지가 법제
화 되어 인공갯벌의 조성이 활발하다. 인공갯벌 조성에서 그 목표는 자연갯벌과 유사한
생태계, 기능을 갖는 것이다. Miyoshi 등 (1990) 은 갯벌의 정화기능의 개념을 제시하
고 인공갯벌과 자연갯벌의 차이를 살펴보았다. 유럽의 경우 갯벌의 생태 및 지화학적
특성에 대한 연구가 활발하다.
북유럽의 바덴해에 연한 독일, 덴마크, 네덜란드 세 나라가 공동으로 추진하고 있는
연안습지 보존계획은 가장 모범적인 성공사례로 주목받고 있다. 1978년에 이 세 나라가
힘을 모아 “The Trilateral Cooperation on the Protection of the Wadden Sea"를 체결
하고 이행한지 25년이 지났다. 자국의 습지조차 개발과 보전의 날카로운 대립을 보이고
있는 우리나라와는 사뭇 다른 부러운 상황이 아닐 수 없다. 바덴해의 거의 모든 해안을
국립공원으로 지정되었다. 드센 개발 압력에도 불구하고 보호가 장기적 안목에서 더 큰
이익을 창출한다는 점을 과학적으로 입증해 보인 것은 실로 대단한 성과이다. 또한 오
랜 정부지원 연구에 힘입어 이들 세 나라가 보유하고 있는 습지연구능력은 가장 우수
한 것으로서 일부만 소개 하자면 퇴적물을 교란시키지 않고 떠내는 방법, 아주 미세한
산소 전극으로 mm 이하 간격으로 산소의 농도변화를 측정하는 기술, 산소요구량

- 40 -
(SOD: sediment oxygen demand)을 정밀하게 측정하는 기술 등이 연구와 긴밀하게 관
련된 첨단 기술을 보유하고 있고 현재도 개발 중이다. 2002년도에는 한국과학재단과 독
일연구협회가 공동으로 황해와 북해의 갯벌 생태학이란 주제로 심포지움을 개최하여
갯벌의 과학을 한 단계 높이려는 노력이 시도된 바 있다. 한편 2003년 5월에 독일에서
개최되었던 워크샵 “Biogeochemistry of Tidal Flats"에서 발표된 연구 내용들의 면면
을 살펴보면 우리나라와의 학문적 격차를 절실하게 느끼게 한다. 이 연구와 관련해서
TRAP (Trilateral Monitoring and Assessment Program)이 제공하고 있는 연구 매뉴
얼은 이 연구에서 어떤 자료를 어떤 방식으로 획득해야 하는지에 대해 아주 훌륭한 지
침이 될 것이다.
제 2절 국내의 연안 하구 습지 연구
새만금 논란이후 갯벌 등 연안 습지의 중요성을 인식하여 갯벌의 기능과 생태계에
관한 연구가 시작되고 있으나(과학기술처, 1989-1991; 한국해양연구소, 1997-1999, 해양
수산부 1999 - 2003, 환경부 1998, 1998-2001) 현장 접근성이 연구의 가장 큰 걸림돌로
여겨진다. 현장에서 재려면 휴대용 계측장비를 가지고 들어가야 하는데 특히 발이 깊이
빠지는 펄 갯벌 같은 경우에는 운신조차 어렵기 때문이다. 운송수단을 개발하기 전 까
지는 현장 계측은 여전히 커다란 진전을 기대하기 어려운 실정이다. 습지의 지화학적
특성이 생물상 또는 생태학과 연계되어 실시된 적은 없다고 보아도 무방하다. 연안 습
지의 환경 특성은 석사논문 지역의 환경을 개괄하는 수준에서 조사된 것이 대부분이고,
국내 학술지에 발표된 관련 논문도 한 편에 불과하다 (최 등, 2000). 유럽의 선진국에서
해양습지에 대해 퇴적물 플럭스를 전공한 전문가는 국내에 없는 것으로 알고 있다. 다
만 이 분야의 중요성을 인식해서 석사 학위논문으로 다루어 진 예가 있고 가장 최근에
있었던 관련 연구투자로는 해양수산부가 지원했던 과제로 “갯벌에서 미생물에 의한 생
지화학적 오염정화능 판정기술 개발“이 있다.
갯벌의 자연정화기능 개념에 혼란이 있었으나, 본 연구팀에서 탈질소화, 생태계를 통
한 계 밖으로의 제거 정화기능임이 밝혀져 이에 대한 연구가 진행되었다 (한국과학재
단 2001-2004). 갯벌 퇴적물의 산소요구량과 탈질소화 기타 미생물과정을 실측과 모델
링을 통해 알아보려는 연구가 진행되었다(해양수산부, “갯벌에서 미생물에 의한 생지화
학적 오염정화능 판정기술 개발“). 2003년부터 해양수산부 지원에 의해 ”대체 습지 조
성 중장기계획수립“연구가 진행 중이다. 국내에서 연안 습지가 복원된 예는 없으나, 육
상의 인공습지는 수질 정화와 습지 생태계 복원을 목적으로 조성된 사례가 있다. 대규
모 사례로서는 한국 수자원 공사가 시공한 시화호 갈대 공원이 있는데, 축산 및 생활하
수를 정화하기 위한 목적으로 약 24만평 규모의 인공습지를 조성하였다. 연안의 인공
습지와는 다른 개념이나 당진군 대호지구 매립 지역 약 800ha를 친 환경적 (환경농업
실천, 환경 친화적이용, 농업교육훈련센터 건립, 농어촌 주택단지, 농어민 교육. 복지센
터 등)으로 이용하기 위한 노력이 진행되고 있다. 조하대 잘피림 조성 연구의 경우 거

- 41 -
의 이루어지지 않았으나, 본 연구팀에서 현재 적정 이식지 선정 방법, 환경차이에 따른
적합 이식방법 개발, 잠수시간을 줄이는 이식방법개발 등에 노력을 기울이고 있다
제 3절 국내외 기술과의 차별성
갯벌 등 연안 습지에 관한 관심이 높아져, 여러 측면에서 연구가 시작되고 있으나,
산발적으로 현상을 파악하는 연구가 주를 이루고 있다. 최근에 시작된 해양연구원의
“대체습지 조성 중장기 계획수립” 연구가 연안 습지 복원의 다양한 측면을 연구하고
있으나, 현재 기초 조사와 문헌 조사 수준에 있으며, 실제 실험용 복원 습지 조성은 불
투명하다. 2002년 이후 사회적으로 큰 논란이 된 새만금 갯벌 및 인근 해양생태계연구
가 해양수산부에서 실시되고 있다. 관심이 큰 공사인 만큼 풍부한 인력과 자원으로 조
사가 이루어지고 있으나, 정책 결정에 필요한 자료를 생산하는데 초점이 맞춰져 있어,
방조제공사로 인한 해양환경악화, 경제적 손실 등에 집중된 연구를 실시하고 있다. 연
구범위도 공사에 따른 환경과 생태계의 구조, 즉 각 생태요소의 양적변화에 초점이 맞
춰져 있으며, 생태기능, 즉 각 생태요소간의 상호 관계 및 상대적인 중요성에 관한 연
구는 없는 실정이다.
본 연구는 자연 연안 습지와 함께 다양한 조건의 실험용 연안습지를 조성하여 비교
분석함으로서 국내 여건에 맞는 연안 습지 복원 파라미터를 찾는 것을 목적으로 하고
있다. 국내에서 연구 개발이 미비한 분야인 저서생태계의 생태, 물질순환 전문가가 연
구팀의 주요 구성원이 되어, 토목공학적인 습지 조성 조건보다는 연안습지의 “건강성”,
“기능성”을 극대화하는 조건을 찾는 연구를 수행하였다. 최근에 연안 생태 분야에 도입
된 첨단 장비 및 기술 (MIMS; Membrane inlet mass spectrometer, 안정동위원소 추적
자, Sediment oxygen demand, 잘피 이식 기술 등)을 이용함으로서 첨단 연구를 진행
하였으며, 이러한 연구가 동시에 진행된 것은 선진국에서도 유래를 찾아볼 수 없다.
본 연구의 결과의 많은 부분은 낙동강 하구에서는 물론 국내 어떤 하구에서도 생산된
적이 없는 첨단 연구 자료로서 향후 국내 연안 생태계 연구의 시금석이 될 것이다.

- 42 -

- 43 -
제 3장 연구개발 수행 내용 및 결과

- 44 -

- 45 -
제 3장 연구개발 수행 내용 및 결과
제 1절 낙동강 하구의 환경 특성
1. 퇴적물 특성
가. 입도 및 퇴적률
(1) 낙동강 하구 갯벌의 퇴적물 입도
퇴적물의 입도분석은 퇴적물에 과산화수소(H2O2)와 염산(HCl)을 넣어 유기물과 탄산
염을 제거한 후 wet sieving에 의해 4Φ으로 퇴적물을 분류하였다. 4Φ이하 조립질 퇴적
물은 sieve shaker로 체질한 후 0.5Φ 간격으로 무게 백분율을 구하였고, 4Φ이상 세립질
퇴적물은 본 연구지역에서는 그 양이 매우 작아 더 이상 퇴적물 분류를 하지 않았다.
퇴적물의 입도 특성을 나타내는 평균입도, 분급도, 왜도, 첨도를 구하였고, 퇴적물의 분
류는 Folk and Word(1957)의 방법을 따랐다. 낙동강하구는 0.4~1.5m의 간조차로 인해
간조시에는 넓은 갯벌을 이루고 만조시에는 바다에 잠기는 갯벌의 형태를 취하고 있다.
실험 결과, 대부분이 사질 퇴적물로 구성되어 있고, 명지주거단지 일대는 다른 연구지
역보다 미사점토 함량이 다소 높게 구성되어 있었다(Table. 1-1).
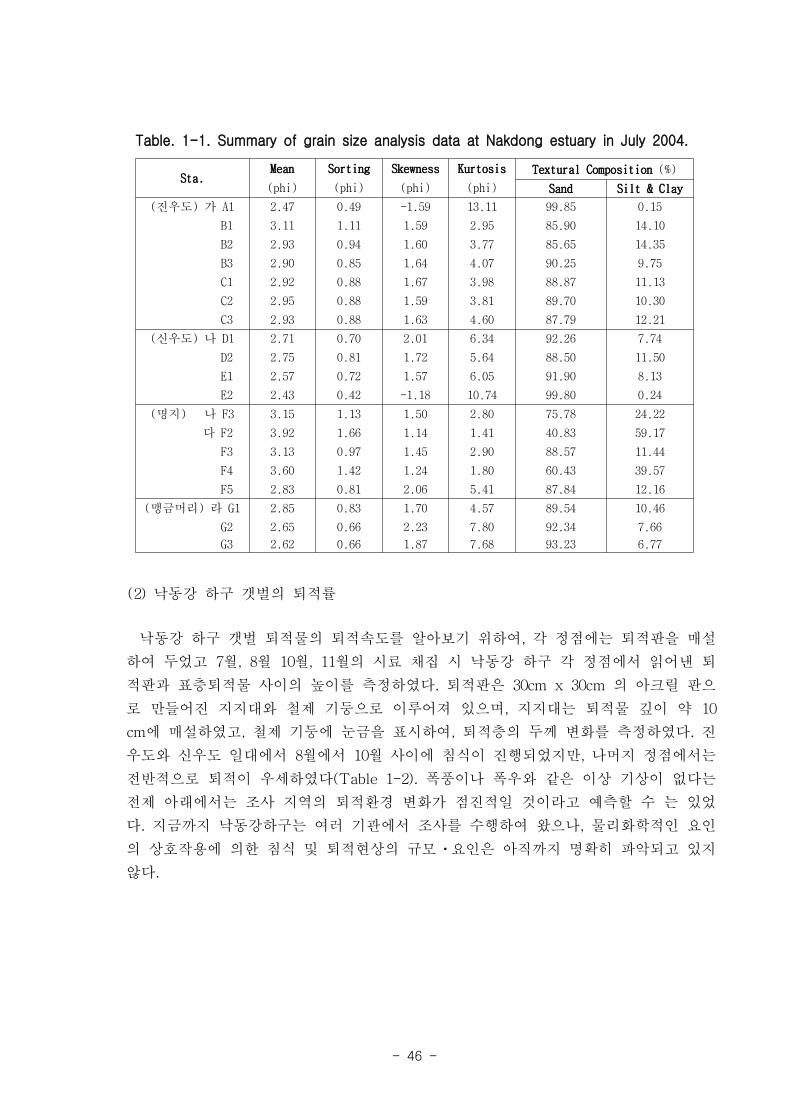
① 전체적으로 모래 퇴적물이 90%이상 우세하였으나, 명지주거단지 정점의 경우 실
트-점토 퇴적물이 다른 정점에서보다 10∼40% 이상 높게 나타났다(Fig. 1-1).
② 표층퇴적물의 평균입도는 2.6Φ~3.9Φ로 slightly muddy sand로 나타났고, 명지주
거단지 정점에서 다른 정점에 비해 다소 세립한 입도를 보였으나, 전체적으로 정점에
따른 평균입도의 큰 차이는 보이지 않았다.
Figure. 1-1. Location map of study area in the
Nakdong estuary.
- 46 -
Sta.Mean
(phi)
Sorting
(phi)
Skewness
(phi)
Kurtosis
(phi)
Textural Composition (%)
Sand Silt & Clay
(진우도) 가 A1 2.47 0.49 -1.59 13.11 99.85 0.15
B1 3.11 1.11 1.59 2.95 85.90 14.10
B2 2.93 0.94 1.60 3.77 85.65 14.35
B3 2.90 0.85 1.64 4.07 90.25 9.75
C1 2.92 0.88 1.67 3.98 88.87 11.13
C2 2.95 0.88 1.59 3.81 89.70 10.30
C3 2.93 0.88 1.63 4.60 87.79 12.21
(신우도) 나 D1 2.71 0.70 2.01 6.34 92.26 7.74
D2 2.75 0.81 1.72 5.64 88.50 11.50
E1 2.57 0.72 1.57 6.05 91.90 8.13
E2 2.43 0.42 -1.18 10.74 99.80 0.24
(명지) 나 F3 3.15 1.13 1.50 2.80 75.78 24.22
다 F2 3.92 1.66 1.14 1.41 40.83 59.17
F3 3.13 0.97 1.45 2.90 88.57 11.44
F4 3.60 1.42 1.24 1.80 60.43 39.57
F5 2.83 0.81 2.06 5.41 87.84 12.16
(맹금머리) 라 G1 2.85 0.83 1.70 4.57 89.54 10.46
G2 2.65 0.66 2.23 7.80 92.34 7.66
G3 2.62 0.66 1.87 7.68 93.23 6.77
Table. 1-1. Summary of grain size analysis data at Nakdong estuary in July 2004.
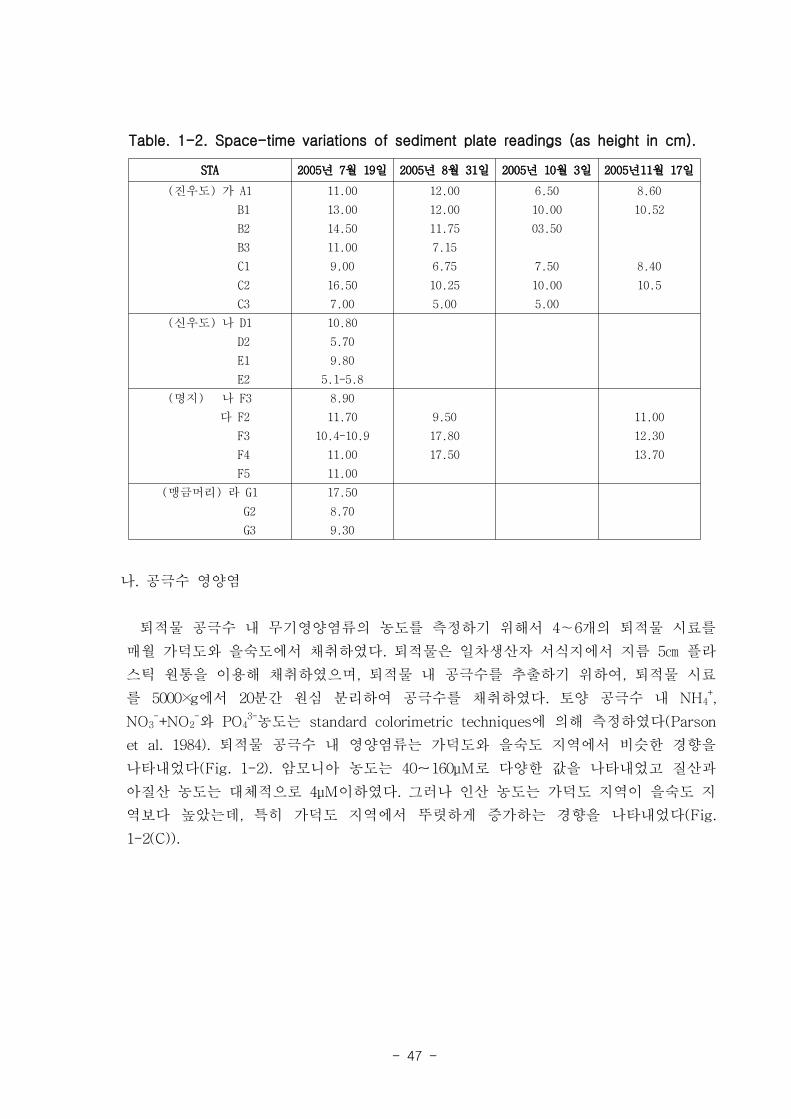
(2) 낙동강 하구 갯벌의 퇴적률
낙동강 하구 갯벌 퇴적물의 퇴적속도를 알아보기 위하여, 각 정점에는 퇴적판을 매설
하여 두었고 7월, 8월 10월, 11월의 시료 채집 시 낙동강 하구 각 정점에서 읽어낸 퇴
적판과 표층퇴적물 사이의 높이를 측정하였다. 퇴적판은 30cm x 30cm 의 아크릴 판으
로 만들어진 지지대와 철제 기둥으로 이루어져 있으며, 지지대는 퇴적물 깊이 약 10
cm에 매설하였고, 철제 기둥에 눈금을 표시하여, 퇴적층의 두께 변화를 측정하였다. 진
우도와 신우도 일대에서 8월에서 10월 사이에 침식이 진행되었지만, 나머지 정점에서는
전반적으로 퇴적이 우세하였다(Table 1-2). 폭풍이나 폭우와 같은 이상 기상이 없다는
전제 아래에서는 조사 지역의 퇴적환경 변화가 점진적일 것이라고 예측할 수 는 있었
다. 지금까지 낙동강하구는 여러 기관에서 조사를 수행하여 왔으나, 물리화학적인 요인
의 상호작용에 의한 침식 및 퇴적현상의 규모 • 요인은 아직까지 명확히 파악되고 있지
않다.
- 47 -
STA 2005년 7월 19일 2005년 8월 31일 2005년 10월 3일 2005년11월 17일
(진우도) 가 A1 11.00 12.00 6.50 8.60
B1 13.00 12.00 10.00 10.52
B2 14.50 11.75 03.50
B3 11.00 7.15
C1 9.00 6.75 7.50 8.40
C2 16.50 10.25 10.00 10.5
C3 7.00 5.00 5.00
(신우도) 나 D1 10.80
D2 5.70
E1 9.80
E2 5.1-5.8
(명지) 나 F3 8.90
다 F2 11.70 9.50 11.00
F3 10.4-10.9 17.80 12.30
F4 11.00 17.50 13.70
F5 11.00
(맹금머리) 라 G1 17.50
G2 8.70
G3 9.30
Table. 1-2. Space-time variations of sediment plate readings (as height in cm).
나. 공극수 영양염
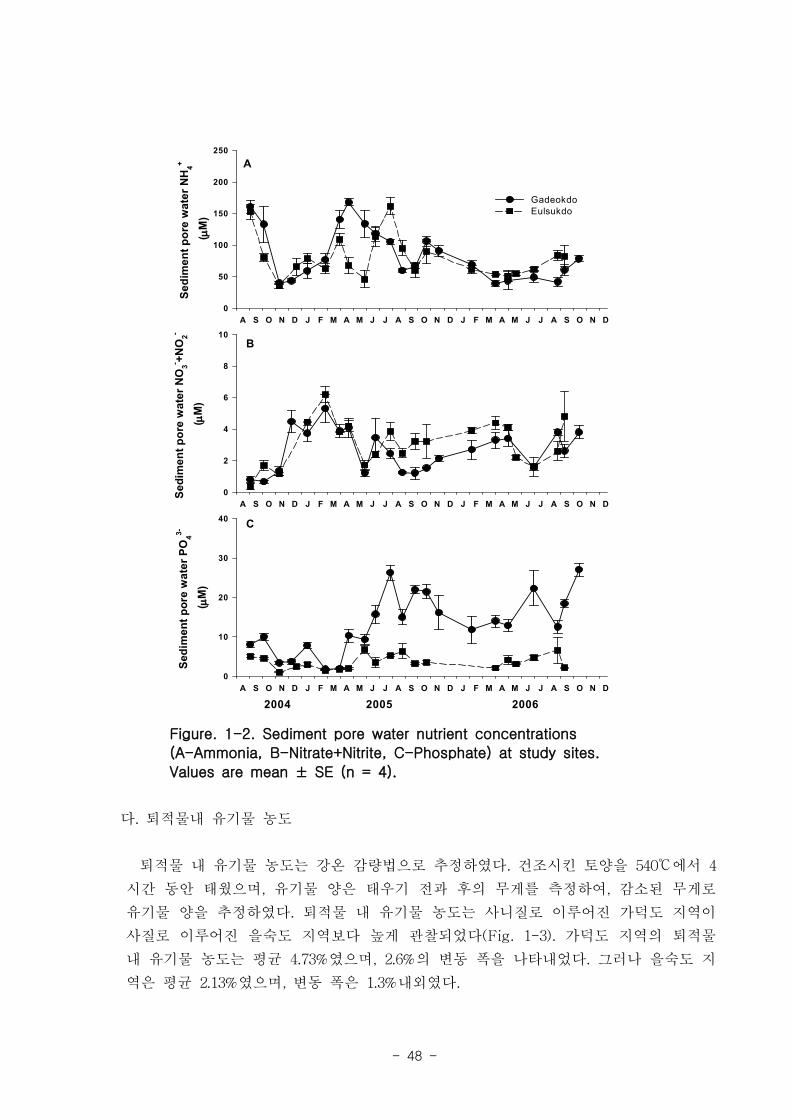
퇴적물 공극수 내 무기영양염류의 농도를 측정하기 위해서 4∼6개의 퇴적물 시료를
매월 가덕도와 을숙도에서 채취하였다. 퇴적물은 일차생산자 서식지에서 지름 5㎝ 플라
스틱 원통을 이용해 채취하였으며, 퇴적물 내 공극수를 추출하기 위하여, 퇴적물 시료
를 5000×g에서 20분간 원심 분리하여 공극수를 채취하였다. 토양 공극수 내 NH4+,
NO3-+NO2
-와 PO43-농도는 standard colorimetric techniques에 의해 측정하였다(Parson
et al. 1984). 퇴적물 공극수 내 영양염류는 가덕도와 을숙도 지역에서 비슷한 경향을
나타내었다(Fig. 1-2). 암모니아 농도는 40~160μM로 다양한 값을 나타내었고 질산과
아질산 농도는 대체적으로 4μM이하였다. 그러나 인산 농도는 가덕도 지역이 을숙도 지
역보다 높았는데, 특히 가덕도 지역에서 뚜렷하게 증가하는 경향을 나타내었다(Fig.
1-2(C)).
- 48 -
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Sedim
ent pore
wate
r N
H4
+
( µM
)
0
50
100
150
200
250
Gadeokdo
Eulsukdo
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Sedim
ent pore
wate
r N
O3
- +N
O2
-
( µM
)
0
2
4
6
8
10
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Sedim
ent pore
wate
r P
O4
3-
( µM
)
0
10
20
30
40
A
B
C
Figure. 1-2. Sediment pore water nutrient concentrations
(A-Ammonia, B-Nitrate+Nitrite, C-Phosphate) at study sites.
Values are mean ± SE (n = 4).
다. 퇴적물내 유기물 농도
퇴적물 내 유기물 농도는 강온 감량법으로 추정하였다. 건조시킨 토양을 540℃에서 4
시간 동안 태웠으며, 유기물 양은 태우기 전과 후의 무게를 측정하여, 감소된 무게로
유기물 양을 추정하였다. 퇴적물 내 유기물 농도는 사니질로 이루어진 가덕도 지역이
사질로 이루어진 을숙도 지역보다 높게 관찰되었다(Fig. 1-3). 가덕도 지역의 퇴적물
내 유기물 농도는 평균 4.73%였으며, 2.6%의 변동 폭을 나타내었다. 그러나 을숙도 지
역은 평균 2.13%였으며, 변동 폭은 1.3%내외였다.

- 49 -
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Sedim
ent org
anic
conte
nt (%
)
0
1
2
3
4
5
6
7 Gadeokdo
Eulsukdo
Figure. 1-3. Sediment organic content at study sites from
August 2004 to September 2006. Values are mean ± SE (n
= 4).
라. 조고 및 노출 특성
(1) DGPS를 이용한 낙동강 하구역 정밀 지형 파악
갯벌의 조위는 특정지역의 물리화학적 환경의 차이를 대변하는 가장 중요한 환경조
건이다. 본 조사에서는 1cm 이하의 정밀도를 갖는 DGPS를 이용하여, 낙동강 하구 각
지역의 조위를 정밀 측정하였다.
Figure. 1-4. DGPS 기지국 설치 및 측정 (2004년 8월 31일, 낙동강 갯벌)

- 50 -
st_name lat(°N)-DGPS long(°E)-DGPS altitude(cm)
진우도 A1 35.07115928 128.86067740
진우도B1 35.07002179 128.86605589 167.100
진우도B2 35.07124301 128.86621895 154.700
진우도B3 35.07300358 128.86679675 135.900
진우도C3 35.07166973 128.87017332 148.100
진우도C2 35.07082458 128.87008660 172.500
진우도C1 35.06979912 128.87007155 189.100
add(1) 35.06946613 128.87348475 186.500
add(2) 35.07122526 128.87414726 131.300
add(3) 35.07465718 128.87598303 139.400
명지 F4 35.07567683 128.93029837 113.500
명지 F3 35.07865932 128.93014579 112.500
명지 F2 35.08147958 128.93078898 127.000
Table. 1-3. Summary DGPS measurement at Nakdong
estuary in Aug. Sept 2004.
조사지역은 평균 해수면 위 112.5~189.1cm 에 걸쳐 위치하였다. 조사 정점 중 진우
도의 가 C1 정점이 가장 높았고 명지의 다 F3 정점이 가장 낮았다 (Table. 1-3).
(2) 각 정점의 연속노출 분석
각 조사정점이 1년 중 얼마만큼 침수되고 얼마만큼 노출되는지를 알아보기 위하여,
인근 가덕도에서 2002년에 실측된 조석 자료를 분석하였다. 각 조고에 따라 1년중 노출
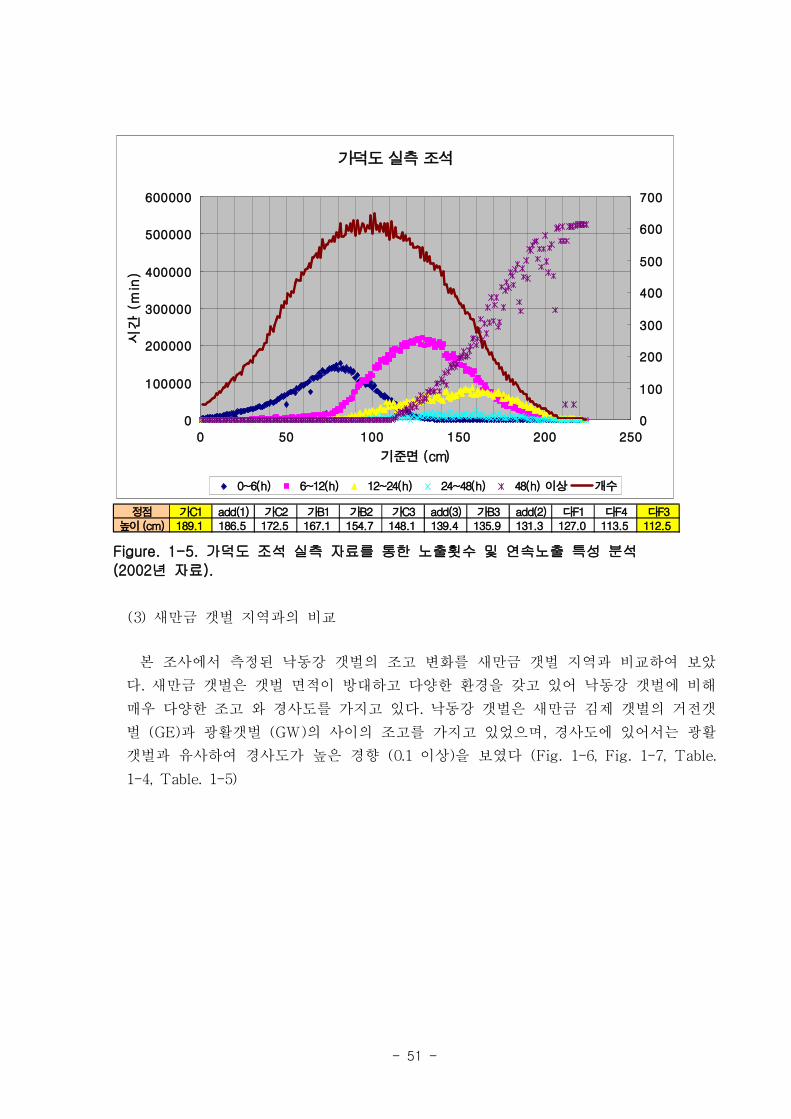
되는 횟수와 연속노출되는 시간을 분석하였다. 노출되는 횟수는 기준면으로부터 100cm
정도에서 가장 높아 650회에 달했는데 이는 1년 365일중 매일 2차례씩 노출됨을 의미
한다(Fig. 1-5). 100cm 보다 낮은 조고와 높은 조고에서는 노출 횟수가 각가 감소하였
다. 그러나 노출 시간은 조고에 따라 연속적으로 증가하였다. 노출시간의 질, 즉 연속노
출을 각 조고에 따라 살펴보면, 조고 0에서 70cm 까지는 0~6시간 연속노출만 일어났
고 70cm이상의 조고에서 6시간이상의 연속노출이 일어나기 시작했다. 12시간 이상의
연속노출은 90cm이상에서 관찰 되었으며, 48시간의 연속노출은 110cm이상의 조고에서
나타났다.
- 51 -
가덕도 실측 조석
0
100000
200000
300000
400000
500000
600000
0 50 100 150 200 250
기준면 (cm)
시간
(m
in)
0
100
200
300
400
500
600
700
0~6(h) 6~12(h) 12~24(h) 24~48(h) 48(h) 이상 개수
정점 가C1 add(1) 가C2 가B1 가B2 가C3 add(3) 가B3 add(2) 다F1 다F4 다F3높이 (cm) 189.1 186.5 172.5 167.1 154.7 148.1 139.4 135.9 131.3 127.0 113.5 112.5
Figure. 1-5. 가덕도 조석 실측 자료를 통한 노출횟수 및 연속노출 특성 분석
(2002년 자료).
(3) 새만금 갯벌 지역과의 비교
본 조사에서 측정된 낙동강 갯벌의 조고 변화를 새만금 갯벌 지역과 비교하여 보았
다. 새만금 갯벌은 갯벌 면적이 방대하고 다양한 환경을 갖고 있어 낙동강 갯벌에 비해
매우 다양한 조고 와 경사도를 가지고 있다. 낙동강 갯벌은 새만금 김제 갯벌의 거전갯
벌 (GE)과 광활갯벌 (GW)의 사이의 조고를 가지고 있었으며, 경사도에 있어서는 광활
갯벌과 유사하여 경사도가 높은 경향 (0.1 이상)을 보였다 (Fig. 1-6, Fig. 1-7, Table.
1-4, Table. 1-5)
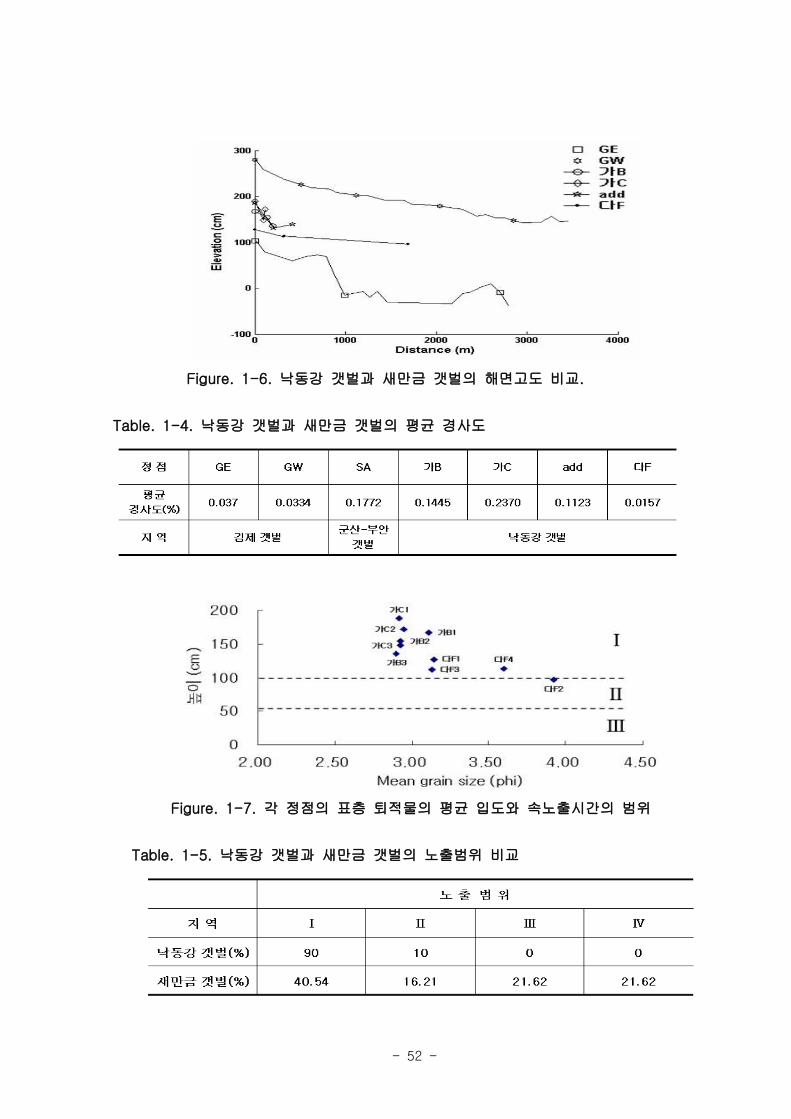
- 52 -
Figure. 1-6. 낙동강 갯벌과 새만금 갯벌의 해면고도 비교.
Table. 1-4. 낙동강 갯벌과 새만금 갯벌의 평균 경사도
Figure. 1-7. 각 정점의 표층 퇴적물의 평균 입도와 속노출시간의 범위
Table. 1-5. 낙동강 갯벌과 새만금 갯벌의 노출범위 비교
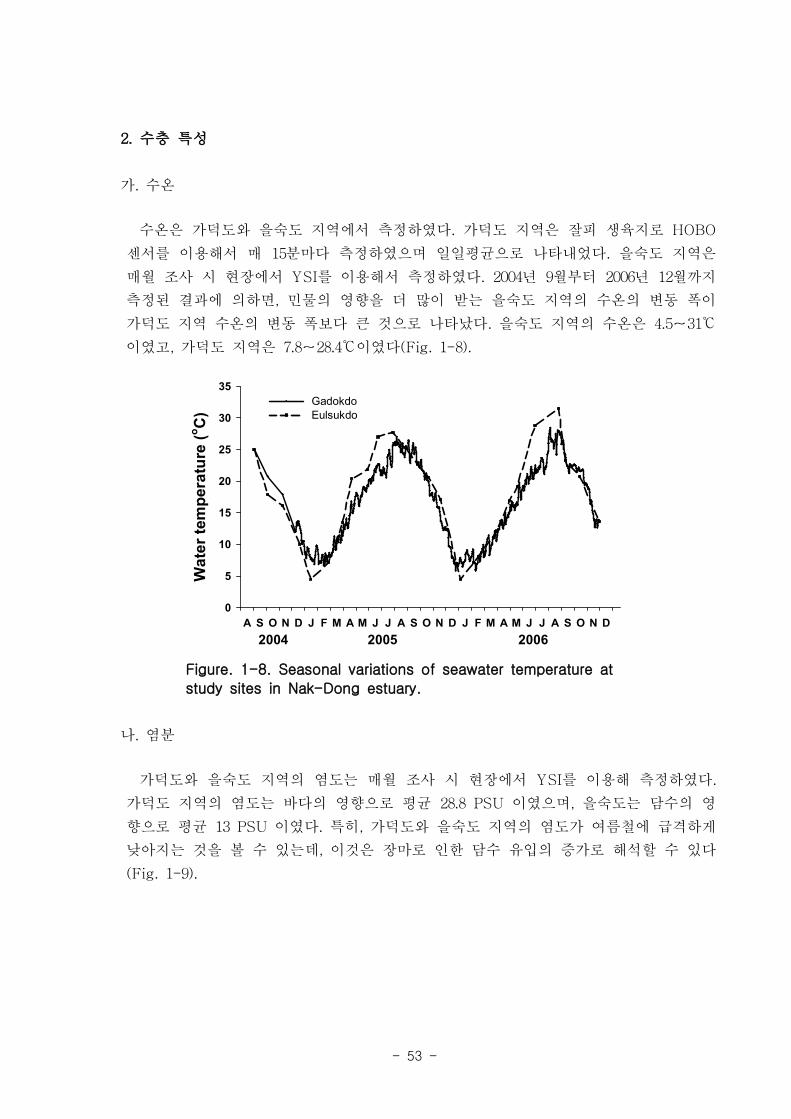
- 53 -
2. 수층 특성
가. 수온
수온은 가덕도와 을숙도 지역에서 측정하였다. 가덕도 지역은 잘피 생육지로 HOBO
센서를 이용해서 매 15분마다 측정하였으며 일일평균으로 나타내었다. 을숙도 지역은
매월 조사 시 현장에서 YSI를 이용해서 측정하였다. 2004년 9월부터 2006년 12월까지
측정된 결과에 의하면, 민물의 영향을 더 많이 받는 을숙도 지역의 수온의 변동 폭이
가덕도 지역 수온의 변동 폭보다 큰 것으로 나타났다. 을숙도 지역의 수온은 4.5~31℃이였고, 가덕도 지역은 7.8~28.4℃이였다(Fig. 1-8).
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Wate
r te
mpera
ture
(oC
)
0
5
10
15
20
25
30
35
Gadokdo
Eulsukdo
Figure. 1-8. Seasonal variations of seawater temperature at
study sites in Nak-Dong estuary.
나. 염분
가덕도와 을숙도 지역의 염도는 매월 조사 시 현장에서 YSI를 이용해 측정하였다.
가덕도 지역의 염도는 바다의 영향으로 평균 28.8 PSU 이였으며, 을숙도는 담수의 영
향으로 평균 13 PSU 이였다. 특히, 가덕도와 을숙도 지역의 염도가 여름철에 급격하게
낮아지는 것을 볼 수 있는데, 이것은 장마로 인한 담수 유입의 증가로 해석할 수 있다
(Fig. 1-9).
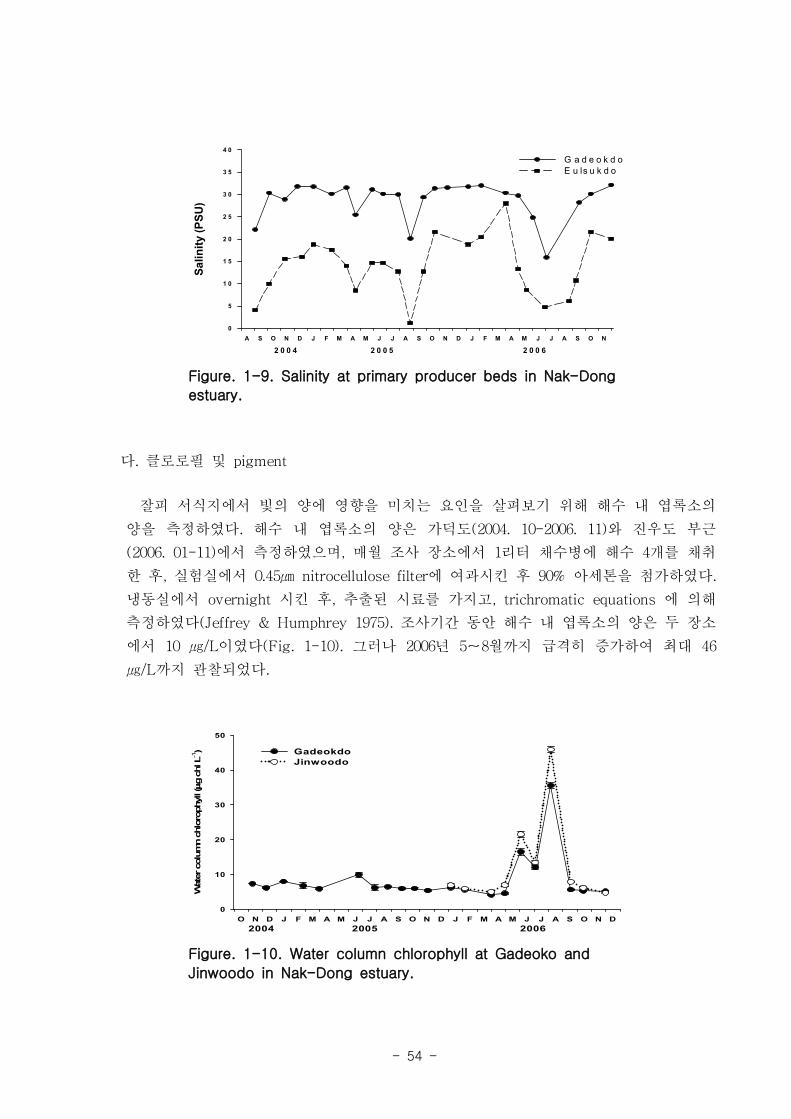
- 54 -
2 0 0 4 2 0 0 5 2 0 0 6
A S O N D J F M A M J J A S O N D J F M A M J J A S O N
Salinity (PSU
)
0
5
1 0
1 5
2 0
2 5
3 0
3 5
4 0
G a d e o k d o
E u ls u k d o
Figure. 1-9. Salinity at primary producer beds in Nak-Dong
estuary.
다. 클로로필 및 pigment
잘피 서식지에서 빛의 양에 영향을 미치는 요인을 살펴보기 위해 해수 내 엽록소의
양을 측정하였다. 해수 내 엽록소의 양은 가덕도(2004. 10-2006. 11)와 진우도 부근
(2006. 01-11)에서 측정하였으며, 매월 조사 장소에서 1리터 채수병에 해수 4개를 채취
한 후, 실험실에서 0.45㎛ nitrocellulose filter에 여과시킨 후 90% 아세톤을 첨가하였다.
냉동실에서 overnight 시킨 후, 추출된 시료를 가지고, trichromatic equations 에 의해
측정하였다(Jeffrey & Humphrey 1975). 조사기간 동안 해수 내 엽록소의 양은 두 장소
에서 10 ㎍/L이였다(Fig. 1-10). 그러나 2006년 5~8월까지 급격히 증가하여 최대 46
㎍/L까지 관찰되었다.
2004 2005 2006
O N D J F M A M J J A S O N D J F M A M J J A S O N D
Wate
r colu
mn chloro
phyll (µg chl L-1)
0
10
20
30
40
50
Gadeokdo
Jinwoodo
Figure. 1-10. Water column chlorophyll at Gadeoko and
Jinwoodo in Nak-Dong estuary.
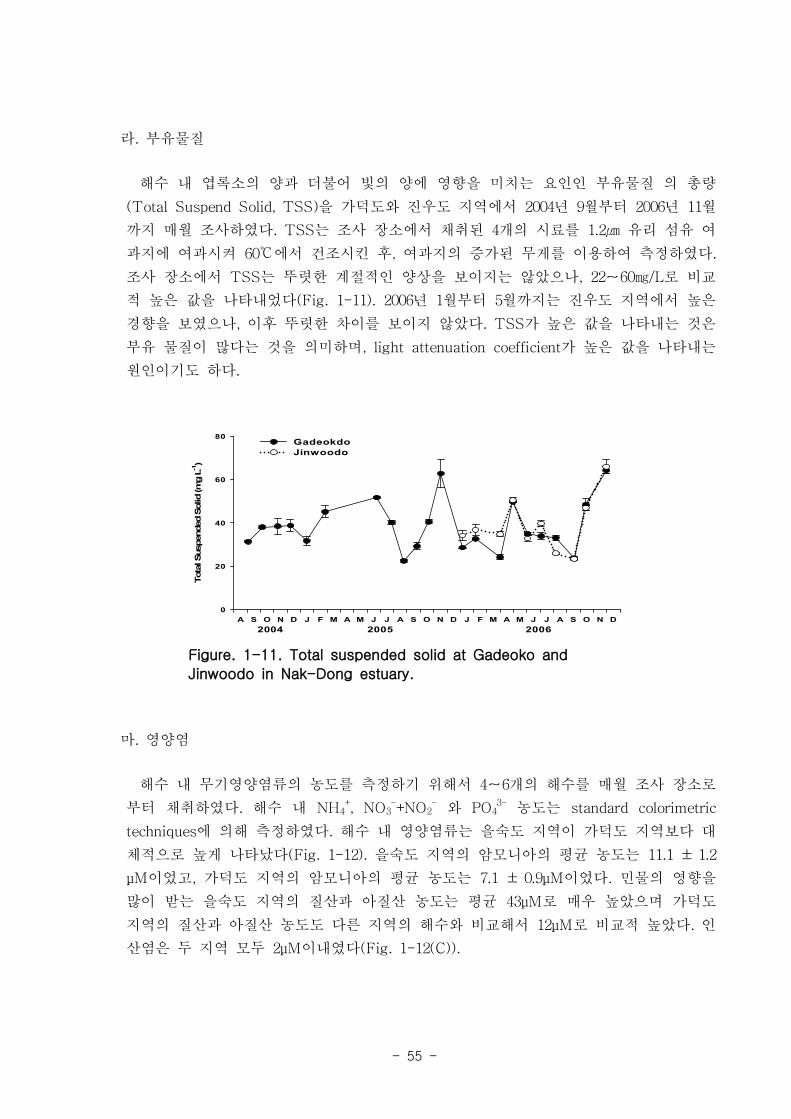
- 55 -
라. 부유물질
해수 내 엽록소의 양과 더불어 빛의 양에 영향을 미치는 요인인 부유물질 의 총량
(Total Suspend Solid, TSS)을 가덕도와 진우도 지역에서 2004년 9월부터 2006년 11월
까지 매월 조사하였다. TSS는 조사 장소에서 채취된 4개의 시료를 1.2㎛ 유리 섬유 여
과지에 여과시켜 60℃에서 건조시킨 후, 여과지의 증가된 무게를 이용하여 측정하였다.
조사 장소에서 TSS는 뚜렷한 계절적인 양상을 보이지는 않았으나, 22~60㎎/L로 비교
적 높은 값을 나타내었다(Fig. 1-11). 2006년 1월부터 5월까지는 진우도 지역에서 높은
경향을 보였으나, 이후 뚜렷한 차이를 보이지 않았다. TSS가 높은 값을 나타내는 것은
부유 물질이 많다는 것을 의미하며, light attenuation coefficient가 높은 값을 나타내는
원인이기도 하다.
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Tota
l Suspended S
olid (mg L
-1)
0
20
40
60
80Gadeokdo
Jinwoodo
Figure. 1-11. Total suspended solid at Gadeoko and
Jinwoodo in Nak-Dong estuary.
마. 영양염
해수 내 무기영양염류의 농도를 측정하기 위해서 4~6개의 해수를 매월 조사 장소로
부터 채취하였다. 해수 내 NH4+, NO3
-+NO2- 와 PO4
3- 농도는 standard colorimetric
techniques에 의해 측정하였다. 해수 내 영양염류는 을숙도 지역이 가덕도 지역보다 대
체적으로 높게 나타났다(Fig. 1-12). 을숙도 지역의 암모니아의 평균 농도는 11.1 ± 1.2
μM이었고, 가덕도 지역의 암모니아의 평균 농도는 7.1 ± 0.9μM이었다. 민물의 영향을
많이 받는 을숙도 지역의 질산과 아질산 농도는 평균 43μM로 매우 높았으며 가덕도
지역의 질산과 아질산 농도도 다른 지역의 해수와 비교해서 12μM로 비교적 높았다. 인
산염은 두 지역 모두 2μM이내였다(Fig. 1-12(C)).
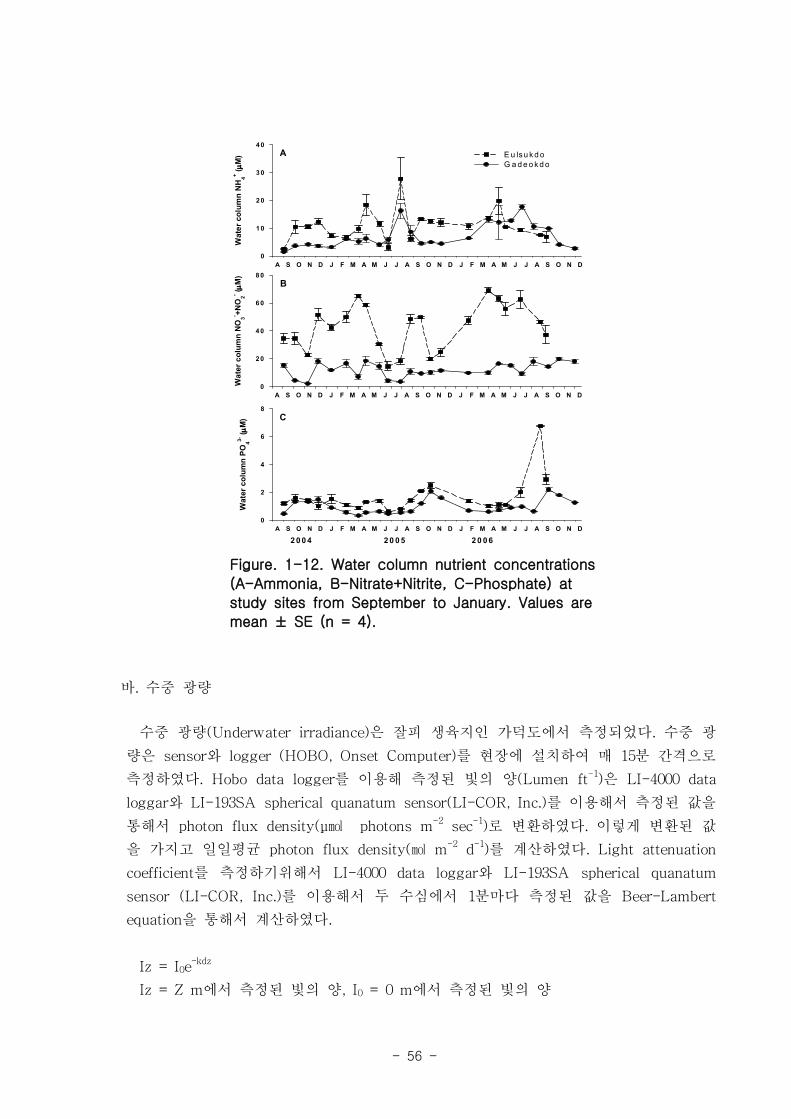
- 56 -
A
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Wate
r colu
mn N
H4
+ (µM
)
0
10
20
30
40
E u lsu kdo
G adeokdo
B
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Wate
r colu
mn N
O3
- +N
O2
- (µM
)
0
20
40
60
80
C
2004 2 0 05 2 0 06
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Wate
r colu
mn P
O4
3- (µM
)
0
2
4
6
8
Figure. 1-12. Water column nutrient concentrations
(A-Ammonia, B-Nitrate+Nitrite, C-Phosphate) at
study sites from September to January. Values are
mean ± SE (n = 4).
바. 수중 광량
수중 광량(Underwater irradiance)은 잘피 생육지인 가덕도에서 측정되었다. 수중 광
량은 sensor와 logger (HOBO, Onset Computer)를 현장에 설치하여 매 15분 간격으로
측정하였다. Hobo data logger를 이용해 측정된 빛의 양(Lumen ft-1
)은 LI-4000 data
loggar와 LI-193SA spherical quanatum sensor(LI-COR, Inc.)를 이용해서 측정된 값을
통해서 photon flux density(μ㏖ photons m-2 sec
-1)로 변환하였다. 이렇게 변환된 값
을 가지고 일일평균 photon flux density(㏖ m-2 d
-1)를 계산하였다. Light attenuation
coefficient를 측정하기위해서 LI-4000 data loggar와 LI-193SA spherical quanatum
sensor (LI-COR, Inc.)를 이용해서 두 수심에서 1분마다 측정된 값을 Beer-Lambert
equation을 통해서 계산하였다.
Iz = I0e-kdz
Iz = Z m에서 측정된 빛의 양, I0 = 0 m에서 측정된 빛의 양
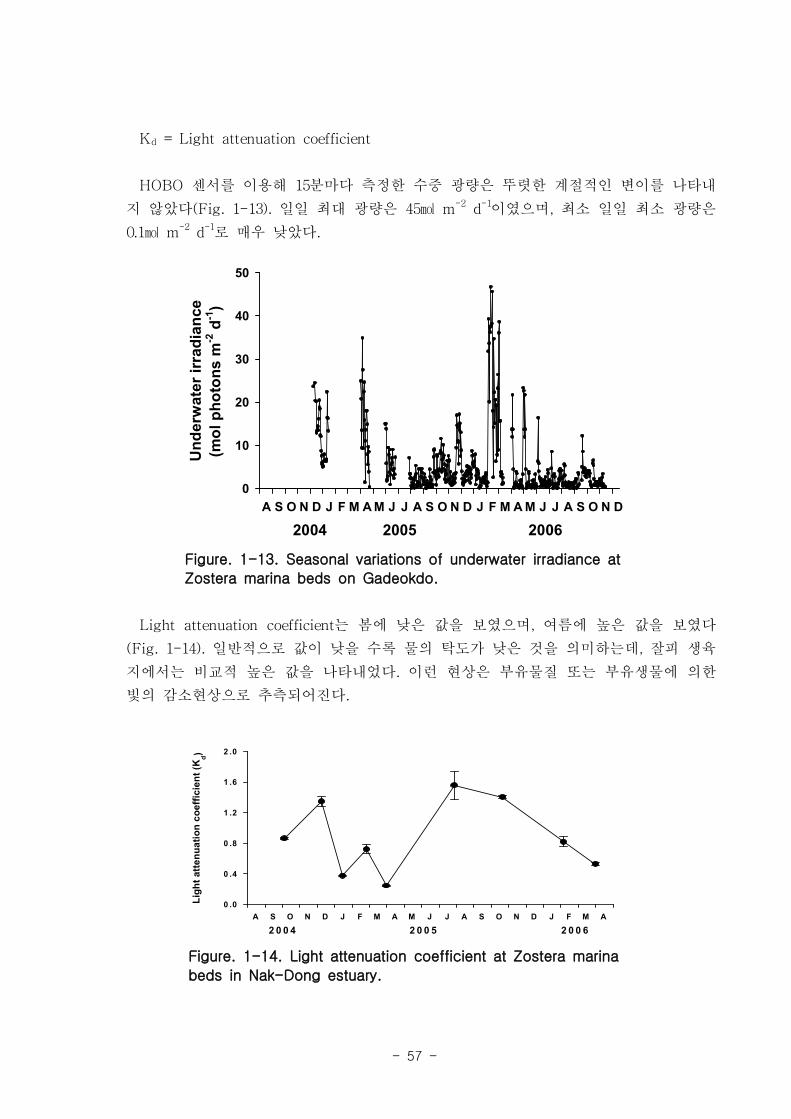
- 57 -
Kd = Light attenuation coefficient
HOBO 센서를 이용해 15분마다 측정한 수중 광량은 뚜렷한 계절적인 변이를 나타내
지 않았다(Fig. 1-13). 일일 최대 광량은 45㏖ m-2 d
-1이였으며, 최소 일일 최소 광량은
0.1㏖ m-2 d
-1로 매우 낮았다.
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Underw
ate
r ir
radia
nce
(m
ol photo
ns m
-2 d
-1)
0
10
20
30
40
50
Figure. 1-13. Seasonal variations of underwater irradiance at
Zostera marina beds on Gadeokdo.
Light attenuation coefficient는 봄에 낮은 값을 보였으며, 여름에 높은 값을 보였다
(Fig. 1-14). 일반적으로 값이 낮을 수록 물의 탁도가 낮은 것을 의미하는데, 잘피 생육
지에서는 비교적 높은 값을 나타내었다. 이런 현상은 부유물질 또는 부유생물에 의한
빛의 감소현상으로 추측되어진다.
2 0 0 4 2 0 0 5 2 0 0 6
A S O N D J F M A M J J A S O N D J F M A
Lig
ht att
enuation c
oeff
icie
nt (K
d)
0 .0
0 .4
0 .8
1 .2
1 .6
2 .0
Figure. 1-14. Light attenuation coefficient at Zostera marina
beds in Nak-Dong estuary.

- 58 -
제 2절 낙동강 하구의 생태 구성 요소
1. 일차 생산자
가. 대형 일차생산자 조사 방법
(1) 조사 장소 및 주요 일차생산자의 분포
연구가 진행된 지역은 낙동강 하구의 가덕도, 진우도, 을숙도와 맹금머리도이며, 이들
지역은 퇴적물의 침식과 퇴적작용이 대단히 활발한 지역이다(Fig. 2-1). 낙동강 하구지
역은 생태계 보전지역으로 지정되었으며, 철새도래지로 유명하다. 거머리말(Zostera
marina)은 진우도와 가덕도 앞 수심 2m 내외에 소규모로 서식하고 있었으며, 매년 서
식지역이 줄어들고 있었다. 거머리말의 조사지역은 비교적 서식 면적이 넓은 가덕도와
진우도 앞을 선정하였다. 애기거머리말(Zostera japonica)과 줄말(Ruppia maritima)은
퇴적 작용에 의해서 일년 중 석달 정도만 확인할 수 있었으며, 뻘질과 사질에 서식하는
세모고랭이(Scirpus trigueter)는 을숙도(뻘질)와 맹금머리도(사질)에서 연구를 진행하
였다. 낙동강 하구지역에서 주요 일차 생산자인 갈대(Phragmites australis)는 을숙도
남단에서 조사하였다. 낙동강 하구에 분포된 일차생산자(식물종 대상)의 분포를 파악하
기 위해서 계절마다 GPS를 이용해서 식물종이 차지하는 면적을 측정하였다. 주요 일차
생산자는 거머리말, 세모고랭이와 갈대이며, 이들 종은 매달 외부형태 생육밀도, 새체량
및 생산성을 측정하였으며, 줄말과 애기거머리말은 조사 대상에서 제외되었다.
Figure. 2-1. Study sites in Nak-Dong estuary on the south
coast of Korean peninsula.

- 59 -
(2) 일차 생산자의 형태학적 특성, 생육밀도 및 생체량
(가) 식물의 외부 형태
거머리말은 매월 10여 개체의 식물들을 채취하여 전체 길이(Shoot height), 엽초의 길
이(Sheath length) 및 잎의 폭(Leaf width) 등을 측정하였으며, 잎 조직 내 엽록소의 함
량도 측정하였다. 갈대와 세모고랭이는 현장에서 전체길이만 측정하였다.
(나) 밀도 및 생체량
주요 식물종의 생육밀도는 매월 방형구 (거머리말 - 35 × 35 ㎝, 갈대 - 20 × 20 ㎝,
세모고랭이 - 50 × 50 ㎝)내에 서식하는 개체수를 측정한 후, 방형구 내 모든 식물들을
채취하였다. 채취된 식물체는 아이스박스를 이용해 실험실로 가져와 민물로 부착생물
및 진흙을 제거한 후, 지상부와 지하부로 분리해서 60℃에서 무게가 일정할 때까지 건
조시켰다. 그런 다음 각각의 무게를 측정, 이를 토대로 단위 ㎡ 당으로 환산하여, 단위
면적 당 생체량(g DW m-2
)을 추정하였다.
(3) 생산성 측정
잘피류의 생산성 측정을 위해서 각 조사장소에서 blade marking technique (Zieman
1974; Kentula and McIntire 1986; Lee and Dunton 1996)를 이용하여 잘피의 생산성을
매달 측정하였다. 실험지역에서 10-15개체에 날카로운 바늘을 이용해 엽초(sheath)에
구멍을 뚫은 다음, 2주내지 4주 후에 채취하였다. 채취한 개체는 새로 자란 부분(new
leaf)과 기존 부분(old leaf)으로 나누어서, 각각의 건조량(dry weight)을 측정하였다. 이
렇게 측정된 값으로 개체 당 하루 동안 생산된 양 (㎎ dry weight sht-1 d
-1)을 계산하
였고, 서식밀도를 곱하여 단위면적당 하루 동안 생산된 양(g dry weight m-1 d
-1)을 계
산하였다.
갈대 및 세모고랭이는 다년생 초본으로 지상부는 봄에 새로 잎이 자라서 가을에 시
들어 사그러진다. 이 때, 가을까지 자란 잎이 그 식물의 총 생산량이라 할 수 있다. 따
라서 갈대와 세모고랭이의 생산성을 측정하기 위해서 매달 단위면적당 생체량을 측정
(거머리말과 동일 방법)하였으며, 매월 생체량의 차이를 통해서 월별 생산성을 측정하
였다. 또한, 성장이 멈춘 시점에 생체량을 측정하여, 연간 생산성을 추정하였다.
(4) 식물조직 내 탄소 및 질소 함량의 계절 변이
거머리말의 잎과 지하경 조직 내 탄소 및 질소의 양을 측정하기 위해서 매월 각 조
사장소에서 6~10개체를 채취하였다. 채취된 개체는 아이스박스에 보관해서 실험실로
옮겨와 부착생물 및 진흙을 제거한 다음, 잎과 지하경으로 나누어서 측정하였는데, 이

- 60 -
때, 잎 조직은 가장 신선하고 충분히 성숙한 2번 또는 3번 잎의 중간부분을 선택하였으
며, 지하경은 두 번째 또는 세 번째 마디를 선택하였다. 또한 갈대와 세모고랭이는 지
상부만 측정하였다. 이 시료들은 건조기(60℃)에서 충분히 건조시킨 후, 막자사발을 이
용해 곱게 갈아 CHN elemental analyzer(Flash EA1112)를 이용해 각 부분의 탄소 및
질소 양과 C:N 비율을 측정하였다.
(5) 주요 일차생산자의 분포
2004년 7월부터 조사된 주요 일차생산자의 분포는 세모고랭이와 갈대가 낙동강 하구
지역에서 가장 우점하는 것으로 나타났으며(Fig. 2-2), 거머리말은 예전에 비해 생육 면
적이 감소한 것으로 나타났다(Fig. 2-3). 정 & 최 (1985)는 낙동강 하구지역에 생육하
는 거머리말의 면적을 약 13.6㎢ 라고 보고했다. 그러나 본 연구 결과 거머리말의 생육
지은 신항만 공사와 공장 매립지 공사로 인해 거의 대부분이 파괴 되었으며, 현재 가덕
도와 진우도 앞 양식장에 소규모로 존재하고 있었다. 낙동강 하구지역에서 차지하는 거
머리말의 면적은 약 0.7㎢이나 매년 그 규모가 줄어들고 있는 실정이다. 따라서 이들
지역에서 거머리말에 대한 보호대책이 매우 절실한 상태이다. 애기거머리말과 줄말은
퇴적작용에 의해서 서식 면적을 평가할 수 없었다. 실험지역에서 가장 우점하는 것으로
관측된 세모고랭이는 봄부터 자라기 시작하여 여름에 가장 넓은 지역을 차지하였다. 그
러나 가을부터 사그러지면서 을숙도는 8~9월초, 맹금머리도는 9~11월초에 완전히 사
그러지는 것으로 나타났다. 11월에 세모고랭이의 지상부는 완전히 자취를 감추었고, 지
하부는 땅 속 깊숙한 곳에 존재하였다. 세모고랭이의 서식면적은 약 5.2㎢이였다. 갈대
도 세모고랭이와 같이 봄에 새싹이 나오기 시작하여 가을에 지는 것으로 나타났다. 서
식 면적은 거의 일정했으며, 전체 면적은 약 1.6㎢이였다.
Figure. 2-2. Seaonal distribution of main primary producers in Nak-Dong
estuary.
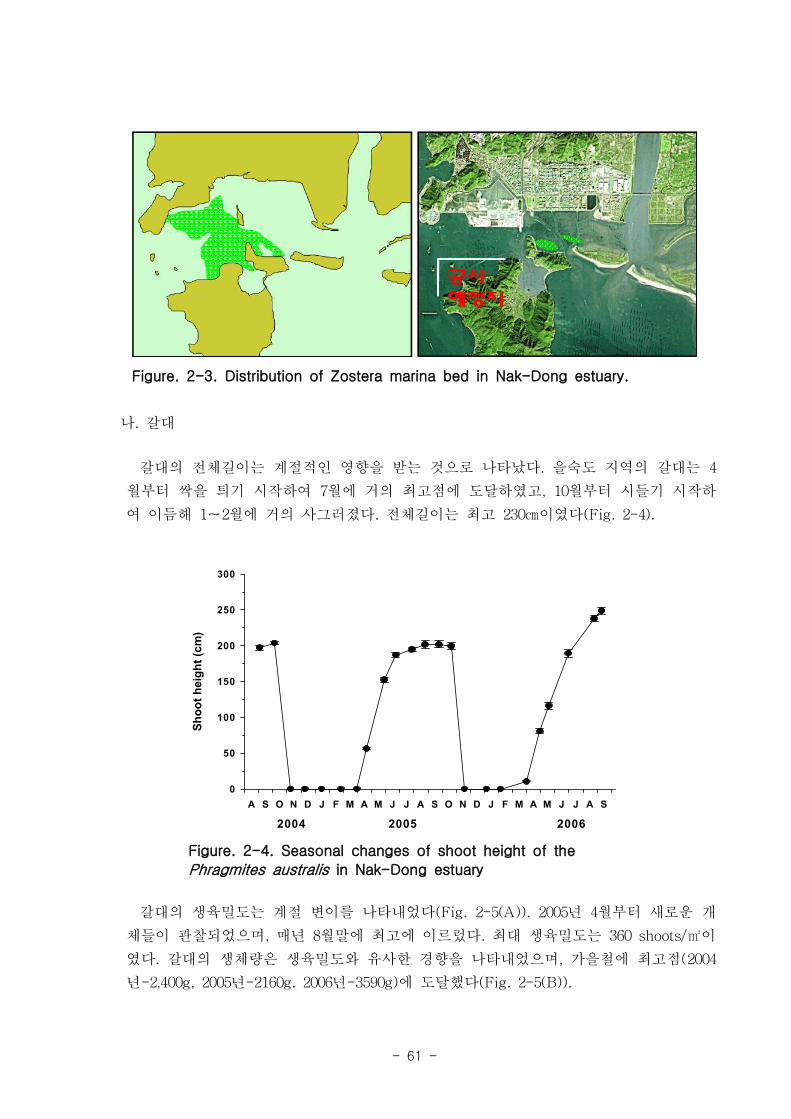
- 61 -
Figure. 2-3. Distribution of Zostera marina bed in Nak-Dong estuary.
나. 갈대
갈대의 전체길이는 계절적인 영향을 받는 것으로 나타났다. 을숙도 지역의 갈대는 4
월부터 싹을 틔기 시작하여 7월에 거의 최고점에 도달하였고, 10월부터 시들기 시작하
여 이듬해 1~2월에 거의 사그러졌다. 전체길이는 최고 230㎝이였다(Fig. 2-4).
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S
Shoot heig
ht (c
m)
0
50
100
150
200
250
300
Figure. 2-4. Seasonal changes of shoot height of the
Phragmites australis in Nak-Dong estuary
갈대의 생육밀도는 계절 변이를 나타내었다(Fig. 2-5(A)). 2005년 4월부터 새로운 개
체들이 관찰되었으며, 매년 8월말에 최고에 이르렀다. 최대 생육밀도는 360 shoots/㎡이
였다. 갈대의 생체량은 생육밀도와 유사한 경향을 나타내었으며, 가을철에 최고점(2004
년-2,400g, 2005년-2160g. 2006년-3590g)에 도달했다(Fig. 2-5(B)).
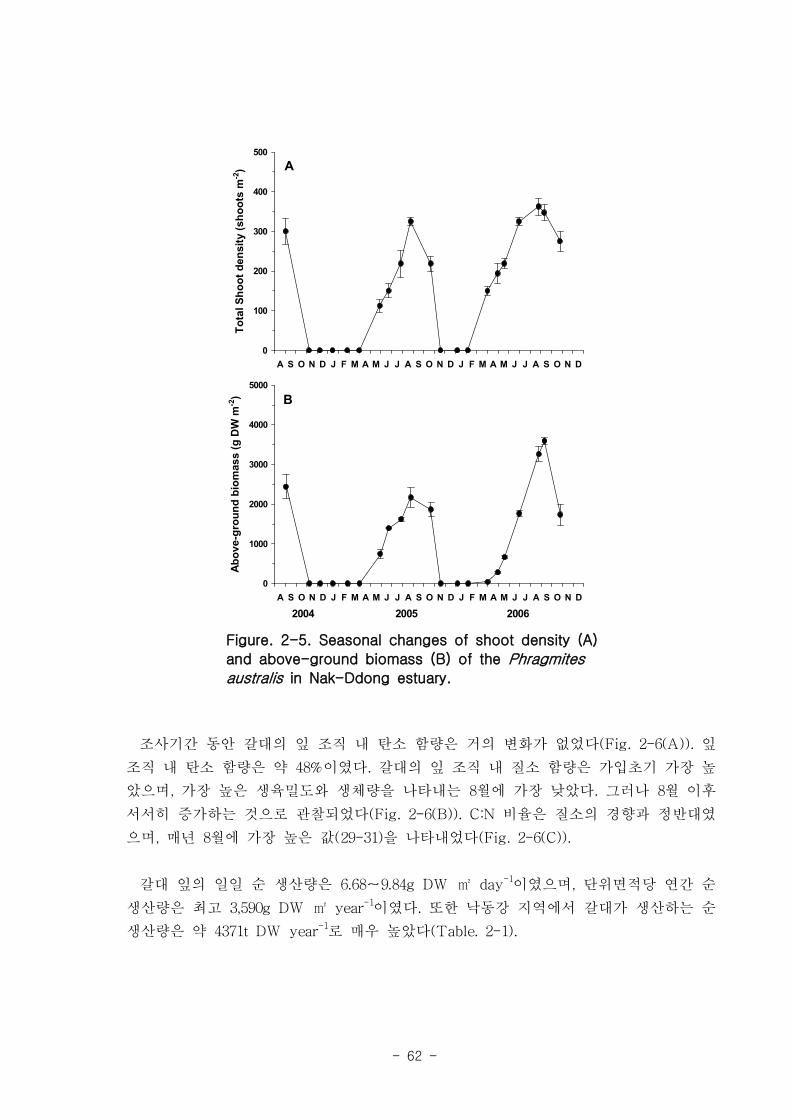
- 62 -
A
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Tota
l Shoot density (shoots
m-2)
0
100
200
300
400
500
B
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Above-g
round b
iom
ass (g D
W m
-2)
0
1000
2000
3000
4000
5000
Figure. 2-5. Seasonal changes of shoot density (A)
and above-ground biomass (B) of the Phragmites australis in Nak-Ddong estuary.
조사기간 동안 갈대의 잎 조직 내 탄소 함량은 거의 변화가 없었다(Fig. 2-6(A)). 잎
조직 내 탄소 함량은 약 48%이였다. 갈대의 잎 조직 내 질소 함량은 가입초기 가장 높
았으며, 가장 높은 생육밀도와 생체량을 나타내는 8월에 가장 낮았다. 그러나 8월 이후
서서히 증가하는 것으로 관찰되었다(Fig. 2-6(B)). C:N 비율은 질소의 경향과 정반대였
으며, 매년 8월에 가장 높은 값(29-31)을 나타내었다(Fig. 2-6(C)).
갈대 잎의 일일 순 생산량은 6.68~9.84g DW ㎡ day-1이였으며, 단위면적당 연간 순
생산량은 최고 3,590g DW ㎡ year-1이였다. 또한 낙동강 지역에서 갈대가 생산하는 순
생산량은 약 4371t DW year-1로 매우 높았다(Table. 2-1).
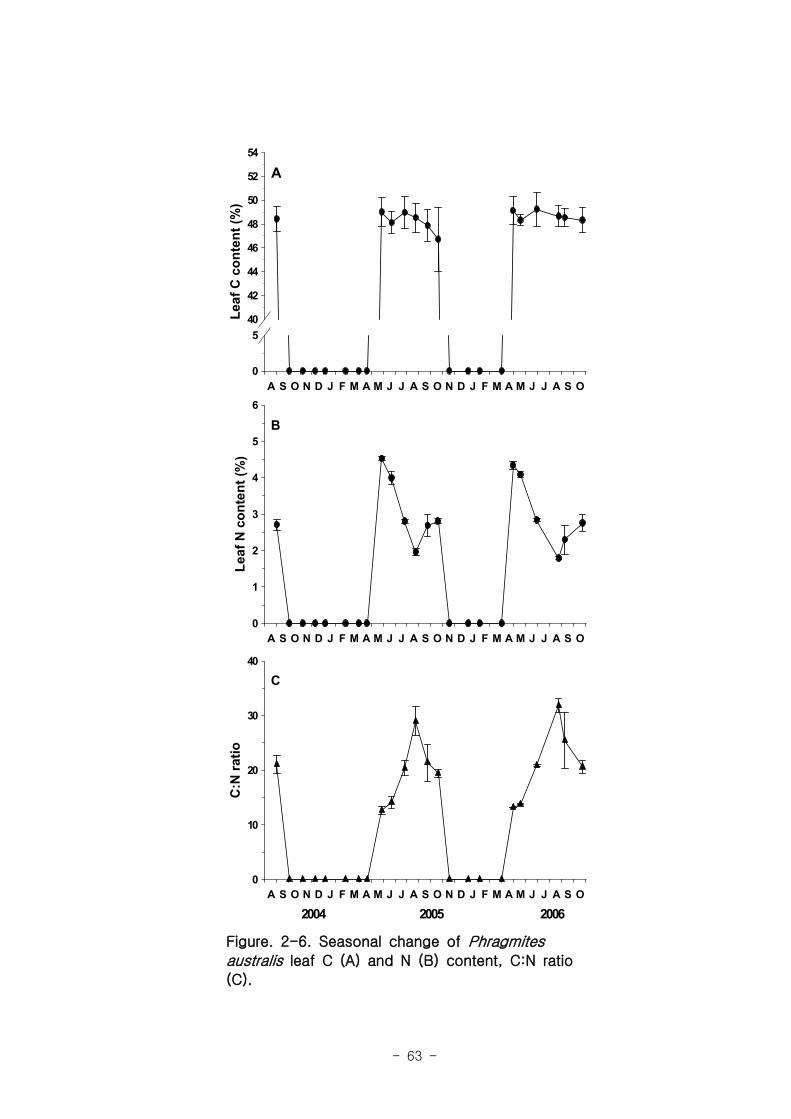
- 63 -
A
A S O N D J F M A M J J A S O N D J F M A M J J A S O
Leaf C
conte
nt (%
)
0
5
40
42
44
46
48
50
52
54
B
A S O N D J F M A M J J A S O N D J F M A M J J A S O
Leaf N
conte
nt (%
)
0
1
2
3
4
5
6
C
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O
C:N
ratio
0
10
20
30
40
Figure. 2-6. Seasonal change of Phragmites australis leaf C (A) and N (B) content, C:N ratio
(C).
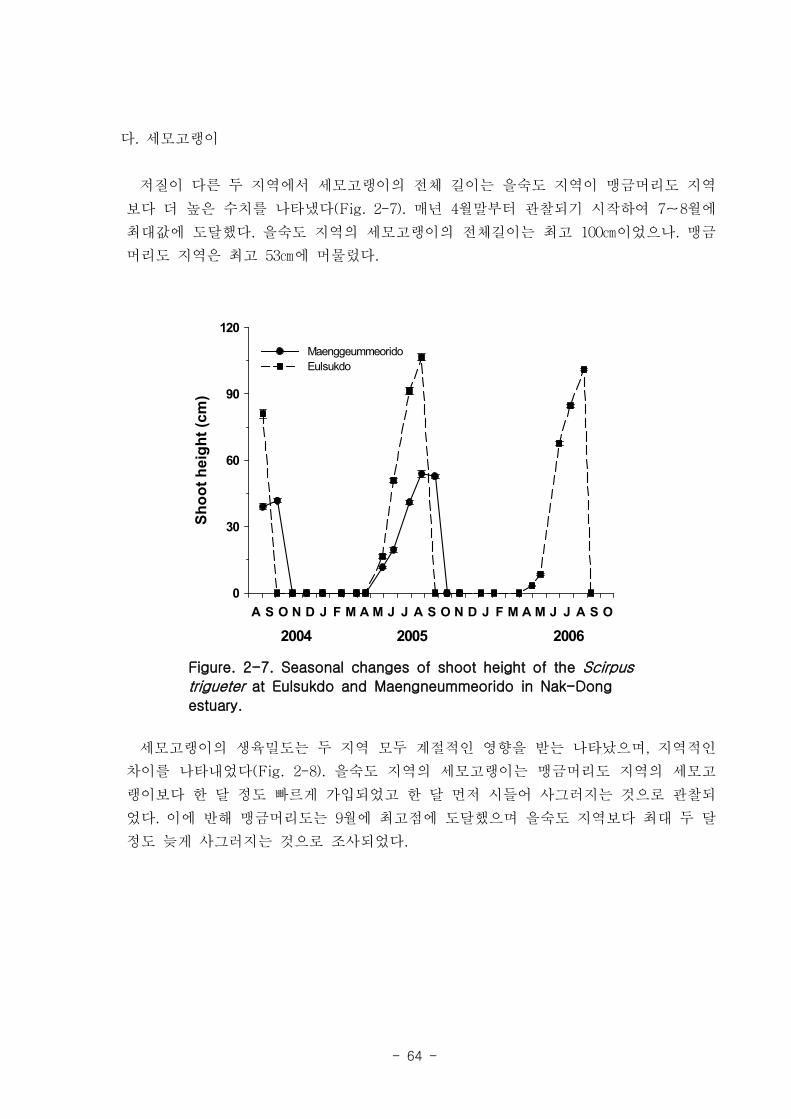
- 64 -
다. 세모고랭이
저질이 다른 두 지역에서 세모고랭이의 전체 길이는 을숙도 지역이 맹금머리도 지역
보다 더 높은 수치를 나타냈다(Fig. 2-7). 매년 4월말부터 관찰되기 시작하여 7~8월에
최대값에 도달했다. 을숙도 지역의 세모고랭이의 전체길이는 최고 100㎝이었으나. 맹금
머리도 지역은 최고 53㎝에 머물렀다.
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O
Shoot heig
ht (c
m)
0
30
60
90
120
Maenggeummeorido
Eulsukdo
Figure. 2-7. Seasonal changes of shoot height of the Scirpus trigueter at Eulsukdo and Maengneummeorido in Nak-Dong
estuary.
세모고랭이의 생육밀도는 두 지역 모두 계절적인 영향을 받는 나타났으며, 지역적인
차이를 나타내었다(Fig. 2-8). 을숙도 지역의 세모고랭이는 맹금머리도 지역의 세모고
랭이보다 한 달 정도 빠르게 가입되었고 한 달 먼저 시들어 사그러지는 것으로 관찰되
었다. 이에 반해 맹금머리도는 9월에 최고점에 도달했으며 을숙도 지역보다 최대 두 달
정도 늦게 사그러지는 것으로 조사되었다.
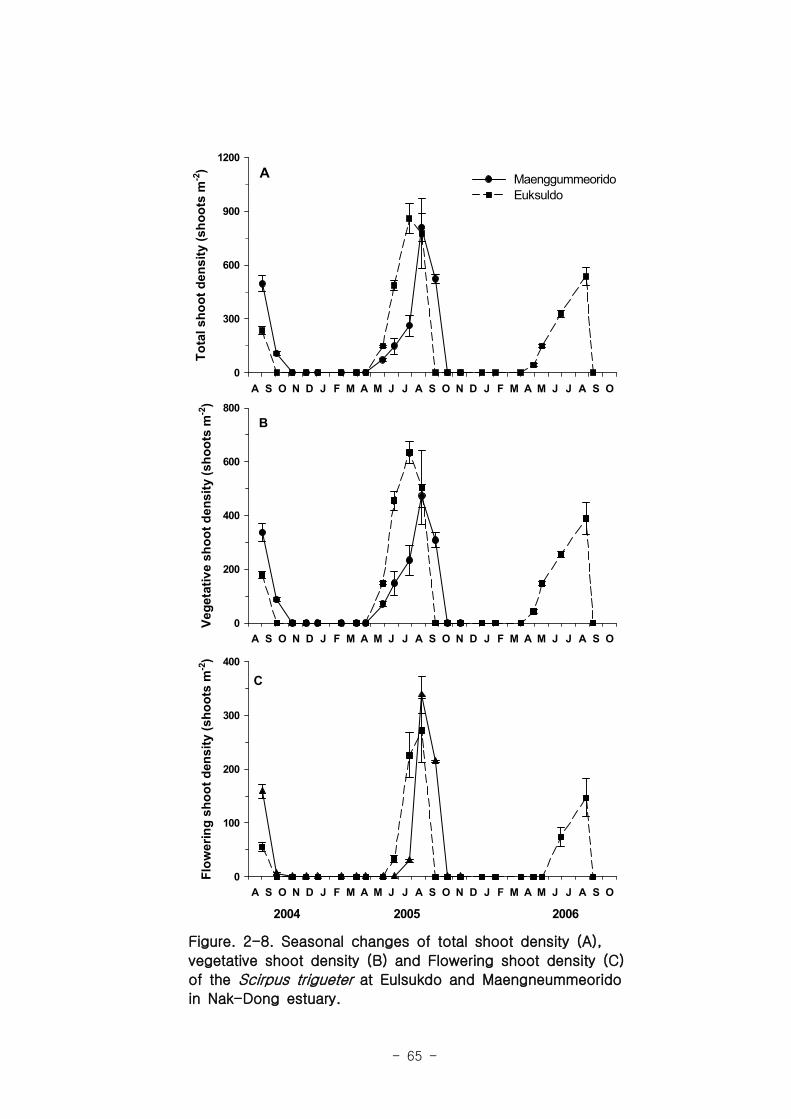
- 65 -
A
A S O N D J F M A M J J A S O N D J F M A M J J A S O
Tota
l shoot density (shoots
m-2)
0
300
600
900
1200
Maenggummeorido
Euksuldo
B
A S O N D J F M A M J J A S O N D J F M A M J J A S O
Vegeta
tive s
hoot density (shoots
m-2)
0
200
400
600
800
C
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O
Flo
wering s
hoot density (shoots
m-2)
0
100
200
300
400
Figure. 2-8. Seasonal changes of total shoot density (A),
vegetative shoot density (B) and Flowering shoot density (C)
of the Scirpus trigueter at Eulsukdo and Maengneummeorido
in Nak-Dong estuary.
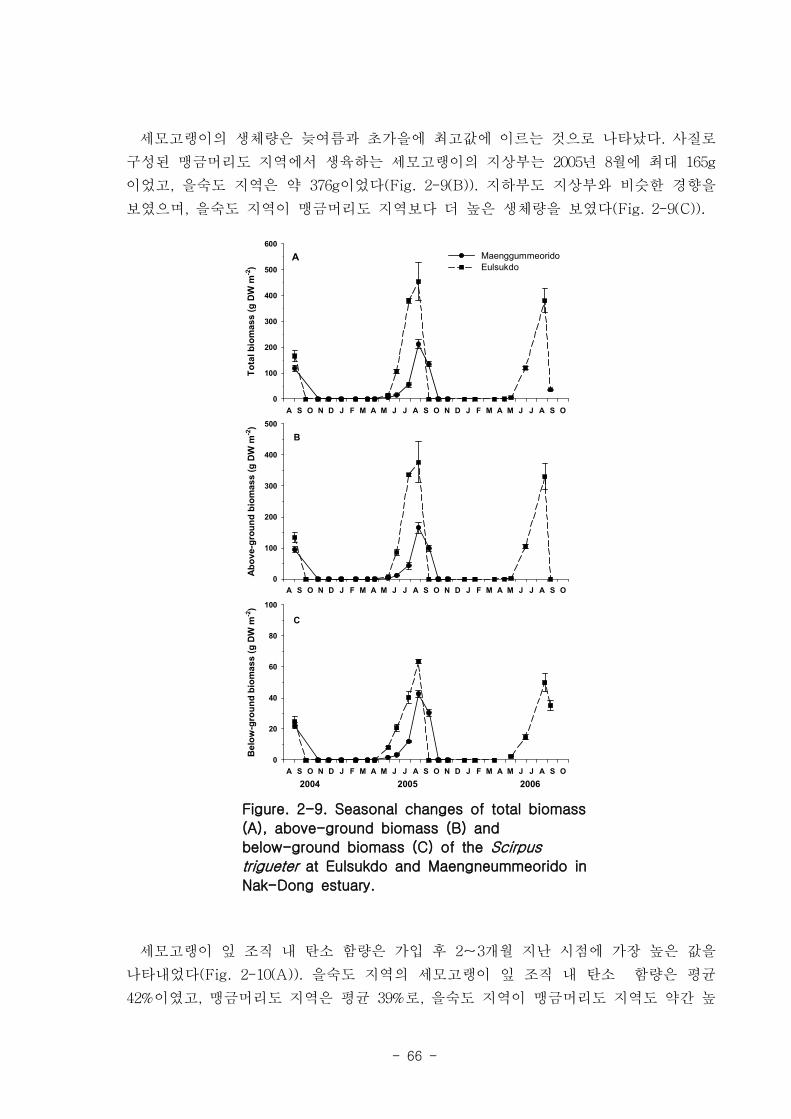
- 66 -
세모고랭이의 생체량은 늦여름과 초가을에 최고값에 이르는 것으로 나타났다. 사질로
구성된 맹금머리도 지역에서 생육하는 세모고랭이의 지상부는 2005년 8월에 최대 165g
이었고, 을숙도 지역은 약 376g이었다(Fig. 2-9(B)). 지하부도 지상부와 비슷한 경향을
보였으며, 을숙도 지역이 맹금머리도 지역보다 더 높은 생체량을 보였다(Fig. 2-9(C)).
A
A S O N D J F M A M J J A S O N D J F M A M J J A S O
Tota
l bio
mass (g D
W m
-2)
0
100
200
300
400
500
600
Maenggummeorido
Eulsukdo
B
A S O N D J F M A M J J A S O N D J F M A M J J A S O
Above-g
round b
iom
ass (g D
W m
-2)
0
100
200
300
400
500
C
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O
Belo
w-g
round b
iom
ass (g D
W m
-2)
0
20
40
60
80
100
Figure. 2-9. Seasonal changes of total biomass
(A), above-ground biomass (B) and
below-ground biomass (C) of the Scirpus trigueter at Eulsukdo and Maengneummeorido in
Nak-Dong estuary.
세모고랭이 잎 조직 내 탄소 함량은 가입 후 2~3개월 지난 시점에 가장 높은 값을
나타내었다(Fig. 2-10(A)). 을숙도 지역의 세모고랭이 잎 조직 내 탄소 함량은 평균
42%이였고, 맹금머리도 지역은 평균 39%로, 을숙도 지역이 맹금머리도 지역도 약간 높
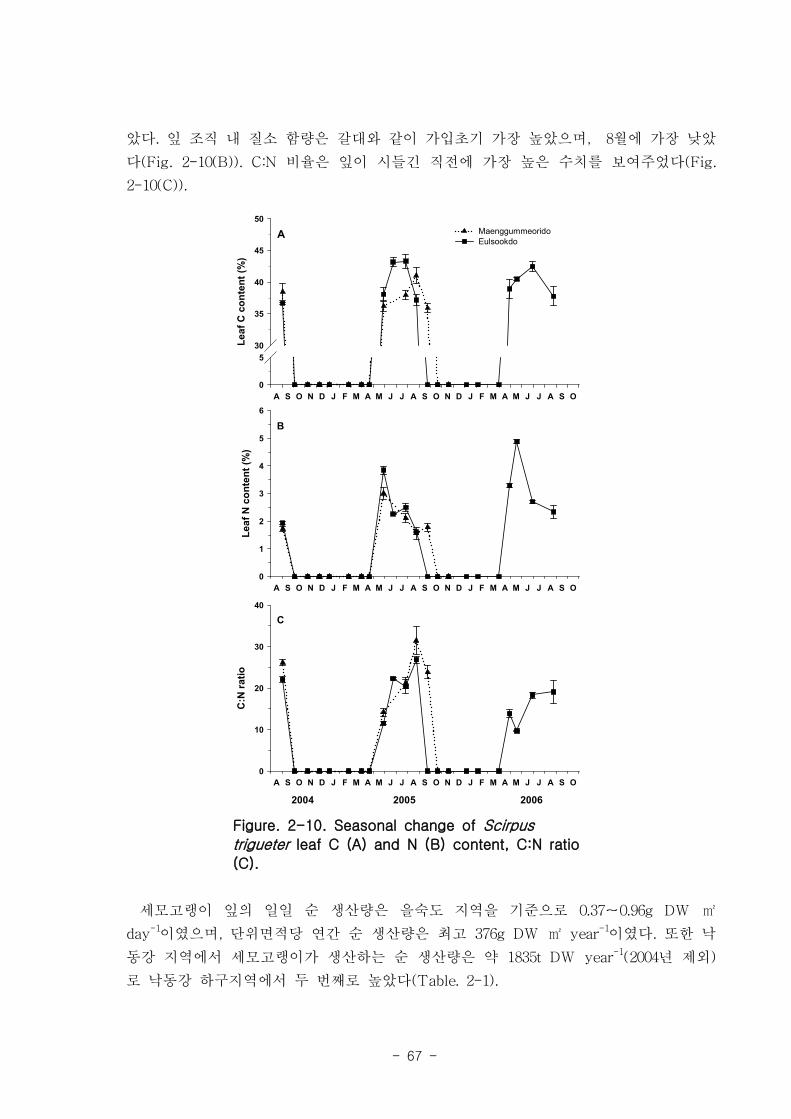
- 67 -
았다. 잎 조직 내 질소 함량은 갈대와 같이 가입초기 가장 높았으며, 8월에 가장 낮았
다(Fig. 2-10(B)). C:N 비율은 잎이 시들긴 직전에 가장 높은 수치를 보여주었다(Fig.
2-10(C)).
A
A S O N D J F M A M J J A S O N D J F M A M J J A S O
Leaf C
conte
nt (%
)
0
5
30
35
40
45
50
Maenggummeorido
Eulsookdo
B
A S O N D J F M A M J J A S O N D J F M A M J J A S O
Leaf N
conte
nt (%
)
0
1
2
3
4
5
6
C
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O
C:N
ratio
0
10
20
30
40
Figure. 2-10. Seasonal change of Scirpus trigueter leaf C (A) and N (B) content, C:N ratio
(C).
세모고랭이 잎의 일일 순 생산량은 을숙도 지역을 기준으로 0.37~0.96g DW ㎡
day-1이였으며, 단위면적당 연간 순 생산량은 최고 376g DW ㎡ year
-1이였다. 또한 낙
동강 지역에서 세모고랭이가 생산하는 순 생산량은 약 1835t DW year-1
(2004년 제외)
로 낙동강 하구지역에서 두 번째로 높았다(Table. 2-1).
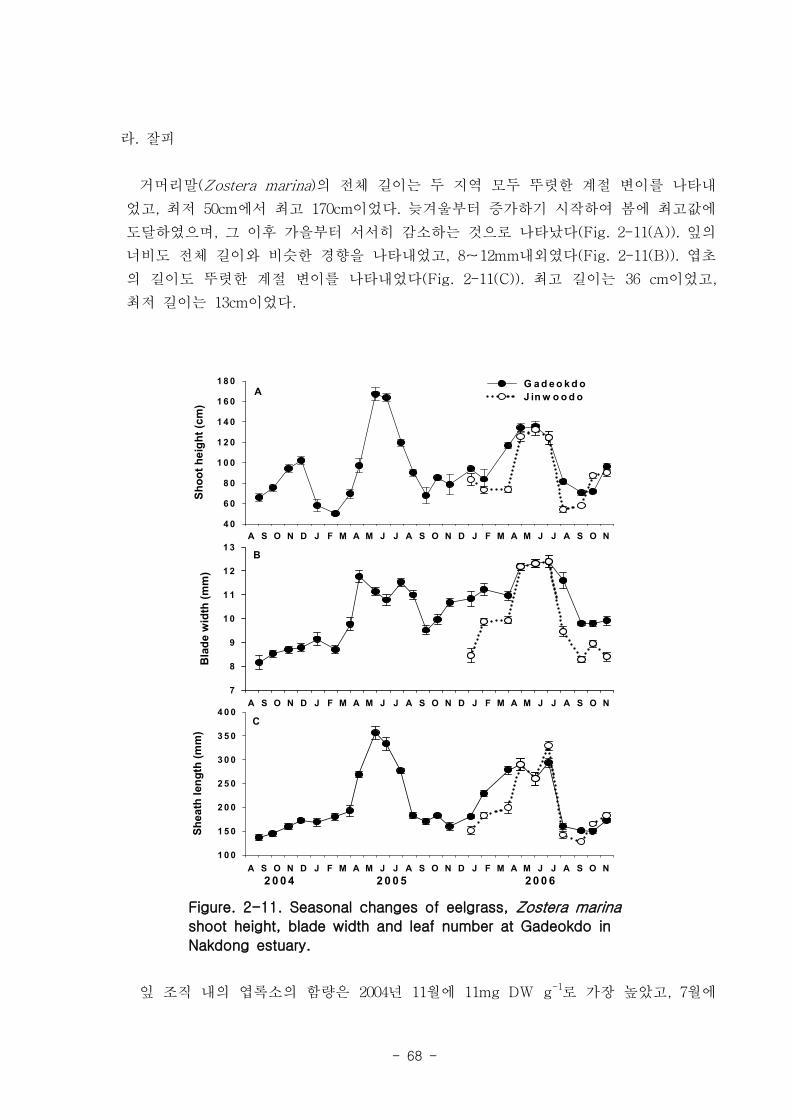
- 68 -
라. 잘피
거머리말(Zostera marina)의 전체 길이는 두 지역 모두 뚜렷한 계절 변이를 나타내
었고, 최저 50cm에서 최고 170cm이었다. 늦겨울부터 증가하기 시작하여 봄에 최고값에
도달하였으며, 그 이후 가을부터 서서히 감소하는 것으로 나타났다(Fig. 2-11(A)). 잎의
너비도 전체 길이와 비슷한 경향을 나타내었고, 8~12mm내외였다(Fig. 2-11(B)). 엽초
의 길이도 뚜렷한 계절 변이를 나타내었다(Fig. 2-11(C)). 최고 길이는 36 cm이었고,
최저 길이는 13cm이었다.
A S O N D J F M A M J J A S O N D J F M A M J J A S O N
Shoot heig
ht (c
m)
40
60
80
100
120
140
160
180 G ad eo kd o
J in w o od o
A S O N D J F M A M J J A S O N D J F M A M J J A S O N
Bla
de w
idth
(m
m)
7
8
9
10
11
12
13
A S O N D J F M A M J J A S O N D J F M A M J J A S O N
Sheath
length
(m
m)
100
150
200
250
300
350
400
2004 2 0 0 5 2 0 0 6
A
B
C
Figure. 2-11. Seasonal changes of eelgrass, Zostera marina
shoot height, blade width and leaf number at Gadeokdo in
Nakdong estuary.
잎 조직 내의 엽록소의 함량은 2004년 11월에 11mg DW g-1로 가장 높았고, 7월에
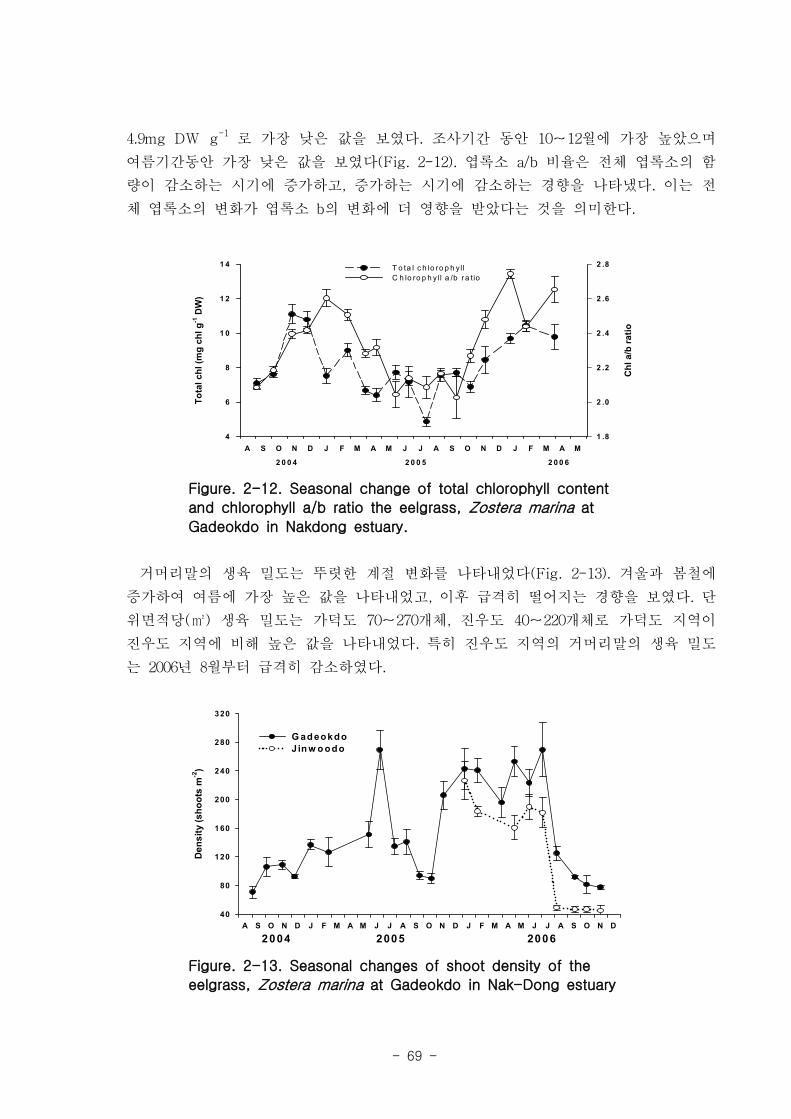
- 69 -
4.9mg DW g-1 로 가장 낮은 값을 보였다. 조사기간 동안 10~12월에 가장 높았으며
여름기간동안 가장 낮은 값을 보였다(Fig. 2-12). 엽록소 a/b 비율은 전체 엽록소의 함
량이 감소하는 시기에 증가하고, 증가하는 시기에 감소하는 경향을 나타냈다. 이는 전
체 엽록소의 변화가 엽록소 b의 변화에 더 영향을 받았다는 것을 의미한다.
2 0 0 4 2 0 0 5 2 0 0 6
A S O N D J F M A M J J A S O N D J F M A M
Tota
l chl (m
g c
hl g
-1 D
W)
4
6
8
1 0
1 2
1 4
Chl a/b
ratio
1 .8
2 .0
2 .2
2 .4
2 .6
2 .8T o ta l c h lo ro p h yl l
C h lo ro p h y ll a /b ra tio
Figure. 2-12. Seasonal change of total chlorophyll content
and chlorophyll a/b ratio the eelgrass, Zostera marina at
Gadeokdo in Nakdong estuary.
거머리말의 생육 밀도는 뚜렷한 계절 변화를 나타내었다(Fig. 2-13). 겨울과 봄철에
증가하여 여름에 가장 높은 값을 나타내었고, 이후 급격히 떨어지는 경향을 보였다. 단
위면적당(㎡) 생육 밀도는 가덕도 70~270개체, 진우도 40~220개체로 가덕도 지역이
진우도 지역에 비해 높은 값을 나타내었다. 특히 진우도 지역의 거머리말의 생육 밀도
는 2006년 8월부터 급격히 감소하였다.
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Density (shoots
m-2)
40
80
120
160
200
240
280
320
G adeokdo
Jinw oodo
Figure. 2-13. Seasonal changes of shoot density of the
eelgrass, Zostera marina at Gadeokdo in Nak-Dong estuary
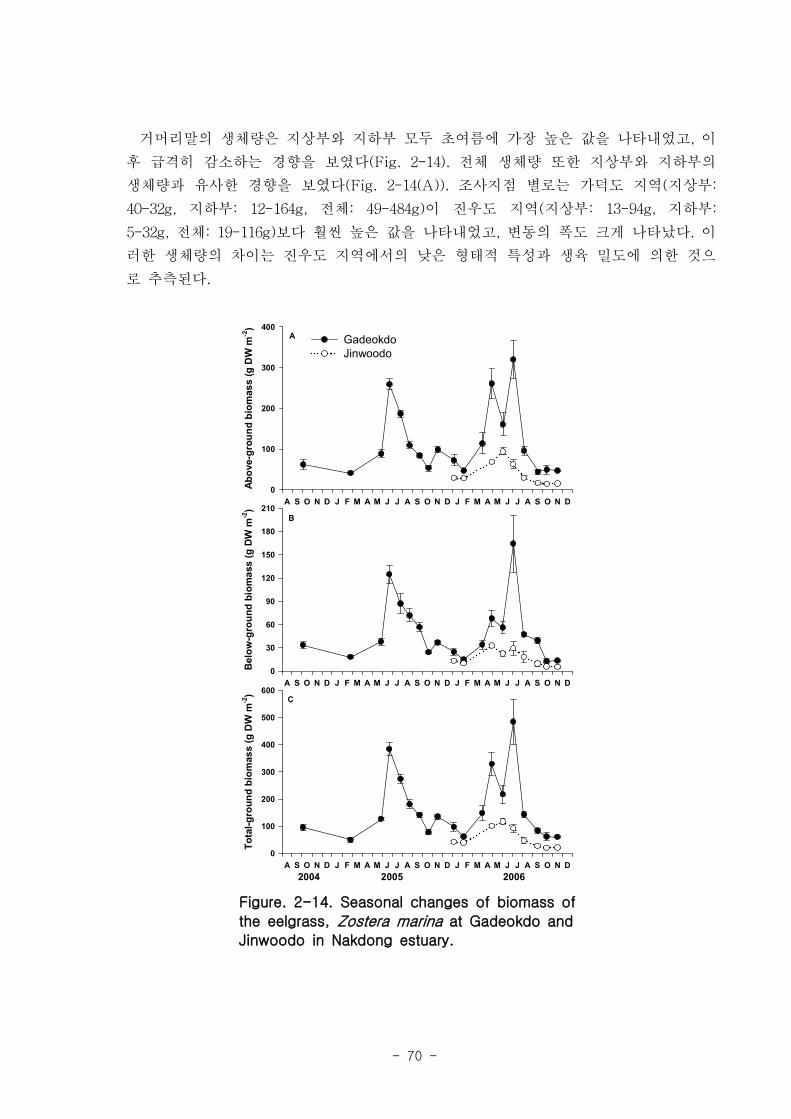
- 70 -
거머리말의 생체량은 지상부와 지하부 모두 초여름에 가장 높은 값을 나타내었고, 이
후 급격히 감소하는 경향을 보였다(Fig. 2-14). 전체 생체량 또한 지상부와 지하부의
생체량과 유사한 경향을 보였다(Fig. 2-14(A)). 조사지점 별로는 가덕도 지역(지상부:
40-32g, 지하부: 12-164g, 전체: 49-484g)이 진우도 지역(지상부: 13-94g, 지하부:
5-32g, 전체: 19-116g)보다 훨씬 높은 값을 나타내었고, 변동의 폭도 크게 나타났다. 이
러한 생체량의 차이는 진우도 지역에서의 낮은 형태적 특성과 생육 밀도에 의한 것으
로 추측된다.
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Above-g
round b
iom
ass (g D
W m
-2)
0
100
200
300
400
Gadeokdo
Jinwoodo
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Belo
w-g
round b
iom
ass (g D
W m
-2)
0
30
60
90
120
150
180
210
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Tota
l-gro
und b
iom
ass (g D
W m
-2)
0
100
200
300
400
500
600
A
B
C
Figure. 2-14. Seasonal changes of biomass of
the eelgrass, Zostera marina at Gadeokdo and
Jinwoodo in Nakdong estuary.
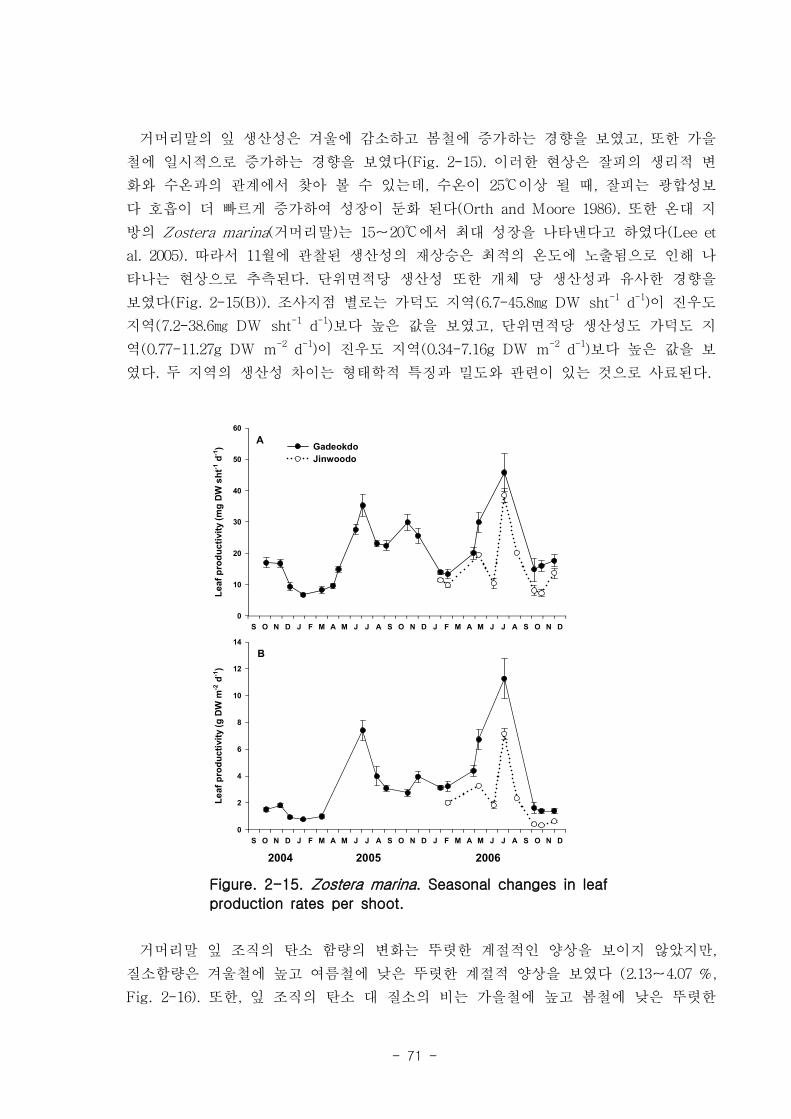
- 71 -
거머리말의 잎 생산성은 겨울에 감소하고 봄철에 증가하는 경향을 보였고, 또한 가을
철에 일시적으로 증가하는 경향을 보였다(Fig. 2-15). 이러한 현상은 잘피의 생리적 변
화와 수온과의 관계에서 찾아 볼 수 있는데, 수온이 25℃이상 될 때, 잘피는 광합성보
다 호흡이 더 빠르게 증가하여 성장이 둔화 된다(Orth and Moore 1986). 또한 온대 지
방의 Zostera marina(거머리말)는 15~20℃에서 최대 성장을 나타낸다고 하였다(Lee et
al. 2005). 따라서 11월에 관찰된 생산성의 재상승은 최적의 온도에 노출됨으로 인해 나
타나는 현상으로 추측된다. 단위면적당 생산성 또한 개체 당 생산성과 유사한 경향을
보였다(Fig. 2-15(B)). 조사지점 별로는 가덕도 지역(6.7-45.8㎎ DW sht-1 d
-1)이 진우도
지역(7.2-38.6㎎ DW sht-1 d
-1)보다 높은 값을 보였고, 단위면적당 생산성도 가덕도 지
역(0.77-11.27g DW m-2 d
-1)이 진우도 지역(0.34-7.16g DW m
-2 d-1
)보다 높은 값을 보
였다. 두 지역의 생산성 차이는 형태학적 특징과 밀도와 관련이 있는 것으로 사료된다.
S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Leaf pro
ductivity (m
g D
W s
ht-1
d-1)
0
10
20
30
40
50
60
Gadeokdo
Jinwoodo
2004 2005 2006
S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Leaf pro
ductivity (g D
W m
-2 d
-1)
0
2
4
6
8
10
12
14
A
B
Figure. 2-15. Zostera marina. Seasonal changes in leaf
production rates per shoot.
거머리말 잎 조직의 탄소 함량의 변화는 뚜렷한 계절적인 양상을 보이지 않았지만,
질소함량은 겨울철에 높고 여름철에 낮은 뚜렷한 계절적 양상을 보였다 (2.13~4.07 %,
Fig. 2-16). 또한, 잎 조직의 탄소 대 질소의 비는 가을철에 높고 봄철에 낮은 뚜렷한
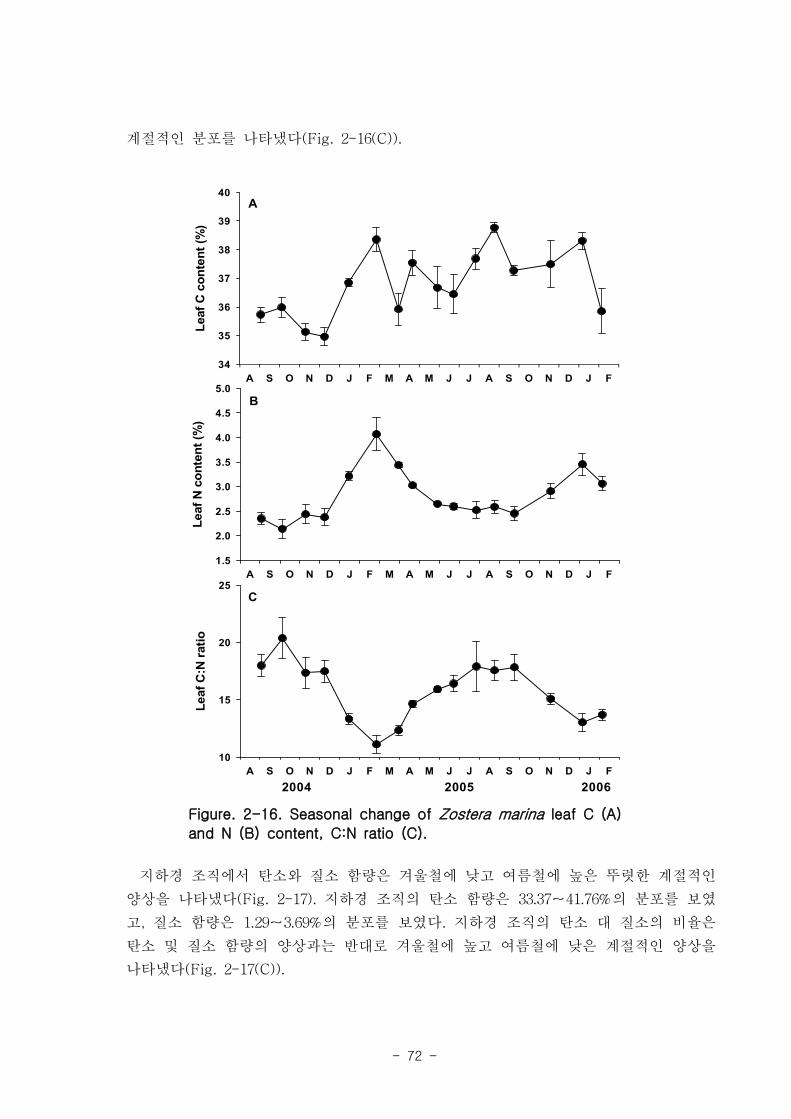
- 72 -
계절적인 분포를 나타냈다(Fig. 2-16(C)).
A S O N D J F M A M J J A S O N D J F
Leaf C
conte
nt (%
)
34
35
36
37
38
39
40
A S O N D J F M A M J J A S O N D J F
Leaf N
conte
nt (%
)
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
2004 2005 2006
A S O N D J F M A M J J A S O N D J F
Leaf C
:N ratio
10
15
20
25
A
B
C
Figure. 2-16. Seasonal change of Zostera marina leaf C (A)
and N (B) content, C:N ratio (C).
지하경 조직에서 탄소와 질소 함량은 겨울철에 낮고 여름철에 높은 뚜렷한 계절적인
양상을 나타냈다(Fig. 2-17). 지하경 조직의 탄소 함량은 33.37~41.76%의 분포를 보였
고, 질소 함량은 1.29~3.69%의 분포를 보였다. 지하경 조직의 탄소 대 질소의 비율은
탄소 및 질소 함량의 양상과는 반대로 겨울철에 높고 여름철에 낮은 계절적인 양상을
나타냈다(Fig. 2-17(C)).
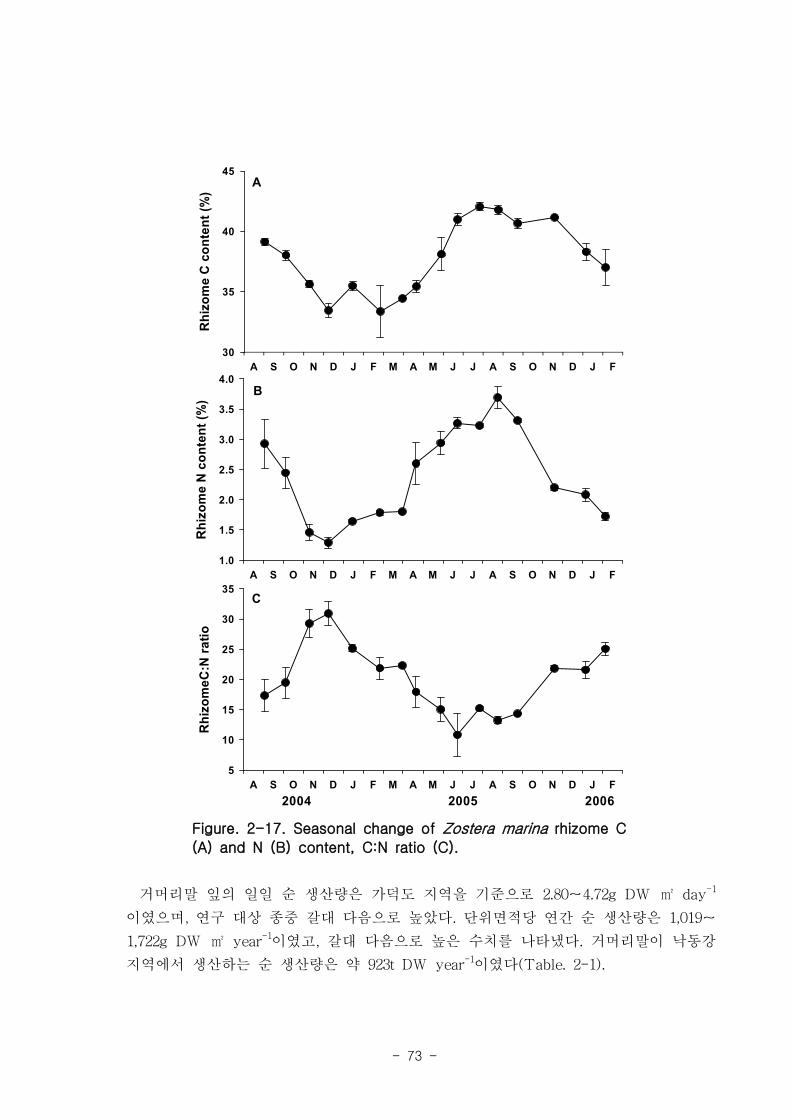
- 73 -
A S O N D J F M A M J J A S O N D J F
Rhiz
om
e C
conte
nt (%
)
30
35
40
45
A S O N D J F M A M J J A S O N D J F
Rhiz
om
e N
conte
nt (%
)
1.0
1.5
2.0
2.5
3.0
3.5
4.0
2004 2005 2006
A S O N D J F M A M J J A S O N D J F
Rhiz
om
eC
:N r
atio
5
10
15
20
25
30
35
A
B
C
Figure. 2-17. Seasonal change of Zostera marina rhizome C
(A) and N (B) content, C:N ratio (C).
거머리말 잎의 일일 순 생산량은 가덕도 지역을 기준으로 2.80~4.72g DW ㎡ day-1
이였으며, 연구 대상 종중 갈대 다음으로 높았다. 단위면적당 연간 순 생산량은 1,019~
1,722g DW ㎡ year-1이였고, 갈대 다음으로 높은 수치를 나타냈다. 거머리말이 낙동강
지역에서 생산하는 순 생산량은 약 923t DW year-1이였다(Table. 2-1).
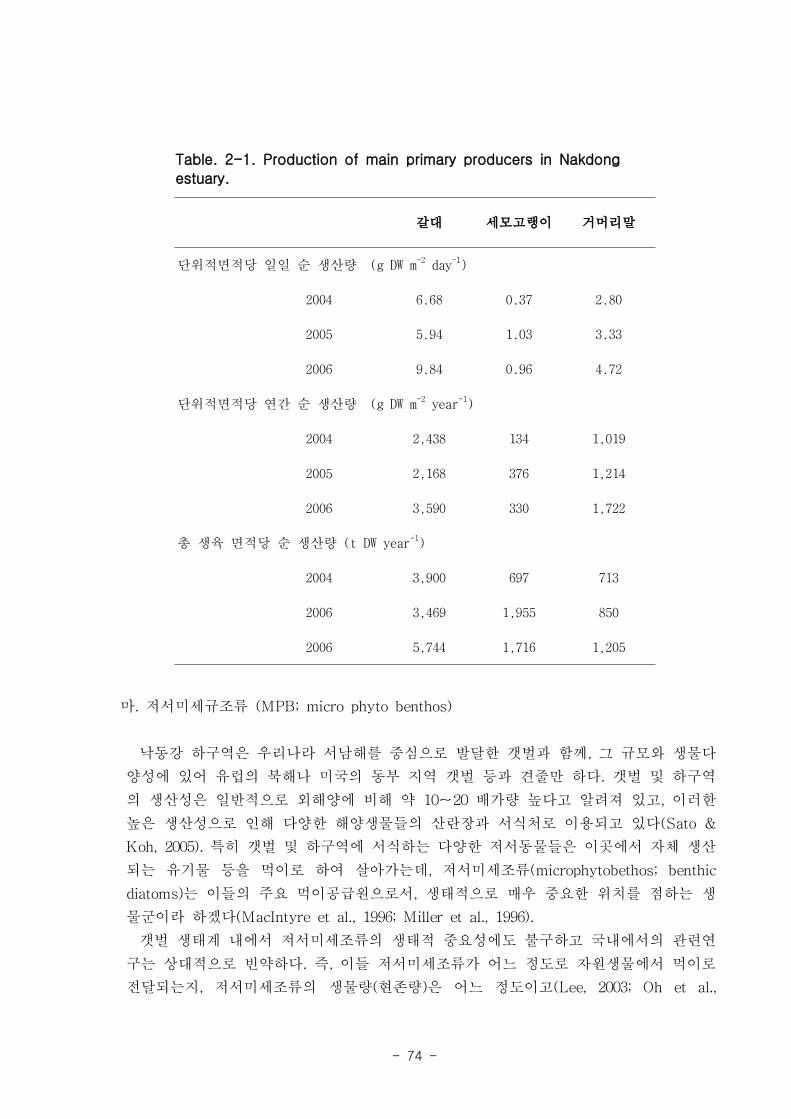
- 74 -
갈대 세모고랭이 거머리말
단위적면적당 일일 순 생산량 (g DW m-2 day
-1)
2004 6.68 0.37 2.80
2005 5.94 1.03 3.33
2006 9.84 0.96 4.72
단위적면적당 연간 순 생산량 (g DW m-2 year
-1)
2004 2,438 134 1,019
2005 2,168 376 1,214
2006 3,590 330 1,722
총 생육 면적당 순 생산량 (t DW year-1)
2004 3,900 697 713
2006 3,469 1,955 850
2006 5,744 1,716 1,205
Table. 2-1. Production of main primary producers in Nakdong
estuary.
마. 저서미세규조류 (MPB; micro phyto benthos)
낙동강 하구역은 우리나라 서남해를 중심으로 발달한 갯벌과 함께, 그 규모와 생물다
양성에 있어 유럽의 북해나 미국의 동부 지역 갯벌 등과 견줄만 하다. 갯벌 및 하구역
의 생산성은 일반적으로 외해양에 비해 약 10~20 배가량 높다고 알려져 있고, 이러한
높은 생산성으로 인해 다양한 해양생물들의 산란장과 서식처로 이용되고 있다(Sato &
Koh, 2005). 특히 갯벌 및 하구역에 서식하는 다양한 저서동물들은 이곳에서 자체 생산
되는 유기물 등을 먹이로 하여 살아가는데, 저서미세조류(microphytobethos; benthic
diatoms)는 이들의 주요 먹이공급원으로서, 생태적으로 매우 중요한 위치를 점하는 생
물군이라 하겠다(MacIntyre et al., 1996; Miller et al., 1996).
갯벌 생태계 내에서 저서미세조류의 생태적 중요성에도 불구하고 국내에서의 관련연
구는 상대적으로 빈약하다. 즉, 이들 저서미세조류가 어느 정도로 자원생물에서 먹이로
전달되는지, 저서미세조류의 생물량(현존량)은 어느 정도이고(Lee, 2003; Oh et al.,
- 75 -
2004; Yoo & Choi, 2005), 계절에 따라 생물량 및 생산량은 얼마나 다른지(Oh et al.,
2004; Hwang & Cho, 2005) 등에 관한 연구가 국외에 비하여 상대적으로 매우 빈약한
현실이다. 나아가, 이상의 연구들에서 저서미세조류의 종조성 및 생물다양성 측면이 깊
이 있게 다루어지지 않아, 생물과 환경과의 관계 등을 규명함에 있어, 그 한계가 있다
고 하겠다.
본 연구에서는 낙동강 하구 갯벌의 생태구성요소 중 일차생산을 담당하는 주요 식물
군인 저서미세조류의 생물량, 생산량, 그리고 종조성 등을 파악하였다. 2005~2006년까
지 총 7차례에 걸쳐 낙동강 하구 갯벌 퇴적물내 저서미세조류의 표층 현존량, 깊이별
생물량, 종조성, 및 일차생산량 등을 측정하였다(Table. 2-2). 갯벌이 노출되는 동안 시
간에 따른 깊이별 생물량도 병행 측정하여 저서미세조류의 거동에 관한 기본적인 자료
도 생산하였다. 2년간의 조사는 계절별로 이루어져 저서미세조류의 생물량이 계절에 따
라 어떻게 다른지에 대해서 알아볼 수 있도록 계획되었고, 나아가 인공갯벌이 조성된
이후에 퇴적물 표층의 저서미세조류의 생물량 및 종조성이 어떻게 달라지는가에 대하
여 상세한 결과를 제시하였다.
Table. 2-2. 저서미세조류 연구조사 개괄 (항목별 시료수)

- 76 -
(1) 표층 퇴적물 클로로필 농도 변화
(가) 시료의 채취
표층 퇴적물에 서식하는 저서미세조류의 생물량 측정을 위하여, 퇴적물 채취용 주사
기 코어(지름 2.8cm)를 이용하여 표층 0.5cm 깊이까지 3회 반복하여 채취하였다. 채취
한 표층 퇴적물 시료는 50ml 플라스틱 튜브에 넣어 냉동상태로 실험실로 운반하였다.
(나) 시료의 추출 및 엽록소 측정
① 표층 퇴적물 시료내 엽록소(Chl-a 및 peopigments) 농도를 측정하기 위하여, 채취
한 퇴적물 시료의 무게를 측정한 후, 100% 아세톤 15 ml을 이용하여 엽록소를 추출하
였다. 아세톤을 넣은 퇴적물 시료를 약 30분간 초음파로 처리한 후 4℃ 암실에서 24시
간 동안 냉장 추출하였다.
② 추출이 완료된 시료는 1500rpm에서 약 5분간 원심분리를 한후, 상등액 약 10 ml
을 Scintillation vial로 옮겨서 냉장 보관하였다.
③ 시료의 Chl-a 농도를 측정하기 위해, 분광광도계(spectrophotometer)를 이용하여
665, 750 nm에서 상등액 시료의 흡광도를 측정하였다. Pheopigments의 측정을 위해 시
료에 1N HCl 2방울을 첨가하고 5분 후에 665, 750nm에서 흡광도를 다시 측정하였다.
이때, 시료가 산과 충분히 반응하여, pheopigments가 효과적으로 추출될 수 있도록 시
료를 잘 혼합하여 주었다.
④ 퇴적물 시료내 Chl a 및 peopigments의 농도는 다음의 Lorenzen의 식(1967)을 이
용하여 계산하였다. 다른 연구와의 비교를 위하여 단위무게(ug g-1
) 및 단위면적당(mg
m-2
) 농도를 동시에 계산하였다(Table. 2-3).

- 77 -
Calculation:
Chl a = A×K×(665o-665a)×v/(Ws)×L
Pheopigments = A×K×(R[665a]-665o)×v/(Ws)×L
(ug g-1)
Chl a = A×K×(665o-665a)×v/(S/10)×L
Pheopigments = A×K×(R[665a]-665o)×v/(S/10)×L
(mg m-2)
Factor & Description:Value
A: absorption coefficient of Chl a
A: absorption coefficient of Chl a
K: Factor to equate the reduction in the absorbancy
to initial chlorophyll concentration, 1.7:0.7
665o: absorbance before acidification
665a: absorbance after acidification
v: volume of acetone used for extraction (ml)
S: surface area of sediment sampler (cm2)
Ws: sample weight (wet or dry weight)
L: path length of cuvette (cm)
R: maximum ratio of 665o: 665a in the absence of pheopigments
11
11
2.43
variable
variable
variable
variable
variable
variable
1
1.7
Table. 2-3. Chl-a 및 Pheopigments 농도 계산식
(다) 주요 결과 및 토의
낙동강 하구 갯벌, 명지 지역의 저서미세조류의 현존량(생물량)을 파악하기 위해,
2005~2006년 동안 총 6회에 걸쳐 퇴적물 Chl-a 및 pheopigments의 농도를 측정하였
다(Table. 2-4).
2005년 명지 지역 상부(original sediment)의 퇴적물 표층내 존재하는 Chl-a 및
pheopigments의 농도는 5월, 8월, 12월에 각각 18, 122, 11mg m-2로 여름철에 가장 높
고 겨울철에 가장 낮았다. 퇴적물 표층 pheopigmnents의 농도는 여름철에 가장 높았고
(26mg m-2), 상대적으로 저서동물의 활동이 낮은 겨울철에는 여름철에 비해 10배 이상
낮게 나타났다(1.7mg m-2
). 하지만 조고가 낮은 중부 및 하부 지역에서는 이와는 달리

- 78 -
겨울철에 퇴적물 표층 Chl-a 및 pheopigments의 농도가 모두 높은 것으로 확인되었다.
이는 상부와 중하부의 저서미세조류 및 저서동물의 생물상이 다름을 시사한다고 하겠
다. 아울러, pheopigmnts/Chl-a의 비는 상부 및 중부에서 대부분 <1.0 으로 하부에서는
>1.0 것으로 나타나 하부 지역에서의 저서동물의 활동이 보다 활발한 것으로 확인되었
다.
명지 지역 상부, 중부, 하부에 각각 설치된 인공갯벌은 서로 다른 퇴적상(sand,
mixed, mud sediment)을 보이는 3개의 방형구가 각각 설치되어있다. 이중 mixed
sediment는 명지 지역 원래의 퇴적물(original sediment)이며, sand 및 mud sediment는
외부로부터 가져온 퇴적물인데, 인공갯벌 방형구는 모두 4월에 설치된 것이다. 5월의
조사에 따르면, 세가지 서로 다른 퇴적상을 보이는 인공갯벌에서의 Chl-a의 농도는
mud에서 가장 높고(41mg m-2
), mixed (18mg m-2
), sand (14mg m-2
) 순으로 나타났으
나, 8월(122mg m-2
) 및 12월(11mg m-2
)에는 mixed sediment에서 모두 가장 높은 값을
보였다. 그러나 명지 상부, 중부, 하부에 각각 설치된 인공갯벌 모두에서, 퇴적상과 관
계없이 겨울철로 가면서 표층 퇴적물내 Chl-a 및 pheopigments의 농도차가 여름에 비
해 줄어드는 경향을 보였다. 이는 인공갯벌이 설치된 후 시간이 지남에 따라 표층 퇴적
물의 퇴적 등으로 표층의 퇴적상이 유사해지면서, 갯벌 퇴적물내 서식하는 미세조류의
종류나 양이 비슷해짐을 시사한다고 하겠다.
2006년도에는 명지 지역 상부, 중부, 하부 각각에서 서로 다른 세 퇴적상 이외에 추
가로 인공갯벌 방형구 바깥쪽(outer sediment)에서도 추가로 조사가 이루어졌다. 대체
로 세 지역 모두에서 퇴적상에 따라 퇴적물 표층내 존재하는 Chl-a 및 pheopigments의
농도는 유의하게 다른 것으로 나타나 퇴적상이 저서미세조류 및 저서동물의 서식에 영
향을 주는 것으로 확인되었다. 그리고 표층 저서미세조류 생물량은 인공갯벌 방형구 바
깥쪽이 내부에 비해서 2배 이상 높은 것으로 확인되어 자연갯벌이 인공갯벌에 비해 보
다 안정적인 서식환경임을 시사하였다. 대체로 2006년 명지 지역 저서미세조류 생물량
은 2005년과 비교하여 크게 다르지 않은 것으로 나타나 연변화가 계절변화에 비해 상
대적으로 적은 것으로 확인되었다. 인공갯벌 조성 후의 저서미세조류 생물량의 변화를
정확하게 파악하기 위해서는 보다 장기적인 모니터링 자료가 필요하다고 하겠다.
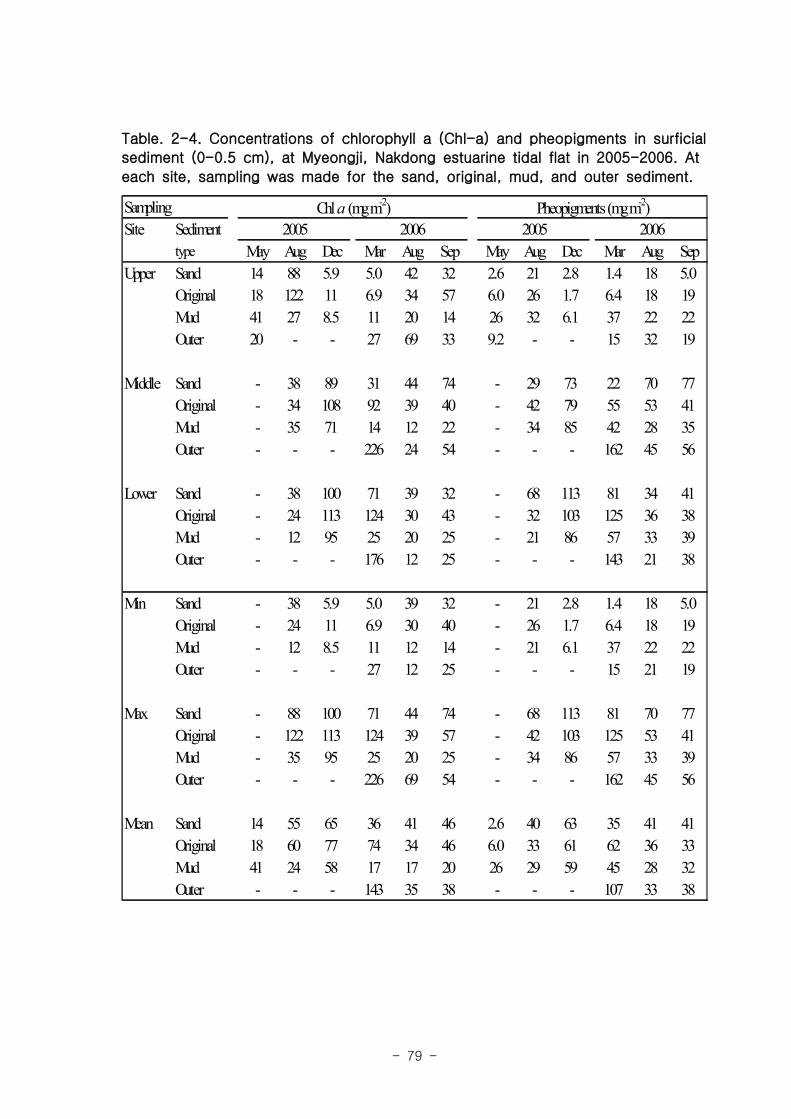
- 79 -
Sampling
Site Sediment 2005 2006 2005 2006
type May Aug Dec Mar Aug Sep May Aug Dec Mar Aug Sep
Upper Sand 14 88 5.9 5.0 42 32 2.6 21 2.8 1.4 18 5.0
Original 18 122 11 6.9 34 57 6.0 26 1.7 6.4 18 19
Mud 41 27 8.5 11 20 14 26 32 6.1 37 22 22
Outer 20 - - 27 69 33 9.2 - - 15 32 19
Middle Sand - 38 89 31 44 74 - 29 73 22 70 77
Original - 34 108 92 39 40 - 42 79 55 53 41
Mud - 35 71 14 12 22 - 34 85 42 28 35
Outer - - - 226 24 54 - - - 162 45 56
Lower Sand - 38 100 71 39 32 - 68 113 81 34 41
Original - 24 113 124 30 43 - 32 103 125 36 38
Mud - 12 95 25 20 25 - 21 86 57 33 39
Outer - - - 176 12 25 - - - 143 21 38
Min Sand - 38 5.9 5.0 39 32 - 21 2.8 1.4 18 5.0
Original - 24 11 6.9 30 40 - 26 1.7 6.4 18 19
Mud - 12 8.5 11 12 14 - 21 6.1 37 22 22
Outer - - - 27 12 25 - - - 15 21 19
Max Sand - 88 100 71 44 74 - 68 113 81 70 77
Original - 122 113 124 39 57 - 42 103 125 53 41
Mud - 35 95 25 20 25 - 34 86 57 33 39
Outer - - - 226 69 54 - - - 162 45 56
Mean Sand 14 55 65 36 41 46 2.6 40 63 35 41 41
Original 18 60 77 74 34 46 6.0 33 61 62 36 33
Mud 41 24 58 17 17 20 26 29 59 45 28 32
Outer - - - 143 35 38 - - - 107 33 38
Chl a (mg m-2) Pheopigments (mg m
-2)
Table. 2-4. Concentrations of chlorophyll a (Chl-a) and pheopigments in surficial
sediment (0-0.5 cm), at Myeongji, Nakdong estuarine tidal flat in 2005-2006. At
each site, sampling was made for the sand, original, mud, and outer sediment.

- 80 -
(2) 시간에 따른 깊이별 클로로필 농도 변화
(가) 시료의 채취
퇴적물 깊이별 미세조류의 생물량 측정을 위한 퇴적물 주상시료의 채취는 깊이 10
cm 까지 1회 혹은 3회 반복하여 채취하였다. 주상시료의 경우는 주사기 코어를 알루미
늄 호일로 쌓아 냉동 보관하여 실험실로 운반한 후, 실험실에서 절단기를 이용하여 깊
이별로(0-2 cm까지는 0.5 cm 간격, 그 이하는 1cm 간격) 12개의 부시료(sub-sample)
를 얻었다.
(나) 시료의 추출 및 엽록소 측정
① 퇴적물 주상 시료내 엽록소(Chl a 및 pheopigments) 농도를 측정하기 위하여, 채
취한 퇴적물 시료의 무게를 측정한 후, 100% 아세톤 15 ml을 이용하여 엽록소를 추출
하였다. 아세톤을 넣은 퇴적물 시료를 약 30분간 초음파로 처리한 후 4℃ 암실에서 24
시간 동안 냉장 추출하였다.
② 추출이 완료된 시료는 1500rpm에서 약 5분간 원심분리를 한후, 상등액 약 10 ml
을 Scintillation vial로 옮겨서 냉장 보관하였다.
③ 시료의 Chl a 농도를 측정하기 위해, 분광광도계(Spectrophotometer)를 이용하여
665, 750 nm에서 상등액 시료의 흡광도를 측정하였다. Pheopigments의 측정을 위해 시
료에 1N HCl 2방울을 첨가하고 5분 후에 665, 750 nm에서 흡광도를 다시 측정하였다.
이때, 시료가 산과 충분히 반응하여, pheopigments가 효과적으로 추출될 수 있도록 시
료를 잘 혼합하여 주었다.
④ 퇴적물 시료내 Chl a 및 pheopigments의 농도는 위의 Lorenzen의 식(1967)을 이
용하여 계산하였다. 다른 연구와의 비교를 위하여 단위무게(ug g-1
) 및 단위면적당(mg
m-2
) 농도를 동시에 계산하였다.(Table. 2-3)
(다) 주요 결과 및 토의
낙동강 하구, 명지 지역의 퇴적물 깊이에 따른 저서미세조류의 생물량 조사는 2005년
에 집중적으로 이루어졌다. 2005년 5월, 8월 12월에 명지 지역 상부의 sand, mixed,
mud sediment로부터 깊이 10 cm까지의 코어 퇴적물을 채취하여 깊이별로 퇴적물내
Chl a의 농도를 측정하였다(Fig. 2-18). 일반적으로 퇴적물 Chl a의 농도는 표층 1 cm
내에 가장 높았고, 깊이 10cm까지의 Chl a 총농도의 약 30% 이상을 차지하는 것으로
확인되었다. 5월과 12월의 퇴적물 Chl a의 수직분포는 퇴적상에 따라 크게 다르지 않았
으나, 8월의 경우는 mud sediment에서 sand 및 mixed sediments와 다른 양상을 보였
다. Sand(88 mg m-2
) 및 mixed sediments(122 mg m-2
)의 경우, 퇴적물 표층의 Chl a
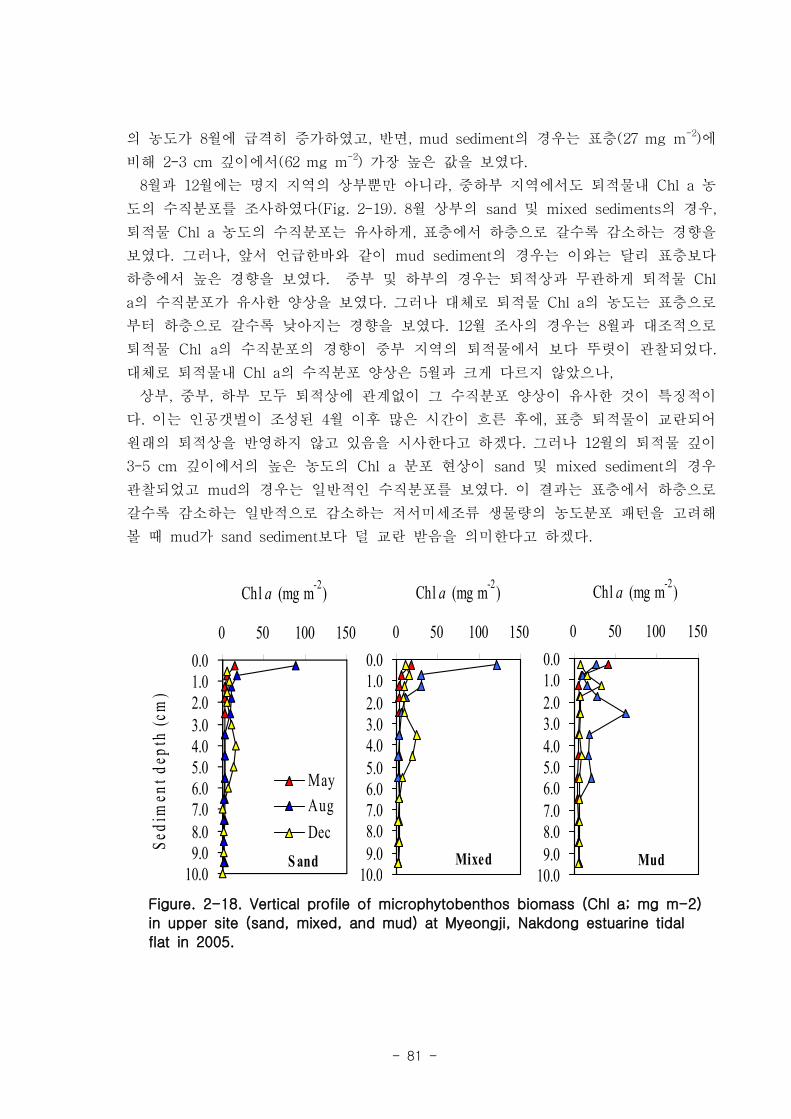
- 81 -
의 농도가 8월에 급격히 증가하였고, 반면, mud sediment의 경우는 표층(27 mg m-2
)에
비해 2-3 cm 깊이에서(62 mg m-2
) 가장 높은 값을 보였다.
8월과 12월에는 명지 지역의 상부뿐만 아니라, 중하부 지역에서도 퇴적물내 Chl a 농
도의 수직분포를 조사하였다(Fig. 2-19). 8월 상부의 sand 및 mixed sediments의 경우,
퇴적물 Chl a 농도의 수직분포는 유사하게, 표층에서 하층으로 갈수록 감소하는 경향을
보였다. 그러나, 앞서 언급한바와 같이 mud sediment의 경우는 이와는 달리 표층보다
하층에서 높은 경향을 보였다. 중부 및 하부의 경우는 퇴적상과 무관하게 퇴적물 Chl
a의 수직분포가 유사한 양상을 보였다. 그러나 대체로 퇴적물 Chl a의 농도는 표층으로
부터 하층으로 갈수록 낮아지는 경향을 보였다. 12월 조사의 경우는 8월과 대조적으로
퇴적물 Chl a의 수직분포의 경향이 중부 지역의 퇴적물에서 보다 뚜렷이 관찰되었다.
대체로 퇴적물내 Chl a의 수직분포 양상은 5월과 크게 다르지 않았으나,
상부, 중부, 하부 모두 퇴적상에 관계없이 그 수직분포 양상이 유사한 것이 특징적이
다. 이는 인공갯벌이 조성된 4월 이후 많은 시간이 흐른 후에, 표층 퇴적물이 교란되어
원래의 퇴적상을 반영하지 않고 있음을 시사한다고 하겠다. 그러나 12월의 퇴적물 깊이
3-5 cm 깊이에서의 높은 농도의 Chl a 분포 현상이 sand 및 mixed sediment의 경우
관찰되었고 mud의 경우는 일반적인 수직분포를 보였다. 이 결과는 표층에서 하층으로
갈수록 감소하는 일반적으로 감소하는 저서미세조류 생물량의 농도분포 패턴을 고려해
볼 때 mud가 sand sediment보다 덜 교란 받음을 의미한다고 하겠다.
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0
9.0
10.0
0 50 100 150
Chl a (mg m-2)
Sedim
ent depth
(cm
)
May
Aug
Dec
S and
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0
9.0
10.0
0 50 100 150
Chl a (mg m-2)
Mixed
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0
9.0
10.0
0 50 100 150
Chl a (mg m-2)
Mud
Figure. 2-18. Vertical profile of microphytobenthos biomass (Chl a; mg m-2)
in upper site (sand, mixed, and mud) at Myeongji, Nakdong estuarine tidal
flat in 2005.
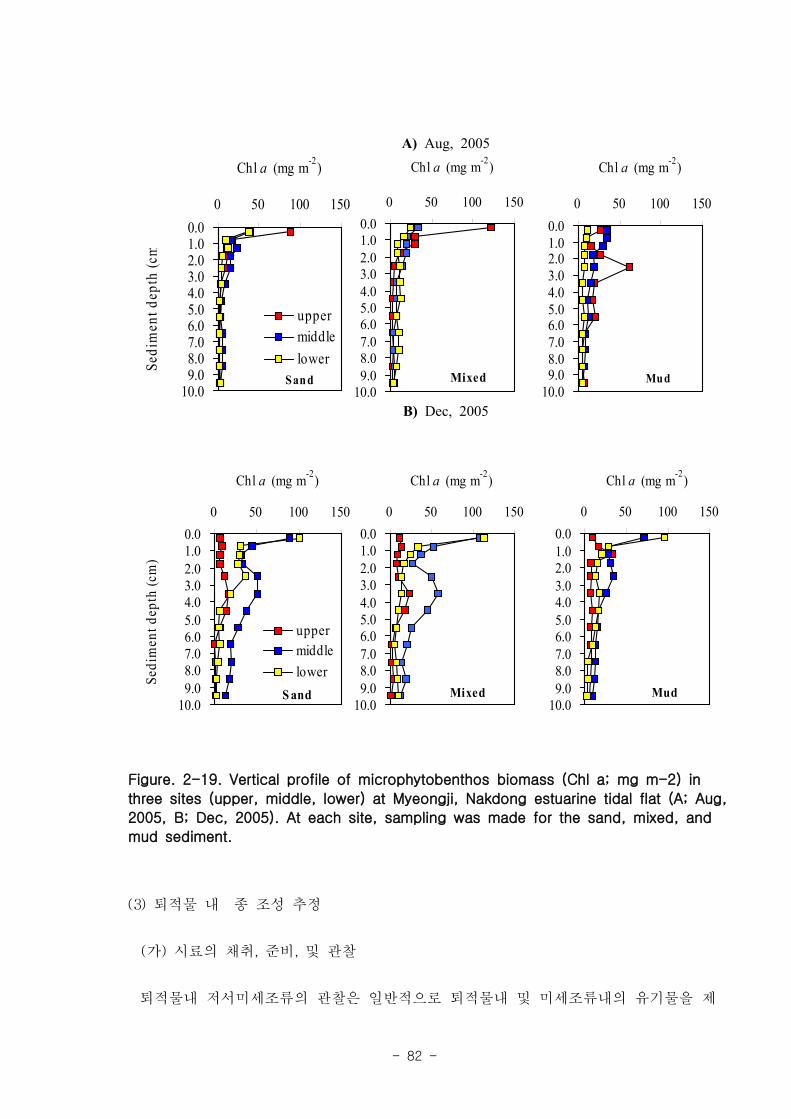
- 82 -
A) Aug, 2005
0.01.02.03.04.05.06.07.08.09.010.0
0 50 100 150
Chl a (mg m-2)
Sedim
ent depth (cm)
upper
middle
lower
Sand
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0
9.0
10.0
0 50 100 150
Chl a (mg m-2)
Mixed
0.0
1.02.0
3.0
4.0
5.06.0
7.0
8.09.0
10.0
0 50 100 150
Chl a (mg m-2)
Mud
B) Dec, 2005
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0
9.0
10.0
0 50 100 150
Chl a (mg m-2)
Sedim
ent depth (cm)
upper
middle
lower
Sand
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0
9.0
10.0
0 50 100 150
Chl a (mg m-2)
Mixed
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0
9.0
10.0
0 50 100 150
Chl a (mg m-2)
Mud
Figure. 2-19. Vertical profile of microphytobenthos biomass (Chl a; mg m-2) in
three sites (upper, middle, lower) at Myeongji, Nakdong estuarine tidal flat (A; Aug,
2005, B; Dec, 2005). At each site, sampling was made for the sand, mixed, and
mud sediment.
(3) 퇴적물 내 종 조성 추정
(가) 시료의 채취, 준비, 및 관찰
퇴적물내 저서미세조류의 관찰은 일반적으로 퇴적물내 및 미세조류내의 유기물을 제

- 83 -
거한 후 미세조류의 세부 구조의 관찰을 바탕으로 이루어진다. 그러나, 유기물 제거 및
산처리 과정은 매우 복잡하고, 살아있는 저서미세조류의 직접적인 관찰에 비해 부적절
할 수 있다. 따라서 본 연구에서는, 살아있는 저서미세조류의 관찰을 위하여 현장에서
채취한 퇴적물 시료를 산처리 과정 없이 분리하여 광학현미경으로 관찰 및 동정하였다.
퇴적물 시료는 표층 상층부 0.5 cm까지 채취하여 scintillation vial에 보관한 후, 냉장상
태로 실험실로 운반하였다.
저서미세조류 관찰용 퇴적물 시료는 다음에 따라 분리하는 과정을 거쳤다.
① 퇴적물 시료가 들어 있는 scintillation vial에 인공 해수를 채운다. 잘 섞일 때까지
흔들어 준 뒤 시료의 절반가량을 plastic bottle(100 ml 용량, 반투명)에 넣고 나머지 반
은 커버 슬라이드 테크닉으로 처리하기 위해 따로 놓아둔다.
② 파라포르말린 10% 용액을 시료에 첨가한다. 이때 첨가하는 양은 시료에 첨가한
인공 해수의 10% 이상이 되도록 하여, 파라포르말린의 최종 농도가 1% 이상이 되도록
한다(이 때 시료 부피는 20 ml 가량임).
③ 시료가 고정될 때까지 몇 시간 동안 기다린 후, 증류수 10 ml을 첨가한다. 시료를
잘 흔들어 3-5초가량 침전 시킨 뒤 100 ml 매스실린더에 상등액만을 따라낸다.
④ 남는 퇴적물에 증류수를 적당량(약 10 ml 정도) 첨가하여 1분간 sonication을 실
시한다. 시료를 잘 흔들어 3-5초가량 침전 시킨 뒤 상등액을 매스실린더에 따라 낸다.
증류수를 조금 더 첨가한 뒤 같은 방법으로 상등액을 한 번 더 따라 낸다.
⑤ 증류수를 첨가하여 5분간 sonication을 실시한다. 시료를 잘 흔들어 3-5초가량 침
전시킨 뒤 상등액을 따라낸다. 상등액이 맑아질 때 까지, 증류수를 첨가하여 상등액을
따라내는 과정을 반복한다. 이 때 매스실린더에 따라낸 시료의 총량이 100 ml을 넘지
않도록 주의한다.
⑥ 매스실린더에 약간의 파라포르말린 용액을 넣어 다시 고정시킨다. 이미 한번 고정
되었기 때문에 적은 양만을 넣어도 충분하다. Plastic bottle에 남아 있는 퇴적물에도 약
간의 파라포르말린 용액을 첨가한다. 후에 이 퇴적물을 과산화수소로 처리하여
episammic diatom을 분리해 낸다.
⑦ 매스실린더에 종이 등을 덮어 먼지가 들어가지 않도록 한 뒤 하루 정도 놓아둔다.
하루가 지난 뒤, 매스실린더를 바닥에 세 번 정도 약하게 내리친다. 이 때 물방울 등과
함께 벽에 붙어 있던 일부 시료가 침전하게 되며, 경우에 따라서는 다음 과정으로 넘어
가지 전에 30분 가량 기다려야 한다.
⑧ 피펫을 이용하여 상등액을 최대한 따라낸다. 시료를 잘 흔들어 준 뒤 최대한 빠른
속도로 scintillation vial에 따라낸다. 아주 약간의 증류수를 첨가한 뒤 매스실린더에
남은 시료를 씻어 내어 scintillation vial에 따라낸다. scintillation vial에 파라포르말린
용액을 넣어 고정시킨다.
(나) 주요 결과 및 토의

- 84 -
낙동강 하구, 명지 지역에서 퇴적물 표층에 서식하는 저서미세조류의 관찰을 위해,
2005년 5월과 8월, 그리고 2006년 3월과 9월에 표층퇴적물(0-0.5 cm)을 채취하였다. 광
학현미경을 이용하여 살아있는 저서미세조류를 관찰 하고, 우점종 리스트(Table. 2-5)
와 도판(Fig. 2-20, Fig. 2-21, Fig. 2-22, Fig. 2-23)을 제시하였다. 중심돌말류 (centric
diatom)는 Thalassiosira bramaputrae와 Thalassiosira sp.1의 두 개 종이, 깃털돌말류
(pennate diatom)는 Achnanthes brevipes, Amphora coffeaeformis, Amphora proteus,
Bremis ambigua, Caloneis crassa, Cylindrotheca closterium, Cylindrotheca gracilis,
Cymbella sp.1, Diploneis smithii, Entomoneis paludosa, Fallacia cf. littoralis,
Gomphonema sp.1, Gyrosigma fasciola, Gyrosigam wansbeckii, Hanzschia
amphioxys, Hippodonta sp.1, Navicula digitoconvergence, Navicula salinarum var.
minima, Navicula sp.1, Navicula sp.2, Navicula sp.3, Navicula sp. 4, Nitzshcia
hungarica, Nitzschia lacunarum, Nitzschia palea, Nitzschia cf. pusilla, Nitzschia
spathulata, Nitzshcia sp.1, Nitzschia sp.2, Parilbellus sp.1, Petroneis marina,
Pleurosigma delictulum, Seminavis strigosa, Surirella fastuosa var. recedens 그리고
Tryblionella levidensis의 35종이 출현하였다. 중심돌말류는 흔히 부유성으로 여겨지며
깃털돌말류는 흔히 저서성으로 여겨지므로(Round et al., 1990), 명지 지역에서 관찰된
저서미세조류는 저서성규조류가 매우 우점함을 알 수 있다.
명지 지역에서 관찰된 우점종의 목록은 전반적으로 큰 변화를 보이는데, (Table. 2-5)
37개 종 중 7개 종만이 4번 이상 우점종으로 기록되었다. 이들 7개 종 중에 Amphora
coffeaeformis, Navicula digitoconvergence, 그리고 Navicula sp.2의 세 종은 총 8번 우
점종으로 기재되어, 계절 및 퇴적상에 관계없이 명지 갯벌에 널리 우점하는 것으로 나
타났다. 이 외에 Amphora proteus 및 Navicula sp.1이 각각 7번 및 6번 우점종으로 기
록되었다. 계절에 따른 우점종 수의 관점에 있어서는 3월이 17개 종, 5월 11개 종, 8월
16개 종, 9월 14개 종으로 나타났다. 이는 일반적으로 식물플랑크톤이 봄에 가장 번성
하는 경향과 대체로 일치한다고 하겠다.
퇴적상과 관련하여서 2005년에는 입도와 종 조성간에 관련성이 있는 것으로 파악되
었다. sand 및 mixed 퇴적상에서는 Navicula 종들이 많이 우점한 반면 mud 퇴적상에
서는 Nizschia 종들이 많이 우점하였다. 그러나 이러한 경향은 2006년에 들어서 전혀
관찰되지 않는다. 이는 2006년 9월에 관찰된 Navicula 및 Nizschia 종의 수 자체가 적
었기 때문인데, 시료 채취 이전에 발생하였던 태풍에 의한 담수의 대량 유입 및 퇴적물
교란과 상관성이 있는 것으로 여겨진다. 이 이외에도 2005년과 2006년의 우점종은 몇몇
종의 출현에 있어 뚜렷한 차이를 보이는데, 2005년에만 각각 세 번 우점종으로 기록되
는 등 비교적 널리 분포하였던 Fallacia cf. littoralis, Hippodonta sp.1, 그리고
Nizschia cf. pusilla는 2006년에는 우점종으로서 관찰된 바가 없었다. 반면
Cylindrotheca gracilis, Navicula sp.4, Petroneis marina의 경우는 2005년에는 우점종
으로 관찰된 바가 없었으나 2006년에는 각각 세번씩 우점종으로서 기록되었다.
명지 갯벌 정점에 인공적으로 조성된 방형구와 기존의 갯벌 흙을 채워 넣은 방형구,

- 85 -
그리고 방형구 바깥쪽의 기존 갯벌 구역에서의 우점종은 그 수에 있어서 상당한 차이
를 보였다. 전반적으로 기존의 갯벌 흙을 이용하여 조성한 방형구 및 방형구 바깥쪽 갯
벌 구역에서의 우점종의 수가 훨씬 많았으며, 인공적으로 조성된 방형구에 있어서는 우
점종의 수가 현저히 적은 것이 관찰되었다. 단 2006년 9월의 경우에는 이러한 경향이
관찰되지 않았으며 각 방형구에서의 우점종의 수가 비교적 일정하였다. 이 역시 앞서
언급한 바와 같이 2006년 여름 태풍에 의한 담수 유입 및 퇴적물 교란으로 인해 우점
종의 수 자체가 변한 것에 기인하는 것이라 생각된다.
전반적으로 명지 갯벌에서의 우점종의 변화는 항상 우점하는 몇몇 종의 경우를 제외
하고는 매우 불규칙하였다. 조사가 이루어진 정점 자체가 인위적으로 조성된 인공 갯벌
임을 감안하면, 이러한 불규칙성은 저서규조류 군집 자체의 속성이라기 보다는 변화가
많은 주변 환경의 영향에 의한 것으로 여겨진다.

- 86 -
Species May Aug Aug Mar Sep Sep
Sand Original Mud Sand Original Mud
Centric diatoms
Thalassiosira bramaputrae +
Thalassiosira sp. 1
Pennate diatoms
Achnanthes brevipes + +
Amphora coffeaeformis + + + + + + + +
Amphora proteus + + + + + + +
Bremis ambigua + +
Caloneis crassa +
Cylindrotheca closterium +
Cylindrotheca gracilis + + +
Cymbella sp. 1 +
Diploneis smithii +
Entomoneis paludosa +
Fallacia cf. littoralis + + +
Gomphonema sp. 1 +
Gyrosigma fasciola
Gyrosigma wansbeckii +
Hantzschia amphioxys +
Hippodonta sp. 1 + + +
#avicula digitoconvergence + + + + + + + +
#avicula salinarum var. minima +
#avicula sp. 1 + + + + + +
#avicula sp. 2 + + + + + + + +
#avicula sp. 3 + +
#avicula sp. 4 + + +
#itzschia hungarica +
#itzschia lacunarum + + + +
#itzschia palea + + + +
#itzschia cf. pusilla + + +
#itzschia spathulata +
#itzschia sp. 1 + +
#itzschia sp. 2 + +
Parlibellus sp. 1 + + +
Petroneis marina + + +
Pleurosigma delicatulum +
Seminavis strigosa +
Surirella fasctuosa var. recedens
Tryblionella levidensis +
Year 2005 Year 2006
Outer Outer
Table. 2-5. List of the major benthic diatoms found in surficial sediment
(0-0.5 cm), at Myeongji (upper site only), Nakdong estuarine tidal flat in 2005
- 2006. (+ indicates the occurrence of benthic diatoms in corresponding
sites)
- 87 -
Figure. 2-20. Benthic diatoms found in surficial sediment (0-0.5 cm), at
Myeongji, Nakdong estuarine tidal flat in 2005-2006. Scale bar = 10 um.
- 88 -
Figure. 2-21. Benthic diatoms found in surficial sediment (0-0.5 cm), at
Myeongji, Nakdong estuarine tidal flat in 2005-2006 (continued). Scale bar =
10 um.

- 89 -
Figure. 2-22. Benthic diatoms found in surficial sediment (0-0.5 cm), at
Myeongji, Nakdong estuarine tidal flat in 2005-2006 (continued). Scale bar
= 10 um.
- 90 -
Figure. 2-23. Benthic diatoms found in surficial sediment (0-0.5 cm), at
Myeongji, Nakdong estuarine tidal flat in 2005-2006 (continued). Scale
bar = 10 um.

- 91 -
(4) 낙동강 하구역 저서 미세식물 군집의 시공간 변동
(가) 조사 개괄
조간대 지역은 육상 환경과 해양환경이 만나는 곳으로 우리나라의 서해안과 남해안
에는 국토 면적의 2.8%에 해당하는 광대한 조간대 갯벌이 발달되어 있다(해양수산부,
1998). 갯벌 조간대에는 육상으로부터 많은 양의 유기물과 영양염이 공급되어 비옥한
영양조건을 갖고 있으며 적절한 수분유지와 다양한 서식환경으로 다양한 해양생물들이
높은 밀도로 서식하고 있어 면적에 비해 동 • 식물의 생산성이 매우 높은 곳으로 알려
져 있으며(Warwick and Price, 1975), 이러한 갯벌조간대의 생산성은 일반적으로 외해
양에 비해 약 10-20배 가량 높고, 이를 바탕으로 다양한 해양생물들의 산란장과 서식처
로 이용되고 있다(조, 2005).
갯벌의 높은 생산성의 바탕에는 이를 뒷받침하는 다양한 식물군이 있으며, 특히 펄
갯벌에서는 저서미세조류군집이 있다. 펄 갯벌 표면에 서식하는 저서미세조류로는 돌말
류, 남조세균, 유글레나류, 녹조류 등 다양한 미세조류가 출현하고 있으나 주로 저서성
돌말류가 우점한다. 저서성 돌말류는 퇴적물 내에서 먹이그물을 연결하는 중요한 탄소
공급원이며(Sullivan and Moncreiff, 1988), 퇴적물을 안정화 시키고 퇴적물과 수층사이
의 영양염 순환을 조절하는 구성원으로 다양한 기능과 역할을 담당하고 있다
(Nybakken, 2001).
이러한 저서미세조류군집의 생태 역할과 지위에 대한 연구는 저서 돌말류 개체군을
중심으로 이루어져 왔다(Sullivan, 1975). 생산력에 관한 연구는 갯벌 조간대에서의 일
차생산력에 대한 평가가 이루어지고 있으나 전반적으로 미비한 실정이다. 우리나라 서
해안 조간대에서의 저서미세조류 연구는 인천연안 갯벌(심과 조, 1984), 금강하구 조간
대(김과 조, 1985), 송도 갯벌(고, 1991) 등에서 주로 현미경 관찰을 바탕으로 한 저서규
조류 군집 연구가 주로 이루어져 왔고, 최근에 곰소만 갯벌(이, 2001), 강화도 갯벌(최,
2002), 새만금 갯벌(오 등, 2004) 등에서도 연구가 진행되었다. 본 연구는 많은 갯벌들
가운데 최근 생태계 훼손에 대한 연구가 이루어지고 있는 낙동강 하구역의 습지 생태
계의 돌말류 군집에 대해 진우도, 장자도, 하구언, 백합 의 네 정점에서 저서미세조류의
현미경 관찰을 바탕으로 정점별, 계절별, 퇴적물 깊이별 특징을 조사하였다. 확인된 종
의 상대적인 분포와 변동을 바탕으로 저서미세조류군집의 생태특성을 파악하고자 하였
다.
(나) 재료와 방법
① 현장 조사
조간대 퇴적물에 서식하는 저서미세조류 군집의 생태 특성을 분석하기 위하여, 낙동
강 하구역의 4개 정점에서 계절별로 조사하였다 (Fig. 2-24). 지정한 장소에서 지름(내

- 92 -
경) 8.3cm, 길이 34cm 플라스틱 원통을 사용하여 표면에서 연직 방향으로 주상시료를
채집하였다. 채집한 시료는 현장에서 1cm씩 원반으로 잘라 최대한 빨리 냉장 운반하여
깊이별 엽록소 a 함량과 종 조성을 분석하였다.
Figure. 2-24. 낙동강 하구역 조사 정점 - 진우도(가), 장자도(나),
명지(다), 백합등(라)
② 엽록소 a 측정
퇴적물의 엽록소 a는 90% 아세톤으로 24시간 동안 냉암소에서 추출하였다. 엽록소 a
함량은 분광 광도계로 흡광도를 측정하여 엽록소 함량과 phaeo-pigments(분해된 엽록
소) 함량(μg chlorophyll a cm-3)을 추정하였다(Parsons et al.. 1984).
③ 현미경 관찰
광학 현미경을 이용하여 조간대 표층 퇴적물의 저서미세조류 군집의 정량적인 계수
와 정성적인 특성을 조사하였다. 현미경 시료는 깊이별로 잘라 얻은 원반 시료의 일부
를 지름 7.2mm 길이 10mm 소형 원반시료 채취기로 취한 퇴적물 중 63㎛ 표준체를 통
과한 시료의 일부를 검경 계수하였다. 계수용 슬라이드에서 무작위로 계수한 개체수를
면적으로 환산하여 조사정점의 개체군 밀도를 추정하였다. 개체의 종 동정은 가능한 한
종 수준까지 확인하였다. 종의 동정은 Jensen(1985), Round(1990), Yamaji(1991),
Thomas(1996), 심(1994) 등을 참고하였다.
④ 군집 생태 특성
광학 현미경으로 계수하여 환산한 개체군의 현존량 자료를 바탕으로 풍부도 지수, 다
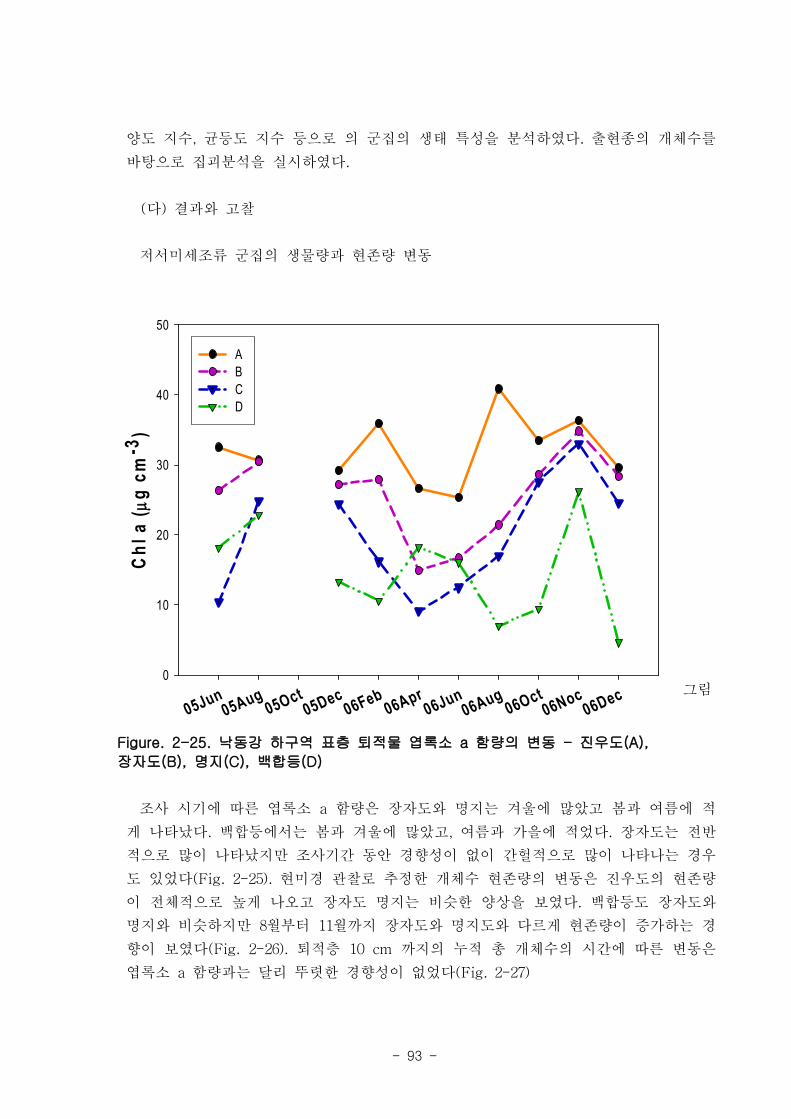
- 93 -
양도 지수, 균등도 지수 등으로 의 군집의 생태 특성을 분석하였다. 출현종의 개체수를
바탕으로 집괴분석을 실시하였다.
(다) 결과와 고찰
저서미세조류 군집의 생물량과 현존량 변동
05Jun05Aug
05Oct05Dec
06Feb06Apr
06Jun06Aug
06Oct06Noc
06Dec
Ch
l a
(µ
g c
m-3
)
0
10
20
30
40
50
A
B
C
D
Figure. 2-25. 낙동강 하구역 표층 퇴적물 엽록소 a 함량의 변동 - 진우도(A),
장자도(B), 명지(C), 백합등(D)
그림
조사 시기에 따른 엽록소 a 함량은 장자도와 명지는 겨울에 많았고 봄과 여름에 적
게 나타났다. 백합등에서는 봄과 겨울에 많았고, 여름과 가을에 적었다. 장자도는 전반
적으로 많이 나타났지만 조사기간 동안 경향성이 없이 간헐적으로 많이 나타나는 경우
도 있었다(Fig. 2-25). 현미경 관찰로 추정한 개체수 현존량의 변동은 진우도의 현존량
이 전체적으로 높게 나오고 장자도 명지는 비슷한 양상을 보였다. 백합등도 장자도와
명지와 비슷하지만 8월부터 11월까지 장자도와 명지도와 다르게 현존량이 증가하는 경
향이 보였다(Fig. 2-26). 퇴적층 10 cm 까지의 누적 총 개체수의 시간에 따른 변동은
엽록소 a 함량과는 달리 뚜렷한 경향성이 없었다(Fig. 2-27)
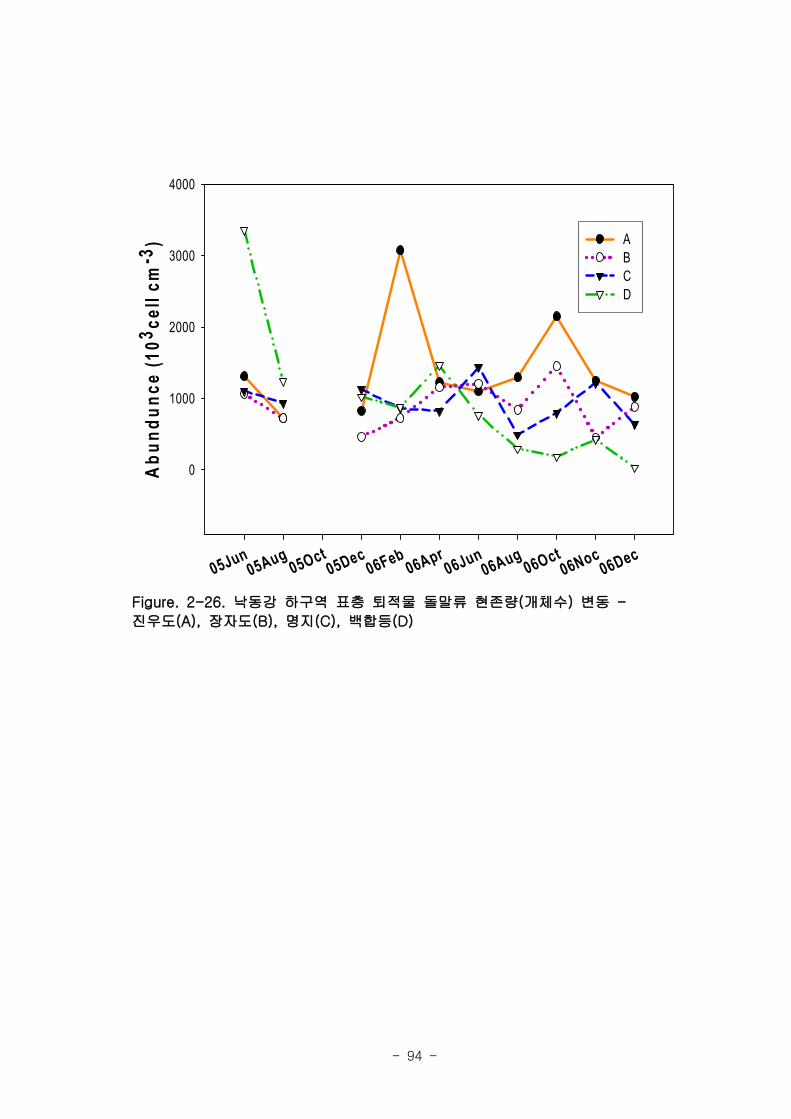
- 94 -
05Jun05Aug
05Oct05Dec
06Feb06Apr
06Jun06Aug
06Oct06Noc
06Dec
Ab
un
du
nc
e (
10
3c
ell
cm
-3)
0
1000
2000
3000
4000
A
B
C
D
Figure. 2-26. 낙동강 하구역 표층 퇴적물 돌말류 현존량(개체수) 변동 -
진우도(A), 장자도(B), 명지(C), 백합등(D)

- 95 -
A
0 200 400 600 800 1000
Depth
s (c
m)
0
1
2
3
4
5
6
7
8
9
10
0 2 4 6 8 10
B
Abundance (103cell cm-3)0 200 400 600 800 1000
chla (µg cm-3)
0 2 4 6 8 10 12 14
C
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
D
0 500 1500 2000
0 2 4 6 8 10 12 14
2005.06
A
0 200 400 600 800 1000
Depth
s (c
m)
0
1
2
3
4
5
6
7
8
9
10
0 2 4 6 8 10 12
B
Abundance (103cell cm-3)0 200 400 600 800 1000
chla (µg cm-3)
0 2 4 6 8 10 12 14
C
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
D
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
2005.08
A
0 200 400 600 800 1000
Depth
s (c
m)
0
1
2
3
4
5
6
7
8
9
10
1.5 2.0 2.5 3.0 3.5 4.0 4.5
B
Abundance (103cell cm-3)0 200 400 600 800 1000
chla (µg cm-3)
0 2 4 6 8 10 12 14
C
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
D
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
2005.12
A
0 200 400 600 1200 1400
Depth
s (c
m)
0
1
2
3
4
5
6
7
8
9
10
0 2 4 6 8 10 12 14
B
Abundance (103cell cm-3)0 200 400 600 800 1000
chla (µg cm-3)
0 2 4 6 8 10 12 14
C
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
D
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
2006.02
0 200 400 600 800 1000
Depth
s(c
m)
0
1
2
3
4
5
6
7
8
9
100 2 4 6 8 10 12 14
0 200 400 600 800 1000
Chl a (µg cm-3)
0 2 4 6 8 10 12 14
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
2006.04
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
Abundance (103cells cm-3)
A B C D A
0 200 400 600 800 1000
Depth
s (c
m)
0
1
2
3
4
5
6
7
8
9
10
Chl a(µg cm-3)
0 2 4 6 8 10 12 14
B
Abundance(103cell cm-3)0 200 400 600 800 1000
0 2 4 6 8 10 12 14
C
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
D
0 2 4 6 8 10 12 14
0 200 400 600 800 1000
2006.06
A
0 200 400 600 800 1000
Depth
s (c
m)
0
1
2
3
4
5
6
7
8
9
10
0 2 4 6 8 10 12 14
B
Abundance (103cell cm-3)0 200 400 600 800 1000
chla (µg cm-3)
0 2 4 6 8 10 12 14
C
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
D
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
2006.08
A
Abundance (103cell cm-3)0 200 400 600 800 1000
Depth
s (c
m)
0
1
2
3
4
5
6
7
8
9
10
0 2 4 6 8 10 12 14
B
0 200 400 600 800 1000
chla (µg cm-3)
0 2 4 6 8 10 12 14
C
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
D
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
2006.9
A
0 200 400 600 800 1000
Depth
s (c
m)
0
1
2
3
4
5
6
7
8
9
10
Chl a(µg cm-3)
0 2 4 6 8 10 12 14
B
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
C
Abundance(103cell cm
-3)
0 2 4 6 8 10 12 14
0 200 400 600 800 1000
D
0 2 4 6 8 10 12 14
0 200 400 600 800 1000
2006.11
A
0 200 400 600 800 100012001400
Depth
s (c
m)
0
1
2
3
4
5
6
7
8
9
10
0 2 4 6 8 10 12 14
B
Abundance (103cell cm-3)0 200 400 600 800 1000
chla (µg cm-3)
0 2 4 6 8 10 12 14
C
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
D
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
2006.12
A
0 200 400 600 800 1000
Depth
s (c
m)
0
1
2
3
4
5
6
7
8
9
10
Chl a(µg cm-3)
0 2 4 6 8 10 12 14
B
0 200 400 600 800 1000
0 2 4 6 8 10 12 14
C
Abundance(103cell cm
-3)
0 2 4 6 8 10 12 14
0 200 400 600 800 1000
D
0 2 4 6 8 10 12 14
0 200 400 600 800 1000
2006.10
Figure. 2-27. 낙동강 하구역 표층 퇴적물에 분포하는 깊이별 저서 돌말류의
개체수와 엽록소 a 함량의 분포 - 진우도(A), 장자도(B), 명지(C), 백합등(D)

- 96 -
엽록소 a함량과 개체 수 모두 깊이가 깊어질수록 감소하는 경향을 보였다. 깊이별 개
체수와 엽록소 a함량의 분포이동은 다르게 나타났다.
개체 수는 백합등 정점에서 가장 적게 나오고 깊이가 깊어질수록 줄어드는 경향이
가장 뚜렷하게 나타났다. 진우도, 장자도 정점에서 개체수가 많으며 깊이 변화에 대한
차이가 뚜렷하게 나타나지 않았다.
엽록소 a함량이 깊이별로 깊어질수록 줄어드는 경향은 대부분의 지역에서 보이며 명
지 정점에서 가장 뚜렷이 나타난다. 진우도, 장자도 정점에서 엽록소 a함량 값이 높게
나타났고 백합등에서는 낮게 나타났다. 11월의 백합등정점에서 다른 조사 시기와 비교
하여 엽록소 a함량 값이 높게 나타나고 엽록소 a함량이 중간 깊이에서 높아지는 다른
양상을 보였다.
조사기간 동안 주요 우점종은 Amphora coffeaformis, Navicula fortis, Thalassionema
sp., Navicula sp. 등 이었다. 조사시기와 정점에 따라 Caloneis sp., Coscinodiscus sp.,
Navicula digitoradiata, N. ramosissima, Leptocylindrys sp. 도 우점종으로 분포하였
다(Table. 2-6).
진우도 정점의 우점하는 종류의 수는 적었으며 백합등에서 우점종의 종 수가 상대적
으로 많았다. 시기별로 보면 8월에 우점종의 종수는 상대적으로 작았다. Amphora
coffeaformis는 모든 조사 정점에서 확인 되었으며 특히 진우도, 장자도 정점에서 뚜렷
하게 우점하고 있었고. 명지, 백합등 정점에서는 Navicula fortis 가 가장 빈번하게 나
타났다.
주사전자현미경(SEM) 사진Amphora coffeaeformis Navicula sp.
광학현미경(LM) 사진Navicula fortis Amphora coffeaeformis
Table. 2-6. 낙동강 하구역 우점종
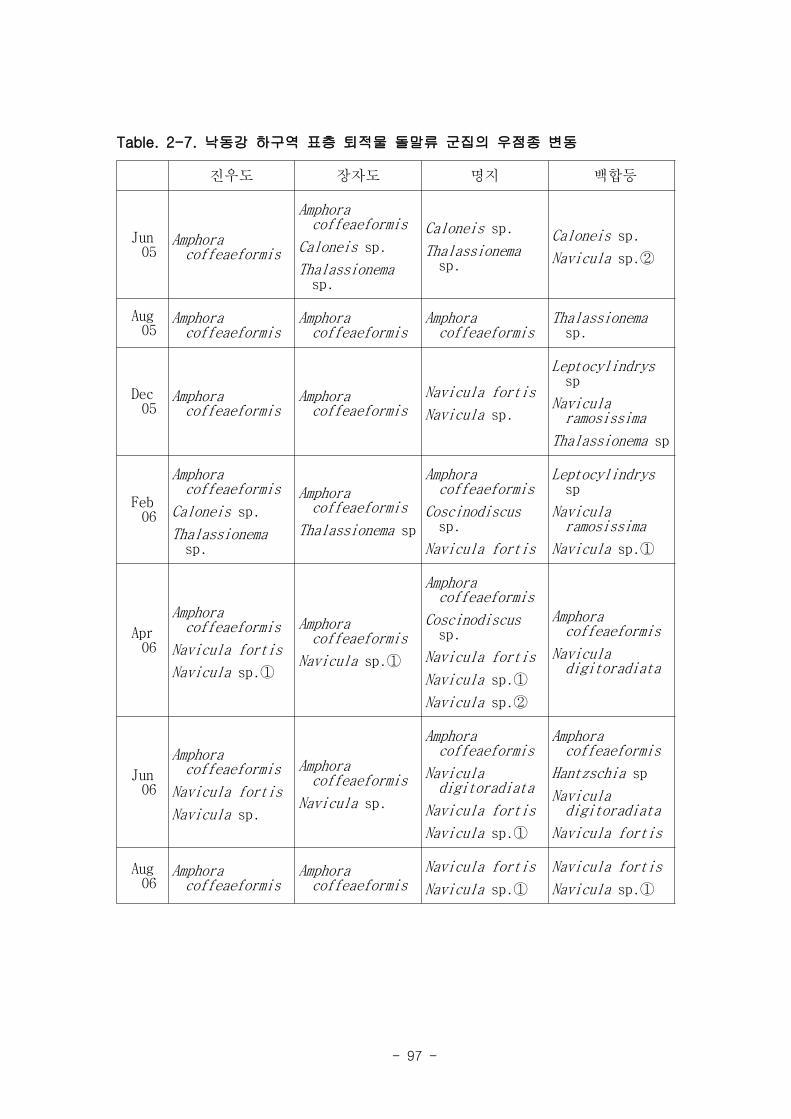
- 97 -
진우도 장자도 명지 백합등
Jun 05
Amphora coffeaeformis
Amphora coffeaeformis
Caloneis sp.
Thalassionema sp.
Caloneis sp.
Thalassionema sp.
Caloneis sp.
Navicula sp.②
Aug 05
Amphora coffeaeformis
Amphora coffeaeformis
Amphora coffeaeformis
Thalassionema sp.
Dec 05
Amphora coffeaeformis
Amphora coffeaeformis
Navicula fortis
Navicula sp.
Leptocylindrys sp
Navicula ramosissima
Thalassionema sp
Feb 06
Amphora coffeaeformis
Caloneis sp.
Thalassionema sp.
Amphora coffeaeformis
Thalassionema sp
Amphora coffeaeformis
Coscinodiscus sp.
Navicula fortis
Leptocylindrys sp
Navicula ramosissima
Navicula sp.①
Apr 06
Amphora coffeaeformis
Navicula fortis
Navicula sp.①
Amphora coffeaeformis
Navicula sp.①
Amphora coffeaeformis
Coscinodiscus sp.
Navicula fortis
Navicula sp.①
Navicula sp.②
Amphora coffeaeformis
Navicula digitoradiata
Jun 06
Amphora coffeaeformis
Navicula fortis
Navicula sp.
Amphora coffeaeformis
Navicula sp.
Amphora coffeaeformis
Navicula digitoradiata
Navicula fortis
Navicula sp.①
Amphora coffeaeformis
Hantzschia sp
Navicula digitoradiata
Navicula fortis
Aug 06
Amphora coffeaeformis
Amphora coffeaeformis
Navicula fortis
Navicula sp.①
Navicula fortis
Navicula sp.①
Table. 2-7. 낙동강 하구역 표층 퇴적물 돌말류 군집의 우점종 변동
- 98 -
Sep 06
Amphora coffeaeformis
Amphora coffeaeformis
Navicula fortis
Amphora coffeaeformis
Navicula sp.②
Amphora lineata
Navicula fortis
Navicula sp.①
Navicula sp.②
Oct 06
Amphora coffeaeformis
Amphora lineata
Amphora sp.
Amphora coffeaeformis
Navicula fortis
Navicula sp.①
Amphora coffeaeformis
Navicula fortis
Navicula ramosissima
Navicula sp.①
Nov.06
Amphora sp.1
A m p h o r a
coffeaeformis
A m p h o r a
coffeaeformis
Navicula fortis
A m p h o r a
coffeaeformis
Navicula fortis
Navicula sp.①
H a n t z s c h i a
amphioxys
Navicula fortis
N a v i c u l a
ramosissima
Navicula sp.①
Dec.06
A m p h o r a
coffeaeformis
Navicula fortis
A m p h o r a
coffeaeformis
Navicula sp.②
A c h n a n t h e s
brevipes
Navicula sp.①
A m p h o r a
coffeaeformis
Hantzschia sp.①
간조 시기와 맞추어 깊이별 생물량 혹은 돌말류의 개체수 변동을 2005년 6월과 2006
년 8월에 2회 조사하였다. 조석 주기에 따라 퇴적층에 서식하는 돌말류의 깊이별 생물
량과 개체수의 변화는 확인되었지만, 주기성을 보여주는 조석과 빛과 같은 다른 물리
요인의 변화와 관련하여 변하는 경향성은 본 조사에서는 나타나지 않았다. 조석과 파도
에 의한 부유와 퇴적, 해수의 유동과 파도에 의한 침식 등과 같은 물리적인 교란 요소
는 물론, 저서동물의 생물교란과 조식 등에 따라 상당한 변화가 예상되지만 자료의 부
족으로 일부 제한된 해석만이 가능하였다. 간조시 완전히 노출된 시간을 기점으로 전후
2 - 3 시간동안의 엽록소a 함량의 깊이별 변화는 뚜렷한 경향성이 없었지만, 간조 후
표층이 노출되는 시점에서 포층의 생물량이 증가하는 경향이 다소 있는 것으로 나타났
다. 조사 시기의 자료를 각각 독립적으로 분석하면 대체적으로 낮 시간대 간조시기의
연직 분포의 변화가 밤 시간대의 간조 시기의 분포보다 복잡하게 나타났다. 중간층
5cm 깊이의 증가가 관찰되었다. 계수한 돌말류의 개체수와 엽록소 a 함량과의 상관관
계는 거의 없었다.
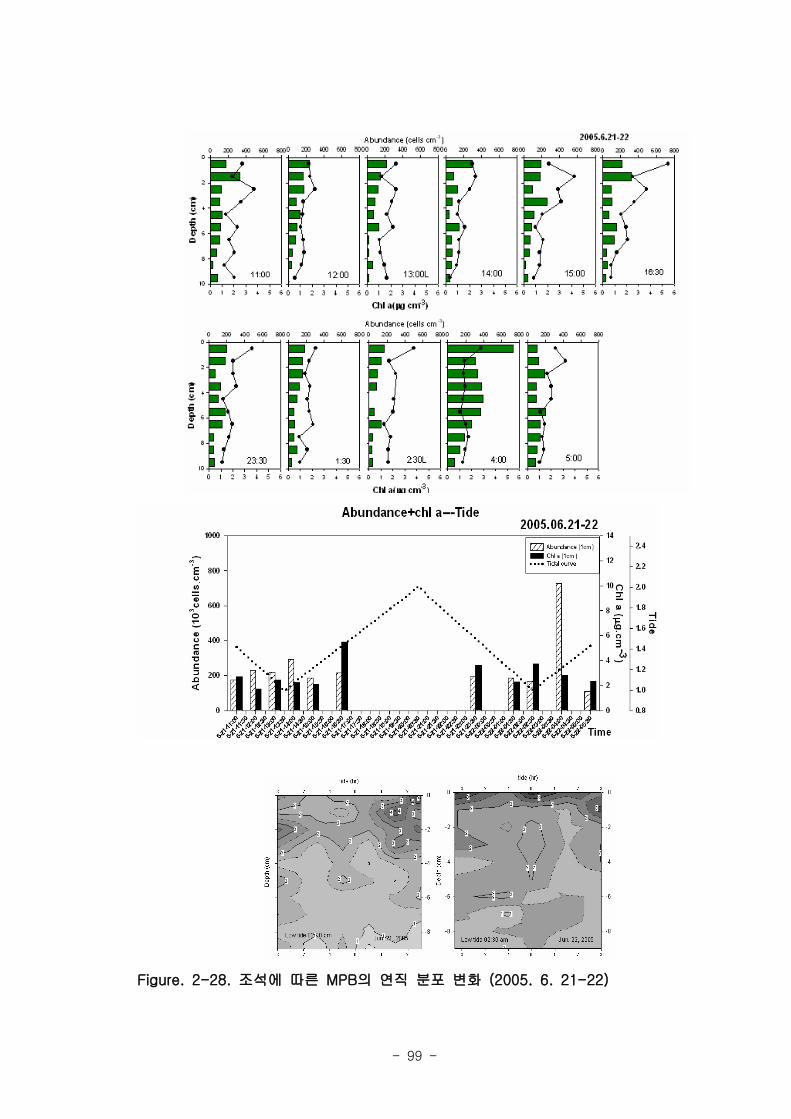
- 99 -
Figure. 2-28. 조석에 따른 MPB의 연직 분포 변화 (2005. 6. 21-22)
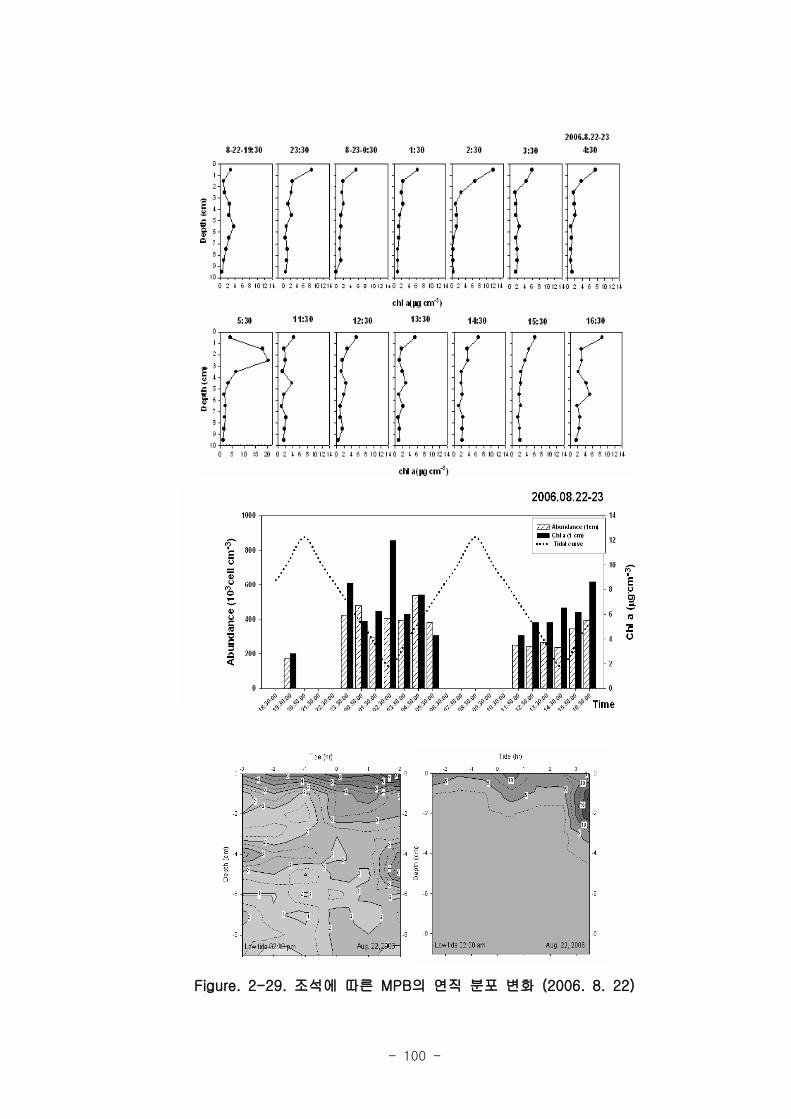
- 100 -
Figure. 2-29. 조석에 따른 MPB의 연직 분포 변화 (2006. 8. 22)
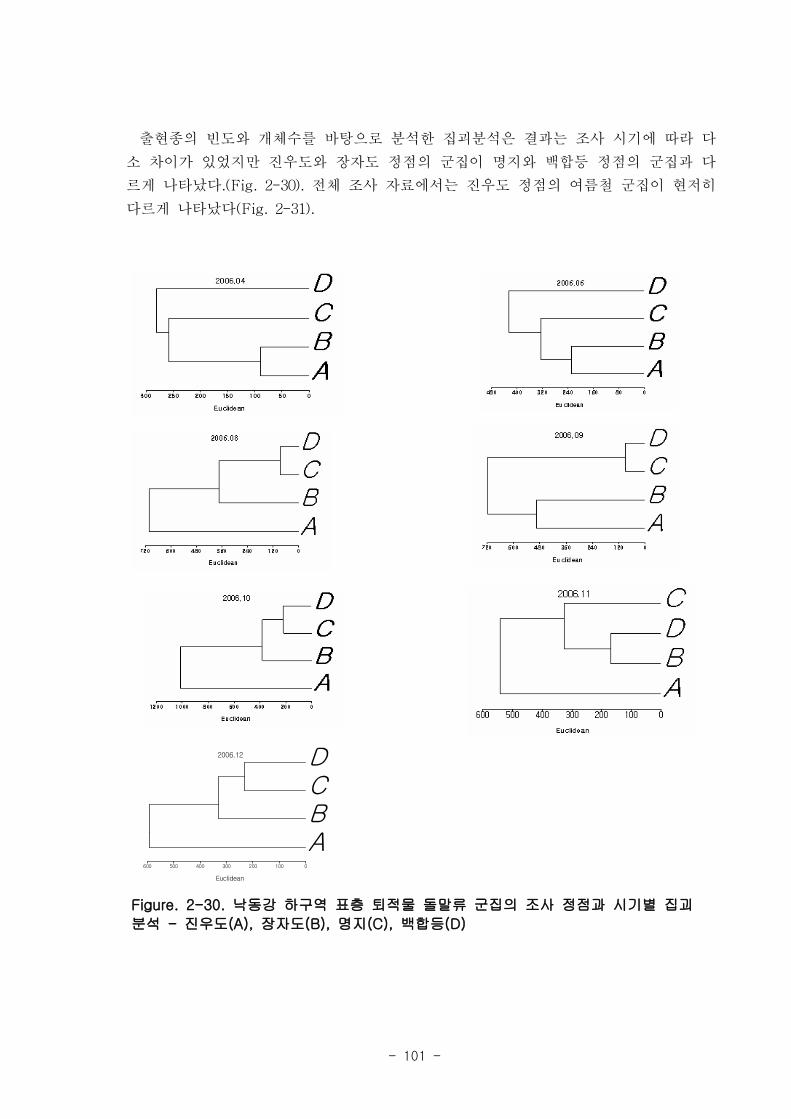
- 101 -
출현종의 빈도와 개체수를 바탕으로 분석한 집괴분석은 결과는 조사 시기에 따라 다
소 차이가 있었지만 진우도와 장자도 정점의 군집이 명지와 백합등 정점의 군집과 다
르게 나타났다.(Fig. 2-30). 전체 조사 자료에서는 진우도 정점의 여름철 군집이 현저히
다르게 나타났다(Fig. 2-31).
2006.12
Euclidean
ABCD
600 500 400 300 200 100 0
Figure. 2-30. 낙동강 하구역 표층 퇴적물 돌말류 군집의 조사 정점과 시기별 집괴
분석 - 진우도(A), 장자도(B), 명지(C), 백합등(D)

- 102 -
Figure. 2-31. 낙동강 하구역 표층 퇴적물 돌말류 군집의 집괴 분석 - 진우도(A),
장자도(B), 명지(C), 백합등(D)

- 103 -
2. 소비자
가. 저서동물
하구역은 육지로부터 공급되는 담수와 해수의 혼합으로 희석된 염분농도를 가지는
반폐쇄의 연안수역(Pritchard, 1967)이며, 다양한 물질이 집적되고 유출되는 육상과 해
양간의 여과장치로서의 역할을 수행한다(Schubel, 1984). 일반적으로 하구역의 수질 특
성은 조석, 담수 유입의 정도 및 바람 등에 의해 연속적으로 변하는 수괴가 완전히 혼
합되지 않는 이질적인 상태를 유지함으로서 염분, 수온 및 용존산소 등과 같은 환경요
인의 시・공간적인 변화가 심하게 나타난다(Kikuchi, 1981). 따라서, 하구역 생물상은 인
접한 해양 및 담수 생태계에 비하여 생물 다양성이 낮지만, 일단 적응한 특정 생물종의
개체군은 매우 잘 발달하여 단위면적당 생물량이 높은 특징이 있다(Remane and
Schlieper, 1958; McLusky, 1981). 한편, 해양생태계를 구성하는 중요한 축의 하나인 저
서동물 군집은 복잡한 시‧ 공간적 패턴을 보이며, 생물학적 다양성 측면에서 매우 높은
가치를 가지고 있다(van der Maarel, 1990). 저서동물은 비교적 이동성이 적다는 이유
로 인하여 플랑크톤이나 유영성 어류의 군집과 비교하여 해역의 환경을 모니터링하는
데 유용하게 이용되어 왔다. 특히, 저서환경의 변화에 매우 민감한 특징을 가지고 있어,
이들 군집의 다양도와 개체수는 생태계의 건강도를 측정하는 요소로 알려져 있다
(Pearson and Rosenburg, 1978; Thouzeau et al., 1991). 국내외의 연구를 통해서 하구
역 환경에 서식하는 저서동물 군집은 염분도(Bae and Yoon, 1989b; Platell and Potter,
1996), 오염부하량이 높은 담수의 유입과 부영양화(Probert, 1984), 퇴적물의 조성
(Gambi and Giangrande, 1986), 퇴적율(Schaffiner et al., 1987), 홍수기 담수의 대량유
입에 따른 퇴적상의 변화(Hong and Yoo, 1996) 등과 같은 다양한 요인에 의해 영향을
받고 있음이 제기된 바 있다. 또한, 이 밖에도 저서동물은 생태계의 먹이사슬에서 일차
소비자인 동시에 2차 생산자로서 역할을 수행하며, 저어류나 조류(鳥類)의 먹이원 및
상위단계로의 에너지 전달원으로서 매우 중요한 부분을 차지한다(Cohen et al., 1982).
따라서 본 조사는 상기에서 언급한 바와 같이 생태계에서 중요한 기능적 역할을 수행
하는 저서동물의 생물다양성과 군집구조를 일차적으로 밝히고, 향후 낙동강 하구역 생
태계의 먹이망과 물질순환의 모식도를 구축하는데 있어 기초자료를 제공하는데 목적이
있다.
(1) 재료 및 방법
(가) 연구해역 개황
낙동강은 강원도 삼척시 황지에서 발원하여 전장 525㎞를 유하하면서 23,656㎢에 달
하는 집수역의 물을 모아 최종적으로 남해로 유입되는 남한 제2의 강이다. 또한, 지형

- 104 -
학적 특성상 조석과 육지로부터 유입되는 담수로 인하여 염분농도의 변화가 비교적 심
한 독특한 환경을 이루고 있다(산업기지개발공사, 1981). 한편, 서・남해안으로 열려있는
강과 비교해 상대적으로 넓은 기수역을 갖는 낙동강 하구역에는 삼각주, 모래등(진우
도, 대마등, 장자도, 새등, 맹금머리등, 백합등, 도요등) 및 갯벌에 다양한 기질이 존재하
며, 저서동물의 밀도가 매우 높은 것으로 알려져 있다(김 등, 1982; Bae and Yoon,
1988; Jang and Kim, 1992). 또한, 저서동물은 계절에 따라 낙동강을 찾는 조류(鳥類)의
중요한 먹이원으로 이용되고 있다(원, 1985). 이러한 이유로 하구역 일원은 1966년 7월
에 “낙동강 하류 철새도래지”-천연기념물 제 179호로 지정되었고(문화공보부, 1971), 이
후 시간의 경과에 따라 자연환경보전지역(1987년, 국토이용관리법), 자연생태계보전지역
(1989년, 자연환경보전법), 습지보호지역(1999년, 습지보전법) 및 부산연안특별관리해역
(1982년, 해양오염방지법)으로 지정, 고시되는 등 환경적으로 매우 중요한 생태계로 고
려되고 있다. 한편, 주민의 삶의 질 향상과 산업발달의 요구에 따른 용수공급, 교통개
선, 매립을 통한 국토확장을 목적으로 하구언이 1983년 9월에 착공되어 1987년 11월에
완공되었다(부산지방환경청, 1992). 그러나, 하구언 완공이후 해수와 담수의 혼합이 매
우 제한적으로 이루어지며, 일부 호수화된 하류지역은 지속적인 물의 정체와 상・중류부
터 유입되는 오염원으로 인해 부영양화가 가속화되는 현상을 보이고 있다. 또한, 오염
부하량이 높은 담수가 수문의 개폐작용을 통해 하구역으로 유입되면 인근의 외양까지
영향을 미칠 것으로 추정되어(부산광역시, 2003), 생태계 보전이나 관리 측면에서 매우
중요하게 다루어야 할 생태계이다.
(나) 현장조사
낙동강 하구역에 서식하는 저서동물의 생물다양성과 군집구조를 파악하기 위하여
2004년 7월부터 동년 9월까지 갯벌에 총 7개 정선, 28개 정점과 조하대 36개 정점을 선
정하여 현장조사를 실시하였다(Fig. 2-32).
- 105 -
JW T 1JW T 2
SH T U ST
J JT� JT
B H T
3 5
3 6
1 6
1 5
1 2
1 4
1 8
1 3
1 7
1 9
2 0
2 1
2 4
7
8
69
4
35
1 0
1 1
1
2
2 8
2 9
3 0
3 1
2 7
3 2
3 3
3 4
2 62 5
2 5
2 2
2 3
3 5
3 6
1 6
1 5
1 2
1 4
1 8
1 3
1 7
1 9
2 0
2 1
2 4
7
8
69
4
35
1 0
1 1
1
2
2 8
2 9
3 0
3 1
2 7
3 2
3 3
3 4
2 62 5
2 5
2 2
2 3
Figure. 2-32. Map of Nakdong River estuary showing the
macozoobenthos sampling locations of this study. (Index: 1;
Jinwoo Is., 2; Shinja Is., 3; DaemaDeung, 4; Jangja Is., 5; Nameless
Is., 6; Saja Is., 7; Maengkeummeori, 8; Ulsuk Is.)

- 106 -
저서동물의 채집을 위하여 갯벌에서는 rectangular can corer를 이용하여 8회를, 조하
대는 van Veen grab으로 2회를 실시하였고, 각 정점에서 공히 0.2㎡의 퇴적물을 채취
하였다. 채취된 퇴적물은 현장에서 1㎜ 망목의 체를 이용하여 저서동물을 분리하였고,
10%의 중성 포르말린으로 고정하여 실험실로 운반하였다. 그 후 분류군별로 선별 및
동정을 실시하였고, 개체수를 계수하고 생물량을 측정하였다.
(다) 환경요인 분석
무기 환경요인 중, 퇴적물의 입도 분석과 유기물 함량(ignition loss)분석을 위해 생물
채집과 동시에 퇴적물을 채취하였다. 이중 퇴적물의 조성은 SYMPA TEC HELOS
Particle Size Analysis WINDOX4를 이용하여 측정하였다. 한편 유기물 함량의 측정은
건조된 퇴적물 시료의 일부를 분말화한 후, 0.001 mm 체로 거른 후, 이 중 5 g을 추출
하여 550℃에서 2시간 연소시켰다. 그 후 무게의 손실량을 백분율로 표시하였다(해양수
산부, 1998).
(라) 저서동물 군집구조 분석
저서동물의 군집구조를 파악하기 위하여, 종 다양도(Shannon and Weaver, 1949)와
균등도(Pielou, 1975) 등의 생태학적 제 지수를 분석하였다. 한편, 출현 종과 개체수의
자료를 이용한 집괴분석은 각 정점간의 유사도(similarity)를 파악하기 위하여, Bray
and Curtis(1957)의 similarity index를 이용하였다. 유사도 지수 행렬로부터 각 조사 정
점과 출현 종을 연결하는 방법으로는 group-average를 이용하였다. 군집분석은
PRIMER(Plymouth Routines Multivariate Ecological Research) computer package를
이용하여 수지도(dendrogram)와 다차원배열법(MDS ordination)으로 나타내었다. 우점
종 서열분석을 위해서는 LeBris index(1988)를 이용하였고, 식은 아래와 같다.
D' ij = Fij × Dij × 100 = Pj
[(Pij / Pj) × 100] × [(∑ (Nik / Nk)) / Pj] × 100 k=1
Pij = j번째 군집에서 i종의 출현횟수
Pj = j번째 군집의 전체 정점수
Nik = j군집 군집에서 k번째 정점에 출현하는 i종의 개체수 또는 생물량
Nk = k번째 정점에서 전체 개체수 또는 생물량
(마) 이매패류 생태조사
낙동강 하구 갯벌에 서식하는 패류 중 가장 높은 출현빈도를 나타내어 철새들의 중
요한 먹이로 여겨지는 3개 종 (Laternula marilina, Corbicula japonica, Nuttallia japon
ica)에 대해 매월 6개 정점에 대해서 생물량 변동조사를 실시하였는데 동시에 약 50개

- 107 -
체를 무작위로 채집하여 생태조사를 실시하였다. 채집된 시료는 실험실로 운반하여 각
장, 각고, 각폭 그리고 전중량을 측정하고 조심스럽게 개각하여 육질부를 회수하여 동
결건조 한 후 건중량과 패각중량을 측정하였으며, 육질부는 미세한 분말을 만들어 일정
량의 분말을 550℃에서 2시간 동안 연소 후 중량을 측정하여 회분제외중량을 구하였다.
또한 이들의 계절에 따른 환경변이에 대한 생태생리 (ecophysiology)를 이해하기 위하
여 건조 육질부의 분말 시료를 이용하여 각 생물군의 월별 단백질, 지방, 탄수화물 그
리고 글리코겐 양을 측정하였다. 이때 단백질은 Folin-Ciocalteu를 이용하여 발색하는
Lowry et al. (1951)의 방법으로 bovine serum albumin을 표준용액으로 하여 분석하였
다. 탄수화물을 glucose로 표준용액을 조제하고 Dubois et al. (1956)의 phenol-sulphuri
c acid 방법으로 분석하였다. 지질의 추출은 chloroform과 methanol 혼합액으로 추출하
고 (Bligh and Dyer, 1959), 지질함량은 Marsh and Weinstein (1966)의 방법으로 측정
하였다. 또한 이들의 먹이원의 계절변동을 분석하기 위하여 육질부 분말시료를 이용한
안정동위원소 분석을 시도하였다.
(2) 연구결과
(가) 환경요인
① 퇴적물 조성
갯벌의 총 22개의 정점을 대상으로 퇴적물의 조성(모래 및 펄함량)을 파악한 결과,
모래함량은 91%~13.38%의 범위에 평균 70.97%를 차지하여 대부분의 정점에서 높았
다. 그러나, 진우도(정선 2의 정점 2)와 백합등(정점 4, 5, 6)의 일부 정점과 장자도의
정점 2에서는 펄함량이 59.8%~86.62% 범위로 높은 특징이 있었다(Fig. 2-33). 각 정선
별로 보면, 진우도 정선(JWT1, 2)은 모래함량이 평균 73.05%를 차지하였으나, 이 가
운데 진우도 정선 2의 정점 2는 상대적으로 펄함량이 67.85%로 높은 특징이 있었다.
무명도와 장자도 사이의 정선(NJT), 장자도 정선(JJT)과 신호도 정선(SHT)도 모래함
량이 각각 평균 88.75%, 67.67%와 84.43%로 높았다. 반면, 백합등 정선(BHT)은 다른
정선과 비교하여 모래와 펄함량이 각각 53.58%와 46.42%로 비슷하였다.
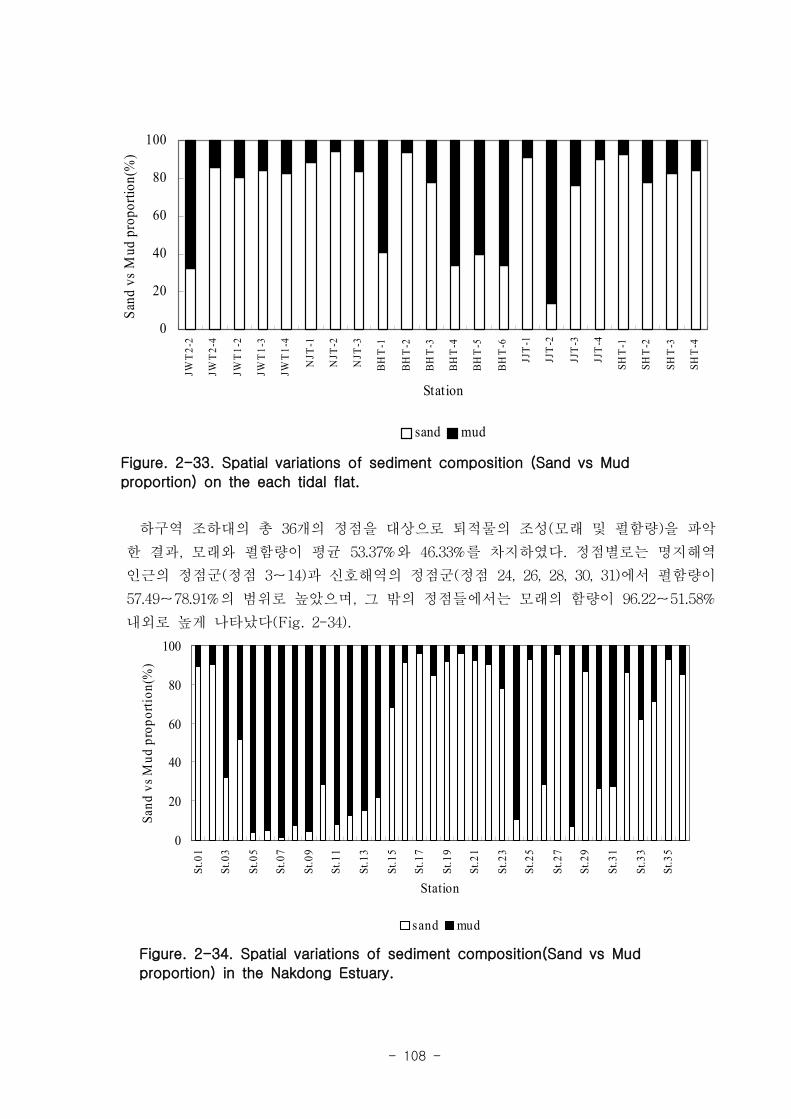
- 108 -
0
20
40
60
80
100
JWT2-2
JWT2-4
JWT1-2
JWT1-3
JWT1-4
NJT
-1
NJT
-2
NJT
-3
BHT-1
BHT-2
BHT-3
BHT-4
BHT-5
BHT-6
JJT-1
JJT-2
JJT-3
JJT-4
SHT-1
SHT-2
SHT-3
SHT-4
Station
San
d vs M
ud proportion(%
)
sand mud
Figure. 2-33. Spatial variations of sediment composition (Sand vs Mud
proportion) on the each tidal flat.
하구역 조하대의 총 36개의 정점을 대상으로 퇴적물의 조성(모래 및 펄함량)을 파악
한 결과, 모래와 펄함량이 평균 53.37%와 46.33%를 차지하였다. 정점별로는 명지해역
인근의 정점군(정점 3~14)과 신호해역의 정점군(정점 24, 26, 28, 30, 31)에서 펄함량이
57.49~78.91%의 범위로 높았으며, 그 밖의 정점들에서는 모래의 함량이 96.22~51.58%
내외로 높게 나타났다(Fig. 2-34).
0
20
40
60
80
100
St.01
St.03
St.05
St.07
St.09
St.11
St.13
St.15
St.17
St.19
St.21
St.23
St.25
St.27
St.29
St.31
St.33
St.35
Station
Sand vs M
ud proportion(%
)
s and mud
Figure. 2-34. Spatial variations of sediment composition(Sand vs Mud
proportion) in the Nakdong Estuary.
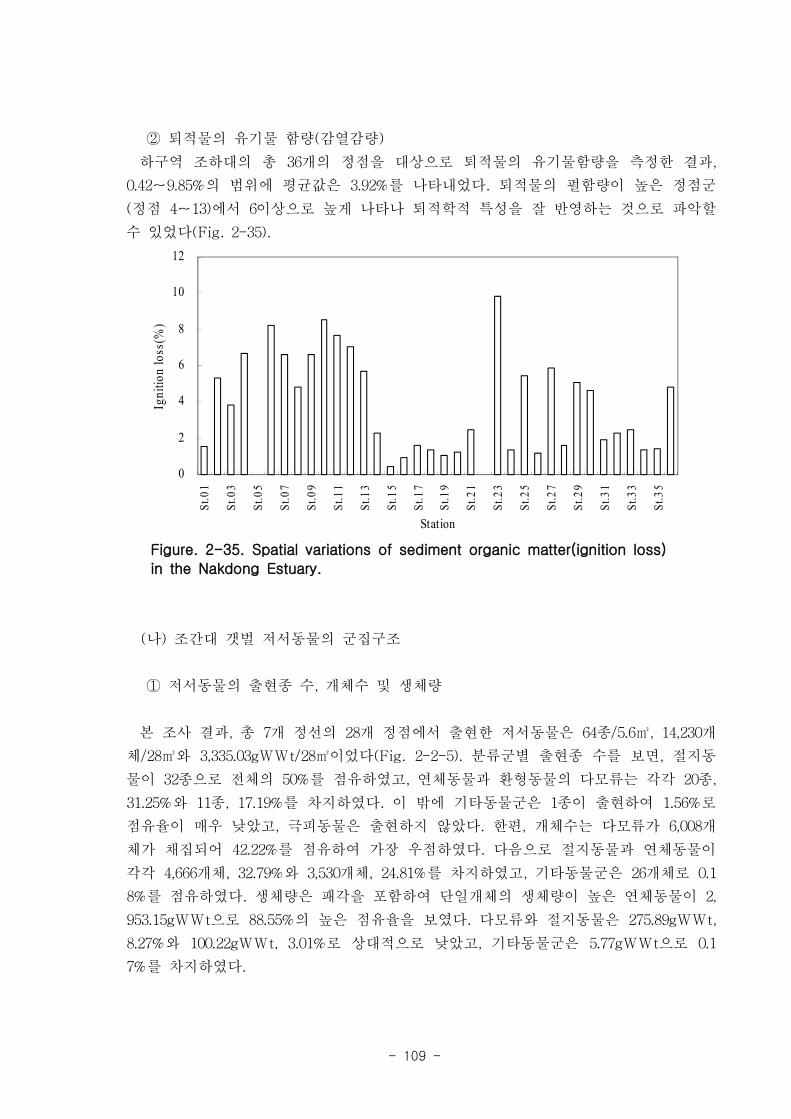
- 109 -
② 퇴적물의 유기물 함량(감열감량)
하구역 조하대의 총 36개의 정점을 대상으로 퇴적물의 유기물함량을 측정한 결과,
0.42~9.85%의 범위에 평균값은 3.92%를 나타내었다. 퇴적물의 펄함량이 높은 정점군
(정점 4~13)에서 6이상으로 높게 나타나 퇴적학적 특성을 잘 반영하는 것으로 파악할
수 있었다(Fig. 2-35).
0
2
4
6
8
10
12
St.01
St.03
St.05
St.07
St.09
St.11
St.13
St.15
St.17
St.19
St.21
St.23
St.25
St.27
St.29
St.31
St.33
St.35
Station
Ignition loss(%
)
Figure. 2-35. Spatial variations of sediment organic matter(ignition loss)
in the Nakdong Estuary.
(나) 조간대 갯벌 저서동물의 군집구조
① 저서동물의 출현종 수, 개체수 및 생체량
본 조사 결과, 총 7개 정선의 28개 정점에서 출현한 저서동물은 64종/5.6㎡, 14,230개
체/28㎡와 3,335.03gWWt/28㎡이었다(Fig. 2-2-5). 분류군별 출현종 수를 보면, 절지동
물이 32종으로 전체의 50%를 점유하였고, 연체동물과 환형동물의 다모류는 각각 20종,
31.25%와 11종, 17.19%를 차지하였다. 이 밖에 기타동물군은 1종이 출현하여 1.56%로
점유율이 매우 낮았고, 극피동물은 출현하지 않았다. 한편, 개체수는 다모류가 6,008개
체가 채집되어 42.22%를 점유하여 가장 우점하였다. 다음으로 절지동물과 연체동물이
각각 4,666개체, 32.79%와 3,530개체, 24.81%를 차지하였고, 기타동물군은 26개체로 0.1
8%를 점유하였다. 생체량은 패각을 포함하여 단일개체의 생체량이 높은 연체동물이 2,
953.15gWWt으로 88.55%의 높은 점유율을 보였다. 다모류와 절지동물은 275.89gWWt,
8.27%와 100.22gWWt, 3.01%로 상대적으로 낮았고, 기타동물군은 5.77gWWt으로 0.1
7%를 차지하였다.

- 110 -
Species numberPolychaeta
17.19%
Mollusca
31.25%
Others
1.56%
Crustacea
50.00%
Abundance
Polychaeta
42.22%
Mollusca
24.81%
Crustacea
32.79%
Others
0.18%
BiomassPolychaeta
8.27%
Mollusca
88.55%
Crustacea
3.01%
Others
0.17%
Figure. 2-36. Total number of species, abundance and biomass on
the tidal flat, Nakdong Estuary.
㉮ 정선별 저서동물의 출현종 수, 개체수 및 생체량
㉠ 진우도 정선(JWT1, 2)
총 2개의 정선을 선정하였고, 정선 1과 2를 대상으로 조위에 따라 각각 3개와 4개 정
점을 선정하여 현장조사를 실시하였다. 정선 1은 총 14종/0.6㎡, 388개체/3㎡, 10.378gW
Wt/3㎡가 채집되었다. 분류군별 출현종 수를 보면, 절지동물과 환형동물만이 채집되어
각각 8종, 57.14%와 6종, 42.86%를 차지하였다. 개체수와 생체량은 다모류가 244개체(6
2.89%)와 10.22gWWt(98.48%)으로 거의 대부분을 차지하였고, 절지동물은 144개체(37.1
1%)와 1.52gWWt(1.52%)을 나타내었다. 정점에 따라서는 갯벌의 중・상부에 해당하는
정점 2에서 출현종 수, 개체수 및 생체량이 상대적으로 높았다. 일반적으로 조위에 따
른 노출의 정도와 퇴적상에 의해 저서동물 밀도에 차이가 발생하는데, 본 조사지역은
갯벌의 하부로 갈수록 생물량이 감소하는 상반되는 결과를 보이는 점이 특징이었다. 그
러나, 이것은 현장조사 시에 갯벌의 하부지역을 대표하는 생물채집을 하지 못했다는 점
에서 원인이 있다고 추정된다. 한편, 정선 2는 총 36종/0.8㎡, 1,862개체/4㎡, 708.18gW
Wt/4㎡가 채집되어 정선 1과 비교하여 생물량이 상대적으로 높았다. 분류군별 출현종
수를 보면, 절지동물과 연체동물이 각각 19종, 52.78%와 12종, 33.33%를 차지하여 전체
의 88%를 점유하였다. 다음으로 다모류와 기타동물군은 4종과 1종이 출현하여 점유율
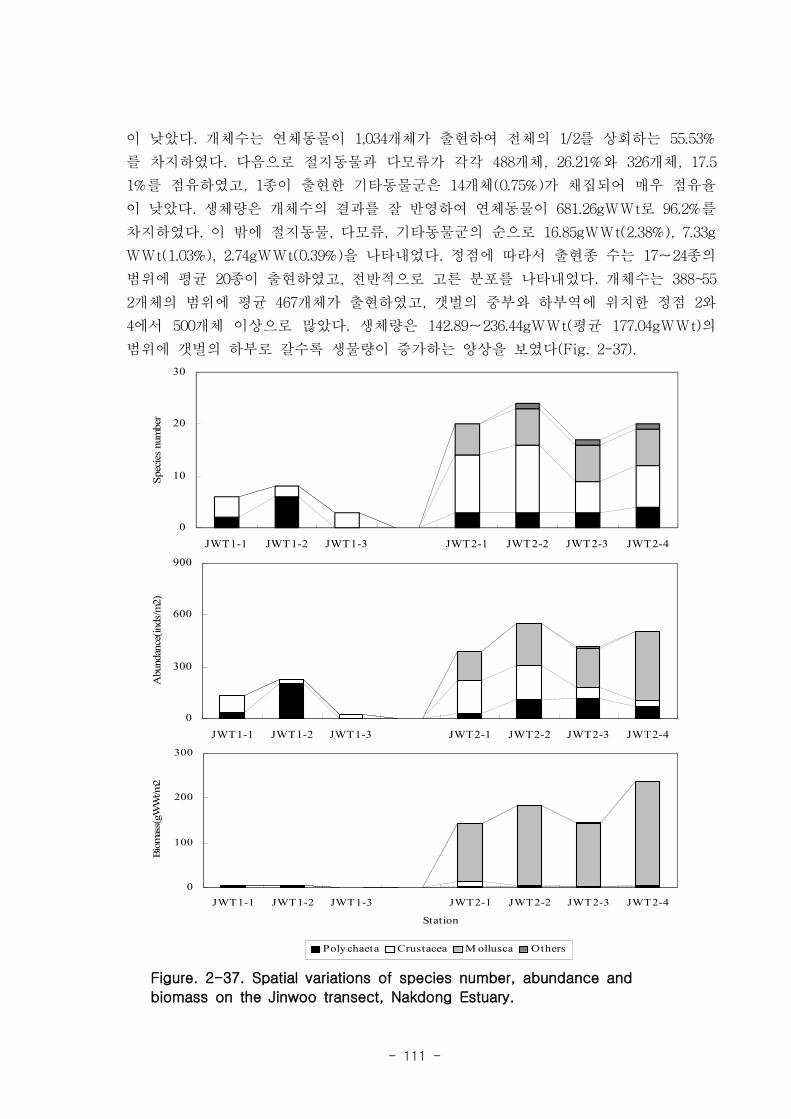
- 111 -
이 낮았다. 개체수는 연체동물이 1,034개체가 출현하여 전체의 1/2를 상회하는 55.53%
를 차지하였다. 다음으로 절지동물과 다모류가 각각 488개체, 26.21%와 326개체, 17.5
1%를 점유하였고, 1종이 출현한 기타동물군은 14개체(0.75%)가 채집되어 매우 점유율
이 낮았다. 생체량은 개체수의 결과를 잘 반영하여 연체동물이 681.26gWWt로 96.2%를
차지하였다. 이 밖에 절지동물, 다모류, 기타동물군의 순으로 16.85gWWt(2.38%), 7.33g
WWt(1.03%), 2.74gWWt(0.39%)을 나타내었다. 정점에 따라서 출현종 수는 17~24종의
범위에 평균 20종이 출현하였고, 전반적으로 고른 분포를 나타내었다. 개체수는 388~55
2개체의 범위에 평균 467개체가 출현하였고, 갯벌의 중부와 하부역에 위치한 정점 2와
4에서 500개체 이상으로 많았다. 생체량은 142.89~236.44gWWt(평균 177.04gWWt)의
범위에 갯벌의 하부로 갈수록 생물량이 증가하는 양상을 보였다(Fig. 2-37).
0
10
20
30
JWT1-1 JWT1-2 JWT1-3 JWT2-1 JWT2-2 JWT2-3 JWT2-4
Species number
0
300
600
900
JWT1-1 JWT1-2 JWT1-3 JWT2-1 JWT2-2 JWT2-3 JWT2-4
Abundance(inds/m2)
0
100
200
300
JWT1-1 JWT1-2 JWT1-3 JWT2-1 JWT2-2 JWT2-3 JWT2-4
Station
Biomass(gWWt/m2)
Polychaeta Crustacea M ollusca Others
Figure. 2-37. Spatial variations of species number, abundance and
biomass on the Jinwoo transect, Nakdong Estuary.

- 112 -
㉡ 신호도 정선(SHT)
총 1개의 정선에서 4개의 정점을 선정하였다. 총 36종/0.8㎡, 1,586개체/4㎡, 380.73gW
Wt/4㎡가 채집되었다. 분류군별 출현종 수를 보면, 절지동물과 연체동물이 각각 15종,
41.67%와 11종, 30.56%를 차지하여 가장 우점하는 생물군이었다. 다음으로 다모류가 9
종으로 25%를 점유하였고, 기타동물군은 1종, 2.78%로 낮았다. 개체수는 다모류가 860
개체가 출현하여 전체의 54.22%를 점유하였다. 출현종이 상대적으로 다양하였던 연체
동물과 절지동물이 각각 436개체, 27.49%와 282개체, 17.78%를 차지하였다. 반면 기타
동물군은 8개체(0.5%)로 밀도가 매우 적었다. 생체량은 연체동물이 215.43gWWt으로 5
6.58%를 점유하였고, 다모류는 144.62gWWt, 37.98%를 차지하였다. 이 밖에 절지동물과
기타동물군은 각각 19.53gWWt, 5.13%와 1.15gWWt, 0.3%로 상대적으로 낮았다. 정점
에 따라서 출현종 수는 11~27종의 범위에 평균 18종이 출현하였고, 갯벌의 상부인 정점
1에서 가장 많았고, 하부의 정점 4에서 가장 적었다. 특히, 현장조사 시의 관찰에 의하
면, 정점 1은 갈파래류(Enteromorpha sp.)의 일종이 mat를 형성하는 지역으로 퇴적물
내 뿐만 아니라 갈파래의 사이 또는 표면에 다양한 생물이 부착하여 서식하는 것을 관
찰할 수 있었고, 이러한 원인으로 생물다양성이 높을 수 있음을 추정할 수 있었다. 개
체수는 318~528개체의 범위에 평균 397개체가 출현하였고, 출현종의 경향과 동일하게
갯벌의 상부에서 밀도가 가장 높았다. 반면, 생체량은 85.62~98.53gWWt(평균 95.18gW
Wt)의 범위에 전반적으로 고른 분포를 나타내었다(Fig. 2-38).
0
10
20
30
SHT-1 SHT-2 SHT-3 SHT-4
Species number
0
200
400
600
800
1000
SHT-1 SHT-2 SHT-3 SHT-4
Abundance(inds/m2)
0
20
40
60
80
100
SHT-1 SHT-2 SHT-3 SHT-4
Station
Biomass(gWWt/m2)
Polychaeta Crustacea M ollusca Others
Figure. 2-38. Spatial variations of species number, abundance and
biomass on the Shinho transect, Nakdong Estuary.
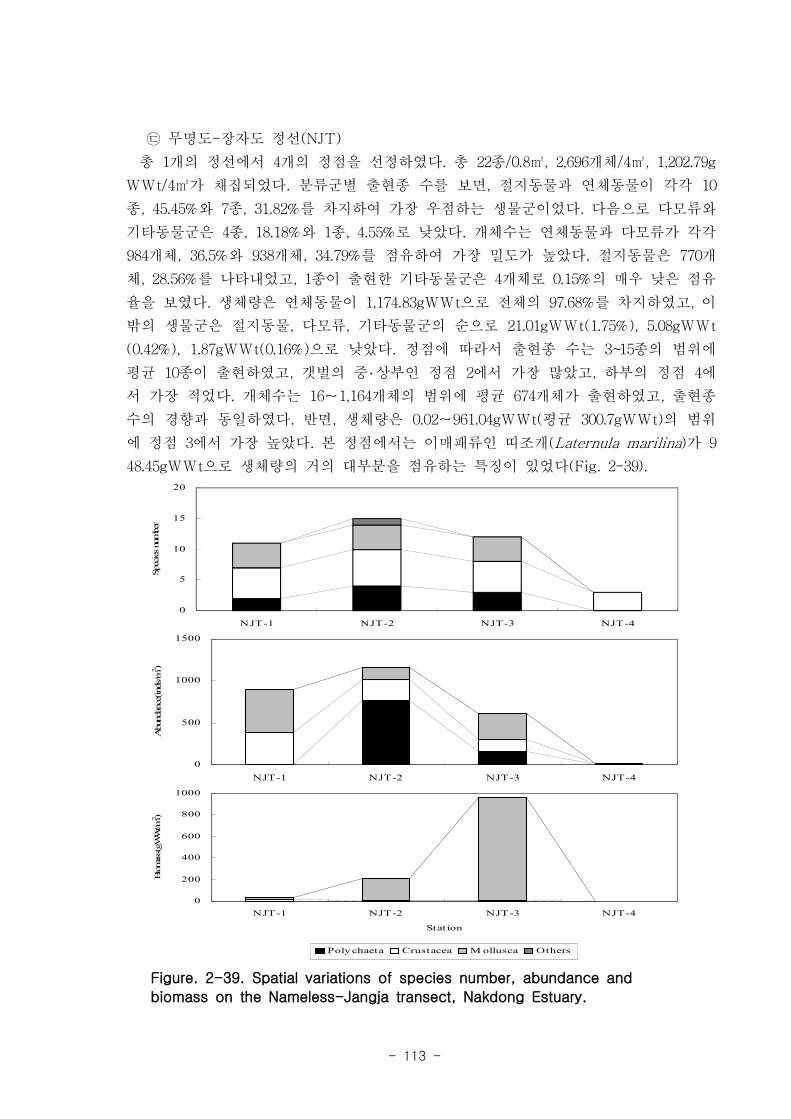
- 113 -
㉢ 무명도-장자도 정선(NJT)
총 1개의 정선에서 4개의 정점을 선정하였다. 총 22종/0.8㎡, 2,696개체/4㎡, 1,202.79g
WWt/4㎡가 채집되었다. 분류군별 출현종 수를 보면, 절지동물과 연체동물이 각각 10
종, 45.45%와 7종, 31.82%를 차지하여 가장 우점하는 생물군이었다. 다음으로 다모류와
기타동물군은 4종, 18.18%와 1종, 4.55%로 낮았다. 개체수는 연체동물과 다모류가 각각
984개체, 36.5%와 938개체, 34.79%를 점유하여 가장 밀도가 높았다. 절지동물은 770개
체, 28.56%를 나타내었고, 1종이 출현한 기타동물군은 4개체로 0.15%의 매우 낮은 점유
율을 보였다. 생체량은 연체동물이 1,174.83gWWt으로 전체의 97.68%를 차지하였고, 이
밖의 생물군은 절지동물, 다모류, 기타동물군의 순으로 21.01gWWt(1.75%), 5.08gWWt
(0.42%), 1.87gWWt(0.16%)으로 낮았다. 정점에 따라서 출현종 수는 3~15종의 범위에
평균 10종이 출현하였고, 갯벌의 중・상부인 정점 2에서 가장 많았고, 하부의 정점 4에
서 가장 적었다. 개체수는 16~1,164개체의 범위에 평균 674개체가 출현하였고, 출현종
수의 경향과 동일하였다. 반면, 생체량은 0.02~961.04gWWt(평균 300.7gWWt)의 범위
에 정점 3에서 가장 높았다. 본 정점에서는 이매패류인 띠조개(Laternula marilina)가 9
48.45gWWt으로 생체량의 거의 대부분을 점유하는 특징이 있었다(Fig. 2-39).
0
5
10
15
20
NJT-1 NJT-2 NJT-3 NJT-4
Species number
0
500
1000
1500
NJT-1 NJT-2 NJT-3 NJT-4
Abundance(inds/m2)
0
200
400
600
800
1000
NJT-1 NJT-2 NJT-3 NJT-4
Station
Biomass(gWWt/m2)
Polychaeta Crustacea M ollusca Others
Figure. 2-39. Spatial variations of species number, abundance and
biomass on the Nameless-Jangja transect, Nakdong Estuary.
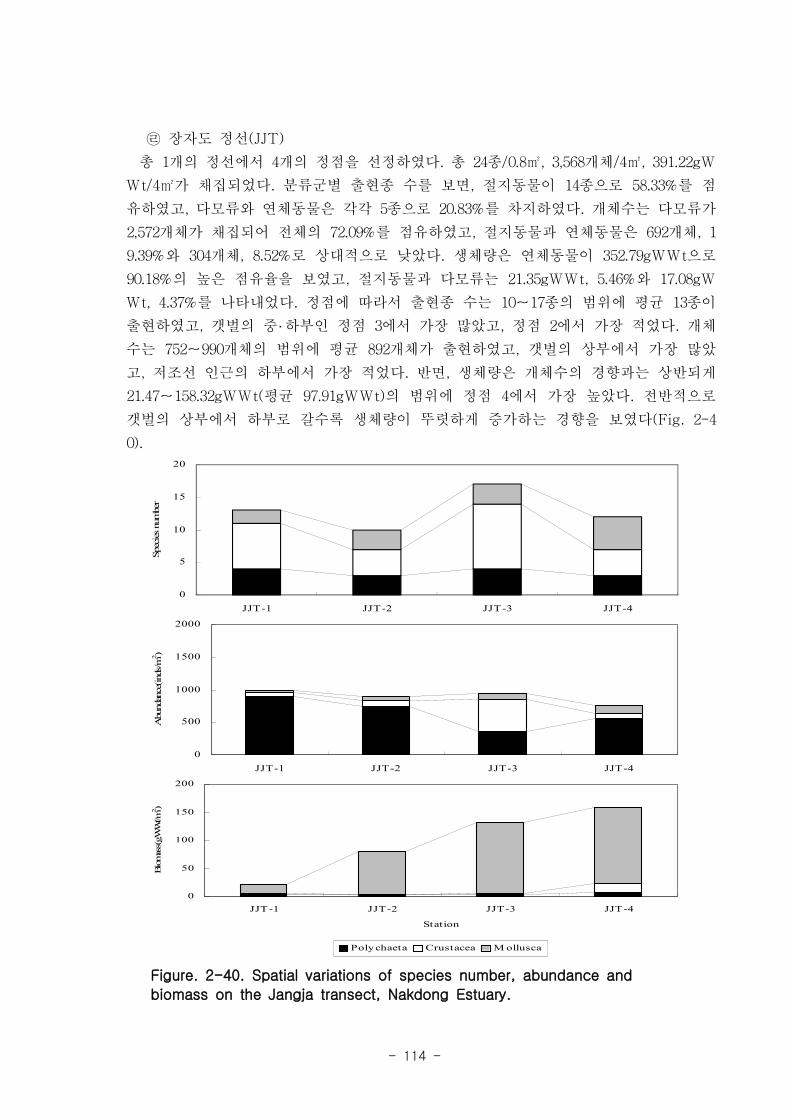
- 114 -
㉣ 장자도 정선(JJT)
총 1개의 정선에서 4개의 정점을 선정하였다. 총 24종/0.8㎡, 3,568개체/4㎡, 391.22gW
Wt/4㎡가 채집되었다. 분류군별 출현종 수를 보면, 절지동물이 14종으로 58.33%를 점
유하였고, 다모류와 연체동물은 각각 5종으로 20.83%를 차지하였다. 개체수는 다모류가
2,572개체가 채집되어 전체의 72.09%를 점유하였고, 절지동물과 연체동물은 692개체, 1
9.39%와 304개체, 8.52%로 상대적으로 낮았다. 생체량은 연체동물이 352.79gWWt으로
90.18%의 높은 점유율을 보였고, 절지동물과 다모류는 21.35gWWt, 5.46%와 17.08gW
Wt, 4.37%를 나타내었다. 정점에 따라서 출현종 수는 10~17종의 범위에 평균 13종이
출현하였고, 갯벌의 중・하부인 정점 3에서 가장 많았고, 정점 2에서 가장 적었다. 개체
수는 752~990개체의 범위에 평균 892개체가 출현하였고, 갯벌의 상부에서 가장 많았
고, 저조선 인근의 하부에서 가장 적었다. 반면, 생체량은 개체수의 경향과는 상반되게
21.47~158.32gWWt(평균 97.91gWWt)의 범위에 정점 4에서 가장 높았다. 전반적으로
갯벌의 상부에서 하부로 갈수록 생체량이 뚜렷하게 증가하는 경향을 보였다(Fig. 2-4
0).
0
5
10
15
20
JJT-1 JJT-2 JJT-3 JJT-4
Species number
0
500
1000
1500
2000
JJT-1 JJT-2 JJT-3 JJT-4
Abundance(inds/m2)
0
50
100
150
200
JJT-1 JJT-2 JJT-3 JJT-4
Station
Biomass(gWWt/m2)
Polychaeta Crustacea M ollusca
Figure. 2-40. Spatial variations of species number, abundance and
biomass on the Jangja transect, Nakdong Estuary.
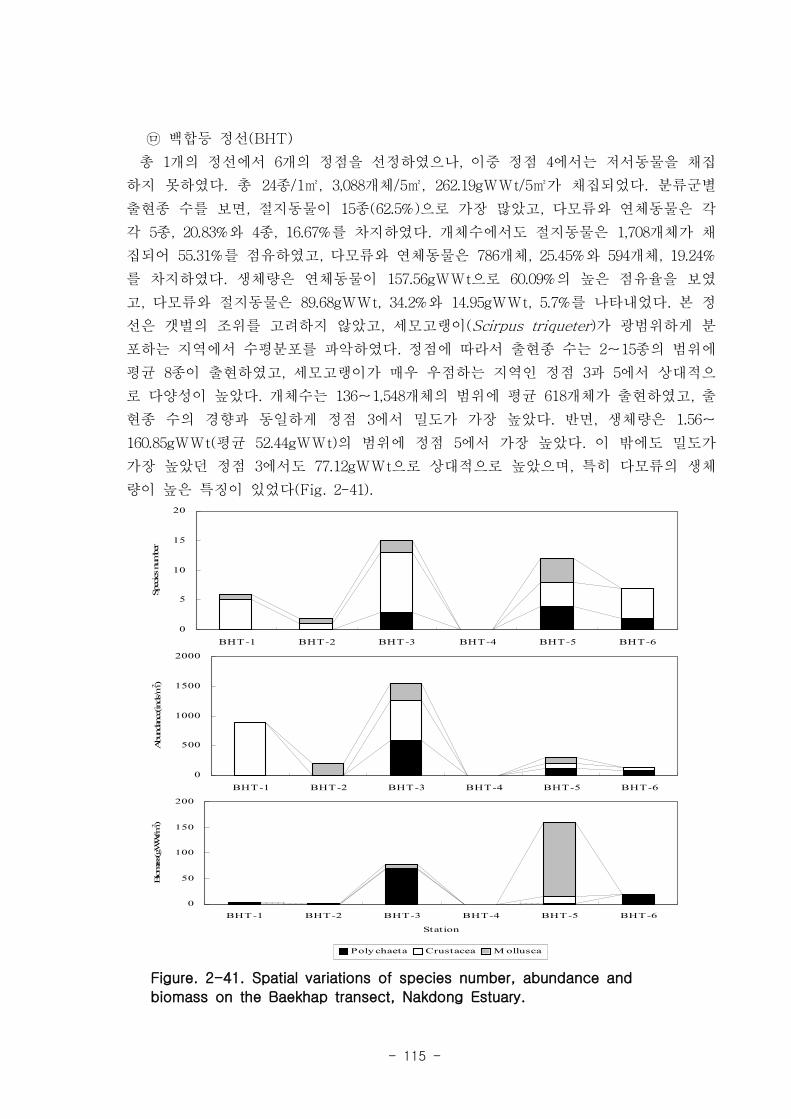
- 115 -
㉤ 백합등 정선(BHT)
총 1개의 정선에서 6개의 정점을 선정하였으나, 이중 정점 4에서는 저서동물을 채집
하지 못하였다. 총 24종/1㎡, 3,088개체/5㎡, 262.19gWWt/5㎡가 채집되었다. 분류군별
출현종 수를 보면, 절지동물이 15종(62.5%)으로 가장 많았고, 다모류와 연체동물은 각
각 5종, 20.83%와 4종, 16.67%를 차지하였다. 개체수에서도 절지동물은 1,708개체가 채
집되어 55.31%를 점유하였고, 다모류와 연체동물은 786개체, 25.45%와 594개체, 19.24%
를 차지하였다. 생체량은 연체동물이 157.56gWWt으로 60.09%의 높은 점유율을 보였
고, 다모류와 절지동물은 89.68gWWt, 34.2%와 14.95gWWt, 5.7%를 나타내었다. 본 정
선은 갯벌의 조위를 고려하지 않았고, 세모고랭이(Scirpus triqueter)가 광범위하게 분
포하는 지역에서 수평분포를 파악하였다. 정점에 따라서 출현종 수는 2~15종의 범위에
평균 8종이 출현하였고, 세모고랭이가 매우 우점하는 지역인 정점 3과 5에서 상대적으
로 다양성이 높았다. 개체수는 136~1,548개체의 범위에 평균 618개체가 출현하였고, 출
현종 수의 경향과 동일하게 정점 3에서 밀도가 가장 높았다. 반면, 생체량은 1.56~
160.85gWWt(평균 52.44gWWt)의 범위에 정점 5에서 가장 높았다. 이 밖에도 밀도가
가장 높았던 정점 3에서도 77.12gWWt으로 상대적으로 높았으며, 특히 다모류의 생체
량이 높은 특징이 있었다(Fig. 2-41).
0
5
10
15
20
BHT-1 BHT-2 BHT-3 BHT-4 BHT-5 BHT-6
Species number
0
500
1000
1500
2000
BHT-1 BHT-2 BHT-3 BHT-4 BHT-5 BHT-6
Abundance(inds/m2)
0
50
100
150
200
BHT-1 BHT-2 BHT-3 BHT-4 BHT-5 BHT-6
Station
Biomass(gWWt/m2)
Polychaeta Crustacea M ollusca
Figure. 2-41. Spatial variations of species number, abundance and
biomass on the Baekhap transect, Nakdong Estuary.
- 116 -
㉥ 을숙도 정선(UST)
총 1개의 정선에서 4개의 정점을 선정하였고, 15종/0.8㎡, 1,042개체/4㎡, 379.54gWWt
/5㎡가 채집되었다. 분류군별 출현종 수를 보면, 다모류, 절지동물과 연체동물이 공통적
으로 5종씩(33.33%) 출현하였다. 개체수는 절지동물이 582개체로 55.85%를 점유하였고,
다모류와 연체동물은 각각 282개체, 27.06%와 178개체, 17.08%를 차지하였다. 생체량은
연체동물이 371.28gWWt으로 97.82%의 높은 점유율을 보였고, 절지동물과 다모류는 6.
38gWWt, 1.68%와 1.88gWWt, 0.5%로 낮았다. 정점에 따라서 출현종 수는 4~10종의
범위에 평균 7종이 출현하였고, 정점 2에서 많았고, 정점 1에서 낮았다. 개체수는 160~5
18개체의 범위에 평균 261개체가 출현하였고, 정점 3에서 밀도가 가장 높았고, 정점 1
과 4에서 적었다. 반면, 생체량은 39.15~140.87gWWt(평균 94.88gWWt)의 범위에 정점
2에서 가장 높았고, 이 밖에도 정점 4에서 120.09gWWt으로 생체량이 높게 나타났다(Fi
g. 2-42).
0
5
10
15
20
UST-1 UST-2 UST-3 UST-4
Species number
0
200
400
600
UST-1 UST-2 UST-3 UST-4
Abundance(inds/m2)
0
50
100
150
200
UST-1 UST-2 UST-3 UST-4
Station
Biomass(gWWt/m2)
Polychaeta Crustacea M ollusca
Figure. 2-42. Spatial variations of species number, abundance and
biomass on the Ulsuk transect, Nakdong Estuary.
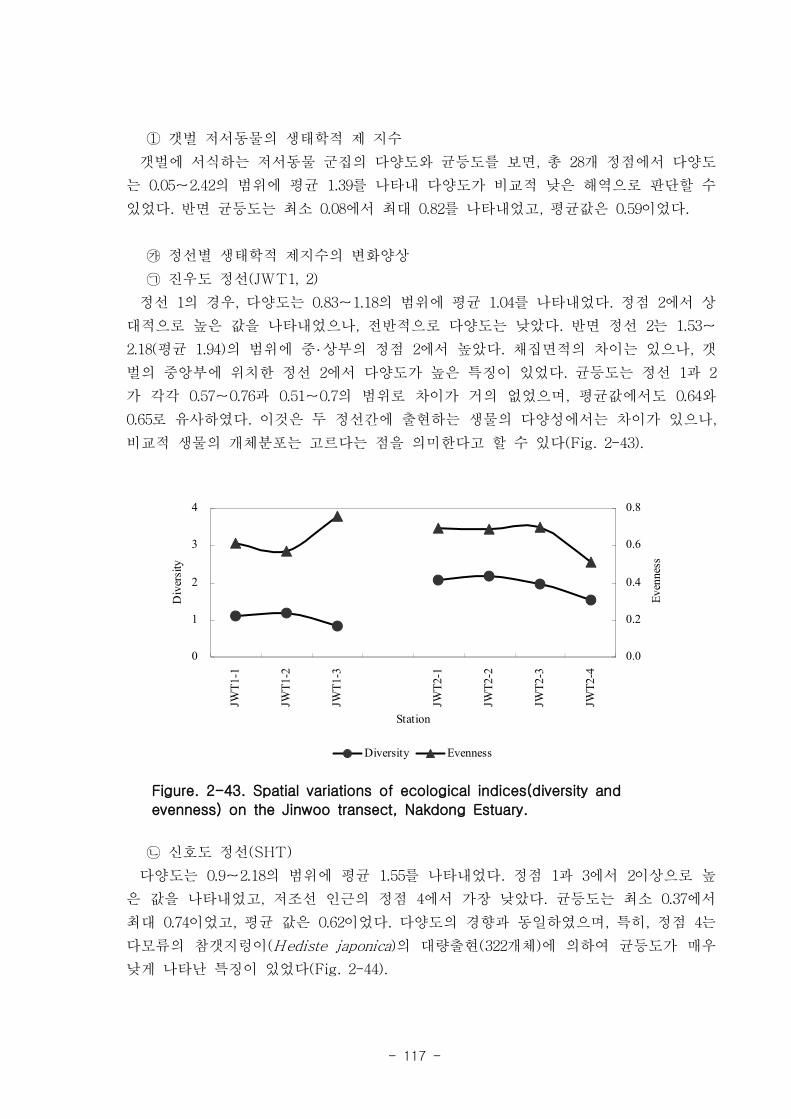
- 117 -
① 갯벌 저서동물의 생태학적 제 지수
갯벌에 서식하는 저서동물 군집의 다양도와 균등도를 보면, 총 28개 정점에서 다양도
는 0.05~2.42의 범위에 평균 1.39를 나타내 다양도가 비교적 낮은 해역으로 판단할 수
있었다. 반면 균등도는 최소 0.08에서 최대 0.82를 나타내었고, 평균값은 0.59이었다.
㉮ 정선별 생태학적 제지수의 변화양상
㉠ 진우도 정선(JWT1, 2)
정선 1의 경우, 다양도는 0.83~1.18의 범위에 평균 1.04를 나타내었다. 정점 2에서 상
대적으로 높은 값을 나타내었으나, 전반적으로 다양도는 낮았다. 반면 정선 2는 1.53~
2.18(평균 1.94)의 범위에 중・상부의 정점 2에서 높았다. 채집면적의 차이는 있으나, 갯
벌의 중앙부에 위치한 정선 2에서 다양도가 높은 특징이 있었다. 균등도는 정선 1과 2
가 각각 0.57~0.76과 0.51~0.7의 범위로 차이가 거의 없었으며, 평균값에서도 0.64와
0.65로 유사하였다. 이것은 두 정선간에 출현하는 생물의 다양성에서는 차이가 있으나,
비교적 생물의 개체분포는 고르다는 점을 의미한다고 할 수 있다(Fig. 2-43).
0
1
2
3
4
JWT1-1
JWT1-2
JWT1-3
JWT2-1
JWT2-2
JWT2-3
JWT2-4
Station
Diversity
0.0
0.2
0.4
0.6
0.8
Evenness
Diversity Evenness
Figure. 2-43. Spatial variations of ecological indices(diversity and
evenness) on the Jinwoo transect, Nakdong Estuary.
㉡ 신호도 정선(SHT)
다양도는 0.9~2.18의 범위에 평균 1.55를 나타내었다. 정점 1과 3에서 2이상으로 높
은 값을 나타내었고, 저조선 인근의 정점 4에서 가장 낮았다. 균등도는 최소 0.37에서
최대 0.74이었고, 평균 값은 0.62이었다. 다양도의 경향과 동일하였으며, 특히, 정점 4는
다모류의 참갯지렁이(Hediste japonica)의 대량출현(322개체)에 의하여 균등도가 매우
낮게 나타난 특징이 있었다(Fig. 2-44).
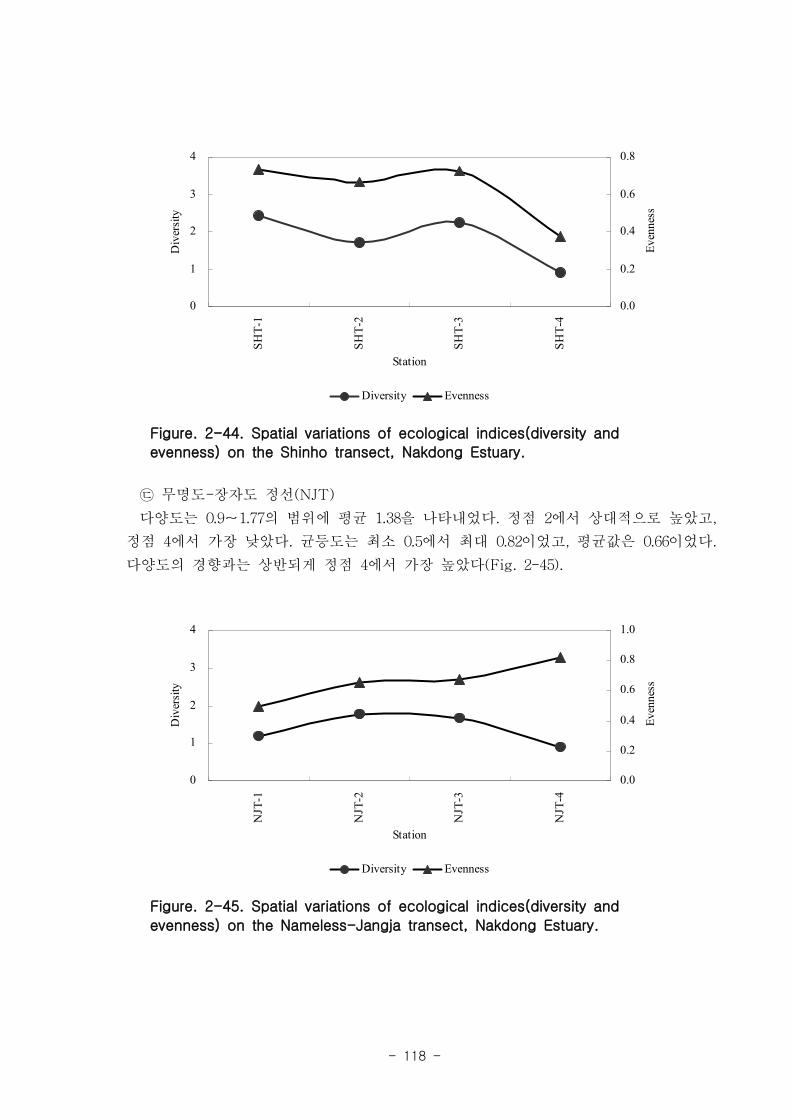
- 118 -
0
1
2
3
4
SHT-1
SHT-2
SHT-3
SHT-4
Station
Diversity
0.0
0.2
0.4
0.6
0.8
Evenness
Diversity Evenness
Figure. 2-44. Spatial variations of ecological indices(diversity and
evenness) on the Shinho transect, Nakdong Estuary.
㉢ 무명도-장자도 정선(NJT)
다양도는 0.9~1.77의 범위에 평균 1.38을 나타내었다. 정점 2에서 상대적으로 높았고,
정점 4에서 가장 낮았다. 균등도는 최소 0.5에서 최대 0.82이었고, 평균값은 0.66이었다.
다양도의 경향과는 상반되게 정점 4에서 가장 높았다(Fig. 2-45).
0
1
2
3
4
NJT-1
NJT-2
NJT-3
NJT-4
Station
Diversity
0.0
0.2
0.4
0.6
0.8
1.0
Evenness
Diversity Evenness
Figure. 2-45. Spatial variations of ecological indices(diversity and
evenness) on the Nameless-Jangja transect, Nakdong Estuary.
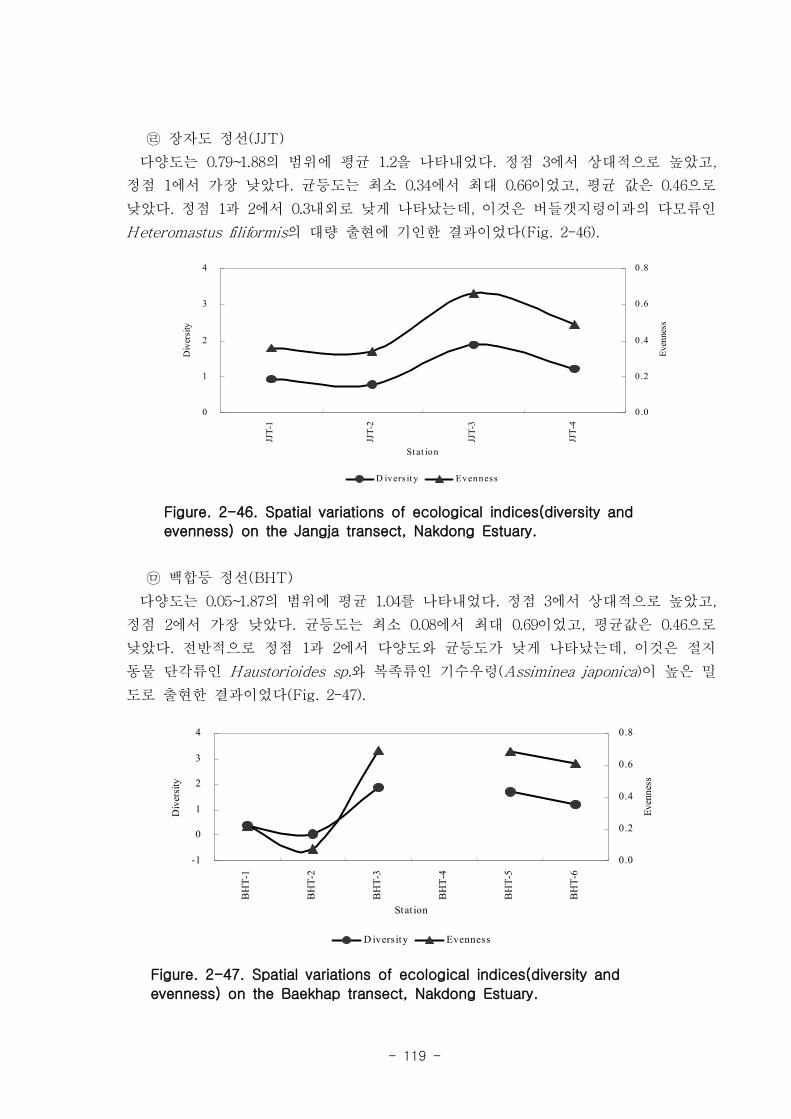
- 119 -
㉣ 장자도 정선(JJT)
다양도는 0.79~1.88의 범위에 평균 1.2을 나타내었다. 정점 3에서 상대적으로 높았고,
정점 1에서 가장 낮았다. 균등도는 최소 0.34에서 최대 0.66이었고, 평균 값은 0.46으로
낮았다. 정점 1과 2에서 0.3내외로 낮게 나타났는데, 이것은 버들갯지렁이과의 다모류인
Heteromastus filiformis의 대량 출현에 기인한 결과이었다(Fig. 2-46).
0
1
2
3
4JJT-1
JJT-2
JJT-3
JJT-4
St at ion
Diversity
0 .0
0 .2
0 .4
0 .6
0 .8
Evenness
D ivers it y Evenness
Figure. 2-46. Spatial variations of ecological indices(diversity and
evenness) on the Jangja transect, Nakdong Estuary.
㉤ 백합등 정선(BHT)
다양도는 0.05~1.87의 범위에 평균 1.04를 나타내었다. 정점 3에서 상대적으로 높았고,
정점 2에서 가장 낮았다. 균등도는 최소 0.08에서 최대 0.69이었고, 평균값은 0.46으로
낮았다. 전반적으로 정점 1과 2에서 다양도와 균등도가 낮게 나타났는데, 이것은 절지
동물 단각류인 Haustorioides sp.와 복족류인 기수우렁(Assiminea japonica)이 높은 밀
도로 출현한 결과이었다(Fig. 2-47).
-1
0
1
2
3
4
BHT-1
BHT-2
BHT-3
BHT-4
BHT-5
BHT-6
Stat ion
Diversity
0.0
0.2
0.4
0.6
0.8
Evenness
D iversity Evenness
Figure. 2-47. Spatial variations of ecological indices(diversity and
evenness) on the Baekhap transect, Nakdong Estuary.
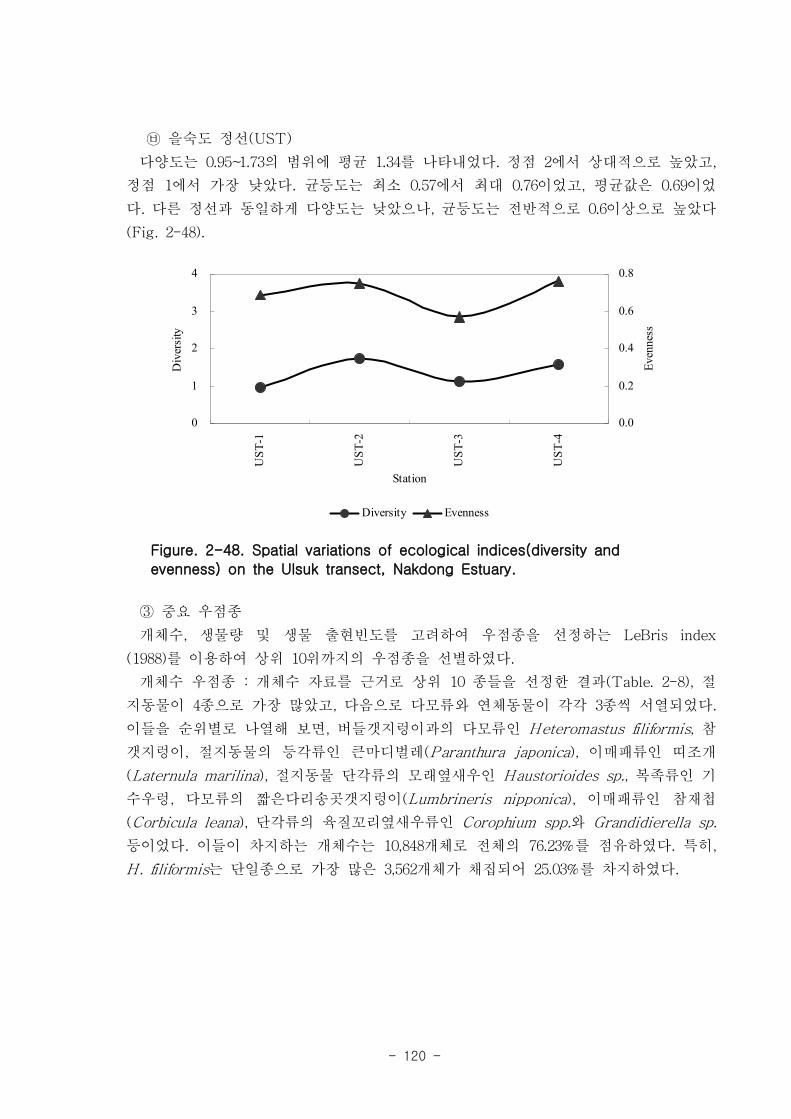
- 120 -
㉥ 을숙도 정선(UST)
다양도는 0.95~1.73의 범위에 평균 1.34를 나타내었다. 정점 2에서 상대적으로 높았고,
정점 1에서 가장 낮았다. 균등도는 최소 0.57에서 최대 0.76이었고, 평균값은 0.69이었
다. 다른 정선과 동일하게 다양도는 낮았으나, 균등도는 전반적으로 0.6이상으로 높았다
(Fig. 2-48).
0
1
2
3
4
UST-1
UST-2
UST-3
UST-4
Station
Diversity
0.0
0.2
0.4
0.6
0.8
Evenness
Diversity Evenness
Figure. 2-48. Spatial variations of ecological indices(diversity and
evenness) on the Ulsuk transect, Nakdong Estuary.
③ 중요 우점종
개체수, 생물량 및 생물 출현빈도를 고려하여 우점종을 선정하는 LeBris index
(1988)를 이용하여 상위 10위까지의 우점종을 선별하였다.
개체수 우점종 : 개체수 자료를 근거로 상위 10 종들을 선정한 결과(Table. 2-8), 절
지동물이 4종으로 가장 많았고, 다음으로 다모류와 연체동물이 각각 3종씩 서열되었다.
이들을 순위별로 나열해 보면, 버들갯지렁이과의 다모류인 Heteromastus filiformis, 참
갯지렁이, 절지동물의 등각류인 큰마디벌레(Paranthura japonica), 이매패류인 띠조개
(Laternula marilina), 절지동물 단각류의 모래옆새우인 Haustorioides sp., 복족류인 기
수우렁, 다모류의 짧은다리송곳갯지렁이(Lumbrineris nipponica), 이매패류인 참재첩
(Corbicula leana), 단각류의 육질꼬리옆새우류인 Corophium spp.와 Grandidierella sp.
등이었다. 이들이 차지하는 개체수는 10,848개체로 전체의 76.23%를 점유하였다. 특히,
H. filiformis는 단일종으로 가장 많은 3,562개체가 채집되어 25.03%를 차지하였다.
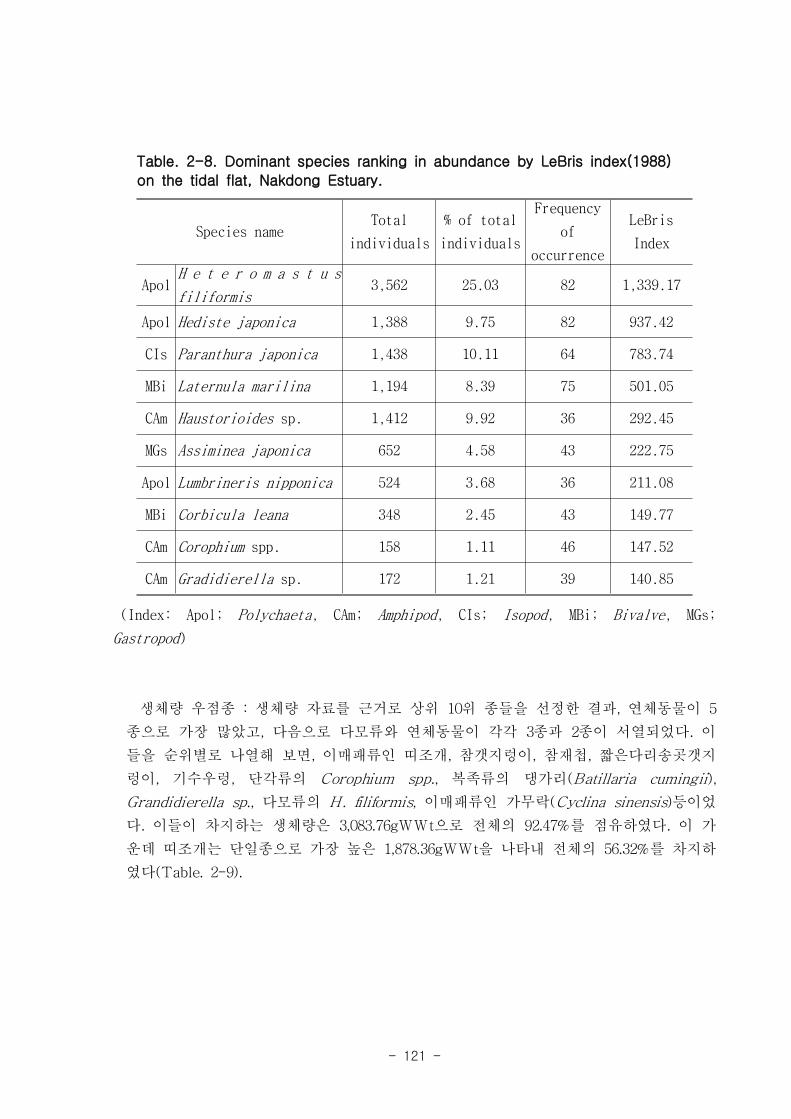
- 121 -
Species nameTotal
individuals
% of total
individuals
Frequency
of
occurrence
LeBris
Index
ApolH e t e r o m a s t u s
filiformis3,562 25.03 82 1,339.17
Apol Hediste japonica 1,388 9.75 82 937.42
CIs Paranthura japonica 1,438 10.11 64 783.74
MBi Laternula marilina 1,194 8.39 75 501.05
CAm Haustorioides sp. 1,412 9.92 36 292.45
MGs Assiminea japonica 652 4.58 43 222.75
Apol Lumbrineris nipponica 524 3.68 36 211.08
MBi Corbicula leana 348 2.45 43 149.77
CAm Corophium spp. 158 1.11 46 147.52
CAm Gradidierella sp. 172 1.21 39 140.85
Table. 2-8. Dominant species ranking in abundance by LeBris index(1988)
on the tidal flat, Nakdong Estuary.
(Index: Apol; Polychaeta, CAm; Amphipod, CIs; Isopod, MBi; Bivalve, MGs;
Gastropod)
생체량 우점종 : 생체량 자료를 근거로 상위 10위 종들을 선정한 결과, 연체동물이 5
종으로 가장 많았고, 다음으로 다모류와 연체동물이 각각 3종과 2종이 서열되었다. 이
들을 순위별로 나열해 보면, 이매패류인 띠조개, 참갯지렁이, 참재첩, 짧은다리송곳갯지
렁이, 기수우렁, 단각류의 Corophium spp., 복족류의 댕가리(Batillaria cumingii),
Grandidierella sp., 다모류의 H. filiformis, 이매패류인 가무락(Cyclina sinensis)등이었
다. 이들이 차지하는 생체량은 3,083.76gWWt으로 전체의 92.47%를 점유하였다. 이 가
운데 띠조개는 단일종으로 가장 높은 1,878.36gWWt을 나타내 전체의 56.32%를 차지하
였다(Table. 2-9).
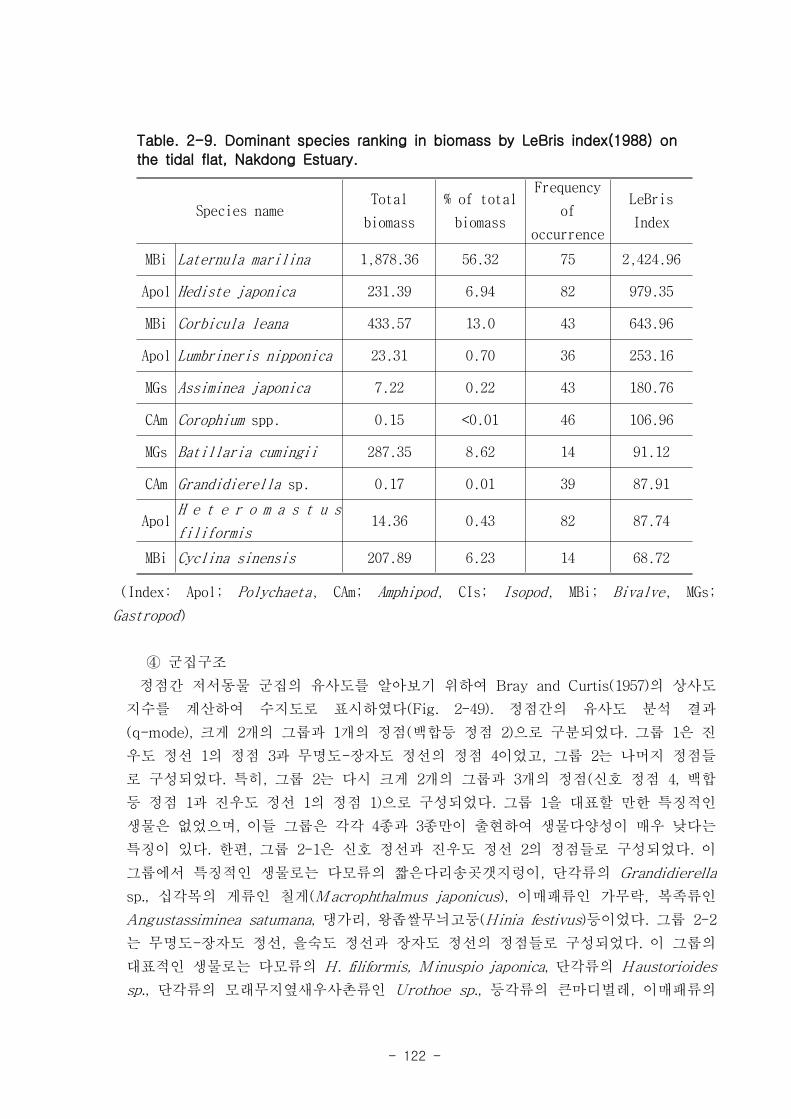
- 122 -
Species nameTotal
biomass
% of total
biomass
Frequency
of
occurrence
LeBris
Index
MBi Laternula marilina 1,878.36 56.32 75 2,424.96
Apol Hediste japonica 231.39 6.94 82 979.35
MBi Corbicula leana 433.57 13.0 43 643.96
Apol Lumbrineris nipponica 23.31 0.70 36 253.16
MGs Assiminea japonica 7.22 0.22 43 180.76
CAm Corophium spp. 0.15 <0.01 46 106.96
MGs Batillaria cumingii 287.35 8.62 14 91.12
CAm Grandidierella sp. 0.17 0.01 39 87.91
ApolH e t e r o m a s t u s
filiformis14.36 0.43 82 87.74
MBi Cyclina sinensis 207.89 6.23 14 68.72
Table. 2-9. Dominant species ranking in biomass by LeBris index(1988) on
the tidal flat, Nakdong Estuary.
(Index: Apol; Polychaeta, CAm; Amphipod, CIs; Isopod, MBi; Bivalve, MGs;
Gastropod)
④ 군집구조
정점간 저서동물 군집의 유사도를 알아보기 위하여 Bray and Curtis(1957)의 상사도
지수를 계산하여 수지도로 표시하였다(Fig. 2-49). 정점간의 유사도 분석 결과
(q-mode), 크게 2개의 그룹과 1개의 정점(백합등 정점 2)으로 구분되었다. 그룹 1은 진
우도 정선 1의 정점 3과 무명도-장자도 정선의 정점 4이었고, 그룹 2는 나머지 정점들
로 구성되었다. 특히, 그룹 2는 다시 크게 2개의 그룹과 3개의 정점(신호 정점 4, 백합
등 정점 1과 진우도 정선 1의 정점 1)으로 구성되었다. 그룹 1을 대표할 만한 특징적인
생물은 없었으며, 이들 그룹은 각각 4종과 3종만이 출현하여 생물다양성이 매우 낮다는
특징이 있다. 한편, 그룹 2-1은 신호 정선과 진우도 정선 2의 정점들로 구성되었다. 이
그룹에서 특징적인 생물로는 다모류의 짧은다리송곳갯지렁이, 단각류의 Grandidierella
sp., 십각목의 게류인 칠게(M acrophthalmus japonicus), 이매패류인 가무락, 복족류인
Angustassiminea satumana, 댕가리, 왕좁쌀무늬고둥(Hinia festivus)등이었다. 그룹 2-2
는 무명도-장자도 정선, 을숙도 정선과 장자도 정선의 정점들로 구성되었다. 이 그룹의
대표적인 생물로는 다모류의 H. filiformis, Minuspio japonica, 단각류의 Haustorioides
sp., 단각류의 모래무지옆새우사촌류인 Urothoe sp., 등각류의 큰마디벌레, 이매패류의
- 123 -
띠조개 등이었다. 집괴분석 결과, 낙동강 하구역의 갯벌 및 모래등에 서식하는 저서동
물의 군집구조는 지역적으로 뚜렷하게 구분되지 않았다. 이것은 상대적으로 퇴적상이
유사하고, 또한 소조차 해역으로서 조위에 따른 영향이 상대적으로 크지 않음을 의미한
다고 볼 수 있다. 한편, 집괴분석 결과는 MDS 분석 결과와도 잘 일치하였다(Fig.
2-49).
BH
T-2
JWT1-3
NJT
-4
SH
T-1
SH
T-3
SH
T-2
BH
T-3
JWT2-1
JWT2-2
JWT2-3
JWT2-4
NJT
-2
NJT
-3
JJT-3
NJT
-1
UST-4
BH
T-5
UST-2
UST-3
JJT-2
JJT-4
JJT-1
UST-1
BH
T-6
JWT1-2
SH
T-4
BH
T-1
JWT1-1
Station
100
80
60
40
20
0
Sim
ilarity
(Pre
sence-Absence)
BHT-1
BHT-2
BHT-3
BHT-5
BHT-6JJT-1JJT-2
JJT-3
JJT-4
JWT1-1
JWT1-2
JWT1-3
JWT2-1
JWT2-2JWT2-3
JWT2-4NJT-1NJT-2NJT-3
NJT-4
SHT-1SHT-2
SHT-3
SHT-4
UST-1UST-2UST-3
UST-4
Stress: 0.14
Figure. 2-49. Dendrogram for hierarchical clustering and
2-dimensional MDS configuration, using group-average
linking of Bray-Curtis similarities calculated on
presence-absence transformed abundance data(Intertidal
area).

- 124 -
(다) 낙동강 하구역 조하대 저서동물의 군집구조
① 저서동물의 출현종 수, 개체수 및 생체량
본 조사 결과, 총 36개 정점에서 출현한 저서동물은 145종/7.2㎡, 35,554개체/36㎡와
2,036gWWt/36㎡이었다(Fig. 2-50).
Species number
Mollusca
26.21%
Polychaeta
44.83%
Others
3.45%
Crustacea
25.52%
Abundance
Mollusca
7.90%
Polychaeta
80.00%
Others
0.23%Crustacea
11.87%
Biomass Polychaeta
9.09%
Mollusca
88.17%
Crustacea
1.92%
Others
0.81%
Figure. 2-50. Total number of species, abundance and biomass in the
Nakdong Estuary.
분류군별 출현종 수를 보면, 다모류가 65종으로 44.83%를 점유하였고, 연체동물과 절
지동물이 각각 38종, 26.21%와 37종, 25.52%를 차지하였다. 이 밖에 기타동물군은 5종
으로 3.45%를 점유하였고, 극피동물은 출현하지 않았다. 개체수는 출현종 수와 동일하
게 다모류가 28,444개체가 채집되어 전체의 80%를 점유하여 가장 우점하였다. 다음으
로 절지동물과 연체동물이 각각 4,220개체, 11.87%와 2,810개체, 7.9%를 차지하였고, 기
타동물군은 80개체로 0.23%의 낮은 점유율을 보였다. 생체량은 단일개체의 생체량이
높은 연체동물이 1,795.2gWWt으로 88.17%의 높은 점유율을 보였다. 다음으로 다모류
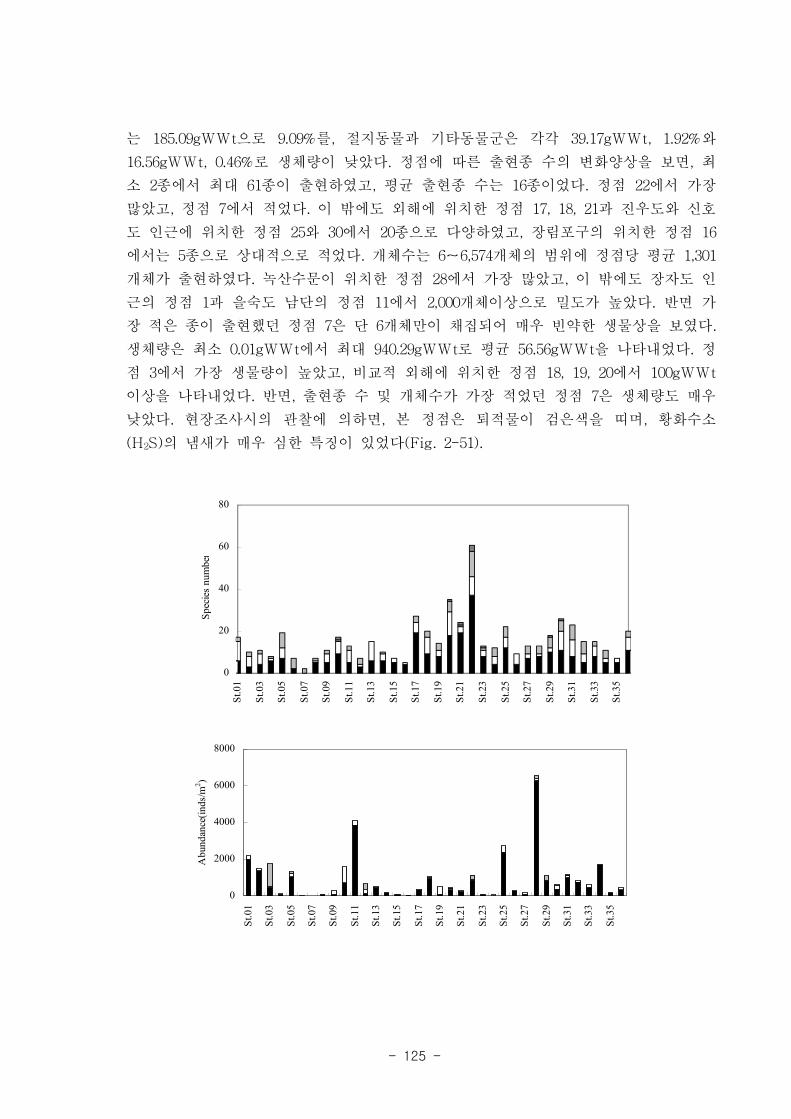
- 125 -
는 185.09gWWt으로 9.09%를, 절지동물과 기타동물군은 각각 39.17gWWt, 1.92%와
16.56gWWt, 0.46%로 생체량이 낮았다. 정점에 따른 출현종 수의 변화양상을 보면, 최
소 2종에서 최대 61종이 출현하였고, 평균 출현종 수는 16종이었다. 정점 22에서 가장
많았고, 정점 7에서 적었다. 이 밖에도 외해에 위치한 정점 17, 18, 21과 진우도와 신호
도 인근에 위치한 정점 25와 30에서 20종으로 다양하였고, 장림포구의 위치한 정점 16
에서는 5종으로 상대적으로 적었다. 개체수는 6~6,574개체의 범위에 정점당 평균 1,301
개체가 출현하였다. 녹산수문이 위치한 정점 28에서 가장 많았고, 이 밖에도 장자도 인
근의 정점 1과 을숙도 남단의 정점 11에서 2,000개체이상으로 밀도가 높았다. 반면 가
장 적은 종이 출현했던 정점 7은 단 6개체만이 채집되어 매우 빈약한 생물상을 보였다.
생체량은 최소 0.01gWWt에서 최대 940.29gWWt로 평균 56.56gWWt을 나타내었다. 정
점 3에서 가장 생물량이 높았고, 비교적 외해에 위치한 정점 18, 19, 20에서 100gWWt
이상을 나타내었다. 반면, 출현종 수 및 개체수가 가장 적었던 정점 7은 생체량도 매우
낮았다. 현장조사시의 관찰에 의하면, 본 정점은 퇴적물이 검은색을 띠며, 황화수소
(H2S)의 냄새가 매우 심한 특징이 있었다(Fig. 2-51).
0
20
40
60
80
St.01
St.03
St.05
St.07
St.09
St.11
St.13
St.15
St.17
St.19
St.21
St.23
St.25
St.27
St.29
St.31
St.33
St.35
Species number
0
2000
4000
6000
8000
St.01
St.03
St.05
St.07
St.09
St.11
St.13
St.15
St.17
St.19
St.21
St.23
St.25
St.27
St.29
St.31
St.33
St.35
Abundance(inds/m
2)
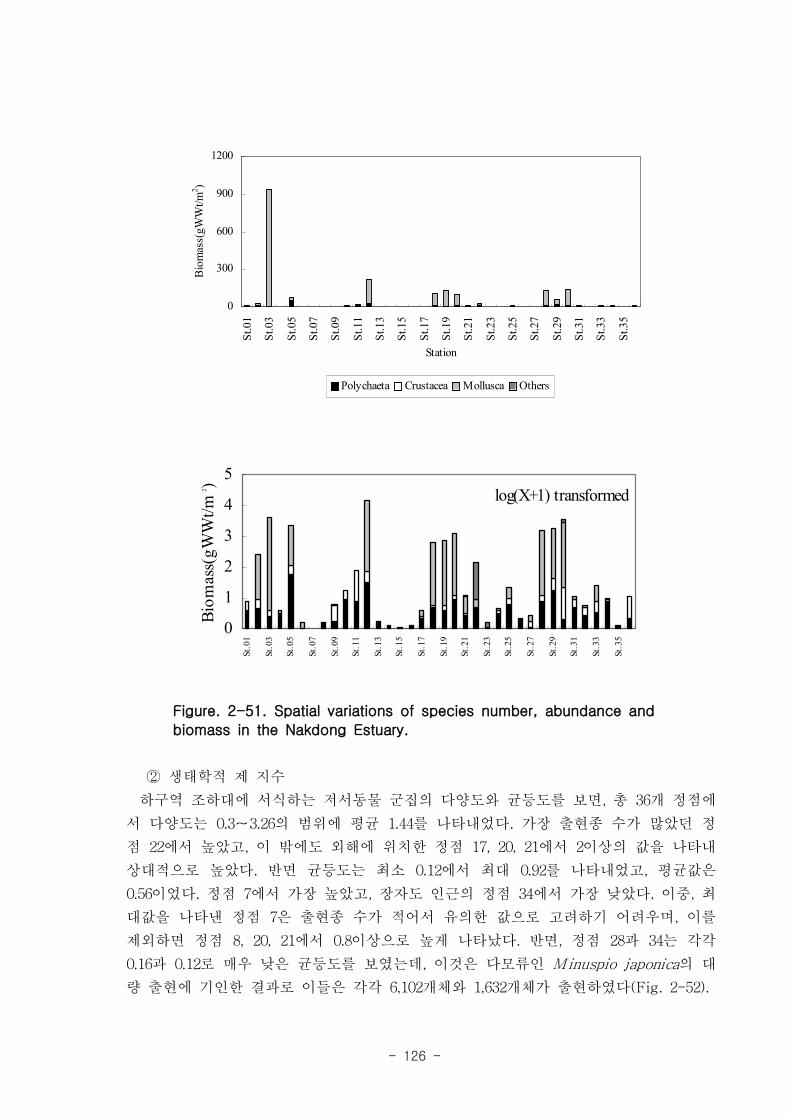
- 126 -
0
300
600
900
1200
St.01
St.03
St.05
St.07
St.09
St.11
St.13
St.15
St.17
St.19
St.21
St.23
St.25
St.27
St.29
St.31
St.33
St.35
Station
Biomass(gWWt/m
2)
Polychaeta Crustacea Mollusca Others
0
1
2
3
4
5
St.01
St.03
St.05
St.07
St.09
St.11
St.13
St.15
St.17
St.19
St.21
St.23
St.25
St.27
St.29
St.31
St.33
St.35
log(X+1) transformed
Biomass(gWWt/m
2
)
Figure. 2-51. Spatial variations of species number, abundance and
biomass in the Nakdong Estuary.
② 생태학적 제 지수
하구역 조하대에 서식하는 저서동물 군집의 다양도와 균등도를 보면, 총 36개 정점에
서 다양도는 0.3~3.26의 범위에 평균 1.44를 나타내었다. 가장 출현종 수가 많았던 정
점 22에서 높았고, 이 밖에도 외해에 위치한 정점 17, 20, 21에서 2이상의 값을 나타내
상대적으로 높았다. 반면 균등도는 최소 0.12에서 최대 0.92를 나타내었고, 평균값은
0.56이었다. 정점 7에서 가장 높았고, 장자도 인근의 정점 34에서 가장 낮았다. 이중, 최
대값을 나타낸 정점 7은 출현종 수가 적어서 유의한 값으로 고려하기 어려우며, 이를
제외하면 정점 8, 20, 21에서 0.8이상으로 높게 나타났다. 반면, 정점 28과 34는 각각
0.16과 0.12로 매우 낮은 균등도를 보였는데, 이것은 다모류인 Minuspio japonica의 대
량 출현에 기인한 결과로 이들은 각각 6,102개체와 1,632개체가 출현하였다(Fig. 2-52).
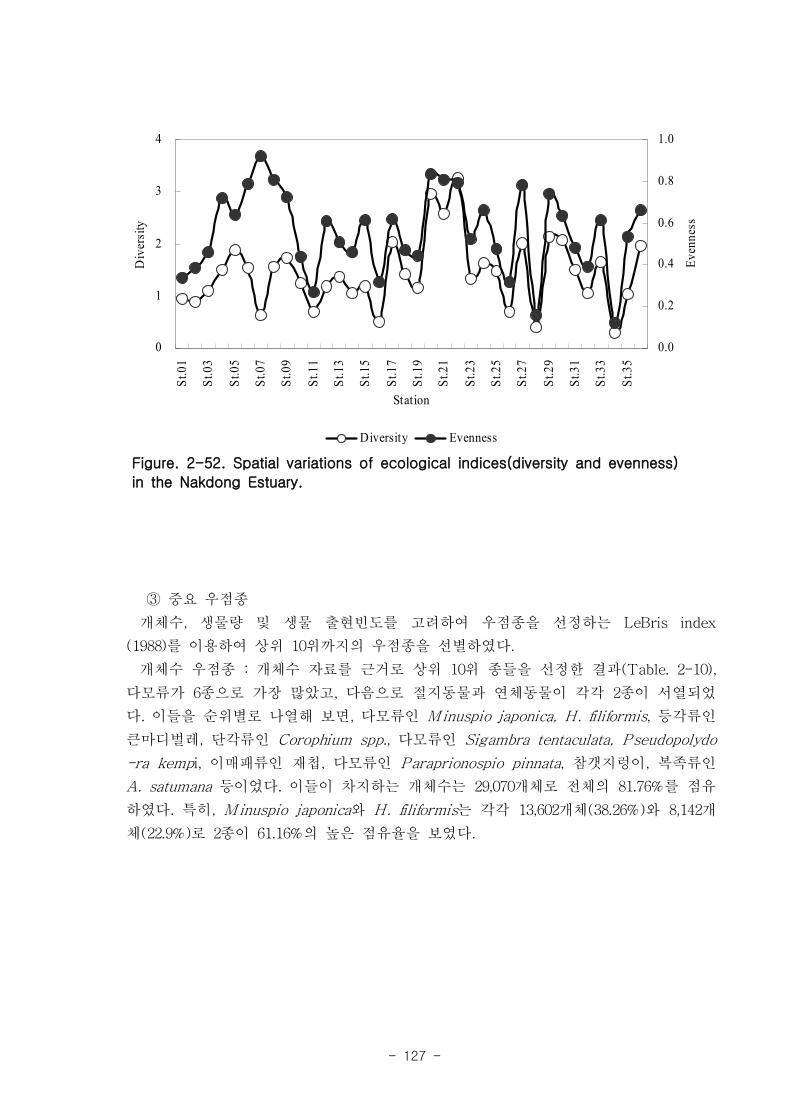
- 127 -
0
1
2
3
4
St.01
St.03
St.05
St.07
St.09
St.11
St.13
St.15
St.17
St.19
St.21
St.23
St.25
St.27
St.29
St.31
St.33
St.35
Station
Diversity
0.0
0.2
0.4
0.6
0.8
1.0
Even
ness
Diversity Evenness
Figure. 2-52. Spatial variations of ecological indices(diversity and evenness)
in the Nakdong Estuary.
③ 중요 우점종
개체수, 생물량 및 생물 출현빈도를 고려하여 우점종을 선정하는 LeBris index
(1988)를 이용하여 상위 10위까지의 우점종을 선별하였다.
개체수 우점종 : 개체수 자료를 근거로 상위 10위 종들을 선정한 결과(Table. 2-10),
다모류가 6종으로 가장 많았고, 다음으로 절지동물과 연체동물이 각각 2종이 서열되었
다. 이들을 순위별로 나열해 보면, 다모류인 Minuspio japonica, H. filiformis, 등각류인
큰마디벌레, 단각류인 Corophium spp., 다모류인 Sigambra tentaculata, Pseudopolydo
-ra kempi, 이매패류인 재첩, 다모류인 Paraprionospio pinnata, 참갯지렁이, 복족류인
A. satumana 등이었다. 이들이 차지하는 개체수는 29,070개체로 전체의 81.76%를 점유
하였다. 특히, M inuspio japonica와 H. filiformis는 각각 13,602개체(38.26%)와 8,142개
체(22.9%)로 2종이 61.16%의 높은 점유율을 보였다.
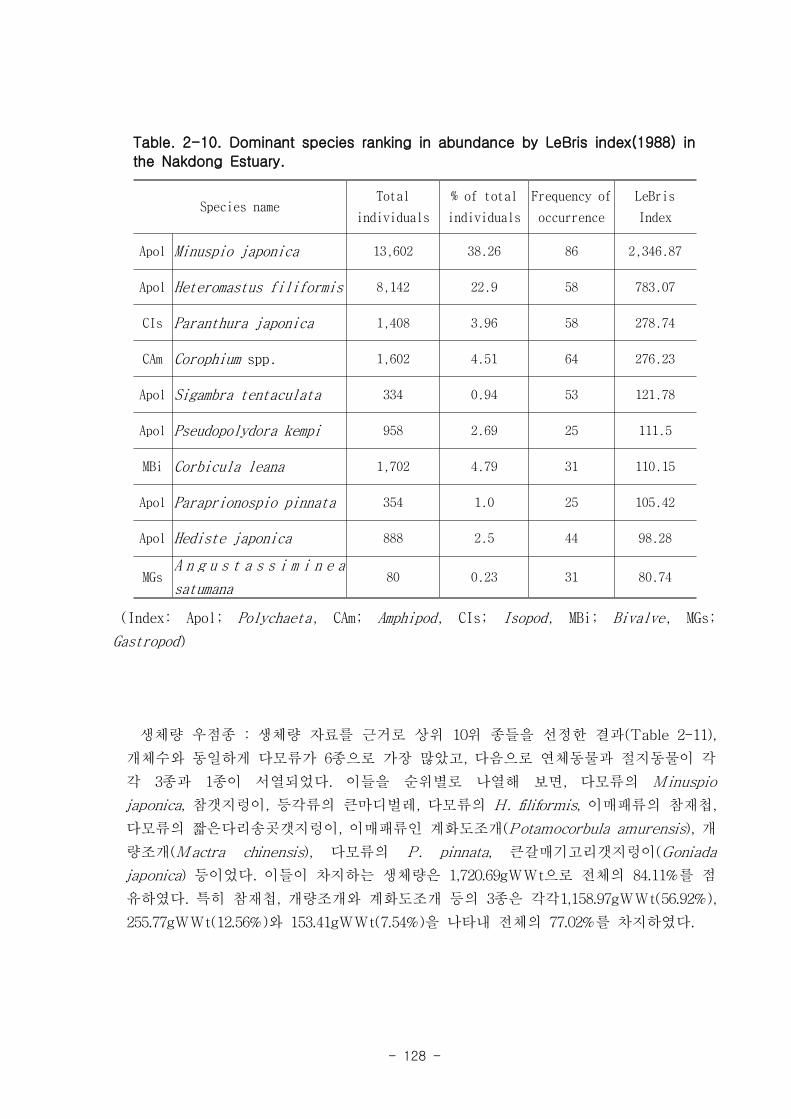
- 128 -
Species nameTotal
individuals
% of total
individuals
Frequency of
occurrence
LeBris
Index
Apol Minuspio japonica 13,602 38.26 86 2,346.87
Apol Heteromastus filiformis 8,142 22.9 58 783.07
CIs Paranthura japonica 1,408 3.96 58 278.74
CAm Corophium spp. 1,602 4.51 64 276.23
Apol Sigambra tentaculata 334 0.94 53 121.78
Apol Pseudopolydora kempi 958 2.69 25 111.5
MBi Corbicula leana 1,702 4.79 31 110.15
Apol Paraprionospio pinnata 354 1.0 25 105.42
Apol Hediste japonica 888 2.5 44 98.28
MGsA n g u s t a s s i m i n e a
satumana80 0.23 31 80.74
Table. 2-10. Dominant species ranking in abundance by LeBris index(1988) in
the Nakdong Estuary.
(Index: Apol; Polychaeta, CAm; Amphipod, CIs; Isopod, MBi; Bivalve, MGs;
Gastropod)
생체량 우점종 : 생체량 자료를 근거로 상위 10위 종들을 선정한 결과(Table 2-11),
개체수와 동일하게 다모류가 6종으로 가장 많았고, 다음으로 연체동물과 절지동물이 각
각 3종과 1종이 서열되었다. 이들을 순위별로 나열해 보면, 다모류의 Minuspio
japonica, 참갯지렁이, 등각류의 큰마디벌레, 다모류의 H. filiformis, 이매패류의 참재첩,
다모류의 짧은다리송곳갯지렁이, 이매패류인 계화도조개(Potamocorbula amurensis), 개
량조개(M actra chinensis), 다모류의 P. pinnata, 큰갈매기고리갯지렁이(Goniada
japonica) 등이었다. 이들이 차지하는 생체량은 1,720.69gWWt으로 전체의 84.11%를 점
유하였다. 특히 참재첩, 개량조개와 계화도조개 등의 3종은 각각1,158.97gWWt(56.92%),
255.77gWWt(12.56%)와 153.41gWWt(7.54%)을 나타내 전체의 77.02%를 차지하였다.
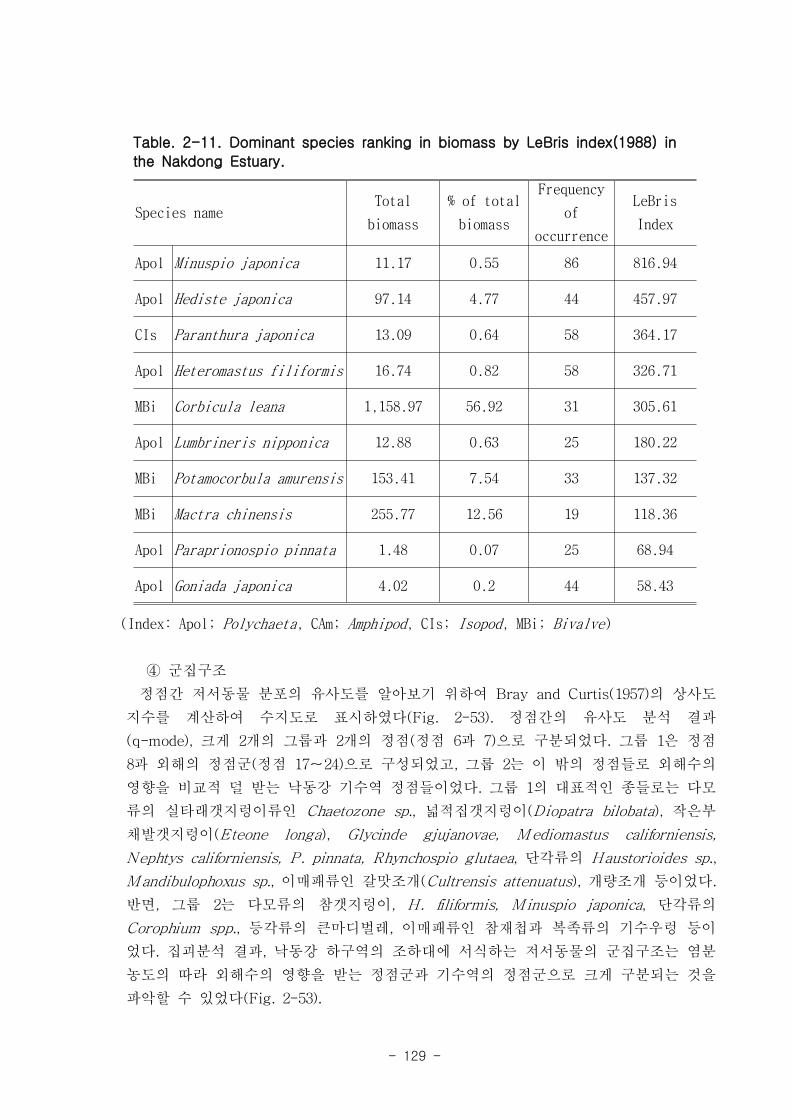
- 129 -
Species nameTotal
biomass
% of total
biomass
Frequency
of
occurrence
LeBris
Index
Apol Minuspio japonica 11.17 0.55 86 816.94
Apol Hediste japonica 97.14 4.77 44 457.97
CIs Paranthura japonica 13.09 0.64 58 364.17
Apol Heteromastus filiformis 16.74 0.82 58 326.71
MBi Corbicula leana 1,158.97 56.92 31 305.61
Apol Lumbrineris nipponica 12.88 0.63 25 180.22
MBi Potamocorbula amurensis 153.41 7.54 33 137.32
MBi Mactra chinensis 255.77 12.56 19 118.36
Apol Paraprionospio pinnata 1.48 0.07 25 68.94
Apol Goniada japonica 4.02 0.2 44 58.43
Table. 2-11. Dominant species ranking in biomass by LeBris index(1988) in
the Nakdong Estuary.
(Index: Apol; Polychaeta, CAm; Amphipod, CIs; Isopod, MBi; Bivalve)
④ 군집구조
정점간 저서동물 분포의 유사도를 알아보기 위하여 Bray and Curtis(1957)의 상사도
지수를 계산하여 수지도로 표시하였다(Fig. 2-53). 정점간의 유사도 분석 결과
(q-mode), 크게 2개의 그룹과 2개의 정점(정점 6과 7)으로 구분되었다. 그룹 1은 정점
8과 외해의 정점군(정점 17~24)으로 구성되었고, 그룹 2는 이 밖의 정점들로 외해수의
영향을 비교적 덜 받는 낙동강 기수역 정점들이었다. 그룹 1의 대표적인 종들로는 다모
류의 실타래갯지렁이류인 Chaetozone sp., 넓적집갯지렁이(Diopatra bilobata), 작은부
채발갯지렁이(Eteone longa), Glycinde gjujanovae, M ediomastus californiensis,
Nephtys californiensis, P. pinnata, Rhynchospio glutaea, 단각류의 Haustorioides sp.,
Mandibulophoxus sp., 이매패류인 갈맛조개(Cultrensis attenuatus), 개량조개 등이었다.
반면, 그룹 2는 다모류의 참갯지렁이, H. filiformis, Minuspio japonica, 단각류의
Corophium spp., 등각류의 큰마디벌레, 이매패류인 참재첩과 복족류의 기수우렁 등이
었다. 집괴분석 결과, 낙동강 하구역의 조하대에 서식하는 저서동물의 군집구조는 염분
농도의 따라 외해수의 영향을 받는 정점군과 기수역의 정점군으로 크게 구분되는 것을
파악할 수 있었다(Fig. 2-53).
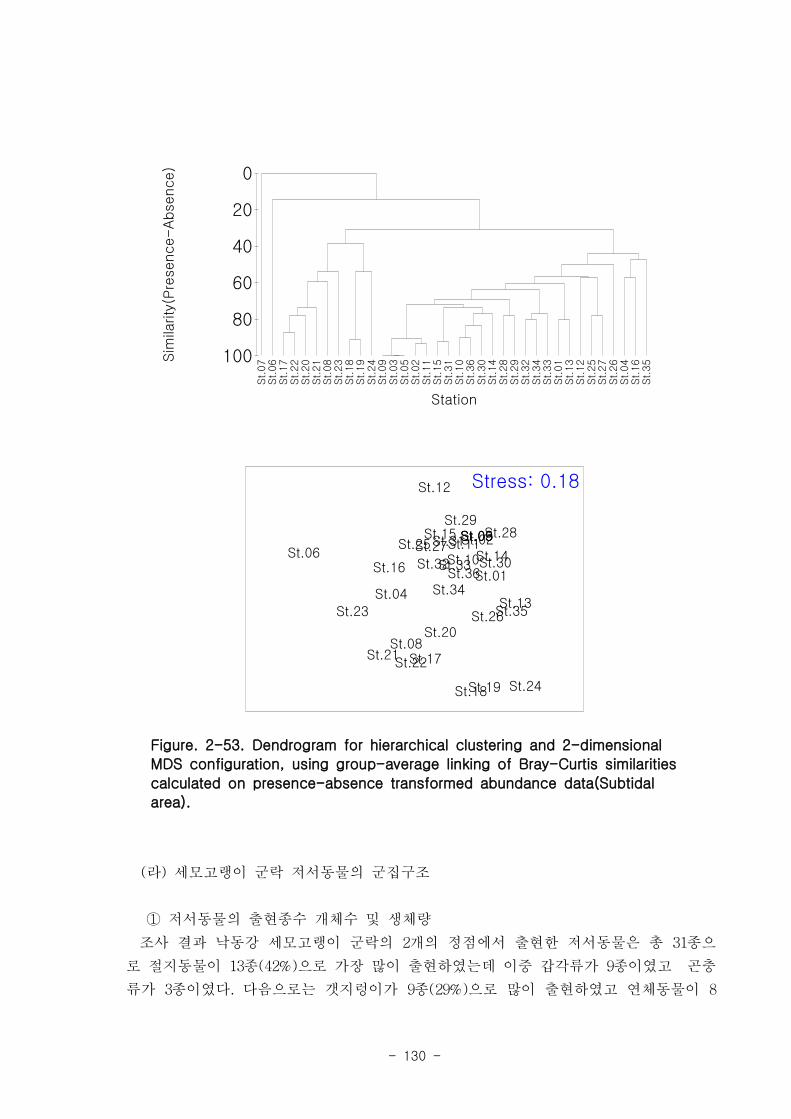
- 130 -
St.07
St.06
St.17
St.22
St.20
St.21
St.08
St.23
St.18
St.19
St.24
St.09
St.03
St.05
St.02
St.11
St.15
St.31
St.10
St.36
St.30
St.14
St.28
St.29
St.32
St.34
St.33
St.01
St.13
St.12
St.25
St.27
St.26
St.04
St.16
St.35
Station
100
80
60
40
20
0Sim
ilarity
(Pre
sence-Absence)
St.01
St.02St.03
St.04
St.05St.06
St.08
St.09
St.10St.11
St.12
St.13
St.14
St.15
St.16
St.17
St.18St.19
St.20
St.21St.22
St.23
St.24
St.25
St.26
St.27St.28
St.29
St.30
St.31
St.32St.33
St.34
St.35
St.36
Stress: 0.18
Figure. 2-53. Dendrogram for hierarchical clustering and 2-dimensional
MDS configuration, using group-average linking of Bray-Curtis similarities
calculated on presence-absence transformed abundance data(Subtidal
area).
(라) 세모고랭이 군락 저서동물의 군집구조
① 저서동물의 출현종수 개체수 및 생체량
조사 결과 낙동강 세모고랭이 군락의 2개의 정점에서 출현한 저서동물은 총 31종으
로 절지동물이 13종(42%)으로 가장 많이 출현하였는데 이중 갑각류가 9종이였고 곤충
류가 3종이였다. 다음으로는 갯지렁이가 9종(29%)으로 많이 출현하였고 연체동물이 8
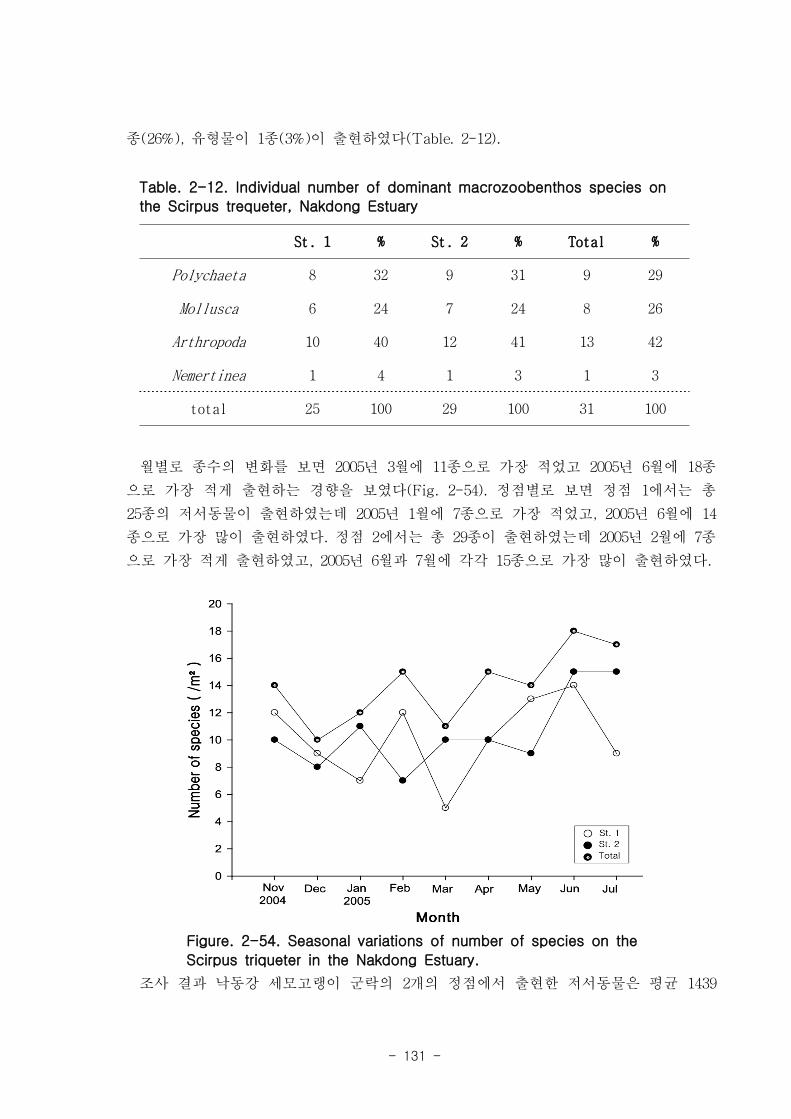
- 131 -
종(26%), 유형물이 1종(3%)이 출현하였다(Table. 2-12).
St. 1 % St. 2 % Total %
Polychaeta 8 32 9 31 9 29
Mollusca 6 24 7 24 8 26
Arthropoda 10 40 12 41 13 42
Nemertinea 1 4 1 3 1 3
total 25 100 29 100 31 100
Table. 2-12. Individual number of dominant macrozoobenthos species on
the Scirpus trequeter, Nakdong Estuary
월별로 종수의 변화를 보면 2005년 3월에 11종으로 가장 적었고 2005년 6월에 18종
으로 가장 적게 출현하는 경향을 보였다(Fig. 2-54). 정점별로 보면 정점 1에서는 총
25종의 저서동물이 출현하였는데 2005년 1월에 7종으로 가장 적었고, 2005년 6월에 14
종으로 가장 많이 출현하였다. 정점 2에서는 총 29종이 출현하였는데 2005년 2월에 7종
으로 가장 적게 출현하였고, 2005년 6월과 7월에 각각 15종으로 가장 많이 출현하였다.
Figure. 2-54. Seasonal variations of number of species on the
Scirpus triqueter in the Nakdong Estuary.
조사 결과 낙동강 세모고랭이 군락의 2개의 정점에서 출현한 저서동물은 평균 1439
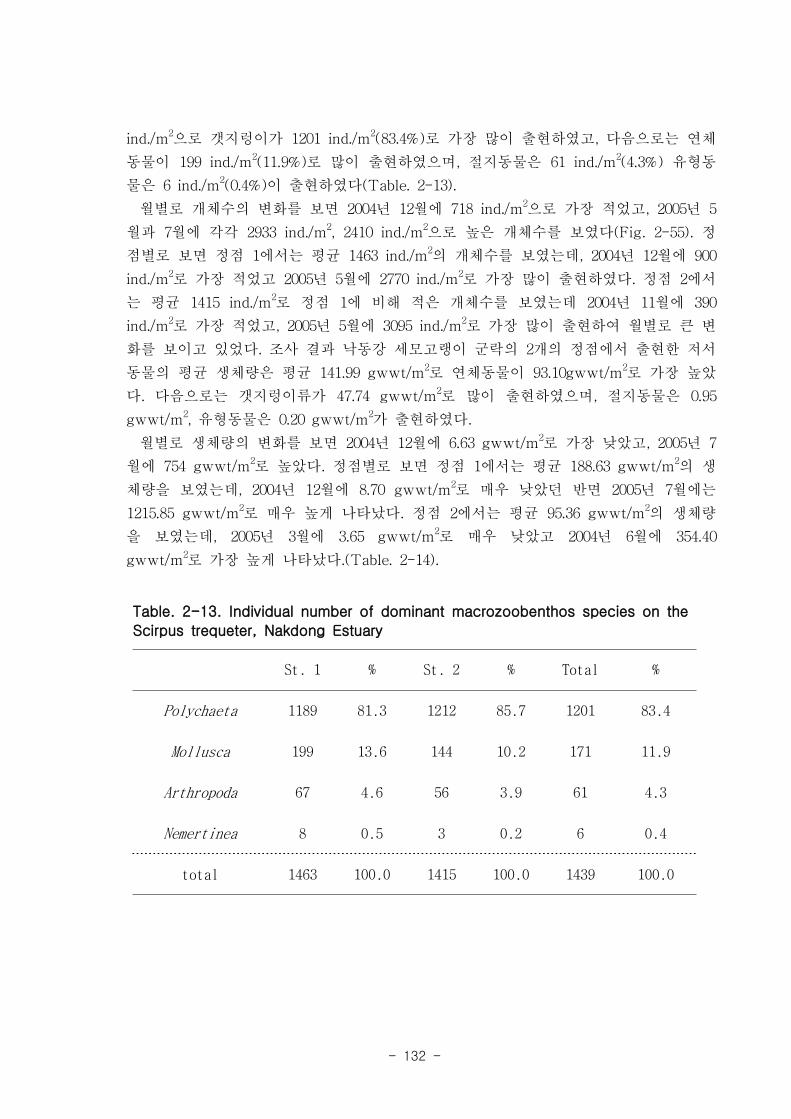
- 132 -
ind./m2으로 갯지렁이가 1201 ind./m
2(83.4%)로 가장 많이 출현하였고, 다음으로는 연체
동물이 199 ind./m2(11.9%)로 많이 출현하였으며, 절지동물은 61 ind./m
2(4.3%) 유형동
물은 6 ind./m2(0.4%)이 출현하였다(Table. 2-13).
월별로 개체수의 변화를 보면 2004년 12월에 718 ind./m2으로 가장 적었고, 2005년 5
월과 7월에 각각 2933 ind./m2, 2410 ind./m
2으로 높은 개체수를 보였다(Fig. 2-55). 정
점별로 보면 정점 1에서는 평균 1463 ind./m2의 개체수를 보였는데, 2004년 12월에 900
ind./m2로 가장 적었고 2005년 5월에 2770 ind./m
2로 가장 많이 출현하였다. 정점 2에서
는 평균 1415 ind./m2로 정점 1에 비해 적은 개체수를 보였는데 2004년 11월에 390
ind./m2로 가장 적었고, 2005년 5월에 3095 ind./m
2로 가장 많이 출현하여 월별로 큰 변
화를 보이고 있었다. 조사 결과 낙동강 세모고랭이 군락의 2개의 정점에서 출현한 저서
동물의 평균 생체량은 평균 141.99 gwwt/m2로 연체동물이 93.10gwwt/m
2로 가장 높았
다. 다음으로는 갯지렁이류가 47.74 gwwt/m2로 많이 출현하였으며, 절지동물은 0.95
gwwt/m2, 유형동물은 0.20 gwwt/m
2가 출현하였다.
월별로 생체량의 변화를 보면 2004년 12월에 6.63 gwwt/m2로 가장 낮았고, 2005년 7
월에 754 gwwt/m2로 높았다. 정점별로 보면 정점 1에서는 평균 188.63 gwwt/m
2의 생
체량을 보였는데, 2004년 12월에 8.70 gwwt/m2로 매우 낮았던 반면 2005년 7월에는
1215.85 gwwt/m2로 매우 높게 나타났다. 정점 2에서는 평균 95.36 gwwt/m
2의 생체량
을 보였는데, 2005년 3월에 3.65 gwwt/m2로 매우 낮았고 2004년 6월에 354.40
gwwt/m2로 가장 높게 나타났다.(Table. 2-14).
St. 1 % St. 2 % Total %
Polychaeta 1189 81.3 1212 85.7 1201 83.4
Mollusca 199 13.6 144 10.2 171 11.9
Arthropoda 67 4.6 56 3.9 61 4.3
Nemertinea 8 0.5 3 0.2 6 0.4
total 1463 100.0 1415 100.0 1439 100.0
Table. 2-13. Individual number of dominant macrozoobenthos species on the
Scirpus trequeter, Nakdong Estuary
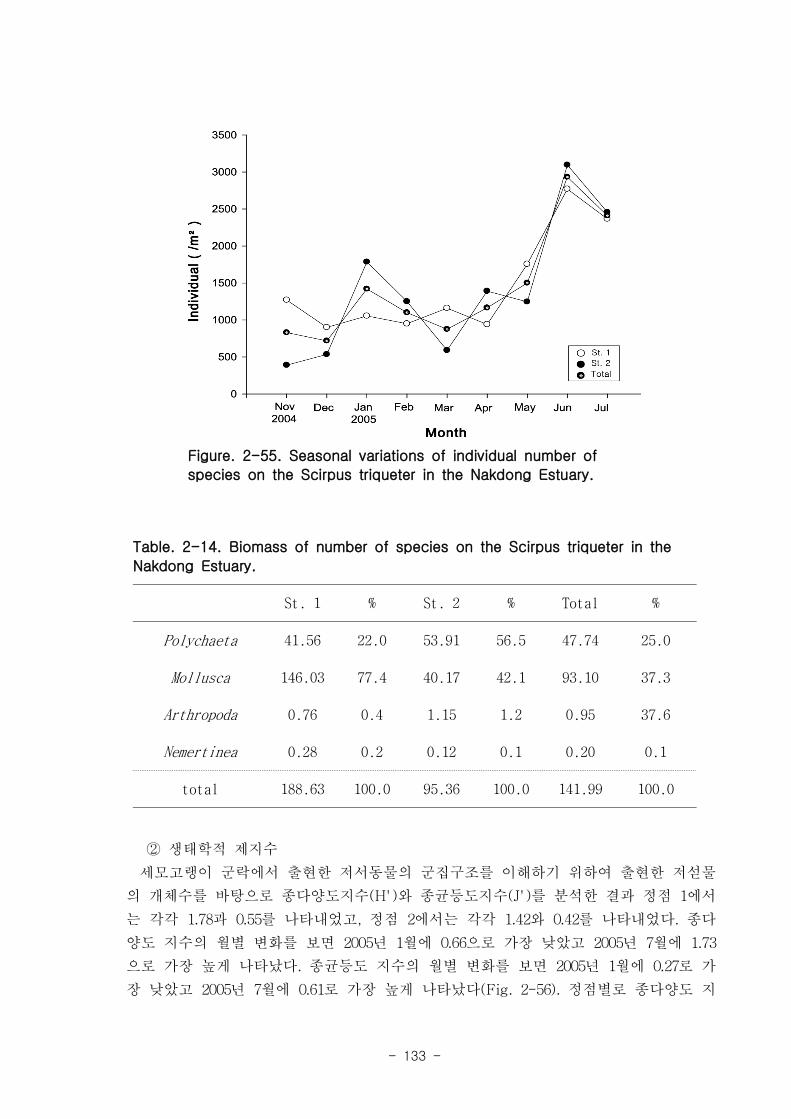
- 133 -
Figure. 2-55. Seasonal variations of individual number of
species on the Scirpus triqueter in the Nakdong Estuary.
St. 1 % St. 2 % Total %
Polychaeta 41.56 22.0 53.91 56.5 47.74 25.0
Mollusca 146.03 77.4 40.17 42.1 93.10 37.3
Arthropoda 0.76 0.4 1.15 1.2 0.95 37.6
Nemertinea 0.28 0.2 0.12 0.1 0.20 0.1
total 188.63 100.0 95.36 100.0 141.99 100.0
Table. 2-14. Biomass of number of species on the Scirpus triqueter in the
Nakdong Estuary.
② 생태학적 제지수
세모고랭이 군락에서 출현한 저서동물의 군집구조를 이해하기 위하여 출현한 저섣물
의 개체수를 바탕으로 종다양도지수(H' )와 종균등도지수(J' )를 분석한 결과 정점 1에서
는 각각 1.78과 0.55를 나타내었고, 정점 2에서는 각각 1.42와 0.42를 나타내었다. 종다
양도 지수의 월별 변화를 보면 2005년 1월에 0.66으로 가장 낮았고 2005년 7월에 1.73
으로 가장 높게 나타났다. 종균등도 지수의 월별 변화를 보면 2005년 1월에 0.27로 가
장 낮았고 2005년 7월에 0.61로 가장 높게 나타났다(Fig. 2-56). 정점별로 종다양도 지
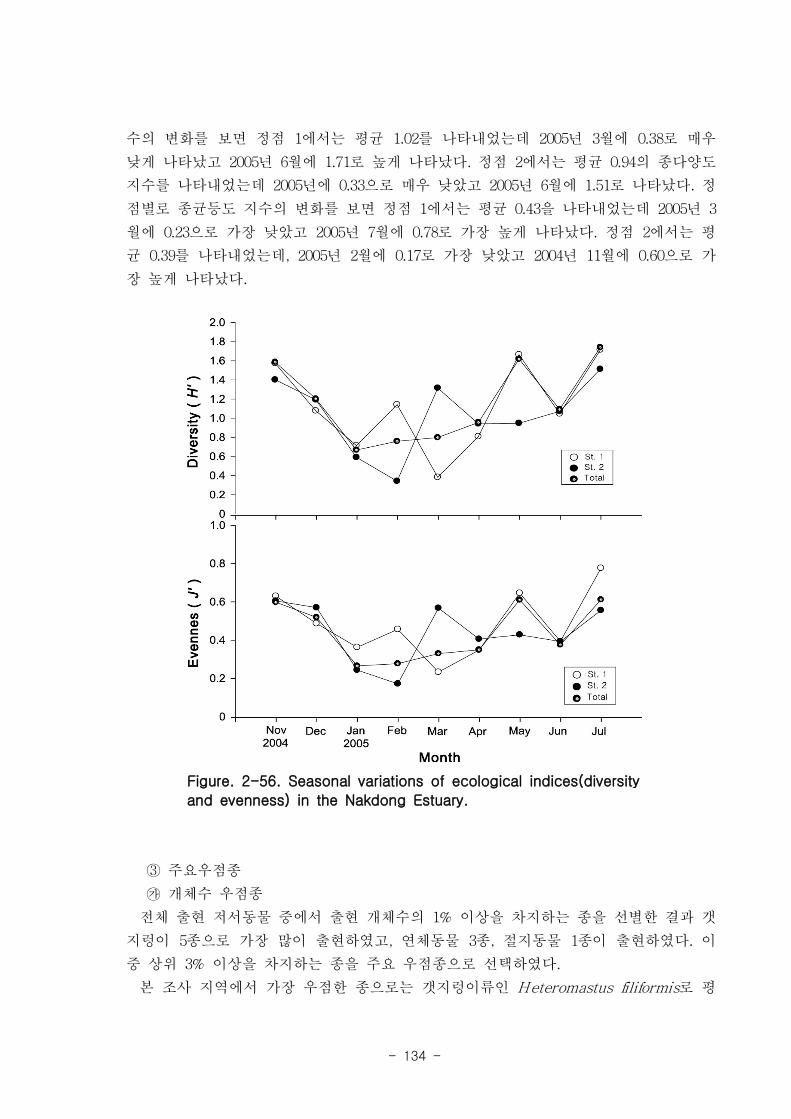
- 134 -
수의 변화를 보면 정점 1에서는 평균 1.02를 나타내었는데 2005년 3월에 0.38로 매우
낮게 나타났고 2005년 6월에 1.71로 높게 나타났다. 정점 2에서는 평균 0.94의 종다양도
지수를 나타내었는데 2005년에 0.33으로 매우 낮았고 2005년 6월에 1.51로 나타났다. 정
점별로 종균등도 지수의 변화를 보면 정점 1에서는 평균 0.43을 나타내었는데 2005년 3
월에 0.23으로 가장 낮았고 2005년 7월에 0.78로 가장 높게 나타났다. 정점 2에서는 평
균 0.39를 나타내었는데, 2005년 2월에 0.17로 가장 낮았고 2004년 11월에 0.60으로 가
장 높게 나타났다.
Figure. 2-56. Seasonal variations of ecological indices(diversity
and evenness) in the Nakdong Estuary.
③ 주요우점종
㉮ 개체수 우점종
전체 출현 저서동물 중에서 출현 개체수의 1% 이상을 차지하는 종을 선별한 결과 갯
지렁이 5종으로 가장 많이 출현하였고, 연체동물 3종, 절지동물 1종이 출현하였다. 이
중 상위 3% 이상을 차지하는 종을 주요 우점종으로 선택하였다.
본 조사 지역에서 가장 우점한 종으로는 갯지렁이류인 Heteromastus filiformis로 평
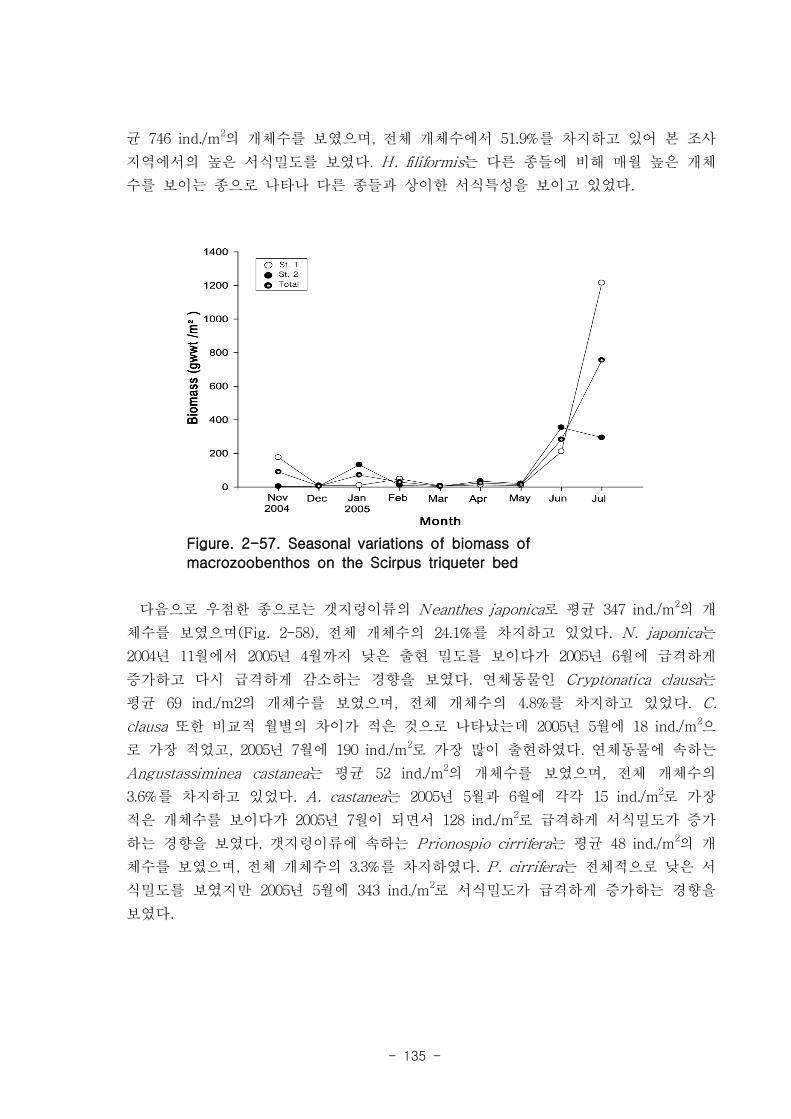
- 135 -
균 746 ind./m2의 개체수를 보였으며, 전체 개체수에서 51.9%를 차지하고 있어 본 조사
지역에서의 높은 서식밀도를 보였다. H. filiformis는 다른 종들에 비해 매월 높은 개체
수를 보이는 종으로 나타나 다른 종들과 상이한 서식특성을 보이고 있었다.
Figure. 2-57. Seasonal variations of biomass of
macrozoobenthos on the Scirpus triqueter bed
다음으로 우점한 종으로는 갯지렁이류의 Neanthes japonica로 평균 347 ind./m2의 개
체수를 보였으며(Fig. 2-58), 전체 개체수의 24.1%를 차지하고 있었다. N. japonica는
2004년 11월에서 2005년 4월까지 낮은 출현 밀도를 보이다가 2005년 6월에 급격하게
증가하고 다시 급격하게 감소하는 경향을 보였다. 연체동물인 Cryptonatica clausa는
평균 69 ind./m2의 개체수를 보였으며, 전체 개체수의 4.8%를 차지하고 있었다. C.
clausa 또한 비교적 월별의 차이가 적은 것으로 나타났는데 2005년 5월에 18 ind./m2으
로 가장 적었고, 2005년 7월에 190 ind./m2로 가장 많이 출현하였다. 연체동물에 속하는
Angustassiminea castanea는 평균 52 ind./m2의 개체수를 보였으며, 전체 개체수의
3.6%를 차지하고 있었다. A. castanea는 2005년 5월과 6월에 각각 15 ind./m2로 가장
적은 개체수를 보이다가 2005년 7월이 되면서 128 ind./m2로 급격하게 서식밀도가 증가
하는 경향을 보였다. 갯지렁이류에 속하는 Prionospio cirrifera는 평균 48 ind./m2의 개
체수를 보였으며, 전체 개체수의 3.3%를 차지하였다. P. cirrifera는 전체적으로 낮은 서
식밀도를 보였지만 2005년 5월에 343 ind./m2로 서식밀도가 급격하게 증가하는 경향을
보였다.
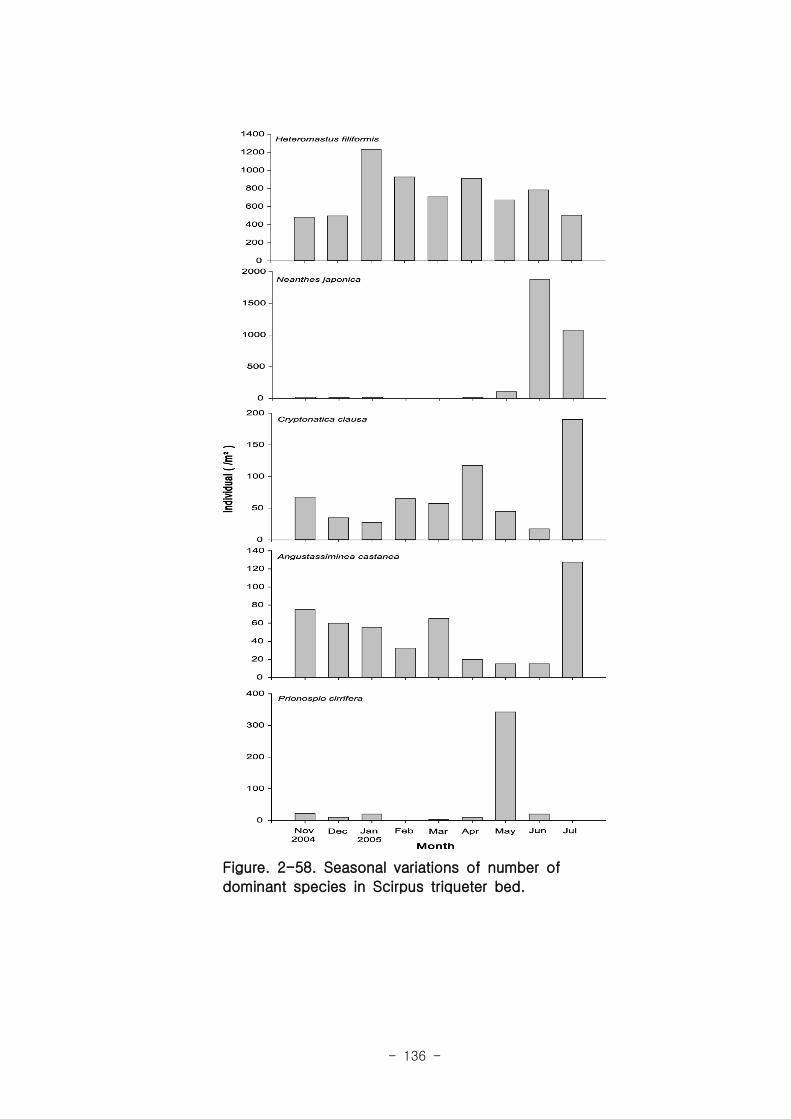
- 136 -
Figure. 2-58. Seasonal variations of number of
dominant species in Scirpus triqueter bed.
- 137 -
Table. 2-15. Individual number of species in stn. 1 of the Scirpus triqueter bed
(ind./m2)
Species Nov Dec Jan Feb Mar Apr May Jun Jul 합계
PolychaetaTylorrhynchus
heterochaetus0 0 0 0 0 5 0 50 0 6
Neanthes japonica 15 0 5 5 0 0 185 1750 890 317
Prionospio cirrifera 30 5 40 0 0 20 680 20 0 88
Eteone sp. 5 30 5 0 0 15 320 15 0 43
Heteromastus
filiformis730 650 880 685 1065 775 395 785 435 711
Polychaeta unid. 0 105 40 0 0 0 0 0 0 16
Nectoneanthes sp. 0 0 0 0 0 0 0 0 0 0
Pseudopotamilla
occelata30 0 0 30 0 0 0 0 0 7
Polydora ligni 0 0 0 0 0 0 0 5 0 1
Nemertinea Nemertea unid. 0 15 0 25 0 0 30 0 0 8
Mollusca Laternula marilina 0 0 0 15 0 0 10 5 485 57
Corbicula sp. 0 0 0 0 0 0 0 0 150 17
Moerella rutila 0 0 0 0 0 0 0 5 0 1
Corbicula japonica 25 0 0 15 0 0 0 5 30 8
Heteromacoma irus 0 0 0 0 0 0 0 0 0 0
Myadora sp. 0 0 0 0 0 0 0 0 0 0
Angustassiminea
castanea95 30 55 40 20 20 10 25 125 47
Cryptonatica clausa 100 35 30 100 55 55 50 20 180 69
Crustacea Cumacea unid. 0 0 0 0 10 5 0 0 0 2
Ilyoplx pusilla 100 0 0 20 0 0 5 10 0 15
Scopimera globosa 0 0 0 0 0 0 5 0 15 2
Tanaidacea unid. 15 10 0 0 0 0 0 0 0 3
Isopoda unid. 20 0 0 5 0 0 5 0 55 9
Gammaridea unid.1 0 0 0 0 0 0 0 0 0 0
Gammaridea unid.2 0 0 0 5 0 0 0 0 0 1
Gammaridea unid.3 0 0 0 0 0 0 0 0 0 0
Dacapoda larvae 0 0 0 0 0 15 0 0 0 2
Insecta Insecta unid.1 105 20 0 5 10 0 55 15 0 23
Insecta unid.2 0 0 0 0 0 0 0 0 0 0
Insecta unid.3 0 0 0 0 0 25 0 0 0 3
Insecta unid.4 0 0 0 0 0 5 5 60 0 8
Total 1270 900 1055 950 1160 940 1755 2770 2365 1463
- 138 -
Table. 2-16. Individual number of species in stn. 2 of the Scirpus triqueter bed
(ind./m2)
Species Nov Dec Jan Feb Mar Apr May Jun Jul 합계
PolychaetaTylorrhynchus
heterochaetus5 0 0 10 0 15 20 0 35 9
Neanthes japonica 20 30 25 0 5 30 20 1995 1265 377
Prionospio cirrifera 15 15 0 0 5 0 5 20 0 7
Eteone sp. 5 5 45 5 10 0 65 65 0 22
Heteromastus
filiformis235 345 1580 1170 350 1045 950 785 575 782
Polychaeta unid. 0 10 20 0 5 70 0 0 15 13
Nectoneanthes sp. 0 0 0 5 0 0 0 0 0 1
Pseudopotamilla
occelata0 0 0 0 0 0 0 10 0 1
Polydora ligni 0 0 0 0 0 0 0 5 0 1
Nemertinea Nemertea unid. 0 0 15 0 0 0 0 15 0 3
Mollusca Laternula marilina 0 0 0 0 0 0 0 105 25 14
Corbicula sp. 0 0 0 0 0 0 0 5 0 1
Moerella rutila 0 0 0 0 0 0 0 0 0 0
Corbicula japonica 0 0 5 0 0 5 0 0 5 2
Heteromacoma irus 0 0 5 0 0 0 0 0 0 1
Myadora sp. 0 0 0 0 0 10 0 0 0 1
Angustassiminea
castanea55 90 55 25 110 20 20 5 130 57
Cryptonatica clausa 35 35 25 30 60 180 40 15 200 69
Crustacea Cumacea unid. 0 0 5 0 0 5 0 0 0 1
Ilyoplx pusilla 0 0 0 5 0 10 0 55 0 8
Scopimera globosa 0 0 0 0 0 0 5 0 0 1
Tanaidacea unid. 5 0 0 0 0 0 0 0 5 1
Isopoda unid. 0 0 0 0 0 0 0 0 5 1
Gammaridea unid.1 5 0 0 0 0 0 0 0 0 1
Gammaridea unid.2 0 0 0 0 0 0 0 5 5 1
Gammaridea unid.3 0 0 0 0 5 0 0 0 0 1
Dacapoda larvae 0 0 0 0 0 0 0 0 0 0
Insecta Insecta unid.1 10 5 5 0 10 0 120 5 105 29
Insecta unid.2 0 0 0 0 0 0 0 0 65 7
Insecta unid.3 0 0 0 0 0 0 0 0 5 1
Insecta unid.4 0 0 0 0 30 0 0 5 15 6
합계 390 535 1785 1250 590 1390 1245 3095 2455 1415

- 139 -
㉯ 생체량 우점종
전체 출현한 저서생물의 생체량중 1% 이상을 차지하는 종을 선별한 결과 연체동물이
5종으로 가장 많았고, 다음으로는 갯지렁이류가 2종으로 많았다. 7종의 생체량의 합은
전체 생체량의 97.9%를 차지할 정도로 세모고랭이 군락에서 우점하는 것으로 나타났는
데 이들 중에서 전체 생체량의 5% 이상을 차지하는 주요 우점종을 선별한 결과 연체
동물이 3종, 갯지렁이류가 2종으로 나타났는데 이들의 합은 전체 생체량의 92.5%를 차
지하였다.
본 조사지역에서 가장 생체량에서 가장 우점한 종은 Laternula marilina로 평균
65.36 gwwt/m2로 전체 생체량의 46.0%를 차지하였으며 2004년 11월에서 2005년 5월까
지는 매우 적은 생체량을 보였지만 2005년 6월, 7월에 각각 77.18 gwwt/m2, 509.93
gwwt/m2로 나타나 여름이 되면서 생체량이 급격히 상승하는 경향을 나타냈다. 다음으
로 우점한 종은 갯지렁이류의 Neanthes japonica로 평균 39.47 gwwt/m2로 전체 생체
량의 27.8 %를 차지하고 있었다. 월별변화를 보면 L. marilina와 마찬가지로 2004년 11
월에서 2005년 5월까지는 매우 낮은 생체량을 보이다가 2005년 6월과 7월에 각각
181.20 gwwt/m2, 171.10 gwwt/m
2을 나타내어 여름이 되면서 생체량이 급격히 증가하
는 경향을 나타냈다. 연체동물인 Corbicula japonica는 평균 11.61 gwwt/m2로 전체 생
체량의 8.2 %를 차지하였다. 월별변화를 보면 2004년 12월과 2005년 3월, 5월에는 전혀
출현하지 않았고, 2004년 11월과 2005년 7월에 각각 36.75 gwwt/m2, 46.75 gwwt/m
2로
높은 생체량을 나타내었다. 연체동물인 Angustassiminea castanea는 평균 7.74
gwwt/m2의 생체량을 보여 전체 생체량의 5.5%를 차지하였다. 월별변화를 보면 2005년
5월과 6월에는 전혀 출현하지 않았지만 2005년 11월과 2005년 2월에 각각 47.7
gwwt/m2, 20.03 gwwt/m
2로 높은 생체량을 보였다. 갯지렁이류의 Heteromastus
filiformis는 평균 7.21 gwwt/m2로 출현하여 전체 생체량의 5.1%를 차지하였으며, 다른
종들에 비해 월별 변동이 작은 것으로 나타났다.
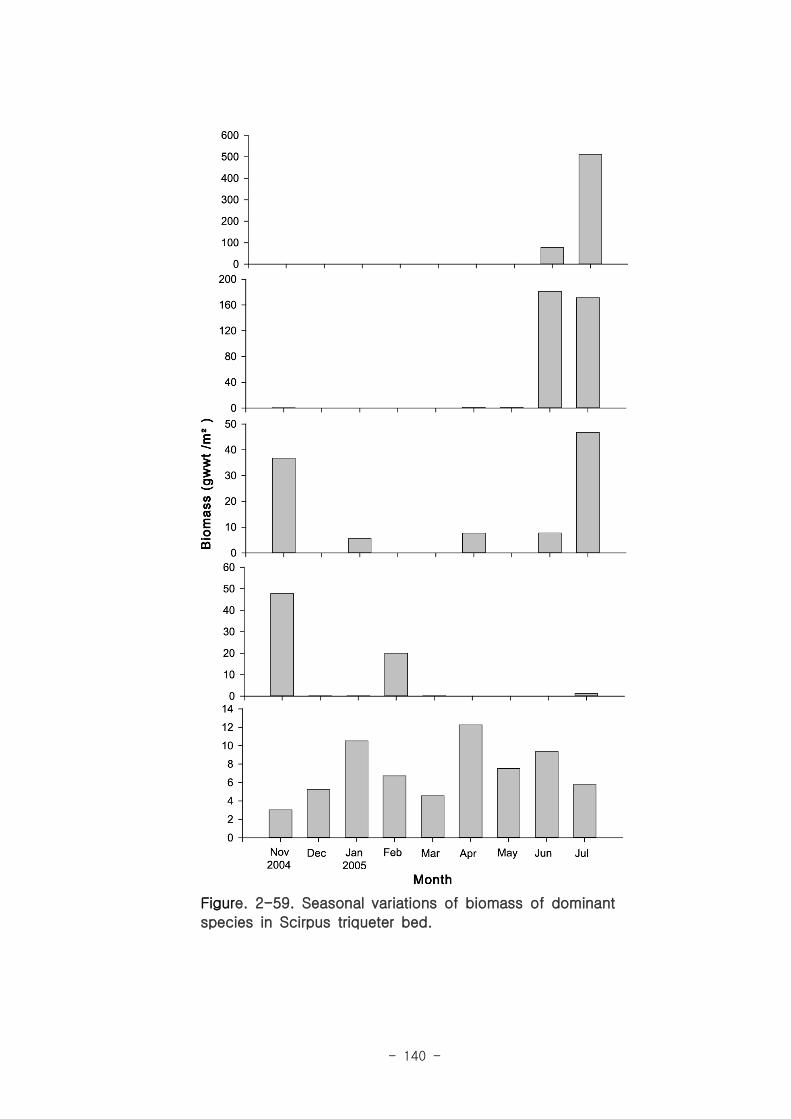
- 140 -
Figure. 2-59. Seasonal variations of biomass of dominant
species in Scirpus triqueter bed.
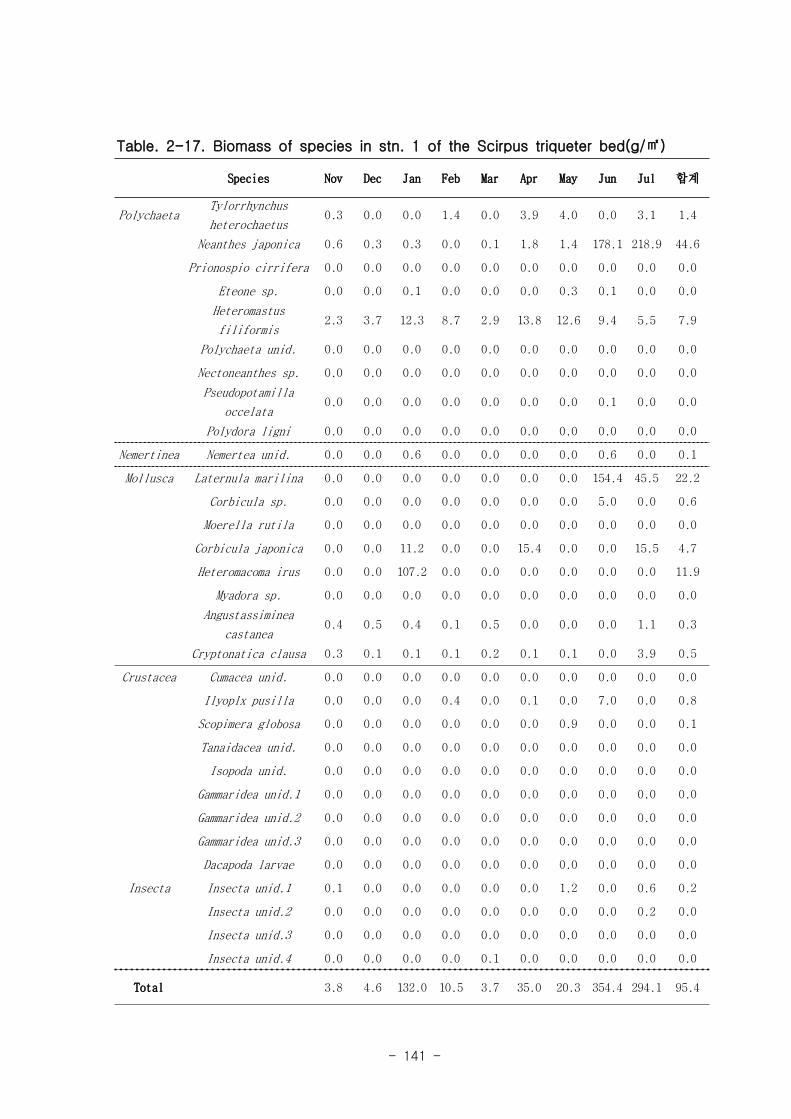
- 141 -
Table. 2-17. Biomass of species in stn. 1 of the Scirpus triqueter bed(g/㎡)
Species Nov Dec Jan Feb Mar Apr May Jun Jul 합계
PolychaetaTylorrhynchus
heterochaetus0.3 0.0 0.0 1.4 0.0 3.9 4.0 0.0 3.1 1.4
Neanthes japonica 0.6 0.3 0.3 0.0 0.1 1.8 1.4 178.1 218.9 44.6
Prionospio cirrifera 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Eteone sp. 0.0 0.0 0.1 0.0 0.0 0.0 0.3 0.1 0.0 0.0
Heteromastus
filiformis2.3 3.7 12.3 8.7 2.9 13.8 12.6 9.4 5.5 7.9
Polychaeta unid. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Nectoneanthes sp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Pseudopotamilla
occelata0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0
Polydora ligni 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Nemertinea Nemertea unid. 0.0 0.0 0.6 0.0 0.0 0.0 0.0 0.6 0.0 0.1
Mollusca Laternula marilina 0.0 0.0 0.0 0.0 0.0 0.0 0.0 154.4 45.5 22.2
Corbicula sp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 5.0 0.0 0.6
Moerella rutila 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Corbicula japonica 0.0 0.0 11.2 0.0 0.0 15.4 0.0 0.0 15.5 4.7
Heteromacoma irus 0.0 0.0 107.2 0.0 0.0 0.0 0.0 0.0 0.0 11.9
Myadora sp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Angustassiminea
castanea0.4 0.5 0.4 0.1 0.5 0.0 0.0 0.0 1.1 0.3
Cryptonatica clausa 0.3 0.1 0.1 0.1 0.2 0.1 0.1 0.0 3.9 0.5
Crustacea Cumacea unid. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Ilyoplx pusilla 0.0 0.0 0.0 0.4 0.0 0.1 0.0 7.0 0.0 0.8
Scopimera globosa 0.0 0.0 0.0 0.0 0.0 0.0 0.9 0.0 0.0 0.1
Tanaidacea unid. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Isopoda unid. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Gammaridea unid.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Gammaridea unid.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Gammaridea unid.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Dacapoda larvae 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Insecta Insecta unid.1 0.1 0.0 0.0 0.0 0.0 0.0 1.2 0.0 0.6 0.2
Insecta unid.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.2 0.0
Insecta unid.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Insecta unid.4 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0
Total 3.8 4.6 132.0 10.5 3.7 35.0 20.3 354.4 294.1 95.4
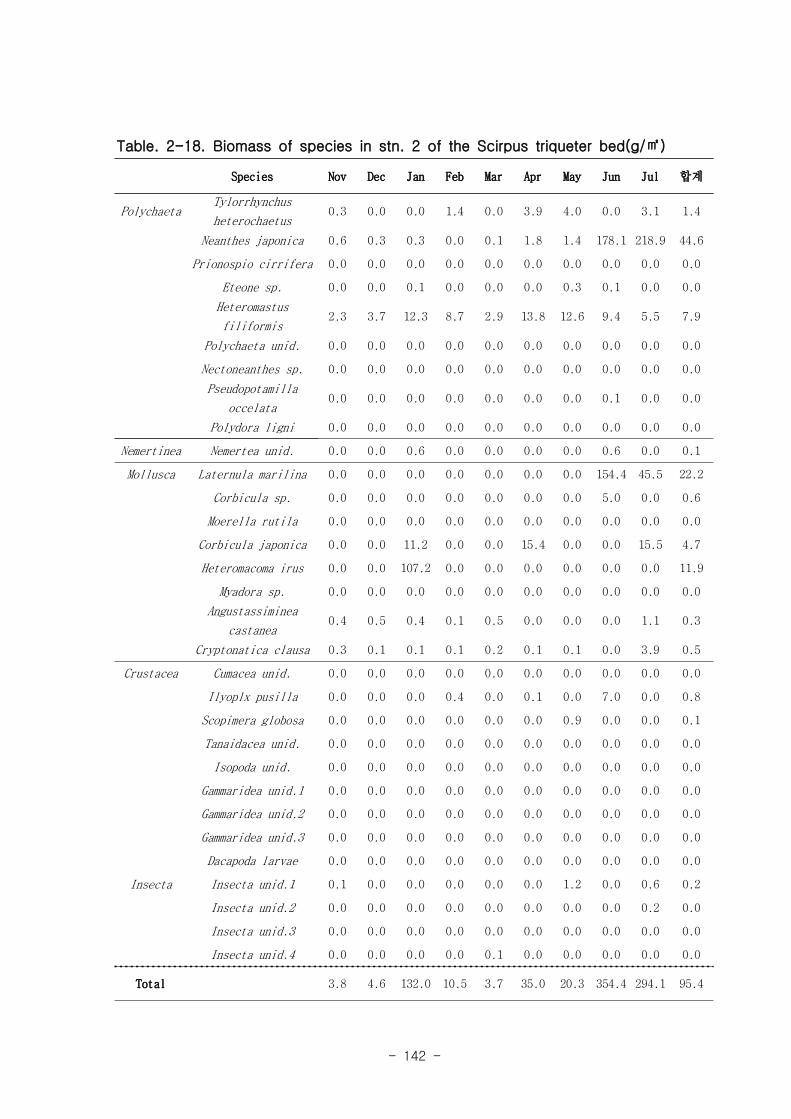
- 142 -
Table. 2-18. Biomass of species in stn. 2 of the Scirpus triqueter bed(g/㎡)
Species Nov Dec Jan Feb Mar Apr May Jun Jul 합계
PolychaetaTylorrhynchus
heterochaetus0.3 0.0 0.0 1.4 0.0 3.9 4.0 0.0 3.1 1.4
Neanthes japonica 0.6 0.3 0.3 0.0 0.1 1.8 1.4 178.1 218.9 44.6
Prionospio cirrifera 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Eteone sp. 0.0 0.0 0.1 0.0 0.0 0.0 0.3 0.1 0.0 0.0
Heteromastus
filiformis2.3 3.7 12.3 8.7 2.9 13.8 12.6 9.4 5.5 7.9
Polychaeta unid. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Nectoneanthes sp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Pseudopotamilla
occelata0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0
Polydora ligni 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Nemertinea Nemertea unid. 0.0 0.0 0.6 0.0 0.0 0.0 0.0 0.6 0.0 0.1
Mollusca Laternula marilina 0.0 0.0 0.0 0.0 0.0 0.0 0.0 154.4 45.5 22.2
Corbicula sp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 5.0 0.0 0.6
Moerella rutila 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Corbicula japonica 0.0 0.0 11.2 0.0 0.0 15.4 0.0 0.0 15.5 4.7
Heteromacoma irus 0.0 0.0 107.2 0.0 0.0 0.0 0.0 0.0 0.0 11.9
Myadora sp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Angustassiminea
castanea0.4 0.5 0.4 0.1 0.5 0.0 0.0 0.0 1.1 0.3
Cryptonatica clausa 0.3 0.1 0.1 0.1 0.2 0.1 0.1 0.0 3.9 0.5
Crustacea Cumacea unid. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Ilyoplx pusilla 0.0 0.0 0.0 0.4 0.0 0.1 0.0 7.0 0.0 0.8
Scopimera globosa 0.0 0.0 0.0 0.0 0.0 0.0 0.9 0.0 0.0 0.1
Tanaidacea unid. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Isopoda unid. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Gammaridea unid.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Gammaridea unid.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Gammaridea unid.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Dacapoda larvae 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Insecta Insecta unid.1 0.1 0.0 0.0 0.0 0.0 0.0 1.2 0.0 0.6 0.2
Insecta unid.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.2 0.0
Insecta unid.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Insecta unid.4 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0
Total 3.8 4.6 132.0 10.5 3.7 35.0 20.3 354.4 294.1 95.4
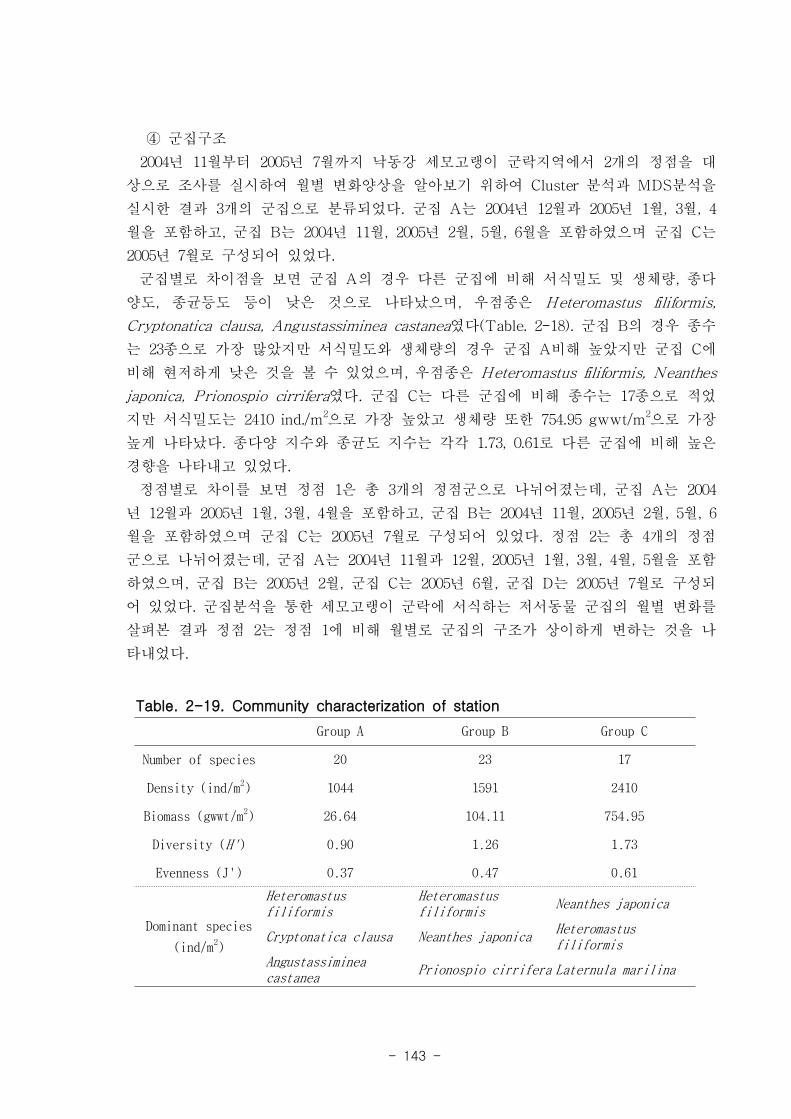
- 143 -
④ 군집구조
2004년 11월부터 2005년 7월까지 낙동강 세모고랭이 군락지역에서 2개의 정점을 대
상으로 조사를 실시하여 월별 변화양상을 알아보기 위하여 Cluster 분석과 MDS분석을
실시한 결과 3개의 군집으로 분류되었다. 군집 A는 2004년 12월과 2005년 1월, 3월, 4
월을 포함하고, 군집 B는 2004년 11월, 2005년 2월, 5월, 6월을 포함하였으며 군집 C는
2005년 7월로 구성되어 있었다.
군집별로 차이점을 보면 군집 A의 경우 다른 군집에 비해 서식밀도 및 생체량, 종다
양도, 종균등도 등이 낮은 것으로 나타났으며, 우점종은 Heteromastus filiformis,
Cryptonatica clausa, Angustassiminea castanea였다(Table. 2-18). 군집 B의 경우 종수
는 23종으로 가장 많았지만 서식밀도와 생체량의 경우 군집 A비해 높았지만 군집 C에
비해 현저하게 낮은 것을 볼 수 있었으며, 우점종은 Heteromastus filiformis, Neanthes
japonica, Prionospio cirrifera였다. 군집 C는 다른 군집에 비해 종수는 17종으로 적었
지만 서식밀도는 2410 ind./m2으로 가장 높았고 생체량 또한 754.95 gwwt/m
2으로 가장
높게 나타났다. 종다양 지수와 종균도 지수는 각각 1.73, 0.61로 다른 군집에 비해 높은
경향을 나타내고 있었다.
정점별로 차이를 보면 정점 1은 총 3개의 정점군으로 나뉘어졌는데, 군집 A는 2004
년 12월과 2005년 1월, 3월, 4월을 포함하고, 군집 B는 2004년 11월, 2005년 2월, 5월, 6
월을 포함하였으며 군집 C는 2005년 7월로 구성되어 있었다. 정점 2는 총 4개의 정점
군으로 나뉘어졌는데, 군집 A는 2004년 11월과 12월, 2005년 1월, 3월, 4월, 5월을 포함
하였으며, 군집 B는 2005년 2월, 군집 C는 2005년 6월, 군집 D는 2005년 7월로 구성되
어 있었다. 군집분석을 통한 세모고랭이 군락에 서식하는 저서동물 군집의 월별 변화를
살펴본 결과 정점 2는 정점 1에 비해 월별로 군집의 구조가 상이하게 변하는 것을 나
타내었다.
Table. 2-19. Community characterization of station
Group A Group B Group C
Number of species 20 23 17
Density (ind/m2) 1044 1591 2410
Biomass (gwwt/m2) 26.64 104.11 754.95
Diversity (H') 0.90 1.26 1.73
Evenness (J') 0.37 0.47 0.61
Dominant species
(ind/m2)
Heteromastus filiformis
Heteromastus filiformis
Neanthes japonica
Cryptonatica clausa Neanthes japonicaHeteromastus filiformis
Angustassiminea castanea
Prionospio cirrifera Laternula marilina
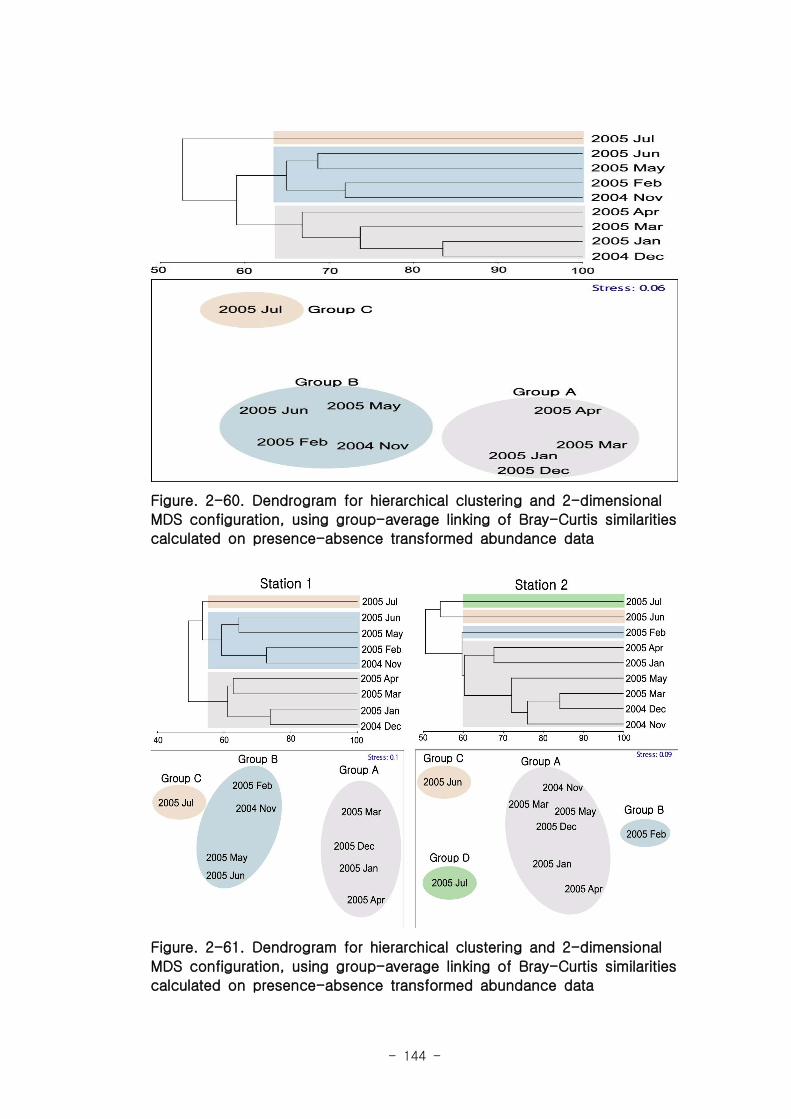
- 144 -
Figure. 2-60. Dendrogram for hierarchical clustering and 2-dimensional
MDS configuration, using group-average linking of Bray-Curtis similarities
calculated on presence-absence transformed abundance data
Figure. 2-61. Dendrogram for hierarchical clustering and 2-dimensional
MDS configuration, using group-average linking of Bray-Curtis similarities
calculated on presence-absence transformed abundance data
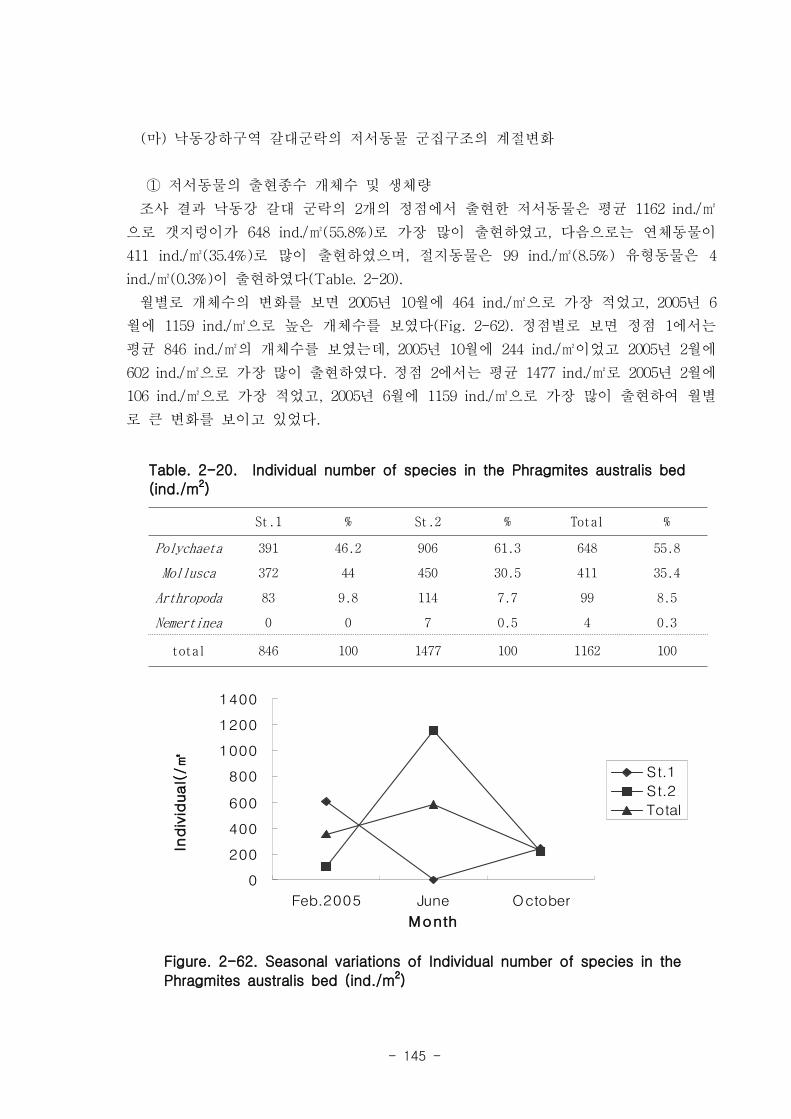
- 145 -
(마) 낙동강하구역 갈대군락의 저서동물 군집구조의 계절변화
① 저서동물의 출현종수 개체수 및 생체량
조사 결과 낙동강 갈대 군락의 2개의 정점에서 출현한 저서동물은 평균 1162 ind./㎡
으로 갯지렁이가 648 ind./㎡(55.8%)로 가장 많이 출현하였고, 다음으로는 연체동물이
411 ind./㎡(35.4%)로 많이 출현하였으며, 절지동물은 99 ind./㎡(8.5%) 유형동물은 4
ind./㎡(0.3%)이 출현하였다(Table. 2-20).
월별로 개체수의 변화를 보면 2005년 10월에 464 ind./㎡으로 가장 적었고, 2005년 6
월에 1159 ind./㎡으로 높은 개체수를 보였다(Fig. 2-62). 정점별로 보면 정점 1에서는
평균 846 ind./㎡의 개체수를 보였는데, 2005년 10월에 244 ind./㎡이었고 2005년 2월에
602 ind./㎡으로 가장 많이 출현하였다. 정점 2에서는 평균 1477 ind./㎡로 2005년 2월에
106 ind./㎡으로 가장 적었고, 2005년 6월에 1159 ind./㎡으로 가장 많이 출현하여 월별
로 큰 변화를 보이고 있었다.
St.1 % St.2 % Total %
Polychaeta 391 46.2 906 61.3 648 55.8
Mollusca 372 44 450 30.5 411 35.4
Arthropoda 83 9.8 114 7.7 99 8.5
Nemertinea 0 0 7 0.5 4 0.3
total 846 100 1477 100 1162 100
Table. 2-20. Individual number of species in the Phragmites australis bed
(ind./m2)
0
200
400
600
800
1000
1200
1400
Feb.2005 June October
Month
Indiv
idual(
/㎡)
St.1
St.2
Total
Figure. 2-62. Seasonal variations of Individual number of species in the
Phragmites australis bed (ind./m2)
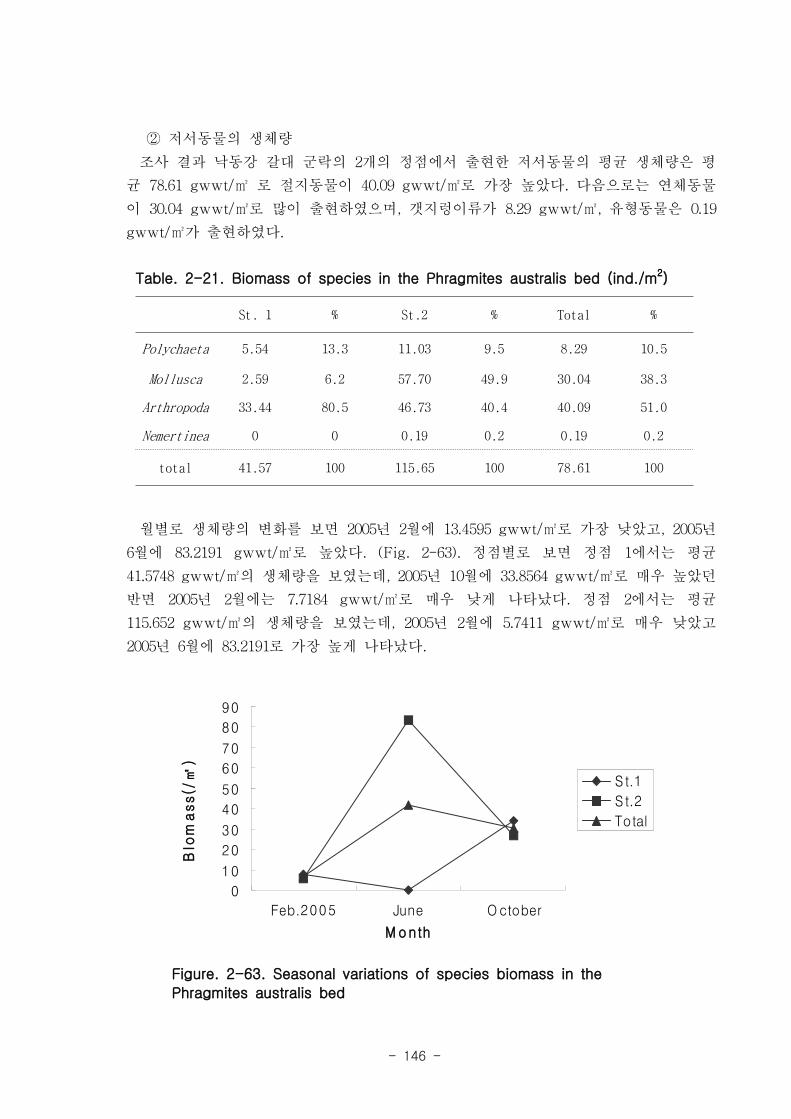
- 146 -
② 저서동물의 생체량
조사 결과 낙동강 갈대 군락의 2개의 정점에서 출현한 저서동물의 평균 생체량은 평
균 78.61 gwwt/㎡ 로 절지동물이 40.09 gwwt/㎡로 가장 높았다. 다음으로는 연체동물
이 30.04 gwwt/㎡로 많이 출현하였으며, 갯지렁이류가 8.29 gwwt/㎡, 유형동물은 0.19
gwwt/㎡가 출현하였다.
St. 1 % St.2 % Total %
Polychaeta 5.54 13.3 11.03 9.5 8.29 10.5
Mollusca 2.59 6.2 57.70 49.9 30.04 38.3
Arthropoda 33.44 80.5 46.73 40.4 40.09 51.0
Nemertinea 0 0 0.19 0.2 0.19 0.2
total 41.57 100 115.65 100 78.61 100
Table. 2-21. Biomass of species in the Phragmites australis bed (ind./m2)
월별로 생체량의 변화를 보면 2005년 2월에 13.4595 gwwt/㎡로 가장 낮았고, 2005년
6월에 83.2191 gwwt/㎡로 높았다. (Fig. 2-63). 정점별로 보면 정점 1에서는 평균
41.5748 gwwt/㎡의 생체량을 보였는데, 2005년 10월에 33.8564 gwwt/㎡로 매우 높았던
반면 2005년 2월에는 7.7184 gwwt/㎡로 매우 낮게 나타났다. 정점 2에서는 평균
115.652 gwwt/㎡의 생체량을 보였는데, 2005년 2월에 5.7411 gwwt/㎡로 매우 낮았고
2005년 6월에 83.2191로 가장 높게 나타났다.
0
10
20
30
40
50
60
70
80
90
Feb.2005 June O ctober
M onth
BIo
ma
ss(/
㎡)
S t.1
St.2
Total
Figure. 2-63. Seasonal variations of species biomass in the
Phragmites australis bed

- 147 -
(바) 진우도 갯벌 저서동물의 군집구조
① 저서동물의 출현종수 개체수
조사 결과 진우도 Bare tide 군락의 2개의 정점에서 출현한 저서동물은 평균 410
ind./㎡으로 갯지렁이가 279 ind./㎡(68%)로 가장 많이 출현하였고, 다음으로는 연체동
물이 84 ind./㎡(20.5%)로 많이 출현하였으며, 절지동물은 47 ind./㎡(11.5%) 유형동물은
1 ind./㎡(0%)이 출현하였다. (Table. 2-22).
월별로 개체수의 변화를 보면 2005년 10월에 38 ind./㎡으로 가장 적었고, 2005년 2월
에 249 ind./㎡으로 높은 개체수를 보였다(Fig. 2-64). 정점별로 보면 정점 1에서는 평
균 391 ind./㎡의 개체수를 보였는데, 2005년 10월에 34 ind./㎡로 가장 적었고 2005년 2
월에 261 ind./㎡로 가장 많이 출현하였다. 정점 2에서는 평균 428 ind./㎡로 정점 1에
비해 많은 개체수를 보였는데 2005년 10월에 41 ind./㎡로 가장 적었고, 2005년 2월에
236 ind./㎡로 가장 많이 출현하여 월별로 큰 변화를 보이고 있었다.
St. 1 % St. 2 % Total %
Polychaeta 273 69.8 284 66.4 279 68
Mollusca 76 19.4 92 21.5 84 20.5
Arthropoda 41 10.5 52 12.1 47 11.5
Nemertinea 1 0.3 0 0 0 0
total 391 100 428 100 410 100
Table. 2-22. Individual number of species in the bare tidal flat (ind./m2)
0
50
100
150
200
250
300
Feb.2005 M ay October
M on th
Ind
ivid
ua
l(/㎡
)
St.1
St.2
Total
Figure. 2-64. Seasonal variation of individual number of species in the
bare tidal flat
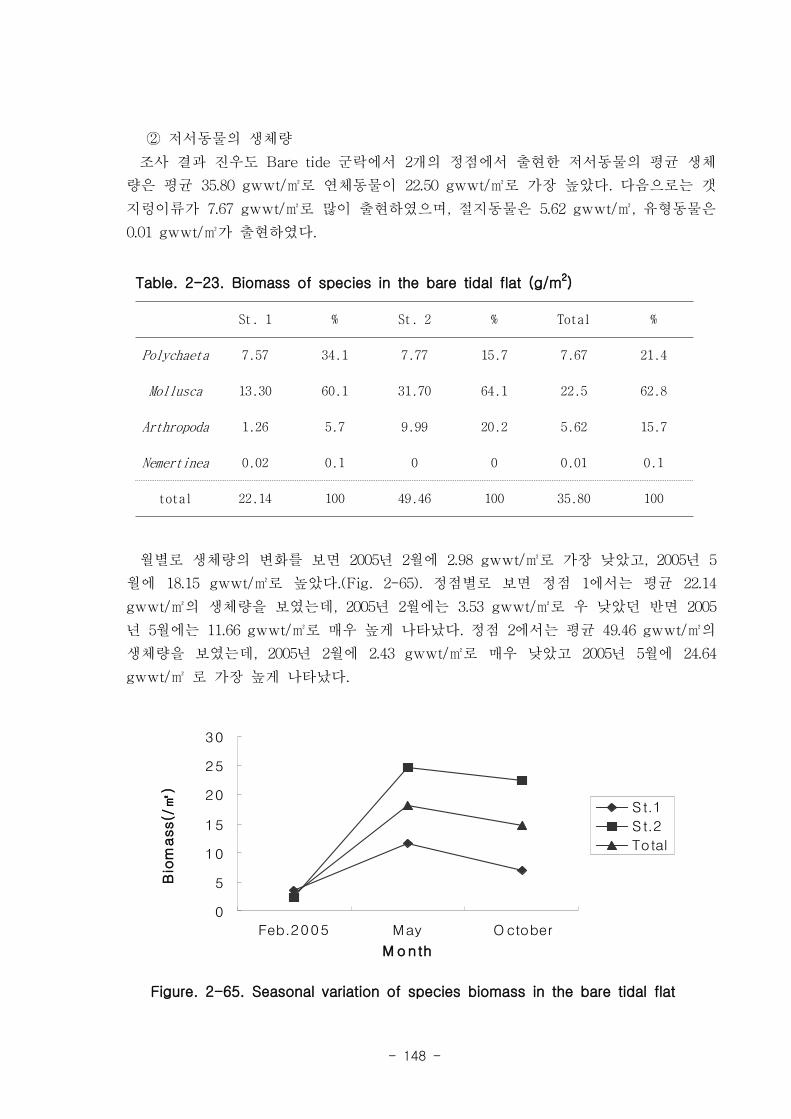
- 148 -
② 저서동물의 생체량
조사 결과 진우도 Bare tide 군락에서 2개의 정점에서 출현한 저서동물의 평균 생체
량은 평균 35.80 gwwt/㎡로 연체동물이 22.50 gwwt/㎡로 가장 높았다. 다음으로는 갯
지렁이류가 7.67 gwwt/㎡로 많이 출현하였으며, 절지동물은 5.62 gwwt/㎡, 유형동물은
0.01 gwwt/㎡가 출현하였다.
St. 1 % St. 2 % Total %
Polychaeta 7.57 34.1 7.77 15.7 7.67 21.4
Mollusca 13.30 60.1 31.70 64.1 22.5 62.8
Arthropoda 1.26 5.7 9.99 20.2 5.62 15.7
Nemertinea 0.02 0.1 0 0 0.01 0.1
total 22.14 100 49.46 100 35.80 100
Table. 2-23. Biomass of species in the bare tidal flat (g/m2)
월별로 생체량의 변화를 보면 2005년 2월에 2.98 gwwt/㎡로 가장 낮았고, 2005년 5
월에 18.15 gwwt/㎡로 높았다.(Fig. 2-65). 정점별로 보면 정점 1에서는 평균 22.14
gwwt/㎡의 생체량을 보였는데, 2005년 2월에는 3.53 gwwt/㎡로 우 낮았던 반면 2005
년 5월에는 11.66 gwwt/㎡로 매우 높게 나타났다. 정점 2에서는 평균 49.46 gwwt/㎡의
생체량을 보였는데, 2005년 2월에 2.43 gwwt/㎡로 매우 낮았고 2005년 5월에 24.64
gwwt/㎡ 로 가장 높게 나타났다.
0
5
10
15
20
25
30
Feb.2005 May O ctober
M onth
Bio
mass(/
㎡)
S t.1
St.2
Total
Figure. 2-65. Seasonal variation of species biomass in the bare tidal flat
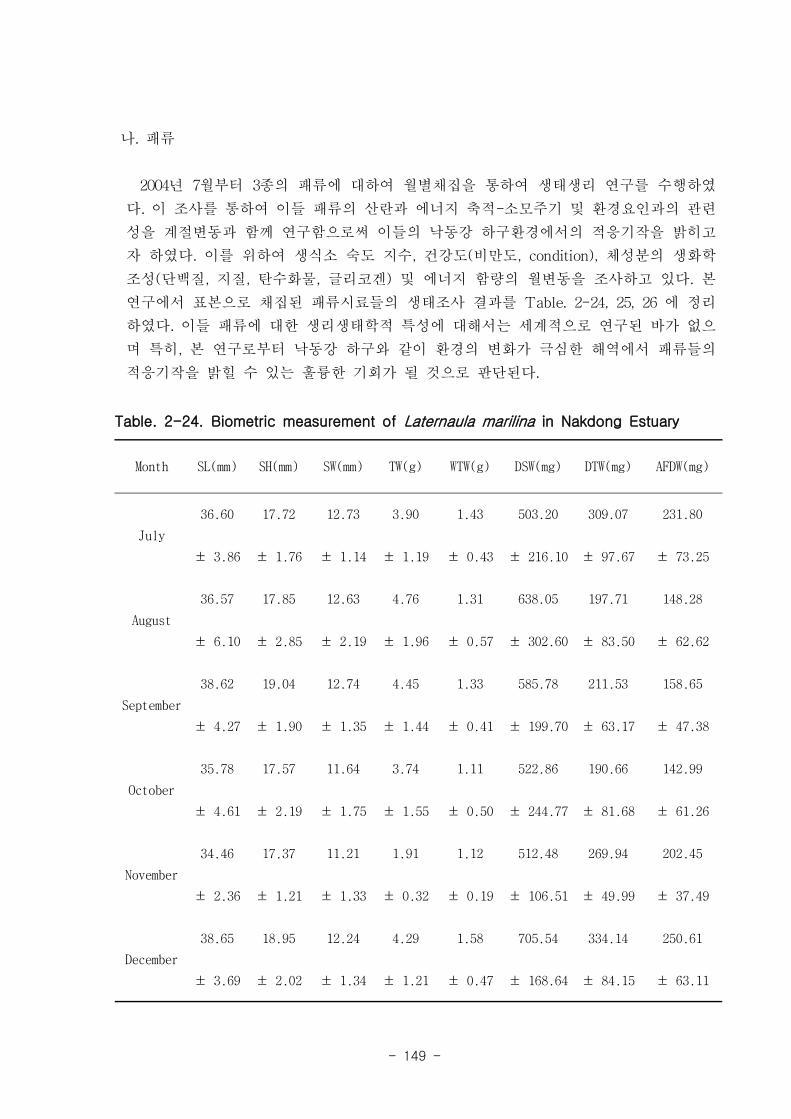
- 149 -
나. 패류
2004년 7월부터 3종의 패류에 대하여 월별채집을 통하여 생태생리 연구를 수행하였
다. 이 조사를 통하여 이들 패류의 산란과 에너지 축적-소모주기 및 환경요인과의 관련
성을 계절변동과 함께 연구함으로써 이들의 낙동강 하구환경에서의 적응기작을 밝히고
자 하였다. 이를 위하여 생식소 숙도 지수, 건강도(비만도, condition), 체성분의 생화학
조성(단백질, 지질, 탄수화물, 글리코겐) 및 에너지 함량의 월변동을 조사하고 있다. 본
연구에서 표본으로 채집된 패류시료들의 생태조사 결과를 Table. 2-24, 25, 26 에 정리
하였다. 이들 패류에 대한 생리생태학적 특성에 대해서는 세계적으로 연구된 바가 없으
며 특히, 본 연구로부터 낙동강 하구와 같이 환경의 변화가 극심한 해역에서 패류들의
적응기작을 밝힐 수 있는 훌륭한 기회가 될 것으로 판단된다.
Month SL(mm) SH(mm) SW(mm) TW(g) WTW(g) DSW(mg) DTW(mg) AFDW(mg)
July
36.60 17.72 12.73 3.90 1.43 503.20 309.07 231.80
± 3.86 ± 1.76 ± 1.14 ± 1.19 ± 0.43 ± 216.10 ± 97.67 ± 73.25
August
36.57 17.85 12.63 4.76 1.31 638.05 197.71 148.28
± 6.10 ± 2.85 ± 2.19 ± 1.96 ± 0.57 ± 302.60 ± 83.50 ± 62.62
September
38.62 19.04 12.74 4.45 1.33 585.78 211.53 158.65
± 4.27 ± 1.90 ± 1.35 ± 1.44 ± 0.41 ± 199.70 ± 63.17 ± 47.38
October
35.78 17.57 11.64 3.74 1.11 522.86 190.66 142.99
± 4.61 ± 2.19 ± 1.75 ± 1.55 ± 0.50 ± 244.77 ± 81.68 ± 61.26
November
34.46 17.37 11.21 1.91 1.12 512.48 269.94 202.45
± 2.36 ± 1.21 ± 1.33 ± 0.32 ± 0.19 ± 106.51 ± 49.99 ± 37.49
December
38.65 18.95 12.24 4.29 1.58 705.54 334.14 250.61
± 3.69 ± 2.02 ± 1.34 ± 1.21 ± 0.47 ± 168.64 ± 84.15 ± 63.11
Table. 2-24. Biometric measurement of Laternaula marilina in Nakdong Estuary
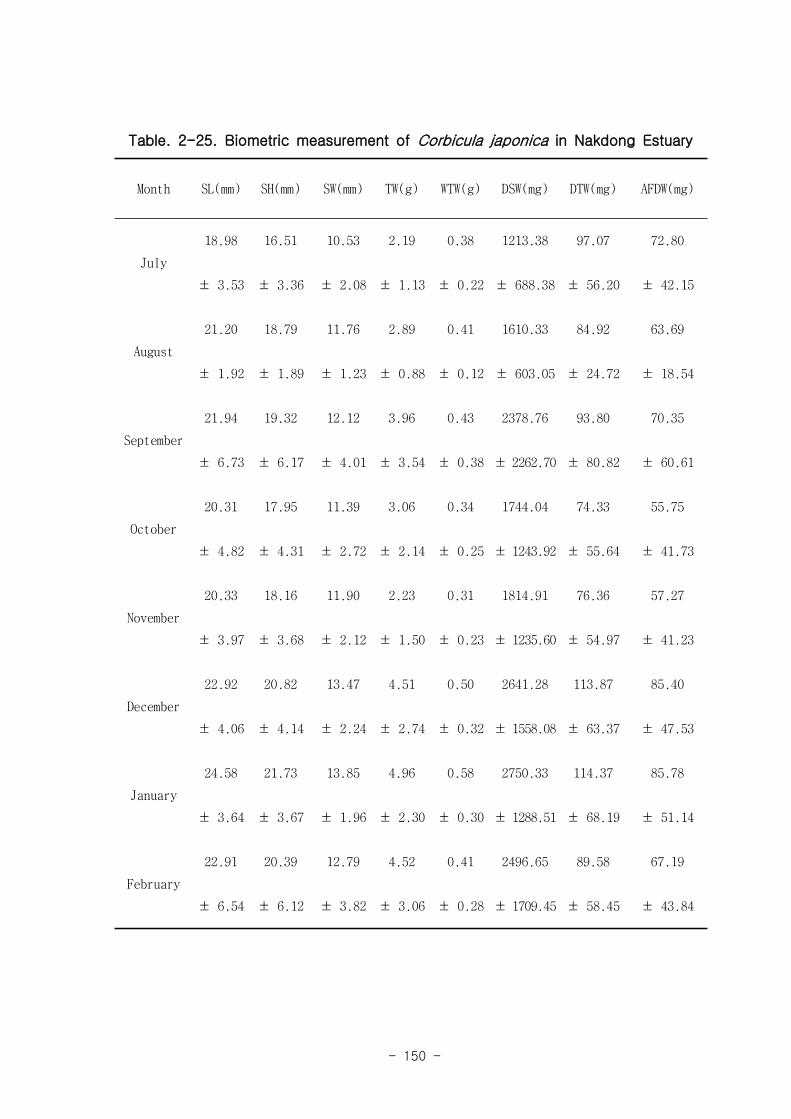
- 150 -
Month SL(mm) SH(mm) SW(mm) TW(g) WTW(g) DSW(mg) DTW(mg) AFDW(mg)
July
18.98 16.51 10.53 2.19 0.38 1213.38 97.07 72.80
± 3.53 ± 3.36 ± 2.08 ± 1.13 ± 0.22 ± 688.38 ± 56.20 ± 42.15
August
21.20 18.79 11.76 2.89 0.41 1610.33 84.92 63.69
± 1.92 ± 1.89 ± 1.23 ± 0.88 ± 0.12 ± 603.05 ± 24.72 ± 18.54
September
21.94 19.32 12.12 3.96 0.43 2378.76 93.80 70.35
± 6.73 ± 6.17 ± 4.01 ± 3.54 ± 0.38 ± 2262.70 ± 80.82 ± 60.61
October
20.31 17.95 11.39 3.06 0.34 1744.04 74.33 55.75
± 4.82 ± 4.31 ± 2.72 ± 2.14 ± 0.25 ± 1243.92 ± 55.64 ± 41.73
November
20.33 18.16 11.90 2.23 0.31 1814.91 76.36 57.27
± 3.97 ± 3.68 ± 2.12 ± 1.50 ± 0.23 ± 1235.60 ± 54.97 ± 41.23
December
22.92 20.82 13.47 4.51 0.50 2641.28 113.87 85.40
± 4.06 ± 4.14 ± 2.24 ± 2.74 ± 0.32 ± 1558.08 ± 63.37 ± 47.53
January
24.58 21.73 13.85 4.96 0.58 2750.33 114.37 85.78
± 3.64 ± 3.67 ± 1.96 ± 2.30 ± 0.30 ± 1288.51 ± 68.19 ± 51.14
February
22.91 20.39 12.79 4.52 0.41 2496.65 89.58 67.19
± 6.54 ± 6.12 ± 3.82 ± 3.06 ± 0.28 ± 1709.45 ± 58.45 ± 43.84
Table. 2-25. Biometric measurement of Corbicula japonica in Nakdong Estuary
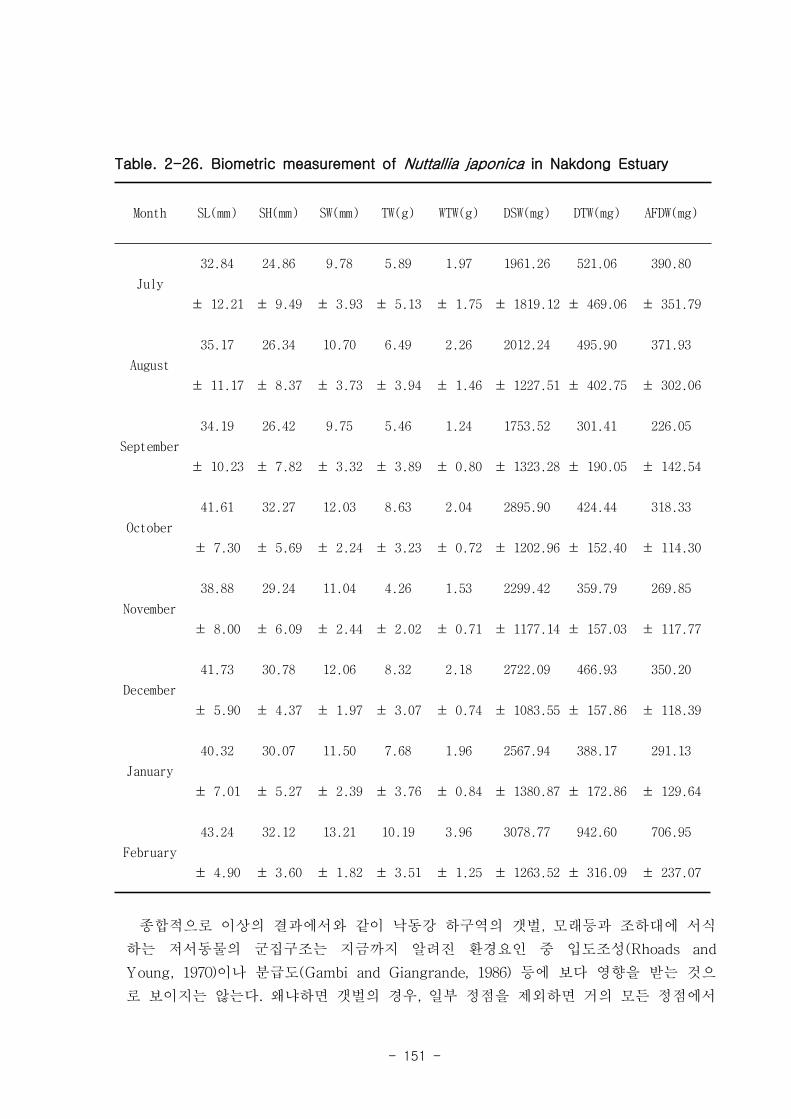
- 151 -
Month SL(mm) SH(mm) SW(mm) TW(g) WTW(g) DSW(mg) DTW(mg) AFDW(mg)
July
32.84 24.86 9.78 5.89 1.97 1961.26 521.06 390.80
± 12.21 ± 9.49 ± 3.93 ± 5.13 ± 1.75 ± 1819.12 ± 469.06 ± 351.79
August
35.17 26.34 10.70 6.49 2.26 2012.24 495.90 371.93
± 11.17 ± 8.37 ± 3.73 ± 3.94 ± 1.46 ± 1227.51 ± 402.75 ± 302.06
September
34.19 26.42 9.75 5.46 1.24 1753.52 301.41 226.05
± 10.23 ± 7.82 ± 3.32 ± 3.89 ± 0.80 ± 1323.28 ± 190.05 ± 142.54
October
41.61 32.27 12.03 8.63 2.04 2895.90 424.44 318.33
± 7.30 ± 5.69 ± 2.24 ± 3.23 ± 0.72 ± 1202.96 ± 152.40 ± 114.30
November
38.88 29.24 11.04 4.26 1.53 2299.42 359.79 269.85
± 8.00 ± 6.09 ± 2.44 ± 2.02 ± 0.71 ± 1177.14 ± 157.03 ± 117.77
December
41.73 30.78 12.06 8.32 2.18 2722.09 466.93 350.20
± 5.90 ± 4.37 ± 1.97 ± 3.07 ± 0.74 ± 1083.55 ± 157.86 ± 118.39
January
40.32 30.07 11.50 7.68 1.96 2567.94 388.17 291.13
± 7.01 ± 5.27 ± 2.39 ± 3.76 ± 0.84 ± 1380.87 ± 172.86 ± 129.64
February
43.24 32.12 13.21 10.19 3.96 3078.77 942.60 706.95
± 4.90 ± 3.60 ± 1.82 ± 3.51 ± 1.25 ± 1263.52 ± 316.09 ± 237.07
Table. 2-26. Biometric measurement of Nuttallia japonica in Nakdong Estuary
종합적으로 이상의 결과에서와 같이 낙동강 하구역의 갯벌, 모래등과 조하대에 서식
하는 저서동물의 군집구조는 지금까지 알려진 환경요인 중 입도조성(Rhoads and
Young, 1970)이나 분급도(Gambi and Giangrande, 1986) 등에 보다 영향을 받는 것으
로 보이지는 않는다. 왜냐하면 갯벌의 경우, 일부 정점을 제외하면 거의 모든 정점에서

- 152 -
퇴적물의 모래함량이 높고, 퇴적물의 노출시간을 결정하는 조위도 많은 지역에서 유사
하게 나타나고 있기 때문이다. 한편, 우리나라 서해안으로 열린 한강과 금강 하구역의
저서동물 군집은 일차적으로는 염분도 구배에 의해, 또한 국소적으로는 퇴적상에 영향
을 받는 것으로 파악되었다(Hong and Yoo, 1996; Choi and Koh, 1994). 그러나 낙동강
하구역은 하구언 건설로 인해 저서동물 뿐만 아니라 저서환경에 있어서도 하구언을 중
심으로 상부와 하부가 격리 또는 차단되어 있고(부산광역시, 2003), 을숙도에서 외양역
에 이르는 해역에서도 기수역 정점군과 해양역 정점군으로 구별되는 특징을 보이고 있
다. 따라서, 하구역의 특징 중의 하나인 퇴적상과 염분농도에 따른 생물상의 분포결정
보다는 홍수기의 일시적인 담수유입, 그에 따른 퇴적율의 변화, 또는 계절에 따른 포식
생물(조류나 유영성 어류)의 밀도 등 다각적인 요인에 의해 영향을 받는 것으로 추정할
수 있다. 이 밖에 하구언 건설 이후, 가장 주목할만한 군집의 변화 중의 하나는 오염지
표종의 출현에 있다. 본 조사에서 출현하고 있는 다모류의 H. filiformis, Capitella
capitata, P. pinnata, P. kempi등은 유기물 오염에 대한 지표종으로서 전 세계적으로
잘 알려져 있다(Pearson and Rosenberg, 1978). 본 조사에서 이들 종들은 주로 하구역
전반에 결쳐 출현하는 H. filiformis 제외하면 외양역에서 주로 밀도가 높은 특징이 있
다. 특히 이 가운데 H. filiformis는 유기물 오염이 상대적으로 심한 우리나라의 만이나
갯벌에서 높은 밀도로 출현하고 있으며, 본 조사에서도 8,142개체가 출현하여 관심이
요구된다. 한편, 현장조사 시의 관찰에 의하면, 명지포구 일원해역의 정점군(정점4, 6~
8)은 저질이 검은색을 띠고 있으며, 황화수소(H2S)의 냄새가 심하였다. 특히, 정점 7에
서는 단 2종만이 출현하였고, 출현종 역시 생물학적 부패의 정도가 심한 상태로 채집되
는 오염역(polluted area)의 특징을 보이고 있다. 따라서, 저질토의 적극적인 개선이 없
는 한 무생물구역으로의 천이가 발생할 수 있을 것으로 예상된다. 결국, 생태계 보전
측면에서 향후 저서동물 군집구조, 생물분포와 다양성에 영향을 미치는 인자 뿐만 아니
라 이들 오염지표종의 동태에 대한 보다 세밀한 모니터링이 요구된다.
다. 어류
명지지역에서 채집된 어류는 9목 27과 34속 38종 3,536마리였다(양 등 2001)(Table.
2-27). 이를 월별로 보면 6월에 25종 1,005개체가 채집되었고 이 중 T. rubrifes 가 780
개체로 가장 많았다. 8월에는 20종 1,520개체가 채집되었고, 10월에는 14종 675개체, 11
월에는 13종 336 개체가 각각 채집되었는데, 이 3개월 동안에는 A.flavimanus 가 각각
738, 535, 116 개체로 그 수가 가장 많았다.
사상지역에서는 7목 9과 18속 20종 3,100개체가 채집되었다. 6월에는 12종 490개체가
채집, 8월에는 9종 134개체가 채집되었고 이 두 달에는 H. labeo 가 각각 262, 102개체
로 가장 많았다. 10월에는 8종 1,044개체가 어획되었고 S. chankaensis tsuchigae가 571
개체로 가장많았다. 11월에는 8종 1,432개체 중 A.macropterus 의 개체수가 가장 많았
다.
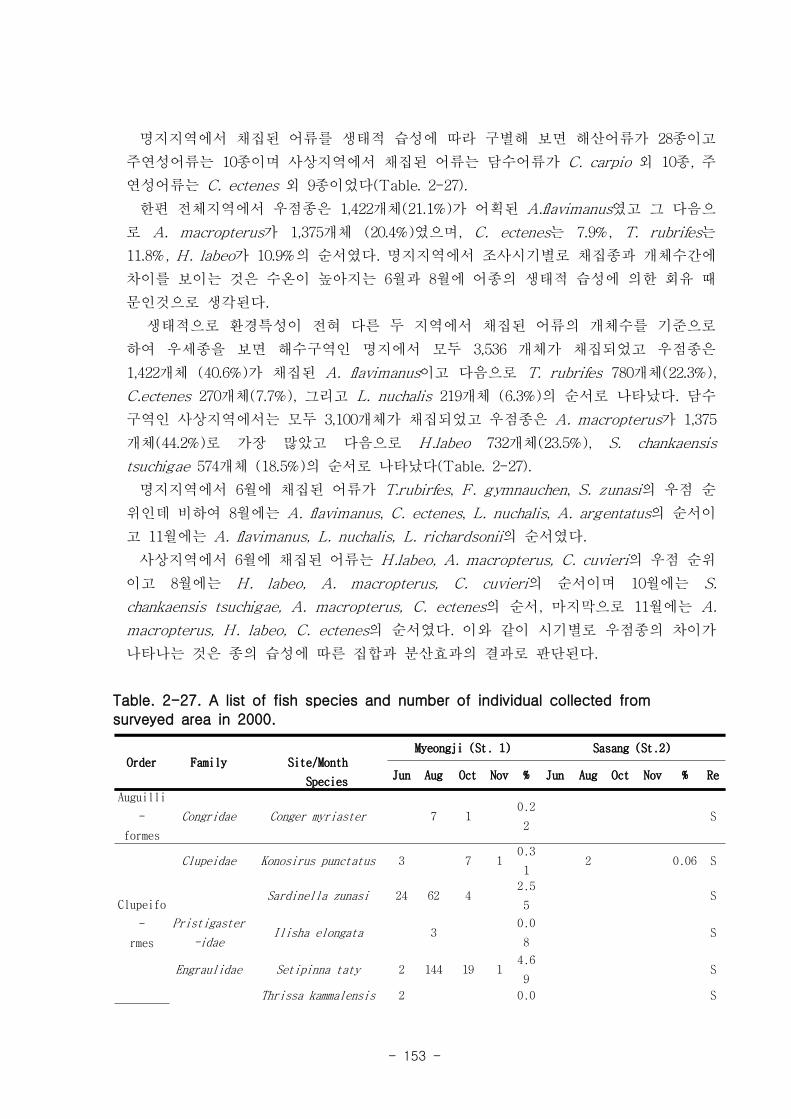
- 153 -
Order Family
Site/Month
Species
Myeongji (St. 1) Sasang (St.2)
Jun Aug Oct Nov % Jun Aug Oct Nov % Re
Auguilli
-
formes
Congridae Conger myriaster 7 10.2
2S
Clupeifo
-
rmes
Clupeidae Konosirus punctatus 3 7 10.3
12 0.06 S
Sardinella zunasi 24 62 42.5
5S
Pristigaster
-idaeIlisha elongata 3
0.0
8S
Engraulidae Setipinna taty 2 144 19 14.6
9S
Thrissa kammalensis 2 0.0 S
명지지역에서 채집된 어류를 생태적 습성에 따라 구별해 보면 해산어류가 28종이고
주연성어류는 10종이며 사상지역에서 채집된 어류는 담수어류가 C. carpio 외 10종, 주
연성어류는 C. ectenes 외 9종이었다(Table. 2-27).
한편 전체지역에서 우점종은 1,422개체(21.1%)가 어획된 A.flavimanus였고 그 다음으
로 A. macropterus가 1,375개체 (20.4%)였으며, C. ectenes는 7.9%, T. rubrifes는
11.8%, H. labeo가 10.9%의 순서였다. 명지지역에서 조사시기별로 채집종과 개체수간에
차이를 보이는 것은 수온이 높아지는 6월과 8월에 어종의 생태적 습성에 의한 회유 때
문인것으로 생각된다.
생태적으로 환경특성이 전혀 다른 두 지역에서 채집된 어류의 개체수를 기준으로
하여 우세종을 보면 해수구역인 명지에서 모두 3,536 개체가 채집되었고 우점종은
1,422개체 (40.6%)가 채집된 A. flavimanus이고 다음으로 T. rubrifes 780개체(22.3%),
C.ectenes 270개체(7.7%), 그리고 L. nuchalis 219개체 (6.3%)의 순서로 나타났다. 담수
구역인 사상지역에서는 모두 3,100개체가 채집되었고 우점종은 A. macropterus가 1,375
개체(44.2%)로 가장 많았고 다음으로 H.labeo 732개체(23.5%), S. chankaensis
tsuchigae 574개체 (18.5%)의 순서로 나타났다(Table. 2-27).
명지지역에서 6월에 채집된 어류가 T.rubirfes, F. gymnauchen, S. zunasi의 우점 순
위인데 비하여 8월에는 A. flavimanus, C. ectenes, L. nuchalis, A. argentatus의 순서이
고 11월에는 A. flavimanus, L. nuchalis, L. richardsonii의 순서였다.
사상지역에서 6월에 채집된 어류는 H.labeo, A. macropterus, C. cuvieri의 우점 순위
이고 8월에는 H. labeo, A. macropterus, C. cuvieri의 순서이며 10월에는 S.
chankaensis tsuchigae, A. macropterus, C. ectenes의 순서, 마지막으로 11월에는 A.
macropterus, H. labeo, C. ectenes의 순서였다. 이와 같이 시기별로 우점종의 차이가
나타나는 것은 종의 습성에 따른 집합과 분산효과의 결과로 판단된다.
Table. 2-27. A list of fish species and number of individual collected from
surveyed area in 2000.
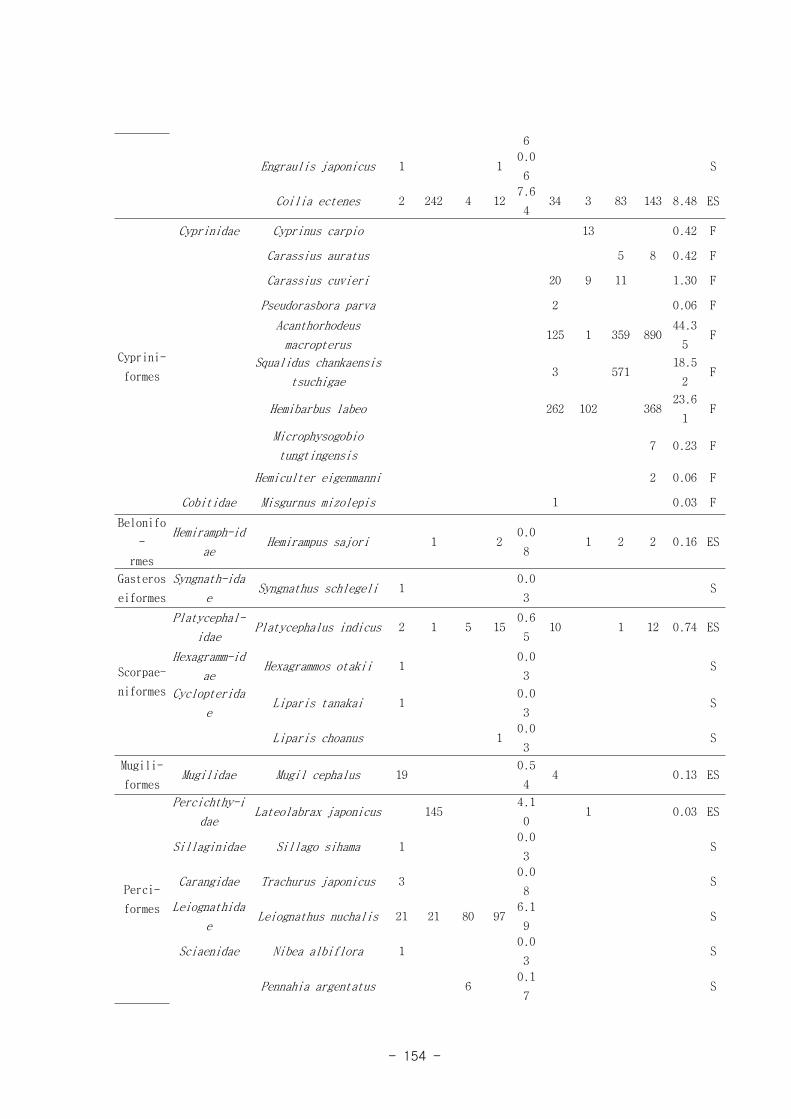
- 154 -
6
Engraulis japonicus 1 10.0
6S
Coilia ectenes 2 242 4 127.6
434 3 83 143 8.48 ES
Cyprini-
formes
Cyprinidae Cyprinus carpio 13 0.42 F
Carassius auratus 5 8 0.42 F
Carassius cuvieri 20 9 11 1.30 F
Pseudorasbora parva 2 0.06 F
Acanthorhodeus
macropterus125 1 359 890
44.3
5F
Squalidus chankaensis
tsuchigae3 571
18.5
2F
Hemibarbus labeo 262 102 36823.6
1F
Microphysogobio
tungtingensis7 0.23 F
Hemiculter eigenmanni 2 0.06 F
Cobitidae Misgurnus mizolepis 1 0.03 F
Belonifo
-
rmes
Hemiramph-id
aeHemirampus sajori 1 2
0.0
81 2 2 0.16 ES
Gasteros
eiformes
Syngnath-ida
eSyngnathus schlegeli 1
0.0
3S
Scorpae-
niformes
Platycephal-
idaePlatycephalus indicus 2 1 5 15
0.6
510 1 12 0.74 ES
Hexagramm-id
aeHexagrammos otakii 1
0.0
3S
Cyclopterida
eLiparis tanakai 1
0.0
3S
Liparis choanus 10.0
3S
Mugili-
formesMugilidae Mugil cephalus 19
0.5
44 0.13 ES
Perci-
formes
Percichthy-i
daeLateolabrax japonicus 145
4.1
01 0.03 ES
Sillaginidae Sillago sihama 10.0
3S
Carangidae Trachurus japonicus 30.0
8S
Leiognathida
eLeiognathus nuchalis 21 21 80 97
6.1
9S
Sciaenidae Nibea albiflora 10.0
3S
Pennahia argentatus 60.1
7S
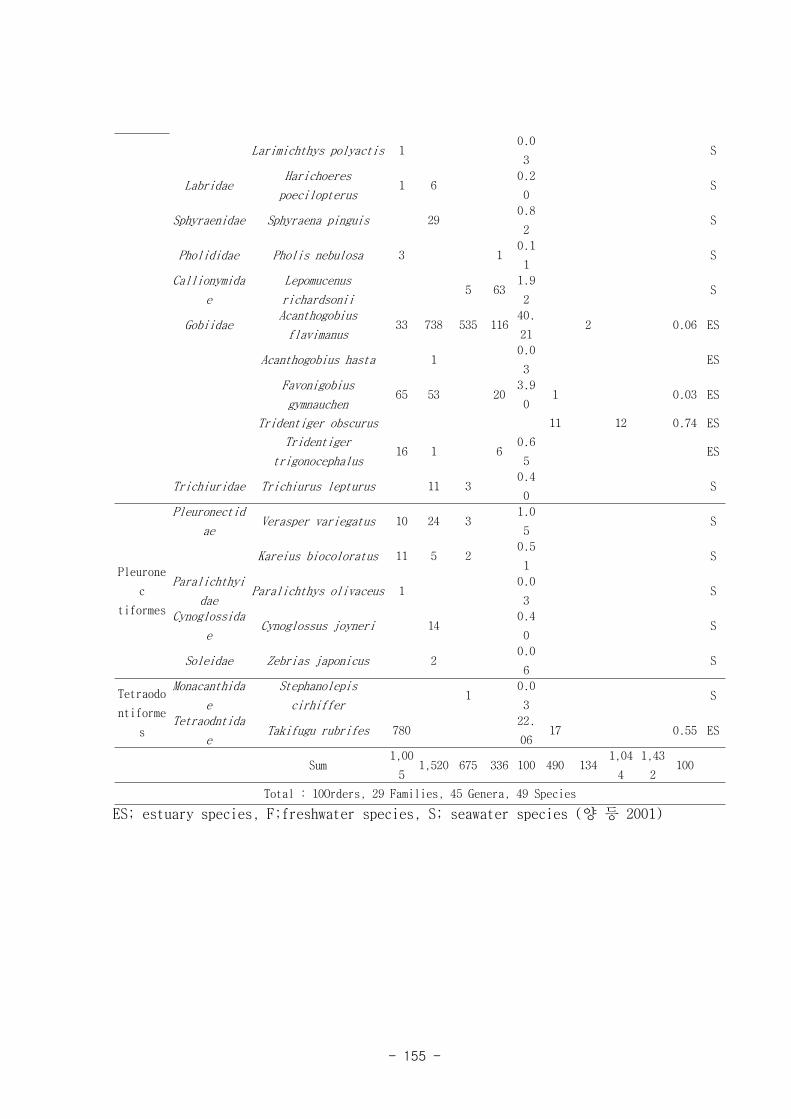
- 155 -
Larimichthys polyactis 10.0
3S
LabridaeHarichoeres
poecilopterus1 6
0.2
0S
Sphyraenidae Sphyraena pinguis 290.8
2S
Pholididae Pholis nebulosa 3 10.1
1S
Callionymida
e
Lepomucenus
richardsonii5 63
1.9
2S
GobiidaeAcanthogobius
flavimanus33 738 535 116
40.
212 0.06 ES
Acanthogobius hasta 10.0
3ES
Favonigobius
gymnauchen65 53 20
3.9
01 0.03 ES
Tridentiger obscurus 11 12 0.74 ES
Tridentiger
trigonocephalus16 1 6
0.6
5ES
Trichiuridae Trichiurus lepturus 11 30.4
0S
Pleurone
c
tiformes
Pleuronectid
aeVerasper variegatus 10 24 3
1.0
5S
Kareius biocoloratus 11 5 20.5
1S
Paralichthyi
daeParalichthys olivaceus 1
0.0
3S
Cynoglossida
eCynoglossus joyneri 14
0.4
0S
Soleidae Zebrias japonicus 20.0
6S
Tetraodo
ntiforme
s
Monacanthida
e
Stephanolepis
cirhiffer1
0.0
3S
Tetraodntida
eTakifugu rubrifes 780
22.
0617 0.55 ES
Sum1,00
51,520 675 336 100 490 134
1,04
4
1,43
2100
Total : 10Orders, 29 Families, 45 Genera, 49 Species
ES; estuary species, F;freshwater species, S; seawater species (양 등 2001)

- 156 -
제 3절 낙동강 하구의 생태 기능 요소
1. 생태 과정 (ecological processes)
가. 일차생산
(1) 저서미세조류(MPB)의 일차생산
① 일차생산의 측정
저서미세조류의 일차생산을 측정하는 방법으로는 방사성 동위 원소(H14CO3-)를 이용
한 방법(Leach, 1970), Bell jar를 이용하여 용존 산소 농도의 변화를 측정하는 방법
(Pomeroy, 1959), 산소 미세전극을 이용하는 방법(Revsbech et al., 1981) 등이 있다. 본
연구에서는 산소 미세전극을 이용하여 퇴적물 표층의 저서미세조류에 의한 생산량을
측정하였다. 산소 미세전극을 이용한 방법은 다른 방법들에 비해 짧은 시간 내(수 분
이내)에 광합성 측정이 용이하며, 전극의 tip의 미세하여 (<10㎛ i.d.) 갯벌 퇴적물과 같
은 미세한 입자내 존재하는 저서미세조류의 일차생산량 측정시 시료의 교란을 최소화
할 수 있다는 장점이 있다.(Revsbech & Jørgensen, 1986)
산소 미세전극이란 전극의 화학적 반응과 전기적 신호의 결합을 이용한 전기화학적
분석기기로, 일반적으로 환원전극(cathode, working electrode), 산화전극(anode,
reference electrode), 그리고 전해질(electrolyte)로 구성되어 있다. 산소 미세전극의 작
동은 환원전극(백금전극) 표면에서 산소의 화학적 환원에 기초를 두고 있다. 산소 환원
전류는 산소분압에 비례하므로 전류치로부터 산소농도를 구할 수 있다. 산화전극(기준
전극, 음극)에 특정 음(-)전위(-0.8 V)를 가하여 환원전극에 비해 음(-)의 전위차를 만
들면 환원전극 표면에서는 전극 내로 확산 유입된 산소분자에 전자를 제공하게 된다.
이때 전해질은 환원전극과 산화전극의 회로에 화학적 반응이 지속적으로 일어나게 하
는 역할을 한다. 공극수내 존재하는 산소와 같은 환원력이 큰 물질이 산소 미세전극의
실리콘 반투과성막을 통과하여 전극 내로 유입되면 환원전극에서 산소는 환원반응을
일으킨다. 이때 발생하는 전위차를 이용하여 용존산소의 농도를 측정하는 것이다.
② 시료의 채취 및 준비
퇴적물 표층에 서식하는 저서미세조류의 일차생산을 측정하기 위해, 현장에서 퇴적물
박스 코어(지름 10cm)를 이용하여 표층 5cm 정도까지 채취한 후에, 상온 상태로 실험
실로 운반하였다. 현장 해수를 채취한 퇴적물과 함께 작은 수조에 넣고, 작은 수조를
큰 수조의 중앙부에 위치시킨 후에, 큰 수조와 작은 수조 사이의 공간에는 냉수 또는
온수를 첨가하여 작은 수조의 해수를 현장수온으로 유지시켰다. 작은 수조내의 용존산
소량을 포화시키기 위해, 산소 기포발생 장치를 설치하였고, 퇴적물 표층의 광량은 현
장에서 측정된 광량에 준하도록 할로겐 램프를 이용하여 30분 이상 광적을 시켰다. 시

- 157 -
료의 준비가 끝난 후에, 산소 미세전극을 이용하여 퇴적물내 공극수의 용존산소 농도를
측정하였다.
③ 용존산소 측정 및 일차생산량 추정
실험에 사용된 산소 미세전극은 전극 크기가 <10um인 Clark type 전극으로, 전극의
검량은 깊이에 따라 더 이상의 전류 감소가 나타나지 않는 무산소 퇴적층에서의 전류
값(산소 농도=0)과 산소에 100% 포화된 표층 해수에서의 전류값을 이용하였다
(Revsbech & Jørgensen, 1983). 포화 용존 산소농도는 현장 수온과 염분 자료로부터
계산하였고, 각 퇴적물 깊이에서의 용존 산소 농도는 검출된 전극의 전류와 산소 농도
간의 선형관계를 이용하여 계산하였다. 미세 전극은 10um의 수직 이동해상도를 갖는
micromanipulator에 장착하여 100um 간격으로 표층 해수로부터 퇴적물에 삽입하며 산
소 농도를 측정하였다. 전극에서 검출된 전류는 picoammeter를 거쳐 strip-chart
recorder에 기록되었다.
순광합성율은 산소농도의 미세수직분포로부터 추정하였다. 퇴적물의 유광층에서 광합
성에 의해 생산된 산소는 확산에 의해 상층과 저층으로 수직이동하기 때문에 상부로
확산되는 산소 플럭스와 하부로 확산되는 산소 플럭스로부터 순광합성률을 추정할 수
있다. 물질의 확산 플럭스는 Fick의 확산 제1법칙에 의하여 구하였다.(Revsbech &
Jørgensen, 1986)
Figure. 3-1. Sampling and experimental set-up for the measurement of
microphytobenthos production. Samples were transported to laboratory on ice
without disturbance and oxygen profiles in upper sediment were obtained
immediately.
④ 주요 결과 및 토의
낙동강 하구, 명지 지역의 퇴적물내 서식하는 저서미세조류의 생산량을 측정하기 위
해, 2005년 8월과 2006년 12월에 명지 지역 상부의 original sediment로부터 퇴적물을
채취하여 산소미세전극을 이용하여 일차생산량을 측정하였다.(Fig. 3-1) 산소 농도의
미세 수직 분포는 Fig. 3-2 에 제시하였다.
퇴적물 시료 표층 0.1mm 깊이에서 용존 산소 농도는 최대값(265~305umol L-1
)을 보
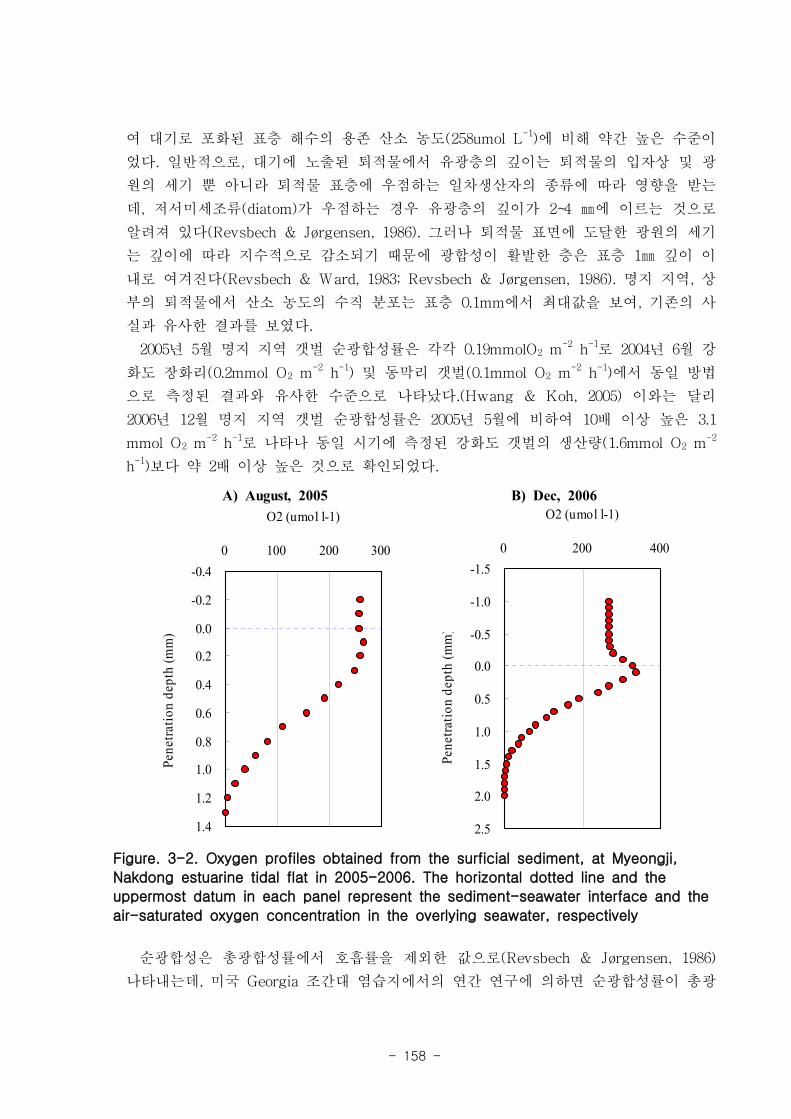
- 158 -
여 대기로 포화된 표층 해수의 용존 산소 농도(258umol L-1
)에 비해 약간 높은 수준이
었다. 일반적으로, 대기에 노출된 퇴적물에서 유광층의 깊이는 퇴적물의 입자상 및 광
원의 세기 뿐 아니라 퇴적물 표층에 우점하는 일차생산자의 종류에 따라 영향을 받는
데, 저서미세조류(diatom)가 우점하는 경우 유광층의 깊이가 2~4 ㎜에 이르는 것으로
알려져 있다(Revsbech & Jørgensen, 1986). 그러나 퇴적물 표면에 도달한 광원의 세기
는 깊이에 따라 지수적으로 감소되기 때문에 광합성이 활발한 층은 표층 1㎜ 깊이 이
내로 여겨진다(Revsbech & Ward, 1983; Revsbech & Jørgensen, 1986). 명지 지역, 상
부의 퇴적물에서 산소 농도의 수직 분포는 표층 0.1mm에서 최대값을 보여, 기존의 사
실과 유사한 결과를 보였다.
2005년 5월 명지 지역 갯벌 순광합성률은 각각 0.19mmolO2 m-2 h
-1로 2004년 6월 강
화도 장화리(0.2mmol O2 m-2 h
-1) 및 동막리 갯벌(0.1mmol O2 m
-2 h-1
)에서 동일 방법
으로 측정된 결과와 유사한 수준으로 나타났다.(Hwang & Koh, 2005) 이와는 달리
2006년 12월 명지 지역 갯벌 순광합성률은 2005년 5월에 비하여 10배 이상 높은 3.1
mmol O2 m-2 h
-1로 나타나 동일 시기에 측정된 강화도 갯벌의 생산량(1.6mmol O2 m-2
h-1
)보다 약 2배 이상 높은 것으로 확인되었다.
A) August, 2005 B) Dec, 2006
-0.4
-0.2
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
0 100 200 300
O2 (umol l-1)
Penetration depth (mm)
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0
2.5
0 200 400
O2 (umol l-1)
Penetration depth (mm)
Figure. 3-2. Oxygen profiles obtained from the surficial sediment, at Myeongji,
Nakdong estuarine tidal flat in 2005-2006. The horizontal dotted line and the
uppermost datum in each panel represent the sediment-seawater interface and the
air-saturated oxygen concentration in the overlying seawater, respectively
순광합성은 총광합성률에서 호흡률을 제외한 값으로(Revsbech & Jørgensen, 1986)
나타내는데, 미국 Georgia 조간대 염습지에서의 연간 연구에 의하면 순광합성률이 총광
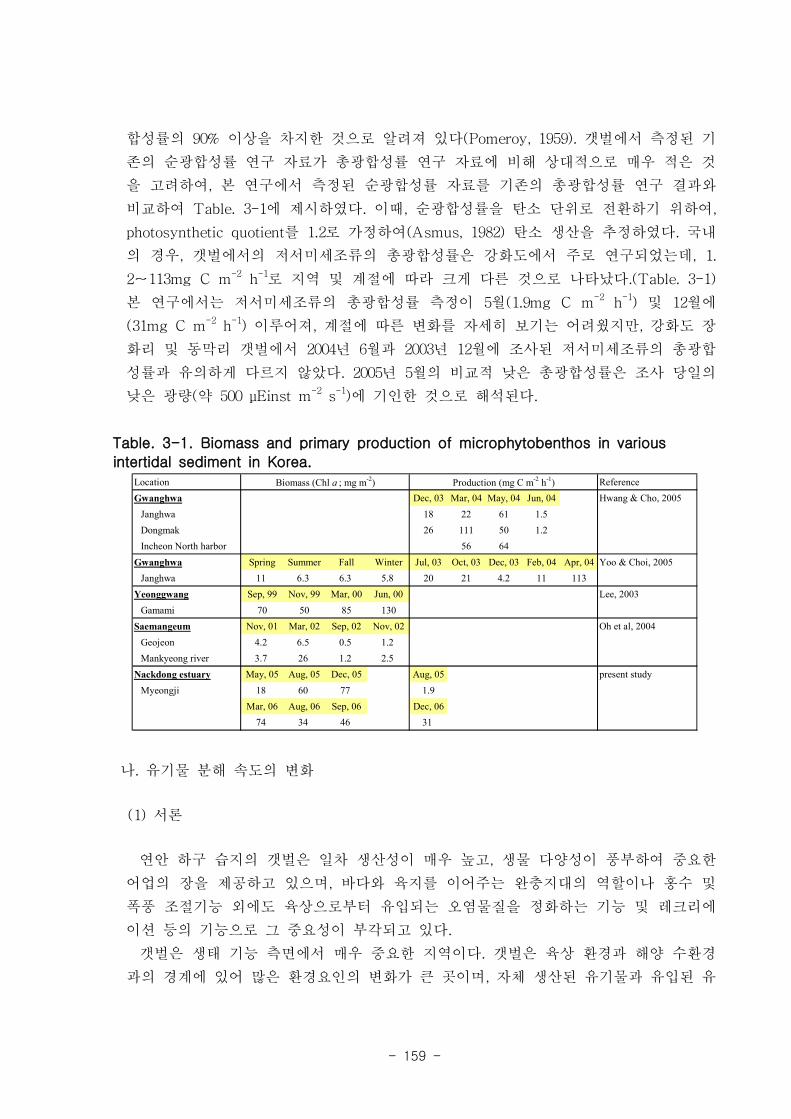
- 159 -
합성률의 90% 이상을 차지한 것으로 알려져 있다(Pomeroy, 1959). 갯벌에서 측정된 기
존의 순광합성률 연구 자료가 총광합성률 연구 자료에 비해 상대적으로 매우 적은 것
을 고려하여, 본 연구에서 측정된 순광합성률 자료를 기존의 총광합성률 연구 결과와
비교하여 Table. 3-1에 제시하였다. 이때, 순광합성률을 탄소 단위로 전환하기 위하여,
photosynthetic quotient를 1.2로 가정하여(Asmus, 1982) 탄소 생산을 추정하였다. 국내
의 경우, 갯벌에서의 저서미세조류의 총광합성률은 강화도에서 주로 연구되었는데, 1.
2~113mg C m-2 h
-1로 지역 및 계절에 따라 크게 다른 것으로 나타났다.(Table. 3-1)
본 연구에서는 저서미세조류의 총광합성률 측정이 5월(1.9mg C m-2 h
-1) 및 12월에
(31mg C m-2 h
-1) 이루어져, 계절에 따른 변화를 자세히 보기는 어려웠지만, 강화도 장
화리 및 동막리 갯벌에서 2004년 6월과 2003년 12월에 조사된 저서미세조류의 총광합
성률과 유의하게 다르지 않았다. 2005년 5월의 비교적 낮은 총광합성률은 조사 당일의
낮은 광량(약 500 µEinst m-2 s-1
)에 기인한 것으로 해석된다.
Table. 3-1. Biomass and primary production of microphytobenthos in various
intertidal sediment in Korea.Location Reference
Gwanghwa Dec, 03 Mar, 04 May, 04 Jun, 04 Hwang & Cho, 2005
Janghwa 18 22 61 1.5
Dongmak 26 111 50 1.2
Incheon North harbor 56 64
Gwanghwa Spring Summer Fall Winter Jul, 03 Oct, 03 Dec, 03 Feb, 04 Apr, 04 Yoo & Choi, 2005
Janghwa 11 6.3 6.3 5.8 20 21 4.2 11 113
Yeonggwang Sep, 99 Nov, 99 Mar, 00 Jun, 00 Lee, 2003
Gamami 70 50 85 130
Saemangeum Nov, 01 Mar, 02 Sep, 02 Nov, 02 Oh et al, 2004
Geojeon 4.2 6.5 0.5 1.2
Mankyeong river 3.7 26 1.2 2.5
�ackdong estuary May, 05 Aug, 05 Dec, 05 Aug, 05 present study
Myeongji 18 60 77 1.9
Mar, 06 Aug, 06 Sep, 06 Dec, 06
74 34 46 31
Biomass (Chl a ; mg m-2) Production (mg C m
-2 h
-1)
나. 유기물 분해 속도의 변화
(1) 서론
연안 하구 습지의 갯벌은 일차 생산성이 매우 높고, 생물 다양성이 풍부하여 중요한
어업의 장을 제공하고 있으며, 바다와 육지를 이어주는 완충지대의 역할이나 홍수 및
폭풍 조절기능 외에도 육상으로부터 유입되는 오염물질을 정화하는 기능 및 레크리에
이션 등의 기능으로 그 중요성이 부각되고 있다.
갯벌은 생태 기능 측면에서 매우 중요한 지역이다. 갯벌은 육상 환경과 해양 수환경
과의 경계에 있어 많은 환경요인의 변화가 큰 곳이며, 자체 생산된 유기물과 유입된 유

- 160 -
기물이 풍부하여, 많은 생물이 살고 있는 곳이다. 갯벌에서 일어나는 일차 생산 및 분
해과정은 갯벌과 주변 해역의 영양염 순환과 물질수지를 조절하는 기본 과정이다. 육지
로부터 유입된 유기물은 갯벌 내에 서식하는 미생물에 의해 분해되고 이 과정에서 생
성된 영양염은 다시 갯벌 표면에 서식하는 저서성규조류의 일차 생산에 이용될 수 있
으며, 저서성 규조류에 의해 생성된 유기물은 갯벌의 저서생물의 먹이로 이용되는 등,
복잡한 먹이사슬을 통하여 하구 생태계 물질 순환이 이루어진다.
특히 육상이나, 주변 집수역으로부터 유입된 유기물이 미생물에 의해 분해 흡수되어
무기물화 되는 과정을 생지화학적 관점에서 재광물화 (remineralization)라고 한다. 재광
물화는 퇴적물내 미생물에 의해 유기물이 분해되는 과정으로 산화제로서 산소나 산소
이외의 물질(NO₃⁻, MnO₂, FeO₂H, SO4
2-)을 이용하게 되는데, 이때 무기 형태의 질소
나 인등이 영양염으로 재순환 (recycling)된다.
본 연구에서는 재광물화를 정량적으로 파악하기 위해 퇴적물 산소 요구량(Sediment
oxygen demand : SOD)을 이용하였다. 미생물은 O2를 소모하면서 유기물을 분해하는
데, 이 때 퇴적물의 면적당 시간당 소모되는 O2농도를 환산하여 유기물 분해율, 즉
SOD를 구할 수 있다. 본 연구의 목적은 낙동강 하구 생태계의 물질순환 중 미생물의
활동 즉, 유기물 분해율을 정량적으로 파악하기 위한 것이다.
(2) 재료 및 방법
연구가 진행된 지역은 낙동강 하구 명지 주거 앞 갯벌에 설치된 실험용 갯벌이다
(Fig. 3-3). 2004년의 지형 정밀 측정결과 낙동강 하구갯벌에 위치한 명지 지역이 실험
용 갯벌 적지로 판단되었고, 2005년 4월 1차 실험용 갯벌이 완성되었다( 가로: 1m, 세
로:1m, 깊이: 1m). 실험용 갯벌은 평균 해수면 위 85.1, 142.9, 175cm에 서로 다른 3개
의 조고(상, 중, 하)를 갖도록 설치하였으며, 각 조고에는 3가지 다른 타입의 퇴적물(사
질, 혼합-명지 본래의 퇴적물, 니질)을 채워 넣었다. 2006년 2월에는 1차 갯벌 인근에 1
차 갯벌과 유사한 조건이나, 면적이 4배로 늘어난 실험용 갯벌(가로: 2m, 세로: 2m, 깊
이: 2m)을 설치하였다.
Figure. 3-3. Artificial intertidal flat in Nakdong estuary.

- 161 -
Figure. 3-4. Batch incubation
실험용 갯벌로부터 2005년 7월에서 11월까지 동안 매달 유기물 분해율 측정을 위한
현장 시료를 채취하였다. 실험용 갯벌의 각 treatment에서 3개의 코어 (지름=4cm, 길이
=22cm), 총 27 ~ 36개의 코어(각 treatment에서 3개씩) 와 현장 해수 약 100L 를 채취
하여 1시간 이내에 실험실로 가져와 배양시스템에 설치하였다(Fig. 3-4).
퇴적물 배양 시스템은 55cm * 55cm * 35cm크기의 항온 수조이다. 퇴적물 코어는
7cm까지 퇴적물을 취하고 윗부분에 overlying water를 두고 현장 온도를 유지하며 퇴
적물을 24시간 동안 전배양을 하였다(Fig. 3-4). 퇴적물 배양은 약 5일 동안 진행되었
는데 전배양 24시간 후에 수조 내 해수와의 유동을 차단해 주기 위해 코어 윗부분에
stopper를 설치하여, 해수 교환을 차단하였다. 초기의 용존 산소 농도를 알기 위하여,
임의로 빈 코어 3개를 넣어 물만 채취하여 측정하였다. 해수 차단 후 각 treatment 당
1시간 후에 1개의 코어를 분석 하고, 24시간이 지난 후 2개의 코어로부터 head-space
의 해수에 용존 산소의 농도를 MIMS(membrane inlet mass spectrometer; 질량분석기)
를 이용하여 측정 하였다. 각 배양 시간의 농도 측정값은 코어의 단면적을 고려하여
SOD로 환산하였다. 퇴적물 산소요구량은 시간에 따른 용존 산소 농도를 보고 시간과
용존 산소 농도 간에 회귀 분석을 통해 계산되었다. 이 감소율로부터 SOD를 측정하는
식은 다음과 같다.
SOD Flux (mmoleㆍm²ㆍday)= (Z×10×24) / S
Z= (V×X)/ 1000(mL)
V= headspace의 부피(cm³)
X= 시간에 따른 DO 의 감소율 (μM/h)
S= core면적(cm²)

- 162 -
(3) 주요 결과 및 토의
실험용 갯벌에서 시간에 따른 유기물 분해율을 알아보기 위하여 2005년에 만들어진
실험용 갯벌에서는 2005년 7월부터 2006년 9월까지, 2006년에 만들어진 실험용 갯벌에
서는 2006년 3월부터 10월까지 퇴적물 코어 배양을 실시하였으며 이로부터 SOD, 영양
염 flux을 측정하였다. 또한, SOD에 영향을 미치는 환경 factor, 즉 실험용 갯벌의 깊이
별 퇴적물 입도, 퇴적물내 유기물 함량을 측정하였다.
(가) 계절 변화
낙동강 실험용 갯벌에서의 SOD를 분석한 결과 여름에는 높은 값을 보였고, 겨울에는
낮은 값을 보였다(Fig. 3-5). 05년 실험용 갯벌의 SOD 값의 범위는 -0.7~35.3mmole •
m-2
• d-1 였고, 하절기에 가장 높은 SOD 값(mean: 30 mmole • m
-2• d
-1)을 보였으며,
동절기에는 산소가 소비되기 보다는 광합성에 의해 산소가 생성되는 것을 나타났다
(mean :-0.5mmole • m-2
• d-1
). 06년 실험용 갯벌의 SOD 값의 범위는 -0.5~38.2
mmole • m-2
• d-1 였고, 05년 실험용 갯벌과 유사하게 하절기 (mean :37mmole • m
-2•
d-1
), 동절기 (mean :0.1mmole • m-2
• d-1
)의 SOD 값을 나타냈다. 이는 미생물 활동을
조절하는 큰 요인은 계절적인 수온 변화임을 시사한다. 설치한 후 1년이 지난 05년 실
험용갯벌에서 SOD의 2005년에 측정된 값의 범위와 2006년에 측정된 값의 범위는 유사
한 것으로 나타났다.
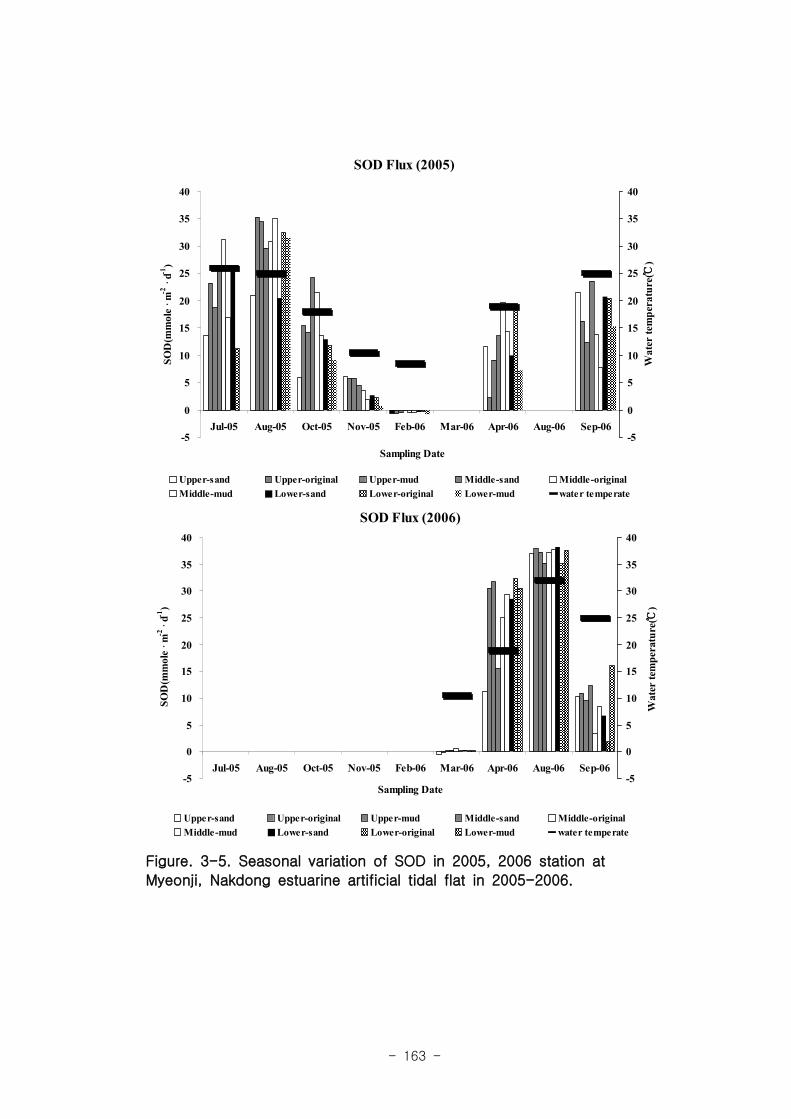
- 163 -
SOD Flux (2005)
-5
0
5
10
15
20
25
30
35
40
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling Date
SOD(m
mole · m
-2 · d-1)
-5
0
5
10
15
20
25
30
35
40
Water temperature(℃
)
Upper-sand Upper-original Upper-mud Middle-sand Middle-original
Middle-mud Lower-sand Lower-original Lower-mud water temperate
SOD Flux (2006)
-5
0
5
10
15
20
25
30
35
40
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling Date
SOD(m
mole · m
-2 · d-
1)
-5
0
5
10
15
20
25
30
35
40
Water temperature(℃
)
Upper-sand Upper-original Upper-mud Middle-sand Middle-original
Middle-mud Lower-sand Lower-original Lower-mud water temperate
Figure. 3-5. Seasonal variation of SOD in 2005, 2006 station at
Myeonji, Nakdong estuarine artificial tidal flat in 2005-2006.

- 164 -
다. 탈질소화의 계절변화
(1) 서론
해양에서 무기 질소 성분은 일차생산을 제한하는 영양분이므로 (Ryther and
Dunston, 1971; Capone, 1988), 무기질소성분의 변환과정의 속도 및 생성물의 성격은
일차생산에 의한 유기물 생산에 직접적으로 영향을 미칠 것이다 (Rowe et al., 1975;
Capone, 1988). 탈질소화(denitification)는 혐기성 미생물에 의해 질산이 질소가스로 변
화되는 과정으로써, 탈질소화의 산물인 질소 가스는 일차생산에 이용되기 힘들다
(Seitzinger, 1988; Seitzinger, 1990). 따라서 탈질소화는 시스템 내에서 총질소량을 감
소시키고 결국 유기물을 제거하는 기작이다. 탈질소화가 상세히 연구된 연안이나 하구
에서는 이 과정이 플랑크톤에 의한 일차생산성을 제한하거나 부영양화를 감소시키는데
중요하다는 것이 밝혀졌다 (Howes et al., 1984; An and Joye, 2001). 일반적으로 갯벌
은 염습지와 마찬가지로 유기물 공급이 높아 탈질소화가 높을 가능성이 있으나 현재
세계적으로 갯벌에서 탈질소화를 측정 자료는 매우 희귀한 실정이다 (Kaplan et al.,
1979). 그 이유 중의 하나는 측정법이 비교적 힘들기 때문이다.
탈질화를 측정하는 가장 이상적인 방법은 시간에 따른 탈질소화의 최종산물 (질소가
스)의 농도변화를 측정하는 것이다. 하지만 대기 중 (78%) 이나 수중 (용존 기체의
61%)에 질소가스 농도가 높아서, 탈질소화에 의한 질소가스 변화를 측정하는 것은 매
우 어렵다 (Lamontagne and Valiela, 1995; Seitzinger et al., 1994). 이러한 문제점을
해결하기 위해 다양한 방법들이 제시되었는데 널리 사용된 예로서는 아세틸렌을 탈질
소화의 저해물질로 이용하는 방법과 안정동위원소를 이용하는 방법 등이 있다
(Seitzinger, 1988; Seitzinger et al., 1994). 아세틸렌은 탈질소화의 최종단계인 아산화질
소 (N2O)에서 질소가스로의 변환을 저해한다. 아세틸렌을 주입한 후 아산화질소의 생
산량을 측정하여 탈질소화를 측정하였는데 비교적 간단하고 저렴하게 측정할 수 있는
장점이 있으나 여러 문제점이 지적되어 왔다 (Knowles, 1990). 안정동위원소를 이용한
측정법은 고가의 장비와 복잡한 용존 가스 추출단계를 필요로 하여 제한적으로 사용되
어왔다 (Koike and Hattori, 1979; Seitzinger, 1990). Seitzinger et al. (1980)는 탈질소
화 측정이전에 수층의 질소가스를 모두 제거 한 후 질소가스 농도변화를 측정하였으나,
7-10일간의 전배양 (preincubation) 기간 동안 탈질소화를 비롯한 생지화학적 과정들이
변할 수 있어 탈질소화가 실제보다 적게 측정될 수 있다고 지적되어 왔다 (Christensen
et al., 1987; Brandes and Devol, 1995; An and Joye, 2001).
최근 들어 매우 정밀하게 질소가스 농도를 측정하는 방법 (Brandes and Devol, 1995;
Lamontagne and Valiela, 1995), 아르곤 가스를 내부 표준기체 (internal standard)로 이
용하는 방법 (Kana et al., 1994; An and Joye, 1997), 질소동위원소를 이용한 방법
(Nielson, 1992)등이 개발되었다. 이 중 Kana et al. (1994)가 개발한 MIMS (membrane
inlet mass spectrometer) 시스템과 Nielson (1992)이 개발한 isotope pairing technique

- 165 -
등이 비교적 간단하고 정확하게 탈질소화를 측정하는 것이 가능하여 널리 사용되고 있
다.
An et. al (2001)는 MIMS 시스템의 기능을 확장시켜, MIMS 시스템을 이용한
isotope pairing technique이 가능하게 하였다. MIMS 시스템에서는 용존 질소가 실리콘
막을 통해 직접 질량분석기의 검출기로 유입되기 때문에 측정 과정이 간편하고 다른
방법에서 용존 가스 추출 시 발생하는 오차를 크게 줄일 수 있었다 (Kana et al.,
1994). 이러한 MIMS 시스템의 장점을 이용하면서, MIMS 시스템에 안정동위원소를 가
진 질소가스들 (29N2,
30N2)을 측정할 수 있는 기능을 더함으로서, isotope paring
technique 이 가능했다 (An et al., 2001). 본 연구에서는 MIMS system 과 isotope
paring technique을 이용하여 탈질소화를 측정하였다.
(2) 재료 및 방법
현장에서 퇴적물 코어는 퇴적물을 7cm까지 취하고 윗부분에 overlying water를 두고
현장 온도를 유지하며 퇴적물을 배양을 하였다. 퇴적물 배양은 약 5일 동안 진행되었는
데 SOD를 먼저 측정하고 다시 전배양 24시간 후 배양수에 15NO3
- (D.W 100ml -
Na15NO3 400mg : 최종 15
NO3-농도~ 400μM)를 인위로 공급하였다.
탈질소화는 29N2,
30N2의 농도변화로부터 isotope paring technique(Nielson, 1992)을
이용하여 구하였고, 다음과 같은 식을 이용하였다.
D15 ; 2n (15, 15) + n (14, 15)
D14 ; n (14, 15) / 2n (15, 15)* D15
D15 ; 15NO3을 이용한 탈질소화율
D14 ; 14NO3을 이용한 탈질소화율
n(14, 15); 29N2 발생율 (N-equivalent)
n(15, 15); 30N2 발생율 (N-equivalent)
(3) 주요 결과 및 토의
(가) 계절 변화
낙동강 실험용 갯벌에서의 탈질소화를 분석한 결과 여름에 높은 값이, 겨울에는 낮은
값이 나타났다. 2005년 실험용 갯벌의 탈질소화 값의 범위는 평균 5.9~2652μmole • m-
2• d
-1 (mean)였고, 하절기에 가장 높은 탈질소화 값(4564.6μmole • m-2
• d-1
)을 보였으
며, 동절기에는 거의 탈질소화가 발생하지 않았다(1.6μmole • m-2
• d-1
). 06년 실험용 갯
벌의 탈질소화 값의 범위는 3.9~2082μmole • m-2
• d-1였고, 05년 실험용 갯벌과 유사하
게 하절기 (2626.5μmole • m-2
• d-1
), 동절기 (0.01μmole • m-2
• d-1
)의 탈질소화 값을 나
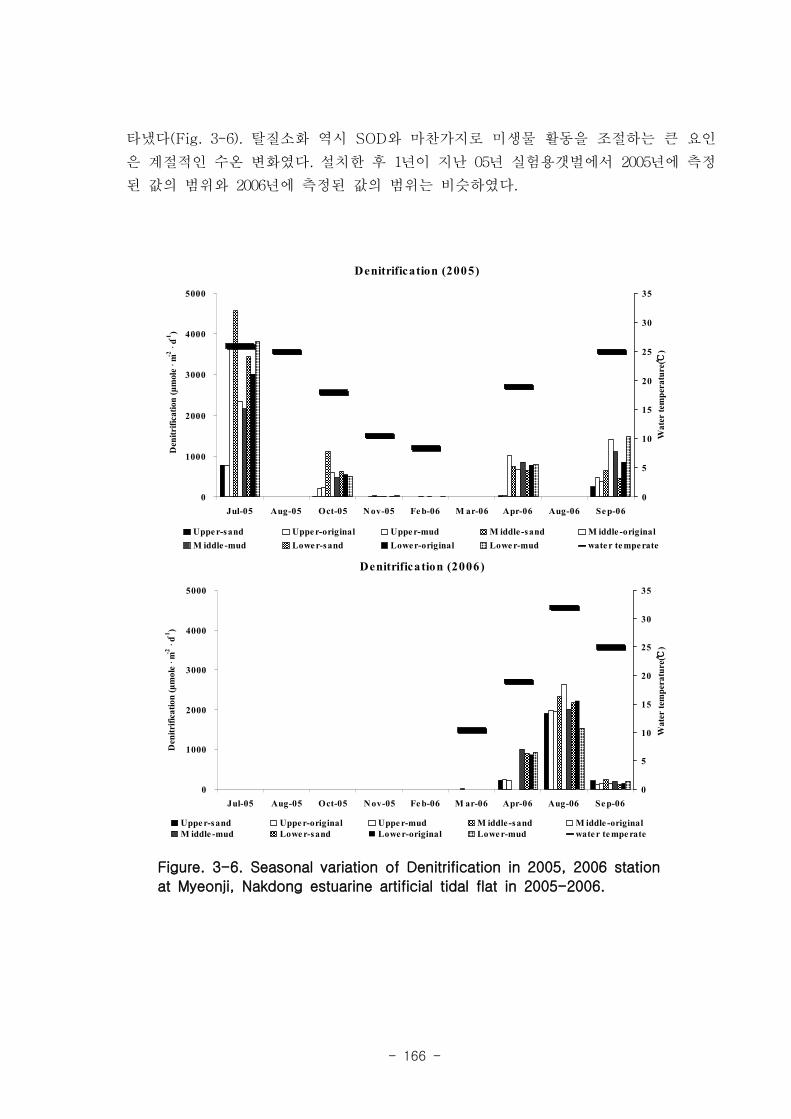
- 166 -
Denitrification (2005)
0
1000
2000
3000
4000
5000
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 M ar-06 Apr-06 Aug-06 Sep-06
Denitrification (µmole · m
-2 · d-1)
0
5
10
15
20
25
30
35
Water temperature(℃
)
Upper-sand Upper-original Upper-mud M iddle-sand M iddle -original
M iddle -mud Lower-sand Lower-original Lower-mud water temperate
Denitrification (2006)
0
1000
2000
3000
4000
5000
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 M ar-06 Apr-06 Aug-06 Sep-06
Denitrification (µmole · m
-2 · d-1)
0
5
10
15
20
25
30
35
Water tem
perature(℃
)
Upper-sand Upper-original Upper-mud M iddle-sand M iddle -original
M iddle -mud Lower-sand Lower-original Lower-mud water temperate
Figure. 3-6. Seasonal variation of Denitrification in 2005, 2006 station
at Myeonji, Nakdong estuarine artificial tidal flat in 2005-2006.
타냈다(Fig. 3-6). 탈질소화 역시 SOD와 마찬가지로 미생물 활동을 조절하는 큰 요인
은 계절적인 수온 변화였다. 설치한 후 1년이 지난 05년 실험용갯벌에서 2005년에 측정
된 값의 범위와 2006년에 측정된 값의 범위는 비슷하였다.
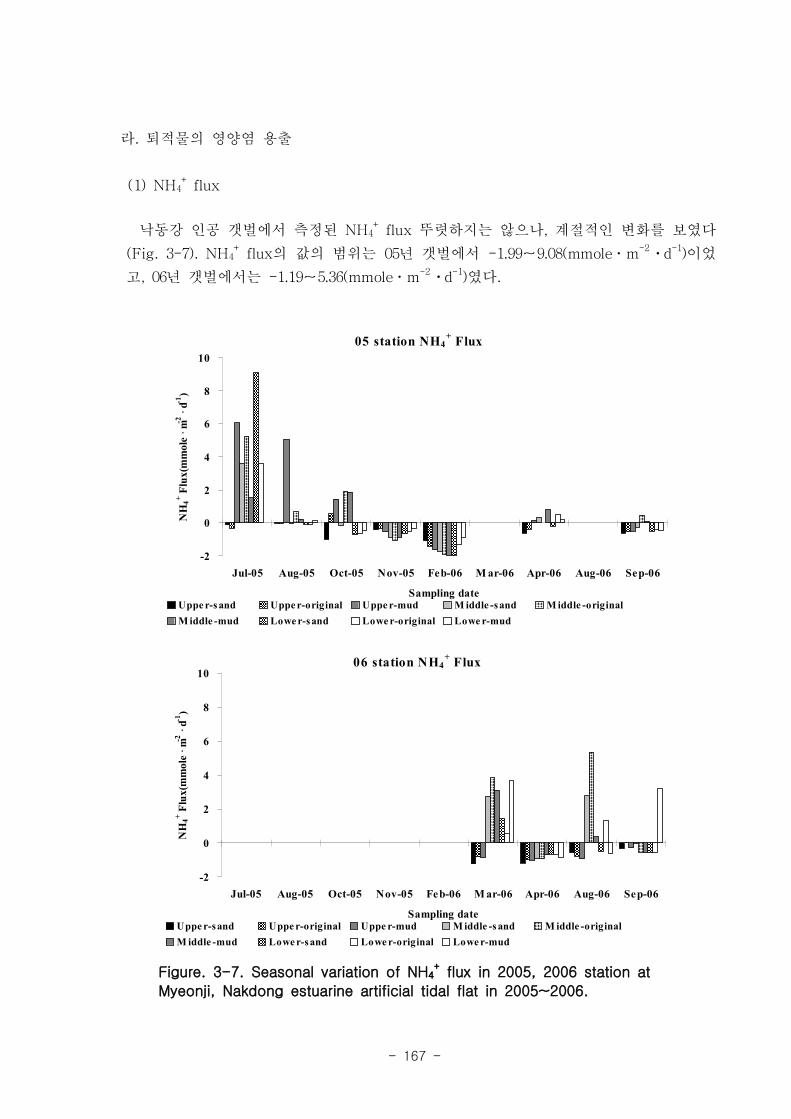
- 167 -
05 station �H4+ Flux
-2
0
2
4
6
8
10
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
�H
4+ Flux(m
mole · m
-2 · d
-1)
Upper-sand Upper-original Upper-mud M iddle -sand Middle -original
M iddle -mud Lower-sand Lower-original Lowe r-mud
06 station �H4+ Flux
-2
0
2
4
6
8
10
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
�H
4+ Flux(m
mole · m
-2 · d
-1)
Upper-sand Upper-original Uppe r-mud Middle -sand M iddle -original
M iddle -mud Lower-sand Lower-original Lower-mud
Figure. 3-7. Seasonal variation of NH4+ flux in 2005, 2006 station at
Myeonji, Nakdong estuarine artificial tidal flat in 2005~2006.
라. 퇴적물의 영양염 용출
(1) NH4+ flux
낙동강 인공 갯벌에서 측정된 NH4+ flux 뚜렷하지는 않으나, 계절적인 변화를 보였다
(Fig. 3-7). NH4+ flux의 값의 범위는 05년 갯벌에서 -1.99~9.08(mmoleㆍm
-2• d
-1)이었
고, 06년 갯벌에서는 -1.19~5.36(mmoleㆍm-2
• d-1
)였다.

- 168 -
계절별로 각각의 조고 및 퇴적물 조건에 따라 서로 다른 양상을 보였다. 05년 갯벌에
서 여름철에는 대부분의 treatment에서 퇴적층에서 수층으로의 용출이 관찰되었다. 이
는 수온이 높아지는 여름철 미생물에 의한 높은 유기물 분해율에 의해 재광물화가 활
발하고 생성되는 NH4+의 양이 많아짐에 따라 수층으로 확산된 것으로 생각된다. 또 다
른 원인으로는 퇴적물내 미생물인 질산화 박테리아의 활동이 작았음을 예상할 수 있다.
질산화 박테리아의 활동을 제한하는 요인으로 크게 두 가지가 있는데, NH4+의 양과 용
존 산소의 농도이다(Nixon et al., 1976; Seitzinger et al., 1983). 여기서 여름철 높은 유
기물 분해율 때문에 수층의 용존산소가 고갈되었고 이에 따라 질산화 박테리아의 활동
이 제한되었으며, 질산화에 의한 NH4+사용이 줄어들어, 남은 NH4
+이 퇴적층 밖으로 용
출된 것을 예상할 수 있다.
봄과 가을에 측정된 NH4+ flux는 그 값이 작아지면서 용출과 소비 두 가지 패턴을
모두 나타내었다. 여기서 소비 flux의 경향은 유기물 분해에 의해 퇴적물 내에서 생성
되는 NH4+이 바로 질산화 박테리아에 이용되었거나, 퇴적물 표층에 서식하는
MPB(Microphytobenthos)의 광합성 작용에 의해 흡수되었을 가능성을 예상할 수 있다.
봄과 가을의 작은 값의 용출 flux는 유기물 분해에 의해 퇴적물내 NH4+의 농도가 수층
의 NH₄⁺ 농도보다 높아져 퇴적물로부터 수층으로 확산에 의해 이동된 것이라 생각되
며 또는, 우리가 측정한 코어내 대형저서동물의 excretion(배설)에 의해 수층의 NH4+의
농도가 증가하여 용출된 것으로 나타났을 수도 있다. 06년 갯벌의 경우 여름과 봄/가을
의 계절에 관계없이 NH4+ flux가 발생되었다.
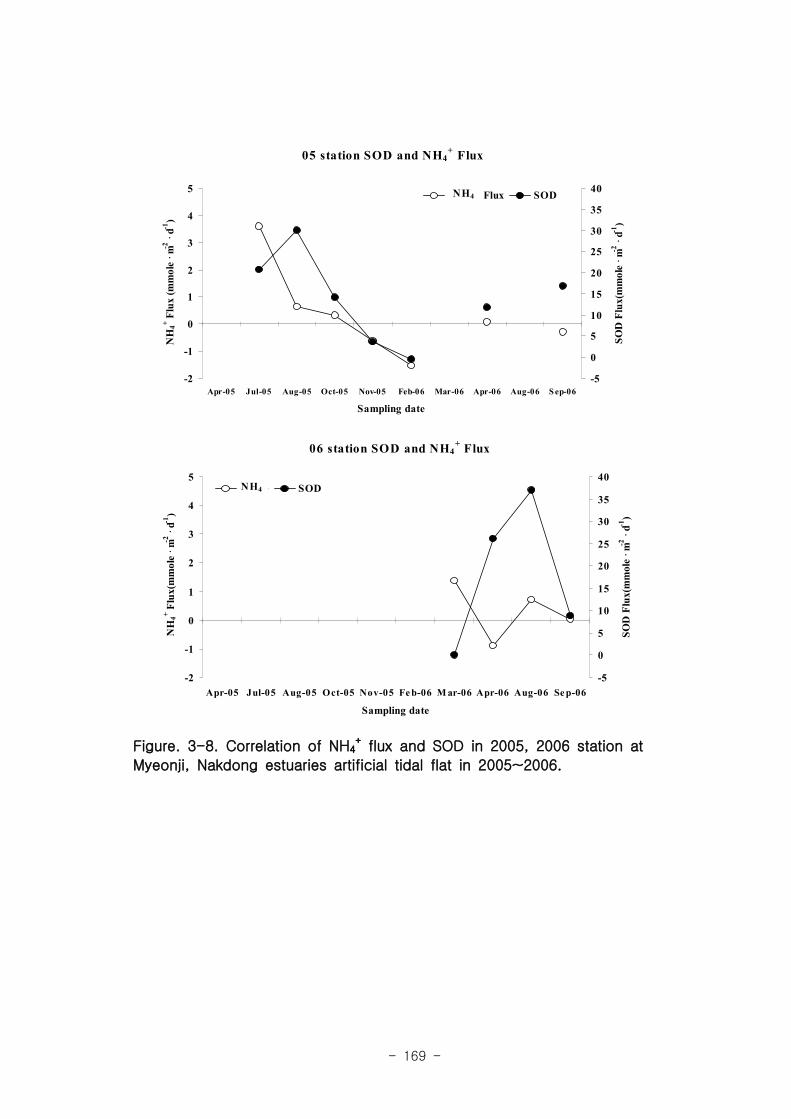
- 169 -
05 station SOD and �H4+ Flux
-2
-1
0
1
2
3
4
5
Apr-05 Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
�H
4+ Flux (mmole · m
-2 · d
-1)
-5
0
5
10
15
20
25
30
35
40
SOD Flux(m
mole · m
-2 · d
-1)
�H4+ Flux SOD�H4
06 station SOD and �H4+ Flux
-2
-1
0
1
2
3
4
5
Apr-05 Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
�H
4+ Flux(m
mole · m
-2 · d
-1)
-5
0
5
10
15
20
25
30
35
40
SOD Flux(m
mole · m
-2 · d
-1)
�H4+ SOD�H4
Figure. 3-8. Correlation of NH4+ flux and SOD in 2005, 2006 station at
Myeonji, Nakdong estuaries artificial tidal flat in 2005~2006.
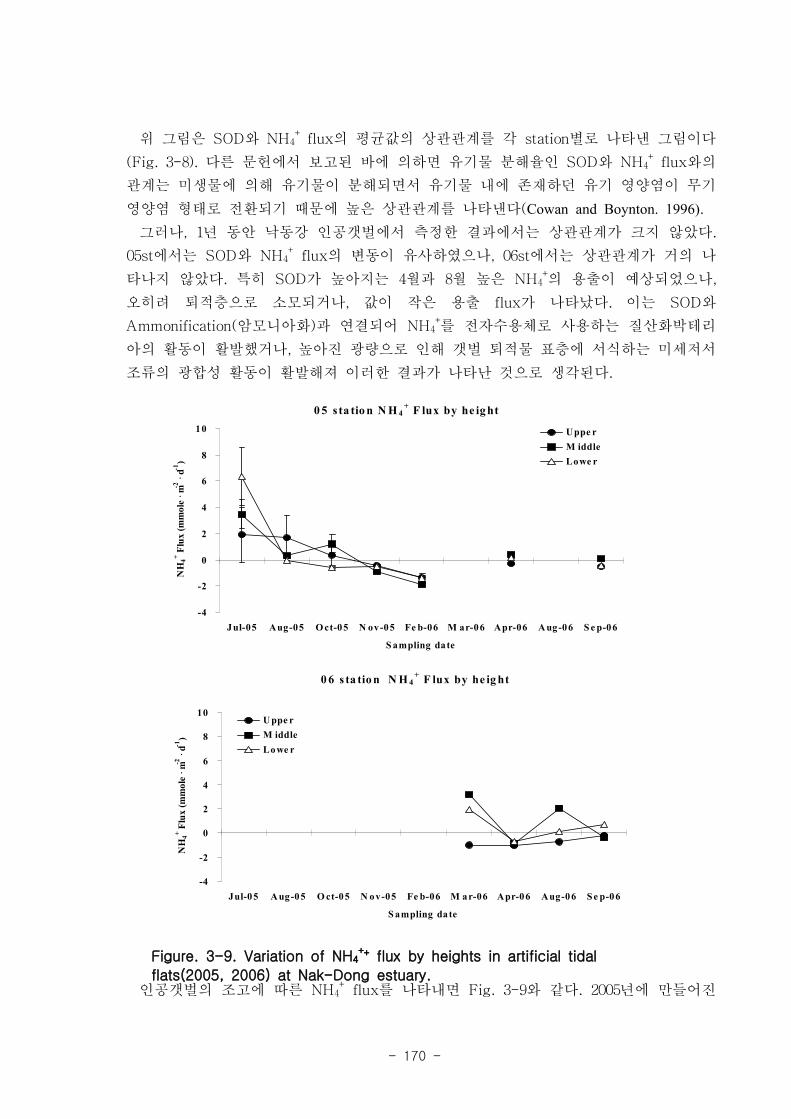
- 170 -
05 s ta tion �H 4+ F lux by he ight
-4
-2
0
2
4
6
8
10
Jul-05 Aug-05 Oct-05 � ov-05 Fe b-06 M ar-06 Apr-06 Aug-06 Se p-06
Sampling date
�H
4+ Flux (mmole · m
-2 · d-1)
Uppe r
M iddle
Lowe r
06 s ta tion �H 4+ F lux by he ight
-4
-2
0
2
4
6
8
10
Jul-05 Aug-05 Oct-05 � ov-05 Fe b-06 M ar-06 Apr-06 Aug-06 Se p-06
Sampling date
�H
4+ Flux (mmole · m
-2 · d-1)
Uppe r
M iddle
Lowe r
Figure. 3-9. Variation of NH4+⁺ flux by heights in artificial tidal
flats(2005, 2006) at Nak-Dong estuary.
위 그림은 SOD와 NH4+ flux의 평균값의 상관관계를 각 station별로 나타낸 그림이다
(Fig. 3-8). 다른 문헌에서 보고된 바에 의하면 유기물 분해율인 SOD와 NH4+ flux와의
관계는 미생물에 의해 유기물이 분해되면서 유기물 내에 존재하던 유기 영양염이 무기
영양염 형태로 전환되기 때문에 높은 상관관계를 나타낸다(Cowan and Boynton. 1996).
그러나, 1년 동안 낙동강 인공갯벌에서 측정한 결과에서는 상관관계가 크지 않았다.
05st에서는 SOD와 NH4+ flux의 변동이 유사하였으나, 06st에서는 상관관계가 거의 나
타나지 않았다. 특히 SOD가 높아지는 4월과 8월 높은 NH4+의 용출이 예상되었으나,
오히려 퇴적층으로 소모되거나, 값이 작은 용출 flux가 나타났다. 이는 SOD와
Ammonification(암모니아화)과 연결되어 NH4+를 전자수용체로 사용하는 질산화박테리
아의 활동이 활발했거나, 높아진 광량으로 인해 갯벌 퇴적물 표층에 서식하는 미세저서
조류의 광합성 활동이 활발해져 이러한 결과가 나타난 것으로 생각된다.
인공갯벌의 조고에 따른 NH4+ flux를 나타내면 Fig. 3-9와 같다. 2005년에 만들어진

- 171 -
05 station �H4+ Flux by Grain size
-4
-2
0
2
4
6
8
10
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
�H
4+ Flux (mmole · m
-2 · d-1)
Sand
Original
Mud
06 station �H4+ Flux by Grain size
-4
-2
0
2
4
6
8
10
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
�H
4+ Flux (mmole · m
-2 · d-1)
Sand
Original
Mud
Figure. 3-10. Variation of NH4+ flux by grain size in artificial tidal
flats(2005, 2006) at Nak-Dong estuary.인공갯벌에서 3개의 조고 중 높은 NH4
+ flux 값(용출)이 나타난 곳은 MIDDLE과
LOWER station이었고, 낮은 NH4+ flux 값(소비)이 나타난 곳은 UPPER station이었다.
2006년에 만든 인공갯벌에서도 같은 경향을 보였다.
인공갯벌의 퇴적물 type에 따른 NH4+ flux는 Fig. 3-10과 같다. 2005년에 만들어진
인공갯벌에서 3개의 조고 중 높은 NH4+ flux(용출) 값이 나타난 곳은 Original 퇴적물
이었고, 낮은 NH4+ flux 값(소비)이 나타난 곳은 sand 퇴적물이었다. 2006년에 만든 인
공갯벌에서도 같은 경향을 보였다.
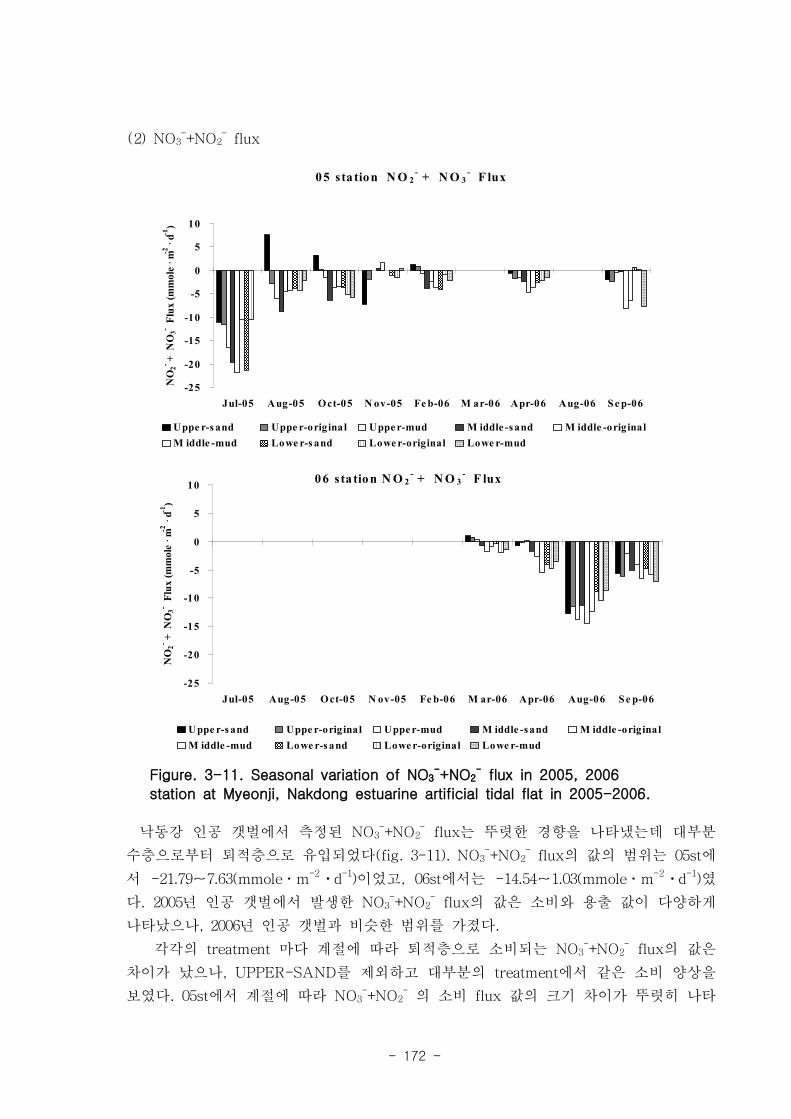
- 172 -
05 sta tion �O 2- + �O 3
- F lux
-25
-20
-15
-10
-5
0
5
10
Jul-05 Aug-05 Oct-05 � ov-05 Feb-06 M ar-06 Apr-06 Aug-06 Sep-06
�O
2- + �O
3- Flux (mmole · m
-2 · d-1
)
Uppe r-s and Uppe r-original Uppe r-mud M iddle -sand M iddle -original
M iddle -mud Lowe r-s and Lower-original Lowe r-mud
06 sta tion �O 2- + �O 3
- F lux
-25
-20
-15
-10
-5
0
5
10
Jul-05 Aug-05 Oct-05 � ov-05 Fe b-06 M ar-06 Apr-06 Aug-06 Se p-06
�O
2- + �O
3- Flux (mmole · m
-2 · d-1)
Uppe r-s and Uppe r-original Uppe r-mud M iddle -sand M iddle -original
M iddle -mud Lowe r-s and Lower-original Lowe r-mud
Figure. 3-11. Seasonal variation of NO3-+NO2
- flux in 2005, 2006
station at Myeonji, Nakdong estuarine artificial tidal flat in 2005-2006.
(2) NO3-+NO2
- flux
낙동강 인공 갯벌에서 측정된 NO3-+NO2
- flux는 뚜렷한 경향을 나타냈는데 대부분
수층으로부터 퇴적층으로 유입되었다(fig. 3-11). NO3-+NO2
- flux의 값의 범위는 05st에
서 -21.79~7.63(mmoleㆍm-2
• d-1
)이었고, 06st에서는 -14.54~1.03(mmoleㆍm-2
• d-1
)였
다. 2005년 인공 갯벌에서 발생한 NO3-+NO2
- flux의 값은 소비와 용출 값이 다양하게
나타났으나, 2006년 인공 갯벌과 비슷한 범위를 가졌다.
각각의 treatment 마다 계절에 따라 퇴적층으로 소비되는 NO3-+NO2
- flux의 값은
차이가 났으나, UPPER-SAND를 제외하고 대부분의 treatment에서 같은 소비 양상을
보였다. 05st에서 계절에 따라 NO3-+NO2
- 의 소비 flux 값의 크기 차이가 뚜렷히 나타

- 173 -
났다. 여름인 7월에 NO3-+NO2
- 의 소비 flux 값이 가장 크게 나타났으며, 가을, 봄과
겨울 순서로 퇴적층으로 소비되는 경향이 나타났다. 06st의 경우도 마찬가지 경향이 나
타났으며, 05st의 소비 flux값보다는 약간 작게 발생되었다.
이러한 flux의 값은 대부분의 treatments에서 나타났는데, 이는 퇴적물내 NO3-+NO2
-
농도보다 수층의 NO3-+NO2
- 농도가 더 높아 수층으로부터 퇴적물로의 이동(확산)을 의
미하며, 탈질소화(denitrification)의 source가 되는 NO3-+NO2
- 의 퇴적물로의 공급이 매
우 활발하여 잠재적 탈질소화를 높게 예상해 볼 수 있다. 또 한편, 수층에서의
NO3-+NO2
- 의 공급이 많다면, 퇴적물 내에서 미생물이 NO3-+NO2
- 를 생성하는 과정인
질산화(nitrification)과정이 제한되었음도 예상할 수 있다.
대부분의 treatments에서 소비 경향이 나타났지만, 그 반대의 flux도 종종 나타남을
확인할 수 있는데, 특히 UPPER-SAND에서는 4계절 모두 NO3-+NO2
- 의 용출 flux 값
이 나타났다. 여름철 낙동강 담수의 유입으로 수층의 NO3-+NO2
- 의 농도가 높음에도
불구하고 퇴적층으로부터의 용출이 발생하였는데, 이는 여러 가지 상황을 예측해 볼 수
있다. UPPER-SAND는 퇴적물의 공극이 매우 커 퇴적물로의 산소공급이 활발하여 유
기물 분해(SOD)에 따른 암모니아의 재생산 후 다시 표층 퇴적물내에서 질산화 과정과
의 coupling(연결)으로 인해 NO3-+NO2
- 가 퇴적물내 많이 축적되어졌으나, 퇴적물내
denitrifier(탈질소화 미생물)의 분포가 적어 NO3-+NO2
- 를 소모하지 않아 퇴적물 밖으
로 확산된 것으로 예상할 수 있다. 실제로 UPPER-SAND는 거의 SAND 로 구성되어
있었으며, 퇴적물 코어를 채취하였을 때도 육안으로 혐기층을 찾아볼 수 없었다. 이는
탈질소화 미생물이 살 수 있는 환경인 혐기층이 존재하지 않았기 때문에 탈질소화가
거의 발생하지 않아 NO3-+NO2
- 가 용출되었다고 추측할 수 있다.
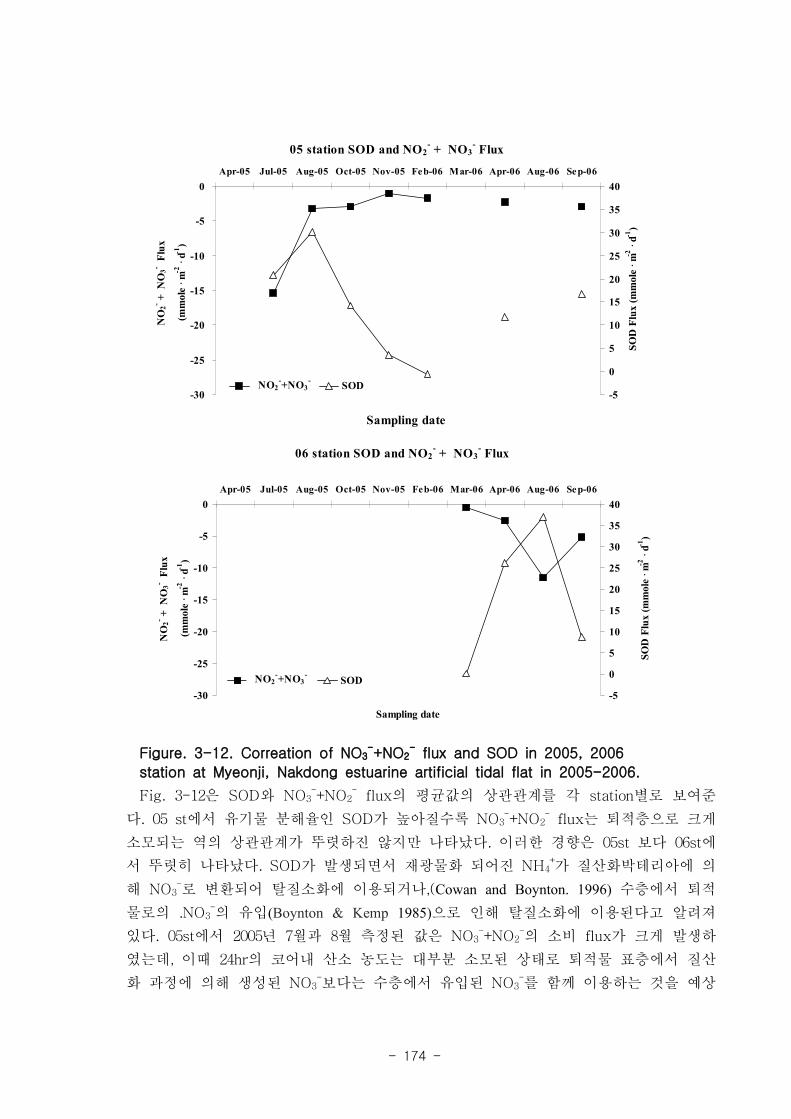
- 174 -
05 station SOD and �O2- + �O3
- Flux
-30
-25
-20
-15
-10
-5
0
Apr-05 Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
�O
2- + �O
3- Flux
(mmole · m
-2 · d
-1)
-5
0
5
10
15
20
25
30
35
40
SOD Flux (mmole · m
-2 · d
-1)
�O2- + �O3- SOD�O2-+�O3
-
06 station SOD and �O2- + �O3
- Flux
-30
-25
-20
-15
-10
-5
0
Apr-05 Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
�O
2- + �O
3- Flux
(mmole · m
-2 · d
-1)
-5
0
5
10
15
20
25
30
35
40
SOD Flux (mmole · m
-2 · d
-1)
�O2- + �O3- SOD�O2-+�O3
-
Figure. 3-12. Correation of NO3-+NO2
- flux and SOD in 2005, 2006
station at Myeonji, Nakdong estuarine artificial tidal flat in 2005-2006.
Fig. 3-12은 SOD와 NO3-+NO2
- flux의 평균값의 상관관계를 각 station별로 보여준
다. 05 st에서 유기물 분해율인 SOD가 높아질수록 NO3-+NO2
- flux는 퇴적층으로 크게
소모되는 역의 상관관계가 뚜렷하진 않지만 나타났다. 이러한 경향은 05st 보다 06st에
서 뚜렷히 나타났다. SOD가 발생되면서 재광물화 되어진 NH4+가 질산화박테리아에 의
해 NO3-로 변환되어 탈질소화에 이용되거나,(Cowan and Boynton. 1996) 수층에서 퇴적
물로의 .NO3-의 유입(Boynton & Kemp 1985)으로 인해 탈질소화에 이용된다고 알려져
있다. 05st에서 2005년 7월과 8월 측정된 값은 NO3-+NO2
-의 소비 flux가 크게 발생하
였는데, 이때 24hr의 코어내 산소 농도는 대부분 소모된 상태로 퇴적물 표층에서 질산
화 과정에 의해 생성된 NO3-보다는 수층에서 유입된 NO3
-를 함께 이용하는 것을 예상
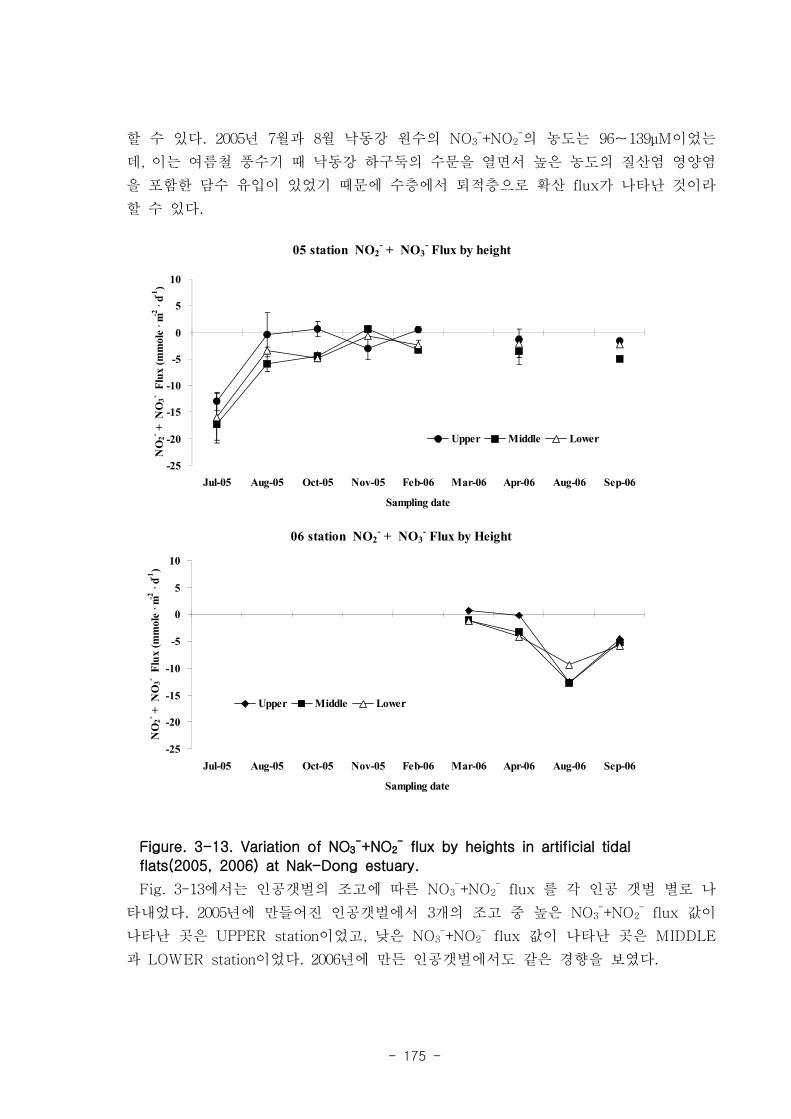
- 175 -
할 수 있다. 2005년 7월과 8월 낙동강 원수의 NO3-+NO2
-의 농도는 96~139μM이었는
데, 이는 여름철 풍수기 때 낙동강 하구둑의 수문을 열면서 높은 농도의 질산염 영양염
을 포함한 담수 유입이 있었기 때문에 수층에서 퇴적층으로 확산 flux가 나타난 것이라
할 수 있다.
05 station �O2- + �O3
- Flux by height
-25
-20
-15
-10
-5
0
5
10
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
�O
2- + �O
3- Flux (mmole · m
-2 · d-
1)
Upper Middle Lower
06 station �O2- + �O3
- Flux by Height
-25
-20
-15
-10
-5
0
5
10
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
�O
2- + �O
3- Flux (mmole · m
-2 · d
-1)
Upper Middle Lower
Figure. 3-13. Variation of NO3-+NO2
- flux by heights in artificial tidal
flats(2005, 2006) at Nak-Dong estuary.
Fig. 3-13에서는 인공갯벌의 조고에 따른 NO3-+NO2
- flux 를 각 인공 갯벌 별로 나
타내었다. 2005년에 만들어진 인공갯벌에서 3개의 조고 중 높은 NO3-+NO2
- flux 값이
나타난 곳은 UPPER station이었고, 낮은 NO3-+NO2
- flux 값이 나타난 곳은 MIDDLE
과 LOWER station이었다. 2006년에 만든 인공갯벌에서도 같은 경향을 보였다.
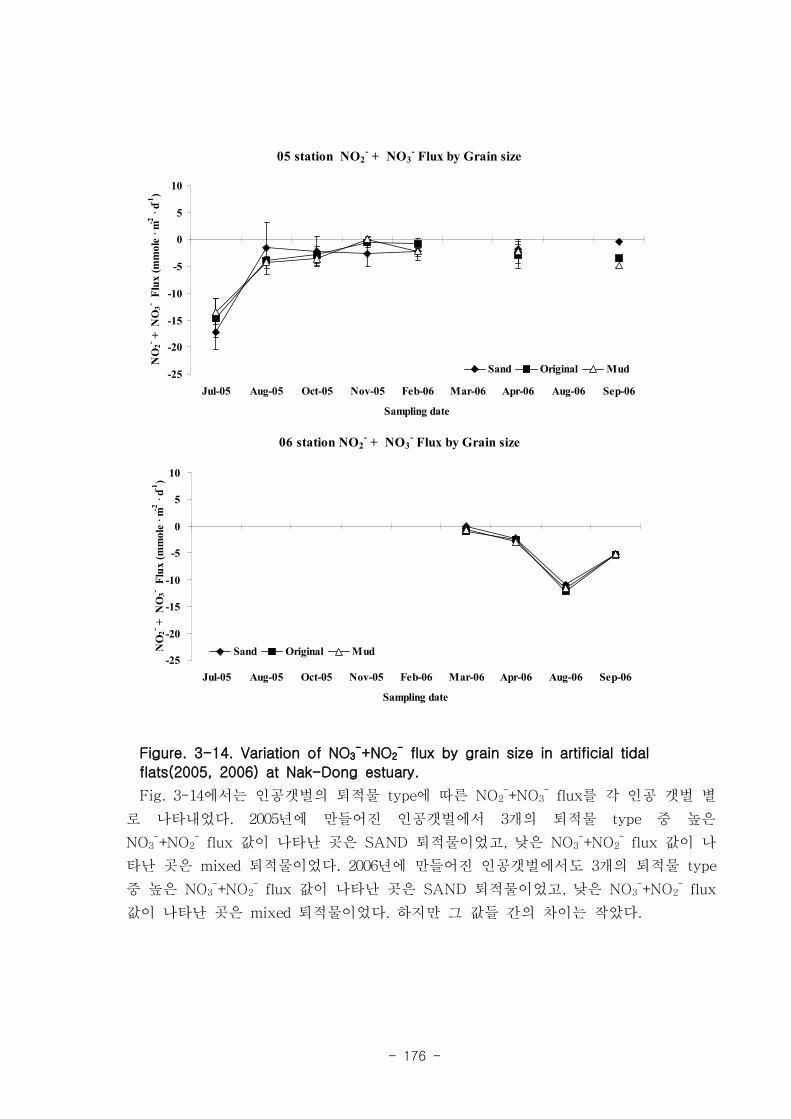
- 176 -
05 station �O2- + �O3
- Flux by Grain size
-25
-20
-15
-10
-5
0
5
10
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
�O
2- + �O
3- Flux (mmole · m
-2 · d
-1)
Sand Original Mud
06 station �O2- + �O3
- Flux by Grain size
-25
-20
-15
-10
-5
0
5
10
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
�O
2- + �O
3- Flux (mmole · m
-2 · d
-1)
Sand Original Mud
Figure. 3-14. Variation of NO3-+NO2
- flux by grain size in artificial tidal
flats(2005, 2006) at Nak-Dong estuary.
Fig. 3-14에서는 인공갯벌의 퇴적물 type에 따른 NO2-+NO3
- flux를 각 인공 갯벌 별
로 나타내었다. 2005년에 만들어진 인공갯벌에서 3개의 퇴적물 type 중 높은
NO3-+NO2
- flux 값이 나타난 곳은 SAND 퇴적물이었고, 낮은 NO3-+NO2
- flux 값이 나
타난 곳은 mixed 퇴적물이었다. 2006년에 만들어진 인공갯벌에서도 3개의 퇴적물 type
중 높은 NO3-+NO2
- flux 값이 나타난 곳은 SAND 퇴적물이었고, 낮은 NO3-+NO2
- flux
값이 나타난 곳은 mixed 퇴적물이었다. 하지만 그 값들 간의 차이는 작았다.
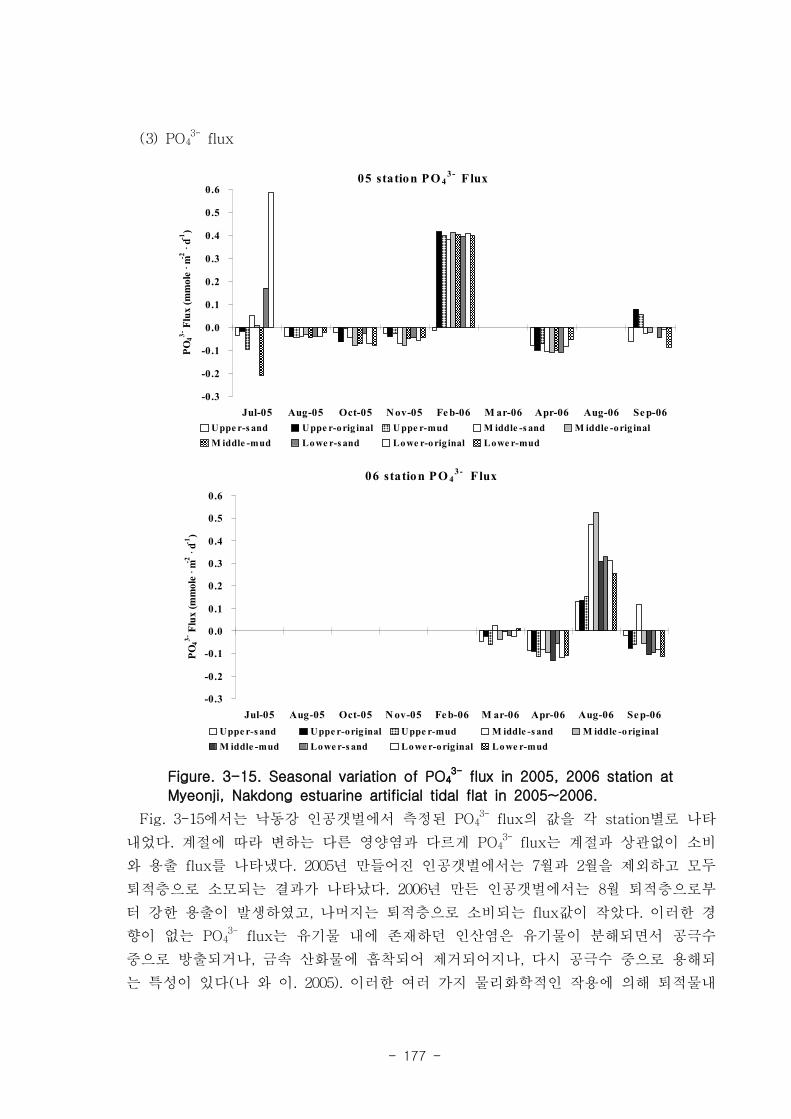
- 177 -
(3) PO43- flux
05 station PO 43- F lux
-0.3
-0.2
-0.1
0.0
0.1
0.2
0.3
0.4
0.5
0.6
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 M ar-06 Apr-06 Aug-06 Sep-06
PO
43- Flux (mmole · m
-2 · d
-1)
Uppe r-s and Uppe r-orig inal Uppe r-mud M iddle -s and M iddle -orig inal
M iddle -mud Lowe r-s and Lowe r-orig inal Lowe r-mud
06 station PO 43- Flux
-0.3
-0.2
-0.1
0.0
0.1
0.2
0.3
0.4
0.5
0.6
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 M ar-06 Apr-06 Aug-06 Sep-06
PO43- Flux (mmole · m
-2 · d-1)
Uppe r-s and Uppe r-orig inal Uppe r-mud M iddle -s and M iddle -original
M iddle -mud Lowe r-s and Lowe r-original Lowe r-mud
Figure. 3-15. Seasonal variation of PO43- flux in 2005, 2006 station at
Myeonji, Nakdong estuarine artificial tidal flat in 2005~2006.
Fig. 3-15에서는 낙동강 인공갯벌에서 측정된 PO43-
flux의 값을 각 station별로 나타
내었다. 계절에 따라 변하는 다른 영양염과 다르게 PO43- flux는 계절과 상관없이 소비
와 용출 flux를 나타냈다. 2005년 만들어진 인공갯벌에서는 7월과 2월을 제외하고 모두
퇴적층으로 소모되는 결과가 나타났다. 2006년 만든 인공갯벌에서는 8월 퇴적층으로부
터 강한 용출이 발생하였고, 나머지는 퇴적층으로 소비되는 flux값이 작았다. 이러한 경
향이 없는 PO43-
flux는 유기물 내에 존재하던 인산염은 유기물이 분해되면서 공극수
중으로 방출되거나, 금속 산화물에 흡착되어 제거되어지나, 다시 공극수 중으로 용해되
는 특성이 있다(나 와 이. 2005). 이러한 여러 가지 물리화학적인 작용에 의해 퇴적물내
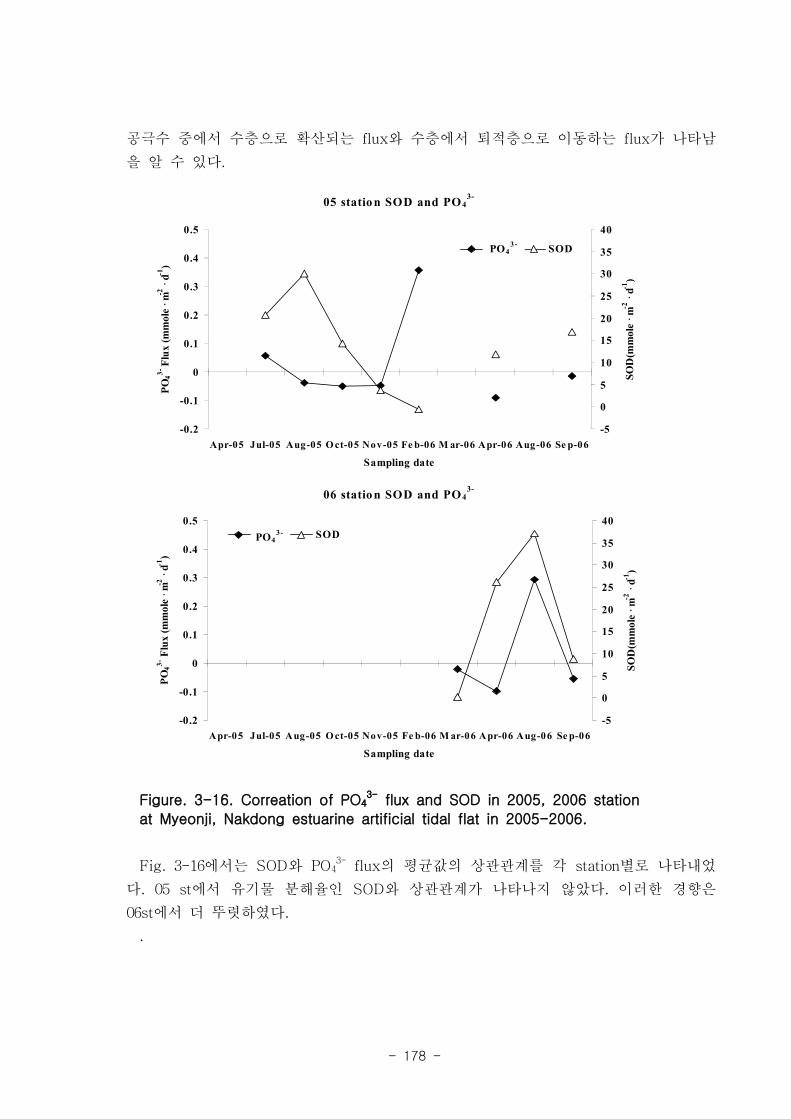
- 178 -
공극수 중에서 수층으로 확산되는 flux와 수층에서 퇴적층으로 이동하는 flux가 나타남
을 알 수 있다.
05 station SOD and PO43-
-0.2
-0.1
0
0.1
0.2
0.3
0.4
0.5
Apr-05 Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 M ar-06 Apr-06 Aug-06 Se p-06
Sampling date
PO
43- Flux (mmole · m
-2 · d
-1)
-5
0
5
10
15
20
25
30
35
40
SOD(m
mole · m
-2 · d
-1)
PO43- SODPO43-
06 station SOD and PO43-
-0.2
-0.1
0
0.1
0.2
0.3
0.4
0.5
Apr-05 Jul-05 Aug-05 Oct-05 �ov-05 Fe b-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
PO
43- Flux (mmole · m
-2 · d
-1)
-5
0
5
10
15
20
25
30
35
40
SOD(m
mole · m
-2 · d
-1)
PO43- SODPO43-
Figure. 3-16. Correation of PO43- flux and SOD in 2005, 2006 station
at Myeonji, Nakdong estuarine artificial tidal flat in 2005-2006.
Fig. 3-16에서는 SOD와 PO43- flux의 평균값의 상관관계를 각 station별로 나타내었
다. 05 st에서 유기물 분해율인 SOD와 상관관계가 나타나지 않았다. 이러한 경향은
06st에서 더 뚜렷하였다.
.
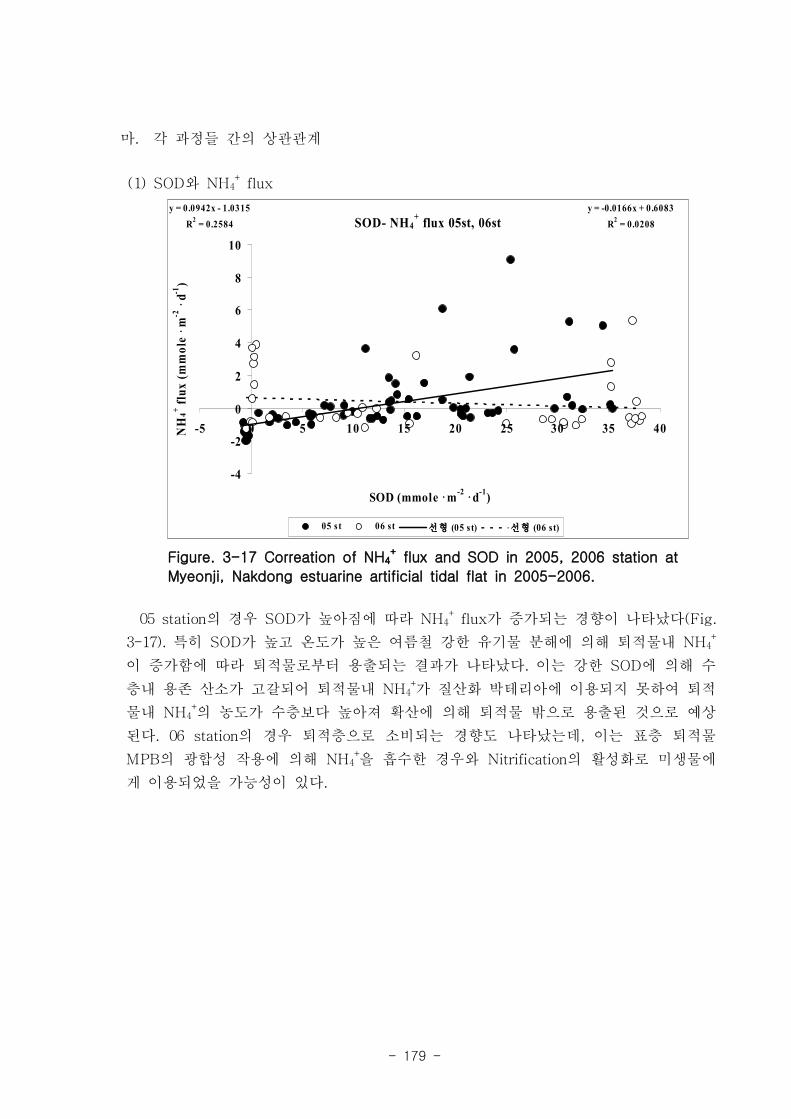
- 179 -
마. 각 과정들 간의 상관관계
(1) SOD와 NH4+ flux
SOD- �H4+ flux 05st, 06st
y = 0.0942x - 1.0315
R2 = 0.2584
y = -0.0166x + 0.6083
R2 = 0.0208
-4
-2
0
2
4
6
8
10
-5 0 5 10 15 20 25 30 35 40
SOD (mmole · m-2 · d
-1)
�H
4+ flux (mmole · m
-2 · d
-1)
05 s t 06 st 선형 (05 s t) 선형 (06 st)
Figure. 3-17 Correation of NH4+ flux and SOD in 2005, 2006 station at
Myeonji, Nakdong estuarine artificial tidal flat in 2005-2006.
05 station의 경우 SOD가 높아짐에 따라 NH4+ flux가 증가되는 경향이 나타났다(Fig.
3-17). 특히 SOD가 높고 온도가 높은 여름철 강한 유기물 분해에 의해 퇴적물내 NH4+
이 증가함에 따라 퇴적물로부터 용출되는 결과가 나타났다. 이는 강한 SOD에 의해 수
층내 용존 산소가 고갈되어 퇴적물내 NH4+가 질산화 박테리아에 이용되지 못하여 퇴적
물내 NH4+의 농도가 수층보다 높아져 확산에 의해 퇴적물 밖으로 용출된 것으로 예상
된다. 06 station의 경우 퇴적층으로 소비되는 경향도 나타났는데, 이는 표층 퇴적물
MPB의 광합성 작용에 의해 NH4+을 흡수한 경우와 Nitrification의 활성화로 미생물에
게 이용되었을 가능성이 있다.
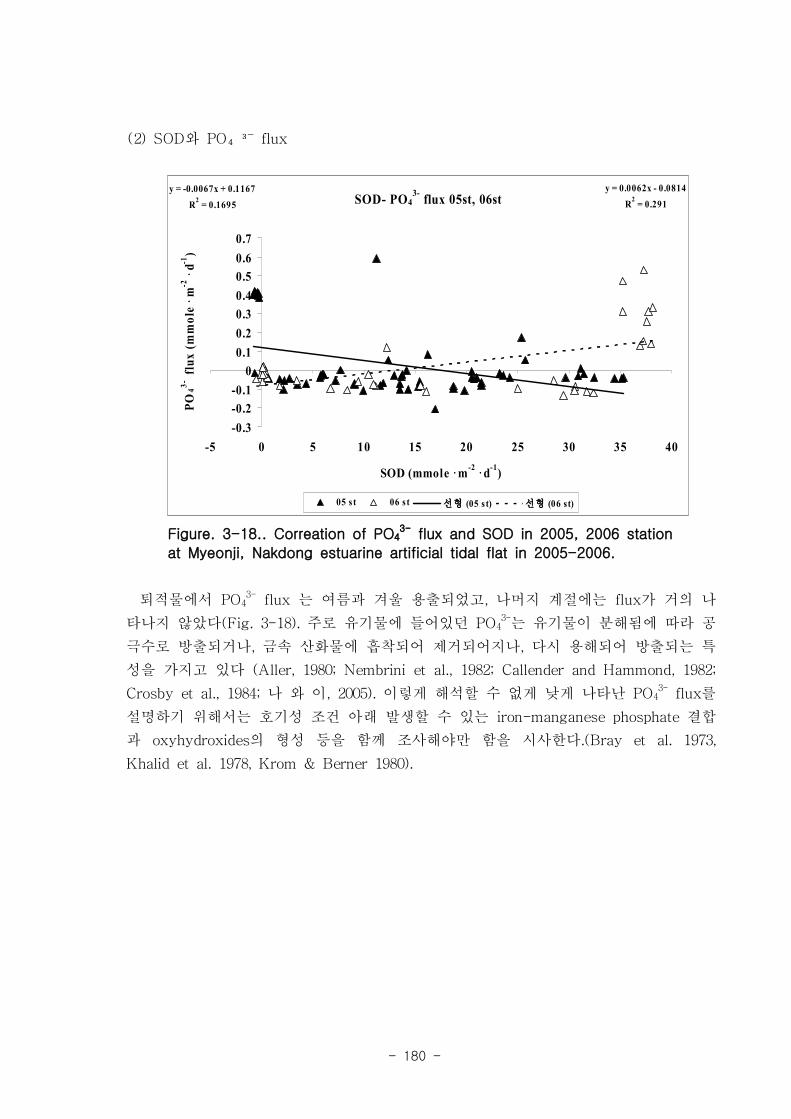
- 180 -
(2) SOD와 PO₄³⁻ flux
SOD- PO43- flux 05st, 06st
y = -0.0067x + 0.1167
R2 = 0.1695
y = 0.0062x - 0.0814
R2 = 0.291
-0.3
-0.2
-0.1
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
-5 0 5 10 15 20 25 30 35 40
SOD (mmole · m-2 · d
-1)
PO
43- flux (mmole · m
-2 · d
-1)
05 s t 06 s t 선형 (05 s t) 선형 (06 st)
Figure. 3-18.. Correation of PO43- flux and SOD in 2005, 2006 station
at Myeonji, Nakdong estuarine artificial tidal flat in 2005-2006.
퇴적물에서 PO43- flux 는 여름과 겨울 용출되었고, 나머지 계절에는 flux가 거의 나
타나지 않았다(Fig. 3-18). 주로 유기물에 들어있던 PO43-는 유기물이 분해됨에 따라 공
극수로 방출되거나, 금속 산화물에 흡착되어 제거되어지나, 다시 용해되어 방출되는 특
성을 가지고 있다 (Aller, 1980; Nembrini et al., 1982; Callender and Hammond, 1982;
Crosby et al., 1984; 나 와 이, 2005). 이렇게 해석할 수 없게 낮게 나타난 PO43- flux를
설명하기 위해서는 호기성 조건 아래 발생할 수 있는 iron-manganese phosphate 결합
과 oxyhydroxides의 형성 등을 함께 조사해야만 함을 시사한다.(Bray et al. 1973,
Khalid et al. 1978, Krom & Berner 1980).
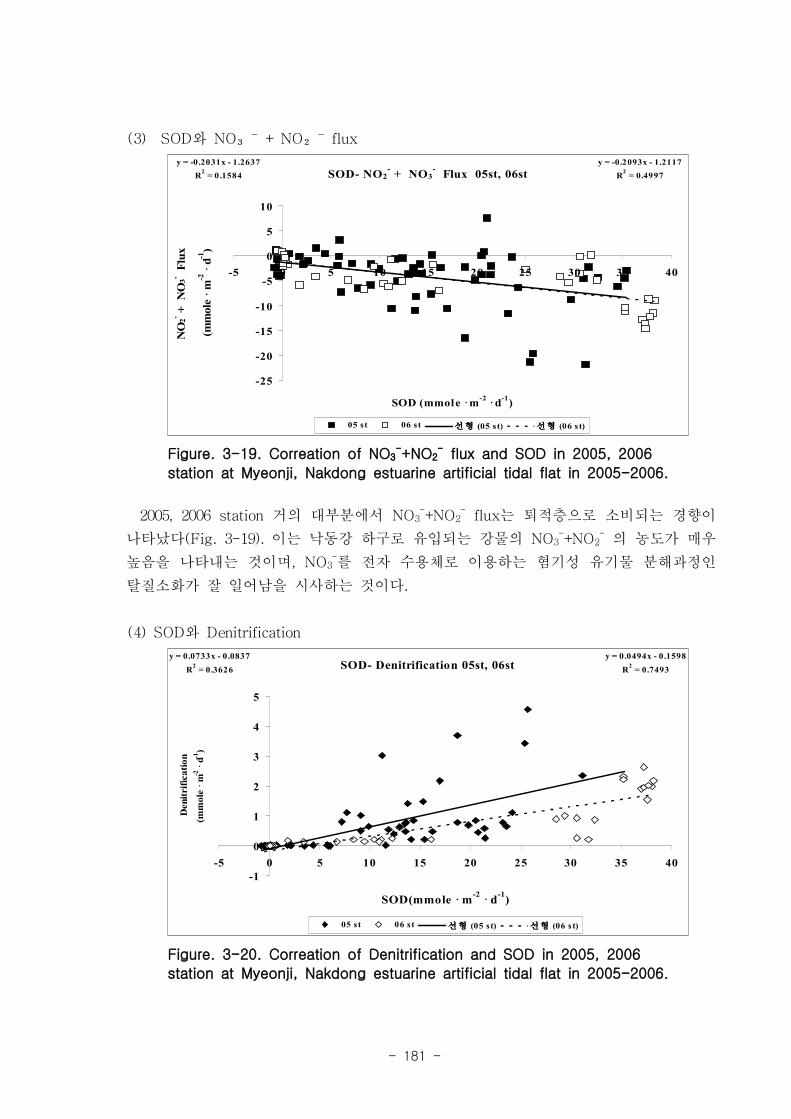
- 181 -
(3) SOD와 NO₃⁻ + NO₂⁻ flux
SOD- �O2- + �O3
- Flux 05st, 06st
y = -0.2031x - 1.2637
R2 = 0.1584
y = -0.2093x - 1.2117
R2 = 0.4997
-25
-20
-15
-10
-5
0
5
10
-5 0 5 10 15 20 25 30 35 40
SOD (mmole · m-2 · d
-1)
�O
2- + �O
3- Flux
(mmole · m
-2 · d
-1)
05 st 06 st 선형 (05 st) 선형 (06 s t)
Figure. 3-19. Correation of NO3-+NO2
- flux and SOD in 2005, 2006
station at Myeonji, Nakdong estuarine artificial tidal flat in 2005-2006.
2005, 2006 station 거의 대부분에서 NO3-+NO2
- flux는 퇴적층으로 소비되는 경향이
나타났다(Fig. 3-19). 이는 낙동강 하구로 유입되는 강물의 NO3-+NO2
- 의 농도가 매우
높음을 나타내는 것이며, NO3-를 전자 수용체로 이용하는 혐기성 유기물 분해과정인
탈질소화가 잘 일어남을 시사하는 것이다.
(4) SOD와 Denitrification
SOD- Denitrification 05st, 06sty = 0.0733x - 0.0837
R2 = 0.3626
y = 0.0494x - 0.1598
R2 = 0.7493
-1
0
1
2
3
4
5
-5 0 5 10 15 20 25 30 35 40
SOD(mmole · m-2 · d
-1)
Denitrification
(mmole · m
-2 · d
-1)
05 st 06 s t 선형 (05 st) 선형 (06 s t)
Figure. 3-20. Correation of Denitrification and SOD in 2005, 2006
station at Myeonji, Nakdong estuarine artificial tidal flat in 2005-2006.
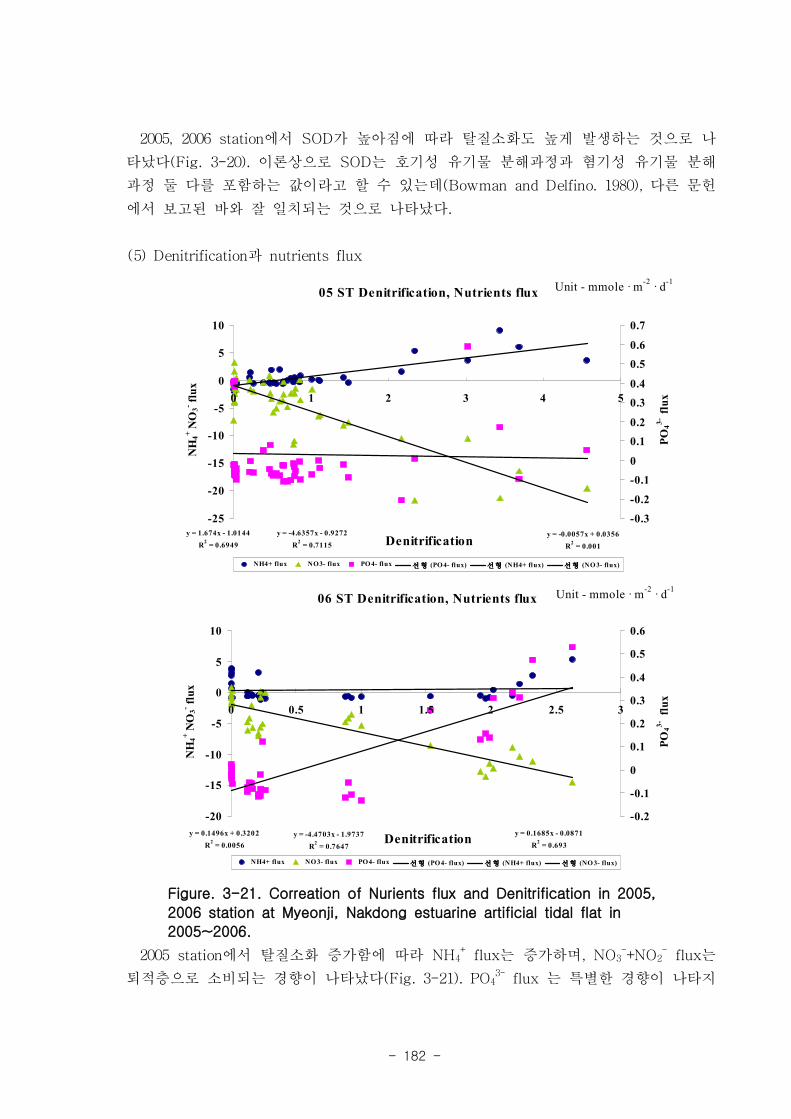
- 182 -
2005, 2006 station에서 SOD가 높아짐에 따라 탈질소화도 높게 발생하는 것으로 나
타났다(Fig. 3-20). 이론상으로 SOD는 호기성 유기물 분해과정과 혐기성 유기물 분해
과정 둘 다를 포함하는 값이라고 할 수 있는데(Bowman and Delfino. 1980), 다른 문헌
에서 보고된 바와 잘 일치되는 것으로 나타났다.
(5) Denitrification과 nutrients flux
05 ST Denitrification, �utrients flux
y = -0.0057x + 0.0356
R2 = 0.001
y = 1.674x - 1.0144
R2 = 0.6949
y = -4.6357x - 0.9272
R2 = 0.7115
-25
-20
-15
-10
-5
0
5
10
0 1 2 3 4 5
Denitrification
�H
4+ �O
3- flux
-0.3
-0.2
-0.1
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
PO
43- flux
�H4+ flux �O3- flux PO4- flux 선 형 (PO4- flux) 선형 (�H4+ flux) 선형 (�O3- flux)
Unit - mmole · m-2 · d
-1
06 ST Denitrification, �utrients flux
y = 0.1685x - 0.0871
R2 = 0.693
y = 0.1496x + 0.3202
R2 = 0.0056
y = -4.4703x - 1.9737
R2 = 0.7647
-20
-15
-10
-5
0
5
10
0 0.5 1 1.5 2 2.5 3
Denitrification
�H
4+ �O
3- flux
-0.2
-0.1
0
0.1
0.2
0.3
0.4
0.5
0.6
PO
43- flux
�H4+ flux �O3- flux PO4- flux 선형 (PO4- flux) 선 형 (�H4+ flux) 선형 (�O3- flux)
Unit - mmole · m-2 · d
-1
Figure. 3-21. Correation of Nurients flux and Denitrification in 2005,
2006 station at Myeonji, Nakdong estuarine artificial tidal flat in
2005~2006.
2005 station에서 탈질소화 증가함에 따라 NH4+ flux는 증가하며, NO3
-+NO2
- flux는
퇴적층으로 소비되는 경향이 나타났다(Fig. 3-21). PO43- flux 는 특별한 경향이 나타지

- 183 -
않았다.
2005 station과 다르게 2006 station에서 탈질소화 증가함에 따라 PO43- flux 가 증가
하는 경향이 나타났다(Fig. 3-21). 이는 혐기성 유기물 분해과정의 하나인 탈질소화에
의해 유기물이 분해되면서 PO43-가 공극수내로 방출되어 퇴적물내 농도가 높아져 확산
에 의해 퇴적층에서 수층으로 이동된 것으로 생각된다. NH4+
flux는 특별한 경향이 없
었다.
(6) SOD, Denitrification, NH4+ flux, NO3
-+NO2
- flux
(가) Data 분석
낙동강 하구 인공갯벌에서 유기물 분해율과 탈질소화가 어떠한 속도로 발생되며, N
(질소 성분) cycling 과의 상관관계를 파악하고, 이들에게 영향을 미치는 요인이 무엇인
지 알기 위해서 각 flux data를 분석하였다. SOD, Denitrification, NH4+ flux,
NO3-+NO2
- flux 를 2005년 7월부터 2006년 9월까지 각 계절별로 2개의 인공갯벌(05,
06 station - 각 9개 treatments)에서 측정하였고, station에 따라 분석하였다.
SOD, Denitrification은 각 station에서 사계절 모두를 측정하여 사계절 각 평균값을
기준으로 하여 높고 낮음(↑,↓)을 설정하였고, NH4+ flux, NO3
-+NO2
- flux 는 계절별
평균값은 기준으로 부적합하여, 값의 양수와 음수 즉, 용출과 소비(↑,↓)를 기준으로
하였다.
위 기준으로 SOD, Denitrification, NH4+ flux, NO3
-+NO2
- flux 값을 가지고 나누어
총 16가지의 경우가 나왔으며, 16가지 조건에 맞추어 4가지 값들을 선택하였다. 총 16
가지 경우들 중 비율을 계산하여 높은 비율을 가지는 story( F type-13.42%, K
type-39.93% , L type-13.42% , N type-13.42% )를 선택하여 분석하였다(Fig. 3-22~
25, Table 3-2 ~ 4).
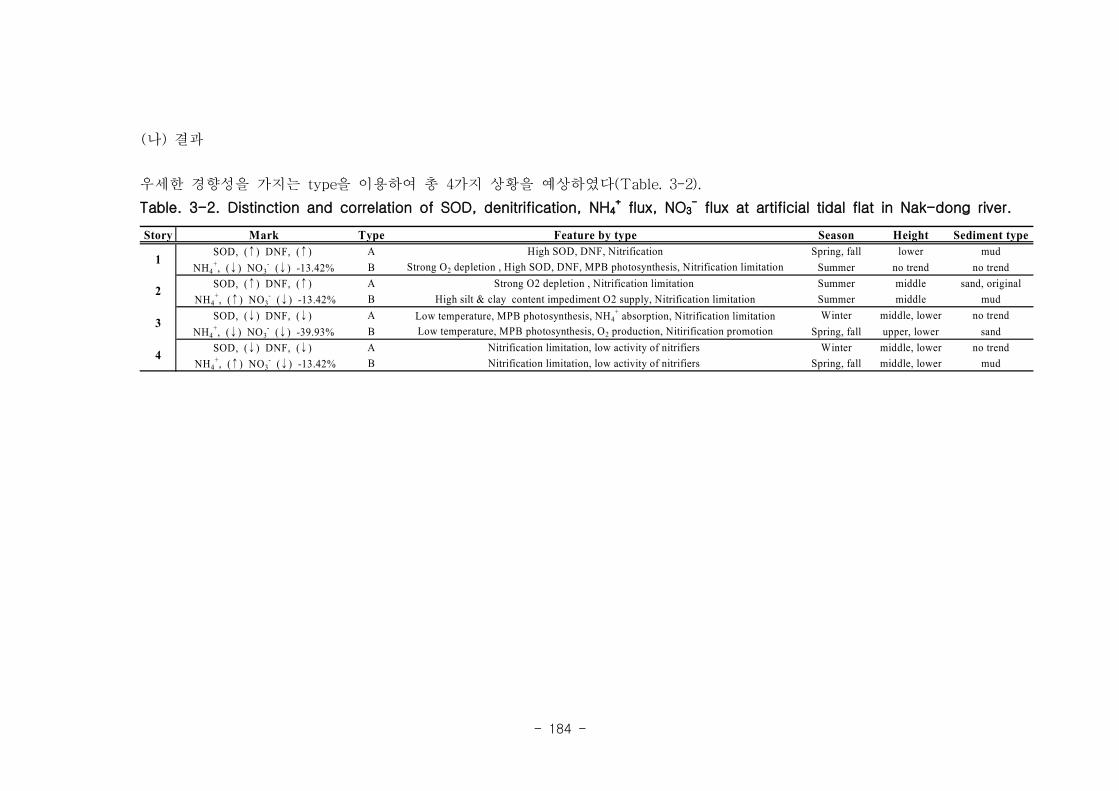
- 184 -
(나) 결과
우세한 경향성을 가지는 type을 이용하여 총 4가지 상황을 예상하였다(Table. 3-2).
Story Mark Type Feature by type Season Height Sediment type
SOD, (↑) DNF, (↑) A High SOD, DNF, Nitrification Spring, fall lower mud
NH4+, (↓) NO3
- (↓) -13.42% B Strong O2 depletion , High SOD, DNF, MPB photosynthesis, Nitrification limitation Summer no trend no trend
SOD, (↑) DNF, (↑) A Strong O2 depletion , Nitrification limitation Summer middle sand, original
NH4+, (↑) NO3
- (↓) -13.42% B High silt & clay content impediment O2 supply, Nitrification limitation Summer middle mud
SOD, (↓) DNF, (↓) A Low temperature, MPB photosynthesis, NH4+ absorption, Nitrification limitation Winter middle, lower no trend
NH4+, (↓) NO3
- (↓) -39.93% B Low temperature, MPB photosynthesis, O2 production, Nitirification promotion Spring, fall upper, lower sand
SOD, (↓) DNF, (↓) A Nitrification limitation, low activity of nitrifiers Winter middle, lower no trend
NH4+, (↑) NO3
- (↓) -13.42% B Nitrification limitation, low activity of nitrifiers Spring, fall middle, lower mud
1
2
3
4
Table. 3-2. Distinction and correlation of SOD, denitrification, NH4+ flux, NO3
- flux at artificial tidal flat in Nak-dong river.
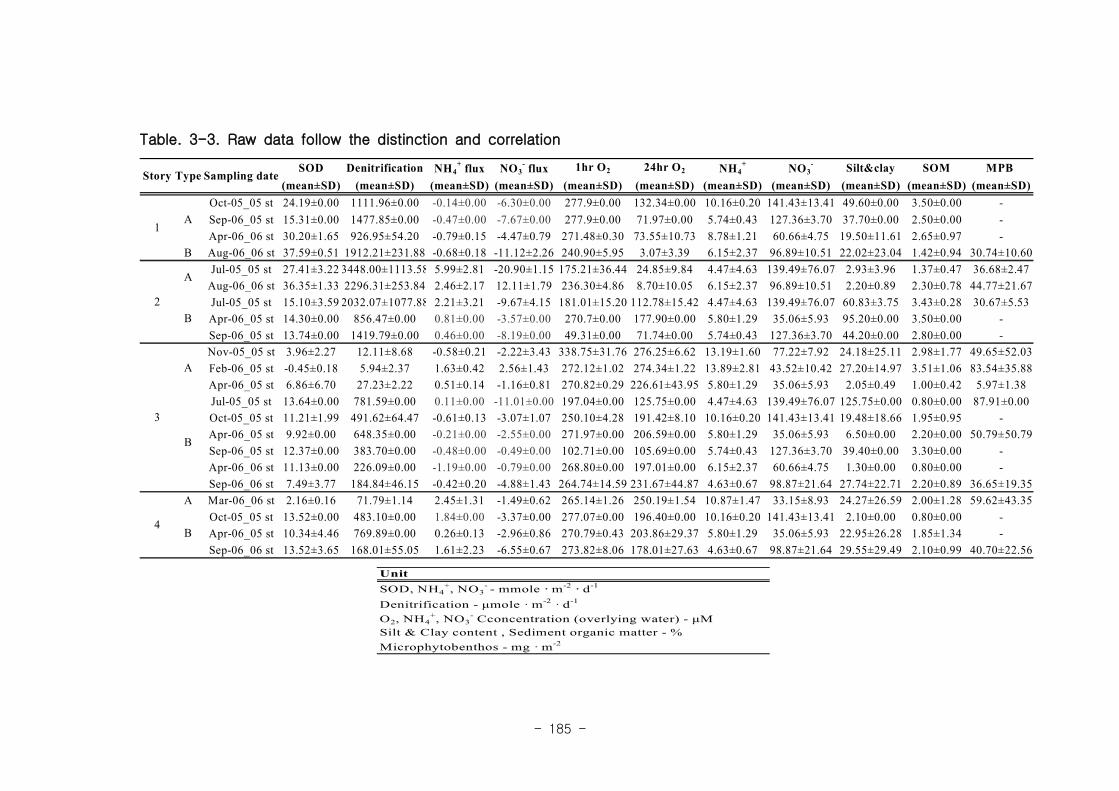
- 185 -
SOD Denitrification �H4+ flux �O3
- flux 1hr O2 24hr O2 �H4
+�O3
-Silt&clay SOM MPB
(mean±SD) (mean±SD) (mean±SD) (mean±SD) (mean±SD) (mean±SD) (mean±SD) (mean±SD) (mean±SD) (mean±SD) (mean±SD)
Oct-05_05 st 24.19±0.00 1111.96±0.00 -0.14±0.00 -6.30±0.00 277.9±0.00 132.34±0.00 10.16±0.20 141.43±13.41 49.60±0.00 3.50±0.00 -
Sep-06_05 st 15.31±0.00 1477.85±0.00 -0.47±0.00 -7.67±0.00 277.9±0.00 71.97±0.00 5.74±0.43 127.36±3.70 37.70±0.00 2.50±0.00 -
Apr-06_06 st 30.20±1.65 926.95±54.20 -0.79±0.15 -4.47±0.79 271.48±0.30 73.55±10.73 8.78±1.21 60.66±4.75 19.50±11.61 2.65±0.97 -
B Aug-06_06 st 37.59±0.51 1912.21±231.88 -0.68±0.18 -11.12±2.26 240.90±5.95 3.07±3.39 6.15±2.37 96.89±10.51 22.02±23.04 1.42±0.94 30.74±10.60
Jul-05_05 st 27.41±3.22 3448.00±1113.58 5.99±2.81 -20.90±1.15 175.21±36.44 24.85±9.84 4.47±4.63 139.49±76.07 2.93±3.96 1.37±0.47 36.68±2.47
Aug-06_06 st 36.35±1.33 2296.31±253.84 2.46±2.17 12.11±1.79 236.30±4.86 8.70±10.05 6.15±2.37 96.89±10.51 2.20±0.89 2.30±0.78 44.77±21.67
Jul-05_05 st 15.10±3.59 2032.07±1077.88 2.21±3.21 -9.67±4.15 181.01±15.20 112.78±15.42 4.47±4.63 139.49±76.07 60.83±3.75 3.43±0.28 30.67±5.53
Apr-06_05 st 14.30±0.00 856.47±0.00 0.81±0.00 -3.57±0.00 270.7±0.00 177.90±0.00 5.80±1.29 35.06±5.93 95.20±0.00 3.50±0.00 -
Sep-06_05 st 13.74±0.00 1419.79±0.00 0.46±0.00 -8.19±0.00 49.31±0.00 71.74±0.00 5.74±0.43 127.36±3.70 44.20±0.00 2.80±0.00 -
Nov-05_05 st 3.96±2.27 12.11±8.68 -0.58±0.21 -2.22±3.43 338.75±31.76 276.25±6.62 13.19±1.60 77.22±7.92 24.18±25.11 2.98±1.77 49.65±52.03
Feb-06_05 st -0.45±0.18 5.94±2.37 1.63±0.42 2.56±1.43 272.12±1.02 274.34±1.22 13.89±2.81 43.52±10.42 27.20±14.97 3.51±1.06 83.54±35.88
Apr-06_05 st 6.86±6.70 27.23±2.22 0.51±0.14 -1.16±0.81 270.82±0.29 226.61±43.95 5.80±1.29 35.06±5.93 2.05±0.49 1.00±0.42 5.97±1.38
Jul-05_05 st 13.64±0.00 781.59±0.00 0.11±0.00 -11.01±0.00 197.04±0.00 125.75±0.00 4.47±4.63 139.49±76.07 125.75±0.00 0.80±0.00 87.91±0.00
Oct-05_05 st 11.21±1.99 491.62±64.47 -0.61±0.13 -3.07±1.07 250.10±4.28 191.42±8.10 10.16±0.20 141.43±13.41 19.48±18.66 1.95±0.95 -
Apr-06_05 st 9.92±0.00 648.35±0.00 -0.21±0.00 -2.55±0.00 271.97±0.00 206.59±0.00 5.80±1.29 35.06±5.93 6.50±0.00 2.20±0.00 50.79±50.79
Sep-06_05 st 12.37±0.00 383.70±0.00 -0.48±0.00 -0.49±0.00 102.71±0.00 105.69±0.00 5.74±0.43 127.36±3.70 39.40±0.00 3.30±0.00 -
Apr-06_06 st 11.13±0.00 226.09±0.00 -1.19±0.00 -0.79±0.00 268.80±0.00 197.01±0.00 6.15±2.37 60.66±4.75 1.30±0.00 0.80±0.00 -
Sep-06_06 st 7.49±3.77 184.84±46.15 -0.42±0.20 -4.88±1.43 264.74±14.59 231.67±44.87 4.63±0.67 98.87±21.64 27.74±22.71 2.20±0.89 36.65±19.35
A Mar-06_06 st 2.16±0.16 71.79±1.14 2.45±1.31 -1.49±0.62 265.14±1.26 250.19±1.54 10.87±1.47 33.15±8.93 24.27±26.59 2.00±1.28 59.62±43.35
Oct-05_05 st 13.52±0.00 483.10±0.00 1.84±0.00 -3.37±0.00 277.07±0.00 196.40±0.00 10.16±0.20 141.43±13.41 2.10±0.00 0.80±0.00 -
Apr-06_05 st 10.34±4.46 769.89±0.00 0.26±0.13 -2.96±0.86 270.79±0.43 203.86±29.37 5.80±1.29 35.06±5.93 22.95±26.28 1.85±1.34 -
Sep-06_06 st 13.52±3.65 168.01±55.05 1.61±2.23 -6.55±0.67 273.82±8.06 178.01±27.63 4.63±0.67 98.87±21.64 29.55±29.49 2.10±0.99 40.70±22.56
2
A
B
3
Story Type Sampling date
4B
A1
A
B
Table. 3-3. Raw data follow the distinction and correlation
Unit
Denitrification - µmole · m-2 · d
-1
SOD, NH4+, NO3
- - mmole · m
-2 · d
-1
O2, NH4+, NO3
- Cconcentration (overlying water) - µM
Silt & Clay content , Sediment organic matter - %
Microphytobenthos - mg · m-2
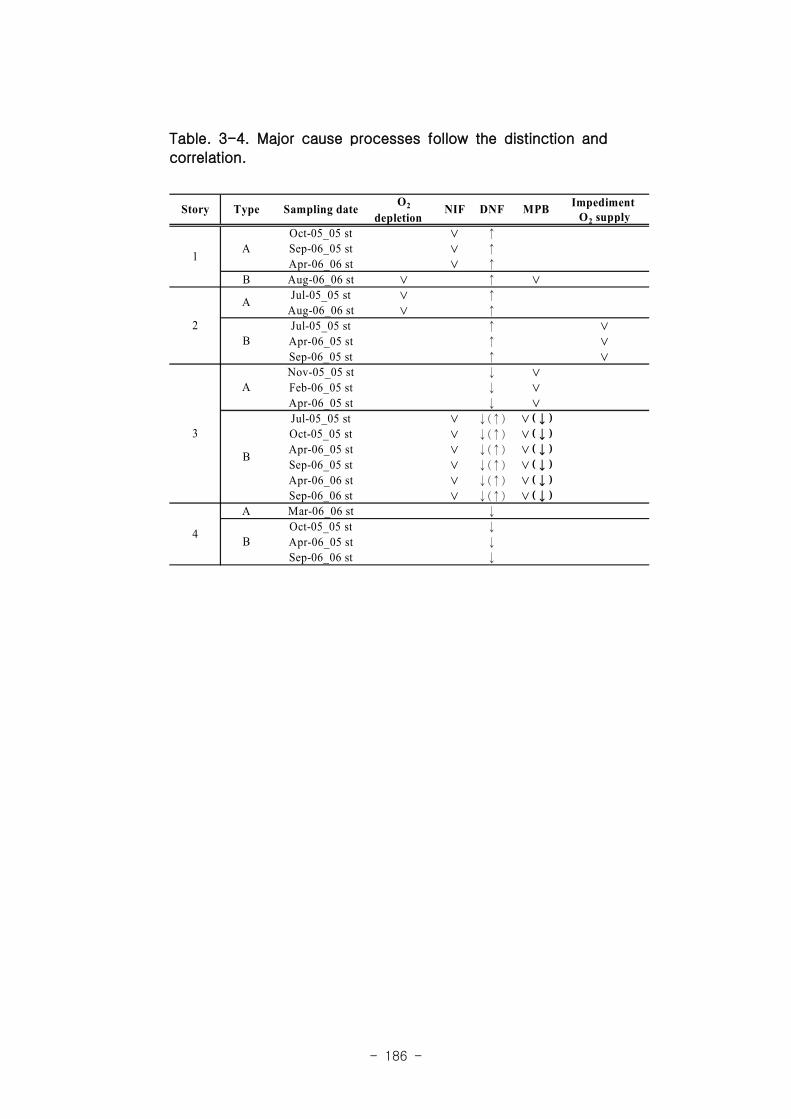
- 186 -
O2 Impediment
depletion O2 supply
Oct-05_05 st ∨ ↑Sep-06_05 st ∨ ↑Apr-06_06 st ∨ ↑
B Aug-06_06 st ∨ ↑ ∨Jul-05_05 st ∨ ↑Aug-06_06 st ∨ ↑Jul-05_05 st ↑ ∨Apr-06_05 st ↑ ∨Sep-06_05 st ↑ ∨Nov-05_05 st ↓ ∨Feb-06_05 st ↓ ∨Apr-06_05 st ↓ ∨Jul-05_05 st ∨ ↓(↑) ∨(↓)Oct-05_05 st ∨ ↓(↑) ∨(↓)Apr-06_05 st ∨ ↓(↑) ∨(↓)Sep-06_05 st ∨ ↓(↑) ∨(↓)Apr-06_06 st ∨ ↓(↑) ∨(↓)Sep-06_06 st ∨ ↓(↑) ∨(↓)
A Mar-06_06 st ↓Oct-05_05 st ↓Apr-06_05 st ↓Sep-06_06 st ↓
MPB
4B
�IF D�F
2
A
B
3
A
B
Story Type Sampling date
1A
Table. 3-4. Major cause processes follow the distinction and
correlation.
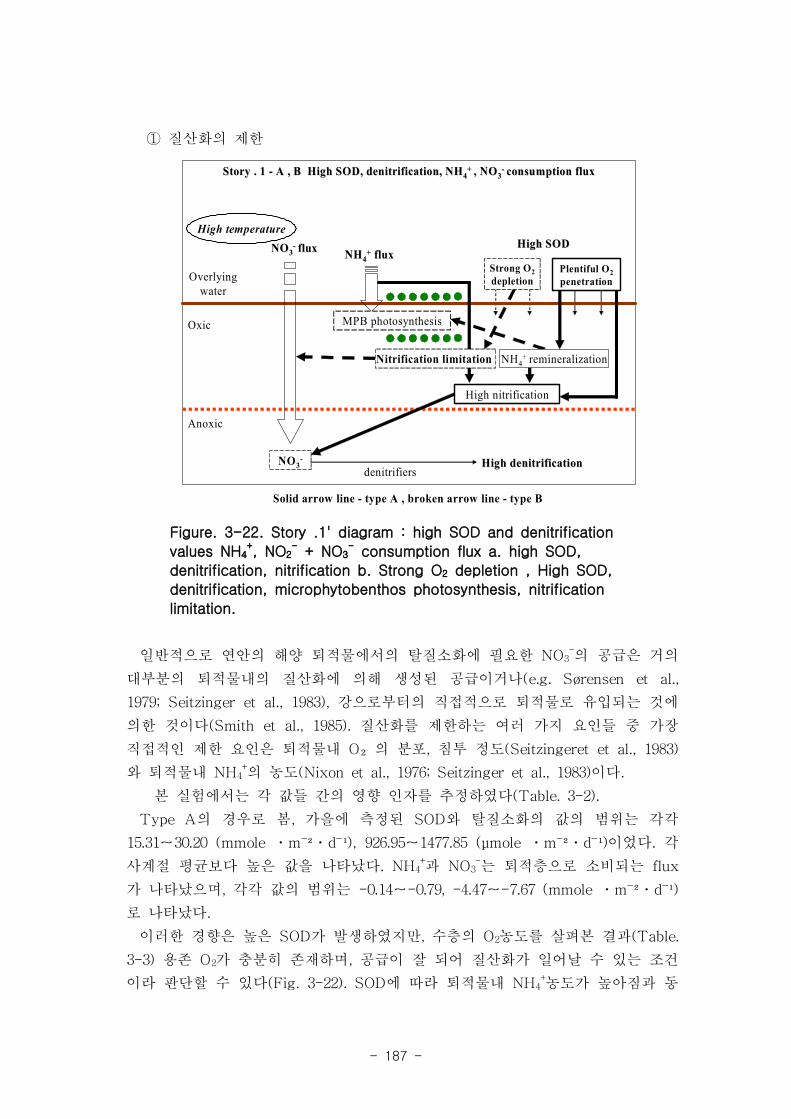
- 187 -
① 질산화의 제한
High SODHigh SOD
Overlying
water
Oxic
Anoxic
Story . 1 Story . 1 -- A , B High SOD, A , B High SOD, denitrificationdenitrification, �H, �H44+ + , �O, �O
33-- consumption fluxconsumption flux
�H�H44++ fluxflux
�O�O33-- fluxflux
�O3-
High High denitrificationdenitrificationdenitrifiers
Solid arrow line - type A , broken arrow line - type B
Strong O2
depletion
High nitrification
NH4+ remineralization�itrification limitation
MPB photosynthesis
Plentiful O2
penetration
High temperature
Figure. 3-22. Story .1' diagram : high SOD and denitrification
values NH4+, NO2
- + NO3- consumption flux a. high SOD,
denitrification, nitrification b. Strong O2 depletion , High SOD,
denitrification, microphytobenthos photosynthesis, nitrification
limitation.
일반적으로 연안의 해양 퇴적물에서의 탈질소화에 필요한 NO3-의 공급은 거의
대부분의 퇴적물내의 질산화에 의해 생성된 공급이거나(e.g. Sørensen et al.,
1979; Seitzinger et al., 1983), 강으로부터의 직접적으로 퇴적물로 유입되는 것에
의한 것이다(Smith et al., 1985). 질산화를 제한하는 여러 가지 요인들 중 가장
직접적인 제한 요인은 퇴적물내 O₂의 분포, 침투 정도(Seitzingeret et al., 1983)
와 퇴적물내 NH4+의 농도(Nixon et al., 1976; Seitzinger et al., 1983)이다.
본 실험에서는 각 값들 간의 영향 인자를 추정하였다(Table. 3-2).
Type A의 경우로 봄, 가을에 측정된 SOD와 탈질소화의 값의 범위는 각각
15.31~30.20 (mmole ㆍm⁻²ㆍd⁻¹), 926.95~1477.85 (μmole ㆍm⁻²ㆍd⁻¹)이었다. 각
사계절 평균보다 높은 값을 나타났다. NH4+과 NO3
-는 퇴적층으로 소비되는 flux
가 나타났으며, 각각 값의 범위는 -0.14~-0.79, -4.47~-7.67 (mmole ㆍm⁻²ㆍd⁻¹)로 나타났다.
이러한 경향은 높은 SOD가 발생하였지만, 수층의 O2농도를 살펴본 결과(Table.
3-3) 용존 O2가 충분히 존재하며, 공급이 잘 되어 질산화가 일어날 수 있는 조건
이라 판단할 수 있다(Fig. 3-22). SOD에 따라 퇴적물내 NH4+농도가 높아짐과 동

- 188 -
시에 빠르게 질산화 일어나서 NO3-로 전환되어 탈질소화 박테리아에게 많이 이용
(nitrification - denitrification coupling)된 것으로 예상할 수 있다. NH4+ flux는
퇴적물로 소비되는 경향이 나타났는데, 이는 퇴적물내 유기물 분해에 의해서 생성
된 NH4+가 동시에 질산화 과정을 거쳐 NO3
-로 빠르게 변환되고, 활성화된 질산화
의 결과로 퇴적물 내에서 NH4+가 부족함에 따른 결과이다. 결과에 따라 퇴적물내
낮아진 NH4+의 농도로 인해 수층으로부터 유입되어 질산화 과정을 거쳐 NO3
-로
변환되었다고 할 수 있다. 이렇게 두 가지 소스로 생성된 NO3-를 탈질소화 박테
리아가 사용하였다고 볼 수 있다. 아니면 2005년 10월과 2006년 9월의 경우 수층
의 NO3-의 농도는 각각 141, 127μM(Table. 3-3)로 다른 달에 비해 현저하게 높은
것으로 보아, 외부 (수층)에서 공급된 NO₃⁻를 미생물이 이용한 것으로 예상할
수 있다(Fig. 3-22).
이 결과 값에 따른 계절별, 조고별, 퇴적물 type별 분석 결과 우세한 경향이 나
타난 곳은 봄과 가을, LOWER station과 MUD 퇴적물이었다. LOWER station의
경우 가장 긴 침수 시간에 의해 유기물은 잘 공급되는 상황이며, 호기성 호흡보다
혐기성 호흡이 더 클 것이라 예상되었다. 하지만 다른 달에 비해 2006년 4월
(06st)과 2006년 9월(06st)의 표층 퇴적물의 silt & clay 함량은 6 ~ 37%인 것을
미루어 용존 O₂ 공급이 잘 일어날 수 있는 상황이라고 볼 수 있다(Table. 3-3).
Type B의 경우 여름철 높은 온도에 의해 큰 SOD 값이 발생하였고, 시간이 흐
를수록 강한 유기물 분해에 의해 퇴적물내 산소가 고갈되었다(Table. 3-3.). 유기
물 분해에 의해 재광물화된 NH4+는 고갈된 산소 농도로 인해 질산화의 저해
(Table. 3-4)로 탈질소화의 소스인 NO3-로 전환되지 못했고, 외부 수층으로부터
유입된 NO3-가 탈질소화에 사용되었다. NH4
+도 소모 flux 값이 나타났는데, 이는
여름철 높은 온도로 인한 저서 미세조류의 활발한 활동으로 NH4+소모 flux가 나
타난 것으로 생각된다.
NO3- flux는 큰 소비 경향을 나타났고, 퇴적물내 산소가 빠르게 고갈되어 질산
화가 제한되어 수층으로부터 공급되는 NO3-를 탈질 박테리아가 이용하는 상황이
발생했다고 예상할 수 있다. 탈질소화에 소스가 되는 NO3-가 퇴적물내 많이 공급
되었고, 높은 온도로 인해 탈질소화 박테리아 활동 또한 높게 일어나 탈질소화가
크게 일어났다고 예상할 수 있다(Fig. 3-22.).
이 결과 값에 따른 계절별, 조고별, 퇴적물 타입별 분석 결과 우세한 경향이 나
타난 곳은 여름, Upper였고, 퇴적물 type은 경향이 보이지 않았다.

- 189 -
② 용존 O₂의 공급
High SODHigh SOD
Overlying
water
Oxic
Anoxic
Story. 2 Story. 2 -- A , B High SOD, A , B High SOD, denitrificationdenitrification, �H, �H44++ production, �Oproduction, �O
33-- consumption fluxconsumption flux
�H�H44++ fluxflux
�O�O33-- fluxflux
�O3-
High High denitrificationdenitrificationdenitrifiers
Solid arrow line - type A , broken arrow line - type B
�itrification limitation
High silt & clay content
impediment O2 supplyStrong O2
depletion
NH4+ remineralization
High temperature
Figure. 3-23. Story .2' diagram : high SOD and denitrification
values NH4+ production flux, NO2
- + NO3- consumption flux a.
high SOD, denitrification, Strong O2 depletion, nitrification
limitation b. Strong O2 depletion , High SOD, denitrification, high
silt & clay content impediment O2 supply, nitrification limitation.
두 번째 story로 여름에 나타난 SOD와 탈질소화의 값의 범위는 각각 13.7~
37.7 (mmole ㆍm⁻²ㆍd⁻¹), 856.5~4564.6 (μmole ㆍm⁻²ㆍd⁻¹)이었다. 각 사계절 평
균보다 높은 값을 나타났다. NH4+ flux는 퇴적층으로부터 수층으로 용출되었으며,
그 값의 범위는 0.5~9.1 (mmole ㆍm⁻²ㆍd⁻¹)이었다. NO3-는 퇴적층으로 강하게
소비되는 flux가 나타났으며, 값의 범위는 -21.8~3.6 (mmole ㆍm⁻²ㆍd⁻¹)로 나타
났다. Type A의 경우 수층과 표층 퇴적물에 존재하는 용존 O₂가 고갈되어 질산
화 가 일어날 수 없어 수층으로부터 퇴적층으로 확산된 NO3-를 이용하여 탈질소
화가 발생한 것으로 예상된다(fig. 3-23, Table. 3-2). SOD가 높게 일어날 수 있
는 조건인 온도가 높고, 퇴적물내 유기물 함량 또한 많아 매우 높은 값들이 나타
난 것으로 해석할 수 있으며, 재광물화가 잘 일어났다고 볼 수 있다(Table. 3-3).
NH4+는 퇴적층으로부터 용출 flux 값이 발생하였는데, 이는 SOD에 의해 퇴적물
내 NH4+농도는 높아졌지만, 수층과 퇴적층의 용존산소의 고갈로 질산화과정이 차
단되었으며, 그에 따라 수층의 NO3-가 퇴적층으로 확산(diffusion)되어 탈질소화의
소스가 된 것으로 판단할 수 있다.
Type B의 경우 표층 퇴적물의 높은 silt & clay 함량 때문에 질산화 제한이 발

- 190 -
생된 경우이다(Fig. 3-23.). 실제로 표층 퇴적물의 높은 silt & clay(41~95%) 함량
때문에 호기층이 매우 얇을 것이며, 이러한 특성으로 인해 퇴적물내로 산소가 공
급되기 어려운 상황 때문에 산소 공급에 영향을 받는 질산화가 제한되어졌을 것
으로 예상할 수 있다. 또한 SOD에 의해 퇴적물내 NH4+농도가 높아짐과 동시에
빠르게 질산화 일어나서 NO3-로 전환되어 표층 퇴적물내 산소가 고갈된 상황도
함께 예상할 수 있다. 실제로 24hr 때의 core 속 용존 산소 농도를 살펴보면, 5
2~227μM로 질산화가 발생하기에는 충분한 양이 존재하는 것을 발견할 수 있었
다.(Table 2.). 표층퇴적물의 높은 silt & clay(41~95%) 함량으로 인해 질산화 저
해되어 수층에서 공급된 NO3-을 이용하여 높은 탈질소화 일어났을 것으로 예상된
다. 이 결과 값에 따른 계절별, 조고별, 퇴적물 타입별 분석 결과 우세한 경향이
나타난 곳은 여름이었고 조고는 MIDDLE이었고, 퇴적물 type은 SAND와 mixed
에서 경향이 뚜렷히 나타났다.
③ 저서미세조류(MPB)의 영향
Low SOD
Overlying
water
Oxic
Anoxic
Story . 3 Story . 3 -- A , B Low SOD, A , B Low SOD, denitrificationdenitrification, �H, �H44+ + , �O, �O
33-- consumption fluxconsumption flux
�H�H44++ fluxflux
�O�O33-- fluxflux
�O3-
Low Low denitrificationdenitrificationdenitrifiers
Solid arrow line - type A , broken arrow line - type B
�itrification limitation
Low temperature
NH4+absorption
Nitrification promotion
Plentiful O2
penetration
O2
production
production
NH4+ remineralization
MPB photosynthesis
Figure. 3-24. Story .3' diagram : low SOD and denitrification
values NH4+ , NO2
- + NO3- consumption flux a. low SOD,
denitrification, Plentiful O2 penetration, MPB photosynthesis,
ammonium absorption, nitrification limitation b. Plentiful O2
penetration , low SOD, denitrification, nitrification promotion, NO2-
+ NO3- by nitrifier .
세 번째 story로 봄과 가을 그리고 겨울에 나타난 SOD와 탈질소화의 값의 범

- 191 -
위는 각각 -0.8~13.6 (mmole ㆍm⁻²ㆍd⁻¹), 0.2~781.6 (μmole ㆍm⁻²ㆍd⁻¹)이었다.
각 사계절 평균보다 낮은 값을 나타났다. NH4+ flux는 퇴적층으로부터 수층으로
소비되었으며, 그 값의 범위는 -0.7~-2.0 (mmole ㆍm⁻²ㆍd⁻¹)이었다. NO3-는 퇴
적층으로 소비되는 flux가 나타났으며, 값의 범위는 -0.0~-11.0 (mmole ㆍm⁻²ㆍd⁻¹)로 나타났다. 전체적으로 낮은 온도와 MPB의 광합성에 영향을 받았다(Fig.
3-24).
Type A의 경우 겨울철의 낮은 수온의 영향으로 전체적으로 SOD와 탈질소화의
값이 작게 나타났으며, 특히 2005년에 만들어진 인공갯벌에서 2005년 11월과 2006
년 2월에 측정된 SOD는 거의 발생하지 않았고, 1hr 때 측정된 core 속 용존 산소
농도를 살펴보면 초기보다 증가된 것을 알 수 있다(Table. 3-3). 이는 퇴적물 표
층에 서식하는 MPB의 광합성으로 인해 산소가 고갈되기보다는 생산되었기 때문
이다. NH4+ flux는 퇴적층으로 소모되는 경향이 나타났는데, 이는 표층 퇴적물에
서식하는 MPB의 광합성 활동으로 인해 용존 무기 질소인 NH4+가 흡수되었기 때
문이라고 예상할 수 있다. 이때의 MPB의 생체량을 살펴본 결과 여름철보다 더
높은 단위면적당 약 100mg 이상의 값으로 나타났다. 또한, 이때의 수층의 NH4+
의 농도 13μM로 계절 중 가장 높은 농도를 나타내서 확산에 의한 퇴적층으로의
소모도 함께 예상할 수 있다. 보고된 바에 의하면 MPB의 광합성은 탈질소화를
촉진시킬 수도 저해할 수도 있다(An. 2005)
Type A의 경우는 MPB의 활발한 활동으로 인해 NH4+가 흡수되고, 질산화에
이용될 소스가 없어 질산화가 제한되어 탈질소화가 거의 발생하지 않았다고 예상
할 수 있다. 이 결과 값에 따른 계절별, 조고별, 퇴적물 타입별 분석 결과 우세한
경향이 나타난 곳은 겨울이었고 조고는 MIDDLE과 LOWER이었고, 퇴적물 type
은 경향이 나타나지 않았다.
Type B의 경우는 봄과 가을철 나타난 경향인데, 이때의 SOD와 탈질소화는 평
균값보다 낮지만, 겨울철보다는 높은 값이었다. 퇴적층으로 소모되는 NH4+ flux가
나타났는데, MPB의 광합성 작용으로 NH4+가 소모되었다고 예상할 수 있으며, 이
는 Type A의 경우와 다르게, 유기물 생산과 산소 발생으로 퇴적물의 산화환원환
경을 바뀌게 만들어 질산화 과정을 촉진시켜 탈질소화가 작지만 발생한 것이라
예상된다. 이 결과 값에 따른 계절별, 조고별, 퇴적물 타입별 분석 결과 우세한 경
향이 나타난 곳은 봄과 가을이었고 조고는 UPPER와 LOWER이었고, 퇴적물
type은 SAND퇴적물에서 경향이 나타났다.

- 192 -
④ 온도에 따른 영향 (DNRA?)
Low SOD
Overlying
water
Oxic
Anoxic
Story . 4 Story . 4 -- A , B Low SOD, A , B Low SOD, denitrificationdenitrification, �H, �H44++ production, �Oproduction, �O
33-- consumption fluxconsumption flux
�H�H44++ fluxflux
�O�O33-- fluxflux
�O3- Low Low denitrificationdenitrification
low activity of denitrifiers
Solid arrow line - type A , type B
NH4+ remineralization
D�RA?D�RA?
�itrification limitation,
low activity of nitrifiers
Low temperature
Plentiful O2
penetration
Figure. 3-25. Story .4' diagram : low SOD and denitrification
values NH4+ production flux, NO2
- + NO3- consumption flux - low
SOD, denitrification, Plentiful O2 penetration, low activity of
nitrifiers, nitrification limitation, low activity of denitrifiers, low
denitrification.
네 번째 story로 봄과 가을 그리고 겨울에 나타난 SOD와 탈질소화의 값의 범
위는 각각 0.1~16.1mmole ㆍm-2
ㆍd-1
, 1.6~798.1μmole ㆍm-2
ㆍd-1이었다. 각 사계
절 평균보다 낮은 값을 나타났다. NH4+ flux는 퇴적층으로부터 수층으로 소비되
었으며, 그 값의 범위는 0.0~3.9mmole ㆍm-2
ㆍd-1이었다. NO3
-는 퇴적층으로 소
비되는 flux가 나타났으며, 값의 범위는 -0.8~-7.0mmole ㆍm-2
ㆍd-1로 나타났다.
전체적으로 낮은 온도의 영향을 받았다(Fig. 3-25).
네 번째 story의 경우 여러 가지 상황을 예상할 수 있다. 작은 값이지만 SOD가
발생되었다. 또한 NH4+ 재광물화 되어 퇴적물내 그 농도가 증가하여 퇴적물로부
터 수층으로 NH4+가 확산(diffusion) 되어 용출 flux가 나타났을 것이라 예상된다.
또한 퇴적층내 NO3-농도보다 수층의 NO3
-의 농도가 높아 수층에서 퇴적층으로
이동(diffusion)되는 NO3-의 양도 있다. 또는 낮은 수온으로 인해 질산화 과정은
nitrifiers 활동도가 작아져 제한될 수 있을 것을 예상할 수 있다. 이러한 상황과
동시에 수층에서 유입된 NO3-는 낮은 denitrifiers 활동도의 영향으로 매우 적은
양만이 탈질소화에 소스로 이용되었고, 남은 NO3-는 퇴적층내 NO3
-환원과정에 의
해 다시 NH4+로 바뀌어 수층으로 용출되는 결과라고도 예상할 수 있다. 이 결과

- 193 -
값에 따른 계절별, 조고별, 퇴적물 타입별 분석 결과 우세한 경향이 나타난 곳은
겨울이었고 조고는 MIDDLE과 LOWER이었고, 퇴적물 type은 SAND퇴적물에서
경향이 나타났다.
2. 먹이망 분석
연안역에서는 외양과는 달리 인접한 해양이나 육상으로부터 외래기원의 유기물
(allochthnous source)을 공급받을 뿐만 아니라 연안역 자체에서 저서미세조류, 해
조류, 해초류 및 염습식물과 같은 다양한 기초생산자 (local sources)들의 존재에
의해 풍부한 먹이를 공급받기 때문에 가장 생산력이 높은 생태계로 간주되고 있
다. 이와 같은 다양한 먹이원은 연안생태계에서 높은 생물 다양성과 더욱 복잡한
먹이망 (food web)을 형성하게 한다. 연안생태계의 영양기저 (trophic base)와 먹
이망을 통한 유기물 혹은 에너지의 흐름경로에 대한 정보는 연안생태계에서 다양
한 유기물의 상대적인 중요성이 어느 정도인가를 평가하고 각각의 생물군의 그
생태계 내에서의 역할을 평가할 수 있도록 하기 때문에 연안역의 관리 (coastal
management)에서 관리 기준을 제시할 수 있는 기본적인 자료이다. 즉, 어떤 연안
생태계 먹이망이 다양한 기원의 유기물 중 어떤 기원의 유기물을 영양기저로 하
고 있는지를 알게 된다면 생물자원회복을 위해 관리 • 보존하거나 복원해야할 생
태구성요소를 설정할 수가 있게 되어서 생태계 관리전략을 수립할 수 있게 되는
것이다.
그러나 연안역에서의 다양한 지형구조와 식물분포에서의 이질성 및 복잡한 유
기물의 순환 경로, 소비자 동물의 먹이원을 평가하는 방법상의 어려움 때문에 이
들 생태계 먹이망의 영양기저를 일반화하여 정의하기는 대단히 어렵다. 실제로,
다양한 어류 군집을 대상으로 위 내용물 분석의 방법을 이용하여 어류들의 이용
먹이에 대한 연구들을 광범위하게 수행되어 왔지만, 이 연구들은 동물들이 먹이를
취하는 생태계내 먹이망의 영양기저에 대한 어떤 정보도 제공해 주지 못할뿐더러
소화관내에서 발견되는 먹이생물의 단순한 % 조성만을 제시한다. 이와 같은 전통
적인 먹이연구 방법상의 문제점들을 보완하고 실제로 소비자 동물들이 이용하는
유기물의 기원을 평가할 수 있는 한 기법으로서 잠재적인 유기물 기원물질인 식
물과 소비자 동물들의 생체 내 안정동위원소 조성을 이용용되고 있다.
타가영양생물(heterotroph)이 무엇을하는 방법이 널리 이 먹는가를 결정하는 전
통적인 방법은 위속의 내용물을 관찰 • 조사하는 것이었다. 그러나 이 방법은 특히
크기가 작거나 기생해서 사는 생물, 부식영양자(세균, 균류, 사해식자 등)이나 미
세조류식자에게는 적용할 수가 없다. 또한 이 방법들은 많은 시료들을 분석해야
하기 때문에 많은 시간과 경제성 및 노력이 요구될 뿐만 아니라 먹이생물로 다양
한 생물종을 대상으로 연구를 수행해야 하기 때문에 생물 분류에 대한 어려움에
직면하게 된다. 그래서 어떤 경우에는 많은 종들이 상호작용하는 자연생태계에서

- 194 -
의 먹이망을 추적하는데 동위원소 추적자가 사용될 수 있다. 예를 들면 어떤 곤충
이 어떤 식물을 먹으며 어떤 포식자가 어떤 피식자를 먹는가를 결정하는데 동위
원소 추적자가 유용하게 이용될 수 있다.
탄소안정동위원소 비를 측정하게 되면 다른 방법으로는 어려운 먹이연쇄에 있
어서의 에너지 흐름을 규명하는데 특히 유용하게 이용될 수 있다. C-3식물과
C-4식물 및 조류는 13C/
12C의 비가 다르며 이것은 어떤 종류의 생물이 특정 식물
또는 식물성 쇄설물(detritus)을 먹더라도 이 비값이 그대로 전이된다. 따라서 어
떤 동물체의 안정동위원소 조성을 분석해 본다면 그 동물이 실제로 이용하는 먹
이의 기여도를 평가할 수 있다. 또한, 체성분 중 안정동위원소 조성은 상당 기간
동안 그들의 섭식활동이 반영된 결과라는 것을 고려한다면 이들이 서식하고 있는
먹이환경의 변화를 해석하는 데에도 이용할 수 있다. 한편, 먹이사슬의 각 영양단
계를 거치는 동안 안정동위원소의 비값은 한 영양단계 당 탄소에서는 약 1‰, 질
소는 약 3~4‰ 높아지는 동위원소 분별효과가 일어나는 것이 일반적으로 알려져
있다. 질소안정동위원소의 이와 같이 비교적 큰 분별효과는 각 소비자 동물들의
그 생태계내에서의 영양단계를 해석하는데 이용된다.
따라서 전통적인 먹이 연구방법인 소화관 내용물 분석법이 주로 상위단계의 먹
이망 분석에 이용될 수 있는 단점을 보완할 수 있도록 하위단계의 영양구조 분석
에 유용한 안정동위원소 응용기법을 함께 적용하여 연안생태계 먹이망 분석을 시
도함으로써 연안생태계의 전체적인 먹이그물을 완성해 낼 수 있고, 이로부터 더욱
정확한 물질순환 경로와 생태계 구성요소들 사이의 연결고리를 해석할 수 있다.
이와 같은 정보는 생태계 구성요소들의 기능을 평가할 수 있는 기본적인 자료가
되어 연안의 생물생산에 대한 이들 생태계 구성요소 각각의 역할과 기여를 평가
할 수 있게 될 것이다. 이렇게 평가된 결과를 근거로 이후의 서식지 복원전략 이
행에 따른 연안 천해 생태계 안정화과정의 파악은 연안의 생물종 다양성과 생물
생산의 증대를 위해 필수적으로 요구되는 기본정보라 할 수 있다.
(1) 재료 및 방법
(가) 시료의 채집과 분석준비
낙동강 하구에서 실시한 이전의 연구들로부터 기존의 물리, 화학, 생물, 지질학
적 특성을 파악하여 조사해역을 가장 잘 대변할 수 있는 정점군을 설정하여 2004
년 9월부터 2005년 11월일 사이에 현장조사와 시료채집을 실시하였다. 본 조사에
서 각 정점별로 수행한 조사항목과 채집시료는 다음과 같다.
① 이화학 조사와 입자물질 및 퇴적물 채취
수온과 용존산소는 CTD meter로 현장에서 측정하였다. 안정동위원소 분석을

- 195 -
위하여 현장 해수 약 5 L를 채수하여 여과지를 이용하여 부유입자물질을 여과하
였으며, 10% 염산(HCl) 2~3 방울을 가해서 탄산칼슘을 제거하고 증류수로 세척
한 후 동결건조하여 냉동 보관하였다. 퇴적물은 소형 core를 이용하여 현장에서
채취하고 동결하여 실험실로 운반한 후 일정량(약 200g)의 퇴적물을 10% 염산
(HCl)으로 처리하여 탄산칼슘 등의 무기탄소를 제거하고 증류수로 세척하여 동결
건조한 후 전기분쇄기를 이용하여 고운 분말을 만들어 안정동위원소 분석을 위해
서 냉동 보관하였다.
② 대형식물과 미세조류시료
조사지역에서 조사된 기존의 자료를 참조하여 인근 육상에서 우점하는 나뭇잎
(terrestrial plants)과 동물의 먹이로서 가용성이 높은 하구 수변의 우점식물군
(macrophytes)을 대상으로 현장채집을 실시하고, 냉장하여 실험실로 운반, 분류
동정 후 깨끗하게 세척하여 동결 저장하였다. 퇴적물 표면에 부착하여 갈색의 띠
를 이루는 부분을 퇴적물 채취용 칼을 이용하여 표면부분을 상당량 채취한 후 냉
장하여 실험실로 운반, Silicagel과 식물플랑크톤 채집 망지를 이용하는 방법을 이
용하여 부착미세조류 (periphyton)를 분리하고 냉동보관 하였다. 식물플랑크톤은
식물플랑크톤용 채집망(net)로 채집하였다.
③ 대형저서무척추동물
각 조사정점에서 Grab 채니기를 이용하여 퇴적물을 채취하고 Seive로 걸러서
남는 잔여물로부터 저서동물을 분리하고, 냉장 보관하여 실험실로 운반한 후 현미
경으로 검경하여 종동정을 실시하였다. 안정동위원소 분석용 시료는 고정과정이
없이 현장에서 동결하여 실험실로 운반한 후 깨끗하게 세척하여 냉동 보관하였다.
④ 동물플랑크톤과 어류시료 채집
동물플랑크톤은 각 조사정점에서 동물플랑크톤 net를 이용하여 플랑크톤을 채
집하고 냉장하여 실험실로 운반한 후 현미경으로 대분류 분급하여 냉동 보관하였
다. 어류시료는 각 조사정점에서 표영성 어류와 저서성 어류를 모두 채집하기에
용이한 반두 어구와 투망 어구 그리고 정치망 어구를 이용하여 어류를 채집하고
10% Formalin 용액으로 고정하여 실험실로 운반하여 어류 종동정을 하였고, 안정
동위원소 분석용 어류 시료는 고정과정이 없이 현장에서 동결하여 실험실로 운반
하여 근육부위를 절개 채집한 후 동결 보관하였다.
(나) 입자물질, 퇴적물, 동 • 식물의 C(탄소)와 N(질소) 안정동위원소 분석
채집된 모든 시료들은 현장에서 아이스박스에 얼음과 함께 채워 실험실로 운반
하였다. 실험실에서 먼저 입자물질 및 플랑크톤 등의 시료는 원심분리를 하여 포

- 196 -
집하였고, 생물시료는 가능하면 종 단위까지 생물을 분류한 후 잘 세척하여 식물
의 잎은 절단하고 동물체는 조심스럽게 근육부위를 절개 혹은 절단하여 동결건조
를 하였으며, 건조된 시료는 잘 갈아서 미세한 분말을 만들고 분석 때까지 데시케
이터에 보관한 후 이 분말들을 약 1~2mg 분취하여 분석에 이용하였다.
안정동위원소조성은 CNS 원소분석기와 연결된 Micromass사의 IsoPrime급 안
정동위원소 질량분석기(IRMS, Isotope Ratio-Mass Spectrometer)를 이용하여 분
석하였다. 동 • 식물체, 입자물질과 퇴적물 등의 각종 시료가 가지는 안정동위원소
비값은 아래 식과 같이 표준물질에 대한 시료의 그 비값 변위를 천분율 (‰)로 나
타내어 δ 기호로 표현하게 되는데, 여기서 사용한 표준물질은 탄소의 경우는
PDB (Pee Dee Belmnite)와 질소의 경우 air N2 등 국제표준 기준을 사용하였다.
δ13C (‰) = 〔 (
13C12C
) sample
(13C12C
) standard
- 1 〕 × 1000
δ15N (‰) = 〔 (
15N14N
) sample
(15N14N
) standard
- 1 〕 × 1000
본 연구에서 동위원소 분석을 위한 carrier gas로는 헬륨 (He) 개스를 이용하였
고, 원소분석기의 온도를 1030℃로 조절하여 모든 유기물을 완전 연소시킨 후
CO2와 N2 개스를 헬륨으로 안정동위원소 질량분석기로 주입하여 각 원소를 측정
하고 위의 식에 따라 국제 표준에 맞는 동위원소 비값을 계산하였다. 이를 위해서
내부 표준물질로 동위원소 비값을 알고 있는 peptone을 이용하였다. 한편, 각각
총 20개의 peptone과 lysine을 이용하여 안정동위원소를 분석한 결과 standard
deviation은 탄소에서 ±0.1‰과 질소에서 ± 0.2‰을 나타내어 매우 높은 정밀도와
재현성을 나타내었다.
(2) 연구 결과
본 연구에서는 세모고랭이 서식지에서 대형저서동물의 잠재적인 먹이원으로서
가용성이 높은 기초생산자 식물군과 입자유기물질 그리고 이들에 대한 소비자 동
물들의 탄소와 질소 안정동위원소를 분석하여 생산자와 소비자 사이의 주요 영양
연결고리를 평가하려는 시도를 하였다.
Fig. 3-26 은 저서미세조류, 하구 조하대의 부유입자유기물, 세모고랭이, 강물을
통해서 유입되는 입자유기물의 생물량과 탄소안정동위원소 비값의 계절변동을 나
타낸다. 전체적으로 겨울과 봄철에 다소 값이 증가하는 경향을 보이지만 각 기원

- 197 -
물질별로 잘 구별되는 양상을 나타내어 주었다. river discharge는 낙동강 하구댐
으로 방류되는 담수로써 계절적인 변이가 매우 크다. 가장 높은 방류량을 보이는
시기는 6~7월로써 하계의 몬순기후와 일치하는 경향이다. river POM의 탄소안정
동위원소비값은 평균 -24.5±1.9‰로 나타났다. 수중의 클로로필의 함량은 8월에
가장 높게 나타났으며 탄소안정동위원소비의 평균은 -23.1±0.7‰이고 저서미세조
류는 -16.5±2.1‰로 비교적 무거운 탄소안정동위원소비값을 보였다. 염습지식물인
세모고랭이와 갈대의 생물량은 여름에 가장 많았으며 탄소안정동위원소비값은
-28.0±1.0‰, -26.6±0.9‰로 나타났다. Fig. 3-26 는 저서미세조류, 하구수로의 부
유입자유기물, 세모고랭이, 강물을 통해서 유입되는 입자유기물 및 퇴적물의 탄소
안정동위원소 비값의 계절변동을 나타낸다. 전체적으로 겨울과 봄철에 다소 값이
증가하는 경향을 보이지만 각 기원물질별로 잘 구별되는 양상을 나타내어 주었다.
Fig. 3-27 에 나타낸 바와 같이 전체 식물기원 유기물을 탄소와 질소안정동위원
소 비값을 동시에 한 좌표에 나타내었을 때 그 구분은 더욱 뚜렷해짐을 알 수 있
다.
Figure. 3-26. Seasonal dynamics of biomass and
carbon isotope rations in potential food sources (river
POM, channel POM, microphytobenthos, Scirpus triqueter, Phragmites australis) on Nakdong River
estuary
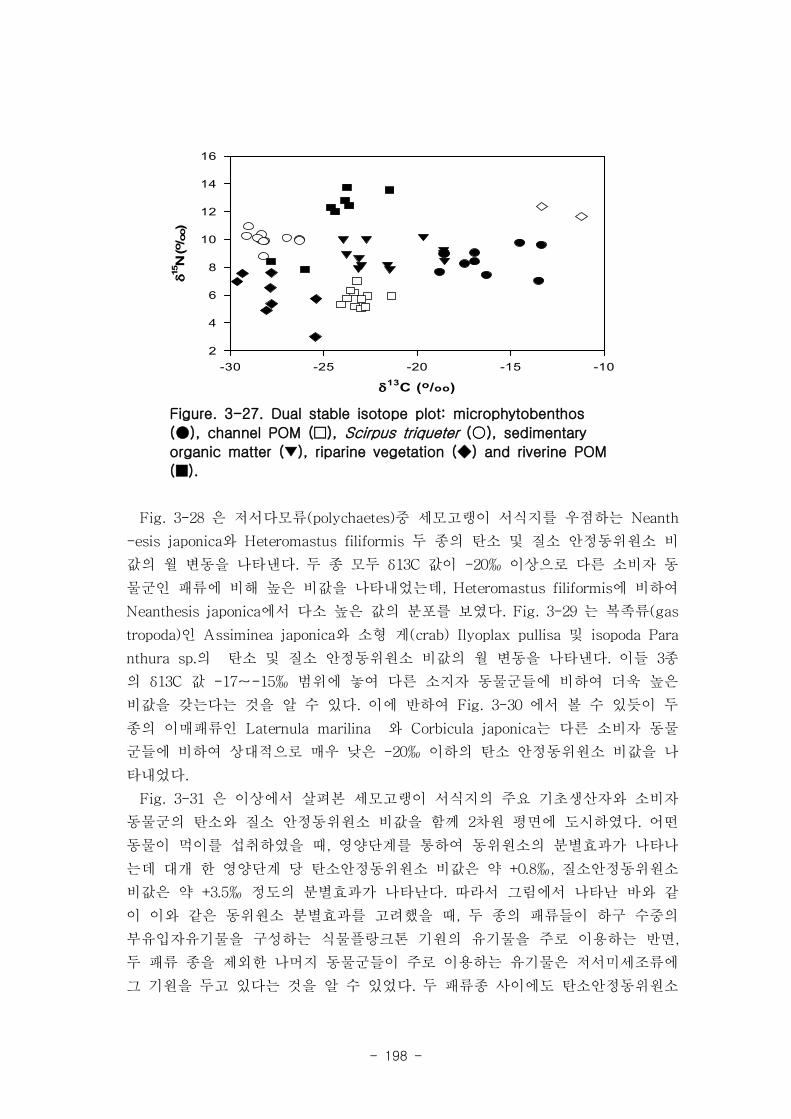
- 198 -
δ13
C (o/oo)
-30 -25 -20 -15 -10
δ15N (o/ o
o)
2
4
6
8
10
12
14
16
Figure. 3-27. Dual stable isotope plot: microphytobenthos
(●), channel POM (□), Scirpus triqueter (○), sedimentary
organic matter (▼), riparine vegetation (◆) and riverine POM
(■).
Fig. 3-28 은 저서다모류(polychaetes)중 세모고랭이 서식지를 우점하는 Neanth
-esis japonica와 Heteromastus filiformis 두 종의 탄소 및 질소 안정동위원소 비
값의 월 변동을 나타낸다. 두 종 모두 δ13C 값이 -20‰ 이상으로 다른 소비자 동
물군인 패류에 비해 높은 비값을 나타내었는데, Heteromastus filiformis에 비하여
Neanthesis japonica에서 다소 높은 값의 분포를 보였다. Fig. 3-29 는 복족류(gas
tropoda)인 Assiminea japonica와 소형 게(crab) Ilyoplax pullisa 및 isopoda Para
nthura sp.의 탄소 및 질소 안정동위원소 비값의 월 변동을 나타낸다. 이들 3종
의 δ13C 값 -17~-15‰ 범위에 놓여 다른 소지자 동물군들에 비하여 더욱 높은
비값을 갖는다는 것을 알 수 있다. 이에 반하여 Fig. 3-30 에서 볼 수 있듯이 두
종의 이매패류인 Laternula marilina 와 Corbicula japonica는 다른 소비자 동물
군들에 비하여 상대적으로 매우 낮은 -20‰ 이하의 탄소 안정동위원소 비값을 나
타내었다.
Fig. 3-31 은 이상에서 살펴본 세모고랭이 서식지의 주요 기초생산자와 소비자
동물군의 탄소와 질소 안정동위원소 비값을 함께 2차원 평면에 도시하였다. 어떤
동물이 먹이를 섭취하였을 때, 영양단계를 통하여 동위원소의 분별효과가 나타나
는데 대개 한 영양단계 당 탄소안정동위원소 비값은 약 +0.8‰, 질소안정동위원소
비값은 약 +3.5‰ 정도의 분별효과가 나타난다. 따라서 그림에서 나타난 바와 같
이 이와 같은 동위원소 분별효과를 고려했을 때, 두 종의 패류들이 하구 수중의
부유입자유기물을 구성하는 식물플랑크톤 기원의 유기물을 주로 이용하는 반면,
두 패류 종을 제외한 나머지 동물군들이 주로 이용하는 유기물은 저서미세조류에
그 기원을 두고 있다는 것을 알 수 있었다. 두 패류종 사이에도 탄소안정동위원소
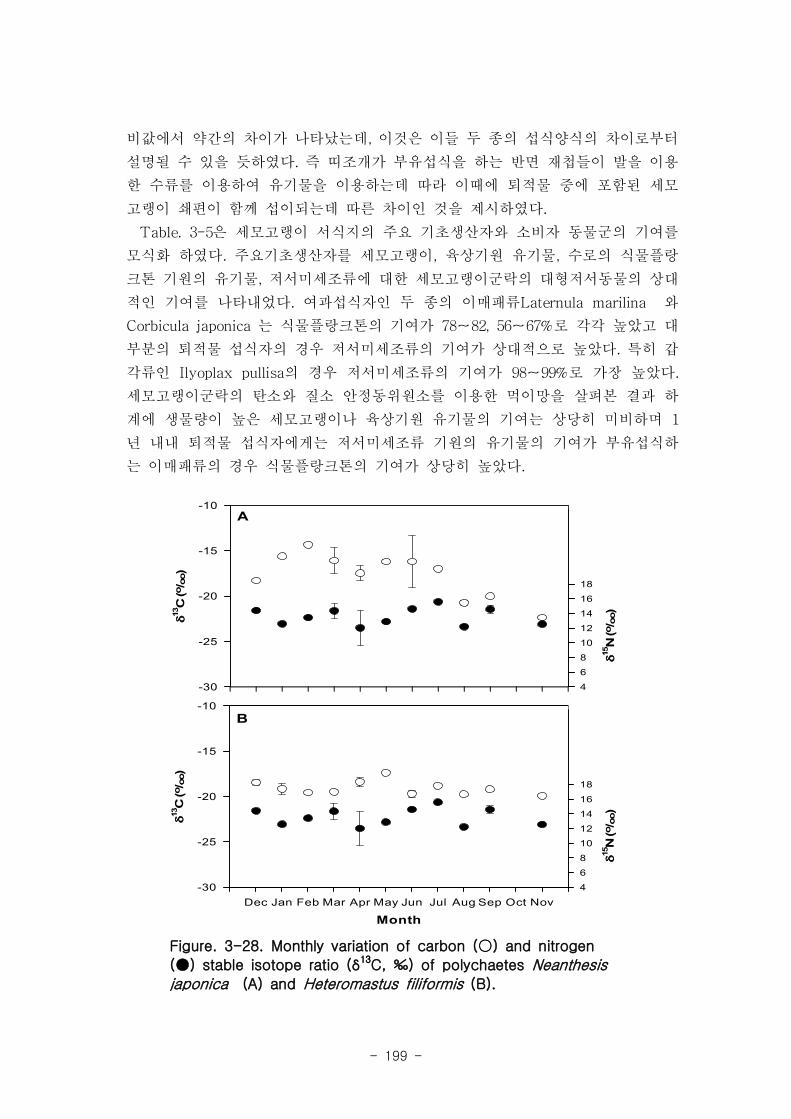
- 199 -
비값에서 약간의 차이가 나타났는데, 이것은 이들 두 종의 섭식양식의 차이로부터
설명될 수 있을 듯하였다. 즉 띠조개가 부유섭식을 하는 반면 재첩들이 발을 이용
한 수류를 이용하여 유기물을 이용하는데 따라 이때에 퇴적물 중에 포함된 세모
고랭이 쇄편이 함께 섭이되는데 따른 차이인 것을 제시하였다.
Table. 3-5은 세모고랭이 서식지의 주요 기초생산자와 소비자 동물군의 기여를
모식화 하였다. 주요기초생산자를 세모고랭이, 육상기원 유기물, 수로의 식물플랑
크톤 기원의 유기물, 저서미세조류에 대한 세모고랭이군락의 대형저서동물의 상대
적인 기여를 나타내었다. 여과섭식자인 두 종의 이매패류Laternula marilina 와
Corbicula japonica 는 식물플랑크톤의 기여가 78~82, 56~67%로 각각 높았고 대
부분의 퇴적물 섭식자의 경우 저서미세조류의 기여가 상대적으로 높았다. 특히 갑
각류인 Ilyoplax pullisa의 경우 저서미세조류의 기여가 98~99%로 가장 높았다.
세모고랭이군락의 탄소와 질소 안정동위원소를 이용한 먹이망을 살펴본 결과 하
계에 생물량이 높은 세모고랭이나 육상기원 유기물의 기여는 상당히 미비하며 1
년 내내 퇴적물 섭식자에게는 저서미세조류 기원의 유기물의 기여가 부유섭식하
는 이매패류의 경우 식물플랑크톤의 기여가 상당히 높았다.
δ13C (o/ o
o)
-30
-25
-20
-15
-10
δ15N (o/ o
o)
4
6
8
10
12
14
16
18
30
Month
Dec Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov
δ13C (o/ o
o)
-30
-25
-20
-15
-10
δ15N (o/ o
o)
4
6
8
10
12
14
16
18
30
A
B
Figure. 3-28. Monthly variation of carbon (○) and nitrogen
(●) stable isotope ratio (δ13C, ‰) of polychaetes Neanthesis japonica (A) and Heteromastus filiformis (B).
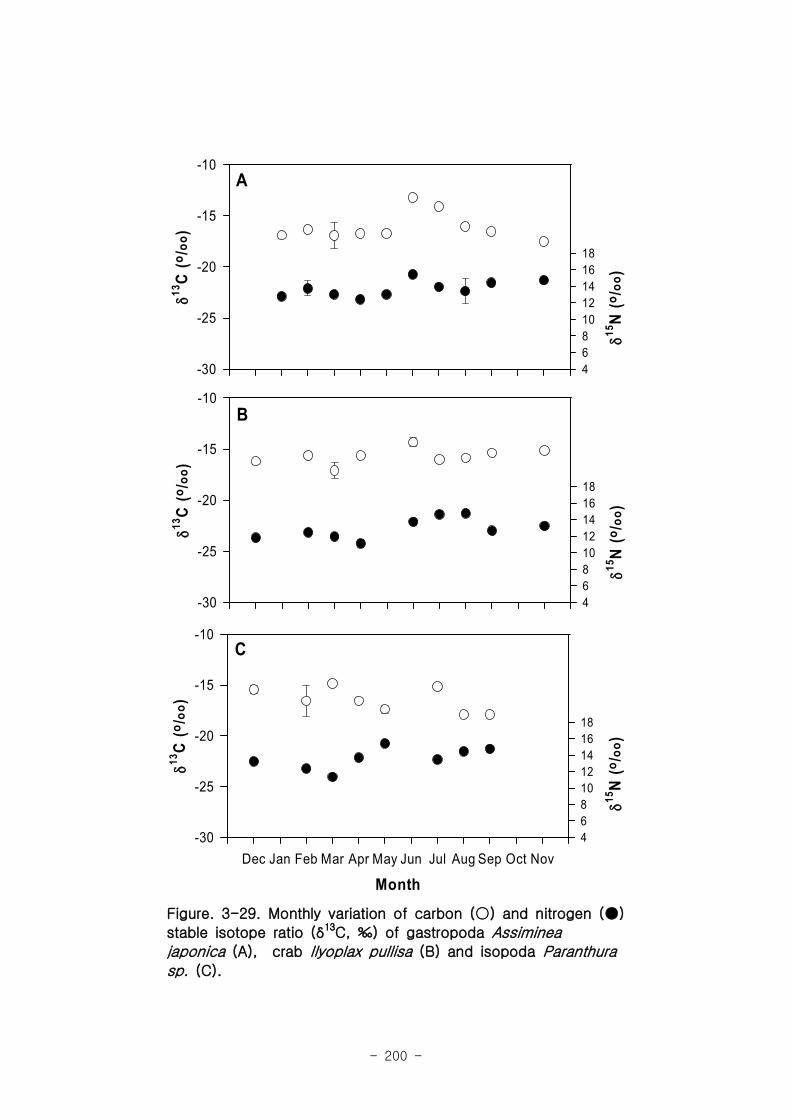
- 200 -
Month
Dec Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov
δ13C
(o/o
o)
-30
-25
-20
-15
-10
δ15N
(o/o
o)
4
6
8
10
12
14
16
18
30
δ13C
(o/o
o)
-30
-25
-20
-15
-10
δ15N
(o/o
o)
4
6
8
10
12
14
16
18
30
δ13C
(o/o
o)
-30
-25
-20
-15
-10
δ15N
(o/o
o)
4
6
8
10
12
14
16
18
30
A
B
C
Figure. 3-29. Monthly variation of carbon (○) and nitrogen (●)
stable isotope ratio (δ13C, ‰) of gastropoda Assiminea japonica (A), crab Ilyoplax pullisa (B) and isopoda Paranthura sp. (C).
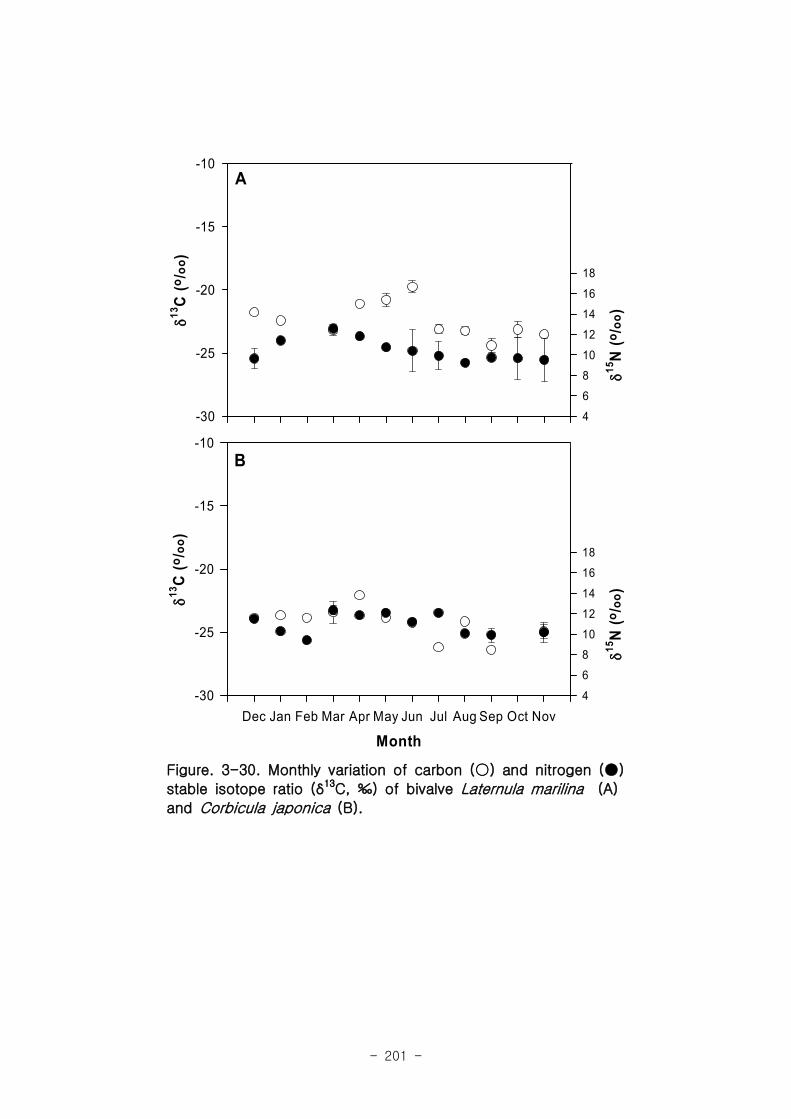
- 201 -
δ13C
(o/o
o)
-30
-25
-20
-15
-10
δ15N
(o/o
o)
4
6
8
10
12
14
16
18
30
Month
Dec Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov
δ13C
(o/o
o)
-30
-25
-20
-15
-10
δ15N
(o/o
o)
4
6
8
10
12
14
16
18
30
A
B
Figure. 3-30. Monthly variation of carbon (○) and nitrogen (●)
stable isotope ratio (δ13C, ‰) of bivalve Laternula marilina (A)
and Corbicula japonica (B).
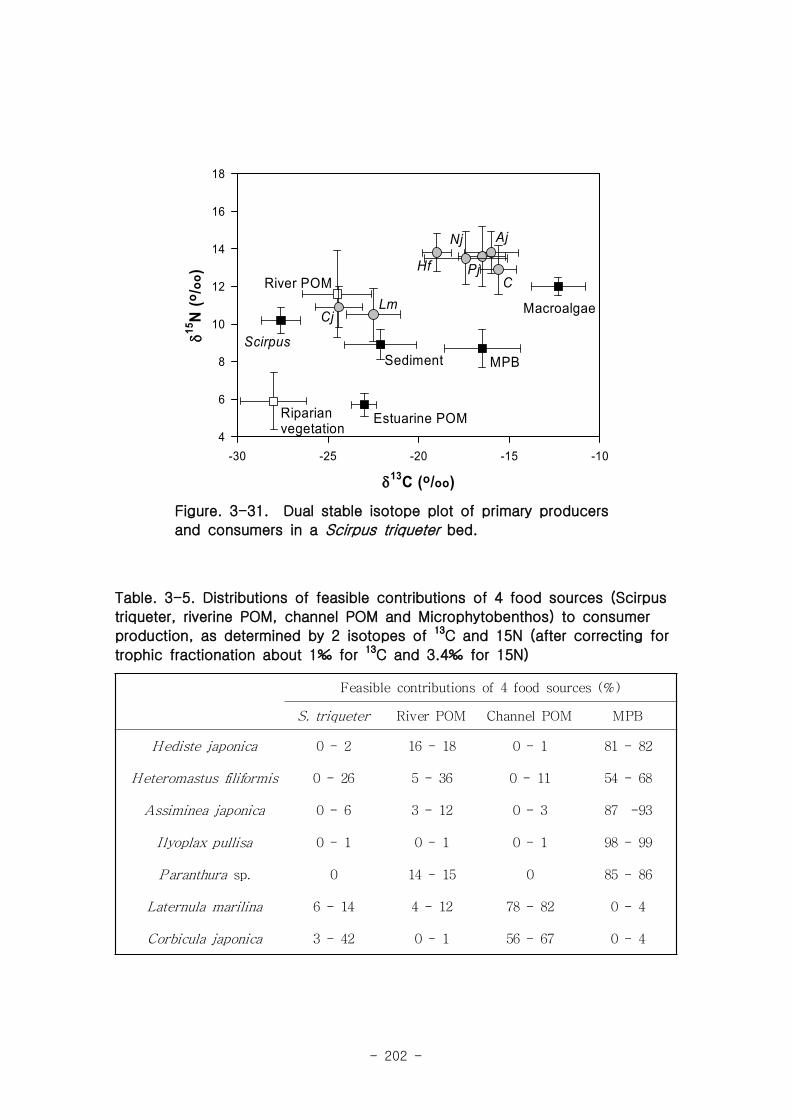
- 202 -
δ13
C (o/oo)
-30 -25 -20 -15 -10
δ15N
(o/o
o)
4
6
8
10
12
14
16
18
Scirpus
MPB
Estuarine POM
Sediment
Macroalgae
Riparian vegetation
River POM
CjLm
Aj
PjC
Hf
Nj
Figure. 3-31. Dual stable isotope plot of primary producers
and consumers in a Scirpus triqueter bed.
Feasible contributions of 4 food sources (%)
S. triqueter River POM Channel POM MPB
Hediste japonica 0 - 2 16 - 18 0 - 1 81 - 82
Heteromastus filiformis 0 - 26 5 - 36 0 - 11 54 - 68
Assiminea japonica 0 - 6 3 - 12 0 - 3 87 -93
I lyoplax pullisa 0 - 1 0 - 1 0 - 1 98 - 99
Paranthura sp. 0 14 - 15 0 85 - 86
Laternula marilina 6 - 14 4 - 12 78 - 82 0 - 4
Corbicula japonica 3 - 42 0 - 1 56 - 67 0 - 4
Table. 3-5. Distributions of feasible contributions of 4 food sources (Scirpus
triqueter, riverine POM, channel POM and Microphytobenthos) to consumer
production, as determined by 2 isotopes of 13C and 15N (after correcting for
trophic fractionation about 1‰ for 13C and 3.4‰ for 15N)

- 203 -
가. 갈대(Phragmites australis) 생태계 먹이망
낙동강 하구역에서 육지와 바다의 경계역에서 좁은 띠를 이루고 광활하게 분포
하고 있는 갈대는 하구전체에서 대단히 높은 생물량을 가진다. 이들로부터 유래한
유기물의 생태계내에서의 유용성을 평가하기 위하여 대형저서동물의 잠재적인 먹
이원으로서 가용성이 높은 기초생산자 식물군과 입자유기물질 그리고 이들에 대
한 소비자 동물들의 탄소와 질소 안정동위원소를 분석하였다.
Fig. 3-32 에서 볼 수 있듯이 갈대 엽체의 탄소안정동위원소 비값은 -25‰ 이
하로 다른 식물군들에 비하여 상대적으로 낮다는 것을 알 수 있는데, 이 비값들은
세모고랭이나 육상현화식물들이 가지는 값들과 유사하였다. 앞서 언급한 영양단계
에 따른 동위원소 분별효과를 고려해서 각 동물군들의 질소 안정동위원소 비값에
대하여 그들의 먹이원이 가질 것으로 예상되는 동위원소 비값을 구하기 위하여,
탄소와 질소 안정동위원소 비값에 대하여 각각 한 영양단계 당 0.8과 3.5‰을 빼
어서 추산했을 때, 대부분의 소비자 동물군들의 먹이원이 가질 것으로 예상되는
탄소와 질소 안정동위원소 비값의 위치는 그림에서 저서미세조류나 식물플랑크톤
값과 일치하거나 이들 두 먹이원이 가지는 동위원소 비값의 사이에 놓이는 것을
알 수 있다. 이와 같은 결과는 갈대서식지 내에서도 대부분의 동물군들이 갈대에
기원을 둔 유기물이 아니라 저서미세조류와 식물플랑크톤에 기원을 둔 유기물을
주로 이용하고 있다는 것을 잘 나타내어 준다. 갈대의 높은 생물량과 생산력에도
불구하고 여기서 유래한 탄소의 이용도가 낮다는 것은 조류와 고등식물사이의 영
양가(food value)의 차이에 기인한다는 것은 잘 알려져 있다.
Fig. 3-34는 이상에서 살펴본 세모고랭이 서식지의 주요 기초생산자와 소비자
동물군의 탄소와 질소 안정동위원소 비값을 함께 2차원 평면에 도시하였다. 어떤
동물이 먹이를 섭취하였을 때, 영양단계를 통하여 동위원소의 분별효과가 나타나
는데 대개 한 영양단계 당 탄소안정동위원소 비값은 약 +0.8‰, 질소안정동위원소
비값은 약 +3.5‰ 정도의 분별효과가 나타난다. 따라서 그림에서 나타난 바와 같
이 이와 같은 동위원소 분별효과를 고려했을 때, 두 종의 패류들이 하구 수중의
부유입자유기물을 구성하는 식물플랑크톤 기원의 유기물을 주로 이용하는 반면,
두 패류 종을 제외한 나머지 동물군들이 주로 이용하는 유기물은 저서미세조류에
그 기원을 두고 있다는 것을 알 수 있었다. 특히 세모고랭이 생태계에서는 달리
Erimacrus isenbeckii 같이 갈대의 의존하여 서식하는 몇몇 개체의 출현으로 아
주 다양한 유기물을 섭식하는 종이 공존하는 생태계가 형성되었다. 하지만 세모고
랭이 군락에서도 출현한 두 이매패류의 겨우 여전히 식물플랑크톤의 의존이 상당
히 높은 것으로 나타났다.
Table. 3-6은 갈대 서식지의 주요 기초생산자와 소비자 동물군의 기여를 모식
화 하였다. 주요기초생산자를 갈대, 수로의 식물플랑크톤 기원의 유기물, 저서미세
조류에 대한 세모고랭이군락의 대형저서동물의 상대적인 기여를 나타내었다. 여과
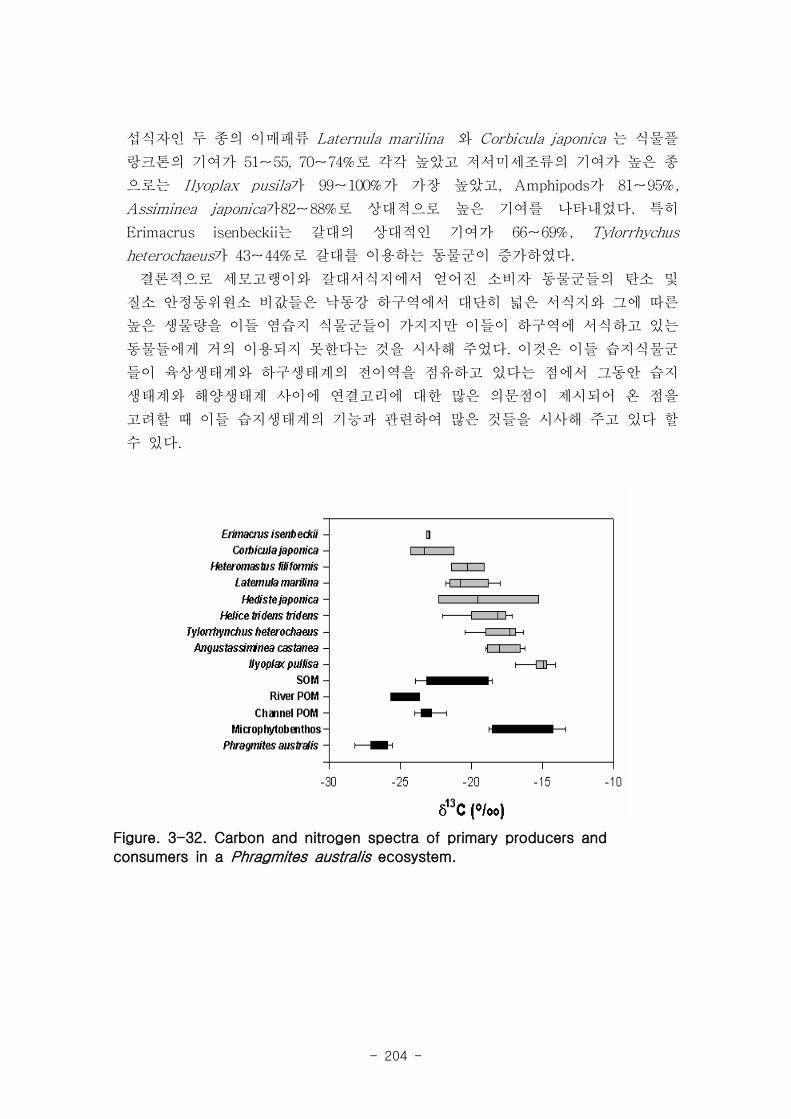
- 204 -
섭식자인 두 종의 이매패류 Laternula marilina 와 Corbicula japonica 는 식물플
랑크톤의 기여가 51~55, 70~74%로 각각 높았고 저서미세조류의 기여가 높은 종
으로는 I lyoplax pusila가 99~100%가 가장 높았고, Amphipods가 81~95%,
Assiminea japonica가82~88%로 상대적으로 높은 기여를 나타내었다. 특히
Erimacrus isenbeckii는 갈대의 상대적인 기여가 66~69%, Tylorrhychus
heterochaeus가 43~44%로 갈대를 이용하는 동물군이 증가하였다.
결론적으로 세모고랭이와 갈대서식지에서 얻어진 소비자 동물군들의 탄소 및
질소 안정동위원소 비값들은 낙동강 하구역에서 대단히 넓은 서식지와 그에 따른
높은 생물량을 이들 염습지 식물군들이 가지지만 이들이 하구역에 서식하고 있는
동물들에게 거의 이용되지 못한다는 것을 시사해 주었다. 이것은 이들 습지식물군
들이 육상생태계와 하구생태계의 전이역을 점유하고 있다는 점에서 그동안 습지
생태계와 해양생태계 사이에 연결고리에 대한 많은 의문점이 제시되어 온 점을
고려할 때 이들 습지생태계의 기능과 관련하여 많은 것들을 시사해 주고 있다 할
수 있다.
Figure. 3-32. Carbon and nitrogen spectra of primary producers and
consumers in a Phragmites australis ecosystem.
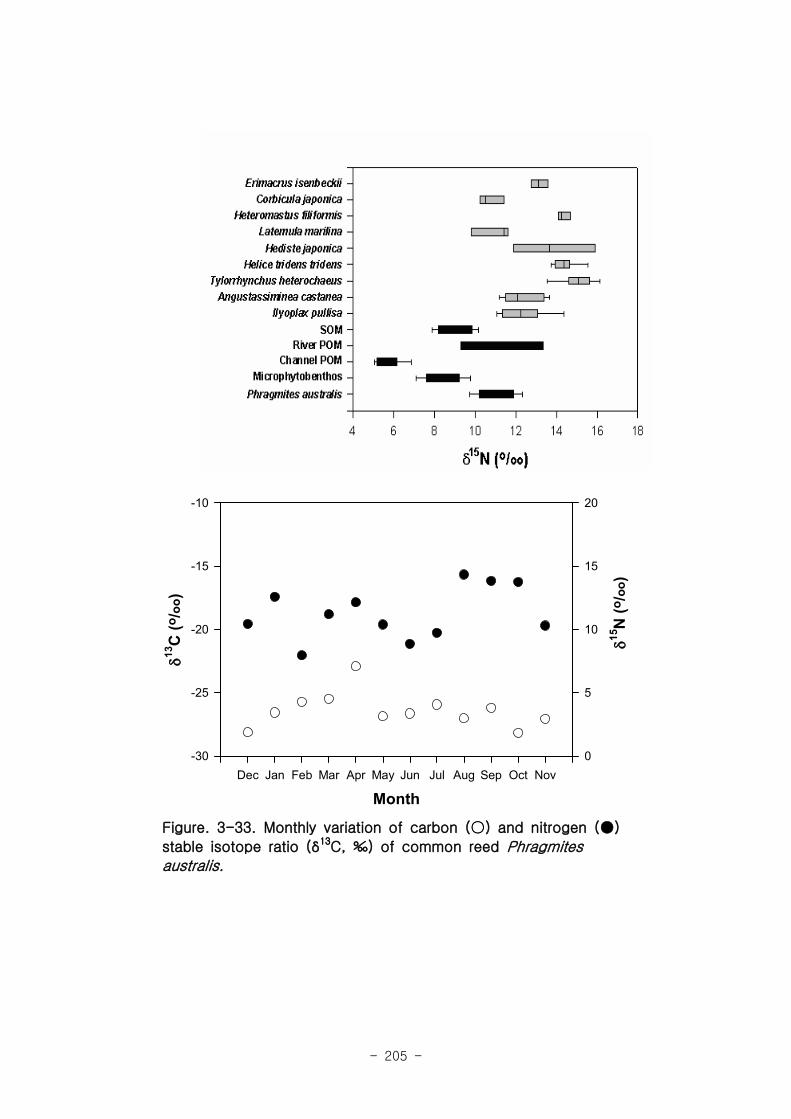
- 205 -
Month
Dec Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov
δ13C
(o/o
o)
-30
-25
-20
-15
-10
δ15N
(o/o
o)
0
5
10
15
20
Figure. 3-33. Monthly variation of carbon (○) and nitrogen (●)
stable isotope ratio (δ13C, ‰) of common reed Phragmites australis.
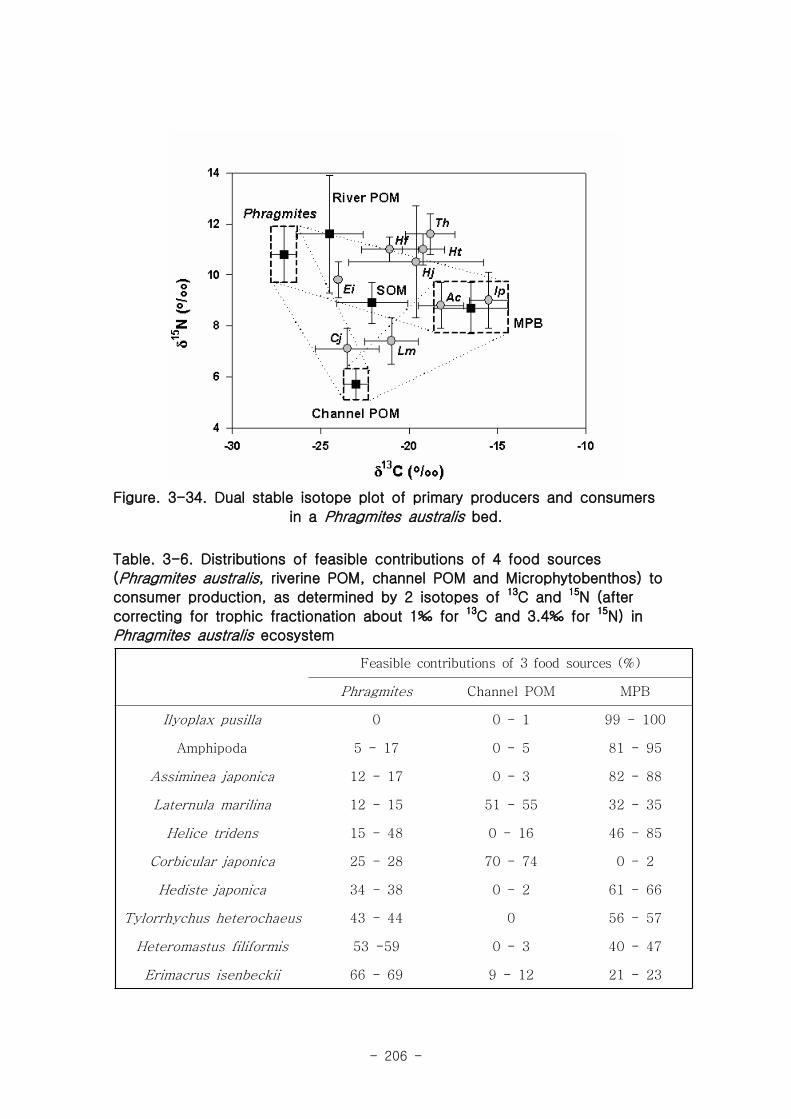
- 206 -
Figure. 3-34. Dual stable isotope plot of primary producers and consumers
in a Phragmites australis bed.
Table. 3-6. Distributions of feasible contributions of 4 food sources
(Phragmites australis, riverine POM, channel POM and Microphytobenthos) to
consumer production, as determined by 2 isotopes of 13C and 15N (after
correcting for trophic fractionation about 1‰ for 13C and 3.4‰ for 15N) in
Phragmites australis ecosystem
Feasible contributions of 3 food sources (%)
Phragmites Channel POM MPB
Ilyoplax pusilla 0 0 - 1 99 - 100
Amphipoda 5 - 17 0 - 5 81 - 95
Assiminea japonica 12 - 17 0 - 3 82 - 88
Laternula marilina 12 - 15 51 - 55 32 - 35
Helice tridens 15 - 48 0 - 16 46 - 85
Corbicular japonica 25 - 28 70 - 74 0 - 2
Hediste japonica 34 - 38 0 - 2 61 - 66
Tylorrhychus heterochaeus 43 - 44 0 56 - 57
Heteromastus filiformis 53 -59 0 - 3 40 - 47
Erimacrus isenbeckii 66 - 69 9 - 12 21 - 23

- 207 -
나. 갯벌생태계와 조하대
세모고랭이와 갈대로 이루어진 하구 습지와 대형식물이 존재하지 않는 갯벌사
이의 연결고리를 파악하기 위하여 갯벌에 서식하고 있는 대형무척추동물과 수로
에서 어획된 어류의 탄소 및 질소 안정동위원소 비값을 분석하여 낙동강 하구 생
태계에서 높은 생물량을 가지는 식물군과 입자물질 기원물질에 대한 각각의 비값
을 비교하였다. 1차년도 연구에서 공간적인 분포를 조사한데 이어 2차년도 본 연
구에서는 특정 생물종들의 동위원소 비값의 연변동을 조사하여 유기물 이용에 있
어서 계절적인 변동이 나타나는지를 평가하고 수로에서 어획된 어류에 대한 분석
을 수행하였다.
Fig. 3-35 은 부유물 섭식자 이매패류인 빛조개에 대한 탄소와 질소안정동위원
소 비값의 월별변동을 나타낸 것이다. 이 조개는 탄소안정동위원소 비값에서 약
-20‰내외의 값으로 대단히 안정된 비값을 연중 유지하는 것으로 나타났다. Fig.
3-36에서 볼 수 있듯이 이들이 가지는 안정동위원소 비값은 영양단계에 따른 분
별효과를 고려했을 때 저서미세조류와 식물플랑크톤을 거의 반반씩 이용하고 있
는 것으로 나타났다.
Fig. 3-37는 이상에서 살펴본 갯벌생태계 서식지의 주요 기초생산자와 소비자
동물군의 탄소와 질소 안정동위원소 비값을 함께 2차원 평면에 도시하였다. 어떤
동물이 먹이를 섭취하였을 때, 영양단계를 통하여 동위원소의 분별효과가 나타나
는데 대개 한 영양단계 당 탄소안정동위원소 비값은 약 +0.8‰, 질소안정동위원소
비값은 약 +3.5‰ 정도의 분별효과가 나타난다. 따라서 그림에서 나타난 바와 같
이 이와 같은 동위원소 분별효과를 고려했을 때, 두 종의 패류들이 하구 수중의
부유입자유기물을 구성하는 식물플랑크톤 기원의 유기물을 주로 이용하는 반면,
Calliansa japonica 의 탄소 및 질소 안정동위원소 비 값은 각각 -15.2 ± 2.4, 12.2
± 0.7‰로 저서미세조류의 의존이 상당히 높음을 알 수 있다. 특히 Batillaria
cimingii 는 탄소 및 질소 안정동위원소 비 값은 각각 -9.1 ± 0.5, 14.9 ± 0.6‰,
Bullacta exarata 탄소 및 질소 안정동위원소 비 값은 각각 -7.9 ± 1.1, 13.7 ± 0.
7‰로 매우 낮은 탄소정동위원소비 값을 가지는 제 3의 source의 기여가 상당히
높음을 알 수 있다. 추정컨대 많은 연구결과에서 -9~-14‰의 아주 낮은 값을 가
지는 cyanobacteria의 기여가 있는 것으로 판단된다.
한편 낙동강 하구역에 서식하는 어류의 탄소안정동위원소와 질소안정동위원소
의 비 값을 보면(Table. 3-7), 탄소안정동위원소의 평균값은 -18.5 ± 2.9‰이고,
질소안정동위원소의 경우 평균 -14.5 ± 2.5‰이었다. 우점하는 어류의 경우 어체
장에 따라서 임의로 소, 중, 대로 나누어 분석하였다. 낙동강 하구역에 서식하는
문절망둑의 경우 높은 평균 14.5‰의 탄소안정동위원소를 보여 하구역에서 생산되
는 자체유기물에 상당히 의존함을 알 수 있으나, 대부분의 어류는 식물플랑크톤과
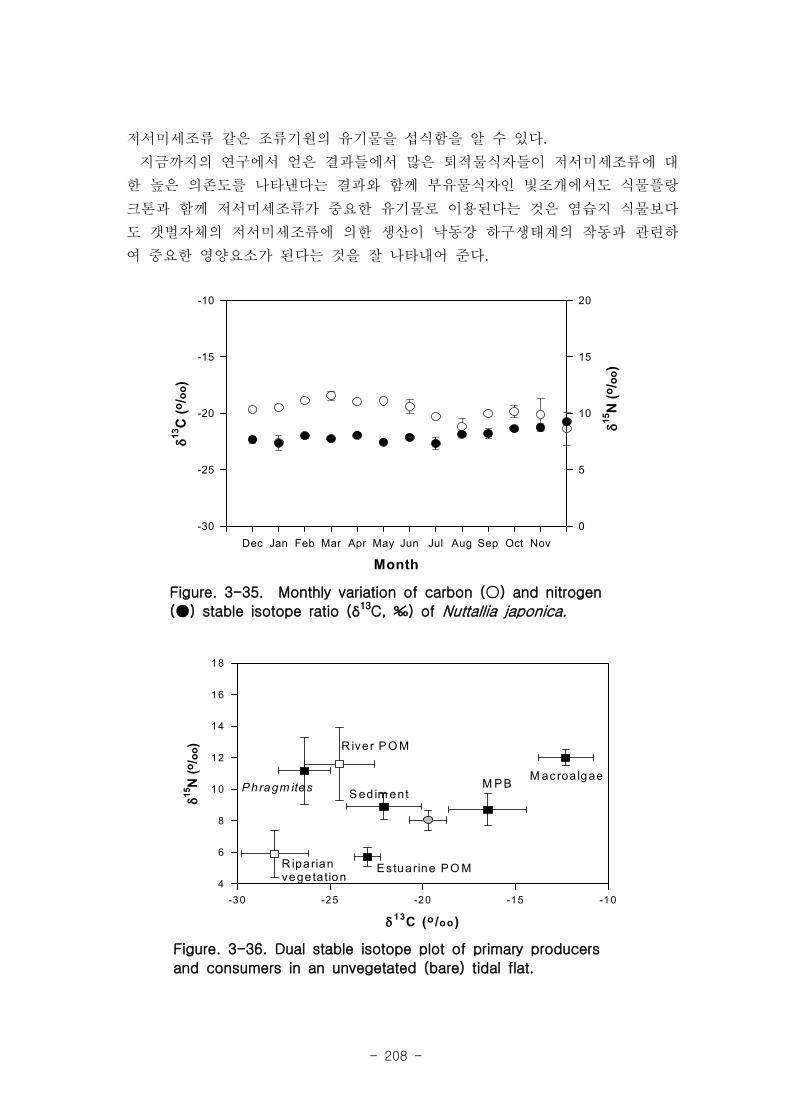
- 208 -
저서미세조류 같은 조류기원의 유기물을 섭식함을 알 수 있다.
지금까지의 연구에서 얻은 결과들에서 많은 퇴적물식자들이 저서미세조류에 대
한 높은 의존도를 나타낸다는 결과와 함께 부유물식자인 빛조개에서도 식물플랑
크톤과 함께 저서미세조류가 중요한 유기물로 이용된다는 것은 염습지 식물보다
도 갯벌자체의 저서미세조류에 의한 생산이 낙동강 하구생태계의 작동과 관련하
여 중요한 영양요소가 된다는 것을 잘 나타내어 준다.
Month
Dec Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov
δ13C
(o/ o
o)
-30
-25
-20
-15
-10
δ15N
(o/ o
o)
0
5
10
15
20
Figure. 3-35. Monthly variation of carbon (○) and nitrogen
(●) stable isotope ratio (δ13C, ‰) of Nuttallia japonica.
δ13
C (o /oo )
-30 -25 -20 -15 -10
δ15N
(o/ o
o)
4
6
8
10
12
14
16
18
Phragm ites MPB
Estuarine POM
Sediment
M acroa lgae
R iparian vegetation
R iver POM
Figure. 3-36. Dual stable isotope plot of primary producers
and consumers in an unvegetated (bare) tidal flat.
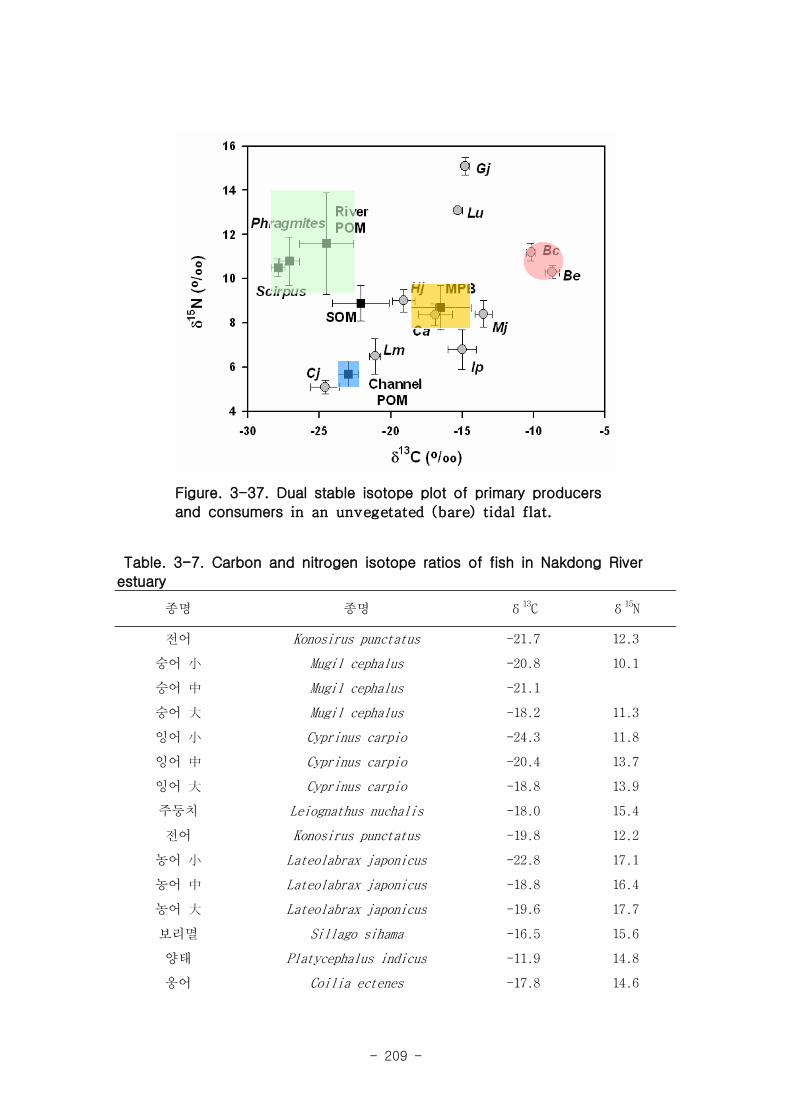
- 209 -
Table. 3-7. Carbon and nitrogen isotope ratios of fish in Nakdong River
estuary
종명 종명 δ13C δ
15N
전어 Konosirus punctatus -21.7 12.3
숭어 小 Mugil cephalus -20.8 10.1
숭어 中 Mugil cephalus -21.1
숭어 大 Mugil cephalus -18.2 11.3
잉어 小 Cyprinus carpio -24.3 11.8
잉어 中 Cyprinus carpio -20.4 13.7
잉어 大 Cyprinus carpio -18.8 13.9
주둥치 Leiognathus nuchalis -18.0 15.4
전어 Konosirus punctatus -19.8 12.2
농어 小 Lateolabrax japonicus -22.8 17.1
농어 中 Lateolabrax japonicus -18.8 16.4
농어 大 Lateolabrax japonicus -19.6 17.7
보리멸 Sillago sihama -16.5 15.6
양태 Platycephalus indicus -11.9 14.8
웅어 Coilia ectenes -17.8 14.6
Figure. 3-37. Dual stable isotope plot of primary producers
and consumers in an unvegetated (bare) tidal flat.

- 210 -
동갈치 Ablennes anastomella -16.6 15.6
몰개 Squalidus japonicus Koreanus -23.1 15.3
황복 Fugu ocellatus obcurus -18.8 13.4
갯장어 Muraenesox cinereus -18.0 8.0
점농어 Lateolabrax sp. -16.6 15.3
양태 Platycephalus indicus -19.3 15.9
밴댕이 Harengula zunasi -17.1 13.9
문절망둑 小 Acanthogobius flavimanus -14.3 17.6
문절망둑 中 Acanthogobius flavimanus -14.2 18.3
문절망둑 大 Acanthogobius flavimanus -14.9 17.8
Fig. 3-38는 이상에서 살펴본 낙동강 하구의 세모고랭이 군락, 갈대군락, 갯벌,
조하대 서식지의 주요 소비자 동물군의 탄소와 질소 안정동위원소 비값을
Hierarchical clustering (Bray and Curtis index method)한 결과를 도시하였다.
그 결과 크게 A, B, C의 세 그룹으로 나누어 졌고, A 그룹에서 3가지 subgroup
으로 나누어 졌다. B group은 갯벌의 Goniada japonica와 Lumbrinerris sp.와 어
류인 Acanthogobius flavimanus의 3종이 속했다. 이 세 종은 질소 안정동위원소
비 값이 매우 높은 top predator로써 분류학적으로도 두 종의 다모류는 강한 이빨
을 이용하여 육식성을 하는 종으로 알려져 있다. 특이하게 Acanthogobius
flavimanus는 낙동강 하구에 주로 서식하는 토착종으로 알려져 있는 종으로 다른
어류의 질소 안정 동위원소 비값이 다른 대형 저서동물에 비해 낮거나 비슷한 것
으로 보아 낙동강 하구에서의 섭이활동은 거의 하지 않는 회유종인 것으로 판단
된다. C group에 속한 Bullacta exarata, Batillaria cumingii, M acrophthalmus
japonicus는 갯벌의 아주 높은 탄소 안정동위원소 기원의 source에 강하게 의존하
는 종으로 나타났다. A group은 세 개의 subgroup으로 이루어 졌다. a1에 속한는
종들은 여과섭식성 이매패류들로써 Laternula marilina나 Corbicular sp.로 서식처
에 관계없이 모두 이 그룹에 속하였다. a3에 속한 종들은 Hediste japonica(c)와
Cesarma dehaani(p), Diopatra sp.를 제외하고는 대부분이 어류가 이 그룹에 속
하였고, 이 그룹에 속한 종들은 대부분 omnivore인 것으로 보인다. 가장 많은 종
이 속한 a2 group은 퇴적물 섭식자가 여기에 속했다.
이와 같이 전체 생태계내에 먹이망을 연구하는 데에 있어 구성 소비자 동물군
의 feeding mode에 따라 나누어서 해석하는 것이 중요하며 퇴적물식자들이 저서
미세조류에 대한 높은 의존도를 나타내며 대부분의 퇴적물 섭식자들이 B group
와 같은 고차 소비자동물군으로 연결되는 역할을 하는 것으로 판단된다.
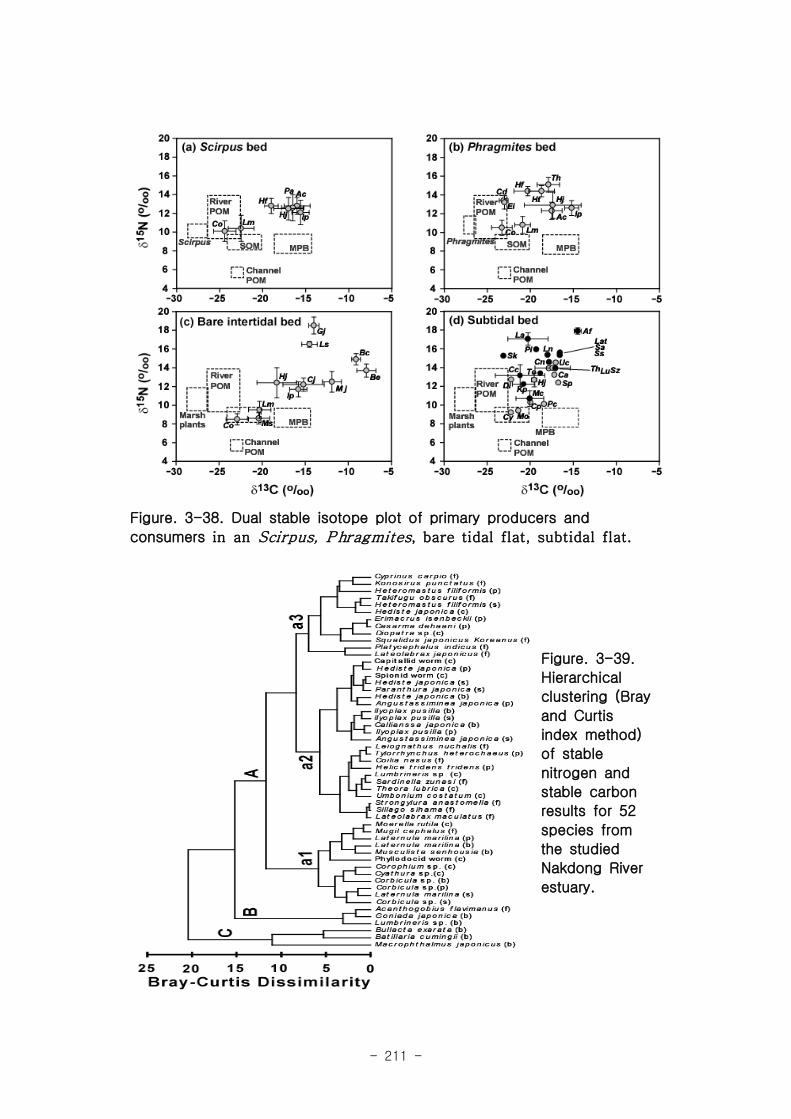
- 211 -
Figure. 3-38. Dual stable isotope plot of primary producers and
consumers in an Scirpus, Phragmites, bare tidal flat, subtidal flat.
Figure. 3-39.
Hierarchical
clustering (Bray
and Curtis
index method)
of stable
nitrogen and
stable carbon
results for 52
species from
the studied
Nakdong River
estuary.

- 212 -
3. 물질수지
하구내로의 주요 물질 유입원은 강이다. 하구는 육상기원 물질의 운반 통로로서
무기물(토양, 중금속, 영양염)이나 유기물(식물체나 동물체의 사체, 하수)등이 하구
를 통해 연안으로 유입된다. 하구는 단순히 통로로서의 역할을 할 뿐만 아니라 자
체가 풍부한 생태계로서 여러 변화과정이 일어나게 된다 이러한 생지화학적 순환
과정은 하구 생태계 및 연안 생태계에 큰 영향을 줄 뿐만 아니라, 향후 환경압력
에 하구 생태계가 어떻게 반응할 것인가를 알 수 있게 한다. 본 연구에서는 낙동
강 하구의 물질 유입과 생지화학적인 물질 변환 속도를 추정해 보려고 하였다.
낙동강 하구를 하나의 시스템으로 보았을 때 유기물 및 영양염의 주요 유입원
은 담수를 통한 유입이다. 유입량을 추정하여 이들이 낙동강 하구생태계에 어떻게
영향을 끼칠 수 있는지를 알아보기 위하여, 한국수자원공사의 협조를 얻어, 200
4~5년 동안 낙동강 하구둑의 일간 방류량 자료를 구하였으며, 해양수산부의 원격
측정망 자료로부터 일간 수질 변화 자료를 얻어 담수와 유기물 및 영양염 유입량
을 확인하였다. 이와 함께 생산자, 소비자, 분해자의 양과 변환속도를 추정하였으
며, 계절변화를 살펴보았다.
가. 낙동강을 통한 물질의 유입
낙동강 하구는 하구둑과 사구 섬사이의 지역을 말하며, 면적은 96km2에 이르며,
평균수심은 5m 정도이다 (Table. 3-8). 낙동강 하구의 담수유입은 7~9월에 집중
되어, 일간 200~600 x 106 입방미터에 이른다. 이 계절을 제외한 건기에는 10 x
106 입방미터 이하의 담수 유입량을 보인다 (Fig. 3-40). 부유물질, 영양염, COD
등의 유입도 담수 유입과 같은 계절변화를 보여, 7~9월사이에 집중되게 되며 이
시기에 부유물질 유입량은 하루에 20,000ton 에 달하기도 한다. 7~9월사이 COD
유입량은 연간변화를 보이나 일간 1000~2000 톤 정도이다.
1160 mm/yrPrecipitation
2000 x103 m3d-1Sewage input
41,000 x103 m3d-1Water input
5 m Depth
96 km2Area
1160 mm/yrPrecipitation
2000 x103 m3d-1Sewage input
41,000 x103 m3d-1Water input
5 m Depth
96 km2Area
Table. 3-8. 낙동강 하구 면적과 평균수심,
강수량과 담수유입량 (한국수자원공사)
- 213 -
0
100
200
300
400
500
600
700
Jan-05 May-05 Sep-05 Jan-06 May-06 Sep-06
Wat
er d
isch
arge
(x 1
06 m
3d
-1) .
0
2
4
6
8
COD (m
g L
-1).
Water
COD
0
5000
10000
15000
20000
25000
Jan-05 May-05 Sep-05 Jan-06 May-06 Sep-06
SS d
isch
arg (to
n d
-1) .
0
10
20
30
40
50
60
70
80
TP d
isch
arge
(ton
day
-1).
SS
TP
C. COD discharge
0
500
1000
1500
2000
2500
3000
Jan-05 May-05 Sep-05 Jan-06 May-06 Sep-06
COD d
isch
arg (to
n d
-1) .
0
100
200
300
400
500
600
700
Jan-05 May-05 Sep-05 Jan-06 May-06 Sep-06
Wat
er d
isch
arge
(x 1
06 m
3d
-1) .
0
2
4
6
8
COD (m
g L
-1).
Water
COD
0
5000
10000
15000
20000
25000
Jan-05 May-05 Sep-05 Jan-06 May-06 Sep-06
SS d
isch
arg (to
n d
-1) .
0
10
20
30
40
50
60
70
80
TP d
isch
arge
(ton
day
-1).
SS
TP
C. COD discharge
0
500
1000
1500
2000
2500
3000
Jan-05 May-05 Sep-05 Jan-06 May-06 Sep-06
COD d
isch
arg (to
n d
-1) .
Figure. 3-40. 2005~6년 동안 낙동강 하구로 유입된 일간 담수, 부유물질(SS),
총인 (TP), COD 유입량의 계절변화
나. 각 생태 과정과의 비교
낙동강 하구로의 물질 유입량의 계절변화를 낙동강 하구에서 일어나는 여러 생
태과정과 비교해 보았다. 물질 유입이 집중되는 7~9월동안 하구 퇴적물에서 일어
나는 유기물 분해과정도 최대값을 보이며, 세모고랭이와 갈대와 같은 대형일차생
산자의 생체량도 최대값을 보이고 있다 (Fig. 3-41)
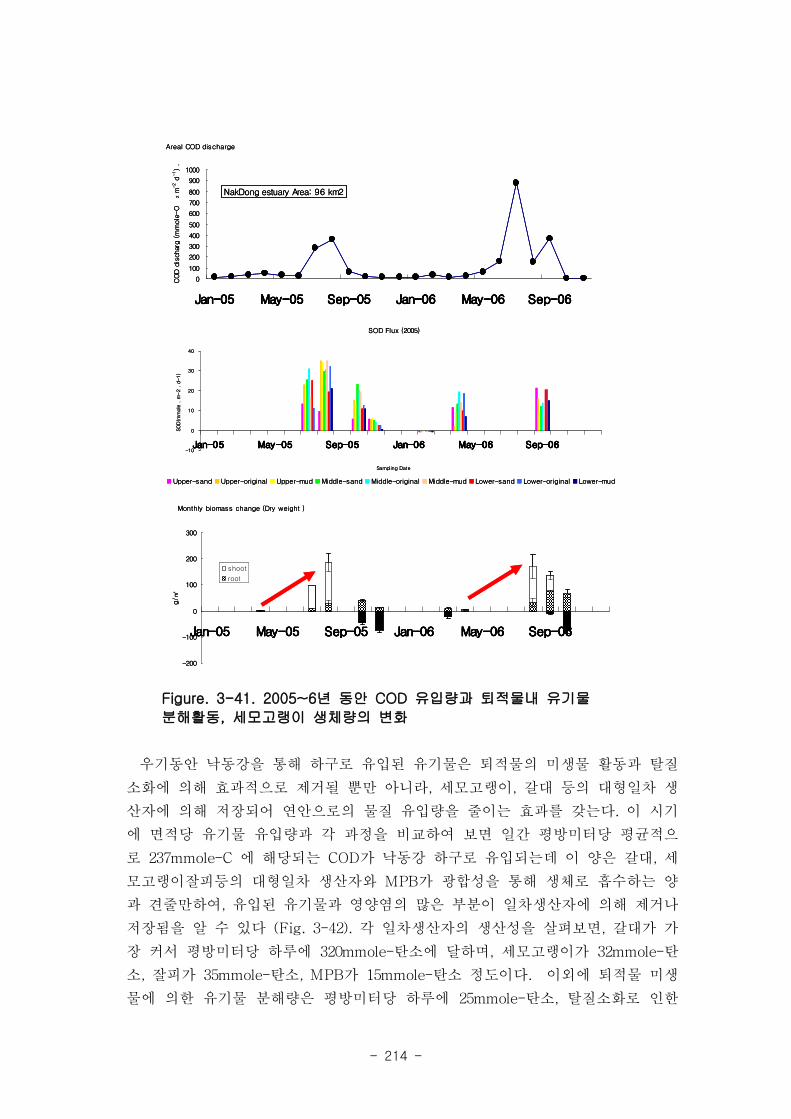
- 214 -
Areal COD discharge
0
100
200
300
400
500
600
700
800
900
1000
Jan-05 May-05 Sep-05 Jan-06 May-06 Sep-06
CO
D d
ischarg
(m
mole
-O
2 m
-2 d
-1)
.
NakDong estuary Area: 96 km2
SOD Flux (2005)
-10
0
10
20
30
40
Jan-05 May-05 Sep-05 Jan-06 May-06 Sep-06
Sampling Date
SO
D(m
mole
. m
-2 .
d-1)
Upper-sand Upper-original Upper-mud Middle-sand Middle-original Middle-mud Lower-sand Lower-original Lower-mud
Monthly biomass change (Dry weight )
-200
-100
0
100
200
300
Jan-05 May-05 Sep-05 Jan-06 May-06 Sep-06
g/㎡
shoot
root
Areal COD discharge
0
100
200
300
400
500
600
700
800
900
1000
Jan-05 May-05 Sep-05 Jan-06 May-06 Sep-06
CO
D d
ischarg
(m
mole
-O
2 m
-2 d
-1)
.
NakDong estuary Area: 96 km2
SOD Flux (2005)
-10
0
10
20
30
40
Jan-05 May-05 Sep-05 Jan-06 May-06 Sep-06
Sampling Date
SO
D(m
mole
. m
-2 .
d-1)
Upper-sand Upper-original Upper-mud Middle-sand Middle-original Middle-mud Lower-sand Lower-original Lower-mud
Monthly biomass change (Dry weight )
-200
-100
0
100
200
300
Jan-05 May-05 Sep-05 Jan-06 May-06 Sep-06
g/㎡
shoot
root
Areal COD discharge
0
100
200
300
400
500
600
700
800
900
1000
Jan-05 May-05 Sep-05 Jan-06 May-06 Sep-06
CO
D d
ischarg
(m
mole
-O
2 m
-2 d
-1)
.
NakDong estuary Area: 96 km2
SOD Flux (2005)
-10
0
10
20
30
40
Jan-05 May-05 Sep-05 Jan-06 May-06 Sep-06
Sampling Date
SO
D(m
mole
. m
-2 .
d-1)
Upper-sand Upper-original Upper-mud Middle-sand Middle-original Middle-mud Lower-sand Lower-original Lower-mud
Monthly biomass change (Dry weight )
-200
-100
0
100
200
300
Jan-05 May-05 Sep-05 Jan-06 May-06 Sep-06
g/㎡
shoot
root
Figure. 3-41. 2005~6년 동안 COD 유입량과 퇴적물내 유기물
분해활동, 세모고랭이 생체량의 변화
우기동안 낙동강을 통해 하구로 유입된 유기물은 퇴적물의 미생물 활동과 탈질
소화에 의해 효과적으로 제거될 뿐만 아니라, 세모고랭이, 갈대 등의 대형일차 생
산자에 의해 저장되어 연안으로의 물질 유입량을 줄이는 효과를 갖는다. 이 시기
에 면적당 유기물 유입량과 각 과정을 비교하여 보면 일간 평방미터당 평균적으
로 237mmole-C 에 해당되는 COD가 낙동강 하구로 유입되는데 이 양은 갈대, 세
모고랭이잘피등의 대형일차 생산자와 MPB가 광합성을 통해 생체로 흡수하는 양
과 견줄만하여, 유입된 유기물과 영양염의 많은 부분이 일차생산자에 의해 제거나
저장됨을 알 수 있다 (Fig. 3-42). 각 일차생산자의 생산성을 살펴보면, 갈대가 가
장 커서 평방미터당 하루에 320mmole-탄소에 달하며, 세모고랭이가 32mmole-탄
소, 잘피가 35mmole-탄소, MPB가 15mmole-탄소 정도이다. 이외에 퇴적물 미생
물에 의한 유기물 분해량은 평방미터당 하루에 25mmole-탄소, 탈질소화로 인한
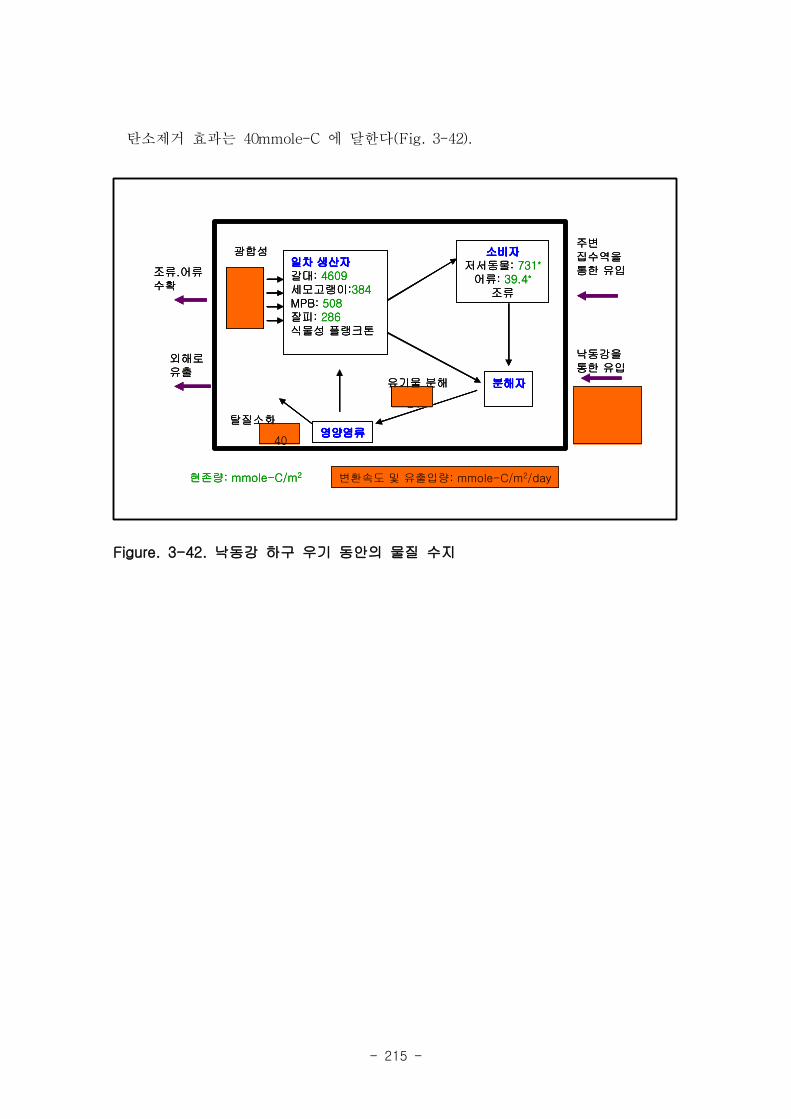
- 215 -
일차 생산자갈대: 4609세모고랭이:384MPB: 508잘피: 286식물성 플랭크톤
소비자저서동물: 731*
어류: 39.4*
조류
분해자
영양염류
낙동강을통한 유입
주변집수역을통한 유입
외해로유출
24
광합성
COD: 237TN: 274TP: 92
현존량: mmole-C/m2 변환속도 및 유출입량: mmole-C/m2/day
유기물 분해
40
탈질소화
조류.어류수확 320
32
3515
일차 생산자갈대: 4609세모고랭이:384MPB: 508잘피: 286식물성 플랭크톤
소비자저서동물: 731*
어류: 39.4*
조류
분해자
영양염류
낙동강을통한 유입
주변집수역을통한 유입
외해로유출
24
광합성
COD: 237TN: 274TP: 92
현존량: mmole-C/m2 변환속도 및 유출입량: mmole-C/m2/day
유기물 분해
40
탈질소화
조류.어류수확 320
32
3515
일차 생산자갈대: 4609세모고랭이:384MPB: 508잘피: 286식물성 플랭크톤
소비자저서동물: 731*
어류: 39.4*
조류
분해자
영양염류
낙동강을통한 유입
주변집수역을통한 유입
외해로유출
24
광합성
COD: 237TN: 274TP: 92
COD: 237TN: 274TP: 92
현존량: mmole-C/m2 변환속도 및 유출입량: mmole-C/m2/day
유기물 분해
40
탈질소화
조류.어류수확 320
32
3515
320
32
3515
탄소제거 효과는 40mmole-C 에 달한다(Fig. 3-42).
Figure. 3-42. 낙동강 하구 우기 동안의 물질 수지

- 216 -
제 4절 낙동강 하구의 생태 구성 요소의 복원
1. 잘피
가. 인공 잘피림 조성을 통한 하구 생태계 복원
(1) 인공 잘피림 조성을 위한 적지 조사
낙동강 하구지역에서 인공 잘피림을 조성에 적합한 장소를 선정하기 위해 수온
과 수중 광량을 진우도 부근에서 측정하였다. 수온은 HOBO 센서를 이용해서 매
15분마다 측정하였으며 일일평균으로 나타내었다. 수중 광량(Underwater
irradiance)은 sensor와 logger (HOBO, Onset Computer)를 현장에 설치하여 매
15분 간격으로 측정하였다. Hobo data logger를 이용해 측정된 빛의 양 (Lumen
ft-1
)은 LI-4000 data loggar와 LI-193SA spherical quanatum sensor (LI-COR,
Inc.)를 이용해서 측정된 값을 통해서 photon flux density(μ㏖ photons m-2
sec-1
)로 변환하였다. 이렇게 변환된 값을 가지고 일일평균 photon flux density(㏖
m-2 d-1
)를 계산하였다.
(2) 인공 잘피림 조성
인공 잘피림 조성을 위해 예비조사를 통해서 진우도 앞 자연 잘피림과 가까운
곳에 이식 장소를 선정하였고 2차년에 걸쳐 진행되었다. 2004년 11월 수행된 이식
은 staple method를 이용해서 2 × 2 m (1unit)내에 20㎝ 간격으로 잘피 2500 개
체를 이식하였으며, 총 13 units를 조성하였다. 2005년 12월에 수행된 2차 이식도
1차 이식과 동일한 방법으로 수행하였고, 2 × 2 m (1unit)내에 30㎝ 간격으로 잘
피 2000개체를 이식하였으며, 총 10 units를 조성하였다.
(3) 이식 개체와 자연 개체와의 광합성 특성 비교
인공적으로 조성된 잘피 개체들이 자연 잘피림과 비교해서 안정적으로 이식되
어 정착되었는지를 알아보기 위해서 Diving-PAM을 이용해서 광합성 특성을 조
사하였다. 잘피의 형광 측정은 현장에서 측정하는 것을 원칙으로 하나, 불가피하
게 현장조사가 불가능할 시, 샘플을 실험실에 옮겨와 수행하였다. 이것은 해조류
와 잘피의 광합성 특성은 증가하는 광 단계에 대한 잎의 광합성 반응으로서 단시
간 내에 측정된다면, 실험실에서 측정해도 크게 영향을 받지 않는다(Hemminga
and Duarte 2000). 실험 재료는 이식된 잘피림과 자연 잘피림에 각각 10개체씩
채집하여 잘피 잎에 붙어 있는 부착생물의 영향을 최소화하기 위해서 가장 신선

- 217 -
한 두 번째 잎을 선택하였고, 잎의 위치를 일정하게 유지하기 위해서 sheath로부
터 10㎝위를 측정하였다. 엽록소형광을 측정하기 전 잘피 잎에 붙어 있는 부착생
물의 영향을 최소화하기 위해 각 부위의 부착생물은 부드러운 솔과 끌을 이용해
서 제거하였다. 광량은 0~2160μmol photons m-2 sec
-1 범위내에서 진행되었으며
총 8단계의 빛을 비추어 측정되었고(Rapid Light Curves), 8㎜ fiberoptics를 사용
하였다. 모든 개체는 ' dark leaf clip'를 이용해서 약 10분간 암적응 시켰으며,
RLC를 측정하는 동안, 각 단계마다 10초씩 빛에 노출시킨 후, 약 0.8초씩 강한 빛
에 노출시켰다. 일반적으로 육상식물의 전자전달속도(Electron Transport Rate -
ETR)은 다음과 같은 식으로 계산되어 진다(Schreiber et al. 1997; Ralph et al.
1998).
ETR = [(Fm-Fo) / Fm] × PAR × 0.84 × 0.5
Fo는 암적응 상태에서의 초기형광, Fm는 암적응된 상태에서 포화광에 의한 최
대형광을 나타내고 두 값을 이용해서 암적응 상태에서 광계Ⅱ (photosynthesis
system Ⅱ)의 광화학 반응에 대한 최대 양자수율 (maximum quantum yield -
[(Fm-Fo) / Fm])을 도출하였다. PAR는 광량 (photosynthetic active radiation: μ
mol photons m-2 s
-1)이고, 0.84는 식물 엽체에 흡수된 빛의 비율을 의미한다. 엽
록소형광 측정은 광계Ⅱ에서만 진행되므로 0.5를 곱하였다(보통 엽록소에 흡수된
빛은 광계Ⅰ과 광계Ⅱ에 똑같은 비율로 나눠진다고 가정하기 때문에 0.5를 곱한
다). 본 연구에서는 대상 종의 엽체에서 흡수된 빛을 직접 측정하지 않고 일반적
으로 고등 식물에서 통용되는 0.84를 이용하였으므로, 이하에서는 상대 전자전달
속도(rETR; relative electron transport rate)로 표현한다. Diving-PAM을 이용하
여 측정된 광도에 따른 전자전달율(relative ETR)은 비선형 회귀법으로 적합화하
여 매개변수들을 구했다. RLCs는 Platt et al. (1980)에 의해 고안된 방법을 이용
해서 나타낼 수 있다.
P = Ps [1-exp(-αEd/Ps)]exp(-βEd/Ps) ⑴
P는 구하고자하는 총 일차생산력, Ed는 광합성에 이용되는 파장의 광량
(wavelength 400~700㎚), Ps는 광저해 현상이 없을 때의 최대 잠재 전자전달속
도로서 정의되는 매개변수, α는 광합성의 최대 양자수율(maximum quantum
yield)과 관련된 매개변수로서 세포가 입사광을 모을 수 있는 능력을 나타내는 값
으로, 포화광 이하에서 광합성-광도 모델의 초기 기울기를 나타내고 β는 광저해
정도를 나타내는 매개변수이다. 광저해 현상이 없을 때 (β = 0) 식 ⑴은 다음과
같이 요약할 수 있다.
P = Pm [1-exp(-αEd/Pm)] ⑵
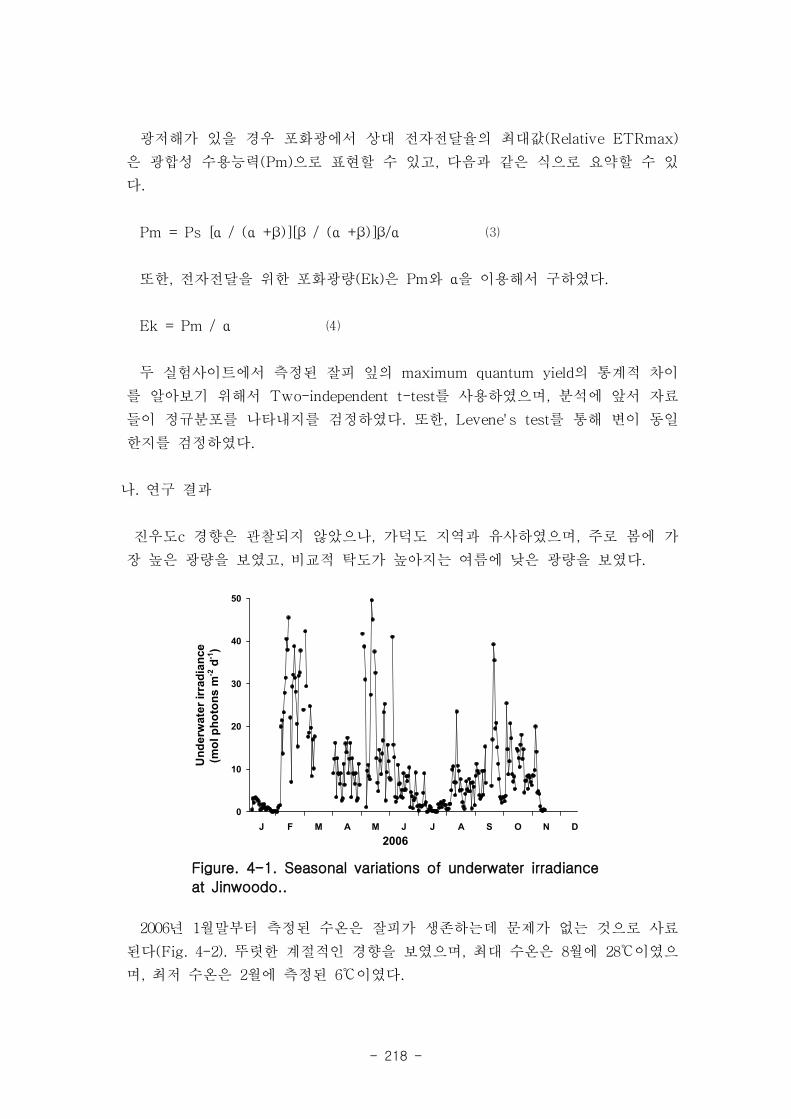
- 218 -
광저해가 있을 경우 포화광에서 상대 전자전달율의 최대값(Relative ETRmax)
은 광합성 수용능력(Pm)으로 표현할 수 있고, 다음과 같은 식으로 요약할 수 있
다.
Pm = Ps [α / (α +β)][β / (α +β)]β/α ⑶
또한, 전자전달을 위한 포화광량(Ek)은 Pm와 α을 이용해서 구하였다.
Ek = Pm / α ⑷
두 실험사이트에서 측정된 잘피 잎의 maximum quantum yield의 통계적 차이
를 알아보기 위해서 Two-independent t-test를 사용하였으며, 분석에 앞서 자료
들이 정규분포를 나타내지를 검정하였다. 또한, Levene' s test를 통해 변이 동일
한지를 검정하였다.
나. 연구 결과
진우도c 경향은 관찰되지 않았으나, 가덕도 지역과 유사하였으며, 주로 봄에 가
장 높은 광량을 보였고, 비교적 탁도가 높아지는 여름에 낮은 광량을 보였다.
2006
J F M A M J J A S O N D
Underw
ate
r irra
dia
nce
(m
ol photo
ns m
-2 d
-1)
0
10
20
30
40
50
Figure. 4-1. Seasonal variations of underwater irradiance
at Jinwoodo..
2006년 1월말부터 측정된 수온은 잘피가 생존하는데 문제가 없는 것으로 사료
된다(Fig. 4-2). 뚜렷한 계절적인 경향을 보였으며, 최대 수온은 8월에 28℃이였으
며, 최저 수온은 2월에 측정된 6℃이였다.
- 219 -
2006
J F M A M J J A S
Wate
r te
mpera
ture
(oC
)
0
5
10
15
20
25
30
Figure. 4-2. Seasonal variations of water
temperature at Jinwoodo.
2004년 11월 16일과 2005월 12월 16일에 수행된 두 번의 이식 실험은 비교적
성공적이었다(Fig. 4-3). 이식 한 달 후부터 이식된 개체의 안정화 과정에 대한
조사가 시작되었다. 2004년에 이식한 이식 장소는 굴 양식장으로 배가 활발하게
운행되고 있었다. 그 결과, 배의 운행으로 이식된 개체의 대부분이 엽초를 제외한
나머지 부분이 잘라져 나갔으며, 더 이상의 실험이 불가능하였다. 그러나 이식된
개체의 초기 생존율은 3개월 동안 50%가 넘었으며, 이것은 이식 실험의 성공 가
능성을 증명해준다.
Figure. 4-3. Photograph of experimental sites before
and after transplanting
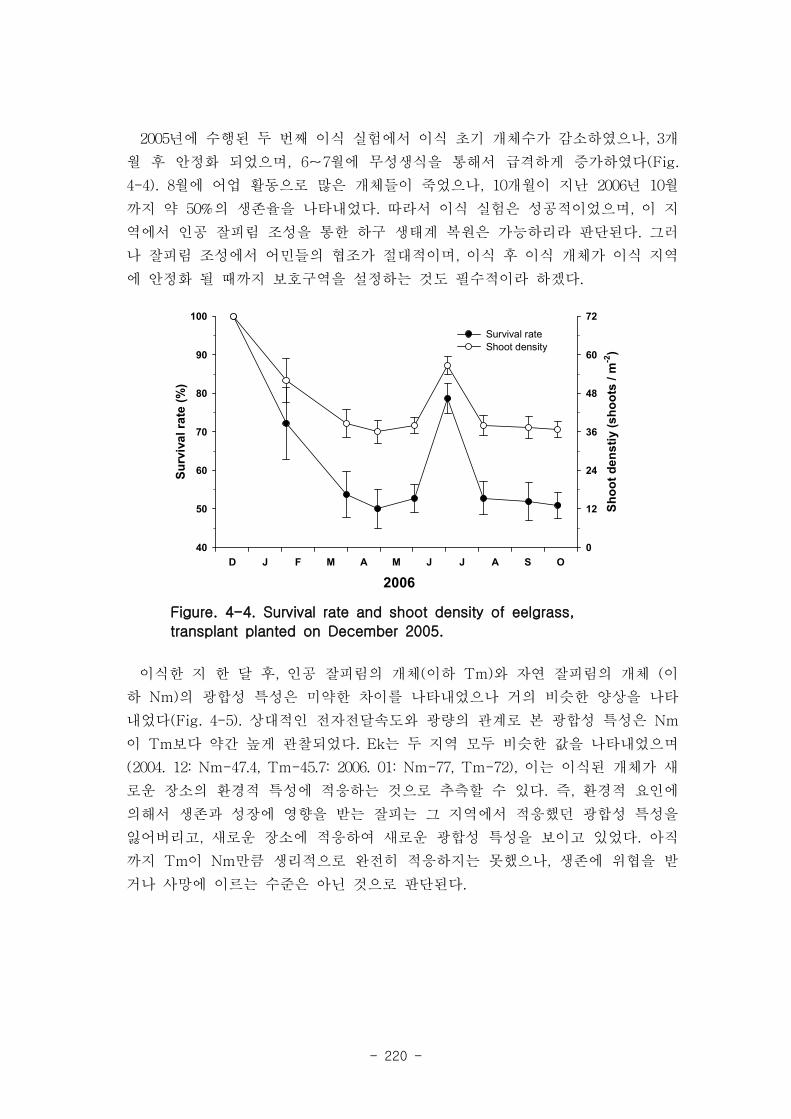
- 220 -
2005년에 수행된 두 번째 이식 실험에서 이식 초기 개체수가 감소하였으나, 3개
월 후 안정화 되었으며, 6~7월에 무성생식을 통해서 급격하게 증가하였다(Fig.
4-4). 8월에 어업 활동으로 많은 개체들이 죽었으나, 10개월이 지난 2006년 10월
까지 약 50%의 생존율을 나타내었다. 따라서 이식 실험은 성공적이었으며, 이 지
역에서 인공 잘피림 조성을 통한 하구 생태계 복원은 가능하리라 판단된다. 그러
나 잘피림 조성에서 어민들의 협조가 절대적이며, 이식 후 이식 개체가 이식 지역
에 안정화 될 때까지 보호구역을 설정하는 것도 필수적이라 하겠다.
2006
D J F M A M J J A S O
Surv
ival ra
te (%
)
40
50
60
70
80
90
100
Shoot denstiy (shoots
/ m
-2)
0
12
24
36
48
60
72
Survival rate
Shoot density
Figure. 4-4. Survival rate and shoot density of eelgrass,
transplant planted on December 2005.
이식한 지 한 달 후, 인공 잘피림의 개체(이하 Tm)와 자연 잘피림의 개체 (이
하 Nm)의 광합성 특성은 미약한 차이를 나타내었으나 거의 비슷한 양상을 나타
내었다(Fig. 4-5). 상대적인 전자전달속도와 광량의 관계로 본 광합성 특성은 Nm
이 Tm보다 약간 높게 관찰되었다. Ek는 두 지역 모두 비슷한 값을 나타내었으며
(2004. 12: Nm-47.4, Tm-45.7: 2006. 01: Nm-77, Tm-72), 이는 이식된 개체가 새
로운 장소의 환경적 특성에 적응하는 것으로 추측할 수 있다. 즉, 환경적 요인에
의해서 생존과 성장에 영향을 받는 잘피는 그 지역에서 적응했던 광합성 특성을
잃어버리고, 새로운 장소에 적응하여 새로운 광합성 특성을 보이고 있었다. 아직
까지 Tm이 Nm만큼 생리적으로 완전히 적응하지는 못했으나, 생존에 위협을 받
거나 사망에 이르는 수준은 아닌 것으로 판단된다.
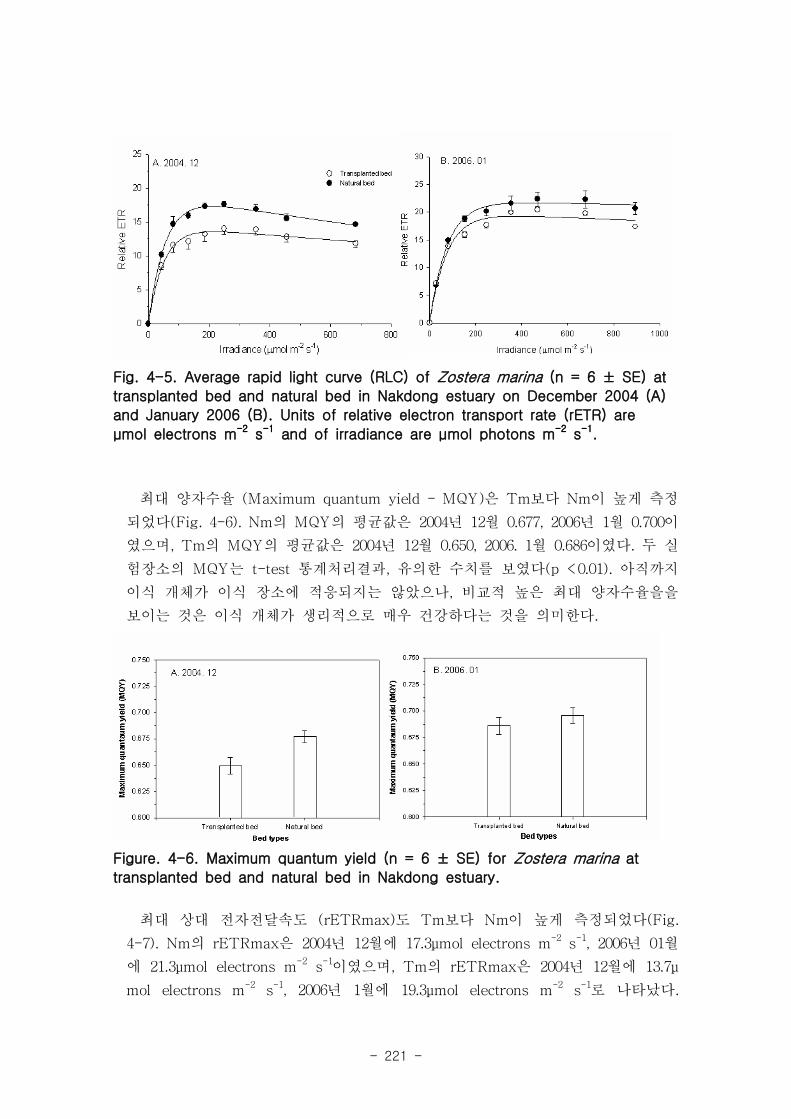
- 221 -
Fig. 4-5. Average rapid light curve (RLC) of Zostera marina (n = 6 ± SE) at
transplanted bed and natural bed in Nakdong estuary on December 2004 (A)
and January 2006 (B). Units of relative electron transport rate (rETR) are
μmol electrons m-2 s-1 and of irradiance are μmol photons m-2 s-1.
최대 양자수율 (Maximum quantum yield - MQY)은 Tm보다 Nm이 높게 측정
되었다(Fig. 4-6). Nm의 MQY의 평균값은 2004년 12월 0.677, 2006년 1월 0.700이
였으며, Tm의 MQY의 평균값은 2004년 12월 0.650, 2006. 1월 0.686이였다. 두 실
험장소의 MQY는 t-test 통계처리결과, 유의한 수치를 보였다(p <0.01). 아직까지
이식 개체가 이식 장소에 적응되지는 않았으나, 비교적 높은 최대 양자수율을을
보이는 것은 이식 개체가 생리적으로 매우 건강하다는 것을 의미한다.
Figure. 4-6. Maximum quantum yield (n = 6 ± SE) for Zostera marina at
transplanted bed and natural bed in Nakdong estuary.
최대 상대 전자전달속도 (rETRmax)도 Tm보다 Nm이 높게 측정되었다(Fig.
4-7). Nm의 rETRmax은 2004년 12월에 17.3μmol electrons m-2 s
-1, 2006년 01월
에 21.3μmol electrons m-2 s
-1이였으며, Tm의 rETRmax은 2004년 12월에 13.7μ
mol electrons m-2 s
-1, 2006년 1월에 19.3μmol electrons m
-2 s-1로 나타났다.
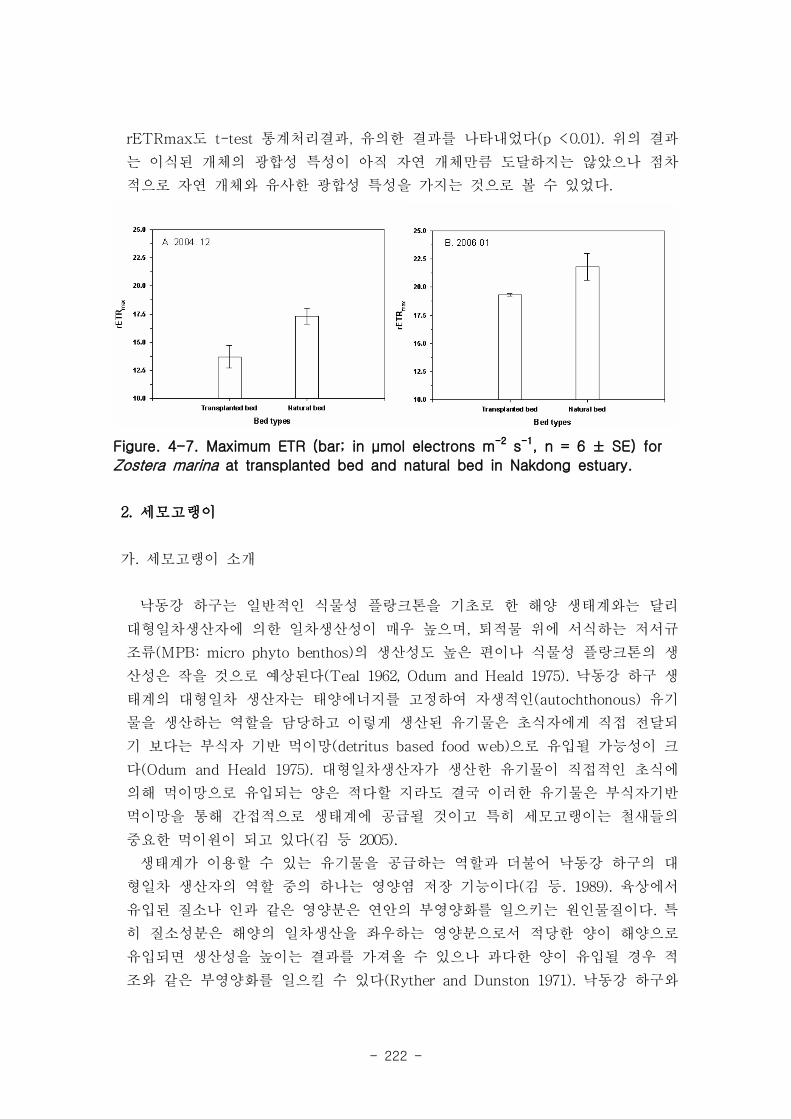
- 222 -
rETRmax도 t-test 통계처리결과, 유의한 결과를 나타내었다(p <0.01). 위의 결과
는 이식된 개체의 광합성 특성이 아직 자연 개체만큼 도달하지는 않았으나 점차
적으로 자연 개체와 유사한 광합성 특성을 가지는 것으로 볼 수 있었다.
Figure. 4-7. Maximum ETR (bar; in μmol electrons m-2 s-1, n = 6 ± SE) for
Zostera marina at transplanted bed and natural bed in Nakdong estuary.
2. 세모고랭이
가. 세모고랭이 소개
낙동강 하구는 일반적인 식물성 플랑크톤을 기초로 한 해양 생태계와는 달리
대형일차생산자에 의한 일차생산성이 매우 높으며, 퇴적물 위에 서식하는 저서규
조류(MPB: micro phyto benthos)의 생산성도 높은 편이나 식물성 플랑크톤의 생
산성은 작을 것으로 예상된다(Teal 1962, Odum and Heald 1975). 낙동강 하구 생
태계의 대형일차 생산자는 태양에너지를 고정하여 자생적인(autochthonous) 유기
물을 생산하는 역할을 담당하고 이렇게 생산된 유기물은 초식자에게 직접 전달되
기 보다는 부식자 기반 먹이망(detritus based food web)으로 유입될 가능성이 크
다(Odum and Heald 1975). 대형일차생산자가 생산한 유기물이 직접적인 초식에
의해 먹이망으로 유입되는 양은 적다할 지라도 결국 이러한 유기물은 부식자기반
먹이망을 통해 간접적으로 생태계에 공급될 것이고 특히 세모고랭이는 철새들의
중요한 먹이원이 되고 있다(김 등 2005).
생태계가 이용할 수 있는 유기물을 공급하는 역할과 더불어 낙동강 하구의 대
형일차 생산자의 역할 중의 하나는 영양염 저장 기능이다(김 등. 1989). 육상에서
유입된 질소나 인과 같은 영양분은 연안의 부영양화를 일으키는 원인물질이다. 특
히 질소성분은 해양의 일차생산을 좌우하는 영양분으로서 적당한 양이 해양으로
유입되면 생산성을 높이는 결과를 가져올 수 있으나 과다한 양이 유입될 경우 적
조와 같은 부영양화를 일으킬 수 있다(Ryther and Dunston 1971). 낙동강 하구와

- 223 -
같은 연안하구에서 유기물 및 질소 영양염의 저장 및 저감 기능에 관한 정량적인
연구는 하구의 자연정화 능력을 평가하는 중요한 기준이 될 것이다(Capone 1988,
Howarth et al. 1988, Seizinger 1990). 그러나 단순히 대형일차생산자 흡수 능력
을 하구 갯벌의 영양염 제거나 자연정화능력으로 평가하는 것은 어렵다(안 2005).
성장 기간 동안 흡수된 영양염은 주변 환경에서 그 양을 감소시키는 역할을 할
수 있으나 완전히 제거된 것은 아니며, 대형일차생산자의 생체가 유기쇄설물로 변
하고 이를 미생물이 분해할 때 다시 환경에 유출될 것이다. 이 경우 대형일차생산
자의 흡수를 “제거”라고 보기는 힘들며, 결국 이러한 흡수가 생태학적으로 어떤
의미를 갖는 가는 영양분의 저장과 변환, 그 시스템으로 부터의 제거 등 종합적
인 물질순환의 측면에서 판단하는 것이 필요하다(안 2005).
낙동강 하구의 주요 일차생산자인 세모고랭이의 성장특성과 생체량 변화를 살
펴보았다. 낙동강 하구 세모고랭이는 10월 이후 지상부가 사라지고, 잎 부분은 퇴
적물에 묻혀 유기쇄설물로 존재하는데, 유기쇄설물의 생체량 변화도 함께 살펴봄
으로써, 미생물에 의한 분해활동 및 세모고랭이에 저장되었던 영양염의 방출량을
추정하였으며, 생체량 측정에서 건중량과 함께 강열감량, 탄소 및 질소함량을 조
사하여, 에너지 및, 영양염 축적과 방출량의 정량화하려 하였다.
나. 조사방법
(1) 조사지역 및 현장조사
조사지역은 낙동강 하구, 명지주거단지 앞에 위치한 세모고랭이 군락이다(Fig.
1). 이 지역은 을숙도 지역과 함께 낙동강 하구에서 세모고랭이가 대규모로 서식
하는 지역으로서 을숙도 보다는 약간 낮으나, 여름철에는 평방미터당 200 개체에
이르는 높은 밀도가 보고되었다(김 등 2005). 명지지역 인근에는 수로가 발달해
있어 수로 부근으로 경사가 급한 갯벌이 발달해 있으며, 비교적 짧은 거리에서 다
양한 조고(tidal height)를 가진 지형을 가지고 있다. 명지지역의 세모고랭이 군락
은 평균해수면 위 190 cm 정도에 위치하고 있으며, 평균입도 2.6~3.9 phi 의 니
사질 퇴적물로 이루어져 있다(환경부 2005).
현장 조사는 2005~6년에 걸쳐 총 10회가 실시되었는데, 2005년에는 4월, 7월, 8
월, 10월, 11월, 2006년에는 3월, 4월, 8월, 9월, 10월에 이루어 졌으며, 주로 대조
시에 현장 조사가 실시되었다. 세모고랭이 군락이 대기 중에 노출 되었을 때 현장
을 방문하여, 무작위로 30cm x 30cm 넓이의 방형구를 설치한 후 삽과 손등을 이
용하여, 방형구내의 세모고랭이의 지상부와 지하부(뿌리 및 괴경)를 채취하였다.
지하부 채취를 위해서는 방형구에 해당하는 퇴적물 내 60cm 깊이까지 파내어, 손
으로 뿌리와 괴경 등을 골라내었다. 한 조사에서 반복 시료수는 2~3번 이었다.
세모고랭이 시료는 플라스틱 백에 보관하였으며, 채취 후 4 시간 내에 실험실로

- 224 -
운반하여 측정이 이루어 졌다.
(2) 실험실 측정
실험실로 가져온 세모고랭이는 증류수로 세척하여 퇴적물과 이물질을 제거하
였다. 수건을 이용하여 표면의 수분을 제거하고 상온에서 30분간 방치한 후 줄기,
뿌리, 괴경으로 구분하여 습중량을 측정하였다. 건중량 측정을 위하여, 세모고랭의
의 각 부위는 60도 온도의 건조기에서 하루 동안 건조 되었다. 각 조사 시기 마다
살아있는 세모고랭이 개체와 죽어서 부패 과정에 있는(유기쇄설물) 세모고랭이를
구분 하였다. 생체내 유기물 측정에는 강열감량(IL: ignition loss)법과 원소분석기
(EA; elecmental analyzer)가 이용되었다. 강열감량은, 약 5g의 건조 시료를 도가
니에 넣고 550도로 맞추어진 전기로에서 2시간 동안 연소 시킨 후 연소 전후의
무게차를 본래 건중량의 무게비로서 나타내었다. 원소분석기 분석에서도 건조 시
료가 이용되었으며 분쇄된 시료를 EA 분석기(Elemental Analysen systeme,
Vario EL)로 분석하였으며 C, N의 함량을 건중량에 대한 무게비로 나타내었다.
다. 결과
(1) 세모고랭이 성장 특성
세모고랭이는 다년해살이풀로서 종자를 생산하나 종자에 의한 번식은 낮은 편
이고 주로 괴경을 통해 번식 한다(윤 1991, 김 등 2005). 세모고랭이의 계절변화를
살펴보면 겨울에 퇴적물에 묻혀있던 괴경 들로부터 4~5월경에 싹 올라온다. 어느
정도 성장하면 땅속줄기를 사방으로 뻗어 다시 새로운 싹이 땅위로 올라온다(김
등 2005). 8월경에는 개화가 시작되고 이때 괴경도 형성되는데, 본격적인 개화는
가을에 일어나고 식물체가 시들기 시작할 때 땅속에서는 괴경 형성이 활발해진
다(김 등 2005). 10월경이 되면 세모고랭이의 지상부가 갑자기 사라지게 되는데,
수금류가 날아와 괴경을 섭식하고 이때 세모고랭이의 지상부를 잘라내기 때문이
다(김 등 2005). 수금류의 섭식을 피한 괴경은 겨울동안 땅속에 뭍혀 있다가 이듬
해 봄에 다시 싹을 틔운다.
본 조사에서는 낙동강 하구 명지 지역에서 4월 싹이 올라온 이후 세모고랭이
의 성장특성을 알아보기 위하여, 세모고랭이의 지상부와 지하부 길이 변화를 살펴
보았다(Fig. 4-8(A)). 4월에 새모고랭이 줄기의 길이는 6cm 내외였으나 7월이 되
면 56cm 까지 자라며, 8월경에는 63.5cm에 이르게 된다. 4월에서 7월까지 빠른
성장을 보이고 7월 이후 포화되어 성장률이 낮았다. 뿌리의 길이도 유사한 성장특
성을 보여 4월의 평균 2.5cm 에서 8월에는 9.1cm로 증가 하였다. 본 조사에서 8

- 225 -
월의 지상부 길이는 김 등(2005)의 결과(30~60 cm)와 유사하였으나 윤(1991)의
결과(26~38 cm) 보다는 큰 편 이었다. 지상부의 넓이는 4월에 0.21로 가장 작았
고 8월에 0.66 cm 로 최대값을 보였다(Fig. 4-8(B)). 한 개체 당 순의 수(number
of shoot)도 4월에 2개에서 8월에는 3개로서 8월에 최댓값을 보이고 더 이상 증가
하지 않았다(Fig. 4-8(B)).
계절에 따른 생체량의 변화도 길이 및 폭의 변화와 유사하여 4월 이후 8월에
최댓값을 보였다(Fig. 4-9). 개체 당 습중량은 2005년 4월에 0.1 g에서 2006년 8월
에 6.5g 으로 증가 하였다(Fig. 4-9(A)). 2005년 8월에는 7월에 비해 감소하였는
데, 이는 계절에 따른 수분함량의 차이로 생각되며, 건중량은 거의 차이를 보이지
않았다(Fig. 4-9(A),(B)). 건중량은 2005년 4월에 0.03g으로 최소값을 보였고,
2006년 8월에 1.1g 으로 최대값을 나타냈다. 2006년 4월에는 2005년 4월에 비해
습중량과 건중량에서 많은 차이를 보였는데, 2006년 4월에는 2005년 4월에 비하여
습중량은 약 16배, 건중량은 약 20배 가까이 높은 값을 보였다. 이러한 차이는 뿌
리 부분의 괴경 중량의 차이에 의한 것으로 여겨진다.(Fig. 4-9(C)). 2006년 4월에
괴경 건중량은 0.2g 으로 2005년 4월의 0.04 g에 비해 5배 이상이었는데 이러한
차이는 괴경으로부터 새싹(turion)이 나와 얼 만큼 성장했는지에 따른 차이로 때
문이었다. 즉 2005년 4월에는 싹이 나온 후 상당기간 성장하여, 괴경이 작아 졌
으며, 2006년 4월에는 새싹이 나온 후 얼마 지나지 않아 괴경 중량이 유지된 것으
로 생각된다. 2005년 4월의 줄기부분의 평균 길이는 약 6cm정도였고 2006년도에
는 약 1cm이었다(Fig. 4-8(A)). 윤(1991)의 조사에서도 3월 새싹이 나오기 전에는
괴경 중량이 전체중량의 55.8 %를 차지하였으나 싹이 튼 이후에는 지상부 중량이
51.8 %로 더 커졌음을 보고하고 있다.
A. Individual shoot and root length
0
20
40
60
80
Apr-05 Jul-05 Aug-05 Aug-06 Sep-06
Length
(cm
))
shoot
root
B. Width and numver of shoot
0.0
0.2
0.4
0.6
0.8
Apr-05 Jul-05 Aug-05 Aug-06 Sep-06
Width (cm)
0
1
2
3
4
# of shoot
Width
# of shoot
Figure. 4-8. 계절에 따른 세모고랭이 개체의 지상부와 지하부의 길이(A)와 폭 및
너비(B)의 변화
- 226 -
A. Wet weight (total)
0
1
2
3
4
5
6
7
8
Apr-05 Jul-05 Aug-05 Apr-06 Aug-06 Sep-06
g/indv.
shoot
root
C. Tuber dry weight
0.0
0.1
0.2
0.3
Apr-05 Aug-05 Apr-06 Oct-06
g/indv.
B. Dry weight
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Apr-05 Jul-05 Aug-05 Apr-06 Aug-06 Sep-06
g/indv
shoot
root
Figure. 4-9. 계절에 따른 세모고랭이 한
개체의 습중량(A), 건중량(B) 변화.
계절에따른 괴경의 무게변화(C)

- 227 -
(2) 단위 면적 당 세모고랭이 생체 및 고사체 양 변화
단위 면적당 세모고랭이의 생체량 변화에 영향을 끼치는 요인은 한 개체의 성
장특성과 세모고랭이의 밀도 변화 등 일 것이다. 단위면적당 생체량은 8월에 가장
높은 값을 보였는데, 개체의 성장특성과 유사하여 계절에 따른 밀도 변화 보다는
개체의 성장이 단위면적당 생체량을 좌우함을 알 수 있었다(Fig. 4-10). 2005년 4
월에 습중량과 건중량은 각각 5.7g m-2
, 1.3g m-2 으로 가장 작았고, 생체량이 최
대인 2006년 8월에는 각각 1084g m-2
, 171g m-2 에 이르렀다. 10월달에 철새에
활동에 의해서 세모고랭이의 지상부가 완전히 사라진 후에도 가을과 겨울동안 상
당한 양의 세모고랭이 유기쇄설물이 퇴적물내에 존재하였다. 고사체 무게는 2006
년 10월에 습중량과 건중량에서 각각 1015g m-2
, 145g m-2 로 상당량이 퇴적물속
에 묻혀 있었다. 2005년 10, 11월에 습중량은 8월보다 유사하거나 오히려 증가하
는 경향을 보였다(Fig. 4-10(A)). 이는 지상부가 퇴적물에 묻힌 상태에서 수분 함
량이 높아진데서 기인한 것으로서 건중량의 경우에는 2005년 10월에 83g m-2 ,
11월에 94g m-2 으로서 8월달의 186g m
-2 에 비하여 약 50%에 해당하는 양을 나
타내었다. 2005년 10월에서 2006년 3월까지의 건중량 변화를 보면, 2005년 10월과
11월에는 거의 변화가 없다가 2006년 3월에 37g m-2 으로 반이상 감소하였음을
알 수 있었다. 퇴적물에 묻힌 세모고랭이의 지상부는 미생물에 의한 분해, 저서동
물에 속하는 초식자 및 잡식자에 의한 섭식 등에 의해 그 양이 감소할 수 있다
(김 2005, Odum and Heald 1975). Kang et al.(submitted)이 탄소 및 질소 동위원
소 분석을 통해 밝힌 바에 따르면, 저서동물이 직접 세모고랭이를 섭식하지는 않
는 것 같으며, 이 지역에 서식하는 저서동물의 주요 먹이는 저서성 규조류(MPB;
micro phyto benthos)였다. 따라서 퇴적물로 유입된 세모고랭이의 생체는 미생물
에 의한 분해 과정에서 재광물화(remineralization) 되어 그 양이 감소할 것으로
여겨진다(Rowe et al. 1975, Howes et al. 1984,). 재광물화 과정에서 생성된 무기
질소나 인등의 영양성분은 저서성 규조류의 일차생산에 사용되어 간접적으로 저
서생물의 먹이양을 증가시키는 역할을 담당하였을 것이다(안 2005, Pickeny et al.
1997). 2005년 11월에서 2006년 3월 사이에 지상부의 건중량은 73g m-2 에서 21g
m-2 으로 70% 정도 감소한 것에 비하여, 뿌리와 괴경을 포함하는 지하부의 건중
량은 20g m-2 에서 15g m
-2 으로 28% 감소하는데 그쳐 괴경을 포함한 뿌리부분
은 퇴적물에 묻혀있는 가을 겨울 동안에도 부패하지 않고 살아있음을 알 수 있었
다(Fig. 4-10(B)).
건조 중량과 더불어 강열감량(IL: Ignition loss)과 원소분석에 의한 생체량을
Table. 4-1 과 Fig. 4-11, 12에 나타내었다. 낙동강 명지지역 세모고랭이의 지상부
와 지하부를 합한 전체의 강열감량은 연평균 82.9g m-2이었다(Table. 4-1). 2006
년 4월에 2g m-2 로 가장 작았고 2006년 8월에 154g m-2 으로 가장 높게 나타났
다. 계절변화가 심하였으나 전체 IL 중 약 67%가 지상부에 존재하였으며 나머지

- 228 -
33%가 지하부에 존재하였다. 2006년 8월에 IL은 154g m-2 으로서 건중량과 거의
유사한 값을 보였으나, 2005년 10월과 11월에는 IL과 건중량이 차이를 보였다. IL
은 건중량의 60~80%에 달해, 연소되지 않는 부분이 매우 커졌다. 한 가지 원인
으로 생각해 볼 수 있는 것은 퇴적물에서 세모고랭이를 분리가 완벽하지 못했다
는 것이다. 퇴적물 속에서 부패 상태에 있는 세모고랭이를 최대한 세척하여 건조
한 후 건중량을 측정하였으나, 일부 퇴적물이 세모고랭이와 결합되어 분리되지 않
았을 가능성이 있다. 이러한 경향이 반영되어 IL에서는 2005년 8월 세모고랭이가
생존했을 때의 유기물 값이 2005년 10월 이후 퇴적물에 묻혔을 때에 비해 약 3배
에 달했다. 이는 건중량에서 같은 시기에 2배 정도의 차이가 있었던 것을 고려하
면, 새모고랭이에서 생체와 사체의 유기물(유기쇄설물) 양을 비교하는 데는 건중
량 보다는 IL이 효과적임을 나타낸다. 원소분석기를 이용하여 구한 평방미터당 세
모고랭이의 탄소량과 질소량도 건중량과 비슷한 경향을 보였다. 각각 33.8과 1.1g
m-2 이었다(Table. 4-1).
2005년 4월과 8월동안 건중량의 변화를 바탕으로 세모고랭이의 순생산성을 추
정하여 보면, 538g DW m-2 yr-1 였다. 이는 세모고랭이 지상부의 탈락이나 초식
동물에 의한 소비를 고려치 않은 것으로서 순생산성의 최소값에 해당할 것이다
(윤 1991). 이 값은 일반적인 염습지 대형일차생산자의 생산성이 0.1~1Kg DW
m-2 yr-1 에 포함되고 있으나 약간 낮은 편이다(Mann 1982, 윤 1991). 또한 조사
장소나 시기가 달라 직접적인 비교는 힘드나, 윤(1991)이 1984년도에서 1989년까
지 낙동강 하구의 세모고랭이를 대상으로 측정한 값(80~150g DW m-2 yr-1
) 보
다는 4~5배 높은 값이다. 원소 분석을 기초로 2005년 4월과 8월동안 순생산성을
추정하여보면 탄소량으로는 240g-C m-2 yr-1
1(54mmole-C m-2 d
-1), 질소량으로는
8.2g-N m-2 yr
-1 에 해당한다. 즉 세모고랭이에 의해 4~8월동안 평방미터당 약
2.7g 의 질소(약 200mmole에 해당)가 새모고랭의의 생체로 유입되었다. 일간 변
화로 환산하면 1.6mmole-N m-2 d
-1 에 해당하는 양이다. 2005년 11월에서 2006
년 3월 동안의 세모고랭이 쇄설물의 탄소 및 질소 함량 변화를 기초로 세모고랭
이의 생체가 분해되는 속도를 추정하여 보았다(Fig. 6). 이 기간 동안 탄소 분해
율은 198mg-C m-2․d-1(16mmole-C m
-2 d-1
) 이고 질소 분해율은 2.8mg-N m-2
d-1
(0.23mmole-N m-2 d
-1)이었다. 본 조사에서 추정된 세모고랭이의 탄소분해율은
강화도 갯벌에서 측정된 퇴적물 산소요구량(SOD; sediment oxygen demand) 유
사한 값 이었다(안 2005). 본 조사결과는 전체 퇴적물을 대상으로한 유기물 분해
율(SOD)이 아니고 단지 세모고랭이의 탄소함량 감소만을 추정한 것으로서 본 조
사지역의 SOD는 이 결과보다는 클 것으로 예상된다.
- 229 -
A. Monthly biomass change (Wet weight )
0
200
400
600
800
1000
1200
1400
1600
Apr-05 Jul-05 Aug-05 Oct-05 Nov-05 Mar-06 Apr-06 Aug-06 Sep-06 Oct-06
g/㎡
B. Monthly biomass change (Dry weight )
0
50
100
150
200
250
Apr-05 Jul-05 Aug-05 Oct-05 Nov-05 Mar-06 Apr-06 Aug-06 Sep-06 Oct-06
g/㎡
shoot
root
Figure. 4-10. 계절에 따른 단위면적당 세모고랭이
생체량. 습중량(A)과 건중량(B).
B. Ignition loss (IL) to dry weight ratio
0
50
100
Aug-05 Oct-05 Nov-05 Mar-06 Apr-06 Aug-06 Sep-06 Oct-06
%
A. Ignition loss (IL)
0
40
80
120
160
200
240
Aug-05 Oct-05 Nov-05 Mar-06 Apr-06 Aug-06 Sep-06 Oct-06
g/㎡
shoot
root
Figure. 4-11. 계절에 따른 Ignition loss(A)와 Ignition loss / 건중량(B)
B. Nitogrn biomass
0
0.5
1
1.5
2
2.5
3
Aug-05 Oct-05 Nov-05 Mar-06 Aug-06 Sep-06 Oct-06
Bio
mass(g
/㎡)
Shoot
Root
A. Carbon biomass
0
20
40
60
80
100
Aug-05 Oct-05 Nov-05 Mar-06 Aug-06 Sep-06 Oct-06
Bio
mass(g
/㎡)
Shoot
Root
Figure. 4-12. 계절에 따른 단위면적당 세모고랭이의 탄소량(A)과 질소량(B)
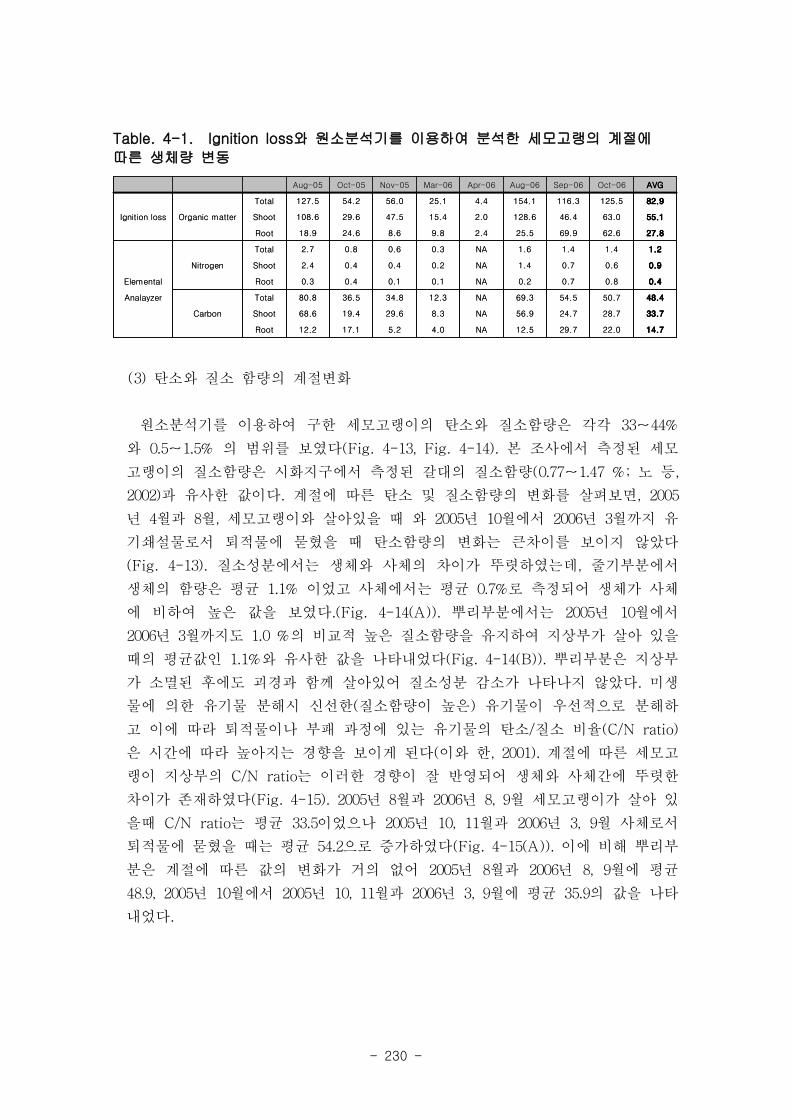
- 230 -
14.7 22.0 29.7 12.5 NA4.0 5.2 17.1 12.2 Root
33.7 28.7 24.7 56.9 NA8.3 29.6 19.4 68.6 ShootCarbon
48.4 50.7 54.5 69.3 NA12.3 34.8 36.5 80.8 TotalAnalayzer
0.4 0.8 0.7 0.2 NA0.1 0.1 0.4 0.3 RootElemental
0.9 0.6 0.7 1.4 NA0.2 0.4 0.4 2.4 ShootNitrogen
1.2 1.4 1.4 1.6 NA0.3 0.6 0.8 2.7 Total
27.8 62.6 69.9 25.5 2.4 9.8 8.6 24.6 18.9 Root
55.1 63.0 46.4 128.6 2.0 15.4 47.5 29.6 108.6 ShootOrganic matterIgnition loss
82.9 125.5 116.3 154.1 4.4 25.1 56.0 54.2 127.5 Total
AVGOct-06Sep-06Aug-06Apr-06Mar-06Nov-05Oct-05Aug-05
14.7 22.0 29.7 12.5 NA4.0 5.2 17.1 12.2 Root
33.7 28.7 24.7 56.9 NA8.3 29.6 19.4 68.6 ShootCarbon
48.4 50.7 54.5 69.3 NA12.3 34.8 36.5 80.8 TotalAnalayzer
0.4 0.8 0.7 0.2 NA0.1 0.1 0.4 0.3 RootElemental
0.9 0.6 0.7 1.4 NA0.2 0.4 0.4 2.4 ShootNitrogen
1.2 1.4 1.4 1.6 NA0.3 0.6 0.8 2.7 Total
27.8 62.6 69.9 25.5 2.4 9.8 8.6 24.6 18.9 Root
55.1 63.0 46.4 128.6 2.0 15.4 47.5 29.6 108.6 ShootOrganic matterIgnition loss
82.9 125.5 116.3 154.1 4.4 25.1 56.0 54.2 127.5 Total
AVGOct-06Sep-06Aug-06Apr-06Mar-06Nov-05Oct-05Aug-05
Table. 4-1. Ignition loss와 원소분석기를 이용하여 분석한 세모고랭의 계절에
따른 생체량 변동
(3) 탄소와 질소 함량의 계절변화
원소분석기를 이용하여 구한 세모고랭이의 탄소와 질소함량은 각각 33~44%
와 0.5~1.5% 의 범위를 보였다(Fig. 4-13, Fig. 4-14). 본 조사에서 측정된 세모
고랭이의 질소함량은 시화지구에서 측정된 갈대의 질소함량(0.77~1.47 %; 노 등,
2002)과 유사한 값이다. 계절에 따른 탄소 및 질소함량의 변화를 살펴보면, 2005
년 4월과 8월, 세모고랭이와 살아있을 때 와 2005년 10월에서 2006년 3월까지 유
기쇄설물로서 퇴적물에 묻혔을 때 탄소함량의 변화는 큰차이를 보이지 않았다
(Fig. 4-13). 질소성분에서는 생체와 사체의 차이가 뚜렷하였는데, 줄기부분에서
생체의 함량은 평균 1.1% 이었고 사체에서는 평균 0.7%로 측정되어 생체가 사체
에 비하여 높은 값을 보였다.(Fig. 4-14(A)). 뿌리부분에서는 2005년 10월에서
2006년 3월까지도 1.0 %의 비교적 높은 질소함량을 유지하여 지상부가 살아 있을
때의 평균값인 1.1%와 유사한 값을 나타내었다(Fig. 4-14(B)). 뿌리부분은 지상부
가 소멸된 후에도 괴경과 함께 살아있어 질소성분 감소가 나타나지 않았다. 미생
물에 의한 유기물 분해시 신선한(질소함량이 높은) 유기물이 우선적으로 분해하
고 이에 따라 퇴적물이나 부패 과정에 있는 유기물의 탄소/질소 비율(C/N ratio)
은 시간에 따라 높아지는 경향을 보이게 된다(이와 한, 2001). 계절에 따른 세모고
랭이 지상부의 C/N ratio는 이러한 경향이 잘 반영되어 생체와 사체간에 뚜렷한
차이가 존재하였다(Fig. 4-15). 2005년 8월과 2006년 8, 9월 세모고랭이가 살아 있
을때 C/N ratio는 평균 33.5이었으나 2005년 10, 11월과 2006년 3, 9월 사체로서
퇴적물에 묻혔을 때는 평균 54.2으로 증가하였다(Fig. 4-15(A)). 이에 비해 뿌리부
분은 계절에 따른 값의 변화가 거의 없어 2005년 8월과 2006년 8, 9월에 평균
48.9, 2005년 10월에서 2005년 10, 11월과 2006년 3, 9월에 평균 35.9의 값을 나타
내었다.
- 231 -
A. Carbon content in shoot (%)
0
10
20
30
40
50
Aug-05 Oct-05 Nov-05 Mar-06 Aug-06 Sep-06 Oct-06
%
B. Carbon content in root (%)
0
10
20
30
40
50
Aug-05 Oct-05 Nov-05 Mar-06 Aug-06 Sep-06 Oct-06
%
Figure. 4-13. 계절에 따른 세모고랭이의 유기 탄소 함량 변화
A. Nitrogen content in shoot (%)
0
0.3
0.6
0.9
1.2
1.5
1.8
Aug-05 Oct-05 Nov-05 Mar-06 Aug-06 Sep-06 Oct-06
%
B. Nitrogen content in root (%)
0
0.3
0.6
0.9
1.2
1.5
Aug-05 Oct-05 Nov-05 Mar-06 Aug-06 Sep-06 Oct-06
%
Figure. 4-14. 계절에 따른 세모고랭이의 유기 질소 함량 변화
A. C/N ratio in shoot
0
20
40
60
80
Aug-05 Oct-05 Nov-05 Mar-06 Aug-06 Sep-06 Oct-06
C/N
B. C/N ratio in root
0
20
40
60
80
Aug-05 Oct-05 Nov-05 Mar-06 Aug-06 Sep-06 Oct-06
C/N
Figure. 4-15. 계절에 따른 세모고랭이의 C/N ratio 변화
라. 세모고랭이의 성장특성 요약
낙동강 하구 갯벌에 명지지역에 서식하는 세모고랭이의 성장특성을 살펴보면 4
월에 싹이 튼 후 7월까지 빠르게 성장하며, 7~8월에 최대성장을 이루었다. 최대
성장시 지상부 길이는 63.5cm, 폭은 0.66cm, 뿌리길이는 9.1cm였다. 한 개체의 생
체량도 길이와 유사한 증가 양상을 보였는데 최대 성장시 습중량은 6.5g ind-1
, 건
중량은 1.1g DW ind-1 였다. 2005년 4월과 2006년 4월 사이에 줄기부분의 건중량
은 큰 차이를 보였는데 이는 괴경의 중량차이에서 기인하였다. 괴경의 중량차이는
새싹이 나와 얼마만큼 성장하였는가에 따라 달라졌는데, 2005년 4월에는 싹이 나
온 후 상당기간 성장하여, 괴경이 작아 졌으며, 2006년 4월에는 새싹이 나온 후
얼마 지나지 않아 괴경 중량이 유지되었다.
명지지역의 단위면적당 세모고랭이의 생체량은 8월에 최대값을 보였다(건중량
= 171g DW m-2
). 10월 이후 세모고랭이의 지상부가 퇴적물 속 유기쇄설물로 바
뀐 이후에도 상당한 양의 생체량이 유지 되었는데, 2005년 10월에 83g DW m-2 ,

- 232 -
11월에 94g DW m-2 으로 최대값의 50 %에 달했다. 겨울동안 유기쇄설물은 미생
물 분해에 의하여 그 양이 계속 감소하였으며, 2006년 3월에 생체량은 36g DW
m-2 이 되었다. 퇴적물에 묻힌 지상부 유기쇄설물의 양이 미생물 분해에 의하여
급격히 감소함에 비하여, 뿌리와 괴경을 포함한 지하부는 20g m-2 에서 15g
m-2 으로 28% 감소하는데 그쳐 괴경을 포함한 뿌리부분은 퇴적물에 묻혀있는 가
을 겨울 동안에도 부패하지 않고 살아있음을 알 수 있었다.
2005년 4월과 8월동안 건중량의 변화를 바탕으로 세모고랭이의 순생산성을 추
정하여 보면, 538g DW m-2 yr
-1 였다. 원소 분석을 기초로 2005년 4월과 8월동안
순생산성을 추정하여보면 탄소량으로는 240g-C m-2 yr
-1(54 mmole-C m
-2 d-1
),
질소량으로는 8.2g-N m-2 yr
-1 에 해당한다. 즉 세모고랭이에 의해 4~8월동안 평
방미터당 약 2.7g 의 질소(약 200mmole에 해당)의 질소가 새모고랭이의 생체로
유입되었다. 2005년 11월에서 2006년 3월 동안의 세모고랭이 쇄설물의 탄소 및 질
소 함량 변화를 기초로 세모고랭이의 생체가 분해되는 속도를 추정하면 탄소 분
해 율은 198mg-C m-2 d
-1(16mmole-C m
-2 d-1
)이고 질소 분해율은 2.8mg-N m-2
d-1
(0.23mmole-N m-2 d
-1)이었다.
세모고랭이가 살아있을 때 와 퇴적물에서 유기쇄설물 형태로 존재할 때 탄소
함량을 비교하면, 줄기 부분에서는 살아있을 때 높은 값을 보인반면, 뿌리에서는
차이를 보이지 않았다. 이러한 경향은 질소함량에서 더 뚜렷하게 나타나서, 살아
있을 때 지상부의 질소 함량은 유기쇄설물의 질소함량에 비해 2배에 달했다. 뿌리
부분에서는 지상부가 사라진 겨울동안에도 질소함량의 변화는 나타나지 않았다.
탄소와 질소의 비율(C/N ratio)을 살펴보면 유기쇄설물에서 생체에 비해 C/N
ratio가 높아, 미생물 분해시 질소성분이 우선적으로 사용됨을 알 수 있었다.
마. 세모고랭이밭의 퇴적물 특성 변화
세모고랭이를 복원하기 위해서는 무엇보다도 식물에 대한 생태학적 자료의 확
보가 시급하며, 특히 세모고랭이 생육지의 환경적 요인에 대한 정보가 매우 중요
하다. 본 연구에서는 세모고랭이 생육지의 퇴적물 공극수 내 암모니아 농도와 유
기물 농도에 대해서 조사하였다. 조사 방법은 앞에서 언급한 것과 동일하다. 낙동
강 지역에서 세모고랭이의 대표적 서식지인 을숙도와 맹금머리도는 생육 환경에
있어서 약간의 차이를 나타내었다. 우선, 퇴적물 공극수 내 암모니아는 유사한 경
향을 나타내었으나, 사니질로 이루어진 을숙도 지역이 더 높은 것으로 나타났다
(Fig. 4-16) 생육 밀도가 가장 높은 여름에 가장 낮은 암모니아 농도를 나타내었
으며, 엽체가 시들어 떨어지는 겨울에 가장 높은 암모니아 농도를 보였다. 을숙도
지역의 암모니아 농도는 55~214μM이였으며, 맹금머리도 지역은 13~158μM이였
다.
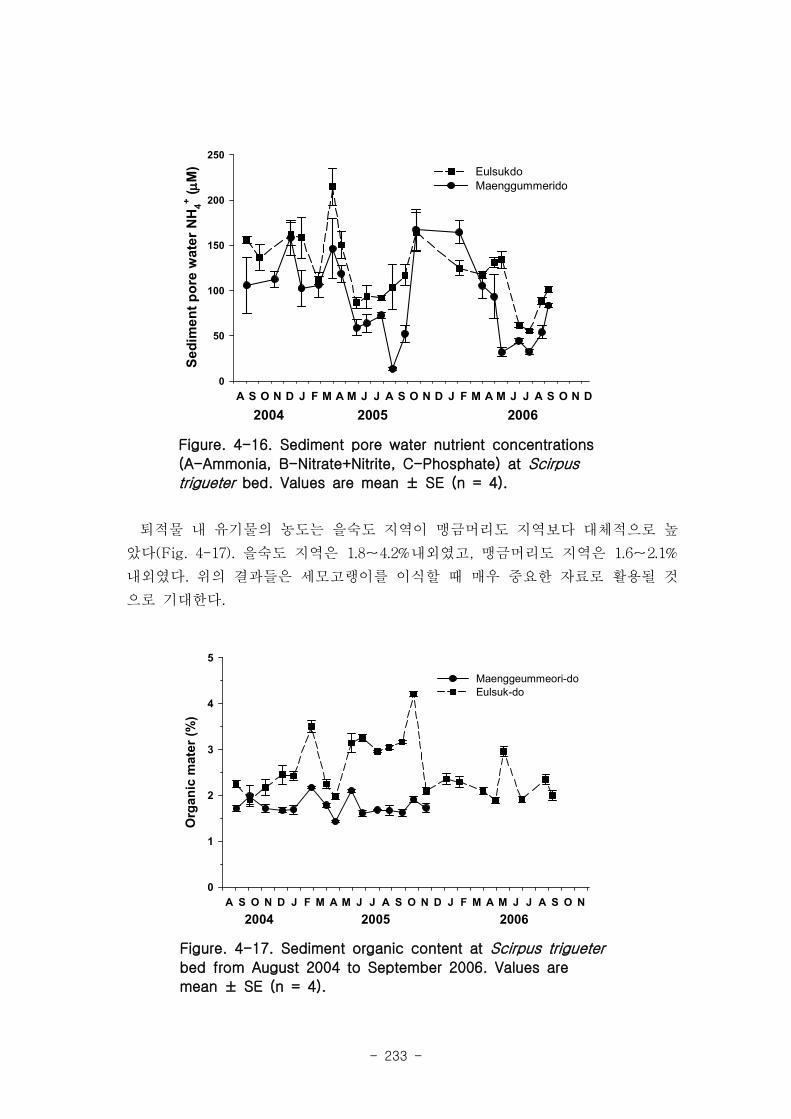
- 233 -
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O N D
Sedim
ent pore
wate
r N
H4
+ (µM
)
0
50
100
150
200
250
Eulsukdo
Maenggummerido
Figure. 4-16. Sediment pore water nutrient concentrations
(A-Ammonia, B-Nitrate+Nitrite, C-Phosphate) at Scirpus trigueter bed. Values are mean ± SE (n = 4).
퇴적물 내 유기물의 농도는 을숙도 지역이 맹금머리도 지역보다 대체적으로 높
았다(Fig. 4-17). 을숙도 지역은 1.8~4.2%내외였고, 맹금머리도 지역은 1.6~2.1%
내외였다. 위의 결과들은 세모고랭이를 이식할 때 매우 중요한 자료로 활용될 것
으로 기대한다.
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O N
Org
anic
mate
r (%
)
0
1
2
3
4
5
Maenggeummeori-do
Eulsuk-do
Figure. 4-17. Sediment organic content at Scirpus trigueter bed from August 2004 to September 2006. Values are
mean ± SE (n = 4).

- 234 -
2006년8월 2006년9월
Mud
Mixed
sand
Mud Mixed Sand
2006년8월 2006년9월
Mud
Mixed
sand
Mud Mixed Sand
바. 세모고랭이 복원 조건
앞에서 살펴 본 바와 같이 낙동강에서 세모고랭이는 4월에서 10월동안 최대성
장을 보여, 낙동강을 통해 유입되는 영양염 이나 유기물을 일시적으로 저장하거나
그 형태를 바꾸어, 하구의 기능 중 자정능력에 핵심적인 역할을 할 뿐만 아니라
, 전반적인 하구의 생태기능에 중요한 역할을 담당하고 있다. 이러한 특징은 낙동
강이나 기타 하구 지역에서 세모고랭이를 복원이 중요함을 시사한다. 본 연구에서
설치한 실험용 갯벌중 06년 갯벌에서 세모고랭이가 자라났는데 이를 토대로 세모
고랭이의 복원 조건을 문헌조사와의 비교를 통해 알아보았다. 세모고랭이의 생존
에 중요한 환경요인으로는 염분, 퇴적물 입도, 노출시간이 있다 (Kim 2006). 생존
가능한 염분범위는 5~15 ppt 정도이며, 하루에 2번 침수와 노출이 반복되는 조간
대 지역에서 주로 나타난다 (Kim 2006). 본 연구에서는 2006년 8월 중갯벌의
mixed 퇴적물에서 111개체/m2으로 가장 높은 세모고랭이 밀도가 나타났으며
sand 퇴적물에서, 26개체/m2로 낮았고, mud 퇴적물에서는 전혀 나타나지 않았다.
중갯벌의 조고는 평균 해수면 위 142.9 cm 에 위치하고 있는데, 이 위치에서는
6~12시간 연속노출이 전체 노출에 50%정도를 차지하며, 48시간 이상의 연속노출
이 25%, 그리고 12~24시간 연속노출이 15%정도 나타나는 지역이다 (1절 조고
및 노출특성 참조). mud 퇴적물에서 세모고랭이가 전혀 나타나지 않은 것은 조고
와 더불어 퇴적물 타입이 세모고랭이의 생존에 중요함을 나타낸다. 세모고랭이가
가장 높게 나타난 mixed 퇴적물은 니질 비율이 24~33 %정도 였으며, 전혀 나타
나지 않은 mud 퇴적물의 경우 니질 비율이 57~61 %였다 (Table. 4-2). 낙동강
에서 세모고랭이는 적당한 염분조건과 조고에 이식한다 할지라도 니질함량이
50% 이상이면 생존하기 힘들 것임을 시사한다. 반대로 니질 함량이 너무 낮아도
최적 생존조건이 아님을 시사 하는데, 본 실험에서 sand 퇴적물의 경우 니질함량
은 4~12%정도 였다. 본 실험의 결과는 Kim (2006)과 유사한 경향을 나타내나,
최적 니질 함량에서 차이를 보이는데, Kim (2006)의 실험에서는 니질함량이 60%
에서 가장 높은 서식밀도를 보였다.
Figure. 4-18. 2006년 8~9월동안 중갯벌 세모고랭이 분포
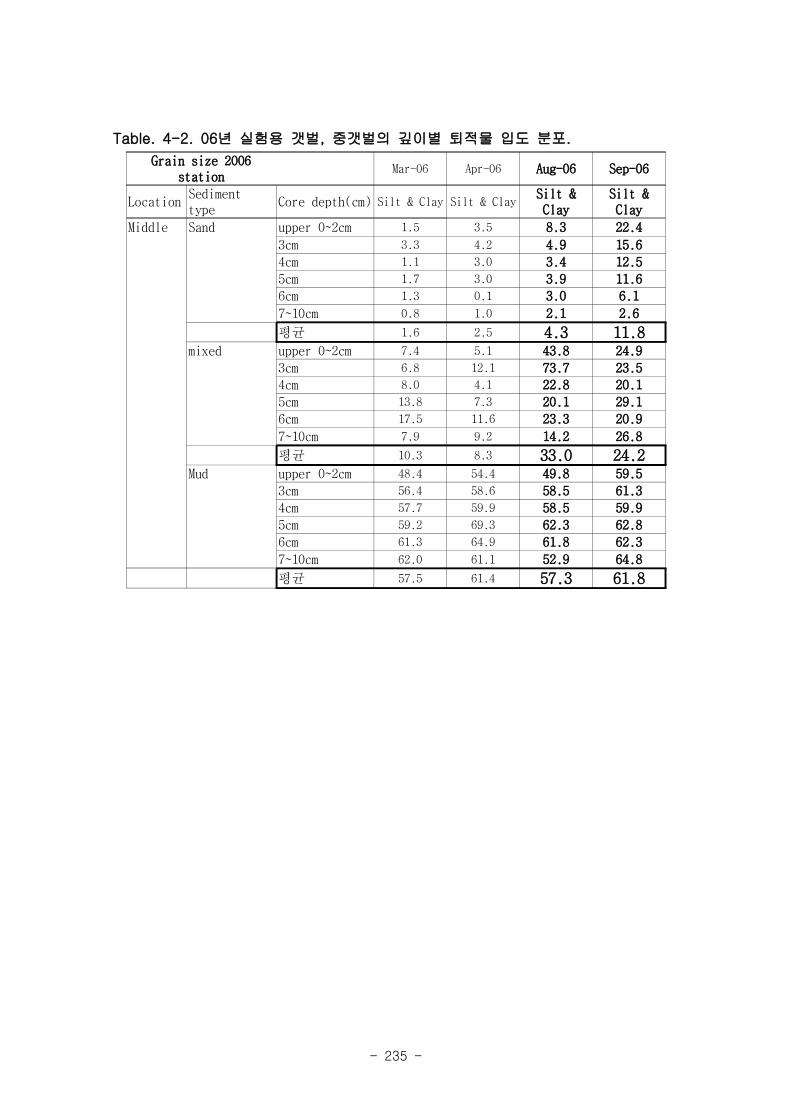
- 235 -
Table. 4-2. 06년 실험용 갯벌, 중갯벌의 깊이별 퇴적물 입도 분포.
Grain size 2006 station
Mar-06 Apr-06 Aug-06 Sep-06
LocationSediment type
Core depth(cm) Silt & Clay Silt & ClaySilt & Clay
Silt & Clay
Middle Sand upper 0~2cm 1.5 3.5 8.3 22.4
3cm 3.3 4.2 4.9 15.6
4cm 1.1 3.0 3.4 12.5
5cm 1.7 3.0 3.9 11.6
6cm 1.3 0.1 3.0 6.1
7~10cm 0.8 1.0 2.1 2.6
평균 1.6 2.5 4.3 11.8 mixed upper 0~2cm 7.4 5.1 43.8 24.9
3cm 6.8 12.1 73.7 23.5
4cm 8.0 4.1 22.8 20.1
5cm 13.8 7.3 20.1 29.1
6cm 17.5 11.6 23.3 20.9
7~10cm 7.9 9.2 14.2 26.8
평균 10.3 8.3 33.0 24.2 Mud upper 0~2cm 48.4 54.4 49.8 59.5
3cm 56.4 58.6 58.5 61.3
4cm 57.7 59.9 58.5 59.9
5cm 59.2 69.3 62.3 62.8
6cm 61.3 64.9 61.8 62.3
7~10cm 62.0 61.1 52.9 64.8
평균 57.5 61.4 57.3 61.8

- 236 -
Figure. 4-19. Artificial intertidal flat in Nakdong estuary.
3. 실험용 갯벌의 환경과 기능변화
가. 실험용갯벌의 설치
연구가 진행된 지역은 낙동강 하구 명지 주거 단지 앞쪽에 명지지역에 설치한
인공 갯벌이다. 2004년의 지형 정밀 측정결과 낙동강 하구갯벌에 위치한 명지 지
역이 실험용 갯벌 적지로 판단되었고, 2005년 4월 실험용 갯벌이 완성되었다(가
로: 1m, 세로:1m, 깊이: 1m). 실험용 갯벌은 해수면 위 85.1, 142.9, 175cm에 서로
다른 3개의 조고(상, 중, 하)를 갖도록 설치하였으며, 각 조고에는 3가지 다른 타
입의 퇴적물(사질, 혼합-명지 본래의 퇴적물, 니질)을 채워 넣었다. 2006년 2월,
05년 실험용갯벌과 같은 조건으로 면적이 4배 늘어난 새로운 인공 갯벌(가로:
2m, 세로: 2m)을 추가 설치하여, 연간 안정화과정을 살펴 보았다.
나. 실험용 갯벌의 입도, 퇴적물 내 유기물 농도 변화
실험용 갯벌을 설치한후 시간의 경과에 따른 퇴적물 특성변화를 앞에서 실험용
갯벌에서 알아보았다
(1) 재료 및 방법
(가) 강열감량
실험용갯벌의 각 treatment별로 깊이별 퇴적물내 유기물 함량을 측정하였다. 현
장에서 코어(지름=4cm, 길이=22cm)를 이용하여 퇴적층 10cm를 채취하여 cm별로
절단하여 실험실 내에서 퇴적물을 고온으로 가열하여 그 무게 차이를 이용하여

- 237 -
퇴적물내 유기물 함량을 측정하였다. 실험 방법 순서는 다음과 같다.
- 25mL 도가니의 무게를 0.001g까지 측정한다. 이 도가니를 건조기에서 110。C
로 말려 항량으로 될 때까지 건조한 후 시험에 사용한다.
- 시료 약 5g의 무게를 정확히 측정하여 25mL 도가니에 담는다. 시료가 담긴
도가니를 역시 110℃에서 항량으로 될 때까지 건조한다.
- 전기로에 시료를 넣고, 550℃의 온도로 2시간 동안 가열한다.
- 가열이 끝나면 전기로의 문을 열고, 도가니를 꺼낸다. 도가니는 데시케이터
안에서 실온으로 식힌다.
- 다 식으면 시료가 담긴 도가니의 무게를 측정한다. 실온으로 완전히 식지 않
으면 무게를 측정하는 도중에 무게가 달라질 수 있다.
위의 결과를 이용하여 강열감량은 다음과 같이 구한다.
강열감량(%)= (가열전 시료 무게-가열 후 시료 무게)/ 가열 전 시료 무게 *
100
(나) 입도 분석
퇴적물의 입도분석은 퇴적물에 과산화수소(H2O2)와 염산(HCl)을 넣어 유기물과
탄산염을 제거한 후 wet sieving에 의해 4Φ으로 퇴적물을 분류하였다. 4Φ이하 조
립질 퇴적물은 sieve shaker로 체질한 후 0.5Φ 간격으로 무게 백분율을 구하였고,
4Φ이상 세립질 퇴적물은 본 연구지역에서는 그 양이 매우 작아 더 이상 퇴적물
분류를 하지 않았다.
(2) 주요 결과 및 토의
(가) 퇴적물내 유기물 함량의 계절 변화
실험용 갯벌 내 퇴적물 내 유기물의함량은 0.7~17.4% 내외였다. 유기물 농도는
입자의 크기와 상관관계가 높아서 Mud에서 높고, Sand에서 낮았다(Table. 4-3~
4). 그러나 시간에 따라 표층 퇴적물은 세립화되고, 각 실험용갯벌에서 유사해 지
는 경향을 보였다. 2005년 8월부터 2006년 9월까지 MIDDLE부분의 퇴적 코어를
각 cm별로 잘라 강열감량 실험을 행했다. 8월, 10월, 11월의 모든 MUD 퇴적물
부분에서 표층으로부터 3~4cm부분에 유기물의 함량이 높아지는 것을 알 수 있
었다. 2005년과 2006년의 실험용 갯벌에서 강열감량으로 측정한 퇴적물내 유기물
함량 결과는 일정한 계절적인 pattern을 나타내지 않고, 조사 월별 변화가 큰 특

- 238 -
징을 보여 하구 갯벌 환경의 급격한 변동을 잘 반영하는 듯하였다. 실험용 갯벌의
조고별 퇴적물내 유기물 함량의 결과 퇴적작용을 거의 받지 않는 조고가 가장 높
은 upper station은 실험용 갯벌 조성 당시의 퇴적물 특성을 그대로 유지하고 있
었다(Fig. 4-20).
그와 반대 경향으로 lower station의 경우는 2006년 2월에 전반적으로 높은 퇴
적물내 유기물 함량이 나타났다(Fig. 4-22). 이는 유기물 함량이 높은 퇴적물이
퇴적되었다고 예상할 수 있는데, 가을에 사그러든 세모고랭이가 퇴적물내 사체 형
태(detritus)로 묻혀 있다가 퇴적 작용으로 가장 높은 조고인 lower station 표층
에 유입되었다고 생각된다. 또한, 2006년 4월과 9월 퇴적물 내 유기물 함량이 낮
아지는 경향이 나타났다. 이는 가을, 겨울 동안 세모고랭이의 죽음으로 인해 퇴적
물내 유기물이 많이 존재는 하지만 유기물이 퇴적물내에서 분해되기보다는 축적
되어 있음을 시사한다. 겨울에는 온도가 낮아 분해되기는 어려우며, 온도가 올라
가는 봄과 여름에 그 축적된 유기물이 분해되어 퇴적물내 유기물 함량이 낮아지
는 것으로 생각된다. middle station은 lower station 보다는 그 경향이 뚜렷하지
는 않지만, 2005년 11월에 퇴적물내 유기물 함량이 가장 높은 것으로 나타났으며,
봄에서 여름으로 갈수록 그 값이 낮아지는 경향을 나타냈다(Fig. 4-21). 또한,
2005년 11월에 육안으로 표층이 퇴적물로 완전히 덮인 것을 확인할 수 있었는데,
그 새로운 퇴적물이 표층을 덮음으로 인해 초기 상태에 실험용으로 넣어준 퇴적
물 특성이 사라졌다고 할 수 있다. 2005년 middle station의 깊이별 퇴적물내 유
기물 함량 결과에서도 시간이 흐를수록 표층 퇴적물 부분의 유기물 함량의 변화
폭이 큰 것으로 나타났다(Fig. 4-21).
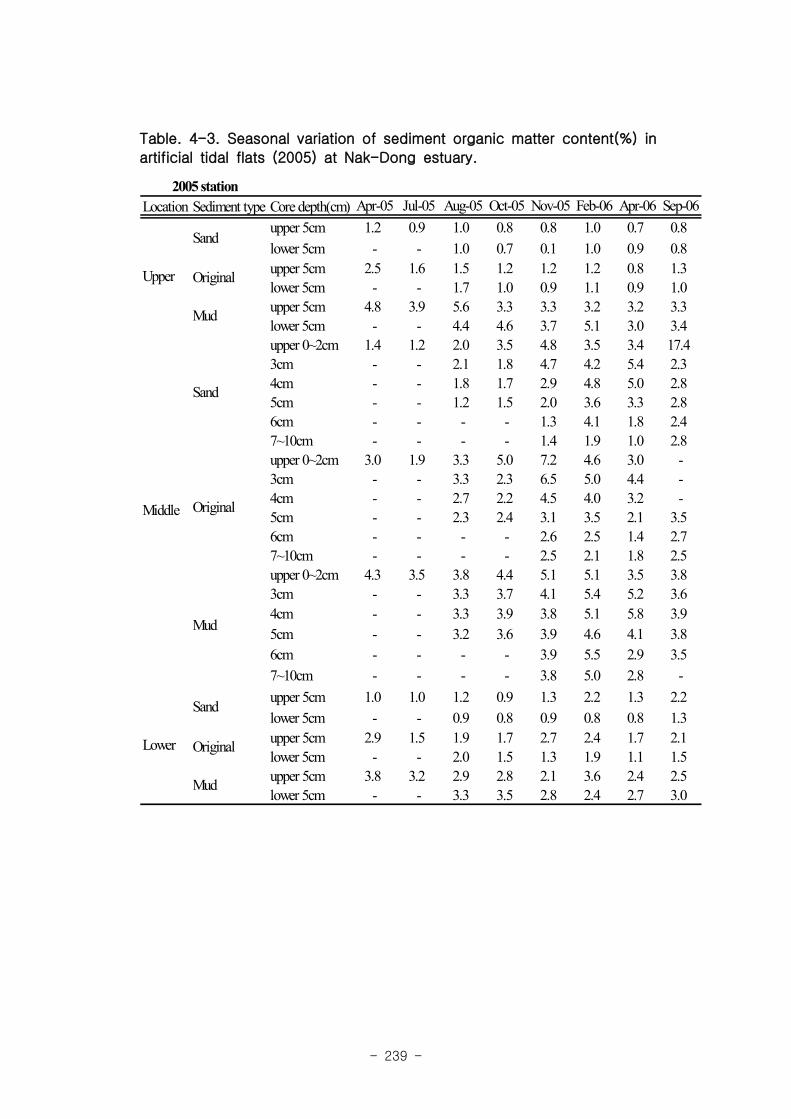
- 239 -
Location Sediment type Core depth(cm) Apr-05 Jul-05 Aug-05 Oct-05 Nov-05 Feb-06 Apr-06 Sep-06
upper 5cm 1.2 0.9 1.0 0.8 0.8 1.0 0.7 0.8
lower 5cm - - 1.0 0.7 0.1 1.0 0.9 0.8
upper 5cm 2.5 1.6 1.5 1.2 1.2 1.2 0.8 1.3
lower 5cm - - 1.7 1.0 0.9 1.1 0.9 1.0
upper 5cm 4.8 3.9 5.6 3.3 3.3 3.2 3.2 3.3
lower 5cm - - 4.4 4.6 3.7 5.1 3.0 3.4
upper 0~2cm 1.4 1.2 2.0 3.5 4.8 3.5 3.4 17.4
3cm - - 2.1 1.8 4.7 4.2 5.4 2.3
4cm - - 1.8 1.7 2.9 4.8 5.0 2.8
5cm - - 1.2 1.5 2.0 3.6 3.3 2.8
6cm - - - - 1.3 4.1 1.8 2.4
7~10cm - - - - 1.4 1.9 1.0 2.8
upper 0~2cm 3.0 1.9 3.3 5.0 7.2 4.6 3.0 -
3cm - - 3.3 2.3 6.5 5.0 4.4 -
4cm - - 2.7 2.2 4.5 4.0 3.2 -
5cm - - 2.3 2.4 3.1 3.5 2.1 3.5
6cm - - - - 2.6 2.5 1.4 2.7
7~10cm - - - - 2.5 2.1 1.8 2.5
upper 0~2cm 4.3 3.5 3.8 4.4 5.1 5.1 3.5 3.8
3cm - - 3.3 3.7 4.1 5.4 5.2 3.6
4cm - - 3.3 3.9 3.8 5.1 5.8 3.9
5cm - - 3.2 3.6 3.9 4.6 4.1 3.8
6cm - - - - 3.9 5.5 2.9 3.5
7~10cm - - - - 3.8 5.0 2.8 -
upper 5cm 1.0 1.0 1.2 0.9 1.3 2.2 1.3 2.2
lower 5cm - - 0.9 0.8 0.9 0.8 0.8 1.3
upper 5cm 2.9 1.5 1.9 1.7 2.7 2.4 1.7 2.1
lower 5cm - - 2.0 1.5 1.3 1.9 1.1 1.5
upper 5cm 3.8 3.2 2.9 2.8 2.1 3.6 2.4 2.5
lower 5cm - - 3.3 3.5 2.8 2.4 2.7 3.0
Upper
Sand
Original
Mud
Middle
Sand
Original
Original
Mud
2005 station
Mud
Lower
Sand
Table. 4-3. Seasonal variation of sediment organic matter content(%) in
artificial tidal flats (2005) at Nak-Dong estuary.
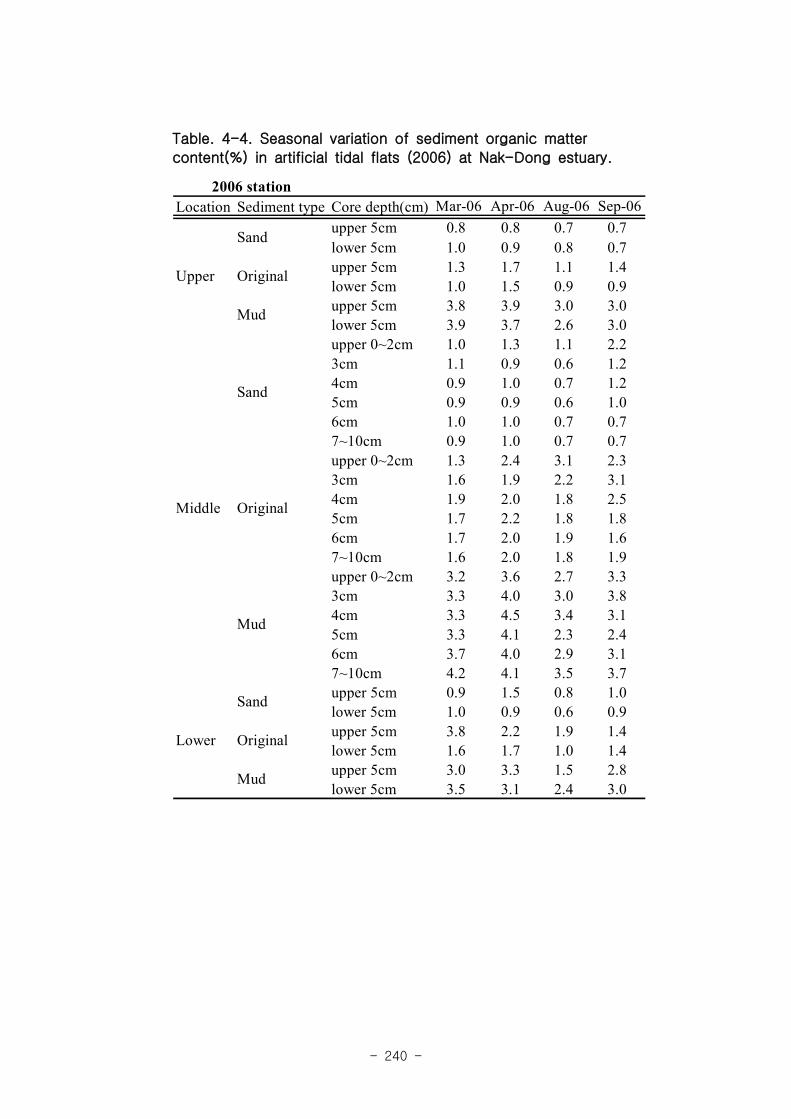
- 240 -
Location Sediment type Core depth(cm) Mar-06 Apr-06 Aug-06 Sep-06
upper 5cm 0.8 0.8 0.7 0.7
lower 5cm 1.0 0.9 0.8 0.7
upper 5cm 1.3 1.7 1.1 1.4
lower 5cm 1.0 1.5 0.9 0.9
upper 5cm 3.8 3.9 3.0 3.0
lower 5cm 3.9 3.7 2.6 3.0
upper 0~2cm 1.0 1.3 1.1 2.2
3cm 1.1 0.9 0.6 1.2
4cm 0.9 1.0 0.7 1.2
5cm 0.9 0.9 0.6 1.0
6cm 1.0 1.0 0.7 0.7
7~10cm 0.9 1.0 0.7 0.7
upper 0~2cm 1.3 2.4 3.1 2.3
3cm 1.6 1.9 2.2 3.1
4cm 1.9 2.0 1.8 2.5
5cm 1.7 2.2 1.8 1.8
6cm 1.7 2.0 1.9 1.6
7~10cm 1.6 2.0 1.8 1.9
upper 0~2cm 3.2 3.6 2.7 3.3
3cm 3.3 4.0 3.0 3.8
4cm 3.3 4.5 3.4 3.1
5cm 3.3 4.1 2.3 2.4
6cm 3.7 4.0 2.9 3.1
7~10cm 4.2 4.1 3.5 3.7
upper 5cm 0.9 1.5 0.8 1.0
lower 5cm 1.0 0.9 0.6 0.9
upper 5cm 3.8 2.2 1.9 1.4
lower 5cm 1.6 1.7 1.0 1.4
upper 5cm 3.0 3.3 1.5 2.8
lower 5cm 3.5 3.1 2.4 3.0
2006 station
Upper
Sand
Original
Mud
Middle
Sand
Original
Mud
Lower
Sand
Original
Mud
Table. 4-4. Seasonal variation of sediment organic matter
content(%) in artificial tidal flats (2006) at Nak-Dong estuary.
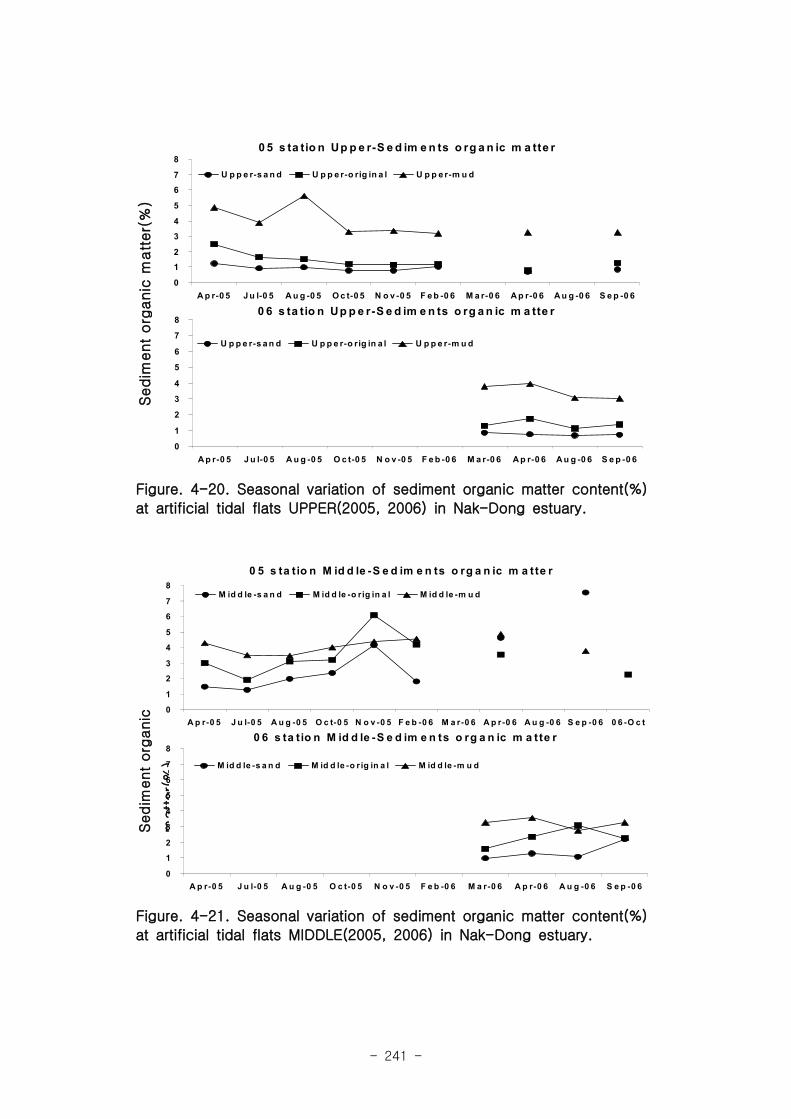
- 241 -
05 s ta tio n Up p e r-S e d im e n ts o rg a n ic m a tte r
0
1
2
3
4
5
6
7
8
A p r-05 Ju l-0 5 A u g -0 5 O c t-05 N o v -0 5 F eb -0 6 M a r-0 6 Ap r-0 6 Au g -0 6 S ep -0 6
U p p e r-s an d U p p e r-o rig in a l U p p e r-m u d
0 6 s ta tio n Up p e r-S e d im e n ts o rg an ic m a tte r
0
1
2
3
4
5
6
7
8
Ap r-0 5 Ju l-0 5 A u g -0 5 O c t-0 5 N o v -0 5 F eb -0 6 M a r-0 6 Ap r-0 6 Au g -0 6 S ep -0 6
U p p e r-s an d U p p e r-o rig in a l U p p e r-m u d
Sedim
ent
org
anic
matt
er(
%)
Figure. 4-20. Seasonal variation of sediment organic matter content(%)
at artificial tidal flats UPPER(2005, 2006) in Nak-Dong estuary.
0 5 s ta tio n M id d le -S e d im e n ts o rg a n ic m a tte r
0
1
2
3
4
5
6
7
8
A p r-0 5 J u l-0 5 A u g -0 5 O c t-0 5 N o v -0 5 F e b -0 6 M a r-0 6 A p r-0 6 A u g -0 6 S e p -0 6 0 6 -O c t
M id d le -s a n d M id d le -o rig in a l M id d le -m u d
0 6 s ta tio n M id d le -S e d im e n ts o rg a n ic m a tte r
0
1
2
3
4
5
6
7
8
A p r-0 5 J u l-0 5 A u g -0 5 O c t-0 5 N o v -0 5 F e b -0 6 M a r-0 6 A p r-0 6 A u g -0 6 S e p -0 6
M id d le -s a n d M id d le -o r ig in a l M id d le -m u d
Sedim
ent
org
anic
matter(
%)
Figure. 4-21. Seasonal variation of sediment organic matter content(%)
at artificial tidal flats MIDDLE(2005, 2006) in Nak-Dong estuary.
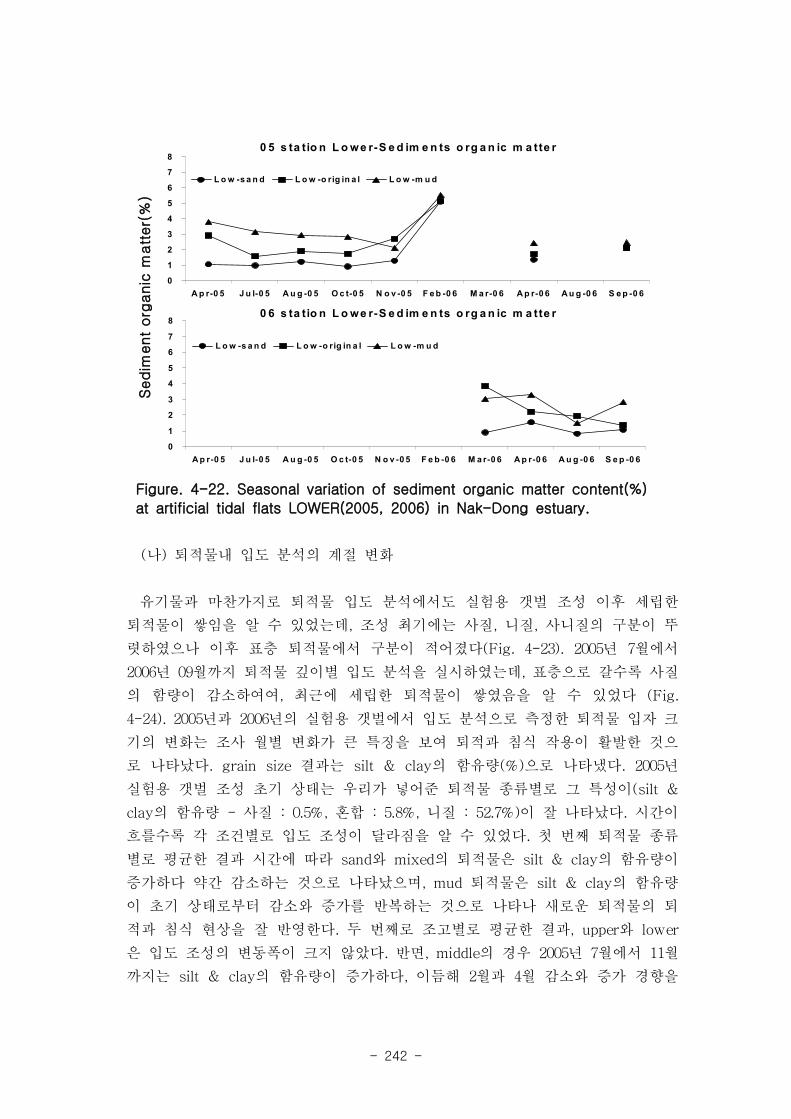
- 242 -
0 5 s ta tio n L o we r-S e d im e n ts o rg a n ic m a tte r
0
1
2
3
4
5
6
7
8
A p r-0 5 J u l-0 5 A u g -0 5 O c t-0 5 N o v -0 5 F eb -0 6 M a r-0 6 Ap r-06 Au g -0 6 S ep -0 6
L o w -sa n d L o w -o rig in a l L o w -m u d
0 6 s ta tio n L o we r-S e d im e n ts o rg a n ic m a tte r
0
1
2
3
4
5
6
7
8
Ap r-0 5 J u l-0 5 A u g -0 5 O c t-0 5 N o v -05 F e b -0 6 M a r-06 Ap r-0 6 A u g -0 6 S e p -0 6
L o w -s a n d L o w -o rig in a l L o w -m u d
Sedim
ent
org
anic
matter(
%)
Figure. 4-22. Seasonal variation of sediment organic matter content(%)
at artificial tidal flats LOWER(2005, 2006) in Nak-Dong estuary.
(나) 퇴적물내 입도 분석의 계절 변화
유기물과 마찬가지로 퇴적물 입도 분석에서도 실험용 갯벌 조성 이후 세립한
퇴적물이 쌓임을 알 수 있었는데, 조성 최기에는 사질, 니질, 사니질의 구분이 뚜
렷하였으나 이후 표층 퇴적물에서 구분이 적어졌다(Fig. 4-23). 2005년 7월에서
2006년 09월까지 퇴적물 깊이별 입도 분석을 실시하였는데, 표층으로 갈수록 사질
의 함량이 감소하여여, 최근에 세립한 퇴적물이 쌓였음을 알 수 있었다 (Fig.
4-24). 2005년과 2006년의 실험용 갯벌에서 입도 분석으로 측정한 퇴적물 입자 크
기의 변화는 조사 월별 변화가 큰 특징을 보여 퇴적과 침식 작용이 활발한 것으
로 나타났다. grain size 결과는 silt & clay의 함유량(%)으로 나타냈다. 2005년
실험용 갯벌 조성 초기 상태는 우리가 넣어준 퇴적물 종류별로 그 특성이(silt &
clay의 함유량 - 사질 : 0.5%, 혼합 : 5.8%, 니질 : 52.7%)이 잘 나타났다. 시간이
흐를수록 각 조건별로 입도 조성이 달라짐을 알 수 있었다. 첫 번째 퇴적물 종류
별로 평균한 결과 시간에 따라 sand와 mixed의 퇴적물은 silt & clay의 함유량이
증가하다 약간 감소하는 것으로 나타났으며, mud 퇴적물은 silt & clay의 함유량
이 초기 상태로부터 감소와 증가를 반복하는 것으로 나타나 새로운 퇴적물의 퇴
적과 침식 현상을 잘 반영한다. 두 번째로 조고별로 평균한 결과, upper와 lower
은 입도 조성의 변동폭이 크지 않았다. 반면, middle의 경우 2005년 7월에서 11월
까지는 silt & clay의 함유량이 증가하다, 이듬해 2월과 4월 감소와 증가 경향을
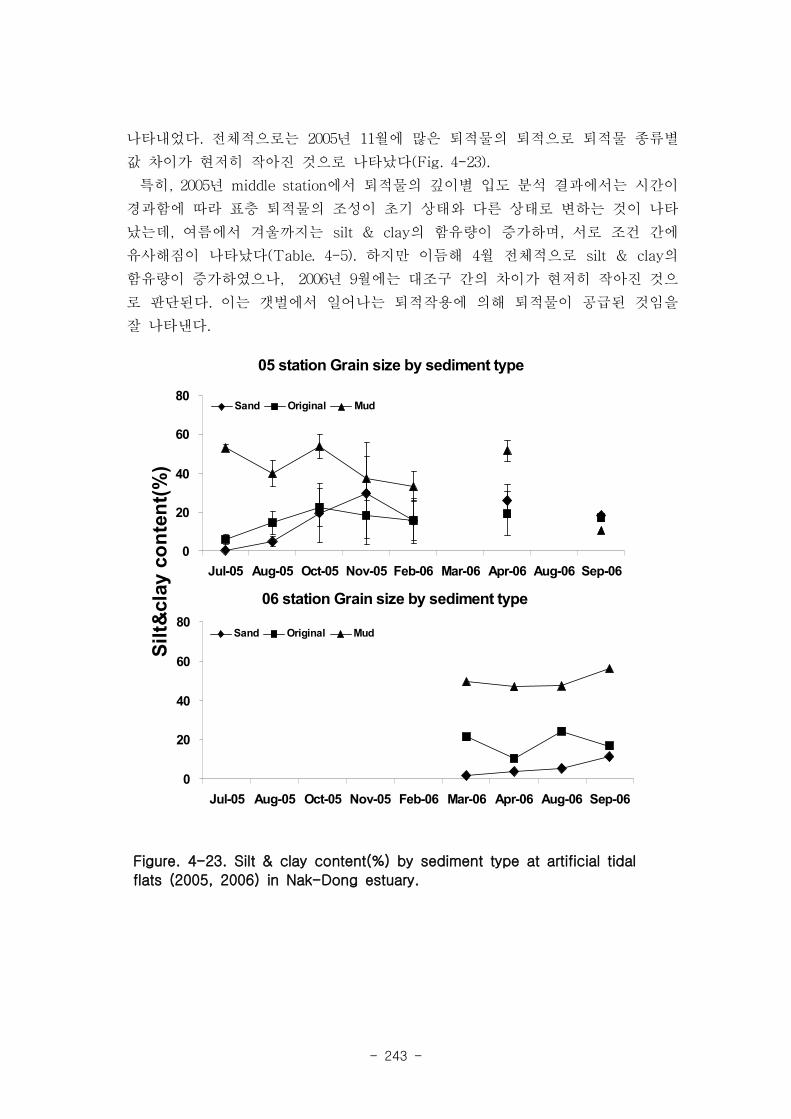
- 243 -
05 station Grain size by sediment type
0
20
40
60
80
Jul-05 Aug-05 Oct-05 Nov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sand Original Mud
06 station Grain size by sediment type
0
20
40
60
80
Jul-05 Aug-05 Oct-05 Nov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sand Original Mud
Silt&
cla
y c
onte
nt(
%)
Figure. 4-23. Silt & clay content(%) by sediment type at artificial tidal
flats (2005, 2006) in Nak-Dong estuary.
나타내었다. 전체적으로는 2005년 11월에 많은 퇴적물의 퇴적으로 퇴적물 종류별
값 차이가 현저히 작아진 것으로 나타났다(Fig. 4-23).
특히, 2005년 middle station에서 퇴적물의 깊이별 입도 분석 결과에서는 시간이
경과함에 따라 표층 퇴적물의 조성이 초기 상태와 다른 상태로 변하는 것이 나타
났는데, 여름에서 겨울까지는 silt & clay의 함유량이 증가하며, 서로 조건 간에
유사해짐이 나타났다(Table. 4-5). 하지만 이듬해 4월 전체적으로 silt & clay의
함유량이 증가하였으나, 2006년 9월에는 대조구 간의 차이가 현저히 작아진 것으
로 판단된다. 이는 갯벌에서 일어나는 퇴적작용에 의해 퇴적물이 공급된 것임을
잘 나타낸다.
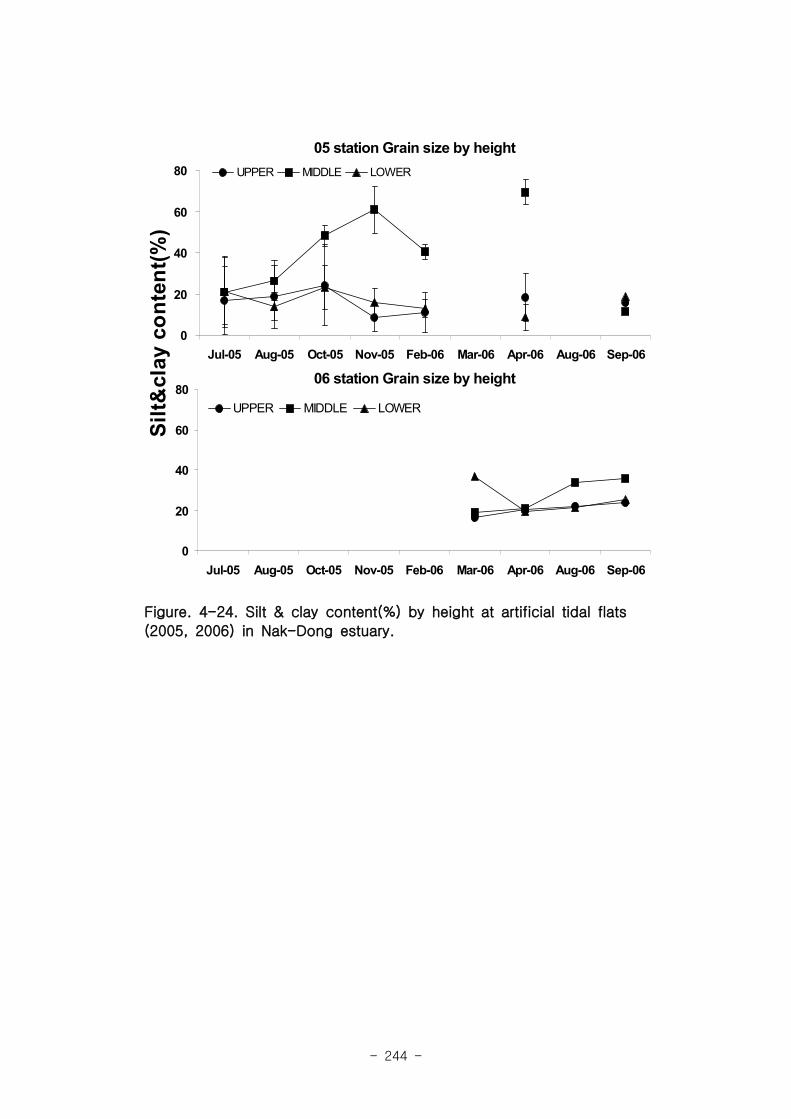
- 244 -
05 station Grain size by height
0
20
40
60
80
Jul-05 Aug-05 Oct-05 Nov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
UPPER MIDDLE LOWER
06 station Grain size by height
0
20
40
60
80
Jul-05 Aug-05 Oct-05 Nov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
UPPER MIDDLE LOWER
Silt&
cla
y c
onte
nt(%
)
Figure. 4-24. Silt & clay content(%) by height at artificial tidal flats
(2005, 2006) in Nak-Dong estuary.
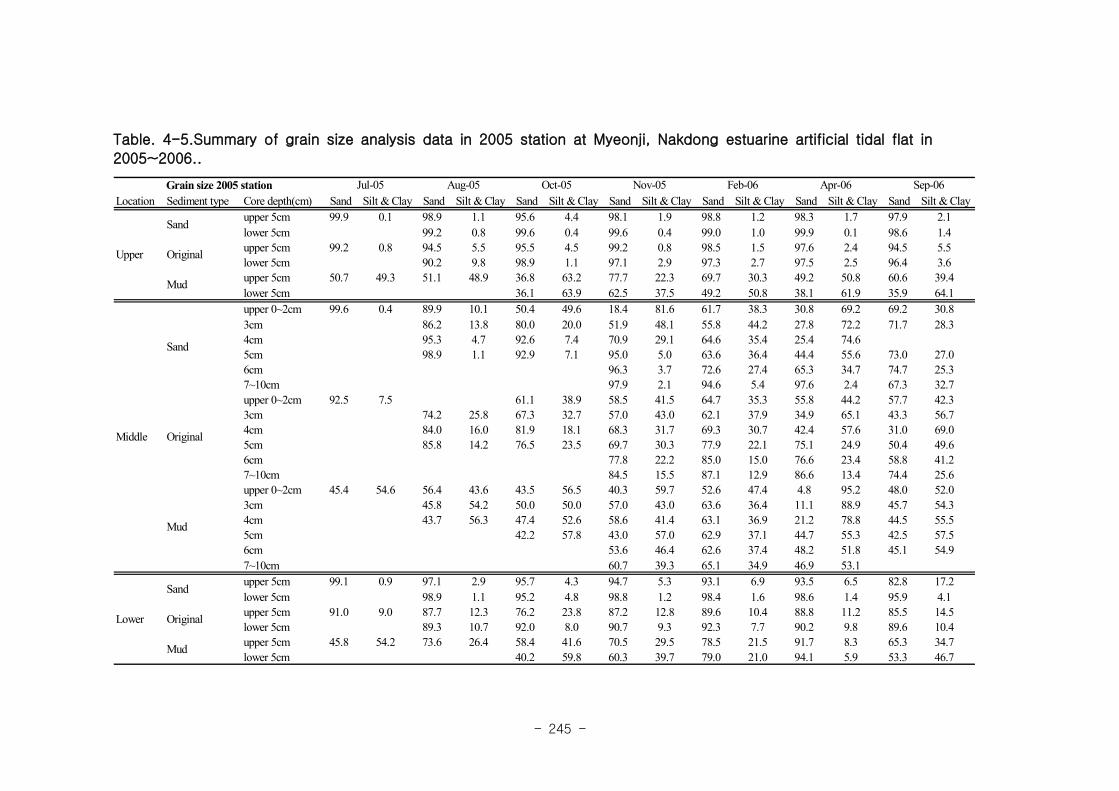
- 245 -
Location Sediment type Core depth(cm) Sand Silt & Clay Sand Silt & Clay Sand Silt & Clay Sand Silt & Clay Sand Silt & Clay Sand Silt & Clay Sand Silt & Clay
upper 5cm 99.9 0.1 98.9 1.1 95.6 4.4 98.1 1.9 98.8 1.2 98.3 1.7 97.9 2.1
lower 5cm 99.2 0.8 99.6 0.4 99.6 0.4 99.0 1.0 99.9 0.1 98.6 1.4
upper 5cm 99.2 0.8 94.5 5.5 95.5 4.5 99.2 0.8 98.5 1.5 97.6 2.4 94.5 5.5
lower 5cm 90.2 9.8 98.9 1.1 97.1 2.9 97.3 2.7 97.5 2.5 96.4 3.6
upper 5cm 50.7 49.3 51.1 48.9 36.8 63.2 77.7 22.3 69.7 30.3 49.2 50.8 60.6 39.4
lower 5cm 36.1 63.9 62.5 37.5 49.2 50.8 38.1 61.9 35.9 64.1
upper 0~2cm 99.6 0.4 89.9 10.1 50.4 49.6 18.4 81.6 61.7 38.3 30.8 69.2 69.2 30.8
3cm 86.2 13.8 80.0 20.0 51.9 48.1 55.8 44.2 27.8 72.2 71.7 28.3
4cm 95.3 4.7 92.6 7.4 70.9 29.1 64.6 35.4 25.4 74.6
5cm 98.9 1.1 92.9 7.1 95.0 5.0 63.6 36.4 44.4 55.6 73.0 27.0
6cm 96.3 3.7 72.6 27.4 65.3 34.7 74.7 25.3
7~10cm 97.9 2.1 94.6 5.4 97.6 2.4 67.3 32.7
upper 0~2cm 92.5 7.5 61.1 38.9 58.5 41.5 64.7 35.3 55.8 44.2 57.7 42.3
3cm 74.2 25.8 67.3 32.7 57.0 43.0 62.1 37.9 34.9 65.1 43.3 56.7
4cm 84.0 16.0 81.9 18.1 68.3 31.7 69.3 30.7 42.4 57.6 31.0 69.0
5cm 85.8 14.2 76.5 23.5 69.7 30.3 77.9 22.1 75.1 24.9 50.4 49.6
6cm 77.8 22.2 85.0 15.0 76.6 23.4 58.8 41.2
7~10cm 84.5 15.5 87.1 12.9 86.6 13.4 74.4 25.6
upper 0~2cm 45.4 54.6 56.4 43.6 43.5 56.5 40.3 59.7 52.6 47.4 4.8 95.2 48.0 52.0
3cm 45.8 54.2 50.0 50.0 57.0 43.0 63.6 36.4 11.1 88.9 45.7 54.3
4cm 43.7 56.3 47.4 52.6 58.6 41.4 63.1 36.9 21.2 78.8 44.5 55.5
5cm 42.2 57.8 43.0 57.0 62.9 37.1 44.7 55.3 42.5 57.5
6cm 53.6 46.4 62.6 37.4 48.2 51.8 45.1 54.9
7~10cm 60.7 39.3 65.1 34.9 46.9 53.1
upper 5cm 99.1 0.9 97.1 2.9 95.7 4.3 94.7 5.3 93.1 6.9 93.5 6.5 82.8 17.2
lower 5cm 98.9 1.1 95.2 4.8 98.8 1.2 98.4 1.6 98.6 1.4 95.9 4.1
upper 5cm 91.0 9.0 87.7 12.3 76.2 23.8 87.2 12.8 89.6 10.4 88.8 11.2 85.5 14.5
lower 5cm 89.3 10.7 92.0 8.0 90.7 9.3 92.3 7.7 90.2 9.8 89.6 10.4
upper 5cm 45.8 54.2 73.6 26.4 58.4 41.6 70.5 29.5 78.5 21.5 91.7 8.3 65.3 34.7
lower 5cm 40.2 59.8 60.3 39.7 79.0 21.0 94.1 5.9 53.3 46.7
Lower
Sand
Original
Mud
Middle
Sand
Original
Mud
Upper
Sand
Original
Mud
Nov-05 Feb-06 Apr-06 Sep-06Grain size 2005 station Jul-05 Aug-05 Oct-05
Table. 4-5.Summary of grain size analysis data in 2005 station at Myeonji, Nakdong estuarine artificial tidal flat in
2005~2006..
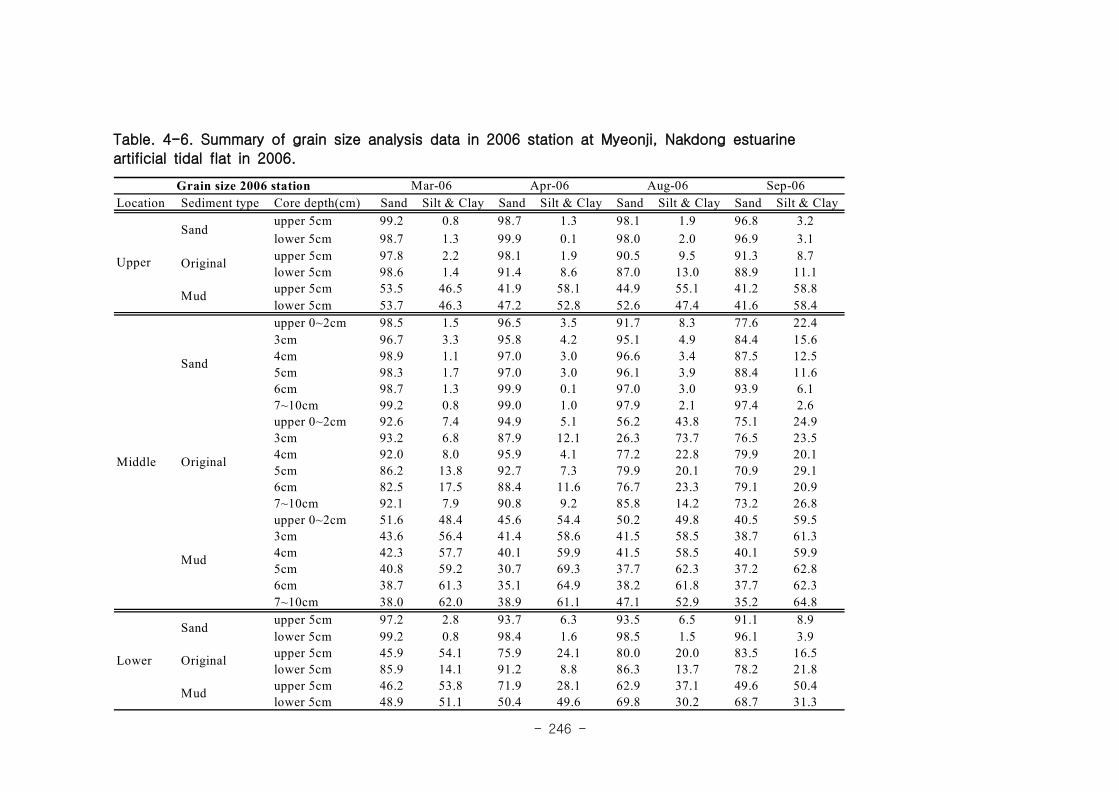
- 246 -
Location Sediment type Core depth(cm) Sand Silt & Clay Sand Silt & Clay Sand Silt & Clay Sand Silt & Clay
upper 5cm 99.2 0.8 98.7 1.3 98.1 1.9 96.8 3.2
lower 5cm 98.7 1.3 99.9 0.1 98.0 2.0 96.9 3.1
upper 5cm 97.8 2.2 98.1 1.9 90.5 9.5 91.3 8.7
lower 5cm 98.6 1.4 91.4 8.6 87.0 13.0 88.9 11.1
upper 5cm 53.5 46.5 41.9 58.1 44.9 55.1 41.2 58.8
lower 5cm 53.7 46.3 47.2 52.8 52.6 47.4 41.6 58.4
upper 0~2cm 98.5 1.5 96.5 3.5 91.7 8.3 77.6 22.4
3cm 96.7 3.3 95.8 4.2 95.1 4.9 84.4 15.6
4cm 98.9 1.1 97.0 3.0 96.6 3.4 87.5 12.5
5cm 98.3 1.7 97.0 3.0 96.1 3.9 88.4 11.6
6cm 98.7 1.3 99.9 0.1 97.0 3.0 93.9 6.1
7~10cm 99.2 0.8 99.0 1.0 97.9 2.1 97.4 2.6
upper 0~2cm 92.6 7.4 94.9 5.1 56.2 43.8 75.1 24.9
3cm 93.2 6.8 87.9 12.1 26.3 73.7 76.5 23.5
4cm 92.0 8.0 95.9 4.1 77.2 22.8 79.9 20.1
5cm 86.2 13.8 92.7 7.3 79.9 20.1 70.9 29.1
6cm 82.5 17.5 88.4 11.6 76.7 23.3 79.1 20.9
7~10cm 92.1 7.9 90.8 9.2 85.8 14.2 73.2 26.8
upper 0~2cm 51.6 48.4 45.6 54.4 50.2 49.8 40.5 59.5
3cm 43.6 56.4 41.4 58.6 41.5 58.5 38.7 61.3
4cm 42.3 57.7 40.1 59.9 41.5 58.5 40.1 59.9
5cm 40.8 59.2 30.7 69.3 37.7 62.3 37.2 62.8
6cm 38.7 61.3 35.1 64.9 38.2 61.8 37.7 62.3
7~10cm 38.0 62.0 38.9 61.1 47.1 52.9 35.2 64.8
upper 5cm 97.2 2.8 93.7 6.3 93.5 6.5 91.1 8.9
lower 5cm 99.2 0.8 98.4 1.6 98.5 1.5 96.1 3.9
upper 5cm 45.9 54.1 75.9 24.1 80.0 20.0 83.5 16.5
lower 5cm 85.9 14.1 91.2 8.8 86.3 13.7 78.2 21.8
upper 5cm 46.2 53.8 71.9 28.1 62.9 37.1 49.6 50.4
lower 5cm 48.9 51.1 50.4 49.6 69.8 30.2 68.7 31.3
Lower
Sand
Original
Mud
Grain size 2006 station Mar-06 Apr-06
Middle
Sand
Original
Mud
Upper
Sand
Original
Mud
Aug-06 Sep-06
Table. 4-6. Summary of grain size analysis data in 2006 station at Myeonji, Nakdong estuarine
artificial tidal flat in 2006.
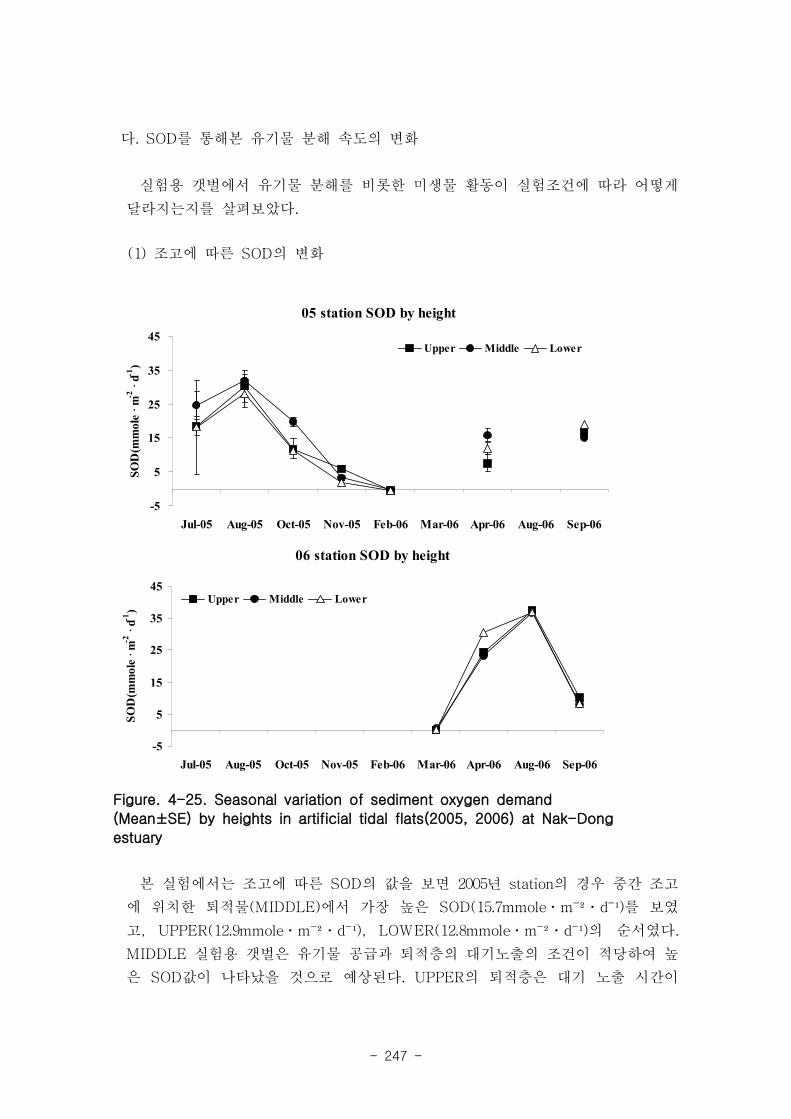
- 247 -
다. SOD를 통해본 유기물 분해 속도의 변화
실험용 갯벌에서 유기물 분해를 비롯한 미생물 활동이 실험조건에 따라 어떻게
달라지는지를 살펴보았다.
(1) 조고에 따른 SOD의 변화
05 station SOD by height
-5
5
15
25
35
45
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
SOD(m
mole · m
-2 · d
-1)
Upper Middle Lower
06 station SOD by height
-5
5
15
25
35
45
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
SOD(m
mole · m
-2 · d-1)
Upper Middle Lower
Figure. 4-25. Seasonal variation of sediment oxygen demand
(Mean±SE) by heights in artificial tidal flats(2005, 2006) at Nak-Dong
estuary
본 실험에서는 조고에 따른 SOD의 값을 보면 2005년 station의 경우 중간 조고
에 위치한 퇴적물(MIDDLE)에서 가장 높은 SOD(15.7mmoleㆍm⁻²ㆍd⁻¹)를 보였
고, UPPER(12.9mmoleㆍm⁻²ㆍd⁻¹), LOWER(12.8mmoleㆍm⁻²ㆍd⁻¹)의 순서였다.
MIDDLE 실험용 갯벌은 유기물 공급과 퇴적층의 대기노출의 조건이 적당하여 높
은 SOD값이 나타났을 것으로 예상된다. UPPER의 퇴적층은 대기 노출 시간이
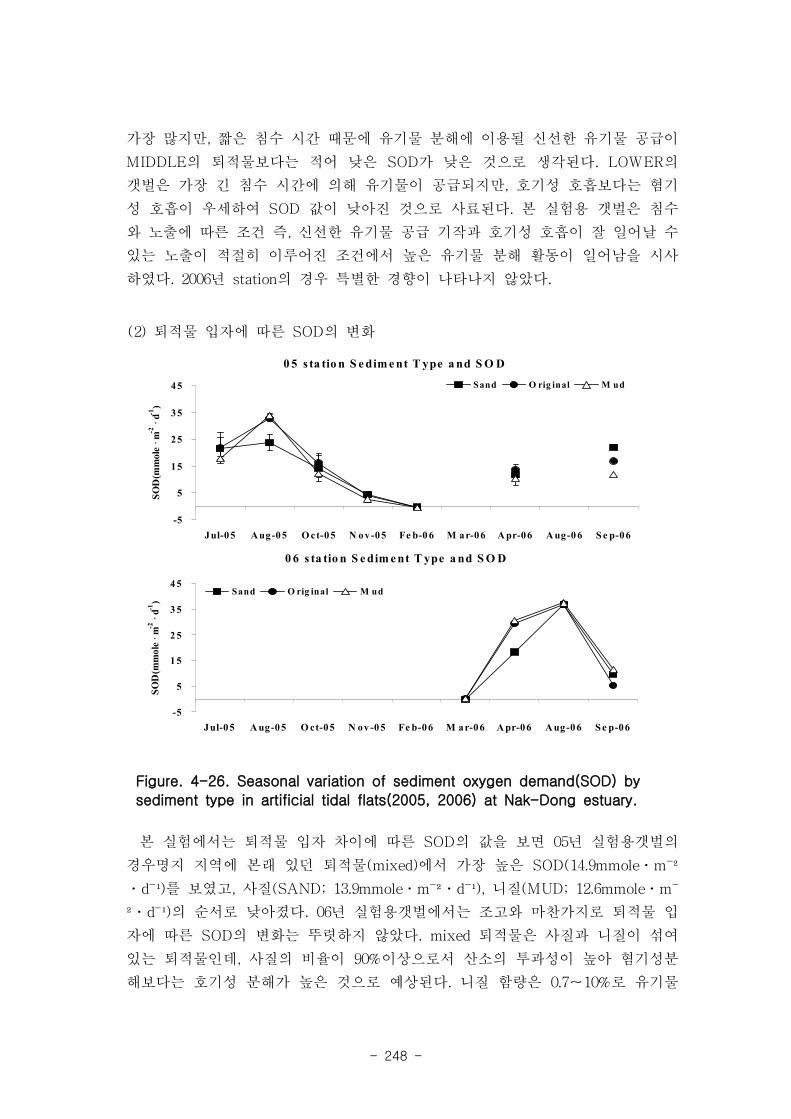
- 248 -
가장 많지만, 짧은 침수 시간 때문에 유기물 분해에 이용될 신선한 유기물 공급이
MIDDLE의 퇴적물보다는 적어 낮은 SOD가 낮은 것으로 생각된다. LOWER의
갯벌은 가장 긴 침수 시간에 의해 유기물이 공급되지만, 호기성 호흡보다는 혐기
성 호흡이 우세하여 SOD 값이 낮아진 것으로 사료된다. 본 실험용 갯벌은 침수
와 노출에 따른 조건 즉, 신선한 유기물 공급 기작과 호기성 호흡이 잘 일어날 수
있는 노출이 적절히 이루어진 조건에서 높은 유기물 분해 활동이 일어남을 시사
하였다. 2006년 station의 경우 특별한 경향이 나타나지 않았다.
(2) 퇴적물 입자에 따른 SOD의 변화
0 5 s ta tio n S ediment T ype a nd SO D
-5
5
15
25
35
45
Jul-05 Aug-05 Oct-05 � ov-05 Fe b-06 M ar-06 Apr-06 Aug-06 Se p-06
SOD(m
mole · m
-2 · d-1)
Sand O rig inal M ud
0 6 s ta tio n S ediment T ype a nd SOD
-5
5
15
25
35
45
Jul-05 Aug-05 Oct-05 � ov -05 Fe b-06 M ar-06 Apr-06 Aug-06 Se p-06
SOD(m
mole · m
-2 · d
-1)
Sand O rig inal M ud
Figure. 4-26. Seasonal variation of sediment oxygen demand(SOD) by
sediment type in artificial tidal flats(2005, 2006) at Nak-Dong estuary.
본 실험에서는 퇴적물 입자 차이에 따른 SOD의 값을 보면 05년 실험용갯벌의
경우명지 지역에 본래 있던 퇴적물(mixed)에서 가장 높은 SOD(14.9mmoleㆍm⁻²ㆍd⁻¹)를 보였고, 사질(SAND; 13.9mmoleㆍm⁻²ㆍd⁻¹), 니질(MUD; 12.6mmoleㆍm⁻²ㆍd⁻¹)의 순서로 낮아졌다. 06년 실험용갯벌에서는 조고와 마찬가지로 퇴적물 입
자에 따른 SOD의 변화는 뚜렷하지 않았다. mixed 퇴적물은 사질과 니질이 섞여
있는 퇴적물인데, 사질의 비율이 90%이상으로서 산소의 투과성이 높아 혐기성분
해보다는 호기성 분해가 높은 것으로 예상된다. 니질 함량은 0.7~10%로 유기물

- 249 -
농도도 SAND 퇴적물보다는 높았다. mixed 퇴적물에서는 유기물 공급이 원활한
상태에서 호기성 분해가 활발하여, 높은 SOD를 보인 것으로 추정된다(Seiki et
al., 1994; Jorgensen and Revsbech, 1986). 유기물 농도가 높았던 MUD 퇴적물
보다 유기물 농도가 가장 낮은 SA퇴적물이 높은 SOD를 나타내어, 본 지역에서
퇴적물에 존재하는 유기물이 주로 난분해성 유기물로 구성되어 있음을 알 수 있
었다. 강화도 갯벌에서 실시한 퇴적물 배양 실험에서도 신선한 유기물의 첨가가
SOD 및 탈질소화를 촉진하여, 갯벌 퇴적물에 존재하는 유기물이 미생물 분해에
활발하게 이용되지 않음을 시사하였다(안 2006).
(3) 퇴적물내 유기물 농도(I.L.)에 따른 SOD의 변화
퇴적물내 유기물 함량의 농도가 SOD에 미치는 영향은 2가지 측면에서 살펴볼
수 있는데, 첫째는 유기물 양이고, 둘째는 유기물의 질이다(Boon et al., 1999;
Seiki et al., 1994; 안 2005). 일반적으로 연안 하구 지역 갯벌 퇴적물내 유기물의
양을 달라지게 만드는 공급기작은 강우와 조석 등에 따라 달라진다(Boon et al.,
1999). 안(2005)은 마산만 갯벌에서 측정한 SOD결과와 마산만으로 유입되는 유기
물 유입량(담수 유입량)과 높은 상관관계를 보여, 갯벌의 유기물 분해가 유기물
유입량에 의해 달라짐을 보고하였다. 낙동강 실험용 갯벌로 유기물이 유입되는 시
기인 7, 8월에는 온도와 광량의 조건이 일차생산량을 증가하게 만들 것이다. 결국
식물 플랑크톤에 의해 일차 생산물의 수층 유입(미생물에 의해 잘 분해될 수 있
는 신선한 유기물)이 SOD에 중요한 영향을 미칠 것으로 예상된다(Boon et al.,
1999; 현 등 2003; Seiki et al., 1994). 위 그림은 퇴적물내 유기물 함량과 SOD를
각 계절별로 나타낸 그림이다. 2005년 station의 경우 우기인 7월과 8월의 퇴적물
내 유기물 함량은 각각 2.07%, 2.51%로 8월에 더 높은 값을 나타냈다(Table.
4-3.). 7월과 8월에는 많은 강우의 영향으로 낙동강 하구 수문을 개방함에 따라
육상기원 유기물이 유입이 되고, 또한 온도가 높아 미생물 분해 활동이 활발히 일
어난 것으로 예상된다. 건기인 10월과 11월의 유기물 함량은 각각 2.25%, 2.87%
로 우기보다 약간 더 높은 유기물 농도가 나타났다(Table. 4-3). 이는 10월 중순
쯤 실험용 갯벌 근처 세모고랭이 군락이 갯벌 생태계 내에서 탈락하는 시기인데,
이때 세모고랭이 잎과 줄기 등이 철새의 먹이로 이용되면서 잘라져 외해로 유출
되거나 퇴적물내에 묻히기 때문에 퇴적물내 유기물 함량이 높게 나타났다고 할
수 있다. 하지만 유기물 함량과 SOD의 상관관계가 높았던 8월(30.1mmoleㆍm⁻²ㆍd⁻¹)과는 달리 11월(2.9mmoleㆍm⁻²ㆍd⁻¹)에는 낮은 상관관계가 나타나는데, 이는
미생물 분해 활동도에 크게 영향을 미치는 온도가 SOD 값에 더 큰 영향을 미쳤
음을 시사한다.
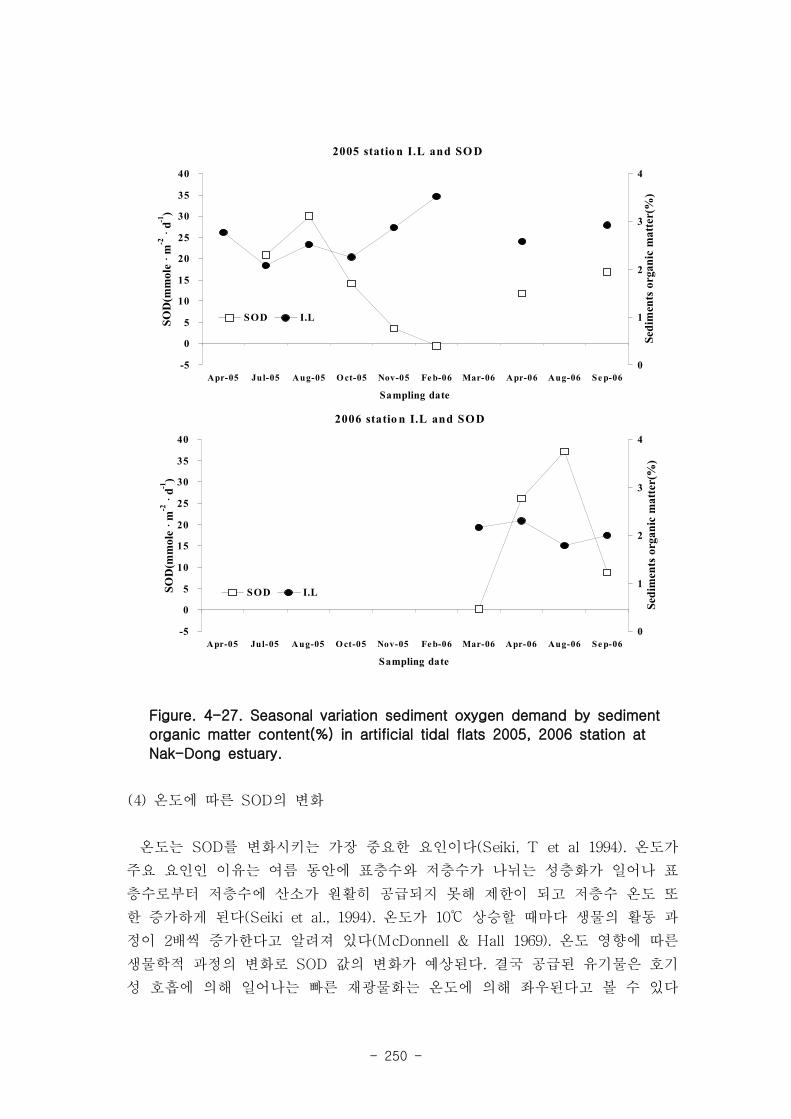
- 250 -
2005 station I.L and SOD
-5
0
5
10
15
20
25
30
35
40
Apr-05 Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
SOD(m
mole · m
-2 · d
-1)
0
1
2
3
4
Sedim
ents organic m
atter(%
)
SOD I.L
2006 statio n I.L and SOD
-5
0
5
10
15
20
25
30
35
40
Apr-05 Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
SOD(m
mole · m
-2 · d
-1)
0
1
2
3
4
Sedim
ents organic m
atter(%)
SOD I.L
Figure. 4-27. Seasonal variation sediment oxygen demand by sediment
organic matter content(%) in artificial tidal flats 2005, 2006 station at
Nak-Dong estuary.
(4) 온도에 따른 SOD의 변화
온도는 SOD를 변화시키는 가장 중요한 요인이다(Seiki, T et al 1994). 온도가
주요 요인인 이유는 여름 동안에 표층수와 저층수가 나뉘는 성층화가 일어나 표
층수로부터 저층수에 산소가 원활히 공급되지 못해 제한이 되고 저층수 온도 또
한 증가하게 된다(Seiki et al., 1994). 온도가 10℃ 상승할 때마다 생물의 활동 과
정이 2배씩 증가한다고 알려져 있다(McDonnell & Hall 1969). 온도 영향에 따른
생물학적 과정의 변화로 SOD 값의 변화가 예상된다. 결국 공급된 유기물은 호기
성 호흡에 의해 일어나는 빠른 재광물화는 온도에 의해 좌우된다고 볼 수 있다
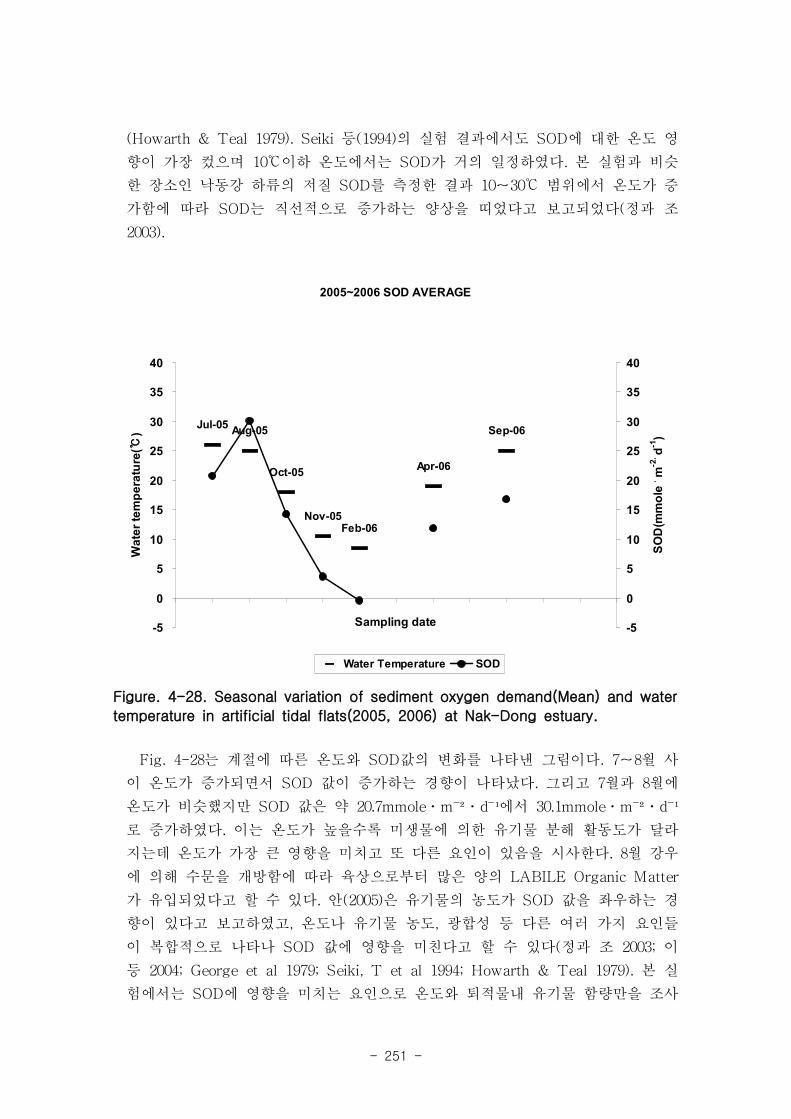
- 251 -
(Howarth & Teal 1979). Seiki 등(1994)의 실험 결과에서도 SOD에 대한 온도 영
향이 가장 컸으며 10℃이하 온도에서는 SOD가 거의 일정하였다. 본 실험과 비슷
한 장소인 낙동강 하류의 저질 SOD를 측정한 결과 10~30℃ 범위에서 온도가 증
가함에 따라 SOD는 직선적으로 증가하는 양상을 띠었다고 보고되었다(정과 조
2003).
2005~2006 SOD AVERAGE
Jul-05Aug-05
Oct-05
Nov-05Feb-06
Apr-06
Sep-06
-5
0
5
10
15
20
25
30
35
40
Sampling date
Wate
r te
mpera
ture
(℃)
-5
0
5
10
15
20
25
30
35
40
SO
D(m
mole
. m-2
. d
-1)
Water Temperature SOD
Figure. 4-28. Seasonal variation of sediment oxygen demand(Mean) and water
temperature in artificial tidal flats(2005, 2006) at Nak-Dong estuary.
Fig. 4-28는 계절에 따른 온도와 SOD값의 변화를 나타낸 그림이다. 7~8월 사
이 온도가 증가되면서 SOD 값이 증가하는 경향이 나타났다. 그리고 7월과 8월에
온도가 비슷했지만 SOD 값은 약 20.7mmoleㆍm⁻²ㆍd⁻¹에서 30.1mmoleㆍm⁻²ㆍd⁻¹로 증가하였다. 이는 온도가 높을수록 미생물에 의한 유기물 분해 활동도가 달라
지는데 온도가 가장 큰 영향을 미치고 또 다른 요인이 있음을 시사한다. 8월 강우
에 의해 수문을 개방함에 따라 육상으로부터 많은 양의 LABILE Organic Matter
가 유입되었다고 할 수 있다. 안(2005)은 유기물의 농도가 SOD 값을 좌우하는 경
향이 있다고 보고하였고, 온도나 유기물 농도, 광합성 등 다른 여러 가지 요인들
이 복합적으로 나타나 SOD 값에 영향을 미친다고 할 수 있다(정과 조 2003; 이
등 2004; George et al 1979; Seiki, T et al 1994; Howarth & Teal 1979). 본 실
험에서는 SOD에 영향을 미치는 요인으로 온도와 퇴적물내 유기물 함량만을 조사
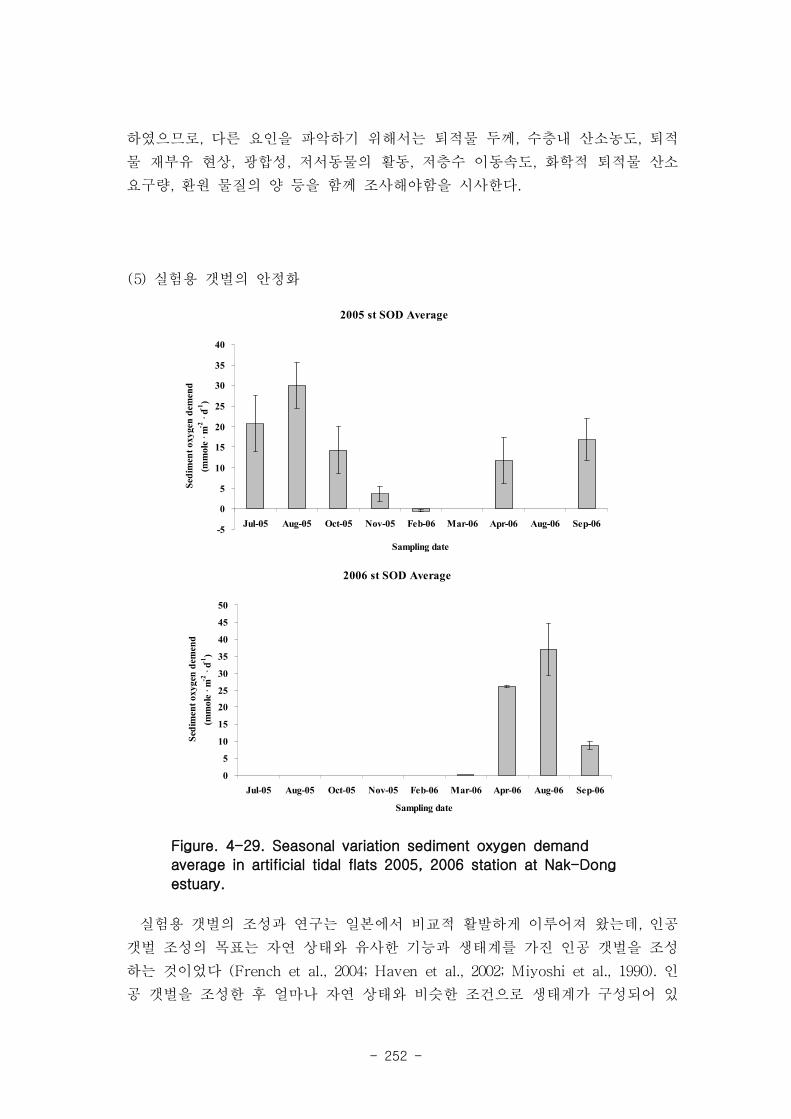
- 252 -
하였으므로, 다른 요인을 파악하기 위해서는 퇴적물 두께, 수층내 산소농도, 퇴적
물 재부유 현상, 광합성, 저서동물의 활동, 저층수 이동속도, 화학적 퇴적물 산소
요구량, 환원 물질의 양 등을 함께 조사해야함을 시사한다.
(5) 실험용 갯벌의 안정화
2005 st SOD Average
-5
0
5
10
15
20
25
30
35
40
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
Sed
iment oxygen
dem
end
(mmole · m
-2 · d-1)
2006 st SOD Average
0
5
10
15
20
25
30
35
40
45
50
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Sampling date
Sedim
ent oxygen dem
end
(mmole · m
-2 · d-1)
Figure. 4-29. Seasonal variation sediment oxygen demand
average in artificial tidal flats 2005, 2006 station at Nak-Dong
estuary.
실험용 갯벌의 조성과 연구는 일본에서 비교적 활발하게 이루어져 왔는데, 인공
갯벌 조성의 목표는 자연 상태와 유사한 기능과 생태계를 가진 인공 갯벌을 조성
하는 것이었다 (French et al., 2004; Haven et al., 2002; Miyoshi et al., 1990). 인
공 갯벌을 조성한 후 얼마나 자연 상태와 비슷한 조건으로 생태계가 구성되어 있

- 253 -
는가를 자연의 갯벌과 비교함으로써, 안정화 정도를 평가할 수 있다(Haven et al.,
2002; Lee et al 1998). 여러 지역에서 인공 갯벌조성 후 약 10년 뒤 인공 갯벌의
안정화된 결과로 생물 서식지와 은신처의 역할이 자연갯벌과 유사하였고, 비안정
화된 경우로는 미국 버지니아에 조성된 인공갯벌의 물리 화학적 조건이 자연 갯
벌과 다르다는 결과가 보고되었다(Miyoshi et al., 1990; French et al., 2004;
Haven et al., 2002; Lee et al., 1998). 인공갯벌에서 서식환경으로 퇴적물의 입자
구성, 산화-환원 상태, 그리고 생물학적 요소로써 미생물의 호흡율, 미생물 및 대
형 저서동물 생체량, 식물 종구성, 개체군 밀도 등을 자연갯벌과 비교함으로써 안
정화와 비안정화를 구분할 수 있다(Haven et al., 1995; Lee et al., 1998).
인공 갯벌이 자연 갯벌과 유사하게 변하는 것을 안정화라고 정의하였을 때 안
정화의 예로는 미국 버지니아에 12년 전 조성된 인공 갯벌을 들 수 있다. 이 인공
갯벌은 어류, 갑각류, 저서성 내서동물, 미생물의 생체량, 식물 군집 유형, 염생 식
물 군락, 식물 줄기 밀도, 염분도, 온도, 용존 산소 농도 등 대부분의 생태항목이
자연갯벌과 유사하였고, 일부 생태 요소 즉, 조류가 먹이로서 저서생물을 소비하
는 정도, 퇴적물내 유기탄소 함량, 염생식물의 밀도 등은 자연 갯벌과 차이를 보
인다고 보고되었다(Haven et al., 2002). 자연 습지와 비교하여 즉 생물의 서식지
로써의 위의 조건이 유사할 때 안정되었다고 할 수 있다(Haven et al., 2002).
1987년 일본 도쿄만에 조성된 인공갯벌에서는 퇴적물내 박테리아 개체군과 유
기물, 세립질 점토의 함량은 자연갯벌과 큰 차이가 있었지만, 퇴적물내 미생물 호
흡율은 주목할만한 큰 차이가 없었다고 보고되었다(Lee et al., 1998). 그 이유는
인공 갯벌 조성 장소를 세립질의 점토가 퇴적되고, 조석활동이 약하며, 퇴적율이
높으며, 경사가 완만한 지역에 조성하였기 때문이다(Lee et al., 1998). 이 연구 결
과에서 자연 갯벌과 유사한 퇴적 및 지형 특성을 가진 인공 갯벌에서는 미생물의
활동이 비슷한 경향을 나타낸다는 것을 알 수 있었다. Middelburg et al.(1996)는
강하구에 위치한 갯벌에서 퇴적물내 미생물 호흡율과 유기물 농도는 관련이 없다
고 보고하였다. 이는 퇴적물내 존재하는 전체 유기물 양에 미생물 분해율이 변하
는 것이 아니라, 퇴적물내 미생물 분해활동에 잘 이용될 수 있는 유기물의 양이
변화 요인이라는 것이다. 결국 미생물 호흡율은 조석활동에 의해 운반될 수 있는,
미생물에 의해 잘 분해될 수 있는, 유기물 공급에 따라 좌우된다고 할 수 있다
(Lee et al., 1998). 즉 인공갯벌의 퇴적작용이나 조석작용에 의해 퇴적물로 공급되
는 유기물의 질에 따라 미생물 분해활동인 SOD값과 상관성을 가진다고 할 수 있
다.
Fig. 4-29은 2005년 낙동강 하구 명지 지역에 실험용 갯벌을 설치한 후 시간이
경과함에 따라 서로 다른 조건의 대조군에서 측정한 SOD를 나타낸 그림이다. 7,
8월은 평균±SD는 각 20.7±6.7mmoleㆍm⁻²ㆍd⁻¹ , 30.1±5.6mmoleㆍm⁻²ㆍd⁻¹ 의 값
으로 각 대조군 간의 격차가 컸으나 10, 11월, 2월은 각각 14.3±5.6mmoleㆍm⁻²ㆍd⁻¹, 3.7±1.9mmoleㆍm⁻²ㆍd⁻¹, -0.5±0.2mmoleㆍm⁻²ㆍd⁻¹ 로 시간이 흐를수록 실험

- 254 -
용 갯벌의 대조구 간 SOD값 차이가 작아짐을 나타낸다. SOD 값으로부터 실험용
갯벌의 조건이 초기 상태에는 각각 틀린 조건이었으나, 시간이 흐름에 따라 여러
가지 물리적 작용에 의해 같은 조건으로 변하였음을 추정해볼 수 있다. 시간이 경
과함에 따라 표층 퇴적물의 조성이 유사해짐이 나타나는데 이는 갯벌에서 일어나
는 퇴적작용에 의해 퇴적물이 공급된 것이다. 결국 실험용 갯벌의 퇴적물의 공급
은 SOD를 변화시킬 수 있는 유기물 공급 조건이 유사하게 변한 것을 의미한다.
이러한 유사한 SOD값을 유발시키는 요인으로는 퇴적물내 유기물의 질이다
(Cramer, 1990; Van Duyl & Kop, 1994 ; Boon et al., 1998). 이러한 결과는 도쿄
만에서 유기물 공급 패턴이 유사할 때 인공 갯벌과 자연 갯벌의 미생물호흡율의
차이가 없었던 결과와 일치한다. 즉 유기물 공급 조건이 유기물 농도, 조고 등의
다른 환경 요인보다 미생물 활동에 중요한 것으로 생각된다. 퇴적물내 유기물의
질을 분석하기 위해서는 유기물의 성분 즉, 지방산과 핵산, 클로로필, 그리고 식물
색소 등을 분석함으로써 평가할 수 있다(Boon et al., 1999).
본 조사에서 각 계절별 SOD 및 표층 입도 조성, 퇴적물내 유기물 함량만을 측
정하고, 유기물의 질을 분석하지 않아 정확한 요인을 말하기는 어렵다. 이외에도
표층 퇴적물의 퇴적작용, 조석의 변화, 온도의 변화 등이 SOD에 영향을 미쳤을
수도 있다. 따라서 실험용 갯벌의 안정화 분석을 위해서는 저서동물의 가입, 식물
군락의 가입, 물리-생지화학적 조건, 생태적 조건 그리고 SOD와 SOD에 영향을
미치는 요인 등이 우선적으로 고려되어야 할 부분임을 시사한다.
라. 탈질소화의 계절변화
(1) 조고에 따른 탈질소화의 변화
본 실험에서는 조고에 따른 탈질소화의 값을 보면 05년 station의 경우 중
LOWER 퇴적물에서 가장 높은 탈질소화(949.2μmoleㆍm⁻²ㆍd⁻¹)를 보였고,
MIDDLE(932.7μmoleㆍm⁻²ㆍd⁻¹), UPPER(438.2μmoleㆍm⁻²ㆍd⁻¹)의 순서였다.
LOWER 실험용 갯벌은 혐기성 유기물 분해 과정인 탈질소화에 이용될 수 있는
NO₃⁻가 수층을 통해 잘 공급이 되며, 가장 긴 침수 시간에 의해 유기물이 공급
또한 좋은 조건이어 호기성 호흡보다는 혐기성 호흡이 우세하여 높은 탈질소화
값이 나타났을 것으로 예상된다. UPPER의 퇴적층은 대기 노출 시간이 가장 길어
표층 퇴적물 내부로 산소가 잘 공급되어 혐기층의 깊이가 깊을 것으로 추측된다.
짧은 침수 시간 때문에 유기물 분해에 이용될 신선한 유기물 공급과 NO3-공급이
LOWER 퇴적물보다는 적어 낮은 탈질소화가 나타났다. 2006년 station의 경우
MIDDLE(881μmoleㆍm⁻²ㆍd⁻¹), LOWER(760.3μmoleㆍm⁻²ㆍd⁻¹), UPPER(591.9μ
moleㆍm⁻²ㆍd⁻¹)의 순서였다. MIDDLE, LOWER 의 탈질소화 값은 거의 비슷한
편인데, MIDDLE, LOWER 실험용 갯벌에서는 혐기성 유기물 분해 과정인 탈질
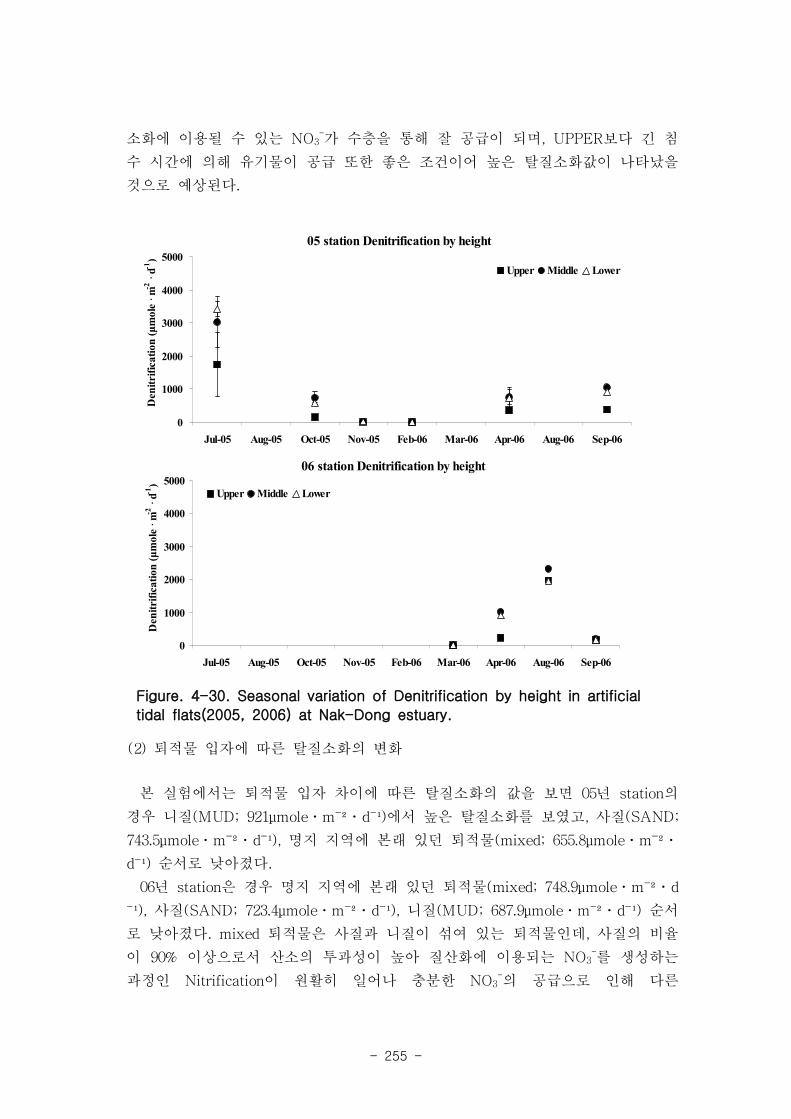
- 255 -
05 station Denitrification by height
0
1000
2000
3000
4000
5000
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Den
itrification (µmole · m
-2 · d
-1)
Upper Middle Lower
06 station Denitrification by height
0
1000
2000
3000
4000
5000
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Den
itrification (µmole · m
-2 · d
-1)
Upper Middle Lower
Figure. 4-30. Seasonal variation of Denitrification by height in artificial
tidal flats(2005, 2006) at Nak-Dong estuary.
소화에 이용될 수 있는 NO3-가 수층을 통해 잘 공급이 되며, UPPER보다 긴 침
수 시간에 의해 유기물이 공급 또한 좋은 조건이어 높은 탈질소화값이 나타났을
것으로 예상된다.
(2) 퇴적물 입자에 따른 탈질소화의 변화
본 실험에서는 퇴적물 입자 차이에 따른 탈질소화의 값을 보면 05년 station의
경우 니질(MUD; 921μmoleㆍm⁻²ㆍd⁻¹)에서 높은 탈질소화를 보였고, 사질(SAND;
743.5μmoleㆍm⁻²ㆍd⁻¹), 명지 지역에 본래 있던 퇴적물(mixed; 655.8μmoleㆍm⁻²ㆍd⁻¹) 순서로 낮아졌다.
06년 station은 경우 명지 지역에 본래 있던 퇴적물(mixed; 748.9μmoleㆍm⁻²ㆍd
⁻¹), 사질(SAND; 723.4μmoleㆍm⁻²ㆍd⁻¹), 니질(MUD; 687.9μmoleㆍm⁻²ㆍd⁻¹) 순서
로 낮아졌다. mixed 퇴적물은 사질과 니질이 섞여 있는 퇴적물인데, 사질의 비율
이 90% 이상으로서 산소의 투과성이 높아 질산화에 이용되는 NO3-를 생성하는
과정인 Nitrification이 원활히 일어나 충분한 NO3-의 공급으로 인해 다른
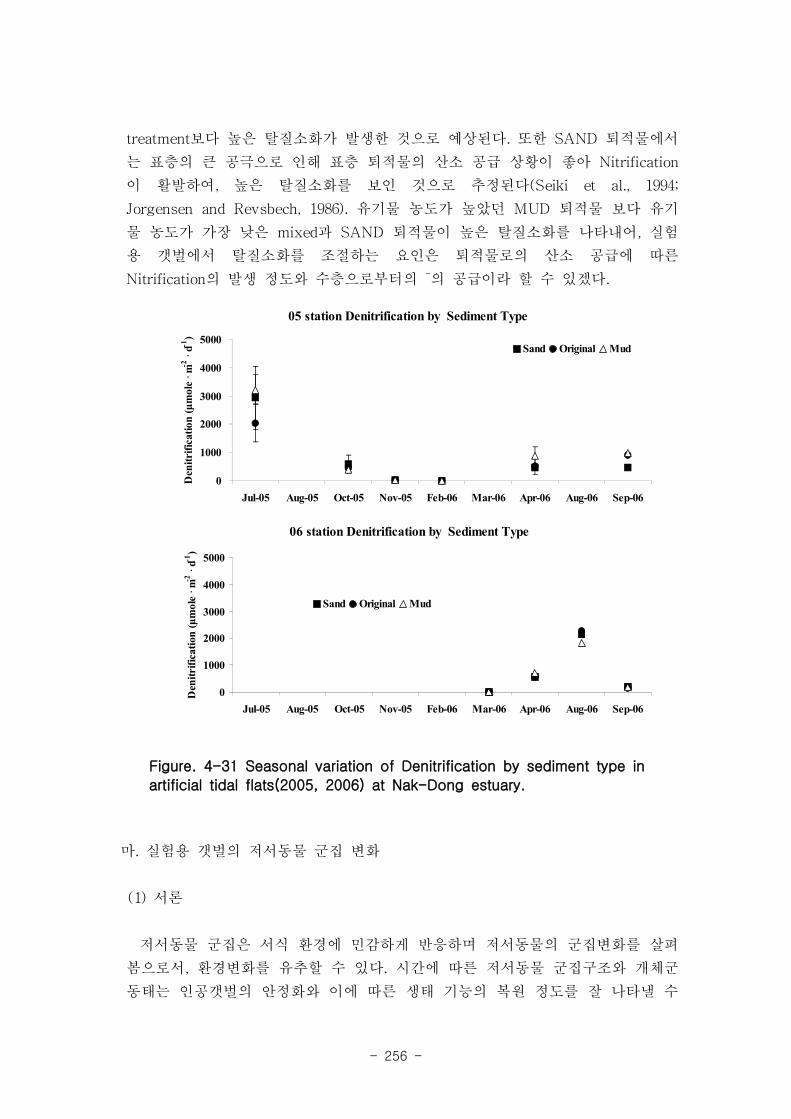
- 256 -
treatment보다 높은 탈질소화가 발생한 것으로 예상된다. 또한 SAND 퇴적물에서
는 표층의 큰 공극으로 인해 표층 퇴적물의 산소 공급 상황이 좋아 Nitrification
이 활발하여, 높은 탈질소화를 보인 것으로 추정된다(Seiki et al., 1994;
Jorgensen and Revsbech, 1986). 유기물 농도가 높았던 MUD 퇴적물 보다 유기
물 농도가 가장 낮은 mixed과 SAND 퇴적물이 높은 탈질소화를 나타내어, 실험
용 갯벌에서 탈질소화를 조절하는 요인은 퇴적물로의 산소 공급에 따른
Nitrification의 발생 정도와 수층으로부터의 -의 공급이라 할 수 있겠다.
05 station Denitrification by Sediment Type
0
1000
2000
3000
4000
5000
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Denitrification (µmole · m
-2 · d-1)
Sand Original Mud
06 station Denitrification by Sediment Type
0
1000
2000
3000
4000
5000
Jul-05 Aug-05 Oct-05 �ov-05 Feb-06 Mar-06 Apr-06 Aug-06 Sep-06
Denitrification (µmole · m
-2 · d-1)
Sand Original Mud
Figure. 4-31 Seasonal variation of Denitrification by sediment type in
artificial tidal flats(2005, 2006) at Nak-Dong estuary.
마. 실험용 갯벌의 저서동물 군집 변화
(1) 서론
저서동물 군집은 서식 환경에 민감하게 반응하며 저서동물의 군집변화를 살펴
봄으로서, 환경변화를 유추할 수 있다. 시간에 따른 저서동물 군집구조와 개체군
동태는 인공갯벌의 안정화와 이에 따른 생태 기능의 복원 정도를 잘 나타낼 수

- 257 -
있다 (Cofer and Niering 1992, Havens et al. 1995). 일반적으로 저서 태계가
교란된 후 안정화 기까지는 6개월에서 3년의 시간이 소요되는 것으로 알려져 있
다 (Bonsdorff 1980, Hilly 1983). 인공갯벌 조성에 따른 저서생물군집의 천이과정
에 대한 연구는 그다지 많지 않으며, 인공갯벌에 얼마의 시간이 흐른 후에 대형
저서동물에 의한 자연적인 군집이 형성될 것인지와 인공갯벌의 환경이 새로운 저
서동물군집에 어떠한 영향을 줄 것인지에 대한 현장 연구는 극소수에 불과하다
(Evans et al. 1998). 본 연구에서는 조고와 퇴적물 형태가 다른 9개의 인공 갯벌
을 조성하여 각각에서 시간에 따른 저서생태계의 변화를 살펴보았다. 저서동물은
생물의 종류가 다양하고 각각의 종이 환경변화에 반응하는 민감도도 다르며 장기
적으로 축적된 환경 변화 혹은 악화에 반응하는 등 환경변화의 지시자로서 알맞
은 특징들을 많이 가지고 있다. 실험용 습지의 저서생물군집 변화를 살펴봄으로서
갯벌 환경의 안정도를 알아 볼 수 있을 것이다. 실험용 습지의 조성은 2005년
2006년,총 2회에 설치되었고 이들을 비교함으로써, 새로 만든 습지와 1년 동안 안
정화된 습지의 차이를 보려 하였다.
(2) 조사내용 및 방법
(가) 현장조사
실험용 습지에서 대형저서동물군집 조사는 2005년 9월에서 2006년 10월까지 총
9번에 걸쳐 이루어 졌다. 2005년에 조성된 실험용 갯벌에서는 2005년 9월과 2006
년 4월, 9월등 총 3회에 걸쳐 조사를 실시하였고, 2006년에 조성된 실험용 갯벌에
서는 2006년 3, 4, 8, 9, 10월(2회) 등 총6회에 걸쳐 조사가 실시되었다. 각각의 실
험용 습지로부터 채집면적이 0.025m2인 주상시료채니기를 이용하여 퇴적물과 저
서동물을 채집하였다. 주상시료채니기를 이용하여 30cm 깊이까지 퇴적물을 파내
었으며, 채집된 퇴적물은 현장에서 1mm 체로 걸러 남은 잔존물만을 비닐에 담아
해수로 중화된 10%의 포르말린으로 고정하였다.
(나) 생물채집 및 동정
고정된 시료는 실험실에서 동물군별로 선별한 후, 가능하한 종 수준까지 동정하
였고, 개체수를 계수하였다. 한편 종 동정을 위하여 김(1973, 1977), 김(1998), 손과
홍(2003), 권 등(1993), 국립수산진흥원(1999), 해양수산부 국립수산진훙원(2001),
Nishimuta(1992, 1995)와 Okutani(1994, 2000)등을 참고하였다. 각 종의 국명은 한
국동물분류학회(1997)를 기준으로 명기하였다.
(3) 연구 결과

- 258 -
(가) 출현 종수 및 개체수
조사 기간 중 출현한 저서동물의 종수는 05년 갯벌이 22~23종으로 06년 갯벌
의 12~22종보다 높은 경향을 보였다. 특히 동시에 실시된 2006년 4월조사에서 05
갯벌에서는 22종이 출현한데 반하여, 06년 갯벌에서는 14종만 출현하였다. 2006년
4월 당시 06년 갯벌은 조성한지 1달 밖에 지나지 않아 안정화되지 못하였음을 시
사한다. 출현 개체수에 있어서도 05년 갯벌의 경우 8400~8800 개체 m-2 로 06년
갯벌의 평균 출현개체수 (~4500 개체 m-2
) 보다 높았다. 06년 갯벌의 경우 갯벌
이 조성된 직후인 2006년 3월 약 2500 로 낮은 출현 개체수를 보이다가 증가하
는 경향을 보인다. 2006년 10월에는 11,000 개체 m-2 로 05년 갯벌보다 높은 수준
을 보여주고 있다.
05년도 갯벌에서 조고 및 퇴적물 타입별 종수와 서식밀도의 차이를 살펴보면,
2006년 4월과 9월 모두 상갯벌(U)에서 낮은 종수를 보였고, 하갯벌 (L)에서 출현
종수가 높은 경향이 나타났다 (Fig. 4-32). 서식밀도에 있어서는 퇴적물 타입별로
차이를 보이고 있는데, 상갯벌 (U)에서 가장 낮은 서식밀도를 보였고, 중 하로 갈
수록 높아지며, 전반적으로는 mix에서 높은 값을 보인다. 05 갯벌은 조성한 후 1
년이 지나, 성숙된 생태계가 이루어 졌을 것이며, 조고에 따른 차이는 노출시간의
차이를 반영한 것으로 볼 수 있으나, 퇴적물 타입의 차이는 앞에서 “ 실헝용 갯벌
표층퇴적물의 입도와 유기물 농도 변화”에서 본 바와 같이 각 treatment 간의 차
이가 적어져 큰 차이를 보지 못한 것으로 생각된다.
2006년 갯벌의 종수는 2006년 3, 4, 8월에는 하갯벌에서 높은 경향을 나타냈으
나 9, 10월에는 갯벌의 높이에 따른 차이는 없어졌다. 특히 9월에는 상 mix에서
가장 높은 종수를 보이고 있다 (Fig. 4-33). 개체수에 있어서는 각 조고간의 차이
가 좀더 뚜렷했는데, 2006년 10월 중갯벌 mix에서 다모류인 Heteromastus
filiformis 가 대량으로 나타난 것을 제외하면, 대체적으로 상갯벌에서 하갯벌로
갈수록 개체수가 증가하는 경향을 보이고 있다.
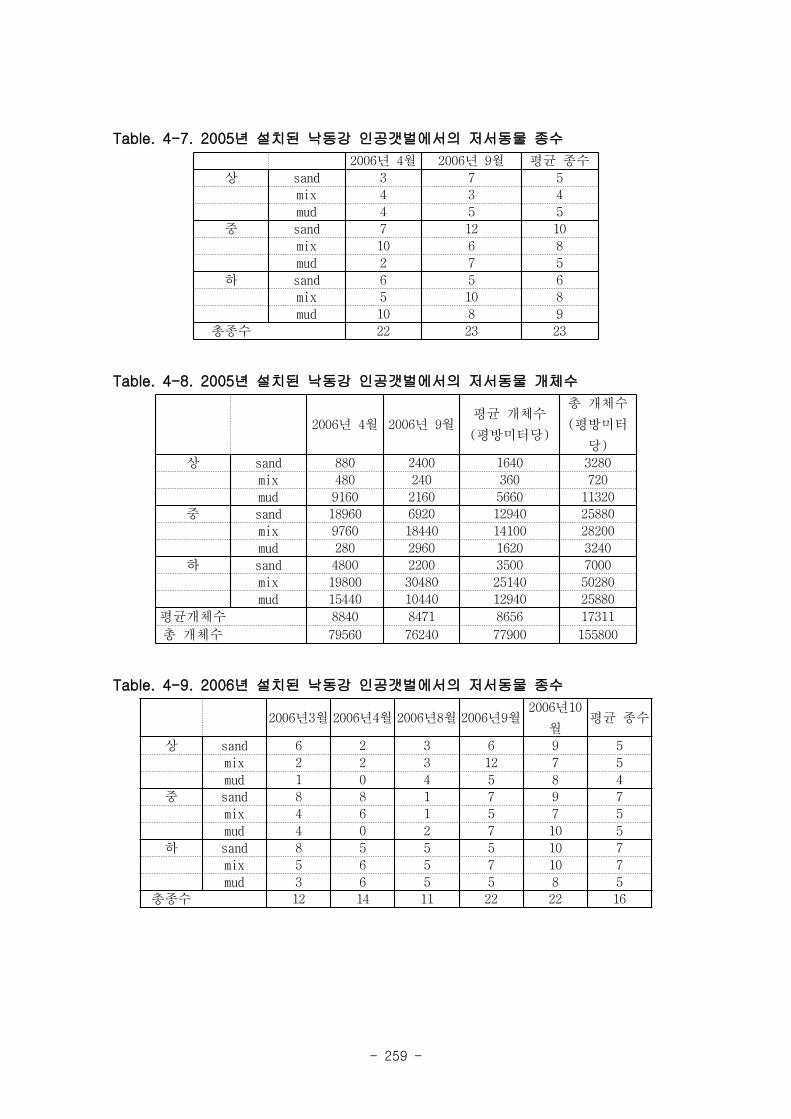
- 259 -
Table. 4-7. 2005년 설치된 낙동강 인공갯벌에서의 저서동물 종수
2006년 4월 2006년 9월 평균 종수
상 sand 3 7 5
mix 4 3 4
mud 4 5 5
중 sand 7 12 10
mix 10 6 8
mud 2 7 5
하 sand 6 5 6
mix 5 10 8
mud 10 8 9
총종수 22 23 23
Table. 4-8. 2005년 설치된 낙동강 인공갯벌에서의 저서동물 개체수
2006년 4월 2006년 9월평균 개체수
(평방미터당)
총 개체수
(평방미터
당)
상 sand 880 2400 1640 3280
mix 480 240 360 720
mud 9160 2160 5660 11320
중 sand 18960 6920 12940 25880
mix 9760 18440 14100 28200
mud 280 2960 1620 3240
하 sand 4800 2200 3500 7000
mix 19800 30480 25140 50280
mud 15440 10440 12940 25880
평균개체수 8840 8471 8656 17311
총 개체수 79560 76240 77900 155800
Table. 4-9. 2006년 설치된 낙동강 인공갯벌에서의 저서동물 종수
2006년3월 2006년4월 2006년8월 2006년9월2006년10
월평균 종수
상 sand 6 2 3 6 9 5
mix 2 2 3 12 7 5
mud 1 0 4 5 8 4
중 sand 8 8 1 7 9 7
mix 4 6 1 5 7 5
mud 4 0 2 7 10 5
하 sand 8 5 5 5 10 7
mix 5 6 5 7 10 7
mud 3 6 5 5 8 5
총종수 12 14 11 22 22 16
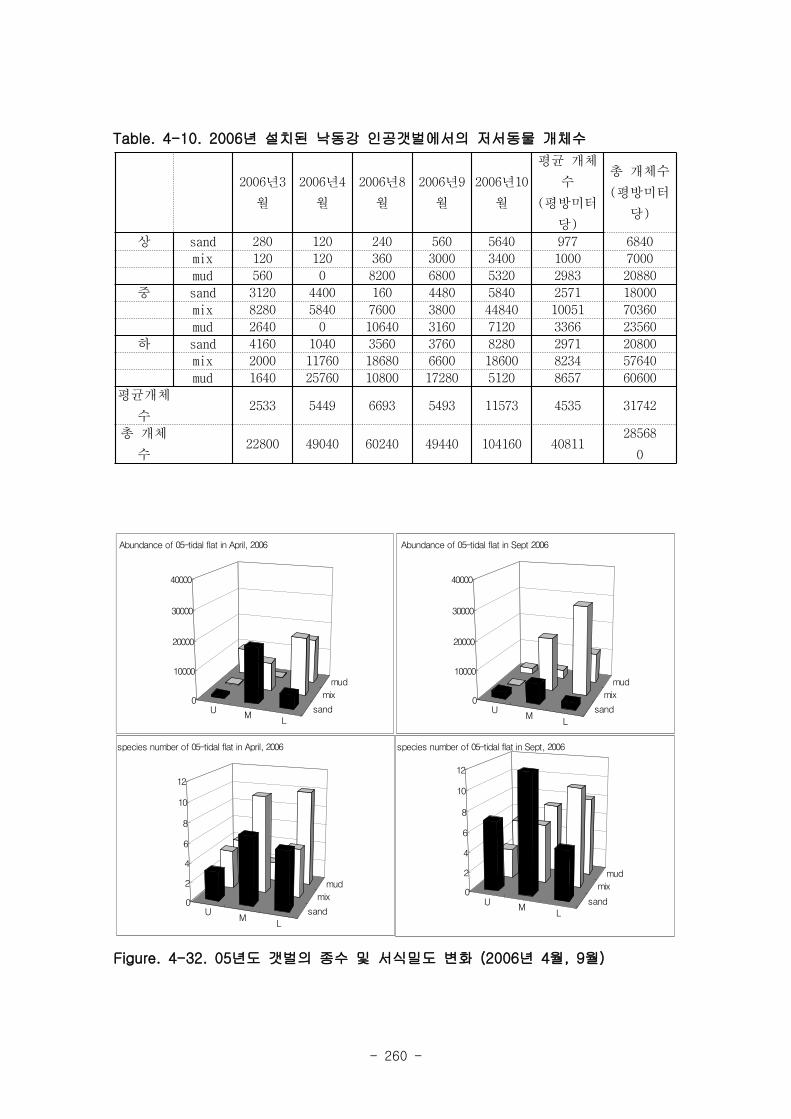
- 260 -
Table. 4-10. 2006년 설치된 낙동강 인공갯벌에서의 저서동물 개체수
2006년3
월
2006년4
월
2006년8
월
2006년9
월
2006년10
월
평균 개체
수
(평방미터
당)
총 개체수
(평방미터
당)
상 sand 280 120 240 560 5640 977 6840
mix 120 120 360 3000 3400 1000 7000
mud 560 0 8200 6800 5320 2983 20880
중 sand 3120 4400 160 4480 5840 2571 18000
mix 8280 5840 7600 3800 44840 10051 70360
mud 2640 0 10640 3160 7120 3366 23560
하 sand 4160 1040 3560 3760 8280 2971 20800
mix 2000 11760 18680 6600 18600 8234 57640
mud 1640 25760 10800 17280 5120 8657 60600
평균개체
수2533 5449 6693 5493 11573 4535 31742
총 개체
수22800 49040 60240 49440 104160 40811
28568
0
UM
Lsand
mix
mud
0
10000
20000
30000
40000
Abundance of 05-tidal flat in April, 2006
UM
L
sand
mix
mud
0
10000
20000
30000
40000
Abundance of 05-tidal flat in Sept 2006
UM
L
sand
mix
mud
0
2
4
6
8
10
12
species number of 05-tidal flat in April, 2006
UM
Lsand
mix
mud
0
2
4
6
8
10
12
species number of 05-tidal flat in Sept, 2006
Figure. 4-32. 05년도 갯벌의 종수 및 서식밀도 변화 (2006년 4월, 9월)
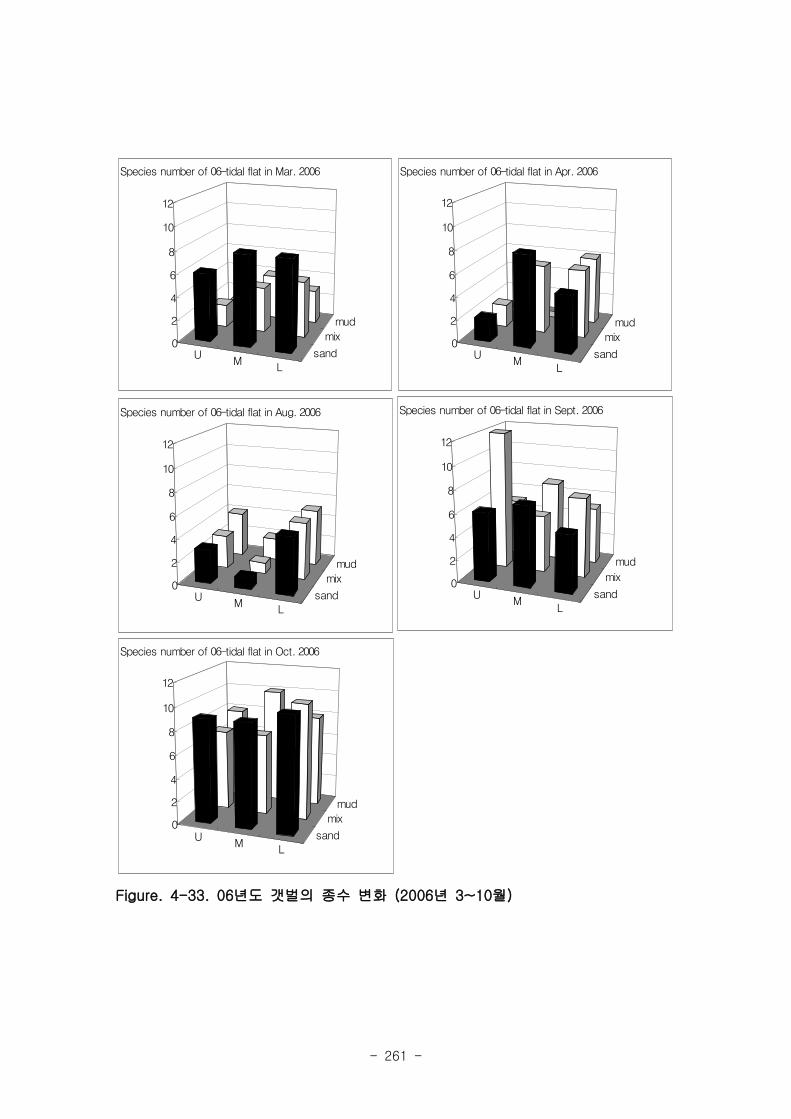
- 261 -
UM
Lsand
mix
mud
0
2
4
6
8
10
12
Species number of 06-tidal flat in Mar. 2006
UM
Lsand
mix
mud
0
2
4
6
8
10
12
Species number of 06-tidal flat in Apr. 2006
UM
Lsand
mix
mud
0
2
4
6
8
10
12
Species number of 06-tidal flat in Aug. 2006
UM
Lsand
mix
mud
0
2
4
6
8
10
12
Species number of 06-tidal flat in Sept. 2006
UM
Lsand
mix
mud
0
2
4
6
8
10
12
Species number of 06-tidal flat in Oct. 2006
Figure. 4-33. 06년도 갯벌의 종수 변화 (2006년 3~10월)
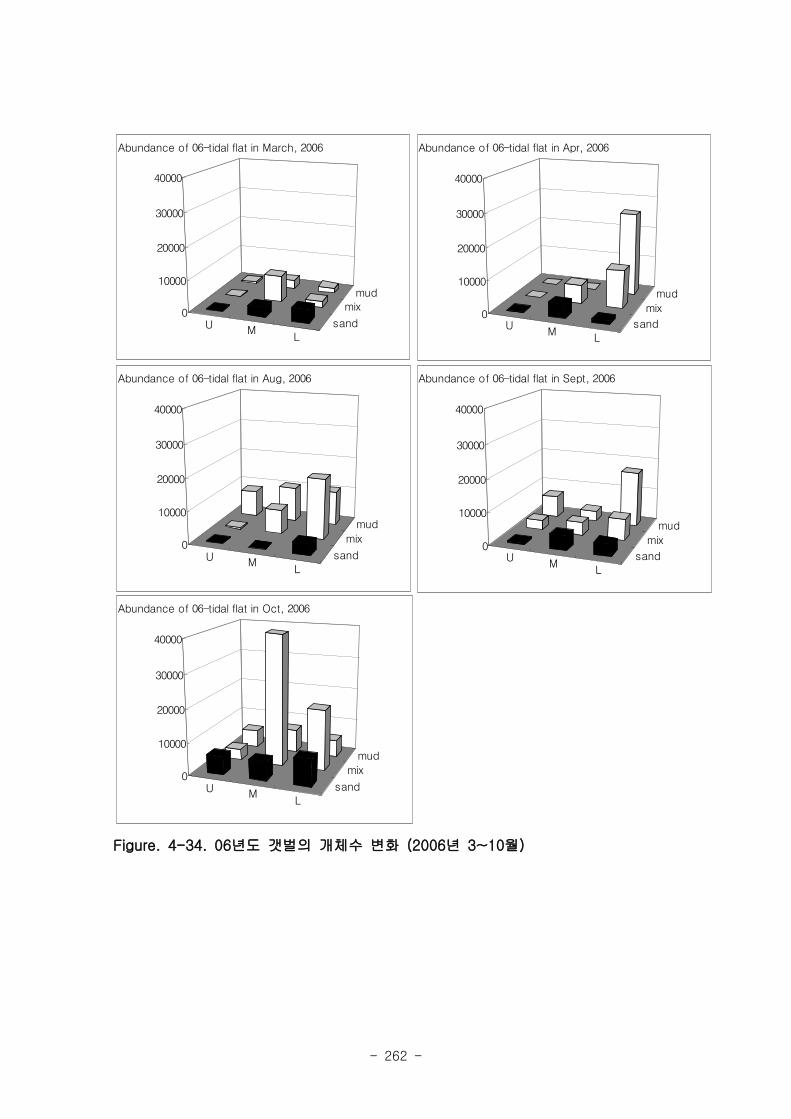
- 262 -
UM
Lsand
mixmud
0
10000
20000
30000
40000
Abundance of 06-tidal flat in March, 2006
UM
Lsand
mix
mud
0
10000
20000
30000
40000
Abundance of 06-tidal flat in Apr, 2006
UM
Lsand
mix
mud
0
10000
20000
30000
40000
Abundance of 06-tidal flat in Aug, 2006
UM
Lsand
mix
mud
0
10000
20000
30000
40000
Abundance of 06-tidal flat in Sept, 2006
UM
Lsand
mix
mud
0
10000
20000
30000
40000
Abundance of 06-tidal flat in Oct, 2006
Figure. 4-34. 06년도 갯벌의 개체수 변화 (2006년 3~10월)
- 263 -
UM L
sand
mixmud
0
0.5
1
1.5
2
2.5
Divers ity of 05-tidal flat in April, 2006
UM L
sand
mixmud
0
0.5
1
1.5
2
2.5
Divers ity of 05-tidal flat in Sept, 2006
UM L
sand
mixmud
0
0.5
1
1.5
2
2.5
Divers ity of 06-tidal flat in Mar., 2006
UM L
sand
mixmud
0
0.5
1
1.5
2
2.5
I. deschampsi of 06-tidal flat in Apr., 2006
UM L
sand
mixmud
0
0.5
1
1.5
2
2.5
Divers ity of 06-tidal flat in Aug., 2006
UM L
sand
mixmud
0
0.5
1
1.5
2
2.5
Divers ity of 06-tidal flat in Sept., 2006
UM
L
sand
mixmud
0
0.5
1
1.5
2
2.5
Divers ity of 06-tidal flat in Oct., 2006
Figure. 4-35.

- 264 -
(나) 우점종
조사 기간 동안 낙동강 실험용 갯벌에서 출현한 저서동물 중에서 개체수를 기
준으로 상위 15종을 살펴보면 절지동물문이 6종, 다모류와 연체동물문이 각 4종
그리고 기타동물문이 한 종이 포함되었다. 다모류인 Heteromastus filiformis가 전
체 출현 개체수의 56.1%로 가장 많이 나타났고 연체동물문의 기수우렁이
(Assiminea japonica), 빈모류인 Oligichaeta, 다모류의 Prionospio japonica, 절지
동물문의 Corophium sp., 다모류의 Neanthes japonica, unid, 연체동물문의 회색
구슬우렁이(Cryptonatica clausa), 절지동물문의 I lyoplax pusilla, Insectra larvae,
I lyoplax deschampsi, Scopomera globosa, 연체동물문의 Laternula marilina,
Corbicula japonica, 절지동물문의 Isopoda spp. 등이 나타났다. 상위 15종이 차
지하는 개체수는 전체의 97.8%를 점유하였다. 출현 빈도에 있어서도
Heteromastus filiformis가 가장 높아 낙동강 갯벌에서 흔히 나타나는 개체수임을
알 수 있다.
순위 분류군 종명 총합계 점유율(%) 출현빈도
1 Apol Heteromastus filiformis 313000 56.1 59
2 MGs Assiminea japonica 64240 11.5 32
3 other Oligochaeta 59160 10.6 29
4 Apol Prionospio japonica 57920 10.4 38
5 Cam Corophium spp. 17680 3.2 45
6 Apol #eanthes japonica 8560 1.5 35
7 Apol un id I 6360 1.1 9
8 MGs Cryptonatica clausa 3440 0.6 7
9 CDB Ilyoplax pusilla 3040 0.5 14
10 C Insecta larvae 2720 0.5 22
11 CDB Ilyoplax deschampsi 2360 0.4 14
12 CDB Scopimera globosa 2360 0.4 13
13 MBi Laternula marilina 2240 0.4 18
14 MGs Corbicula japonica 1640 0.3 16
15 Cis Isopoda spp. 1360 0.2 10
Table. 4-11. 낙동강 실험용 갯벌에서 우점한 저서동물
각 조사 시기에 출현한 종을 개체수를 이용하여 조사시기별로 나타내었다. 각
조사 기간 동안 나타는 우점종을 보면, 다모류인 Heteromastus filiformis가 거의
모든 조사 기간 동안 50%이상의 우점도를 나타내고 있었다. 다모류인 Prionospio
japonica, 절지동물문인 Corophium spp.등도 높은 시기에 따라 높은 출현 빈도
를 보이고 있었다.
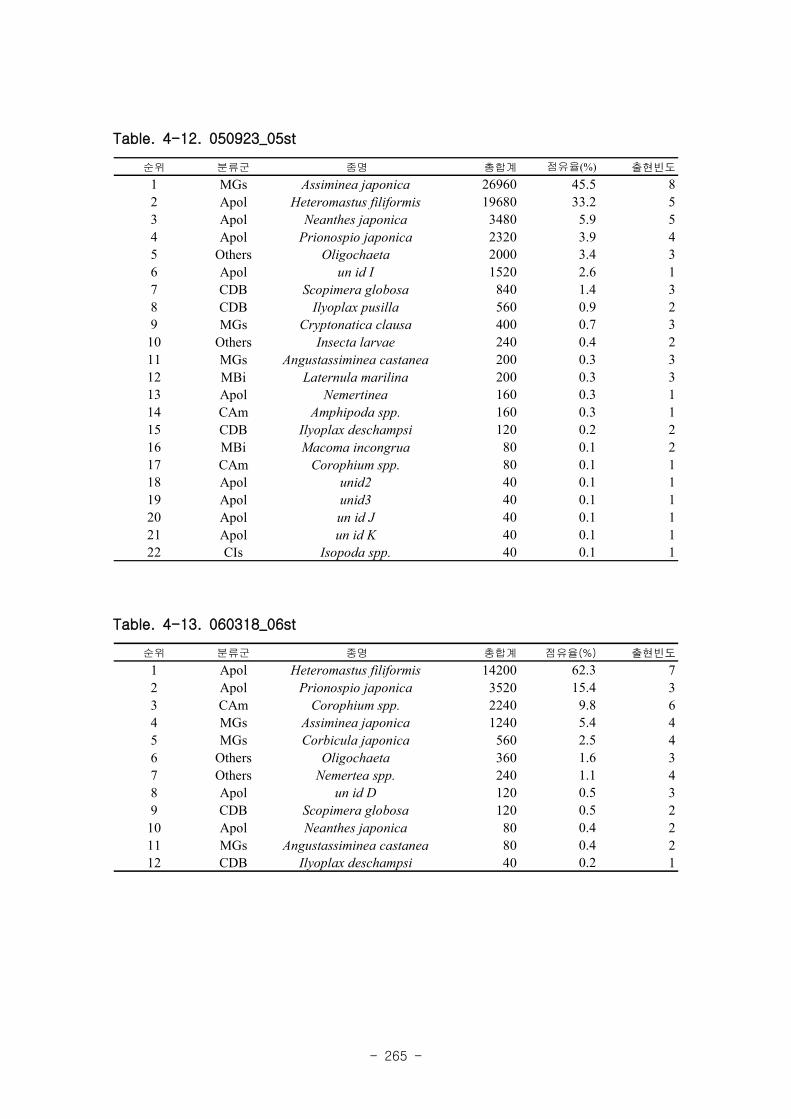
- 265 -
순위 분류군 종명 총합계 점유율(%) 출현빈도
1 MGs Assiminea japonica 26960 45.5 8
2 Apol Heteromastus filiformis 19680 33.2 5
3 Apol #eanthes japonica 3480 5.9 5
4 Apol Prionospio japonica 2320 3.9 4
5 Others Oligochaeta 2000 3.4 3
6 Apol un id I 1520 2.6 1
7 CDB Scopimera globosa 840 1.4 3
8 CDB Ilyoplax pusilla 560 0.9 2
9 MGs Cryptonatica clausa 400 0.7 3
10 Others Insecta larvae 240 0.4 2
11 MGs Angustassiminea castanea 200 0.3 3
12 MBi Laternula marilina 200 0.3 3
13 Apol #emertinea 160 0.3 1
14 CAm Amphipoda spp. 160 0.3 1
15 CDB Ilyoplax deschampsi 120 0.2 2
16 MBi Macoma incongrua 80 0.1 2
17 CAm Corophium spp. 80 0.1 1
18 Apol unid2 40 0.1 1
19 Apol unid3 40 0.1 1
20 Apol un id J 40 0.1 1
21 Apol un id K 40 0.1 1
22 CIs Isopoda spp. 40 0.1 1
Table. 4-12. 050923_05st
순위 분류군 종명 총합계 점유율(%) 출현빈도
1 Apol Heteromastus filiformis 14200 62.3 7
2 Apol Prionospio japonica 3520 15.4 3
3 CAm Corophium spp. 2240 9.8 6
4 MGs Assiminea japonica 1240 5.4 4
5 MGs Corbicula japonica 560 2.5 4
6 Others Oligochaeta 360 1.6 3
7 Others #emertea spp. 240 1.1 4
8 Apol un id D 120 0.5 3
9 CDB Scopimera globosa 120 0.5 2
10 Apol #eanthes japonica 80 0.4 2
11 MGs Angustassiminea castanea 80 0.4 2
12 CDB Ilyoplax deschampsi 40 0.2 1
Table. 4-13. 060318_06st
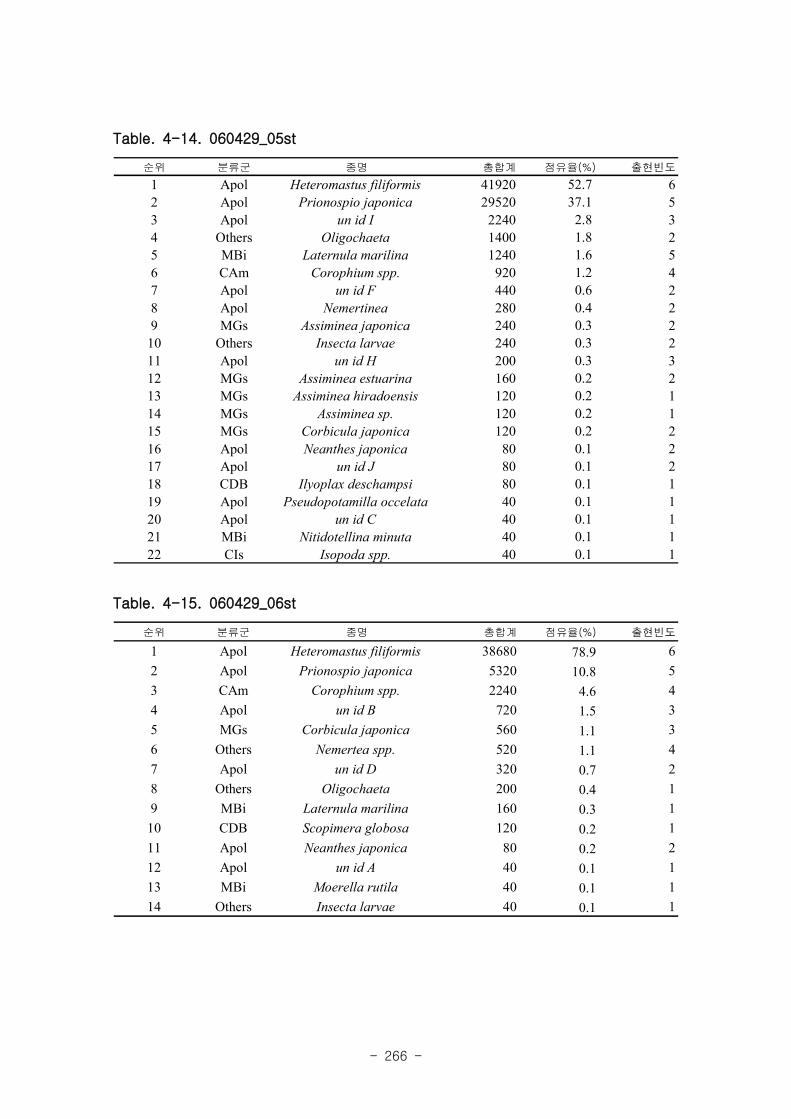
- 266 -
순위 분류군 종명 총합계 점유율(%) 출현빈도
1 Apol Heteromastus filiformis 41920 52.7 6
2 Apol Prionospio japonica 29520 37.1 5
3 Apol un id I 2240 2.8 3
4 Others Oligochaeta 1400 1.8 2
5 MBi Laternula marilina 1240 1.6 5
6 CAm Corophium spp. 920 1.2 4
7 Apol un id F 440 0.6 2
8 Apol #emertinea 280 0.4 2
9 MGs Assiminea japonica 240 0.3 2
10 Others Insecta larvae 240 0.3 2
11 Apol un id H 200 0.3 3
12 MGs Assiminea estuarina 160 0.2 2
13 MGs Assiminea hiradoensis 120 0.2 1
14 MGs Assiminea sp. 120 0.2 1
15 MGs Corbicula japonica 120 0.2 2
16 Apol #eanthes japonica 80 0.1 2
17 Apol un id J 80 0.1 2
18 CDB Ilyoplax deschampsi 80 0.1 1
19 Apol Pseudopotamilla occelata 40 0.1 1
20 Apol un id C 40 0.1 1
21 MBi #itidotellina minuta 40 0.1 1
22 CIs Isopoda spp. 40 0.1 1
Table. 4-14. 060429_05st
순위 분류군 종명 총합계 점유율(%) 출현빈도
1 Apol Heteromastus filiformis 38680 78.9 6
2 Apol Prionospio japonica 5320 10.8 5
3 CAm Corophium spp. 2240 4.6 4
4 Apol un id B 720 1.5 3
5 MGs Corbicula japonica 560 1.1 3
6 Others #emertea spp. 520 1.1 4
7 Apol un id D 320 0.7 2
8 Others Oligochaeta 200 0.4 1
9 MBi Laternula marilina 160 0.3 1
10 CDB Scopimera globosa 120 0.2 1
11 Apol #eanthes japonica 80 0.2 2
12 Apol un id A 40 0.1 1
13 MBi Moerella rutila 40 0.1 1
14 Others Insecta larvae 40 0.1 1
Table. 4-15. 060429_06st
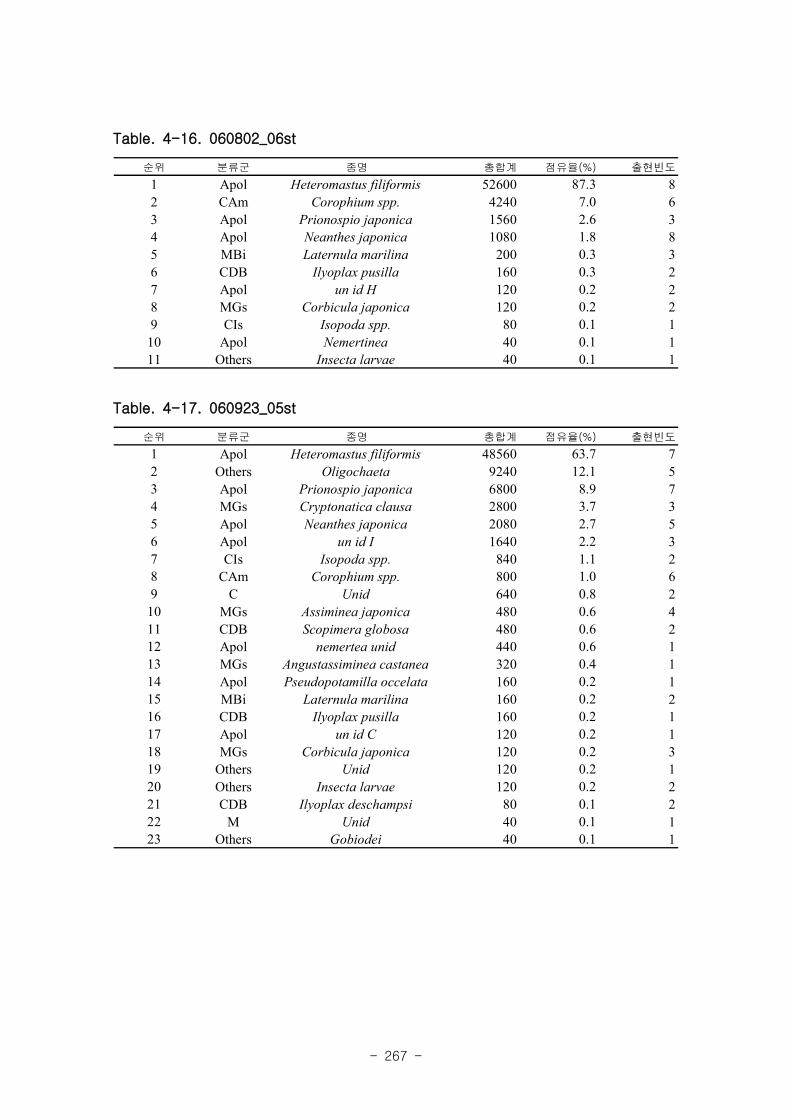
- 267 -
순위 분류군 종명 총합계 점유율(%) 출현빈도
1 Apol Heteromastus filiformis 52600 87.3 8
2 CAm Corophium spp. 4240 7.0 6
3 Apol Prionospio japonica 1560 2.6 3
4 Apol #eanthes japonica 1080 1.8 8
5 MBi Laternula marilina 200 0.3 3
6 CDB Ilyoplax pusilla 160 0.3 2
7 Apol un id H 120 0.2 2
8 MGs Corbicula japonica 120 0.2 2
9 CIs Isopoda spp. 80 0.1 1
10 Apol #emertinea 40 0.1 1
11 Others Insecta larvae 40 0.1 1
Table. 4-16. 060802_06st
순위 분류군 종명 총합계 점유율(%) 출현빈도
1 Apol Heteromastus filiformis 48560 63.7 7
2 Others Oligochaeta 9240 12.1 5
3 Apol Prionospio japonica 6800 8.9 7
4 MGs Cryptonatica clausa 2800 3.7 3
5 Apol #eanthes japonica 2080 2.7 5
6 Apol un id I 1640 2.2 3
7 CIs Isopoda spp. 840 1.1 2
8 CAm Corophium spp. 800 1.0 6
9 C Unid 640 0.8 2
10 MGs Assiminea japonica 480 0.6 4
11 CDB Scopimera globosa 480 0.6 2
12 Apol nemertea unid 440 0.6 1
13 MGs Angustassiminea castanea 320 0.4 1
14 Apol Pseudopotamilla occelata 160 0.2 1
15 MBi Laternula marilina 160 0.2 2
16 CDB Ilyoplax pusilla 160 0.2 1
17 Apol un id C 120 0.2 1
18 MGs Corbicula japonica 120 0.2 3
19 Others Unid 120 0.2 1
20 Others Insecta larvae 120 0.2 2
21 CDB Ilyoplax deschampsi 80 0.1 2
22 M Unid 40 0.1 1
23 Others Gobiodei 40 0.1 1
Table. 4-17. 060923_05st
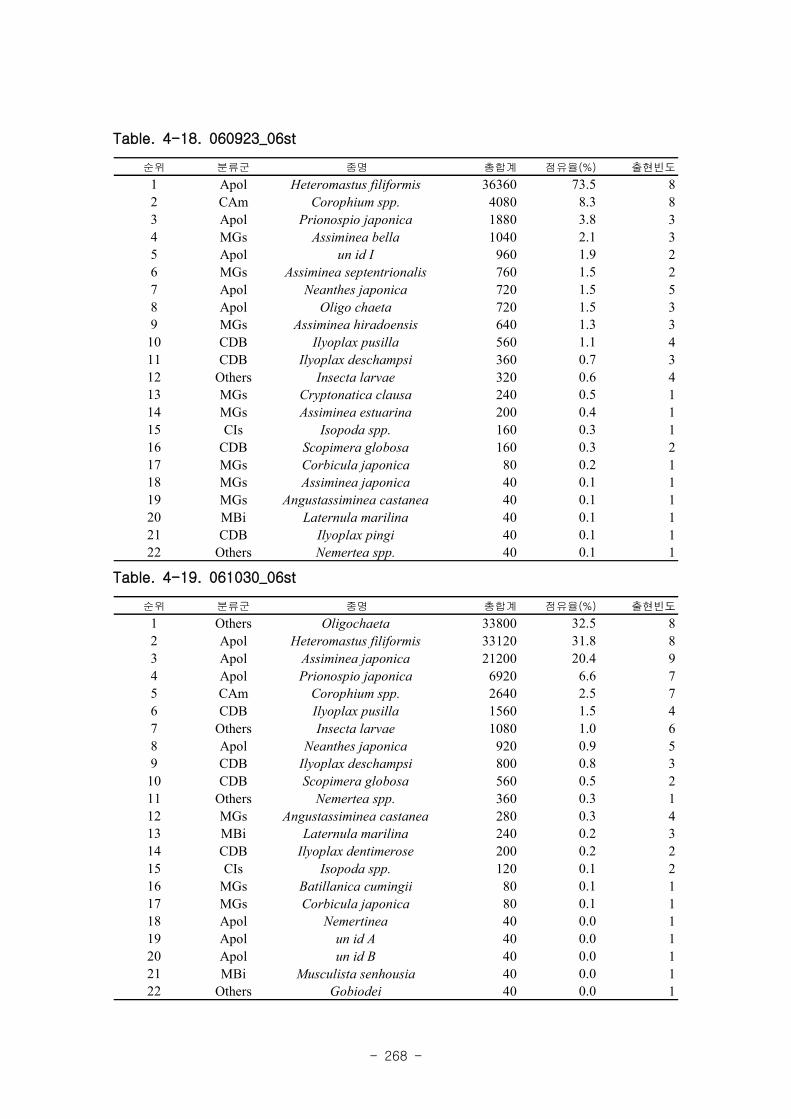
- 268 -
순위 분류군 종명 총합계 점유율(%) 출현빈도
1 Apol Heteromastus filiformis 36360 73.5 8
2 CAm Corophium spp. 4080 8.3 8
3 Apol Prionospio japonica 1880 3.8 3
4 MGs Assiminea bella 1040 2.1 3
5 Apol un id I 960 1.9 2
6 MGs Assiminea septentrionalis 760 1.5 2
7 Apol #eanthes japonica 720 1.5 5
8 Apol Oligo chaeta 720 1.5 3
9 MGs Assiminea hiradoensis 640 1.3 3
10 CDB Ilyoplax pusilla 560 1.1 4
11 CDB Ilyoplax deschampsi 360 0.7 3
12 Others Insecta larvae 320 0.6 4
13 MGs Cryptonatica clausa 240 0.5 1
14 MGs Assiminea estuarina 200 0.4 1
15 CIs Isopoda spp. 160 0.3 1
16 CDB Scopimera globosa 160 0.3 2
17 MGs Corbicula japonica 80 0.2 1
18 MGs Assiminea japonica 40 0.1 1
19 MGs Angustassiminea castanea 40 0.1 1
20 MBi Laternula marilina 40 0.1 1
21 CDB Ilyoplax pingi 40 0.1 1
22 Others #emertea spp. 40 0.1 1
Table. 4-18. 060923_06st
순위 분류군 종명 총합계 점유율(%) 출현빈도
1 Others Oligochaeta 33800 32.5 8
2 Apol Heteromastus filiformis 33120 31.8 8
3 Apol Assiminea japonica 21200 20.4 9
4 Apol Prionospio japonica 6920 6.6 7
5 CAm Corophium spp. 2640 2.5 7
6 CDB Ilyoplax pusilla 1560 1.5 4
7 Others Insecta larvae 1080 1.0 6
8 Apol #eanthes japonica 920 0.9 5
9 CDB Ilyoplax deschampsi 800 0.8 3
10 CDB Scopimera globosa 560 0.5 2
11 Others #emertea spp. 360 0.3 1
12 MGs Angustassiminea castanea 280 0.3 4
13 MBi Laternula marilina 240 0.2 3
14 CDB Ilyoplax dentimerose 200 0.2 2
15 CIs Isopoda spp. 120 0.1 2
16 MGs Batillanica cumingii 80 0.1 1
17 MGs Corbicula japonica 80 0.1 1
18 Apol #emertinea 40 0.0 1
19 Apol un id A 40 0.0 1
20 Apol un id B 40 0.0 1
21 MBi Musculista senhousia 40 0.0 1
22 Others Gobiodei 40 0.0 1
Table. 4-19. 061030_06st
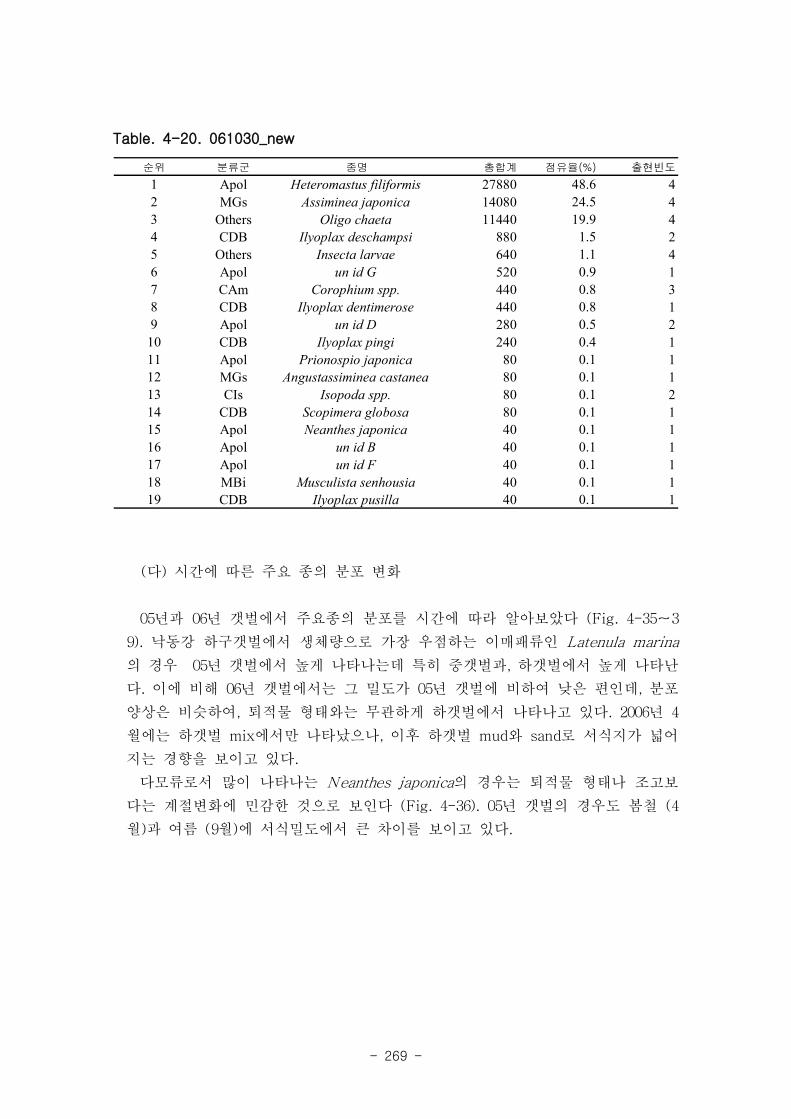
- 269 -
순위 분류군 종명 총합계 점유율(%) 출현빈도
1 Apol Heteromastus filiformis 27880 48.6 4
2 MGs Assiminea japonica 14080 24.5 4
3 Others Oligo chaeta 11440 19.9 4
4 CDB Ilyoplax deschampsi 880 1.5 2
5 Others Insecta larvae 640 1.1 4
6 Apol un id G 520 0.9 1
7 CAm Corophium spp. 440 0.8 3
8 CDB Ilyoplax dentimerose 440 0.8 1
9 Apol un id D 280 0.5 2
10 CDB Ilyoplax pingi 240 0.4 1
11 Apol Prionospio japonica 80 0.1 1
12 MGs Angustassiminea castanea 80 0.1 1
13 CIs Isopoda spp. 80 0.1 2
14 CDB Scopimera globosa 80 0.1 1
15 Apol #eanthes japonica 40 0.1 1
16 Apol un id B 40 0.1 1
17 Apol un id F 40 0.1 1
18 MBi Musculista senhousia 40 0.1 1
19 CDB Ilyoplax pusilla 40 0.1 1
Table. 4-20. 061030_new
(다) 시간에 따른 주요 종의 분포 변화
05년과 06년 갯벌에서 주요종의 분포를 시간에 따라 알아보았다 (Fig. 4-35~3
9). 낙동강 하구갯벌에서 생체량으로 가장 우점하는 이매패류인 Latenula marina
의 경우 05년 갯벌에서 높게 나타나는데 특히 중갯벌과, 하갯벌에서 높게 나타난
다. 이에 비해 06년 갯벌에서는 그 밀도가 05년 갯벌에 비하여 낮은 편인데, 분포
양상은 비슷하여, 퇴적물 형태와는 무관하게 하갯벌에서 나타나고 있다. 2006년 4
월에는 하갯벌 mix에서만 나타났으나, 이후 하갯벌 mud와 sand로 서식지가 넓어
지는 경향을 보이고 있다.
다모류로서 많이 나타나는 Neanthes japonica의 경우는 퇴적물 형태나 조고보
다는 계절변화에 민감한 것으로 보인다 (Fig. 4-36). 05년 갯벌의 경우도 봄철 (4
월)과 여름 (9월)에 서식밀도에서 큰 차이를 보이고 있다.

- 270 -
UM
L
sand
mix
mud
0
50
100
150
200
250
300
350
400
L. marina of 05-tidal flat in April, 2006
UM
L
sand
mix
mud
0
50
100
150
200
250
300
350
400
L. marina of 05-tidal flat in sept, 2006
UM
Lsand
mix
mud
0
50
100
150
200
250
300
350
400
L. marina of 06-tidal flat in Apr., 2006
UM
L
sand
mix
mud
0
50
100
150
200
250
300
350
400
L. marina of 06-tidal flat in Aug., 2006
UM
L
sand
mix
mud
0
50
100
150
200
250
300
350
400
L. marina of 06-tidal flat in Sept., 2006
UM
Lsand
mix
mud
0
50
100
150
200
250
300
350
400
L. marina of 06-tidal flat in Sept., 2006
Figure. 4-35. 실험용 갯벌에서 시간에 따른 Latenula marina 의 분포 변화.
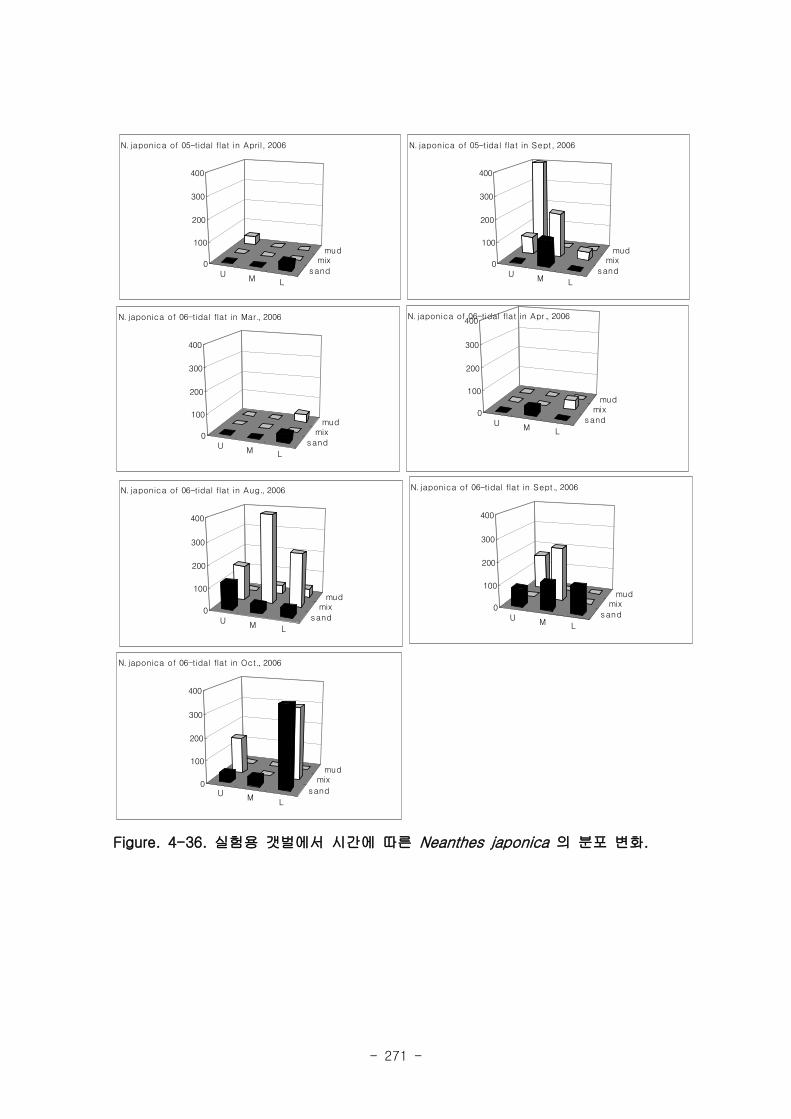
- 271 -
UM L
sand
mixmud
0
100
200
300
400
N. japonica of 05-tidal flat in April, 2006
UM L
sand
mixmud
0
100
200
300
400
N. japonica of 05-tidal flat in Sept, 2006
UM L
sand
mixmud
0
100
200
300
400
N. japonica of 06-tidal flat in Mar., 2006
UM L
sand
mixmud
0
100
200
300
400N. japonica of 06-tidal flat in Apr., 2006
UM L
sand
mixmud
0
100
200
300
400
N. japonica of 06-tidal flat in Aug., 2006
UM L
sand
mixmud
0
100
200
300
400
N. japonica of 06-tidal flat in Sept., 2006
UM
L
sand
mixmud
0
100
200
300
400
N. japonica of 06-tidal flat in Oct., 2006
Figure. 4-36. 실험용 갯벌에서 시간에 따른 Neanthes japonica 의 분포 변화.
- 272 -
UM L
sand
mixmud
0
100
200
300
400
C. japonica of 05-tidal flat in April, 2006
UM L
sand
mixmud
0
100
200
300
400
C. japonica of 05-tidal flat in Sept, 2006
UM L
sand
mixmud
0
100
200
300
400
C. japonica of 06-tidal flat in Mar., 2006
UM L
sand
mixmud
0
100
200
300
400
C. japonica of 06-tidal flat in Apr., 2006
UM L
sand
mixmud
0
100
200
300
400
C. japonica of 06-tidal flat in Aug., 2006
UM L
sand
mixmud
0
100
200
300
400
C. japonicaof 06-tidal flat in Sept., 2006
UM
L
sand
mixmud
0
100
200
300
400
C. japonica of 06-tidal flat in Oct., 2006
Figure. 4-37. 실험용 갯벌에서 시간에 따른 Corbicula japonica 의 분포 변화.

- 273 -
UM L
sand
mixmud
0
5000
10000
15000
H. filiformis of 05-tidal flat in April, 2006
UM L
sand
mixmud
0
5000
10000
15000
H. filiformisof 05-tidal flat in Sept, 2006
UM L
sand
mixmud
0
5000
10000
15000
H. filiformisof 06-tidal flat in Mar., 2006
UM L
sand
mixmud
0
5000
10000
15000
H. filiformis of 06-tidal flat in Apr., 2006
UM L
sand
mixmud
0
5000
10000
15000
H. filiformis of 06-tidal flat in Aug., 2006
UM L
sand
mixmud
0
5000
10000
15000
H. filiformis of 06-tidal flat in Sept., 2006
UM
L
sand
mixmud
0
5000
10000
15000
H. filiformis of 06-tidal flat in Oct., 2006
Figure. 4-38. 실험용 갯벌에서 시간에 따른 Heteromastus filiformis의 분포 변화.
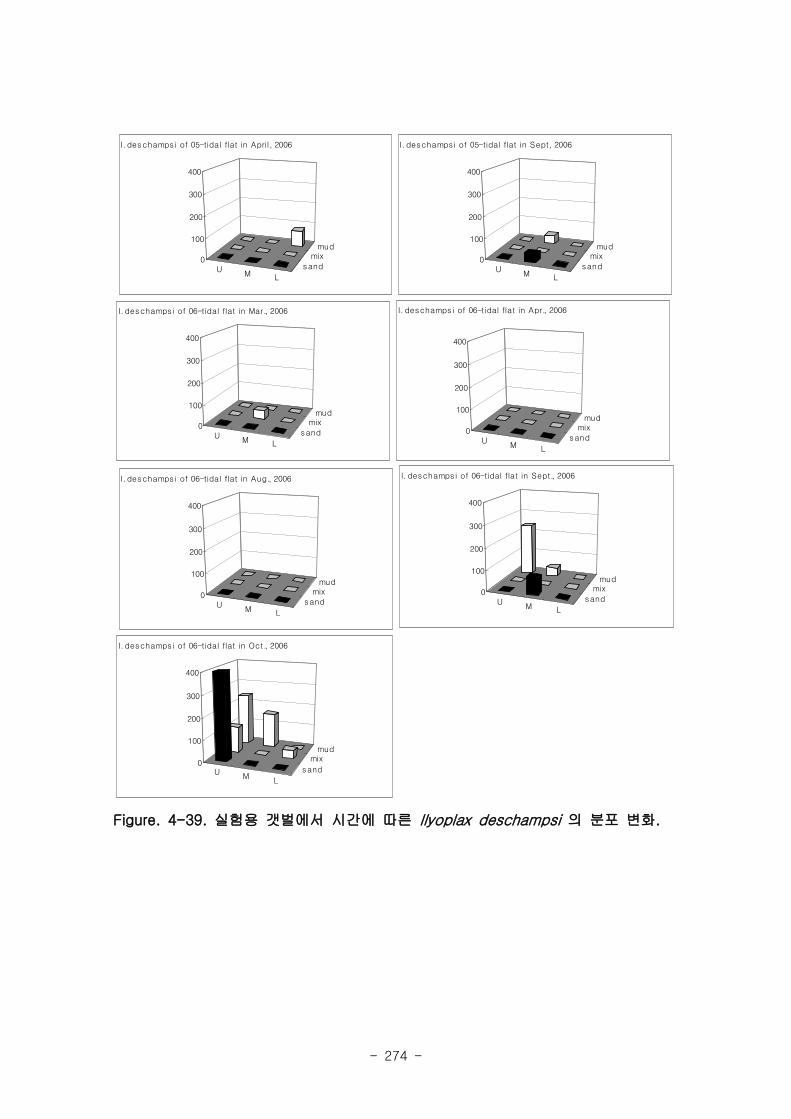
- 274 -
UM L
sand
mixmud
0
100
200
300
400
I. deschamps i of 05-tidal flat in April, 2006
UM L
sand
mixmud
0
100
200
300
400
I. deschampsi of 05-tidal flat in Sept, 2006
UM L
sand
mixmud
0
100
200
300
400
I. deschampsi of 06-tidal flat in Mar., 2006
UM L
sand
mixmud
0
100
200
300
400
I. deschampsi of 06-tidal flat in Apr., 2006
UM L
sand
mixmud
0
100
200
300
400
I. deschamps i of 06-tidal flat in Aug., 2006
UM L
sand
mixmud
0
100
200
300
400
I. deschampsi of 06-tidal flat in Sept., 2006
UM
L
sand
mixmud
0
100
200
300
400
I. deschampsi of 06-tidal flat in Oct., 2006
Figure. 4-39. 실험용 갯벌에서 시간에 따른 Ilyoplax deschampsi 의 분포 변화.

- 275 -
제 5절 낙동강 하구의 자정 능력
1. 인공 갯벌의 유기물 분해와 탈질소화를 통한 유기물 제거
가. 유기물 분해와 탈질소화 측정을 통한 자정능력의 정량화
갯벌은 육지와 해양의 중간에 위치하고 있어 육상기원물질이 해양으로 유입되
는 통로 역할을 한다. 단순한 통로 역할 이외에 자연정화조로서 자정능력을 갖고
있을 것으로 여겨지고 있으며, 개벌의 경제적 가치를 평가하는 기준으로 자주 인
용되고 있다. 현재 논의되고 있는 개벌의 자연 정화 능력의 핵심은 개벌로 유입된
유기물이 미생물에 의해 분해되는, 갯벌의 유기물분해 능력이다. 갯벌은 노출과
침수로 인한 수층과 표면 퇴적물의 교란과 저서동물의 활동에 의한 퇴적층의 생
교란(bioturbation) 등으로 인하여 퇴적층 내로의 산소 공급이 원활하며, 많은 유
기물의 유입에도 불구하고, 혐기성 분해과정보다 유기물 분해에 있어서 좀더 효과
적인 호기성 분해과정의 비중이 높을 것으로 예상되며 따라서 갯벌에서는 육상에
서 기원한 유기물의 분해가 매우 활발할 것으로 예상된다 (Howe et al., 1984;
Boudreau, 1997). 그러나 갯벌에서 미생물에 의한 유기물의 분해 능력 전체를 유
기오염 자연정화능력으로 간주하는데는 어려움이 있다. 현재 갯벌의 자연정화능
력에 대한 논란의 일부는 “자연정화의 개념”을 유기물 분해 능력으로 잘못 이해
한데서 비롯된다. 유기물은 탄소성분과 질소성분으로 이루어져 있는데, 미생물이
유기물을 분해할 경우 탄소성분은 이산화탄소로서 시스템 내에서 제거되나 질소
성분은 무기질소로 변하여 남아 있게 되며, 이러한 무기질소는 일차 생산을 제한
하는 제한 요소로서, 광합성에 의한 유기물 생산에 이용될 것이다 (Jorgensen,
1977). 결국 탄소성분이 제거되었다 하더라도 질소성분은 남아 다시 유기물을 생
산하게 될 것이며, 미생물에 의한 유기물의 분해 능력 전체를 시스템 내에서 유기
물의 제거능력으로 간주하는 것은 잘못이다. 갯벌의 유기정화 능력을 파악하기
위해서는 미생물에 의한 유기물의 분해 능력과 더불어 일차생산에 의한 유기물의
합성과정, 합성된 유기물의 소비과정을 파악하는 것이 중요할 것이다. 갯벌에서
일차 생산에 의해 생산된 유기물은 먹이망에서 상위 단계에 있는 생물들에게 소
비되거나, 입자성 유기물 (POM; particulated organic matter)로서 미생물에 의한
무기화과정에 이용될 것이다. 어류나 기타 대형 저서동물들이 일차 생산에서 생
산된 유기물을 소비하여 생체량 (biomass) 을 만들고, 인간이나 조류 등이 이러
한 어류나 대형 저서동물을 소비하면, 갯벌내에서 유통되는 유기물의 양은 감소할
것이고 따라서 갯벌의 유기정화능력을 증가시키는데 기여할 것이다 (Fig. 5-1).
갯벌에 의한 유기물의 분해와 미세조류에 의한 재생산과정은 유용하지 않은 유기
물 혹은 생태계에 부정적인 영향을 주는 유기물 (오염유기물; 생활폐수등) 이 인
간이나 조류 등이 이용할 수 있는 자원(자원유기물; 저서동물, 어패류)으로 변할

- 276 -
저서 미세조류(C/N)
오염유기물유입 (C/N)
유기물(C/N) 무기질소 (N)
자원유기물(C/N) 탈질소화 (N)
유기물정화
퇴적층
수층
수확 (C/N)
저서 미세조류(C/N)
오염유기물유입 (C/N)
유기물(C/N) 무기질소 (N)
자원유기물(C/N) 탈질소화 (N)
유기물정화
퇴적층
수층
수확 (C/N)
Figure. 5-1. 갯벌의 유기오염 정화의 개념도 유기물과
저서미세조류는 탄소 (C)와 질소(N) 성분을 가진다.
수 있는 기작을 보여준다 (Fig. 5-1). 이 기작이 중요하다면 갯벌은 유기오염 자
연 정화와 자원생성이 동시에 이루어지는 곳이 될 것이다.
갯벌의 유기정화 능력을 파악하기 위해서는 미생물에 의한 유기물의 분해 능력
과 더불어 일차생산에 의한 유기물의 합성과정, 합성된 유기물의 소비과정을 파악
하는 것이 중요할 것이다. 이와 더불어 시스템 내에서 유기물을 제거하는 기작으
로 중요한 것은 탈질소화 (denitrification)가 있다. 탈질소화는 미생물에 의한 혐기
성 호흡의 하나로 이 과정에서 질산이 가스형태(N2 나 N2O)로 변환된다. 탈질소
화의 산물인 질소 가스는 일차생산에 이용될 수 없으며, 따라서 탈질소화는 시스
템 내에서 일차생산에 이용 될 수 있는 총질소 (무기질소와 유기질소)를 감소시
키고 결과적으로 유기물을 제거할 수 있는 기작이다(Seitzinger, 1990).탈질소화가
상세히 연구된 연안이나 하구에서는 이 과정이 플랑크톤에 의한 일차생산성을 제
한하거나 부영양화를 감소시키는데 중요하다는 것이 밝혀졌다 (Howarth et al.,
1988; An and Joye, 2001). 본 연구에서는 낙동강하구 갯벌에서 퇴적물에 의한
유기물 분해와 탈질소화를 자세히 연구한바 있다. 이를 바탕으로 낙동강 하구전체
퇴적물에서 일어나는 유기물 정화능력을 추정해 보았다.
나. 낙동강 하구 갯벌에서 유기물 분해 및 탈질소화에 의한 유기물 제거
낙동강 실험용 갯벌에서의 SOD를 분석한 결과 일정한 계절적인 경향이 나타났

- 277 -
다. 05년 실험용 갯벌의 SOD 값의 범위는 -0.7~35.3 mmole m-2
d-1 였고, 하절기
에 가장 높은 SOD 값(평균: 30mmole m-2
d-1
)을 보였으며, 동절기에는 산소가 소
비되기 보다는 광합성에 의해 산소가 생성되는 것을 나타났다(mean :-0.5 mmole
m-2
d-1
). 06년 실험용 갯벌의 SOD 값의 범위는 -0.5~38.2mmole m-2
d-1였고, 2005
년 실험용 갯벌과 유사하게 하절기 (평균: 37mmole m-2
d-1
), 동절기 (평균:
0.1mmole m-2
d-1
)의 SOD 값을 나타냈다. 이는 미생물 활동을 조절하는 큰 요인
은 계절적인 수온 변화임을 시사한다. 이러한 유기물 분해율을 낙동강 전체에 적
용하여보면 탄소량으로 년간 평균 7.1 x 106ton이 된다. 이는 년간 낙동강 하구
로 유입되는 유기물 총량인 38.3 x 106ton (화학적산소요구량 COD에서 환산)의
약 19%에 해당하는 값이다. 탈질소화의 경우 05년 실험용 갯벌의 탈질소화 값의
범위는 5.9~2652μmole m-2
d-1 (mean)였고, 하절기에 가장 높은 탈질소화 값
(4564.6μmole m-2
d-1
)을 보였으며, 동절기에는 거의 탈질소화가 발생하지 않았다
(1.6μmole m-2
d-1
). 06년 실험용 갯벌의 탈질소화 값의 범위는 3.9~2082 μmole
m-2
d-1였고, 05년 실험용 갯벌과 유사하게 하절기 (2626.5μmole m
-2d
-1), 동절기
(0.01μmole m-2
d-1
)의 탈질소화 값을 나타냈다. 탈질소화에 의해 제거되는 질소영
양염을 탄소/질소비율 6.6 (redfield ratio )을 적용하여, 탄소량으로 환산하면, 낙
동강 전체에서 3.9 x 106 ton이 된다. 이는 COD 유입량의 약 10%에 달하는 양이
다.
Table. 5-1. 낙동강 전체 면적당 퇴적물 산소요구량 (SOD)와 탈질소화
SOD 탈질소화 COD 유입량
낙동강 전체면적당 (백만 ton-C year-1)
하절기 14.3 6.5 64.4
동절기 0 15.0 12.2
평균 7.1 3.9 38.3
2. 잘피에 의한 영양염 제거
연안 및 하구 생태계에서 가장 큰 오염원은 양영분의 과다 유입으로 인한 해양
의 부영양화 현상이 대표적이라 할 수 있다. 이 중 대표적인 영양염류는(암모니
아, 질산염, 인산염) 질소로서, 해수 내 질소의 제거는 대표적인 해수 정화의 한
방법이다. 본 연구에서는 매월 잎에서 생산되는 생산량과 잎 조직 내 질소 함량을
이용해서 매월 잘피 잎 조직이 해수에서 제거되는 질소 양을 추정하였다.
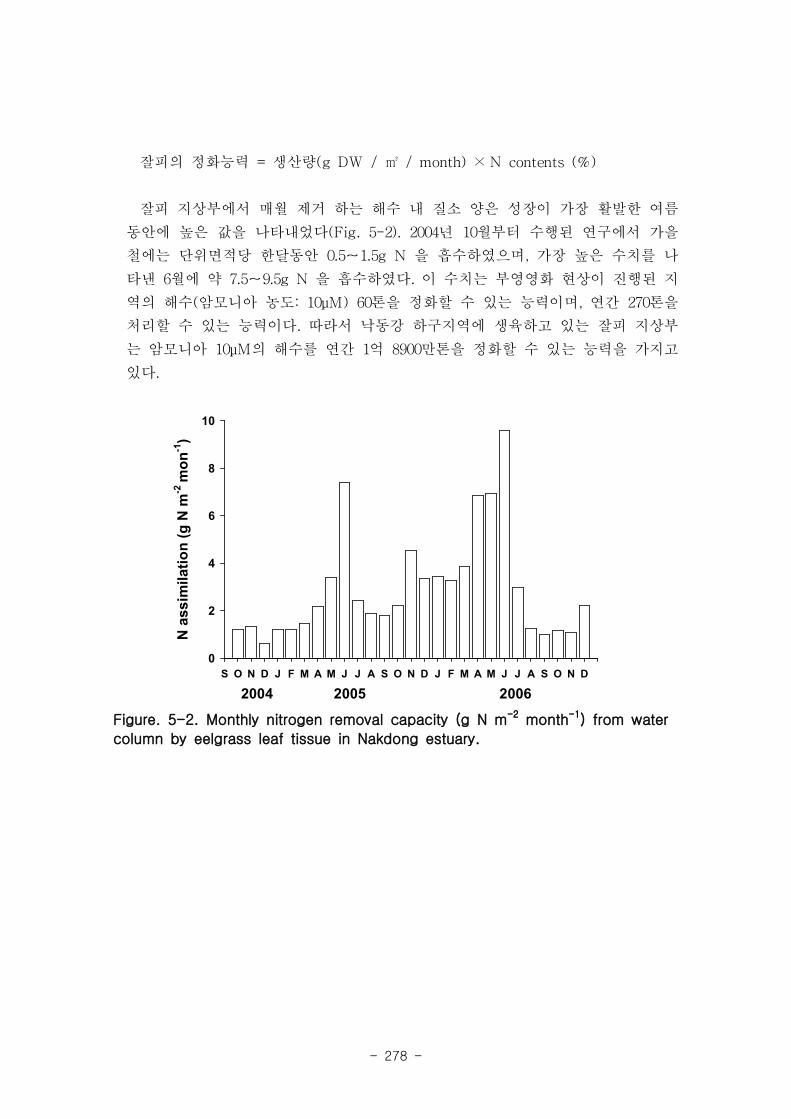
- 278 -
잘피의 정화능력 = 생산량(g DW / ㎡ / month) × N contents (%)
잘피 지상부에서 매월 제거 하는 해수 내 질소 양은 성장이 가장 활발한 여름
동안에 높은 값을 나타내었다(Fig. 5-2). 2004년 10월부터 수행된 연구에서 가을
철에는 단위면적당 한달동안 0.5~1.5g N 을 흡수하였으며, 가장 높은 수치를 나
타낸 6월에 약 7.5~9.5g N 을 흡수하였다. 이 수치는 부영영화 현상이 진행된 지
역의 해수(암모니아 농도: 10μM) 60톤을 정화할 수 있는 능력이며, 연간 270톤을
처리할 수 있는 능력이다. 따라서 낙동강 하구지역에 생육하고 있는 잘피 지상부
는 암모니아 10μM의 해수를 연간 1억 8900만톤을 정화할 수 있는 능력을 가지고
있다.
2004 2005 2006
S O N D J F M A M J J A S O N D J F M A M J J A S O N D
N a
ssim
ilation (g N
m-2
mon
-1)
0
2
4
6
8
10
Figure. 5-2. Monthly nitrogen removal capacity (g N m-2 month-1) from water
column by eelgrass leaf tissue in Nakdong estuary.
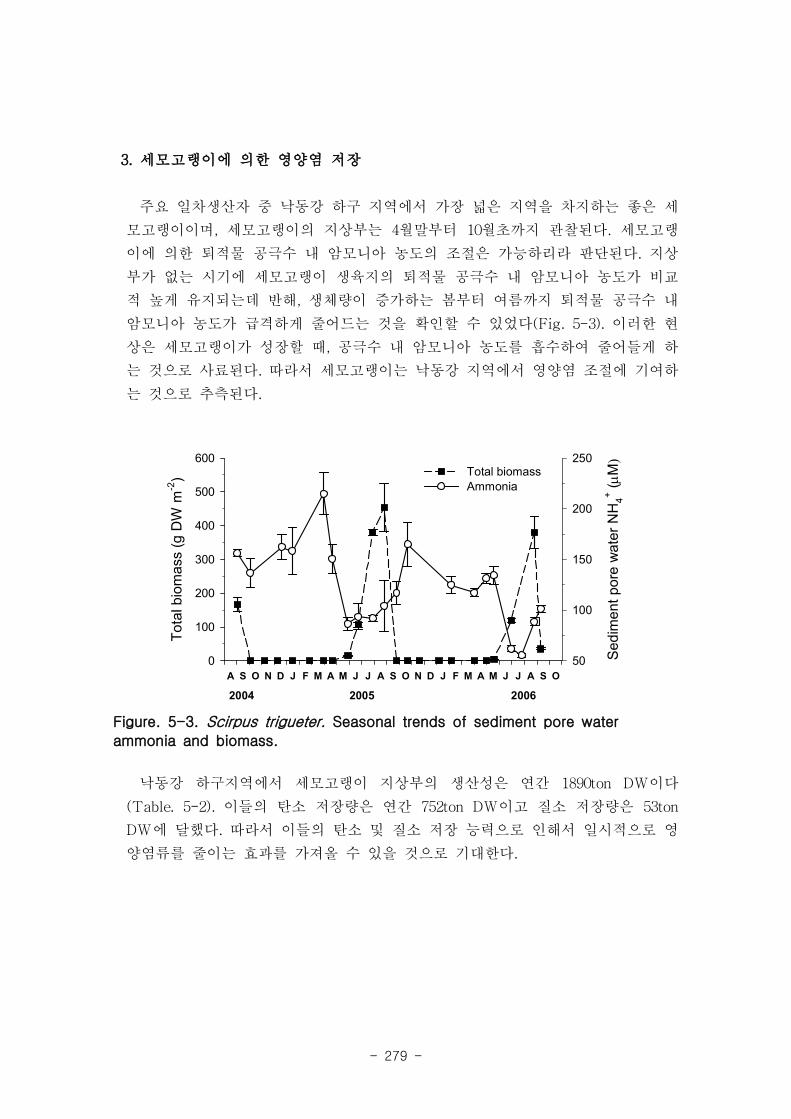
- 279 -
3. 세모고랭이에 의한 영양염 저장
주요 일차생산자 중 낙동강 하구 지역에서 가장 넓은 지역을 차지하는 좋은 세
모고랭이이며, 세모고랭이의 지상부는 4월말부터 10월초까지 관찰된다. 세모고랭
이에 의한 퇴적물 공극수 내 암모니아 농도의 조절은 가능하리라 판단된다. 지상
부가 없는 시기에 세모고랭이 생육지의 퇴적물 공극수 내 암모니아 농도가 비교
적 높게 유지되는데 반해, 생체량이 증가하는 봄부터 여름까지 퇴적물 공극수 내
암모니아 농도가 급격하게 줄어드는 것을 확인할 수 있었다(Fig. 5-3). 이러한 현
상은 세모고랭이가 성장할 때, 공극수 내 암모니아 농도를 흡수하여 줄어들게 하
는 것으로 사료된다. 따라서 세모고랭이는 낙동강 지역에서 영양염 조절에 기여하
는 것으로 추측된다.
2004 2005 2006
A S O N D J F M A M J J A S O N D J F M A M J J A S O
Total biomass (g DW m
-2)
0
100
200
300
400
500
600
Sediment pore water NH4
+ (µM)
50
100
150
200
250Total biomass
Ammonia
Figure. 5-3. Scirpus trigueter. Seasonal trends of sediment pore water
ammonia and biomass.
낙동강 하구지역에서 세모고랭이 지상부의 생산성은 연간 1890ton DW이다
(Table. 5-2). 이들의 탄소 저장량은 연간 752ton DW이고 질소 저장량은 53ton
DW에 달했다. 따라서 이들의 탄소 및 질소 저장 능력으로 인해서 일시적으로 영
양염류를 줄이는 효과를 가져올 수 있을 것으로 기대한다.
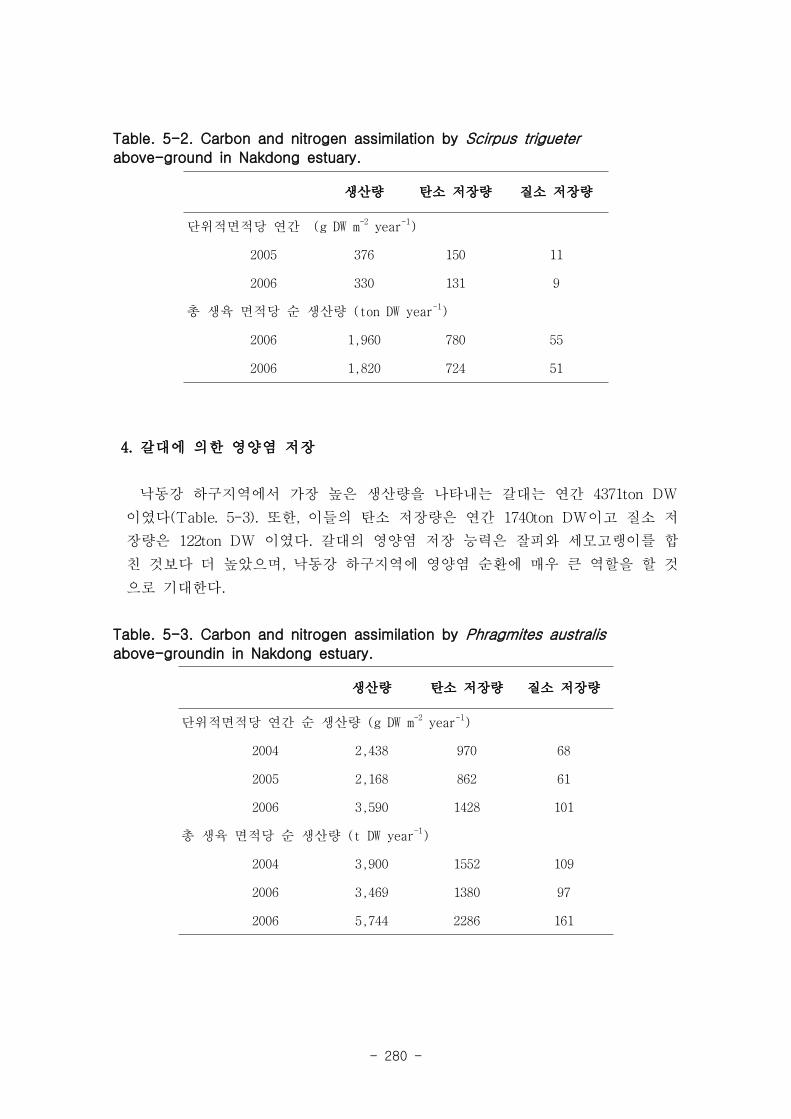
- 280 -
Table. 5-2. Carbon and nitrogen assimilation by Scirpus trigueter above-ground in Nakdong estuary.
생산량 탄소 저장량 질소 저장량
단위적면적당 연간 (g DW m-2 year
-1)
2005 376 150 11
2006 330 131 9
총 생육 면적당 순 생산량 (ton DW year-1)
2006 1,960 780 55
2006 1,820 724 51
4. 갈대에 의한 영양염 저장
낙동강 하구지역에서 가장 높은 생산량을 나타내는 갈대는 연간 4371ton DW
이였다(Table. 5-3). 또한, 이들의 탄소 저장량은 연간 1740ton DW이고 질소 저
장량은 122ton DW 이였다. 갈대의 영양염 저장 능력은 잘피와 세모고랭이를 합
친 것보다 더 높았으며, 낙동강 하구지역에 영양염 순환에 매우 큰 역할을 할 것
으로 기대한다.
Table. 5-3. Carbon and nitrogen assimilation by Phragmites australis above-groundin in Nakdong estuary.
생산량 탄소 저장량 질소 저장량
단위적면적당 연간 순 생산량 (g DW m-2 year
-1)
2004 2,438 970 68
2005 2,168 862 61
2006 3,590 1428 101
총 생육 면적당 순 생산량 (t DW year-1)
2004 3,900 1552 109
2006 3,469 1380 97
2006 5,744 2286 161

- 281 -
제 4장 목표 달성도 및 관련분야에의
기여도

- 282 -

- 283 -
제 4장 목표 달성도 및 관련분야에의 기여도
본 연구의 연도별 연구 개발 목표, 세부목표, 평가착안점 및 개발 내용 및 달성
도를 Table. 나에 요약하였다. 1차년도에는 낙동강 하구 습지의 퇴적물 및 수층
환경 특성 그리고 생산자, 소비자, 분해자등의 각 생태 요소의 실태를 파악하는데
중점을 두었으며, 낙동강 하구 갯벌의 전반적인 퇴적물과 수층의 특성을 파악하
고, 생태 요소를 조사하는 등 목표를 달성하였다. DGPS를 이용하여, 조고를 정밀
측정하였으며 이를 토대로 실험용 습지 조성 적지를 선정하였다. 2차년도에는 1차
년도의 환경 및 생물 특성조사를 계속하여, 계절 및 연간 변화를 추적하였으며,
실험용 갯벌의 조성하여, 안정화 과정 중에 일어나는 환경 및 생물변화를 조사하
는 등 2차년도의 연구 목표를 달성하였다. 3차 년도에도 각종 환경 및 생태요인들
을 계속 조사하여, 장기적인변화를 살펴보았으며, 2차로 실험용갯벌을 조성하여, 1
차 갯벌과 비교를 통해 연간 변화와 안정화과정을 살피었다. 잘피 이식실험을 추
가로 실시하였고, 안정동위원소 추적 실험을 통해 잘피 및 갈대밭에서 먹이망을
파악하였다. 낙동강을 통해 하구로 유입되는 영양염 및 오염물질의 유입량의 계절
변화를 정량화 하고 이를 낙동강 하구에서 일어나는 여러 생태과정의 속도와 비
교하여, 시스템 버젯을 계산하였다. 이러한 계산을 통하여, 각 생태요소가 낙동강
하구 생태계에서 얼마나 중요한 역할을 하는지 알 수 있었으며, 낙동강 생태계가
자정 작용 등에 얼마나, 어떤 방식으로 기여하는지 알 수 있었다.
- 284 -
구분연구개발의
목표세부목표 평가 착안점 개발 내용 및 달성도
1차연도(200
4)
. 낙동강 하구 습지의 환경 특성 및 생태 요소 현황 파악
. 실험용 습지 조성 조건 파악
낙동강 하구
습지의 환경
생태 환경에 중요한 환경
요인 공간/시간 범위, 조사
방법의 적절성
-퇴적물 입도, 영양염 농도, 유기물농도, 조고 및 지형변화측정 완료;
- 수층의 특성측정 완료
저서미세조류분포퇴적물 형태별, 지역별
종조성
- 종조성, 다양성, 서식밀도, 클로로필량 측정완료
저서 동물 분포조고, 퇴적물 형태별,
지역별 종조성
- 낙동강 갯벌 저서동물의 종조성, 다양성, 서식밀도, 습중량, 건중량, 탄소, 질소량 파악 완료
퇴적물 내
미생물 분해
환경요인의 차이에 따른
분해율
- 자연 및 인공갯벌의 퇴적물산소요구량 (SOD), 탈질소화, 영양염 flux 측정 완료
잘피 및
대형일차생산자
의 분포
분포 파악 및 잘피
이식가능성
- 주요 일차생산자의 현황 및 분포도 작성, - 인공잘피림과 자연 잘피림의 일일 광량
및 온도 측정완료
2차연도(200
5)
.실험용 습지의 안정화 과정
. 조고 및 퇴적물 타입에 따른 생태 반응
.대형일차생산자와 물질 순환 패턴
실험용 습지조성
및 안정화과정
파악
생태 환경에 중요한 환경
요인 공간/시간 범위, 조사
방법의 적절성
-1m x 1m,크기의 실험용 갯벌 조성; 3개의 조고, 3개의 다른 입도 조건 (총 9개)
-실험용 갯벌의 안정화 과정 및 생태 변화 조사 완료
저서미세조류에
의한 생산성
퇴적물 형태별, 지역별
종조성 생산성
- 실험용갯벌의 미세저서 조류 및 저서생물의 가입 및 종다양성 변화조사 완료
저서 동물
먹이망저서생물의 먹이망 분석
- 세모고랭이밭과 같대밭의 먹이망분석- 주요 패류자원의 생리생태 조사완료
퇴적물 내
미생물 분해 및
탈질소화
실험용 갯벌에서 각
환경요인의 차이에 따른
분해율 및 탈질소화
- 각 상중하, 사질, 니질, 사니질 갯벌에서 SOD 및 탈질소화 측정 완료
대형일차생산자
의 분포 및
생산성
대형일차 생산자 생산성 및
물질 순환에서의 역할 규명
- 갈대 및 잘피 서식 면적 파악 완료- 생산성 파악 완료- 1차년도에 이어 이식실험 실시 완료
3차연도(200
6)
.실험용 습지의 연간 변화
.시스템 budget을 통한 낙동강 하구 의 생태계 특성 파악. 낙동강 하구의 생태계를 통한 자정능력 파악.습지 복원 파라미터 개발
실험용습지의 추가
조성으로 연간변화
파악
자료의 범위, 본 조사
자료와의 적절한 비교 분석
.-2m x 2m,크기의 실험용 갯벌 추가 조성 (총 9개)
- 1차 갯벌과의 비교를 통해 연간 변화파악
대형일차 생산자
및 MPB의 생산성
생산성의 계절 변화, 다른
생태요소와의 비교
- 갈대, 잘피, MPB의 생산성 파악- 생체량과 생산성간의 관계 파악
시스템 budget
파악
유입량과 각 생태과정의
비교의 적절성
. 담수 및 오염 물질의 계절성
. 각 생태요인의 상대적 중요성 파악
생태계를 통한
자정능력
하구 갯벌의 자정능력 개념
정립 및 정량화
- 유기물 분해와 자정 능력 개념- 탈질소화의 정량화- 유입량과의 비교분석 완료
복원 파라미터
개발
잘피, 세모고랭이, MPB,
유기물 분해를 극대화하는
복원 조건 파악
. 종다양성, 생물량을 높이는 조성 조건 파악
. 유기물 분해, 영양염 제거 상관관계
. 하구 각 생태요소의 기능 파악
Table. 나. 낙동강 하구연구의 개발 목표, 평가착안점 및 개발 내용 및 달성도

- 285 -
제 5장 연구개발결과의 활용계획

- 286 -

- 287 -
제 5장 연구개발결과의 활용계획
해양수산부는 연안 하구역 조사사업을 장기적으로 추진할 예정이며, 1단계로서
2007년에 “하구역의 고유 기능과 가치를 과학적이고 종합적으로 평가할 수 있는 체
계화된 조사지침서를 작성”연구를 수행하고 있다. 이 조사의 목적은 지금까지 실시
되어온 생태계 구조연구에서 벋어나, 하구의 생태계의 기능과 가치를 알아보려는 것
으로서 본 연구의 결과가 좋은 모델로서 사용될 수 있을 것이다. 물론 조사사업의
경우 본 연구의 모든 부분을 대상으로 삼기는 힘들 것이나, 향후 자료가 축적되고,
조사 재원이 확보되었을 때 어떠한 연구를 해야 하는가에 대한 방향타 역할을 본 연
구가 할 수 있을 것이다.
이처럼 정부나 민간의 습지복원/관리 투자가 이루어지면 산업화가 촉진되어 벤처
기업화가 가능할 것이다. 유럽 및 미국의 경우 상업적으로 잘피를 이식해 주는 많은
기업들이 있다. 여러 가지 요인에 의해 연안습지를 훼손할 경우, 훼손한 주체 (기업
또는 지자체 등)에서 훼손한 습지와 동일한 면적의 인공습지를 조성해야한다는 법령
(예, No net loss of wetland policy)이 제정될 경우, 상업적인 잘피이식이 필요하게
될 것이다. 그 이외에 본 연구의 결과는 다음과 같은 분야에서 이용될 수 있다.
- 대형 복원공사에서 각 지역의 여건에 맞는 복원 목표를 설정, 경제적인 시공
및 관리 기법을 선택.
- 해양환경의 기준설정이나 해양환경 영향평가에 구체적인 평가 자료를 제공.
- 연안 수산자원관리 조성사업에 대한 정책설정의 기본적인 방향을 자연과학적
측면에서 평가할 수 있는 틀 제시.
-현재 대형 연안개발에서 나타나는 논쟁에서 “개발” 혹은 “보존” 이외의 제 3의
방안으로 복원이 검토되어 막대한 사회비용을 절감할 수 있으며 복원 산업이 활
성화.
-조하대 잘피림을 조성하면 1) 수산어족자원 및 해양생물의 다양성 보존 2) 짧
은 기간 내 환경친화적으로 수산생산성 향상을 꾀할 수 있음. 3) 조성된 잘피밭에
의한 무기영양염류 및 오염물질의 흡수제거에 따른 연안 생태계 정화 등에 기여.
- 연안생태계의 물질순환 모델을 통하여 얻어지는 결과들로부터 인위적 연안환
경의 변동이 연안생태계의 생물다양성과 생물생산 특히 수산자원생물의 생산 변
동에 미치는 직접적인 효과에 대한 구체적인 정보를 제공.
- 연안생태계의 물질순환 모델을 통한 각 연안생태계 구성요소들의 역할에 대한 정
확한 평가를 바탕으로 합리적인 연안역의 보호와 관리를 위한 해양환경 보전정책수립.
본 연구에서는 하구 습지 생태계의 구조와 기능을 전반적으로 살펴 그 상호 관
계를 살펴본 것으로서 본 연구 결과로 갖추어진 틀 안에서 각각의 개별 연구가
추가적으로 진행되어야 할 것이다.

- 288 -

- 289 -
제 6장 연구개발과정에서 수집한
해외과학기술정보

- 290 -

- 291 -
제 6장 연구개발과정에서 수집한 해외과학기술정보
연관 분야 도서 수집
- Batzer, Darold P. 저, 2007. Ecology of Freshwater And Estuarine
Wetlands. ISBN-13 : 9780520247772 ,568쪽
- Boudreau, Bernard P 저, 2001. The Benthic Boundary Layer. ISBN-13 :
9780195118810, 404쪽
-Bianchi, Thomas S 저, 2006, Biogeochemistry of Estuaries. ISBN-13 :
9780195160826 , 706쪽

- 292 -

- 293 -
제 7장 참고문헌

- 294 -

- 295 -
제 7장 참고문헌
고철환, 1991. 한국 서해 펄 갯벌에서의 미세조류에 의한 제1차 생산 및 생물량,
KOSEF 891-0505-015-2.
국립수산진흥원, 1999. 한국연근해 유용연체동물도감. 도서출판구덕, 부산, 215pp.
권오길, 박갑만, 이준상, 1993. 원색한국패류도감. 아카데미서적, 445pp.
김구연, 이찬우, 윤해순, 주기재, 2005. 낙동강 하구의 수생관속식물의 분포 변화
와 수금류(고니류)의 먹이식물인 세모고랭이의 성장변화. 한국생태학
회지 28(5):335~345.
김준호, 김훈수, 이인규, 김종원, 문병태, 서규홍, 권도헌, 유순애, 서영배, 김영상,
1982. 낙동강하구생태계의 구조와 기능에 관한 연구. 서울대학교
자연과학대학교논문집, 7(2), 121~163.
김준호, 문형태, 민병미, 조경재, 1989. 낙동강 하구 염습지 식물군락의 질소 및
인의 동태. 한국생태학회지 12(1):1~7.
김준호, 조경재, 문형태, 민병미, 1986. 낙동강 하구 사주의 갈대, 천일사초 및 갯
잔디 군락의 생활동태. 한국생태학회지 9(2):59~71.
김준호, 조경제, 1985. 금강 하구 조간대 저토의 물리화학적 특성, 저생규조의 종
조성 및 생물량. 한국생태학회지, 8:21-29.
김훈수, 1973. 한국동식물도감, 제14권 동물편(집게・게류). 삼화출판사, 서울, pp.
694.
김훈수, 1977. 한국동식물도감, 제19권 동물편(새우류). 삼화출판사, 서울, 414pp.
노희명, 최우정, 이은주, 윤석인, 최영대, 2002. 시화지구 인공습지에서 갈대에 의
한 질소 및 인의 흡수. 한국생태학회지 25(5):359~364.
문화공보부, 1971. 지정문화재목록. 118pp.
박병권, 이광식, 1994. 서해안 곰소만 조간대 유공충의 안정동위원소 성분. 한국
해양학회지, 29: 183―187.
부산광역시, 2003. 낙동강 에코센터 전시기본계획 설계. 174pp.
부산지방환경청, 1992. 낙동강 하구언 축조 전후 주요환경 변화조사. 308pp.
산업기지개발공사, 1981. 낙동강 하구언건설공사 환경영향평가서. 298pp.
손민호, 홍성윤, 2003. 바위해변에 사는 해양생물. 풍등출판사, 서울, 143pp.
심재형, 1994. 한국 동식물 도감 제 34권 식물편(해양식물플랑크톤), 국정교과서
주식회사
심재형, 조병철, 1984. 인천 부근 조간대의 저생규조류 군집. 서울대학교 자연과
학대학 논문집,135~150.
안태영, 1985. 진해만 생태계에 있어서 미생물 분포에 영향을 미치는 환경요인의
분석, 박사학위논문, 서울대학교
안순모, 2005. 강화도 갯벌 퇴적물의 산소요구량과 탈질소화의 계절 변화. 한국해

- 296 -
양학회지. 10(1):63-70
양홍준, 김구환, 금지돈, 2001. 낙동강 하구의 어류상과 댐의 어도에서 어류의 이
동 Korean J. Lim., 34(3): 251-258.
오승진, 문창호, 박미옥, 2004. 한국 서해 새만금 갯벌에서 저서미세조류의 생체
량과 군집조성에 대한 HPLC분석. 한국수산학회지. 37:215~225.
우한준, 장진호, 한상준, 1995. 곰소만 조간대의 현생 유공충과 표층 퇴적물의 특
성: 고환경 해석에 적용 가능성. 한국해양학회지, 30: 184―196.
원병오, 1985. ‘84 낙동강 하구 생태계조사-8, 조류(鳥類), 산공. 273~328.
윤춘경, 권순국, 우선호, 권태영, 1999. 농촌지역 수질개선을 위한 인공습지실험시
설의 3년간 실험결과 검토. 한국물환경학회지. 15(4) 581-589.
윤해순, 1991. 낙동강 하구 간석지의 수생관속식물에 관한 연구. 하구둑 건설 전
후의 간석지 식생의 비교. 한국생태학회지 14(1):63~73.
이강현, 한명우, 2001. 퇴적물내 유기물의 초기 속성 작용에 나타난 유기질소의
선택적 분해. 한국해양학회지. 6(2):63-70
이용우, 2001. HPLC를 이용한 곰소만 퇴적물 중의 저서미세조류의 색소 연구,
부경대학교 석사학위 논문, 69pp.
이찬원, 권영택, 1995. 퇴적오니 준설과 해양환경변화.
임현식, 홍개상, 1994. 진해만 저서동물의 군집생태1. 저서환경. 한국 수산학회지
27(2). 200-214.
임현식, 홍개상, 1997. 진해만 저서동물의 군집생태2. 우점종의 분포. 한국 수산학
회지 27(2). 200-214.
장진호, 박용안, 한상준, 1996. 한국 서해안 곰소만 조간대의 제 4기 층서와 해수
면 변화. 한국해양학회지「바다」, 1: 59―72.
장진호, 전승수, 권수재, 신동혁, 한상준, 박용안, 1993. 황해 곰소만 조간대에 발
달한 Chenier의 퇴적학적 특성과 진화. 한국해양학회지, 28: 212―228.
19.
조병철, 2005. 경기만 부근 갯벌의 생지화학적 연구, 한국해양학회지. 바다. 10:1-
5.
최휴창, 2002. 강화 펄 조간대에서의 저서성 미세조류 연구. 인하대학교 석사학위
논문. 67pp.
한국해양연구원, 2001. 폐쇄해역의 생물천이과정 및 환경평가. BSPE 00791-00-1325-
3, 155.
해양수산부, 1998. 우리나라의 갯벌.
해양수산부, 1998. 해양환경공정시험방법. 317pp.
해양연구소, 1993. 폐쇄성 연안역의 수질관리기술 연구(I).
환경부, 2005. 낙동강 하구역 습지생태계 훼손지역 복원 및 관리기술(I). 차세대
핵심환경기술개발사업 연구보고서. 117 p

- 297 -
환경부, 2006. 낙동강 하구역 습지생태계 훼손지역 복원 및 관리기술(II). 차세대
핵심환경기술개발사업 연구보고서. 149p
An, S., and S. B. Joye, 1997. An improved chromatographic method to measur
e nitrogen, oxygen, argon and methane in gas or liquid samples. M
ar. Chem. 59:63-70.
An, S., and S. B. Joye, 2001. Enhancement of coupled nitrification-denitrificatio
n by benthic photosynthesis in shallow estuarine sediments. Limnol.
Oceanogr. 46(1) 62-74.
An, S., and W. S. Gardner. submitted. Dissimilatory nitrate reduction to amm
onium (DNRA) as nitrogen link versus denitrification as a sink in
a shallow estuary (Laguna Madre/Baffin Bay, Texas). Mar. Ecol. P
rog. Ser.
An, S., W. S. Gardner. and Kana T, 2001. Simultaneous measurement of denitrifi
cation and nitrogen fixation using isotope pairing with membrane inle
t mass spectrometer (MIMS) analysis. App. Env. Micro. 67(3) 1171-1
178.
Asmus, R, 1982. Field measurements of seasonal variation of the activity of pr
imary producers on a sandy tidal flat in the Northern Wadden Sea.
Netherland Journal of Sea Research 16: 389-402.
Bae, K. S. and I. B. Yoon, 1988. A study on the characteristics of substratum
and benthic macroinvertebrates community in Nakdong Estuary. Ko
rean J. Lim., 21(2): 93~116.
Bae, K. S. and I. B. Yoon, 1989a. Ecological studies of benthic macroinverteb
rates in Nakdong estuary, Korea. Korean J. Lim., 22: 337~356.
Bae, K. S. and I. B. Yoon, 1989b. Study on the environmental characteristics
and dynamics of benthic macroinvertebrates in Nakdong estuary. K
orean J. Lim., 22(1): 11~27.
Bligh, E. G. & W. F. Dyer, 1959. A rapid method of total lipid extraction and
purification. Can. J. Biochem. Physiol. 37:911-917.
Bonsdorff E, 1980. Macrobenthic recolonization of dredged brackish water ba
y in SW Finland. Ophelia. Suppl. 1. 145-155
Borja, A., J. Franco, and V. Perez, 2000. A marine biotic index to establish th
e ecological quality of soft-bottom benthos within European estuari
ne and coastal environments. Mar. Pollut. Bull., 40: 1100-1114.
Brandes, J. A. and A. H. Devol., 1995. A marine isotopic nitrogen budget: Im
plications for present and past nutrient balances. EOS. 76(3): 134-1
38.

- 298 -
Bray, J. R. and Curtis, J. T., 1957. An ordination of the upland forest commu
nities of Southern Wisconsin, Ecol.. Monogr. 27: 325~349.
Capone DG, 1988. Benthic Nitrogen fixation. In Blackburn, T. H. and Sørensen, J.
(eds.), Nitrogen Cycling in Coastal Marine Environments. Wiley.
Choi, J. W. and C. H. Koh, 1994. Macrobenthos community in Keum-Manky
ung-Dongjin estuaries and its adjacent coastal region, Korea. J. K
orean Soc. Oceanogr., 29(3): 304~318.
Christensen, J. P., J. W. Murray, A. H. Devol and L.A. Codispoti. 1987. Deni
trification in continental shelf sediments has major impact on the
oceanic nitrogen budget. Glob. Biogeochem. Cyc. 1(2): 97-116.
Chung YH, Choi HK, 1985. Flora and standing stock of vascular hydrohpytes
from the water course of Naktong River. Korean J. Environ. Biol.
1: 29-44.
Cofer, S.R., Niering, W. A., 1992 Comparison of created and natural freshw
ater emerged wetland in connecticut (U.S.A). Wetlands Ecol. Ma
nagement. 2(3) 143-156.
Cohen, E. B., Grosslein, M. D., Sissenwine, M. P., Steimle, F. W., Wright, W. R.,
1982. An energy budget of Georges Bank. In: Mercer, M. C. (ed.) M
ulti-species approaches to fisheries management advice. Can. Spec.
Pub. Fish. Aquat. Sci., 59: 95~107.
Drake, P., F. Baldo, V. Saenz, and A.M. Arias, 1999. Macrobenthic commu
nity structure in estuarine pollution assessment on the Gulf of c
adiz (SW Spain): is the phylum-level meta-analysis approach ap
plicable? Mar. Pollut. Bull., 38: 1038-1047.
Dubois, M., K. A. Gilles, J. K. Hamilton, P. A. Rebecs & F. Smith. 1956. Colo
rimetric method for the determination of sugars and related substan
ces. Anal. Chem. 28:350-356. et al., 1956.
Evans, P. R., R. M. Ward, M. Bone, and M. Leakey, 1998. Creation of tempe
rate-climate intertidal mudflats: factors affecting colonization and
use by benthic invertebrates and their bird predators. Mar. Pollut.
Bull., 37: 535-545.
Gambi, M. C. and A. Giangrande, 1986. Distribution of soft-bottom polychaete
s in two coastal areas of the Tyrrhenian Sea(Italy): structural analy
sis. Estuar. Coast. Shelf Sci., 23: 847~862.
Gardner WS, Bootsma HA, Evans C, St. John PA., 1995. Improved chromatog
raphic analysis of 15N: 14N ratios in ammonium or nitrate for isoto
pe addition experiments. Mar Chem 48:271-382

- 299 -
Gedik CM et al., 1992. International Journal of Radiation Biology 62, 313-320
Havens K. J. Varnell, L. M., and Bradshaw, J. G., 1995. An assessment of ec
ological conditions in a constructed tidal marsh and two natural ref
erence tidal marshes in coastal Virginia. Ecol. Eng. 4. 117-141
Hemminga M. A. and Duarte C. M., 2000. Seagrass ecology. Cambridge Univ
ersity Press, Cambridge. 298 pp.
Hilly, C., 1983. Macrozoobenthic recolonization after dredging in a sandy mud
area of the Bay of Brest enriched by organic matter. Oceanol. Acta
Proc. 17th Europ. Mar. Biol. Symp. 1982. 113-120.
Hong, J. S. and J. W. Yoo, 1996. Salinity and sediment types as sources of vari
ability in the distribution of the benthic macrofauna in Han Estuary
and Kyonggi Bay, Korea. J. Kor. Soc. Oceanogr., 31(4):217~231.
Howarth, RW., R. Marino, J. Lane and JJ. Cole, 1988. Nitrogen fixation in fr
esh-water, estuarine and marine ecosystems. I. Rates and Importa
nce. Limnol Oceanogr., 33: 669- 687.
Howes, B. L., J. W. H. Dacey and G. M. King. 1984. Carbon flow through ox
ygen and sulfate reduction pathways in salt marsh sediments. Limno
l. Oceanogr. 29(5):1037-1051.
Hwang , C. H., Cho, B. C. 2005. Measurement of Net Photosynthetic Rates in Int
ertidal flats of Ganghwa-gun and Incheon North Harbor using Oxygen
Microsensors. The Sea, Journal of the Korean Society of Oceanograph
y 10: 31-37
Jang, I. K. and C. H. Kim, 1992. A study on the changes of molluscan and cr
ustacean fauna after the construction of the Nakdong estuary barrag
e. Bull. korean Fish. Soc., 25(4): 265~281.
Jeffrey S. W. and Humphrey G. F., 1975. New spectrophotometric equations f
or determining chlorophylls a, b, c1, c2 in higher plants, algae and
natural phytoplankton. Biochem. Physiol. Pflanzen. 167: 191-194
Jensen NG, 1985. The pennate diatoms. A translation of Hustedt' s "Die Kiesela
lgen, 2. Teil." Koeltz Sci. Books, Koenigstein. 918 pp.
Jørgensen, B. B., 1977. The sulfur cycle of coastal marine sediment (Limfjord
en, Denmark). Limnol. Oceanogr. 22(3): 814-832.
Kana TM, Darkangelo C, Hunt MD, Oldham JB, Bennett GE, Cornwell. 1994.
Membrane inlet mass spectrometer for rapid high-precision determi
nation of N2, O2, and Ar in environmental water samples. Anal Che
m 66(23):4166-4170
Kang CK, Choy EJ, Paik SK, Park HJ, Lee KS, An S. submitted.Trophic im

- 300 -
portance of algal carbon to macroinvertebrate production in an inter
tidal salt marsh Scirpus triqueter ecosystem. Mar. Ecolo. Prog. Ser.
Kaplan, W.A., Valiel, I and Teal, J. M., 1979. Denitrification in a marsh ecosy
stem. Limnol. Oceanogr. 24. 726-734.
Kentula M. E. and McIntire C. D., 1986. The autecology and production dynam
ics of eelgrass (Zostera marina L.) in Netarts Bay, Oregon. Estuarie
s 9: 188-199
Kikuchi, T., 1981. Some morphological aspects and distribution of four types
of Paraprionospio(Polychaeta: Spionidae) found from adjacent water
s to western part of japan. Bull. Nansei Reg. Fish. res. Lab., 13: 4
1~58.
Kim GB et al., 2000. Marine Biology 137, 675-682
Knowles, R., 1990. Acetylene inhibitions technique: Development, advantage, and p
otential problems. 1151-1166. In N.P. Revsbech & Sørensen, J. [eds.], D
enitrification in soils and sediment. pp. FEMS Symp. No. 56, Plenum P
ress.
Koike, I. and A. Hattori., 1979. Estimates of denitrification in sediments of Be
ring Sea shelf. Deep-Sea Res. 26:409-15.
Lamontagne, M.G. and I. Valiela., 1995. Denitrification measurement by a dire
ct N2 flux method in sediment of Waquoit Bay, MA. Biogeochem.
31(2): 63-83.
Leach, J. H., 1970. Epibenthic algal production in an intertidal mudflat. Limn
ology and Oceanography 15: 514-521.
LeBris, H., 1988. Fonctionnement des ecosystemes benthiques Cotiers au co
ntact d' estuaries : la rade de Lorient et la baie de Vilaine. These
doc., Univ. Bretagne Occidentale Brest.
Lee K-S, Dunton KH, 1996. Production and carbon reserve dynamics of the sea
grass Thalassia testudinum in Corpus Christi Bay, Texas, USA. Ma
r. Ecol. Prog. Ser. 143: 201-210
Lee K-S, Park SR, Kim J-B, 2005. Production dynamics of the eelgrass, Zost
era marina in two bay systems on the south coast of the Korean
peninsula. Mar. Biol. 147: 1091-1108
Lee, H. Y., 2003. Studies on the distribution of the microalgae in the tidal flat
s of Gamami beach, Young-Gwang, Korea. Journal of the Environ
mental sciences 12:715-724.
Lee, J.G, Nishijima, W., Mukai, T., Takimoto, K., Seiki, T., Hiraoka, K., and
Okada, M., 1998. Factors to determine the functions and structure i

- 301 -
n natural and constructed tidal flats. Wat. Res. 32(9) 2601-2606.
Lorenzen, C. J., 1967. Determination of chlorophyll and pheopigments: Spectrophoto
metric equations. Limnology and Oceanography 12, 343-346
Lowry, O. M., N. I. Roseborough, A. L. Farrand & R. J. Randall. 1951. Protei
n measurement with the folin phenol reagent. J. Biol. Chem. 193:26
3-275.
MacIntyre, H. L., Geider, R. J., Miller, D. C., 1996. Microphytobenthos: the ecolo
gical role of the "Secret Garden" of unvegetated, shallow-water mari
ne habitats. I. Distribution, abundance and primary production. Estuar
ies 19: 186-201
Mann, KH., 1982. Ecology of coastal waters. A system approach. Blackwell Scie
ntific Publication. Oxford. 18-22.
Marsh, J. B. & D. B. Weinstein., 1966. Simple charring method for determinat
ion of lipid. J. Lipid. Res. 7:574-576.
McLusky, D. S., 1981. The Estuarine Ecosystem. Blackie and Son Limited, Glasgow.
215pp.
Miller, D. C., Geider, R.J., MacIntyre, H. L., 1996. Microphytobenthos: the ecol
ogical role of the "Secret Garden" of unvegetated, shallow-water m
arine habitats. II. Role in sediment stability and shallow-water food
web. Estuaries 19: 202-212
Miyoshi, k., Shimati, T, Kimura, K., 1990 A capacity of purification at construc
ted tidal flat. Tokyo Metropolitan Reserach Institute of Environment
al Science (Annual report). 120-125.
Nacci DE et al., 1996. Aquatic Toxicology 35, 197-210
Nielson LP., 1992. Denitrification in sediment determined from nitrogen isotope p
airing. FEMS Microb Ecol 86:357-362
Nishimura, S. (ed.)., 1992. Guide to seashore animals of Japan with color pict
ures and keys, Vol. I. Hoikusha, Tokyo, 663pp.
Nybakken JW, 2001. Marine biology. An ecological approach. Harpper & Row. New
York.
Odum WE. and Heald EJ., 1975. The detritus-based food web of an estuarine
mangrove community. In Estuarine Research. L.E. Cronin(Ed.), Vol
1. pp. 265-286. New York: Academic Press.
Oh, S.J., Moon, C. H., Park, M.O., 2004. HPLC Analysis of Biomass and Com
munity Composition of Microphytobenthos in the Saemankeum Tidal
flat, West Coast of Korea. Journal of the Korean Fisheries Society
37: 215-225

- 302 -
Okutani, T. (ed.)., 2000. Marine mollusks in japan. Tokai Univ. Pre., Tokyo, 1
173pp.
Okutani, T., 1994. Field books, Vol. 8. Yama-kei Publishers Co., Ltd., Tokyo, 3
67pp.
Orth RJ, Morre KA, 1986. Seasonal and year-to-year variations in the growt
h of Zostera marina L. (eelgrass) in the lower Chesapeake Bay. A
quat. Bot. 24: 335-341.
Parsons T. R., Maita, Y. and Lalli C.M., 1984. Annual of chemical and biologi
cal methods for seawater analysis. Pergamon Press, New York.
Pearson, T. H. and R. Rosenberg, 1978. Macrobenthic succession in relation to
organic enrichment and pollution of the marine environment. Ocean
ogr. mar. Biol. Ann. Rev., 16: 229-311.
Pielou, E. C., 1975. Ecological Diversity. Wiley, New York.
Pinckney, J., D. Millie, B. Vinyard and H. Paerl, 1997. Environmental controls
of phytoplankton bloom dynamics in the Neuse River Estuary, NC,
USA. Canadian Journal of Fisheris and Aquatic science., 54: 2491-2
501.
Platell, M. E. and I. C. Potter, 1996. Influence of water depth, season, habitat a
nd estuary location on the macrobenthic fauna of a seasonality close
d estuary. J. Mar. Biol. Ass. U. K., 76: 1~21.
Platt T., Gallegos C. and Harrison W. G., 1980. Photoinhibition of photosynthe
sis in natural assemblage of marine phytoplankton. J. Mar. Res. 38:
687-701.
Pomeroy, L. R., 1959. Algal productivity in salt marshes of Georgia. Limnolog
y and Oceanography 4: 386-397.
Pritchard, D. W., 1967. What is an estuary : Physical viewpoint. In: Lauff, G. H.
(ed.). Estuaries. Amer. Assoc. Adv. Sci. Pub. 83, Washinton D. C., 3~
5.
Probert, P. K., 1984. Disturbance, sediment stability, and trophic structure of soft-
bottom communities. J. Mar. Res., 42: 893~921.
Ralph P. J., Gademann R. and Dennison W. C., 1998. In situ seagrass photosy
nthesis measured using a submersible, pulse-amplitude modulated fl
uorometer. Mar Biol 132: 367-373
Remane, A. and C. Schlieper., 1958. Die biologie des brackwassers, E.Schwiezerbar
t'sche verlagsbuchhandlung, Stuttgart., 348pp.
Revsbech, N.P., Jørgensen, B.B., 1983. Photosynthesis of benthic microflora mea
sured with high spatial resolution by the oxygen microprofile metho

- 303 -
d: capabilities and limitation of the method. Limnology and Oceanogr
aphy 28: 749-756.
Revsbech, N.P., Jørgensen, B. B., 1986. Microelectrodes: their use in microbial ec
ology. Advanced Microbial Ecology 9: 293-352.
Revsbech, N. P., Jørgensen, B. B. Brix, O., 1981. Primary production of micro
algae in sediments measured by oxygen microprofile, H14CO3- fixa
tion and oxygen exchange methods. Limnology and Oceanography
26: 717-730.
Revsbech, N. P., Ward, D. M., 1983. Oxygen microelectrode that is insensitive
to medium chemical composition: use in an acid microbial mat domi
nated by Cyanidium caldarium. Applied Environmental Microbiology
45: 755-759.
Rhoads, D. C. and D. K. Young, 1970. The influence of deposit-feeding organi
sms on the sediment stability and community trophic structure. J.
Mar. Res., 28: 150~178.
Roberts, R. D., M. R. Greory, and B. A. Foster, 1998. Developing an efficient macrof
auna monitoring index from an impact study - a dredge spoil example.
Mar. Pollut. Bull., 36: 231-235.
Round FE, Crawfoord RM and Mann DG, 1990. The diatoms. Biology and morp
hology of the genera. Cambridge Univ. Press, Cambridge. 747pp.
Rowe, G. T., C. H. Clifford, J. K. Smith Jr. and P. L. Hamilton, 1975. Benthic nu
trient regeneration and its coupling to primary productivity in coastal
waters. Nature. 255: 215-217.
Ryther J. M. and W. M. Dunston. 1971. Nitrogen, phosphorus and eutrophicati
on in the coastal marine environment. Science. 171:1008-1013
Santos, S. L. and J. L. Simon, 1980. Marine soft-bottom community establishmen
t following annual defaunation: larval or adult recruitment? Mar. Ecol.
Prog. Ser., 2: 235-241.
Sato N., Koh C.-H., 2004. Biological richness of the Asian tidal flats and its cris
is by human impacts, In: Hong SK, Lee JA, Ihm BS, Farina A, Son
YH, Kim ES, Choi JC (eds) Ecological issues in a changing world-St
atus, response, and strategy. Kluwer Academic Publishers, Dordrecht,
the Netherlands
Schaffiner, L. C., R. J. Diaz, C. R. Olson and I. L. Larsen, 1987. Faunal characte
ristics and sediment accumulation processes in the James River Estua
ry, Virginia. Estuar. Coast. Shelf Sci., 25: 211~226.
Schreiber U., Gademann R., Ralph P.J. and Larkum, A.W.D., 1997. Assessment

- 304 -
of photosynthetic performance of prechloron in Lissoclinum patella in
hospite by chlorophyll fluorescence measurements. Pl. Cell Physiol.
Tokyo 38: 945-951.
Schubel, J. R., 1984. The estuary as a filter : an introduction. In : The estuar
y as a filter. Ed. by V. S. Kennedy, Academic Press., 1~11.
Seitzinger, S. P., L. P. Nielson, J.M. Caffrey and P.B. Christen., 1994. Denitrifi
cation measurements in aquatic sediments: A comparison of three
methods. Biogeochem. 23: 147-167.
Seitzinger, S. P., 1990. Denitrification in aquatic sediments. 301-322. In Revsb
ech, N. P. and Sørensen, J. [eds.], Denitrification in soil and sedime
nt. FEMS Symposium No. 56. Plenum Press
Seitzinger, S. P., 1988. Denitrification in freshwater and coastal marine ecosys
tem: Ecological and geochemical significance. Limnol. Oceanogr. 33:
702-724.
Seitzinger, S. P., 1990. Denitrification in aquatic sediments. In: FEMS Sympos
ium on Denitrification in soil and sediment edited by Revsbech, N.
P. and J. Sorensen, Plenum, 56: 301-322.
Seitzinger, S. P., S. W. Nixon, M. E. Q. Pilson and S. Burke., 1980. Denitrific
ation and nitrous oxide production in near shore marine sediments.
Geochimica Cosmochimica Acta. 44:1853-1860.
Shannon, C. E. and W. Weaver, 1949. The mathematical theory of communica
tion Univ. Illinois Press, Urbana, Illinoise, 125pp.
Somerfield, P. J. and K. R. Clarke, 1995. Taxonomic levels in marine commun
ity studies revisited. Mar. Ecol. Prog. Ser., 127: 113-119.
Steinert SA et al., 1998. Marine Environmental Research 46, 355-358
Sullivan M and Moncreiff C, 1988. Primary production of euphytic algal comm
unities in a Mississippi salt marsh. J. phycol. 24:49~58.
Sullivan MJ, 1975. Diatom communities from a Delaware salt marsh. J. Phycol.11:38
4-390.
Teal, J.M., 1962. Energy flow in the salt marsh ecosystem of Georgia. Ecolog
y 43:614~624.
Thouzeau, G., Robert, G., Ugarte, R., 1991. Faunal assemblages of benthic m
egainvertebrates inhabiting sea scallop grounds from eastern G
eorges Bank, in relation to environmental factors. Mar. Ecol.
Prog. Ser. 74: 6 1~82.
Tiedje, J. M., S. Simkins and P. M. Groffman., 1989. Perspectives on measure
ment of denitrification in the field including recommended protocols

- 305 -
for acetylenebased methods. Plant Soil. 115: 261-284.
Tomas CR, 1996. Identifying marine Diatoms and dinoflagellates. Academic Pr
ess, San Diego. 598pp.
van der Maarel, E., 1990. Ecotones and ecoclines are different. J. Veg. Sci. 1: 13
5~138.
Ward, J. Q., 1978. The infaunal trophic index. In: Coastal water research Proj
ect. Annual Report, pp. 19-39.
Warwick RM and Price R, 1975. Macrofauna production in an estuarine mud-
flat. J. Mar. Biol. Ass. U.K.,55:1-18.
Warwick, R.M., 1988. The level of taxonomic discrimination required to detect
pollution effects on marine benthic community. Mar. Pollut. Bull., 1
9: 259-268.
Wollast R. 1993. Interactions of carbon and nitrogen cycles in the coastal zone. 19
5-210. In Wollast R., F. T. Mackenzie and L. Chou [eds.], Interactions
of C, N, P and biogeochemical cycles and global change. Springer-Verl
ag.
Yamaji I, 1984. Illustrations of the marine plankton of Japan. 3rd ed. Hoiku
sha Publ. Co. Ltd., Osaka. 537pp. (in Japanese)
Yoo, M.H., Choi, J.K. 2005. Seasonal distribution and primary produciton of mi
crophytobenthos on an intertidal mud flat of the Janghwa in Gangh
wa Island, Korea. The Sea, Journal of the Korean Society of Ocean
ography 10: 8-18
Zieman J. C., 1974. Methods for the study of the growth and production of t
urtle grass, Thalassia testudinum König. Aquaculture 4: 139-14.

- 306 -
주 의
1. 이 보고서는 환경부에서 시행한 차세대 핵심환경기
술개발 사업의 연구보고서입니다.
2. 이 보고서 내용을 발표할 때에는 반드시 환경부에서
시행한 차세대 핵심환경기술개발의 연구개발결과임을
밝혀야 합니다.
3. 국가과학기술 기밀유지에 필요한 내용은 대외적으
로 발표 또는 공개하여서는 아니됩니다.